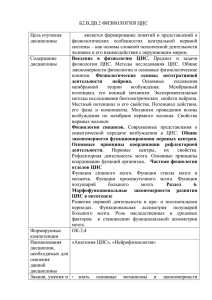
В.М. Смирнов, А.В. Смирнов ФИЗИОЛОГИЯ Учебник Рекомендовано Учебно-методическим объединением по классическому университетскому образованию в качестве учебника для студентов высших учебных заведений, обучающихся по направлению 06.03.01 «Биология» и смежным направлениям и специальностям Медицинское информационное агентство Москва 2016 УДК 612(075.8) ББК 28.707+28.073я73 С50 Рецензенты: Дьяконова И.Н. — профессор, заведующая учебной частью кафедры физиологии РНИМУ им. Н.И. Пирогова. Трошин В.И. — профессор, заведующий кафедрой физиологии Российского университета дружбы народов. Смирнов, В.М. Физиология : учебник / В.М. Смирнов, А.В. Смирнов. — МоС50 сква : ООО «Издательство «Медицинское информационное агентство», 2016. — 448 с. : ил. ISBN 978-5-9986-0283-2 Учебник написан в соответствии с действующим Госстандартом для медицинских вузов и отражает многолетний опыт преподавания на кафедре нормальной физиологии РНИМУ им. Н.И. Пирогова. Особое внимание уделено дискуссионным вопросам, в том числе взаимодействию клеток с помощью электрических полей. Прочному усвоению материала способствуют иллюстрации и таблицы. Для студентов и аспирантов медицинских вузов, практикующих врачей. Студентам обеспечит быструю и хорошую подготовку к зачетам и экзамену, будет полезен в лечебной деятельности. УДК 612(075.8) ББК 28.707+28.073я73 ISBN 978-5-9986-0283-2 © Смирнов В.М., Смирнов А.В., 2016 © Оформление. ООО «Издательство «Медицинское информационное агентство», 2016 Все права защищены. Никакая часть данной книги не может быть воспроизведена в какой-либо форме без письменного разрешения владельцев авторских прав. Оглавление Сокращения ..................................................................................8 Предисловие .................................................................................9 Глава 1. Общая физиология возбудимых тканей. Транспорт веществ.....................................................10 1.1. Функции клетки и ее органелл .......................................11 1.2. Транспорт веществ ..........................................................15 1.3. Свойства биологической ткани и ее раздражители ............................................................23 1.4. Потенциалы клетки ........................................................25 1.5. Возбудимость и лабильность. Использование электрического тока в медицине ....................................32 Глава 2. Частная физиология возбудимых тканей ............ 38 2.1. Рецепторы .......................................................................38 2.2. Физиология нервных волокон........................................43 2.3. Физиология нервно-мышечного синапса......................48 2.4. Скелетная мышца ...........................................................52 2.5. Гладкая мышца ................................................................62 Глава 3. Характеристика регуляции функций организма ....................................................................64 3.1. Нервный механизм регуляции........................................64 3.2. Гуморальная и миогенная регуляция..............................67 3.3. Единство и особенности регуляторных механизмов .....................................................................68 3.4. Системный принцип регуляции и его типы ..................70 Глава 4. Физиология центральной нервной системы.........74 4.1. Структурно-функциональная характеристика ЦНС ......................................................74 4.2. Медиаторы и рецепторы ЦНС. Возбуждение нейронов ЦНС ................................................................81 4 Оглавление 4.3. Основные принципы организации деятельности ЦНС ........................................................... 85 4.4. Свойства нервных центров .............................................. 89 4.5. Торможение ЦНС и координационная деятельность ЦНС............................................................ 93 4.6. Интегративная роль нервной системы............................ 99 4.7. Двигательные системы спинного мозга........................ 100 4.8. Двигательные системы ствола мозга ............................. 112 4.9. Интегративные системы ствола мозга .......................... 118 4.10. Двигательные функции мозжечка ............................... 120 4.11. Функции промежуточного мозга ................................ 122 4.12. Функции базальных ядер и лимбической системы ......................................................................... 125 4.13. Нейронная организация новой коры. Функциональные зоны ................................................ 127 4.14. Методы исследования ЦНС ........................................ 135 Глава 5. Вегетативная нервная система ................................ 138 5.1. Общая характеристика ВНС ......................................... 138 5.2. Характеристика отделов ВНС ....................................... 142 5.3. Центры ВНС и их тонус. Трофическое действие нервной системы ............................................................ 150 5.4. Точки действия фармакологических препаратов и системное воздействие ................................................ 154 5.5. Характеристика ВНС детей ........................................... 156 Глава 6. Гормональная регуляция функций организма ...................................................................158 6.1. Характеристика гормонов и регуляция их выработки .................................................................. 158 6.2. Гормоны гипофиза ......................................................... 162 6.3. Функции гормонов щитовидной железы ...................... 165 6.4. Гормоны паращитовидных желез, эпифиза и тимуса. Кальцитриол................................................... 167 6.5. Гормоны поджелудочной железы .................................. 168 6.6. Гормоны надпочечников................................................ 171 6.7. Половые гормоны .......................................................... 173 Глава 7. Система крови ............................................................ 176 7.1. Кровь как внутренняя среда организма........................ 176 7.2. Физиология эритроцитов .............................................. 179 7.3. Физиология лейкоцитов и тромбоцитов ....................... 184 7.4. Группы крови .................................................................. 188 Оглавление 5 7.5. Система ГПФ (гемостаза, противосвертывания и фибринолиза) .............................................................. 191 Глава 8. Система дыхания .......................................................202 8.1. Функции внешнего звена системы дыхания ................203 8.2. Механизм вдоха и выдоха. Расход энергии ...................207 8.3. Объемы и емкости легких (вентиляция) ....................... 211 8.4. Газообмен между альвеолами и кровью организма ...... 213 8.5. Транспорт газов кровью ................................................. 215 8.6. Регуляция дыхания. Защитные дыхательные рефлексы ........................................................................ 219 Глава 9. Система кровообращения и лимфатическая система .......................................224 9.1. Цикл сердечной деятельности .......................................225 9.2. Особенности свойств сердечной мышцы и ее энергетического обеспечения........................................227 9.3. Автоматия сердца ........................................................... 231 9.4. Основные методы исследования деятельности сердца..............................................................................234 9.5. Регуляция деятельности сердца .....................................238 9.6. Гемодинамика .................................................................243 9.7. Регуляция тонуса сосудов ..............................................257 9.8. Регуляция артериального давления............................... 261 9.9. Кровобращение при изменениях положения тела, физическом и эмоциональном напряжении................. 266 9.10. Лимфатическая система ...............................................267 Глава 10. Система пищеварения ............................................272 10.1. Общая характеристика системы пищеварения ...........272 10.2. Пищеварение в полости рта. Акт глотания .................277 10.3. Пищеварение в желудке ...............................................282 10.4. Пищеварение в двенадцатиперстной кишке ..............288 10.5. Физиология печени ......................................................290 10.6. Пищеварение в тонкой кишке .....................................292 10.7. Всасывание питательных веществ...............................295 10.8. Пищеварение в толстой кишке....................................297 Глава 11. Обмен и терморегуляция........................................ 301 11.1. Обмен веществ и питание ............................................ 301 11.2. Обмен энергии в организме ......................................... 314 11.3. Терморегуляция ............................................................ 321 6 Оглавление Глава 12. Выделительная система ..........................................326 12.1. Структурно-функциональная характеристика почки.............................................................................327 12.2. Роль различных отделов нефрона в образовании мочи ..............................................................................330 12.3. Методы исследования выделительной функции почек .............................................................................335 12.4. Регуляция мочеобразования и физиологических показателей организма .................................................337 12.5. Количество, состав и выведение мочи ........................342 12.6. Водно-солевой баланс ..................................................344 Глава 13. Сенсорные системы ................................................346 13.1. Структурно-функциональная характеристика сенсорных систем .........................................................346 13.2. Свойства СС, их значение, критерии для их оценки ...............................................................350 13.3. Кодирование информации в нервной системе ...........354 13.4. Система зрения.............................................................356 13.5. Система слуха ............................................................... 366 13.6. Системы положения тела .............................................370 13.7. Сенсорные системы кожи ...........................................373 13.8. Сенсорные системы внутренней среды организма......................................................................378 13.9. Система вкуса и обоняния ...........................................379 Глава 14. Система боли ............................................................384 14.1. Основные понятия. Классификация боли..................384 14.2. Болевая система............................................................386 14.3. Обезболивающая система ............................................388 Глава 15. Высшая нервная деятельность ..............................392 15.1. Память ..........................................................................392 15.2. Уcловный рефлекс как научение .................................398 15.3. Другие виды научения ..................................................404 15.4. Типы высшей нервной деятельности и темпераменты личности............................................ 409 15.5. Бодрствование и сон. Сновидения. Гипноз................. 411 15.6. Методы исследования ВНД ......................................... 415 Глава 16. Психическая деятельность и поведение ............. 417 16.1. Сознание и мышление ................................................. 417 16.2. Осознаваемая и подсознательная деятельность организма......................................................................420 16.3. Эмоции ......................................................................... 421 Оглавление 7 16.4. Другие виды психической деятельности .....................423 16.5. Особенности сознания и психической деятельности человека .................................................424 16.6. Потребности и мотивация как фактор поведения ......429 16.7. Функциональная система поведенческого акта .........432 16.8. Механизм произвольных движений ............................435 Предметный указатель .............................................................439 Литература ..................................................................................444 Сокращения АД — артериальное давление АДГ — антидиуретический гормон АТФ — аденозинтрифосфат ВНС — вегетативная нервная система ВПСП — возбуждающий постсинаптический потенциал ВСБ — водно-солевой баланс ГАМК — гамма-аминомасляная кислота ГП — генераторный потенциал ГЭБ — гематоэнцефалический барьер ЖКТ — желудочно-кишечный тракт КУД — критический уровень деполяризации ПД — потенциал действия ПКП — потенциал концевой пластинки ПП — потенциал покоя РП — рецепторный потенциал РФ — ретикулярная формация СПР — саркоплазматический ретикулум ТПСП — тормозной постсинаптический потенциал ЦНС — центральная нервная система ЧСС — частота сердечных сокращений ЭТЛ — эластическая тяга легких ЭЭГ — электроэнцефалограмма Предисловие Объем учебника небольшой, но его содержание соответствует программам лечебного, педиатрического и стоматологического факультетов, за исключением профильных вопросов, которые изучаются по кратким дополнительным учебным пособиям. Краткость достигнута благодаря четкости изложения материала, исключению его повторений в различных главах, исключению громоздких, перегруженных информацией рисунков. В учебнике излагаются современные представления по всем разделам физиологии, он содержит много оригинальных (авторских) иллюстраций. Последовательность глав в учебнике такова, что изучение каждого предыдущего раздела способствует лучшему пониманию и усвоению следующего материала. В.М. Смирнов — основной автор и титульный редактор девяти учебников для студентов различных факультетов медицинских, педагогических и спортивных вузов. Вместе со своими учениками он доказал наличие в организме ранее неизвестного серотонинергического отдела вегетативной нервной системы, что подтверждено его дипломом на открытие серотонинергических нервов у млекопитающих. Главная цель авторов и издателей — это обеспечение студентов компактным и недорогим учебником, который они могли бы приобретать в личное пользование, получая при этом возможность не только глубоко изучить предмет в период обучения и быстрее подготовиться к коллоквиуму и экзамену, но и использовать учебник в дальнейшем — в процессе врачебной деятельности. Авторы ГЛАВА 1 Физиология — это научный стержень, на котором держатся все медицинские науки. Клод Бернар Общая физиология возбудимых тканей. Транспорт веществ Физиология (от греч. phisis — природа, 1оgos — наука) — наука о функциях органов и систем организма и механизмах их регуляции. Глубокое знание физиологии обеспечивает успешное изучение других предметов — патологической физиологии, фармакологии, клинических дисциплин. Врач, знающий современную физиологию, может поставить более точный диагноз и понять, что произойдет в организме, если ввести ему то или иное лекарство. Организм животного — это открытая макромолекулярная, саморегулирующаяся, самовосстанавливающаяся и самовоспроизводящаяся с помощью непрерывного обмена веществ и энергии система, способная чувствовать, активно целенаправленно передвигаться и адаптироваться в окружающей среде. Организм существует как единое целое благодаря взаимодействию клеток, органов и систем, которое осуществляется с помощью нейрогуморальной регуляции. Физиологическая система — это наследственно закрепленная совокупность органов и тканей, выполняющих определенную функцию. Число физиологических систем ограничено. К ним относятся: покровная (кожа), нервная, эндокринная, иммунная, сердечно-сосудистая, системы крови, дыхания, пищеварения, выделения, воспроизведения. Функциональная система, согласно П.К. Анохину, — динамическая совокупность отдельных органов и физио- Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 11 логических систем, формирующаяся для достижения полезного для организма приспособительного результата. Функциональные системы поддерживают оптимальные физиологические показатели, обеспечивающие удовлетворение биологических, социальных, бытовых и других потребностей, их количество огромно. Число органов и физиологических систем организма, входящих в те или иные функциональные системы, вариабельно. Например, для восстановления нормальной величины снизившегося артериального давления может оказаться достаточно учащения и усиления сердечных сокращений и сужения кровеносных сосудов с помощью нервной системы. После существенного изменения артериального давления для его восстановления до нормы формируется более сложная функциональная система, включающая не только сердечно-сосудистую и нервную системы, но и системы выделения (почки выделяют больше или меньше воды), эндокринную, системы крови и пищеварения. 1.1. Функции клетки и ее органелл Понятие о клетке. Клетка — структурно-функциональная единица органа (ткани), способная самостоятельно существовать, выполнять специфическую функцию в малом объеме, расти, размножаться, активно реагировать на раздражение. Цитоскелет — совокупность белковых волокон, расположенных по всей цитоплазме (микротрубочки, промежуточные филаменты, актиновые филаменты). Клеточная мембрана — оболочка клетки, образующая замкнутое пространство, содержащее протоплазму. Протоплазма — совокупность всех внутриклеточных элементов (гиалоплазмы, органелл и включений). Цитоплазма — это протоплазма, за исключением ядра. Гиалоплазма (цитозоль) — гомогенная внутренняя среда клетки, содержащая питательные вещества (глюкозу, аминокислоты, белки, фосфолипиды, депо гликогена) и обеспечивающая взаимодействие всех органелл клетки. Ядро — самая большая органелла клетки, выполняет генетическую (хранение и передачу информации в ряду 12 Физиология поколений) и метаболическую функции (управление синтезом белка). У каждой клетки имеются специфические функции, свойственные в основном данному виду клеток, и общие (основные) функции, подобные функциям других клеток. Общие функции клетки обеспечивают жизнедеятельность самой клетки. Среди них можно выделить следующие. 1. Выработка энергии происходит в результате катаболизма — процессов расщепления (сгорания) мономеров, поступающих в клетки из крови, при этом высвобождается энергия. Энергия необходима для обеспечения жизнедеятельности каждой живой клетки и организма в целом. 2. Синтез тканевых и клеточных структур и необходимых для жизнедеятельности соединений. 3. Размножение клеток способствует росту, развитию молодого организма, восстановлению нарушенной части органа или ткани у зрелого организма. 4. Трансмембранный перенос обеспечивает поступление в клетку необходимых ей веществ и выделение из клетки метаболитов и секрецию соединений, используемых другими клетками организма. 5. Рецепторная функция обеспечивает восприятие раздражителей, изменений внешней и внутренней среды организма. 6. Детоксикация продуктов метаболизма реализуется с помощью биохимических превращений. Специфические функции клеток 1. Сократительная функция — характерна для мышечных клеток. 2. Секреторная функция. Клетки эндокринных желез вырабатывают гормоны, а клетки секреторного аппарата желудочно-кишечного тракта — ферменты. Биологически активные вещества синтезируются и другими клетками. 3. Главными функциями нервных клеток являются восприятие, передача сигнала другим клеткам, усвоение и хранение информации. 4. Эритроциты выполняют газообменную функцию. Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 13 5. У клеток соединительной ткани (костная ткань, связ- ки, сухожилия, фасции, строма многих внутренних органов, стенки сосудов) главной функцией является опорная. 6. Клетки иммунной системы выполняют защитную функцию. Функции органелл клеток 1. Эндоплазматический ретикулум представляет собой систему канальцев, уплощенных цистерн и мелких пузырьков. Строение мембраны ретикулума подобно строению клеточной мембраны. Функции эндоплазматического ретикулума: а) является резервуаром для различных ионов, в том числе Са 2+; б) обеспечивает синтез и транспорт различных веществ, в том числе молекул белков, липидов; в) обеспечивает детоксикацию (в клетках печени) ядовитых веществ, попадающих в организм с пищей или вдыхаемым воздухом, а также метаболитов. Эти вещества в результате превращений выводятся с мочой и желчью в виде глюкуроновых и сульфуроновых соединений. 2. Ядро — это самая большая органелла клетки. Оно несет генетическую информацию и обеспечивает регуляцию синтеза белка в клетке. 3. Митохондрии представляют собой округлые, овальные или удлиненные образования с двойной мембраной. Они являются энергетическими станциями клеток, так как в них вырабатывается (высвобождается) основное количество энергии из поступающих в организм питательных веществ. 4. Рибосомы — плотные частицы, состоящие из рибосомных РНК (рРНК) и белка, их функцией является синтез различных по функции белков (ферменты, белки-переносчики, рецепторы, компоненты цитоскелета). Структура клеточной мембраны. Ее основу (матрикс) составляет бимолекулярный слой фосфолипидов, являющихся барьером для заряженных частиц и молекул водорастворимых веществ. 14 Физиология Молекулы белков, встроенные в фосфолипидный матрикс клеточной мембраны, называют интегральными. Обновление белков мембраны происходит очень быстро — в течение 2–5 дней (срок их жизни). Большинство интегральных белков пронизывают всю толщу клеточной мембраны и образуют каналы, обеспечивающие транспорт веществ через клеточную мембрану (рис. 1.1). Белки, прикрепленные к поверхности клеточной мембраны (в основном к внутренней ее части), называют периферическими. Они, как правило, являются ферментами: это фосфатазы, ацетилхолинэстераза, протеинкиназы, аденилатциклаза. Клеточные мембраны обладают избирательной проницаемостью и текучестью: отдельные части мембраны могут перемещаться из одного участка к другому. Функции клеточной мембраны 1. Барьерная (защитная) функция выполняется клеточными мембранами всех тканей организма. 2. Транспортная функция клеточной мембраны в совокупности с барьерной функцией формирует состав внутриклеточной среды, наиболее благоприятный для метаболических реакций. Транспорт частиц и воды (че- Рис. 1.1. Структурно-функциональная организация клеточной мембраны (по A. Vander et al., 2004) Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 15 рез биологические мембраны, внутри клеток, через стенки сосудов, протоков, канальцев, внутри сосудов и канальцев) — жизненно важный для организма процесс. Если энергия расходуется непосредственно на перенос частиц, транспорт следует называть первично активным. Если же на транспорт частиц расходуется ранее запасенная энергия, например концентрационный градиент, такой транспорт следует называть вторично активным. Поскольку транспорт в обоих случаях является активным (с затратой энергии), обоснованно использовать термины «первичный и вторичный транспорт веществ». 3. Рецепторная функция — восприятие изменений внешней и внутренней среды организма с помощью специальных структур — рецепторов, обеспечивающих распознавание различных раздражителей и реагирование на них клеток. Рецепторы клеточной мембраны обладают специфической чувствительностью к различным химическим и физическим агентам, в том числе медиаторам, гормонам, лекарствам. 4. Создание электрического заряда и возникновение потенциала действия в возбудимых тканях (возбуждения) и проведение его. Электрический заряд имеют все живые клетки. 5. Выработка биологически активных веществ (тромбоксаны, лейкотриены, простагландины и др.). 1.2. Транспорт веществ Транспорт веществ делят на первично активный (первичный) и вторично активный (вторичный). Если энергия затрачивается непосредственно на движение вещества — это первично активный транспорт. Если на транспорт веществ расходуется ранее затраченная энергия на создание электрохимических градиентов — это вторичный транспорт. После блокады выработки энергии сразу же прекращается первичный транспорт. Вслед за этим прекращается и вторичный транспорт, что доказывает его активный характер. Активность транспорта определяется тем, что расходуется энергия АТФ. Частицы же всегда движутся пассивно (у них нет транспортного средства), их либо пере- 16 Физиология носит насос, либо двигает электрохимический градиент, который создают ионные насосы. Первичный транспорт осуществляется вопреки концентрационному и электрическому градиентам и включает три варианта. 1. Транспорт с помощью насосов (помп). Насосы представляют собой белковые молекулы, обладающие свойствами переносчика и АТФазной активностью. Непосредственным источником энергии является АТФ. В нервных клетках 60–70% потребляемой энергии расходуется на работу Na/К-насоса. Достаточно хорошо изучены Na/К-, Саи Н-насосы. Есть данные, свидетельствующие о наличии С1-насоса. Насосы локализуются на клеточных мембранах или на мембранах клеточных органелл и представляют собой интегральные белки. Механизм работы ионных насосов. Принцип работы всех насосов одинаков. Nа/К-насос (Nа/К-АТФаза) электрогенный, поскольку за один цикл выводится из клетки три иона Nа+ и возвращается в клетку два иона К+. На один цикл работы Nа/К-насоса расходуется одна молекула АТФ. Натрий-калиевый насос — интегральный белок (пронизывает всю толщу клеточной мембраны), состоит из четырех полипептидов, имеет центры связывания Nа+ и К+. Центр связывания Nа+ находится на внутренней стороне клеточной мембраны, для К+ — на внешней стороне мембраны. Когда ионы Nа+ и К+ связываются со своими центрами, высвобождается энергия АТФ, под действием которой изменяется конформация белка-переносчика. При этом центр связывания Nа+ оказывается с наружной, а К+ — с внутренней стороны клеточной мембраны. Сродство центров к ионам теряется, и Nа+ оказывается снаружи, а К+ — внутри клетки. Переносчик возвращается в исходное состояние. Затем цикл повторяется. Главными активаторами насоса являются гормоны (альдостерон, тироксин); его специфическими блокаторами являются строфантины, особенно уабаин. Работа Na/К-насоса после удаления К+ из среды сильно нарушается. Недостаток энергии (кислородное голодание) ингибирует Na/К-насос. Кальциевые насосы (Са-АТФазы) работают таким же образом, с тем лишь различием, что переносятся только ионы Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 17 Са 2+ и в одном направлении — из гиалоплазмы в сарко- или эндоплазматический ретикулум, а также из гиалоплазмы наружу клетки. Протонный насос (Н-АТФаза) локализуется в канальцах почек, в мембране обкладочных клеток в желудке, где играет важную роль в выработке соляной кислоты; этот насос постоянно работает во всех митохондриях; в почке он участвует в регуляции рН внутренней среды организма. Насосы специфичны. Это проявляется в том, что они обычно переносят какой-то определенный ион или два иона. Например, Са-насос не переносит ионы Na, Nа/К-насос не способен переносить другие ионы. 2. Микровезикулярный транспорт. С помощью этого механизма переносятся крупномолекулярные вещества (полисахариды, белки, нуклеиновые кислоты), которые не могут транспортироваться по каналам или с помощью насосов. Различают три вида этого транспорта. Эндоцитоз — перенос веществ в клетку. Он характерен для синапса — обратное захватывание медиатора в пресинаптическое окончание наблюдается в проксимальных извитых канальцах почек. При эндоцитозе клеточная мембрана образует впячивания или выросты внутрь клетки, которые, отшнуровываясь, превращаются в пузырьки. Последние затем обычно сливаются с первичными лизосомами, образуя вторичные лизосомы, в которых содержимое подвергается гидролизу — внутриклеточному перевариванию. Продукты гидролиза используются клеткой. Различают два типа эндоцитоза — фагоцитоз (поглощение твердых частиц) и пиноцитоз — поглощение жидкого материала (суспензия, коллоидный раствор, в том числе и белков). Экзоцитоз — это транспорт веществ из клетки, наиболее распространенный механизм секреции: из секреторных клеток путем экзоцитоза выводится их жидкий секрет (слизь, гормоны, ферменты), из гепатоцитов — альбумины, из пресинаптических окончаний — медиаторы, из пищеварительных вакуолей удаляются оставшиеся непереваренными частицы. Трансцитоз — это совокупность эндо- и экзоцитоза (перенос частиц через клетку, например молекул белка в виде 18 Физиология везикул через эндотелиальную клетку капилляров на другую ее сторону). 3. Фильтрация — первичный транспорт, при котором переход раствора через полупроницаемую мембрану, через межклеточные соединения (например, через стенку сосуда) осуществляется под действием градиента гидростатического давления между жидкостями по обе стороны этой мембраны. При этом с водой проходят и частицы, для которых проницаема мембрана. Градиент гидростатического давления создается либо деятельностью сердца (фильтрация в артериальном конце капилляра всех органов и тканей организма, образование первичной мочи в почке), либо гладкими мышцами желудочно-кишечного тракта. Это обеспечивает повышение гидростатического давления в полости желудка и кишечника, что способствует всасыванию веществ в кровь. При отсутствии градиента гидростатического давления фильтрация прекращается. Например, прекращается или замедляется мочеобразование и развиваются отеки при недостаточной работе сердца. Вторичный транспорт веществ (диффузия) — переход различных частиц и молекул воды за счет ранее запасенной (потенциальной) энергии, которая создается в виде электрического и концентрационного градиентов ионными насосами. Осуществляется по законам диффузии — частицы перемещаются из области с высокой концентрацией в область с низкой концентрацией, частицы с одноименными электрическими зарядами отталкиваются, с разноименными зарядами — притягиваются друг к другу. Направление диффузии определяется взаимодействием концентрационного (химического) и электрического градиентов (их совокупность называют электрохимическим градиентом). Если частицы не заряжены, то направление их диффузии определяется только градиентом концентрации. Многие авторы этот вид транспорта называют пассивным. Но это необоснованно, поскольку после блокады выработки энергии он прекращается, т.к. не поддерживаются электрохимические градиенты ионов — не работают ионные насосы. Различают несколько видов диффузии. Простая диффузия. Она происходит либо через каналы, либо непосредственно через липидный бислой, сквозь Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 19 который проходят жирорастворимые частицы. Если они находятся в воде по одну сторону мембраны, то могут внедряться в липидную оболочку благодаря тепловому движению (при этом необходимо освободиться от гидратной оболочки). Частицы-неэлектролиты обычно легко освобождаются от гидратной оболочки (разрыв водородных связей). Примером простой диффузии через липидный слой может служить диффузия стероидных гормонов и других липидов, тироксина, мочевины, а также чужеродных клетке веществ, в частности ядов и лекарств, малых незаряженных полярных молекул (этанол, кислород, углекислый газ). Ионы диффундируют через ионные каналы, однако ионные насосы постоянно возвращают их на свои места и поддерживают постоянный заряд клетки (рис. 1.2). Облегченная диффузия характерна для частиц-неэлектролитов, способных образовывать комплексы с другими молекулами (молекулами-переносчиками). Согласно общепризнанному мнению, переносчиками при облегченной диффузии являются интегральные белки мембран (пронизывающие всю толщу клеточной мембраны). Перенос молекул вещества осуществляется с помощью изменения конформации белка-переносчика, но движущей силой является химический градиент частиц, т.е. вторично активно (за счет ранее запасенной энергии). Транспортируемые вещества — в основном сахара и аминокислоты. Этот транспорт осуществляется очень быстро и регулируется гор- Рис. 1.2. Расположение основных ионов внутри и вне клетки. Размер букв Na+ и K+ пропорционален концентрации этих ионов внутри и вне клетки. Крупномолекулярные анионы белковой природы (А–) из клетки не выходят: ~ — Na/K насос; +/– — электрические заряды; — диффузия (вторичный транспорт); — первичный транспорт 20 Физиология монами, например, инсулин увеличивает проницаемость клеточной мембраны для аминокислот и глюкозы, способствуя встраиванию белков-переносчиков в клеточную мембрану. При отсутствии концентрационного градиента вещества облегченная диффузия, как и простая диффузия, прекращается. Осмос — это частный случай диффузии: движение воды (растворителя) через полупроницаемую мембрану в область с большей концентрацией частиц, т.е. с большим осмотическим давлением. Осмотическое давление одномолярного раствора чрезвычайно велико: 22,4 атм, в плазме крови оно существенно ниже — 7,6 атм, несколько больше оно внутри клетки, что обеспечивает ее упругость вследствие поступления воды в клетку и растяжения ее мембраны. Вода поступает в клетку через водные каналы (аквапорионы), через которые могут проходить также малые незаряженные молекулы: этанол, мочевина, кислород, углекислый газ. Натрийзависимый транспорт — вид диффузии, осуществляющийся за счет градиента концентрации Nа+, на создание которого затрачивается энергия. Имеется два варианта данного транспорта. Котранспорт — направление движения транспортируемого вещества совпадает с направлением движения Nа+ согласно его электрохимическому градиенту (совокупность концентрационного и электрического градиентов). Например, перенос глюкозы и аминокислот в проксимальных канальцах нефрона в клетки канальца из первичной мочи или из просвета кишки. Это осуществляется с помощью интегральных белков мембраны клетки, на которых с внешней стороны имеются центры связывания для глюкозы и Nа+ или для аминокислоты и Nа+. При связывании белка-переносчика, например, с глюкозой и Nа+ изменяется конформация переносчика, в результате чего центры связывания оказываются на внутренней стороне клеточной мембраны, при этом сродство между переносчиками и Nа+ с глюкозой теряется, а Nа+ и глюкоза остаются внутри клетки. Переносчик принимает прежнее (исходное) состояние и готов к следующему циклу. Транспортируемые вещества диффундируют в интерстиций согласно своим градиентам, а Nа+ Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 21 выкачивается из клетки Nа/К-насосом, именно поэтому данный вид транспорта и называют вторично активным. При отсутствии градиента Na+ этот вид транспорта невозможен. Контртранспорт (противотранспорт) — это перемещение транспортируемых частиц, направленное в противоположную сторону по отношению к движению Nа+. Этим обменным механизмом регулируется, например, содержание Са 2+ в клетке, рН внутри клетки за счет выведения иона Н+ в обмен на внеклеточный Nа+. Контртранспорт осуществляется так же, как и котранспорт, за исключением того, что центр связывания в белке-переносчике для Nа+ находится снаружи, а центр связывания Са2+ — изнутри клеточной мембраны. В результате связывания ионов с переносчиком изменяется его конформация и Nа+, согласно своему электрохимическому градиенту, оказывается с внутренней стороны клеточной мембраны, а Са2+ — с наружной стороны. При этом комплекс распадается, Nа+ выкачивается из клетки Nа/К-насосом. Как видно, и здесь движущей силой является электрохимический градиент Nа+ благодаря постоянной работе Nа/К-насоса. Если заблокировать этот обменный механизм сердечными гликозидами (например, уабаином), количество Са 2+ в кардиомиоцитах возрастает, что ведет к усилению сердечных сокращений — это используется в клинической практике). Ионы транспортируются через ионные каналы. Ионные каналы. Известно более 50 видов каналов, причем каждая нервная клетка имеет более пяти из них. Состояние активации управляемого ионного канала (ворота открыты) обычно длится около 1 мс, иногда до 3 мс и значительно больше, при этом через один канал может пройти 12–20 млн ионов. Механизм функционирования ионных каналов определяется их строением, особо важную роль играет наличие или отсутствие управляемых ворот ионных каналов. Структурно-функциональная характеристика ионных каналов. Каналы имеют устье и селективный фильтр, а управляемые каналы — и ворота. Каналы заполнены жидкостью, их размеры — 0,3–0,8 нм. Селективность ионных каналов определяется их размером и размером частиц. 22 Физиология Число ионных каналов на клеточной мембране огромно (на 1 мкм2 насчитывают примерно 50 Nа+-каналов). Классификация ионных каналов проводится по нескольким признакам. 1. По наличию или отсутствию механизмов управления функцией канала различают управляемые и неуправляемые каналы. Управляемые каналы имеют ворота с механизмами их управления, поэтому ионы через них могут проходить только при открытых воротах. Через неуправляемые каналы ионы перемещаются постоянно, но медленно, естественно, при наличии электрохимического градиента, как и в случае быстрого перемещения ионов по управляемым каналам. 2. По механизму управления различают: электро-, хемои механоуправляемые каналы. При взаимодействии медиатора (лиганда) с рецепторами, расположенными на поверхности клеточной мембраны, может происходить открытие ворот хемоуправляемых каналов, поэтому их называют также рецепторуправляемыми каналами. Лиганд — это биологически активное вещество или фармакологический препарат, активирующий или блокирующий рецептор. Открытие хемоуправляемых каналов происходит в результате конформационных изменений рецепторного комплекса. Ворота потенциалуправляемых каналов открываются и закрываются при изменении величины мембранного потенциала. В конструкции их ворот имеются частицы, несущие электрический заряд — сенсор напряжения. Механоуправляемые каналы имеются у сенсорных механорецепторов, они активируются и инактивируются деформацией клеточной мембраны (сдавливание, растяжение, сгибание микроворсинок рецепторов слуховых, вестибулярных и др.). 3. По скорости открытия и закрытия ионные каналы делятся на быстрые и медленные. Например, потенциал действия в скелетной мышце возникает вследствие активации быстрых Nа- и К-каналов. В развитии потенциала действия сердечной мышцы наряду с быстрыми каналами для Nа+ и К+ важную роль играют медленные кальциевые каналы. 4. Различают также ионоселективные каналы, пропускающие только один ион, и каналы, не обладающие селективностью. Наиболее высока степень селективности Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 23 потенциалуправляемых каналов, несколько ниже она у хемоуправляемых каналов. Для одного и того же иона может быть несколько видов каналов. Например, постоянно открытые (неуправляемые) К- и Nа-каналы, играющие важную роль в формировании потенциала покоя, и быстрые электроуправляемые К- и Nаканалы, обеспечивающие возникновение потенциала действия, что изучается ниже. Особенно важно для клинической практики то, что ионные каналы блокируются фармакологическими препаратами. Это широко используется с лечебной целью. Новокаин, например, как местный анестетик снимает болевые ощущения потому, что он, блокируя Nа-каналы, прекращает проведение возбуждения по нервным волокнам. 1.3. Свойства биологической ткани и ее раздражители Основные свойства биологической ткани: • раздражимость — общее свойство живой материи активно изменять характер своей жизнедеятельности при действии раздражителя (изменение интенсивности обмена веществ, рН, ускорение клеточного деления и роста, биоэлектрические явления). По степени выраженности последних все ткани делят на возбудимые (нервная и мышечная) и невозбудимые (эпителиальная и соединительная); • возбудимость — это способность некоторых клеток (нервных и мышечных) генерировать потенциал действия (ПД) при раздражении. Возбудимость является частным случаем наиболее общего свойства всех клеток — раздражимости; • проводимость — это способность ткани и клетки передавать возбуждение; • сократимость присуща мышечной ткани и выражается в изменении ее длины и/или напряжения. Для того чтобы клетка возбуждалась, на нее должен подействовать раздражитель. Раздражитель — это изменение внешней или внутренней среды организма, воспринимаемое клетками и вызы- 24 Физиология вающее ответную реакцию. В зависимости от природы раздражители делят на физические (электрические, механические, температурные, световые) и химические. В зависимости от степени специфической чувствительности клеток к тому или иному раздражителю их подразделяют на адекватные и неадекватные. Адекватный раздражитель — это такой раздражитель, к которому клетка в процессе эволюции приобрела наибольшую чувствительность вследствие развития специальных структур, воспринимающих этот раздражитель. Например, рецепторы сетчатки глаза обладают наибольшей чувствительностью к свету, вкусовые рецепторы — к химическим веществам и т.д. Неадекватные раздражители — это раздражители, к которым клетка не обладает высокой чувствительностью. Например, фоторецептор может возбуждаться и при действии механического раздражителя, но его величина должна быть огромной по сравнению со световым раздражителем. При действии раздражителя на возбудимые клетки возникает возбуждение. Сущность процесса возбуждения можно сформулировать следующим образом (Ходжкин А., Хаксли Э., Катц Б.) — все клетки организма имеют электрический заряд, создаваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для разных ионов и работы ионных насосов. При действии раздражителя на возбудимую клетку очень быстро повышается проницаемость ее мембраны для Nа+ и быстро возвращается к норме, затем для К+ и тоже быстро, но с некоторым отставанием возвращается к норме. Вследствие этого ионы быстро перемещаются в клетку и из клетки согласно электрохимическому градиенту (совокупность концентрационного и электрического градиентов) — это и есть процесс возбуждения. Возбуждение возможно только в том случае, если клетка постоянно поддерживает потенциал покоя (мембранный потенциал) — характерный признак живой клетки. Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 25 1.4. Биопотенциалы клетки Потенциал покоя (ПП) — это разность электрических потенциалов между внутренней и наружной средами невозбужденной клетки. При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом скачком отклоняется вниз и показывает отрицательный заряд внутри клетки (рис. 1.3, А). Величина ПП обычно варьирует в пределах 40–90 мВ. Органеллы клеток имеют собственные заряды. Непосредственная причина существования ПП, согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц, 1902–1952), — неодинаковая концентрация анионов и катионов внутри (К+) и вне клетки (Nа+ и Cl–). Неравномерное расположение ионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для различных ионов и работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки электрохимическому градиенту. Терминология. Проницаемость клеточной мембраны — это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильтрации. Термин «проводимость» следует использовать лишь применительно к заряженным частицам. Роль проводимости клеточной мембраны для различных ионов в формировании ПП. В состоянии покоя из клетки К+ Рис. 1.3. Потенциал покоя возбудимой клетки (А) и схема опыта его регистрации (Б): 1 — клетка; 2 — микроэлектрод; 3 — регистратор; А– — крупномолекулярные анионы белковой природы 26 Физиология выходит в значительно большем количестве, чем входит Nа+ в клетку, поскольку проводимость клеточной мембраны для К+ примерно в 100 раз больше проводимости для Nа+. Органические анионы из-за своих больших размеров вообще не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. По этой причине клетка внутри имеет отрицательный заряд, а снаружи — положительный. Таким образом, калий является основным ионом, обеспечивающим формирование ПП. Основные анионы, находящиеся в клетке, — это глутамат, аспартат, органические фосфаты, сульфаты. Вклад С1– и Nа+ в создание ПП невелик, т.к. проводимость клеточной мембраны в покое для этих ионов очень низкая. Роль ионов Са 2+ в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными фиксированными зарядами мембраны клетки и отрицательными карбоксильными группами интерстиция, нейтрализуя их, что ведет к увеличению и стабилизации ПП. При недостатке Са 2+ в организме ПП снижается, могут возникать потенциал действия (ПД) и судороги. Роль поверхностных зарядов клеточной мембраны в формировании ПП. Наружная и внутренняя поверхности клеточной мембраны имеют собственные электрические заряды преимущественно с отрицательным знаком. Это полярные молекулы клеточной мембраны — гликопротеиды, гликолипиды, фосфолипиды. Фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП. Фиксированные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Таким образом, ПП — это алгебраическая сумма всех положительных и отрицательных зарядов ионов вне и внутри клетки и поверхностных зарядов клеточной мембраны. Роль ионных насосов в формировании ПП заключается в том, что они постоянно поддерживают неодинаковую концентрацию различных ионов в клетке и вне клетки с помощью энергии АТФ. Если заблокировать высвобождение Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 27 энергии АТФ, то в течение одного часа ПП уменьшится до нескольких милливольт (установится доннановское равновесие). С другой стороны, если уравнять проводимость клеточной мембраны для всех ионов, то ПП будет составлять также несколько милливольт за счет работы Nа/К-помпы (электрогенный насос). Нормальная величина ПП является необходимым условием возникновения процесса возбуждения клетки, т.е. возникновения ПД. Потенциал действия (ПД) — это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала вследствие изменения проводимости клеточной мембраны и диффузии ионов в клетку и из клетки. Величина ПД вариабельна — 80–130 мВ, длительность ПД нервного волокна — 2 мс, волокна скелетной мышцы — до 10 мс. Длительность ПД сердечной мышцы — около 300 мс. Механизм возникновения ПД. Наиболее часто встречаемый вариант возникновения ПД — это быстрая диффузия Nа+ в клетку, а К+ — из клетки вследствие фазовых изменений проводимости клеточной мембраны под влиянием самого ПД. Величина мембранного потенциала при этом сначала быстро уменьшается, а затем снова восстанавливается до исходного уровня. Фазы ПД классифицируются по изменению величины и знака заряда клетки. Большинство авторов названия фазам ПД дают вне связи между собой, что не является классификацией и затрудняет усвоение материала. Рассмотрим три фазы ПД. 1. Фаза деполяризации (уменьшение заряда клетки до нуля, рис. 1.4, 1) развивается при действии деполяризующего раздражителя на клетку (электрический ток, локальное возбуждение, распространяющийся по нервному или мышечному волокну электрический импульс). Начальная частичная деполяризация клеточной мембраны происходит без изменения ее проводимости для ионов. Поэтому, несмотря на наличие движущей силы (концентрационный и электрический градиенты), движение Nа+ в клетку через быстрые потенциалуправляемые Nа-каналы отсутствует. Дальнейшая деполяризация ведет к открыванию ворот Na-каналов, движению Nа+ в клетку и достижению деполяризации клетки до КУД (критический уровень деполяри- 28 Физиология Рис. 1.4. Процесс возбуждения клетки (фазы ПД): 1 — деполяризация; 2 — инверсия; 3 — реполяризация зации) — величина мембранного потенциала, при которой начинается регенеративная часть ПД во время деполяризации клетки. КУД обычно составляет 50 мВ (возможны и другие величины), при этом проводимость мембраны для Nа+ резко возрастает — открывается дополнительно большое число потенциалуправляемых ворот Nа-каналов, и Nа+ лавиной устремляется в клетку согласно концентрационному и электрическому градиентам. В итоге мембранный потенциал исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается. Во время деполяризации мембраны увеличивается проницаемость ее и для Са 2+, он также идет в клетку, но в нервных волокнах, нейронах и волокнах скелетной мускулатуры роль Са 2+ в развитии ПД мала. В клетках гладкой мышцы и миокарда его роль существенна. 2. Фаза инверсии — изменение знака заряда клетки на противоположный. Вся восходящая часть пика ПД в большинстве случаев обеспечивается в основном диффузией Nа+ в клетку. Во время восходящей части фазы инверсии диффузия Nа+ в клетку продолжается, поэтому число положительных ионов в клетке превосходит число отрицательных, заряд внутри клетки становится положительным. Напомним, что крупномолекулярные анионы выйти из клетки не могут, а ворота для К+ еще закрыты. Теперь электрический градиент препятствует входу Nа+ внутрь клетки (положительные Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 29 заряды отталкиваются друг от друга), Nа-проводимость снижается. Тем не менее некоторый период времени (доли миллисекунды) Nа+ продолжает входить в клетку, о чем свидетельствует продолжающееся нарастание величины ПД. Это означает, что концентрационный градиент, обеспечивающий движение Nа+ в клетку, сильнее электрического, препятствующего входу Nа+ в клетку. Примерно через 0,5–2 мс и более после начала деполяризации (это время зависит от вида клетки) рост ПД прекращается вследствие закрытия Nа-каналов и прекращения поступления Nа+ в клетку (рис. 1.4, 2). Нисходящая часть фазы инверсии начинается с открытия ворот К-каналов и быстрого возрастания выхода К+ из клетки. Поскольку К+ находится преимущественно внутри клетки, он согласно концентрационному градиенту быстро выходит из клетки после открытия ворот К-каналов, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. В период нисходящей части фазы инверсии выходу К+ из клетки способствует также электрический градиент. К+ выталкивается из клетки положительным зарядом внутри нее. Так продолжается до полного исчезновения положительного заряда внутри клетки (до конца нисходящей части фазы инверсии (рис. 1.4, 2, пунктирная линия)). Таким образом, амплитуда ПД складывается из величины ПП и величины фазы инверсии, составляющей у разных клеток 10–50 мВ. Если ПП мал, амплитуда ПД этой клетки небольшая. 3. Фаза реполяризации (рис. 1.4, 3) — восстановление заряда клетки. Она обеспечивается также продолжающимся быстрым выходом К+ из клетки согласно концентрационному градиенту. Проводимость клеточной мембраны для К+ все еще высока. Поскольку клетка теперь уже снова внутри имеет отрицательный заряд, а снаружи — положительный (рис. 1.4, 3), электрический градиент препятствует выходу К+ из клетки, что снижает его проводимость, хотя он продолжает выходить из клетки. Это означает, что действие концентрационного градиента выражено сильнее электрического градиента. Таким образом, вся восходящая часть ПД обусловлена вхо- 30 Физиология дом Nа+ в клетку, нисходящая — выходом К+ из клетки. Главную роль в возникновении ПД играет Nа+, входящий в клетку при повышении проводимости клеточной мембраны — в безнатриевой среде ПД не возникает. Причина последовательного открытия электроуправляемых ворот ионных каналов — это изменение величины мембранного потенциала и знака заряда клетки. Все фазы регенеративные — необходимо только достичь КУД, далее ПД развивается за счет потенциальной энергии клетки в виде электрохимических градиентов, т.е. вторично активно. Особо важно отметить тот факт (это видно из анализа механизмов формирования ПД), что Na/К-помпа непосредственно за фазы потенциала действия не отвечает, хотя она работает непрерывно в покое и продолжает работать во время развития ПД. Необходимо хорошо запомнить: ПД развивается благодаря диффузии ионов с огромной скоростью согласно их электрохимическому градиенту, т.е. потенциальной энергии, запасенной ранее в результате работы ионных насосов. Следовые гиперполяризация и деполяризация. Следовая гиперполяризация клеточной мембраны — это увеличение мембранного потенциала после фазы реполяризации, она обычно является результатом еще сохраняющейся повышенной проводимости клеточной мембраны для К+, характерна для нейронов (рис. 1.5, Б1). Ворота К-каналов еще не полностью закрыты, поэтому К+ продолжает выходить из клетки согласно концентрационному градиенту, что и ведет к гиперполяризации клеточной мембраны. Следовая деполяризация развивается после гиперполяризации, она также характерна для нейронов, но может быть зарегистрирована и в волокнах скелетной мышцы (рис. 1.5, Б2). Механизм ее изучен недостаточно. Роль ПД. Он обеспечивает передачу информации от клетки к клетке на большие расстояния, при этом осуществляется связь нейронов друг с другом, а также передача команд на эффекторы (рабочие органы). Мышца, например, произвольно сокращается только в том случае, если к ней приходит нервный импульс из ЦНС, отсутствие ПД в сердечной мышце также ведет к остановке сердца и т.д. Вся информация о внешней среде организма поступает в кору Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 31 Рис. 1.5. Два варианта развития ПД двух клеток: А — исчерченного мышечного волокна, выражено замедление фазы реполяризации; Б — нейрона: 1 — следовая гиперполяризация; 2 — следовая деполяризация большого мозга с помощью ПД. При слабых раздражениях возникает локальный потенциал. Локальный потенциал. В эксперименте локальный потенциал можно вызвать слабым (подпороговым) раздражением. В условиях натуральной деятельности нервной системы локальными потенциалами являются: возбуждающий и тормозной постсинаптический потенциалы, рецепторные потенциалы. Величина локальных потенциалов весьма вариабельна, она может достигать 10–40 мВ в зависимости от рода клеток и силы стимула. Свойства такого ответа существенно отличаются от импульсного (табл. 1.1). Таблица 1.1 Сравнительная характеристика локального потенциала и ПД Свойство Распространение Зависимость величины от силы стимула Локальный потенциал Не распространяется, затухает там, где возникает Возрастает с увеличением силы стимула, т.е. подчиняется закону «силы» Потенциал действия Распространяется без затухания (декремента) на большие расстояния по всей длине нервного волокна Не зависит (подчиняется закону «все или ничего») ª 32 Физиология Окончание табл. 1.1 Свойство Явление суммации Амплитуда Возбудимость ткани при возникновении потенциала Локальный Потенциал потенциал действия Суммируется — воз- Не суммируется растает при повторных частых подпороговых раздражениях 10–40 мВ 80–130 мВ Увеличивается Уменьшается вплоть до полной невозбудимости (рефрактерность) Амплитуда ПД не зависит от силы раздражения потому, что он возникает вследствие регенеративного процесса. Величина ПД складывается из величин потенциала покоя и фазы инверсии ПД. Следует также отметить, что возбудимость клетки сильно изменяется в процессе ее возбуждения и связана с фазами ПД. Повышение возбудимости клетки во время локального потенциала объясняется тем, что клеточная мембрана оказывается частично деполяризованной, поэтому для достижения КУД во время локального потенциала нужен раздражитель значительно меньшей силы. Возбудимость клетки при частичной деполяризации повышается также и потому, что начинают открываться ворота Na-каналов. 1.5. Возбудимость и лабильность. Использование электрического тока в медицине Изменения возбудимости клетки во время ПД. Различают четыре основных фазы изменения возбудимости, каждая из которых соответствует определенной фазе ПД и так же, как и фазы ПД, определяется состоянием проводимости клеточной мембраны для ионов. 1. Кратковременное повышение возбудимости в начале развития ПД (рис. 1.6, Б1) наблюдается при частичной деполяризации клеточной мембраны. Если деполяризация не достигает критического уровня (КУД), то регистрируется локальный потенциал. Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 33 Возбудимость повышена потому, что клетка частично деполяризована, мембранный потенциал приближается к КУД. Кроме того, начинают открываться электроуправляемые Nа-каналы. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла КУД, когда возникает ПД, развивающийся независимо от действия раздражителя. 2. Абсолютная рефрактерная фаза (полная невозбудимость клетки) соответствует пику ПД и продолжается 1–2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефрактерная фаза (рис. 1.6, Б2). Клетка в этот период на раздражения любой силы не отвечает, т.к. ПД развивается вследствие открывания и закрывания ионных каналов под влиянием изменения величины и знака заряда самой клетки. Величина ПД подчиняется закону «все или ничего». Абсолютный рефрактерный период ограничивает максимальную частоту генерации ПД. Продолжительность абсолютной рефрактерной фазы вариабельна, у нервных волокон она невелика и составляет 2–3 мс, у сер- КУД Рис. 1.6. Изменения возбудимости клетки во время потенциала действия: А — ПП нейрона; Б — фазы изменения возбудимости: 1, 4 — возбудимость повышена; 2 — абсолютная рефрактерная фаза; 3 — относительная рефрактерная фаза; Н — уровень нормальной возбудимости 34 Физиология дечной мышцы достигает 270 мс (столько же длится и ПД сердечной мышцы). 3. Относительная рефрактерная фаза — это период восстановления возбудимости клетки, когда сильное раздражение может вызвать новое возбуждение (рис. 1.6, Б3). Она соответствует конечной части фазы реполяризации (начиная от КУД) и следовой гиперполяризации клеточной мембраны при ее наличии. В этот период состояние ворот ионных каналов и движения ионов еще не восстановилось до состояния покоя клетки. 4. Фаза экзальтации (период повышенной возбудимости, рис. 1.6, Б4) соответствует следовой частичной деполяризации клеточной мембраны вслед за гиперполяризацией, что свойственно нейронам ЦНС. В этот период очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько меньше обычного и оказывается ближе к КУД, что объясняют повышенной проницаемостью клеточной мембраны для ионов Nа+. Скорость фазовых изменений возбудимости клетки определяет ее лабильность (функциональную подвижность). Лабильность (Введенский Н.Е.) — это скорость развития одного цикла возбуждения, т.е. ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая, в свою очередь, зависит от скорости изменения проводимости клеточной мембраны. При этом особое значение имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани, поскольку в период рефрактерности клетка не отвечает на раздражение. Мерой лабильности является максимальное число ПД, которое ткань может воспроизвести в 1 с. Лабильность нервного волокна равна 200–400, мышцы — 100–200 ПД/с, нервно-мышечного синапса — порядка 40 передач в секунду. При длительном бездействии органа и при утомлении, а также в случае нарушения иннервации лабильность ткани понижается. Критерии для оценки возбудимости. Возбудимость различных тканей неодинакова: она выше у нервных клеток, чем у мышечных, что используется в клинической Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 35 практике, например при выяснении причины двигательных нарушений. Возбудимость изменяется в процессе возбуждения, при отклонении химического состава внеклеточной жидкости, например в результате высокой длительной активности клеток, изменений показателей внутренней среды в патологических случаях. Имеется три основных показателя состояния возбудимости ткани. 1. Пороговый потенциал (ΔV) — это минимальная величина, на которую надо уменьшить ПП, чтобы вызвать ПД. Небольшая величина ΔV свидетельствует о высокой возбудимости клетки. При ΔV = 5–10 мВ возбудимость клетки высокая, при ΔV = 30–40 мВ возбудимость клетки низкая (рис. 1.7). Чаще всего возбудимость ткани оценивается по пороговой силе раздражителя, несмотря на то что ΔV — наиболее точный показатель (но его сложно определять). 2. Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД) при неограниченом времени ее действия на ткань (рис. 1.8). Следует, однако, отметить, что это время мало — несколько миллисекунд. Сила раздражителя — понятие собирательное, она отражает степень выраженности раздражающего воз- КУД Рис. 1.7. Зависимость возбудимости клетки от величины потенциала покоя при одинаковой величине КУД: ΔV — пороговый потенциал; ΔV1 = 10 мВ; ΔV2 = 30 мВ 36 Физиология действия стимула на ткань. Например, концентрация химического вещества выражается в ммоль/л, сила звука — в децибелах (дБ), температура среды — в градусах Цельсия (°С), сила электрического тока — в амперах (А). При использовании в качестве раздражителя электрического тока предложенное определение пороговой силы совпадает с понятием «реобаза». Реобаза — наименьшая сила тока, способная вызвать импульсное возбуждение. Большая пороговая сила свидетельствует о низкой возбудимости ткани. Чем выше возбудимость, тем ниже пороговая сила. При сверхпороговых раздражениях могут возникать болевые ощущения. 3. Пороговое время — это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение (рис. 1.8 — проекция точки А на абсциссу). В эксперименте и в клинической практике для оценки свойств возбудимой ткани чаще используют не пороговое время, а хронаксию. Это связано с тем, что определение порогового времени затруднено (пологая часть кривой на рис. 1.8). Хронаксия — наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение (рис. 1.8 — проекция точки Б на абсциссу). Хронаксия соответствует более крутой части кривой «сила–время»; она колеблется от сотых долей до сотен миллисекунд. Рис. 1.8. Кривая «сила–длительность». Точка А отражает пороговую силу раздражителя и пороговое (полезное) время раздражения; точка Б отображает удвоенную реобазу и хронаксию Глава 1. Общая физиология возбудимых тканей. Транспорт веществ 37 Измерение хронаксии в клинической практике позволяет уточнить характер повреждений мышцы при травмах. Взаимозависимость между сверхпороговой силой раздражителя и временем его действия, необходимым для вызова возбуждения, показана также на рис. 1.8. Кривая в виде гиперболы (кривая Гоорвега–Вейса–Лапика) демонстрирует, что с увеличением сверхпороговой силы раздражителя время его действия, необходимое для вызова возбуждения, уменьшается, но, если раздражитель действует очень короткое время, возбуждение не успевает возникнуть. Использование электрического тока в медицине. Высокочастотный переменный ток (>10 кГц) опасности для организма не несет: при сверхкоротком воздействии на ткань импульс электрического тока дает лишь тепловой эффект, что используется в клинической практике для глубокого прогревания тканей при различных патологических процессах. Низкочастотный переменный синусоидальный ток (50 Гц) стимулирует возбудимые ткани. Стимулы синусоидального тока частотой 50 Гц большого напряжения опасны для жизни — они могут вызвать фибрилляцию сердца с летальным исходом! ГЛАВА 2 Разная концентрация анионов и катионов внутри и вне клетки — это ее электрический заряд и признак жизни. Исчезновение электрических зарядов возбудимых клеток свидетельствует о смерти организма. Частная физиология возбудимых тканей В данной главе рассмотрим физиологию рецепторов, нервных волокон, мышц и нервно-мышечного синапса. 2.1. Рецепторы Рецепторы (от лат. receptum — принимать) — структурные элементы клеточных мембран, воспринимающие действие определенного вида раздражителя и запускающие формирование ответной реакции клетки. Рецепторы для гормона могут находиться и внутри клетки. Существует несколько классификаций рецепторов. Классификация рецепторов по назначению. Сенсорные рецепторы (лат. sensus — чувство) — рецепторы, воспринимающие действие адекватных физических или химических раздражителей внешней или внутренней среды организма и запускающие процесс формирования сигнала (информации, ПД) для посылки в ЦНС или вегетативный ганглий. Это чувствительные нервные окончания или специальные рецепторные клетки, в мембранах которых имеются рецепторные молекулы, являющиеся интегральным белком, имеющим ионный канал. При действии раздражителя активируется ионный канал многих рецепторных молекул, ионы Na (преимущественно) входят в клетку и деполяризуют ее, то есть возникает РП. Таким образом, восприятие раздражителя осуществляется с помощью преоб- Глава 2. Частная физиология возбудимых тканей 39 разования энергии раздражения в рецепторный потенциал (РП), который посредством своего электрического поля обеспечивает возникновение ПД в окончании дендрита чувствительного (афферентного) нейрона. Сенсорные рецепторы запускают только процессы информации об изменении внешней или внутренней среды организма. Нейронные рецепторы — рецепторы нейронов, воспринимающие действие медиатора и запускающие процесс обработки информации и ее хранения в нейронах, посылку сигналов (ПД) к другим нейронам и центрам. Эффекторные рецепторы — рецепторы, воспринимающие действие гормона или медиатора и запускающие процесс формирования ответной реакции самой клетки. Это рецепторы рабочих (эффекторных) клеток — мышечных и секреторных. В свою очередь, нейронные и эффекторные рецепторы классифицируются по медиатору: адрено-, холино-, серотонинорецепторам и др. Классификация сенсорных рецепторов, в свою очередь, проводится по нескольким критериям (рис. 2.1). В зависимости от вида воспринимаемого раздражителя выделяют четыре типа сенсорных рецепторов. Рецепторы Соматосенсорные Первичные Вторичные Мышечный Обонятельный Слуховой Вестибулярный Вкусовой Зрительный Рис. 2.1. Различные виды рецепторов. Пунктирными стрелками показаны участки действия раздражителей на рецептор, сплошными — места возникновения потенциала действия (по Г. Шеперду, 1987, с изменениями) 40 Физиология Хеморецепторы — вкусовые и обонятельные рецепторы, часть сосудистых и тканевых рецепторов (реагирующих на изменение химического состава крови, лимфы, межклеточной жидкости). Хеморецепторы имеются в гипоталамусе (например, в центре пищеварения) и продолговатом мозге (дыхательный центр). Механорецепторы расположены в коже и слизистых оболочках, опорнодвигательном аппарате, сосудах, внутренних органах, слуховой, вестибулярной и тактильной сенсорных системах. Терморецепторы (они подразделяются на тепловые и холодовые) находятся в коже, сосудах, внутренних органах, различных отделах ЦНС (гипоталамусе, среднем, продолговатом и спинном мозге). Фоторецепторы расположены в сетчатке глаза, они воспринимают световую (электромагнитную) энергию. По структурно-функциональной организации различают первичные и вторичные сенсорные рецепторы. Первичные рецепторы представляют собой чувствительные окончания дендрита афферентного нейрона. К ним относятся: обонятельные, тактильные, температурные, болевые рецепторы, проприорецепторы, а также хемо- и терморецепторы внутренних органов. Тело нейрона расположено в спинальных ганглиях или в ганглиях черепных нервов. Вторичные сенсорные рецепторы имеют специальную клетку, синаптически связанную с окончанием дендрита сенсорного нейрона (см. рис. 2.1). К вторичным рецепторам относятся: вкусовые, фото-, фоно-, вестибуло- и хеморецепторы аортальной и синокаротидных зон. По скорости адаптации различают быстро адаптирующиеся рецепторы вибрации — тельца Пачини и прикосновения — тельца Мейснера; медленно адаптирующиеся проприорецепторы — часть болевых рецепторов, механорецепторы легких; адаптирующиеся со средней скоростью — фоторецепторы, терморецепторы кожи. В зависимости от способности воспринимать один или более видов раздражителей выделяют моносенсорные рецепторы (они обладают максимальной чувствительностью к одному виду раздражителей, например рецепторы сетчатки) и полисенсорные рецепторы (они воспринимают несколько адекватных раздражителей, например механи- Глава 2. Частная физиология возбудимых тканей 41 ческого и температурного или механического, химического и болевого). Примером являются болевые рецепторы. По ощущениям рецепторы подразделяются на зрительные, слуховые, вкусовые, обонятельные, терморецепторы, тактильные, болевые рецепторы (ноцицепторы) — это свободные нервные окончания, которые имеются в зубах, коже, мышцах, сосудах, внутренних органах. Они возбуждаются при действии механических, термических и химических (гистамин, брадикинин, К+, Н+ и др.) раздражителей. По расположению в организме рецепторы делят на экстеро- и интерорецепторы (внутренних органов — висцерорецепторы, сосудов и ЦНС). Разновидностью интерорецепторов являются рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулярные рецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек (например, слизистой рта) и органов чувств: зрительные, слуховые, вкусовые, терморецепторы, обонятельные. Свойства сенсорных рецепторов. Возбудимость сенсорных рецепторов очень высокая и превышает чувствительность новейших технических аппаратов, регистрирующих соответствующие сигналы. В частности, для возбуждения фоторецептора сетчатки достаточно нескольких квантов световой энергии, для обонятельного рецептора — одной молекулы пахучего вещества. Спонтанная активность многих рецепторов (фоно-, вестибуло-, термо-, хемо-, проприорецепторов) без действия на них раздражителя связана с повышенной проницаемостью клеточной мембраны для ионов, приводящей к снижению ПП до КУД и генерации ПД в нервном волокне. Возбудимость таких рецепторов выше, чем рецепторов без фоновой активности — даже очень слабый раздражитель способен значительно повысить частоту импульсации нейрона. Фоновая активность рецепторов в условиях физиологического покоя участвует в поддержании тонуса ЦНС и бодрствующего состояния организма. Адаптация рецепторов — это уменьшение их возбудимости при длительном действии раздражителя, выражающееся в снижении амплитуды рецепторного потенциала (РП) и, как следствие, — частоты импульсации в афферент- 42 Физиология ном нервном волокне. Механизм адаптации изучен недостаточно. Значение адаптации рецепторов заключается в том, что она защищает организм от избыточного потока импульсов, иногда — от неприятных ощущений. Каждый вид рецепторов в процессе эволюции приспособлен к восприятию одного или нескольких видов раздражителей. Такие раздражители называются адекватными. К ним рецепторы имеют наибольшую чувствительность (например, рецепторы сетчатки возбуждаются при действии нескольких квантов световой энергии). К другим, неадекватным раздражителям, рецепторы малочувствительны. Неадекватные раздражители также могут возбудить сенсорные рецепторы, но энергия неадекватного раздражителя должна быть в миллионы и миллиарды раз больше адекватного. Механизм возбуждения сенсорных рецепторов. При действии адекватного раздражителя в первичном рецепторе возникает рецепторный потенциал, представляющий собой деполяризацию клеточной мембраны обычно вследствие движения ионов Na+ внутрь клетки, К+ — из клетки. РП — это локальный потенциал, он за счет своего электрического поля обеспечивает возникновение ПД: в мякотных волокнах — в первом перехвате Ранвье, в безмякотных — ПД РП ПП КУД Рис. 2.2. Типичные взаимоотношения между амплитудой рецепторного потенциала (РП) и частотой ПД, возникающих в афферентном нервном волокне при сверхпороговых уровнях РП: КУД — критический уровень деполяризации, ПП — потенциал покоя (по А. Гайтону, 1985) Глава 2. Частная физиология возбудимых тканей 43 в непосредственной близости от рецептора. Во вторичных рецепторах при действии раздражителя сначала также возникает РП в рецепторной клетке вследствие движения Na+ или К+ в клетку. Под влиянием РП в синаптическую щель высвобождается медиатор, который, действуя на постсинаптическую мембрану, обеспечивает формирование генераторного потенциала (ГП, тоже локальный). Последний за счет своего электрического поля обеспечивает возникновение ПД в нервном окончании, как и в окончаниях с первичными рецепторами. Зависимость частоты ПД в афферентном нервном волокне от величины РП и ГП показана на рис. 2.2. 2.2. Физиология нервных волокон Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и эффекторными клетками. Имеются два вида нервных волокон: миелинизированные и немиелинизированные (безмиелиновые). Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100 слоев), а в ЦНС — олигодендроциты (глиальные клетки, это те же шванновские клетки). Миелиновая оболочка нервного волокна через равные участки (около 1,0 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78% сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают проведение возбуждения и аксонный транспорт, выполняющий трофическую функцию нейрона. Классификация нервных волокон по толщине и миелинизации, от которых зависит скорость проведения возбуждения. Имеется несколько подтипов, их диаметр (d) — до 20 мкм, скорость проведения возбуждения (V) — до 160 м/с. 44 Физиология Волокна типа А и В являются миелинизированными. А-волокна — это афферентные и эфферентные волокна соматической нервной системы (V — до 160 м/с). К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы (d — до 10 мкм, V — до 10 м/с). С-волокна немиелинизированные — это постганглионарные волокна вегетативной нервной системы, а также афферентные волокна от некоторых болевых, тепловых и висцеральных рецепторов. Аксонный транспорт обеспечивает доставку многих веществ к нервному окончанию, его скорость — 1–400 мм/сут. Основная масса веществ образуется в трофическом центре нейрона, расположенном преимущественно около ядра, а используются они в различных участках тела нейрона и его отростках. В аксонных окончаниях также происходит синтез медиаторов, АТФ и повторное использование мембраны пузырьков после высвобождения медиатора. Путем обратного транспорта в тело нейрона доставляются фрагменты мембран, факторы роста и др. В патологических случаях к телу нейрона могут поступать столбнячный токсин, вирусы полиомиелита, герпеса, бешенства. При разрыве нервного отростка его периферическая часть дегенерирует, центральная часть может расти и снова проводить возбуждение. Механизм непрерывного проведения ПД. Это происходит в мышечных волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалуправляемых ионных каналов по всей длине волокна, участвующих в генерации ПД. В нервном и мышечном волокнах ПД возникают под влиянием ВПСП. Проведение нервного импульса, как и мышечного, начинается с распространения колеблющегося по величине электрического поля ПД, которое может деполяризовать мембрану соседнего участка до КУД на расстояние от 0,1 до 1 мм. Это означает, что на данном участке (0,1–1,0 мм) одновременно генерируются новые ПД, обусловленные движением ионов Na+ в клетку, К+ — из клетки, т.е. перпендикулярно относительно волокна (на распространение электрического поля время не затрачивается). Число одновременно возникающих ПД ограничивается длиной возбужденного участка — для безмиелинового волокна — 0,1–1,0 мм (ПД возникают Глава 2. Частная физиология возбудимых тканей 45 рядом друг с другом в непосредственной близости, их число достигает, согласно нашим расчетам, нескольких сотен). Возникающие ПД в соседнем участке волокна за счет своего электрического поля обеспечивают генерацию ПД в новых участках и т.д. (рис. 2.3). Расстояние, которое успевают пройти ионы во время одного ПД из клетки в клетку (перпендикулярно волокну) и вдоль волокна, составляет всего лишь несколько нанометров (толщина клеточной мембраны), что на несколько порядков меньше, чем расстояние, на котором возникают новые ПД, — до 1 мм. Механизм сальтаторного проведения ПД реализуется по миелинизированным волокнам (типы А и В), у которых электроуправляемые ионные каналы имеются только в небольших участках мембраны (в перехватах Ранвье). В области миелиновых муфт, обладающих хорошими изолирующими свойствами, электроуправляемых каналов почти нет, поэтому ПД здесь не возникают. Покрытый миелиновой муфтой участок нервного волокна между перехватами Ранвье в механизме проведения ПД выполняет роль изолятора. В этих условиях ПД, возникший в одном перехвате Ранвье, за счет своего электрического поля деполяризует мембрану соседних перехватов волокна до КУД, что приводит к возникновению в них новых ПД, т.е. возбуждение возникает как бы скачкообразно — только в перехватах, одновременно максимально в 5 перехватах (расстояние до 5 мм). Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только пер- Na+ Na+ Na+ 1 Na+ K+ K+ 2 3 Na+ Na+ K+ K+ 4 Na+ K+ K+ 5 6 K+ 7 8 Рис. 2.3. Непрерывное проведение возбуждения (ПД) в безмиелиновом нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1–7 — состояние возбуждения (ПД); 8 — состояние покоя (пунктиром обозначены промежуточные ПД, вертикальные стрелки указывают направление движения Na+ в клетку и K+ — из клетки) 46 Физиология пендикулярно относительно длины волокна — в клетку и из клетки (вдоль волокна они успевают смещаться во время ПД лишь на несколько нанометров). Электрическое поле ПД, возникших сзади переднего (промежуточные ПД), суммируется с электрическим полем переднего ПД, как и при непрерывном распространении возбуждения (рис. 2.4). 1 2 3 4 Рис. 2.4. Сальтаторное проведение ПД в миелинизированном нервном волокне. Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля переднего ПД, инициирующего возбуждение соседнего участка волокна: 1–3 — состояние возбуждения (ПД); 4 — состояние покоя, пунктиром обозначены промежуточные ПД; вертикальные стрелки указывают направление движения Na+ в клетку и K+ — из клетки Таким образом, посредством электрического поля возбуждение проводится очень быстро потому, что время затрачивается только на возникновение ПД, т.е. на перпендикулярное относительно волокна движение ионов в клетку и из клетки. Длительность одного ПД, как известно, всего лишь несколько миллисекунд. Простые расчеты показали, что скорость проведения ПД по нервному волокну за счет движения ионов вдоль волокна была бы слишком низкой — на преодоление расстояния в 1 м пришлось бы затратить более суток. Распространение возбуждения электротонически, как считают некоторые авторы, вообще невозможно, т.к. электротон — явление физическое, без активации ионных каналов и возникает только в лабораторных условиях при действии катода постоянного тока, чего в организме нет. Сравнение механизмов непрерывного и сальтаторного проведения возбуждения показывает, что различие между ними не принципиально. Характеристика проведения возбуждения по нервному волокну 1. Изолированное проведение возбуждения в отдельных волокнах нервного ствола обусловлено тем, что влияние элек- Глава 2. Частная физиология возбудимых тканей 47 трического поля ПД соседнего волокна не возбуждает другие волокна нерва вследствие изолирующего эффекта их оболочек и интерстиция. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияние функционально разнородных волокон нерва. Если одновременно возбуждается значительное количество волокон, то возникает достаточно сильное электрическое поле, способное открыть ворота натриевых каналов соседних волокон (прежде всего, высоковозбудимых) и таким образом усилить нервное влияние на эффекторные клетки или нейроны в регуляторных процессах. 2. Двустороннее проведение возбуждения, что можно продемонстрировать в эксперименте при нанесении раздражения в любом участке нерва или нервного волокна, при этом возбуждение регистрируется как в одном, так и в другом направлении от места раздражения. Но в естественных условиях возбуждение проводится только в одном направлении: по афферентным волокнам к телу нейрона, по аксону — от тела нейрона. 3. Большая скорость проведения возбуждения (до 160 м/с в нервных волокнах типа Аα). Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока — от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (время полного кругооборота крови в организме — около 22 с). Большая скорость распространения ПД обеспечивает быстрое влияние на другие нейроны, рабочие органы, а также получение обратной информации, играющей важную роль в регуляторных процессах. 4. Малая утомляемость нервного волокна впервые была показана Н.Е. Введенским (1883) — в его опытах нерв сохранял способность к проведению возбуждения в течение 6–8 ч непрерывного раздражения несильными импульсами тока при условии наличия кислорода в окружающей среде и поддержании влажного состояния нерва. Это обусловлено тем, что при проведении ПД по нервным волокнам используется незначительная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для их восстановления. Расход энергии в нервном волокне на единицу массы в миллион раз меньше, чем в работающей мышце. 48 Физиология 5. Возможен функциональный блок проведения возбуждения. Н.Е. Введенский (1901) показал, что при действии на нерв различных факторов, вызывающих длительную деполяризацию клеточной мембраны, возникает полный блок проведения нервных импульсов (состояние парабиоза). Нарушение физиологической непрерывности нервных волокон возникает при действии анестетиков, гипоксии, воспалении, охлаждении, что широко используется в клинической практике. После прекращения действия этих факторов проведение возбуждения по нервным волокнам восстанавливается, если не произошли грубые структурные изменения. Возбуждение от нервного волокна передается к другой клетке с помощью синапса. 6. Высокая лабильность — нервное волокно может проводить до 200–400 импульсов в 1 с. 7. Бездекрементное проведение (от лат. decrementum — убывание, уменьшение) — проведение без затухания (ПД проводится по всей длине нервного волокна). 2.3. Физиология нервно-мышечного синапса Синапс (от греч. synapsis — соединение) — это совокупность структурных элементов, обеспечивающих передачу сигнала от клетки к клетке с помощью химических веществ (медиаторов) или электрического поля. Посредством синапса реализуется действие многих фармакологических препаратов. Структурно-функциональная организация синапса. Каждый синапс имеет пре- и постсинаптическую мембрану и синаптическую щель (рис. 2.5). Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется высвобождение (экзоцитоз) медиатора (от лат. mediator — посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пре- Глава 2. Частная физиология возбудимых тканей 49 синаптическое окончание, где заполняются медиатором и АТФ. Постсинаптическая мембрана (концевая пластинка) в нервно-мышечном синапсе находится около середины мышечного волокна, что свидетельствует о двустороннем проведении возбуждения в нем — это часть клеточной мембраны иннервируемой клетки, содержащая рецепторы, способные связывать молекулы медиатора. Ее особенностью является наличие множества мелких складок, увеличивающих площадь постсинаптической мембраны и количество рецепторов на ней в нервно-мышечном синапсе (до 10–20 млн). Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, образующих в совокупности базальную мембрану, соединяющую преи постсинаптическую мембраны. Механизмы синаптической передачи включают пять основных этапов. Первый этап — процесс выброса медиатора в синаптическую щель, который запускается посредством ПД пресинаптического окончания. Деполяризация его мембраны ведет к открытию потенциал управляемых Са-каналов. Са 2+ входит в нервное окончание согласно 1 3 5 2 4 6 7 Рис. 2.5. Нервно-мышечный синапс скелетной мышцы: 1 — ветвь аксона; 2 — пресинаптическое окончание аксона; 3 — митохондрия; 4 — синаптические пузырьки, содержащие ацетилхолин; 5 — синаптическая щель; 6 — молекулы медиатора в синаптической щели; 7 — постсинаптическая мембрана мышечного волокна с N-холинорецепторами 50 Физиология электрохимическому градиенту, активирует экзоцитозный аппарат пресинапса, представляющий собой совокупность белков, активация которых обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель. Второй этап — диффузия ацетилхолина в течение 0,1–0,2 мс к постсинаптической мембране и действие его на N-холинорецепторы (стимулируются также никотином, вследствие чего и получили свое название). Третий этап — взаимодействие ацетилхолина с N-холинорецепторами постсинаптической мембраны, в результате чего открываются ионные каналы на 1 мс и, вследствие преобладания входа Nа+ в клетку, происходит деполяризация постсинаптической мембраны (концевой пластинки). Эту деполяризацию в нервно-мышечном синапсе называют потенциалом концевой пластинки (ПКП в других синапсах — ВПСП — возбуждающий постсинаптический потенциал) (рис. 2.6). Четвертый этап — возникновение ПД на мембране мышечного волокна. Особенностью нервно-мышечного синапса скелетного мышечного волокна является то, что при одиночной его активации формируется ПКП большой амплитуды (30–40 мВ), электрическое поле которого вызывает генерацию ПД на мембране мышечного волокна вблизи синапса. Большая амплитуда ПКП обусловлена тем, что нервные окончания делятся на многочисленные веточки, при этом каждая из них выбрасывает медиатор. Пятый этап (удаление ацетилхолина из синаптической щели) осуществляется путем разрушения его под действием ацетилхолинэстеразы, расположенной в базальной мембране синаптической щели, в течение нескольких десятых долей миллисекунды. Около 60% холина захватывается обратно пресинаптическим окончанием, что делает синтез медиатора более экономичным, часть ацетилхолина рассеивается. В промежутках между ПД из пресинаптического окончания происходит спонтанное высвобождение 1–2 квантов медиатора в синаптическую щель в течение 1 с, формируя так называемые миниатюрные потенциалы (0,4–0,8 мВ). Они поддерживают высокую возбудимость иннервируемой Глава 2. Частная физиология возбудимых тканей мВ +20 51 ПД 0 –40 КУД КП ПКП ПКП А –80 Б 0 5 10 0 5 10 мс Рис. 2.6. Потенциал концевой пластинки (ПКП) (по Р. Шмидту, 1996, с изменениями): А — ПКП в нормальной мышце; Б — ослабленный ПКП в курарезированной мышце; стрелками указан момент нанесения стимула клетки в условиях функционального покоя и выполняют трофическую роль. Имеется несколько особенностей проведения возбуждения в химических синапсах. 1. Одностороннее проведение возбуждения от нервного волокна к нервной или эффекторной клетке, поскольку на пресинаптической мембране нет рецептора к медиатору, секретируемому нервным окончанием, поэтому не возникает ВПСП. 2. Для синапса характерна задержка в передаче сигнала к другой клетке (в нервно-мышечном синапсе — 0,5–1,0 мс), что связано с высвобождением медиатора из нервного окончания, диффузией его к постсинаптической мембране и возникновением ПКП, способного вызвать ПД в мышечном волокне. 3. Низкая лабильность синапса (в нервно-мышечном синапсе — 30–50 Гц), что в несколько раз ниже лабильности нервного волокна (до 400 Гц). Это объясняется синаптической задержкой. 4. Неизолированное возбуждение — возбуждение рядом расположенных постсинаптических мембран суммируется. 5. Утомляемость синапса (синаптическая депрессия) — ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапса (главная причина — истощение медиатора в пресинаптическом окончании). 52 Физиология 6. Проводимость нервно-мышечного синапса (как и химических синапсов ЦНС) угнетается или стимулируется различными веществами, что используется в медицине. 7. Декрементность (затухание) возбуждения в химических синапсах при недостаточном высвобождении медиатора из пресинаптического окончания в синаптическую щель. 2.4. Скелетная мышца Морфофункциональная характеристика. Мышечная ткань подразделяется на поперечно-полосатую (скелетную и сердечную) и гладкую, образующую стенки внутренних органов и сосудов. Функции скелетной мышцы: • обеспечение всех видов двигательной активности организма, в том числе дыхания, устной и письменной речи; • механическая защита внутренних органов и костей; • участие в процессах теплообразования: терморегуляторный тонус, мышечная дрожь. Свойства мышечной ткани. Специфическим свойством поперечнополосатых и гладких мышц является сократимость — способность сокращаться, т.е. укорачиваться или развивать напряжение. Они обладают возбудимостью и проводимостью, а также растяжимостью и эластичностью. Строение скелетной мышцы. Она состоит из мышечных волокон, каждое из которых формируется в онтогенезе из многих клеток, поэтому включает много ядер. Толщина волокна составляет в среднем 50 мкм (от 10 до 100 мкм), длина колеблется от нескольких миллиметров до нескольких сантиметров. От мембраны мышечного волокна вглубь отходят многочисленные поперечные инвагинации сарколеммы (Т-трубочки), которые обеспечивают ее взаимодействие с саркоплазматическим ретикулумом (СПР) (рис. 2.7). Внутри мышечного волокна тянется масса нитей — миофибрилл, с которыми связана способность мышцы к сокращению, — это толстые параллельно расположенные нити миозина и более тонкие нити актина, частично пере- Глава 2. Частная физиология возбудимых тканей 53 крывающие друг друга. Участок мышечного волокна, состоящий из группы нитей миозина и двух групп нитей актина, называют саркомером. Саркомеры отграничены друг от друга вертикальными полосками, с которыми связаны с обоих сторон нити актина. Существуют белые (быстрые) и красные (медленные) мышечные волокна. Белые мышечные волокна содержат больше миофибрилл, в них относительно мало митохондрий, миоглобина и жиров. Капиллярная сеть, окружающая эти волокна, является относительно редкой. Для этих волокон характерна высокая скорость сокращения, в результате которого развивается значительное усилие. Эти мышцы обладают малой выносливостью. Красные мышечные волокна имеют меньший диаметр. В цитоплазме клеток много митохондрий, жирных кислот, миоглобина (это придает волокнам характерную окраску, вследствие чего их называют красными). Эти волокна окружены густой сетью кровеносных капилляров. Скорость сокращения меньше по сравнению с быстрыми волокнами, но утомление в них развивается медленнее. Функцию поддержания тонуса выполняют в основном медленные волокна. Имеются мышечные волокна, которые занимают промежуточное положение между быстрыми и медленными. Двигательная (нейромоторная) единица — комплекс, состоящий из α-мотонейрона и группы мышечных волокон, иннервируемых коллатералями его аксона. Число мышечных волокон, входящих в состав различных двигательных 1 3 3 2 1 3 3 4 2 5 5 А Z Z Б Z Z Рис. 2.7. Взаимоотношения клеточной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплазматического ретикулума, сократительных белков (5): А — в состоянии покоя; Б — при сокращении мышечного волокна; точками обозначены ионы Ca2+ 54 Физиология единиц, варьирует в широких пределах. Так, в мышцах, совершающих быстрые и точные движения (например, в глазодвигательных), нейромоторные единицы состоят из 3–5 мышечных волокон. В мышцах, осуществляющих менее точные движения (например, мышцы туловища и конечностей), двигательные единицы велики и включают сотни и тысячи мышечных волокон. Механизм сокращения скелетной мышцы. Укорочение мышцы является результатом скольжения актиновых нитей относительно миозиновых, в результате чего длина мышечного волокна уменьшается. При этом длина самих нитей остается неизменной. Миозиновые нити имеют поперечные выступы (поперечные мостики) длиной около 20 нм. Каждый выступ имеет головку, которая соединена с миозиновым стержнем — шейкой. Саркомер миоцита представлен на рис. 2.8. При расслабленном состоянии мышцы головки поперечных мостиков не могут взаимодействовать с актиновыми нитями, поскольку их активные участки (места взаимного контакта с головками) изолированы тропомиозином. Пять основных этапов последовательности процессов, обеспечивающих сокращение мышцы. 1. После генерации ПД на мембране мышечного волокна вблизи нервно-мышечного синапса (вследствие действия электрического поля ПКП) возбуждение распространяется в обе стороны по поверхностной мембране волокон, в том числе по мембранам поперечных трубочек Т-системы. A-диск Н-зона Z-пластинка Филамент белка титина Тонкий филамент М-линия Саркомер Z-пластинка Толстый филамент Рис. 2.8. Саркомер миоцита скелетной мышцы (по A. Vander, J. Sherman, D. Luciano, 2004) Глава 2. Частная физиология возбудимых тканей 55 2. ПД Т-трубочек за счет своего электрического поля активирует электроуправляемые кальциевые каналы на мембране СПР, вследствие чего Са2+ выходит из цистерн СПР согласно электрохимическому градиенту. 3. Попав в межфибриллярное пространство, Са 2+ контактирует с тропонином, что приводит к изменению его конфигурации и смещению тропомиозина. В итоге на нитях актина обнажаются активные участки, с которыми соединяются головки миозиновых мостиков. 4. В результате взаимодействия с актином АТФазная активность головок миозиновых нитей усиливается, обеспечивая высвобождение энергии АТФ. 5. Энергия АТФ расходуется на сгибание шейки миозинового мостика. Внешне это напоминает движение весел при гребле, в связи с чем изменения конфигурации поперечных мостиков сравнивают с «гребковыми движениями» (рис. 2.9). Такие движения — это непосредственная причина скольжения актиновых нитей вдоль миозиновых. Расслабление мышцы: описанные выше процессы проходят в обратной последовательности (рис. 2.10): • реполяризация сарколеммы и Т-трубочек, что ведет к закрытию кальциевых каналов мембраны СПР и к включению ее Са 2+-насосов, возвращающих Са 2+ в цистерны СПР; • снижение концентрации Са 2+ в межфибриллярном пространстве, что вызывает обратную конформацию тропонина, в результате чего тропомиозиновые нити изолируют активные участки актиновых фила40 нм Актин-мономер Шейка Головка Тропонин Тропомиозин Миозиновая нить Рис. 2.9. Пространственная организация сократительных и регуляторных белков в исчерченной мышце. Показано положение миозинового мостика (гребковый эффект, шейка согнута) в процессе взаимодействия сократительных белков в мышечном волокне (сокращение волокна) 56 Физиология Сокращение Z Расслабление Z Толстый филамент (миозин) Z Z Тонкий филамент (актин) Рис. 2.10. Взаимное расположение структурных элементов миофибрилл при их расслаблении и сокращении ментов, и это делает невозможным взаимодействие с ними головок поперечных мостиков миозина; • скольжение актиновых нитей вдоль миозиновых в обратном направлении (под действием веса мышцы или груза и тяги эластических элементов) и восстановление исходных размеров саркомеров. Энергетическое обеспечение мышечного сокращения. Обеспечение деятельности скелетных мышц связано с достаточно значительными энерготратами. Даже в состоянии максимального физиологического покоя (основной обмен) на функционирование мускулатуры организм тратит около 25% всех своих энергоресурсов. Энергия АТФ расходуется для осуществления следующих процессов: • работы натрий-калиевого насоса, обеспечивающего поддержание градиентов концентрации К+ и Na+; • цикла «гребкового движения»; • работы кальциевого насоса, активация которого вызывает понижение концентрации Са2+ в саркоплазме, что приводит к расслаблению мышечного волокна (рис. 2.11). Запасы АТФ в клетке незначительны и могут обеспечить не более 10 одиночных сокращений, поэтому происходит постоянный ресинтез АТФ. Виды мышечного сокращения по длительности. Различают два основных варианта — одиночное и тетаническое. Одиночное сокращение мышцы — это однократное сокращение, которое возникает при действии на мышцу одиночного электрического или нервного им- Глава 2. Частная физиология возбудимых тканей 57 пульса. На кривой одиночного сокращения выделяют три основных фазы (периода): • латентный период — время от нанесения раздражения до начала сокращения; • фаза укорочения (или развития напряжения); • фаза расслабления (см. рис. 2.11). Обычно мышца укорачивается на 5–10% от исходной длины. В латентный период возникает возбуждение мышечных волокон и его распространение вдоль мембраны. Продолжительность латентного периода варьирует до 10 мс. Тетаническое сокращение — это длительное укорочение мышцы, возникающее под действием ритмического раздражения. В его основе лежит явление суммации одиночных мышечных сокращений (рис. 2.12). При нанесении на мышечное волокно или целую мышцу двух и более быстро следующих друг за другом раздражений возникает сокращение, которое имеет бÓльшую амплитуду и длительность, чем одиночное сокращение. При этом раздражения должны поступать в период предыдущего сокращения. Увеличение амплитуды сокращений объясняется тем, что повторные ПД возникают прежде, чем Сапомпа СПР сможет удалить ранее высвобожденный Са 2+. ПД Внутриклеточная концентрация Са2+ Сокращение Время, мс Рис. 2.11. Соотношение времени возникновения ПД, внутриклеточной концентрации Ca2+ и одиночного сокращения скелетной мышцы 58 Физиология Рис. 2.12. Суммация сокращений икроножной мышцы лягушки: 1 и 2 — одиночные сокращения на отдельные раздражения; 3 — суммированное сокращение на спаренное раздражение сокращающейся мышцы (обозначено стрелками) Поэтому уровень Са 2+ в гиалоплазме повышается, активное состояние сократительных компонентов продлевается (последовательная суммация), увеличивается также количество сокращающихся мышечных волокон (пространственная суммация). При повторных раздражениях в фазу расслабления, что бывает при сравнительно низких частотах стимуляции, наступает зубчатый тетанус. При большей частоте стимуляции повторные раздражения приходятся на фазу укорочения, что вызывает гладкий тетанус (рис. 2.13). Амплитуда сокращения и величина напряжения, развиваемые мышечными волокнами при гладком тетанусе, обычно в 2–4 раза больше, чем при одиночном сокращении. Тетаническое сокращение мышечных волокон, в отличие от одиночного сокращения, быстрее вызывает их утомление и поэтому не может поддерживаться длительное время. При некоторой оптимальной (достаточно высокой) частоте стимуляции нерва амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума. При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва — пессимум Введенского. Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной (рис. 2.13). В естественных условиях пессимум не наблюдается, т.к. α-мотонейроны посылают к мышце серии потенциалов с частотой 20–50 имп./с, что должно вызывать сокращения мышечных волокон, близкие к зубчатому тетанусу. При Глава 2. Частная физиология возбудимых тканей Сила сокращения, усл. ед. 10 59 5 4 3 5 2 6 1 0 Раздражение нерва Рис. 2.13. Сокращения икроножной мышцы лягушки при различной частоте раздражения седалищного нерва: 1 — одиночное сокращение (частота 1 Гц); 2, 3 — зубчатый тетанус (15–20 Гц); 4, 5 — гладкий тетанус (25–50 Гц); 6 — расслабление при пессимальной частоте раздражения (60 Гц) этом сокращение целой мышцы напоминает гладкий тетанус. Причина этого — асинхронность работы мотонейронов и обусловленная этим асинхронность сократительной реакции нейромоторных единиц. В зависимости от характера мышечного сокращения различают три вида: • изотоническое — это сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным. Этот режим возникает, например, когда сокращается ненагруженная мышца с одним закрепленным сухожилием, не поднимая (не перемещая) никакого внешнего груза, либо поднимает груз без ускорения. В реальных условиях чисто изотоническое сокращение практически отсутствует; • изометрическое — сокращение мышцы, при котором ее длина не изменяется, но развиваемое ею напряжение возрастает. Такой режим возникает, когда оба конца мышцы зафиксированы и отсутствует возможность их сближения (например, при попытке поднять груз, который больше силы мышцы); • ауксотоническое — это сокращение мышцы, при котором одновременно изменяются длина и напря- 60 Физиология жение. Этот режим характерен для натуральных двигательных актов — мышца, поднимая груз, не только укорачивается, но и изменяет свое напряжение вследствие реальной нагрузки. Сила мышц, их работа и утомление, мощность. В системе СИ сила выражается в ньютонах (H). В физиологической практике силу мышцы, как правило, определяют по максимальной величине веса груза, который может быть поднят при ее сокращении. Используют динамометры, при помощи которых определяют «кистевую», «становую» силу, силу сгибателей и т.д. Факторы, влияющие на силу мышцы. Анатомическое строение — перистые мышцы (волокна расположены косо под углом к продольной оси) способны развивать гораздо большее напряжение, чем мышцы с параллельным расположением волокон. В связи с этим принято определять так называемое физиологическое сечение мышцы, т.е. сумму поперечных сечений всех волокон, из которых состоит мышца. У перистых мышц физиологическое сечение значительно превосходит анатомическое (геометрическое). При соотношении входящих в ее состав быстрых (белых) и медленных (красных) волокон в случае преобладания красных волокон сила мышцы меньше. При умеренном предварительном растяжении мышцы ее сократительный эффект увеличивается, но при сильном растяжении уменьшается. Оказывается, если мышца перерастянута так, что тонкие и толстые нити ее саркомеров не перекрывают друг друга, то сила мышцы равна нулю, т.к. мышца вообще не сокращается из-за отсутствия зон зацепления между нитями актина и миозина. По мере приближения к оптимальной длине покоя, при которой все головки миозиновых нитей способны контактировать с актиновыми филаментами, сила мышечного сокращения вырастает до максимума. Сила сокращения любой мышцы увеличивается с увеличением числа возбужденных мышечных волокон (пространственная суммация) и синхронизацией их сокращения (в естественных условиях это определяется числом возбужденных мотонейронов, в эксперименте — силой Глава 2. Частная физиология возбудимых тканей 61 стимула). Сила тетанического сокращения возрастает с увеличением частоты раздражений или импульсов, поступающих к мышце (последовательная суммация). Работа мышцы. В механике работа (А) определяется как произведение силы (F), приложенной к телу, на расстояние (L) его перемещения под воздействием данной силы: A = F × L (Дж). Утомление мышцы выражается в уменьшении силы ее сокращения, скорости укорочения и расслабления, в результате работа и мощность также уменьшаются. Ослабление мышечных сокращений является следствием ионных сдвигов, т.к. ионные помпы не успевают поддерживать исходные градиенты ионов. В частности, Na+ накапливается в клетке, а К+ — вне клетки, что и ведет к деполяризации миоцитов и снижению их ПД. Наряду с утомлением самих мышечных волокон утомление развивается в нервно-мышечном синапсе (расходование медиатора, закисление среды). В условиях целостного организма физическое утомление развивается и в ЦНС (центральное утомление) — в настоящее время считается второстепенным. При физической работе увеличивается выброс в кровь адреналина и норадреналина, которые стимулируют работу Na/K-насоса, что тормозит развитие утомления. Скорость развития утомления зависит от ритма работы и величины груза. Большой груз или слишком частый ритм работы приводят к быстрому развитию утомления. Наибольшей бывает работа при некотором среднем, оптимальном для данного человека, ритме работы и среднем, оптимальном грузе (правило средних нагрузок). И.М. Сеченов показал, что работа в период утомления других мышц ускоряет восстановление работоспособности утомленных мышц. Мощность мышцы — работа, совершаемая в единицу времени. В системе СИ выражается в ваттах (Дж/с2). Максимальная мощность соответствует выполнению наибольшего объема работы в течение минимального отрезка времени. Но в этом случае быстро развивается утомление. 62 Физиология 2.5. Гладкая мышца Структурные особенности гладких мышц. Сократительные белки (актин и миозин) гладких мышц ориентированы не столь упорядоченно, саркомеры в них отсутствуют, поэтому при микроскопическом исследовании не выявляется характерная для соматической мускулатуры поперечная исчерченность. В связи с этим такие мышцы называют гладкими. Кроме того, гладкие мышцы принято обозначать как непроизвольные, поскольку управление их деятельностью по желанию индивидуума невозможно. Саркоплазматический ретикулум (СПР) организован гораздо проще и представлен плоскими везикулами, расположенными в непосредственной близости от внутренней поверхности клеточной мембраны. В структурном отношении различают унитарные и мультиунитарные гладкие мышцы. У унитарных мышечные волокна объединены в пучки (до 1000 волокон), волокна соединены друг с другом щелевыми контактами (электрические синапсы). Унитарные мышцы образуют стенки полых внутренних органов и многих сосудов. Мультиунитарные мышечные волокна не связаны друг с другом, в организме их мало (например, ресничная мышца радужной оболочки, значительная часть мышц сосудов, мышцы, поднимающие волосы). Функциональные особенности гладких мышц. Автоматия присуща клеткам — водителям ритма (пейсмекерам). В ее основе лежит повышенная проницаемость клеточной мембраны для Ca2+, ведущая к поступлению его в клетку. Когда деполяризация клетки достигает КУД, возникает ПД. Возбудимость ПП большинства гладкомышечных клеток составляет 60–70 мВ. Для миоцитов, обладающих спонтанной электрической активностью, характерны более низкие значения — 30–60 мВ. ПД более продолжительные (10–50 мс). У некоторых миоцитов после начальной быстрой реполяризации формируется плато, которое удлиняет ПД до 500 мс (рис. 2.14). Деполяризация мембраны в основном обусловлена открытием кальциевых каналов и диффузией Са2+ в клетку. Формирование плато связано с поступлением в клетку Nа2+ и Са2+. Проводимость и сократимость гладких мышц зависят от структурных особенностей. Унитарные мышечные во- Глава 2. Частная физиология возбудимых тканей 63 локна образуют пучки (функциональные единицы). Возбуждение в пучке через щелевые контакты передается с помощью электрического поля ПД одного волокна к другому — функциональный синцитий. Поэтому в пучке одновременно возбуждаются и сокращаются все мышечные волокна. Мультиунитарные мышечные волокна возбуждаются и сокращаются изолированно, каждое мышечное волокно получает отдельную иннервацию. Механизм проведения возбуждения у мультиунитарных и унитарных мышечных волокон одинаковый — непрерывный, как и у скелетной мышцы. Расход энергии при сокращении гладкой мышцы мал, поэтому она не утомляется во время продолжительной активности. Пластичность. Большинство гладких мышц, в отличие от скелетных, ведут себя не как эластическое, а как вязкоэластическое (пластическое) образование. При растяжении гладкой мышцы ее напряжение первоначально увеличивается, затем снижается до исходного уровня. В этом проявляется ее свойство пластичности: гладкая мышца может не развивать напряжения как в укороченном, так и в растянутом состоянии. Такая особенность гладкой мышцы предотвращает избыточный рост давления в полых внутренних органах (мочевой пузырь, желудок и др.) при их наполнении. Имеются фазные и тонические мышцы ЖКТ. Длительность одного цикла сокращения фазных мышц — около 2 с, что в 30 раз больше токового у скелетной мышцы. Кроме того, гладкая мышца способна находиться в состоянии длительного тонуса, практически не затрачивая энергии. Энергетическое обеспечение сокращения гладких мышц также осуществляется за счет молекул АТФ. Ресинтез ее в гладкомышечных клетках в основном является результатом анаэробного гликолиза. мВ мВ 0 0 –50 –50 0 0 Сокращение Сокращение Рис. 2.14. Потенциалы действия и сокращения гладких мышц (по Б.И. Ткаченко, 2005) ГЛАВА 3 Медицина — родина и источник материализма. Л. Фейербах Характеристика регуляции функций организма Регуляция функций — это изменение интенсивности работы органов и систем для достижения полезного результата согласно потребностям организма в различных условиях его жизнедеятельности. Механизмы регуляции обеспечивают взаимодействие клеток, органов и систем организма, объединяя его в единое целое. Классифицировать регуляцию целесообразно по двум основным критериям: по механизму ее осуществления (три механизма: нервный, гуморальный и миогенный) и по времени ее включения относительно момента изменения величины регулируемого показателя организма (два типа регуляции: по отклонению и по опережению). Ведущим является нервный механизм регуляции функций организма. 3.1. Нервный механизм регуляции Выделяют пусковое и модулирующее (корригирующее) влияние нервной системы на орган. Пусковое влияние вызывает деятельность органа, находящегося в покое. Примером такого влияния может служить инициация сокращений покоящейся скелетной мышцы при поступлении к ней импульсов от мотонейронов спинного мозга или от мотонейронов ствола мозга по эфферентным (двигательным) нервным волокнам. Пусковое влияние нервной системы обычно осуществляется с помощью электрофизиологических процессов. После Глава 3. Характеристика регуляции функций организма 65 прекращения поступления импульсов по нервному волокну к мышце сокращение также прекращается, мышца расслабляется. Модулирующее (корригирующее) влияние вызывает изменение интенсивности деятельности органа. Модулирующее влияние реализуется: • с помощью изменения интенсивности биохимических процессов — обмена веществ в органе (трофическое действие нервной системы); • посредством изменения характера электрофизиологических процессов в органе (гиперполяризация, деполяризация — электрофизиологическое действие); • за счет изменения кровоснабжения органа (сосудодвигательный эффект). Все они взаимосвязаны. Модулирующее влияние, например, блуждающего нерва на сердце выражается в угнетении его сокращений, но этот же нерв может оказывать пусковое влияние на покоящуюся гладкую мышцу желудка. Рефлекторный принцип нервной регуляции. Рефлекс — подсознательная ответная реакция организма на раздражение сенсорных рецепторов, осуществляемая с помощью нервной системы. Ответная реакция может быть в виде активации или торможения деятельности органа. Каждый рефлекс осуществляется посредством рефлекторной дуги. Рефлекторная дуга — это совокупность структур, при помощи которых осуществляется рефлекс. Схематично рефлекторную дугу вегетативного и соматического рефлексов можно представить состоящей из пяти звеньев (рис. 3.1). 1. Воспринимающее звено (рецептор) обеспечивает восприятие изменений внешней или внутренней среды организма, что достигается посредством трансформации энергии раздражения в рецепторный потенциал. Совокупность рецепторов, раздражение которых вызывает рефлекс, называют рефлексогенной зоной. Конкретные механизмы восприятия раздражителя (света, звука, изменения химического состава крови и др.) различны у каждого вида рецепторов, но во всех случаях возникают рецепторные 66 Физиология потенциалы, обеспечивающие посылку нервных импульсов в ЦНС с помощью второго звена. 2. Афферентное звено. Для соматической нервной системы это афферентный нейрон с его отростками — тело его расположено в спинномозговых ганглиях или ганглиях черепных нервов. Импульс поступает по дендриту афферентного нейрона, а затем по его аксону в ЦНС. Роль этого звена заключается в передаче сигнала в ЦНС к третьему звену рефлекторной дуги. 3. Управляющее звено — совокупность центральных (для ВНС) и периферических нейронов, формирующих ответную реакцию организма. 4. Эфферентное звено — аксон эффекторного нейрона (для соматической нервной системы — мотонейрона). 5. Эффектор (рабочий орган). В простейшем случае рефлекторная дуга может быть двухнейронной — без вставочного нейрона. Эффекторным нейроном соматической нервной системы является мотонейрон, вегетативной — нейрон ганглия. 2 3 1 4 5 Рис. 3.1. Рефлекторная дуга соматического рефлекса: 1 — воспринимающее звено; 2 — афферентное звено; 3 — управляющее звено; 4 — эфферентное звено; 5 — эффектор Глава 3. Характеристика регуляции функций организма 67 Нервизм — это концепция о ведущей роли нервной системы в регуляции функций всех органов и тканей организма (физиологический нервизм). Концепция нервизма прошла весьма длинный исторический путь развития. Основоположником этой концепции является Р. Декарт (1596–1650), выдвинувший идею о рефлекторном принципе деятельности нервной системы. Особо важную роль в развитии концепции нервизма сыграли И.М. Сеченов (1829–1905), С.П. Боткин (1832–1889) и И.П. Павлов (1849–1936). Нарушение деятельности нервной системы может привести к заболеванию (клинический нервизм). Нервный центр — это скопление нейронов, достаточное для регуляции функции органа согласно потребностям организма. Нервный центр реализует свое влияние на эффекторы либо непосредственно с помощью эфферентных импульсов соматической и вегетативной нервной системы, либо с помощью активации выработки соответствующих гормонов. 3.2. Гуморальная и миогенная регуляция Гормональное звено в случае рефлекторной регуляции функций организма включается с помощью вегетативной нервной системы. Виды влияний гормонов. Гормональное влияние на органы, ткани и системы организма подразделяется на функциональное (изменение интенсивности работы организма) и морфогенетическое — обеспечивает морфогенез (рост, физическое и половое развитие, созревание структур ЦНС). Функциональное влияние гормонов бывает трех видов: пусковое, модулирующее и пермиссивное. • Пусковое влияние — это способность гормона запускать деятельность эффектора, например, только в присутствии АДГ стенки дистального сегмента нефрона проницаемы для воды. • Модулирующее влияние — изменение интенсивности деятельности органа. Например, стимуляция адреналином деятельности сердца, которая осуществляется и без адреналина. 68 Физиология • Пермиссивное влияние — способность одного гормона обеспечивать реализацию эффекта другого гормона. Например, инсулин необходим для проявления действия соматотропного гормона. Морфогенетическое влияние гормонов на рост, физическое и половое развитие. Оба вида влияний гормонов (морфогенетическое и функциональное) реализуются с помощью метаболических процессов, запускаемых посредством клеточных ферментных систем. Регуляция с помощью метаболитов — продуктов, образующихся в организме в процессе обмена веществ (угольная, молочная, пировиноградная, адениловая кислоты, аминокислоты, нуклеотиды). Например, накопление СО2 в организме стимулирует дыхание, действие угольной и молочной кислот ведет к расширению кровеносных сосудов. Миогенный механизм регуляции. В процессе эволюции этот вид регуляции бурно прогрессировал с развитием мышечной ткани, что связано с интенсификацией двигательной активности животных. Организм человека примерно на 50% состоит из мышц по своей массе. Это скелетная мускулатура (40% массы тела), мышца сердца и гладкие мышцы внутренних органов (сосудов, стенки желудочнокишечного тракта и мочевого пузыря и др.). Сущность миогенного механизма регуляции состоит в том, что предварительное умеренное растяжение скелетной или сердечной мышц увеличивает силу их сокращений. Сократительная активность гладкой мышцы также зависит от степени ее растяжения. При увеличении наполнения органа тонус гладкой мышцы сначала возрастает, а затем возвращается к исходному уровню (пластичность гладкой мышцы), что обеспечивает регуляцию тонуса сосудов и наполнение внутренних полых органов без существенного повышения давления в них. 3.3. Единство и особенности регуляторных механизмов Единство регуляторных механизмов заключается в их взаимодействии. Так, при действии холодного воздуха на Глава 3. Характеристика регуляции функций организма 69 терморецепторы кожи увеличивается поток афферентных импульсов в ЦНС, это ведет к выбросу гормонов, увеличивающих интенсивность обмена веществ и к увеличению теплопродукции. Ядра гипоталамуса вырабатывают нейрогормоны, регулирующие функцию эндокринных желез с помощью аденогипофиза. В случае увеличения содержания углекислого газа в крови возбуждаются хеморецепторы аортальной и синокаротидной рефлексогенных зон, при этом увеличивается поток импульсов по соответствующим нервам в ЦНС, а оттуда — к дыхательной мускулатуре, что ведет к учащению и углублению дыхания. Углекислый газ действует на дыхательный центр с помощью рефлексогенных зон и непосредственно на дыхательный центр, что также вызывает усиление дыхания. Основные особенности нервного и гуморального механизмов регуляции функций организма. Нервная регуляция — ведущая, нервной системе подчиняется эндокринная система. Различная скорость связи. Нервный импульс распространяется с огромной скоростью — до 160 м/с; химические вещества распространяются с током крови относительно медленно, частица крови проходит один раз через весь организм за 22 с. У гуморального и нервного механизмов регуляции функций различные способы связи: у нервной системы — нервный импульс как универсальный сигнал, а у гуморального механизма связь с регулируемым органом или тканью осуществляется с помощью различных химических веществ (гормоны, медиаторы, метаболиты и парагормоны). Различная точность связи. Нервная система может оказывать точное, локальное влияние на отдельный орган или даже на группу клеток этого органа. Так, нервная система может вызывать сокращения мышц указательного или другого пальца руки, не вызывая сокращения мышц всей конечности или других пальцев. Химические вещества (например, адреналин, тироксин), попадая в кровь, разносятся по всему организму и действуют нередко на многие органы и ткани — генерализованный характер влияния. Следует заметить, что и у гуморального механизма нередко имеется точный адресат воздействия. Кортикотропин хотя и разно- 70 Физиология сится с кровью по всему организму, но действует только на кору надпочечников. У гуморального механизма регуляции нередко наблюдается противоположное влияние биологически активных веществ на один и тот же орган в зависимости от точки приложения действия этого химического вещества. Так, например, угольная кислота, действуя прямо на кровеносные сосуды, вызывает их расширение, а посредством возбуждения центра кровообращения — сужение. Это наблюдается в тех случаях, когда вещество проникает через гематоэнцефалический барьер (ГЭБ) — совокупность структур, препятствующих проникновению микробов и ряда веществ из крови в мозг. 3.4. Системный принцип регуляции и его типы Cистемный принцип регуляции заключается в том, что различные показатели организма поддерживаются на оптимальном уровне с помощью многих органов и систем. Например, Ро2 и Рсо2 обеспечиваются деятельностью систем кровообращения, дыхания, нервно-мышечной, крови. Они объединяются в различные функциональные системы. Представление о функциональных системах разработал П.К. Анохин в середине XX в. В последние годы теория функциональных систем успешно развивается К.В. Судаковым и сотрудниками. Функциональная система — это динамическая совокупность различных органов и физиологических систем, объединяющихся для достижения приспособительного (полезного для организма) результата (рис. 3.2). Различают гомеостатические и поведенческие функциональные системы. Гомеостатические функциональные системы обеспечивают поддержание на оптимальном для метаболизма уровне различных показателей организма (например, pH, АД). Это достигается посредством изменения интенсивности работы внутренних органов. Полезный результат поведенческой функциональной системы — бытовой, социальный успех (получение образования, достижение выдающихся спортивных результатов и т.п.), удовлетворение биологической потребности. Глава 3. Характеристика регуляции функций организма 71 Поведенческая регуляция РП Эффектор-1 Нервный центр Рецепторы регулируемого показателя Эффектор-2 Эффектор-3 Гормональная регуляция Эффектор-4 Метаболиты Гуморальные влияния Нервные влияния Обратная связь Рис. 3.2. Общая схема гомеостатической функциональной системы — регуляции физиологических показателей (по П.К. Анохину, с изменениями): РП — регулируемый показатель Любые функциональные системы формируются из физиологических систем. Физиологическая система — генетически сформированная совокупность органов и тканей организма, выполняющая определенную функцию (например, система дыхания). Структура физиологической системы включает следующие элементы: • управляющий центр, представляющий объединение ядер различных уровней ЦНС; • его выходные каналы (нервы и гормоны); • исполнительные органы — эффекторы, обеспечивающие в ходе физиологической деятельности поддержание регулируемого показателя (РП) на некотором оптимальном уровне; • каналы обратной связи (входные каналы), информирующие нервный центр с помощью афферентной импульсации от рецепторов результата (сенсорных рецепторов) о достигнутом результате. По такому же принципу формируется и функциональная гомеостатическая система, за исключением того, что она включает несколько физиологических систем. 72 Физиология Типы регуляции функций организма. Как отмечалось выше, имеется два типа регуляции — по отклонению и по опережению. Рассмотрим их подробнее. Регуляция по отклонению — ведущая в организме и заключается в том, что всякое отклонение регулируемого показателя от оптимального уровня мобилизует физиологические системы к восстановлению его на прежнем уровне. Регуляция по отклонению осуществляется с помощью обратной отрицательной связи, обеспечивающей разнонаправленное влияние — усиление стимулирующих механизмов управления (в случае ослабления показателей организма) или их ослабление (в случае чрезмерного усиления показателей организма). В отличие от отрицательной обратной связи, положительная обратная связь встречается в организме редко; ее пример — усиление начавшейся секреции пищеварительных соков в желудке после приема пищи с помощью продуктов гидролиза, всосавшихся в кровь. Положительная обратная связь оказывает только однонаправленное влияние на развитие процесса, находящегося под контролем управляющего центра. Поэтому положительная обратная связь делает систему неустойчивой, неспособной обеспечить стабильность регулируемого показателя в пределах физиологического оптимума. Например, если бы АД регулировалось по принципу положительной обратной связи, то в случае понижения АД действие регуляторных механизмов привело бы к еще большему его снижению, а в случае повышения — к еще большему его увеличению. Вся безусловно-рефлекторная регуляция (в том числе с включением гормонов) осуществляется по отклонению. Регуляция по опережению заключается в том, что регулирующие механизмы включаются до реального изменения показателя на основе информации, поступающей в ЦНС. Регуляция по опережению встречается в организме редко. Например, усиление вентиляции легких при физической нагрузке начинается раньше изменений газового состава крови организма. Это происходит благодаря афферентной импульсации от проприорецепторов активно работающих мышц к моторным центрам и взаимодействию их с дыхательным центром. Но и здесь регуляция по от- Глава 3. Характеристика регуляции функций организма 73 клонению ведущая, поскольку главными стимуляторами дыхания при физической работе являются изменение газового состава крови, рН внутренней среды и накопление метаболитов. По опережению (на 1–2 с) расширяются сосуды мышц при планировании движения. Главную роль в регуляции кровоснабжения работающих мышц играют метаболические сдвиги, которые развиваются с началом сократительной деятельности мышц (рабочая гиперемия), причем местное действие метаболитов выражено сильнее нервных влияний, в том числе и вазоконстрикторов. Таким образом, данный пример демонстрирует ведущую роль регуляции по отклонению. Механизм условного рефлекса, несомненно, обеспечивает регуляцию по опережению. Например, у спортсмена на старте заранее усиливается деятельность ряда систем (дыхания, сердечно-сосудистой, эндокринной). После начала физической деятельности (например, бег, гонка на лыжах) ведущим типом регуляции является также регуляция по отклонению (метаболический фактор). Регуляция по опережению дополняет регуляцию по отклонению и делает единую функциональную систему — организм — весьма устойчивой в различных условиях жизнедеятельности. Есть жесткие показатели организма (колеблющиеся в узком диапазоне) и гибкие (колеблющиеся в широком диапазоне). Примером первых являются осмотическое давление плазмы крови, кислотно-основное состояние организма (КОС), характеризующееся рН крови (7,35–7,45), сдвиг которого на 0,3 несовместим с жизнью. Примером гибких показателей являются артериальное давление, частота сердечных сокращений: в покое равна 75 в минуту, при большой физической нагрузке может достигать 150 в минуту и больше. ГЛАВА 4 Центральная нервная система новорожденного ничего не умеет, но она может всему научиться. Л.О. Бадалян Физиология центральной нервной системы ЦНС человека содержит свыше 100 млрд нервных клеток, взаимодействие между которыми осуществляется посредством множества синапсов, которых в тысячи раз больше самих клеток (1015–1016), т.к. их аксоны многократно делятся дихотомически. Нейроны оказывают влияние на органы и ткани также посредством синапсов. Ряд заболеваний у человека сопровождается повреждением нервной ткани. Например, развитие неврологических нарушений у больных СПИДом связано с гибелью 40–50% нейронов коры большого мозга. 4.1. Структурно-функциональная характеристика ЦНС Нервная система в организме выполняет интегрирующую роль. Она объединяет в единое целое все ткани, органы, координируя их специфическую активность в составе целостных гомеостатических и поведенческих функциональных систем (Анохин П.К.). Основные частные функции нервной системы: • обеспечение восприятия и хранения информации (памяти) и психической деятельности; • управление деятельностью опорно-двигательного аппарата; • восприятие изменений внешней и внутренней среды организма и формирование приспособительных реакций; Глава 4. Физиология центральной нервной системы 75 • регуляция работы внутренних органов в различных условиях жизнедеятельности, что осуществляется с помощью автономной (вегетативной) нервной системы и эндокринных желез. Любая деятельность самой ЦНС реализуется в конечном итоге с помощью функционирования отдельных нервных клеток. Нервная клетка (нейрон) — структурная и функциональная единица ЦНС, она состоит из сомы (тела клетки с ядром) и отростков, представляющих собой большое число дендритов и один аксон (рис. 4.1). Сома (тело нейрона) синтезирует макромолекулы, которые транспортируются по аксону и дендритам, выполняет трофическую функцию по отношению к своим отросткам и клеткам-эффекторам. Отросток, лишенный связи с телом нейрона, дегенерирует. Нейроны ЦНС образуют скопления нейронов (ядра и слои), выполняющие определенные функДендриты Пресинаптические окончания Интеграция Тело ВПСП и ПД Аксонный холмик Проведение импульса Аксон Узловой перехват Миелиновая оболочка Секреция медиатора Рис. 4.1. Структурно-функциональные элементы мотонейрона спинного мозга (по Р. Эккерту, Д. Ренделлу, Дж. Огастину, 1991, с изменениями) 76 Физиология ции. Сома и дендриты каждого нейрона покрыты нервными окончаниями — синаптическими бутонами других нервных клеток и отростками глиальных клеток. На одном мотонейроне число синаптических бутонов может достигать 10–20 тыс. (см. рис. 4.1). Аксон начинается от наиболее возбудимого участка тела клетки, получившего название аксонного холмика. Потенциал покоя (ПП) нейрона составляет 60–80 мВ, потенциал действия (ПД) равен 80–110 мВ. Отдельные структурные элементы нейрона выполняют определенные функции: • структурами, воспринимающими импульсы от других нервных клеток, являются тело и дендриты нейрона с шипиками и расположенными на них постсимпатическими мембранами. Если шипики не получают синаптических сигналов, они исчезают. Сигналы через синапсы могут поступать и к аксонам (аксоаксонные синапсы), например в случае пресинаптического торможения. Число дендритов у нейрона весьма вариабельно — от 1 до 1500 (рис. 4.2); • структуры, где возникает ПД, — аксонный холмик и тело нейрона, причем зарождается ПД на аксонном холмике (генераторный пункт); • структура, проводящая возбуждение к другому нейрону или к эффекторной клетке, — аксон; • структура, передающая сигналы на другую клетку, — это пресинаптический аппарат, представляющий со- Б А Клетка Пуркинье Пирамидный нейрон В Г Нейрон ядра таламуса Нейрон ядра тройничного нерва Рис. 4.2. Множество дендритов на теле нейрона (А, В) и многократное деление аксона нейрона (Б, Г) различных отделов ЦНС Глава 4. Физиология центральной нервной системы 77 бой окончание аксона с пресинаптической мембраной. В нем синтезируется и запасается медиатор. Нейроны ЦНС классифицируют по нескольким признакам: • по виду синтезируемого медиатора различают нейроны адренергические, серотонинергические, холинергические и др.; • по влиянию на другие клетки — возбуждающие и тормозящие нейроны; • по специфичности воспринимаемой сенсорной информации нейроны высших отделов ЦНС могут быть моно-, би- и полисенсорными. Моносенсорными являются нейроны первичных сенсорных зон в коре большого мозга, например центров слуха; • по активности — фоновоактивные и молчащие нейроны, возбуждающиеся только в ответ на действие раздражителя. В зависимости от отдела нервной системы выделяют нейроны соматической и автономной (вегетативной) нервной системы (регулирующей функции внутренних органов) и др. Синапсы ЦНС также классифицируют по нескольким признакам: • в зависимости от местоположения на теле или отростках нейронов синапсы делятся на аксосоматические, аксодендритные, аксоаксонные, дендродендритные, дендросоматические; • по механизму передачи сигнала к другим клеткам имеются химические синапсы — их большинство, электрические и смешанные (электрохимические) — их мало. Химические синапсы (рис. 4.3) имеют относительно широкую синаптическую щель, составляющую 20–50 нм. В пресинаптической терминали содержится большое число пузырьков — пресинаптических везикул диаметром около 50 нм, заполненных медиатором — химическим передатчиком. Медиатор либо образуется в теле нейрона и транспортируется в пресинаптическое окончание аксонным транспортом, либо синтезируется непосредственно в пресинаптическом окончании и накапливается в везикулах. 78 Физиология – Рис. 4.3. Структурно-функциональная организация химического синапса ЦНС (по А.К. Гайтону, Д.Э. Холлу, 2008) Для синтеза медиатора нужны ферменты, образующиеся в теле клетки на рибосомах. По механизму активации постсинаптические рецепторы химических синапсов делятся на два класса: • ионотропные (например, N-холинорецепторы), активация ионных каналов которых осуществляется непосредственно медиатором, в результате чего возникает возбуждение; • метаботропные (их большинство), когда через мембранный рецептор, связанный с G-белком, ионные каналы активируются с помощью вторых посредников (цАМФ, цГМФ, инозитолтрифосфат, диацилглицерол и др.), в результате чего формируются биохимические реакции, а потом и электрофизиологический ответ. Глиальные клетки (нейроглия — нервный клей, от греч. neuron — нерв, glia — клей) более многочисленны, чем нейроны, составляют около 50% объема ЦНС, они способны к делению в течение всей жизни. Их размеры в 3–4 раза меньше, чем у нервных клеток, а число в 10 раз больше, чем нейронов, с возрастом оно увеличивается (число нейронов уменьшается). Глиальные клетки окружают тела нейронов и их аксоны. Глиальные клетки выполняют несколько функций: • электроизолирующую (глиоциты окутывают нейроциты в виде своеобразного футляра и продуцируют миелин), мембранный потенциал клеток нейроглии составляет 70–90 мВ, ПД они не генерируют, возни- Глава 4. Физиология центральной нервной системы 79 кают лишь локальные потенциалы, их электрические поля взаимодействуют с постсинаптическими потенциалами (ВПСП и ТПСП) и ПД нейронов. При некоторых заболеваниях функция нейроглии нарушается. Например, при рассеянном склерозе вследствие повреждения миелиновой оболочки происходит утечка ионов из нервных волокон, поэтому нарушается их возбудимость и проводимость; • опорную; • защитную (выработка цитокинов, фагоцитоз); • обменную функцию (синтез белков памяти, снабжение нейронов питательными веществами, поддержание оптимальной концентрации ионов К+ в интерстиции; • продвижение аксоплазмы. Микроглиальные клетки способны к ритмическому изменению своего объема (период «сокращения» — 1,5 мин, «расслабления» — 4 мин). Циклы изменения объема повторяются через каждые 2–20 ч. Полагают, что пульсация способствует продвижению аксоплазмы в нейронах и влияет на ток межклеточной жидкости. Глиоциты, охватывая снаружи капилляры ЦНС, участвуют в формировании гематоэнцефалического барьера. Гематоэнцефалический барьер (ГЭБ) — барьер, препятствующий проникновению из крови в мозг микробов и ряда веществ. Наибольший вклад в развитие представлений о гематоэнцефалическом барьере внесла Л.С. Штерн. Регулирующая функция ГЭБ заключается в том, что он формирует особую внутреннюю среду мозга, обеспечивающую оптимальный режим деятельности нервных клеток, и избирательно пропускает многие вещества, содержащиеся во внутренней среде организма, например воду, ионы, глюкозу, аминокислоты, газы. При этом ГЭБ задерживает многие физиологически активные вещества: дофамин, инсулин, адреналин, тироксин, серотонин. Проникающие в ЦНС вещества могут оказывать противоположное влияние на орган, по сравнению с местным действием. Углекислый газ, например, действует на сердечно-сосудистый центр, вызывает сужение сосудов, а при местном действии — их расширение. 80 Физиология ГЭБ формируется особыми структурами стенок капилляров мозга — их эндотелием, а также базальной мембраной, окружающей капилляры снаружи. Эндотелий капилляров, входящих в состав ГЭБ, имеет очень мало пор, узкие щелевые контакты, между клеток почти нет окошек. Составная часть барьера — глиальные клетки, образующие своеобразные футляры вокруг капилляров, покрывающие около 90% их поверхности. ГЭБ выполняет защитную функцию — предотвращает попадание микробов, чужеродных или токсичных веществ экзо- и эндогенной природы в межклеточные пространства мозга. ГЭБ не пропускает многие лекарственные вещества, что необходимо учитывать в клинической практике. При воздействии на ЦНС с помощью лекарственных средств следует подбирать препараты, которые проникают через ГЭБ. Ликвор — бесцветная прозрачная жидкость, заполняющая мозговые желудочки, спинномозговой канал и субарахноидальное пространство. Ее происхождение связано с интерстициальной жидкостью мозга, значительная часть ликвора образуется сосудистыми сплетениями желудочков мозга. Интерстициальная жидкость является непосредственно питательной средой клеток мозга, в нее клетки выделяют также продукты своего обмена. Таким образом, ликвор представляет собой совокупность фильтрата плазмы крови и интерстициальной жидкости. Его объем — 100–200 мл, за сутки вырабатывается около 600 мл. Обновление этой жидкости происходит 4–8 раз в сутки, давление ликвора при горизонтальном положении человека составляет 7–14 мм рт. ст., в вертикальном положении — в 2 раза больше. Функции ликвора: • защищает ЦНС от микробов (в случае их попадания в кровь), т.к. обладает бактерицидными свойствами, содержит иммуноглобулины G и А, систему комплемента, моноциты и лимфоциты; • является своеобразной гидравлической «подушкой» мозга; • выполняет питательную функцию для нейронов ЦНС и дренажную (препятствие оттоку ликвора в кровь может сопровождаться тяжелыми нарушениями в деятельности ЦНС). Глава 4. Физиология центральной нервной системы 81 4.2. Медиаторы и рецепторы ЦНС. Возбуждение нейронов ЦНС Медиаторы ЦНС. Медиатор (от лат. mediator — посредник) — химическое вещество, с помощью которого передается сигнал от одной клетки к другой в химическом синапсе. Медиатор является первичным посредником в метаботропных рецепторах (см. раздел 2.4). Рецептор (от лат. receptor — воспринимающий). Постсинаптический рецептор — это совокупность рецепторных молекул, которые являются интегральными белками постсинаптической мембраны. Рецепторная молекула имеет ионный канал. Он специфически взаимодействует с медиаторами и химическими блокаторами, широко применяемыми в медицине. Рассмотрим основные медиаторы и рецепторы ЦНС. На одну клетку могут действовать несколько медиаторов (рис. 4.4). Основные биогенные амины (дофамин, норадреналин, серотонин, гистамин) встречаются в разных отделах ЦНС, в значительных количествах содержатся в нейронах ствола мозга. Амины обеспечивают возникновение процессов возбуждения и торможения, например в промежуточном мозге, в среднем мозге, в лимбической системе, в полосатом теле. Ацетилхолин как медиатор в ЦНС встречается довольно широко. Обнаружены М- и N-холинорецепторы. N-холинорецепторы — холинорецепторы, чувствительные к алколоиду, выделенному из листьев табака, — никотину. ГАМК Глицин Норадреналин Глутамат Серотонин ДНК Ацетилхолин ↓ РНК ↓ Белок Рис. 4.4. Функция нейрона может стимулироваться или угнетаться несколькими медиаторами 82 Физиология M-холинорецепторы — холинорецепторы, чувствительные к алкалоиду, выделенному из красного мухомора, — мускарину. Установлено 7 подтипов М-холинорецепторов. Ацетилхолин активирует как возбуждающие, так и тормозные нейроны, что и определяет его эффект. Аминокислоты. Глицин и гамма-аминомасляная кислота (ГАМК) являются тормозными медиаторами в синапсах ЦНС и действуют на соответствующие рецепторы: глицин — в основном в спинном мозге, ГАМК — в коре большого мозга, мозжечке, стволе мозга, спинном мозге, в интернейронах таламуса. Глутамат и аспартат передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гиппокампа, коры большого мозга. Глутамат — это основной возбуждающий медиатор ЦНС (75% возбуждающих синапсов мозга). Глутамат — медиатор первичных болевых афферентов. Нейропептиды встречаются в синапсах различных отделов ЦНС. Энкефалины и эндорфины — опиоидные медиаторы нейронов, блокирующих, например, болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиатных рецепторов, которые особенно плотно располагаются на клетках лимбической системы, много их также в ядрах промежуточного мозга и др. Вещество Р является медиатором нейронов, передающих сигналы боли. Особенно много этого нейропептида содержится в дорсальных корешках спинного мозга. Физиологические эффекты действия некоторых медиаторов головного мозга. Дофамин участвует в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, участвует в механизмах формирования некоторых фаз сна, сновидений. Серотонин ускоряет процессы обучения, угнетения болевых ощущений, сенсорное восприятие, засыпание. Эндорфины и энкефалины вызывают антиболевые эффекты, повышение устойчивости к стрессу. Пептид, вызывающий дельта-сон, участвует в меха- Глава 4. Физиология центральной нервной системы 83 низмах ортодоксального сна. Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны и вторичных посредников. Ацетилхолин, например, в коре мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца — только торможение, в синапсах гладкой мускулатуры желудочно-кишечного тракта — только возбуждение. Катехоламины (норадреналин, адреналин, дофамин) тормозят сокращения желудка и кишечника, но стимулируют сердечную деятельность. Механизм возбуждения нейронов ЦНС. Передача сигнала в химических синапсах ЦНС подобна таковой в нервно-мышечном синапсе, но имеется ряд отличительных особенностей, основные из которых приведены ниже. Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных импульсов и их взаимодействие. Это объясняется тем, что один ПД, пришедший в пресинаптическое окончание, обеспечивает высвобождение небольшого количества медиатора и, соответственно, небольшого возбуждающего постсинаптического потенциала (ВПСП) (рис. 4.5). Необходимо учесть, что одновременно на теле клетки с расположенными на ней синапсами могут возникать не только возбуждающие, но и тормозные потенциалы. Поэтому требуется большое число ПД, приходящих к нейрону для его возбуждения. Механизм выброса медиатора из нервного окончания подобен таковому в нервно-мышечном синапсе (см. раздел 2.3). Поступивший в пресинаптическое окончание Са2+ удаляется за его пределы с помощью Са-насоса. Прекращение –64 ВПСП 7 мс ПП = –70 –74 Рис. 4.5. Возбуждающий постсинаптический потенциал (ВПСП) нейрона 84 Физиология действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично — с помощью разрушения специальными ферментами. В частности, норадреналин расщепляется моноаминоксидазой и катехолметилтрансферазой, ацетилхолин гидролизуется ацетилхолинэстеразой, имеющейся в синаптической щели и встроенной в постсинаптическую мембрану. Место возникновения ВПСП — тело нейрона и дендриты. Однако зарождается ПД нейрона на аксонном холмике, хотя синапсов на нем нет. Близость ВПСП, возникающих на теле нейрона, к аксонному холмику обеспечивает за счет их электрического поля деполяризацию аксонного холмика до КУД. Это объясняется тем, что возбудимость аксонного холмика (он не имеет миелиновой оболочки) в 3–4 раза выше возбудимости тела нейрона. Развивающееся возбуждение на аксонном холмике обеспечивает деполяризацию до КУД и тела нейрона. Далее идет одновременный регенеративный процесс деполяризации аксонного холмика и тела нейрона. ПД не распространяется с аксонного холмика на тело нейрона. ПД нейрона — это единый процесс аксонного холмика и тела нейрона. Об этом, в частности, свидетельствует перегиб на восходящей части ПД нейрона — задержка развития ПД из-за более низкой возбуждаемости тела нейрона по сравнению с возбудимостью аксонного холмика (рис. 4.6). 2 мВ +25 0 –25 1 3 КУД –50 5 мс ПП–75 4 Рис. 4.6. Типичный ПД нейронов ЦНС: 1 — деполяризация; 2 — инверсия; 3 — реполяризация; 4 — следовая гиперполяризация; 5 — следовая деполяризация Глава 4. Физиология центральной нервной системы 85 Роль дендритов в возникновении возбуждения нейрона. Большинство синапсов нейрона находится на дендритах. Эти синапсы получили название модуляторных, т.к. они находятся на значительном расстоянии от генераторного пункта нейрона — аксонного холмика. Поэтому их ВПСП не могут вызвать достаточную деполяризацию аксонного холмика и обеспечить возникновение ПД, а определяют лишь возбудимость нейрона. При возбуждении нейронов потребление О2 значительно возрастает (ЦНС потребляет 20% О2, расходуемого организмом в покое). Источником энергии является в основном глюкоза крови, собственных небольших запасов гликогена достаточно лишь на 3–5 мин работы нейрона. 4.3. Принципы организации деятельности ЦНС Принципы организации деятельности ЦНС — это варианты взаимодействия нейронов и центров ЦНС. В организации деятельности ЦНС имеется ряд принципов: рефлекторный, принципы сетей, связей и др. Нейронные сети. По распространенности выделяют локальные и распространенные сети. Локальные сети — это сети, обеспечивающие возникновение возбуждения или торможения в ограниченных участках ЦНС. Примерами локальных сетей являются синаптически связанные нейроны, обеспечивающие циркуляцию возбуждения (рис. 4.7), варианты пре- и постсинаптического торможения, например реципрокного торможения, межнейронные связи в лимбической системе, где выражена циркуляция возбуждения. Вместо термина «локальные 1 2 3 Рис. 4.7. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто-де-Но: 1, 2, 3 — возбуждающие нейроны 86 Физиология сети» некоторое авторы использует также термин «контуры», что нецелесообразно, т.к. это создает терминологическую путаницу. Распространенные сети. По принципу распространенных сетей построена ретикулярная формация, нейроны которой имеют множество внутренних связей и связей с другими отделами ЦНС. Нейронные связи. По характеру различают жесткие и гибкие связи. Жесткие связи — это связи, с помощью которых нервное влияние осуществляется строго по одному и тому же пути. Например, управление функцией α-мононейронов спинного мозга с помощью ядер ствола мозга или с помощью пирамидных клеток двигательной коры большого мозга. Примером жесткой связи в ЦНС являются также рефлекторные дуги безусловных рефлексов, замыкающиеся на уровне спинного мозга. Ярким примером являются проприоцептивные безусловные рефлексы, которые осуществляются всего лишь с помощью двух нейронов — чувствительного и α-мононейрона (моносимпатическая рефлекторная дуга). Жесткие связи обеспечиваются односторонним распространением возбуждения в химических синапсах. Одностороннее распространение возбуждения (от аксона одного нейрона к телу или дендритам другого нейрона) объясняется тем, что на пресинаптическом окончании нет рецепторов для медиатора, который секретируется в синаптическую щель. По жесткому принципу связи построены локальные сети. Гибкие связи — это связи, в которых может участвовать различное число нейронов — сотни и тысячи. Они широко встречаются в различных отделах ЦНС. Это обеспечивается дивергенцией и конвергенцией нервных путей. Дивергенция возбуждения в ЦНС (лат. divergere — направляться в разные стороны, менее точный термин — иррадиация, от лат. irradiare — озарять, освещать) объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.8, А). Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов. Дивергенцию возбуждения можно легко наблюдать Глава 4. Физиология центральной нервной системы 87 в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное — энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Конвергенция возбуждения (принцип общего конечного пути) — схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному модулю (принцип шеррингтоновской воронки). Это тоже пример гибкой связи. Объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС могут располагаться до 10 000 синапсов (на мотонейронах — до 20 000). Например, мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чихания и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг (рис. 4.8, Б). Временные связи, формирующиеся в процессе выработки условного рефлекса, — тоже пример гибких связей. По направлению информации различают прямые, обратные, возвратные и реципрокные связи (см. разд. «Координационная деятельность ЦНС»). Рефлекторный принцип деятельности ЦНС — один из важных принципов организации работы ЦНС. Он является базисным в безусловно-рефлекторной регуляции функций внутренних органов, в формировании ответных реакций организма на внешние воздействия, в сохранении естественной позы и восстановлении нарушенной позы. Скопления различного количества нейронов образуют модули и нервные центры. Спинальные нейроны Афференты Рецептор А Кора большого мозга Ядра ствола мозга Ретикулярная формация ствола мозга α-мотонейрон Б Афференты Рис. 4.8. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (А), и конвергенция эфферентных путей от различных отделов ЦНС на α-мотонейрон спинного мозга (Б) 88 Физиология Принцип нервных центров. Нервный центр — это скопление взаимосвязанных нейронов, обеспечивающих регуляцию функции органа. Например, центр сгибания или разгибания нижней конечности локализован в спинном мозге (L2–S5). Дыхательный центр расположен в продолговатом мозге и т.д. Более широкое понятие — «ядро». Ядро — совокупность связанных нейронов, выполняющих общую функцию. Отличается от нервного центра только в том, что центр обеспечивает регуляцию функций органов, посылая к ним эфферентные импульсы. Ядра могут быть сенсорными и двигательными (например, ядра тройничного нерва). Имеются двигательные ядра без выхода на эффекторы (например, красное ядро и ядро Дейтерса). Нейронный модуль — часть нервного центра или ядра, выполняющая определенную роль в их деятельности. Например, модуль ранних инспираторных нейронов в дыхательном центре обеспечивает начало вдоха. Модуль поздних инспираторных нейронов — прекращение вдоха. Модуль депрессорных нейронов центра кровообращения (продолговатый мозг) обеспечивает снижение артериального давления при активной импульсации от сосудистых рефлексогенных зон при высоком артериальном давлении. Колонки коры большого мозга, воспринимающие сенсорную информацию от определенной части тела, сетчатки и др., также являются примером нейронного модуля. Авторы используют три термина («нейронный пул», «нейронный ансамбль», «нейронный модуль») вне связи их друг с другом, что затрудняет восприятие материала. С нашей точки зрения, более удачным является термин «нейронный модуль». Термин «ансамбль» воспринимается как более крупное объединение. Например, ансамбль нервных ядер ствола мозга, регулирующих тонус скелетной мускулатуры. Иерархический принцип организации деятельности ЦНС наблюдается в эфферентных и афферентных системах мозга. В эфферентных системах мозга иерархия заключается в подчинении нижележащих отделов ЦНС вышележащим и в усложнении двигательных реакций с помощью вышележащих отделов ЦНС. Например, ядра ствола (красное ядро, ядро Дейтерса) управляют функцией мотонейронов спинно- Глава 4. Физиология центральной нервной системы 89 го мозга, функция самих ядер ствола мозга регулируется базальными ядрами (стриопалидарной системой). Двигательные области коры большого мозга управляют функциями всех нижележащих двигательных систем ЦНС (например, при осуществлении произвольных движений). Чем больше вышестоящих структур ЦНС участвует в организации движения, тем совершеннее реакция организма. Например, двигательные системы спинного мозга обеспечивают в основном сгибательные и разгибательные рефлексы конечностей. У мезэнцефального организма возможно сохранение естественной позы и восстановление нарушенной позы, но ходьба невозможна. При наличии подкорковых центров и коры большого мозга возможны любые сложные двигательные акты, в том числе ходьба, бег, прыжки и т.д. В афферентных системах иерархия заключается в передаче информации от первичных сенсорных нейронов (чувствительных нейронов) к сенсорным нейронам второго порядка, от них — к нейронам третьего порядка, от последних — к сенсорным зонам коры большого мозга, где осуществляется более совершенная оценка поступающей информации в вышележащих отделах ЦНС. Так, в рецепторах происходит лишь трансформация энергии раздражения в рецепторный потенциал, обеспечивающий возникновение нервного импульса, поступающего в ЦНС. В таламической области начинают формироваться ощущения. С помощью первичной сенсорной зоны коры большого мозга формируется восприятие о действующем раздражителе. Однако наиболее полное комплексное восприятие об объекте или явлении формируется только с участием первичной, вторичной и третичной (ассоциативной) сенсорных зон коры большого мозга. Принцип структурности заключается в том, что каждая реакция организма осуществляется с помощью определенных структур, и чем больше структур ЦНС участвует в ее осуществлении, тем она совершеннее (см. выше иерархический принцип). 4.4. Свойства нервных центров Свойства нервных центров связаны с некоторыми особенностями распространения возбуждения в ЦНС, особы- 90 Физиология ми свойствами химических синапсов и свойствами мембран нервных клеток. Нервные центры обладают постоянной (фоновой) активностью (тонусом), что объясняется: • спонтанной активностью нейронов ЦНС; • гуморальными влияниями биологически активных веществ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и влияющих на возбудимость нейронов; • афферентной импульсацией от различных рефлексогенных зон; • суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах; • циркуляцией возбуждения в ЦНС. Значение фоновой активности нервных центров заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов. Этот уровень может увеличиваться или уменьшаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора. В последнее время все больше подчеркивается роль пейсмекерных нейронов в генезе ритмов мозга. Пейсмекерный нейрон порождает градуальные потенциалы, которые, достигая порога, запускают генерацию ПД. У такого нейрона ритмический ПД регистрируется даже после полной изоляции нейрона. Это свидетельствует о том, что у таких нейронов повышена проницаемость клеточной мембраны для ионов Na+, которые постоянно поступают в клетку и деполяризуют ее. Когда деполяризация достигает критического уровня, возникает ПД. Na/K-помпа поддерживает ионные градиенты, поэтому циклы повторяются. Нервным центрам свойственно явление суммации возбуждения, которое открыл И.М. Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождаются ответной реакцией — лягушка совершает прыжок — это последовательная суммация, и пространственная — когда к центру поступают импульсы с нескольких рефлексогенных зон одновременно. Глава 4. Физиология центральной нервной системы 91 Для нервного центра характерно последействие — это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Главной причиной последействия является циркуляция возбуждения по замкнутым нейронным цепям (см. рис. 4.8), которая может продолжаться минуты и даже часы. Таким образом, особенность распространения возбуждения (его циркуляция) обеспечивает другое явление в ЦНС — последействие. Последействие играет важнейшую роль в процессах обучения — это начальный этап процесса запоминания информации. Нервным центрам присуща трансформация ритма возбуждения — изменение числа импульсов, возникающих в нейронах центра, на выходе относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна в сторону как увеличения, так и уменьшения. Уменьшение числа импульсов в нервном центре объясняется снижением его возбудимости за счет процессов пре- и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных импульсов, когда уже все нейроны центра или нейронного модуля возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов и нервных импульсов на выходе («окклюзия» по Шеррингтону — закупорка, от лат. occlusum — закрывать). Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют дивергенция процесса возбуждения и последействие. Важнейшим свойством нервных центров является их пластичность — способность к перестройке функциональных свойств. Она включает феномены: • синаптическая потенциация — улучшение проведения возбуждения в синапсах после кратковременной их активации, которая ведет к увеличению амплитуды постсинаптических потенциалов. Она может иметь пре- и постсинаптическую локализацию, может быть кратковременной (от нескольких секунд до 5 минут) и долговременной (несколько часов); • доминанта (от лат. dominans — господствующий) — это стойкий господствующий очаг возбуждения 92 Физиология в ЦНС, подчиняющий себе функции других нервных центров (Ухтомский А.А., 1923). Доминантное состояние нервных центров может быть вызвано метаболическими сдвигами показателей внутренней среды, например жаждой, возникающей при повышении осмолярности крови. Главные свойства доминантного очага возбуждения следующие: инерционность, повышенная возбудимость, способность реагировать на дивергирующие по ЦНС возбуждения, стойкость возникшего возбуждения, способность оказывать угнетающее влияние на центры-конкуренты и другие нервные центры. Биологическая роль доминантного очага возбуждения в ЦНС заключается в том, что на его основе формируется конкретная приспособительная деятельность, ориентированная на достижение полезного результата. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение, на базе доминантного состояния центра жажды побуждается поведение, направленное на поиск воды. Доминантные взаимодействия двух возбужденных центров лежат в основе замыкания временной связи при выработке условных рефлексов. Доминантное состояние двигательных центров обеспечивает выполнение автоматизированных реакций (например, работа на конвейере, выполнение комплекса гимнастических упражнений и т.п.). Синаптическая депрессия (утомляемость нервных центров) и восстановление работоспособности после отдыха также являются одним из вариантов пластичности ЦНС. Утомление объясняется расходованием медиатора, снижением чувствительности постсинаптической мембраны к медиатору вследствие накопления метаболитов, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям. Особо важный вариант пластичности нервных центров — компенсация нарушенных функций после повреждения того или иного центра. Хорошо известны клинические наблюдения за больными, у которых после кровоизлияний в вещество мозга повреждались центры регуляции мышечного тонуса и акта ходьбы. Тем не менее со временем отмечалось, что парализованная конечность у больных посте- Глава 4. Физиология центральной нервной системы 93 пенно начинает вовлекаться в двигательную активность, при этом нормализуется тонус ее мышц. Нарушенная двигательная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и увеличения ветвления их аксона (спраутинг), а также вовлечения в эту функцию других «рассеянных» нейронов в коре большого мозга с подобными функциями. В случае дополнительного ветвления аксона нейрона возрастает возможность улучшения иннервации мышечных волокон. Этому способствуют регулярные пассивные и активные движения органа, функция которого нарушена. ЦНС весьма чувствительна к изменениям внутренней среды, например к изменению газового состава крови, содержания глюкозы в крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы могут возникнуть судороги. Тяжелые последствия для организма вызывает недостаток кислорода в крови. Прекращение кровотока всего лишь на 10 с приводит к очевидным нарушениям функций мозга, человек теряет сознание. Прекращение кровотока на 8–12 мин вызывает необратимые нарушения деятельности мозга: погибают многие нейроны, в первую очередь корковые. Это ведет к тяжелым последствиям, что необходимо учитывать во время хирургических вмешательств на сердце, при реанимации пострадавшего. Наряду с возбуждением в ЦНС постоянно возникают процессы торможения. 4.5. Торможение ЦНС и координационная деятельность ЦНС Торможение ЦНС. Торможение — активный нервный процесс, результатом которого являются прекращение, ослабление или предотвращение возникновения возбуждения. Торможение всегда возникает как следствие возбуждения. При этом тормозные нейроны активируются медиаторами возбуждающих нейронов. Торможение в ЦНС 94 Физиология открыл И.М. Сеченов (1863). Известны две разновидности торможения: пре- и постсинаптическое торможение. Постсинаптическое торможение (Экклс Д., 1952). Под действием тормозных медиаторов (ГАМК и глицина) на постсинаптических мембранах возникает тормозной постсинаптический потенциал — ТПСП (рис. 4.9). Торможение — это следствие алгебраической суммации ТПСП и ВПСП, в связи с чем деполяризация постсинаптического нейрона не достигает КУД. В спинном мозге глицин высвобождается особыми тормозными клетками (клетки Реншоу) в синапсах, образуемых этими клетками на мембране мотонейронов. Действуя на ионотропный рецептор постсинаптической мембраны, глицин yвeличивaeт ee проницаемоcть для Сl–, при этом Сl– поступает в клетку согласно концентрационному градиенту, но вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В бесхлорной среде тормозная роль глицина не реализуется. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа Сl– в клетку или выхода К+ из клетки, что зависит от вида ГАМК-рецепторов. Основные разновидности постсинаптического торможения представлены на рис. 4.10. Пресинаптическое торможение. В основе механизма пресинаптического торможения лежит деполяризация пресинаптических окончаний (тормозная пресинаптическая деполяризация — ТПД), в результате чего возбуждение не проходит в них, постсинаптический нейрон не возбуждается. ТПД вызывают специальные тормозные –64 ВПСП 7 мс ПП = –70 –74 ТПСП Рис. 4.9. Возбуждающий и тормозной постсинаптические потенциалы (ВПСП и ТПСП) Глава 4. Физиология центральной нервной системы А 95 Б В Г Рис. 4.10. Разновидности постсинаптического торможения: А — параллельное; Б — возвратное; В — латеральное; Г — прямое. Нейроны: светлые — возбуждающие; темные — тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона-мишени. Снижение ПД в пресинаптической терминали (частичная деполяризация) всего лишь на 5% уменьшает ВПСП на постсинаптической мембране на 50%. ТПД после одного афферентного залпа продолжается 300–400 мс, медиатор — гамма-аминомасляная кислота (ГАМК), которая действует на ГАМК-рецепторы и повышает их проницаемость для Сl–, который выходит из клетки, что сопровождается развитием ТПД. Этот факт свидетельствует о том, что в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий первичный транспорт Сl– внутрь клетки вопреки электрическому градиенту. Разновидности пресинаптического торможения представлены на рис. 4.11. Здесь проходят только первые импульсы А ТПД Б ТПД Рис. 4.11. Разновидности пресинаптического торможения: А — параллельное; Б — латеральное. Нейроны: светлые — возбуждающие; темные — тормозные; ТПД — тормозная пресинаптическая деполяризация 96 Физиология Значение торможения. Выполняет охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС. Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пресинаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть заблокированы отдельные синаптические входы. К одному возбуждающему нейрону могут подходить сотни и тысячи импульсов по разным терминалям. Вместе с тем число дошедших до нейрона импульсов определяется пресинаптическим торможением. Торможение латеральных путей обеспечивает выделение существенных сигналов из фона. Торможение является важным фактором обеспечения координационной деятельности ЦНС. Об этом свидетельствует тот факт, что блокада торможения ведет к широкой дивергенции возбуждения и судорогам (например, при выключении постсинаптического торможения столбнячным токсином). Координационная деятельность ЦНС — это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Специальных центров координационной деятельности ЦНС не существует. Она осуществляется благодаря взаимодействию процессов возбуждения и торможения. Если выключить один из этих процессов, деятельность организма нарушается. Если выключить процесс торможения в ЦНС, например, введением в опыте на лягушке стрихнина (блокатора постсинаптического торможения), развиваются судороги. При блокаде же процессов возбуждения в ЦНС с помощью эфира лягушка становится обездвиженной, ее мышцы теряют тонус, активность лягушки полностью нарушается. Имеется ряд факторов, обеспечивающих взаимодействие центров между собой, центров и рабочих органов, обеспечивающих приспособительную деятельность организма. Фактор структурно-функциональной связи — это наличие связи между отделами ЦНС, между ЦНС и различными органами, обеспечивающей преимущественное распространение возбуждения между ними. Подобная связь включает несколько вариантов (рис. 4.12): Глава 4. Физиология центральной нервной системы 97 Реципрокная (сочетанная) Спинной мозг Возвратная Обратная Прямая Мышца Коленный сустав Мышечный рецептор Сухожильный рецептор Кость Раздражение Рис. 4.12. Структурно-функциональная связь между нервными центрами и рабочими органами, обеспечивающая координационную деятельность ЦНС. Уровень — спинной мозг (нижняя конечность, коленный сустав). Центры: С — сгибания; Р — разгибания. Нейроны: светлые — возбуждающие; темные — тормозные • прямая связь — это управление другим центром (ядром) или рабочим органом с помощью посылки к ним эфферентных импульсов (команд). Например, нейроны дыхательного центра продолговатого мозга посылают импульсы к α-мотонейронам спинного мозга, от которых нервные импульсы поступают к дыхательным мышцам; • обратная связь (обратная афферентация) — управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них, — принцип обратной связи в регуляции функций организма. Ее нарушение ухудшает функцию органа; • возвратная связь — вид функциональной связи, обеспечивающий торможение нейронов вслед за их возбуждением. Яркий пример этого вида связи — торможение мотонейронов (иннервирующих мышцы конечностей) с помощью возвратного постсинаптического торможения. При этом вслед за возбуждением мотонейронов, обеспечивающих сокращение мышцы, развивается торможение нейронов этого же центра с помощью тормозных клеток Реншоу. Это обеспечивает расслабление мышцы и возможность очередного ее сокращения, на- 98 Физиология пример, при шагательном рефлексе и других повторяющихся сокращениях и расслаблениях мышц; • реципрокная (взаимная) связь — вид связи, обеспечивающий торможение центра-антагониста при возбуждении центра-агониста. Например, при вызове сгибательного рефлекса конечности импульсы от рефлексогенной зоны (кожа) поступают через вставочные нейроны к мотонейронам центра мышц-сгибателей, а также одновременно к центру-антагонисту (мышцразгибателей), но с включением на пути тормозного нейрона, который образует тормозной синапс на нейронах центра-разгибателя. Мышцы-разгибатели поэтому не сокращаются и не препятствуют сгибанию конечности. У другой конечности, напротив, центр сгибания тормозится, а центр разгибания возбуждается, что обеспечивает сохранение позы. Реципрокные взаимоотношения между центрами встречаются довольно широко. Так, при возбуждении центра глотания тормозится центр жевания, возбуждение центра вдоха тормозит центр выдоха, рефлекс глотания тормозит вдох. Принцип иерархии в деятельности ЦНС, в данном случае — субординация — подчинение нижележащих отделов ЦНС вышележащим. Например, ядро Дейтерса (продолговатый мозг), нейроны красного ядра (средний мозг) управляют активностью α- и γ-мотонейронов спинного мозга. В процессе эволюции наблюдается тенденция к увеличению роли вышележащих отделов головного мозга в обеспечении координированной деятельности нижележащих центров (цефализация), причем с преобладанием тормозных влияний. Фактор силы процесса возбуждения также проявляется в согласовании деятельности нервных центров. При поступлении импульсов к одному и тому же центру от различных рефлексогенных зон (принцип общего конечного пути) центр реагирует на более сильное возбуждение. В борьбе за общий конечный путь побеждает более сильное возбуждение — выполняется более важная команда в биологическом отношении, что является важным приспособительным результатом. Одностороннее проведение возбуждения в химических синапсах ЦНС обеспечивает упорядоченное распростра- Глава 4. Физиология центральной нервной системы 99 нение возбуждения. В случае двустороннего проведения в синапсах наблюдалась бы дивергенция возбуждения в различных направлениях. Синаптическая потенциация участвует в процессах обеспечения координационной деятельности ЦНС при выработке навыков, поскольку возбуждение распространяется в ЦНС быстрее и точнее по проторенным путям, возбудимость которых повышена. Недостаточно координированные движения в начале выработки навыка постепенно становятся более точными — координированными. Дополнительные, ненужные движения постепенно устраняются. Доминанта также играет важную роль в координационной деятельности ЦНС, т.к. доминантное состояние двигательных центров обеспечивает автоматизированное выполнение многократно повторявшихся ранее двигательных актов, например в процессе трудовой деятельности человека, при выполнении гимнастических элементов. При этом движения точные, а также экономные в энергетическом и эмоциональном отношениях. Изучив общие закономерности деятельности ЦНС, рассмотрим функции различных отделов ЦНС. 4.6. Интегративная роль нервной системы Интегративная роль нервной системы (от лат. integrum — цельный, объединение каких-либо частей) заключается в объединении органов и систем в единую функциональную систему — организм. Такое объединение становится возможным благодаря участию ЦНС в управлении опорно-двигательным аппаратом с помощью соматической нервной системы, благодаря регуляции функций всех тканей и внутренних органов с помощью вегетативной нервной системы и эндокринной системы, благодаря наличию обширнейших афферентных связей ЦНС со всеми соматическими и вегетативными эффекторами. Обоснованно выделить четыре основных уровня ЦНС, каждый из которых вносит свой вклад в обеспечение интегративной деятельности ЦНС. Первый уровень — нейрон. Взаимодействие возбуждающих и тормозящих входов, взаимодействие субсинаптических нейрохимических процессов в протоплазме 100 Физиология в конечном итоге определяют, возникнет ли та или иная последовательность ПД на выходе нейрона или нет, т.е. будет ли дана команда другому нейрону, рабочему органу или нет и возникнет ли соответствующая реакция. Благодаря множеству возбуждающих и тормозящих синапсов на нейроне он превратился в ходе эволюции в решающее устройство. Второй уровень — нейронный модуль (нейронный ансамбль, нейронный пул), обладающий качественно новыми свойствами, отсутствующими у отдельных нейронов. Один и тот же модуль может участвовать в деятельности различных центров и отделов ЦНС и обеспечивать возникновение более сложных реакций ЦНС. Третий уровень — нервный центр. Они часто выступают как автономные командные устройства, реализующие управление тем или иным процессом на периферии в составе саморегулирующейся, самовосстанавливающейся, самовоспроизводящейся системы — организма. Это достигается за счет множественных прямых, обратных, реципрокных связей в ЦНС, наличия прямых и обратных связей с периферическими органами. Четвертый уровень интеграции — вся ЦНС, объединяемая корой большого мозга в единую регулирующую систему, а отдельные органы и системы — в единую физиологическую систему — организм, подсистемы которого постоянно формируют функциональные системы. Это достигается взаимодействием главных систем ЦНС: лимбической, ретикулярной формации, подкорковых образований и неокортекса как высшего отдела ЦНС, организующего поведенческие реакции и их вегетативное обеспечение в различных условиях жизнедеятельности организма. 4.7. Двигательные системы спинного мозга Структурно-функциональная характеристика спинного мозга включает структуры, обеспечивающие формирование фазных и тонических (позных) рефлексов спинального организма. Спинной мозг имеет сегментарное строение (31–33 сегмента) — каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный Глава 4. Физиология центральной нервной системы 101 (СI–СVIII), грудной (ТhI–ТhXII), поясничный (LI–LV), крестцовый (SI–SV) и копчиковый (СоI–СоII). В спинном мозге насчитывают около 13 млн нейронов (3% из них — мотонейроны, 97% — вставочные нейроны, относящиеся в том числе и к вегетативной нервной системе). При этом выделяют: • нейроны соматической и вегетативной нервной системы; • эфферентные, афферентные, вставочные, ассоциативные; • возбуждающие и тормозные. Центры спинного мозга участвуют в регуляции большинства внутренних органов и скелетных мышц, в частности центр зрачкового рефлекса локализуется в сегментах СVIII–ТhII, регуляции деятельности сердца — в ТhI–ТhV, слюноотделения — в ТhII–ТhV, регуляции функции желудка — в ТhVI–ТhIX, почек — в ТhV–LIII. Сегментарно расположены центры гладких мышц внутренних органов, центры пиломоторных рефлексов, центры, регулирующие функции потовых желез и сосудов. Парасимпатическую иннервацию получают из спинного мозга (SII–SIV) все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. У женщин парасимпатическая иннервация обеспечивает сосудистые реакции клитора, влагалища; у мужчин — рефлекторный компонент эрекции. Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу мышцы шеи (СI–СIV), диафрагмы (СIII–СV), верхних конечностей (СV–ТhII), туловища (ТhIII–LI) и нижних конечностей (LII–SV). Повреждения определенных сегментов спинного мозга или его проводящих путей вызывают специфические расстройства чувствительности и двигательные нарушения. Каждый сегмент спинного мозга участвует в чувствительной иннервации трех дерматомов. Имеется дублирование двигательной иннервации скелетных мышц, что повышает надежность функционирования опорно-двигательного аппарата и восприятия внешних раздражителей. 102 Физиология Ретикулярная формация спинного мозга обнаруживается на уровне шейных сегментов. Она участвует в регуляции тонуса мышц, вегетативных функций, афферентной импульсации. Спинной мозг имеет собственный ассоциативный аппарат, устанавливающий связь между сегментами и внутри сегментов. Ассоциативный аппарат спинного мозга участвует в регуляции движений конечностей и туловища, тонуса мышц, поддержании позы. Спинной мозг выполняет проводниковую и рефлекторную функции. Проводниковая функция спинного мозга и движения организма. С помощью проводниковой функции спинного мозга вышележащие отделы ЦНС получают информацию от организма и внутренних органов, управляют скелетной мускулатурой тела и конечностей, регулируют функции внутренних органов. Проводниковая функция спинного мозга (табл. 4.1) осуществляется с помощью восходящих и нисходящих путей. Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляются через передние корешки (закон Белла–Мажанди). Таблица 4.1 Основные проводящие пути спинного мозга, обеспечивающие связь с вышележащими отделами ЦНС, и их роль Восходящие Физиологическое значение (чувствительные) пути Клиновидный пучок Проприорецептивная импульсация (пучок Бурдаха), проходит от верхней части туловища и рук в задних столбах, импульсация поступает в кору Тонкий пучок (пучок Голля), Проприорецептивная импульсация проходит в задних столбах, от нижней части туловища и ног импульсация поступает в кору Передний спинно-мозжеч- Проприорецептивная импульсация ковый (Говерса) поступает в мозжечок Дорсальный спинно-мозТо же жечковый (Флексига) Глава 4. Физиология центральной нервной системы Восходящие (чувствительные) пути Латеральный спинно-таламический Передний спинно-таламический Нисходящие (двигательные) пути Латеральный кортико-спинальный (пирамидный) Передний кортико-спинальный (пирамидный) Рубро-спинальный (Монакова), проходит в боковых столбах Ретикуло-спинальный, передние столбы 103 Физиологическое значение Болевая и температурная чувствительность Тактильная чувствительность — прикосновение, давление Физиологическое значение Импульсы к скелетным мышцам, произвольные движения То же Участие в регуляции тонуса скелетных мышц Регуляция тонуса скелетных мышц с помощью возбуждающих и тормозящих влияний на α- и γ-мотонейроны, а также регулирующие состояние спинальных вегетативных центров Вестибуло-спинальный, Регуляция тонуса мышц, поддержание передние столбы позы и восстановление нарушенной позы Текто-спинальный, проходит Импульсы, обеспечивающие осув передних столбах ществление зрительных и слуховых двигательных рефлексов (рефлексов четверохолмия) Значение афферентной импульсации, поступающей в спинной мозг: • несет информацию об изменениях окружающей среды; • участвует в координационной деятельности ЦНС по управлению скелетной мускулатурой — при выключении афферентной импульсации от рабочего органа управление им становится несовершенным; • способствует поддержанию тонуса ЦНС — при выключении афферентной импульсации уменьшается суммарная тоническая активность ЦНС; • участвует в процессах регуляции функций внутренних органов. Двигательные системы спинного мозга обеспечивают формирование тонических, фазных рефлексов и ритмических рефлексов (классификация рефлексов по характеру ответной реакции) — это сегментарные рефлексы. Надсег- 104 Физиология ментарные рефлексы осуществляются только с помощью шейного отдела. По рецепторам, раздражение которых вызывает рефлексы, выделяют проприоцептивные, висцероцептивные и кожные рефлексы (защитные). Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцероцептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины. Эти рефлексы изучают в эксперименте у спинального организма с перерезанным спинным мозгом ниже сегментов С3–С5, где локализуются центры диафрагмальных нервов. В этом случае сохраняется диафрагмальное дыхание (центры диафрагмальных нервов связаны с головным мозгом). Фазные рефлексы конечностей спинального организма. Рефлексы конечностей (рис. 4.13) в клинической практике исследуют наиболее часто. Фазные рефлексы — это однократное сгибание или разгибание конечности при однократном раздражении кожных рецепторов или проприорецепторов. Сгибательные рефлексы конечностей. Рефлексы, возникающие с рецепторов кожи, полисинаптические, они 1 Рис. 4.13. Рефлексы нижних конечностей: А — дуга сгибательного (защитного) рефлекса; Б — дуга перекрестного разгибательного рефлекса; В — коленные суставы с мышцами; Г — сегмент спинного мозга; → — раздражение кожи; ↑ — афферентный путь; ↓ — эфферентные пути от α-мотонейронов центров сгибания (С) и разгибания (Р). Нейроны: светлые — возбуждающие; темные — тормозные Глава 4. Физиология центральной нервной системы 105 имеют защитное значение. Сгибательные (как и разгибательные) рефлексы, возникающие с проприорецепторов, — моносинаптические. Фазные проприоцептивные рефлексы участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. Поэтому в клинической практике исследуют несколько сгибательных фазных рефлексов — ахиллов (пяточный) рефлекс, подошвенный рефлекс — кожный рефлекс, локтевой рефлекс — проприоцептивный. При возникновении фазных сгибательных рефлексов одновременно с возбуждением мотонейронов мышцсгибателей происходит торможение мотонейронов мышцразгибателей этой же конечности — иначе сгибание будет невозможным. Разгибательные рефлексы конечностей — моносинаптические, возникают только с проприорецепторов мышцразгибателей. Фазные разгибательные рефлексы, в отличие от фазных сгибательных рефлексов, возникают в ответ на однократное раздражение только мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенника. При этом наблюдается коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся с помощью постсинаптического реципрокного торможения посредством вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного разгибательного рефлекса замыкается в поясничных сегментах (LII–LIV). Фазные разгибательные рефлексы, как и сгибательные, участвуют в формировании акта ходьбы (перемещение тела в пространстве). Ритмические рефлексы. Характерным примером ритмических рефлексов является шагательный рефлекс. Шагательные рефлексы вызываются у спинальной собаки (перерезан спинной мозг ниже сегментов С3–С5 — центр диафрагмального нерва, что обеспечивает диафрагмальное дыхание и жизнь организму). Они вызываются однократным раздражением кожи задней конечности, 106 Физиология что вызывает ее сгибание и разгибание противоположной конечности — перекрестный и разгибательный рефлекс. Затем согнутая конечность разгибается и опускается, а разогнутая — сгибается и поднимается и т.д. Конечности не касаются опоры (собака зафиксирована в станке с помощью лямок), и без дополнительного раздражения конечности продолжают ритмично поочередно сгибаться и разгибаться — «шагать» в воздухе. Шагательный рефлекс (локомоция) может продолжаться часами, т.к. устранено тормозное влияние коры большого мозга. Импульсы к центрам сгибания и разгибания конечностей поступают от их проприорецепторов. Роль рецепторов Мышечные веретена (мышечные рецепторы, они первичные) расположены параллельно скелетной мышце — своими концами они крепятся к соединительнотканной оболочке (перимизию) пучка экстрафузальных мышечных волокон с помощью напоминающих сухожилия полосок соединительной ткани. В связи с этим, когда мышца расслабляется (удлиняется), растягиваются (раздражаются) и мышечные рецепторы, что и ведет к их возбуждению. Сухожильные рецепторы (рецепторы Гольджи) локализуются в сухожилиях скелетных мышц, т.е. крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при сокращении мышцы и натяжении сухожилия. Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы — что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее). Механизм шагательного рефлекса. Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные рецепторы. Импульсы от них поступают к своим α-мотонейронам спинного мозга и возбуждают их (рис. 4.14). Поэтому α-мотонейроны, в свою очередь, посылают импульсы к этой же скелетной мышце, что ведет к ее сокраще- Глава 4. Физиология центральной нервной системы 107 нию. В результате сокращения мышцы возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы, поскольку сухожилие натягивается (рис. 4.14, Б). Импульсы от сухожильных рецепторов поступают тоже в свой центр в спинном мозге, но к тормозным клеткам. Возбуждение этих клеток вызывает торможение α-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Ее расслабление (удлинение) ведет снова к возбуждению мышечных рецепторов и α-мотонейронов — мышца вновь сокращается. Вследствие сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что ведет к очередному расслаблению скелетной мышцы и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разги- А Б Рис. 4.14. Механизм возбуждения и торможения α-мотонейронов спинного мозга во время сокращения и расслабления скелетной мышцы: А — мышца растянута и расслаблена, возбуждаются мышечные рецепторы; Б — мышца сокращена (напряжена) и укорочена, возбуждаются сухожильные рецепторы; 1 — мышечные рецепторы (мышечные веретена); 2 — сухожилия мышцы и их рецепторы (рецепторы Гольджи); 3 — полусегмент спинного мозга 108 Физиология бателю, при этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу, что наблюдается при ходьбе и беге или любом повторяющемся действии. При ходьбе и беге, если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, иначе нарушится поза. Это обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров-сгибателей и центров-разгибателей другой конечности — перекрестный разгибательный рефлекс. Шагательные координированные движения возможны при отсутствии обратной афферентации от проприорецепторов. Их программа формируется в онтогенезе. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга и генератора локомоции каждой конечности. О наличии межсегментарных связей на уровне спинного мозга свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей при достаточно длительном и сильном раздражении одной конечности. Произвольные движения конечностей после разрыва спинного мозга невозможны — нет связи с корой большого мозга. Спотыкательный рефлекс. У спинального организма во время шагательного рефлекса можно наблюдать спотыкательный рефлекс — при движении ноги вперед и столкновении ее с препятствием она поднимается и переступает через препятствие. Тонические рефлексы спинального организма (рефлексы позы). Сегментарные рефлексы позы — это тонические сгибательные и разгибательные рефлексы конечностей, обеспечивающие сохранение естественной позы. Они возникают при длительном растяжении мышц и возбуждении мышечных рецепторов. Тоническое сокращение скелетных мышц является фоновым для всех двигательных актов, осуществляемых с помощью фазных сокращений мышц. Изменение тонуса мышц конечностей свидетельствует о нарушениях в ЦНС. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц Глава 4. Физиология центральной нервной системы 109 спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибания и разгибания) называют также миотатическими. Таким образом, главная роль тонических рефлексов — поддержание естественной позы организма. Надсегментарные рефлексы позы спинального организма — это шейные рефлексы. Они заключаются в перераспределении тонуса мышц конечностей при изменении положения шеи относительно туловища. Эти рефлексы осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р. Магнус (1924) в опытах на кошке. Имеется две группы этих рефлексов. Первая группа шейных позных рефлексов возникает при наклоне головы вперед или назад. При наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние — разгибаются (рис. 4.15). При наклоне головы вверх (кзади) возникают противоположные реакции — передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения А Б Рис. 4.15. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом: А — при пассивном опускании; Б — при пассивном поднятии головы 110 Физиология тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях естественного поведения животных они увеличивают шансы достать пищу, находящуюся выше или ниже головы, — это автоматизированная приспособительная реакция, рефлексы являются надсегментарными. Шейные рефлексы позы верхних конечностей у человека, по-видимому, утрачены, а рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы. Вторая группа шейных позных рефлексов (тонических) возникает с шейных проприорецепторов при поворотах или наклонах головы вправо или влево. Рефлекс выражается в повышении тонуса мышц-разгибателей обеих конечностей на стороне, в которую повернута (наклонена) голова, и в повышении тонуса мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота (наклона) головы. Центр тяжести смещается в сторону поворота головы — именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей, что повышает устойчивость организма. Подобная ситуация наблюдается и у человека. Спинальный шок. В случае повреждения спинного мозга у человека вследствие травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния вышележащих отделов ЦНС на спинной мозг. Об этом свидетельствует тот факт, что повторная перерезка спинного мозга в эксперименте на животных ниже первой шока не вызывает. Спинальный шок у лягушек длится несколько минут, у собак — несколько дней, у человека — около 2 мес. Тонус мышц спинального организма. После исчезновения спинального шока тонус мышц (иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки) резко повышается. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются (рис. 4.16). Глава 4. Физиология центральной нервной системы 111 Гипертонус имеет рефлекторную природу: деафферентация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста). Произвольные движения невозможны. Афферентная импульсация от мышечных рецепторов по задним корешкам спинного мозга (рис. 4.16), которая ведет к возбуждению α-мотонейронов спинного мозга, сокращению скелетных мышц и развитию гипертонуса, поддерживается двумя способами: • спонтанной активностью мышечных рецепторов, что обеспечивает поступление импульсов от них к α-мотонейронам, возбуждение последних и сокращение мышц; • спонтанной активностью γ-мотонейронов. Возбуждение γ-мотонейронов вызывает возбуждение и сокращение иннервируемых ими интрафузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы 1 2 3 Рис. 4.16. Механизм регуляции тонуса мышц на уровне спинного мозга и развития гипертонуса мышц нижних конечностей в случае повреждения спинного мозга: 1 — полусегмент спинного мозга; 2 — скелетная мышца; 3 — мышечное веретено; α — мотонейрон, иннервирующий миоциты скелетной мышцы; γ — мотонейрон, иннервирующий миоциты мышечного рецептора 112 Физиология на скелетной мышце. Это вызывает раздражение и возбуждение мышечных рецепторов, импульсы от которых поступают к α-мотонейронам и возбуждают их. В свою очередь, α-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение, вследствие чего и развивается гипертонус у спинального организма. Если спинальное животное поставить на ноги, то оно будет стоять; если же положить на бок, оно будет лежать, не делая попытки встать на ноги. Функции центров спинного мозга управляются центрами головного мозга, и в частности ствола мозга. Животное с сохраненным средним мозгом называют мезэнцефальным организмом. 