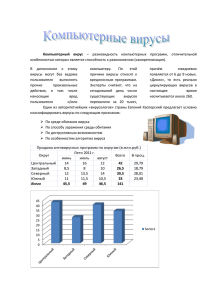
2 Содержание Введение…………………………………………………………………..………..3 Основная часть……………………………………………………………….……4 Вывод………………………………………………………………..…………….17 Список литературы…………………………………………………………….....18 3 Введение Согласно одному из определений вирусы представляют собой форму жизни, согласно другому вирусы являются комплексами органических молекул, взаимодействующими с живыми организмами. Вирусы характеризуют как «организмы на границе живого». Вирусы похожи на живые организмы в том, что они имеют свой набор генов и эволюционируют путём естественного отбора, и кроме этого в том, что способны размножаться, создавая собственные копии путём самосборки. Вирусы имеют генетический материал, в тоже время лишены клеточного строения, а именно эту черту обычно рассматривают как фундаментальное свойство живой материи. У вирусов нет собственного обмена веществ, и для синтеза собственных молекул им необходима клетка-хозяин. По этой причине они не способны размножаться вне клетки. В то же время такие бактерии, как риккетсии и хламидии, несмотря на то, что не могут размножаться вне клеток хозяина, считаются живыми организмами. Общепризнанные формы жизни размножаются делением клетки, в то время как вирусные частицы самопроизвольно собираются в инфицированной клетке. От роста кристаллов размножение вирусов отличается тем, что вирусы наследуют мутации и находятся под давлением естественного отбора. Самосборка вирусных частиц в клетке даёт дополнительное подтверждение гипотезы, что жизнь могла зародиться в виде самособирающихся органических молекул. Опубликованные в 2013 году данные о том, что некоторые бактериофаги обладают собственной иммунной системой, способной к адаптации, являются дополнительным доводом в пользу определения вируса как формы жизни. 4 Вирусы – это особая форма жизни, объединяющая организмы с неклеточным строением. Вирусы способны существовать в двух формах: вне клеток и внутри клеток. Вне клеток существуют свободные вирусы – вирионы. Вирионы не проявляют свойств биологических систем: у них отсутствует обмен веществ, и они неспособны к самовоспроизведению. Вирионы состоят из нуклеиновых кислот (ДНК или РНК), заключенных в белковую оболочку – капсид. В состав капсида входит строго определенное количество повторяющихся белковых субъединиц – капсомеров. Например, у вируса полиомиелита в состав капсида входит 60 капсомеров, у аденовируса – 252, у вируса табачной мозаики – 2000. Размеры вирусов колеблются от 20 до 350 нм. По морфологии различают следующие формы вирусов: сферическую, палочковидную, кубоидальную, сперматозоидную. По характеру симметрии капсида различают вирусы со спиральным, кубическим (икосаэдрическим) и комбинированным типом симметрии. Степень сложности вириона может быть различной. У простых вирусов в состав вириона входит только нуклеиновая кислота и белки, которые связаны в единую нуклеопротеиновую структуру – нуклеокапсид. У сложных вирусов имеется дополнительная липопротеиновая оболочка – суперкапсид. В состав сложных вирионов могут входить углеводы и некоторые ферменты. Однако вирусы никогда не содержат метаболических систем, обеспечивающих обмен веществ. Для собственного воспроизведения вирусы должны проникнуть в клетку. Поверхность капсида включает химические вещества, способствующие прикреплению вириона к поверхности клетки и проникновению всего вириона или нуклеиновой кислоты в клетку. Сначала происходит адсорбция (фиксация) вирионов на поверхности клетки, а затем внутрь клетки проникает или весь вирион или только вирусная нуклеиновая кислота. В большинстве случаев вирусы проникают в клетку путем виропексиса (этот механизм проникновения вирусов в 5 клетку сходен с фагоцитозом). Organism type Virus Virus Virus Virus Bacterium Bacterium Bacterium – cyanobacterium Amoeboid Plant Plant Fungus – yeast Fungus Nematode Nematode Insect Insect Mammal Mammal Fish Organism Genome size (base pairs) 3,569 9,749 48,502 1,259,197 4,600,000 9,970,000 Bacteriophage MS2 HIV Phage λ Megavirus Escherichia coli Solibacter usitatus (strain Ellin 6076) Nostoc punctiforme 3.5kb 9.7kb 48kb 1.3Mb 4.6Mb 10Mb 9,000,000 9Mb Polychaos dubium ("Amoeba" dubia) Arabidopsis thaliana Fritillaria assyrica Saccharomyces cerevisiae Aspergillus nidulans Pratylenchus coffeae Caenorhabditis elegans Drosophila melanogaster (fruit fly) Apis mellifera (honey bee) Mus musculus Homo sapiens Protopterus aethiopicus (marbled lungfish) 670,000,000,000 670Gb 157,000,000 130,000,000,000 12,100,000 30,000,000 20,000,000 100,300,000 130,000,000 157Mb 130Gb 12.1Mb 30Mb 20Mb 100Mb 130Mb 236,000,000 2,700,000,000 3,200,000,000 130,000,000,000 236Mb 2.7Gb 3.2Gb 130Gb Как и любое чужеродное вещество, нуклеиновые кислоты вирусов, проникшие в клетку, должны разрушаться защитными системами клетки. Основу защитных систем составляют нуклеазы – ферменты, расщепляющие нуклеиновые кислоты. Однако существует множество способов защиты вирусных нуклеиновых кислот от разрушения ферментными системами клетки. Например, у крупных ДНК-содержащих бактериофагов в состав вирусной ДНК входят модифицированные основания: окисленные и метилированные. Такая модифицированная ДНК не разрушается клеточными нуклеазами. После проникновения в клетку вирусы вступают в вегетативно- репродуктивную фазу, то есть приобретают способность к обмену веществ и воспроизведению, причем, метаболизм вирусов неразрывно связан с 6 метаболизмом клетки-хозяина. Таким образом, вирусы являются облигатными (обязательными) специализированными внутриклеточными паразитами на молекулярно-генетическом уровне. Многие вирусы приступают к репродукции сразу же поле проникновения в цитоплазму клетки-хозяина. Однако в ряде случаев нуклеиновые кислоты вирусов встраиваются (интегрируются) в состав хромосом хозяина. В интегрированном состоянии вирус называется провирусом. Провирусы неотличимы от генетического материала хозяина и воспроизводится вместе с ним. В интегрированном (вирогенном) состоянии вирусы могут находиться долгое время. Но в ряде случаев (при изменении физиологического состояния клетки, например, при облучении) начинается репродукция вируса. С помощью ферментов и пластических веществ клетки идет репликация вирусных нуклеиновых кислот и вирусных белков. Путем самосборки из этих молекул формируется множество вирионов, которые покидают клетку. При этом клетка может погибнуть или сохраниться. Жизненный цикл Вирусы не размножаются клеточным делением, поскольку не имеют клеточного строения. Вместо этого они используют ресурсы клетки-хозяина для образования множественных копий самих себя, и их сборка происходит внутри клетки. Условно жизненный цикл вируса можно разбить на несколько взаимоперекрывающихся этапов (обычно выделяют 6 этапов): Прикрепление представляет собой образование специфичной связи между белками вирусного капсида и рецепторами на поверхности клетки-хозяина. Это специфичное связывание определяет круг хозяев вируса. К примеру, ВИЧ поражает только определённый тип человеческих лейкоцитов. Это объясняется тем, что оболочечный гликопротеин вируса gp120 специфично связывается с молекулой CD4 — хемокиновым рецептором, который обычно встречается на поверхности CD4-положительных T-лимфоцитов. Этот механизм обеспечивает инфицирование вирусом только тех клеток, которые способны осуществить его 7 репликацию. Связывание с рецептором может вызвать конформационные изменения белка оболочки (или белка капсида в случае безоболочечного вируса), что в свою очередь служит сигналом к слиянию вирусной и клеточной мембран и проникновению вируса в