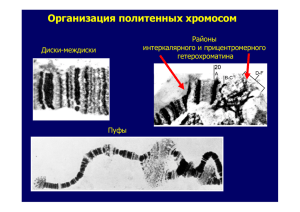
Генетика для студентов КРИ Электронный лекционный курс – мультимедийная презентация Анна Михайловна Гусаченко Часть 3. Хромосомная теория наследственности Хромосомная теория наследственности В начале 20 в. было известно о том, что число хромосом организма постоянно, открыты митоз и мейоз. Сэттон и Бовери отмечали сходство в поведении генов и хромосом (19021903). В это время многие цитологи исследовали хромосомные числа различных животных и растений. Половые хромосомы В начале 20 в. цитологи исследовали мейоз у насекомых. У клопа Protenor в гаметах 6 или 7 хромосомотличаются Х-хромосомой – Х0 определение пола. У клопа Lygaeus одна пара отличается: Y меньше, чем Х, в гаметах Х или Y- XY определение пола. Хромосомная теория наследственности Использовал плодовую мушку Drosophila melanogaster как модельный объект генетики Ученики: А. Стертевант, К. Бриджес, Г. Мёллер Томас Морган (1866-1945) Цикл развития дрозофилы Взрослая муха яйцо При 25 С от яйца до яйца 14 дней куколка эмбрион личинка Drosophila melanogaster, дикий тип самка самец •Половые хромосом дрозофилы Нашли половые хромосомы у дрозофилы: X и Y Х Х Х Y Метафазные хромосомы Хромосомы Drosophila melanogaster X Y самец X X самка Ген white (1910 г.). Нашли самца с белыми глазами, получили линию. Морган с учениками провели два реципрокных скрещивания : в одном мутантный ген был у самца, в другом- у самки белые глаза х красные гл. х красные гл. белые глаза Мендель показывал, что реципрокные скрещивания дают одинаковые результаты P: w/w белые глаза G: x w F1: w/w+ красные 1 w+/ Y красные гл. w+ x : G: w w+ F2: w/w w+/w x белые красные 1 : 1 : Y w/ Y белые 1 w Y w/ Y w+/ Y белые красные 1 : 1 P: w/w белые глаза G: x w F1: w/w+ красные 1 w+/ Y красные гл. w+ x : G: w w+ F2: w/w w+/w x белые красные 1 : 1 : Y w/ Y белые 1 w Y Крисс-кросснаследование: сыновья наследуют признак матери, а дочери - отца w/ Y w+/ Y белые красные 1 : 1 P: w+/w+ красные глаза G: F1: x w+ w w/w+ красные x w+ F2: w+/w+ w+/w x красные красные : Y w+/ Y красные G: w w+ 1 w/ Y белые глаза 1 : Y w/ Y w+/ Y белые красные 1 : 1 Реципрокные скрещивания различались и не соответствовали менделевскому расщеплению Морган предположил, что ген white находится в Х-хромосоме, а Y не содержит ген white У самца Х-хромосома одна, ген white в гемизиготе У дрозофилы самки- гомогаметный пол, самцы – гетерогаметный пол Нерасхождение половых хромосом • Ученик Моргана Бриджес обнаружил, что в скрещиваниях • белоглазые х красноглазые • с частотой 0.001-0.1 % появляются • белоглазые красноглазые • Предположил, что хромосомы не разошлись в мейозе - произошла ошибка, мутация Нерасхождение половых хромосом XX w X w X w+ w 0 w w w+ w+ w+ Y w w 0.001-0.1 % 0.001-0.1 % Нерасхождение половых хромосом вторичное X X Y XY XX Y Нерасхождение хромосом постоянно: сцепленные Х-хромосомы • Мутация yellow (желтое тело) – в Ххромосоме • В 1922 г. Л.В.Морган обнаружила в скрещивании желтой самки yy с серым самцом исключительную (единственную) желтую самку. P yy x y+ / Y желтая серый F1 yy желтая y+ / Y серый Нерасхождение хромосом постоянно: сцепленные Х-хромосомы • Эта самка далее давала потомство, сходное с родителями: желтые самки и серые самцы. • Цитологический анализ показал, что 2 акроцентрические Х-хромосомы у таких самок оказались соединенными в одну метацентрическую. Это тоже мутация - хромосомная перестройка. Еще эти самки имеют Y-хромосому: желтая серый Напишите схему скрещивания Выводы Гены, расположенные в Х- хромосоме, наследуются не по Менделю. Видно, что у дрозофилы пол определяет число Х-хромосом, Y на развитие пола не влияет. Бриджес исследовал это далее и вывел балансовую теорию определения пола Хромосомная теория наследственности Сэттон заметил, что признаков больше, чем хромосом. Одна хромосома должна определять несколько признаков. Тогда должен нарушаться закон Менделя о независином наследовании признаков. Стали искать такие нарушения • В 1906 г. У.