4.8. Двигательные системы ствола мозга Характеристика мезэнцефального организма. Ствол мозга включает продолговатый мозг, мост и средний мозг (рис. 4.17), где локализуются ядра черепных нервов продолговатого мозга (IX–XII пары), моста (V–VIII пары) и среднего мозга (III, IV пары), а также другие ядра, управляющие мускулатурой туловища и конечностей. Перечислим черепные нервы: добавочный нерв (XI, n. accessories), языкоглоточный (IX, n. glossopharyngeus), подъязычный (XII, n. hypoglossus), преддверно-улитковый (VIII, Большие полушария Промежуточный мозг (таламус и гипоталамус) Средний мозг Мост Продолговатый мозг Мозжечок Рис. 4.17. Основные отделы головного мозга Глава 4. Физиология центральной нервной системы 113 n. vestibulocochlearis), блуждающий (X, n. vagus), лицевой (VII, n. facialis), отводящий (VI, n. abducens), тройничный (V, n. trigeminus), глазодвигательный (III, n. oculomotorius), блоковой (IV, n. trochlearis). У мезэнцефального организма тонус мышц нормальный, он может сохранять позу и восстанавливать ее при нарушении, но произвольные движения отсутствуют. Центры ствола мозга обеспечивают регуляцию тонуса мышц и функции внутренних органов. Все центры ствола мозга целесообразно объединить в три группы: • жизненно важные вегетативные центры регуляции систем: дыхания, сердечно-сосудистой, пищеварения. Поражение продолговатого мозга ведет к остановке дыхания и смерти; • центры защитных рефлексов, в осуществлении которых участвуют по несколько черепных нервов, — это рефлексы чихания, кашля, мигания, слезоотделения, рвоты; • ядра ствола мозга, управляющие мускулатурой конечностей и туловища, которые включают несколько ядер: – ядра шва (продолговатый мозг). Аксоны его нейронов поступают в спинной мозг, многие из них содержат серотонин. На интернейронах заднего рога этот путь образует тормозные синапсы (угнетение передачи сигналов), а на мотонейронах — возбуждающие. В целом моноаминонергические пути модулируют реактивность нейронных цепей спинного мозга, а не конкретные движения; – вестибулярные ядра (мост) оказывают возбуждающее влияние на α- и γ-мотонейроны спинного мозга мышц-разгибателей конечностей, туловища, шеи и тормозное влияние на мотонейроны сгибателей этих частей тела. Подобный эффект вызывают и ретикулярные ядра моста (медиальный ретикулоспинальный путь); – красные ядра (средний мозг, руброспинальный путь), напротив, оказывают тормозное влияние на мышцы-разгибатели и возбуждающее — на мышцы-сгибатели. Такое же влияние оказывает и рети- 114 Физиология кулярное гигантоклеточное ядро продолговатого мозга (латеральный ретикулоспинальный путь). В случае нарушения соотношения возбуждающих и тормозных влияний на мотонейроны спинного мозга (нарушение связи продолговатого и среднего мозга) развивается децеребрационная ригидность (рис. 4.18). У мезэнцефального организма при внезапных раздражениях возникает сторожевая реакция (ориентировочный рефлекс). При действии света наблюдаются поворот головы и глаз в сторону света, фиксация взора, слежение за движущимся объектом (осуществляются с помощью верхних холмиков четверохолмия — первичного зрительного центра). При действии звукового раздражителя наблюдаются прислушивание, поворот ушных раковин у животных (реализуются с помощью нижних холмиков четверохолмия — первичного слухового центра). В обоих случаях наблюдается готовность к действию — повышается тонус мышц-сгибателей конечностей. Ориентировочный рефлекс — это запрограммированная реакция, как и рефлексы шагания, сосания, глотания. Двигательные системы ствола мозга обеспечивают с помощью тонических рефлексов сохранение естественной позы и восстановление позы при ее нарушении. Рефлексы возникают с вестибулярных, проприоцептивных, тактильных и зрительных рецепторов и осуществляются двигательными центрами ствола мозга, включая ядра ретикулярной формации. Афферентные (восходящие) пути см. в табл. 4.1. Эфферентные влияния из двигательных центров ствола мозга распространяются на мускулатуру конечностей и туловища по четырем нисходящим путям (табл. 4.1). Механизм возникновения децеребрационной ригидности и Рис. 4.18. Децеребрационная ригидность у кошки при перерезке ствола мозга между мостом и средним мозгом Глава 4. Физиология центральной нервной системы 115 сохранения нормального тонуса мышц на уровне среднего мозга (мезэнцефальный организм) показан на рис. 4.19. Классификация тонических (позных) рефлексов организма по Р. Магнусу с изменениями. Все тонические рефлексы (рефлексы позы) осуществляются с помощью одних и тех же двигательных ядер — красных, вестибулярных и ретикулярных ядер ствола мозга, ядер спинного мозга — и отличаются друг от друга только рефлексогенными зонами. Позные рефлексы включают три группы: статические, статокинетические и кинетические. Они выражаются в перераспределении мышечного тонуса. Статические рефлексы (от греч. statos — стоящий) — это тонические рефлексы, обеспечивающие сохранение естественной позы организма в покое. Человек стоит или сидит, в том числе и при поворотах или наклонах головы и движениях рук, а сидя — и ног, вызывающих смещение центра тяжести. К ним относятся сегментарные и надсегментарные (шейные) тонические рефлексы спинного мозга и рефлексы ствола мозга, возникающие с проприо- и вестибулорецепторов. При вертикальной позе человека (голова вверх, шея наклонена несколько вперед) шейные и вестибулярные рефлексы усиливают друг друга, в результате Средний мозг КЯ Мост РФ Рис. 4.19. Механизмы регуляции тонуса мышц и возникновения децеребрационной ригидности при перерезке ствола мозга между средним мозгом и мостом (пунктирная линия): РФ — ретикулярная формация; Д — ядро Дейтерса; α, γ — мотонейроны; вставочные нейроны спинного мозга: светлый — возбуждающий; темный — тормозной; КЯ — красное ядро Д РФ Продолговатый мозг α γ Спинной мозг Мышечный рецептор Мышца- 116 Физиология чего происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей. Статические тонические рефлексы открыл Бронжест в опыте на спинальной лягушке. Он перерезал задние корешки спинного мозга и подвесил лягушку на крючок. Нижняя конечность на стороне перерезки корешков была выпрямлена, а другая конечность слегка согнута, что свидетельствует о наличии рефлекторного тонуса мышц (см. раздел 4.10). Статокинетические рефлексы — это тонические рефлексы, обеспечивающие восстановление нарушенной позы. Они возникают с кожных, проприо- (шея) и вестибулорецепторов, при нарушении позы (организм находится в горизонтальном положении). К ним относятся выпрямительные (установочные) рефлексы — выпрямление головы и туловища. Выпрямление головы запускается с вестибулярных и кожных рецепторов, выпрямление туловища — с проприорецепторов шеи (они раздражаются при движении головы) и кожных рецепторов — по две рефлексогенных зоны. Именно поэтому при выключении только рецепторов кожи или вестибулярного аппарата у мезэнцефального организма рефлекс выпрямления головы не нарушается. При выключении же обеих названных рефлексогенных зон этот рефлекс не возникает. Рефлекс выпрямления туловища также исключается, если выключить проприорецепторы шеи (накладывание гипса на шею) и рецепторы кожи. Если одна из названных зон сохраняется, выпрямление туловища не нарушается. Вестибулярный аппарат в эксперименте выключается путем его разрушения или анестезией. Это надсегментарные рефлексы ствола мозга и спинного мозга. Особо важную роль играют красные ядра — при их разрушении нарушенная поза животного не восстанавливается. Р. Магнус назвал эти рефлексы статическими. Однако они имеют две составляющих — статическую (нет перемещения тела в пространстве или вращения тела вокруг вертикальной оси на месте) и кинетическую (от греч. kinetikos — относящийся к движению) — животное поднимается на ноги и принимает естественную позу. У человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии коры большого мозга, при ее выключении выпрямительные рефлексы не возникают. Глава 4. Физиология центральной нервной системы 117 Кинетические рефлексы (от греч. kinema — движение) — это тонические рефлексы, обеспечивающие сохранение естественной позы в движении — при ходьбе и беге, вращательном движении на месте, при изменении скорости движения в транспорте и в лифте. Поэтому называть их статокинетическими нет оснований — никакой статики, только движение. Эти рефлексы возникают с проприо- и вестибулорецепторов. Сокращения мышц направлены на преодоление сил гравитации, изменяющихся по величине вследствие ускорения или замедления движения. Важную роль играет система зрения. В процессе вращательных движений наблюдаются также повороты головы и нистагм глаз (пилообразные движения глаз). Глазной нистагм способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При вращении тела сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению тела. Затем глаза быстро движутся обратно — по ходу вращения «догоняют организм». Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). Во врачебной практике направление нистагма обозначают по быстрому компоненту. Исследование нистагма (степени его выраженности, длительности) используется для оценки функционального состояния вестибулярной системы и дифференциальной диагностики поражения ЦНС. Лифтные рефлексы. При остановке движущегося вниз лифта и в начале его быстрого подъема в нижних конечностях повышается тонус мышц-разгибателей, что обеспечивает преодоление сил инерции хотя наблюдается некоторое сгибание конечностей под влиянием массы тела. В начале опускания лифта и при остановке движущегося вверх лифта наблюдаются противоположные явления. Существует и другая точка зрения, учитывающая только внешние признаки — пассивное сгибание или разгибание конечностей вследствие действия сил гравитации. Таким образом, ствол мозга обеспечивает сохранение естественной позы, восстановление позы при ее нарушении, нормальный тонус мышц. 118 Физиология Совокупность проприоцептивной и вестибулярной сенсорных систем и ядер ствола мозга и спинного мозга образует систему положения тела. 4.9. Интегративные системы ствола мозга Интегративные системы ствола мозга включают ретикулярную формацию (РФ), голубое пятно, ядра шва, центральное серое вещество и черную субстанцию. Эти структуры имеют обширные нисходящие и восходящие связи со всеми отделами ЦНС. Ретикулярная формация представляет собой скопления нейронов, различных по функции и размерам, связанных множеством нервных волокон, проходящих в разных направлениях и образующих сеть на всем протяжении ствола мозга, что и определяет ее название. Нейроны либо расположены диффузно, либо образуют ядра. Каждый нейрон РФ устанавливает связь с множеством других нейронов ЦНС (до 25 000). Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, которые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая — восходящее направление. Свойства нейронов РФ: • обладают спонтанной активностью, частота их разрядов в покое равна 5–10 имп./с; • полисенсорность — они принимают коллатерали от нескольких сенсорных путей, идущих от разных рецепторов; • им свойственна высокая чувствительность к некоторым веществам крови (например, адреналину, СО2) и лекарствам (барбитуратам, аминазину и др.); • они обладают высокой лабильностью — до 400 имп./с; • нейроны РФ более возбудимы по сравнению с другими нейронами. Нейроны и ядра РФ входят в состав центров, регулирующих функции внутренних органов (кровообращения, дыхания, пищеварения), регулируют тонус скелетной мускулатуры, активность коры большого мозга. Обширны связи ретикулярной формации с другими отделами ЦНС Глава 4. Физиология центральной нервной системы 119 и рефлексогенными зонами — РФ получает импульсацию от различных сенсорных рецепторов организма и отделов ЦНС и, в свою очередь, посылает импульсы во все отделы мозга. При этом выделяют восходящие и нисходящие влияния ретикулярной формации. Нисходящие влияния РФ на моторные спинальные центры показаны на рис. 4.12 (мышцы-разгибатели). На сгибатели влияния противоположные. С помощью нисходящих влияний на спинной мозг РФ принимает участие в регуляции тонуса мышц, в формировании всех двигательных актов. Восходящие влияния РФ на большой мозг могут быть как активирующими (они преобладают), так и тормозными. Импульсы от ретикулярных нейронов поступают к неспецифическим ядрам таламуса и после переключения в них проецируются в различные области коры, в задний гипоталамус, стриатум. Вегетативные функции РФ заключаются в поддержании тонуса вегетативных центров, интеграции симпатических и парасимпатических влияний для реализации потребностей организма, в передаче модулирующих влияний от гипоталамуса и мозжечка к органам. РФ является главным структурным элементом жизненно важных центров продолговатого мозга — сердечно-сосудистого, дыхательного, жевательного, глотательного. Черная субстанция участвует в регуляции мышечного тонуса, координации жевания и глотания, мелких движений конечностей (например, при печатании на машинке, письме), в осуществлении эмоций. В случае повреждения черной субстанции наблюдается бедность движений. Ее нейроны — дофаминергические, поэтому черную субстанцию выделяют как дофаминергическую систему. Голубое пятно (ГП) расположено в среднем мозге, его нейроны являются адренергическими, как и вентролатерального ядра ретикулярной формации продолговатого мозга. В совокупности они составляют адренергическую систему ствола мозга. Нейроны ГП имеют фоновую импульсную активность (10–20 Гц). На их мембранах имеются рецепторы к адреналину, норадреналину, серотонину, ГАМК, глицину, опиоидам, активация которых вызывает преимущественно торможение, а также рецепторы к аце- 120 Физиология тилхолину, пептиду Р, глутамату, активация которых вызывает преимущественно возбуждение. В свою очередь, нейроны ГП оказывают тормозное влияние на другие нейроны посредством β-адренорецепторов, реже — возбуждающее (например, в ядрах VII черепного нерва), посредством α-адренорецепторов. Импульсная активность нейронов голубого пятна повышена в фазе быстрого сна, что выключает мышечный тонус и фазические движения мускулатуры туловища и конечностей. Иннервируя ядра гипоталамуса, нейроны ГП участвуют в развитии общего адаптационного синдрома при действии на организм стрессоров. Норадренергическая система контролирует болевую и неболевую импульсацию сенсорных и двигательных систем, угнетая фоновую активность нейронов их ядер. Оно участвует в регуляции деятельности висцеральных систем, эмоциональных состояний (тоска, страх), механизмов памяти и внимания, индуцирует фазу быстрого сна. Серотонинергическая система ЦНС включает центральное серое вещество и ретикулярные магноцеллюлярные ядра, ядра шва, имеющие афферентные и эфферентные связи со всеми отделами ЦНС. Ее нейроны наиболее активны во время бодрствования, менее активны во время медленного сна, их возбуждение прекращается в фазе быстрого сна. Эта система тормозит нейроны сенсорных, особенно болевых, путей спинного мозга, ствола и таламуса. Серотонинергические нейроны тормозят агрессивное поведение, потребление воды и пищи. 4.10. Двигательные функции мозжечка Структурно-функциональная характеристика мозжечка. Главная функциия мозжечка — координация автоматизированных движений. Мозжечок содержит больше половины всех нейронов ЦНС, но его масса составляет всего лишь 10% массы головного мозга. Выделяют три основных структуры мозжечка: древний мозжечок, старый мозжечок и новый мозжечок. Афферентные входы и эфферентные выходы мозжечка осуществляются посредством ножек: нижних — с продолговатым мозгом (афферентная импульсация в мозжечок посту- Глава 4. Физиология центральной нервной системы 121 пает от вестибуло- и проприорецепторов), средних — с мостом и корой большого мозга, верхних — со средним мозгом (рис. 4.20). Роль мозжечка в организации движений. Мозжечок участвует в регуляции тонуса мышц — он оказывает тормозное влияние посредством красных и вестибулярных ядер, ретикулярной формации ствола мозга. Поддержание равновесия (позы) осуществляется с помощью перераспределения мышечного тонуса. Этому способствует импульсация от вестибулярных рецепторов, проприорецепторов, кожных, зрительных и слуховых рецепторов. Наиболее характерным симптомом поражения древнего мозжечка является нарушение равновесия. Координация выполняемых движений реализуется старым и новым мозжечком. Сюда поступает информация от проприорецепторов и от моторной коры большого мозга (программа движения). Координация достигается с помощью анализа этой информации и реализуется через свои промежуточные ядра, имеющие выходы на красное ядро и моторную кору. При этом регулируются амплитуда, направление и темп движения. Наиболее характерным симптомом нарушения функции промежуточной зоны мозжечка является нарушение координации движения (атаксия). Еще одна функция мозжечка — коррекция быстрых движений, КП Рис. 4.20. Схема основных межнейронных связей мозжечка (по В.Н. Яковлеву): КП — клетки Пуркинье; КЗ — клетки зерна; КК — корзинчатые клетки; ЗК — звездчатые клетки; кГ — клетки Гольджи; «+», «–» — возбуждающие и тормозные влияния 122 Физиология которая из-за большой скорости их развития не может быть осуществлена в процессе движения с помощью импульсаций от проприорецепторов. Мозжечок получает копию эфферентной программы движения от моторной коры большого мозга до начала движения. Кора мозжечка обучена в фило- и онтогенезе исправлять возможные ошибки, с учетом которых изменяется эфферентная программа движения (т.е. мозжечок выполняет роль, аналогичную акцептору результата действия в функциональной системе). К таким движениям относятся многие быстрые спортивные движения, игра на музыкальных инструментах и др. Программирование целенаправленных движений осуществляется корой нового мозжечка, которая получает импульсацию преимущественно из ассоциативных зон коры большого мозга через ядра моста. Эта информация характеризует замысел движения. Инициация движения обеспечивается лимбической системой и ассоциативной корой. В коре нового мозжечка (а также в базальных ядрах, в моторной коре) она перерабатывается в программу движения и через таламус поступает в премоторную кору. Там осуществляется дальнейшая обработка информации, и через пирамидную и экстрапирамидную системы она реализуется в виде сложного целенаправленного движения. Вегетативные функции мозжечка заключаются в том, что он, как и гипоталамус, участвует в согласовании вегетативного обеспечения соматической деятельности организма (модулирующее влияние на деятельность внутренних органов) посредством РФ, вегетативной нервной системы и эндокринной системы. После удаления мозжечка в эксперименте гомеостазис становится неустойчивым. 4.11. Функции промежуточного мозга Промежуточный мозг (diencephalon) расположен между средним и конечным мозгом. Он включает таламическую область (таламус, метаталамус и эпиталамус — эпифиз) и гипоталамус. Таламус (thalamus — зрительный бугор) и метаталамус (коленчатые тела). Таламус представляет собой парный ядерный комплекс, включающий до 60 ядер, которые Глава 4. Физиология центральной нервной системы 123 подразделяют на несколько групп: релейные (переключательные — сенсорные и несонсорные), специфические и неспецифические и ассоциативные. Изобилие терминов затрудняет усвоение материала. В частности термины «специфические» и «неспецифические» — некорректные. Каждое ядро или центр выполняет свою специфическую функцию. Поэтому термины «специфические» и «неспецифические» ядра целесообразно исключить. По функции основные ядра таламуса следует объединить в две группы: переключательные — сенсорные и двигательные (релейные, реле-переключатель) — и ассоциативные (объединяющие, интегрирующие). Причем, переключательные ядра не только посылают информацию к коре большого мозга, но и получают обратные входы от соответствующих зон коры большого мозга, обеспечивающих взаимодействие между таламусом и корой большого мозга. Переключательные ядра таламуса Переключательные сенсорные ядра (вентральные задние ядра) направляют потоки афферентной импульсации в сенсорные зоны коры большого мозга. В них переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая импульсации. В этих ядрах имеется топографическая проекция периферии; при этом функционально более тонко организованные ,части тела (например, язык, кисть) имеют большую зону представительства. В таламусе начинают формироваться ощущения — электростимуляция вентральных задних ядер вызывает парестезии (ложные ощущения) в разных частях тела. Импульсация от вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины, где формируются соответствующие ощущения. В латеральном коленчатом теле переключается зрительная импульсация и направляется в затылочную кору. Медиальное коленчатое тело переключает слуховую импульсацию, поступающую в височную кору. Переключательные двигательные ядра таламуса (вентральные) направляют программы движений, сформированные в базальных ядрах и в мозжечке, в двигательную кору большого мозга (поля 4 и 6). Ассоциативные ядра таламуса — это, во-первых, ядра РФ (ретикулярные ядра). Они имеют многочисленные вхо- 124 Физиология ды от других ядер головного мозга, сенсорных систем и коры большого мозга. В свою очередь, они посылают импульсы ко всем структурам ЦНС. Благодаря этим связям ретикулярные ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ядрами — с другой, объединяя их в единый функциональный комплекс. На кору мозга они оказывают преимущественно модулирующее (изменяющее состояние) влияние. Поэтому нарушение его функции не вызывает грубых расстройств. Во-вторых, ассоциативными являются группа ядер таламуса (подушка, медиодорсальное ядро, латеральные ядра — ассоциативные ядра таламуса), которые получают импульсацию от других ядер самого таламуса, от зрительных и слуховых путей и других ядер головного мозга и направляют ее в основном в ассоциативные зоны коры большого мозга. В свою очередь, кора большого мозга посылает импульсы к ассоциативным ядрам, регулируя их интегративную функцию. Гипоталамус включает преоптическую область, сосцевидные (мамиллярные) тела, область перекреста зрительных нервов, серый бугор и воронку. В гипоталамусе выделяют несколько десятков парных ядер, которые объединяют в несколько групп. Связи гипоталамуса. Его ядра образуют многочисленные связи друг с другом, а также с выше- и нижележащими структурами ЦНС. Главные входы в гипоталамус: от лимбической системы, коры большого мозга, базальных ядер и ретикулярной формации ствола мозга. Основные выходы гипоталамуса: в ствол мозга (ретикулярную формацию, моторные и вегетативные центры), в вегетативные центры спинного мозга, в лимбическую систему, от супраоптического и паравентрикулярного ядер — к нейрогипофизу, от вентромедиального и аркуатного ядер — к аденогипофизу, а также к стриатуму и лобной коре. Функции гипоталамуса. Он является многофункциональной системой, обладающей широкими регулирующими и интегрирующими влияниями в регуляции деятельности внутренних органов, обмена веществ и энергии. При этом обычно отдельно взятое ядро выполняет несколько функций, а отдельно взятая функция локализуется в не- Глава 4. Физиология центральной нервной системы 125 скольких ядрах. Свои влияния на функции внутренних органов гипоталамус осуществляет посредством гормонов эндокринных желез, симпатической (задние ядра) и парасимпатической (передние ядра) систем. Функции эпифиза описаны в разделе 6.5. 4.12. Функции базальных ядер и лимбической системы Основные структуры базальных ядер (стриопаллидарная система): полосатое тело (corpus striatum), называемое также striatum (хвостатое ядро и скорлупа), бледный шар и субталамическое ядро. Двигательные функции базальных ядер. В целом базальные ядра, имея двусторонние связи с корой большого мозга, таламусом, ядрами ствола мозга, участвуют в создании программ целенаправленных движений с учетом доминирующей мотивации. При этом нейроны стриатума оказывают тормозное влияние (медиатор — ГАМК) на нейроны черного вещества. В свою очередь, нейроны черного вещества (медиатор — дофамин) оказывают модулирующее влияние (тормозное и возбуждающее) на фоновую активность нейронов стриатума. При нарушении дофаминергических влияний на базальные ядра наблюдаются двигательные расстройства типа паркинсонизма, при которых резко падает концентрация дофамина в обоих ядрах стриатума. Наиболее важные функции базальных ядер выполняют стриатум и бледный шар. Функции стриатума. Участвует в осуществлении поворота головы и туловища и ходьбы по кругу, которые входят в структуру ориентировочного поведения. Поражение хвостатого ядра при заболеваниях и при разрушении в эксперименте ведет к насильственным, избыточным движениям (гиперкинезы: хорея и атетоз). Функции бледного шара. Оказывает модулирующее влияние на двигательную кору, мозжечок, РФ, красное ядро. При стимуляции бледного шара у животных преобладают элементарные двигательные реакции в виде сокращения мышц конечностей, шеи и лица, активация пищевого поведения. Разрушение бледного шара сопровождается сни- 126 Физиология жением двигательной активности — возникает адинамия (бледность двигательных реакций), а также развитие сонливости, «эмоциональной тупости», что затрудняет реализацию имеющихся условных рефлексов и ухудшает выработку новых (ухудшает кратковременную память). Структурно-функциональная организация лимбической системы (от лат. limbus — край) — это функциональное объединение различных структур, среднего, промежуточного и конечного мозга, обеспечивающее мотивацию поведенческих реакций и их вегетативное обеспечение. Мотивации формируют также эмоциональную окраску поведения. Структуры. Корковые: обонятельная извилина, гиппокамп (аммонов рог), зубчатая, поясная и парагиппокампальная извилины; подкорковые: миндалевидное тело, область перегородки, ограда, гипоталамус, переднее таламическое ядро. Входы в лимбическую систему осуществляются от различных областей головного мозга, от обонятельных рецепторов по волокнам обонятельного нерва (I, n. olfactorii). Главным источником возбуждения лимбической системы является РФ ствола мозга. Выходы из лимбической системы осуществляются через гипоталамус к нижележащим вегетативным и соматическим центрам ствола мозга и спинноПоясная извилина Парагиппокампова извилина А Таламус, передние ядра ГТ Гиппокамп МТ Б СМ Миндалина Рис. 4.21. Схема основных внутренних связей лимбической системы (по В.Н. Яковлеву): А — круг Пейпеца; Б — круг через миндалину; ГТ/МТ — мамиллярные тела гипоталамуса; СМ — средний мозг (лимбическая область) Глава 4. Физиология центральной нервной системы 127 го мозга (нисходящие влияния). Восходящая импульсация в новую кору направляется преимущественно в ассоциативные области. Кольцевые нейронные связи объединяют различные ее структуры и делают возможным длительную циркуляцию (реверберацию) возбуждения, которая является механизмом его пролонгирования (рис. 4.21). Функции лимбической системы. Получая информацию о внешней и внутренней средах организма, лимбическая система, после сравнения и обработки этой информации, запускает вегетативные и соматические (поведенческие) реакции, обеспечивающие адекватное приспособление организма к внешней среде и сохранение гомеостазиса. Частные функции лимбической системы: • регуляция функций внутренних органов, осуществляется преимущественно через гипоталамус; • формирование мотиваций, эмоций и поведенческих реакций (агрессивно-оборонительных, пищевых и сексуальных; • играет важную роль в обучении. 4.13. Нейронная организация новой коры. Функциональные зоны Новая кора (неокортекс) — это слой серого вещества, общая площадь которого за счет складок достигает 2000 см2, она покрывает большие полушария и составляет около 70% всей площади коры. Общая характеристика коры. В новой коре имеются поли- и монофункциональные поля. Свойство полифункциональности позволяет данной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию (Андрианов О.С., 1976). Степень полифункциональности различных корковых структур неодинакова: например, поля ассоциативной коры полисенсорны, первичные сенсорные зоны моносенсорны, во вторичных сенсорных зонах локализуются преимущественно бисенсорные нейроны. В основе мультифункциональности лежит многоканальность поступления в кору мозга афферентного возбуждения. Детальное деление коры на различные поля 128 Физиология проведено на основе цитоархитектонических признаков (формы и расположения нейронов) К. Бродманом (1909), который выделил 52 поля, многие из них характеризуются функциональными и нейрохимическими особенностями. В направлении с поверхности вглубь неокортекс имеет шесть горизонтальных слоев. В слоях I и IV новой коры происходят восприятие и обработка поступающих сигналов. Нейроны II и III слоев осуществляют кортикокортикальные ассоциативные связи. Нейроны V и VI слоев формируют нисходящие пути. Функциональные нейронные колонки новой коры большого мозга. В коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5–1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов (нейронные колонки). Об этом, в частности, свидетельствуют электрофизиологические исследования с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры. При этом все встречаемые на пути нейроны отвечают на раздражитель только одного вида (например, свет). При погружении электрода под углом на его пути попадались нейроны разной сенсорности. Колонки обнаружены в моторной коре и различных зонах сенсорной коры. Нейроны колонки могут осуществлять саморегуляцию по типу возвратного торможения. Соседние нейронные колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения. Кортикализация функций — это возрастание в филогенезе роли коры большого мозга в регуляции функций организма и подчинение нижележащих отделов ЦНС, в обеспечении психической деятельности организма. Например, регуляция локомоторных двигательных функций (прыжки, ходьба, бег) и выпрямительных рефлексов у низших позвоночных полностью обеспечивается стволом мозга, удаление больших полушарий практически их не изменяет. У кошек после перерезки ствола между средним и промежуточным мозгом локомоция сохраняется лишь частично. Выключение коры большого мозга в эксперименте у обезьян и в патологических случаях у человека ведет к потере не только произвольных движений, но и выпрямительных рефлексов. Глава 4. Физиология центральной нервной системы 129 Функциональные зоны новой коры. Функциональными отделами коры большого мозга являются сенсорные, ассоциативные и двигательные зоны (рис. 4.22). Сенсорные зоны коры (проекционная кора, корковые отделы сенсорных систем). Характеристика сенсорной коры. В эти зоны поступает информация от сенсорных рецепторов. Они расположены в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса. Участки сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения определенной чувствительности организма, называют первичными сенсорными зонами. Они состоят преимущественно из моносенсорных нейронов и формируют ощущения одного вида (качества). В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, от которых поступает импульсация (рис. 4.23). Вокруг первичных сенсорных зон локализуются вторичные сенсорные зоны, нейроны которых преимущественно бисенсорные — они реагируют обычно на два раздражителя, например на свет и звук. Во вторичные сенсорные зоны поступает также информация от первичных, что способствует формированию образа объекта или явления. Первичная моторная Первичная кора соматосенсорная кора Префронтальная ассоциативная кора Вторичная зрительная кора Первичная зрительная кора Рис. 4.22. Функциональные области коры большого мозга ть Ки с ль цы ль па шой ле ц Па Бо Стопа Половые органы Рук а Суст авы кист и Нога Шея Голова Физиология Таз Торс 130 з Гла с Но о Лиц Губы Челюсть и зубы Язык Гло тка Рис. 4.23. Чувствительный гомункулус — проекция общей чувствительности в коре (постцентральная извилина) (по У. Пенфилду, Т. Расмуссену, 1950) Главные сенсорные зоны 1. Соматосенсорная область (SI) — это постцентральная извилина теменной доли и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1–3). При этом поле 3 представляет собой первичное сенсорное поле, а поля 1–2 — вторичные сенсорные поля. В области SI имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной и проприоцептивной чувствительности. Проекции наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеют относительно большие зоны по сравнению с другими частями тела. Предполагают, что в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности. Имеются также меньших размеров вторичная соматосенсорная область (SII), расположенная в стенке боковой борозды, на границе ее пересечения с центральной бороздой. Лобная и теменная доли разделены центральной бороздой. Зона SII, по-видимому, участвует в сенсорной и моторной коор- Глава 4. Физиология центральной нервной системы 131 динации двух сторон тела, локализация поверхности тела в ней менее четкая, чем в области SI. 2. Височная область — здесь в верхней височной извилине расположена слуховая сенсорная кора. Первичной сенсорной зоной является поле 41, в глубине боковой борозды с четкой топической проекцией различных участков кортиева органа. Вторичная сенсорная зона расположена в верхней височной извилине (поле 42), где происходит более сложная обработка звуковой (частично и речевой) информации; 3. Затылочная область (поле 17), где локализуется первичная сенсорная зона зрительной системы с топическим представительством рецепторов сетчатки, особенно желтого пятна (основная часть поля 17), рядом располагается вторичная сенсорная зона (поля 18, 19). Обработанная в первичной и вторичной сенсорной коре информация передается для дальнейшей обработки в ассоциативную кору — третичную сенсорную зону. Взаимодействие трех сенсорных зон обеспечивает с помощью мышления формирование комплексного восприятия и формирование соответствующего поведения. Ассоциативные области коры. Следует заметить, что описание ассоциативных областей несколько различается у разных авторов. Теменная ассоциативная кора (поля 5, 7, 40) получает основные афферентные входы от задней группы ассоциативных ядер таламуса, имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы. Основными функциями теменной ассоциативной коры являются гнозис и праксис. Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, узнавание их на ощупь, понимание речи, познание процессов, закономерностей и др. Под праксисом понимают целенаправленное автоматизированное действие (например, печатание на клавиатуре компьютера). Понимание сложных логико-грамматических конструкций обеспечивается нижнетеменными и теменно-затылочными областями коры большого мозга. Эти области обеспечивают ориентировку в пространстве. 132 Физиология Лобная (фронтальная) ассоциативная кора имеет основной афферентный вход от теменной ассоциативной коры и ассоциативного медиодорсального ядра таламуса. Главная функциия лобной ассоциативной коры — формирование программ целенаправленного поведения (особенно в новой для человека обстановке), под влиянием доминирующей мотивации, обеспечивающей направление поведения человека. Эта функция основана на тесных двусторонних связях лобной коры с лимбической системой. Причем лобная кора обеспечивает вероятностное прогнозирование, что выражается изменением поведения в ответ на изменения обстановки окружающей среды и доминирующей мотивации. В лобной коре осуществляется самоконтроль действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата предвидения (акцептора) результата действия в функциональной системе по П.К. Анохину. В результате проведения по медицинским показаниям префронтальной лоботомии, при которой пересекаются связи между лобной долей и таламусом, наблюдается развитие «эмоциональной тупости», отсутствие мотивации, твердых намерений и планов, основанных на прогнозировании. План поведения, формируемый ассоциативной корой, реализуется с помощью двигательной коры. Некоторые авторы выделяют теменно-затылочно-височную ассоциативную область, функции которой в основном совпадают с вышеописанными, а также — лимбическую область, участвующую в формировании мотиваций, эмоций и поведения. Двигательные зоны коры — это первичная двигательная (моторная) кора прецентральной извилины (поле 4) и вторичная двигательная кора (поле 6), которая включает прилегающую к ней рострально-премоторную область и дополнительную двигательную область на медиальной стороне полушария. Пирамидные нейроны V слоя этих областей (формирующие пирамидные пути) составляют двигательные колонки и иннервируют α- и γ-мотонейроны двигательных центров как непосредственно (моносинаптически), так и через интернейроны двигательных центров (полисинаптически). Пирамидные нейроны двигательной Глава 4. Физиология центральной нервной системы 133 сть Ки Рук а Сус та кис вы ти Таз Нога Стопа Торс колонки получают импульсы от проприо- и тактильных рецепторов, формируют двигательные команды и посылают их через нейроны верхних слоев к стволовым и спинальным центрам, последние управляют мышцами одного сустава или одной мышцы, но мышцы туловища могут иннервироваться от нескольких колонок. Соседние колонки в функциональном плане перекрываются. В двигательные области информация поступает по восходящим афферентам через таламус, непосредственно от таламуса, и интегрированная информация от корковых нейронов различных сенсорных систем, ассоциативной коры и лимбической системы. Первичная двигательная кора. Ее пирамидные клетки, иннервируют мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (рис. 4.24). Нейроны коры управляют функцией мышц противоположной стороны тела. При поражениях этой корковой зоны нарушается способность к тонким координированным движениям конечностей, исключаются тонкие движения пальцев руки на противоположной стороне, движение языка и нижней части лица. Па цы ль ой льш ц Бо пале Лоб з Гла ко и ве о Лиц Мимические мышцы Челюст ь Язы к ка от Гл Рис. 4.24. Двигательный гомункулус — проекция двигательной системы в коре (прецентральная извилина) (по У. Пенфилду, Т. Расмуссену, 1950) 134 Физиология Вторичная двигательная кора имеет главенствующее значение по отношению к первичной двигательной коре и обеспечивает осуществление более сложных координированных движений, играет важную роль в планировании движения. Поражения вторичной двигательной зоны ведут к резкому ограничению движений у человека. В премоторной коре локализуется моторный центр речи (центр Брока), управления движениями глаз. При раздражении вторичной коры возникают более сложные движения (конечностей, туловища, головы), чем при раздражении первичной двигательной коры, например одного сустава, движения пальцев рук. Двигательная кора осуществляет свои функции через пирамидную и экстрапирамидную системы. Пирамидная система — совокупность нейронов пирамидного пути (от первичной двигательной коры), αи γ-мотонейронов ствола мозга и спинного мозга, синаптически связанных друг с другом непосредственно и с помощью вставочных нейронов. Пирамидный путь образуют аксоны пирамидных клеток V слоя коры большого мозга (клетки Беца, первичная двигательная кора). Пирамидная система играет особо важную роль в выполнении тонких движений пальцев рук и кистей. Экстрапирамидная система (ЭПС) — совокупность двигательных ядер головного мозга, большинство из которых не имеют прямой связи с α- и γ-мотонейронами спинного мозга и ствола мозга или со вставочными нейронами спинного мозга. Ядра ЭПС — двигательная кора (кроме первичной), базальные ядра, черная субстанция, красные ядра, люисово тело (субталамическое ядро), ретикулярная формация, центры четверохолмия, голубое пятно, вестибулярные ядра и мозжечок. Функция — регуляция мышечного тонуса, сохранение естественной позы, восстановление нарушенной позы, участие в осуществлении произвольных движений. Межполушарные функциональные связи осуществляются через мозолистое тело, а также комиссуры (спайки), они играют важную роль в деятельности организма, что показано в эксперименте на животных с рассечением мозолистого тела и в результате подобной операции у пациентов по медицинским показаниям (чтобы прекратить межполушарное распространение эпилептических судорог). При Глава 4. Физиология центральной нервной системы 135 этом возникают различные нарушения — двигательные, речевые, узнавания с помощью зрительной и тактильной систем. Такие больные могут рассказать о соматосенсорных стимулах, нанесенных на правую сторону тела, но не на левую, т.к. информация, поступающая в правые сенсомоторные области коры (от левой половины тела), не достигает речевых центров (они расположены в левом полушарии), но полушария могут выполнять отдельные функции независимо друг от друга. У здорового же организма мозг работает как единое целое. 4.14. Методы исследования ЦНС Наибольшее распространение получили методы регистрации биоэлектрической активности отдельных нейронов, суммарной активности нейронного пула или головного мозга в целом (электроэнцефалография), компьютерная томография (позитронно-эмиссионная томография, магнито-резонансная томография). Электроэнцефалография (ЭЭГ) — это регистрация c поверхности кожи головы суммарного электрического поля, генерируемого нейронами мозга при их возбуждении. Это поле — ПД нейронов и нервных волокон, ВПСП (не достигших величины КУД), ТПСП, следовых гиперполяризации и деполяризации нейронов. На ЭЭГ регистрируется четыре основных физиологических ритма: α-, β-, θ- и δ-ритмы, частота и амплитуда которых отражают степень активности ЦНС (рис. 4.25). Метод вызванных потенциалов (ВП) заключается в регистрации изменений электрической активности мозга, возникающих в ответ на раздражение сенсорных рецепторов или афферентных путей (рис. 4.26). Метод ВП используется для изучения функций сенсорных систем, в частности локализации сенсорных зон в коре большого мозга. Позитронно-эмиссионная томография — метод функционального изотопного картирования мозга. Основан на введении в кровоток изотопов (N13, F18, О15) в соединении с дезоксиглюкозой. Чем активнее участок мозга, тем больше он поглощает меченой глюкозы. Ее радиоактивное излучение регистрируется детекторами, расположенными вокруг голо- 136 Физиология α 8–13 Гц, до 70 мкВ 50 мкВ β 14–30 Гц, до 30 мкВ θ 4–7 Гц, до 200 мкВ А δ 0,5–3 Гц, до 300 мкВ Б 1с 1 2 Рис. 4.25. Основные ритмы электроэнцефалограммы (ЭЭГ): А — затылочной области коры большого мозга; Б — реакция десинхронизации ЭЭГ затылочной области коры большого мозга при открывании глаз (↑) и восстановление альфа-ритма при закрывании глаз (↓) Н1 П1 0 Н3 Н2 П2 100 Н4 П3 П4 200 300 2 мкВ 400 мс Рис. 4.26. Вызванные потенциалы (ВП) у человека на вспышку света: П — позитивные; Н — негативные компоненты ВП; цифровые индексы означают порядок следования позитивных и негативных компонентов в составе ВП. Начало записи совпадает с моментом включения вспышки света (стрелка) вы. Информация от детекторов поступает на компьютер, который создает «срезы» мозга на регистрируемом уровне, отражающие неравномерность распределения изотопа в связи с метаболической активностью мозговых структур, что позволяет судить о возможных поражениях ЦНС. Магнитно-резонансная томография позволяет выявить активно работающие участки мозга. Методика основана на том, что после диссоциации оксигемоглобина ге- Глава 4. Физиология центральной нервной системы 137 моглобин приобретает парамагнитные свойства. Чем выше метаболическая активность мозга, тем больше объемный и линейный кровоток в данном участке мозга и тем меньше отношение парамагнитного дезоксигемоглобина к оксигемоглобину. В мозге существует много очагов активации, что отражается в неоднородности магнитного поля. Стереотаксический метод. Этот метод позволяет с помощью специального устройства точно вводить макро- и микроэлектрод, термопару в различные структуры головного мозга. Координаты структур мозга приведены в стереотаксических атласах. Через введенные электроды можно регистрировать биоэлектрическую активность данной структуры, раздражать или разрушать ее, через микроканюли — вводить химические вещества в нервные центры или желудочки мозга, с помощью микроэлектродов (d ~ 1 мкм), подведенных вплотную к клетке, — регистрировать импульсную активность отдельных нейронов и судить об их участии в рефлекторных, регуляторных и поведенческих реакциях, а также о возможных патологических процессах и применении соответствующих лечебных воздействий фармакологическими препаратами. Пальцево-носовая проба. Испытуемому предлагают дотронуться указательным пальцем до кончика носа с открытыми, а затем с закрытыми глазами. В норме отмечается попадание (дотрагивание до кончика носа). При травмах головного мозга, переутомлении, перетренированности, неврозах наблюдается промахивание (непопадание) и дрожание (тремор) указательного пальца или кисти. Запись тремора до и после тренировочного занятия позволяет корригировать тренировочный процесс, поскольку улучшение тренированности сопровождается, как правило, снижением величины тремора. Имеются и другие пробы (например, проба Ромберга, которая изучается в неврологии). ГЛАВА 5 Знание современной физиологии дает врачу возможность понять, что произойдет в организме пациента, если ввести ему то или иное лекарство. Вегетативная нервная система Вегетативная (автономная) нервная система (ВНС) — это часть нервной системы, регулирующая работу внутренних органов, обмен веществ и энергии, обеспечивая гомеостазис организма в различных условиях жизнедеятельности. 5.1. Общая характеристика ВНС Отделы ВНС. В настоящее время общепризнанными являются два отдела вегетативной нервной системы — симпатический и парасимпатический; влияния их на органы, как правило, противоположны (табл. 5.1). Таблица 5.1 Основные симпатические и парасимпатические влияния Органы Симпатический нерв, эффекторные рецепторы Парасимпатический нерв, эффекторные рецепторы Гладкомышечные 1. Все гладкомыРасслабление, α1, β2 шечные внутренние органы, кроме сердца 2. Cфинктеры Сокращение, α1 3. Cосуды В основном сужение, α1; коронарные и мышечные: α1 — сужение, β2 — расширение 4. ЖКТ (секреция) Снижение, α1 Сердце Стимуляция, β1 Мужские половые Эякуляция, α1 органы Сокращение, М2 Расслабление, М3 Не иннервируются (за исключением некоторых органов) Стимуляция, М3 Торможение, М2 Эрекция, М2 Глава 5. Вегетативная нервная система Органы Жировые клетки 139 Симпатический нерв, эффекторные рецепторы Липолиз, α, β1 Парасимпатический нерв, эффекторные рецепторы Влияние изучено недостаточно В последние годы получены убедительные факты, доказывающие наличие серотонинергического отдела вегетативной нервной системы, усиливающего сокращения гладких мышц желудочно-кишечного тракта. Серотонинергические волокна идут в составе симпатических нервов, их там в несколько раз больше, чем адренергических. Дуга вегетативного рефлекса построена по общему принципу (рис. 5.1). Но имеются особенности ее организации. 1. Главное отличие заключается в том, что рефлекторная дуга ВНС может замыкаться вне ЦНС. Эффекторный нейрон для симпатического отдела нервной системы расположен экстраорганно — в симпатических ганглиях, а для парасимпатической, как правило, — внутри органа или в непосредственной близости от него. 2. Афферентное звено дуги вегетативного рефлекса может быть образовано как собственными — вегетативными, так и соматическими афферентными волокнами. 3 3 2 1 4 5 Рис. 5.1. Рефлекторная дуга вегетативного рефлекса: 1 — воспринимающее звено, рецептор; 2 — афферентное звено, чувствительный нейрон; 3 — управляющее звено, совокупность центральных и периферических нейронов; 4 — эфферентное звено, аксон эффекторного нейрона; 5 — эффектор, рабочий орган 140 Физиология 3. В дуге вегетативного рефлекса слабее выражена сегментарность, что повышает надежность вегетативной иннервации. Общая схема ВНС представлена на рис. 5.2. Функциональные отличия ВНС от соматической нервной системы связаны со структурными ее особенностями и степенью выраженности влияния на ВНС коры большого мозга. Имеется четыре основных отличия. 1. Регуляция функций внутренних органов с помощью ВНС может осуществляться (хотя и менее совершенно) при полном нарушении связи с ЦНС, поскольку эффекторный нейрон ВНС находится за пределами ЦНС: либо в экстра-, либо в интраорганных вегетативных ганглиях, образующих периферические экстра- и интраорганные рефлекторные дуги. При нарушении же связи скелетных мышц с ЦНС III Средний мозг VII Мост IX Продолговатый мозг Сонный синус X Слезные и слюнные железы Трахея и бронхи Сердце CVII–ThI–XII Скелетные мышцы Глаз LI–III Артериола Печень с желчными протоками и желчным пузырем Желудок Тонкая кишка Проксимальный отдел толстой кишки Мозговое вещество надпочечников Почки с мочеточниками Дистальный отдел толстой кишки Прямая кишка SII–IV Мочевой пузырь Холинергические нейроны Адренергические нейроны Чувствительные (афферентные) нейроны Рис. 5.2. Общая схема холинергической и адренергической иннервации внутренних органов Глава 5. Вегетативная нервная система 141 соматические рефлексы утрачиваются, поскольку все эффекторные мотонейроны находятся в ЦНС. 2. Влияние ВНС на органы и ткани организма не контролируется непосредственно сознанием (человек не может произвольно управлять частотой и силой сердечных сокращений, сокращений желудка, секрецией кишечных желез и т.д.). 3. Генерализованный (диффузный) характер влияния в симпатическом отделе ВНС объясняется двумя основными факторами. Во-первых, большинство адренергических нейронов имеет длинные постганглионарные тонкие аксоны, многократно ветвящиеся в органах и образующие так называемые адренергические сплетения. Во-вторых, в генерализации эффектов симпатической нервной системы постганглионарных волокон в 50–100 раз больше, чем преганглионарных (в ганглиях нейронов больше, чем преганглионарных волокон). В парасимпатических узлах каждое преганглионарное волокно контактирует только с однойдвумя ганглионарными клетками. В органах с двойной иннервацией эффекторные клетки получают симпатическую и парасимпатическую иннервацию. Мышечные клетки желудочно-кишечного тракта имеют тройную экстраорганную иннервацию — симпатическую (адренергическую), парасимпатическую (холинергическую) и серотонинергическую, а также от нейронов интраорганной нервной системы. Некоторые из них, например мочевой пузырь, получают в основном парасимпатическую иннервацию, ряд органов имеют только симпатическую (потовые железы, мышцы, поднимающие волосы, селезенка, надпочечники). Преганглионарные волокна симпатической и парасимпатической нервной системы — холинергические и образуют синапсы с ганглионарными нейронами с помощью ионотропных N-холинорецепторов (медиатор — ацетилхолин, рис. 5.3). Рецепторы получили свое название (Д. Ленгли) из-за чувствительности к никотину: малые его дозы возбуждают нейроны ганглия, большие — блокируют. Симпатические ганглии расположены экстраорганно, парасимпатические, как правило, — интраорганно или в непосредственной близости от органа. 142 Физиология Симпатическая нервная система N-холинорецепторы Ганглии α Парасимпатическая нервная система М β Эффекторная клетка α-, β-адренорецепторы М-холинорецепторы преганглионарные волокна постганглионарные волокна Рис. 5.3. Нейроны и рецепторы симпатической и парасимпатической нервной системы: А — адренергические нейроны; Х — холинергические нейроны 5.2. Характеристика отделов ВНС Симпатическая нервная система. Преганглионарные симпатические нервные волокна выходят из спинного мозга в составе передних корешков, затем отделяются от них и прерываются в узлах симпатического ствола или в других экстраорганных ганглиях. Преганглионарные нейроны симпатической нервной системы расположены в боковых рогах спинного мозга (сегменты CVII–LII — центр Якобсона). От нейронов симпатических ганглиев идут постганглионарные волокна, иннервирующие все органы и сосуды организма, включая ЦНС и сенсорные рецепторы (см. рис. 5.1). Медиатор постганглионарных симпатических волокон — норадреналин, около 90% (адреналин — около 7%, дофамин — около 3%). Синтезируется норадреналин из аминокислоты тирозина, в основном в терминальных симпатических окончаниях и только около 1% — в теле нейрона. Поскольку в окончаниях симпатических ганглионарных нейронов медиатор — норадреналин, их называют адренергическими (см. рис. 5.3). Инактивирование медиатора, выброшенного в синаптическую щель, осуществляется за счет обратного его захвата (75–80%) пресинаптической мембраной. Не попавший в пузырьки норадреналин разрушается ферментами, локализованными в пресинаптической терминали, — мо- Глава 5. Вегетативная нервная система 143 ноаминоксидазой (МАО) и катехол-0-метил-трансферазой (КОМТ). Часть норадреналина, высвободившегося в синаптическую щель, диффундирует в интерстиций и кровь. В большинстве органов, реагирующих на катехоламины, содержатся α- и β-адренорецепторы, причем одна гладкомышечная клетка может иметь оба рецептора. Эффекты активации α- и β-адренорецепторов зависят от преобладания каждого из них в данном органе и различной чувствительности α- и β-рецепторов к медиатору. При возбуждении симпатических (адренергических) нервных волокон активация α1- и α2-адренорецепторов клеток-эффекторов в разных органах вызывает различные реакции (см. табл. 5.1). В частности, сосуды суживаются, моторика желудочно-кишечного тракта угнетается. Активация β-адренорецепторов также вызывает разнонаправленные эффекты в различных органах: в желудочнокишечном тракте — тормозные; в сердце (β1-рецепторы) — стимуляторные. Если высвободившийся из нервного окончания норадреналин вызывает ВПСП, функция органа усиливается, если ТПСП — функция органа тормозится. Все эффекторные рецепторы метаботропные (активируются с помощью вторых посредников, т.е. вторых медиаторов, с помощью биохимических реакций). Они являются ферментами (цАМФ, цГМФ и др.), запускают биохимические реакции, что ведет к возникновению ВПСП или ТПСП. Преганглионарные симпатические волокна образуют возбуждающие холинергические синапсы также и на клетках мозгового вещества надпочечников. Высвобождение катехоламинов из мозгового вещества надпочечников регулируется исключительно нервными влияниями (перерезка преганглионарных симпатических волокон прекращает высвобождение катехоламинов). При возбуждении преганглионарных симпатических волокон у человека из надпочечников в кровоток обычно выбрасывается смесь катехоламинов, состоящая из адреналина (80–90%) и норадреналина (10–20%). Выброс катехоламинов имеет важное приспособительное значение, обеспечивая мобилизацию систем организма при физическом и эмоциональном напряжении. Точки приложения для продуцируемых надпочечниками катехоламинов те же, что и у симпатической нервной системы. 144 Физиология Парасимпатическая нервная система. Парасимпатическую иннервацию (cм. рис. 5.1) получают не все органы и сосуды. В частности, ее не имеют подавляющее большинство кровеносных сосудов (кожи, органов брюшной полости, мышц, мозга), органы чувств и мозговое вещество надпочечников, скелетные мышцы, ЦНС. Парасимпатические нервные волокна имеются в черепных нервах. Парасимпатические волокна III пары (глазодвигательный нерв) иннервируют глазные мышцы (m. sphincter pupillae и m. ciliaris), регулируя диаметр зрачка и степень аккомодации. Парасимпатические веточки VII пары (лицевой нерв): n. petrosus major — секреторный нерв, иннервирует слизистую оболочку носа, нёба, слезную железу; n. chorda tympani — подчелюстную и подъязычную слюнные железы. Парасимпатические секреторные волокна IX пары (языкоглоточный нерв) подходят к околоушной железе в составе n. auriculotemporalis — от третьей ветви тройничного нерва. Преганглионарные волокна блуждающего нерва (X пара) берут начало от дорсального и двойного ядер продолговатого мозга и иннервируют глотку, гортань, трахею, бронхи, сердце, пищевод, печень, поджелудочную железу и большую часть желудочно-кишечного тракта, мягкое нёбо. Парасимпатические нервы сакрального отдела спинного мозга (S2–S4) иннервируют нисходящую часть ободочной кишки и тазовые органы (прямую кишку, мочевой пузырь, половые органы). Парасимпатические ганглии (в отличие от симпатических, локализующихся экстраорганно) расположены внутри органов, а в области головы и тазовых органов — в непосредственной близости от органов. От нервных клеток парасимпатических ганглиев идут короткие постганглионарные парасимпатические волокна, иннервирующие эффекторные клетки органов. Медиатор преганглионарных волокон — ацетилхолин, как и преганглионарных симпатических (см. рис. 5.2). Медиатор действует на ионотропные N-холинорецепторы постсинаптической мембраны ганглионарного нейрона. Ионотропные рецепторы — это рецепторы, ионные каналы которых активируются непосредственно медиато- Глава 5. Вегетативная нервная система 145 ром. Во всех эффекторных рецепторах медиатор активирует G-белок (ГТФ-связывающий белок). Медиатором постганглионарных парасимпатических волокон также является ацетилхолин. Высвобождению квантов медиатора способствуют ионы Са2+. Ацетилхолин синтезируется в цитоплазме окончаний холинергических нейронов, депонируется в везикулах по несколько тысяч молекул в каждой из них. Инактивирование медиатора. Часть ацетилхолина разрушается ферментом ацетилхолинэстеразой с образованием холина и уксусной кислоты, которые захватываются пресинаптической мембраной и вновь используются для синтеза ацетилхолина, часть диффундирует в интерстиций. Эффекторные рецепторы. Ацетилхолин, высвобождаемый постганглионарными терминалями, реализует свое влияние посредством М-холинорецепторов (см. рис. 5.3), которые получили название от мускарина — токсина мухомора, активирующего эти рецепторы и вызывающего такой же эффект, как и ацетилхолин. Активация М-холинорецепторов эффекторных клеток разных органов вызывает противоположные реакции по сравнению с симпатическими, в частности торможение работы сердца, сокращение гладких мышц желудочно-кишечного тракта — перистальтика усиливается, сокращение мышц бронхов — их просвет сужается и т.д. (табл. 5.1). Парасимпатическая нервная система способствует поддержанию гомеостазиса, обеспечивая трофотропный эффект (анаболизм). Особо важно помнить врачу тот факт, что блокада любых рецепторов может привести к разнонаправленным реакциям различных органов. Например, инъекция атропина, который, попадая в кровь, разносится по всему организму и блокирует все М-холинорецепторы, ведет к учащению сокращений сердца, но к торможению сокращений желудка и кишечника. Экстраорганные вегетативные нервы действуют на органы как непосредственно, так и с помощью интраорганной нервной системы, что особенно характерно для желудочно-кишечного тракта. 146 Физиология Механизм реализации симпатических и парасимпатических влияний на органы: тормозные влияния осуществляются посредством гиперполяризации постсинаптических мембран рабочих клеток, стимуляторные — посредством их деполяризации — ВПСП и ПД. Подавляющее число эффекторных α- и β-адрено-, М-холино- и серотонинорецепторов — метаботропные: они активируются с помощью вторых посредников, т.е. вторых медиаторов (цАМФ, цГМФ и др.). Это ферменты, запускающие биохимические (метаболические) реакции, обеспечивающие возникновение ВПСП или ТПСП и соответствующую реакцию органа. У симпатической и парасимпатической нервных систем только N-холинорецептор ионотропный: ацетилхолин без вторых посредников действует непосредственно на рецептор, обеспечивает возникновение ВПСП или ТПСП. В последние годы получены убедительные факты о наличии серотонинергического отдела ВНС. Серотонинергические пути в ЦНС идут от ядер шва продолговатого мозга к симпатическим центрам спинного мозга и образуют синапсы с их нейронами. На периферии преганглионарные серотонические волокна идут в составе симпатических нервов, их там в несколько раз больше, чем адренергических. Серотонинергические нервные волокна усиливают сокращения гладких мышц ЖКТ. Они образуют синапсы с серотонинергическими нейронами вегетативных ганглиев, иннервирующих гладкомышечные волокна органов ЖКТ. Эффекты их раздражения подчиняются закону силы, их возбудимость и скорость проведения возбуждения ниже таковых симпатических нервов, а сила стимуляторного влияния на орган значительно превосходит тормозное влияние симпатических нервов (рис. 5.4. и 5.5). Серотонинергический нерв (как и симпатический) — понятие собирательное, это симпатические стволы, их разветвления и веточки, иннервирующие внутренние органы. Схема эфферентного отдела ВНС предоставлена на рис. 5.6. Регуляция функции синапсов. Центральная регуляция. Эффективность передачи в синапсах ВНС определяется числом импульсов, пришедших к нервному окончанию: Глава 5. Вегетативная нервная система 147 20 мм рт. ст. А Б В Рис. 5.4. Усиление сокращений желудка (запись давления в его полости) при раздражении симпатического ствола в грудной полости у собаки: А — до введения фармакологических препаратов; Б — в условиях блокады симпатического нерва; В — в условиях блокады серотонинергического нерва. Отметка раздражения нерва — жирная часть нулевой линии (30 с). Опыт В.М. Смирнова, И.Л. Мясникова, Д.С. Свешникова 1 2 А 20 мм рт. ст 1 2 Б Рис. 5.5. Усиление сокращений двенадцатиперстной кишки с увеличением силы раздражения правого симпатического ствола в грудной полости у собаки (в начале раздражения — слабое угнетение): А — раздражение 5 В, 20 Гц, 0,5 мс; Б — раздражение 10 В, 20 Гц, 1,5 мс; 1 — гидростатическое давление в полости кишки (мм рт. ст.); 2 — нулевая линия и отметка раздражения (жирная часть линии, 30 с). Опыт В.М. Смирнова, Д.С. Свешникова, И.Л. Мясникова с увеличением импульсации высвобождение медиатора до определенного значения увеличивается, при ослаблении импульсации — уменьшается. Ауторегуляция с помощью медиатора. На окончаниях нервных волокон ВНС имеются пресинаптические рецепторы к серотонину, гистамину, ацетилхолину и др. Активация одних из этих рецепторов облегчает передачу в синап- 148 Физиология 1 2 Рис. 5.6. Функциональная организация эфферентного отдела ВНС (на примере желудка и тонкой кишки). Нейроны: А — адрен-, С — серотонин-, ВИПергические и соответствующие их медиаторам рецепторы: α-, β-, N-, M- и ВИП-рецепторы (ВИП — вазоактивный интестиальный пептид). Нервные волокна: 1 — серотонинергические; 2 — симпатические се, других — угнетает. Степень активации этих рецепторов связана с высвобождением медиатора по принципу обратной отрицательной связи и разной чувствительностью рецепторов к медиатору: малые дозы медиатора стимулируют его высвобождение, большие — угнетают (рис. 5.7). Ауторегуляция с помощью синтеза рецепторов. При уменьшении высвобождения медиатора синтез рецепторов постсинаптической мембраны возрастает, чувствительность ее повышается (сенситизация); при увеличении выхода медиатора синтез рецепторов постсинаптической мембраны уменьшается, чувствительность ее снижается (десенситизация), что также является важным регуляторным механизмом. Имеется перекрестное взаимное торможение активности симпатических и парасимпатических терминалей (см. рис. 5.6). Глава 5. Вегетативная нервная система Адренергическое окончание α β М НА Холинергическое окончание М α АХ N НА α 149 АХ β М Эффекторная клетка Рис. 5.7. Ауторегуляция выделения медиатора симпатическими и парасимпатическими постганглионарными нервными окончаниями с помощью пресинаптических αи β-адренорецепторов, M- и N-холинорецепторов и их реципрокные взаимодействия. Влияния: — угнетение; — увеличение высвобождения медиатора; НА — норадреналин; АХ — ацетилхолин Экстраорганные нервы взаимодействуют с метасимпатической нервной системой. Особенно это характерно для ЖКТ. Метасимпатическая нервная система (А.Д. Ноздрачев). Структурно-функциональная характеристика. Многие внутренние органы продолжают функционировать, будучи извлеченными из организма: сокращаются перфузируемое сердце, мочеточники, желчный пузырь, кишечник. В пищеварительном тракте после перерезки симпатических и парасимпатических путей сохраняются все виды двигательной активности. Двигательная активность пищеварительных органов после перерезки их нервов регулируется рефлекторными дугами, замыкающимися в пределах мышечного и подслизистого сплетений его стенок, а также в экстраорганных ганглиях. Дуги рефлексов, обусловливающих перистальтику, лежат в пределах стенки кишечника. Возбуждающие нейроны высвобождают ацетилхолин и серотонин, хотя не исключено, что их действие может быть опосредовано и другими медиаторами. Возбуждение афферентных нейронов кишечной стенки под действием пищевого комка приводит к возникновению рефлексов, при которых пищевой комок продви- 150 Физиология гается в каудальном направлении путем сокращения мышц орального участка кишки и одновременного расслабления каудального участка. Тормозные нейроны этих дуг — ВИП-нейроны (медиатор — вазоактивный интестинальный пептид). Комедиатор (модулятор) ВИП-нейронов — АТФ. Имеются спонтанно активные эфферентные нейроны; на них могут конвергировать интра- и экстраорганные нервные волокна. Таким образом, метасимпатическая нервная система — важный фактор надежности регуляторных механизмов организма. Метасимпатические нейроны взаимодействуют между собой и с экстраорганными нервами. Это особенно характерно для желудочно-кишечного тракта. Парасимпатические преганглионарные волокна образуют синапсы на возбуждающих холинергических и тормозных ВИПергических нейронах. Рефлекторные дуги метасимпатической нервной системы содержат все необходимые для осуществления рефлексов элементы: афферентный, вставочный и эффекторный нейроны. Медиаторы и рецепторы интраорганной нервной системы весьма разнообразны — в энтеральной ее части функционирует около 20 различных медиаторов и модуляторов. Так, серотонин стимулирует сокращение гладких мышц. Вазоактивный интестинальный пептид (ВИП) — медиатор интраорганных тормозных нейронов желудочно-кишечного тракта, ацетилхолин — возбуждающих нейронов. 5.3. Центры ВНС и их тонус. Трофическое действие нервной системы Особо важное значение для вегетативной регуляции имеет гипоталамус. Он содержит до 50 пар ядер. Роль различных групп ядер гипоталамуса определяется их связью с симпатическим или парасимпатическим отделами ВНС. Передние ядра регулируют деятельность парасимпатической нервной системы, задние — симпатической нервной системы. Поэтому разрушение задних ядер гипоталамуса сопровождается нарушением обмена веществ, механизмов теплообразования и теплоотдачи. Передние ядра гипоталамуса стимулируют половое созревание, задние, напротив, тормозят. Функции средней группы ядер гипоталамуса: ла- Глава 5. Вегетативная нервная система 151 теральное ядро — это центр голода, вентромедиальное — центр насыщения. Нейроны гипоталамуса обладают рецепторной функцией — они способны непосредственно улавливать изменения химического состава крови и цереброспинальной жидкости и при необходимости устранять их посредством вегетативных нервов и гормональных сдвигов. Гипоталамус вырабатывает гормоны (окситоцин, вазопрессин) и факторы, регулирующие функции эндокринных желез. В гипоталамусе имеется центр, регулирующий цикл сон–бодрствование. Лимбическая система участвует в приспособительной регуляции деятельности сердечно-сосудистой, дыхательной, пищеварительной и других систем согласно потребностям организма. Особенно это проявляется при формировании мотиваций и возникновении эмоций. Влияние лимбической системы осуществляется с помощью эндокринных желез и ВНС. Роль ретикулярной формации в регуляции вегетативных функций заключается в том, что ее нейроны формируют жизненно важные центры продолговатого мозга — дыхательный, кровообращения. Они реализуют свое влияние посредством активации симпатической нервной системы. Серотонинергические нейроны ядер шва (продолговатый мозг) образуют серотонинергические пути, аксоны которых поступают к нейронам, расположенным вместе с симпатическими нейронами в боковых рогах спинного мозга (Колас А. и др., 1989–2002), откуда выходят серотонинергические нервные волокна, стимулирующие сокращения желудка и кишки (Смирнов В.М. и др., 1986–2012). Мозжечок. Раздражение структур мозжечка вызывает сужение кровеносных сосудов, расширение зрачка, учащение сердцебиения, изменение интенсивности дыхания, кроветворения, терморегуляторные реакции. Мозжечок стабилизирует гомеостазис — при удалении мозжечка гомеостазис становится неустойчивым, в частности угнетается активность кишечных желез и моторики желудочно-кишечного тракта. Мозжечок реализует свое влияние с помощью симпатической нервной системы и эндокринных желез. Кора большого мозга с помощью соиннервации (управление двигательными функциями и их вегетативным 152 Физиология обеспечением) осуществляет высшую интеграцию в деятельности всех систем организма. В конце ХIХ в. В.Я. Данилевский установил, что раздражение лобных долей электрическим током ведет к изменению сердечной деятельности и дыхательного ритма. Позже было показано, что раздражение различных участков коры большого мозга, особенно лобных долей, может изменить деятельность любого органа, имеющего вегетативную иннервацию. Стимуляция двигательной зоны коры большого мозга вызывает такие же изменения деятельности сердечно-сосудистой системы (увеличение минутного объема сердца, усиление кровообращения в мышцах), как и активная мышечная деятельность. Выработка условных рефлексов на изменение интенсивности функционирования внутренних органов также свидетельствует о важной роли коры большого мозга в регуляторной функции внутренних органов (Быков К.М.), интенсивность деятельности которых зависит также и от эмоционального состояния организма. Тонус вегетативных центров. Вегетативные центры, как и соматические, обладают постоянной активностью, называемой тонусом. Он поддерживается рядом факторов, основными из которых являются следующие: • спонтанная активность вегетативных центров, которая особенно характерна для нейронов ретикулярной формации, являющихся составной частью центров ствола мозга (дыхательного, кровообращения и др.); • поток афферентных импульсов в ЦНС от сосудистых рефлексогенных зон, от внутренних органов, от проприорецепторов, от экстерорецепторов; • действие биологически активных веществ и метаболитов непосредственно на клетки центра: СО2, например, возбуждает клетки дыхательного центра, прессорного отдела центра кровообращения. Значение тонуса вегетативных центров иллюстрируется следующими примерами. В результате наличия тонуса сосудосуживающих симпатических нервов гладкие мышцы сосудов постоянно находятся в состоянии некоторого сокращения, поэтому уменьшение тонуса сосудосуживающих нервов ведет к расширению сосудов и увеличению кровотока в них, к снижению АД. При системном повышении Глава 5. Вегетативная нервная система 153 тонуса сосудов АД повышается. Ярко выражен тонус блуждающего нерва для сердца. Поскольку этот нерв оказывает тормозящее влияние на деятельность сердца, он постоянно сдерживает частоту сердечных сокращений. Уменьшение тонуса блуждающего нерва сопровождается увеличением частоты сердцебиений. Таким образом, при наличии тонуса один и тот же центр с помощью одних и тех же эфферентных нервных волокон может вызвать двоякий эффект в деятельности органа и обеспечивать изменение интенсивности его деятельности согласно потребностям организма. Трофическое действие нервной системы. В экспериментах на собаках И.П. Павлов обнаружил веточку симпатического нерва, идущую к сердцу, раздражение которой вызывает усиление сердечных сокращений без изменения их частоты (усиливающий нерв Павлова). Впоследствии было показано, что раздражение симпатического нерва действительно усиливает в сердце обменные процессы. Развивая идею И.П. Павлова, Л.А. Орбели и А.Г. Гинецинский в 20-х гг. XX в. обнаружили усиление сокращений утомленной скелетной мышцы при раздражении идущего к ней симпатического нерва (феномен Орбели–Гинецинского, рис. 5.8). Усиление сокращений утомленной мышцы в опыте Орбели–Гинецинского связано с активацией в ней обменных (трофических) процессов под влиянием норадреналина. Раздражение симпатических нервов не только улучшает функциональные характеристики скелетных мышц, 1 2 3 Рис. 5.8. Повышение работоспособности утомленной изолированной икроножной мышцы лягушки (1) при раздражении симпатических волокон (2) по Орбели–Гинецинскому. Сокращения утомляемой мышцы вызываются ритмическим (30 импульсов в минуту) раздражением двигательных нервных волокон (3). Моменты раздражения симпатического нерва обозначены поднятием сигнальной линии 154 Физиология но и повышает возбудимость периферических рецепторов и структур ЦНС. Такое действие симпатической нервной системы Л.А. Орбели назвал адаптационно-трофическим. Трофическое действие на ткань присуще всем нервам и осуществляется с помощью химических веществ, в первую очередь с помощью медиаторов, но наиболее ярко оно выражено у симпатической нервной системы. 5.4. Точки действия фармакологических препаратов и системное воздействие 1. Пресинаптическое окончание постганглионарного нервного волокна: а) активация или блокада синтеза медиатора, результат — усиление или ослабление влияния нерва на эффекторные клетки; б) угнетение секреции медиаторов из нервного окончания в синаптическую щель, результат — ослабление деятельности эффекторных клеток; в) угнетение обратного захвата медиатора, результат — усиление действия медиатора на постсинаптическую мембрану. 2. Синаптическая щель (блокада действия фермента, разрушающего медиатор, результат — усиление действия медиатора и соответствующего эффектора). 3. Рецептор постсинаптической мембраны эффекторной клетки (активация или блокада, результат — усиление или ослабление функции эффекторной клетки). 4. Нервное волокно (блокада проведения возбуждения, например действие новокаина). 5. Ионные каналы возбудимых клеток (например, блокада Са-каналов гладких мышц сосудов коринфаром ведет к расширению сосудов и снижению АД у пациента с высоким АД). 6. Секреторные клетки — активация или угнетение выработки секрета (например, угнетение выработки соляной кислоты, при избыточной ее секреции). Врачу следует помнить — введенный в организм препарат (независимо от способа его введения) попадает в кровь и, циркулируя по сосудам всего организма, действует на все клетки организма. Ответные реакции различных клеток могут быть противоположными. Например, бускопан, введенный в организм пациента с гиперацидным гастритом с целью блокады М-холинорецепторов секреторных клеток и угне- Глава 5. Вегетативная нервная система 155 тения выработки HCl, действует на М-холинорецепторы всех клеток организма, в том числе и в сердце. Блокада М-холинорецепторов сердца ведет к снятию тормозного влияния блуждающего нерва на сердце и резкому ускорению и усилению сердечных сокращений. У пациентов с пороками сердца или гипертонической болезнью это может привести к тяжелым последствиям. Поэтому, прежде чем выписать пациенту рецепт, необходимо взвесить все обстоятельства. При этом необходимо системное воздействие на организм в целом. Следует выделить несколько мероприятий в этом направлении. Интенсивные ежедневные физические нагрузки на все мышцы организма (полезны и через день). Степень их выраженности должна определяться пациентом и здоровым человеком индивидуально согласно своему самочувствию. Продолжительность около часа. Во время физических упражнений активируется деятельность всех систем организма, особенно системы кровообращения и дыхания. Повышается обмен веществ и энергии вследствие увеличения окислительных процессов, в кровь выбрасываются гормоны и другие биологически активные вещества (катехоламины, кортикоиды, тиреоидные гормоны, гепарин и др.). В частности, гепарин уменьшает свертываемость крови. Повышенный обмен веществ способствует самообновлению организма вследствие преобладания процессов диссимиляции над процессами ассимиляции. После таких оздоравливающих нагрузок человек ощущает свое тело более легким («мышечная радость», по выражению И.П. Павлова). Физический «прогрев» должен быть до пота. В процессе систематических упражнений число движений должно постепенно возрастать (в легких упражнениях до нескольких десятков) — до усталости. В результате пациент или здоровый человек получит удовольствие. Закончить физическую культуру следует водными процедурами по «вкусу». Затем в течение 15–20 мин сохранить тепло в теле. Физическое воздействие является очень важным дополнением в хирургическом, фармакологическом или гипнотическом лечении. Оно особенно важно для профилактики заболеваний. В заключение сделаем вывод: сохранение здоровья и долголетия в жизни — это тяжелый труд! 156 Физиология 5.5. Характеристика ВНС детей ВНС новорожденных незрелая: небольшой мембранный потенциал нейронов вегетативных ганглиев — 20 мВ (у взрослых — 70–90 мВ), медленное проведение возбуждения, автоматизм симпатических нейронов. Медиатором преганглионарных симпатических нейронов (спинной мозг) является адреноподобное вещество (у взрослых — ацетилхолин, то есть дифференцировка преганглионарных симпатических нейронов несколько отстает от парасимпатических), отмечается поливалентная чувствительность нейронов вегетативных ганглиев (к ацетилхолину, норадреналину); N-холинергические синапсы появляются со 2-й недели жизни; развитие холинергической передачи в ганглиях идет одновременно с процессом миелинизации преганглионарных волокон. В онтогенезе число холинергических синапсов в структурах ВНС постепенно увеличивается. Автоматизм клеток симпатических ганглиев и низкий мембранный потенциал симпатических нейронов новорожденных объясняются функциональными особенностями мембраны нейронов, обладающей высокой проницаемостью для Na+, что приводит также к спонтанной активности этих нейронов. ВНС в первые годы жизни ребенка. Наблюдаются также повышенная возбудимость, непостоянство вегетативных реакций, значительная их выраженность и легкая генерализация возбуждения. Поэтому у детей, особенно грудного возраста, наблюдается неустойчивость показателей вегетативных функций, например частоты дыхания, пульса. Устойчивость вегетативных показателей начинает формироваться на 2-м году жизни ребенка. В постнатальном онтогенезе в метасимпатической нервной системе сначала выявляются холинергические, серотонинергические, неадренергические и нехолинергические тормозные нейроны, затем — адренергические. Парасимпатический отдел начинает включаться в рефлекторные реакции сердца с 3-го месяца жизни. К 3 годам тонус блуждающего нерва уже выражен, о чем свидетельствует появление дыхательной аритмии. Однако уже в период новорожденности может наблюдаться глазосердечный рефлекс Данини–Ашнера. Описаны Глава 5. Вегетативная нервная система 157 случаи прекращения деятельности сердца при введении носоглоточных тампонов недоношенным детям. Максимальное замедление пульса от 150 до 30 в минуту было отмечено у них при надавливании на передний родничок. Брадикардия отмечалась у недоношенных детей при зондировании для питания, при икоте, зевании, дефекации. У физиологически зрелых детей увеличение степени выраженности тонуса блуждающего нерва в начале его формирования сочетается с увеличением степени тонического возбуждения сосудодвигательного центра. В регуляцию желудочно-кишечного тракта также сначала включается парасимпатическая нервная система, симпатическая регуляция начинает осуществляться в период отнятия от груди. Таким образом, в онтогенезе созревание парасимпатической нервной системы опережает созревание симпатической. Сроки созревания серотонинергической нервной системы не изучены. Механизм формирования тонуса. В формировании тонуса блуждающего нерва важную роль играет афферентная импульсация от различных рефлексогенных зон, в том числе и от проприорецепторов. Об этом, в частности, свидетельствует тот факт, что недостаточная двигательная активность детей сопровождается недостаточной степенью выраженности тонуса блуждающего нерва. Важное значение в становлении тонуса блуждающего нерва играет импульсация от баро- и хеморецепторов сосудистых рефлексогенных зон. Созревание центральных и периферических отделов ВНС и соматической нервной системы ведет к становлению тонуса всех отделов ЦНС, в том числе симпатических и парасимпатических центров. Следует, однако, отметить, что тонус симпатических центров для сердца не выражен и у взрослого человека. Афферентация от периферических отделов слуховой и зрительной систем также способствует развитию тонуса ЦНС. Для оценки степени выраженности тонуса блуждающего нерва в детском возрасте используют глазосердечный рефлекс (Данини–Ашнера) — давление на боковые поверхности глаз в течение 20–60 с вызывает замедление пульса, падение артериального давления, замедление дыхания. Рефлекс проявляется быстро (через 3–5 с) или медленно (через 8–10 с). Эффект считается положительным, если пульс замедляется на 4–12 в минуту, резко положительным — более чем на 12 сокращений в минуту. ГЛАВА 6 Соответствующее количество женских и мужских половых гормонов в организме детей обеспечивает формирование тела по женскому или мужскому типу подобно деятельности скульптора в его творчестве. Гормональная регуляция функций организма Гормоны (от греч. hormao — приводить в движение, возбуждать) — биологически активные вещества, вырабатываемые эндокринными железами либо отдельными клетками или группами клеток (например, в поджелудочной железе, желудочно-кишечном тракте). 6.1. Характеристика гормонов и регуляция их выработки Основные положения. Эндокринными называют железы, которые не имеют выводных протоков и высвобождают свой секрет (инкрет, гормон) путем экзоцитоза непосредственно в межклеточное пространство, оттуда они попадают в кровь. Гормоны вырабатываются также нервными клетками (нейрокриния) — это нейрогормоны, например гормоны гипоталамуса (либерины и статины), регулирующие функцию гипофиза. Биологически активные вещества вырабатываются также неспециализированными клетками — это тканевые гормоны (гормоны местного действия, паракринные факторы, парагормоны). Гормоны участвуют в регуляции всех функций развивающегося и зрелого организма, особо важную роль они играют в регуляции обменных процессов (поддерживают гомеостазис организма в целом), определяют рост и развитие детей и подростков. Действие биологически активных веществ непосредственно на соседние клетки, минуя кровь, называют паракринным (паракриния), действие Глава 6. Гормональная регуляция функций организма 159 таких веществ преимущественно на рецепторы выделившей их клетки называют аутокринным (например, медиатором нервного окончания на само окончание — аутокриния). Действие гормонов специфично, гормоны не могут быть заменены другими биологически активными веществами. Например, недостаток йодированных гормонов щитовидной железы сопровождается недоразвитием ЦНС, и никакое другое биологически активное вещество не может заменить гормоны щитовидной железы. Гормоны обладают чрезвычайно большой биологической активностью — их концентрация в крови измеряется пикограммами и микромолями. Классификация гормонов осуществляется по нескольким признакам: • по эффекту различают стимулирующие и тормозящие гормоны; • по локализации рецепторов на клетках-мишенях гормоны подразделяют на две группы: действующие посредством мембранных рецепторов (липофобные гормоны) и действующие с помощью внутриклеточных рецепторов (липофильные гормоны); • по месту действия гормоны также делят на две группы: – эффекторные гормоны, действующие на клеткиэффекторы (например, инсулин, регулирующий обмен веществ в организме, увеличивает синтез гликогена в клетках печени, увеличивает транспорт глюкозы и других веществ через клеточную мембрану, повышает интенсивность синтеза белка); – тропные гормоны (тропины), действующие на другие эндокринные железы и регулирующие их функции (например, АКТГ регулирует выработку гормонов корой надпочечников, а ТТГ — функцию щитовидной железы); • по химической природе гормоны делят на три группы: – белки и полипептиды (гормоны гипоталамуса, гипофиза, поджелудочной железы и тимуса); – производные аминокислот (гормоны щитовидной железы, эпифиза, катехоламины); – стероиды (кортикоиды, половые гормоны). 160 Физиология Механизм действия гормонов. Для каждого гормона в клетках-мишенях имеется рецептор. Влияние любого гормона на клетку-мишень начинается с взаимодействия его с рецептором и реализуется с помощью биохимических реакций. Существуют два основных варианта действия гормона на клетку. Если клеточная мембрана непроницаема для гормона, то рецепторы расположены в самой мембране (для белково-полипептидных гормонов гипоталамуса, гипофиза, поджелудочной железы, а также катехоламинов мозгового слоя надпочечников, которые водорастворимы, но нерастворимы в липидах). Взаимодействие этих гормонов с мембранным рецептором активирует внутриклеточные процессы, ведущие к образованию вторых посредников, реализующих свое действие в основном через ядерный аппарат клетки. Например, аденилилциклаза — циклический аденозинмонофосфат (цАМФ). Другие гормоны проникают через клеточную мембрану (стероидные гормоны — кортикоиды и половые гормоны, а также гормоны щитовидной железы, растворимые в липидах), поэтому они взаимодействуют с внутриклеточными рецепторами. В обоих случаях запускаются метаболические процессы. В крови гормоны находятся в двух формах: свободной — готовой к действию (активная форма) и связанной с белками (неактивная форма) — это своеобразное депо гормонов. Период полураспада гормонов обычно около часа. Действие гормонов прекращается с помощью тканевых ферментов и ферментов самих эндокринных желез, печени, почек. Понятие о тканевых гормонах. Они представляют собой биогенные амины (гистамин, серотонин), простагландины, кинины (калликреин, брадикинин) и др. Занимают промежуточное положение между гормонами и метаболитами как гуморальные факторы регуляции. Тканевые гормоны образуются неспециализированными клетками, но действуют они посредством специализированных клеточных рецепторов. Паракринные вещества оказывают свое регулирующее влияние на клетки тканей посредством изменения их биофизических свойств (проницаемости мембран, их возбудимости), изменения интенсивности обмен- Глава 6. Гормональная регуляция функций организма 161 ных процессов, чувствительности клеточных рецепторов, образования вторых посредников. В результате этого изменяется чувствительность клеток к нервным и гуморальным влияниям. Поэтому тканевые гормоны называют модуляторами регуляторных сигналов: они оказывают модулирующее влияние. Регуляция выработки гормонов осуществляется на четырех уровнях. 1. Регуляция с помощью ЦНС. Высшим уровнем регуляции выработки гормонов является лимбическая система и новая кора, при этом особо важную роль играет гипоталамус (рис. 6.1), как одна из структур лимбической системы. Гипоталамус с помощью своих нейрогормонов (либерины-стимуляторы, статины-ингибиторы) регулирует выработку тропных гормонов гипофизом. Симпатические нервные волокна, идущие от верхнего шейного ганглия, усиливают выработку тропных гормонов гипофизом, пара- Рис. 6.1. Регуляторные взаимодействия гипоталамуса и эндокринных желез по выработке гормонов и регуляции функций — стимуляция; — угнетение организма: 162 Физиология симпатические нервные волокна, идущие от языкоглоточного нерва, напротив, угнетают их синтез. 2. Регуляция деятельности эндокринных желез с помощью гипофиза осуществляется посредством тропных гормонов по принципу отрицательной обратной связи. Если уровень эффекторного гормона низкий, выработка тропного гормона гипофизом возрастает, что ведет к увеличению выработки соответствующего эффекторного гормона. Если уровень последнего высок, развиваются противоположные реакции. 3. Регуляция на эффекторном уровне (эндокринные железы — параметры регулируемого показателя) осуществляется также по принципу обратной отрицательной связи. Если, например, концентрация глюкозы в крови снижена, выработка инсулина уменьшается, а выработка глюкагона возрастает, уровень глюкозы нормализуется. 4. Внутриклеточная саморегуляция деятельности клеток эндокринных желез по выработке гормонов генетически запрограммирована. При этом цепь реакций по выработке секрета может стимулироваться или тормозиться по принципу обратной отрицательной связи продуктами последующих реакций: недостаток синтезируемых элементов стимулирует их образование, избыток — угнетает. 6.2. Гормоны гипофиза Гипофиз состоит из передней доли (аденогипофиза) и задней доли (нейрогипофиза). В передней доле гипофиза вырабатываются тропные гормоны (тиреотропный гормон — тиреотропин, адренокортикотропный гормон — кортикотропин и гонадотропные гормоны — гонадотропины) и эффекторные гормоны (гормон роста — соматотропин и пролактин). В задней доле гипофиза гормоны не вырабатываются. Функция тропных гормонов аденогипофиза заключается в регуляции выработки эффекторных гормонов эндокринными железами (см. рис. 6.1). Адренокортикотропный гормон (кортикотропин, АКТГ) является стимулятором образования глюкокортикоидов в пучковой зоне коры надпочечников. В меньшей степени Глава 6. Гормональная регуляция функций организма 163 выражено влияние АКТГ на клубочковую и сетчатую зоны. Кортикотропин, стимулируя образование глюкокортикоидов, приводит к тем же эффектам, что и глюкокортикоиды. Нарушение выработки АКТГ ведет к тяжелым заболеваниям. В частности, при избытке АКТГ развивается болезнь Иценко–Кушинга (вследствие чрезмерной стимуляции функции надпочечников). Ее характерными признаками являются ожирение, полнокровие, акроцианоз, тенденция к появлению пурпуры, багровые полосы на животе, гирсутизм, дистрофия половой системы, гипертония, остеопороз. Тиреотропный гормон (ТТГ, тиреотропин) стимулирует функцию щитовидной железы, действие ТТГ на белковый, жировой, углеводный, минеральный и водный обмен осуществляется через гормоны щитовидной железы (см. раздел 6.3). К гонадотропным гормонам (гонадотропинам) относятся фолликулостимулирующий (ФСГ, фоллитропин) и лютеинизирующий (ЛГ, лютропин), имеющийся у мужчин и женщин (см. раздел 6.7). Снижение выработки кортикотропных гормонов сопровождается быстрой утомляемостью и слабостью мышц вследствие угнетения секреции эффекторных гормонов. Механизм регуляторных взаимодействий представлен на рис. 6.2. Функции эффекторных гормонов аденогипофиза. Гормон роста (ГР, соматотропин, СТГ) назван так потому, что стимулирует рост организма. СТГ угнетает окисление углеводов в тканях. Это действие в значительной мере опосредовано и через поджелудочную железу. СТГ вызывает задержку в организме фосфора, натрия, калия, кальция. Одновременно увеличивается распад жира, о чем свидетельствует нарастание в крови количества свободных жирных кислот. Гормон роста увеличивает выработку лимфоцитов. Пролактин стимулирует образование и выделение молока (лактация) у матери, чему способствуют эстрогены и прогестерон (эстрогены вызывают рост протоков молочной железы, прогестерон — развитие ее альвеол). Пролактин обладает также лютеотропным действием, т.е. способствует продолжительному функционированию желтого 164 Физиология Рис. 6.2. Регуляторные взаимодействия гипоталамуса и аденогипофиза по выработке эффекторных гормонов и регуляции функций организма: — стимуляция; — угнетение тела и образованию им гормона прогестерона. Секреция пролактина увеличивается во время акта сосания (рефлекторный механизм). В задней доле гипофиза (нейрогипофизе) гормоны не вырабатываются. В ней активируются окситоцин и вазопрессин (рис. 6.3), которые вырабатываются в неактивном состоянии нейронами супраоптического и паравентрикулярного ядер гипоталамуса и накапливаются в клетках задней доли гипофиза. Нейросекреты (нейрогормоны) транспортируются по аксонам нейронов этих ядер (по так называемому гипоталамо-гипофизарному тракту). Окситоцин стимулирует сокращение матки и миоэпителиальных клеток, способствующих выделению молока из молочных желез (секреция регулируется пролактином). Окситоцин во время беременности не действует на матку, т.к. под влиянием гормона желтого тела прогестерона она становится нечувствительной к окситоцину. Если механически раздражать шейку матки, происходит рефлекторное высвобождение окситоцина, чему способствует также акт сосания. Главная функция вазопрессина (антидиуретический гормон, АДГ) связана с его способностью усиливать ре- Глава 6. Гормональная регуляция функций организма 165 Рис. 6.3. Регуляторные взаимодействия задней зоны гипоталамуса и гипофиза по выработке и активации гормонов и регуляции функций организма: — стимуляция; — угнетение абсорбцию воды в собирательных трубках почек — это антидиуретическое действие гормона. При недостаточности нейрогипофиза развивается синдром несахарного диабета, при котором с мочой теряется огромное количество воды (до 15 л/сут), поскольку ее реабсорбция в собирательных трубках снижается или прекращается. Вследствие невыносимой жажды больные постоянно пьют воду. Другая функция вазопрессина связана с влиянием его на гладкие мышцы артериол. Но второе название АДГ, вазопрессин, не полностью соответствует свойству суживать сосуды. Дело в том, что в нормальных физиологических концентрациях он сосудосуживающим действием не обладает. Сужение сосудов под влиянием АДГ может наблюдаться при экзогенном введении или большом выбросе его гипофизом в случае значительной кровопотери. 6.3. Функции гормонов щитовидной железы Щитовидная железа вырабатывает йодированные гормоны тироксин (Т4) и трийодтиронин (Т3) и нейодированный гормон тиреокальцитонин. 166 Физиология Тироксин и трийодтиронин. В клетках всех тканей организма под влиянием тиреоидных гормонов синтезируется большое количество ферментов, структурных и транспортных белков, что стимулирует обмен белков, жиров и углеводов, рост и развитие организма, созревание ЦНС. При гиперфункции щитовидной железы возникают нервозность, тревожность, раздражительность. Отмечается высокая утомляемость, мышечный тремор и другие симптомы, свидетельствующие о нарушении функции ЦНС, отмечаются также нарушения сна, памяти, эмоциональная неустойчивость и другие патологические изменения высшей нервной деятельности. При гипофункции у детей развивается кретинизм, у взрослых снижается работоспособность. Регуляция функции щитовидной железы. Возбуждение ретикулярной формации ствола мозга ведет к повышению функциональной активности щитовидной железы, за счет усиления выработки тиреолиберина гипоталамусом. Возбуждение постганглионарных симпатических нервных волокон, идущих от средних шейных симпатических ганглиев, приводит к повышению активности щитовидной железы, а стимуляция блуждающего нерва снижает гормонообразовательную функцию железы. Тиреокальцитонин — нейодированный гормон, принимает участие в фосфорно-кальциевом обмене, выступая антагонистом паратгормона. Осуществляет защиту организма от избыточного поступления кальция, уменьшая реабсорбцию кальция в канальцах почек, всасывание кальция из кишечника и увеличивая фиксацию кальция в костной ткани. Выработка тиреокальцитонина регулируется уровнем кальция в крови и изменениями секреции гастрина при приеме пищи, богатой кальцием (коровье молоко). Повышение концентрации Са 2+ в крови быстро приводит к увеличению секреции тиреокальцитонина путем экзоцитоза, что сопровождается повышением его содержания в крови. Физическая работоспособность уменьшается при снижении функции щитовидной железы. Глава 6. Гормональная регуляция функций организма 167 6.4. Гормоны паращитовидных желез, эпифиза и тимуса. Кальцитриол Паращитовидные железы расположены на задней поверхности щитовидной железы или погружены внутрь нее. Гормон паращитовидных желез (паратгормон, паратирин) увеличивает содержание Са 2+ в крови, обеспечивая его всасывание в кишечнике, реабсорбцию в канальцах почки, мобилизацию из костей и активацию остеокластов костной ткани, тормозит реабсорбцию фосфатов канальцами почек и способствует выведению фосфора с мочой, т.е. оказывает противоположное тиреокальцитонину влияние, участвуя тем самым в обеспечении оптимального содержания Са 2+ в организме и перемоделирования костной ткани. Кости имеют огромный запас кальция, поэтому даже значительное выведение его из костей не нарушает их структуру. Активация паращитовидных желез возникает через несколько минут в ответ на снижение йодированного кальция в крови. Увеличение выброса паратгормона в ответ на этот стимул способствует быстрой мобилизации кальция из костной ткани и включению более медленных механизмов, повышению реабсорбции кальция в почках и увеличению всасывания кальция из кишечника. Кальциевое голодание или нарушение всасывания витамина D, лежащее в основе рахита у детей, всегда сопровождается гиперплазией паращитовидных желез и функциональными проявлениями гиперпаратиреоидизма, но все эти изменения суть проявление нормальной регуляторной реакции. При повышенной концентрации Са 2+ активность и размер паратиреоидных желез уменьшаются, а его содержание в крови нормализуется (отрицательная обратная связь). Кальцитриол — производное витамина D3. В печени образуется кальцидиол, который в почках превращается в кальцитриол под влиянием паратгормона. В тонкой кишке гормон стимулирует всасывание Ca 2+ и фосфатов, в костях — мобилизацию Ca2+. Таким образом, паратгормон и витамин D3 являются синергистами в отношении мобилизации Ca2+ из костей и повышения его концентрации в крови. 168 Физиология Гормон эпифиза (мелатонин) — это производное аминокислоты триптофана, промежуточный продукт синтеза мелатонина — серотонин. Мелатонин участвует в регуляции пигментного обмена за счет обесцвечивания меланофоров кожи. Основной эффект мелатонина — торможение секреции гонадотропинов, что сдерживает половое созревание. Поэтому при гипофункции эпифиза наступает раннее половое созревание, а при гиперфункции — явления гипогенитализма и ожирения. Вилочковая железа (тимус) вырабатывает несколько гормонов: тимозин, тимусовый гуморальный фактор, тимопоэтины, гомеостатический тимусовый гормон, стимулирующий образование антител и, по-видимому, рост развивающегося организма. После полового созревания происходит быстрая атрофия тимуса под влиянием главным образом эстрогенов и в меньшей степени — андрогенов. В тимусе происходит дифференцировка лимфоидных стволовых клеток в Т-лимфоциты. В случае недоразвития тимуса у детей нарушаются клеточные иммунные реакции, в результате чего возникают гнойные инфекции. 