клетку. Проникновение в клетку. На следующем этапе вирусу нужно доставить внутрь клетки свой генетический материал. Некоторые вирусы также переносят внутрь клетки собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза и попадают в кислую среду лизосом, где происходит депротеинизация вирусной частицы, после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различают по тому, где происходит их репликация: часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре. Процесс инфицирования вирусами клеток грибов и растений отличается от инфицирования клеток животных. Растения имеют прочную клеточную стенку, состоящую из целлюлозы, а грибы — из хитина, так что большая часть вирусов могут проникнуть в них только после повреждения клеточной стенки. Однако почти все вирусы растений (включая вирус табачной мозаики) могут перемещаться из клетки в клетку в форме одноцепочечных нуклеопротеиновых комплексов через плазмодесмы. Бактерии, как и растения, имеют крепкую клеточную стенку, которую вирусу, чтобы попасть внутрь, приходится повредить. Но в связи с тем, что клеточная стенка бактерий намного тоньше таковой у растений, некоторые вирусы выработали механизм впрыскивания генома в бактериальную клетку через толщу клеточной стенки, при котором капсид остаётся снаружи. Лишение оболочек представляет собой процесс потери капсида. Это достигается при помощи вирусных ферментов или ферментов клетки-хозяина, а 8 может быть и результатом простой диссоциации. В конечном счёте вирусная геномная нуклеиновая кислота освобождается. Репликация вирусов подразумевает, прежде всего, репликацию генома. Репликация вируса включает синтез мРНК ранних генов вируса (с исключениями для вирусов, содержащих положительную РНК), синтез вирусных белков, возможно, сборку сложных белков и репликацию вирусного генома, которая запускается после активации ранних или регуляторных генов. Вслед за этим может последовать (у комплексных вирусов с крупными геномами) ещё один или несколько кругов дополнительного синтеза мРНК: «поздняя» экспрессия генов приводит к синтезу структурных или вирионных белков. Вслед за этим происходит самосборка вирусных частиц, позже происходят некоторые модификации белков. У вирусов, таких как ВИЧ, такая модификация (в некоторых случаях называемая созреванием) происходит после выхода вируса из клетки-хозяина. Выход из клетки. Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки, если такая есть. Эта особенность есть у многих бактериальных и некоторых животных вирусов. Некоторые вирусы подвергаются лизогенному циклу, где вирусный геном включается путём генетической рекомбинации в специальное место хромосомы клетки-хозяйки. Тогда вирусный геном называется провирусом, или, в случае бактериофага, профагом. Когда клетка делится, вирусный геном также удваивается. В пределах клетки вирус в основном не проявляет себя; в тоже время в некоторый момент провирус или профаг может вызвать активацию вируса, который может вызвать лизис клеток-хозяев. Быстро размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы, в том числе ВИЧ, обычно отделяются от клетки путём отпочковывания. В ходе этого процесса вирус обзаводится своей оболочкой, которая представляет собой модифицированный фрагмент клеточной мембраны хозяина или другой внутренней мембраны. Таким образом, клетка может продолжать жить и продуцировать вирус. 