6.5. Гормоны поджелудочной железы Вырабатываются гормоны островковым аппаратом поджелудочной железы, локализующимся преимущественно в ее хвостовой части. Основную массу островков Лангерганса (около 60%) составляют β-клетки, которые секретируют инсулин. Примерно 25% общего количества клеток островкового аппарата приходится на долю α-клеток, секретирующих глюкагон. Дельта-клетки (δ-клетки), которых примерно 10%, секретируют соматостатин. Клетки РР, которых немного в железе, секретируют гормон неясной функции, называемый панкреатическим полипептидом, G-клетки (их количество менее 5%) вместе с РР-клетками продуцируют гастрин. Главная функция гормонов поджелудочной железы — регуляция обмена углеводов (поддерживает оптимальный уровень глюкозы в крови). Функции инсулина. В крови инсулин циркулирует в основном в свободном виде, период полужизни — примерно 6 мин. Он принимает участие в регуляции углеводного, бел- Глава 6. Гормональная регуляция функций организма 169 кового и жирового обмена. Стимулирует гликогенез (особенно в печени) и увеличивает число переносчиков для глюкозы и аминокислот. Стимулирует синтез белков на фоне угнетения их распада, а также торможение глюконеогенеза. Инсулин способствует образованию жирных кислот (липогенез) из продуктов углеводного обмена, а также тормозит мобилизацию жира из жировой ткани (липолиз). Главным регулятором секреции инсулина является концентрация глюкозы в крови (рис. 6.4). Гипергликемия ведет к увеличению его секреции, гипогликемия — к уменьшению. Глюкоза стимулирует секрецию инсулина и за счет непосредственного воздействия на β-клетки островков Лангерганса, при этом ионы Са2+ запускают высвобождение инсулина. Нервная регуляция осуществляется рефлекторно при действии глюкозы на хеморецепторы каротидного синуса и возбуждении их, а также под действием глюкозы на глюкорецепторы гипоталамуса. Симпатический нерв тормозит образование инсулина, увеличивая содержание глюкозы в крови. Блуждающий нерв усиливает образование инсулина, что ведет к снижению уровня глюкозы в крови в результате увеличения его потребления клетками и усиления гликогенеза. Белковое питание мг/дл 120 100 80 120 100 80 мг/дл Углеводное питание Глюкоза 120 80 80 Инсулин 40 40 0 0 200 200 пг/мл пг/мл Глюкагон 100 А мкг/мл мкг/мл 120 100 0 1 2 Часы 3 4 0 1 2 Часы 3 4 Б Рис. 6.4. Секреция инсулина и глюкагона после приема глюкозы (А) и белковой пищи (Б) (по K. Voigt, 2004) 170 Физиология Стимулируют образование инсулина гормоны соматотропин (посредством соматомединов), секретин и холецистокинин-панкреозимин, простагландин Е за счет повышения аденилатциклазной активности β-клеток поджелудочной железы. Соматостатин, напротив, тормозит образование инсулина, действуя непосредственно на β-клетки островков Лангерганса. Глюкагон — это антагонист инсулина, он увеличивает содержание глюкозы в крови с помощью гликогенолиза в печени. Глюкагон и инсулин поддерживают оптимальную концентрацию глюкозы в крови и снабжение ею клеток организма. При повышении содержания глюкозы в крови происходит торможение образования и секреции глюкагона, а при понижении — увеличение (рис. 6.5). Высокие концентрации аминокислот в крови стимулируют секрецию инсулина и глюкагона. Взаимодействие инсулина и глюкагона стабилизирует концентрацию глюкозы в крови, глюкагон при этом стимулирует глюконеогенез и Соматотропин Соматостатин Соматомедин α-клетки островкового аппарата Глюкагон Гликогенолиз Гипергликемия (более 6,5 ммоль/л) Гипогликемия (менее 3,5 ммоль/л) Активирующее влияние Тормозное влияние Рис. 6.5. Регуляция секреции глюкагона (по В.Ф. Киричуку, с изменениями) Глава 6. Гормональная регуляция функций организма 171 гликогенез, защищает организм от снижения содержания глюкозы в крови в результате действия инсулина. Соматостатин, секретируемый δ-клетками островкового аппарата поджелудочной железы, тормозит образование и секрецию глюкагона и инсулина. Гормон роста (соматотропин) посредством соматомединов повышает активность α-клеток, и они продуцируют больше глюкагона. Таким образом, только инсулин снижает уровень глюкозы в крови, катехоламины, глюкагон, тироксин и гормон роста, увеличивают содержание глюкозы в крови. Соматостатин (третий из основных гормонов поджелудочной железы) участвует также в регуляции выработки других гормонов (см. раздел 6.3). При ослаблении функции поджелудочной железы физическая работоспособность снижается. 6.6. Гормоны надпочечников Надпочечники имеют два слоя — корковый и мозговой. Корковый слой состоит из клубочковой, пучковой и сетчатой зон. Мозговой слой надпочечников представляет собой хромаффинную ткань, с двумя видами клеток, образующих адреналин и норадреналин. Удаление обоих надпочечников в эксперименте ведет к смерти животных в течение 1–2 недель в результате выключения функции коркового слоя надпочечников. Гормоны коркового слоя надпочечников: • глюкокортикоиды, основным из которых является кортизол (гидрокортизон); • минералокортикоиды, главный из которых альдостерон; • половые гормоны, в основном мужские (андрогены); • небольшое количество женских (эстрогены, прогестерон). Образуются половые гормоны в сетчатой, минералокортикоиды — в клубочковой зоне. Глюкокортикоиды (образуются в пучковой зоне) участвуют в регуляции обмена углеводов (глюконеогенез — образование глюкозы из аминокислот), повышают отложение гликогена в печени, тормозят утилизацию глюкозы в тка- 172 Физиология нях и при передозировке могут привести к гипергликемии и глюкозурии, вызывают распад тканевого белка, задерживают включение аминокислот в белки, что тормозит формирование грануляций, последующее образование рубца, заживление ран. Оказывают тормозное влияние на функцию иммунной системы, подавляя продукцию лимфоцитов, особенно Т-лимфоцитов, угнетают развитие воспалительных процессов, что связано в основном с понижением проницаемости стенок сосудов. Гидрокортизон подавляет также синтез антител и тормозит реакцию взаимодействия чужеродных белков (антигенов) с антителами. При недостаточности глюкокортикоидов снижается сопротивляемость организма вследствие нарушения обмена углеводов и белков, ослабевает умственное и физическое развитие. Регуляция образования глюкокортикоидов осуществляется кортикотропином (см. рис. 6.1). Адреналин усиливает образование глюкокортикоидов. Минералокортикоиды примерно на 90% представлены альдостероном, который усиливает реабсорбцию Na+ в почечных канальцах и секрецию К+. Альдостерон усиливает воспалительные реакции посредством повышения проницаемости капилляров и серозных оболочек. Содержание альдостерона в крови регулируется по принципу обратной отрицательной связи. В частности, снижение Na+ в крови стимулирует выработку альдостерона, повышение — тормозит, калий оказывает противоположное влияние, что ведет к нормализации содержания Na+ и К+ в организме (иначе — альдостерон задерживает Na+ в организме, способствует выделению К+). Тормозит секрецию альдостерона увеличение объема тканевой жидкости и плазмы крови, что сопровождается увеличением выделения Na+ и связанной с ним воды с мочой. Гормоны мозгового слоя надпочечников — адреналин (80–90%), норадреналин (10–20%) и дофамин (1–2%). Эти гормоны образуются в хромаффинных клетках из аминокислоты тирозина. Адреналин быстро разрушается под действием фермента моноаминооксидазы. Адреналин поступает из надпочечников в кровь постоянно, но при кровопотерях, охлаждении, эмоциях, боли, гипогликемии, мышечной деятельности образование его увеличивается. Глава 6. Гормональная регуляция функций организма 173 Функции адреналина — он ускоряет расщепление гликогена в печени и в мышцах, в результате чего повышается содержание глюкозы в крови. В целом влияние адреналина на внутренние органы подобно таковому симпатической нервной системы. Возбуждение симпатической нервной системы увеличивает выброс катехоламинов надпочечниками, так же действует гипогликемия. Физическая работоспособность снижается при уменьшении выработки гормонов надпочечниками. 6.7. Половые гормоны Особенность половых желез (семенники — у мужчин, яичники — у женщин) — это выработка половых клеток (у мужчин — сперматозоиды, у женщин — яйцеклетки) и половых гормонов. Мужские половые гормоны (андрогены — тестостерон и андростерон) вырабатываются в интерстициальных клетках семенников. Они необходимы для созревания мужских половых клеток — сперматозоидов, для проявления полового инстинкта и связанного с ним полового поведения. При их отсутствии не образуются подвижные зрелые сперматозоиды. Гормоны повышают основной обмен веществ. Уменьшают содержание жира в организме, увеличивают образование белка в различных тканях, особенно в мышцах. Интенсивные физические упражнения приводят к повышению продукции андрогенов, которые способствуют развитию мышечной гипертрофии. Применение андрогенов в спорте недопустимо, т.к. это приводит к серьезным осложнениям: поражение печени (цирроз, рак и др.), у женщин страдает детородная функция, нарушается менструальный цикл. Женские половые гормоны (эстрогены — эстрон, эстрадиол) образуются в оболочке фолликула яичника. В желтом теле яичника, которое развивается на месте лопнувшего фолликула, вырабатываются прогестины, в частности гормон прогестерон. Эстрогены стимулируют рост половых органов, способствуют развитию вторичных женских половых признаков и проявлению половых рефлексов, стиму- 174 Физиология лируют сокращения матки, повышают чувствительность к окситоцину. Прогестерон обеспечивает нормальное протекание беременности. Под его воздействием разрастается эндометрий матки, что способствует имплантации оплодотворенной яйцеклетки, развитию децидуальной ткани вокруг имплантированной яйцеклетки и поддерживает нормальное течение беременности. Половые гормоны (особенно в период полового созревания) стимулируют рост тела и половых органов, формируют строение тела согласно полу, увеличивают основной обмен, ускоряют созревание ЦНС. Эритропоэз стимулируется андрогенами, эстрогенами подавляется. Уровень половых гормонов в крови определяется гипоталамусом, до определенного возраста вырабатывается мало либеринов и, соответственно, гонадотропных гормонов. Регуляция образования половых гормонов отражена на рис. 6.6. Лютропин стимулирует образование андрогенов в семенниках, эстрогенов и прогестерона в яичниках. Пролактин усиливает синтез прогестерона в желтом теле. МеОвуляция 0,2 1 12–14 Дни Тестостерон Тестостерон, нг/мл 0,4 Эстрадиол 500 28 Время жизни 10 СТГ 5 СТГ, нг/мл 50 ЛГ Месяцы Эстрадиол, нг/мл Менструация ЛГ, мЕД/мл 100 100 0 20 40 Годы 60 80 Рис. 6.6. Динамика секреции половых гормонов: у женщин — в зависимости от менструального цикла; у мужчин — от возраста (по K. Voigt, 2004) Глава 6. Гормональная регуляция функций организма 175 латонин (гормон эпифиза) тормозит деятельность половых желез. При гипофункции эпифиза наступает раннее половое созревание, а при гиперфункции — явления гипогенитализма и ожирения. Корковые влияния реализуются посредством гипоталамуса–гипофиза–гонадотропных гормонов. При сильных отрицательных эмоциях у женщин может произойти нарушение полового цикла. Нарушение образования половых гормонов — одна из причин остеопороза, что наблюдается в пожилом возрасте и при патологиях. Температура среды, в которой находятся гонады (мужские и женские), оказывает весьма сильное влияние на выработку половых гормонов: в условиях низкой температуры секретируются в основном мужские половые гормоны (яички расположены в мошонке), в условиях высокой температуры вырабатываются преимущественно женские половые гормоны (яичники расположены в брюшной полости). Поэтому, если у мальчика яички не опустились в мошонку (подобная патология иногда встречается), то операцию необходимо проводить в раннем онтогенезе, иначе яички нормально функционировать не будут. ГЛАВА 7 Медицине нельзя научить, медицине можно только научиться. М.Я. Мудров Система крови Система крови — это совокупность органов кроветворения, периферической крови и органов кроверазрушения (Ланг Г.Ф.). 7.1. Кровь как внутренняя среда организма Внутренняя среда организма — это совокупность жидкостей организма, включающая кровь, лимфу, тканевую и цереброспинальную жидкости. Из нее ткани получают все необходимые вещества для своей жизнедеятельности и отдают в нее продукты обмена веществ (метаболиты). Система крови является составной частью иммунной системы организма. Иммунитет (от лат. immunitas — свобода от чего-либо) — совокупность свойств и механизмов, обеспечивающих постоянство состава организма и его защиты от инфекционных и других чужеродных для него агентов и от веществ и клеток эндогенного (мутагенного) происхождения. Согласно одной из классификаций различают неспецифический и специфический иммунитет. Неспецифическими называют механизмы защиты против любых чужеродных веществ — это неспецифическая резистентность организма (врожденный иммунитет). Специфические механизмы защиты — приобретенный активный иммунитет (постинфекционный и поствакцинальный) и пассивный (введение антител) — направлены против конкретных чужеродных агентов и обеспечивают им специфическое противодействие. Структурно-функциональная характеристика крови. Кровь — это жидкая ткань организма. Она состоит из формен- Глава 7. Система крови 177 ных элементов (40–45%) — эритроцитов, лейкоцитов, тромбоцитов и плазмы (жидкая часть крови — 55–60%). Процент форменных элементов называют гематокритным числом (в норме 40–45). Его определяют при помощи прибора гематокрита. Количество крови в организме человека составляет 5–9% от массы тела (4,5–6 л у человека массой около 70 кг). В состоянии покоя до 45–50% всей массы крови находится в кровяных депо (селезенке, подкожном сосудистом сплетении печени и легких). Кровь представляет собой суспензию — взвесь (от лат. suspensio — подвешивание). Это объясняется тем, что форменные элементы, белки и эндотелий сосудов имеют отрицательный заряд снаружи и отталкиваются друг от друга. Функции крови: • транспортная — разносит по организму различные вещества, благодаря чему выполняются следующие функции: – дыхательная; – питательная; – экскреторная; – регуляторная (вследствие переноса биологически активных веществ); • участие в процессах терморегуляции; • защитная функция; • стабилизация рН внутренней среды организма. Плазма крови (рис. 7.1) на 90% состоит из воды, содержит около 1% неорганических и около 9% органических веществ. Неорганические вещества включают минеральные соли. Они создают осмотическое давление, участвуют в стабилизации рН крови, в процессе свертывания крови, в создании электрических зарядов клеток, в транспорте воды — вода движется в сторону более высокого осмотического давления. Органические вещества плазмы. Белки (67–75 г/л), среди них альбумины (37–41 г/л), глобулины (30–34 г/л), фибриноген (3,0–3,3 г/л). Значение белков: • участвуют в процессе свертывания крови (фибриноген и другие плазменные факторы свертывания крови); • создают коллоидно-осмотическое (онкотическое) давление (25–30 мм рт. ст.); 178 Физиология • стабилизируют рН крови (белковый буфер); • участвуют в реакциях иммунной системы; • выполняют транспортную функцию. Азотсодержащие вещества небелковой природы — это промежуточные продукты обмена белка: мочевина, аминокислоты, мочевая кислота. Они формируют остаточный азот. Безазотистые органические вещества: глюкоза (4,4– 6,6 ммоль/л), липиды (фосфолипиды, жирные кислоты, лецитин, холестерин — общее количество 3,5–5,2 мМ/л), молочная и пировиноградная кислоты. Биологически активные вещества: витамины, гормоны, ферменты, простагландины и др. Физико-химические показатели крови. Осмотическое давление плазмы крови в норме составляет около 7,6 атмосферы — это фактор, участвующий в транспорте веществ в организме и формировании тургора клеток. Раствор, имеющий одинаковое осмотическое давление с кровью, называется изотоническим (0,85–0,9% раствор NaCl). Раствор, осмотическое давление которого выше осмотического давления крови, называют гипертоническим, раствор с более низким давлением — гипотоническим. Часть осмотического давления, создаваемого белками, называется онкотическим давлением (25–30 мм рт. ст.). Кислотно-основное состояние (КОС) организма характеризуется рН крови (7,35–7,45). Этот показатель определяет активность ферментов (скорость метаболических реакций). Сдвиг рН крови даже на 0,1 за указанные границы ведет к нарушению функций клеток; сдвиг на 0,3 может вызвать коматозное состояние, а на 0,4 — несовместим с жизнью. Постоянство рН организма регулируется выделительной систеПлазма крови Вода — 90% Белки: альбумины, глобулины, фибриноген Органические вещества — 9% Азотсодержащие вещества: мочевина, мочевая кислота, аминокислоты Неорганические вещества — 1% Безазотистые вещества: глюкоза, липиды, фосфолипиды, молочная, пировиноградная и жирные киcлоты Биологически активные вещества: ферменты, гормоны, витамины, медиаторы Рис. 7.1. Состав плазмы крови Катионы: Na+, K+, Ca2+, Mg2+ Анионы: Cl–, HCO3–, фосфаты, сульфаты Глава 7. Система крови 179 мой, стабилизируется буферными системами плазмы крови, которые способны соединяться с ионами Н+ или ОН, образующимися в организме. Основные буферные системы крови — бикарбонатная (NaHCO3/H2CO3), фосфатная (Na2HPO4/ NaH2PO4) и белковая. В эритроцитах действует гемоглобиновый буфер (ННb/ НbO2) (ион Н+ связывается Hb). Реологические свойства крови определяются ее вязкостью, которая зависит в основном от количества форменных элементов и белка, с увеличением их содержания вязкость возрастает. В норме вязкость крови 4–5 усл. ед. (вязкость воды условно 1 ед.). Кроме того, агрегация эритроцитов увеличивает вязкость крови. 7.2. Физиология эритроцитов Основные показатели. Структурные элементы и форма эритроцитов показаны на рис. 7.2. В зависимости от размеров различают нормоциты, микроциты и макроциты. Эритроциты не имеют ядра, митохондрий, белоксинтезирующей системы, для них характерна гомогенная цитоплазма. В эритроцитах содержится до 60% воды (в других клетках 80% и более) и 40% сухого остатка, который на 90–95% состоит из гемоглобина. Продолжительность жизни эритроцита в кровяном русле — около 120 дней. Разрушение старых эритроцитов происходит в селезенке (кладбище эритроцитов) и в печени. Количество эритроцитов у мужчин колеблется в пределах 4,5–5,5 × 1012/л, у женщин — 3,7–4,7 × 1012/л. Количество эритроцитов в крови изменяется как в норме, так и при па- Рис. 7.2. Зрелые эритроциты 180 Физиология тологии. Эритроцитоз — увеличение количества эритроцитов в периферической крови. Эритропения — уменьшение количества эритроцитов в единице объема крови. Свойства эритроцитов. Эритроциты обладают высокой пластичностью (способностью к обратимой деформации), что облегчает прохождение через капилляры диаметром до 2,5–3 мкм. Свойство пластичности выражено у дискоидных эритроцитов, их размеры составляют 7,2–7,5 мкм. По мере старения эритроцитов их пластичность снижается, они превращаются в сфероциты (имеющие форму шара), вследствие чего эритроциты не могут проходить через капилляры диаметром до 3 мкм и задерживаются в селезенке, где и разрушаются («кладбище» эритроцитов). Пластичность оболочки эритроцитов обеспечивает их осмотическую стойкость. Осмотическое давление в эритроцитах несколько выше, чем в плазме крови, что обеспечивает тургор клеток вследствие поступления воды в эритроциты. Повышенное осмотическое давление — это результат более высокой внутриклеточной концентрации белков по сравнению с плазмой крови. При помещении эритроцитов в гипотонический раствор вода поступает в эритроциты, они набухают, при этом может наступить осмотический гемолиз (разрушение эритроцитов), в результате чего гемоглобин выходит в окружающую среду. При помещении эритроцитов в гипертонический раствор происходит их сморщивание, что связано с выходом воды из эритроцита в среду. Эритроциты способны к агрегации — образованию конгломератов при замедлении движения крови и повышению ее вязкости, что может наблюдаться при патологиях. Вначале агрегация эритроцитов носит обратимый характер, при этом образуются ложные агрегаты или монетные столбики. В случае быстрого восстановления кровотока они распадаются на полноценные клетки. Свойства эритроцитов определяются особенностями их строения. Пластичность эритроцитов — это способность к обратимой деформации при прохождении через узкие извитые капилляры. Эритроциты способны удлиняться при Глава 7. Система крови 181 деформации в 2–3 раза, поэтому дисковидный эритроцит легко проходит через капилляры диаметром 3 мкм. Старые неэластичные эритроциты задерживаются в синусах селезенки и подвергаются деструкции и в конечном счете лизируются. Деформация эритроцитов при прохождении через капилляры усиливает внутриклеточную конвекцию кислорода, дезокси- и оксигемоглобина, что способствует газообмену между эритроцитами и плазмой, между плазмой и интерстицием. Осмотическая стойкость эритроцитов. Осмотическое давление в эритроцитах несколько выше, чем в плазме крови. Оно создается высокой внутриклеточной концентрацией белков по сравнению с плазмой крови. При помещении эритроцитов в 0,30% раствор NaCl они разрушаются (осмотический гемолиз). Это обусловлено тем, что вода поступает в эритроциты до того момента, пока не разрывается мембрана, поэтому гемоглобин выходит в раствор. В умеренно гипотоническом растворе эритроциты приобретают сферическую форму. В гипертоническом растворе происходит сморщивание эритроцитов, что связано с выходом воды из эритроцита. В условиях организма может происходить химический гемолиз (например, действие антималярийных препаратов), биологический (переливание несовместимой группы крови, змеиный яд) и др. Агрегация эритроцитов — способность эритроцитов создавать в цельной крови конгломераты. Движущаяся кровь содержит как одиночные эритроциты, так и агрегаты. Среди агрегатов имеются отдельные цепочки эритроцитов («монетные столбики») и цепочки в виде выростов. Для агрегации эритроцитов необходим фибриноген, адсорбция которого на мембране этих клеток приводит к образованию мостиков между эритроцитами. Прочность образовавшихся агрегатов прямо пропорциональна концентрации фибриногена. Агрегация эритроцитов обратима: агрегаты клеток способны деформироваться и разрушаться при движении крови. Скорость оседания эритроцитов (СОЭ). Удельный вес эритроцитов (1,096) выше, чем плазмы крови (1,027), поэтому эритроциты в пробирке с кровью, лишенной возможности свертываться, медленно оседают на дно. 182 Физиология СОЭ зависит от соотношения альбуминовой и глобулиновой фракций крови, т.е. от белкового коэффициента (БК). В норме БК равен 1,5–1,7. При нормальном белковом коэффициенте СОЭ составляет у мужчин 5–10 мм/час, у женщин — 5–12 мм/час. При воспалениях СОЭ повышается, так как увеличивается количество белков глобулиновой фракции. Механизм увеличения СОЭ. Молекулы белков плазмы крови имеют отрицательный заряд. Причем самый большой заряд у альбуминов (–18 усл. ед.). Их электрические заряды взаимодействуют между собой и с электрическими отрицательными зарядами мембран эритроцитов (–35 мВ), что создает отталкивающую силу. Поскольку отрицательные заряды глобулинов меньше, отталкивающая сила уменьшается, в результате чего эритроциты быстрее оседают. Кроме того, в этих условиях легче создаются агрегаты эритроцитов. Суммарное сопротивление таких агрегатов трению меньше, чем суммарное сопротивление отдельных эритроцитов, и они быстрее оседают. При эритропении СОЭ возрастает (меньше одноименных зарядов), при эритроцитозе — уменьшается (больше одноименных зарядов — больше отталкивающая сила). Метаболизм эритроцитов. В эритроцитах почти полностью выключены окислительные процессы, что весьма экономично в энергетическом отношении. Энергетическое обеспечение эритроцита осуществляется только за счет утилизации глюкозы в результате анаэробного гликолиза. Основные функции эритроцитов: • главная — дыхательная (транспорт О2 и СО2). Эритроциты переносят и другие вещества; • участие в стабилизации КОС крови за счет гемоглобина и наличия фермента карбоангидразы; • участие в процессах свертывания крови (эритроциты заполняют сгусток крови); • участие в иммунных реакциях организма, в реакциях преципитации, лизиса, опсонизации; • дезинтоксикация веществ обеспечивается адсорбцией токсичных продуктов эндогенного и экзогенного происхождения и их инактивацией. Глава 7. Система крови 183 Физиология гемоглобина (от греч. haema — кровь и лат. glomus — шарик). Гемоглобин — сложный белок (он относится к классу так называемых хромопротеидов), состоит из железосодержащих групп гема и белка глобина. Основные показатели: у мужчин содержание гемоглобина в среднем составляет 130–160 г/л, у женщин — 120–140 г/л. Гем — это комплексное соединение протопорфирина с железом, именно гем обеспечивает транспорт О2 и СО2), а также выполняет буферную функцию — он способен связывать ионы Н+. Физиологические соединения гемоглобина — оксигемоглобин, карбогемоглобин (соединение гемоглобина с СО2) и восстановленный гемоглобин, т.е. дезоксигемоглобин. В составе этих соединений гемоглобина сохраняется двухвалентное железо, благодаря гему он способен обратимо связывать О2 и выполнять дыхательную функцию, чему способствует также двояковогнутая форма — увеличение площади контакта с О2 и СО2 и возможности деформации эритроцитов при прохождении его через узкие капилляры. Патологические соединения гемоглобина описаны в разделе 8.5. Регуляция эритропоэза (образования эритроцитов, рис. 7.3). Продукция форменных элементов крови (гемоцитопоэз) осуществляется в гемопоэтических тканях: миелоидной (костный мозг) и лимфоидной (тимус, селезенка, лимфатические узлы). В миелоидной ткани образуются эритроциты, тромбоциты, гранулоциты, моноциты, предГИПОКСИЯ Клетки почечного интерстиция Гипоталамогипофизарнонадпочечниковая система Эритропоэтин Красный костный мозг Эритроциты Рис. 7.3. Пути влияния гипоксии на образование эритропоэтина в почках 184 Физиология шественники лимфоцитов. В лимфоидной ткани продуцируются лимфоциты, плазматические клетки. Нервная регуляция. Количество эритроцитов в крови может рефлекторно изменяться при раздражении рецепторов слизистой полости рта. Поэтому исследование содержания эритроцитов в крови проводят до принятия пищи. Регуляция с помощью биологически активных веществ. Основной орган синтеза эритропоэтина (гликопротеид) — почки, его образование стимулируется гипоксией. При избытке кислорода вырабатывается ингибитор эритропоэза. Гормоны. Стимулируют эритропоэз соматотропин, тиреоидные гормоны; глюкагон оказывает ингибирующее влияние на эритропоэз. Для выработки и функционирования эритроцитов нужны микроэлементы (железо, медь, цинк, кобальт), фолиевая кислота и витамин В12. 7.3. Физиология лейкоцитов и тромбоцитов Физиология лейкоцитов. Количество лейкоцитов в крови составляет 4 × 109–9 × 109/л. Лейкоциты содержат ядра, их размеры — 4–20 мкм. Увеличение количества лейкоцитов в крови называют лейкоцитозом, уменьшение — лейкопенией. Имеется две группы лейкоцитов: гранулоциты (нейтрофилы, эозинофилы, базофилы) и агранулоциты (моноциты, лимфоциты). Продолжительность жизни гранулоцитов и моноцитов составляет 4–20 дней, лимфоцитов — 100–120 дней. Лейкограмма (лейкоцитарная формула) — это процентное соотношение различных видов лейкоцитов в крови: нейтрофилы — 46–78%, эозинофилы — 1–5%, базофилы — 0–1%, моноциты — 2–10%, лимфоциты — 18–40%. Свойства лейкоцитов: фагоцитоз, амебовидная подвижность, миграция (диапедез) — способность проникать через стенку неповрежденных капилляров. Функции лейкоцитов: • защитная (фагоцитоз микробов, бактерицидное и антитоксическое действие, участие в иммунных реакциях) (рис. 7.4); • участие в процессах свертывания крови и фибринолиза; Глава 7. Система крови 185 • регенеративная — способствуют заживлению поврежденных тканей; • транспортная — являются носителями ряда ферментов. Функциональные особенности гранулоцитов. Нейтрофилы. Зрелые сегментоядерные нейтрофилы находятся в синусах костного мозга в течение 3–4 дней (в кровеносном русле — 6–8 ч), далее мигрируют в слизистые оболочки, ткани, где превращаются в микрофаги. Покинувшие сосудистое русло нейтрофилы в кровоток не возвращаются и разрушаются в тканях. Часть нейтрофилов удаляется из организма через ЖКТ. В лизосомах нейтрофилов происходит гидролитическое расщепление бактерий. Лизоцим нейтрофилов оказывает бактериостатическое и бактериолитическое действие, участвует в разрушении поврежденных при воспалении тканей и клеток организма. Интерферон нейтрофилов — один из элементов противовирусной защиты. Эозинофилы. Имеют цитоплазму, подобную содержимому лизосом; в крови циркулируют около 5 ч и постепенно переходят в ткани, где и разрушаются. Их количество повышается при аллергических реакциях, глистных инвазиях и аутоиммунных заболеваниях. Обладают фагоцитарной активностью (микрофаги), участвуют в процессах свертывания крови и фибринолиза (синтезируют плазминоген). Базофилы созревают в костном мозге в течение 1,5 сут, в кровь выходят через 2–7 дней, где циркулируют около 6 ч; содержат много гистамина, гепарина, гиалуроновой кислоты. Базофилы участвуют в аллергических и воспалительных реакциях, в процессах фагоцитоза, свертывания крови и фибринолиза. Функциональные особенности агранулоцитов. Моноциты — наиболее крупные клетки (12–18 мкм) по сравнению с другими лейкоцитами, они — предшественники Рис. 7.4. Механизм защитной функции системы крови 186 Физиология тканевых макрофагов, центральное звено мононуклеарнофагоцитарной системы. После миграции в ткани макрофаги живут более месяца. Фагоцитарная функция обеспечивается выраженной способностью этих клеток к миграции и накоплению в очагах воспаления, регионарных лимфатических узлах, селезенке, печени. Моноциты обеспечивают также противоопухолевый иммунитет. Лимфоциты. Основная их функция — участие в реакциях специфического иммунитета. Обучение клетокпредшественниц Т-лимфоцитов происходит в тимусе в результате контакта клеток с его стромой под действием гормонов, вырабатываемых в нем. По выполняемой функции выделяют несколько видов Т-лимфоцитов: Т-киллеры — осуществляют иммунный лизис клеток-мишеней (возбудителей инфекционных заболеваний, актиномицетов, микобактерий, опухолевых клеток), участвуют в реакциях отторжения трансплантата; Т-эффекторы (хелперы) — осуществляют реакции гиперчувствительности замедленного типа при многих инфекционных заболеваниях, участвуют в синтезе антител; Т-супрессоры — обеспечивают саморегуляцию системы иммунитета — подавляют иммунный ответ на антигены и предотвращают возможность развития аутоиммунных реакций; Т-клетки иммунной памяти — обеспечивают возможность воспроизведения иммунного ответа при повторном контакте организма с антигеном. Процесс обучения В-лимфоцитов происходит в лимфатических узлах кишечника, костном мозге, миндалинах. Различают В1- и В2-лимфоциты, В-супрессоры, подавляющие иммунный ответ, В-клетки иммунной памяти, В-киллеры (natural killers — естественные киллеры, NK-клетки), осуществляющие цитолиз клеток-мишеней (например, клеток, пораженных вирусом, клеток опухолей). В-лимфоциты обеспечивают реакции гуморального иммунитета, и среди них выделяют клеткипродуценты антител, причем каждая лимфоидная клетка способна продуцировать антитела одной специфичности. Имеются и другие виды лимфоцитов. Регуляция лейкопоэза. Различают лимфопоэз — процесс образования лимфоцитов и миелопоэз — процесс образования гранулоцитов и моноцитов. Глава 7. Система крови 187 Регуляция миелопоэза. Количество лейкоцитов в крови изменяется рефлекторно при раздражении пищей различных рецепторов слизистой рта. Лейкопоэтины стимулируют гранулоцитопоэз и моноцитопоэз. АКТГ, глюкокортикоиды и катехоламины стимулируют миелопоэз. Андрогены стимулируют миелопоэз, действуя на стволовые клетки и, возможно, стимулируя активность всех клеток костного мозга, участвующих в гемопоэзе. Регуляция лимфоцитопоэза. Стимулируют лимфопоэз лимфокины, интенсивно продуцируемые на фоне антигенного воздействия на организм. Лимфоцитарные кейлоны (вид гликопротеидов) — ингибиторы клеточного деления, что осуществляется с помощью подавления синтеза ДНК. Лимфопоэтины регулируют процессы дифференцировки лимфоцитов. Простагландин Е1 усиливает пролиферацию и дифференцировку Т-лимфоцитов. Простагландин Е2 подавляет митогенный ответ Т-клеток, но не В-лимфоцитов. Преимущественно подавляют лимфопоэз липопротеиды с низкой плотностью, ненасыщенные жирные кислоты, α2-глобулин (С-реактивный белок). Свойства и функции тромбоцитов. Тромбоциты — маленькие кровяные пластинки (диаметр 2–5 мкм), не имеют ядра, содержат гранулы. При соприкосновении с чужеродной поверхностью распластываются и выпускают псевдоподии. В 1 л крови содержится 180 × 109–320 × 109 тромбоцитов. Свойства тромбоцитов заключаются в их способности к: • фагоцитозу; • амебовидной подвижности; • секреторной активности; • адгезии; • агрегации. Адгезия — прилипание к чужеродной поверхности, в частности к измененной сосудистой стенке, например, коллаген, обнаженная базальная мембрана сосуда. Способствуют адгезии фибриноген, ионы кальция. При этом тромбоциты активируются, чему способствует их агрегация (рис. 7.5). Агрегация — склеивание тромбоцитов друг с другом, которое вызывает ряд веществ: адреналин, серотонин, 188 Физиология Б А Рис. 7.5. Тромбоциты: А — в покое; Б — активированные, взаимодействуют друг с другом (по Ch. Bauer, 2004) АДФ, тромбин, коллаген, простагландины, фактор, активирующий тромбоциты, ионы кальция. Функции тромбоцитов: • участие в свертывании крови; • ангиотрофическая — питание эндотелия капилляров, благодаря чему поддерживаются структура и функция сосудов микроциркуляторного русла; • регуляция тонуса сосудистой стенки — с помощью серотонина, содержащегося в гранулах тромбоцитов; • участие в защитных реакциях организма (фагоцитоз). Главным регулятором тромбоцитопоэза является тромбоцитопоэтин, образующийся в печени, почках и костном мозге и поступающий в кровь при тромбоцитопении. 7.4. Группы крови Группы крови обнаружили австрийский ученый К. Ландштейнер и чешский врач Я. Янский в 1901–1907 гг. при смешивании крови различных людей и наблюдении склеивания (агглютинации) эритроцитов. Они установили в эритроцитах людей наличие особых антигенов — агглютиногенов и предположили существование в сыворотке крови соответствующих им антител — агглютининов. Групповую принадлежность крови обусловливают изоантигены. У человека описано около 200 эритроцитарных изоантигенов. Они объединяются в групповые антигенные системы, в частности АВ0, Rh-hr и другие. Изоантигены передаются по наследству, постоянны в течение всей жизни. Как известно, в результате взаимодействия антигена с ан- Глава 7. Система крови 189 тителом происходит склеивание антигенных частиц, то же самое наблюдается и с эритроцитами. Нормальные антитела (α- и β-агглютинины системы АВ0) находятся в сыворотке крови людей и не связаны с иммунными антигенами. Имеется несколько систем групп крови. Система АВ0. Известны четыре основные группы крови системы АВ0: 0 α β (I), Аβ (II), В α (III), АВ00 (IV). Агглютиногены А и В — полисахариды, находятся в мембране эритроцитов. В эритроцитах может содержаться также антиген 0, но его антигенные свойства выражены слабо и в крови нет одноименных ему агглютининов. В эритроцитах обнаружены разновидности агглютиногенов А и В: А1–7 и В1–6, различающиеся по антигенным свойствам. Агглютины α и β находятся в плазме крови. Обнаружены α1- и β2-агглютинины (экстраагглютинины). Врожденные одноименные агглютиногены и агглютинины в крови одного и того же человека не встречаются, но могут находится иммунные анти-А- и анти-В-антитела. Особенностью крови системы АВ0 является наличие врожденных агглютининов. Все другие системы крови не содержат врожденных агглютинов. Система резус (Rh-hr) открыта в 1941 г. К. Ландштейнером и А. Винером при иммунизации кроликов кровью обезьяны макака-резус. Антигены системы резус (Rh) — липопротеиды, известно шесть разновидностей антигенов этой системы. Rh-фактор содержится в эритроцитах 85% людей (резус-положительная кровь, Rh+). У остальных 15% людей Rh-фактор отсутствует (резус-отрицательная кровь, Rh–). Антирезус-агглютинины могут сформироваться только при попадании резус-отрицательному человеку резус-положительной крови (переливание крови, беременность). В этом случае при повторном переливании резус-отрицательному человеку (реципиенту) резус-положительной крови может произойти агглютинация, т.к. у реципиента уже выработаются иммунные антирезус-антитела. Беременность и Rh-фактор. Если мать резус-отрицательна, а отец резус-положителен, то плод может быть резус-положительным. В конце беременности, особенно при нарушении гематоплацентарного барьера, небольшое количество крови плода (0,1–0,2 мл) может проникать в кровь 190 Физиология матери. В ответ в организме матери начинают вырабатываться антирезус-агглютинины. Первая беременность проходит сравнительно спокойно, но при повторной беременности титр антирезус-антител у матери нарастает, они проникают в кровь плода (их размеры малы) и происходит агглютинация и гемолиз эритроцитов плода (гемолитическая желтуха) — в тяжелых случаях с летальным исходом. Резус-положительная кровь плода может попасть в кровь резус-отрицательной матери во время родов. В этом случае опасность возникает при второй беременности, поскольку у женщины в крови будут Rh-антитела. Есть и другие антигенные системы, наиболее значимы из которых Келл, Даффи, Кидд и MNSs. Все они не имеют, как и Rh-hr, врожденных антител, но могут появиться иммунные антитела. Правила переливания крови: • необходимо переливать только одногруппную кровь по антигенной системе АВ0, чтобы предупредить встречу одноименных агглютиногенов и агглютининов; • нельзя переливать резус-положительную (Rh+) кровь резус-отрицательному (Rh–) реципиенту, т.к. при повторном переливании крови может развиться гемагглютинация. Если же резус-отрицательной женщине будет перелита Rh+ кровь, то вынашивание плода будет под вопросом и при первой беременности! Перед переливанием крови необходимо: • определить группу крови донора и реципиента по системе АВ0 и Rh-фактору; • провести пробу на совместимость (плазма реципиента + кровь донора), а также биологическую пробу (реципиенту вводят по 5–10 мл донорской крови 3 раза с 3-минутными интервалами), если не возникает осложнений — вводят всю порцию крови. По жизненным показаниям допускается переливание небольшого количества крови I (0) группы, не содержащей групповых агглютиногенов, реципиентам других групп. Также в исключительных случаях реципиентам группы IV (АВ), не имеющей групповых агглютининов, допускается переливание крови другой группы. Количество переливае- Глава 7. Система крови 191 мой крови в обоих случаях должно быть не более 1/10 объема циркулирующей крови. Детям можно переливать только одногруппную кровь! Кровь обладает замечательным свойством — свертываемостью, которая ярко проявляется при повреждении сосудов. При некоторых заболеваниях, передаваемых по наследству (гемофилии), жизнь человека обречена на страдания — физические и психические. Различают первичный и вторичный гемостаз. 7.5. Система ГПФ (гемостаза, противосвертывания и фибринолиза) Сосудисто-тромбоцитарный (первичный) гемостаз. В организме существует единая система, которая поддерживает жидкое состояние крови и растворение тромба после прекращения кровотечения, — система гемостаза, противосвертывания и фибринолиза (система ГПФ). Эта система останавливает кровотечение при повреждении сосудов и тканей, предотвращает массивное (лавинообразное) свертывание крови и восстанавливает проходимость сосудов после прекращения кровотечения. Факторами, запускающими процессы гемостаза, являются повреждение ткани и боль. Боль почти всегда связана с высокой вероятностью травмы. Поэтому в эволюции сформировалась безусловная рефлекторная реакция, направленная на предотвращение потери крови, повышение ее свертываемости при болевых воздействиях. Гиперкоагулемия, в свою очередь, усиливает риск тромбоза, что обеспечило формирование механизмов фибринолиза, позволяющего организму быстро очистить кровеносное русло от образовавшегося тромба. Различают два механизма гемостаза: сосудисто-тромбоцитарный (первичный, микроциркулярный) и коагуляционный (вторичный, свертывание крови), они взаимодействуют и перекрывают друг друга. Фактор, запускающий процессы гемостаза, — травма, разрушающая ткани. Сосудисто-тромбоцитарный гемостаз реализуется в сосудах микроциркуляторного русла (низкое кровяное давление и малый просвет сосудов) за счет спазма сосудов и образова- 192 Физиология ния тромбоцитарной пробки, активации факторов тромбоцитов, которые обозначают буквами Р (Р1–Р5) (рис. 7.6). Роль спазма сосудов. При травме тканей происходит спазм сосудов за счет их рефлекторного сокращения — кратковременный (первичный) спазм, затем присоединяется действие на стенку сосудов биологически активных веществ (серотонин, адреналин, норадреналин, тромбоксан А2), которые высвобождаются из тромбоцитов и поврежденных тканей. Этот спазм (вторичный) более продолжителен. Образование тромбоцитарной пробки, закрывающей просвет поврежденного сосуда. В результате адгезии тромбоцитов из них высвобождаются тромбоцитарные факторы (реакция высвобождения тромбоцитов). Они запускают ферментативный процесс свертывания крови (внешний механизм), вследствие которого образуется небольшое количество тромбина и фибрина. Нити фибрина оплетают тромбоциты, что ведет к образованию тромбоцитарной пробки. Из тромбоцитов высвобождается особый белок — тромбостенин, под влиянием которого происходит сокращение тромбоцитарной пробки, после чего она прочно закрывает просвет микрососуда и кровотечение останавливается. В норме эндотелий сосудов препятствует образованию тромбов в основном за счет выработки ингибитора агрегации тромбоцитов — простациклина (простагландина) Рис. 7.6. Сосудисто-тромбоцитарный гемостаз (основные стадии): 1 — 1-я стадия; 2 — 2-я стадия; 3 — 3-я стадия Глава 7. Система крови 193 и препятствует контактной активации, фиксации на эндотелии комплекса гепарин–антитромбин III. Коагуляционный гемостаз. Характеристика факторов свертывания крови. Гомеостаз с помощью свертывания крови (т.е. гемокоагуляции) заключается в переходе растворимого белка плазмы крови фибриногена в нерастворимое состояние — фибрин, что ведет к образованию тромба, закрывающего просвет поврежденного сосуда, и остановке кровотечения. Факторы свертывания крови обозначаются римскими цифрами с буквами F (например FIIа, буква «а» означает активный). Факторы содержатся в плазме крови, в форменных элементах и в тканях — это в основном белки многие из них являются ферментами, но находятся в крови в неактивном состоянии. Большинство плазменных факторов свертывания крови образуются в печени, для образования некоторых из них необходим витамин К. Приведем краткую характеристику факторов свертывания крови. Фактор I (фибриноген) под влиянием тромбина переходит в фибрин; принимает участие в агрегации тромбоцитов. Фактор II (протромбин) под влиянием протромбиназы переходит в тромбин. Фактор III (тканевой фактор — TF, внешний тромбопластин) входит в состав мембран клеток всех тканей и форменных элементов крови; активирует фактор VII и, вступая с ним в комплекс, активирует фактор Х. Фактор IV (Са 2+) участвует в образовании ряда комплексов факторов свертывания крови, входит в состав тромбиназы, способствует агрегации тромбоцитов, связывает гепарин; принимает участие в ретракции сгустка и тромбоцитарной пробки, тормозит фибринолиз. Фактор V (акцелератор — глобулин) активируется тромбином, усиливает действие фактора Ха на протромбин (входит в состав протромбиназы). Фактор VII (конвертин) принимает участие в формировании протромбиназы по внешнему механизму, активируется факторами IIa, III, IXa, Ха. Фактор VIII (антигемофильный глобулин А) образует комплексную молекулу с фактором Виллебранда, активируется тромбином, совместно с фактором IХа способствует активации фактора Х (внутренний механизм). Фактор IХ (фактор Кристмаса, антигемофильный глобулин В) активируется тромбином. Фактор Х (Стюарта–Прауэра) является ос- 194 Физиология новной частью протромбиназы. Активируется по внешнему и внутреннему механизму. Фактор ХI (предшественник внутреннего тромбопластина) активируется факторами IIa и ХIIа, необходим для активации фактора IХ. Фактор ХII (Хагемана, или контакта) активируется отрицательно заряженными поверхностями, адреналином, калликреином; запускает внутренний механизм образования протромбиназы и фибринолиза, активирует факторы ХI и VII и переводит прекалликреин в калликреин. Фактор ХIII (фибринстабилизирующий фактор, фибриназа) содержится практически во всех тканях и форменных элементах; стабилизирует фибрин, участвует в восстановлении стенок сосудов. Фактор ХIV (фактор Флетчера — прекалликреин) участвует в активации факторов ХII, IХ и плазминогена; переводит кининоген в кинин; активируется фактором ХIIa. Фактор ХV (фактор Фитцжеральда, Фложек, Вильямса) высокомолекулярный кининоген (ВМК) образуется в тканях; активируется калликреином; принимает участие в активации факторов ХI, ХII и переводе плазминогена в плазмин. Основными плазменными факторами свертывания крови являются: I — фибриноген; II — протромбин; III — тканевой фактор (TF); IV — ионы Са 2+. Факторы с V по ХIII — это дополнительные факторы, ускоряющие процесс свертывания крови, — акцелераторы. Кальций участвует в большинстве реакций процесса свертывания крови. Кровь, лишенная кальция, не свертывается! Процесс свертывания крови происходит на поврежденном эндотелии стенок сосудов и на поврежденных мембранах тромбоцитов и представляет собой каскад, в котором проферменты, переходя в активное состояние, способны активировать другие факторы свертывания крови. Обычно выделяют три фазы свертывания крови. Фазы назвали по получающимся результатам. Первая фаза — образование небольших количеств протромбиназы и тромбина по внешнему механизму. Механизм назвали внешним потому, что запускает его TF (тканевой фактор — липопротеиновый комплекс), отсутствующий в крови. Он появляется из поврежденных тканей, т.е. TF находится вне крови. Однако все другие Глава 7. Система крови 195 факторы свертывания постоянно имеются в крови (рис. 7.7). Свертывание крови осуществляется следующим образом. Структурной основой клеточных мембран является бимолекулярный слой фосфолипидов. Наружный слой фосфолипидов тромбоцитов и клеток эндотелия сосудов электронейтрален, что и обеспечивает атромбогенность. Фосфолипиды внутреннего слоя мембран тромбоцитов и клеток эндотелия сосудов имеют отрицательный электрический заряд. При повреждении наружного слоя тромбоцитов и эндотелия сосудов Ca 2+ плазмы крови и интерстиция взаимодействует с отрицательно заряженными фосфолипидами, в результате чего образуются активные зоны, к которым прикрепляются факторы свертывания крови. 1-я фаза 2-я фаза 3-я фаза Рис. 7.7. Основные процессы коагуляционного гемостаза: в рамках — активные комплексы; TF — тканевый фактор (липопротеиновый комплекс) Таким образом, инициируется процесс свертывания крови поврежденным эндотелием сосудов и поврежденными мембранами тромбоцитов (активированные тромбоциты). Далее тканевой фактор (TF) взаимодействует с фактором VII. Образуется активный комплекс TF + VIIa, под 196 Физиология воздействием которого активируются факторы Х и IX. Комплекс TF + VIIa назвали внешняя теназа (от англ. ten — десять, активирует фактор X). Еще раз подчеркнем, что все факторы в крови находятся в неактивном состоянии. Только 1% фактора VII находится в активном состоянии (VIIa). Факторы Va и Ха в присутствии ионов Са2+ на поверхности активированного тромбоцита образуют протромбиназу по внешнему механизму (Va + Xa + Са2+). Фактор Va в первой фазе высвобождается из активированных тромбоцитов. Под воздействием протромбиназы образуется небольшое количество тромбина (IIa). Вторая фаза — образование основного количества протромбиназы и тромбина по внутреннему механизму (участвуют только факторы, находящиеся «внутри» крови). Их активация запускается активными факторами, получившимися в первой фазе. С появлением первой порции тромбина (когда уже обнажен субэндотелий) возникают условия для активации тромбоцитов. На их мембране фиксируется фактор VIII, который под воздействием тромбина (фактор IIa), образовавшегося в первую фазу гемокоагуляции, переходит в фактор VIIIа. Последний, взаимодействуя с фактором IXа (см. рис. 7.7), образует вначале небольшое количество активного комплекса (VIIIa + IXa) — этот комплекс назвали внутренняя теназа (внутри крови). По мере появления все больших концентраций фактора IIa дополнительно активируются факторы VIII, V и XI, что приводит к увеличению содержания внутренней теназы, активирующий дополнительные порции фактора X и образование протромбиназы по внутреннему механизму (Va + Xa + Са 2+), переводящей протромбин в тромбин (основное количество тромбина). Третья фаза — преобразование фибриногена в фибрин и образование тромба, закрывающего поврежденный сосуд. Когда появляется значительное количество активированных тромбоцитов в месте локального повреждения сосуда, связываются активные и неактивные факторы II, V и VIII–XI. При этом продолжается образование внутренней теназы, протромбиназы (см. рис. 7.7) и тромбина (IIa), Глава 7. Система крови 197 переводящего фибриноген в фибрин, после чего формируется тромб. Переход фибриногена в фибрин осуществляется также в три фазы (рис. 7.8). Рис. 7.8. Фазы преобразования фибриногена в фибрин Далее формируется тромб — кровяной сгусток (скопление форменных элементов, в основном эритроцитов, в нитях фибрина). Тромб прикрепляется к стенке сосудов и уплотняется (ретракция) вследствие активации тромбином актинмиозиновой системы тромбоцитов. Некоторые авторы выделяют четвертую стадию гемостаза — локализацию сгустка (препятствие распространению свертывания крови в неповрежденных участках сосуда) с помощью действия естественных антикоагулянтов. Однако, с нашей точки зрения, это не стадия свертывания крови, а включение противосвертывающих механизмов, препятствующих обширному свертыванию крови, что могло бы привести к тяжелым последствиям Противосвертывающие механизмы При нарушении биологических механизмов регуляции жидкого состояния крови может произойти внутрисосудистое свертывание крови (ДВС-синдром). Имеется две группы факторов, обеспечивающих жидкое состояние крови. Факторы, предотвращающие свертывание крови: 1) неактивное состояние плазменных факторов свертывания крови; 2) антитромбогенное свойство эндотелия стенки сосудов, предотвращающего активацию тромбоцитов (они активируются при контакте с поврежденной стенкой сосудов). Кроме того, тромбоциты имеют отрицательный электрический заряд (–15 мВ), эндотелиоциты — –25 мВ. Это создает отталкивающую силу их друг от друга и исключает активацию тромбоцитов, пока не нарушен эндотелий сосудов. 198 Физиология Факторы, тормозящие свертывание крови — естественные антикоагулянты. Они делятся на две группы: первичные (синтезируемые в печени, легких и других органах, постоянно циркулирующие в крови) и вторичные (образуются в процессе свертывания крови и фибринолиза). Первичные антикоагулянты. Антитромбин III (АТIII) — гликопротеид, локализован преимущественно в интиме крупных сосудов. Его биосинтез осуществляется в печени и в эндотелиальных клетках. АТIII тормозит активность тромбина, факторов IХа, Ха, ХIа, ХIIа, плазмина и калликреина. Гепарин — мукополисахарид. Синтезируется в базофилах и во всех тканях организма, тормозит все фазы гемокоагуляции. Антикоагулянтный эффект гепарина реализуется с помощью образовывания комплексных соединений протромбином, тромбином, фибриногеном. Протеины (С, S, Ζ), основным из которых является протеин С. Он образуется в печени и секретируется в кровь в неактивном виде, является гликопротеидом. Активация неактивной формы протеина С осуществляется комплексом тромбомодулин-тромбин. Протеин С инактивирует активные плазменные факторы Vа и VIIIа и этим тормозит образование тромбина и фибрина в процессе свертывания крови. Имеются и другие факторы, тормозящие свертывание крови. Вторичные антикоагулянты — отработанные факторы свертывания (фибрин, активные факторы ХIа и Vа, фибринпептиды, отщепляемые от фибриногена), а также продукты фибринолиза. Регуляция свертывания крови Гуморальная регуляция осуществляется с помощью факторов свертывания крови, образуемых в различных клетках организма, особенно в клетках печени, легких, в сосудистой стенке. Синтез и выход в кровоток факторов гемокоагуляции регулируются по принципу обратной отрицательной связи (тип регуляции по отклонению): чем меньше факторов свертывания содержится в крови, тем больше их образуется в клетках, и наоборот. Гормоны по их влиянию на процесс гемокоагуляции можно разделить Глава 7. Система крови 199 на три группы: 1) стимулируют процесс свертывания крови (вазопрессин, окситоцин, глюкокортикоиды, минералокортикоиды, адреналин, половые гормоны); 2) тормозят процесс свертывания крови (инсулин); 3) оказывают двухфазное действие — вначале стимулируют процесс гемокоагуляции, а затем вызывают гипокоагуляцию (тироксин). Нервная регуляция. Возбуждение симпатического отдела автономной нервной системы (кровопотеря, боль, гипоксия, мышечная деятельность, стрессы и др.) вызывает гиперкоагуляцию за счет образования микровезикул, несущих и не несущих тканевой фактор, и активации тромбоцитов, усиленного выхода в кровоток факторов свертывания крови, ведущую роль при этом играет тканевой фактор. При болевом воздействии наблюдается гиперкоагуляция вследствие активации симпатоадреналовой системы (главную роль играет адреналин). Рефлексогенные зоны — хеморецепторы сосудов, возбуждаемые образующимся в кровтоке тромбином, рефлекторно стимулирующим выход АТIII из сосудистой стенки. Кроме того, тромбин, взаимодействуя с тромбомодулином, экспрессируемом на эндотелии, активирует РС, переводя его в деятельное состояние (АРС), что способствует возникновению вторичной гипокоагуляции. Корковые влияния (эмоции, предстартовое состояние) сопровождаются гиперкоагуляцией вследствие активации симпатоадреналовой системы. После остановки кровотечения включаются механизмы фибринолиза и восстановления кровотока в поврежденном сосуде. Фибринолиз Фибринолиз — совокупность ферментативных процессов, обеспечивающих расщепление нитей фибрина, образовавшихся в процессе свертывания крови, на растворимые комплексы и восстановление просвета кровеносного сосуда. Основные компоненты 1. Фермент плазмин (фибринолизин) находится в крови в неактивном состоянии в виде плазминогена, является протеолитическим ферментом, расщепляет фибрин, фибриноген, некоторые плазменные факторы свертывания крови и другие белки плазмы крови. 200 Физиология 2 . Активаторы плазминогена (кислые и щелочные фосфатазы, трипсин, урокиназа и другие). Процесс ферментативного фибринолиза протекает в три фазы (рис. 7.9). Кроме того, в ферментативном фибринолизе участвует урокиназа, образующаяся в почках, на долю которой приходится около 15% общей фибринолитической активности, она напрямую активирует плазминоген, превращая его в плазмин. Урокиназа широко используется в качестве лечебного препарата. Рис. 7.9. Фазы фибринолиза (1, 2, 3): ВМК — высокомолекулярный кининоген Неферментативный фибринолиз обусловлен фибринолитическим эффектом комплексных соединений гепарина с некоторыми гормонами, биогенными аминами и тромбогенными белками крови. Неферментативным фибринолизом осуществляется процесс расщепления нестабилизированного фибрина (фибрин S), в результате чего он из фибриллярной формы превращается в глобулярную, и тем самым происходит очищение сосудистого русла от промежуточных продуктов образования фибрина. Глава 7. Система крови 201 Лейкоциты также способны захватывать фибрин, лизировать его и высвобождать в окружающую среду продукты его деградации. Лизис фибрина в лейкоцитах осуществляется содержащимися в гранулах фибринолитическими протеазами до высокомолекулярных фрагментов без участия плазмина. ГЛАВА 8 Дыхание — жизненная повседневная неощущаемая функция. О том, что мы дышим, каждый из нас узнает лишь при затруднении дыхания. В.А. Сафонов Система дыхания Система дыхания — это совокупность структур, обеспечивающих потребление организмом О2 и выделение образовавшегося при этом СО2. Она состоит из внешнего звена системы дыхания (легкие с воздухоносными путями, грудная клетка, мышцы, приводящие ее в движение) и внутреннего звена системы дыхания, включающего кровь, сердечно-сосудистую систему, органеллы клеток, которые обеспечивают тканевое (внутреннее) дыхание. Дыхание — совокупность процессов, обеспечивающих поступление в организм кислорода, использование его для окисления органических веществ с высвобождением энергии и выделением углекислого газа в окружающую среду. Термином «дыхание» обозначают также экскурсию грудной клетки и легких. В среднем в состоянии покоя человек потребляет в течение минуты 250 мл О2 и выделяет 230 мл СО2. Различают пять основных этапов дыхания: • газообмен между легкими и окружающей средой — вентиляция легких; • газообмен между кровью организма и газовой смесью, находящейся в легких; • транспорт газов кровью — О2 от легких к тканям, СО2 от тканей организма к легким; • газообмен между кровью и тканями организма — О2 поступает к тканям, а СО2 — из тканей в кровь; • потребление О2 тканями и выделение СО2 — тканевое (внутреннее) дыхание. Глава 8. Система дыхания 203 Совокупность первого и второго этапов дыхания — это внешнее дыхание, обеспечивающее газообмен между окружающей средой и кровью. Главная роль системы дыхания — снабжение организма О2 и выделение СО2. Дыхание обеспечивает высвобождение энергии. Энергия высвобождается на последнем этапе — тканевом дыхании, при окислении органических соединений. Энергия необходима для деятельности всех клеток, органов, тканей, организма в целом. Весьма важна роль системы дыхания в регуляции рН внутренней среды организма, что обеспечивается за счет выделения постоянно образующейся в организме Н2СО3 → → СО2 + Н2О (СО2 покидает организм через легкие, Н2О — с мочой). 8.1. Функции внешнего звена системы дыхания Функции легких. Легкие в процессе дыхания выполняют газообменную и негазообменные функции. Газообменная функция — главная. Структурно-функциональная единица легкого — ацинус, конечный элемент которого — альвеола (рис. 8.1). Диаметр альвеол — 0,3–0,4 мм, суммарная площадь всех альвеол достигает 80 м2, их число — около 300–350 млн. Негазообменные функции легких: • участвуют в процессах выделения, причем газообменная функция также выделительная (удаление СО2 из организма); посредством легких осуществляется выделение воды и некоторых летучих веществ: ацетона, этанола, эфира, закиси азота; • инактивируют и активируют биологически активные вещества. В легких инактивируется 90–95% простагландинов группы E и F; происходит превращение ангиотензина I в ангиотензин II под влиянием ангиотензиназы; • участвуют в выработке биологически активных веществ: гепарина, простагландинов, тромбопластина, факторов свертывания крови VII и VIII, гистамина, серотонина и др.; 204 Физиология • защитная функция — барьер между внутренней и внешней средой организма, где образуются антитела, осуществляется фагоцитоз с помощью альвеолярных фагоцитов, вырабатываются лизоцим, интерферон, иммуноглобулины; в капиллярах задерживаются и разрушаются микробы; • участвуют в процессах терморегуляции — в них вырабатывается большое количество тепла; кроме того, легкие участвуют в процессе теплоотдачи; • резервуар воздуха для голосообразования. Функции воздухоносных путей. Газообменная функция — доставка атмосферного воздуха в дыхательную зону и проведение газовой смеси из легких в атмосферу. Воздухоносные пути включают полость рта (при ротовом дыхании), носоглотку, гортань, трахею, бронхи и бронхиолы до 16 генераций включительно (рис. 8.1). Негазообменные функции воздухоносных путей осуществляются в основном слизистой оболочкой полости носа (носовое дыхание): • очищение вдыхаемого воздуха от пылевых частиц осуществляется главным образом в волосяном фильтре, где задерживаются пылевые частицы в преддверии полости носа и носовых ходах (верхний, средний и нижний); • увлажнение вдыхаемого воздуха — начинается еще в верхних дыхательных путях, и в первую очередь в полости носа, насыщением воздуха влагой слизистой оболочки. За сутки может испариться до 0,5 л воды; • согревание воздуха — уже на уровне 10-й генерации воздух нагревается до 37 °С; • периферический аппарат генерации звуков; • воздухоносные пути участвуют в процессах терморегуляции организма за счет теплопродукции, теплоиспарения и конвекции. Роль грудной клетки в процессе дыхания — обеспечение сужения и расширения легких, а значит, и их вентиляции. Грудная клетка предохраняет легкие от высыхания и механического повреждения. Давление в плевральной щели несколько ниже атмосферного. В норме на 4 мм рт. ст. в конце выдоха Глава 8. Система дыхания 205 Трахея Генерация 0 3 150 мл 2 Бронхи Воздухоносный путь 1 4 17 18 19 20 21 22 2500 мл Альвеолярные Дыхательные ходы бронхиолы Альвеолярные мешочки Переходная зона Дыхательная зона Газообменная область 16 Бронхиолы 15 200 мл 14 23 Рис. 8.1. Воздухоносный путь, газообменная область и их объемы после спокойного выдоха и 8 мм рт. ст. в конце вдоха. Таким образом, реальное давление в плевральной щели очень близко к атмосферному (752–756 мм рт. ст.). Значение пониженного давления в плевральной щели: • предотвращает спадение альвеол при выдохе, т.к. легкое все время растянуто; • играет важную роль в осуществлении вдоха (см. раздел 8.2). Происхождение пониженного давления. В процессе развития организма рост легких отстает от роста грудной клетки. Поскольку на легкие атмосферный воздух действует только с одной стороны (через воздухоносные пути), они растянуты и прижаты к внутренней стороне грудной клетки. Вследствие растянутого состояния легких возникает сила, стремящаяся вызвать спадение 206 Физиология легких. Эта сила называется эластической тягой легких (ЭТЛ). На рис. 8.2 она показана короткими стрелками, направленными внутрь. Плевральная щель 8 8 760 760 1 1 4 А 4 760 — ЭТЛ (мм рт. ст.) 2 760 2 Б Пневмоторакс Рис. 8.2. Состояние легких и грудной клетки в конце спокойного выдоха (А), расширение грудной клетки и спадение легких при пневмотораксе (Б): 1 — легкие; 2 — грудная клетка; ЭТЛ — эластическая тяга легких О том, что легкие находятся в растянутом состоянии, свидетельствует факт спадения их при пневмотораксе (от греч. рneuma — воздух, torax — грудь) — патологическом состоянии, возникающем при нарушении герметичности плевральной щели, в результате чего ее заполняет атмосферный воздух, оказываясь между висцеральным и париетальным листками плевры. Составные элементы ЭТЛ: • эластиновые и коллагеновые волокна; • гладкие мышцы сосудов легких; • поверхностное натяжение пленки жидкости, покрывающей внутреннюю поверхность альвеол. Силы поверхностного натяжения составляют 2/3 величины ЭТЛ. Величина поверхностного натяжения альвеолярной пленки существенно уменьшается в присутствии сурфактанта — активного вещества легких, покрывающего изнутри альвеолы, альвеолярные ходы, мешочки и бронхиолы. Сурфактант (от англ. surface active agents — поверхностно-активные вещества) содержит фосфолипиды, триглицериды, холестерин, протеины и углеводы. Значение сурфактанта. Уменьшает поверхностное натяжение жидкости, покрывающей альвеолы (почти Глава 8. Система дыхания 207 в 10 раз), что облегчает вдох, т.к. при вдохе легкие необходимо растянуть, и предотвращает ателектаз (слипание) альвеол при выдохе. Облегчает диффузию кислорода из альвеол в кровь вследствие хорошей растворимости кислорода в нем. Выполняет защитную роль: • защищает стенки альвеол от повреждающего действия окислителей и перекисей; • обладает бактериостатической активностью; • обеспечивает обратный транспорт пыли и микробов по воздухоносному пути; • уменьшает проницаемость легочной мембраны, а это профилактика развития отека легких в результате уменьшения выпотевания жидкости из крови в альвеолы. У курильщиков чаще встречаются заболевания легких, поскольку защитные свойства сурфактанта ослабевают. 8.2. Механизм вдоха и выдоха. Расход энергии Вдох осуществляется с помощью трех одновременно происходящих процессов. 1. Расширение грудной клетки. Оно обеспечивается сокращением инспираторных мышц — диафрагмы, наружных межреберных и межхрящевых. Грудная клетка при вдохе расширяется в трех направлениях: в вертикальном (рис. 8.3), фронтальном и сагиттальном. Рис. 8.3. Положение главной дыхательной мышцы — диафрагмы при вдохе и выдохе 208 Физиология Наружные межреберные мышцы при своем сокращении с одинаковой силой (F) верхнее ребро тянут вниз, а нижнее поднимают вверх, но система из каждой пары ребер поднимается вверх (рис. 8.4). Это связано с тем, что момент силы F2, направленной вверх, больше момента силы F1, направленной вниз, поскольку плечо действия силы у нижнего ребра (L2) больше плеча у верхнего ребра (L1), т.е. F1 = F2, но L2 > L1; поэтому F2 × L2 > F1 × L1. Расширению грудной клетки способствуют также силы ее упругости, т.к. грудная клетка в процессе выдоха сильно сжимается с помощью ЭТЛ, вследствие чего стремится расшириться. 2. Увеличение объема легких при вдохе обеспечивает одностороннее атмосферное давление, действующее через воздухоносные пути и постоянно прижимающее легкие к грудной клетке. Это означает, что увеличение отрицательного давления в плевральной щели не причина, а следствие расширения легких. Расширению легких при вдохе способствует также сила сцепления (адгезии) между висцеральным и париетальным листками плевры. Но она крайне мала по сравнению с атмосферным давлением, действующим на легкие через воздухоносные пути. Об этом свидетельствует тот факт, L1 F1 M L2 F2 Рис. 8.4. Механизм поднятия ребер при вдохе в результате сокращения наружных межреберных мышц (М): F1 — сила, опускающая ребро; F2 — сила, поднимающая ребро; L1, L2 — плечи действия сил F1 и F2 Глава 8. Система дыхания 209 что легкие при открытом пневмотораксе спадаются (отрываются от париетальной плевры), когда воздух поступает в плевральную щель и на легкие с обеих сторон (и со стороны альвеол, и со стороны плевральной щели) действует одинаковое атмосферное давление (см. рис. 8.2). 3. Воздух поступает в легкие при их расширении вследствие некоторого (на 2 мм рт. ст.) падения давления в них. Этого незначительного градиента давления достаточно, поскольку воздухоносные пути имеют большой просвет и не оказывают существенного сопротивления движению воздуха. Выдох осуществляется также вследствие одновременно проходящих трех процессов. 1. Легкие сжимаются при выдохе под действием ЭТЛ и эластической тяги брюшной стенки (напомним, что легкие и брюшная стенка при вдохе были растянуты). Поэтому на обеспечение выдоха в спокойном состоянии дополнительная энергия не затрачивается. 2. Сужение грудной клетки при выдохе также происходит под действием ЭТЛ и эластической тяги брюшной стенки. Это достигается следующим образом. Напомним, что после прекращения поступления импульсов к мышцам вдоха по диафрагмальным и межреберным нервам прекращается возбуждение мышц вдоха, вследствие чего они расслабляются. После этого грудная клетка сужается под влиянием ЭТЛ и эластической тяги брюшной стенки. При этом органы брюшной полости сжимаются и оказывают давление на диафрагму, что способствует ее поднятию (при наполненном желудке дышать труднее). Сужению грудной клетки (опусканию ребер) способствует также ее масса, но главную роль играет ЭТЛ. ЭТЛ передается на грудную клетку и сжимает ее, обеспечивая выдох за счет уменьшения давления атмосферного воздуха на грудную клетку изнутри через воздухоносные пути и легкие (см. рис. 8.2). Уменьшение давления равно силе ЭТЛ, т.к. с внутренней стороны реальное давление, оказываемое воздухом на грудную клетку, равно Ратм–Рэтл, а снаружи на грудную клетку действует Ратм. Этот перепад давлений действует и на вдохе, и на выдохе, но вдоху он препятствует (преодоление ЭТЛ), а выдоху способствует. ЭТЛ 210 Физиология сжимает грудную клетку как пружину при расслабленных мышцах. Сила сцепления (адгезии) висцерального и париетального листков плевры мала, поэтому в сужении грудной клетки в процессе выдоха практической роли не играет. 3. Воздух изгоняется из легких вследствие повышения давления в них (при спокойном выдохе +2 мм рт. ст.), т.к. объем легких при выдохе уменьшается, что ведет к сжатию воздуха и выталкиванию его из легких. Расход энергии на дыхание. При спокойном дыхании на работу дыхательных мышц затрачивается лишь около 1% потребляемого организмом кислорода. Для сравнения заметим, что ЦНС потребляет 20% O2, Na/K-помпа расходует 30% всей энергии организма в покое. При тяжелой физической работе расход энергии на обеспечение вентиляции легких может возрастать до 20% от общего расхода энергии организмом в основном из-за возрастания аэродинамического сопротивления вдоху и выдоху. Расход энергии на вентиляцию легких в покое так мал потому, что внешнее звено системы дыхания работает подобно качелям, для поддержания качания которых затрачивается весьма мало энергии. Это объясняется тем, что значительная часть энергии сокращения мышц, обеспечивающей расширение грудной клетки при вдохе, переходит в потенциальную энергию ЭТЛ и эластической тяги брюшной стенки, поэтому они растягиваются. Эта накопившаяся потенциальная энергия эластической тяги при вдохе обеспечивает выдох — сжатие грудной клетки как пружины после расслабления мышц вдоха. Вследствие сжатия грудной клетки при выдохе возрастают силы упругости грудной клетки, способствующие ее расширению при очередном вдохе. Подобный переход одного вида энергии в другой и обратно происходит в каждом цикле дыхания, что мы и называем дыхательными качелями. Именно поэтому мы так легко дышим, не замечая усилия. Дыхание происходит на подсознательном уровне, как и деятельность внутренних органов (сердца, желудка и т.д.). И только при фиксации внимания дыхательные движения осознаются. Глава 8. Система дыхания 211 8.3. Объемы и емкости легких (вентиляция) Объемы вентиляции легких непосредственно зависят от глубины вдоха и выдоха и частоты дыхания. Вентиляция легких — это газообмен между атмосферным воздухом и легкими. Повышение метаболических потребностей организма сопровождается непроизвольным усилением дыхания (гиперпноэ). Произвольное усиление дыхания, не связанное с метаболическими потребностями организма, называют гипервентиляцией. Различают легочные объемы и емкости, при этом под термином «емкость» понимают совокупность нескольких объемов (рис. 8.5). Общая емкость легких 4–6 л Емкость вдоха 2–3 л РО вдоха 1,5–2,5 л до 0,4–0,5 л ФОЕ 2–3 л РО выдоха 1,0–1,5 л ЖЕЛ М: 3,5– 4,5 л Ж: 3,0– 4,0 л Остаточный объем 1,0–1,5 л Рис. 8.5. Спирограмма (легочные объемы и емкости): ЖЕЛ — жизненная емкость легких; РО — резервный объем; ФОЕ — функциональная остаточная емкость Легочные объемы: • дыхательный объем (ДО) — это объем воздуха, который человек вдыхает и выдыхает при спокойном дыхании, при этом продолжительность одного цикла дыхания составляет 4–6 с, акт вдоха проходит несколько быстрее выдоха. Такое дыхание называется эйпноэ (хорошее дыхание); • резервный объем вдоха (РО вдоха) — максимальный объем воздуха, который человек может дополнительно вдохнуть после спокойного вдоха; • резервный объем выдоха (РО выдоха) — максимальный объем воздуха, который можно выдохнуть после спокойного выдоха; • остаточный объем (ОО) — объем воздуха, остающийся в легких после максимального выдоха. 212 Физиология Емкости легких: • емкость вдоха — максимальный объем воздуха, который можно вдохнуть после спокойного выдоха. Жизненная емкость легких (ЖЕЛ) — это наибольший объем воздуха, который можно выдохнуть после максимального вдоха. У молодых людей должную величину ЖЕЛ можно рассчитать по формуле: ЖЕЛ = Рост (м) × 2,5 (л); • функциональная остаточная емкость (ФОЕ) — количество воздуха, остающееся в легких после спокойного выдоха, равна сумме остаточного объема и резервного объема выдоха; • общая емкость легких (ОЕЛ) — объем воздуха, содержащийся в легких на высоте максимального вдоха, равна сумме ЖЕЛ и остаточного объема. Общая емкость легких, как и другие объемы и емкости, весьма вариабельна и зависит от пола, возраста и роста. Так, у молодых людей в возрасте 20–30 лет она составляет в среднем около 6 л, у мужчин в 50–60 лет — в среднем около 5,5 л. Минутный объем воздуха (МОВ) — это объем воздуха, проходящего через легкие за 1 мин. Он составляет в покое 6–8 л, частота дыхания — 14–18 в 1 мин. При интенсивной мышечной нагрузке МОВ может достигать 100 л. Максимальная вентиляция легких (МВЛ) — это объем воздуха, который проходит через легкие за 1 мин при максимально возможной глубине и частоте дыхания. Но дышит испытуемый 10 с, поскольку могут развиться респираторный алкалоз, сужение сосудов мозга, головокружение вследствие гипоксии мозга, т.к. Н2СО3 (вещество, расширяющее сосуды) вымывается из крови. Полученный результат умножают на 6. МВЛ может достигать у молодого человека 120–150 л/мин, а у спортсменов — до 180 л/мин, она зависит от возраста, роста, пола. При прочих равных условиях МВЛ характеризует проходимость дыхательных путей, а также упругость грудной клетки и растяжимость легких. МВЛ — один из вариантов форсированного дыхания. Тип дыхания зависит от рода трудовой деятельности и пола. У мужчин более выражен брюшной тип дыхания, Глава 8. Система дыхания 213 у женщин — преимущественно грудной тип. При брюшном типе в результате мощного сокращения диафрагмы органы брюшной полости смещаются вниз, поэтому при вдохе живот «выпячивается». Грудной тип дыхания в значительной степени обеспечивается за счет работы межреберных мышц. Доминирование того или другого типа дыхания зависит от вида трудовой деятельности (вернее, от положения тела при ее выполнении). Так, если выполнение физической работы затрудняет грудное дыхание, то формируется преимущественно брюшной тип дыхания и у женщин. 8.4. Газообмен между альвеолами и кровью организма Обмен газов между кровью организма и альвеолами осуществляется с помощью диффузии: СО2 поступает из крови в альвеолы, О2 — из альвеол в венозную кровь, пришедшую в легочные капилляры от всех органов и тканей организма. При этом венозная кровь, богатая СО2 и бедная О2, превращается в артериальную, насыщенную О2 и обедненную СО2. Движущая сила, обеспечивающая газообмен в альвеолах, это градиент парциального давления газов — разность парциальных давлений Ро2 и Рсо2 в альвеолярной смеси газов и напряжений этих газов в крови (табл. 8.1). Парциальное давление газа (от лат. partialis — частичный) — это часть общего давления газовой смеси, приходящаяся на долю данного газа. Напряжение газа в жидкости зависит только от парциального давления газа над жидкостью, и они равны между собой. Таблица 8.1 Ро2 и Рсо2 в альвеолах и напряжение этих газов в крови в мм рт. ст. и кПа (цифры в скобках) Альвеоляр- Артериальная ная смесь кровь, оттекагазов ющая от легких Газы Венозная кровь, поступающая в легкие О2 40 (5,3) 100 (13,3) 100 (13,3) СО2 46 (6,1) 40 (5,3) 40 (5,3) 214 Физиология Согласно закону Фика, скорость диффузии газа прямо пропорциональна площади барьера и градиенту парциального давления газа и обратно пропорциональна толщине барьера. Факторы, способствующие диффузии газов в легких: • свойства самого газа — углекислый газ диффундирует в альвеолы в 20–25 раз быстрее, чем кислород, вследствие его лучшей растворимости в жидкости и мембранах. Именно поэтому обмен СО2 в легких происходит достаточно полно, несмотря на небольшой градиент парциального давления этого газа в альвеолах и его напряжения в крови — всего 6 мм рт. ст. (для кислорода — до 60 мм рт. ст.); • тонкая легочная мембрана — выравнивание Pо2 в альвеолах и крови в легких происходит примерно за 0,25 с; кровь находится в капиллярах легких в среднем около 0,5 с, т.е. в 2 раза дольше; • большая поверхность контакта легочных капилляров и альвеол (60–120 м2); • корреляция между кровотоком в данном участке легкого и его вентиляцией — если участок легкого плохо вентилируется, то кровеносные сосуды в этой области сужаются и даже полностью закрываются. Это осуществляется с помощью механизмов местной саморегуляции, посредством реакций гладких мышц: при снижении в альвеолах Ро2 возникает вазоконстрикция; • интенсивность вентиляции и кровообращения различных отделов легких зависит от положения тела: в вертикальном положении лучше вентилируются нижние отделы, в горизонтальном — отделы легких, находящиеся внизу (в положении на спине — дорсальные, на животе — вентральные, на боку — тоже нижней части легких), что необходимо учитывать в клинической практике. Изменение состава газовой смеси в легких при дыхании. В покое человек потребляет около 250 мл О2 и выделяет около 230 мл СО2, поэтому в альвеолярном воздухе уменьшается количество О2 и увеличивается СО2 по сравнению с атмосферным воздухом (табл. 8.2). Глава 8. Система дыхания 215 Таблица 8.2 Состав атмосферного воздуха и газовой смеси в легких (в %) Компоненты Атмосферный воздух Выдыхаемая смесь газов Альвеолярная смесь газов О2 20,93 16,0 14,0 СО2 0,04 4,0 5,5 Азот и инертные газы 78,53 74,5 74,9 Пары воды 0,5 5,5 5,6 8.5. Транспорт газов кровью Транспорт кислорода. Газы транспортируются кровью главным образом в виде химической связи и лишь незначительная часть — в виде физического растворения. Практически весь О2 переносится кровью в виде химического соединения с гемоглобином. Гемоглобин — хромопротеид, обладающий замечательным свойством присоединять О2, когда кровь находится в легком, и отдавать О2, когда кровь проходит по капиллярам всех органов и тканей организма. Гемоглобин состоит из белка глобина и четырех одинаковых групп — гемов. Гем — это протопорфирин, в центре которого расположен ион двухвалентного железа, играющего ключевую роль в переносе О2. Содержание гемоглобина в крови у мужчин — 130–160 г/л, у женщин — 120–140 г/л. Количество О2, которое может быть связано в 1 л крови у мужчин, составляет 180–200 мл/л (кислородная емкость крови), у женщин она на 6–8% меньше, т.к. у них меньше Нb. Физиологические соединения гемоглобина. Кислород образует обратимую связь с гемом (оксигенация), причем валентность железа не изменяется. При этом восстановленный гемоглобин (Нb) становится оксигенированным НbО2 (оксигемоглобин). Гемоглобин соединяется с СО2 — это карбогемоглобин. Патологические соединения гемоглобина. Гемоглобин может соединяться с угарным газом (СО), образуя карбоксигемоглобин. Поскольку сродство гемоглобина к СО больше, чем к кислороду, гемоглобин, связанный с СО, не 216 Физиология может транспортировать О2. Первая помощь пострадавшему заключается в немедленном прекращении его контакта с угарным газом. Специфическое лечение — дыхание чистым кислородом, особенно полезно под повышенным давлением; поступление кислорода в кровь ускоряет диссоциацию НbСО. Гем может подвергаться также истинному окислению (двухвалентное железо превращается в трехвалентное — метгемоглобин). Опасность таких состояний заключается в том, что окисленный гемоглобин очень слабо диссоциирует (не отдает О2 тканям) и, естественно, не может присоединять дополнительно молекулы О2, т.е. он теряет свойства переносчика кислорода. Образование оксигемоглобина. Главный фактор, обеспечивающий образование оксигемоглобина, — высокое парциальное давление О2 в альвеолах (100 мм рт. ст.). Связывание кислорода гемоглобином происходит в капиллярах легких очень быстро. Время полунасыщения гемоглобина кислородом составляет всего лишь 0,01 с (длительность пребывания крови в капиллярах легких в среднем — 0,5 с). Пологий характер кривой образования и диссоциации оксигемоглобина в верхней ее части свидетельствует о том, что в случае значительного падения Ро2 в легких содержание О2 в крови будет сохраняться достаточно высоким (рис. 8.6), что биологически целесообразно. 100 Оксигемоглобин, % 80 60 40 20 0 0 20 40 60 80 100 Ро2, мм рт. ст. Рис. 8.6. Кривая образования и диссоциации оксигемоглобина при pH 7,4 и температуре 37 °С Глава 8. Система дыхания 217 Не только образование оксигемоглобина в легких, но и диссоциация его в тканях проходят в пределах одного и того же верхнего участка кривой (75–96% насыщения гемоглобина кислородом). Имеется ряд вспомогательных факторов, способствующих оксигенации крови: • отщепление от карбгемоглобина СО2 и удаление его; • понижение температуры в легких; • увеличение рН крови. Следует также отметить, что с возрастом связывание О2 гемоглобином ухудшается. Основные процессы образования оксигемоглобина (KHbO2) представлены на рис. 8.7. Диссоциация оксигемоглобина (см. рис. 8.6) происходит в тканевых капиллярах, куда поступает артериальная кровь. При этом гемоглобин не только отдает О2 тканям, но и присоединяет образовавшийся в тканях СО2. Главный фактор, обеспечивающий диссоциацию оксигемоглобина, — падение Ро2, который быстро потребляется тканями. Весьма важно, что имеется ряд вспомогательных факторов, способствующих диссоциации оксигемоглобина в тканях. К ним относятся: • накопление СО2 в тканях; • закисление среды; • повышение температуры; • увеличение количества 2,3-дифосфоглицерата — промежуточного продукта, который образуется в эритроцитах при расщеплении глюкозы. Некоторую роль в обеспечении мышц кислородом играет также и миоглобин (он присоединяет О2). Это особенно важно для сердечной мышцы, где источник энергии — в основном аэробное окисление. Транспорт углекислого газа. Углекислый газ образуется в тканях и переносится, как и кислород, плазмой и эритроцитами (Сеченов И.М., 1859) с помощью различных соединений. Большая часть СО2 транспортируется плазмой крови, причем около 60% всего СО2 находится в виде бикарбоната натрия (NaНСО3), небольшое количество — в виде физически растворенного СО2, и около 15 мл/л СО2 находится в виде Н2СО3. 218 Физиология Значительная часть СО2 находится в эритроците, причем в форме химических соединений — карбгемоглобин (HНbСО2) и бикарбонат калия (КНСО3). Углекислый газ, образующийся в организме, выделяется в основном через легкие (около 98%), и только 0,5% — через почки, около 2% — через кожу в виде НСО3-бикарбонатов. Важно отметить, что некоторое увеличение содержания СО2 в крови оказывает благотворное влияние на организм: увеличивает кровоснабжение мозга и миокарда, стимулирует процессы биосинтеза и регенерацию поврежденных тканей. Образование соединений углекислого газа происходит в эритроцитах и ускоряется карбоангидразой. Небольшое количество Н2СО3 (H2O + CO2 → H2CO3) образуется также в плазме, но этот процесс идет очень медленно, так как в плазме крови нет фермента карбоангидразы, катализирующего образование Н2СО3. В результате окислительных процессов и образования СО2 его напряжение в клетках достигает 60– 70 мм рт. ст., что значительно больше, чем в поступающей к тканям артериальной крови (40 мм рт. ст.). Поэтому СО2 согласно градиенту напряжения из клеток переходит в интерстиций и далее через стенку капилляров в плазму крови, из нее — в эритроцит. Под влиянием карбоангидразы Альвеолярная смесь газов СО2 О2 K+ + HCO3– + О2 + HHb СО2 KA KHbO2 + HCO3 HCO3– Na+ HCO3– K+ Cl– H2 O CO2 Cl– Плазма H2 O Эритроцит Рис. 8.7. Основные процессы газообмена между кровью и альвеолами легких: КА — карбоангидраза Глава 8. Система дыхания 219 СО2 + Н2О → Н2СО3. Ион НСО–3 с К+ образует КНСО3. Большая часть НСО–3 переходит в плазму, образуя NaHCO3. Диссоциация соединений СО2 происходит в процессе движения крови по капиллярам легких. В первую очередь начинается выход в альвеолы физически растворенного СО2 из плазмы крови, поскольку парциальное давление Рсо2 в альвеолах (40 мм рт. ст.) ниже, чем в венозной крови (46 мм рт. ст.). Это ведет к уменьшению напряжения Рсо2 в крови. Причем присоединение О2 к гемоглобину ведет к уменьшению сродства СО2 к гемоглобину и расщеплению карбгемоглобина (ННbСО2). СО2 уходит в альвеолы (подробнее см. рис. 8.7). 8.6. Регуляция дыхания. Защитные дыхательные рефлексы Характеристики дыхательного центра. Дыхательный центр локализуется в продолговатом мозге. Это было установлено с помощью перерезок ЦНС. Перерезка ствола мозга непосредственно над мостом не изменяет характер генерации дыхательного ритма, при перерезке ниже моста вдохи становятся затянутыми, более глубокими. Перерезка спинного мозга под продолговатым мозгом ведет к остановке дыхания. Нейроны дыхательного центра Мотонейроны спинного мозга — исполнительные (эффекторные): они получают импульсы от нейронов продолговатого мозга и посылают их к дыхательным мышцам по межреберным нервам (от сегментов ThIV–Th X) и диафрагмальным нервам (от СIII–CV). Иннервация мышц живота осуществляется ThIV–LIII-сегментами. Нейроны моста при взаимодействии с нейронами продолговатого мозга обеспечивают нормальный цикл дыхания — участвуют в регуляции продолжительности фаз вдоха и выдоха. Роль вышележащих отделов ЦНС в регуляции дыхания. Средний мозг играет важную роль в регуляции тонуса всей мускулатуры организма, в том числе и дыхательной. Гипоталамус выполняет интегративную роль в регуляции частоты и глубины дыхания при физической деятельности. Об участии коры большого мозга в регуляции дыхания, 220 Физиология в частности, свидетельствует тот факт, что частоту и глубину дыхания можно изменять произвольно. Дыхательный центр обладает автоматией. Впервые ее наблюдал с помощью гальванометра на изолированном продолговатом мозге лягушки И.М. Сеченов (1882). С помощью микроэлектродной техники ритмическую активность изолированного продолговатого мозга золотой рыбки зарегистрировал Эдриан (1931). Основная часть нейронов дыхательного центра в продолговатом мозге относится к ретикулярной формации, эти нейроны обладают автоматией. Автоматии дыхательного центра способствует афферентная импульсация от рефлексогенных зон — от хемо- и механорецепторов. В связи с тем, что цикл дыхания у человека состоит из вдоха и выдоха, имеется несколько классификаций нейронов дыхательного центра, основные из них: • инспираторные (ранние, возбуждающиеся в начале вдоха; поздние — в конце и полные — в течение всего вдоха); • экспираторные (соответственно — ранние, поздние и полные); • инспираторно-экспираторные; • экспираторно-инспираторные; • непрерывно активные; • постинспираторные (рис. 8.8). Саморегуляция вдоха и выдоха осуществляется благодаря циркуляции возбуждения в дыхательном центре и взаимного возбуждения и торможения нейронов (рис. 8.9). Дыхательный цикл начинается с возбуждения инспираторных ранних нейронов (ИР). Далее возбуждение переходит на полные инспираторные нейроны (ПИ). В процессе циркуляции возбуждения импульсы по возвратным связям поступают к предшествующим нейронам и тормозят их. Полные инспираторные и экспираторные нейроны по нисходящим путям посылают импульсы к мотонейронам спинного мозга, иннервирующим дыхательную мускулатуру. Данная схема саморегуляции вдоха и выдоха не отражает ряд известных в настоящее время процессов взаимодействия дыхательных нейронов моста, продолговатого мозга и афферентных импульсов от рефлексогенных зон, но она Глава 8. Система дыхания 221 А Б 1 2 3 4 5 6 Рис. 8.8. Активность основных групп дыхательных нейронов продолговатого мозга в связи с фазами дыхательного цикла (по Б.И. Ткаченко, 1994, с изменениями): А — вдох; Б — выдох. Нейроны инспираторные: 1 — ранние; 2 — полные; 3 — поздние; экспираторные: 4 — ранние; 5 — полные; 6 — поздние хорошо иллюстрирует главные процессы саморегуляции вдоха и выдоха. Значение блуждающих нервов в саморегуляции дыхательного цикла установили Геринг и Брейер в опыте с раздуванием легких воздухом в различные фазы дыхательного цикла — раздувание легких воздухом тормозит вдох, после чего наступает выдох. Уменьшение объема легких (забор воздуха) тормозит выдох, ускоряет вдох. После перерезки блуждающих нервов раздувание легких не изменяет характер дыхания. Во время вдоха вследствие растяжения легких возбуждаются их механорецепторы (рецепторы растяжения — локализуются в стенках трахеи и бронхов). Афферентные импульсы по блуждающим нервам поступают к дыхательным нейронам, тормозят вдох и способствуют смене вдоха на выдох (рефлексы Геринга–Брейера) (рис. 8.9). Главный гуморальный стимулятор дыхательного центра — избыток углекислого газа в крови. Влияние СО2 и ионов H+ на дыхание опосредовано главным образом их действием на особые структуры ствола мозга, обладающие хемочувствительностью (центральные хеморецепторы), хеморецепторы локализуются также в дуге аорты и синокаротидной области. 222 Физиология М-мост М ИР ИП Продолговатый мозг ПИ Спинной мозг Блуждающий нерв α Легкое Мышцы вдоха Рис. 8.9. Основные процессы саморегуляции вдоха и выдоха при спокойном дыхании. Нейроны: М — ретикулярной формации моста; ИР — инспираторные ранние; ПИ — инспираторные поздние, прерывающие вдох; светлые нейроны — возбуждающие; темные — тормозящие; α — мотонейрон спинного мозга Сосудистые хеморецепторы преимущественно реагируют на снижение в крови уровня О2, центральные хеморецепторы реагируют преимущественно на изменения в крови и спинномозговой жидкости на рН и Рсо2. Некоторые гормоны (адреналин, прогестерон при беременности) усиливают дыхание. Схема функциональной системы, регулирующей содержание в организме О2 и СО2, представлена на рис. 8.10. Защитные дыхательные рефлексы. Кашлевой рефлекс возникает при раздражении механорецепторов бронхов и трахеи, например, частицами пыли или малыми чужеродными телами. Гортань и бифуркация (место разделения трахеи на бронхи) обладают особо высокой чувствительностью, а конечные бронхиолы и даже альвеолы чувствительны к едким химическим раздражителям, например к газообразному диоксиду серы или хлору. Афферентные нервные импульсы из дыхательных путей передаются по блуждающим нервам в продолговатый мозг. Возбуждается дыхательный центр и осуществляется быстрый мощный вдох, а затем выдох, в результате чего организм освобождается от инородных частиц. Глава 8. Система дыхания Эмоциональное напряжение Легкие Все отделы ЦНС Дыхательный центр Физические нагрузки Вегетативная нейрогормональная регуляция Система крови О2/СО2 крови Сердечно-сосудистая система Почки Кожа Другие метаболиты Хеморецепторы сосудов Внешние воздействия 223 Обратная связь Рис. 8.10. Функциональная система, поддерживающая оптимальное для метаболизма содержание О2 и СО2 в крови (по К.В. Судакову, с изменениями) Чихательный рефлекс во многом похож на кашлевой рефлекс, но он связан с носовыми ходами. Чихательный рефлекс вызывается раздражением носовых ходов; афферентные импульсы проходят по V паре черепных нервов в продолговатый мозг, где и запускается рефлекторный ответ. Появляется серия ответных реакций, похожих на кашлевой рефлекс; при этом язычок нёба направлен вниз так, что большое количество воздуха быстро проходит через нос, помогая таким образом вычистить носовые ходы от инородных пахучих частиц. ГЛАВА 9 Когда я присутствую на вскрытии умершего от заболевания сердца, то в половине случаев не могу понять, от чего он умер, а в другой половине случаев — как он еще мог жить. С.П. Боткин Система кровообращения и лимфатическая система Центральным звеном системы кровообращения является четырехкамерное сердце. Оно состоит из левой и правой половин, каждая из которых включает предсердие и желудочек. Сердце — полый орган, стенки которого представляют собой поперечно-полосатые мышцы. Выделяют большой и малый круги кровообращения, но это деление условное, поскольку два круга включены в кровообращение последовательно, т.е. один — это продолжение другого (рис. 9.1). Две части системы кровообращения названы так потому, что каждая из них начинается в сердце и возвращается в сердце, но в отдельности замкнутых кругов они не образуют. Фактически имеется один общий замкнутый круг кровообращения. Главная функция системы кровообращения — обеспечение движения крови по всему организму. При отсутствии движения крови она не может выполнять транспортную функцию, обеспечивать образование мочи, участвовать в терморегуляции и защитных реакциях. Кроме того, в стенках сердца и сосудов вырабатываются многие биологически активные вещества, например атриопептид (натрийуретический гормон, гепарин, гистамин, серотонин, эндотелин) — сосудосуживающее вещество, антитела, факторы свертывания крови. При остановке деятельности сердца наступает смерть организма. Система кровообращения является интегрирующей системой в организме. Глава 9. Система кровообращения и лимфатическая система Малый круг кровообращения Полые вены 225 ЛВ ЛА Аорта ЛП ПП ПЖ Вены ЛЖ Артерии Большой круг кровообращения Рис. 9.1. Общая схема системы кровообращения: ЛА — легочная артерия; ЛВ — легочная вена; ПП и ЛП — соответственно правое и левое предсердия; ПЖ и ЛЖ — правый и левый желудочки Кровь циркулирует по кругу в результате циклической деятельности сердца и нагнетания крови в артериальную систему. 9.1. Цикл сердечной деятельности Цикл сердечной деятельности включает три фазы: систолу предсердий (от греч. systole — сокращение), систолу желудочков и общую паузу сердца. Общая пауза — это часть сердечного цикла, когда совпадают по времени диастола (от греч. dyastole — расширение) желудочков с диастолой предсердий. Весь цикл сердечной деятельности (при частоте его сокращений 75 в 1 мин1) длится 0,8 с. Графически цикл представлен на рис. 9.2. Рассмотрим каждую фазу цикла сердечной деятельности. Рис. 9.2. Временное соотношения фаз цикла сердечной деятельности при его длительности 0,8 с (75 сокращений в 1 мин): а — систола предсердий; б — систола желудочков; в — общая пауза сердца — совпадение по времени диастолы предсердий с диастолой желудочков 1 Число сокращений сердца у мыши — 600, у этруссканской землеройки — 1000 в минуту. 226 Физиология Систола предсердий начинается после общей паузы сердца и обеспечивает дополнительную подачу крови в желудочки (примерно 1/5 от конечного диастолического объема) (рис. 9.2, а). Значение систолы предсердий заключается еще и в том, что возникающее при этом давление в желудочках обеспечивает дополнительное растяжение миокарда желудочков и последующее усиление их сокращений во время систолы за счет миогенного механизма и эластичности желудочков. К началу систолы предсердий вся мускулатура предсердий и желудочков расслаблена, закрыты полулунные клапаны из-за градиента давления в полостях сердца и в артериальной системе, открыты атриовентрикулярные клапаны — они провисают в желудочки, расслаблены сфинктеры, представляющие собой кольцевые мышцы в области впадения полых вен в предсердия и выполняющие функцию клапанов. Поскольку весь рабочий миокард расслаблен, давление в полостях сердца равно нулю. Возбуждение предсердий начинается в области впадения полых вен, поэтому одновременно с сокращением рабочего миокарда предсердий сокращаются мышцы сфинктеров — они закрываются. Вследствие сокращения предсердий давление в их полостях начинает расти, дополнительная порция крови выталкивается в желудочки. Поскольку сфинктеры закрыты во время систолы предсердий, кровь из них обратно в полые вены не возвращается. К концу систолы давление в левом предсердии возрастает до 10–12 мм рт. ст., в правом — до 4–8 мм рт. ст. Такое же давление к концу систолы предсердий создается и в желудочках. После окончания систолы предсердий начинается систола желудочков. Систола желудочков обеспечивает выброс крови в артериальную систему. Она состоит из двух периодов — напряжения и изгнания (рис. 9.2, б). В период напряжения в сокращение вовлечены все мышечные волокна, поэтому давление в желудочках начинает быстро повышаться, вследствие чего закрываются атриовентрикулярные клапаны. Сокращающиеся вместе с желудочками сосочковые мышцы натягивают сухожильные нити и препятствуют выворачиванию клапанов в предсердия. Кроме того, эластичность и растяжимость сухожиль- Глава 9. Система кровообращения и лимфатическая система 227 ных нитей смягчают удар крови об атриовентрикулярные клапаны, что обеспечивает долговечность их работы. Давление в желудочках быстро нарастает. Как только давление в левом желудочке окажется больше диастолического давления в аорте (70–80 мм рт. ст.), а в правом желудочке — больше диастолического давления в легочной артерии (10– 12 мм рт. ст.), открываются соответствующие полулунные клапаны — начинается период изгнания. В период изгнания крови давление в левом желудочке достигает 120–130 мм рт. ст. (систолическое давление в аорте и крупных артериях большого круга), а в правом желудочке — 25–30 мм рт. ст. Поскольку атриовентрикулярные клапаны закрыты, кровь обратно в предсердия не поступает. Волна их сокращения возникает в верхушке сердца и распространяется вверх, выталкивая кровь из желудочков в аорту и легочный ствол. Вслед за фазой изгнания начинается диастола желудочков, с которой частично совпадает и диастола предсердий, поэтому следующую фазу сердечной деятельности обосновано называть общей паузой сердца. Общая пауза сердца начинается с расслабления мышц желудочков (рис. 9.2, в). Когда давление в желудочках становится несколько ниже, чем в аорте и легочной артерии, полулунные клапаны закрываются. Расслабление желудочков продолжается, давление в них падает до нуля, что приводит к открытию атриовентрикулярных клапанов массой крови, накопившейся во время диастолы в предсердиях. Начинается период наполнения желудочков кровью, поступающей из предсердий и полых вен. Общая пауза сердца (0,37 с) обеспечивает наполнение камер сердца кровью и отдых сердца. Миокард интенсивно снабжается кровью, получает кислород и питательные вещества. Это весьма важно, т.