9 Особенности жизненного цикла различных групп Генетический материал внутри вирусных частиц и способ его репликации, значительно отличается у различных вирусов. ДНК-содержащие вирусы. Репликация генома у большинства ДНК- содержащих вирусов происходит в клеточном ядре. Если клетка имеет соответствующий рецептор на своей поверхности, эти вирусы проникают в клетку либо путём непосредственного слияния с клеточной мембраной (напр. герпесвирусы), либо — что бывает чаще — путём рецептор-зависимого эндоцитоза. Большинство ДНК-содержащих вирусов полностью полагаются на синтетический аппарат клетки-хозяина для производства их ДНК и РНК, а также последующего процессинга РНК. Однако вирусы с крупными геномами могут сами кодировать большую часть необходимых для этого белков. Геному вируса эукариот нужно преодолеть ядерную мембрану для того, чтобы получить доступ к ферментам, синтезирующим ДНК и РНК, в случае же бактерифагов ему достаточно просто проникнуть в клетку. РНК-содержащие вирусы. Репликация таких вирусов обычно происходит в цитоплазме. РНК-содержащие вирусы можно подразделить на 4 группы в зависимости от способа их репликации. Механизм репликации определяется тем, является ли геном вируса одноцепочечным или двухцепочечным, вторым важным фактором в случае одноцепочечного генома является его полярность (может ли он непосредственно служить матрицей для синтеза белка рибосомами). Все РНК-вирусы используют собственную РНКрепликазу для копирования своих геномов. Вирусы, использующие обратную транскрипцию. Эти вирусы содержат одноцепочечную РНК (Retroviridae, Metaviridae, Pseudoviridae) или двухцепочечную ДНК (Caulimoviridae и Hepadnaviridae). РНК-содержащие вирусы, способные к обратной транскрипции (ретровирусы, например, ВИЧ), используют ДНК-копию генома как промежуточную молекулу при репликации, а содержащие ДНК (параретровирусы, например, вирус гепатита B) — РНК. В обоих случаях используется обратная транскриптаза, или РНК-зависимая-ДНК- 10 полимераза. Ретровирусы встраивают ДНК, образующуюся в процессе обратной транскрипции, в геном хозяина, такое состояние вируса называется провирусом. Параретровирусы же этого не делают, не смотря на то, что встроенные копии их генома могут давать начало инфекционным вирусам, особенно у растений. Вирусы, использующие обратную транскрипцию, восприимчивы к противововирусным препаратам, ингибирующим обратную транскриптазу, в том числе к зидовудину и ламивудину. Геном вирусов Геном вирусов включает: – Структурные гены, которые кодируют белки. Занимают примерно 95 % вирусной хромосомы. Белки вирусов можно разделить на несколько групп: структурные, ферменты, регуляторы. – Регуляторные последовательности, которые не кодируют белки: промоторы, операторы и терминаторы. – Прочие некодирующие участки (сайты), в том числе: участок attP, обеспечивающий интеграцию вирусной хромосомы в хромосому клетки–хозяина; участки cos – липкие концевые участки линейных вирусных хромосом, обеспечивающие замыкание линейной хромосомы в кольцевую форму. Гены, кодирующие рРНК и тРНК, в геноме вирусов обычно отсутствуют. Однако в геноме крупного фага Т4 имеются гены, кодирующие несколько тРНК. Геном вирусов отличается высокой плотности упаковки информации. Например, у фага φХ174 в пределах одного гена может располагаться еще один ген. В частности, ген В находится в пределах гена А, а ген Е – в пределах гена D. У мелкого РНК-содержащего фага f2 ген регуляторного белка, блокирующего лизис (созревание вирионов и разрушение клетки), перекрывается с двумя другими генами, удаленными друг от друга. Особенности вирусов эукариот У вирусов эукариот обнаружены следующие особенности: 1. Интрон - экзонная структура генов. 11 2. Модификация белков после синтеза полипротеинов: весь геном транскрибируется в виде одной молекулы мРНК, которая служит матрицей для синтеза полипротеина – одного гигантского инертного белка, и лишь затем происходит расщепление полипротеина на белки, выполняющие определенные функции. 3. Перекрывание генов (обезьяний вирус SV 40, вирус гриппа). ДНК-содержащие вирусы К ДНК-содержащим вирусам относятся многие вирусы бактерий – бактериофаги (или просто фаги ). Некоторые мелкие фаги (например, фаг М13) при репродукции не разрушают клетку. Репродукция крупных фагов (например, фага Т–4) приводит к гибели клетки. Фаг Т–4 – это один из наиболее сложно организованных вирусов. Белковый капсид включает не менее 130 белков, образующих головку, воротничок, сократимый хвост, базальную пластинку и хвостовые нити. Такое строение капсида позволяет впрыскивать ДНК в бактериальную клетку через толстую оболочку, поэтому подобные вирусы образно называют «живыми шприцами». Т–фаги могут существовать в виде профага длительное время. К ДНК-содержащим вирусам относятся возбудители многих заболеваний человека и животных: вирусы оспы, герпеса, гепатита В, аденовирусы млекопитающих и человека (вызывают желудочно-кишечные заболевания,ОРВИ, конъюнктивиты), вирусы бородавок человека. К ДНКсодержащим вирусам относятся и некоторые вирусы растений (вирус золотистой мозаики фасоли, вирус мозаики цветной капусты). Некоторые вирусы используются в генной инженерии для переноса генов от одних организмов к другим, например, обезьяний вирус SV 40. Вирионы ДНК-содержащих вирусов содержат ДНК. Объемом ДНК определяется количество белков в вирионе: один полипептид кодируется отрезком ДНК длиной примерно 1 тысяча нуклеотидов (нуклеотидных пар). После проникновения в клетку вирусная ДНК становится матрицей для синтеза ДНК и РНК. Примеры организации генома ДНК-содержащих вирусов 12 1. Кольцевая двухцепочечная ДНК длиной около 5 тпн. – Обезьяний вирус SV 40. Мелкий эукариотический вирус. Вирионы в виде икосаэдра. Капсид белковый. Используется в генной инженерии как вектор переноса генов. Кодирует 5 белков. – Вирусы бородавок человека. 2. Кольцевая одноцепочечная ДНК длиной около 5 тн; может быть как кодирующей, так и антикодирующей. – Мелкие бактериофаги типа М13. Не разрушают клетку. Капсид включает 8 белков. – Вирус золотистой мозаики фасоли. 3. Линейная двухцепочечная ДНК длиной 30-150 тпн. – Бактериофаги типа Т4. Вирионы крупные. Белковый капсид из 130 белков включает: головку, хвостовой отдел и хвостовые нити. Эти вирусы могут существовать в виде профага длительное время. – Аденовирусы млекопитающих и человека. Вирионы средних размеров в виде икосаэдра. Капсиды белковые. Вызывают ОРВИ, конъюнктивиты, желудочно-кишечные заболевания, иногда обладают онкогенными свойствами. – Вирусы оспы, герпеса и им подобные. Вирионы крупные. Имеется липопротеиновая оболочка. 4. Линейная одноцепочечная ДНК длиной около 5 тн; ДНК может быть как кодирующей, так и антикодирующей. У человека известны как спутники аденовирусов. 5. Двухцепочечная ДНК, замкнутая в кольцо из перекрывающихся сегментов. Длина ДНК – 3-8 тн. – Вирус гепатита В. Вирион сферический, средних размеров. Имеется дополнительная оболочка из вирусных и клеточных белков. Кодирует 5 белков. – Вирус мозаики цветной капусты (CaMV). Промотор 35S-RNA (CaMV35S) этого вируса широко используется в традиционной генной инженерии для создания генетических конструкций. РНК-содержащие вирусы 13 К РНК-содержащим вирусам относятся многие вирусы растений, возбудители заболеваний человека и животных: вирус полиомиелита, вирусы гриппа А, В и С, вирусы паротита (свинки), кори, чумы плотоядных животных (чумки), бешенства, вирус иммунодефицита человека (ВИЧ). В отдельную группу выделяются арбовирусы, которые переносятся членистоногими (клещами, москитами), например, вирусы клещевого энцефалита, желтой лихорадки. Многие РНК-содержащие вирусы вызывают ОРВИ (например, коронавирусы), желудочно-кишечные заболевания (реовирусы птиц, млекопитающих и человека). Некоторые РНК-содержащие вирусы используются в биотехнологии, например, вирусы полиэдроза насекомых. Вирионы РНК-содержащих вирусов содержат РНК. После проникновения в клетку вирусная РНК становится матрицей для синтеза ДНК и РНК. Примеры организации генома РНК-содержащих вирусов 1. Линейная одноцепочечная мРНК (плюс–цепь) длиной около 4 тн; в виде единой молекулы или в виде нескольких разных молекул. Плюс-цепь сразу же может использоваться для трансляции. Вегетативно-репродуктивная фаза этих вирусов протекает в цитоплазме. Вплюс-цепи закодирована РНК-репликаза (РНКзависимая РНК-полимераза). Представители: – Вирус табачной мозаики (ВТМ) – сегментированная РНК. Вирион нитевидный (18х300 нм). ВТМ открыт Д.