к. во время систолы кровоток коронарных сосудов левого желудочка сильно снижен (см. коронарный кровоток). 9.2. Особенности свойств сердечной мышцы и ее энергетического обеспечения В сердце имеются мышечные волокна двух типов — клетки рабочего миокарда, составляющие основную массу сердца (99%), обеспечивающие его нагнетательную дея- 228 Физиология тельность, и атипические мышечные волокна, образующие проводящую систему сердца и обеспечивающие его автоматию. В этих клетках зарождается возбуждение, которое потом распространяется по всему сердцу. Сердечной мышце (рабочему миокарду) присущи те же свойства, что и скелетной мышце, но есть существенные особенности, отличающие их. Особенности возбудимости и возбуждения сердечной мышцы. Параметры потенциалов сердечной мышцы примерно такие же, как и у скелетной мышцы, за исключением длительности ПД — для желудочков она равна 300–350 мс вследствие длительной фазы инверсии (плато), по этой же причине и длительная рефрактерная фаза — до 270 мс. Плато обеспечивается медленным выходом K+ из клетки и входом Ca2+ в клетку (рис. 9.3). А мВ +30 K+ 0 Ca2+ 2 K+ 3 –30 1 K+ Na+ КП КУД –60 –90 Возбудимость N 100 200 300 мс 2 Б 1 Рис. 9.3. Механическая и электрическая активность сердца: А — потенциал действия (сплошная линия), сокращение миокарда (пунктир): 1 — фаза деполяризации; 2 — фаза инверсии; 3 — фаза реполяризации; (→ — медленный ток ионов; ⇒ — быстрый ток ионов); Б — фазовые изменения возбудимости клетки рабочего миокарда в процессе ее возбуждения: N — нормальная возбудимость; 1 — фаза абсолютной рефрактерности; 2 — фаза относительной рефрактерности. Ток ионов: ⇒ — быстрый; → — медленный Глава 9. Система кровообращения и лимфатическая система 229 Фазы деполяризации и реполяризации развиваются примерно так же, как и у скелетной мышцы. Длительная рефрактерная фаза предотвращает круговое распространение возбуждения по миокарду и нарушения деятельности сердца. Особенности проводимости. Возбуждение в сердечной мышце распространяется диффузно (во всех направлениях), в отличие от скелетной мышцы. Электрическое поле ПД возбужденной клетки — раздражитель для соседних клеток (оно обеспечивает деполяризацию их до КУД). Поэтому достаточно возбудить один кардиомиоцит, чтобы возбуждение распространилось на все кардиомиоциты. Это объясняется особенностями строения сердечной мышцы. Клетки сердечной и гладкой мышц, глии и амакринные клетки сетчатки глаза соединены между собой щелевыми контактами (щель около 2 нм). Кардиомиоциты соединяются друг с другом своими концами с помощью вставочных дисков, являющихся продолжением сарколемм клеток. В области вставочных дисков имеются высокопроводимые щелевые контакты (в сердце их называют нексусы), с помощью которых возбуждение передается посредством электрического поля от одной клетки к другой. Кардиомиоциты — цилиндрической формы, они могут ветвиться и образовывать пространственную сеть. Боковые поверхности кардиомиоцитов также соединены между собой нексусами. Таким образом, миокард хотя и состоит из отдельных клеток, но функционирует как единое целое — функциональный синцитий, благодаря быстрому распространению возбуждения по всему миокарду посредством электрических полей ПД. Особенности сократимости сердечной мышцы. Вопервых, сердечной мышце несвойственна суммация сокращений, поэтому она не может сокращаться тетанически. Это объясняется тем, что у нее длительный рефрактерный период — он равен продолжительности ПД и одиночного сокращения мышцы. Это означает, что весь период сокращения мышца сердца невозбудима, именно поэтому невозможны суммация и тетанус. Возникновение тетанического сокращения могло бы нарушить нагнетательную функцию сердца, что можно наблюдать в эксперименте при действии большой концентрации Са 2+, когда сердце сильно сокращается и останавливается в систоле. 230 Физиология Во-вторых, сердечная мышца подчиняется закону «все или ничего»: она либо не отвечает на раздражение, если оно ниже порогового, либо отвечает максимальным сокращением, если раздражитель достигает пороговой или сверхпороговой силы. Увеличение силы раздражения выше пороговой не увеличивает силу сокращения, как при действии на скелетную мышцу. Это объясняется тем, что у сердечной мышцы возбуждение, возникнув в одном месте, распространяется посредством электрических полей ПД диффузно по всем кардиомиоцитам и все они вовлекаются в сокращение. У скелетной же мышцы возбуждение проводится изолированно по отдельным мышечным волокнам, на соседние волокна возбуждение не передается. В-третьих, у сердечной мышцы более длительный период одиночного сокращения — он примерно соответствует длительности ПД: у предсердий — около 100 мс, у желудочков — 300–350 мс. Роль растяжимости и эластичности мышцы сердца в выполнении его функции. Во-первых, эластическая тяга, возникающая вследствие растяжения стенок сердца при его наполнении кровью, увеличивает силу сокращений миокарда — создается некоторый запас потенциальной энергии. Во-вторых, растяжимость и эластичность смягчают гидравлический удар, который мог бы возникнуть в результате несжимаемости жидкости. В-третьих, эластичность структурных элементов сердца обеспечивает возникновение в конце систолы упругих сил, способствующих расправлению — расслаблению сократившегося (сжавшегося) миокарда. Как только прекращается возбуждение клеток, стенки (особенно желудочков) расслабляются и расправляются, как предварительно сдавленная резиновая груша. Способность стенок сердца расправляться объясняется не только эластичностью отдельных элементов, но в большей степени различным направлением отдельных мышечных слоев друг относительно друга. Особенности энергетического обеспечения сердечной мышцы. Анаэробное окисление (анаэробный гликолиз) для сердца, в отличие от скелетной мышцы, играет незначительную роль. Глава 9. Система кровообращения и лимфатическая система 231 Главный источник энергии для сердца — процесс аэробного окисления неуглеводных субстратов. Это свободные жирные кислоты и молочная кислота (около 60%), пировиноградная кислота, кетоновые тела и аминокислоты (менее 10%). При интенсивной мышечной работе в крови накапливается молочная кислота в результате анаэробного гликолиза в мышцах. Лактат — это дополнительный источник энергии для сердца, причем, расщепляя молочную кислоту, сердце способствует поддержанию постоянства рН. Около 30% расходуемой сердцем энергии покрывается за счет глюкозы; при физической нагрузке увеличивается энергетическая доля жирных и молочной кислот при одновременном снижении энергетической доли глюкозы. Коэффициент полезного действия у сердца — около 30%, у скелетной мышцы — 20–25%. 9.3. Автоматия сердца Автоматия сердца — это способность сердца сокращаться под действием импульсов, возникающих в нем самом. Автоматией обладают только атипические мышечные волокна, формирующие проводящую систему. Клетки рабочего миокарда автоматией не обладают. Доказательство автоматии — ритмические сокращения изолированного сердца лягушки, помещенного в раствор Рингера (Станниус Г., 1880). Сердце млекопитающих, помещенное в теплый, снабжаемый кислородом раствор Рингера для теплокровных, также продолжает ритмически сокращаться. Проводящая система сердца (рис. 9.4) представляет собой атипические мышечные клетки, имеет в своем составе узлы, образованные скоплением этих клеток, пучки и волокна, с помощью которых возбуждение передается на клетки рабочего миокарда посредством электрического поля. Градиент автоматии — это убывание частоты генерации возбуждения проводящей системой сердца в направлении от предсердий к верхушке. Его наличие доказал Г. Станниус (1880) в опыте с накладыванием лигатур между различными отделами сердца лягушки и последующим подсчетом сокращений различных 232 Физиология СА-узел Бахмана Пучки Пучок Гиса Венкебаха Тореля Левая ножка Передняя ветвь АВ-узел Правая ножка Задняя ветвь Волокна Пуркинье Рис. 9.4. Проводящая система сердца отделов сердца. Водитель ритма сердца — синоатриальный узел. Находясь под влиянием экстракардиальных нервов, он определяет частоту сокращений сердца 60–80 в 1 мин. В случае повреждения этого узла функцию водителя ритма выполняет атриовентрикулярный узел (40–50 в 1 мин), далее — пучок Гиса (30–40 в 1 мин) и волокна Пуркинье (20 в 1 мин). Активность всех нижележащих отделов проводящей системы сердца проявляется только в патологических случаях, в норме они функционируют в ритме, навязанном им синоатриальным узлом, поскольку частота возникающих в нем импульсов выше. Градиент автоматии — это убывание частоты генерации возбуждения проводящей системой сердца в направлении от предсердий к верхушке. Его наличие доказал Г. Станниус (1880) в опыте с накладыванием лигатур между различными отделами сердца лягушки и последующим подсчетом сокращений различных отделов сердца. Водителем ритма сердца является синоатриальный узел. Находясь под влиянием экстракардиальных нервов, он определяет частоту сокращений сердца 60–80 в 1 мин. В случае повреждения этого узла функцию водителя ритма выполняет атриовентрикулярный узел (40–50 в 1 мин), далее — пучок Гиса (30–40 в 1 мин) и волокна Пуркинье (20 в 1 мин). Активность всех нижележащих отделов проводящей системы сердца проявляется толь- Глава 9. Система кровообращения и лимфатическая система 233 ко в патологических случаях, в норме они функционируют в ритме, навязанном им синоатриальным узлом, поскольку частота возникающих в нем импульсов выше. Механизм автоматии. Активация и инактивация Na-, K- и Ca- каналов П-клеток (пейсмейкерных клеток), как и других возбудимых клеток, осуществляется посредством изменения величины и знака заряда мембранного потенциала их. Дело в том, что у П-клеток повышена проницаемость для Na+ и Са 2+, что делает неустойчивым мембранный потенциал, создаваемый К+. Из-за преобладания входа в клетку Na+ и Са 2+ над выходом К+ из клетки во время диастолы развивается медленная диастолическая деполяризация (МДД) П-клеток (рис. 9.5). Когда МДД достигает КУД, активируются быстрые Саканалы и Са 2+ входит в клетку, обеспечивая дальнейшее развитие ПД П-клеток. Нисходящая часть ПД развивается, как обычно, за счет выхода К+ из клетки. После возникновения каждого ПД П-клеток цикл повторяется. Скорость распространения возбуждения в разных отделах проводящей системы различна. По проводящей системе предсердий и его рабочему миокарду она оди- КУД ДП Рис. 9.5. Потенциалы действия клетки пейсмекера синоатриального узла: ДП — диастолический потенциал; КУД — критический уровень деполяризации; 1а — МДД; 1б — быстрая деполяризация; 2 — инверсия; 3 — реполяризация (по R. Berne, 2004, с изменениями) 234 Физиология наковая — около 1 м/c. Далее возбуждение переходит на атриовентрикулярный узел, где имеет место задержка возбуждения на 0,05 с (скорость проведения возбуждения — 0,05 м/с). Задержка связана с малым диаметром клеток атриовентрикулярного узла по сравнению с клетками проводящей системы и сократительного миокарда предсердий, а также с последовательным уменьшением числа нексусов между клетками в этой области проводящей системы, в результате ПД возникают здесь медленнее. Эта задержка обеспечивает последовательное сокращение предсердий и желудочков. Затем возбуждение по пучку Гиса, его ножкам и волокнам Пуркинье передается на клетки рабочего миокарда, также с помощью электрического поля ПД волокон проводящей системы. Скорость распространения возбуждения по пучкам проводящей системы желудочков и по волокнам Пуркинье — до 4 м/с, что обеспечивает быстрый охват возбуждением всех отделов желудочков. Таким образом, роль проводящей системы сердца заключается в том, что она обеспечивает: • автоматию сердца; • надежность в работе сердца — при повреждении основного водителя ритма его в какой-то степени могут заменить другие отделы проводящей системы сердца, т.к. они тоже обладают автоматией; • последовательность сокращений предсердий и желудочков за счет атриовентрикулярной задержки; • синхронное сокращение всех отделов желудочков, что увеличивает их мощность. Аритмия в сердечной деятельности у здорового человека может проявляться экстрасистолией. Экстрасистола — это внеочередное сокращение сердца. Ее можно получить также в эксперименте. 9.4. Основные методы исследования деятельности сердца Деятельность сердца сопровождается электрическими, механическими и звуковыми явлениями, регистрация которых и их анализ позволяет судить о функциональном состоянии сердца в норме и при патологии. Глава 9. Система кровообращения и лимфатическая система 235 Важнейшим методом исследования состояния сердца является электрокардиография — регистрация в определенных участках тела суммарного электрического поля, генерируемого кардиомиоцитами в процессе их возбуждения. Электрические токи сердца можно наблюдать, если на сокращающееся сердце крысы набросить нерв нервномышечного препарата лягушки: мышца лягушки начинает сокращаться в ритме сердца крысы. Электрокардиограмма (ЭКГ) — кривая, отражающая процесс возникновения, распространения и исчезновения возбуждения в различных отделах сердца. ЭКГ показывает только изменения электрических потенциалов, но не сокращения миокарда. ЭКГ-отведение — это вариант расположения электродов на теле при регистрации ЭКГ. Интервал — совокупность зубцов и изоэлектрической линии, находящейся между ними. Имеется три основных варианта отведения. 1. Стандартные биполярные отведения (Эйнтховена): I отведение — левая рука/правая рука, II отведение — правая рука/левая нога и III отведение — левая рука/левая нога (рис. 9.6). Рис. 9.6. Расположение электродов при стандартных отведениях (Эйнтховена) и ЭКГ, получаемые при этих отведениях: направление электрической оси сердца (стрелки) и их проекции на стороны треугольника Эйнтховена 236 Физиология Эта система отведений регистрирует электрическую активность сердца во фронтальной плоскости. 2. Шесть грудных однополюсных отведений (Вильсона) — V1–6 (справа налево по возрастанию номера). 3. Три усиленных однополюсных отведения от конечностей (Гольдбергера). Анализ ЭКГ стандартных отведений (рис. 9.7). Зубец Р отражает процесс деполяризации и распространения возбуждения правого и левого предсердий. Желудочковый комплекс (интервал QRST) отражает процесс деполяризации распространения возбуждения по миокарду обоих желудочков (комплекс зубцов QRS), периоды полного охвата их возбуждением (сегмент RST, чаще ST) и реполяризации желудочков (зубец Т). Интервал R–R соответствует расстоянию между вершинами двух зубцов R, по времени он равен длительности одного сердечного цикла. Чем больше частота сердечных сокращений, тем короче это время. Этот интервал дает возможность определить частоту кардиоциклов, наличие или отсутствие аритмии в сердечной деятельности. Основные параметры ЭКГ — см. рис. 9.7. Соотношения величин зубцов ЭКГ в норме следующие: Q : R = 1 : 4; Р : T : R = 1 : 3 : 9. Таким образом, различные параметры ЭКГ дают разностороннюю информацию о состоянии электрической R = 1,5 мВ P = 0,15 мВ T = 0,5 мВ Рис. 9.7. Основные усредненные параметры электрокардиограммы во втором стандартном отведении Глава 9. Система кровообращения и лимфатическая система 237 активности сердца и широко используются в клинической и спортивной практике. Аускультация — выслушивание тонов сердца на поверхности грудной клетки. Тоны сердца — это звуки, возникающие при работе сердца. Различают четыре тона. Выслушивают обычно I и II тоны. Все тоны можно зарегистрировать с помощью фонокардиографа. Первый тон (глухой, протяжный, низкий) возникает в начале систолы желудочков, поэтому его называют также систолическим. Главная причина его возникновения — захлопывание атриовентрикулярных клапанов и возникающие при этом вибрации их туго натянутых створок, сухожильных нитей, массы крови в полостях желудочков, их стенок и ближайших сосудов. Второй тон (высокий, кратковременный) возникает при захлопывании полулунных клапанов аорты и легочной артерии и в результате вибрации их стенок и крови. Третий и четвертый тоны в норме, как правило, не выслушиваются, но обычно регистрируются на фонокардиограмме. Фонокардиография (ФКГ) — это методика регистрации тонов сердца с поверхности грудной клетки. Для регистрации фонокардиограммы используют микрофон, который прикладывают к грудной клетке в месте, где лучше выслушиваются тоны сердца. Звуковые колебания преобразуются в электрические, усиливаются и подаются на регистратор — фонокардиограф. Соотношение зубцов ЭКГ и ФКГ представлено на рис. 9.8. R ЭКГ T P Q P S I ФКГ II IV III 0 0,2 0,4 0,6 0,8 с Рис. 9.8. Соотношение зубцов ЭКГ и тонов сердца при их синхронной регистрации (I–IV, ФКГ) 238 Физиология Ультразвуковое исследование (УЗИ). Метод основан на использовании отражения ультразвука от границ двух сред с различной плотностью (кровь–ткань). Он позволяет определить индекс выброса (процент систолического выброса крови от конечнодиастолического объема в левом желудочке), поражение клапанов сердца. В норме этот выброс составляет 50%. 9.5. Регуляция деятельности сердца Сердце (как и система дыхания) быстро реагирует на эмоциональное и физическое напряжение, связанное с изменениями внешней и внутренней среды организма, с трудовой деятельностью, занятиями спортом (реакции других внутренних органов не такие срочные). Механизмы регуляции деятельности сердца, как и любого мышечного органа, — нервный, гуморальный и миогенный. Регуляция с помощью блуждающих нервов. Преганглионарные парасимпатические волокна — это аксоны нейронов ядер блуждающих нервов, расположенных в каудальном отделе продолговатого мозга. Они образуют синаптические контакты с помощью N-холинорецепторов с собственными внутрисердечными эффекторными холинергическими нейронами в сердце и посредством их реализуют свое влияние. Раздражение периферического отрезка блуждающего нерва вызывает торможение деятельности сердца (братья Веберы, 1845) (рис. 9.9). Тонус блуждающих нервов выражен ярко, о чем свидетельствует тот факт, что перерезка их в эксперименте или блокада атропином вызывают значительное повышение ЧСС (рис. 9.10, А). Рис. 9.9. Торможение деятельности сердца и падение АД при раздражении блуждающего нерва у кролика Глава 9. Система кровообращения и лимфатическая система 239 Рис. 9.10. Опыт на сердце, иллюстрирующий наличие тонуса блуждающих нервов и отсутствие тонуса симпатических нервов (регистрация ЭКГ): А — нарастание ЧСС у голубя после двусторонней ваготомии (стрелка); Б — фоновая ЧСС у морской свинки; В — ЧСС этой же морской свинки в условиях блокады симпатической нервной системы орнидом (ЧСС без изменений). Отметка времени — 1 с Поскольку блуждающий нерв все время сдерживает деятельность сердца, уменьшение его тонуса ведет к учащению, а увеличение — к урежению сердечных сокращений, т.е. один и тот же нерв при наличии собственного тонуса обеспечивает двоякий эффект и более совершенную регуляцию. Медиатором парасимпатической нервной системы является ацетилхолин. В сердце он с помощью М2-ионотропных рецепторов активирует непосредственно К+-каналы пейсмекерных клеток, но снижает активность Na- и Са-каналов, поэтому развивается гиперполяризация и МДД замедляется, задерживается и проведение возбуждения в атриовентрикулярном узле, а ЧСС урежается, сила сердечных сокращений ослабевает. Роль симпатических нервов в регуляции деятельности сердца. Симпатические сердечные волокна берут начало в боковых рогах пяти верхних грудных сегментов спинного мозга (Th1–Th5) и прерываются в звездчатом ганглии, от которого идут постганглионарные волокна к сердцу (рис. 9.11). Раздражение симпатического нерва вызывает повышение частоты и силы сердечных сокращений (Цион, 1867). И.П. Павлов (1887) обнаружил нервные веточки звездчатого ганглия, раздражение которых только усиливает сокращения сердца без изменения их частоты (усиливающий нерв, оказывающий трофическое действие). Эффекты раздражения симпатического нерва реализуются с помощью 240 Физиология Продолговатый мозг Ядро блуждающего нерва Спинной мозг Блуждающий нерв Симпатический нерв N X Th1–Th5 Х А N β1 Х M2 Эффекторная клетка Сердце Рис. 9.11. Пути распространения возбуждения по преи постганглионарным волокнам симпатического и блуждающего нервов, с помощью которых осуществляются влияния на деятельность сердца. Нейроны: А — адренергические; Х — холинергические. Нервные волокна: сплошные — преганглионарные; пунктир — постганглионарные. Рецепторы: β1 — адрено-, N- и M2-холинорецепторы β1-адренорецепторов, их активация посредством норадреналина ведет к увеличению проницаемости мембран пейсмекерных клеток для Na+ и Са 2+, поступление которых по медленным каналам в клетки ускоряет их медленная диастолическая деполяризация (МДД) — электрофизиологический процесс, наблюдается хронотропный эффект. Возрастание входа Са 2+ в клетки рабочего миокарда ведет также к усилению сокращений сердца. Тонус симпатического нерва для сердца не выражен (см. рис. 9.10). Роль α-адренорецепторов в регуляции деятельности сердца дискутируется. На рис. 9.12 представлены эффекты раздражения симпатического нерва. Эфферентные влияния симпатического и парасимпатического нервов выражаются не только в изменении частоты сердечных сокращений (хронотропное влияние) и их силы (инотропное влияние), но также в изменении проводимости (дромотропное влияние) и возбудимости миокарда (батмотропное влияние). Все влияния блуждающего нерва на сердце являются отрицательными, а симпатического нерва — положительными. Инотропное влияние па- Глава 9. Система кровообращения и лимфатическая система 241 расимпатического нерва на предсердия выражено лучше, чем на желудочки. В спокойном состоянии ЧСС определяется тонусом блуждающего нерва и гуморальными веществами, циркулирующими в крови, а при эмоциональной и физической нагрузке ЧСС возрастает в результате уменьшения тонуса блуждающего нерва и возбуждения симпатической нервной системы. Миогенный механизм регуляции силы сокращений сердца включает гетеро- и гомеометрический механизмы. Эта регуляция осуществляется за счет изменения интенсивности функционирования кардиомиоцитов при исключении влияния на них экстра- и интраорганной нервной системы, а также гуморальных факторов. Гетерометрический механизм регуляции определяет зависимость силы сокращений сердца от степени предварительного растяжения миокарда непосредственно перед сокращением. Так, предварительное растяжение сердца лягушки увеличивает силу его сокращения (Франк О., 1895). На сердечно-легочном препарате подобные исследования провел Э. Старлинг (1918). Однако растяжение должно быть умеренным, обеспечивающим максимальное число зон зацепления с помощью миозиновых мостиков нитей миозина и актина. В случае чрезмерного растяжения и сильного уменьшения зоны Рис. 9.12. Учащение и усиление сердечной деятельности у кошки при раздражении сердечной веточки звездчатого ганглия электрическим током в остром эксперименте: А — кровяное давление в сонной артерии; Б — давление в левом желудочке сердца; В — скорость нарастания давления в желудочке (первая производная давления ΔP/Δt) 242 Физиология контакта нитей миозина и актина сокращение миокарда, как и скелетной мышцы, будет слабым, а при отсутствии контакта нитей актина и миозина сокращение миокарда вообще невозможно. Значение механизма Франка–Старлинга заключается в усилении сердечной деятельности в случае увеличения притока крови к сердцу (преднагрузки). Гомеометрический механизм регуляции деятельности сердца включается при возрастании ЧСС (ритмоинотропная зависимость), что проявляется и на изолированной полоске миокарда без предварительного растяжения. Это объясняется увеличением запаса Са 2+ в саркоплазматическом ретикулуме кардиомиоцитов, в результате чего больше ионов Са 2+ высвобождается в ответ на каждый следующий ПД. Ионы Са 2+, как известно, обеспечивают взаимодействие нитей актина и миозина при возбуждении мышечного волокна, что и ведет к усилению сокращений миокарда. При учащении сокращений сердца больше Са2+ поступает в клетку и Са-помпа клеточной мембраны не успевает выкачивать Са2+. Роль внутрисердечной нервной системы. При высоком давлении в аорте наблюдается угнетение сердечной деятельности, при низком — усиление, т.е. внутрисердечная нервная система стабилизирует давление в артериальной системе, предупреждая резкие колебания давления в аорте (Косицкий Г.И.). Внутрисердечная нервная система образует периферические рефлекторные дуги, включающие афферентный нейрон, дендрит которого оканчивается рецептором растяжения на кардиомиоцитах и коронарных сосудах, и эфферентный нейрон, аксон которого заканчивается на кардиомиоцитах. Внутрисердечная рефлекторная дуга может иметь вставочный нейрон. Гуморальная регуляция деятельности сердца. Гормональные влияния. Тироксин увеличивает ЧСС, чувствительность сердца к симпатическим воздействиям. Возбуждение симпатоадреналовой системы и выброс катехоламинов в кровь при физической нагрузке и эмоциях ведут к увеличению частоты и силы сердечных сокращений. Кортикоиды, ангиотензин, серотонин, адреналин, норадреналин, вазопрессин, инсулин, глюкагон увеличивают силу сокращений сердца. Глава 9. Система кровообращения и лимфатическая система 243 Адреналин и норадреналин, циркулирующие в крови, увеличивают вход Cа 2+ в клетку в момент развития потенциала действия, вызывая тем самым усиление сердечных сокращений. Изменение концентрации Са2+ и К+ в крови сопровождается различными эффектами. Са 2+ усиливает сердечные сокращения, улучшая электромеханическое сопряжение, активируя фосфорилазу, что способствует высвобождению энергии. Большие концентрации Са 2+ вызывают настолько сильное сокращение сердца в эксперименте, что оно останавливается в систоле. Снижение концентрации К+ вне клетки ниже нормы (4 ммоль/л) или увеличение в 1,5–2 раза может сопровождаться нарушением ритма. Увеличение же концентрации K+ вне клеток в 3–4 раза и более ведет к снижению возбудимости и проводимости кардиомиоцитов и даже к остановке сердца, что используется в хирургической практике при операциях на сердце. Повышение температуры увеличивает ЧСС, а понижение — снижает. Корковые влияния на деятельность сердца весьма выражены, чаще это усиление и учащение сердечной деятельности. Кора реализует свое влияние посредством вегетативной нервной системы и эндокринных желез. Кора большого мозга обеспечивает возникновение более тонких приспособительных реакций сердца по механизму условного рефлекса. Например, в предстартовом состоянии у спортсмена частота и сила сердечных сокращений увеличиваются еще до сигнала стартера, т.е. заблаговременно. Таким образом, корковые влияния обеспечивают более тонкое приспособление деятельности сердца к потребностям организма. 9.6. Гемодинамика В настоящее время термин «гемодинамика» применяют в двух значениях. 1. Гемодинамика — учение о движении крови в сердечно-сосудистой системе; 2. Гемодинамика — движение крови по сосудам. 244 Физиология Непосредственной движущей силой, обеспечивающей кровоток, является разность кровяного давления между проксимальным и дистальным участками сосудистого русла. Давление крови в сосудах создается работой сердца и зависит от тонуса сосудов и количества крови в них. Препятствует движению крови сопротивление в кровеносных сосудах. Для иллюстрации приводят формулу Пуазейля: R = 8Lη/(πr4) [Па•с/см3], где R — гидродинамическое сопротивление; L — длина сосуда; r — радиус сосуда; η — вязкость крови; π — отношение окружности к диаметру. Однако эта формула не применима к эластическим сосудам. Кроме того, в организме расширение сосуда (аорты) происходит за счет ветвления его. Самый широкий просвет «сосуда» в области артериол и капилляров. Согласно формуле Пуазейля здесь должно быть самое маленькое сопротивление. В действительности же оно самое большое, так как сопротивление всех артериол и капилляров суммируется (табл. 9.1). Формула Пуазейля дает лишь представление, от каких факторов зависит сопротивление сосудов. Сердце не преодолевает сопротивление сосудов (см. в разд. 9.6). Таблица 9.1 Сопротивление току крови в различных отделах сосудистой системы организма (из учебника Б.И. Ткаченко, 2005) Сосуд Аорта Терминальные артерии Артериолы Капилляры Венулы Полые вены Сопротивление, дин•с•см–5 64 1,2•105 2•1010 3,9•1011 4•109 26 Поперечное сечение сосудов и кровоток. Наименьшую площадь поперечного сечения всего кровеносного русла имеет аорта — 3–4 см 2. По ходу артериального русла суммарное поперечное сечение его все возрастает, т.к. каждая артерия дихотомически делится. Самая большая Глава 9. Система кровообращения и лимфатическая система 245 суммарная площадь поперечного сечения — у капилляров. После чего она снова уменьшается в направлении к сердцу, поскольку венулы собираются и образуют вены, которые, в свою очередь, сливаются в полые вены. Объемная скорость кровотока (количество крови, протекающее через поперечное сечение сосуда в единицу времени) составляет 4–6 л/мин (МВ) и распределяется по регионам и органам в зависимости от интенсивности их метаболизма. При активном состоянии тканей кровоток в них может возрастать в 2–20 раз. Объем крови, протекающий через поперечное сечение в любом участке большого и малого кругов кровообращения, одинаков. Если нарушена эта закономерность, развиваются нарушения кровоснабжения органов и тканей организма вплоть до летального исхода. Линейная скорость кровотока — это скорость движения частиц крови относительно стенки сосуда (в аорте 20–25 см/с). Она различна в сосудах разного типа и зависит от объемной скорости кровотока и площади поперечного сечения сосудов. При равенстве объемной скорости кровотока в разных отделах сосудистого русла — в аорте, суммарно — в полых венах, капиллярах линейная скорость кровотока наименьшая в капиллярах, где самая большая суммарная площадь поперечного сечения. Методы исследования гемодинамики. Определение линейной скорости кровотока проводят с помощью ультразвукового и индикаторного методов. При этом в практической медицине обычно исследуют время полного кругооборота крови. В норме оно равно 21–23 с. В локтевую вену вводят индикатор (эритроциты, меченные радиоактивным изотопом, раствор метиленовой сини и др.) и отмечают время его первого появления в венозной крови соответствующего участка другой конечности. Ультразвуковое определение скорости кровотока основано на эффекте Доплера — ультразвук посылают через сосуд в диагональном направлении по току крови и против тока. По разнице во времени распространения звуковой волны по току крови и против (в последнем случае она меньше) прибор регистрирует линейную скорость кровотока. 246 Физиология Определение объемной скорости кровотока чаще производится с помощью окклюзионной плетизмографии и реографии. • Окклюзионная плетизмография основана на регистрации увеличения объема части конечности в ответ на прекращение венозного оттока при сохранении артериального притока крови в орган. Это достигается сдавливанием сосудов с помощью манжеты, например наложенной на плечо, и накачиванием в манжету воздуха под давлением выше венозного, но ниже артериального. Конечность помещается в камеру, заполненную жидкостью (плетизмограф), обеспечивающей регистрацию прироста ее объема (используются также воздушные герметически закрытые камеры). • Реография (реоплетизмография) — регистрация изменений сопротивления электрическому току, пропускаемому через ткань. Это сопротивление обратно пропорционально кровенаполнению ткани или органа. • Флоуметрия основана на разных физических принципах. В частности, при электромагнитной флоуметрии датчик плотно накладывают на исследуемый артериальный сосуд и осуществляют непрерывную регистрацию кровотока, основанную на явлении электромагнитной индукции. При этом движущаяся по сосуду кровь выполняет функцию сердечника электромагнита, генерируя напряжение, которое снимается электродами датчика, подается на регистратор, проградуированный в объемных единицах крови. • Метод разведения индикаторов. В артерию региона или органа быстро вводят определенное количество индикатора, не способного диффундировать в ткани (красители или радиоизотопы, фиксированные на белках крови), а в венозной крови через равные промежутки времени в течение одной минуты после введения индикатора определяют его концентрацию, по которой строят кривую разведения, позволяющую рассчитать объемную скорость кровотока. Самым точным методом является метод Фика — МВ определяется по артерио-венозной разности О2 (исследуют кровь и количество потребленного О2 за одну минуту). Классификация сосудов по функциям (по Б.И. Ткаченко в модификации В.Г. Афанасьева). Глава 9. Система кровообращения и лимфатическая система 247 1. Амортизирующие сосуды — аорта, легочная артерия и их крупные ветви, т.е. сосуды эластического типа. Специфическая функция этих сосудов — поддержание движущей силы кровотока в диастолу желудочков сердца и уменьшение колебания давления между систолой и диастолой желудочков за счет эластических свойств стенки сосудов (см. раздел 9.8). 2. Сосуды распределения — средние и мелкие артерии мышечного типа, обеспечивающие распределение потока крови по всем органам и тканям организма. Вклад этих сосудов в общее сосудистое сопротивление небольшой и составляет 10–20%. 3. Сосуды сопротивления: артерии диаметром менее 100 мкм, артериолы, прекапиллярные сфинктеры, сфинктеры магистральных капилляров. На долю этих сосудов приходится около 50–60% общего сопротивления кровотоку, с чем и связано их название. Сосуды сопротивления определяют кровоток системного, регионального и микроциркуляторного уровней. 4. Обменные сосуды — это в основном капилляры, через стенки которых происходит обмен веществ между кровью и интерстициальной жидкостью (газообмен, обмен жидкостью, метаболиты из интерстиция поступают в кровь). 5. Шунтирующие сосуды — артериоло-венулярные анастомозы. Их функция — шунтирование кровотока. Наиболее типичны эти шунты для кожи: при необходимости уменьшить теплоотдачу кровоток по системе капилляров прекращается и кровь (тепло) сбрасывается по шунтам из артериальной системы в венозную. 6. Емкостные (аккумулирующие) сосуды: венулы, мелкие вены, венозные сплетения и специализированные образования — синусоиды селезенки. Их общая емкость составляет около 50% всего объема крови, содержащейся в сердечно-сосудистой системе. Функции этих сосудов связаны со способностью изменять свою емкость, у них ярко выражены анастомозы. При сужении вен увеличивается приток крови к сердцу и систолический выброс. 7. Сосуды возврата крови в сердце: средние, крупные и полые вены, выполняющие роль коллекторов, через кото- 248 Физиология рые обеспечивается региональный отток крови и возврат ее к сердцу. Движение крови по артериям. Непосредственная сила, обеспечивающая движение крови по сосудам, — разность давлений (ΔР) в начале сосудистого русла (Р1 — аорта и легочная артерия) и в конце его (Р2 — вены в области предсердий) — ΔР = Р1 – Р2. Создается же и поддерживается высокое гидростатическое давление в сосудах организма сердцем. Весьма важный вспомогательный фактор движения крови по артериям — эластичность их стенок. Роль эластичности артерий 1. Уменьшает нагрузку на сердце и, естественно, расход энергии на обеспечение движения крови. Это достигается, во-первых, за счет того, что сердце не преодолевает инерционность столба жидкости и предотвращает гидравлический удар во время каждой систолы. Удар возникал бы в силу несжимаемости жидкости и быстрого выброса сердцем очередной порции крови, что приводило бы к разрушению структур сердца и сосудов. Очередная порция крови, выбрасываемая левым желудочком во время систолы, размещается в начальном отделе аорты за счет ее поперечного расширения в силу ее эластичности (рис. 9.13). Во-вторых, сердце не преодолевает общее периферическое сопротивление, оно огромно и сердце не могло бы его преодолеть, растянулось бы и остановилось. Сердце преодолевает только давление крови в аорте и легочной артерии. Энергия сокращения сердца переходит в потенциальную энергию эластической тяги аорты во время ее растяжения. Во время диастолы сердца эластическая тяга аорты сжимает ее и продвигает выброшенную порцию крови дальше от сердца (вазоэффект). Сердце же в период диастолы отдыхает. Обратный ток крови в аорте предотвращают аортальные клапаны. Особенно важно то, что давление крови, создаваемое сердцем в одной точке, передается на все сообщающиеся сосуды по всем разветвлениям, поэтому дополнительного расхода энергии сердцем на это тоже не требуется. 2. Обеспечивает непрерывный ток крови, что увеличивает объемную скорость крови в сосудистой системе и спо- Глава 9. Система кровообращения и лимфатическая система 249 А Б В Рис. 9.13. Механизм продвижения крови по аорте и артериям (по Э. Вицлебу, 1996, с изменениями): А — выброс крови сердцем в аорту, конец систолы; Б — начало диастолы; В — конец диастолы собствует непрерывному и более эффективному обмену веществ между кровью и тканями. 3. Увеличивает емкость сосудов. 4. Поддерживает кровяное давление в сосудах во время диастолы желудочков. Характеристика гемодинамики по артериям. Артериальное давление (АД) пульсирует. Пульсовая волна (область распространения повышенного давления) возникает в начальном сегменте аорты, а затем распространяется дальше. Давление крови в аорте и крупных артериях большого круга во время систолы называют систолоческим (Рс). В норме у взрослых людей в плечевой артерии оно составляет 115–140 мм рт. ст., диастолическое (Рд) — 60–90 мм рт. ст., пульсовое (Рп = Рс – Рд) — 30–60 мм рт. ст., среднее (Рср) — 80–100 мм рт. ст. Величина кровяного давления увеличивается с возрастом, но в норме не выходит за указанные границы. Во время сна давление снижается. У курильщиков систолическое давление больше на 10–15 мм рт. ст. в сравнении с некурящими. В начале систолы давление быстро повышается, а затем снижается, продолжая плавно уменьшаться и в диастоле желудков, но оставаясь достаточно высоким до следующей систолы. Среднее АД (Рср) — это давление, вычисленное 250 Физиология путем интегрирования во времени кривой пульсового колебания давления. Для центральных артерий его ориентировочно вычисляют по формуле: Рср = Рд + 1/3 Рп. Скорость распространения пульсовой волны гораздо больше скорости кровотока и зависит от растяжимости сосудистой стенки и отношения толщины стенки к радиусу сосуда, поэтому данный показатель используют для характеристики упруго-эластических свойств и тонуса сосудистой стенки. При снижении растяжимости стенки с возрастом (атеросклероз) и при повышении тонуса мышечной оболочки сосуда скорость распространения пульсовой волны возрастает. В норме у взрослых людей скорость распространения пульсовой волны в сосудах эластического типа равна 5–8 м/с, в сосудах мышечного типа — 6–10 м/с. Артериальный пульс (колебание стенки артерий вследствие изменения величины АД — систолического и диастолического) доступен для пальпаторного исследования в местах, где артерия располагается близко к поверхности кожи, а под ней находится костная ткань. По артериальному пульсу можно получить предварительное представление о функциональном состоянии системы кровоснабжения. В клинической практике оценивают высоту, скорость, напряжение пульса и его симметричность на обеих конечностях. На кривой регистрации пульса — сфигмограмме (рис. 9.14) отражаются повышение давления в артериях во время систолы желудочка — анакрота, снижение давления при расслаблении желудочков — катакрота — и небольшое увеличение давления под влиянием отраженного удара гидравлической волны о замкнутый полулунный клапан — дикрота. Частота пульса характеризует частоту сокращений сердца. Пульс менее 60 ударов в минуту называется брадикардией, более 90 в минуту — тахикардией. Ритмичный или аритмичный пульс дает представление о водителях ритма сердца. Наблюдается также дыхательная аритмия. На кривой АД, записанного прямым методом, регистрируются кроме пульсовых дыхательные волны кровяного давления (вследствие дыхательной аритмии ЧСС): на вдохе АД и ЧСС выше, чем на выдохе. Увеличение ЧСС является следствием уменьшения импульсации в блуждающих нервах. Это свидетельствует о том, что во время вдоха возбуждение в сер- Глава 9. Система кровообращения и лимфатическая система 251 мм рт. ст. 120 3 100 80 2 1 60 40 Рис. 9.14. Артериальный пульс (сфигмограмма) в сонной артерии: 1 — анакрота; 2 — катакрота (вся нисходящая часть); 3 — дикрота дечном центре блуждающих нервов тормозится. Механизм этого явления не изучен. Тормозная импульсация к центру кровообращения поступает, по-видимому, от нейронов дыхательного центра. Дыхательная аритмия биологически целесообразна — увеличение кровотока в сосудах легких на вдохе и объем свежего воздуха в легких увеличивают газообмен между кровью организма и смесью газов в альвеолах. Методы измерения артериального давления. При прямом измерении давления катетер или иглу вводят в сосуд и соединяют с прибором для измерения кровяного давления (манометром). Непрямые методы разработали Рива–Роччи и Коротков (изучаются на практических занятиях). В настоящее время используют автоматические или полуавтоматические методы измерения АД, основанные на методе Короткова; для диагностических целей применяют мониторирование АД с автоматической регистрацией его величины до 500 раз в сутки. При этом регистратор и датчик фиксируются на теле пациента. Движение крови по капиллярам. Характеристика микроциркуляторного русла (рис. 9.15). Длина капилляров — 0,5–1,0 мм, диаметр — 5–10 мкм, кровяное давление на артериальном конце — 30 мм рт. ст., в венозном — 15 мм рт. ст., средняя скорость кровотока — около 1 мм/с. В капиллярах осуществляется транспорт веществ через их стенку, в результате чего клетки органов и тканей обмениваются с кровью теплом, водой, газами и другими веществами, образуется лимфа. Время прохож- 252 Физиология Рис. 9.15. Схема микроциркуляторного русла (по R. Berne, M. Levy, 2004). Стрелки указывают направление движения крови дения эритроцита через капилляр большого круга кровообращения составляет 2,5 с, в малом круге — 0,3–1 с. Его определяют с помощью биомикроскопии, дополненной кинотелевизионным и другими методами. Транскапиллярный обмен веществ происходит путем фильтрации, простой и облегченной диффузии, трансцитоза и осмоса. • Объем транспорта веществ зависит от количества функционирующих капилляров и их проницаемости, от линейной скорости кровотока, от гидростатического и онкотического давления в капиллярах. • Фильтрация — главный фактор, обеспечивающий переход жидкости из капилляров в интерстиций (рис. 9.16). • Обеспечивает фильтрацию жидкости в артериальном конце капилляра фильтрационное давление (ФД). При этом способствует фильтрации гидростатическое давление крови (ГДК = 30 мм рт. ст.) и онкотическое давление жидкости в тканях (ОДТ = 5 мм рт. ст.). Препятствует фильтрации онкотическое давление плазмы крови (ОДК = 25 мм рт. ст.). Гидростатическое давление в интерстиции колеблется около нуля, поэтому: ФД = ГДК + ОДТ – ОДК = 30 + 5 – 25 = 10 (мм рт. ст.). Глава 9. Система кровообращения и лимфатическая система 253 • Реабсорбция межклеточной жидкости в капилляры. По мере продвижения крови по капилляру ГДК снижается до 15 мм рт. ст., в результате силы, способствующие фильтрации, становятся меньше сил, противодействующих фильтрации, — формируется реабсорбционное давление (РД), обеспечивающее переход жидкости из интерстиция в венозные концы капилляра: РД = ОДК – ГДК – ОДТ = 25 – – 15 – 5 = 5 (мм рт. ст.). Количество фильтрата (20 л/сут) несколько превышает количество реабсорбируемой жидкости (около 18 л/сут), однако эта часть воды (2 л) из тканей удаляется через лимфатическую систему. Между объемом жидкости, фильтрующейся в артериальном конце капилляра, объемом жидкости, реабсорбируемой в венозном конце и удаляемой лимфатическими сосудами, в норме существует динамическое равновесие. В случае накопления жидкости в интерстиции возникает отек тканей. Движение крови по венам. Характеристика венозной гемодинамики. Давление крови в посткапиллярных венулах равно 10–15 мм рт. ст., в полых венах вблизи сердца оно колеблется в соответствии с фазами дыхания от +5 до –5 мм рт. ст. Следовательно, движущая сила (ΔР) составляет в венах 10–15 мм рт. ст. Давление в крупных венах имеет пульсирующий характер, но волны давления распространяются по ним ретроградно — от устья полых вен к периферии. Причиной возникновения этих волн являются затруднения притока крови к сердцу из полых вен во время систолы правых предсердия и желудочка. Рис. 9.16. Обмен жидкости между кровью и интерстицием через стенку капилляра. Стрелками указаны направление движения жидкости и изменения движущей силы 254 Физиология Линейная скорость кровотока в венах, как и в других отделах сосудистого русла, зависит от суммарной площади поперечного сечения, поэтому она наименьшая в венулах (0,3–1,0 см/с), наибольшая — в полых венах (10–15 см/с). Течение крови в венах ламинарное, но в месте слияния двух вен в одну возникают вихревые потоки, перемешивающие кровь, ее состав становится однородным. Факторы обеспечивающие движение крови по венам. Основная движущая сила — разность давлений в начальном и конечном отделах вен, создаваемая работой сердца. Вспомогательные факторы, влияющие на возврат венозной крови к сердцу: • сокращения скелетных мышц, сдавливающих вены и венозные клапаны, обеспечивают движение крови по направлению к сердцу (обратному току препятствуют клапаны). Например, в венах стопы при ходьбе давление равно 15–30 мм рт. ст., а у стоящего человека — 90 мм рт. ст. Мышечный насос уменьшает фильтрационное давление и предупреждает накопление жидкости в интерстициальном пространстве тканей ног. У людей, стоящих длительное время, гидростатическое давление в венах нижних конечностей обычно выше и эти сосуды растянуты сильнее, чем у тех, кто попеременно напрягает мышцы голени, как при ходьбе; • пульсация артерий, ведущая к ритмичному сдавлению вен. Наличие клапанного аппарата в венах предотвращает обратный ток крови в венах при их сдавливании; • присасывающее действие отрицательного давления в грудной полости. При вдохе приток крови к сердцу возрастает больше, чем замедляется при выдохе; • присасывающее действие сердца. В момент открытия атриовентрикулярных клапанов давление в полых венах снижается и кровоток по ним в начальный период диастолы желудочков возрастает. Это результат быстрого поступления крови из правого предсердия и полых вен в правый желудочек (присасывающий эффект диастолы желудочков). Во Глава 9. Система кровообращения и лимфатическая система 255 время периода изгнания атриовентрикулярная перегородка смещается вниз, увеличивая объем предсердий, вследствие чего давление в правом предсердии и прилегающих отделах полых вен снижается. Кровоток увеличивается из-за возросшей разницы давления (присасывающий эффект атриовентрикулярной перегородки). • Гидростатический фактор в венах, расположенных выше сердца, способствует возврату крови к сердцу, а в венах, расположенных ниже сердца, препятствует этому. У лежащего человека давление в венах на уровне стопы равно примерно 5 мм рт. ст. Характеристика кровотока по всем сосудам организма представлена на рис. 9.17. Особенности кровотока в отдельных органах. Системное артериальное давление, т.е. давление в крупных артериях большого круга, обеспечивает одинаковую возможность кровотока в любом органе. В действительности интенсивность кровотока в различных органах весьма вариабельна и может изменяться в соответствии с запросами метаболизма в широком диапазоне. Особенности кровотока в скелетной мышце. Сокращение мышц уменьшает кровоток в артериях и обеспечивает Рис. 9.17. Соотношение линейной скорости кровотока, кровяного давления и суммарной площади поперечного сечения сосудов по ходу кровеносного русла 256 Физиология продвижение крови к сердцу в венах. В сосудах мышц ярко выражена рабочая гиперемия (см. раздел 9.7). Кровоток в легких осуществляется по двум сосудистым системам — по малому кругу кровообращения (основная из них), в нем осуществляется газообмен с альвеолярным воздухом; по сосудам большого круга кровообращения, обеспечивающего кровоснабжение легочной ткани (1–2% МВ). Систолическое давление в легочной артерии составляет 20– 25 мм рт. ст., диастолическое — около 10 мм рт. ст. Низкое АД объясняется высокой растяжимостью сосудов, широким их просветом, меньшей длиной и поэтому малым сопротивлением току крови. Артерии малого круга тонкостенны, им присущи выраженные эластические свойства. Гладкомышечные волокна имеются только в мелких артериях и прекапиллярных сфинктерах, типичных артериол малый круг не содержит. Легочные капилляры короче и шире системных. Давление в капиллярах легких равно 6–7 мм рт. ст., время пребывания эритроцита в капилляре — 0,3–1 с. Кровоток в коронарных сосудах. Во время систолы аортальные клапаны прижимаются кровью к стенкам аорты, закрывают вход в коронарные артерии и почти не пропускают кровь из аорты в коронарные артерии. Это биологически целесообразно, так как заполненные кровью под большим давлением артерии сердца во время каждой систолы превращались бы в жесткий гидравлический каркас (жидкость несжимаема), который препятствовал бы сокращению миокарда в период изгнания крови. В диастолу коронарный кровоток восстанавливается. В правом желудочке кровоток в фазу напряжения страдает меньше. В покое через коронарные артерии протекает около 4–6% МВ (200–250 мл крови в мин). Он может увеличится в 4 раза за счет увеличения линейной скорости и использования резервных капилляров. В покое 50% капилляров функционирует, 50% находятся в резерве. Венозная кровь из бассейна левой коронарной артерии оттекает в венозный синус (80–85% крови), а затем в правое предсердие; 10–15% венозной крови поступает через вены Тебезия в правый желудочек. Кровь из бассейна правой коронарной артерии оттекает через передние сердечные вены в правое предсердие. Глава 9. Система кровообращения и лимфатическая система 257 У коронарных сосудов человека нет анастомозов — в случае нарушения кровотока в сосуде развивается инфаркт в определенном участке миокарда. Важно то, что капилляры в миокарде располагаются весьма плотно — в 3–4 раза больше, чем в скелетной мышце, общая площадь диффузионной поверхности капилляров достигает 20 м2. Это создает хорошие условия для транспорта кислорода к миоцитам. Сердце потребляет в покое 25–30 мл кислорода в минуту, что составляет примерно 10% от общего потребления кислорода организмом. Кровоток в ЦНС постоянный при колебаниях АД 60– 160 мм рт. ст., что обеспечивается гуморальным, миогенным и нервным механизмами регуляции (см. разделы 9.7, 9.8). Кровоток в коже очень вариабелен вследствие терморегуляторных реакций. При понижении температуры окружающей среды закрываются с помощью симпатической нервной системы артерио-венозные шунты (анастомозы). Мало крови попадает в капилляры — сохраняется тепло, при повышении температуры окружающей среды развиваются противоположные реакции. Кроме того, при повышении температуры окружающей среды расширяются сосуды микроциркуляторного сплетения кожи с помощью холинергических нейронов, что ведет также к увеличению потоотделения. 9.7. Регуляция тонуса сосудов Количество крови, протекающее через любой орган, возрастает с увеличением системного АД и просвета сосудов в органе. Тонус сосудов (их просвет) регулируется с помощью нервного, гуморального и миогенного механизмов, что осуществляется в основном за счет артериол, метартериол, прекапиллярных сфинктеров, а также мелких и средних артерий и вен. Нервная регуляция тонуса сосудов. Иннервация сосудов осуществляется в основном симпатической нервной системой, она иннервирует все отделы сосудистого русла, кроме капилляров. Парасимпатическую иннервацию получают только некоторые органы. Возбуждение симпатической нервной системы вызывает сильную вазоконстрикцию в сосудах всего организма, кроме 258 Физиология сердца, мозга и легких. Значение слабой вазоконстрикции этих органов очевидно — сохранение достаточного кровоснабжения в жизненно важных органах при эмоциональном и физическом напряжении. Сужение сосудов во всех органах осуществляется с помощью α-адренорецепторов, расширение — посредством β-адренорецепторов. Расширение кровеносных сосудов осуществляется с помощью различных нервов: • главный нервный механизм вазодилатации — уменьшение тонуса симпатических сосудосуживающих нервов. Наличие тонуса у симпатических сосудосуживателей обеспечивает двоякий эффект: увеличение их тонуса сопровождается сужением сосудов, уменьшение тонуса этих нервов ведет к расширению сосудов; • вазодилатация в скелетных мышцах осуществляется благодаря активации β-адренорецепторов с помощью симпатических нервных волокон. Эта сосудорасширяющая система берет начало от моторной зоны коры большого мозга. Такое расширение сосудов мышц предшествует физической нагрузке (еще при планировании движения) — опережающее обеспечение мышц питательными веществами и кислородом. Расширение сосудов может наблюдаться также в мелких пиальных сосудах мозга, в мелких сосудах сердца; • в некоторых нервах имеются парасимпатические (холинергические) волокна, расширяющие сосуды органов челюстно-лицевой области и органов таза. Парасимпатическая вазодилатация является местной. Миогенная регуляция сосудистого тонуса наиболее выражена в артериолах. Она осуществляется с помощью Са2+ и определяет кровоток в органах. При повышении АД и растяжении мышц сосуда активируются Ca-каналы, в результате чего Са2+ диффундирует в клетки, что ведет к увеличению концентрации Са2+ в миоцитах, они сильнее сокращаются, сосуды суживаются, кровоток в органе уменьшается. При уменьшении давления в сосуде развиваются противоположные процессы. Миогенная регуляция Глава 9. Система кровообращения и лимфатическая система 259 стабилизирует кровоток в органах. Наличие тонуса даже у полностью денервированного сосуда называют миогенным тонусом. Гуморальная регуляция тонуса сосудов осуществляется биологически активными веществами, имеет три варианта по эффектам действия: 1) расширение сосудов: • сосудорасширяющим действием обладают многие биологически активные вещества: гистамин, брадикинин, ацетилхолин, окись азота, простагландины, атриопептид (натрийуретический гормон). Субстанция Р расширяет сосуды слюнных желез, наружных половых органов; • сосуды расширяются под влиянием снижения рO2 и метаболитов — рабочая гиперемия (СО2, угольная и молочная кислоты, снижение рН и др). Сосудорасширяющие вещества вырабатываются в эндотелии сосудов (например оксид азота NO); 2) сужение сосудов: • наиболее сильное вещество, суживающее артерии, артериолы и в меньшей степени вены, — ангиотензин, вырабатывающийся в печени. Ангиотензин не активен, он активируется в крови под действием ренина (получается малоактивный ангиотензин I, который под влиянием АПФ (ангиотензиназы) превращается в ангиотензин II — сильный вазоконстриктор). Норадреналин действует главным образом на α-адренорецепторы и суживает сосуды, в результате увеличивается периферическое сопротивление, но эффекты невелики, поскольку эндогенная концентрация норадреналина мала; 3) двоякий эффект: • некоторые биологически активные вещества обладают двояким действием на сосуды, например адреналин (рис. 9.18). Адреналин обладает двояким действием потому, что он активирует α- и β-адренорецепторы. Причем реакция сосудов кожи, мышц, органов брюшной полости зависит от концентрации адреналина. В низких концентрациях адреналин вызывает расширение сосудов этих органов, в высоких — сужение сосудов. Это объясняется тем, что 260 Физиология Симпатический нерв Активация α- и β-рецепторов НА Сужение А Расширение β Умеренный — расширение α Сосуд Максимальный — сужение Надпочечник: выброс адреналина (А) А α β НА α β Рис. 9.18. Адренергическая регуляция тонуса кровеносных сосудов. β-рецепторы боле чувствительны к адреналину, чем α-рецепторы: А — адреналин; НА — норадреналин чувствительность к адреналину β-рецепторов выше, чем α-рецепторов, поэтому физиологические концентрации адреналина активируют только β-рецепторы, что и ведет к расширению сосудов. По-видимому, эндогенный адреналин всегда или в подавляющем большинстве случаев вызывает расширение всех сосудов организма (покраснение лица при эмоциях). Только при сильном эмоциональном напряжении, кровопотере концентрация адреналина в крови может повыситься настолько, что преобладающим эффектом станет сужение сосудов вследствие одновременной активации β- и α-рецепторов. Преобладающий вазоконстрикторный эффект в этом случае, как и при экзогенном введении больших доз адреналина, объясняется тем, что α-адренорецепторов в различных сосудах организма больше, нежели β-рецепторов. Серотонин (5-гидрокситриптамин), высвобождаемый преимущественно энтерохромаффинными клетками, а также нейронами ствола мозга, при разрушении кровяных пластинок, обладает двояким действием. Вазоконстрикция особенно важна для прекращения кровотечения при повреждении сосудов. Механизм двоякого влияния серотонина изучен недостаточно. Глава 9. Система кровообращения и лимфатическая система 261 9.8. Регуляция артериального давления Величину АД определяют следующие факторы: • работа сердца; • просвет сосудов; • объем циркулирующей крови (ОЦК); • вязкость крови. Скорость изменения этих факторов различна. Работа сердца и просвет сосудов могут изменяться очень быстро, в течение нескольких секунд. Количество крови в организме и ее вязкость изменяются медленно. Естественно, чем больше объем циркулирующей крови, тем больше АД. В зависимости от скорости включения и длительности действия все механизмы поддержания АД можно объединить в три группы: механизмы быстрого, небыстрого и медленного реагирования. Механизмы быстрого реагирования. Эта регуляция осуществляется с помощью рефлекторного механизма (реакции срабатывают в течение нескольких секунд) и реализуется с помощью сосудистых реакций, включая вены и деятельность сердца. Центр кровообращения находится в продолговатом мозге. Нейроны, регулирующие деятельность сердца и просвет сосудов, расположены также в других отделах ЦНС. Важную роль в регуляции деятельности сердечно-сосудистой системы играет гипоталамус, который, как и продолговатый мозг, содержит прессорные и депрессорные зоны, нейроны которых посылают аксоны к соответствующим центрам продолговатого мозга и регулируют их активность. Кора большого мозга реализует свое влияние на сердечно-сосудистую систему в обеспечении приспособительных реакций организма с помощью вегетативной нервной системы (безусловных и условных рефлексов, при эмоциональном напряжении, например в предстартовом состоянии) и гормональных механизмов. Рефлексогенные зоны механизмов быстрого реагирования. Главные сосудистые рефлексогенные зоны расположены в дуге аорты и в области каротидного синуса, они содержат баро- и хеморецепторы. Отклонение параметров кровяного 262 Физиология давления в области этих рефлексогенных зон означает изменение системного АД, что воспринимается барорецепторами, и центр кровообращения вносит соответствующие коррекции. Эти зоны обеспечивают слежение за системным АД и снабжением кровью мозга. Чувствительные волокна от барорецепторов каротидного синуса идут в составе синокаротидного нерва, IX пара черепных нервов от барорецептов дуги аорты — депрессорным нервом. Изменения ЧСС при изменении системного АД представлены на рис. 9.19. Повышение АД вызывает увеличение импульсации, поступающей в центр кровообращения от барорецепторов. В результате усиливается возбуждение нейронов блуждающего нерва и угнетается сердечная деятельность. Усиливается также возбуждение депрессорной части симпатического центра и расширяются резистивные и емкостные сосуды организма, что также ведет к снижению АД (рис. 9.20). В случае уменьшения АД развиваются противоположные реакции, АД повышается, возвращаясь к нормальным значениям. Предсердная рефлексогенная зона. Возбуждение механорецепторов предсердий при растяжении их вследствие переполнения кровью и повышения давления крови в предсердиях ведет к увеличению ЧСС и силы сокращений сердца — рефлекс Бейнбриджа. Рефлекс препятствует застою крови в венах. Легочная рефлексогенная зона. Барорецепторы (механорецепторы) этой зоны локализуются в артериях малого круга Рис. 9.19. Влияние изменения кровяного давления в области каротидного синуса и дуги аорты на ЧСС (запись ЭКГ): А — понижение давления (забор крови из обеих сонных артерий); Б — повышение давления (1, 2 — обратное нагнетание крови через Т-образные канюли). Опыт на собаке, кровоснабжение головного мозга не нарушено Глава 9. Система кровообращения и лимфатическая система 263 кровообращения. Повышение давления в сосудах легких закономерно ведет к урежению сокращений сердца, к падению АД в большом круге кровообращения и увеличению кровенаполнения селезенки (рефлекс В.В. Парина) — защитная реакция, предотвращающая переполнение легких кровью и развитие отека легких. Попадание в сосуды легких (в патологических случаях) пузырьков воздуха, жировых эмболов, вызывающих раздражение механорецепторов сосудов малого круга кровообращения, вызывает настолько сильное угнетение сердечной деятельности, что может привести к летальному исходу — нормальная физиологическая реакция переходит, в случае чрезмерного ее проявления, в патологическую. Рефлексогенная зона ЦНС. При уменьшении кровотока в ЦНС из-за снижения АД кровоток восстанавливается вследствие повышения (нормализации) АД за счет сужения сосудов с помощью симпатической нервной системы. Если же кровоток в ЦНС снизился при нормальном АД (напри- Рис. 9.20. Рефлекторная регуляция системного АД: 1 — рефлекс с барорецепторов; 2 — рефлекс с хеморецепторов; 3 — центр блуждающего (парасимпатического) нерва; 4 — депрессорная часть симпатического центра; 5 — прессорная часть симпатического центра; 6 — ганглионарный холинергический нейрон; 7 — синоатриальный узел; 8 — рабочий миокард; — пре-; — постганглионарные нервные волокна 264 Физиология мер, сдавливание крупных артерий опухолью), АД резко возрастает. Нормальная приспособительная реакция для мозга становится патологической для всего организма. Сопряженные (межсистемные) рефлексы — это рефлекторные влияния на сердечно-сосудистую систему с рефлексогенных зон других органов или с сердечно-сосудистой системы на другие системы организма (Черниговский В.Н.). Большинство из них не принимают прямого участия в регуляции системного АД, однако имеют важное клиническое значение. Рефлекс Гольца — урежение ЧСС или даже полная остановка сердца при поколачивании по желудку и кишечнику лягушки. Рефлекс Даниньи–Ашнера (глазо-сердечный рефлекс) — это снижение ЧСС, возникающее при надавливании на боковую поверхность глаз и др. Эти рефлексы осуществляются посредством блуждающих нервов. Механизмы небыстрой регуляции, участвующие в регуляции АД, включают четыре основных варианта. Реакции развиваются в течение 5–20 мин. Существенную роль в регуляции системного АД играют изменения объема депонированной крови, количество которой составляет 40–50% от общего объема крови. Функцию депо выполняют селезенка (около 0,5 л крови), сосудистые сплетения кожи (около 1 л крови), где кровь течет в 10–20 раз медленнее, печень и легкие. Кровь из депо мобилизуется с помощью возбуждения симпатоадреналовой системы и может включаться в общий кровоток в течение нескольких минут, например при физическом и эмоциональном напряжениях. Регуляция АД посредством изменения скорости транскапиллярного перехода жидкости, что может осуществляться в течение 5–10 мин в значительных количествах. Повышение АД ведет к увеличению фильтрационного давления в капиллярах большого круга кровообращения и, естественно, к увеличению перехода жидкости в межклеточные пространства и нормализации АД. При снижении АД повышается реабсорбция жидкости из тканей в капилляры, в результате АД возрастает. Глава 9. Система кровообращения и лимфатическая система 265 Большое значение в регуляции АД имеет ангиотензин, суживающий сосуды. После снижения АД максимальный эффект развивается в течение 20 мин. Механизм регуляции АД с помощью изменения миогенного тонуса сосудов см. в разделе 9.7, c помощью ангиотензина — на рис. 9.21. Ангиотензин стимулирует выработку альдостерона, что ведет к задержке Na+ и воды и повышению АД (ангиотензин-альдостероновая система). Механизмы медленного реагирования — это регуляция системного АД с помощью изменения количества выводимой из организма воды, подробно рассмотренная в разделе 12.4. Функциональная система, поддерживающая оптимальный уровень АД, представлена на рис. 9.22. Рис. 9.21. Регуляция АД с помощью ренин-ангиотензинальдостероновой системы. Альдостерон задерживает Na+ и воду в организме, что также ведет к повышения АД Физическая нагрузка АД Эмоциональное напряжение Центр кровообращения Гипоталамус Кора Экстремальное воздействие Вегетативная нейрогормональная регуляция Просвет сосудов Работа почек (выведение воды) Переход жидкости в ткань и обратно Депонирование крови Метаболиты Барорецепторы и хеморецепторы сосудов Работа сердца Обратная связь Рис. 9.22. Функциональная система, поддерживающая оптимальный для метаболизма уровень системного АД (по К.В. Судакову, с изменениями) 266 Физиология 9.9. Кровобращение при изменениях положения тела, физическом и эмоциональном напряжении Реакции сердца при физической нагрузке. При максимальной физической нагрузке МВ возрастает в 6–10 раз. При этом объем кровотока в мышцах повышается в 30 раз, увеличивается систолическое АД при неизменном или несколько сниженном диастолическом давлении крови. Такое увеличение притока крови к работающим мышцам возможно только в результате перераспределения крови. Увеличение МВ при физической работе — это результат главным образом возрастания ЧСС (рабочего времени сердца). Увеличение ЧСС необходимо для обеспечения возрастающих потребностей организма в доставке кислорода и питательных веществ к усиленно работающим мышцам и удаления продуктов обмена, прежде всего углекислого газа. Стимуляция деятельности сердца запускается с рефлексогенных зон (при возбуждении хеморецепторов аортальной и каротидной рефлексогенных зон вследствие закисления крови и накопления К+ с проприорецепторов мышц) влиянием возбужденной моторной зоны на гипоталамус, который действует на бульбарный отдел центра кровообращения. Эти влияния реализуются с помощью нескольких механизмов: во-первых, вследствие уменьшения тонуса блуждающих нервов, во-вторых, в результате увеличения возврата крови к сердцу по венам (мышечный насос). Изменения интенсивности деятельности сердечнососудистой системы при эмоциональном напряжении подобны тем, которые возникают при физической нагрузке, — резко возрастают частота и сила сердечных сокращений, но АД повышается в большей степени, поскольку нет рабочей гиперемии в мышцах. Это полезная приспособительная реакция, сформировавшаяся в процессе эволюции. Она способствует мобилизации ресурсов организма. Например, перед стартом у спортсмена вследствие возбуждения симпатоадреналовой системы стимулируется сердечная деятельность, повышается обмен веществ, учащается дыхание — организм заранее готовится выполнить Глава 9. Система кровообращения и лимфатическая система 267 повышенную нагрузку. Эмоциональное напряжение вследствие различных переживаний при отсутствии физической нагрузки нецелесообразно, оно носит негативный характер и при частых повторениях может привести к патологическому состоянию организма. При переходе из горизонтального положения в положение стоя (ортостатическая проба: от лат. ortos — прямой, statos — стоящий) АД на несколько минут снижается, а ЧСС возрастает. В течение первых секунд в результате резкого снижения АД у некоторых людей может наблюдаться головокружение и даже обморок. Это объясняется тем, что вначале кровь депонируется в нижних конечностях — до 10–20% ОЦК. Затем увеличивается фильтрация жидкости в микрососудах, что при отсутствии двигательной активности может привести к развитию «скрытого» отека нижних конечностей. Из-за уменьшения возврата крови к сердцу снижаются на 40% систолический выброс (СВ), минутный выброс (МВ) и как следствие наблюдается снижение АД. Активация рефлекторных регуляторных механизмов ведет к усилению сердечной деятельности и сужению сосудов, ЧСС может повыситься на 30%, что ведет к повышению (нормализации) АД. Этому может способствовать и увеличение выброса катехоламинов надпочечниками. Возврату крови из нижних конечностей при вертикальном положении тела помогает и активация сократительной деятельности скелетной мускулатуры (мышечный насос). Гемодинамические реакции считают нормальными, если через 10 минут после перехода в вертикальное положение систолическое давление находится в пределах ±5% исходной величины, а диастолическое снижается не более чем на 5 мм рт. ст. 9.10. Лимфатическая система Лимфатическая система — это совокупность лимфатических сосудов и расположенных по их ходу лимфатических узлов, обеспечивающая всасывание межклеточной жидкости, веществ и возврат их в кровяное русло. Лимфатические сосуды представляют собой обширную разветвленную сеть мелких тонкостенных сосудов, 268 Физиология неравномерно представленную в разных участках тела. Начинается эта система с тончайших, закрытых с одного конца терминальных лимфатических капилляров. Стенки их обладают высокой проницаемостью, вместе с тканевой жидкостью внутрь лимфатического капилляра легко проходят молекулы белка. Наибольшее скопление лимфатических капилляров отмечено в печени, тонкой кишке. В результате слияния терминальных капилляров образуются лимфатические «вены», снабженные, подобно венам кровеносной системы, клапанами, препятствующими обратному току лимфы. Участки между двумя клапанами (клапанные сегменты) — лимфангионы — обеспечивают насосную функцию лимфатической системы (Орлов Р.С.). Самые крупные лимфатические сосуды впадают в кровеносную систему в месте соединения внутренней яремной и подключичной (плечевой) вен. Центральный коллектор лимфы у человека — грудной проток. В него впадают многочисленные сосуды, собирающие лимфу от нижних конечностей, органов брюшной полости, левой половины груди, от сердца и левого легкого, от левой верхней конечности, от левой половины головы и шеи. Проток впадает в угол, образованный левыми (наружной яремной и подключичной) венами в месте их слияния. Главный коллектор лимфы дополняется правым лимфатическим протоком, формирующимся путем слияния лимфатических сосудов правой половины головы, шеи, груди и правой верхней конечности. Этот проток впадает в правый венозный угол. Таким образом, вся постоянно образующаяся лимфа возвращается в общий кровоток. Лимфатические узлы — ключевые участки системы. Благодаря наличию гладкомышечных элементов они способны сокращаться, особенно при нейрогуморальных воздействиях. Число сосудов, приносящих лимфу в лимфоузлы, больше числа выносящих сосудов. Лимфатические узлы существенно влияют на клеточный состав лимфы. Лимфа очень медленно проходит здесь через узкие и извилистые синусы. Попадающие в нее бактерии фагоцитируются клетками лимфатического узла. При массивном заражении микробы могут пройти через все узлы и вторгнуться в кровеносное русло. Когда микробы попадают в лимфати- Глава 9. Система кровообращения и лимфатическая система 269 ческие узлы, в них развивается воспалительный процесс. Узлы увеличиваются в размерах, становятся болезненными. У людей, которые много курят, лимфатические узлы в области легких наполняются частицами табачного дыма, становятся темно-серыми или черными. Эти частицы могут нарушить функционирование лимфатических узлов, снизить устойчивость организма к легочным инфекциям, способствуют другим заболеваниям, в том числе и развитию рака легких. Функции лимфатической системы. Кроветворная функция. В лимфатических узлах формируются главным образом лимфоциты, продолжают дифференцировку долгоживущие Т-лимфоциты. Защитная функция заключается в транспорте антигенов и антител, в кооперации различных иммунокомпетентных клеток (лимфоцитов, макрофагов), в формировании первичного и вторичного иммунных ответов на антиген, реализации клеточного иммунитета, переносе из лимфоидных органов плазматических клеток для обеспечения гуморального иммунитета. Детоксикация — переход в лимфу из интерстиция патологически измененных белков, токсинов, клеток и обезвреживание их в лимфоузлах. См. также «Лимфатические узлы». Дренажная функция — удаление из интерстиция продуктов обмена и избытка воды, профильтровавшейся из кровеносных капилляров и не полностью реабсорбировавшейся. Лимфатическая система обеспечивает возврат белков, липидов и электролитов в кровь. За сутки в кровь возвращается около 40 г белка. Она транспортирует из пищеварительной системы в кровь продукты гидролиза пищевых веществ, в основном липидов. Лимфа состоит из лимфоплазмы и форменных элементов, в основном лимфоцитов. По химическому составу лимфа близка к плазме крови. Существенно различие только в содержании белков: в плазме крови их в среднем 70 г/л, тогда как в лимфе — около 20 г/л. Лимфа формируется из интерстициальной жидкости, давление в которой колеблется около нуля (оно может быть 270 Физиология выше или ниже атмосферного на несколько миллиметров ртутного столба). Давление в лимфатических капиллярах ниже атмосферного вследствие насосной функции лимфангионов. Пиноцитоз обеспечивает транспорт белков, хиломикронов, других частиц в полость лимфатического капилляра через эндотелиальные клетки. В лимфатические сосуды попадает только небольшая часть жидкости, которая не реабсорбируется обратно из интерстиция в кровеносный капилляр, а в виде лимфы возвращается в кровеносное русло. В лимфатические капилляры попадает 2–4 л жидкости в сутки. Белки в небольшом количестве попадают в интерстиций, затем в лимфокапилляр, за белком движется вода. Непосредственная движущая сила лимфы, как и крови, в любом участке сосудистого русла — это градиент давления, который создается несколькими факторами. Основной из них — сократительная активность лимфатических сосудов и узлов. Сократительная деятельность каждого лимфангиона активируется повышением в нем гидростатического давления вследствие попадания порции лимфы из соседнего лимфангиона. Для функций лимфатических сосудов характерны фазные ритмические сокращения (10–20/мин), медленные волны (2–5/мин) и тонус. Присасывающее действие грудной клетки, по общепринятому мнению, способствует продвижению лимфы в область крупных вен грудной полости, как и движение крови по венам. Способствуют движению лимфы пульсация близлежащих крупных артериальных сосудов, сокращение скелетных мышц, повышение внутрибрюшного давления. Клапанный аппарат обусловливает односторонний ток жидкости по направлению к сердцу при сдавливании лимфатических сосудов и их активном сокращении. Регуляция сократительной активности лимфангионов. Нервная регуляция (Орлов Р.С. и соавт., 1982). Возбуждение симпатической нервной системы ведет к учащению фазных сокращений лимфангионов с помощью активации α-адренорецепторов. Гуморальная регуляция. Катехоламины оказывают разнонаправленные влияния, гистамин, серотонин и вазо- Глава 9. Система кровообращения и лимфатическая система 271 прессин стимулируют сокращения лимфангионов. Окситоцин оказывает противоположные влияния. Миогенная регуляция заключается в том, что заполнение лимфой проксимального лимфангиона ведет к увеличению его растяжения, возрастанию силы сокращения и перекачиванию лимфы в соседний лимфангион и т.д. При этом клапанный аппарат лимфатических сосудов обеспечивает, как и в венах, ток лимфы только к сердцу. ПД миоцитов лимфангиона и одиночные его сокращения подобны таковым у сердечной мышцы (рис. 9.23). Рис. 9.23. Потенциал действия (1) и сокращение стенки лимфангиона (2) (по Г.И. Лобову, 1998, с изменениями) ГЛАВА 10 Пища — источник энергии и пластического материала для организма животного. Система пищеварения 10.1. Общая характеристика системы пищеварения Система пищеварения, включающая пищеварительный тракт (ротовой отдел со слюнными железами, глотка, пищевод, желудок, кишечник), поджелудочную железу и печень, обеспечивает поступление питательных веществ в организм. Пищеварение — это совокупность физико-химических процессов, происходящих в пищеварительном тракте, в результате которых из пищи образуются питательные вещества. Это продукты гидролиза белков, жиров и углеводов — мономеры (они лишены видовой специфичности, но сохраняют энергетическую и пластическую ценность), способные всасываться в кровь и лимфу, а также вода, минеральные соли и витамины, выполняющие только пластическую функцию. Система пищеварения выполняет пищеварительные и непищеварительные функции. Пищеварительные функции: • секреция ферментов, общее количество пищеварительных секретеров в сутки составляет 6–8 л (рис. 10.1); • моторика (двигательная функция) обеспечивает измельчение пищи, перемешивание ее с пищеварительными соками и перемещение содержимого в дистальном направлении; • всасывание (транспорт) продуктов гидролиза пищевых веществ, воды, солей и витаминов из полости пищеварительного тракта в кровь и лимфу. Глава 10. Система пищеварения 273 Поступление Общий объем 9000 мл/сут Отток Слюна 1000 мл Пища 2000 мл Желудок Желудочная секреция 2000 мл Панкреатическая секреция 1000 мл Двенадцатиперстная кишка Желчь 400 мл Секреция в тощей кишке 5000 мл 2000 мл Секреция в подвздошной кишке 600 мл 2900 мл Тощая кишка Подвздошная кишка Толстая кишка 1100 мл 1000 мл 100 мл Рис. 10.1. Баланс жидкости в ЖКТ. Из количества жидкости, поступающей в ЖКТ с пищей (2 л) и эндогенными секретами (7 л), с калом выводится только 100 мл (по К. Эве, У. Карабаху, 1996) Непищеварительные функции желудочно-кишечного тракта: • экскреторная функция осуществляется с помощью выведения из организма небелковых азотсодержащих веществ (мочевина, аммиак, креатинин, креатин), солей тяжелых металлов, лекарственных веществ; • защитная функция (специфические и неспецифические механизмы). Специфические механизмы защиты обеспечиваются иммунокомпетентными Т- и В-лимфоцитами миндалин глоточного кольца, солитарных лимфатических фолликулов стенки кишки и червеобразного отростка. Сюда же следует включить и выработку ОМФантител (антител к облигатной микрофлоре кишечника). Неспецифические механизмы защиты: – барьерная: слизистые оболочки препятствуют проникновению во внутреннюю среду организма бактерий, непереваренных пищевых веществ; 274 Физиология – бактерицидное и бактериостатическое действие пищеварительных соков; – фагоцитарное действие лейкоцитов; • эндокринная функция ЖКТ: ее выполняют специфические клетки диффузной эндокринной системы, вырабатывающие гастроинтестинальные гормоны, которые через кровь или местно (паракринным путем) оказывают регулирующее влияние на пищеварительную и другие системы организма. Основные закономерности деятельности пищеварительной системы. Конвейерный характер деятельности пищеварительного тракта (Павлов И.П.). Частично переваренная пища в ротовой полости по эстафете передается в последующие отделы пищеварительного тракта, где подвергается дальнейшей механической и химической обработке. Адаптация деятельности пищеварительных желез проявляется в выработке пищеварительных ферментов в соответствии с составом и количеством принятой пищи. В деятельности пищеварительной системы имеются базальные биоритмы с частотой 3–12 в минуту и околочасовой биоритм непищеварительной активности 14–16 в сутки (рис. 10.2). Особенности регуляторных процессов ЖКТ (табл. 10.1). Влияние экстраорганных нервов (ЦНС) наиболее выражено в начальном отделе пищеварительного тракта, оно постепенно снижается в дистальном направлении. Напротив, регуляторная роль интраорганной нервной системы в каудальном направлении возрастает. В интраорганной нервной системе содержится большое количество нейронов разной природы, образующих местные рефлекторные дуги. Гуморальные механизмы регуляции с наибольшей отчетливостью выражены в средней части пищеварительной системы (желудок, двенадцатиперстная кишка, поджелудочная 10.45 11.00 11.15 11.30 11.45 12.00 12.15 12.30 12.45 13.00 13.15 13.30 13.45 14.00 14.15 Рис. 10.2. Голодная моторная периодическая деятельность желудка (натощак) Глава 10. Система пищеварения 275 железа, печень), где особенно велика роль гастроинтестинальных гормонов. Натощак пищеварительный тракт находится в состоянии относительного покоя (кроме периодической голодной непищеварительной деятельности). Таблица 10.1 Влияние основных интестинальных гормонов на деятельность различных органов ЖКТ и печени Секреция Моторика ОслабСтимуляция Торможение Усиление ление ВИП, ГИП Желудок ХЦК (пепсин), Секретин (HCl), Гастрин, мотилин, ПП, мотилин (пеп- ХЦК (HCl), серотонин, син), гистаВИП, ГИП, мин, гастрин соматостатин, гистамин нейротензин (HCl–), энкефалин, серотонин (HCl–) ПодГастрин, сеПП, энкефалин желукретин, ХЦК, дочная нейротензин, железа субстанция Р, серотонин, гистамин Тонкая Секретин, Гастрин, ВИП кишка мотилин, ПП, серосеротонин тонин, ХЦК, мотилин, субстанция Р, гистамин Толстая СеротоВИП кишка нин (!), гастрин, ХЦК ЖелчГастрин, ВИП ный ХЦК, пузырь серотонин Печень Секретин, Глюкагон, ХЦК, гастрин ВИП Органы Примечания: (!) — очень сильное влияние на моторику. В скобках указано специфическое влияние гормона на секрецию отдельного вещества пищеварительного сока (например, НCl–). Сокращения: ХЦК — холецистокинин, ГИП — гастроингибирующий пептид; ВИП — вазоактивный интестинальный пептид; ПП — панкреатический пептид. 276 Физиология Во-первых, следует обратить внимание на число гормонов, регулирующих секрецию различных органов ЖКТ. Наибольшее число гормонов вырабатывается для усиления секреции пищеварительного сока поджелудочной железой, а гормонов, тормозящих его секрецию, вырабатывается мало, что биологически целесообразно: эта железа — главный орган, вырабатывающий ферменты для гидролиза пищевых веществ. Для желудка вырабатывается немного гормонов, стимулирующих его секрецию, — здесь только начинается пищеварение. Причем два из них стимулируют секрецию только пепсинов, что также биологически целесообразно, т.к. желудок не вырабатывает липазы и амилазы, — это удивительная «гармония». Но очень много вырабатывается гормонов, тормозящих секрецию желудка. Причем больше половины этих гормонов тормозят секрецию только HCl– (см. табл. 10.1). Это тоже биологически целесообразно, хотя НCl– — очень важное вещество для пищеварения и для выполнения непищеварительных функций. Дело в том, что его избыток может привести к заболеванию желудка (гиперацидный гастрит) и к нарушению пищеварения в двенадцатиперстной кишке (ДПК), т.к. панкреатические и кишечные ферменты оптимально активны в слабощелочной среде. Во-вторых, необходимо обратить внимание также на очень большое количество гормонов, стимулирующих моторику ЖКТ, особенно тонкой кишки, что также биологически целесообразно, т.к. застой химуса может привести к тяжелым последствиям, причем серотонин оказывает очень сильное стимуляторное влияние на толстую кишку, что также биологически целесообразно — предупреждение запоров. Состояние голода формирует поисковое и пищедобывательное поведение, а состояние насыщения прекращает прием пищи. В результате непрерывного расходования организмом питательных веществ формируется пищевая потребность — вызванное метаболическими процессами понижение уровня питательных веществ в крови. При этом возникает ощущение голода и мотивация к поиску пищи. Состояние голода включает две стадии. Первая стадия состояния голода — сенсорная (ощущение голода), она возникает еще при наличии в крови достаточного Глава 10. Система пищеварения 277 количества питательных веществ. Она формируется под влиянием нервных импульсов, поступающих в пищевой центр от механорецепторов пустого желудка и двенадцатиперстной кишки, мышечная стенка которых по мере эвакуации из них химуса приобретает все более повышенный тонус, что ведет к раздражению механорецепторов. Вторая стадия состояния голода — метаболическая, она начинается с момента снижения содержания питательных веществ в крови. Состояние насыщения, как и состояние голода, формируется в две стадии. Первая стадия — сенсорное насыщение (ощущение насыщения) — возникает во время приема пищи в результате потока афферентных импульсов от рецепторов языка, глотки, пищевода и желудка в центр насыщения, который реципрокно тормозит деятельность центра голода, что приводит к снижению ощущения голода. После приема достаточного количества пищи прекращается пищедобывательное поведение и потребление пищи. Сенсорное насыщение предупреждает поступление избыточного количества пищи в организм. Вторая стадия — стадия метаболического насыщения — наступает через 1,5–2 ч от начала приема пищи, когда в кровь начинают поступать питательные вещества. Достаточная концентрация питательных веществ в крови (мономеры — аминокислоты, моносахара — в основном глюкоза, жирные кислоты) улавливается непосредственно соответствующими рецепторами гипоталамуса и рецепторами сосудов, что обеспечивает поддержание ранее сформированного ощущения сытости. Далее рассмотрим функции различных отделов пищеварительной системы. 10.2. Пищеварение в полости рта. Акт глотания Обработка пищи в ротовой полости. Механическая обработка обеспечивается актом жевания — периодическими произвольными движениями нижней челюсти относительно неподвижной верхней. Жевание осуществляется жевательным аппаратом (верхняя и нижняя челюсти 278 Физиология с зубными рядами и мышцы, приводящие в движение нижнюю челюсть). Жевание обеспечивает: • измельчение пищи; • перемешивание ее со слюной, ослизнение и формирование пищевого комка, что облегчает последующее переваривание пищи; • стимуляцию выработки слюны; • моторную и секреторную деятельность ЖКТ. Химическая обработка пищи осуществляется слюной. У человека имеются три пары больших слюнных желез (околоушные, поднижнечелюстные, подъязычные) и многочисленные мелкие слюнные железы, располагающиеся в слизистой оболочке полости рта, языка и глотки. По строению и характеру секрета слюнные железы делят на две основные группы: серозные, т.е. околоушные, продуцирующие жидкую слюну с большим содержанием хлоридов, и смешанные, где большинство малых слюнных желез, поднижнечелюстные и подъязычные. Механизм секреции слюны. По механизму выделения секрета все слюнные железы относятся к мерокриновому типу, при котором секреторный продукт в виде молекул проникает через апикальную мембрану, не вызывая ее повреждений. Электрическая активность секреторных клеток характеризуется не деполяризацией, а гиперполяризацией клеточных мембран — возбуждающая гиперполяризация (ВГП, Гуткин В.И.). ПП секреторных клеток составляет 10–35 мВ. Во время ВГП мембранный потенциал достигает 40–50 мВ. ВГП секреторных клеток обусловлена работой йодной и хлорной помп, закачивающих анионы йода и хлора из крови в клетку. В клетку согласно электрохимическому градиенту поступают ионы Na+. За ионами в секреторную клетку поступает интерстициальная жидкость, что ведет к увеличению в ней гидростатического давления и выходу жидкости через поры апикальной мембраны. Слюна отделяется непрерывно вне приема пищи со скоростью 0,24 мл/мин, при жевании — 3–3,5 мл/мин; за сутки выделяется 0,5–2,0 л. Состав и свойства слюны. Органические вещества сухого остатка слюны включают альбумины, глобулины, ферменты, аминокислоты, мукосахариды, экскреторные азот- Глава 10. Система пищеварения 279 содержащие соединения небелковой природы — мочевину, аммиак, креатинин, мочевую кислоту, — это обусловлено экскреторной функцией слюнных желез. Неорганические вещества составляют 1/3 сухого остатка — это хлориды, бромиды, фториды, гидрокарбонаты, фосфаты, сульфаты, йодиды, Nа+, Са 2+, Mg2+, К+, микроэлементы — литий, железо, медь, никель. Функции слюны. Слюна обеспечивает формирование пищевого комка для проглатывания и создает условия для работы ферментов. Основные пищеварительные ферменты слюны — карбогидразы (α-амилаза и α-глюкозидаза, мальтаза). α-амилаза специфически расщепляет 1,4-глюкозидные связи в молекулах крахмала и гликогена, с образованием декстринов, а затем — мальтозы и глюкозы. Мальтаза расщепляет мальтозу и сахарозу до моносахаридов. В связи с краткостью пребывания пищи во рту (15–20 с) гидролиз углеводов под действием карбогидраз слюны происходит в основном внутри пищевого комка в желудке. Непищеварительные функции слюны: • защитная функция, которую выполняют протеолитические ферменты слюны. Нуклеазы слюны участвуют в деградации нуклеиновых кислот вирусов. Муромидаза (лизоцим) слюны вместе с иммуноглобулинами обладает выраженной бактерицидной активностью. Слюна покрывает слизистую оболочку полости рта и тем самым защищает ее от термических раздражителей, а также от кислых и щелочных продуктов; • слюна обеспечивает поступление ионов, необходимых для реминерализации эмали зубов, в частности фториды слюны проникают в эмаль зубов и защищают их от кариеса; • слюна способствует механическому очищению органов полости рта от остатков пищи, зубного налета и бактерий. Регуляция деятельности слюнных желез. Парасимпатические волокна к подчелюстной и подъязычной железам идут в составе барабанной струны (VII пара), к околоушным железам — в составе языкоглоточного нерва (IX пара). 280 Физиология Их возбуждение вызывает с помощью ацетилхолина и активации М1-холинорецепторов мембран секреторных клеток обильную секрецию жидкой слюны с высокой концентрацией солей и низким содержанием органических веществ («отмывная слюна»). Симпатические нервы к слюнным железам идут от II–V сегментов спинного мозга через верхний шейный симпатический ганглий. Их возбуждение ведет к активации β2-адренорецепторов секреторных клеток норадреналином, что сопровождается выделением небольшого количества густой слюны с низким содержанием солей и высокой концентрацией ферментов и муцина. Гуморальная регуляция деятельности слюнных желез существенной роли не играет. Фазы секреции слюнных желез — сложнорефлекторная и нейрогуморальная. Сложнорефлекторная фаза слюноотделения обусловлена комплексом безусловных и условных рефлексов (на вид и запах пищи), эта фаза регуляции главная. Акт глотания. Глотание обеспечивается согласованными последовательными сокращениями мышц глоточной области, гортани и пищевода, в результате чего пищевой комок переходит из ротовой полости в желудок. Выделяют три фазы глотания. Первая фаза глотания — ротовая (произвольная, быстрая). Импульсация от корковых нейронов инициирует сокращения мышц языка, благодаря которым верхушка языка поднимается, отделяет из пережеванной пищевой массы пищевой комок и переводит его на спинку языка. Наряду с этим эфферентные импульсы от корковых нейронов центра глотания обусловливают сокращения жевательных мышц, поднимающих нижнюю челюсть, что приводит к смыканию зубных рядов. В результате последующих произвольных сокращений мышц языка пищевой комок прижимается к твердому нёбу, продвигается кзади и перемещается на корень языка за передние (нёбноязычные) дужки зева. На этом заканчивается первая фаза глотания. Вторая фаза глотания — глоточная, непроизвольная, быстрая. Пищевой комок, проходя через зев, раздражает механорецепторы слизистой оболочки корня языка, передних и задних дужек, нёбной занавески и ротоглотки, в ре- Глава 10. Система пищеварения 281 зультате чего от них афферентные импульсы поступают по чувствительным волокнам тройничного, языкоглоточного и верхнего гортанного нервов в центр глотания продолговатого мозга и возбуждают его, что реципрокно тормозит центр дыхания. Одновременно эфферентные импульсы от центра глотания по волокнам тройничного, лицевого, языкоглоточного, блуждающего и подъязычного нервов поступают к группе мышц глоточной области и гортани, сокращение которых обеспечивает подъем мягкого нёба, гортани, языка и глотки, что предотвращает возможность поступления пищи в дыхательные пути. Вслед за этим расслабляется верхний пищеводный сфинктер. Далее последовательные сокращения верхнего, среднего и нижнего констрикторов глотки в виде перистальтической волны продвигают комок в пищевод, после чего верхний пищеводный сфинктер закрывается. Это предотвращает пищеводно-глоточный рефлюкс. Взаимодействие нейронов центра глотания между собой, а также центров дыхания и глотания генетически запрограммировано с помощью локальных жестких связей. Мышцы глотки верхней трети пищевода являются поперечно-полосатыми, они управляются соматическими нервами, идущими в составе языкоглоточного и блуждающего нервов. Синаптические контакты, как обычно, осуществляются с помощью мышечных N-холинорецепторов. Третья фаза глотания — пищеводная. Также непроизвольная, медленная, является продолжением глоточной фазы. При поступлении пищевого комка в начальную часть пищевода раздражаются его механорецепторы и возникает рефлекторная перистальтическая волна, обеспечивающая продвижение пищевого комка по пищеводу за счет распространяющегося сокращения циркулярных мышц в области пищевого комка, которому предшествует волна расслабления кольцевых мышц дистальнее пищевого комка. Регуляция продвижения пищевого комка по гладкомышечной части пищевода осуществляется благодаря взаимодействию блуждающего нерва с интраорганной нервной системой. Причем предшествующая волна расслабления кольцевых мышц пищевода, в том числе и нижнего сфинктера, обеспечивается блуждающим нервом с помощью интраорганных ВИП-нейронов. 282 Физиология Перистальтическая волна сокращения в направлении расслабленного нижнего сфинктера обеспечивает переход пищевого комка в желудок, вследствие чего возникает ваго-вагальный рефлекс, ведущий к закрытию сфинктера пищевода. Иногда, вследствие недостаточного закрытия нижнего сфинктера пищевода, в его полость попадает кислое содержимое желудка, вызывающее изжогу. Если после первой перистальтической волны в пищеводе осталась часть пищи, возникает вторая и последующие рефлекторные ваго-вагальные перистальтические волны (в области, где осталась пища), пока вся пища не перейдет из пищевода в желудок. В заключение отметим, что в нижних двух третях пищевода перистальтическая волна может осуществляться за счет рефлексов интраорганной нервной системы после перерезки блуждающих нервов. Роль симпатической нервной системы в регуляции процесса глотания изучена недостаточно. 10.3. Пищеварение в желудке Желудок — это орган депо пищи, он участвует в ее переваривании и обеспечивает поступление содержимого в тонкую кишку. Желудок натощак находится в спавшемся состоянии и содержит около 50 мл сока, близкого к нейтральной реакции. При обычном пищевом режиме желудок вырабатывает 2 л сока в сутки (рис. 10.1). Отделение желудочного сока начинается через 5–10 мин после начала приема пищи, продолжается в течение нескольких часов после прекращения акта еды и зависит от вида и количества принятой пищи. Секреторная функция желудка Состав желудочного сока — это бесцветная прозрачная жидкость с удельным весом 1,002–1,007. Она имеет очень кислую реакцию (рН 1,5) благодаря высокому содержанию соляной кислоты (0,3–0,5%), состоит из воды (99–99,5%) и плотных веществ (1–0,5%). Ферменты желудочного сока. Протеазы желудка — это пепсиногены, активация которых запускается соляной кислотой, а в дальнейшем протекает аутокаталитически под действием уже образовавшихся пепсинов. В белковых мо- Глава 10. Система пищеварения 283 лекулах расщепляются пептидные связи, в результате чего образуются полипептиды. Имеется несколько групп пепсинов, активность которых наибольшая при определенной величине рН, но все пепсиногены активируются при рН < 3,0. Липаза и амилаза в желудке не вырабатываются. Углеводы в центре содержимого перевариваются амилазами слюны, пока они не нейтрализуются HCl. Вся масса пищи в желудке не смешивается с желудочным соком, ее пищеварение происходит в зоне непосредственного контакта пищевого содержимого со слизистой оболочкой («слизывает, как мороженое эскимо»), а в центре — с ферментами слюны. Функции соляной кислоты: • вызывает денатурацию и набухание белков, способствуя их гидролитическому расщеплению; • активирует пепсиногены и создает в желудке кислую среду, оптимальную для действия протеолитических ферментов; • стимулирует образование гастроинтестинальных гормонов (гастрина, секретина); • принимает участие в регуляции эвакуации содержимого желудка в двенадцатиперстную кишку; • обеспечивает антибактериальное действие желудочного сока. Функции желудочной слизи (муцина). Муцин создает слизистый барьер желудка, защищает от действия соляной кислоты и самопереваривания. Нарушению защитной функции слизистого барьера и усилению секреции соляной кислоты в желудке способствует деятельность микроорганизмов Helicobacter pylori, в результате чего может развиваться язвенная болезнь. Слизистый барьер желудка может нарушаться под действием алкоголя, высокой концентрации соляной кислоты. Гастромукопротеид (внутренний фактор Касла) связывает в желудке витамин В12 (внешний фактор кроветворения, необходимый для синтеза глобина), поступающий с пищей, и предохраняет его от расщепления пищеварительными ферментами, тем самым слизь участвует в кроветворении. В тонкой кишке комплекс «гастромукопротеид — витамин В12» взаимодействует со специфическими рецепторами, после чего витамин В12 всасывается в кровь и с помощью транспортных белков 284 Физиология переносится к печени и костному мозгу. Отсутствие этого фактора приводит к развитию заболевания — железодефицитной анемии. Нервная регуляция желудочной секреции. Блуждающий нерв стимулирует секреторную деятельность главных, обкладочных и мукоидных клеток фундальных желез, активируя М-холинорецепторы. Симпатические нервы оказывают преимущественно тормозное действие на секреторную деятельность желудка, однако они способствуют накоплению пепсиногенов и слизи в соответствующих видах гландулоцитов. Эффекты реализуются с помощью α- и β-адренорецепторов. Гуморальная регуляция (см. табл. 10.1). Продукты гидролиза белков (после всасывания их в кровь) стимулируют секрецию желудка посредством увеличения выработки интестинальных гормонов. Фазы желудочной секреции: • сложнорефлекторная (мозговая); • нейрогуморальная. Сложнорефлекторная фаза осуществляется с помощью комплекса условных и безусловных рефлексов на вид и запах пищи и вследствие раздражения пищей рецепторов слизистой оболочки рта, глотки и пищевода. Этот сок богат ферментами, его отделение сопровождается возникновением аппетита, И.П. Павлов назвал его «запальным». Данную фазу демонстрирует опыт «мнимого кормления», проведенный на эзофаготомированной собаке с фистулой желудка, когда проглоченная пища выпадает из перерезанного пищевода, не поступая в желудок (рис. 10.3). Рис. 10.3. Опыт И.П. Павлова — «мнимое кормление» Глава 10. Система пищеварения 285 Через 5–10 мин после начала «мнимого кормления» наблюдается обильная секреция желудочного сока с высоким содержанием ферментов. Эта секреция продолжается до 2 ч. Нейрогуморальная фаза наслаивается на мозговую. Ее условно делят на желудочную и кишечную. Поступление желудочного содержимого в двенадцатиперстную кишку стимулирует желудочную секрецию с помощью уже рассмотренных нервно-гуморальных механизмов. При этом G-клетками двенадцатиперстной кишки дополнительно выделяется гастрин. Продукты гидролиза жиров, образующиеся в двенадцатиперстной кишке, сдерживают чрезмерную секрецию. Моторная функция желудка. Исходное состояние для всех видов сокращений желудка — пластический тонус. В начале приема пищи наблюдается рецептивная релаксация (рефлекторное расслабление мышц фундального отдела желудка, обеспечивающее увеличение его объема и прием поступающей пищи). Релаксация осуществляется с помощью блуждающих нервов, эффект реализуется с помощью ВИПергических нейронов. В зависимости от химического состава и количества принятой пищи она может задерживаться в желудке от 3 до 10 ч. Спустя 5–30 мин после начала приема пищи начинаются сокращения желудка в области малой кривизны вблизи кардии, где локализован пейсмекер — водитель ритма (Кромин А.А.). Второй пейсмекер находится в пилорической части желудка. Виды сокращений наполненного пищей желудка. Перистальтические сокращения (рис. 10.4) — сокращения циркуляторных мышц желудка разной амплитуды, распространяющиеся в дистальном направлении с частотой 3 цикла в минуту благодаря последовательному сокращению зон по окружности желудка и расслаблению ранее сокращенных участков. При этом в кишку переходит небольшая порция, основная часть содержимого смещается в обратном направлении, в результате чего происходят периодические смещения содержимого — перемешивание, перетирание его и гомогенизация. Тонические волны — тоже разной амплитуды. Они могут возникать независимо от перистальтических волн 286 Физиология мм рт. ст. 1 80 2 3 60 40 20 0 1 мин Рис. 10.4. Перистальтические сокращения наполненного пищей желудка человека: 1 — низкоамплитудные; 2 — среднеамплитудные; 3 — высокоамплитудные (сокращения на фоне тонической волны) и продвигают все содержимое желудка из фундального в антральный его отдел. Пластический тонус обеспечивает постоянный контакт содержимого со стенкой желудка, где непосредственно секретируются ферменты, — и в случае уменьшения и при увеличении его наполнения, что улучшает переваривание пищи и способствует переходу содержимого в двенадцатиперстную кишку. Нервная регуляция моторики желудка. Блуждающие нервы с помощью ацетилхолина стимулируют сокращения желудка (рис. 10.5); симпатические (адренергические) — их тормозят. В составе чревных нервов содержатся также серотонинергические волокна, усиливающие моторику желудка (рис. 10.6). Симпатические нервы с помощью α- и β-адренорецепторов повышают тонус пилорического сфинктера; считают, что активация ВИПергических нейронов ведет к его снижению. Корковые влияния реализуются с помощью условных и безусловных рефлексов посредством гипоталамуса, центра пищеварения в продолговатом мозге, симпатического и блуждающего нервов. Рис. 10.5. Усиление сокращений желудка (регистрация гидростатического давления в его полости) при раздражении блуждающего нерва у собаки (отметка — жирная часть линии 30 с, шкала 20 мм рт. ст.) Глава 10. Система пищеварения 287 А Б Рис. 10.6. Реакция желудка у собаки (изменения давления в его полости) на раздражение симпатического ствола в условиях действия различных фармакологических препаратов. На каждом фрагменте — запись давления в полости желудка и нулевая линия, она же (более жирная часть линии) отметка раздражения — 30 с, шкала 20 мм рт. ст. В составе симпатического нерва имеются серотонинергические волокна, усиливающие сокращения желудка. Опыт В.М. Смирнова, И.Л. Мясникова, Д.С. Свешникова: А — до введения фармакологических препаратов; Б — на фоне действия α- и β-адреноблокаторов и лизергола — блокатора серотониновых рецепторов Отрицательные эмоции, возникающие при виде и запахе плохо приготовленной пищи, тормозят, а разговор о вкусной еде усиливает моторику желудка. Местная нервная регуляция осуществляется с помощью возбуждающих и тормозных интраорганных нейронов. Моторика желудка нарушается при воспалительных процессах в толстой кишке. Гуморальная регуляция моторики желудка (см. табл. 10.1). Ряд факторов оказывает существенное влияние на скорость перехода содержимого желудка в двенадцатиперстную кишку: • консистенция — жидкая часть пищевого содержимого подвергается быстрой эвакуации, а твердая часть надолго задерживается в желудке, пока не будет измельчена до частиц размерами менее 2 мм. Средняя порция смешанной пищи эвакуируется из желудка за 4 ч; • состав пищи — быстрее всего эвакуируется из желудка пища, богатая углеводами, медленнее — белковая, еще медленнее — жирная пища; • объем пищи — увеличение объема твердой пищи в желудке ускоряет ее эвакуацию из желудка вследствие усиления раздражения механорецепторов желудка; • раздражение механорецепторов порцией химуса двенадцатиперстной кишки вызывает закрытие 288 Физиология сфинктера, тормозит эвакуацию, но стимулирует сокращения двенадцатиперстной кишки, готовя ее для принятия следующей порции содержимого желудка; • осмотическое давление — тормозное влияние на моторно-эвакуаторную функцию желудка оказывают гипертонические растворы, продукты гидролиза белков и глюкоза, ГИП, энкефалины; • pН среды — действие кислого химуса на хеморецепторы желудка рефлекторно ускоряет, на рецепторы двенадцатиперстной кишки — замедляет эвакуацию содержимого желудка. 10.4. Пищеварение в двенадцатиперстной кишке Двенадцатиперстная кишка — это центральный узел пищеварительной системы (сюда поступают панкреатический и кишечный соки с полным набором ферментов для гидролиза белков, жиров и углеводов, а также желчь, играющая важную роль в переваривании и всасывании жиров). Нервная регуляция сокращений двенадцатиперстной кишки представлена на рис. 10.7 и 10.8; гуморальная регуляция (тонкой кишки) — см. табл. 10.1. pН дуоденального химуса в процессе пищеварения колеблется в пределах 4–8. По мере продвижения кислого желудочного содержимого, попавшего в двенадцатиперстную кишку, происходит его нейтрализация в результате перемешивания с щелочными названными секретами, главный из них — сок поджелудочной железы. За сутки у человека выделяется 1,5– 2,5 л поджелудочного сока, основная его особенность — высокая концентрация в нем бикарбонатов, в 5 раз превышающая их концентрацию в плазме крови, что определяет его щелочные свойства и создает оптимум рН для активности ферментов, поступающих сюда из поджелудочной железы. Ферменты поджелудочной железы Протеазы (трипсин, химотрипсин, эластаза) расщепляют белковые молекулы до олигопептидов. Эластаза особенно эффективно гидролизует белки соединительной ткани. Карбоксипептидазы (А и В) расщепляют олигопептиды до аминокислот. Конечные продукты действия всех пепти- Глава 10. Система пищеварения 289 Рис. 10.7. Усиление сокращений двенадцатиперстной кишки (регистрация гидростатического давления в ее полости) при раздражении блуждающего нерва у собаки (отметка раздражения — жирная часть линии 30 с, шкала 20 мм рт. ст.) Рис. 10.8. Усиление сокращений двенадцатиперстной кишки (увеличение давления в ее полости) с помощью серотонинергических волокон при раздражении правого симпатического ствола в грудной полости у собаки (А) и угнетение — посредством адренергических волокон после выключения серотонинергических (Б). Опыт В.М. Смирнова, Д.С. Свешникова, И.Л. Мясникова, шкала 20 мм рт. ст., отметка раздражения — жирная часть линии (30 с) даз в двенадцатиперстной кишке — олигопептиды (70%) и аминокислоты (30%). Все протеазы выделяются в виде проферментов. Трипсиноген активируется энтерокиназой двенадцатиперстной кишки. Ее освобождение связано с действием желчных кислот. Далее трипсин активирует трипсиноген и другие протеолитические ферменты. Трипсин активирует также процесс освобождения энтерокиназы. Рибо- и дезоксирибонуклеазы, продуцируемые в активном состоянии, расщепляют РНК и ДНК до нуклеотидов. α-амилаза продуцируется в активном состоянии, расщепляет крахмал до декстринов (их образуется мало), мальтозы и мальтотриозы. 290 Физиология Панкреатическая липаза секретируется в активной форме и расщепляет жиры, эмульгированные желчными кислотами и их солями в мельчайшие капельки жира, что ведет к увеличению общей площади капелек жира и обеспечивает доступность к действию фермента. В результате гидролиза триглицеридов (жиров) образуются моноглицериды и свободные жирные кислоты, причем уже в середине двенадцатиперстной кишки жир на 80% оказывается гидролизованным. Фосфолипаза А2 секретируется в виде профермента, активируется трипсином, гидролизует фосфолипиды. Фазы панкреатической секреции и их регуляция — сложнорефлекторная и нейрогуморальная — желудочная и кишечная, основная из них — кишечная. Главный секреторный нерв поджелудочной железы, как показали исследования И.П. Павлова, — блуждающий нерв. Латентный период сокоотделения не превышает 2–3 мин от начала приема пищи. Симпатические волокна, активируя β-адренорецепторы, оказывают тормозное влияние на отделение поджелудочного сока, но усиливают синтез органических веществ в панкреацитах. Активация α-адренорецепторов вследствие сужения кровеносных сосудов угнетает отделение панкреатического сока. Гуморальная регуляция секреции поджелудочной железы (см. табл. 10.1). Важную роль в дуоденальном пищеварении играет печень. 10.5. Физиология печени Печень выполняет пищеварительные (с помощью желчи) и непищеварительные функции. Характеристика желчи и ее образование. Желчь представляет собой золотистую жидкость, изотоничную плазме крови, рН — 7,3–8,0. Она содержит холевую, хенодезоксихолевую кислоту, желчные пигменты, холестерин, неорганические соли, мыла, жирные кислоты, нейтральные жиры, лецитин, мочевину, витамины А, В, С, в небольшом количестве некоторые ферменты (амилазу, фосфатазу, протеазу, каталазу, оксидазу). В сутки образуется примерно 0,6–1,5 л желчи. Образование желчи идет непрерывно, даже при голодании. Блуждающие нервы стимулируют, симпатические Глава 10. Система пищеварения 291 угнетают образование желчи. Прием пищи рефлекторно (с интерорецепторов желудка, кишечника) увеличивает образование желчи через 3–12 мин. Сильные стимуляторы желчеобразования — это яичные желтки, хлеб, молоко, мясо. Эффективно стимулируют желчеобразование сами желчные кислоты, секретин. Желчь вне пищеварения скапливается в желчном пузыре, что обеспечивается закрытием сфинктера общего желчного протока (Одди). Желчевыведение. Нервная регуляция желчевыведения. Блуждающие нервы повышают сократительную активность мышц желчного пузыря, одновременно расслабляют сфинктер Одди, что обеспечивает изгнание желчи в двенадцатиперстную кишку через 7–10 мин после приема пищи. Наибольшее количество желчи выводится при потреблении яичных желтков, жиров, молока, мяса. Рефлексогенные зоны — вкусовые рецепторы слизистой оболочки полости рта, механорецепторы желудка. Симпатические нервы снижают сократительную активность мышц желчного пузыря и желчевыводящих протоков, одновременно усиливая сокращения мышц сфинктера Одди, что способствует накоплению желчи в желчном пузыре. Гуморальная регуляция выведения желчи. Пища, попадая в желудок и двенадцатиперстную кишку, стимулирует выработку гастроинтестинальных гормонов (см. табл. 10.1). Пищеварительные функции желчи: • главная пищеварительная функция желчи — переваривание жиров. Соли желчных кислот обладают мощным эмульгирующим действием на жиры, образуя все более мелкие капельки, подвергающиеся в конечном итоге гидролизу в тонкой кишке под действием липазы панкреатического сока. Желчь ускоряет всасывание жирных кислот и жирорастворимых витаминов D, E, K; • желчь обеспечивает смену желудочного пищеварения на кишечное (Павлов И.П.), что реализуется посредством нейтрализации соляной кислоты желудочного содержимого; • желчь стимулирует моторику кишечника и движения кишечных ворсинок; • угнетает развитие кишечной флоры и предупреждает гнилостные процессы в толстой кишке. 292 Физиология Непищеварительные функции печени: • в печени инактивируются протеиназами белковопептидные гормоны, стероидные (гидроксилазами), дезаминируются катехоламины при участии моноамиоксидазы; • печень участвует в разрушении эритроцитов, биохимических трансформациях гема с образованием желчных пигментов; • защитная функция осуществляется посредством участия печени в иммунных реакциях организма и дезинтоксикации (обезвреживании) токсических веществ, поступающих в кровь из кишечника (индол, фенол, скатол), а также чужеродных веществ. Дезинтоксикация аммиака, образующегося в печени в реакциях дезаминирования продуктов белкового обмена, осуществляется посредством синтеза мочевины, выделяемой в последующем почками; • в печени синтезируются почти все альбумины, подавляющее большинство фракций глобулинов, фибриногена (100%), протромбина, других факторов, обеспечивающих процессы гемостаза и противосвертывания крови; • печень — депо гликогена, крови, белков, жиров, микроэлементов, витаминов (A, D1, D2, K, C, PP); • в гепатоцитах происходит расщепление жира с образованием жирных кислот. 10.6. Пищеварение в тонкой кишке Переваривание химуса. Характеристика кишечного сока. За сутки отделяется 2,5 л кишечного сока, он состоит из плотной и жидкой частей. Плотная часть кишечного сока — это слизистые комочки распадающихся эпителиальных клеток, слизи. Слизь предохраняет кишку от повреждений, ферменты содержатся в основном в плотной части. Жидкая часть кишечного сока также содержит много ферментов. Величина рН в верхнем отделе тонкой кишки составляет 7,2– 8,6. В тонкой кишке имеется свыше 20 ферментов. Протеазы и пептидазы. Энтерокиназа, хорошо растворимая в воде, активирует в полости кишки все проте- Глава 10. Система пищеварения 293 азы поджелудочного сока. Катепсины (от греч. kathepsin — переваривать) — тканевые протеазы, внутриклеточные ферменты. В кишечнике гидролизуют белки десквамированного эпителия и пептиды. В тонкой кишке содержатся нуклеазы, деполимеризующие нуклеиновые кислоты. Липазы тонкой кишки гидролизуют липиды, моноэфиры ортофосфорной кислоты, фосфолипиды самого кишечного сока, холестерин. Амилазы тонкой кишки высокоактивны, осуществляют заключительный гидролиз углеводов: поли- и олигосахаридов — до мальтозы и глюкозы, сахарозы — до глюкозы и фруктозы, лактозы — до глюкозы и галактозы, мальтозы и трегалозы — до глюкозы. В тонкой кишке пищеварение происходит в три этапа. Первый этап — полостное пищеварение, оно обеспечивает гидролиз до 50% углеводов и всего лишь 10% белков, причем до крупных молекул и надмолекулярных агрегаций, в результате чего образуются в основном олигомеры. Второй этап — пристеночное пищеварение (слизистное и гликокаликсное). В слое слизистых наложений сорбировано большое количество ферментов кишечного и поджелудочного пищеварительных соков. Сюда поступают олигомеры из полости кишки и начинают образовываться димеры. В гликокаликсе, представляющем собой скопления мукополисахаридных нитей, находятся панкреатические и кишечные ферменты, также перешедшие из полости тонкой кишки (см. рис. 10.6). Здесь продолжается гидролиз олигомеров, приводящий к образованию в основном димеров. Третий этап — мембранное пищеварение, где образуются мономеры. Его обеспечивают ферменты, представляющие собой интегральные белки клеточной мембраны с каталитическим центром, направленным в полость кишки. На апикальной поверхности каждого энтероцита находится до трех тысяч микроворсинок, благодаря чему площадь, на которой происходит мембранное пищеварение, огромна, что превращает микроворсинки в катализаторы. Скорость мембранного пищеварения возрастает еще и потому, что оно сопряжено с транспортом мономеров через энтероцит в кровь или лимфу, поскольку здесь же располагаются транспортные системы. С помощью мембранного пищева- 294 Физиология рения гидролизуются до 80–90% пептидных и гликозидных связей. Регуляция кишечной секреции осуществляется в основном местными механизмами, воздействие продуктов переваривания на слизистую кишки вызывает отделение кишечного сока, богатого ферментами. Возбуждение блуждающего нерва увеличивает секрецию ферментов тонкой кишкой, не влияя на количество отделяемого сока. Возбуждение симпатического нерва, напротив, угнетает кишечную секрецию. Гуморальная регуляция (см. табл. 10.1). Моторика тонкой кишки. Виды сокращений тонкой кишки. Перистальтические сокращения — это волнообразно распространяющиеся по кишке сокращения циркулярных мышц, которым предшествует волна расслабления со скоростью 1–2 см/с. Начинаются они обычно в двенадцатиперстной кишке в момент поступления в нее желудочного содержимого и обеспечивают продвижение химуса по кишке в дистальном направлении. Ритмическая сегментация заключается в одновременном сокращении циркулярных мышц в нескольких соседних участках кишки, разделяющих ее на сегменты, с последующим их расслаблением и сокращением циркулярных мышц других участков кишки, что обеспечивает перемешивание химуса кишки и более эффективное полостное переваривание. Маятникообразные сокращения — это ритмические сокращения, главным образом продольных мышц при участии циркулярных, приводящих к перемещению химуса вперед-назад, что также обеспечивает перемешивание химуса и улучшение гидролиза. Микродвижения кишечных ворсинок также способствуют перемешиванию химуса и всасыванию продуктов гидролиза. Все виды названных сокращений накладываются на тонические сокращения, являющиеся локальными или медленно перемещающимися. Базальное давление в полости тонкой кишки составляет 8–9 см вод. ст., величина его существенно возрастает при фазных сокращениях мышц любого вида. Регуляция сокращений тонкой кишки. Блуждающие нервы с помощью холинергических нейронов оказывают стимулирующее влияние на тонус и амплитуду перистальтических Глава 10. Система пищеварения 295 сокращений тонкой кишки; посредством ВИПергических нейронов — тормозное (АТФ в ВИП-нейронах — модулятор). Симпатический нерв с помощью норадреналина угнетает моторику кишки (см. рис. 10.8) в основном посредством торможения холиэнергетических влияний, а также и непосредственного действия на миоциты. В составе чревных нервов содержатся серотонинергические волокна, оканчивающиеся непосредственно на миоцитах и стимулирующие моторику тонкой кишки (см. рис. 10.8). Экстраорганные нервы взаимодействуют с интраорганной нервной системой, которая представляет собой совокупность сенсорных нейронов, интернейронов, эндогенных осцилляторов, тонических и эфферентных нейронов, придающих ей черты истинной автономии (Ноздрачев А.Д.). Гуморальная регуляция (см. табл. 10.1). Основной закон рефлекторной регуляции моторики деятельности ЖКТ заключается в том, что адекватное раздражение любого участка ЖКТ (переполнение его химусом) вызывает усиление моторно-эвакуаторной деятельности его нижележащих участков и одновременно торможение вышерасположенных отделов, что обеспечивает оптимальное продвижение содержимого и химуса органов ЖКТ. 10.7. Всасывание питательных веществ Характеристика всасывания в различных отделах ЖКТ. В ротовой полости всасывание незначительно ввиду кратковременности пребывания в ней пищи. Всасывающая способность слизистой оболочки полости рта отчетливо проявляется при помещении некоторых веществ, в том числе лекарственных, под язык, что используется в клинической практике. В желудке всасываются вода и растворимые в ней минеральные соли, глюкоза, алкоголь. В тонкой кишке всасываются быстро все питательные вещества — это основное всасывание. Уже через 1–2 мин после поступления химуса в кишку питательные вещества появляются в крови, а через 5–10 мин их концентрация в крови достигает максимальных значений. Всасывательная 296 Физиология поверхность слизистой оболочки тонкой кишки очень велика за счет круговых складок, ворсинок и микроворсинок и достигает 200 м2. Характерная особенность микроциркуляторного русла ворсинок — это высокая степень фенестрирования эндотелия капилляров и большой размер пор. После приема пищи кровоток в ворсинках возрастает на 30–130%. Всасыванию способствуют ритмические сокращения и расслабление ворсинок, они улучшают контакт их поверхности с химусом и обеспечивает выдавливание крови и лимфы с питательными веществами в общий кровоток. Механизмы всасывания отдельных веществ. Аминокислоты всасываются в основном с помощью натрийзависимого транспорта, частично — посредством диффузии. Основная масса аминокислот всасывается в двенадцатиперстной и тощей кишках (до 80–90%), и только 10% достигает толстой кишки. Эндоцитоз обеспечивает транспорт иммуноглобулинов, ферментов, витаминов, в небольших количествах — интактных белковых молекул. Продукты гидролиза жиров всасываются в основном в двенадцатиперстной и тощей кишках. Они образуют мицеллы с желчными кислотами, которые поступают на мембраны энтероцитов. Липидные компоненты мицелл растворяются в клеточной мембране и согласно концентрационному градиенту поступают в цитоплазму энтероцитов. Желчные кислоты мицелл, остающиеся в полости кишки, поступают в подвздошную кишку, где подвергаются всасыванию по механизму первичного транспорта. На эндоплазматическом ретикулуме энтероцитов происходит ресинтез триглицеридов из жирных кислот и моноглицеридов. Из образованных триглицеридов, холестерина, фосфолипидов и гликопротеинов образуются хиломикроны — мельчайшие жировые частицы, заключенные в тончайшую белковую оболочку. Хиломикроны сливаются с латеральной мембраной энтероцита и через образующееся при этом отверстие выходят в межклеточное пространство, откуда по центральному лимфатическому и грудному протокам поступают в кровь. Вместе с жирами всасываются растворимые в жирах витамины (А, D, Е, К). С калом теряется до 5–7 г жира в сутки. Глюкоза и галактоза всасываются в основном с помощью натрийзависимого транспорта. Моносахариды могут Глава 10. Система пищеварения 297 всасываться путем простой и облегченной диффузии в случае их высокой концентрации в просвете кишечника. Всасывание воды осуществляется с помощью фильтрации и осмоса и наиболее интенсивно происходит в тонкой и толстой кишках (за сутки около 9 л, из них 1,5 л — в толстой кишке). При повышении гидростатического давления в полости ЖКТ всасывание воды возрастает. Всасывание минеральных солей осуществляется с помощью диффузии и Nа-зависимого транспорта. 10.8. Пищеварение в толстой кишке В толстой кишке завершаются переваривание и всасывание, формируются каловые массы, синтезируются некоторые витамины. В гидролизе принимают участие ферменты тонкой кишки и микроорганизмов. Ферментативная активность сока толстой кишки значительно ниже, чем в тонкой кишке, — в ней нет энтерокиназы и сахаразы, а содержание щелочной фосфатазы в 15–20 раз ниже, чем в тонкой кишке, что физиологически целесообразно, поскольку химус, поступающий в толстую кишку, содержит мало непереваренных пищевых веществ. Регуляция сокоотделения в толстой кишке осуществляется в основном местными механизмами. По мере продвижения по толстой кишке химус становится более плотным за счет деятельности бактерий и всасывания воды (1–1,5 л/сут), в результате чего формируются каловые массы. Микрофлора кишечника локализуется в основном в толстой кишке (30–50% кала приходится на долю бактерий). В проксимальном направлении количество микробов становится меньше — этому препятствует моторика пищеварительного тракта (движение химуса в каудальном направлении). Нормальная микрофлора желудочно-кишечного тракта — необходимое условие жизнедеятельности организма. Мало микроорганизмов в верхних отделах тонкой кишки, а желудок практически стерилен, что объясняется губительным воздействием на бактерии соляной кислоты. У новорожденного содержимое толстой кишки стерильно. В течение нескольких месяцев жизни 298 Физиология этот отдел пищеварительного тракта заселяется микрофлорой. Значение микробов организма: • расщепляют содержащиеся в толстой кишке остатки пищевых веществ; • расщепляют растительные волокна, которые не перевариваются амилазами в тонкой кишке; • защищают организм от размножения в нем патогенных микробов — облигатная микрофлора обладает выраженной антагонистической активностью по отношению к патогенным бактериям; • синтезируют витамины К и группы В (В1, В6, В12), которые всасываются в толстой кишке и частично усваиваются организмом; • продуцируют биологически активные вещества, оказывающие влияние на тонус кишечной стенки и всасывание воды и аминокислот; • инактивируют пищеварительные ферменты (частично они разрушаются и реабсорбируются в тонкой кишке). Переход кишечного химуса в толстую кишку происходит через илеоцекальный сфинктер — мышечные утолщения (баугиниева заслонка) и попадает в слепую кишку маленькими порциями (струйками) вследствие периодического (6–8 циклов в минуту) расслабления и последующего сокращения его мышц в ритме сокращений подвздошной кишки. Сокращения начинаются через 1–4 мин после приема пищи. Сфинктер выполняет функцию клапана — пропускает химус кишечника только в одном направлении. За сутки из тонкой в толстую кишку переходит 1–4 л химуса, который начинает поступать в толстую кишку через 3–3,5 ч после приема пищи. Моторика толстой кишки. Двигательная активность толстой кишки, как и тонкой, обеспечивается автоматией гладкомышечных клеток. Перистальтические волны возникают редко, существенной роли в продвижении кишечного содержимого не играют. Маятникообразные сокращения обеспечивают перемешивание содержимого и способствуют всасыванию воды. Антиперистальтические сокращения приводят к созданию дистально-проксимального гради- Глава 10. Система пищеварения 299 ента давления и ретроградного перемещения содержимого, что способствует всасыванию воды. Пропульсивные сокращения (масс-сокращения) обеспечивают быстрое продвижение содержимого на большое расстояние — из поперечной кишки в сигмовидную и прямую кишку. Они возникают 1–3 раза в сутки после еды. Регуляция моторики толстой кишки. Парасимпатические нервы повышают, а симпатические понижают амплитуду и частоту сокращений толстой кишки. Блуждающий нерв иннервирует толстую кишку до сигмовидной, тазовый — сигмовидную и прямую кишку. Интраорганная нервная система тормозит миогенную ритмику посредством ВИП-нейронов. Моторика толстой кишки усиливается во время еды посредством условных и безусловных рефлексов. Раздражение механорецепторов толстой кишки вызывает усиление ее моторной активности с помощью местных рефлексов. Раздражение механорецепторов прямой кишки при ее переполнении рефлекторно тормозит сокращения вышележащих отделов толстой и тонкой кишок, что ограничивает дальнейшее поступление каловых масс в ее полость. Роль гуморальных веществ (см. табл. 10.1). Весь процесс пищеварения у взрослого здорового человека длится около 1–3 сут, из которых наибольшая часть времени приходится на передвижение остатков пищи по толстой кишке. Акт дефекации как составная часть моторики толстой кишки. Проход из прямой кишки закрыт внутренним гладкомышечным и наружным (исчерченные мышцы) сфинктерами. Они открываются только во время дефекации. Симпатические нервы (LI–LIV) повышают тонус внутреннего сфинктера и тормозят моторику прямой кишки, что способствует ее заполнению каловыми массами. Наружный сфинктер (произвольный) иннервируется соматическими мотонейронами (SII–SIV). Его тонус поддерживается рефлекторно — импульсацией, поступающей с экстерорецепторов кожи области анального отверстия и проприорецепторов сфинктера по тазовому и срамному нервам в ЦНС. Афферентные пути — тазовый и срамной нервы. Далее импульсация по спинному мозгу проводится в продолго- 300 Физиология ватый мозг, лимбические структуры, кору большого мозга, в результате чего возникает позыв к дефекации. Когда каловые массы поступают в прямую кишку, стенки ее растягиваются, что приводит к расслаблению внутреннего анального сфинктера (интраорганный рефлекс) и повышению тонуса наружного сфинктера (соматический рефлекс). Позыв к дефекации возникает нередко после еды вследствие пропульсивных движений толстой кишки и поступления каловых масс в прямую кишку. В процессе дефекации наружный сфинктер расслабляется произвольно, внутренний — рефлекторно с помощью парасимпатических нервов (SII–SIV). Кроме того, сокращаются также мышцы диафрагмы и брюшной стенки, что ведет к повышению внутрибрюшного давления до 16 мм рт. ст., сокращаются также гладкие мышцы дистальной части толстой кишки. Совокупность этих эффектов приводит к дефекации. При повреждении спинного мозга выше SII рефлекс дефекации сохраняется, но произвольная дефекация исключена. Разрушение SII–SIV-сегментов устраняет и рефлекс дефекации. ГЛАВА 11 Здоровый нищий счастливее больного короля. А. Шопенгауэр Обмен и терморегуляция 11.1. Обмен веществ и питание Обмен веществ — это совокупность процессов поступления питательных веществ в организм, использования их организмом для синтеза клеточных структур и выработки энергии, а также выделения конечных продуктов распада в окружающую среду. Ассимиляция — совокупность процессов, обеспечивающих поступление питательных веществ во внутреннюю среду организма и использование их для синтеза клеточных структур и секретов клеток, в результате чего накапливается энергия в организме. Пищеварение — первый этап ассимиляции (гидролиз белков, жиров и углеводов пищи). Конечные продукты гидролиза белков в пищеварительном тракте — это аминокислоты, нуклеотиды, углеводы, т.е. моносахариды, жиров — жирные кислоты, моноглицериды. Долю питательных веществ, поступивших из пищеварительного тракта во внутреннюю среду организма (около 90%), называют усвояемостью питательных веществ. При гидролизе в пищеварительном тракте образуются мономеры, практически не теряющие своей энергетической ценности (высвобождается лишь около 1% заключенной в пище энергии). Анаболизм — заключительная часть ассимиляции (совокупность внутриклеточных процессов, обеспечивающих синтез структур и секретов клеток организма). Исходные продукты анаболизма — мономеры (аминокислоты, моносахариды, жирные кислоты, моноглицериды, нуклеотиды), а также вода, минеральные соли и витамины; конечные — полимеры: специфические белки, жиры, углеводы, 302 Физиология нуклеиновые кислоты организма. Анаболизм обеспечивает рост развивающихся организмов, восстановление (обновление) распавшихся в процессе диссимиляции клеточных структур. Диссимиляция — процесс расщепления клеточных структур до мономеров и других соединений без высвобождения энергии. Продукты диссимиляции попадают в кровь и повторно используются клетками в процессах анаболизма вместе с мономерами, поступающими в кровь из ЖКТ, и формирующими общий фонд питательных веществ, циркулирующих в крови. Катаболизм — это процесс расщепления (сгорания) питательных веществ (мономеров и других соединений, попадающих в клетку из крови) до конечных продуктов (Н2О, СО2 и NН3) с высвобождением энергии. У здорового взрослого человека наблюдается равновесие между диссимиляцией и анаболизмом. Эти процессы обеспечивают самообновление клеточных структур. В старости, при истощающих заболеваниях, голодании, стрессовых состояниях диссимиляция превалирует над анаболизмом. В период роста, при беременности, интенсивной физической нагрузке, в период выздоровления или выхода из состояния голодания анаболизм преобладает над диссимиляцией. Обмен белков. Биологическая ценность различных белков определяется набором заменимых и незаменимых аминокислот, содержащихся в их составе. Белки, не содержащие хотя бы одной незаменимой аминокислоты, называют неполноценными, т.к. это ведет к нарушению синтеза белков. Растительные белки являются неполноценными, т.к. они не содержат полного набора аминокислот. Животные белки считаются полноценными для организма — они по аминокислотному составу ближе к белкам человека и содержат полный набор незаменимых аминокислот, не синтезирующихся в организме (аргинин, валин, гистидин, изолейцин, лейцин, лизин, метеонин, треонин, триптофан, фенилаланин). В пищевом рационе должно быть 55–60% животных белков от общего количества белков. Потребность организма в белках. Белковый минимум — минимальное количество белка пищи, при котором воз- Глава 11. Обмен и терморегуляция 303 можно поддержание азотистого равновесия (в условиях покоя — около 40 г белка в сутки). Белковый оптимум — количество белка пищи, которое полностью обеспечивает потребности организма, хорошее самочувствие, высокую работоспособность, достаточную сопротивляемость неблагоприятным воздействиям на организм (около 90 г в сутки, но не менее 1 г/кг массы в сутки). Коэффициент изнашивания — количество белка, распадающегося в организме за сутки при безбелковой диете, но достаточной по калорийности за счет жиров и углеводов (белковое голодание). Он равен около 20 г в сутки. Избыточное поступление белка с пищей ведет к повышению образования мочевины и увеличению нагрузки на почки с последующим их функциональным истощением. Возможно развитие интоксикации в результате накопления в кишечнике продуктов неполного расщепления и гниения белков. Увеличивать количество белков в пищевом рационе нецелесообразно, поскольку у взрослого здорового человека, в отличие от хищников, белки про запас не откладываются — с увеличением поступления белков в организм увеличивается их диссимиляция, что является дополнительной нагрузкой на организм. Избыток белка (аминокислот) преобразуется в жир. При этом устанавливается азотистое равновесие на более высоком уровне. При недостаточном поступлении белков в организм развивается снижение умственной и физической работоспособности, недостаточность защитных функций организма, могут наблюдаться отеки, атрофия мышц и слизистой кишечника. Определение прихода и расхода белка в организме. Для определения суточного расхода белка организмом определяют в суточной моче содержание азота в граммах и умножают результат на 6,25. В навеске пищевого продукта биохимическим методом определяют содержание азота в граммах, умножают результат на 6,25, т.к. белок на 16% состоит из азота, затем пересчитывают на общий вес продукта и вычитают 10%, т.е. количество белка, не усвоенного в пищеварительном тракте. Обмен азота. В процессе обмена белков может наблюдаться азотистое равновесие, положительный или отрица- 304 Физиология тельный азотистый баланс. Положительным азотистым балансом называют состояние азотистого обмена, при котором количество поступившего в организм азота большем, чем выводимого с мочой. Он наблюдается в период роста организма, после голодания, при беременности, физической тренировке, сопровождающейся ростом мышечной массы, при выздоровлении после истощающей болезни. Под отрицательным азотистым балансом понимают состояние азотистого обмена, при котором количество поступившего в организм азота меньше выводимого с мочой. Он наблюдается при голодании, при недостатке количества или биологической ценности белка пищи, при истощающих заболеваниях, в старости. Азотистым равновесием называют состояние азотистого обмена, при котором количество поступившего в организм азота равно количеству азота, выводимого с мочой. При увеличении содержания белка в пище азотистое равновесие вскоре установится на новом, более высоком уровне. Регуляция обмена белка. В гипоталамусе имеются сенсорные рецепторы, улавливающие содержание аминокислот в крови, с помощью чего гипоталамус включает нейрогормональный регуляторный механизм. Гормон щитовидной железы тироксин (Т3) усиливает синтез белков, высокие концентрации Т3, наоборот, подавляют синтез белка. Гормон роста, инсулин, тестостерон, эстроген стимулируют синтез белка в организме. Глюкокортикоиды усиливают распад белков, особенно в мышечной и лимфоидной тканях, но стимулируют синтез белков в печени. Функции белков: • главная функция — пластическая. Белки необходимы для синтеза клеточных структур (рост организма, восстановление поврежденных структурных элементов), для синтеза биологически активных веществ — гормонов, ферментов. Белок — это первооснова жизни, 50% сухого вещества клетки составляют белки. Азот содержится только в белках, поэтому их нельзя заменить углеводами или жирами; • энергетическая роль белков второстепенная — белки при сбалансированном питании поставляют организму около 15% энергии; Глава 11. Обмен и терморегуляция 305 • защитная функция (иммунные белки плазмы крови, антитела); • создают онкотическое давление (часть осмотического давления); • являются компонентами буферных систем крови; • транспорт гормонов, липидов, холестерина, минеральных веществ. Обмен липидов. Липиды организма — в основном нейтральные жиры (триглицериды), а также фосфолипиды, холестерин и жирные кислоты. Жирные кислоты и глицерин поступают с пищей в виде животных жиров. Жиры синтезируются из жирных кислот и глицерина, из аминокислот и моносахаридов. Биологическая ценность различных жиров, поступающих в организм, зависит от соотношения в жирах насыщенных и ненасыщенных (заменимых и незаменимых) жирных кислот. В свою очередь, это зависит от соотношения в пище жиров животного и растительного происхождения, содержания витаминов А, D, Е. Оптимальный вариант соотношения в пищевом рационе жиров животного и растительного происхождения — 70% животных жиров, 30% — растительных. Линолевая и линоленовая кислоты незаменимы, т.к. они не синтезируются в организме человека из других органических соединений, и составляют всего лишь около 1% от общего количества жиров. Они в значительной степени определяют активность ферментов мембран и их проницаемость, являются материалом для синтеза простагландинов — регуляторов многих жизненно важных функций организма (проницаемость клеточных мембран, возбудимость, интенсивность метаболизма). Заменимые жирные кислоты — это кислоты, которые синтезируются в организме (насыщенные — пальметиновая, стеариновая — и ненасыщенные — олеиновая, араидоновая и др.). Потребность организма в жирах составляет около 110 г в сутки. Избыточное поступление в организм растительных жиров (кроме оливкового) способствует развитию опухолей. При недостатке жира в организме наблюдаются поражения кожи и волос, нарушение синтеза простагландинов, страдают все функции организма. При недостаточном поступлении в организм только незаменимых жирных кислот 306 Физиология развиваются подобные нарушения, а также гиперхолестеринемия, что способствует развитию атеросклероза. При избыточном поступлении жиров в организм развивается ожирение, в большей степени, чем обычно, — атеросклероз. Ожирение служит фактором риска развития сердечно-сосудистых заболеваний и их осложнений (инфаркт миокарда, инсульт и др.), ведет к снижению продолжительности жизни. Увеличивается риск развития рака кишечника, молочной и предстательной желез. Уменьшению концентрации холестерина в плазме крови способствуют регулярные физические упражнения. Общее количество холестерина в крови — 3,5–5,2 мМ/л. Функции липидов: • пластическая роль липидов заключается в том, что из жиров образуются элементы клеточных структур, синтезируется ряд биологически активных веществ, например гормоны, простагландины, витамины А и D. Пластическая роль жиров реализуется в основном фосфолипидами и холестерином. Особую роль играют ненасыщенные жирные кислоты. У взрослой женщины доля жировой ткани в организме составляет 20–25% массы тела, что почти вдвое больше, чем у мужчины (12–14%); • энергетическая роль жиров — почти все клетки (в меньшей степени клетки ЦНС) могут использовать жирные кислоты наряду с глюкозой. Около 30% энерготрат организма должно покрываться за счет жиров, в пожилом возрасте — 25%; • защитная функция жиров: – они предохраняют кожу от высыхания и действия воды; – защищают организм от механических воздействий; – защищают организм от переохлаждения; – уменьшают неощутимые потери воды через кожу (в норме 0,3–0,4 л). При ожогах такие потери могут составлять в сутки до 5–10 л; • роль жиров как депо заключается в том, что они составляют резерв энергии и воды. При окислении 100 г жира образуется 110 г воды и высвобождается Глава 11. Обмен и терморегуляция 307 930 ккал энергии. Жировая ткань обеспечивает женщине резерв энергии, необходимый для вынашивания плода и грудного вскармливания; • жиры обеспечивают механическую фиксацию и защиту внутренних органов. Регуляция обмена жиров. Адреналин, норадреналин, тироксин, гормон роста, глюкагон, глюкокортикоиды мобилизуют жиры из жировых депо в организме. Поэтому при физических нагрузках и стрессовых состояниях в результате выброса в кровь адаптивных гормонов (катехоламинов, глюкокортикоидов) расход жиров организмом возрастает, а интенсивный длительный стресс способствует развитию атеросклероза. Запускается выброс адаптивных гормонов в кровь гипоталамусом. При отсутствии углеводов в крови быстрая мобилизация жирных кислот из жировой ткани обеспечивается за счет снижения секреции инсулина. При этом ограничивается и депонирование жира — большая его часть используется для получения энергии. Парасимпатические центры способствуют отложению жира, симпатические влияния тормозят синтез триглицеридов и усиливают их распад. Обмен углеводов. Биологическая ценность различных углеводов. Доля сахаров не должна превышать 10–12% углеводов суточного рациона, что соответствует 50–100 г. Избыточный прием рафинированных углеводов, в частности сахарозы, способствует развитию кариеса зубов. С пищей должны поступать неперевариваемые углеводы (балластные вещества, пищевые волокна — до 20–35 г), полисахариды — целлюлоза, гемицеллюлоза, пектины, содержащиеся в клеточных оболочках растительных тканей. Они способствуют выведению токсинов, канцерогенов и холестерина, стимулируют секрецию и моторику кишечника, что снижает риск развития запоров, геморроя, дивертикулов и рака кишечника. Потребность организма в углеводах составляет около 400 г в сутки и зависит от интенсивности физического труда — с увеличением физической нагрузки потребность организма в углеводах, как и в белках и жирах, возрастает. Концентрация глюкозы в плазме крови — важный параметр гомеостазиса. Она колеблется в пределах 4,4–5,5 ммоль/л че- 308 Физиология рез 3–4 ч после еды. Углеводы поступают в организм в основном в виде полисахаридов растительного (крахмал) и животного (гликоген) происхождения. Депо углеводов — гликоген, он составляет в среднем 400 г, содержится во всех клетках организма, но больше его в миоцитах и гепатоцитах. При избыточном поступлении углеводов в организм развивается ожирение, что способствует возникновению атеросклероза (фактор риска развития сердечно-сосудистых заболеваний и их осложнений); избыточное потребление глюкозы может способствовать развитию аллергических состояний, повышает риск развития недостаточности поджелудочной железы (ослабление выработки инсулина из-за повышенной нагрузки на β-клетки островкового аппарата и развитие сахарного диабета). При недостатке глюкозы в крови возникает чувство голода, снижается умственная и физическая работоспособность. При выраженном уменьшении глюкозы в крови (до 50% от нормы) — потеря сознания и судороги (гипогликемическая кома). Углеводы выполняют преимущественно энергетическую, а также пластическую функцию. Клетчатка улучшает также двигательную и секреторную функции желудочнокишечного тракта, способствует выведению из организма холестерина пищи. Пластическая роль углеводов заключается в том, что они входят в состав нуклеиновых кислот некоторых гормонов, ферментов, витаминов, являются структурным элементом клеточных мембран, разных элементов соединительной ткани; из углеводов синтезируются заменимые амино- и жирные кислоты. Более 90% углеводов расходуется для выработки энергии. Регуляция обмена углеводов осуществляется в основном с помощью гормонов. Инсулин способствует утилизации глюкозы в клетках с помощью повышения проницаемости мембраны клеток для глюкозы, стимулирует синтез гликогена в печени и мышцах, синтез жиров из углеводов, что ведет к уменьшению содержания глюкозы в крови. При гипергликемии происходит повышение секреции инсулина в результате непосредственного стимулирующего действия недостатка глюкозы на β-клетки поджелудочной железы и путем активирующего влияния глюкозы плазмы крови на глюкорецепторы гипоталамуса и последующего повы- Глава 11. Обмен и терморегуляция 309 шения парасимпатических влияний на секрецию инсулина. Адреналин, норадреналин, глюкагон, глюкокортикоиды обеспечивают глюконеогенез. Тироксин, гормон роста увеличивают содержание глюкозы в крови. При снижении концентрации глюкозы в крови (гипогликемии) ускоряется гликогенолиз — превращение гликогена в глюкозу с помощью глюкагона и адреналина. Парасимпатическая нервная система стимулирует процессы анаболизма (в том числе способствует использованию глюкозы клетками организма); симпатическая способствует диссимиляции и катаболизму (увеличивает содержание глюкозы в крови). Обмен воды и минеральных веществ. Вода составляет 55–60% массы тела человека. В тканях женщин объем воды значительно меньше, чем у мужчин, в связи с большим содержанием жира. На биологическую ценность воды могут влиять дополнительные компоненты: содержание микроэлементов, минеральных солей, тяжелого водорода и кристаллической (талой) воды, она содержится также в овощах и фруктах. Питьевая вода — важнейший источник кальция, магния, ряда микроэлементов. Кальций воды усваивается на 90%, а кальций пищевых веществ — только на 30%. Следует учесть, что в кипяченой воде содержание минеральных компонентов снижено, поэтому постоянное ее использование вместо сырой воды нецелесообразно, т.к. ряд минеральных веществ не поступает в организм. Функции воды в организме. Имеется три основных состояния внутриклеточной и внеклеточной воды: • конституционная вода — структурный элемент молекул клеток и тканей организма; • связанная вода, образующая гидратные оболочки макромолекул (коллоиды), делает их активными; • свободная, т.е. ничем не связанная, вода представляет собой растворитель, обеспечивающий протекание биохимических реакций, выделение продуктов обмена, участвует в процессах терморегуляции (см. раздел 13.4), определяет реологические свойства крови. Потребность организма в воде вариабельна. В средних широтах она составляет 2,5–3,0 л в сутки. При недостатке 310 Физиология воды страдают все обменные процессы в организме, нарушается выведение продуктов обмена белков (мочевины, мочевой кислоты, креатинина и др.). При недостатке воды в организме возникает жажда, представляющая собой реакцию организма на повышение осмотического давления и снижение объемов жидкостей. Одной из причин появления жажды может быть высыхание слизистой оболочки рта в результате уменьшения слюноотделения, потери жидкости при разговоре, одышке, курении. Выраженность жажды обычно уменьшается при полоскании рта. Один из факторов, способствующих формированию жажды, — действие ангиотензина. Утоление жажды возникает в процессе питья (сенсорное насыщение) — до всасывания воды. По-видимому, это явление, как и сенсорное насыщение пищей, развивается благодаря растяжению стенок желудка и возбуждению его механорецепторов. Истинное водное насыщение формируется в результате всасывания необходимого организму количества воды. Прекращение поступления воды в организм ведет к смерти. При избыточном поступлении воды в организм наблюдается увеличение объема циркулирующей крови, что увеличивает нагрузку на сердце, наблюдается усиление потоотделения и мочеотделения, что ведет к потере солей, витаминов, к ослаблению организма, возможна водная интоксикация. Регуляция объема жидкостей и осмотического давления в организме — см. раздел 12.4. Минеральные вещества участвуют в осуществлении многих процессов в организме: • регуляции осмотического давления и кислотно-основного состояния; • создании мембранного потенциала покоя и потенциала действия возбудимых клеток; • процессов свертывания крови; • кофакторы в ферментативных реакциях; • натрий играет весьма важную роль в транспорте других веществ. Особая роль ионов Na+ и К+ заключается в том, что они — основные ионы в формировании потенциала покоя и потенциала действия. Na+ обеспечивает транспорт аминокислот, глюкозы и других ионов через мембраны клеток. Глава 11. Обмен и терморегуляция 311 Калий — в основном внутриклеточный катион, в плазме крови его концентрация равна 3,3–4,9 ммоль/л. Натрий составляет 93% всех катионов плазмы крови, его концентрация равна 135–145 ммоль/л. Кальций в организме содержится в виде фосфатов в костях, он участник реакции гемостаза, ионы Са 2+ в возбудимых тканях играют роль фактора электросекреторного и электромеханического сопряжений, стабилизируют мембранный потенциал покоя, поэтому при недостатке Са2+ могут развиться судороги. Фосфор — элемент макроэргических соединений, коферментов тканевого дыхания и гликолиза, входит в состав фосфорно-кальциевых соединений костного вещества, а также анионов внутриклеточной жидкости. Магний служит катализатором многих внутриклеточных процессов, снижает возбудимость нервной системы и сократительную активность скелетных мышц, способствует снижению артериального давления вследствие расширения кровеносных сосудов и уменьшения частоты сердечных сокращений. Основные микроэлементы, необходимые человеку — это медь, цинк, фтор, йод, кобальт, бор, железо. Обычно они поступают в организм в достаточном количестве при сбалансированном питании. Особая роль железа заключается в том, что оно входит в состав гемоглобина и цитохромов митохондрий (необходимо для транспорта кислорода и для окислительных реакций), при окислении железа транспорт кислорода нарушается, наступает смерть. Йод — единственный из известных микроэлементов, участвующий в построении молекул гормонов — до 90% циркулирующего в крови органического йода приходится на долю тироксина и трийодтиронина. Недостаточное поступление в организм йода может быть причиной нарушения функций щитовидной железы. Фтор защищает зубы от кариеса, стимулирует реакции иммунитета и кроветворение, предупреждает развитие старческого остеопороза. Потребность в NaCl — 5–8 г/сут, это количество содержится в обычной пище (обычно потребляют 10–12 г/сут). Избыток Na+ задерживает жидкость в организме, что может приводить к повышению нагрузки на сердце и почки; 312 Физиология накопление NaCl в стенке кровеносных сосудов способствует ее набуханию, утолщению и сужению просвета сосудов. Избыток Na+ в миоцитах сосудов повышает их возбудимость, что ведет к более легкому возбуждению миоцитов и их сокращению, а также повышению тонуса сосудов. Диета со сниженным содержанием натрия и увеличенным содержанием калия может приводить к предупреждению развития инсультов. Особенно богаты калием картофель, чернослив, абрикосы, виноград. Механизм регуляции минерального обмена см. в разделе 6.4; 6.6. Питание — основные положения. Термин «питание» используют в двух значениях. Во-первых, это совокупность питательных веществ, поступающих в организм с пищей. Питательные вещества — это продукты гидролиза жиров, белков и углеводов (мономеры) — пластический и энергетический материал, а также вода, минеральные соли и витамины, являющиеся только пластическим материалом. Во-вторых, это способ поступления питательных веществ во внутреннюю среду организма (энтеральное и парентеральное питание). Питание должно обеспечивать пластические и энергетические потребности организма с учетом возраста, пола, антропометрических данных (рост, масса), трудовой деятельности, климатических условий. Это достигается путем соблюдения четырех основных принципов питания: • общая калорийность пищи должна соответствовать всем перечисленным требованиям; • белки, жиры и углеводы в пищевом рационе взрослого человека должны содержаться в соотношении 1 : 1,2 : 4,6 (сбалансированное питание); • распределение пищевого рациона при четырехразовом питании наиболее целесообразно сделать следующим образом: завтрак — 25%, второй завтрак — 15%, обед — 45%, ужин — 15%; • в организм должно поступать достаточное количество воды, минеральных веществ и витаминов. Критерий соответствия питания человека энергетическому принципу — сохранение у взрослого человека стабильной массы тела. Ориентировочно должную массу тела Глава 11. Обмен и терморегуляция 313 можно вычислить по методу Брока, вычитая 100 из показателя длины тела в сантиметрах (в настоящее время сделана поправка: если длина тела равна 166–175 см, из ее величины вычитают не 100, а 105, если же длина тела превышает 175 см, вычитают 110). Нормы физиологических потребностей в энергии (ккал, в скобках кДЖ) для различных возрастных и профессиональных групп населения (мужчины возраста 30–40 лет): • умственный труд, небольшая физическая нагрузка — 2300 (2145–2475); • легкий физический труд — 2650 (2475–2970); • среднетяжелый физический труд — 3150 (2970–3465); • тяжелый физический труд — 3850 (3675–4200); • очень тяжелый физический труд — 3960 и более. С возрастом расход энергии уменьшается. Расход энергии у женщин на 20% меньше, чем у мужчин. Витамины — это биологически активные вещества, являющиеся компонентом ферментов, поступающие в организм с пищей и необходимые для регуляции биохимических реакций. Некоторые витамины синтезируются в организме в небольших количествах. Носители ряда витаминов, прежде всего аскорбиновой кислоты, — это свежие продукты (при хранении и термической обработке пищи многие витамины разрушаются). Небольшое количество витамина K, а также группы B и биотина синтезируется в кишечнике человека при участии микрофлоры. Роль витаминов в организме. Они участвуют в регуляции метаболизма и клеточного дыхания (витамины группы B и никотиновая кислота), в синтезе жирных кислот, гормонов стероидной природы, в регуляции процессов фоторецепции и размножения, обмена кальция и фосфата, во многих окислительно-восстановительных процессах, в гемопоэзе и синтезе факторов свертывания крови, а также обеспечивают антиоксидантное действие (витамины A, C, E), что особенно необходимо при экстремальных нагрузках. Антиоксидантное действие витаминов обусловлено их способностью инактивировать токсические продукты преобразования в организме кислорода, или так называе- 314 Физиология мые свободные радикалы. Повышенный прием витаминов, в частности A, C и E, рекомендуется для сохранения здоровья у лиц, предрасположенных к стрессам, а также к сердечно-сосудистым и онкологическим заболеваниям, в развитии которых свободные радикалы играют большую роль. Так, относительно высокое потребление цитрусовых и овощей, содержащих значительное количество аскорбиновой кислоты (витамина C), может защищать человека от развития рака желудка. В популяциях, характеризующихся высоким приемом каротинов (провитамина A), снижен риск развития рака легких и органов пищеварения. Потребление же токоферола (витамина E), в 6–7 раз превышающее обычно рекомендуемую дозу, может привести к снижению смертности от ишемической болезни сердца на 40% по сравнению с показателями у людей, потребляющих мало витамина E. 11.2. Обмен энергии в организме Различают пластический, основной и общий обмен. Основной обмен — минимальный расход энергии, обеспечивающий гомеостазис в стандартных условиях: при бодрствовании, максимальном мышечном и эмоциональном покое, натощак (12–16 ч после еды), при температуре комфорта (отсутствие ощущений тепла или холода, что зависит от одежды и температуры окружающей среды). Основной обмен определяют в указанных стандартных условиях потому, что физическая нагрузка, эмоциональное напряжение, прием пищи и изменение температуры окружающей среды увеличивают интенсивность метаболических процессов в организме (расход энергии). Энергия основного обмена в организме расходуется на обеспечение жизнедеятельности всех органов и тканей организма, клеточный синтез и поддержание температуры тела. Примерно половина основного обмена приходится на печень и скелетную мускулатуру. На величину должного (среднестатистического) основного обмена здорового человека влияют пол, возраст (рис. 11.1), рост и масса тела (вес). На величину истинного (реального) основного обмена здорового человека влияют также условия жизнедеятельности, к которым организм Глава 11. Обмен и терморегуляция Интенсивность основного обмена кДж 225 315 ккал 200 175 150 0 10 20 30 40 50 Возраст, лет 60 70 Рис. 11.1. Зависимость относительной интенсивности основного обмена от возраста и пола человека (по Х.-Ф. Ульмеру, Г. Тевсу, 1996) адаптирован: постоянное проживание в холодной климатической зоне увеличивает основной обмен, а длительное вегетарианское питание — уменьшает. Величина основного обмена в сутки у мужчин составляет 1500–1700 ккал (6300–7140 кДж); в расчете на 1 кг массы в сутки равна: 21–24 ккал (88–101 кДж/кг/сут (1 ккал = = 4,19 кДж)). У женщин эти показатели примерно на 10% меньше (меньше мышечная масса, кроме того, мужские половые гормоны повышают основной обмен на несколько процентов, женские половые гормоны таким действием не обладают). Показатели основного обмена при расчете на 1 м2 поверхности тела у теплокровных животных разных видов и человека примерно равны, при расчете на 1 кг массы — сильно различаются: чем меньше организм, тем больше расход энергии. Отклонение от должного основного обмена можно определить по формуле и номограмме Рида, приведенным ниже. Известно, что в условиях основного обмена 9–10% энергии расходуется на работу, совершаемую сердцем. Это дает основание установить взаимосвязь между параметрами, характеризующими нагнетательную функцию сердца (ЧСС, АД), и суммарными энерготратами организма. Такая взаимосвязь может быть наиболее четкой, если 316 Физиология организм находится в стандартных условиях определения основного обмена. Рид на основании многочисленных параллельно исследуемых параметров гемодинамики и значений основного обмена установил зависимость между показателями измеряемых функций. Формула Рида: процент отклонения = 0,75 × (частота пульса + пульсовое давление × 0,74) – 72. Вместо проведения расчетов можно пользоваться номограммой Рида (рис. 11.2). На левой шкале находят измеренные значения частоты пульса, а на правой — пульсовое давление. Найденные точки соединяют линейкой (удобнее пользоваться прозрачной). Точка пересечения линейки со средней шкалой показывает процент отклонения основного обмена. Пластический обмен — это расход энергии, обеспечивающий рост развивающегося организма и восстановление структурных элементов организма после тяжелой болезни или длительного голодания. Общий обмен — это совокупность пластического, основного обмена и дополнительного расхода энергии, обеспечивающего жизнедеятельность организма в различных условиях. Факторы, повышающие расход энергии организмом: Рис. 11.2. Номограмма Рида Глава 11. Обмен и терморегуляция 317 физическая и умственная нагрузка, эмоциональное напряжение, изменение температуры и других условий окружающей среды, специфически динамическое действие пищи (увеличение расхода энергии после приема пищи) — все это включает понятие «общий обмен». Термонейтральная среда — температура воздуха и стен (температура комфорта) для легко одетого человека 25–26 °С, для обнаженного — 26–28 °С. При понижении температуры среды расход энергии увеличивается, что предотвращает охлаждение организма; при повышении температуры среды расход энергии уменьшается. При значительном повышении температуры среды обмен веществ может возрастать вследствие перегревания тканей организма и усиления биохимических реакций. Расход энергии организмом после приема белковой пищи увеличивается на 20–30%, после приема жиров — на 5%, при смешанной пище — на 6–10%. При эмоциональных реакциях увеличение расхода энергии у взрослого человека составляет обычно 40–90% от уровня основного обмена и связано главным образом с вовлечением фазных и тонических мышечных реакций. Даже при небольшой физической нагрузке (например, ходьбе со скоростью 5 км/ч) уровень расхода энергии может повыситься в 3–4 раза. Ежедневная двухкилометровая прогулка (без изменений в питании) может способствовать устранению за месяц 1 кг жира. Для определения эффективности преобразования тепловой энергии в механическую введено понятие «коэффициент полезного действия (КПД) организма при мышечной работе» — это выраженное в процентах отношение энергии, эквивалентной полезной механической работе, ко всей энергии, затраченной на выполнение этой работы. Коэффициент полезного действия у человека при мышечной работе рассчитывают по формуле: КПД = А С–е × 100% , где А — энергия, эквивалентная полезной работе; С — общий расход энергии; е — расход энергии за такой же промежуток времени в состоянии покоя. КПД равен примерно 20%. Источник энергии для организма — продукты гидролиза углеводов, жиров и белков, поступающих с пищей, цир- 318 Физиология кулирующие в крови вместе с продуктами диссимиляции клеточных структур и составляющие общий фонд питательных веществ (в основном мономеры). Высвобождение же энергии в организме происходит в процессе катаболизма мономеров, поступающих в клетки из крови. Химическая энергия белков, жиров и углеводов пищи используется для образования АТФ или для синтеза крупномолекулярных веществ. АТФ — донор свободной энергии в клетках. В клетках АТФ используется в течение одной минуты после ее образования. За сутки количество образующейся и расщепляющейся АТФ в организме в 2–3 раза превышает массу тела. Питание должно обеспечить поступление в организм нужного количества энергии и быть сбалансированным — соотношение белков, жиров и углеводов 1:1/2:4/6. Пища должна содержать также достаточное количество воды, минеральных солей и витаминов. Принципы исследования прихода и расхода энергии 1. Обычно применяется расчет по таблицам, в которых указано содержание белков, жиров и углеводов в каждом продукте, поэтому необходимо только умножить количество принятых белков, жиров и углеводов на их калорические коэффициенты, суммировать полученные результаты и вычесть 10% (потери с калом). 2. Физическая калориметрия — сущность метода заключается в следующем: в калориметре сжигают навеску продукта, а затем по степени нагревания воды и материала калориметра рассчитывают выделившуюся энергию. Количество тепла, выделившегося при сгорании продукта в калориметре, рассчитывают по формуле: Q = Mв × Св (t2 – t1) + Mк × Ск (t2 – t1) – Qо, где Q — количество тепла; М — масса (в — воды, к — калориметра); t2 – t1 — разность температур воды и калориметра после и до сжигания навески; С — удельная теплоемкость; Qо — количество тепла, образуемое окислителем. Количество тепла, высвобождаемое при сгорании 1 г вещества в калориметре, называют физическим калорическим коэффициентом; при окислении 1 г вещества в орга- Глава 11. Обмен и терморегуляция 319 низме — физиологическим калорическим коэффициентом. Основание для расчета прихода энергии в организм по количеству усвоенных белков, жиров и углеводов — закон термодинамики Гесса, который гласит: термодинамический эффект зависит только от теплосодержания начальных и конечных продуктов реакции и не зависит от промежуточных превращений этих веществ. При окислении в организме 1 г белков высвобождается 4,1 ккал (17,2 кДж), 1 г жиров — 9,3 ккал (38,9 кДж), 1 г углеводов — 4,1 ккал (17,2 кДж). При сгорании в калориметре жиров и углеводов выделяется столько же тепла, сколько в организме. При сгорании белка в калориметре энергии выделяется несколько больше, чем в организме, т.к. часть энергии белка при окислении в организме теряется с мочевиной и другими веществами белкового обмена, которые содержат энергию и выводятся с мочой. 3. Физико-химический принцип. Чтобы рассчитать приход энергии в организм с пищей, определяют содержание белков, жиров и углеводов в продуктах питания, умножают их количество на соответствующие физиологические калорические коэффициенты, суммируют и из суммы вычитают 10% — то, что не усваивается в пищеварительном тракте (потери с калом). Основной способ определения расхода энергии организмом — непрямая (косвенная) калориметрия, основанная на расчете количества выделившейся энергии по данным газообмена (поглощенный О2 и выделившийся СО2 за сутки). Количество выделяемой организмом энергии можно рассчитать по показателям газообмена потому, что количество потребленного организмом О2 и выделенного СО2 точно соответствует количеству окисленных белков, жиров и углеводов, а значит, и израсходованной организмом энергии. Для расчета расхода энергии способом непрямой калориметрии используются дыхательный коэффициент и калорический эквивалент кислорода. Дыхательным коэффициентом называют отношение объема выделенного организмом углекислого газа к объему потребленного за это же время кислорода. Величина дыхательного коэффициента зависит от соотношения белков, жиров и углеводов, окислившихся в организме. Ды- 320 Физиология хательный коэффициент при окислении в организме белков равен 0,8, жиров — 0,7, углеводов — 1,0. Дыхательный коэффициент для жиров и белков ниже, чем для углеводов, вследствие того, что на окисление белков и жиров расходуется больше О2, т.к. они содержат меньше внутримолекулярного кислорода, чем углеводы. Дыхательный коэффициент в начале интенсивной физической работы приближается к единице, потому что источник энергии в этом случае — преимущественно углеводы. В первые минуты после интенсивной и длительной физической работы дыхательный коэффициент у человека больше единицы, так как СО2 выделяется больше, чем потребляется О2, поскольку молочная кислота, накопившаяся в мышцах, поступает в кровь и вытесняет СО2 из бикарбонатов. Калорическим эквивалентом кислорода (КЭК) называют количество тепла, высвобождаемое организмом при потреблении 1 л О2. КЭК для белков — 4,48 ккал (18,8 кДж), для жиров — 4,69 ккал (19,6 кДж), для углеводов — 5,05 ккал (21,1 кДж). Известны три основных метода определения расхода энергии по газообмену — метод Дугласа–Холдена, метод Шатерникова и метод Крога. В клинической практике обычно определение расхода энергии проводится по методу Крога. Испытуемый вдыхает кислород из мешка метаболиметра, вдыхаемый воздух возвращается в тот же мешок, предварительно пройдя через поглотитель СО2 (рис. 11.3). По показаниям метаболиметра определяют расход О2 и умножают на калориметрический эквивалент кислорода в условиях основного обмена: 4,86 ккал (20,36 кДж). По методу Дугласа–Холдена расчет энергии проводят по данным полного газового анализа. Способ прямой калориметрии основан на непосредственном измерении количества тепла, выделенного организмом (Пашутан В.В., Этуотор У., Бенедикт Ф.). Камера, в которую помещают испытуемого, термически изолирована от окружающей среды, ее стенки не поглощают тепло, внутри них находятся радиаторы, через которые течет вода. По степени нагрева определенного количества воды Глава 11. Обмен и терморегуляция 321 Клапан Кислород 3 Объем поглощенного О2 2 Мундштук 1 0 Противовес 0 1 Время, мин 2 Рис. 11.3. Принцип определения расхода энергии по методу Крога рассчитывают количество тепла, израсходованного организмом. В клинической практике, как и способ Дугласа– Холдена, не используется. 11.3. Терморегуляция По степени развития терморегуляторных механизмов животные делятся на три группы: • пойкилотермные организмы (от греч. poikilos — изменчивый), не способные поддерживать температуру тела на постоянном уровне, т.к. они вырабатывают мало тепла; • гетеротермные организмы (от греч. heteros — иной, другой) занимают промежуточное положение, колебания температуры их тела значительны. Это характерно, например, для животных с зимней спячкой; • гомойотермные организмы (от греч. homeo — подобный, одинаковый), к которым относится и человек; отличаются относительным постоянством температуры тела, поскольку вырабатывают много тепла. Постоянство температуры тела обеспечивает высокий уровень жизнедеятельности независимо от температуры окружающей среды. Только внутренние части тела человека, названные «ядром», имеют стабильную температуру около 37 °С. При температуре комфорта окружающей среды температура кожи обнаженного человека составляет 33–34 °С, температура в подмышечной впадине при плотном прижатии руки к телу равна 36,0–36,9 °С (рис. 11.4). 322 Физиология Терморегуляция осуществляется с помощью изменения интенсивности теплопродукции и теплоотдачи. Теплопродукция наиболее интенсивно идет в мышцах, легких, печени, почках. В мышцах вырабатывается до 30– 50% тепла при охлаждении тела. Выделяют сократительную и несократительную теплопродукцию. Сократительная теплопродукция наблюдается только в скелетной структуре. Сокращения мышц при этом могут быть произвольными и непроизвольными. Произвольные сокращения — двигательная активность человека, она очень быстро ведет к увеличению теплообразования, при этом повышаются и теплопотери за счет усиления отдачи тепла конвекцией, что неэкономично для организма. Непроизвольная сократительная теплопродукция — это дрожь и терморегуляторный тонус мышц. Теплопродукция при этом возрастает примерно на 40–50%. Дрожь свойственна только человеку и наблюдается при значительном снижении температуры окружающей среды, что ведет к повышению образования тепла в организме в несколько раз. Дрожь — более экономичный способ поддержания температуры тела по сравнению с произвольными сокращениями. 37 °С 36 °С 32 °С 28 °С 34 °С 31 °С А Б Рис. 11.4. Температура различных областей тела человека в условиях холода (А) и тепла (Б) (по К. Брюку, 1996) Глава 11. Обмен и терморегуляция 323 Несократительная теплопродукция при охлаждении организма увеличивается во многих органах (легкие, печень и др.). Специфическая ткань такой теплопродукции — это бурая жировая ткань. После удаления бурой жировой ткани в эксперименте устойчивость организма к холоду существенно снижается. Скорость окисления жирных кислот в бурой жировой ткани в 20 раз превышает скорость в белой жировой ткани. Масса бурой жировой ткани, обычно составляющая 1–2% массы тела, при адаптации к холоду может увеличиваться до 5% массы тела. Ее доля участия в процессах терморегуляции у человека дискутируется. Теплоотдача осуществляется четырьмя способами: • теплоотдача с помощью проведения; • конвекция — происходит в том случае, если температура воздуха (воды, в случае нахождения человека в воде) ниже температуры тела. При движениях воздуха или воды относительно тела интенсивность теплоотдачи значительно повышается. Охлаждающее действие проточной воды в 50–100 раз более действия воздуха; • теплоотдача излучением инфракрасными лучами при наличии на некотором расстоянии от кожи предметов с более низкой температурой — происходит в основном с не покрытой одеждой кожи. Уменьшают теплоотдачу подкожная жировая клетчатка и одежда. Когда жарко, сосуды кожи расширяются вследствие уменьшения тонуса симпатических нервов (теплоотдача возрастает), когда холодно, развиваются противоположные реакции; • теплоотдача с помощью испарения составляет около 20% в условиях комфортной температуры среды. Существуют два варианта испарения воды с поверхности тела: испарение пота в результате его выделения и испарение воды, оказавшейся на поверхности путем диффузии, — неощутимые потери воды. Если температура окружающей среды оказывается равной или выше температуры тела, то теплоиспарение — единственный способ отдачи тепла. Тепло к поверхности кожи проводится с кровью (конвекция), противоположно направленный ток кро- 324 Физиология ви в венах относительно артерий уменьшает теплоотдачу по принципу поворотно-противоточной системы — это внутренние потоки тепла, которые существенно изменяются при сужении или расширении кожных сосудов. Регуляция температуры тела. При понижении температуры окружающей среды выработка тепла (расход энергии) увеличивается, а сосуды кожи суживаются, что уменьшает теплоотдачу и обеспечивает поддержание постоянной температуры тела. При повышении температуры окружающей среды выработка тепла уменьшается, сосуды кожи расширяются, возможно также потоотделение, в результате чего больше тепла отдается в окружающую среду, что также способствует поддержанию постоянной температуры тела. Большинство рецепторов, воспринимающих изменение температуры тела, находятся в коже. Они обеспечивают передачу сигналов в ЦНС об изменениях температуры среды и формирование температурных ощущений. В коже холодовых рецепторов во много раз больше, нежели тепловых. Во внутренних органах и тканях также преобладают холодовые рецепторы. Терморецепторы (термосенсоры) обнаружены в спинном и среднем мозге, в гипоталамусе. Термосенсоры — это нейроны, которые могут возбуждаться при их непосредственном охлаждении или нагревании, они очень чувствительны — улавливают изменение температуры всего лишь на 0,011 °С. Центр терморегуляции локализуется в гипоталамусе. В переднем гипоталамусе происходит сравнение полученных с периферии сигналов с активностью центральных термосенсоров, отражающих температурное состояние мозга. На основе интеграции информации этих двух источников задний гипоталамус обеспечивает выработку сигналов, управляющих процессами теплопродукции и теплоотдачи. Терморегуляторные реакции могут включаться также по механизму условного рефлекса, что доказывает участие коры большого мозга в регуляции температуры тела. Регуляция сократительной теплопродукции осуществляется соматической нервной системой. Сужение и расширение сосудов, обеспечивающие изменение теплоотдачи, осуществляются в основном симпатоадреналовой систе- Глава 11. Обмен и терморегуляция 325 Рис. 11.5. Функция экзокринной потовой железы стимулируется симпатическим нервом (по А.К. Гайтону, Д.Э. Холлу, 2008) мой. Интенсивность потоотделения также регулируется симпатической нервной системой (рис. 11.5), но с помощью холинергических нейронов, при повышении температуры тела потоотделение возрастает. Симпатоадреналовая система и гормоны щитовидной железы (три- и тетрайодтиронины) стимулируют обмен веществ (выработку тепла) в различных органах и тканях при понижении температуры окружающей среды и уменьшают при ее увеличении. Совокупность всех механизмов обеспечивает температурный гомеостазис. При температуре среды выше комфортной (в том числе и между кожей и одеждой) и недостаточности теплоотдачи за счет теплоиспарения теплопродукция может не уменьшаться, а возрастать вследствие повышения температуры тканей организма — наблюдается перегревание организма (тепловой удар). Перегревание тела человека более чем на 6 °С ведет к смерти, т.е. при температуре тела выше 43 °С, но это может происходить только в экстремальных случаях и наблюдается крайне редко. ГЛАВА 12 Выделение конечных продуктов обмена веществ из организма — это способ очищения его от «шлаков». Выделительная система Выделение — освобождение организма от конечных продуктов обмена, чужеродных веществ и избытка питательных веществ (например, глюкозы, NaCl). Это последний этап совокупности процессов обмена веществ, конечными продуктами которого являются Н2О, СО2 и NH3. Аммиак образуется только при окислении белков и выделяется в основном в виде мочевины после соответствующих превращений в печени. Вода и СО2 образуются при окислении углеводов, белков и жиров и выделяются из организма в свободном виде. Лишь небольшая часть СО2 выделяется почками в виде карбонатов. Главные органы, выполняющие выделительную функцию, — почки и легкие. Почки выделяют практически все азотсодержащие вещества, больше половины воды, минеральные соли, избыток питательных веществ, чужеродные вещества (например, продукты распада микроорганизмов, лекарственные вещества). Легкие удаляют практически весь образующийся в организме СО2; они выделяют также воду, некоторые летучие вещества, попавшие в организм (выхлопные газы автотранспорта и выбросы промышленных предприятий, алкоголь, мочевину, продукты деградации сурфактанта). Выделительную функцию кроме почек и легких выполняют также и другие органы. Железы пищеварительного тракта могут выделять соли тяжелых металлов, чужеродные органические соединения, небольшое количество мочевины и мочевой кислоты, лекарственные вещества (морфий, хинин, салицилаты). Экскреторная функция слюнных желез и всей пищеварительной си- Глава 12. Выделительная система 327 стемы возрастает при заболеваниях почек. При этом заметно увеличивается выведение продуктов обмена белков. С помощью печени через желудочно-кишечный тракт удаляются из крови гормоны и продукты их превращений, продукты обмена гемоглобина, конечные продукты обмена холестерина — желчные кислоты. Кожа. Потовые железы выделяют соли натрия, калия, кальция, мочевину (5–10% всей выводимой организмом мочевины), креатинин, мочевую кислоту. При высокой температуре потоотделение и потеря NaCl сильно возрастают, но при этом увеличивается выработка альдостерона, уменьшающего выделение натрия с мочой. Кожа выделяет также небольшое количество СО2 (около 2%). Потовые железы наиболее плотно расположены в подмышечных впадинах, на ладонях, подошвах. Сальные железы выделяют продукты обмена половых гормонов, кортикостероидов, витаминов, ферментов. Выделение воды различными органами распределяется следующим образом: около 1,5 л выводится с мочой, 100– 150 мл — с калом, 0,4 л — через легкие, 0,5–1,0 л воды удаляется в виде паров с поверхности кожи (всего около 2,5 л/сут в жаркие дни — до 10 л/сут). 12.1. Структурно-функциональная характеристика почки Структурно-функциональная единица почки — нефрон. Он начинается с почечного тельца (рис. 12.1), включающего сосудистый клубочек и охватывающую его капсулу. Капсула клубочка похожа на двустенную чашу, образованную внутренним и наружным листками. Внутренний листок ее в виде слепого конца эпителиального канальца покрывает капилляры клубочка, наружная стенка капсулы (ее внешний диаметр равен 0,2 мм) образует небольшую полость вокруг клубочка и переходит в следующий элемент нефрона — проксимальный извитой каналец. Характеристика отдельных нефронов. Различают корковые нефроны (их около 85%) и юкстамедуллярные нефроны (около 15%). Корковые нефроны практически полностью располагаются в корковом слое почки, и лишь петля нефрона 328 Физиология А Б Корковое вещество 5 I 2 II 1 4 6 Наружное 3 мозговое вещество Внутреннее мозговое вещество Рис. 12.1. Основные функциональные элементы нефрона: А — юкстамедуллярный нефрон; Б — корковый нефрон; 1 — почечное тельце; 2 — проксимальный каналец; 3 — петля нефрона; 4 — плотное пятно (macula densa) дистального канальца; 5 — дистальный каналец; 6 — собирательная трубка опускается на небольшую глубину в мозговое вещество почки. Юкстамедуллярные нефроны располагаются в основном в наружной зоне мозгового слоя, а петля нефрона опускается глубоко во внутреннюю зону мозгового вещества почки. Главную роль в мочеобразовательной функции почки играют корковые нефроны (на них приходится 85% почечного кровотока, поэтому от них зависит объем выводимой мочи). При нарушении их функции может возникнуть анурия (прекращение мочеобразования), что наблюдается, например, при краш-синдроме (синдроме раздавливания). Главное назначение юкстамедуллярных нефронов с их длинной петлей нефрона — создание высокого осмотического давления в мозговом слое почки. Важным структурно-функциональным элементом нефрона является так называемый юкстагломерулярный комплекс, вырабатывающий ренин. Особенности кровоснабжения почек 1. В почке самый большой удельный (на единицу массы) кровоток: примерно в 60 раз больше, чем на единицу массы во всем организме. За сутки через почки проходит 1700 л крови! 2. В клубочковых капиллярах высокое кровяное давление (50–60 мм рт. ст.). Это объясняется близостью приносящей артериолы к аорте и широким ее просветом, что сформиро- Глава 12. Выделительная система 329 валось в процессе эволюции под влиянием обильного кровотока через почки. У корковых нефронов диаметр выносящей артериолы почечного клубочка у́же приносящей, что является результатом меньшей функциональной нагрузки на выносящую артериолу. Это объясняется тем, что около 20% жидкости из крови в клубочках переходит в первичную мочу. 3. В корковых клубочках весьма стабильно капиллярное давление и кровоток даже при значительных колебаниях системного АД: от 80 до 180 мм рт. ст. Постоянство кровотока коркового слоя обеспечивается миогенным механизмом регуляции (см. раздел 9.6). 4. Имеются две системы капилляров в корковом слое почки: первичная — в почечных клубочках (она образуется в результате ветвления приносящих артериол почечных клубочков) и вторичная — околоканальцевая (эти капилляры оплетают проксимальные и дистальные извитые канальцы и начальный отдел собирательных трубок). Она образуется в результате ветвления выносящей артериолы корковых нефронов. Назначение этих систем капилляров принципиально отличается: клубочковые капилляры обеспечивают образование первичной мочи, а вторичная сеть капилляров обеспечивает реабсорбцию веществ из первичной мочи, питание и доставку кислорода тканям почки. Выносящая артериола юкстамедулярных нефронов не распадается на околоканальцевую капиллярную сеть, а образует прямые сосуды, которые опускаются в мозговое вещество и поворачивают обратно, образуя свою противоточную систему. Она обеспечивает обмен веществами между кровью и интерстицием мозгового вещества почки — как и капилляры во всех других тканях. Функции почек весьма разнообразны, но их можно объединить в четыре основные группы: • главная и жизненно необходимая — экскреторная функция почек. Особое значение имеет выделение продуктов обмена белков — мочевины, мочевой кислоты, креатинина. Почкой выводятся гормоны и продукты их распада, лекарства, избыток питательных веществ, вода. Острая почечная недостаточность ведет к летальному исходу в течение 1–2 недель вследствие отравления организма продуктами обмена 330 Физиология • • • • белкового происхождения. Экскреторная функция почки выполняется в процессе мочеобразования; почка участвует в метаболических процессах организма. В ней достаточно активно идет глюконеогенез — особенно при голодании, когда 50% глюкозы, поступающей в кровь, образуется в почке. Почка участвует также в обмене липидов. Роль почки в обмене белков заключается в том, что она расщепляет белки, реабсорбируемые из первичной мочи с помощью пиноцитоза. В ней синтезируются важные компоненты клеточных мембран. В почке образуются аминокислоты аланин и серин, которые могут быть использованы в печени для глюконеогенеза. В почке образуются вещества, выделяемые с мочой, — гиппуровая кислота, аммиак, преобразующийся в почке в аммониевые соли (NH4Cl, (NH4)2SO4); почка участвует также в регуляции различных показателей организма; почка продуцирует биологически активные вещества: ренин, тромбопластин, эритропоэтин, тромбопоэтин, серотонин, простагландины, брадикинин и др. Клетки почки, как и печени, превращают витамин D в его активную форму — витамин D3 (кальцитриол) посредством паратиреоидного гормона; защитная функция почки заключается в том, что она благодаря SH-группе обезвреживает чужеродные яды и ядовитые вещества. Функции различных отделов нефрона весьма специфичны. 12.2. Роль различных отделов нефрона в образовании мочи Роль почечных клубочков заключается в образовании первичной мочи с помощью фильтрации плазмы крови, проходящей по капиллярам клубочка в корковом веществе почек (рис. 12.2). Поскольку при прохождении по разным отделам нефрона состав и количество фильтрата значительно меняются, следует различать первичную мочу (в капсуле Шумлянского–Боумена), вторичную (канальцевая жидкость по ходу нефрона) и конечную мочу (почечные лоханки). Глава 12. Выделительная система 331 Проксимальный каналец Клубочковая полость Эфферентная артериола Наружный слой клубочка Афферентная артериола Рис. 12.2. Почечный клубочек Факторы, определяющие состав первичной мочи (фильтрата): • состав плазмы крови: первичная моча — это плазма крови, лишенная белков (форменные элементы и белки не проходят через фильтрующую мембрану); • проницаемость фильтрующей мембраны, которая, в свою очередь, определяется размером ее пор и самих частиц. Частицы с молекулярным весом выше 70 000 Да, как правило, не проходят через фильтрующую мембрану; • электрический заряд фильтрующихся частиц (например, фильтрации отрицательно заряженных белков препятствуют отрицательные заряды базолатеральной мембраны). Факторы, определяющие объем фильтрации: • фильтрационное давление (ФД), которое является движущей силой перехода жидкости из капилляров почечного клубочка в капсулу Шумлянского–Боумена; • проницаемость фильтрующей мембраны: чем она больше, тем больше объем фильтрата; • площадь фильтрующей мембраны: чем она больше, тем больше объем фильтрата. Площадь весьма велика и составляет 1,5–2 м2 (площадь поверхности тела в среднем равна около 1,7 м2): ФД = ГДК – ОДК – ГДП, где ГДК — гидростатическое давление крови в капиллярах почечного клубочка (около 50 мм рт. ст.); ОДК — онкоти- 332 Физиология ческое давление крови, около 25 мм рт. ст.; ГДП — гидростатическое давление в почке, первичной мочи в капсуле Шумлянского–Боумена, около 10 мм рт. ст.), оно препятствует фильтрации. Таким образом, в среднем ФД = 50 – 25 – 10 = 15 мм рт. ст. В сутки образуется около 180 л фильтрата, т.е. первичной мочи. Роль проксимальных извитых канальцев в мочеобразовательной функции почки. Реабсорбция из первичной мочи необходимых организму веществ — главная их функция (облигатная реабсорбция). Полностью реабсорбируются аминокислоты, глюкоза, витамины и микроэлементы, белки, которые в малых количествах попадают в первичную мочу, пептиды, почти полностью реабсорбируются фосфаты, сульфаты, основная часть бикарбоната. В проксимальных извитых канальцах реабсорбируется значительная часть ионов НСО3–, Cl–, 50% мочевины, около 65% натрия и воды. Вещества, в зависимости от степени их реабсорбции, делят на пороговые и беспороговые. Пороговые вещества обычно реабсорбируются полностью и выводятся из организма лишь при высокой их концентрации в плазме крови («порог выведения», «почечный порог»). Это составные части плазмы: аминокислоты, глюкоза. Выделение глюкозы с мочой (гликозурия) происходит лишь тогда, когда ее содержание в крови превышает пороговый уровень, примерно 1,8 г/л плазмы (10 ммоль/л). Беспороговые вещества выводятся из организма при любой их концентрации в плазме крови (нет порога выведения) — это конечные продукты обмена, подлежащие удалению из организма. Одни из них (мочевина, мочевая кислота) реабсорбируются не полностью, другие практически не реабсорбируются (креатинин, сульфаты). Действуя на процессы реабсорбции в почке с помощью фармакологических препаратов, можно уменьшить или, напротив, увеличить выведение воды из организма (например, борьба с отеками при сердечной недостаточности, снижение АД при гипертонической болезни). В проксимальных канальцах секретируются органическое основание — тетраэтиламмоний, холин, тиамин, серо- Глава 12. Выделительная система 333 тонин, гуанидин, хинин, водород и аммиак, как и в других отделах нефрона, но здесь больше. Главная функция петли нефрона (петли Генле) — создание высокого осмотического давления в мозговом веществе почки. Эту функцию выполняют в основном юкстамедуллярные нефроны, петля которых пронизывает весь мозговой слой почки. По мере продвижения от коркового слоя почки к мозговому его осмотическое давление возрастает от 300 мосмоль/л (0,9% — изотонический раствор NaCl) до 1400 мосмоль/л. Это осуществляется в основном с помощью первично активной реабсорбции Na+ и Cl– из восходящего колена петли Генле в интерстиций — до 25% реабсорбируемого Na+, за которым идет Cl– согласно электрическому градиенту. Из нисходящего колена реабсорбируется в интерстиций вода (около 20% объема первичной мочи) согласно закону осмоса. Ионы Na+ и Cl– переходят также из интерстиция в нисходящее колено. Здесь устанавливается динамическое равновесие осмолярности между интерстицием и нисходящим коленом, из которого NaCl движется в восходящее колено. Эти ионы снова перекачиваются Na/K-помпой в интерстиций — циркуляция ионов Na+ и Cl–. Петля Генле вместе с собирательной трубкой нефрона образуют поворотно-противоточную систему почки. Восходящее колено непроницаемо для воды, поэтому переход из него в интерстиций почки электролитов (без эквивалентного количества воды) обеспечивает создание высокой осмолярности в интерстиции мозгового слоя почки. Поскольку жидкость в петле нефрона движется (в результате работы сердца) в нисходящем и восходящем коленах навстречу друг другу, то небольшие поперечные градиенты (200 мосмоль/л) на каждом участке суммируются, в связи с чем формируется большой продольный градиент. Представим, что вторичная моча не движется — в петле по всей ее длине создается только единичный поперечный градиент осмолярности (рис. 12.3, А). Как только вторичная моча начинает двигаться, в нижней части начинается суммация единичных поперечных градиентов, которая распространяется на всю длину петли нефрона (рис. 12.3, Б, В). 334 Физиология 300 300 Н2О NaCl 100 300 300 100 300 300 100 Н2О NaCl 300 100 NaCl 700 700 500 Н2О NaCl 1400 1400 1200 Н2О NaCl NaCl NaCl Н2О Н2 О NaCl 300 NaCl 1400 А Б Диффузия В Работа помпы Рис. 12.3. Механизм работы петли нефрона как поворотнопротивоточной системы: А–В — этапы формирования высокой осмолярности в мозговом слое почки В дистальных извитых канальцах практически завершается реабсорбция электролитов: реабсорбируется около 10% Na+, за Na+ идет вода (около 8% от общего объема фильтрата, выделяется из организма около 1%). Здесь начинается концентрирование конечной мочи — от гипотонической до изотонической. Поскольку реабсорбция воды регулируется, она называется факультативной. Изотоническая моча из дистальных извитых канальцев переходит в собирательные трубки — конечный отдел нефрона. В собирательных трубках завершается формирование небольшого количества (около 1,5 л) концентрированной конечной мочи, что обеспечивается работой петли нефрона, создающей высокое осмотическое давление в мозговом слое почки. В собирательных трубках реабсорбируются вода, электролиты, мочевина. Реабсорбция воды — это главная причина концентрирования конечной мочи. Вторичная моча течет медленно по собирательным трубкам, которые проходят параллельно петлям нефрона в мозговом слое в направлении почечной лоханки в области с постепенно возрастающим осмотическим давлением. Вода, естественно, из собирательных полупроницаемых трубок согласно закону осмоса переходит в интерстиций мозгового слоя почки с высоким осмотическим давлением, а оттуда — в капилляры и уносится с током крови в пропорциональных количествах с электролитами. Количество реабсорбируемой воды определяется АДГ — это факультативная Глава 12. Выделительная система 335 реабсорбция. При отсутствии АДГ выделяется около 15 л мочи в сутки (в клинике это называют «несахарным мочеизнурением»). Реабсорбция электролитов в собирательных трубках играет незначительную роль: в них реабсорбируется менее 1% Na+, мало реабсорбируется Сl–. Реабсорбция мочевины. Всего в нефроне реабсорбируется 50% профильтровавшейся мочевины, а 50% удаляется с мочой. Из собирательной трубки она диффундирует в интерстиций, а оттуда — в восходящее колено петли нефрона, по которому опять попадает в собирательную трубку. Поскольку для мочевины нет специального насоса, она может перемещаться из канальца в интерстиций и обратно только согласно законам диффузии. В мозговом веществе почки 45% осмолярности обеспечивается за счет мочевины. 