И. Ивановским в 1982 г. – Вирус полиомиелита – несегментированная РНК. Вирионы мелкие, в виде икосаэдра. Капсид белковый. – Вирус бешенства. Нитевидный вирион. Имеется дополнительная липопротеиновая оболочка. – Арбовирусы (переносятся членистоногими: клещами, москитами) – вирусы клещевого энцефалита, желтой лихорадки. Морфология и размеры вирионов разнообразны, например, вирус энцефалита содержит 9 белков. Имеется дополнительная липопротеиновая оболочка. – Мелкие бактериофаги (с несегментированной РНК). 2. Линейная одноцепочечная кРНК (минус–цепь, порядок нуклеотидов 14 комплементарен по отношению к мРНК). Минус–цепь не может служить для трансляции и используется как матрица для синтеза плюс–цепи. Плюс-цепь служит для трансляции вирусных белков и используется как матрица для синтеза вирусной кРНК. Вегетативно-репродуктивная фаза этих вирусов также протекает в цитоплазме. – Вирусы гриппа А, В, С. Вирус гриппа А содержит минус-цепь РНК, состоящую из 8 фрагментов. Фрагменты РНК связаны с вирусными белками и образуют спиральный нуклеокапсид. Поверх нуклеокапсида располагается гликолипопротеиновый суперкапсид. В составе вириона 10 белков. В состав суперкапсида входит два белка, определяющих антигенные свойства вируса: гемагглютинин и нейраминидаза. Кроме того, в состав вириона входит уже готовая РНК-репликаза, обеспечивающая синтез плюс-цепи на матрице минусцепи. – Вирусы паротита (свинки), кори, чумы плотоядных животных (чумки). Сферический вирион средних размеров. Имеется дополнительная липопротеиновая оболочка. 3. Линейная двухцепочечная РНК – Мелкие бактериофаги. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. – Вирусы полиэдроза насекомых. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Используются в биотехнологии (для синтеза интерферона). – Реовирусы птиц, млекопитающих и человека. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. ВызываютОРВИ, желудочно-кишечные заболевания. РНК фрагментированная (10...11 фрагментов), кодирует 11 белков. 4. Две линейные одноцепочечные одинаковые молекулы мРНК длиной около 10 тн. Ретровирусы. Способны интегрироваться в ДНК.В состав вирионов входит фермент обратная транскриптаза (ревертаза). Имеется дополнительная липопротеиновая оболочка. Многиеретровирусы вызывают онкологические 15 заболевания: лейкозы, саркомы, опухоли молочных желез. К ретровирусам относится и вирус иммунодефицита человека, вызывающий СПИД. – Вирус иммунодефицита человека (ВИЧ). Содержит одну плюс-цепь РНК, кодирует 13 белков. липопротеиновая Сферический оболочка, вирион. включающая Имеется фрагменты дополнительная мембран человека. Избирательно поражает Т–лимфоциты. Роль вирусов В настоящее время известно свыше 800 видов вирусов (вероятно, миллионы видов еще не открыты). Вирусы классифицируются по носителям наследственной информации (ДНК-содержащие и РНК-содержащие) и по хозяевам (вирусы растений, вирусы грибов, вирусы животных и вирусы прокариот, или бактериофаги). Биномиальная номенклатура в вирусологии не привилась, и обычно каждый вид вируса получает собственное имя. Вирусы – возбудители многих инфекционных заболеваний растений, животных и человека. В то же время, вирусы – возбудители заболеваний у нежелательных для человека организмов («враги наших врагов»). Вирусы широко используются как объекты молекулярно-генетических