12.3. Методы исследования выделительной функции почек Важные показатели для оценки мочеобразовательной функции почек в норме и патологии — это объем первичной мочи и почечный кровоток. Их можно рассчитать, определив почечный клиренс (от англ. clearance — очистка). Определение объема первичной мочи с помощью исследования клиренса какого-либо вещества. Почечный клиренс — это объем плазмы крови в миллилитрах, полностью очищенной от какого-то вещества за 1 мин. Если вещество, попавшее из крови в первичную мочу, не реабсорбируется обратно в кровь, то плазма, профильтровавшаяся в первичную мочу и возвратившаяся с помощью реабсорбции обратно в кровь, полностью очищается от этого вещества. Это вещество не должно также секретироваться и вызывать физиологические эффекты. Такие вещества, например, креатинин, фруктоза, полисахарид инулин (С6Н10О5, молекулярная масса 5200). Указанные свойства этих веществ определяют равную концентрацию их в плазме крови и в первичной моче, а также равное их количество в первичной и конечной моче. Исследуем клиренс по инулину и обозначим: РИН — концентрация инулина в плазме крови; 336 Физиология С — количество первичной мочи, образовавшейся за 1 мин (клиренс по инулину); UИН — концентрация инулина в конечной моче; V — объем конечной мочи за 1 мин: РИН × С = UИН × Vмочи, отсюда: С= UИН × Vмочи . РИН У мужчин (со стандартной поверхностью тела 1,73 м2) образуется суммарно в обеих почках в среднем 125 мл первичной мочи за 1 мин (180 л/сут), у женщин — 110 мл, что составляет примерно 20% плазмы крови, прошедшей через почку. В клинической практике при определении почечного клиренса вместо инулина используют креатинин. Это значительно упрощает процедуру, поскольку его не надо вводить в кровь. Креатинин, попавший в почечный каналец, практически полностью выводится с мочой. Почечный кровоток можно определить, например, с помощью исследования клиренса парааминогиппуровой кислоты (ПАГ). Она свободно фильтруется, а остальная ее часть секретируется в канальцы и обратно не реабсорбируется. В этих условиях кровь за однократное прохождение через почку полностью освобождается от используемого вещества. Клиренс такого вещества показывает почечный плазмоток (ППТ). Принцип расчета тот же, но к плазмотоку надо добавить 45% — объем форменных элементов. В клинической практике используется и ряд других методов. Наиболее широко проводится исследование состава мочи, с помощью чего можно судить о функциях почек и других систем организма. Например, появление глюкозы в моче свидетельствует, что она не полностью реабсорбируется в почках. Это может быть следствием избыточного содержания глюкозы в крови при недостаточной выработке инсулина панкреатической железой (β-клетками островков Лангерганса). Поэтому параллельно необходимо провести исследование крови. Используетсся также УЗИ. Глава 12. Выделительная система 337 12.4. Регуляция мочеобразования и физиологических показателей организма Главным механизмом регуляции мочеобразования является гормональный. Об этом свидетельствует тот факт, что денервированная или пересаженная в эксперименте на шею почка продолжает нормально вырабатывать мочу и адекватно реагировать на эмоциональное напряжение, водные и солевые нагрузки. Процесс регуляции мочеобразования и ряда показателей организма взаимосвязаны. Так, регулируются количество воды в организме, осмоляльность, кислотно-основное состояние (КОС) и АД. Регуляция количества воды в организме осуществляется с помощью регуляции фильтрации и образования первичной мочи в почечных клубочках, а также с помощью реабсорбции и выведения воды. Регуляция с помощью фильтрации • Роль адреналина. Низкие концентрации адреналина увеличивают фильтрацию вследствие расширения сосудов, а высокие уменьшают и снижают мочеобразование вплоть до анурии вследствие сужения сосудов (механизм см. в кровообращении — гуморальная регуляция тонуса сосудов). • Возбуждение симпатической нервной системы уменьшает фильтрацию вследствие спазма сосудов почечных клубочков с помощью ативации α-адренорецепторов, что, естественно, уменьшает мочеобразование и количество выводимой воды из организма. • Роль блуждающего нерва изучена недостаточно. • Натрийуретический гормон (атриопептид) увеличивает фильтрацию и мочеобазование, так как он расширяет сосуды, повышает кровоток в почках и увеличивает проницаемость сосудов. Также действуют простациклины, оксид азота, глюкокортикоиды. Лейкотриены, тромбоксан А2, ангиотензин II уменьшают фильтрацию. Регуляция объема воды в организме с помощью реабсорбции в канальцах • Роль АДГ. Он увеличивает проницаемость для воды конечных отделов дистальных извитых канальцев 338 Физиология (половина их общей длины) и собирательных трубок посредством активации аквапорионов, действуя на Y2-рецепторы. При уменьшении количества воды в организме (объем крови уменьшается), АДГ вырабатывается больше, поэтому больше воды реабсорбируется из канальцев в интерстиций, где высокое осмотическое давление (больше воды задерживается в организме). При отсутствии АДГ из организма выделяется до 15 л мочи/сут. Выработка самого АДГ в данном случае регулируется рефлекторно (рис. 12.4). Симпатический нерв Блуждающий нерв Прямая связь Обратная отрицательная связь Рис. 12.4. Регуляция осмолярности, объема жидкости и АД с помощью почек: СОЯ — супраоптическое ядро; ПВЯ — паравентрикулярное ядро • В случае увеличения объема крови (много воды в организме) возникают противоположные реакции. При этом возрастает импульсация в ЦНС по афферентным волокнам блуждающего нерва от волюморецепторов (барорецепторы низкого давления — предсердия, полые вены). В результате этого выработка АДГ в СОЯ и ПВЯ тормозится, воды выводится больше. • При избытке воды в организме и растяжении предсердий включается также натрийуретический гормон (атриопептид). Он вырабатывается в предсер- Глава 12. Выделительная система 339 диях (больше в правом) и частично в желудочках. Атриопептид увеличивает выведение почками Na+ и Cl−, а значит, и воды. Как отмечалось выше, атриопептид увеличивает также и фильтрацию в почечных клубочках с помощью расширения сосудов почек (как и всего организма) — это самый сильный диуретик. Нервная регуляция реабсорбции. Возбуждение симпатической нервной системы увеличивает в канальцах реабсорбцию Na+, глюкозы, фосфатов, а значит, и воды. Это осуществляется посредством активации β-адренорецепторов. Влияния блуждающего нерва противоположны. Влияние коры большого мозга на процесс мочеобразования осуществляется с помощью условных рефлексов, посредством которых диурез может быть усилен и заторможен. Проявляется в результате эмоций, болевых воздействий. Ведущим фактором при этом является АДГ, а также возбуждение симпатической нервной системы. В частности, при сильных болях мочеобразование тормозится вплоть до анурии. Регуляция осмолярности также осуществляется в основном с помощью АДГ. Запускается процесс с помощью осморецепторов. Они представляют собой клетки, в которых имеются рецепторы, чувствительные к растяжению, т.е. являются механорецепторами. Они расположены в сосудах печени, почек, селезенки, сердца, костного мозга, в интерстиции, в нейронах СОЯ и ПВЯ гипоталамуса. Эти же нейроны вырабатывают АДГ. Часть осморецепторов является хеморецепторами — чувствительны к отдельным ионам, например Na-рецепторы, К-рецепторы и др. Механизм этого процесса следующий: при избытке воды в организме плазма крови становится гипотоничной и вода поступает в клетки, так как осмолярность внутри клеток выше относительно крови, что ведет к набуханию клеток и растяжению их мембран (раздражение осморецепторов и возбуждение их). Это ведет к поступлению импульсов по симпатическим афферентным волокнам в гипоталамус. Так же происходит активация и осморецепторов (нейронов 340 Физиология супраоптического и паравентрикулярных ядер) гипоталамуса, что сопровождается торможением выработки АДГ, увеличением выведения воды и нормализацией осмолярности крови. Роль альдостерона в регуляции осмолярности незначительна, несмотря на то что он задерживает натрий в организме. Это объясняется тем, что при задержке натрия задерживается и вода в пропорциональном количестве согласно закону осмоса, а при выведении натрия в большем количестве больше выводится и воды. Если же накапливается вода в организме, например при избытке NaCl, то возбуждаются волюморецепторы, импульсы поступают в гипоталамус, тормозится выработка АДГ — больше выводится воды. Все регуляторные механизмы взаимодействуют таким образом, что при отклонении от нормы любого показателя, они нормализуют его. Участие почек в регуляции АД осуществляется двумя основными путями. 1. Регуляция АД за счет изменения количества выводимой воды из организма (диуреза). При увеличении объема крови (много воды в организме) увеличивается АД. Оно нормализуется за счет увеличения диуреза (выведение воды). При уменьшении объема воды в организме АД снижается, вода задерживается в организме, АД нормализуется. 2. За счет сосудистых реакций. Это осуществляется с помощью выработки веществ, расширяющих сосуды (простагландины, нейтральный депрессорный липид мозгового вещества почки, кинины), что сопровождается снижением АД, либо веществ, сужающих сосуды (РААС) (см. рис. 9.21). Влияние ренин-ангиотензин-альдостероновой системы (РААС) на АД реализуется и за счет повышения тонуса сосудов, и за счет уменьшения количества выводимой из организма воды с мочой — комбинированная регуляторная система. Ренин вырабатывается в почке эпителиоидными клетками юкстагломерулярного аппарата. При снижении АД секреция ренина возрастает, по мнению одних авторов, под влиянием симпатической нервной системы, по мнению других — в результате активации внутрипо- Глава 12. Выделительная система 341 чечного механизма, который изучен недостаточно. Ренин представляет собой протеазу, под действием которой от α2-глобулина крови (ангиотензиногена) отщепляется малоактивный декапептид ангиотензин I. Последний под действием фермента крови ангиотензиназы (ангиотензинпревращающий фактор, АПФ) превращается в активную форму — ангиотензин II. Это самое мощное из всех известных сосудосуживающих веществ. Он вызывает длительное и значительное сужение сосудов, вследствие чего существенно повышается АД. Кроме того, ангиотензин II вызывает выброс альдостерона из коры надпочечников, что ведет к задержке Na+ и воды и повышению АД. При повышении АД развиваются противоположные реакции. Роль почек в регуляции КОС реализуется двумя путями. 1. Анионы сильных кислот (фосфорной, серной и соляной) выводятся почкой с мочой в виде соответствующих соединений, например NaH2PO4; анионы угольной кислоты (HCO3–) реабсорбируются, что способствует восстановлению буферных систем крови. Сульфаты и фосфаты образуются в результате расщепления белков и нуклеиновых кислот. К нелетучим основаниям относятся главным образом щелочные ионы пищи (бикарбонаты) — их больше содержится в растительных продуктах питания, они тоже выводятся с мочой. 2. Нейтрализация Н2СО3, образующейся в почках, осуществляется с помощью ряда биохимических превращений (рис. 12.5) и выделения их продуктов с мочой. Рис. 12.5. Один из механизмов поддержания постоянства pH внутренней среды организма с помощью почек: — Н-помпа К–а — карбоангидраза; 342 Физиология 12.5. Количество, состав и выведение мочи С помощью почек из организма выводятся практически все азотсодержащие продукты обмена белков, содержание которых в моче, как и некоторых других веществ, например, глюкозы, свидетельствует о состоянии функции не только почек, но и некоторых других систем организма (например наличие глюкозы в моче, т.е. глюкозурия, может быть следствием нарушения эндокринной функции поджелудочной железы). В связи с этим анализ состава мочи представляет большой клинический интерес. С мочой выделяются мочевина, мочевая кислота, креатинин, натрий, калий, пигменты, образующиеся из билирубина, и другие токсические вещества, что и определяет цвет мочи. Белков и глюкозы в моче в норме нет, концентрация натрия в моче и в плазме крови одинакова. За сутки выделяется до 1,5 л мочи. При отсутствии АДГ в крови в течение суток выводится 15 л мочи. Объем диуреза сильно зависит от температуры воздуха: при ее повышении количество выделяемой мочи уменьшается вследствие большого потоотделения; от времени суток — ночью диурез уменьшается. Минимальный суточный объем мочи, достаточный для выполнения экскреторной функции почек, составляет 400 мл. Наполнение мочевого пузыря. В процессе мочеобразования моча продвигается по всем отделам нефрона благодаря деятельности сердца, создающего фильтрационное давление около 15 мм рт. ст., под действием которого моча начинает двигаться в капсуле Шумлянского–Боумена. Градиент давления в системе «капсула — почечная чашечка» составляет 15–0 мм рт. ст. Моча из собирательных трубок по выводным протокам поступает в чашечки, а из них — в лоханку, которая обладает автоматией — периодически сокращается и расслабляется. Заполнение лоханки во время ее расслабления (диастола) длится 4 с, опорожнение лоханки вследствие сокращения гладкой мускулатуры (систола) длится 3 с, при этом порция мочи выдавливается в мочеточник. Моча по мочеточнику движется вследствие наличия градиента давления, создаваемого систолой лоханки, и распространения по мочеточнику перистальтиче- Глава 12. Выделительная система 343 ской волны, которая возникает за счет автоматии гладкой мышцы мочеточника 1–5 раз в минуту. Парасимпатическая иннервация верхней части мочеточников осуществляется из блуждающего нерва, нижней части — из тазового нерва, возбуждение парасимпатических волокон ведет к усилению сокращений стенок мочеточников. Симпатическая иннервация мочеточников осуществляется из почечного, мочеточникового и нижнего подчревного сплетений, ее активация ведет к расслаблению мочеточников. Позыв к мочеиспусканию возникает при объеме мочи 150–200 мл. Если наполнение мочевого пузыря идет быстро, позывы к мочеиспусканию возникают чаще, т.к. быстрое растягивание гладких мышц вызывает более эффективное раздражение рецепторов растяжения. Дальнейшее наполнение мочевого пузыря до 300 мл с обычной скоростью резко усиливает поток афферентных импульсов, возникающих в механорецепторах и поступающих в спинной мозг, усиливается, естественно, и позыв к мочеиспусканию. В результате этого тормозится процесс мочеобразования в почке вследствие сужения сосудов почечных клубочков и уменьшения скорости клубочковой фильтрации. Повышение активности симпатической нервной системы создает условия для наполнения мочевого пузыря вследствие расслабления мышц мочевого пузыря, парасимпатической — для изгнания мочи из мочевого пузыря, вследствие возбуждения и сокращения мышц мочевого пузыря. При этом сфинктер мочевого пузыря расслабляется. Процессом мочеиспускания взрослый человек управляет произвольно. Центр коркового контроля мочеиспускания находится в первой сенсомоторной зоне на медиальной поверхности полушарий и в лимбической части коры мозга (в орбитальной области). Акт мочеиспускания этим центром может стимулироваться и тормозиться и может включаться и прерываться произвольно. Механизм произвольной регуляции мочеиспускания созревает в возрасте 1–2 лет, а в старости этот механизм может нарушаться. Афферентная импульсация от рецепторов уретры, возбуждающихся при мочеиспускании, поступает в спинной мозг по n. pudendus. Мочеиспусканию способствует также 344 Физиология натуживание (сокращение мышц брюшной стенки и дна таза, сужение грудной клетки), ведущее к повышению давления в брюшной полости и сдавливанию мочевого пузыря. После мочеиспускания в мочевом пузыре остается около 30 мл мочи, пузырь спадается, импульсы от него в ЦНС не поступают. По мере очередного наполнения мочевого пузыря вновь возникает афферентная импульсация — цикл повторяется. 12.6.Водно-солевой баланс Водно-солевой баланс (ВСБ) — это соотношение между поступающими в организм водой и солями и выделяемыми в окружающую среду. В норме у взрослого человека поддерживается равенство между поступлением в организм и выведением воды и солей (табл. 12.1), также и других питательных веществ. Показатели ВСБ — важные составляющие гомеостазиса. Однако ВСБ может быть положительным и отрицательным. Например, у растущего организма, беременных женщин этот баланс положительный. Во время голодания и недостаточного поступления воды и солей ВСБ отрицательный. Таблица 12.1 Водный баланс в норме, мл Поступление: 2500 Выделение: 2500 Питье и жидкая пища — 1200 С мочой — 1500 С твердой пищей — 900 С потом — 500, с калом — 100 Эндогенная вода (при окислении) — 400 С выдыхаемым воздухом — 400 Эндогенная вода образуется при окислении веществ: 100 г жира — 100 мл, 100 г белка — 40 мл, 100 г углеводов — 55 мл воды. Следует за метить, что в жаркое время с потом может выделяться несколько литров воды за сутки. Общее количество воды в организме составляет 50–70% от массы тела (38– 42 л): 1/3 от общего объема воды (плазма крови, лимфа, интерстиций, спинно-мозговая жидкость — ликвор) — это внеклеточная вода; 2/3 — внутриклеточная. Глава 12. Выделительная система 345 В тканях от 10 (жировая ткань) до 83–90% в почках и крови. Уменьшение воды в организме происходит с возрастом и при ожирении (жир содержит мало воды). Вся вода в организме обновляется за месяц, внеклеточная за неделю. Избыток воды в организме — это гипергидратация, а недостаток — гипогидратация. При гипергидратации вода накапливается в интерстиции, в результате могут развиться отеки и водная интоксикация: понижается осмотическое давление в интерстиции, набухают клетки, их осмотическое давление понижается, начинается возбуждение нервных центров и, как следствие, — судороги. Понижение содержания воды в интерстиции вызывает сгущение крови и, как следствие, нарушение гемодинамики. За счет жидкости интерстициального сектора происходит компенсация объема плазмы при острой кровопотере. Переливание значительного количества физиологического раствора не сопровождается значительным увеличением объема циркулирующей крови вследствие его перехода в межклеточную жидкость. Потеря 20% воды от массы тела приводит к смерти. Суммарная концентрация ионов солей формирует осмотическое давление, от которого зависит, в частности, количество воды в организме, величина ПП и ПД, возбудимость мышечных и нервных клеток. Например, снижение концентрации Са+2 в организме ведет к снижению величины ПП и повышению возбудимости нервных и мышечных клеток, что может сопровождаться судорогами, ослаблением сокращений сердца. Количество солей и воды в организме регулируется нейрогормональными механизмами с помощью почек (см. раздел 12.4, регуляцию питьевого и пищевого поведения). ГЛАВА 13 Глаз орла видит дальше, а глаз человека видит больше. Ф. Энгельс Сенсорные системы Одна из форм психической деятельности — ощущения, формирующиеся с помощью органов чувств. Сенсорная система (анализатор по И.П. Павлову) — совокупность центральных и периферических образований нервной системы, обеспечивающих восприятие раздражителей (изменения внешней и внутренней среды организма), их анализ и участие в формировании ответной реакции организма. Орган чувств — периферическое образование, воспринимающее и частично анализирующее факторы окружающей среды. Главная часть органа чувств — рецепторы. 13.1. Структурно-функциональная характеристика сенсорных систем Классификация сенсорных систем (CC). Обычно выделяют пять чувств: зрение, слух, вкус, обоняние и осязание. Но в действительности их значительно больше. Например, чувство осязания включает ощущение прикосновения, чувство давления — вибрации, мышечное чувство — температурное чувство и чувство щекотки. Есть и другие ощущения, например жажда. Существует несколько классификаций сенсорных систем по разным признакам, но нет единой классификации по какому-то общему критерию. В качестве такого критерия мы предлагаем роль (значение) той или иной сенсорной системы в жизнедеятельности организма (четыре основные группы): • сенсорные системы внешней среды воспринимают и анализируют изменения окружающей среды (системы Глава 13. Сенсорные системы 347 зрения, слуха, тактильная система, температуры, обоняния и вкуса). Роль этих систем — познание внешнего мира, приспособление к окружающей среде, поддержание тонуса ЦНС, что возможно благодаря импульсации от периферических отделов CC; • сенсорные системы внутренней среды воспринимают и анализируют изменения внутренней среды организма, показателей деятельности различных органов. Возбуждение внутренних CC субъективно обычно не воспринимается. Так, здоровый человек не может субъективно определить величину артериального давления, состояние сфинктеров и т.д. Информация, идущая из внутренней среды, играет важную роль в регуляции функций внутренних органов, в поддержании гомеостазиса, обеспечивая приспособление организма в различных условиях его жизнедеятельности. Таких сенсорных систем несколько: химизма внутренней среды (например, глюкорецепторы — один из вариантов периферического отдела внутренней СС), осмотического давления (осморецепторы), количества жидкости (волюморецепторы), наполнения полых внутренних органов (рецепторы растяжения), кровяного давления в артериях, система температуры (она имеет две части: внешней и внутренней среды); • сенсорные системы положения тела воспринимают и анализируют изменения положения тела в пространстве и частей тела относительно друг друга. Они играют важную роль в регуляции мышечного тонуса и поддержании с помощью этого естественной позы, в восстановлении нарушенной позы, в координации движений. К ним следует отнести вестибулярную и проприоцептивную системы. Поскольку организм оценивает положение тела или его частей относительно друг друга, эта импульсация доходит до сознания; • систему боли также следует выделить согласно ее особому значению — информированию о повреждающих действиях на организм и патологических процессах. 348 Физиология Согласно представлению И.П. Павлова, все анализаторы (сенсорные системы) построены по общему принципу, и каждый из них имеет три отдела. Отделы сенсорных систем: • периферический отдел. Основная часть периферического отдела СС — рецептор, назначение которого — восприятие раздражителей и первичный их анализ (изменений окружающей и внутренней среды организма). Это осуществляется с помощью трансформации энергии раздражения в РП, который обеспечивает с помощью своего электрического поля возбуждение окончания чувствительного нервного волокна; • проводниковый отдел СС, где происходит частичная обработка информации и доставка ее к коре большого мозга по афферентным (чувствительным) путям. Обработка информации осуществляется в ядрах продолговатого и среднего мозга, в гипоталамусе. В таламусе начинают формироваться ощущения: – чувствительный (проекционный) путь — от рецептора по строго обозначенным путям с переключением на различных уровнях ЦНС: на уровне спинного и продолговатого мозга, в зрительных буграх и в соответствующей проекционной зоне коры большого мозга (сенсорной области); – на уровне ствола мозга от чувствительного пути отходят коллатерали к клеткам ретикулярной формации, к которым могут конвергировать различные афферентные возбуждения, обеспечивая взаимодействие СС. При этом афферентные возбуждения теряют свои специфические свойства (сенсорную модальность) и изменяют возбудимость корковых нейронов. Блокада ретикулярной формации ведет к глубокому торможению и невозможности выполнения не только психической деятельности, но и двигательных произвольных реакций; • корковый отдел сенсорной системы. В данном отделе, согласно И.П. Павлову, выделяют первич- Глава 13. Сенсорные системы 349 ную и вторичную зоны. В первичную зону (IV слой коры, нейронные колонки) импульсы поступают от рецепторов одного вида по афферентным путям (зрительные, слуховые и т.д. — от моносенсорных рецепторов к моносенсорным нейронам). Их активация сопровождается ощущениями одной модальности (свет, звук). Во вторичную зону, которая расположена в непосредственной близости от первичной, импульсы поступают от первичной зоны и от других афферентных путей. Нейроны вторичной зоны — преимущественно бисенсорные, они возбуждаются при действии двух раздражителей. Например, нейроны вторичных зон зрительного и слухового анализаторов реагируют и на свет, и на звук. С помощью первичных зон возникают соответствующие раздражителю ощущения (звук, свет и т.д.). Во вторичных зонах осуществляется более сложная обработка информации о действующем раздражителе (например, более точное восприятие высоты, силы звука), формируется в сознании более полный образ. Афферентное возбуждение поступает также через ретикулярную формацию в ассоциативные области коры большого мозга (третичные зоны) и от вторичных зон. Взаимодействие возбуждений первичной, вторичной и третичной зон обеспечивает формирование в сознании как деталей объекта (параметры, форма, запах, температура, консистенция и др.) — анализ отдельных характеристик с помощью возникающих ощущений, так и комплексное представление об объекте — синтез. Возможно формирование и соответствующей поведенческой реакции организма. В целом это аналитико-синтетическая деятельность мозга. Нейроны коры, ответственные за формирование образов, назвали детекторными. Одни нейроны-детекторы отвечают за восприятие простых образов (например, для зрения — это линии, углы). Другие детекторы обеспечивают восприятие образа — например, образ степлера. 350 Физиология 13.2. Свойства СС, их значение, критерии для их оценки Сенсорные системы обеспечивают весьма тонкое приспособление организма к окружающей среде и познание ее, благодаря особым свойствам. Высокая чувствительность сенсорных систем к адекватному раздражителю — важнейшее свойство, обеспечивающее восприятие очень слабых раздражителей. Все отделы СС, и прежде всего рецепторы, обладают чрезвычайно высокой возбудимостью. Так, фоторецепторы сетчатки могут возбуждаться при действии лишь 1–2 квантов света, обонятельные рецепторы информируют организм о появлении единичных молекул пахучих веществ, но не любое возбуждение рецепторов вызывает ощущение. В связи с этим при рассмотрении этого свойства СС необходимо использовать термин не «возбудимость», а «чувствительность», поскольку его оценивают по возникновению ощущений. Возможность повышения чувствительности одной СС при утрате другой также играет важную приспособительную роль, например, у слепых повышается чувствительность системы слуха. Такие люди могут определить местоположение крупных объектов и обойти их, если нет посторонних шумов. Это осуществляется за счет отражения звуковых волн от находящегося впереди объекта и восприятия их. Способность к адаптации СС к постоянной силе длительно действующего раздражителя — также важное приспособительное свойство. Оно заключается в основном в понижении чувствительности. Это свойство присуще всем отделам СС, но наиболее ярко оно проявляется на уровне рецепторов и заключается в уменьшении не только их возбудимости и импульсации, но и в показателей функциональной мобильности, т.е. в изменении числа функционирующих рецепторных структур (Снякин П.Г.). В проводниковом отделе (в ядрах) и в коре адаптация проявляется в уменьшении числа соответственно активированных нервных клеток, что реализуется с помощью пре- и постсинаптического торможения. Способность к сенситизации (повышение чувствительности СС) может наблюдаться при возбуждении сим- Глава 13. Сенсорные системы 351 патической нервной системы, выбросе катехоламинов надпочечниками, при увеличении тироксина в крови, в условиях действия слабых раздражителей (например, темновая адаптация системы зрения, повышение чувствительности системы слуха при действии звука в тишине). Это свойство наряду с высокой чувствительностью и адаптацией также повышает разрешающую способность сенсорных систем. Инерционность сенсорных систем — отставание возникновения ощущений от начала действия раздражителя и их продолжение после выключения раздражения. Латентное время возникновения ощущений определяется латентным периодом возбуждения рецепторов и временем, необходимым для перехода возбуждения в синапсах от одного нейрона к другому, временем возбуждения ретикулярной формации и генерализации возбуждения в коре большого мозга. Сохранение на некоторый период времени ощущений после выключения раздражителя объясняется явлением последействия в ЦНС — в основном циркуляцией возбуждения (см. рис. 4.7). Так, латентный период зрительного ощущения равен 0,1 с, время последействия (последовательные образы) — 0,05 с. Быстро следующие одно за другим световые раздражения (мелькания) могут давать ощущение непрерывного света (феномен слияния мельканий). Максимальная частота вспышек света, которые воспринимаются еще раздельно, называется критической частотой мельканий. Она тем больше, чем сильнее яркость стимула и выше возбудимость ЦНС, и составляет 20–24 мельканий в секунду. Наряду с этим, если два неподвижных стимула последовательно с интервалом 20–200 мс проецировать на разные участки сетчатки, возникает ощущение движения объекта (фи-феномен). Такой эффект наблюдается даже в том случае, когда один стимул несколько отличается по форме от другого. Эти два феномена, слияние мельканий и фи-феномен, лежат в основе кинематографии. В силу инерционности восприятия зрительное ощущение от одного кадра длится до появления другого, отчего и возникает иллюзия непрерывного движения. Системный механизм восприятия раздражителей, выражающийся во взаимодополнении и взаимодействии 352 Физиология нескольких СС. Взаимодополнения выражаются в дополнительной оценке действующих на организм раздражителей с помощью различных СС. Например, качество дольки лимона оценивается с помощью зрительной, обонятельной, тактильной и вкусовой систем. Взаимодействие СС выражается в изменении возбудимости одной системы при возбуждении другой, что проявляется либо в ослаблении, либо в усилении соответствующих ощущений. Например, прослушивание музыки способно вызвать обезболивание при стоматологических процедурах — сверлении, шлифовке, экстракции зуба (аудиоаналгезия), шум ухудшает зрительное восприятие, при слабом освещении хуже воспринимается речь; яркий свет, напротив, повышает восприятие громкости звука и понимание речи. В основе взаимодействия СС лежит процесс формирования двух доминантных очагов возбуждения. Кроме того, взаимодействия реализуются посредством активации нейронов вторичных сенсорных зон, которые являются преимущественно бисенсорными. Например, человек лучше слышит при ярком свете потому, что при этом возбуждаются под действием света бисенсорные нейроны вторичной сенсорной зоны системы слуха. Критерии для оценки чувствительности СС. Порог ощущения (абсолютный порог) — это минимальная сила раздражения, воспринимаемая субъективно в виде ощущения. Пороговая сила стимула для СС очень мала. Например, система слуха способна различать звук, интенсивность которого равна одной трехмиллионной ватта на квадратный метр. Порог различения (дифференциальный порог) — минимальное изменение силы действующего раздражителя, воспринимаемое субъективно в виде изменения интенсивности ощущения. Эту закономерность установил Э. Вебер в опыте с определением по ощущению испытуемого силы давления на ладонь. Оказалось, что при действии груза в 100 г для ощущения прироста давления необходимо было добавить груз в 3 г, при действии груза в 200 г — добавить 6 г, в 400 г — 12 г и т.д. При этом отношение прироста силы раздражения (ΔI) к силе действующего раздражителя (I) есть величина постоянная: ΔI/I = const. Глава 13. Сенсорные системы 353 У разных СС эта величина различна, в данном случае она равна примерно 1/30. Подобная закономерность наблюдается и при уменьшении силы раздражителя. Порог пространства — минимальное расстояние между двумя раздражаемыми точками, при котором они еще воспринимаются раздельно. Пороги пространства на коже весьма вариабельны: на губах и кончиках пальцев — 2,0– 2,5 мм, на ладони — 9 мм, на спине — 55 мм. Интенсивность ощущений также может быть критерием чувствительности СС, поскольку интенсивность ощущения, возникающего при одной и той же силе раздражителя, зависит от состояния возбудимости СС на всех ее уровнях. Эту закономерность изучил Г. Фехнер, показавший, что интенсивность ощущения прямо пропорциональна логарифму силы раздражения. Это положение выражается формулой: E = a × log I + b, где Е — интенсивность ощущений; a и b — константы, различные для разных СС; I — сила действующего раздражителя. Интенсивность ощущений определяется уровнем возбуждения во всех отделах сенсорной системы, в том числе и на уровне рецепторов (рис. 13.1). Рис. 13.1. Торможение сенсорных рецепторов (1) с помощью АТФ, высвобождаемой соседними рецепторами под влиянием импульсов, распространяющихся антидромно по разветвлению дендрита афферентного нейрона (2) 354 Физиология 13.3. Кодирование информации в нервной системе Кодирование — это процесс преобразования информации в условную форму (код), удобную для передачи по каналу связи. Любое преобразование информации в отделах СС является кодированием. Так, в слуховой системе механическое колебание перепонки и других звукопроводящих элементов на первом этапе преобразуется в рецепторный потенциал, последний обеспечивает секрецию медиатора в синаптическую щель и возникновение генераторного потенциала, в результате действия электрического поля которого в афферентном волокне возникает нервный импульс (ПД). ПД достигает следующего нейрона, в синапсе которого электрический сигнал снова преобразуется в химический, химический — снова в электрический, многократно меняется код. Роль структурно-функциональной организации ЦНС в кодировании заключается в том, что она обеспечивает преимущественное распространение возбуждения между определенными отделами ЦНС и частями тела организма с помощью специфических афферентных путей — меченая линия. Коды нервной системы. Универсальный код — нервный импульс, который распространяется по нервным волокнам. Передача сигнала от одной клетки к другой во всех отделах СС осуществляется с помощью химического кода — медиатора. Хранение информации в ЦНС кодируется с помощью нервных импульсов, биохимических процессов и структурных изменений в нейронах (механизмы памяти). Кодируемые характеристики раздражителя. В СС кодируется качественная характеристика раздражителя (его вид), сила раздражителя, время его действия, а также пространство, т.е. место действия раздражителя на организм и локализация его в окружающей среде. Поскольку в кодировании характеристик раздражителя принимают участие все отделы СС, рассмотрим процессы кодирования в каждом отделе. Кодирование в периферическом отделе СС. Кодирование вида раздражителя осуществляется за счет специфичности рецепторов, способности воспринимать раздражитель определенного вида, который рецептор приспособился Глава 13. Сенсорные системы 355 воспринимать в процессе эволюции (адекватный раздражитель). Так, световой луч возбуждает только рецепторы сетчатки, другие рецепторы (обоняния, вкуса, тактильные и т.д.) на него не реагируют, что и обеспечивает организму способность отличать один вид раздражителя от другого. Сила раздражителя кодируется величиной рецепторного потенциала и, следовательно, изменением частоты импульсов, возникающих в окончании чувствительного нервного волокна при изменении силы действующего раздражителя. В конечном итоге от силы раздражителя зависит общее число импульсов, возникающих в единицу времени. Это так называемое частотное кодирование. Как правило, с увеличением силы стимула возрастает число импульсов, возникающих в нервном окончании, и наоборот. При изменении силы раздражителя может изменяться также и число возбужденных рецепторов. Кодирование пространства. Локализация действия раздражителя кодируется тем, что возбуждаются только те рецепторы, на которые действуют раздражители, при этом рецепторы различных участков тела посылают импульсы в определенные зоны коры большого мозга, в этом и заключается пространственное кодирование. Пространство на теле организма кодируется также величиной площади, на которой возбуждаются рецепторы. Например, мы легко определяем, острым или тупым концом карандаш касается поверхности кожи. Время действия раздражителя кодируется тем, что рецепторы начинают возбуждаться с началом действия раздражителя и прекращают возбуждаться сразу после выключения раздражителя (временное кодирование). Также кодируется и частота раздражения. В целом рецепторы обеспечивают формирование структуры ответа, который поступает в проводниковый отдел СС (частота импульсов, формирование их в пачки, число импульсов в пачке, интервалы между пачками и отдельными импульсами в пачке). В структуре ответа закодированы все характеристики действующего раздражителя, вид, сила, пространство и время, а также все оттенки действующего раздражителя (например, высота звука, сплошной или прерывистый звук). Кодирование в проводниковом отделе СС осуществляется только на «станциях переключения», т.е. при передаче 356 Физиология сигнала от одного нейрона к другому, где происходит смена кода (в ядрах). В нервных волокнах информация не кодируется, а только передается к другим клеткам. По мере поступления импульсов к вышележащим отделам ЦНС уменьшается частота разрядов нейронов, длительная импульсация превращается в короткие пачки импульсов (паттерны). Продолжительность разряда большинства нейронов уже не соответствует длительности стимула. В таламусе начинают формироваться ощущения. Кодирование в корковом отделе СС характеризуется тем, что электрофизиологические процессы (кратковременная память) вызывают химические реакции в нейронах (нейрохимическая — промежуточная память), которые, в свою очередь, вызывают структурные изменения в нейронах — информация кодируется с помощью синтеза белка (структурная — долговременная память). Посредством глаза, а не глазом, Смотреть на мир умеет разум. У. Блейк 13.4. Система зрения Система зрения — совокупность структур, обеспечивающих восприятие электромагнитных колебаний определенного участка спектра (волны длиной 390–750 нм), передачу, обработку зрительной информации и формирование зрительных ощущений. Через систему зрения человек получает 80–90% осознаваемой информации об окружающем мире. Система зрения обеспечивает различение цветов, крупных объектов и их деталей, подвижных и неподвижных, разноудаленных от организма и между собой. Отделы системы зрения. Периферический отдел системы зрения — фоторецептор, локализующийся в глазу (орган зрения, который включает также часть проводникового отдела и вспомогательные структуры) (рис. 13.2). Фоторецепторы реагируют на кванты света и преобразуют зрительную информацию в нервные импульсы для передачи ее по афферентным путям зрительного нерва в ЦНС. Глава 13. Сенсорные системы 357 Передняя камера Задняя камера Хрусталик Радужная оболочка Роговица Конъюнктива Ресничное тело Ресничная мышца Сетчатка Склера Сосудистая оболочка Зрительный нерв Ресничные отростки Оптическая ось Сосок зрительного нерва Цинновы связки Зрительная ось Центральная ямка Стекловидное тело Желтое пятно Рис. 13.2. Орган зрения Вспомогательные структуры глаза: • оптическая система — слезная жидкость, роговица, водянистая влага, хрусталик, стекловидное тело. Обеспечивает фокусирование световых лучей на сетчатке и формирование в области ее центральной ямки перевернутого изображения рассматриваемых зрительных объектов и явлений; внутриглазное давление 12– 20 мм рт. ст. При высоком давлении повреждается зрительный нерв, что может привести к слепоте; • глазодвигательный аппарат — наружные мышцы глазного яблока (четыре прямые и две косые), подниматель верхнего века и орбитальная мышца; внутренние мышцы глаза обеспечивают фиксацию, поворот глаз и установку зрительных осей; • защитные органы включают: веки, ресницы, конъюнктиву, слезный аппарат, фасции глазницы. Рецепторы зрительной системы включают четыре вида фоторецепторов: один вид палочек и три — колбочек. Рецепторный слой сетчатки прилежит к слою клеток пигментного эпителия (рис. 13.3). Всего в сетчатке выделяют 10 слоев, и кванты света могут достигнуть фоторецепторов, только пройдя через слои сетчатки, расположенные кпереди от рецепторного слоя (за исключением области центральной ямки, где все эти слои 358 Физиология Пигментный слой Колбочки Палочки Ядра палочек Горизонтальные клетки Биполярные клетки Амакриновые клетки А Ганглиозные клетки Б Рис. 13.3. Фоторецепторы и нейронная организация сетчатки (по А.К. Гайтону, Д.Э. Холлу, 2008): А — периферическая область; Б — область центральной ямки «сдвинуты», и свет сразу попадает на колбочки, составляющие основу этой части сетчатки). Палочки содержат пигмент родопсин, поглощающий электромагнитное излучение в диапазоне 400–620 нм. Колбочки имеются трех типов, в каждом содержится по одному пигменту (сине-голубой, зеленый и красный). Пигменты фоторецепторов состоят из ретиналя (альдегид витамина А, 11-цис-изомер) и гликопротеида опсина. При поглощении квантов света в фоторецепторах происходят фотохимические (распад пигментов) и электрофизиологические процессы, вследствие чего увеличивается афферентный поток импульсов. В условиях постоянного и равномерного освещения фотохимический распад пигментов и их ресинтез находятся в равновесии. При повышении освещенности распад пигментов увеличивается и преобладает над ресинтезом, вследствие чего чувствительность фоторецепторов Глава 13. Сенсорные системы 359 уменьшается. При уменьшении освещенности ресинтез пигментов возрастает и превосходит их распад, что ведет к многократному увеличению светочувствительности фоторецепторов. Эти процессы лежат в основе световой и темновой адаптации фоторецепторов. Световая адаптация осуществляется примерно за 60 с, а темновая проходит в несколько этапов и достигается к 30–60 мин. При недостатке в организме витаминов (А, В1, В2, С), особенно витамина А или его предшественника β-каротина, может развиться гемералопия («куриная слепота») — нарушение темновой адаптации, проявляющееся снижением ночного и сумеречного зрения. Проводниковый отдел (рис. 13.4) обеспечивает доставку информации в корковый отдел и ее обработку в центрах ствола мозга и промежуточного мозга (на «станциях переключения») — особо важную роль играет таламическая область. Процессы взаимодействия нейронов сетчатки идут в двух взаимосвязанных направлениях — горизонтальном и вертикальном (рис. 13.3). На уровне сетчатки осуществляются обработка и передача зрительных сигналов, в которых участвуют многие функциональные типы нейронов, используются различные нейромедиаторы, обеспечивающие взаимные возбуждающие и тормозные влияния. Биполярные нейроны (1-й вертикальный нейрон) получают сигналы с помощью медиатора от фоторецепторов и передают их с помощью ацетилхолина на ганглиозные нейроны, где впервые возникает ПД (2-й вертикальный нейрон). Горизонтальное распространение возбуждения обеспечивает взаимодействие между нейронами сетчатки. Аксоны РП 1 ГП ПД 2 3 4 КБМ Рис. 13.4. Нейронная организация системы зрения: 1 — рецептор (вторичный), сетчатка; 2 — биполярная нервная клетка, сетчатка; 3 — ганглиозная нервная клетка, сетчатка; 4 — нейрон латерального коленчатого тела; РП — рецепторный потенциал; ГП — генераторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (затылочная доля) 360 Физиология ганглиозных клеток формируют зрительный нерв, информация по которому передается в ЦНС. Проводниковый отдел вне сетчатки начинается зрительными нервами; после частичного перекреста их волокон (от внутренних носовых поверхностей сетчатки каждого глаза) — это зрительные тракты, а после латеральных коленчатых тел — зрительная лучистость (рис. 13.5, см. цв. вклейку). Эти зрительные пути проводят афферентные сигналы к коре большого мозга. В латеральных коленчатых телах оценивается освещенность, в верхнем двухолмии четверохолмия — движение объекта. Корковый отдел системы зрения. Первичная сенсорная зона (стриарная кора) локализуется в затылочной области (поле 17 по Бродману), в ней представлена ретинотопическая карта зрительных полей. Она играет главную роль в формировании зрительных образов. Повреждение у человека затылочных долей ведет к слепоте. Название «стриарная кора» (полосатая кора) объясняется тем, что плотный пучок аксонов зрительной лучистости слоя 4 образует полосу. Вторичная сенсорная зона расположена в экстрастриарной области (за пределами) первичной зрительной коры. Зрительная кора на каждой стороне мозга получает сигналы от ипсилатеральных половин сетчатки обоих глаз и контралатеральных половин полей зрения. Совместная работа первичной, вторичной и третичной (ассоциативной) сенсорных зон коры обеспечивает распознавание зрительных объектов, зрительное внимание, выполнение целенаправленных действий под зрительным контролем. Механизмы глаза, обеспечивающие ясное зрение в различных условиях 1. При рассматривании разноудаленных от наблюдателя объектов включаются несколько механизмов, главный из которых аккомодация — это процесс сохранения изображения объекта на сетчатке глаза за счет изменения преломляющей силы хрусталика. Кривизна хрусталика увеличивается при сокращении ресничной мышцы вследствие ослабления натяжения цинновой связки. При этом хрусталик за счет своей эластичности (упругости) становится более выпуклым, преломляющая сила его увеличивается, глаз Глава 13. Сенсорные системы 361 настраивается на видение близко расположенных предметов. При рассматривании более удаленных предметов ресничная мышца расслабляется, циннова связка натягивается, что ведет к натяжению капсулы хрусталика, его уплощению и снижению преломляющей силы хрусталика. Адекватный стимул для изменения степени аккомодации — это нечеткость изображения на сетчатке — ретинальный рефлекс. Иннервируется цилиарная мышца волокнами парасимпатической ветви III пары черепных нервов. Нормальная рефракция называется эмметропией (рис. 13.6). Но нередко встречаются и аномалии рефракции глаза. Миопия (близорукость) — рефракция глаза, при которой лучи от объекта после прохождения через оптический аппарат фокусируются впереди сетчатки (см. рис. 13.6). Эмметропия Гиперметропия Коррекция гиперметропии Миопия Коррекция миопии Рис. 13.6. Виды рефракции и линзы для коррекции ее нарушений (ход лучей через преломляющие среды глаза) 362 Физиология Гиперметропия (дальнозоркость) — рефракция глаза, при которой лучи от далеко расположенных объектов фокусируются за сетчаткой (рис. 13.6). При гиперметропии нечеткость зрения возникает при рассматривании близко расположенных предметов. Астигматизм — нарушение рефракции глаза, обусловленное различной кривизной роговицы и/или хрусталика в разных меридианах. Вергентные движения глаз — это также приспособление для ясного видения разноудаленных предметов. Конвергенция (сведение) зрительных осей происходит в случае рассматривания близко расположенных объектов, дивергенция (разведение) осей — при удалении объекта. Это обеспечивает удержание изображения объекта в центре желтого пятна. Конвергентная рефлекторная реакция зрачков: при рассматривании близко расположенных объектов (их сужение) и при рассматривании более удаленных объектов (расширение) также способствует ясному зрению. Бинокулярное зрение тоже улучшает восприятие разноудаленных предметов. Оно формируется в процессе опыта на основе механизмов зрительной коры; обеспечивает слияние сетчаточных изображений (фузии) от обеих сетчаток в единое целое. 2. В условиях изменения освещенности ясное зрение обеспечивают два основных механизма. Во-первых, темновая и световая адаптация — они главные. Они осуществляются с помощью фотохимических процессов (расщепление зрительных пигментов в колбочках и палочках на свету и их ресинтез в темноте), а также с помощью увеличения размеров рецептивных полей биполярных и ганглиозных клеток. Во-вторых, зрачковый рефлекс. Он может изменять интенсивность светового потока, попадающего на сетчатку, примерно в 30 раз за счет изменения диаметра зрачка. При уменьшении освещенности происходит расширение зрачка (мидриаз) за счет сокращения мышцы-дилататора, иннервируемой волокнами от верхнего шейного симпатического ганглия. При увеличении освещенности и рассматривании близко расположенных предметов происходит сужение зрачка (миоз) за счет сокращения мышечного сфинкте- Глава 13. Сенсорные системы 363 ра, который регулируется парасимпатическим нервом (III пара черепных нервов). 3. Восприятие крупных объектов и их деталей обеспечивается посредством центрального и периферического зрения. Центральное зрение обеспечивается в том случае, если изображение предмета попадает в центральную ямку сетчатки глаза — наибольшая острота зрения, которая определяется наименьшим углом зрения, при котором глаз еще способен раздельно воспринимать две светящиеся точки. В норме он составляет 1 мин. Периферическое зрение обеспечивает видение крупных объектов, что объясняется большим полем зрения (видимое пространство при фиксированном взоре). Используется также произвольное движение глаз, как при чтении — перевод взора с одной точки на другую. 4. При оценке неподвижного объекта возможны два варианта приспособления. Во-первых, непроизвольные движения глаз — при фиксации взора возможна адаптация фоторецепторов. Но она предотвращается с помощью быстрых непроизвольных постоянных содружественных движений глаз (тремор), совершаемых с большой скоростью (частота колебаний — 30–80 Гц) и превращающих постоянный раздражитель в прерывистый, т.к. световые лучи быстро смещаются от одних рецепторов сетчатки к другим, поэтому не успевает произойти их адаптация. Если заблокировать эти движения глаз, то окружающий мир, вследствие адаптации рецепторов сетчатки, станет трудно различимым, каков он, например, у лягушки. Глаза совершают также постоянные медленные движения (медленный дрейф). При этом в случае фиксации взора и попадания изображения вследствие дрейфа на край центральной ямки срабатывает (как и при рассматривании подвижных объектов) фиксационный рефлекс (саккады) — глаз «нашел» объект, изображение переводится на центр ямки. Во-вторых, произвольные движения глаз — при смещении взора с одной детали неподвижного объекта на другую и при чтении используются плавные произвольные движения глаз с помощью глазных мышц (рис. 13.7), а также поворота и наклона головы, но это не приспособление глаз. Произвольные движения глаз осуществляют- 364 Физиология Прямая верхняя Косая нижняя Косая верхняя Прямая нижняя Прямая медиальная III ЧН Прямая латеральная Ядра IV ЧН VI ЧН Медиальный продольный пучок Рис. 13.7. Нервы и мышцы глаза, управляющие движениями глаз ся с помощью корковых центров, а непроизвольные — посредством верхних бугорков четверохолмия. 5. Восприятию движущихся объектов способствуют произвольные и непроизвольные содружественнные движения глаз. Произвольные движения глаз со скоростью движения объекта — влево, вправо, вверх, вниз — осуществляются благодаря содружественной деятельности глазодвигательных мышц и обеспечивают совмещение изображения в центральной ямке сетчатки. Плавные, непроизвольные следящие движения глаз за перемещающимся в поле зрения объектом обеспечивают совмещение изображения на сетчатке с центральной ямкой. Если изображение смещается на край центральной ямки, срабатывает (как и при рассматривании неподвижных объектов) фиксационный рефлекс — непроизвольные быстрые скачкообразные мелкие движения (саккады), обеспечивающие коррекцию совмещения изображения на сетчатке с центральной ямкой. Саккады возникают с частотой 2–3 раза в секунду. 6. Цветовое (хроматическое) зрение — это способность зрительной системы различать электромагнитные Глава 13. Сенсорные системы 365 световые волны по их длине с формированием ощущения цвета. Цветовые оттенки возникают в результате смешения семи чистых тонов спектра. Последние, в свою очередь, образуются из трех основных, чистых цветов (тонов) — красного (700 нм), зеленого (546 нм) и синего (435 нм) (рис. 13.8). Согласно трехкомпонентной теории цветовосприятия Ломоносова–Юнга–Гельмгольца–Лазарева, в сетчатке имеется три вида колбочек, каждый из которых содержит специфический светочувствительный пигмент, обладающий наибольшей чувствительностью к одному из трех основных, чистых цветов. Комбинации возбуждения различных колбочек приводят к ощущению различных цветов и оттенков. Равномерное возбуждение колбочек дает ощущение белого цвета. Встречаются протанопия (дальтонизм) — слепота на красный свет (описана физиком Дж. Дальтоном); дейтеранопия — невосприятие зеленого цвета; тританопия — цветовая слепота на фиолетовый цвет и дефектное зрение на синий и желтый. Исследуют цветовое зрение (что необходимо при профотборе) с помощью приборов (аномалоскопов) и таблиц; из последних наибольшее признание получили таблицы Рабкина. Относительная чувствительность колбочек, % 100 «Синий тип» «Зеленый тип» «Красный тип» 50 0 400 500 600 Длина световой волны, нм 700 Рис. 13.8. Спектры цветовой чувствительности колбочек трех типов (воспринимающих синий, зеленый и красный цвета) в сетчатке глаза человека 366 Физиология 13.5. Система слуха Человек воспринимает звук (механические колебания среды) в диапазоне 20–20 000 Гц. Колебания ниже 20 Гц (инфразвуки) и выше 20 000 Гц (ультразвуки) человек не ощущает. Периферический отдел системы слуха — ухо. Наружное ухо играет роль рупора, который усиливает звуки путем концентрации их в направлении барабанной перепонки, а также защищает последнюю от воздействий внешней среды. Среднее ухо представляет собой небольшую полость, наполненную воздухом, со слуховыми косточками и слуховой (евстахиевой) трубой, включает также ячейки сосцевидного отростка (рис. 13.9). Оно отделено мембраной (барабанной перепонкой) от наружного слухового прохода. Слуховые косточки: рукоятка молоточка вплетена в бараМолоточек Наковальня Овальное окно Базальная мембрана Стремечко Круглое окно Барабанная перепонка А Три канала улитки Овальное окно Вестибулярная лестница Геликотрема Круглое окно Б Тимпаническая лестница Средняя лестница Рис. 13.9. Каналы улитки (по П. Линдсею, Д. Норманну, 1974): А — среднее и внутреннее ухо в разрезе; Б — распространение звуковых колебаний в улитке Глава 13. Сенсорные системы 367 банную перепонку, другой его конец образует суставную поверхность с наковальней, которая, в свою очередь, сочленена со стремечком. Стремечко прилегает к мембране так называемого овального окна и обеспечивает передачу колебаний на слуховые рецепторы внутреннего уха. В результате взаимодействия слуховых косточек, барабанной перепонки и ушной раковины происходит усиление давления звуковых волн на мембрану овального окна в 120 раз. Звуки проводятся также по костям черепа. Исследование воздушной и костной проводимости у пациента помогает врачу в постановке диагноза заболевания. Внутреннее ухо — это система сообщающихся заполненных жидкостью каналов и полостей в лабиринте височной кости (рис. 13.9, А, Б). Здесь расположены два органа: орган слуха — улитка и орган равновесия — вестибулярный аппарат. Улитка — спирально извитой костный канал, имеющий у человека 2,5 оборота (завитка) и разделенный основной (базальной) мембраной и мембраной Рейснера на три узкие части («лестницы»). Между верхним и нижним каналом находится средний (средняя лестница). Он изолирован и заполнен эндолимфой. Слуховые рецепторы расположены внутри среднего канала на базальной мембране (кортиев орган) (рис. 13.10). Звуковые колебания передаются от стремечка на овальное окно, а оттуда сообщаются перилимфе. Колебания перилимфы, в свою очередь, распространяются на базальную мембрану. РП возникает в волосковых клетках (рецепторах) в результате колебания базальной мембраны и деформации волосков (стереоцилий). РП обеспечивает секрецию медиатора (повидимому, глутамат), который инициирует возникновение ГП. Последний с помощью своего электрического поля возбуждает чувствительные окончания волокон слухового нерва. Проводниковый отдел системы слуха начинается с биполярного нейрона, локализующегося в спиральном ганглии (рис. 13.11), дендрит нейрона синаптически связан с рецепторной клеткой кортиева органа (вторичный рецептор). Как отмечалось выше, под действием медиатора на постсинаптической мембране (окончания дендрита 1-го нейрона системы слуха) развивается ГП, который за счет своего электрического поля обеспечивает возник- 368 Физиология Рейснерова мембрана Покровная мембрана Вестибулярная лестница Средняя лестница Волосковые клетки кортиева органа Барабанные лестницы Основная мембрана Спиральный ганглий Рис. 13.10. Поперечный разрез кортиева органа новение ПД в первом перехвате Ранвье этих окончаний (рис. 13.11). Корковый отдел системы слуха локализуется в верхней части височной доли коры большого мозга: первичная слуховая кора — поля 41 и 42, вторичная — поля 21 и 22. Нервные импульсы от медиального коленчатого тела в основном направляются в первичную слуховую кору. Вторичная кора активируется импульсами таламуса и от первичной коры. В результате взаимодействия возбуждений первичной и вторичной слуховых зон и ассоциативной коры (третичная зона для каждого анализатора) формируются соответствующие ощущения. РП ГП 1 ПД 2 3 4 5 6 КБМ Рис. 13.11. Нейронная организация системы слуха: 1 — рецептор (вторичный), кортиев орган — улитка; 2 — чувствительный нейрон спирального ганглия улитки; 3 — нейроны кохлеарного комплекса продолговатого мозга (улитковое ядро); 4 — ядро оливы, мост; 5 — нижнее двухолмие; 6 — нейроны медиального коленчатого тела; РП — рецепторный потенциал; ГП — генераторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (верхняя височная извилина) Глава 13. Сенсорные системы 369 Кодирование высоты звуков осуществляется с помощью изменения частоты и локализации колебаний базальной мембраны в зависимости от частоты звука (частотное и пространственное кодирование). При частотном кодировании звуки низкой частоты (до 400 Гц) приводят в колебательные движения всю мембрану, но особенно сильно в области, ближайшей к геликотреме (верхний отдел улитки, где более длинные волокна основной мембраны). Частотное кодирование заключается в том, что частота афферентных импульсов в слуховом нерве повторяет частоту звуковых колебаний, поскольку она не превышает лабильность этих нервных волокон. Пространственное кодирование начинается с увеличением частоты колебаний звуковых волн выше 400 Гц. При этом максимальная амплитуда колебаний основной мембраны смещается к нижней части улитки (ближе к овальному окну), где расположены более короткие волокна базальной мембраны. В этом случае частота возникающих в слуховом нерве ПД не возрастает (предел лабильности). Характер импульсации меняется только в соответствии с местом наиболее интенсивных колебаний базальной мембраны (теория места). Информация в виде нервных импульсов передается по определенным волокнам слухового нерва, связанным с участком базальной мембраны, колебания которой наибольшие. Кодирование силы звука сочетает в себе частотное и пространственное кодирование. Во-первых, оно осуществляется за счет увеличения амплитуды колебаний базальной мембраны, что ведет к возбуждению менее возбудимых рецепторов и к возрастанию амплитуды РП и, следовательно, частоты ПД, распространяющихся по слуховому нерву в корковый отдел системы слуха (частотное кодирование). Во-вторых, чем сильнее звук, тем больше площадь эпицентра колебаний базальной мембраны и, соответственно, большее число волосковых клеток одновременно вовлекается в колебательный процесс (пространственное кодирование). Определение локализации источника звука обусловлено разницей силы звуковых волн, поступающих к каждому уху, и различием во времени прихода одинаковых фаз звуковой волны к обоим ушам, что обеспечивает бинауральный слух. 370 Физиология 13.6. Системы положения тела Системы положения тела включают вестибулярную и проприоцептивную системы. Вестибулярная система обеспечивает восприятие изменений положения головы и всего тела в пространстве, прямолинейных или вращательных ускорений при движениях тела (акселерационное чувство), участвует в регуляции тонуса мышц, поддерживая тем самым естественную позу и восстанавливая нарушенную позу, а также принимает участие в координации движений. Периферический отдел — вестибулярный орган, расположен вместе с улиткой в лабиринте височной кости и состоит из преддверия и трех полукружных каналов (рис. 13.12), заполненных эндолимфой и содержащих волосковые механорецепторы (вторичные рецепторы). Рецепторы покрыты желеобразной массой. При линейном ускорении, наклонах головы или всего тела, при угловом ускорении различные рецепторы раздражаются желеобразной массой вследствие ее инерционности и некоторого отставания в смещении относительно тела. В рецепторах (они вторичные) возникают РП, секретируется медиатор, под влиянием которого возникает ГП, обеспечивающие возникновение ПД в чувствительном нервном окончании. Рецепторная клетка представлена на рис. 13.13. Проводниковый отдел начинается дендритом биполярного нейрона (первый нейрон), синаптически связанного Ампула Ампула Рис. 13.12. Вестибулярный аппарат (по E. Kandel, J. Schwartz) Глава 13. Сенсорные системы 371 Рис. 13.13. Структурно-функциональные элементы волосковой (рецепторной) клетки вестибулярного аппарата: 1 — эфферентное нервное волокно, регулирующее функцию рецепторной клетки; 2 — афферентное (чувствительное) нервное волокно, на которое рецепторная клетка передает информацию с помощью медиатора с волосковой рецепторной клеткой (медиатор, по-видимому, ацетилхолин) (рис. 13.14). По аксонам биполярных нейронов, расположенных во внутреннем слуховом проходе в вестибулярных ганглиях, возбуждение направляется к четырем вестибулярным ядрам продолговатого мозга (главное из которых ядро Дейтерса). Нейроны этих ядер (второй нейрон) получают дополнительную информацию от проприорецепторов (прежде всего, шейных), рецепторов сетчатки и мышц глаза, мозжечка. В свою очередь, они посылают информацию к спинному мозгу, мозжечку, к ядрам глазодвигательных нервов и к таламусу — третьему нейрону вестибулярной системы. Аксоны этих нейронов образуют прямой путь в кору большого мозга к четвертым нейронам. РП ГП ПД 1 2 3 4 КБМ Рис. 13.14. Нейронная организация вестибулярной системы: 1 — рецептор (вторичный), вестибулярный орган, лабиринт височной кости; 2 — чувствительный нейрон вестибулярного ганглия, дно внутреннего слухового прохода; 3 — нейроны вестибулярных ядер (продолговатый мозг); 4 — таламус; РП — рецепторный потенциал; ГП — генераторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (височная доля) 372 Физиология Корковый отдел представлен нейронами, рассеянными в постцентральной извилине, верхней височной извилине, верхней теменной дольке (поля 20 и 21 по Бродману). В премоторной коре имеется также вторая вестибулярная зона, в которую поступает информация по непрямому вестибулоцеребеллоталамическому пути. С помощью взаимодействия этих возбуждений формируются ответные реакции организма (вестибулярные реакции). Соматические реакции участвуют в сохранении естественной позы с помощью регуляции тонуса мышц, в восстановлении нарушенной позы, в координации движений, в формировании глазодвигательных реакций. Сенсорные реакции возникают при интенсивных раздражениях вестибулорецепторов и проявляются чувством головокружения, воспринимаемым как движения окружающих предметов. Вегетативные реакции — это комплекс разнообразных вегетативных рефлексов, осуществляемых за счет связей вестибулярных ядер с вегетативными центрами гипоталамуса и ствола мозга. В обычных условиях эти реакции участвуют в регуляции обменных процессов в мышцах. Могут возникнуть патологические реакции (тошнота, рвота, саливация, потливость, тахи- или брадикардия) — кинетозы, морская болезнь, воздушная, транспортная, космическая. Проприоцептивная система. Она обеспечивает возникновение ощущений положения тела и его частей в пространстве в результате действия силы гравитации и возбуждения проприорецепторов. Эта система играет важную роль в регуляции тонуса мышц и в координации движений (менее точное название ее — «двигательная»). Периферический отдел представлен первичными рецепторами мышц, сухожилий и суставных сумок (рис. 13.15). Адекватный раздражитель проприорецепторов — сокращения и расслабления мышц (см. в разделе 4.11). Проводниковый отдел. Афферентная импульсация в кору большого мозга направляется по лемнисковому пути от чувствительных нейронов спинномозговых узлов — первый нейрон, их аксоны в составе задних канатиков спинного мозга достигают нейронов ядер Голля и Бурдаха в продолговатом мозге — второй нейрон. Аксоны нейронов в составе медиальной петли (лемниск) после перекреста на- Глава 13. Сенсорные системы 373 Рис. 13.15. Нейронная организация проприоцептивной (двигательной) системы: 1 — проприорецептор (первичный, мышцы, связки, сухожилия, суставные сумки, фасции); 2 — спинальный ганглий (для головы — ганглии черепных нервов); 3 — ствол мозга; 4 — таламус; РП — рецепторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (постцентральная извилина, теменная доля) правляются в сенсорные ядра таламуса, где располагается третий нейрон. Импульсация от проприорецепторов поступает также в мозжечок по путям Флексинга и Говерса. Корковый отдел представлен нейронами 1-й и 2-й соматосенсорных областей (S1 и S1 — четвертый нейрон) постцентральной извилины (поля 1–3), к которым импульсация поступает от нейронов таламуса. Поступающая в кору большого мозга импульсация обеспечивает осознание положения частей тела и их движение, участвует в регуляции мышечного тонуса. 13.7. Сенсорные системы кожи Система температуры участвует в поддержании температурного гомеостазиса с помощью изменения интенсивности теплопродукции и теплоотдачи и поведенческих приспособительных реакций, когда температура внешней среды становится некомфортной. Периферический отдел — это холодовые и тепловые терморецепторы кожи, слизистой оболочки полости рта и ее органов. Тепловые рецепторы представлены тельцами (гроздьями) Руффини, холодовые — колбами Краузе. Кожные рецепторы холода расположены в эпителии или непосредственно под ним, рецепторы тепла залегают глубже — в сосочковом слое дермы. Кроме того, терморецепторы располагаются в слизистой полости рта. Причем холодовые рецепторы преобладают в передних отделах слизистой, а тепловые — в задних. Эти особенности обусловливают более высокую чувствительность кожи и слизистой оболочки к 374 Физиология холоду, чем к теплу, и имеют ведущее значение их в определении температуры внешней среды. Терморецепторы внутренних органов воспринимают изменения температуры внутренней среды организма. Они локализованы в слизистых оболочках пищеварительного тракта, органов дыхания, мочевого пузыря, в серозных оболочках, в стенках артерий и вен, в каротидном синусе, а также в ядрах гипоталамуса. Проводниковый отдел представлен тонкими миелинизированными волокнами типа Аδ — это аксоны чувствительных спинальных ганглиев и ганглиев тройничного нерва, где расположены первые нейроны. Они проводят чувствительную импульсацию от различных участков кожи тела, слизистой оболочки полости рта. Клетки задних рогов спинного мозга и ядер черепных нервов ствола мозга представляют второй нейрон. Нервные волокна, отходящие от вторых нейронов системы внешней температуры в составе латерального спиноталамического тракта, направляются к зрительному бугру, где образуют синаптические контакты с третьими нейронами. Отсюда информация о температуре внешней среды поступает в кору большого мозга (четвертый нейрон) (рис. 13.16). Афферентная импульсация от внутренних органов поступает в спинной мозг в основном по вегетативным нервам. Корковый отдел локализуется в области задней центральной извилины коры большого мозга и соответствует тактильной чувствительности. Локализация 2-го и 3-го нейронов также совпадает с соматосенсорной чувствительностью. Степень выраженности температурных ощущений определяется тремя параметрами: Рис. 13.16. Нейронная организация системы температуры внешней среды: 1 — терморецептор (первичный, кожа); 2 — спинальный ганглий (для головы — ганглии черепных нервов); 3 — задние рога спинного мозга (для головы — ствол мозга); 4 — таламус; РП — рецепторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (постцентральная извилина) Глава 13. Сенсорные системы 375 • величиной поверхности кожи, на которую действует температурный фактор — следствие пространственной суммации на разных уровнях системы температуры; • исходной температурой кожи, ведущей к адаптации или сенситизации сенсорной системы — чем ниже температура кожи, тем выше возбудимость холодовых и ниже — тепловых рецепторов, при высокой температуре кожи ощущения противоположные; • скоростью изменения температуры внешней среды — при большой скорости степень выраженности температурных ощущений больше, т.к. не успевает развиться адаптация рецепторов. Тактильная система обеспечивает формирование ощущений прикосновения, давления, вибрации и щекотки (последнее возникает обычно при раздражении определенных участков кожи, например подмышечной области). Периферический отдел тактильной системы представлен несколькими видами механорецепторов, строение и локализация которых различны. Все они — первичные рецепторы. Часть этих рецепторов — свободные нервные окончания (неинкапсулированные); они иннервируют фолликулы волос, имеются они и в эпидермисе и в сосочковом слое дермы, не обладают высокой возбудимостью. Большинство инкапсулированных рецепторов (тельца Паччини, Мейснера и др.) содержатся в лишенной волос коже (ладони, пальцы и др.) — обеспечивают более тонкую тактильную чувствительность. Проводниковый отдел. Импульсы от тактильных рецепторов проводятся по миелинизированным нервным волокнам быстрее (30–70 м/с), чем по немиелинизированным волокнам (около 1 м/с). Первый нейрон локализуется в спинальных ганглиях или ганглиях черепных нервов. Информация, соответственно, поступает в спинной мозг или ствол мозга. В заднем роге спинного мозга аксоны нейронов спинальных ганглиев в составе задних канатиков спинного мозга (без переключения) восходят к продолговатому мозгу, где в ядрах заднего столба (второй нейрон) происходит первое переключение. От кожи головы и слизистой рта импульсы идут по тройнично-таламическому тракту: первый 376 Физиология нейрон — в тройничном ганглии, второй нейрон — в главном ядре тригеминального комплекса (средний мозг). Далее через медиальную петлю проводящий путь тактильной системы следует к контрлатеральным вентробазальным ядрам зрительного бугра (третий нейрон). Нейроны таламических ядер посылают свои аксоны в корковый отдел системы, где локализуется четвертый нейрон этого пути (рис. 13.17). Рис. 13.17. Нейронная организация тактильной системы: 1 — механорецептор кожи, первичный; 2 — спинальный ганглий (для головы — ганглии тройничного нерва); 3 — продолговатый мозг (для головы — мост); 4 — таламус; РП — рецепторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (постцентральная извилина) Корковый отдел тактильной системы находится в I и II зонах постцентральной извилины (соматосенсорная область коры большого мозга). От проекционных зон коры тактильная информация поступает во фронтальные и задние ассоциативные зоны коры, благодаря которым происходит узнавание объекта или явления, повторно встретившихся, или формирование в сознании образа и объекта, явления, впервые встречаемых. Абсолютная тактильная чувствительность существенно различается в различных частях кожи, что обусловлено неравномерным распределением различных рецепторов. Система зуда. Кожный зуд (pruritus cutaneus) — это кожное ощущение легкого жжения, покалывания, вызывающее потребность чесать (скрести) кожу. Зуд возникает в поверхностном слое кожи. Классификация зуда. По локализации выделяют генерализованный (общий) и локализованный (местный) зуд. По длительности зуд делят на острый и хронический. В зависимости от причины, вызывающей зуд, его делят на физиологический (например, реакция на укус насекомого) и патологический, причиной которого может стать какой-либо Глава 13. Сенсорные системы 377 патологический процесс, например дерматит, поражение периферической нервной системы (нейрогенный зуд). В отдельных случаях возникновение зуда может быть спровоцировано воздействием факторов окружающей среды. Например, кожный зуд при подъеме на высоту, при воздействии ультрафиолетовым облучением, при подводных погружениях. Отделы системы зуда Периферический отдел системы зуда. Согласно мнению одних авторов, зуд возникает при раздражении различных рецепторов — тактильных, температурных и болевых. По мнению других, зуд возникает при раздражении только свободных нервных окончаний — сенсорных рецепторов немиелинизированных нервных волокон. Проводниковый и корковый отделы сенсорной системы зуда практически совпадают с таковыми болевой системы (см. ниже). Данный факт (с нашей точки зрения) свидетельствует о том, что часть болевой системы (которая обеспечивается немиелинизированными нервными волокнами) и система зуда являются общими. При слабых раздражениях возникает зуд, при сильных — боль. Значение зуда. Зуд включает механизмы избавления от слабых раздражений, боль — от сильных. При сильном зуде и сильном чесании кожи зуд может прекратиться в результате включения обезболивающей системы — блокады афферентной импульсации. Однако воздействие на кожу спины душем с горячей водой или веником в парилке сопровождается наслаждением высокой степени. Именно поэтому так много любителей париться в бане с веником. В формировании ощущений зуда принимают различные биологически активные вещества — пруритогены. Концентрация пруритогенов может нарастать на определенном участке кожи (например, в месте укуса насекомого). Пруритогены, взаимодействуя с соответствующими рецепторами, запускают формирование ощущения зуда. Наиболее типичным пруритогеном считается биогенный амин гистамин, который при относительно малых концентрациях вызывает ощущение зуда, а при больших концентрациях участвует в формировании ощущения боли. Пруритогенными свойствами обладают серотонин, трипсин 378 Физиология и хемотрипсин, субстанция P, калликреин, брадикинин, простогландины, лейкотриены, белковые факторы межклеточного взаимодействия (цитокины). 13.8. Сенсорные системы внутренней среды организма Эти системы воспринимают изменения внутренней среды организма и посылают информацию в ЦНС, участвуя таким образом в регуляции работы внутренних органов и поддержании гомеостазиса организма в целом. Целесообразно выделить следующие системы: • химизма внутренней среды организма; • осмотического давления внутренней среды; • давления в кровеносных сосудах; • температуры внутренней среды (см. раздел 13.8); • давления во внутренних полых органах и их наполнения. Периферический отдел представлен соответствующими рецепторами (механо-, хемо-, термо- и осморецепторы), расположенными в различных органах, сосудах, слизистых оболочках и ЦНС. Все рецепторы внутренних СС первичные. Хеморецепторы расположены в аортальном и каротидном клубочках, в слизистых оболочках пищеварительного тракта, органов дыхания, в головном мозге, в серозных оболочках. Механорецепторы расположены в сосудах, сердце, легких, ЖКТ и др. Осморецепторы локализованы в аортальном и каротидном синусах в гипоталамусе, в интерстициальной ткани вблизи капилляров, в других сосудах артериального русла, в печени и других органах. Часть осморецепторов — механорецепторы, часть — хеморецепторы (например, Na+-рецепторы, глюкорецепторы). Проводниковый отдел представлен в основном афферентными волокнами вегетативных нервов, частично — смешанными нервами, в состав которых входят и соматические, и вегетативные волокна. Первые нейроны находятся в спинальных или черепных ганглиях, вторые нейроны — в спинном мозге или стволе мозга. Восходящие пути от них поступают к третьим нейронам (заднемедиального ядра таламуса). Глава 13. Сенсорные системы 379 Горькое Кислое Листовидные сосочки Соленое Сладкое Грибовидные сосочки Желобовидные сосочки Рис. 13.18. Распределение вкусовых почек и рецептивных полей разных вкусовых ощущений на поверхности языка Корковый отдел локализуется в зонах S1 и S2 соматосенсорной области коры — задняя центральная извилина (четвертый нейрон) и в орбитальной области коры большого мозга. Поступление потока висцеральной импульсации к нейронам коры обеспечивает формирование многочисленных натуральных вегетативных условных рефлексов, что является важным приспособительным ответом в различных условиях жизнедеятельности организма. Например, прием пищи в одно и то же время обеспечивает заблаговременное начало выработки пищеварительных соков до очередного приема пищи. Импульсация от внутренних органов, например от сердца, печени, обычно не вызывает ощущений — это сигнал о благополучии в организме. В случае же патологических процессов возникают различные неприятные ощущения, в т.ч. и болевые, что побуждает обратиться к врачу. Наполнение мочевого пузыря, желудка сопровождается возникновением четко локализованных ощущений, при недостатке воды в организме формируется чувство жажды и др. 13.9. Система вкуса и обоняния Система вкуса. Главное ее назначение заключается в определении пригодности и ценности пищи, а также в формировании аппетита. Чувство вкуса многокомпонентно и связано с раздражением хемо-, механо- и терморецеп- 380 Физиология торов, а также рецепторов обоняния. Известно 13 типов хеморецепторов вкусовых клеток, чувствительных к различным веществам (натрий, калий, хлор, ионы водорода, аденозин, инозин и др.). Активация этих рецепторов вызывает четыре типа первичных вкусовых ощущений: сладкого, горького, кислого и соленого. Четкой специфичности рецепторов сладкого и горького вкусов к определенным веществам не выявлено, напротив, вещества, различные по своей химической структуре, обладают одинаковым вкусом. Так, ощущение сладкого вызывают сахара, многие спирты и другие органические растворители. Ощущение горького вызывают все алкалоиды, а также глюкозиды, пикриновая кислота, эфир, хинин, морфин, стрихнин, пилокарпин. Чувствительность вкусового анализатора самая высокая к горьким веществам, например для хинина порог ощущения составляет 0,000008 М, что почти на 5 порядков выше чувствительности к соленому (0,01 М NaCl). Это дает возможность организму предотвратить попадание несъедобных или ядовитых веществ. Периферический отдел системы вкуса. Вкусовые рецепторы вторичны, они локализуются во вкусовых почках языка, включающих также опорные клетки и нервные окончания. Вкусовая почка (их насчитывают от 3000 до 10 000) напоминает луковицу диаметром 0,03 мм, высотой 0,06 мм. Узкая часть представлена микроворсинками рецепторных клеток, на которых расположены рецепторы. Микроворсинки вкусовых рецепторных клеток контактируют с содержимым ротоглотки через небольшое отверстие в слизистой, называемое вкусовой порой (распределение вкусовых почек см. рис. 13.18). Взаимодействие рецепторов ворсинок с химическими веществами ведет к открыванию Naканалов, деполяризации мембраны и формированию РП. Проводниковый отдел начинается внутри вкусовой почки дендритами биполярных нейронов ганглиев черепных нервов (первый нейрон), где они образуют синапсы с рецепторными клетками (рис. 13.19). Вкусовые почки передних двух третей языка получают нервные волокна от барабанной струны, входящей в состав лицевого нерва и язычного нерва; рецепторы вкусовых почек, расположенных в области глотки, надгортанника и гортани, синап- Глава 13. Сенсорные системы 381 Рис. 13.19. Нейронная организация системы вкуса: 1 — вкусовой рецептор слизистой рта (вторичный); 2 — чувствительные ганглии черепных нервов; 3 — продолговатый мозг (ядро одиночного тракта); 4 — таламус; РП — рецепторный потенциал; ГП — генераторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (задняя центральная извилина в области представительства лица и в островке) тически связаны с волокнами от верхнегортанного нерва (порция блуждающего нерва); почки задней трети языка, мягкого и твердого нёба, миндалин — от языкоглоточного нерва. Аксоны первых нейронов обычно входят в состав одиночного пучка продолговатого мозга, в ядрах которого локализуются вторые нейроны вкусового пути. Отсюда нервные волокна в составе медиальной петли подходят к таламусу (третий нейрон). Корковый отдел системы вкуса локализуется в нижней части соматосенсорной зоны коры в области представительства языка (четвертый нейрон). Большая часть нейронов этой области полисенсорна, т.е. реагирует не только на вкусовые, но и на температурные, механические и ноцицептивные раздражители. Вкусовая чувствительность у сытого человека снижается, а у голодного — повышается. Вкусовая чувствительность снижается при переживаниях, например, у студентов перед экзаменом, при заболеваниях органов полости рта (стоматит, глоссит), заболеваниях желудочно-кишечного тракта, органов дыхания, болезнях крови, ЦНС. Особенностью адаптации вкусовой системы является то, что она различна для разных веществ, например, к сладкому и соленому она быстрее, чем к горькому и кислому, — предупреждение попадания вредных веществ в организм. Снижение чувствительности к одному вкусовому веществу не исключает ее сохранения к другому. Система обоняния. Ее значение заключается в том, что она помогает избежать опасности благодаря способности организма различать вредные вещества, способствует ориентации организма в окружающей среде и процессу позна- 382 Физиология ния внешнего мира, оказывает влияние на деятельность желудочно-кишечного тракта. Известно более 100 первичных обонятельных ощущений, например цветочный, мятный, эфирный и др. Периферический отдел системы обоняния — дендриты биполярных обонятельных нейронов, расположенных в слизистой оболочке носовых ходов, которая в этой части называется обонятельной мембраной (рис. 13.20). Короткий булавовидный дендрит (он выглядит как часть тела нейрона) содержит 6–12 ресничек, имеющих мембранные обонятельные рецепторы. Реснички значительно увеличивают площадь контакта рецептора с молекулами пахучих веществ. Каждый обонятельный рецептор возбуждается при действии многих пахучих веществ, но к некоторым он наиболее чувствителен. Активация рецепторов молекулами пахучих веществ приводит к синтезу цАМФ — второго посредника, с помощью которого происходит открытие натриевых каналов и формирование рецепторного потенциала в дендрите обонятельного нейрона. РП с помощью электрического поля обеспечивает деполяризацию аксонного холмика и тела Рис. 13.20. Биполярный обонятельный нейрон Глава 13. Сенсорные системы 383 обонятельного нейрона, которая, достигнув критической величины, ведет к возникновению ПД. Проводниковый отдел. Аксоны обонятельных нейронов пронизывают пластинку решетчатой кости и формируют обонятельный нерв, волокна которого образуют синапсы с крупными митральными клетками (вторые нейроны) обонятельной луковицы, локализующейся на решетчатой кости (рис. 13.21). Рис. 13.21. Нейронная организация системы обоняния: 1 — дендрит с ресничками чувствительного нейрона (первичный рецептор, слизистая носа); 2 — чувствительный нейрон, слизистая носа; 3 — митральные клетки в обонятельной луковице, локализующейся на решетчатой кости; РП — рецепторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга (обонятельная извилина, прегрушевидная область) Аксоны митральных клеток обонятельных луковиц образуют обонятельный тракт, информация по которому доставляется к корковому отделу системы обоняния. Корковый отдел системы обоняния включает прегрушевидную область (первичная обонятельная кора), обонятельную извилину, гиппокамп (аммонов рог), заднебоковой квадрант орбитофронтальной коры. Первичная обонятельная кора играет ведущую роль в научении организма отличать приятную пищу от неприятной. ГЛАВА 14 Боль — это страж, сигнализирующий организму о патологии и разрушающих воздействиях. Система боли 14.1. Основные понятия. Классификация боли Боль — это неприятное ощущение, вызывающее страдания организма. Боль возникает в результате действия на организм сверхсильного раздражителя, повреждения тканей, патологического процесса или кислородного голодания тканей. При кислородном голодании клеток активируется анаэробное дыхание, разрушаются клетки, в межклеточном пространстве накапливаются органические кислоты, продукты распада клеток, протеолитические ферменты. Многие из них оказывают прямое раздражающее действие на нервные элементы, вызывая боль. У болевой системы механизмы регуляции ее чувствительности наиболее развиты, причем особенно выражено нисходящее угнетающее влияние на афферентную болевую импульсацию. Поэтому наряду с болевой системой следует выделить обезболивающую (антиноцицептивную) систему организма, вместе они составляют единую систему — систему боли. В целостной болевой реакции организма выделяют следующие относительно самостоятельные компоненты: • сенсорный компонент, отражающий возникновение ощущения боли; • эмоциональный компонент в виде страдания, тягостных переживаний, формирующихся на основе дивергенции возбуждения по ретикулярной формации к структурам лимбической, теменной и лобной коры большого мозга; Глава 14. Система боли 385 • вегетативный компонент, проявляющийся в изменениях деятельности почти всех вегетативных эффекторов; • двигательный компонент, выступающий в виде рефлекса избавления от болевого раздражения, локального или генерализованного напряжения определенных групп мышц, в других формах двигательной активности организма. Особенно разнообразны вегетативные сдвиги при возникновении боли: ускорение свертывания крови, повышение ее фагоцитарной активности, лейкоцитоз, усиление выработки антител, системное усиление кровотока, легочной вентиляции, местное расширение сосудов в болевом очаге, улучшающее трофику тканей и доставку кислорода клеткам в очаге поражения. Активируется антиноцицептивная система. Классификация боли. По месту возникновения боль разделяют на соматическую и висцеральную. Боль, возникающая в коже, получила название соматической поверхностной боли. Ее пример — это боль, вызываемая введением инъекционной иглы в кожу человека. Боль, возникающая при поражениях суставных связок, мышц, суставов, надкостницы, костей, получила название соматической глубокой боли. Типичный ее пример — зубная боль. Глубокая боль почти всегда плохо локализуется. Висцеральная боль по своим качественным характеристикам во многом совпадает с глубокой соматической болью. Основные причины ее возникновения: • ишемия тканей, приводящая к накоплению метаболитов и нарушению тканевого дыхания; • выход содержимого полых органов при повреждении целостности их стенок (например, выход содержимого аппендикса в брюшную полость при его воспалении и разрыве); • cильные тонические сокращения или растяжения мышц желудка, желчного пузыря, кишечника, мочевого пузыря, мочеточников. В чувственной картине вызванной боли выделяют два этапа. Первый — «быстрый», легко локализуемый этап, отражающий ощущение четкой, яркой боли с латентным 386 Физиология периодом порядка 0,1 с. Второй — «медленный», плохо локализуемый этап, отражающий ощущение тупой боли с латентным периодом порядка 1–3 с. По продолжительности различают боль острую, подострую и хроническую. Острая — внезапно возникающая боль. Ее можно вызвать у больного в ходе пальпации очага воспаления, она имеет сигнальное значение. Хроническая — длительная, долго сохраняющаяся или регулярно повторяющаяся боль. Примером такой боли могут служить определенного вида головные боли, боли в спине при опухолях на поздних стадиях их развития. Подострая боль занимает промежуточное положение между острой и хронической видами болей. Отметим мгновенную, приступообразную, непрерывную, пульсирующую боли, отражающие субъективное восприятие интенсивности болей в пределах некоторого временного интервала. По отношению боли к причинам возникновения различают боль физическую и психогенную. Физическая боль возникает в результате влияния на тканевые структуры организма конкретных внешних или внутренних факторов различной природы. Психогенная боль — это следствие нарушения психических функций человека. Она возникает как страдание по поводу некоего «больного» органа, функциональные характеристики которого при объективной оценке оказываются в норме. 14.2. Болевая система Болевая система (сенсорная часть системы боли) построена по обычному принципу сенсорных систем. Периферический отдел. Болевые рецепторы (от лат. посеrе — разрушать) реагируют на стимулы, угрожающие организму повреждением или вызывающие повреждения. Основные ноцицепторы — механоцепторы тонких миелинизированных волокон (Аδ-механорецепторы) и полисенсорные ноцицепторы немиелинизированных С-волокон (С-ноцицепторы реагируют на механические, химические и температурные раздражители). Аδ-ноцицепторы обычно не реагируют на термические, химические и болевые раздражители, но возбуждаются при сильных механических Глава 14. Система боли 387 воздействиях (щипок пинцетом, укол иглой). Болевые рецепторы всех органов и тканей представляют неинкапсулированные окончания нервных волокон, имеют форму волосков, пластинок, сплетений, спиралей. Их около 30% от всех сенсорных рецепторов организма. На поверхности кожи число ноцицепторов (болевых точек) намного превышает число рецепторов прикосновения, тепла, холода. Наибольшая плотность кожных ноцицепторов у человека отмечается в паховой области (100–200 на 1 см2), на ладони, кончике носа. Основная часть ноцицепторов кожных, мышечных и внутренних органов полисенсорные (Смолин Л.Н.). Моносенсорные ноцицепторы (механо- и термоцепторы) встречаются редко. Медиатор болевых первичных нейронов — глутамат (модуляторы — субстанция Р, ВИП). Выделяют три типа веществ, вызывающих боль (алгогены): • тканевые (серотонин, гистамин, ацетилхолин, простагландины, ионы К и Н); • плазменные (брадикинин, каллидин); • выделяющиеся из нервных окончаний (субстанция P). Эти вещества действуют на болевые рецепторы и участвуют в формировании РП и болевой импульсации в ЦНС. При воспалительных процессах ощущение боли возникает под действием тканевых алгогенов — веществ, появляющихся в результате разрушения тканевых структур (гистамин, серотонин, ионы К+, простагландины и лейкотриены). Болевые рецепторы первичные. Алгогены всех типов отчетливо угнетают клеточное дыхание, что также способствует формированию болевых ощущений. Проводниковый отдел. Болевая импульсация от тела организма поступает по дендритам биполярных нейронов, спинальных ганглиев, аксоны которых в спинном мозге переключаются (в основном) в задних рогах (вторые нейроны). Аксоны этих нейронов в составе латерального и переднего спиноталамических трактов вступают в таламус. В таламусе происходит переключение на нейроны сенсорного комплекса ядер вместе с волокнами дорсальных столбов (третьи нейроны). Они формируют медиальную петлю с другими волокнами. На своем пути в стволе моз- 388 Физиология га Аδ-волокна отдают коллатерали к ядрам стволовой РФ, многократно переключаясь (рис. 14.1). Рис. 14.1. Нейронная организация болевой системы: 1 — болевой рецептор (первичный); 2 — спинальный ганглий (для головы — ганглии черепных нервов); 3 — задние рога спинного мозга (для головы — ствол мозга); 4 — таламус; ГП — генераторный потенциал; ПД — потенциал действия; КБМ — кора большого мозга Болевая импульсация от кожи лица, органов челюстнолицевой области и слизистой полости рта поступает в ЦНС по чувствительным волокнам тройничного нерва, ганглии которого располагаются на височной кости. Аксоны нейронов ганглия (первые нейроны) направляются в ствол мозга, заканчиваются в основном на нейронах ядра тригеминального тракта (вторые нейроны). Часть волокон подходит к ядрам РФ продолговатого мозга, а также к ядрам шва (рис. 14.1), далее — к ассоциативным и переключательным ядрам таламуса (третьи нейроны). Нейроны коркового отдела локализуются в соматосенсорной проекционной коре (постцентральная извилина). Точная идентификация участка кожной поверхности, на которую действует болевой раздражитель, требует параллельного вовлечения в процесс кожных тактильных рецепторов. Таким образом, ход чувствительных нервных волокон и локализация нейронов тактильной, температурной и болевой систем практически одинаковы. 