исследований. В генной инженерии вирусы применяются для создания генетических конструкций и для переноса генетического материала. Происхождение вирусов. Существует ряд теорий происхождения вирусов. Согласно одной из теорий, вирусы – крайне упрощенные прокариотические организмы, утратившие цитоплазму. Противоположные теории рассматривают вирусы как часть генетического материала клетки, вынесенного за ее пределы. Биологическое значение вирусов в первую очередь связывается с их патогенностью – способностью вызывать заболевания. Различают острые вирусные заболевания (например, грипп), хронические и латентные (скрытые). Борьба с вирусными заболеваниями человека и животных ведется с использованием неспецифических препаратов (например, интерферона), специфических сывороток и препаратов, подавляющих репродукцию вирусов. Для профилактики вирусных заболеваний применяют различные вакцины. 16 Антибактериальные препараты (сульфаниламиды, антибиотики) на вирусы не действуют. существуют инфекционные агенты, которые не являются вирусами. К одной из этих групп относятся вироиды – мелкие одноцепочечные кольцевые молекулы РНК длиной до 300 нуклеотидов, не кодирующие собственных белков (не имеющие собственного капсида). Вироиды вызывают многие болезни растений, например, веретеновидность клубней картофеля. К вироидам близки вирусоиды. Это молекулы РНК, которые способны к самовоспроизведению в присутствии вируса-помощника. Вирусоиды также вызывают заболевания растений, например, преходящую полосатость люцерны. К другой группе инфекционных агентов относятся регуляторные белки прионы. Прионы включают те гены, которые не должны функционировать в данное время и в данной ткани. Это нарушает нормальное функционирование клеток, что приводит к развитию тяжелых заболеваний, например, энцефалопатии («бешенство коров»). Некоторые вирусы (аденовирусы, ретровирусы) могут нарушать нормальное функционирование генетического аппарата клетки хозяина, что приводит к развитию онкологических заболеваний. 17 Вывод По определению Х. Френкель-Конрата, "вирусы - это частицы, состоящие из одной или нескольких молекул ДНК или РНК, окруженных белковой оболочкой; вирусы способны передавать свои нуклеиновые кислоты от одной клетки-хозяина к другой и использовать ее ферментативный аппарат для осуществления своей внутриклеточной репликации путем наложения собственной информации на информацию клетки-хозяина. Иногда вирусы могут обратимо включать свой геном в геном хозяина (интеграция), и тогда они либо ведут "скрытое существование", либо так или иначе трансформируют свойства клетки-хозяина". Вирусы, несомненно, являются самыми многочисленными биологическими объектами на Земле, и по этому показателю они превосходят все организмы, вместе взятые. Они поражают все формы клеточных организмов, включая животных, растения, бактерии и грибы. Тем не менее различные типы вирусов могут поражать только ограниченный круг хозяев, многие вирусы видоспецифичны. Некоторые, как, например, вирус оспы, могут поражать только один вид — людей, в подобных случаях говорят, что вирус имеет узкий диапазон хозяев. Напротив, вирус бешенства может поражать различные виды млекопитающих, то есть он имеет широкий диапазон хозяев. Вирусы растений безвредны для животных, а большинство вирусов животных безопасны для человека. Диапазон хозяев некоторых бактериофагов ограничивается одним штаммом бактерий, и они могут использоваться для определения штаммов, вызывающих типирования. вспышки инфекционных заболеваний методом фагового 18 Список литературы 1. Агол В.И., Атабеков И.Г., Крылов В.Н., Тихоненко Т.И. Молекулярная биология вирусов. Изд-во « Наука», 1971. 2. Лурия С., Дарнелл Дж. и др. Общая вирусология. Москва, Мир. 1981 3. Френкель-Конрат Х. Химия и биология вирусов. М.: Мир, 1972