14.3. Обезболивающая система В ЦНС открыты опиатные рецепторы, взаимодействие с которыми препаратов опия (морфий и его производные) вызывает состояние аналгезии. Имеется также опиоидная система аналгезии (медиаторы — энкефалин, динорфин, b-эндорфин), влияние которой реализуется с помощью включения тормозных нейронов, и неопиоидная, медиаторами которой являются серотонин, норадреналин, Глава 14. Система боли 389 адреналин, дофамин, тормозящие ноцицептивные нейроны. Обезболивающая система организма включает три основных уровня. Первый уровень обезболивающей системы (ствол мозга — спинной мозг) включает голубое пятно, центральное серое околоводопроводное вещество, ядра шва и РФ, образующие единый функциональный блок. Изолированная электрическая стимуляция этих ядер блокирует прохождение болевой импульсации на уровне релейных нейронов ядер задних рогов спинного мозга, а также релейных нейронов сенсорных ядер тройничного нерва. Это реализуется нисходящими волокнами, большинство из которых в своих окончаниях высвобождают серотонин, вызывающий ТПСП на релейных нейронах и ВПСП на энкефалинергических вставочных нейронах. Энкефалинергические нейроны включают тормозные интернейроны, реализующие эффект с помощью пост- и пресинаптического торможения, что ведет к дополнительному торможению релейных нейронов. Около 15% нейронов указанных ядер в окончаниях своих аксонов высвобождают энкефалины, вещество Р, ацетилхолин и другие вещества — модуляторы синаптической передачи (рис. 14.2) — такой же механизм обезболивания и в сенсорных ядрах тройничного нерва. Второй уровень (гипоталамус — таламус — ствол мозга) представлен в основном дорсомедиальным и вентромедиальным ядрами гипоталамуса и передней долей гипофиза. Он функционирует самостоятельно и является надстройкой, контролирующей обезболивающие механизмы первого уровня; включает энкефалин-, β-эндорфин-, норадреналин-, дофаминергические нейроны. Раздражение дорсомедиального ядра гипоталамуса (зона положительного подкрепления) вызывает гипоалгезию — частично, за счет энкефалинергических влияний, повышающих активность антиноцицептивных механизмов первого уровня, частично за счет β-эндорфинергических влияний, угнетающих передачу болевой импульсации через неспецифические ядра таламуса с включением тормозных интернейронов (рис. 14.3). Раздражение вентромедиального ядра гипоталамуса (зона отрицательного подкрепления) также сопровождается гипоалгезией, в ряде случаев — аналгезией, как при 390 Физиология - На уровне ствола мозга — нейроны сенсорных ядер тройничного нерва Рис. 14.2. Взаимодействие основных элементов обезболивающей системы организма первого уровня: ствол мозга — спинной мозг; ЭН — энкефалинергический нейрон; «+» — возбуждение; «–» — торможение. Такой же механизм обезболивания и в сенсорных ядрах тройничного нерва стрессовых состояниях, что реализуется в основном с помощью норадренергических и дофаминергических нейронов. При стрессе в кровь из передней доли гипофиза выбрасывается β-эндорфин, который угнетает активность ноцицепторов и нейронов большинства каналов передачи болевой импульсации в ЦНС. Третий уровень обезболивающей системы представлен нейронами лобной коры, различных областей моторной, орбитальной коры (поля S1 и S2). Нисходящие волокна идут к задним рогам спинного мозга, сенсорным ядрам тройничного нерва. Эти нейроны могут оказывать возбуждающие влияния, но преобладают пост- и пресинаптические тормозные влияния, изменения активности стволового комплекса и нейронов таламуса антиноцицептивной системы. Кора большого мозга контролирует обезболивающую функцию гипоталамуса и связанных с ним образований лимбической системы, РФ. Физиологические основы обезболивания, применяемые в клинической практике (местная и общая анестезия). Местная анестезия реализуется за счет блокады проведения болевой импульсации периферических нервных волокон и на уровне ноцицепторов. Местная анестезия обычно длится 20–60 мин; местные анестетики, например новокаин, блокируют Na-каналы рецепторов и нервных волокон, Глава 14. Система боли 391 Рис. 14.3. Механизм работы обезболивающей системы организма второго уровня с помощью опиоидов (гипоталамус — таламус — ствол мозга). Нейроны: светлые — возбуждающие; темный — тормозной что предотвращает возникновение ПД и, естественно, проведение возбуждения (болевой импульсации). Проводниковая анестезия — блокада нервных стволов, в составе которых проходят волокна, проводящие сигналы от ноцицепторов определенного участка. Общая анестезия достигается применением (ингаляционно, внутривенно) наркотических препаратов, которые прерывают проведение болевой импульсации преимущественно на подкорковом уровне; при этом используются также опиоиды, активирующие опиатные рецепторы и, соответственно, эндогенную обезболивающую систему. Нетрадиционные методы обезболивания — например, с помощью механического или электрического воздействия на биологически активные точки тела (скопление сенсорных рецепторов), а также с помощью специальных игл (иглоукалывание — акупунктура). Продолжительность аналгезии — 20–30 мин. Акупунктурная аналгезия осуществляется с помощью рефлекторного выброса в кровь из гипофиза эндорфинов, которые возвращаются в мозг, связываются с опиатными рецепторами, нарушая синаптическую передачу болевой импульсации в ЦНС (Померанц Б.). Более подробно способы анастезии изучаются в курсе фармакологии. ГЛАВА 15 Русская литература родилась из шинели Н.В. Гоголя, учение о высшей нервной деятельности — из сюртука И.П. Павлова. В.В. Шульговский Высшая нервная деятельность Высшая нервная деятельность (ВНД) — это совокупность нейрофизиологических процессов, обеспечивающих память, сознание и психическую деятельность. В начале главы рассмотрим память, которая обеспечивает все виды обучения. 15.1. Память Память — это способность организма приобретать и сохранять информацию и навыки. По механизму формирования память целесообразно подразделять на электрофизиологическую, нейрохимическую и нейроструктурную. Электрофизиологическая память — это кратковременная память, длительность хранения информации с помощью кратковременной памяти составляет от нескольких секунд до 5 минут; объем этой памяти — 7 ± 2 единицы. При очень сильных возбуждениях циркуляция возбуждения может достигать 10 часов. В кратковременной памяти удерживается, например, номер телефона, по которому нам нужно позвонить, адрес знакомых. Но если этой информацией не пользоваться, она быстро забывается. В основе механизма кратковременной памяти лежит кратковременная циркуляция возбуждения (рис. 15.1). Начальная стадия усвоения любой информации — восприятие раздражителя (свет, звук, слова). Информация как раздражитель воспринимается рецепторами органов чувств и кодируется в виде рецепторного потенциала, ко- Глава 15. Высшая нервная деятельность 393 Рис. 15.1. Циркуляция возбуждения по замкнутым нейронным цепям торый с помощью своего электрического поля обеспечивает возникновение нервных импульсов, передающихся в ЦНС. Эта память стирается при различных воздействиях на ЦНС (например, механическое сотрясение при ударе по голове, после чего развивается ретроградная амнезия), что, в частности, подтверждает электрофизиологический механизм этой памяти. Эту память называют первичной электрофизиологической памятью. Вторичная электрофизиологическая память — это результат циркуляции возбуждения благодаря долговременной синаптической потенциации, с помощью NMDAрецепторов (тип глутаматных рецепторов, селективно связывающихся с N-метил-Д-аспартатом). Схема такого синапса представлена на рис. 15.2. NMDA-рецепторы обладают уникальным свойством: в течение длительного времени (10–15 ч) находятся в рабочем состоянии после их активации. Исходное состояние синапса, способного к долговременной потенциации, характеризуется наличием только инактивированных NMDA-рецепторов, когда канал NMDA-рецептора оказывается перекрыт ионом Mg2+ (магниевая пробка). Сильный стимул или эмоциональное возбуждение вызывают высвобождение большого количества глутаминовой кислоты и «выбивание Рис. 15.2. Круг Папеца: 1 — гиппокамп; 2 — свод; 3 — амиллярное тело; 4 — передние ядра таламуса; 5 — поясная извилина; 6 — зубчатая извилина 394 Физиология магниевых пробок» NMDA-рецепторов. Это переводит в рабочее состояние каналы NMDA-рецепторов и к возможности длительной циркуляции возбуждения, в течение которой организм способен вспоминать информацию, полученную 10–15 ч назад. Нейрохимическая память — это процесс перевода кратковременной памяти в долговременную (консолидация памяти). Продолжительность нейрохимической памяти составляет от нескольких минут до нескольких часов и становится устойчивой примерно через 4 часа. По длительности хранения информации ее называют промежуточной памятью (между кратковременной и долговременной-нейроструктурной). В этот период экстремальные воздействия (наркоз, сотрясение мозга и др.) еще способны стереть память. Объем промежуточной памяти не изучен, его можно определить лишь ориентировочно. Например, оценить, в какой степени человек способен пересказать содержание книги, прочитанной 2–3 ч назад. На данном этапе развиваются биохимические реакции, активируется синтез медиаторов, рецепторов, ионных каналов, но запускаются они с помощью электрофизиологических процессов и вторых посредников (цАМФ, ИФ, ДАГ, NO и др.). В частности, активируются Са-зависимые ферменты в результате накопления ионов Са2+ в постсинаптическом нейроне под действием глутамата, увеличивается синтез модуляторных пептидов (энкефалины, эндорфины, ангиотензин). Поэтому данный период по механизму усвоения поступившей информации следует назвать нейрохимической памятью. При этом важную роль играют нейропептиды. Например, ВИП усиливает сродство к ацетилхолину более чем в 10 000 раз. Известно, что пептиды могут находиться в пресинаптических терминалях в качестве комедиатора, они тесно взаимодействуют в синапсах с медиаторами и что после обучения кругооборот катехоламинов в мозге увеличивается. Заметно улучшают память эндорфины и энкефалины — эндогенные опиатные пептиды. Нейроструктурная память формируется с помощью структурных изменений в нейронах (синтеза белка). Ее объем практически безграничен, ее отличает большая длительность (часы, дни и на протяжении всей жизни при Глава 15. Высшая нервная деятельность 395 повторении информации — долговременная память). По механизму она качественно отличается от кратковременной и промежуточной памяти, т.к. не нарушается при экстремальных воздействиях на мозг (механическая травма, электрошок, наркоз); некоторое ухудшение памяти после наркоза постепенно проходит. Нейроструктурная память формируется с помощью механизмов кратковременной и промежуточной памяти. При этом одновременно протекают ультраструктурные и макромолекулярные изменения в синапсах. Ультраструктурная память — хранение информации, сопровождаемое структурными изменениями в нейронах, видимыми с помощью электронного микроскопа.Изменения в синапсах. Под влиянием процесса обучения в ЦНС образуются новые синапсы, увеличивается число постсинаптических рецепторов, увеличиваются их размеры, количество медиаторов. В результате этого в структурах мозга формируются зоны повышенной синаптической проводимости, что и определяет формирование соответствующих энграмм памяти. В отростках нейрона разрастается дендритное дерево, увеличивается число шипиков на дендритах, ветвление аксона нейронов, возрастает число синапсов между нейронами. Все эти структурные изменения в отростках и синапсах нейронов наблюдаются в условиях повышенной активности обучения и исчезают в течение месяца. В частности, наблюдается рост новых синапсов между сенсорными и моторными нейронами. Молекулярная память (хранение информации с помощью синтеза белка в нейронах, невидимого даже электронным микроскопом). Открытие структурных изменений нейронов в ходе формирования памяти дало основание предполагать ключевую роль синтеза белка в консолидации памяти. Участие нейроспецифических белков в формировании следов памяти доказывается усилением их новообразования в процессе обучения и регистрацией расстройства памяти при блокаде их синтеза. Подавление же синтеза белка не влияет (по крайней мере, в опытах на животных) на кратковременную память. Это еще один важный аргумент, подтверждающий, что механизмы кратковременной и долговременной памяти различны. Усиление 396 Физиология синтеза белков при формировании нейроструктурной памяти доказывается увеличением внедрения в них метки после предварительного введения меченых аминокислот. Феномен синтеза нейроспецифических белков начинает проявляться в пределах 1 ч обучения и достигает максимума на 3-й и 6-й ч (Ашмарин И.П.). Нейроанатомия памяти. В процессе запоминания информации мозг работает как единое целое. Но в первую очередь процессы памяти связывают с фронтальной, височной и париетальной корой, мозжечком, базальными ядрами, амигдалой, гиппокампом, неспецифической системой мозга. Процесс формирования следа памяти характеризуется перемещением локусов активности по структурам мозга. Вновь формируемая энграмма в отличие от старого следа памяти представлена в мозге более широкой зоной активации. Некоторые ее области выполняют специфическую роль. В частности, гиппокамп (древний отдел мозга) вместе с медиальной височной корой обеспечивают консолидацию памяти. Гиппокамп анатомически тесно связан с височными долями, прижат к амигдале височной доли. У людей с повреждением медиальной височной коры и гиппокампа отсутствует способность перевода информации из кратковременной в долговременную память, т.е. нарушена консолидация памяти. В памяти существует только та часть жизни, которая прошла до операции или травмы. Текущие события сохраняются в памяти не более 15–20 мин. Амигдала обеспечивает быстрое и прочное запечатление в памяти эмоциональных событий часто после одноразового обучения. Важную роль в процессах запоминания двигательных навыков играет мозжечок, об этом свидетельствует, в частности, следующий опыт. У кролика выработали условный рефлекс моргания одним глазом в ответ на звук. Безусловным подкреплением служила направленная в этот глаз струя воздуха. После закрепления рефлекса кролик начал моргать при одном только звуке. Затем экспериментаторы удалили небольшую часть мозжечка со стороны глаза, на который был выработан рефлекс. Условный рефлекс ис- Глава 15. Высшая нервная деятельность 397 чез полностью, хотя безусловный (моргание глаза на струю воздуха) сохранился. Нужно добавить, что у этого кролика в дальнейшем можно было вырабатывать условные рефлексы только на другой глаз (с неповрежденной стороны мозжечка). Мозжечок играет важную роль в хранении автоматизированных реакций. Долговременная память в нормальных условиях легко доступна для извлечения информации. Роль глиальных клеток в долговременной памяти заключается в синтезе специальных веществ, облегчающих синаптическую передачу. При обучении в глиальных клетках увеличивается содержание РНК. Ретикулярная формация (РФ) оказывает активирующее влияние на структуры, участвующие в фиксации и извлечении информации из памяти, а также непосредственно включается в процессы формирования энграмм. Блокада РФ нейрофармакологическими средствами сопровождается нарушением процесса консолидации памяти. Долговременная память в нормальных условиях легко доступна для извлечения информации. Роль тренировки механизмов памяти. Известно, что у лиц, занимающихся интенсивным умственным трудом, хорошая память сохраняется даже в преклонном возрасте, что связано со структурными изменениями. Косвенное подтверждение этого — опыты на животных. Так, например, у крыс, воспитанных в обогащенной среде с наличием множества раздражителей и обученных выполнению зрительных задач, кора головного мозга значительно толще, чем у крыс, выращенных в обедненной информацией окружающей среде. Вспоминание и забывание. Долговременная память в нормальных условиях легко доступна для извлечения информации (вспоминание), механизмы которого изучены недостаточно, так же как и забывания. Значение забывания заключается в том, что организм может заниматься без помех разнообразной деятельностью, а постоянное вспоминание огромного объема информации и навыков препятствовало бы сосредоточению на текущей деятельности. Значение памяти заключается в приобретении, хранении информации и пользовании ею, а также в научении. 398 Физиология При нарушении памяти человек становится беспомощным. Например, при ожоге о красивое раскаленное железо человек мог бы много раз повторить эту ошибку. Одна из форм научения — это условный рефлекс. 15.2. Уcловный рефлекс как научение Научение — это выработка в онтогенезе приспособительных форм поведения организма, в том числе навыков физического труда. Условный рефлекс — выработанная в онтогенезе реакция организма на раздражитель, ранее индифферентный для этой реакции, т.е. это тоже научение. Выработка условного рефлекса. Основные правила выработки условных рефлексов: • условный рефлекс образуется на базе безусловного (врожденного) рефлекса; • условный стимул должен предшествовать безусловному; • необходимо неоднократное совпадение во времени действия индифферентного раздражителя с безусловным. Факторы, способствующие образованию условных рефлексов: • активное функциональное состояние нервного центра безусловного рефлекса (для пищевых рефлексов — наличие пищевой потребности) и оптимальное состояние организма (здоровье), в первую очередь состояние коры большого мозга; • оптимальное соотношение силы условного и безусловного раздражителей, при очень сильном условном раздражителе развивается оборонительная реакция; • отсутствие значимых посторонних сигналов из внешней и внутренней среды организма (рис. 15.3). Механизм формирования условных рефлексов. Временная связь, согласно И.П. Павлову (1903), образуется между корковыми центрами безусловного и условного раздражителей (рис. 15.4). Импульсы, вызываемые индифферентным (условным) сигналом с любых рецепторов, по нервам поступают в кору большого мозга и обеспечивают там образование очага воз- Глава 15. Высшая нервная деятельность 399 Измеритель объема слюны Станок для фиксации животного Лампа Кормушка – Кнопки включения электродов – Рис. 15.3. Общий вид установки для изучения условных рефлексов у собаки по И.П. Павлову. Условные раздражители — лампочка и звонок, безусловный (подкрепление) — пища в кормушке буждения. Если после индифферентного сигнала подать, например, пищевое подкрепление, то возникает более мощный второй очаг возбуждения в коре большого мозга. Возбуждение от обоих центров дивергирует, между центрами формируются доминантные отношения. Это представление получило позже подтверждение в учении о доминанте А.А. Ухтомского. Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздраI 1 2 II 3 4 Рис. 15.4. Дуга условного рефлекса с двусторонней связью (по Э.А. Асратяну): 1 — корковый центр мигательного рефлекса; 2 — корковый центр пищевого рефлекса; 3, 4 — подкорковые центры мигательного и пищевого рефлексов; I — прямая временная связь; II — обратная временная связь 400 Физиология жителя облегчает прохождение импульсов (синаптическая потенциация) от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса (проторение пути). Индифферентный сигнал не может быть вообще индифферентным (Асратян Э.А.), т.к. в начале своего применения вызывает собственную реакцию в виде ориентировочно-исследовательского рефлекса, который по мере повторения ослабляется. Замыкание происходит между центрами двух безусловных рефлексов — сигнального и подкрепляющего (см. рис. 15.3). Временная связь замыкается с помощью механизмов памяти. Классификация условных рефлексов осуществляется по нескольким критериям: • по безусловным рефлексам, на базе которых вырабатываются условные рефлексы; • по биологическому значению различают рефлексы оборонительные, пищевые и др.; • по виду рецепторов, на которые воздействует условный сигнал (интероцептивные, экстероцептивные, проприоцептивные условные рефлексы (Быков К.М., Черниговский В.Н.); • по отделу нервной системы (соматические, т.е. двигательные, и вегетативные, например слюноотделение на звук); • по условиям выработки (Павлов И.П.) имеется две группы условных рефлексов: – натуральные условные рефлексы вырабатываются на обстановку, на время, на естественные признаки подкрепляющего раздражителя, например вид, запах мяса, на которые вырабатывается условный рефлекс; – искусственные (лабораторные) условные рефлексы вырабатываются на раздражители, которые в природе не имеют отношения к безусловному (подкрепляющему) раздражителю, например выработка слюны на звонок при неоднократном подкреплении его пищей. В зависимости от сигнальной системы различают условные рефлексы на сигналы первой и второй сигнальных систем (на слово). Глава 15. Высшая нервная деятельность 401 Сравнительная характеристика условных и безусловных рефлексов: • условные рефлексы — индивидуальные (приобретаются в течение жизни); безусловные — врожденные (отражают видовые особенности организма); • условные рефлексы осуществляется с помощью функционально организующихся временных (замыкательных) связей, а безусловные — по анатомическим путям, определенным генетически; • условные рефлексы изменчивы (исчезают, когда они не нужны); безусловные — относительно постоянны в течение жизни особи; • условные рефлексы для своего образования и реализации требуют целостности коры большого мозга, особенно у высших млекопитающих; безусловные рефлексы могут осуществляться с помощью любого уровня ЦНС; • условные рефлексы могут образовываться с любого рецептивного поля, на самые разнообразные раздражители; каждый безусловный рефлекс имеет свое специфическое рецептивное поле и специфические раздражители; • условные рефлексы возникают заблаговременно (готовят организм к предстоящим событиям); безусловные возникают в ответ на действие наличного раздражителя, которого уже нельзя избежать; • набор условных рефлексов безграничен; число определенных видов безусловных рефлексов детерминировано генетически. Биологическое значение условных рефлексов состоит в том, что они обеспечивают более тонкое приспособление организма — организм заранее готовится к будущей полезной поведенческой деятельности, они изменчивы и тормозятся, если изменились условия, — исключается бесполезная для организма деятельность, их число не ограничено. Торможение условных рефлексов. Различают врожденное (безусловное) и приобретенное (условное) торможение. 402 Физиология Безусловное торможение условных рефлексов не является научением — оно возникает сразу (его не надо вырабатывать, учиться), включает два варианта (рис. 15.5). 1. Внешнее торможение условных рефлексов — ослабление или прекращение наличной условно-рефлекторной реакции при действии какого-либо постороннего раздражителя, например неожиданного звука. При этом возникает ориентировочно-исследовательская реакция (рефлекс «что такое?» по И.П. Павлову). Рефлекс «что такое?» состоит в реакции настораживания и подготовке организма на случай внезапно возникающей необходимости к действию. Имеется два варианта этого вида торможения. Гаснущий тормоз — когда посторонний раздражитель, который с повторением его действия теряет свое тормозящее влияние (угасает), т.к. не имеет существенного значения для организма (научение). В обыденной жизни на человека действует масса посторонних сигналов, на которые он сначала обращает внимание, а затем перестает их замечать. Постоянный (негаснущий) тормоз, т.е. с повторением не теряет своего тормозного действия (например, болевые раздражители, раздражения от переполненных внутренних органов). Эти раздражители имеют существенное значение для организма и требуют от него принятия решительных мер к их устранению. Механизм внешнего торможения заключается в том, что посторонний раздражитель вызывает появление в коре большого мозга нового очага возбуждения, вне дуги условного рефлекса (отсюда и название — внешнее торможение), который оказывает угнетающее влияние на текущую ус- Рис. 15.5. Виды торможения условных рефлексов Глава 15. Высшая нервная деятельность 403 ловно-рефлекторную деятельность по механизму доминанты. Значение внешнего торможения заключается в том, что организм сосредоточивает свое внимание на новом раздражителе, чтобы решить, как действовать в новой ситуации. 2. Запредельное торможение возникает при действии чрезвычайно сильного или длительного условного раздражителя. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие (закон силы): чем сильнее условный сигнал, тем сильнее условно-рефлекторная реакция. Но закон силы сохраняется до определенной величины силы раздражителя, выше которой эффект начинает уменьшаться, несмотря на увеличение силы или частоты раздражения. Возможно развитие торможения, если условный сигнал включается сразу очень сильным (например, звук). Механизм запредельного торможения. Согласно И.П. Павлову, в данном случае нервный центр условного рефлекса сильно возбужден (предел работоспособности). Значение запредельного торможения играет охранительную роль. Условное торможение условных рефлексов вырабатывается (научение), как и сам условный рефлекс, поэтому оно называется условно-рефлекторным торможением. Согласно И.П. Павлову, оно локализовано в пределах («внутри») центральных структур условно-рефлекторного акта, что и определяет второе его название — внутреннее торможение. Его называют также отрицательным условным рефлексом (термин, с нашей точки зрения, неудачный — рефлекс очень важен для организма, торможение условного рефлекса — это устранение ненужной реакции для организма). Выделяют четыре вида условного торможения: • угасательное торможение в жизнедеятельности организма встречается наиболее часто — формируется при повторном включении условного сигнала без подкрепления; • запаздывательное торможение развивается при отставании подкрепления на 1−3 мин относительно начала действия условного сигнала. При многократном повторении этого опыта условная реакция сдвигается к моменту подкрепления; 404 Физиология • дифференцировочное торможение формируется при дополнительном включении другого раздражителя, близкого к подкрепляемому сигналу, но без подкрепления дополнительного раздражителя. Например, если собаке предъявлять круг и подкреплять его пищей, а эллипс не подкреплять и их чередовать в течение одного опыта, то через некоторое время животное начинает различать оба сигнала: на круг возникает условная реакция, например слюноотделение, а на эллипс слюноотделения не наблюдается; • условный тормоз формируется при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Например, условный сигнал «звонок» подкрепляют пищей, а комбинацию «звонок и свет» не подкрепляют. После этого «свет» становится условным тормозом для любого условного рефлекса. Механизм внутреннего торможения условных рефлексов изучен недостаточно. Установлено, что при этом регистрируется гиперполяризация корковых нейронов. Функциональные изменения при выработке условных рефлексов (изменения возбудимости нейронов, изменения ЭЭГ и др.) и их торможение имеют общие черты (Белов А.Ф.). Значение всех видов внутреннего торможения условных рефлексов заключается в тонком приспособлении организма к окружающей среде — устранение ненужной в данных условиях деятельности организма (экономный характер деятельности). 15.3. Другие виды научения Все виды научения делят на две группы: неассоциативное (привыкание, сенситизация, импринтинг и подражание) и ассоциативное. Неассоциативное — научение реагировать или не реагировать на непосредственное действие раздражителя без сочетания его с действием какого-либо другого сигнала (без ассоциации). Ассоциативное — научение при совпадении безусловно-рефлекторной деятельности организма с каким-либо раздражителем, который приобретает сигнальное значение. При этом формируется Глава 15. Высшая нервная деятельность 405 связь (ассоциация, от лат. associatio — соединение, связь, сочетание) между этим раздражителем (сигналом) и деятельностью организма. Кроме того, целесообразно все формы научения объединить в четыре основные группы преимущественно по степени активности организма в ходе научения. Пассивное (менее удачное название — реактивное) научение формируется само собой при действии на организм внешних факторов, организм не предпринимает усилий для этого научения. В эту группу следует включить следующие варианты. Привыкание (габитуация) — угасание ориентировочной реакции (рефлекса «что такое?» по И.П. Павлову). Оно формируется, если раздражитель многократно повторяется и не имеет особого значения, поэтому организм прекращает на него реагировать. Так, появление звука в лаборатории, где находится подопытное животное (собака), первоначально вызывает у него ориентировочную реакцию: поворот головы, ушей в сторону источника звука, изменение частоты сердечных сокращений, десинхронизацию ЭЭГ, изменение частоты и глубины дыхания. После многократного повторения звука, не сопровождающегося другими воздействиями на животное, все реакции затормаживаются. Привыкание — это самая распространенная форма научения у человека и животных. За счет него организм игнорирует раздражители, не несущие никакой новизны и не имеющие для него значения, сосредоточивает внимание на более важных явлениях. Сенситизация — усиление реакции организма на повторяющийся стимул, если он вызывает каждый раз неприятные ощущения. Например, многократно повторяющийся пронзительный писк комара становится неприятным, непереносимым. В данном случае научение выражается в соответствующих поведенческих реакциях типа «стимул–ответ». Условные рефлексы (кроме инструментальных) и их условное торможение — один из вариантов пассивного научения. Импринтинг (запечатление, от лат. imprint — оставлять след, запечатлевать, фиксировать) — запечатление в памя- 406 Физиология ти новорожденного окружающей действительности. Это особая форма реагирования, занимающая промежуточное положение между условными и безусловными рефлексами, проявляющаяся в раннем онтогенезе (рис. 15.6). Импринтинг проявляется в трех формах: • реакция следования — следование детенышей за своими родителями и даже за любым объектом, встретившимся сразу после рождения; • запечатление образов и объектов — родителей, братьев, сестер, объектов пищи, гнезда и т.д.; • усвоение поведенческих актов — дети повторяют действия родителей. Оперантное научение (от лат. operatio — действие) — это научение, в ходе которого организм добивается полезного результата с помощью активного поведения. Оно включает три вида: • инструментальный условный рефлекс — научение действию по сигналу с помощью вознаграждения (подкрепления). Животное, например крыса, по световому сигналу нажимает на рычаг, чтобы получать пищу; • метод проб и ошибок — американский ученый Э. Торндайк (1890), видный представитель на- Рис. 15.6. Привязанность к К. Лоренцу — объекту импринтинга, который гусята увидели после вылупления, заставляет их относиться к нему как к своей настоящей матери (по Ж. Годфруа, 1992) Глава 15. Высшая нервная деятельность 407 правления бихевиоризма (от англ. behaviour — поведение), помещал голодных кошек в так называемые проблемные клетки, которые открывались в том случае, если кошка предпринимала определенные действия, например приподнимала лапой запорный крючок. Когда кошка выходила из клетки, она получала пищу. При повторении процедуры выхода из клетки число проб и ошибок уменьшалось. Различие в этих двух формах научения заключается в том, что научение с помощью инструментального условного рефлекса происходит по сигналу, а научение методом проб и ошибок — без сигнала; • самораздражение структур мозга для получения удовольствия (положительных эмоций) (рис. 15.7) или избегание самораздражения, которое сопровождается возникновением неприятных ощущений и отрицательных эмоций. Когнитивное научение (научение с помощью мышления). В эту форму научения следует включить научение путем наблюдения и правильное действие организма в ситуации, с которой он впервые встретился. Научение путем наблюдения. Различают два вида подобного научения: • простое подражание, например, обезьяна в общении с исследователями в экспедиции научилась мыть банан перед едой, не понимая, зачем это делается. Особенно часто используют научение путем наблюдения А Б Рис. 15.7. Опыт с самораздражением через вживленные электроды (схема): А — раздражение не включено; Б — электроды замкнуты, раздражение включено 408 Физиология (подражания) дети, причем в раннем онтогенезе оно является просто подражательным; • викарное научение, которое также осуществляется посредством наблюдения, но при этом результат действия оценивается. Например, наблюдение за выполнением гимнастических упражнений и последующее их выполнение, если обучающийся их запомнил. Научение путем мышления в ситуации, впервые встретившейся организму. Это, например, способность животного определять направление движения предмета, другого животного, движущихся по прямой линии, после исчезновения их из поля зрения. Сложное приспособительное поведение строится из трех основных компонентов: инстинктов, условных рефлексов и мышления, благодаря которым сразу вырабатывается программа адаптивного поведения в новых условиях. Подсознательное научение (путем инсайта, от англ. insite — озарение) — внезапное нестандартное правильное решение задачи с помощью подсознательной деятельности мозга. Этот вид научения может сформироваться после отдельных проб и ошибок, когда возникает идея результативного действия, которое может совершенствоваться в процессе достижения цели. Так, например, находящаяся в клетке обезьяна сначала пыталась достать банан одной короткой палкой в виде трубки, потом другой такой же, но безрезультатно. Тогда она, прекратив свои попытки, стала разглядывать находящиеся возле нее предметы и вдруг быстро проделала логическую последовательность действий: схватила обе палки, вставила их одну в другую, просунула между прутьев клетки и достала банан (рис. 15.8). Рис. 15.8. Обезьяна путем инсайта находит способ, позволяющий ей достать находящийся далеко банан с помощью вставленных одна в другую трубок Глава 15. Высшая нервная деятельность 409 В конкретных ситуациях для достижения того или иного полезного приспособительного результата индивид чаще всего использует не один, а несколько видов научения. 15.4. Типы высшей нервной деятельности и темпераменты личности Тип высшей нервной деятельности (ВНД) — это совокупность врожденных и приобретенных свойств нервной системы (силы процессов возбуждения и торможения, их уравновешенности и подвижности), определяющих темперамент личности. Согласно И.П. Павлову, комбинации этих трех основных свойств нервной системы позволили выделить определенные типы по адаптивным способностям и устойчивости к невротизирующим факторам — это учение о реактивности нервной системы, особенно ее высших отделов — коры большого мозга. Определение типов ВНД имеет клиническое значение: одни и те же заболевания по-разному протекают у больных в зависимости от типа ВНД и требуют разного подхода при лечении. Темперамент — это совокупность поведенческих черт личности (индивидуальности), характеризующих ее двигательную, эмоциональную и речевую активность, а также скорость реагирования и переключения с одного вида деятельности на другой. Личность — это отдельный человек, характеризующийся морально-волевыми качествами и мировоззрением, знаниями, навыками и привычками, уровнем умственной и физической работоспособности, темпераментом и стилем поведения. Характеристика свойств нервных процессов. Под силой нервных процессов понимают работоспособность корковых клеток, которая определяется их способностью отвечать на раздражители и тормозить возникающие реакции. Уравновешенность нервных процессов определяется соотношением силы процессов возбуждения и торможения. Подвижность нервных процессов — это скорость перехода процесса возбуждения в процесс торможения и обратно. Внешне это проявляется скоростью переключения с одного рода деятельности на другой. 410 Физиология Типы ВНД и темпераменты личности (от лат. temperamentum — соразмерность, надлежащее соотношение). Древнегреческий врач, основатель медицины Гиппократ (460–377 до н.э.) объяснял неодинаковое течение одной и той же болезни у разных людей разным соотношением «соков тела» в организме человека: крови, слизи, желчи, черной желчи. Согласно Гиппократу, различают четыре темперамента: • сангвинический (от лат. sangius — кровь); • холерический (от лат. chole — желчь); • флегматический (от лат. phlegma — слизь, мокрота); • меланхолический (от греч. melanos + chole — черная, испорченная желчь). Учение о типах ВНД является общим для животных и человека. Согласно И.П. Павлову, сильному, уравновешенному, подвижному типу ВНД соответствует темперамент сангвиник — человек решительный, энергичный, с высокой возбудимостью, подвижный, впечатлительный, с ярким внешним выражением эмоций, легкой их сменяемостью (рис. 15.9). Сильному, уравновешенному, неподвижному типу ВНД соответствует темперамент флегматик — спокойный, медлительный, со слабым проявлением эмоций, ему трудно переключаться с одной деятельности на другую. Сильному, неуравновешенному типу ВНД соответствует темперамент холерик — вспыльчивый, с высоким уровнем активности, раздражительный, энергичный, с сильными, быстро возникающими эмоциями, ярко отражающимися в речи, мимике, жестах. Слабому типу ВНД соответствует темперамент меланхолик — унылый, тоскливый, с низТипы ВНД и соответствующие им темпераменты Сильный Уравновешенный: подвижный (сангвиник), инертный (флегматик) Слабый (меланхолик) Неуравновешенный (холерик) Рис. 15.9. Основные типы ВНД и соответствующие им темпераменты Глава 15. Высшая нервная деятельность 411 ким уровнем нервно-психической активности, с высокой эмоциональной ранимостью, мнительный. В действительности у человека «чистые» темпераменты редки; обычно наблюдаются смешанные варианты с преимуществом одного из них. 15.5. Бодрствование и сон. Сновидения. Гипноз Бодроствование — это циркадный биоритм деятельности или отдыха организма, находящегося в сознании. Сон — циркадный биоритм пониженной активности мозга, обеспечивающий отдых организма, при котором выключены сознание и механизмы поддержания естественной позы, снижена чувствительность сенсорных систем. По происхождению выделяют три вида сна: • естественный сон — периодический суточный и периодический сезонный (последний только у животных); • искусственный — наркотический и гипнотический; • патологический сон — при анемии мозга, поражении структур мозга («сонный энцефалит», летаргический сон). Периодический суточный сон может быть монофазным (1 раз в сутки), дифазным (2 раза в сутки) и полифазным (несколько раз в сутки у новорожденных). Длительность сна зависит от возраста: например, новорожденный спит около 20 ч, взрослые — 7–8 ч. В зависимости от активности коры большого мозга и состояния организма различают две стадии сна: медленный и быстрый. Медленный (ортодоксальный) сон занимает около 80% времени ночного сна, а быстрый (парадоксальный) — 20%. Вначале развивается фаза медленного сна, которая характеризуется расслаблением мышц. В ЭЭГ возникает медленная электрическая активность в виде синхронизации (частота волн уменьшается, амплитуда возрастает). Медленный сон по мере его углубления проходит последовательно четыре фазы 412 Физиология (рис. 15.10). Первая фаза — преобладают альфа-волны (8– 13 Гц); вторая фаза — тета-волны (3–7 Гц); третья фаза — появляются сонные веретена (12–15 Гц) и К-комплексы (серия медленных потенциалов большой амплитуды) в сочетании с высоковольтными дельта-волнами; четвертая фаза — наиболее глубокий сон: в ЭЭГ преобладают дельта-волны (0,5–2 Гц). Быстрый (парадоксальный) сон скачкообразно следует после 4-й фазы медленного сна через 60–90 мин. Возникают быстрые движения глазных яблок, почти исчезает мышечный тонус, отмечается фазное подергивание конечностей, учащаются пульс и дыхание, повышается кровяное давление, возникают сновидения, в 2–3 раза возрастают А 1 θ-волны 2 Сонные веретена K-комплексы 3 4 Б 5 Рис. 15.10. ЭЭГ у человека: А — бодрствование, глаза открыты, взор перемещается; Б — различные фазы сна: 1 — преобладают альфа-волны (8–12 Гц); 2 — тета-волны (3–7 Гц); 3 — появляются сонные веретена и К-комплексы (12–15 Гц); 4 — преобладание дельта-волн (0,5–2 Гц); 5 — парадоксальная фаза сна с быстрыми движениями глаз (БДГ-сон) и десинхронизированной электрической активностью мозга Глава 15. Высшая нервная деятельность 413 пороги сенсорного пробуждения относительно медленного сна. На ЭЭГ появляется электрическая активность, характерная для бодрствующего состояния, — в течение 6–8 ч сна стадии быстрого сна появляются 4–5 раз, делаясь все более продолжительными (особенно перед пробуждением). Механизм сна. Роль гуморальных факторов. В формировании сна участвует пептид аргинин-вазотоцин, экстрагированный из нейрогипофиза, который вызывает дельта-сон. У человека введение аргинин-вазотоцина усиливает показатели парадоксального сна. Вазоактивный пептид (ВИП), бета-эндорфин углубляют состояние сна. Пептид, прерывающий сон, — это тиреотропный гормон. Уровень аргинин-вазотоцина в ликворе человека с возрастом постепенно снижается, что объясняет причины постепенного уменьшения длительности у пожилых именно парадоксального сна. Роль нервной системы. Наблюдения сотрудников П.К. Анохина над сросшимися девочками-близнецами показали, что они могли спать порознь (между их кровеносными сосудами были анастомозы). Показано, что в ответ на низкочастотное раздражение ретикулярных ядер таламуса и субталамуса подопытные кошки засыпают. После прекращения раздражения животные просыпались. При локальном повреждении переднего гипоталамуса животные перестают спать — все указанное свидетельствует о важной роли нервной системы в развитии сна. Ретикулярная формация среднего мозга способствует развитию медленного сна, активация голубого пятна способствует развитию парадоксального сна. Значение сна. Сон обеспечивает отдых организма, парадоксальный сон восстанавливает пластические свойства нейронов головного мозга, медленный сон способствует восстановлению внутренних органов, поскольку через гипоталамус либерины воздействуют на гипофиз, способствуя высвобождению соматотропина (гормон роста), который участвует в биосинтезе белков в организме. Сон — эволюционное приспособление к изменению освещенности (циркадный ритм — день–ночь): в светлое время суток — добыча пищи, ночью — отдых. Важна роль сна 414 Физиология в усвоении информации, решении задач, которые не были решены перед сном. Сновидения — возникающие во сне нереальные образы и явления, воспринимаемые как действительность. Они встречаются в обеих фазах сна, чаще — в парадоксальной фазе. Все люди видят сны. В фазу медленного сна у человека могут иметь место снохождения (сомнамбулизм). Во время медленного сна сновидения менее четки, менее длительны. Факторы, побуждающие сновидения, — предшествующая сну деятельность (например, дети во сне продолжают «играть» с игрушками, действие внешних раздражителей (если к стопам спящего человека приложить горячую грелку, ему может присниться сон, что он идет по раскаленному песку), избыточная импульсация от внутренних органов, возникающая при их патологическом состоянии или в связи с биологическими потребностями (голодному человеку может присниться вкусная еда). Гипноз — это искусственно вызванное (с помощью внушения) состояние человека, при котором не полностью выключено сознание и снижены реакции на все раздражители, кроме слов гипнотизирующего, к которым человек становится чрезвычайно восприимчивым. В ответ на многократные или монотонные стимулы непременно развивается внутреннее торможение. Если такая стимуляция продолжается, то наступает сон. Переходный период между бодрствованием и сном назван гипнотическим состоянием. Выделяют три фазы гипнотического состояния: уравнительную, парадоксальную и ультрапарадоксальную. В парадоксальной фазе сильные раздражители вызывают слабую реакцию, а слабые — сильную. В ультрапарадоксальной фазе реакция может быть получена посредством неадекватного стимула, т.е. стимула, на который клетки головного мозга в состоянии бодрствования не реагируют. Например, обычная холодная монета путем внушения может вызвать ожог кожи. Внушаемость основана на легком переходе в тормозное состояние корковых нейронов. По И.П. Павлову, «основной механизм внушаемости есть разорванность нормальной более или менее объединенной работы всей коры». Она обусловлена отсутствием обычных влияний со стороны остальных частей коры. Основное фи- Глава 15. Высшая нервная деятельность 415 зиологическое условие внушаемости — снижение тонуса коры полушарий большого мозга и легко возникающая при этом функциональная расчлененность корковой деятельности. В гипнозе волны ЭЭГ сходны с ЭЭГ бодрствования, а не сна. Стадии гипноза. Стадия гипноидности, сопровождающаяся психическим и мышечным расслаблением, миганием и закрыванием глаз. Стадия легкого транса, для которой характерна каталепсия конечностей (конечности могут длительное время находиться в необычном положении). Стадия среднего транса, когда возникают неполная амнезия, изменения личности, возможны простые гипнотические внушения. Стадия глубокого транса, которая характеризуется полной амнезией, сомнамбулизмом и фантастическими внушениями. 15.6. Методы исследования ВНД Разнообразие методик. ВНД изучается с помощью выработки условных рефлексов и других видов научения и воздействия на ЦНС или исследования ее состояния. В частности, выработка различных условных рефлексов и других форм научения на фоне холодового выключения, разрушения и раздражения отдельных структур мозга, что позволяет выявить роль отдельных структур в замыкании временной связи. Научение при введении агонистов (ацетилхолина, адреналина, серотонина и др.), антагонистов (блокаторов); на фоне действия молекул ДНК, РНК. Исследования изменений в синаптических структурах нейронов, их отростков с помощью электронной микроскопии после выработки условных рефлексов и формирования других видов научения. Регистрация в процессе научения электроэнцефалограммы (ЭЭГ), вызванных потенциалов, нейронной активности различных отделов коры большого мозга, подкорковых структур, отдельных нейронов на разной стадии выработки условного рефлекса и других видов научения, любой психической деятельности. Характеристика отдельных методик. Электроэнцефалография — это регистрация с поверхности головы сум- 416 Физиология марного электрического поля, создаваемого ПД (нейронов и нервных волокон), ВПСП и ТПСП (более подробно см. раздел 4.21). Электроэнцефалограмма (ЭЭГ) — это кривая, зарегистрированная при электроэнцефалографии. Метод вызванных потенциалов (ВП) — это регистрация колебаний электрической активности (электрического поля) мозга, возникающих на ЭЭГ при однократном раздражении сенсорных рецепторов (зрительных, слуховых, тактильных), афферентных путей, центров переключения афферентной импульсации, поступающей в кору большого мозга. Микроэлектродный метод позволяет изучать активность отдельных нейронов ЦНС. С помощью микроэлектродов, вводимых внутрь нервных клеток, можно измерять мембранные потенциалы покоя, регистрировать постсинаптические потенциалы — возбуждающие и тормозные, а также потенциалы действия, вводить различные вещества и регистрировать с помощью многоканального электрода электрическую активность нейронов. Используются и некоторые другие методы. ГЛАВА 16 Нет ничего сильнее и бессильнее слова. И.С. Тургенев Психическая деятельность и поведение Психическая деятельность — это идеальная, субъективно осознаваемая деятельность организма, осуществляемая с помощью нейрофизиологических процессов, обеспечивающая организму познание и формирование целенаправленного поведения. Видами психической деятельности являются: ощущение, внимание, мышление (узнавание, восприятие, представление, вспоминание, воображение), мотивация и эмоция. Психическая деятельность осуществляется с помощью высшей нервной деятельности — совокупности нейрофизиологических процессов, обеспечивающих память и обучение, сознание и психическую деятельность. Таково соотношение этих понятий. Психическая деятельность осуществляется только тогда, когда организм находится в состоянии сознания; во сне, например, психическая деятельность (в частности, мышление) невозможнa. Но усвоение полученной до наступления сна информации продолжается, т.е. ВНД осуществляется и в период бодрствования, и во сне, когда сознание выключено. 16.1. Сознание и мышление Характеристика сознания. Сознание — это состояние организма, при котором он может дать действительности соответствующую оценку; оно является условием всех видов психической деятельности, в том числе и мышления. ВНД — основа всех видов психической деятельности. Для включения сознания необходима активация большого количества структур, где ведущее значение имеет кора 418 Физиология большого мозга с ближайшей подкоркой, лимбической системой, а также их взаимодействие. Важнейшую роль играют и восходящие активирующие влияния ретикулярной формации. Должен быть определенный уровень активности ЦНС — при слишком низкой нейронной активности (например, при наркозе или коме, во сне) сознание выключается. В то же время сознание невозможно и при чрезмерной активности нейронов, в частности при эпилептических припадках, характеризующихся пиками и волнами на ЭЭГ, оно может выключаться в состоянии ярости (состояние аффекта). Необходим минимальный период времени активации мозговых структур для осознанного восприятия сигнала (100–300 мс). В сознании представлены только конечные результаты процессов обработки информации, которые мы осознаем в виде последовательных картин, эпизодов. Электрофизиологические и биохимические процессы, лежащие в основе формирования эпизодов, не ощущаются. Об их прохождении и локализации (но не о содержании информации) можно косвенно судить с помощью различных методов исследования ЦНС. Мышление — это оперирование (воспоминание) закодированной в головном мозге информацией с помощью механизмов памяти и процесс приобретения новой информации. Классификация мышления осуществляется по двум основным критериям. По сложности мышление может осуществляться на шести уровнях. Первый уровень — узнавание объекта или явления, встретившихся повторно. Это осуществляется с помощью сличения поступающей в данный момент информации от органов чувств с ранее закодированной с помощью механизмов памяти информацией — без сличения узнавание невозможно. Второй уровень мышления — восприятие (формирование в сознании нового образа объекта или явления, также действующего на органы чувств, но встретившегося впервые). В этом случае тоже происходит процесс сличения новой информации с подобной закодированной информацией. Третий уровень мышления — представление, это формирование в сознании образа объекта или явления, не действующих в данный момент на органы чувств (например, воспроизведение в сознании Глава 16. Психическая деятельность и поведение 419 покинутой в беспорядке собственной квартиры). Четвертый — воспоминание (формирование в сознании любой информации, закодированной в головном мозге с помощью механизмов памяти — например, содержание просмотренного фильма). Термины «воспоминание» и «представление» близки по содержанию, но и различие между ними очевидно (представление — частный случай воспоминания). Пятый уровень мышления — принятие решения. Шестой — воображение: формирование в сознании новых образов объектов, явлений или их комбинаций, которые в природе и обществе вообще не существуют, т.е. чем больше информации закодировано в мозге, тем богаче мир воображения. По способу мышления различают образное и абстрактное мышление. Образное (конкретно-предметное) мышление, свойственное человеку и животным, представляет собой мышление в действии, проявляющееся в целесообразном адекватном поведении или оценке объекта, явления. Абстрактное мышление (словесно-логическое — с помощью слов, символов, формул) — это отвлеченное понятийное мышление, появление которого тесно связано с развитием речи. Центральным для процесса абстрактного мышления является употребление слова или знака в качестве средства для расчленения и выделения признаков, их абстрагирования и нового синтеза. В результате образуются понятия, а на основе последних — обобщения. Абстрактное мышление свойственно только человеку. Нейроанатомия мышления. Ведущее значение для мышления имеют лобные доли. Получая разнообразную информацию о состоянии организма и окружающей среды, лобная кора оценивает их значимость в настоящий момент и выбирает информацию, имеющую первостепенное значение. Она ответственна за выбор целей, которые человек ставит перед собой. Лобная кора взаимодействует со всеми отделами коры большого мозга, особенно со структурами мозга, обусловливающими языковые функции. Деятельность лобной коры по составлению планов на будущее связана с извлечением информации из долговременной памяти, которая в основном обеспечивается деятельностью височной коры. Амигдала у человека — комплексное образование, включающее несколько групп ядер, располо- 420 Физиология женных в глубине височной доли и имеющих многочисленные связи со многими структурами мозга. Поражение амигдалы у человека нарушает понимание эмоциональных сигналов, исходящих от других людей. Следует отметить, что мозг работает как единое целое. 16.2. Осознаваемая и подсознательная деятельность организма Неосознаваемая (подсознательная) деятельность включает шесть основных видов: • восприятие подпороговых внешних раздражителей, о чем свидетельствует, например, возможность выработки условного рефлекса на очень слабый (неощущаемый) звук; • усвоение ранее поступившей информации, например, после прочтения текста мозг продолжает обрабатывать эту информацию, но организм этого не ощущает, а значит, и не осознает; • на уровне подсознания осуществляются хорошо отработанные автоматизированные элементы работы, а также ходьба, бег и другая привычная деятельность, глубоко усвоенные нормы поведения (динамический стереотип, рис. 16.1); • деятельность мозга во сне, направленная на обработку информации, поступившей до сна, запоминание ее; • интуиция (догадка, чутье), связанная с процессами творчества, которые не контролируются сознанием. Интуиция — источник гипотез, открытий, возможных благодаря трансформации и рекомбинации следов памяти (энграмм), результат подсознательной деятельности мозга на базе высокого профессионализма; • обработка импульсов от внутренних органов. Обычно организм не ощущает импульсацию от внутренних органов, но при чрезмерном возрастании импульсации от них у человека могут возникнуть ощущения. Например, от переполненного желудка, в случае патологии — болевые ощущения. Значение подсознательной деятельности мозга. Ресурсы сознания во время подсознательной деятельности осво- Глава 16. Психическая деятельность и поведение 1 2 421 5 3 5 5 5 6 5 5 5 А 4 Б Рис. 16.1. Отражение системности работы полушарий большого мозга (по Э.А. Асратяну): А — применение системы разных условных раздражителей; Б — повторение одного и того же раздражителя (света) вместо разных условных раздражителей; 1 — звонок; 2 — метроном с частотой 60 уд./мин; 3 — шипение; 4 — метроном с частотой 120 уд./мин (дифференцировка); 5 — свет; 6 — касалка бождаются и переключаются на другую деятельность. Реакции, проходящие на уровне подсознания, энергетически наиболее выгодны, наиболее экономны. Осознаваемая деятельность организма включает два вида деятельности: • все формы психической деятельности — ощущение, внимание, мышление, мотивация, эмоция; • неавтоматизированная физическая работа, что осуществляется в подавляющем большинстве случаев, т.к. при нарушении концентрации внимания немедленно совершаются ошибки в производственной деятельности, в спортивных упражнениях. Например, при выполнении многих элементов в вольных упражнениях, на перекладине один элемент за другим следует почти на подсознательном уровне, что обеспечивает доминантное состояние соответствующих нервных центров. 16.3. Эмоции Эмоции — это переживания организма в виде удовольствия или неудовольствия (от лат. emoveo, emovere — потрясать, волновать). Классификация эмоций. В зависимости от степени выраженности выделяют: настроение — эмоция — аффект; по 422 Физиология знаку — положительные и отрицательные эмоции. Отрицательные эмоции могут быть стеническими (повышение активности организма — физической, духовной) и астеническими (угнетение деятельности организма). Состояние организма во время эмоций зависит от степени их выраженности и знака эмоций. Например, стеническая отрицательная эмоция обычно сопровождается возбуждением ЦНС, выбросом в кровь катехоламинов, ведущих к активации (мобилизации) ряда систем организма — усилению деятельности сердечно-сосудистой системы, дыхания, повышению тонуса мышц, двигательной активности (рис. 16.2), ярко выраженной десихронизации ЭЭГ. Деятельность желудочно-кишечного тракта при этом, как правило, угнетается. Причины возникновения эмоций. Существует несколько теорий возникновения эмоций, что свидетельствует об отсутствии единого мнения. С нашей точки зрения, обобщающий фактор, вызывающий эмоции, — удовлетворение или, напротив, неудовлетворение любой потребности организма (например, успех или неуспех в спорте, учебе, недостаток пищи и т.п.). Нейроанатомия эмоций. Основные структуры, ответственные за формирование эмоциональных реакций, — это элементы лимбической системы, лобные и височные доли. Поражение лобных долей сопровождается эмоциональной тупостью и растормаживанием биологических реакций. При удалении височных областей мозга у обезьян нарушается нормальная пищевая деятельность. Такие животные без конца обследуют предметы, находящиеся перед Рис. 16.2. Периферические компоненты эмоций (по К.В. Судакову, с изменениями) Глава 16. Психическая деятельность и поведение 423 ними, и притом все время берут их в рот и часто поедают несъедобные, т.е. у животных наблюдается потеря оценки значения пищевого раздражителя. Поражение амигдалы у человека нарушает понимание эмоциональных сигналов, исходящих от других людей. Электрическая стимуляция амигдалы у пациентов вызывает эмоции страха, гнева, ярости и редко — удовольствия. Нейрохимия эмоций. Участки мозга, с которых можно получить самораздражение, почти полностью совпадают с зонами иннервации катехоламинергическими нейронами. С увеличением концентрации серотонина в мозге настроение у человека улучшается, его истощение вызывает состояние депрессии и тревоги. Нарушения функционирования холинергической системы мозга ведут к психозу с преимущественным поражением интеллектуальных (информационных) процессов. В генезе положительных эмоций важная роль, очевидно, принадлежит энкефалинам и эндорфинам — эндогенным морфиноподобным веществам, вырабатываемым особыми энкефалин- и эндорфинпродуцирующими клетками ЦНС. Значение эмоций в жизнедеятельности организма. Мобилизация физических и интеллектуальных ресурсов, особенно это ярко выражено у спортсменов. Коммуникативная роль эмоции реализуется с помощью интонации и громкости голоса, мимических и пантомимических движений, позволяющих человеку передавать свои переживания другим людям, сообщать им о своем отношении к объектам, явлениям. Эмоции влияют на здоровье организма. Положительные эмоции оказывают благотворное влияние на состояние здоровья человека, отрицательные — негативное. 16.4. Другие виды психической деятельности К другим видам относятся ощущение, внимание и мотивация. Узнавание, восприятие, представление и воображение — это разные уровни мышления (см. раздел 16.1). Ощущение — осознание действия на организм пороговых или сверхпороговых раздражителей. Всякое ощущение имеет качество, силу и длительность. В зависимости от 424 Физиология качества раздражителя различают: зрительные, слуховые, тактильные и т.д. С помощью ощущения имеется возможность определить локализацию действующих раздражителей на поверхности тела или в окружающей среде. Появление ощущения как особого свойства живой материи связано с длительной эволюцией нервной системы, развитием сенсорных систем и механизмов рефлекторной деятельности в процессе взаимодействия организма с внешней средой. Так, особенности чувствительности глаза определяются свойствами рассеиваемой на Земле солнечной энергии (С.И. Вавилов). Внимание — состояние активного бодрствования, характеризующееся готовностью к деятельности и сопровождающее саму деятельность. В фило- и онтогенезе внимание развивается на базе врожденного ориентировочного рефлекса, направленного на создание в организме условий для восприятия изменений в окружающей среде. Внимание бывает непроизвольным (пассивным) и произвольным (активным). Непроизвольное внимание формируется на сильный либо новый раздражитель, вызывающий такое возбуждение ЦНС, которое воспринимается субъективно в виде ощущений. Непроизвольное внимание — это ориентировочный рефлекс, автоматически включающий произвольное внимание. Произвольное внимание свойственно в основном человеку и заключается в том, что субъект может намеренно сосредоточить свое внимание то на одном, то на другом объекте, явлении, планировании своей деятельности и т.п. Непроизвольное внимание сразу же переходит в произвольное и человек начинает думать, что необходимо предпринимать в возникшей ситуации. Мотивация — побуждение к действию для удовлетворения существующей потребности (см. раздел 16.6). 16.5. Особенности сознания и психической деятельности человека Особенности человеческого сознания. Во-первых, оно социально детерминировано, по наследству не передается, наследуется лишь возможность появления человеческого сознания в онтогенезе, которое формируется только в условиях Глава 16. Психическая деятельность и поведение 425 человеческого общества. У детей, воспитанных животными, формируется сознание животных. У новорожденного животного, помещенного в общество людей, человеческое сознание не формируется. У него формируется сознание животного. Во-вторых, человеческое сознание носит созидательный характер — дает возможность направить действия индивида и коллектива на производство предметов потребления и орудий труда, необходимых для жизнедеятельности. При этом животные не могут дать адекватную оценку деятельности человека. В-третьих, человеческое сознание (в отличие от сознания животных) способствует сохранению всех видов животного и растительного мира. Особенности психической деятельности человека. У человека есть первая и вторая сигнальные системы, у животных — только первая. Сигнальные системы — это совокупность структур организма, обеспечивающих формирование, передачу и восприятие сигналов, и сами сигналы для обмена информацией между особями. Структуры для передачи информации — ЦНС и управляемые ею мышцы и органы, обеспечивающие формирование сигналов. Это мышцы голосовых связок, языка, оральные, дыхательные, а у человека — и верхних конечностей, обусловливающие формирование письма и рисунков. В передаче сигналов участвуют практически все мышцы организма (мимика, жесты, телодвижения и т.д.). Структуры, воспринимающие информацию от других особей, — системы слуха, зрения и головной мозг в целом. Сигналы первой сигнальной системы (ПСС) — элементы языка животных, т.е. совокупность звуковых сигналов, мимики, жестов, позы, движения глаз и тела. Она свойственна и человеку. Сигналы второй сигнальной системы (ВСС) — специфические элементы языка человека — речь и рисунки. Язык человека сформировался на базе языка животных. Язык человека — средство общения людей друг с другом с помощью устной и письменной речи и рисунков, а также элементов языка животных. Речь — средство общения людей друг с другом с помощью устных и письменных сигналов (в виде слов, формул, 426 Физиология символов), обеспечивающее абстрактное мышление человека. При этом смысловое содержание слова, например «стол», не зависит от звучания его в речи на различных языках. Язык человека (и в первую очередь основная его форма — речь) позволяет передавать информацию о текущих событиях, что было в прошлом и прогнозировать, что может произойти в будущем, способствует накоплению знаний об окружающей природе и безмерно повышает власть человека над окружающим миром. Язык животных информирует только о текущих событиях. Речь — главное отличие языка человека от языка животных. Таким образом, первая сигнальная система — это система, обеспечивающая обмен информацией между особями с помощью элементов языка животных. Она присуща и человеку. Вторая сигнальная система — это система, обеспечивающая обмен информацией между людьми с помощью речи и рисунков. Центры речи. В 1861 г. Брока впервые обнаружил, что левостороннее поражение нижних отделов третьей лобной извилины приводит к потере речи (афазии). Такие больные понимают обращенные к ним слова, однако говорить практически не могут. Если их попросить что-либо сказать, они неуверенно и с большим усилием произносят короткие фразы, состоящие из самых необходимых имен существительных, глаголов и прилагательных («телеграфная речь»). Это называется моторной афазией, а область мозга, поражение которой приводит к такому синдрому, — двигательным центром речи, который расположен непосредственно спереди от участков двигательной коры, управляющих мышцами лица, челюсти, языка, нёба и глотки, т.е. мускулатурой, участвующей в артикуляции. Моторная афазия при поражении центра Брока не связана с параличом этих мышц. Несколько позже К. Вернике описал тип афазии, характеризующийся тяжелыми нарушениями понимания речи (сенсорная афазия) при сохранении у больного способности бегло, хотя и несколько искаженно говорить. Такая сенсорная афазия четко совпадает с поражением левой височной доли, особенно задней части первой височной из- Глава 16. Психическая деятельность и поведение 427 вилины в непосредственной близости от слуховой коры (слуховой центр речи). Эти результаты свидетельствуют о том, что центры сенсорной речи расположены лишь в одном из полушарий, тогда как зоны коры, обеспечивающие артикуляцию, т.е. произнесение слов, — в обоих. Речевые функции у правшей локализованы преимущественно в левом полушарии (только у 1–5% лиц — в правом полушарии); у 70% левшей — тоже в левом полушарии, у 15% — в правом, у 15% — в обоих полушариях. Ведущая роль в вынесении суждений принадлежит речевому, доминантному полушарию (левому). В настоящее время известно более 2500 живых развивающихся языков. Следует отметить, что языковые знания не передаются по наследству. Однако у человека имеются генетические предпосылки к общению с помощью речи — это совокупность соответствующих структур ВСС. Амбидекстры — лица, у которых функциональная асимметрия полушарий менее выражена, — обладают большими языковыми способностями. Процесс понимания речи обеспечивается декодированием поступающей в акустической или оптической форме информации. Центры Вернике и Брока связаны дугообразным пучком (рис. 16.3). В процессе восприятия текста и речи возбуждение от слуховых и зрительных рецепторов поступает соответ- Рис. 16.3. Распространение возбуждения в центре речи по Вернике–Гешвинду 428 Физиология ственно по слуховому и зрительному нервам в первичные, вторичные и третичные (ассоциативные) зоны (как и у всех других сенсорных систем). Кроме того, эти возбуждения поступают в центры речи Вернике и Брока и циркулируют между ними по дугообразному пучку, соединяющему эти два центра (см. рис. 16.13). Взаимодействие всех описанных возбуждений обеспечивает понимание смысла речи или текста. Отличия мышления человека от мышления животных. Во-первых, мышление человека может осуществляться на основании инструкций. Во-вторых, у человека имеется образное (конкретно-предметное) и абстрактное (словесно-логическое) мышление, у животных — только образное. Яркий пример, свидетельствующий об отсутствии абстрактного мышления у животных, — поведение обезьяны в опытах И.П. Павлова. Для достижения полезного результата (получения пищи) обезьяна научилась гасить огонь на плоту, находящемся на озере около берега, заливая его водой из бака, который стоял на берегу. Эта обезьяна каждый раз ходила на берег за водой, не подозревая, что вода из озера, окружающая плот, обладает такими же возможностями для тушения огня, что и вода из бака. Более того, когда плот оттолкнули от берега, обезьяна с трудом добиралась по воде из озера до берега, набирала воду ковшом из бака, снова возвращалась по воде на плот (озеро около берега было не глубокое) и гасила огонь на плоту, чтобы достать пищу. Целенаправленная планируемая трудовая деятельность человека включает предварительное планирование в виде схем, чертежей, расчетов, будущих испытаний продуктов труда (жилищ, орудий производства, машин, самолетов и др.). Человек активно изменяет окружающий мир в соответствии с потребностями общества. Труд человека имеет социально-детерминированную направленность. Поведение животных направлено только на приспособление к природе. Человек изготовляет орудия производства, широко используя их в своей производственной деятельности, а также применяет преемственный характер труда. Глава 16. Психическая деятельность и поведение 429 У человека ярко выражена латерализация психических функций. Характер мышления. Левому полушарию в большей степени присуще абстрактное мышление, а правому — образное. Правое полушарие обрабатывает информацию одновременно, синтетически, по принципу дедукции (от общего к частному), при этом лучше воспринимаются пространственные и относительные признаки предметов. Левое полушарие обрабатывает информацию последовательно, аналитически, расчлененно, по принципу индукции (от частного к общему). В организме оба полушария работают как единое целое и имеют комплементарные отношения, т.е. дополняют друг друга. Эмоции. Левое полушарие обусловливает в основном положительные эмоции, контролирует проявление более слабых эмоций. Правое полушарие обусловливает преимущественно более древние, отрицательные эмоции, контролирует проявления сильных эмоций, в целом оно более «эмоционально». Необходимо отметить, что латерализация психических функций имеется и у животных, но она развита слабее. 16.6. Потребности и мотивация как фактор поведения Потребности — это то, что необходимо организму. Единой классификации потребностей не сформировалось, хотя три главные группы определены: биологические (витальные), социальные и идеальные (познание окружающего мира, творчество и т.п.). Предлагаем обобщающую классификацию потребностей по происхождению их роли в жизнедеятельности организма. 1. Биологические потребности, происхождение которых связано с сохранением жизни индивида и вида, они весьма многочисленны: • потребность в хорошем самочувствии, здоровье, физическом развитии, гомеостазисе, самосохранении и сохранении вида; 430 Физиология • родительские потребности — воспроизведение, включающее все этапы, в том числе охрану и воспитание потомства; • половая потребность; • потребность в труде и отдыхе, в том числе и сне. Движение человеку так же необходимо, как и отдых. 2. Бытовые потребности — одежда, жилье, санитария и т.п. Санитарный инстинкт животных у человека утратился и перешел в разряд бытовых потребностей. Удовлетворение бытовых потребностей способствует сохранению здоровья и обеспечивает плодотворную деятельность человека. 3. Социальные потребности и их происхождение связаны со средой обитания: стремление принадлежать к определенной социальной среде и иметь желанную профессию; соблюдение норм поведения в рамках соответствующего общества; стремление к успеху в любой сфере деятельности — творчество, спортивные достижения и т.д. 4. Потребность в исследовании (в ориентировочно-исследовательской деятельности неизвестной, загадочной среды, обстановки, стремление преодолевать трудности и т.п.). Стремление к исследованию «неизвестности» приводит к открытиям, к развитию науки, техники и др. 5. Потребность в свободе у человека находит свое отражение в таких понятиях, как свобода слова, собраний, выбора профессии, свобода и независимость своей Родины и т.п. Эта потребность особенно ярко выражена у диких животных, живущих в неволе, она играет исключительно важную роль в жизнедеятельности человека. Лишение человека свободы — очень сильное наказание. Значение потребностей заключается в том, что они формируют мотивации, стимулирующие деятельность организма для достижения различных целей. Мотив и мотивация. Мотив (от фр. Motif — причина) — обычно это потребность, побуждающая организм выполнить какое-либо действие. Мотивация — это стремление организма выполнить действие с целью удовлетворения существующей у него потребности. Таким образом, потребность является причиной формирования мотивации, поэтому классификация мотиваций соответствует классификации потребностей. Глава 16. Психическая деятельность и поведение 431 Механизм формирования мотивации изучен в основном на примере возникновения биологических потребностей и изменения гомеостазиса внутренней среды организма. Мотивационное возбуждение, возникнув в центрах гипоталамуса вследствие непосредственного действия на них факторов внутренней среды или же в результате поступления к ним импульсации от различных органов и тканей, постепенно начинает распространяться на образования лимбической системы, на нейронные комплексы передних долей коры большого мозга. Дальнейшее усиление активности в гипоталамусе через таламо-тегментальные связи стимулирует восходящую ретикулярную формацию среднего мозга, что приводит к генерализованному возбуждению всех отделов коры большого мозга, предопределяя, по-видимому, новый — доминантный статус мотивационного возбуждения. С этого момента все механизмы мозга по принципу взаимосодействия объединяются в единый комплекс, направляющий организм к удовлетворению той потребности, на базе которой сформировались доминантная мотивация. Нейроанатомия мотиваций. Гипоталамические центры считают ведущими в развитии мотивационного возбуждения, его инициаторами, поэтому их называют пейсмекерными центрами (от англ. pacemaker — водитель ритма). Разрушение этих центров немедленно устраняет возможность появления той или иной мотивации. Например, состояние голода у подопытной крысы не возникает после разрушения гипоталамического центра голода, расположенного в области латеральных ядер гипоталамуса. Одни нейроны активируются при снижении уровня глюкозы в крови, другие, наоборот, — при возрастании ее содержания. В области супраоптических и паравентрикулярных ядер гипоталамуса обнаружены нейроны, чувствительные к уровню осмотического давления крови, в области заднедорсального гипоталамуса — к норадреналину и т.д. Кора большого мозга обеспечивает поиск, завершение целенаправленного поведения организма во внешней среде, приводящие к удовлетворению потребности. Нейрохимия мотиваций. Мотивация страха, например, блокируется α-адреноблокаторами (аминазином), 432 Физиология пищевые мотивации — холинолитиками (атропином или амизилом). Ангиотензин, брадикинин, вазопрессин изменяют поведенческие реакции, сформированные на базе мотивации страха. Мотивации активируют сенсорные системы. При голоде возрастает возбудимость вкусовых рецепторов, понижается порог дифференцировочных условных рефлексов. При половом возбуждении повышается возбудимость рецепторов эрогенных зон и т.д. Любая мотивация повышает активность моторной системы. Исключение — пассивный страх во время замирания животного. Мотивация побуждает организм к формированию поведения для удовлетворения существующей потребности. 16.7. Функциональная система поведенческого акта Для достижения цели, т.е. полезного для организма результата, формируются функциональные системы. Учение о функциональных системах разработал П.К. Анохин (середина ХХ в.), это учение успешно развивалось К.В. Судаковым. Функциональная система — это динамическая совокупность различных органов и физиологических систем, формирующаяся для достижения приспособительного (полезного) для организма результата. Общая схема формирования функциональных систем включает четыре основных стадии (рис. 16.4). 1. Афферентный синтез заключается в обработке и сопоставлении всей информации, которая используется организмом для принятия решения и формирования наиболее адекватного для данных условий приспособительного поведения с учетом доминирующей мотивации, обстановочной афферентации, пусковой афферентации, взаимодействующих с аппаратом памяти. Мотивационное возбуждение возникает в ЦНС с появлением какой-либо потребности, оно имеет доминирующий характер (доминанта открыта А.А. Ухтомским), при этом подавляются остальные мотивации, поведение организма направляется на достижение полезного результата, который удовлетворяет имеющуюся потребность. Глава 16. Психическая деятельность и поведение 433 Рис. 16.4. Схема центральной архитектоники поведенческого акта (по П.К. Анохину, с изменениями) Воспоминание — мотивационное возбуждение извлекает необходимую информацию из блоков памяти, которая нередко определяет целенаправленную деятельность организма на основании приобретенного ранее опыта по достижению данного полезного результата. Обстановочная афферентация вызывает возбуждение, обусловленное действием на организм окружающей среды: в ЦНС создается разветвленная система возбуждений, своего рода «нервная модель обстановки». Обстановочная афферентация может способствовать либо, напротив, препятствовать реализации мотивации. Например, на лекции студенты не включают музыку. Пусковая афферентация — это специальное возбуждение, обусловленное действием пусковых раздражителей, оцениваемых организмом как сигнал к действию, например, звонок во время урока означает сигнал на перемену. 2. Эфферентный синтез — развитие афферентного синтеза. Он характеризуется тем, что содержит элементы предвидения (что совершится в ближайшее время), поскольку на данной стадии формируются принятие решения, программа действия и акцептор результата действия (АРД). Благодаря принятию решения для совершения определенного акта организм будет выполнять только одну конкретную форму поведения, соответствующую потребности и окружающей обстановке. 434 Физиология Формирование АРД происходит на основе афферентного синтеза. АРД — нервная модель будущего результата, который должен быть получен в соответствии с принятым решением. Например, принято решение взять кусок мела со стола. В АРД возникает возбуждение тех нейронов, которые ранее стимулировались при взятии мела. Они отражают его форму, параметры. Формирование программы действия идет параллельно с формированием АРД. Аппарат программы действия обеспечивает посылку импульсов (эфферентное возбуждение) к эффекторам для совершения определенного действия, приводящего к получению полезного результата. Начало этого пути — пирамидные клетки коры большого мозга. Команды в виде нервных импульсов одновременно направляются по коллатеральным путям и в аппарат АРД, где они взаимодействуют с запрограммированным результатом, вследствие чего осуществляется его коррекция. 3. Действие характеризуется выполнением программы поведения: эфферентное возбуждение доходит до эффекторов и осуществляет действие (рис. 16.5). Стадия находится под контролем АРД за счет информации, постоянно поступающей от проприорецепторов мышц, т.е. обратной афферентации. Поскольку функциональная система формируется для достижения приспособительного результата, его считают системообразующим фактором, которым является и доминирующая мотивация. 4. Оценка достигнутого результата осуществляется с помощью сопоставления обратной афферентации о параметрах достигнутого результата с ранее сформировавшейся нервной моделью результата в АРД. Если афферентная модель реального результата совпадает с нервной моделью запрограммированного результата, то происходит удовлетворение ведущей потребности, что сопровождается положительной эмоцией. Удовлетворение потребности ведет к прекращению мотивационного возбуждения, функциональная система распадается. В случае несовпадения возбуждения от параметров полученного результата с возбуждением от параметров запрограммированного результата возникает ориентировочно-исследовательская реакция, Глава 16. Психическая деятельность и поведение 435 сопровождающаяся отрицательной эмоцией. При этом вовлекаются все механизмы мозга и формируется новая, более совершенная функциональная система для удовлетворения существующей потребности. 16.8. Механизм произвольных движений В условиях свободного поведения животного зарегистрирована активность отдельных нейронов, локализующихся в различных областях мозга (в гипоталамусе, соматосенсорной коре, ретикулярной формации, гиппокампе, таламусе), приуроченная к определенной деятельности организма. Управление двигательными актами строится на двух основных принципах — прямого программного управления и сенсорных коррекциях выполняемого движения (обратной афферентации). Следует выделить три блока структур мозга, обеспечивающих выполнение произвольного двигательного акта (рис. 16.5). 1. Блок принятия решений, планирования движений и их инициация — лобная ассоциативная кора (префронтальная кора) и теменная ассоциативная кора большого мозга. Эти структуры обеспечивают оценку ситуации с помощью информации, поступающей от сенсорных систем внешней и внутренней среды организма, и формирование функциональной системы, обеспечивающей выполнение поведенческой реакции. Ведущая роль в построении новых Рис. 16.5. Три блока структур ЦНС, обеспечивающих выполнение двигательного акта 436 Физиология моторных программ принадлежит префронтальной коре. Поражение в префронтальной коре может привести вообще к распаду произвольной регуляции движений. Этот дефект особенно выражен, когда требуется менять программы движений. Передние отделы коры для построения новых моторных программ используют весь видовой и накопленный в течение жизни индивидуальный опыт. 2. Блок хранения программ движения — вторичная двигательная кора (поле 6, включающее премоторную кору и дополнительную моторную область), мозжечок, базальные ядра. Двигательные программы, сформированные в онтогенезе, хранятся в перечисленных структурах и других компонентах пирамидной и экстрапирамидной систем. В частности, в процессе индивидуальной жизни мозжечок обучается различным программам движения и сохраняет их. Мозжечок получает информацию о планируемом корой большого мозга движении, вносит коррекцию в план будущего движения и посылает ее в кору большого мозга. Быстрые мелкие точные движения вообще осуществляются без сличения корой с импульсами от проприорецепторов. Мозжечок обучается также корригировать выполняемое движение, играет важную роль в регуляции тонуса мышц, поддержании позы. Базальные ядра (стриопаллидарная система) включают полосатое тело (от лат. corpus striatum), называемый также striatum (хвостатое ядро и скорлупа), бледный шар и субталамическое ядро. Базальные ядра имеют отношение к хранению программ врожденных двигательных актов, быстрых стереотипных движений. Врожденные программы имеются также в спинном мозге (шагательный рефлекс) и в стволе мозга (дыхание, глотание, выпрямительный рефлекс — восстановление нарушенной позы, ориентировочный рефлекс). Двигательные программы вырабатываются и в онтогенезе, например каскад гимнастических элементов. 3. Исполнительный блок, включающий пирамидную и экстрапирамидную системы. Пирамидная система — совокупность нейронов пирамидного пути, α- и γ-мотонейронов ствола мозга и спинного мозга, синаптически связанных друг с другом непосредственно и с помощью вставочных нейронов. Пирамидный Глава 16. Психическая деятельность и поведение 437 путь образуют аксоны пирамидных клеток V слоя коры большого мозга (клетки Беца, первичная двигательная кора). Мотонейроны ствола мозга включены в ядра черепных нервов, за исключением I, II, VIII пар. Пирамидная система вместе с экстрапирамидной системой — исполнительный блок произвольных движений. Пирамидная система играет особо важную роль в выполнении тонких движений пальцев рук и кистей. Экстрапирамидная система (ЭПС) — совокупность ядер головного мозга, управляющих α- и γ-мотонейронами спинного мозга и ствола мозга, входящих в пирамидную систему. Ядра ЭПС — двигательная кора (кроме первичной), базальные ядра, черная субстанция, красные ядра, люисово тело (субталамическое ядро), ретикулярная формация, центры четверохолмия, голубое пятно, вестибулярные ядра и мозжечок. Функция — регуляция мышечного тонуса, сохранение естественной позы, восстановление нарушенной позы, участие в осуществлении произвольных движений. Само движение запускается непосредственно α-мотонейронами спинного мозга и ядер черепных нервов ствола мозга, с которыми синаптически связаны пирамидные клетки коры большого мозга (первичной двигательной коры — клетки Беца). Важную роль при выполнении двигательного акта играют γ-мотонейроны, участвующие в регуляции тонуса мышц, и афферентная импульсация. Роль афферентных импульсов в управлении движением. Ч. Шеррингтон впервые указал на существование сенсорных обратных связей, которые регулируют активность мотонейронов. Он ввел термин «проприоцепция» для обозначения сенсорной импульсации от возбуждающихся во время движения проприорецепторов и дающих информацию о собственных движениях организма. Выделяют два вида обратной афферентации при выполнении движения — направляющую и результативную. Направляющая афферентация поступает в ЦНС от проприорецепторов и участвует в регуляции тонуса мышц, сохранении естественной позы и восстановлении нарушений позы, координации движений. Нарушение проприоцептивной импульсации ведет к расстройству движений. Результативная импульсация идет от рецепторов других орга- 438 Физиология нов чувств (главным образом от рецепторов систем зрения и тактильной). С помощью результативной афферентации организм оценивает полученный вследствие двигательного акта результат (более подробно о роли афферентации от проприорецепторов см. в разделе 8.1). Главные отличия формирования произвольных и непроизвольных движений. Непроизвольные движения запускаются посредством возбуждения экстеро- и проприорецепторов, могут осуществляться с помощью любого уровня ЦНС, в том числе и на уровне спинного мозга. Произвольные движения инициируются с помощью ассоциативной коры большого мозга; они осознанно регулируемые и осуществляются с заранее намеченной целью. У человека (в отличие от животных) цели произвольных движений формируются с помощью речевых процессов. Роль речевого опосредования в превращении непроизвольных импульсивных движений у детей в произвольные и осознанно управляемые изучили А.Р. Лурия и его сотрудники, которые показали, что активация детской речи (предварительное планирование в речевой форме предпринимаемых действий) ведет ребенка к овладению своим поведением, тогда как задержка в речевом развитии детей в патологических случаях приводит к снижению уровня произвольной (волевой) регуляции двигательного (поведенческого) акта. Предметный указатель А Автоматия: дыхательного центра 220 сердца 231 Адреналин 172, 259 Азотистое равновесие 303 Аксонный транспорт 44 Альдостерон 172 Анаболизм 301 Антидиуротический гормон (вазопрессин) 164 Антикоагулянты 198 Ассимиляция 301 Ауторегуляция: вдоха и выдоха 220 синапсов 146 Афферентный синтез 422 Б Базальные ядра Белковый минимум Белковый оптимум Бодрствование 125 302 303 411 В Вдох и выдох Вестибулярная система 207 370 Внимание 424 Возбудимость 23, 32 Воздухоносные системы 204 Всасывание питательных веществ 295 Выделение 326 Высшая нервная деятельность 392 Г Газообмен 213 Гематоэнцефалический барьер (ГЭБ) 70, 79 Гемоглобин 183, 215 Гемодинамика: 243 линейная скорость кровотока 245 объемная скорость кровотока 245 особенности кровотока в отдельных органах 255 по артериям 248 по венам 253 по капиллярам 251 Гетерометрический механизм регуляции 241 Гибкие связи 86 440 Предметный указатель Гипноз 414 Гипоталамус 124, 150 Глаз 356 Глотание 280 Голубое пятно 119 Гомеометрический механизм регуляции 242 Гормоны: гипофиза 162 классификация 159 механизм действия 160 надпочечников 171 поджелудочной железы 168 половые 173 тимуса 168 тканевые 160 щитовидной железы 165 Группы крови 188 Д Децеребрационная ригидность Диссимиляция Дыхание 114 302 202 Ж Жесткие связи 86 З Зрение в различных условиях 360 И Иерархический принцип организации деятельности ЦНС 88 Импринтинг 405 Интегративная роль нервной системы 99 К Кальцитриол 167 Катаболизм 302 Клетка: 11 глиальная 78 нервная 75 Кодирование 354 Координационная деятельность ЦНС 96 Кора большого мозга 127–135, 152 Коронарный кровоток 256 Кортикализация функций 128 Коэффициент изнашивания 303 Кровь 176 Л Лабильность Легочные объемы емкости Лейкоциты Лимбическая система Лимфатическая система 34 211 212 184 126 267 М Медиаторы: вегетативной нервной системы 142–149 нервно-мышечного синапса 48 ЦНС 81 Медленная диастолическая деполяризация 233 Мезэнцэфальный организм 112 Межполушарные связи 134 Предметный указатель 441 Метасимпатическая нервная система 149 Методы исследования: ВНД 415 деятельности сердца 234 ЦНС 135 Микрофлора кишечника 297 Мозжечок 120 Мотивация 424, 430 Мышление 418 Мышцы: гладкая 62 сердечная 227 скелетная 52 Н Наполнение мочевого пузыря 342 Научение: 398–409 когнитивное 407 оперантное 406 пассивное 405 подсознательное 408 Нервное волокно 43 Нефрон 327 Новая кора 127 О Обмен веществ: белков воды липидов минеральных веществ углеводов Обмен энергии: общий основной пластический 301 302 309 305 310 307 314 316 314 316 Осмолярность и регуляция 339 Особенности человеческого сознания 424 Осознаваемая деятельность 422 Ощущение 423 П Память 392 Петля Генле 333 Пирамидная система 134 Питание 312 Пищеварение: 272 в двенадцатиперстной кишке 288 в желудке 282 в полости рта 277 в толстой кишке 297 в тонкой кишке 292 закономерности 274 функции 272 Подсознательная деятельность 420 Потенциалы клетки: ВПСП 83 действия 27 локальный 31 покоя 25 пороговый 35 Потребности 429 Почечные клубочки 330 Почечный клиренс 335 Проксимальные извитые канальцы 332 Принципы исследования прихода и расхода энергии 318 Проведение возбуждения 43 442 Произвольные движения 435 Промежуточный мозг 122 Проприоцептивная система 372 Психическая деятельность 417 Р Раздражимость 23 Регуляция: гуморальная 67 по опережению 72 по отклонению 72 Регуляция АД: 261 механизмы быстрого реагирования 261 механизмы медленного реагирования 265 механизмы небыстрого реагирования 264 Регуляция тонуса сосудов: 257 гуморальная 259 миогенная 258 нервная 257 Ретикулярная формация: спинного мозга 102 ствола мозга 118 Рецепторы: 38 ионотропные 144 метаботропные 143 Рефлексы: 65 двигательные 103–110 кашлевой 222 классификация 103–110, 115–117 чихания 223 Рефлекторная дуга 65 вегетативного рефлекса 139 Речь 426 Предметный указатель С Связи в ЦНС: гибкие 86 жесткие 86 локальные 85 Сенсорные системы: 346 вкуса 379 внутренней среды 378 классификация 346 кодирование 354 обоняния 381 отделы 348 свойства 350 тактильная 375 температуры 373 Серотонический нерв 146 Сигнальные системы 425 Синапсы: нервно-мышечный 48 ЦНС 77 Система боли 384 Система зрения: 356 корковый отдел 360 периферический отдел 356 проводниковый отдел 359 Система зуда 376 Система слуха 366 Сновидения 414 Сознание 411 Сон 411 Спинальный шок 110 Спирограмма 211 Стриопаллидарная система 125 Сурфактант 206 Т Таламус 122 Темпераменты личности 410 Терморегуляция 321 Предметный указатель Теплоотдача 323 Теплопродукция 322 Типы ВНД 409 Тонус: блуждающих нервов 153, 157, 238 вегетативных центров 152 мышц 110, 115 Торможение: пессимальное 58 постсинаптическое 94 пресинаптическое 94 условных рефлексов 401 Транспорт веществ 15 Транспорт газов 215 Тромбоциты 187 Ф Феномен Орбели–Гинецинского 153 Фибринолиз 199 Физиологическая система 9 Фонокардиография 237 Функции почек 329 443 Функциональная система 9, 70 Х Хронаксия 36 Ц Цикл сердечной деятельности 225 Ч Черная субстанця 119 Э Экстрапирамидная система Электрокардиография Эмоции Эритроциты Эфферентный синтез 437 235 421 179 433 Я Язык (речь) человека 425 Литература 1. Агаджанян Н.А., Смирнов В.М. Нормальная физиоло- гия: Учебник. — М.: МИА, 2012. 2. Анохин П.К. Биология и нейрофизиология условного рефлекса. — М.: Медицина, 1968. 3. Афанасьев В.Г., Офрихтер Е.Г., Берг М.Д., Кононова М.Л., Смирнова В.М. Курс лекций по общей валеологии. — Пермь: Перм. гос. мед. академия, 2000. 4. Ашмарин И.П. Молекулярные механизмы нейрологической памяти (Механизмы памяти): Руководство по физиологии. — Л.: Наука, 1987. — С. 57–77. 5. Барбараш Н.А. Обмен веществ и энергии, терморегуляция // Нормальная физиология / Под ред. В.М. Смирнова. — М.: Академия, 2012. — С. 304–327. 6. Вейн А.М., Хехт К. Сон человека. Физиология и патология. — М.: Медицина, 1989. 7. Волкова О.В., Елецкий Ю.К. Гистология, цитология и эмбриология: Атлас. — М.: Медицина, 1996. 8. Гайтон А.К., Холл Д.Э. Медицинская физиология: Учебник. — М.: Логосфера, 2008. 9. Годфруа Ж. Что такое психология: Пер. с фр.: В 2 т. Т. 2. — М.: Мир, 1992. 10. Голышенков С.П. Физиология крови. Система гемостаза и при мышечной деятельности и в покое. — Саранск, 2004. 11. Дерягина Л.В., Зашихина В.В. Физиологические особенности челюстно-лицевой области. — Архангельск, 2004. 12. Завьялов А.В. Физиология человека. — СПб.; Самара, 2002. 13. Калюжный Л.В. Физиологические механизмы боли и аналгезии // Физиол. журнал СССР им. И.М. Сеченова. — 1991. — Т. 77. — № 24. — С. 123–133. Литература 445 14. Киричук В.Ф. Железы внутренней секреции. Система крови // Нормальная физиология / Под ред. В.М. Смирнова. — М.: Академия, 2012. — С. 144–193. 15. Кромин А.А. Система пищеварения // Нормальная физиология / Под ред. В.М. Смирнова. — М.: Академия, 2012. — С. 265–303. 16. Кузник Б.И., Максимова О.Г. Общая гематология. Гематология детского возраста: Учебное пособие. — Ростов н/Д: Феникс, 2007. 17. Маркина Л.Д. Физиология высшей нервной деятельности: Учебное пособие. — Владивосток: Медицина ДВ, 2003. 18. Медведев М.А., Хоч Н.С., Низкобудова С.В., Байков А.Н. Физиология высшей нервной деятельности и сенсорных систем: Учебное пособие. — Томск: Изд-во Томского ун-та, 2003. 19. Меерсон Ф.З. Адаптационная медицина: концепция долговременной адаптации. — М.: Медицина, 1993. 20. Механизм проведения возбуждения по нервному волокну // Бюллетень эксп. биологии и медицины. — 2008. — Т. 146. — № 8. — С. 124–127. 21. Мищенко В.П., Мищенко И.В. Физиология системы гемостаза. — Полтава: АСМИ, 2003. 22. Наточин Ю.В. Физиология водно-солевого обмена. — СПб.: Наука, 1993. 23. Ноздрачев А.Д., Баженов Ю.И., Баранникова И.А., Батуев А.С. и др. Начала физиологии. — СПб.: Лань, 2001. 24. Павлов И.П. Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных. — М.: Медгиз, 1951. 25. Правдивцев В.А., Яснецов В.В., Козлов С.Б. и др. Основы системных механизмов высшей нервной деятельности. — Смоленск: Гос. мед. академия, 1997. 26. Розен В.Б. Основы эндокринологии. — М.: Высшая школа, 1980. 27. Рухлова С.А. Основы офтальмологии. — Н. Новгород: Изд-во НГМА, 2001. 28. Савченков Ю.И. Основы психофизиологии: Учебное пособие для психологов. — Ростов н/Д: Феникс, 2007. 446 Литература 29. Сергеев П.В., Шимановский Н.Л., Петров В.И. Рецепторы физиологически активных веществ. — Волгоград: Семь ветров, 1999. 30. Сеченов И.М. Рефлексы головного мозга. — М.: Медгиз, 1961. 31. Симонов П.В. Мозг: эмоции, потребности, поведение. — М.: Наука, 2004. 32. Смирнов В.М. Физиология человека: Учебник. — 4-е изд. — М.: Академия, 2012. 33. Судаков К.В. Нормальная физиология: Учебник. — М.: МИА, 2006. 34. Ткаченко Б.И. Нормальная физиология человека: Учебник. — М.: Медицина, 2005. 35. Камкин А.Г., Каменский А.А. Фундаментальная и клиническая физиология: Учебник. — М.: Академия, 2004. 36. Шеперд Г. Нейробиология. Т. 2. — М.: Мир, 1987. 37. Шмидт Р., Тевс Г. Физиология человека: Руководство: В 3 т. — М.: Мир, 1996. 38. Шульговский В.В. Физиология высшей нервной деятельности с основами нейробиологии. — М.: Академия, 2003. 39. Эккерт Р., Рэнделл Д., Огайн Д. Физиология животных: В 2 т. — М.: Мир, 1992. 40. Яковлев В.Н. Частная физиология центральной нервной системы // Нормальная физиология / Под ред. В.М. Смирнова. — М: Академия, 2012. — С. 92–128. Для заметок Учебное издание Смирнов Виктор Михайлович, Смирнов Андрей Викторович Физиология Оригинал-макет подготовлен ООО «Медицинское информационное агентство» Главный редактор А.С. Петров Санитарно-эпидемиологическое заключение № 77.99.60.953.Д.000945.01.10 от 21.01.2010 г. Подписано в печать 04.07.2016. Формат 84 × 108/32 Бумага офсетная. Гарнитура «NewtonC». Печать офсетная. Объем 14 печ. л. Тираж 1000 экз. Заказ № ООО «Издательство «Медицинское информационное агентство» 119048, Москва, ул. Усачева, д. 62, стр. 1, оф. 6 Тел./факс: (499) 245-45-55 E-mail: [email protected] http://www.medagency.ru Интернет-магазин: www.medkniga.ru Отпечатано в ОАО «Тверской полиграфический комбинат» 170024, г. Тверь, проспект Ленина, д. 5