Бэтсон и Р.Пеннет, работая с растением «душистый горошек» Lathyrus odoratus, обнаружили отклонение от расщепления 9:3:3:1 Признаки: цветки пурпурные -Р, красные -р, пыльцевые зерна удлинённые -L, круглые -l • В 1906 г. У.Бэтсон и Р.Пеннет, работая с растением «душистый горошек» Lathyrus odoratus, обнаружили отклонение от расщепления 9:3:3:1 P PPLL x ppll F1 PpLl F2 P_L_, P_ll, ppL_, ppll %: 69.5: 5.6: 5.6: 19.3 реальные % 56.25 :18.75 : 18.75 : 6.25 ожидаемые Такое расщепление получится, если гаметы образуются в таких соотношениях: 0.44 PL: 0.06 Pl : 0.06 pL : 0.44 pl, вместо 0.25. Гены pl сцеплены – наследуются вместе. Сцепление оказалось не полным, а частичным Расщепление в F2 PL 69.5 56.25 75 Pl 5.6 pL 5.6 18.75 18.75 0 0 pl Фенотипы Расщепления 19.3 Реальное Неполное сцепление 6.25 Независимое 9:3:3:1 25 Полное сцепление 3:1 Позже, в 1919 г. Холдейн рассчитал, какие должны быть частоты гамет, чтобы вышло полученное расщепление: PL 0.44 PL 0.44 Pl 0.06 pL 0.06 pl 0.44 Pl 0.06 pL 0.06 pl 0.44 Работы Моргана с сотрудниками b-(black) – черное тело, b+ - серое тело vg –(vestigial)- короткие редуцированные крылья, vg+ - нормальные крылья Получил F1 и сделал 2 реципрокных анализирующих скрещивания В первом получилось 2 сорта потомков- только родительские, во втором – 4 сорта: в 83 случаях родительские комбинации, в остальных 17% Работы Моргана с сотрудниками Показали: Если гетерозиготен отец F1. в F2 образуется только 2 родительских варианта- полное сцепление. . Работы Моргана с сотрудниками Показали: Если гетерозиготна мать F1, в F2 образуется 4 фенотива: 2 родительских и 2 рекомбинантных Обмен- кроссинговер у дрозофилы происходит только у самок. Потом взяли мух с другим сочетанием признаков: bb vg+vg+ x b+b+ vg vg В анализирующих скрещиваниях получили обратные результаты Стертевант: Теперь 3 гена, в анализирующем скрещивании 8 вариантов: Родительские классы Обмен на 2-м участке Обмен на 1-м участке Обмены на двух участках Получили частоты рекомбинации: b – pr – 6%, pr – vg – 12%, b – vg – 17% Обнаружили, что между одной парой генов частота кроссинговера больше, чем между другой. Стертевант предположил: •гены в хромосоме расположены линейно, •частота кроссинговера зависит от расстояния между генами, •можно построить карту: b pr 6% vg 12% Гипотеза А.Стертеванта: частота рекомбинации пропорциональна расстоянию между генами rf- частота рекомбинации d - расстояние между генами Предложил использовать частоту кроссинговера как меру расстояния между генами на хромосоме Единица расстояния – процент кроссинговера – cантиморганида, сМ 1 сМ = 1 % кроссинговера Предположили, что происходит физический обмен участками хромосом. Хиазмы в мейозе уже видели Генетическая карта Drosophila melanogaster Группа сцепления группа генов, проявляющих сцепленное наследование, группа генов, расположенных в одной хромосоме. К 1980 г. построены генетические карты человека, дрозофилы, мыши, крысы, домашней мухи, таракана, курицы, пшеницы, риса, кукурузы и пр.. Расстояние между генами: число кроссоверов х100% = 17 % общее число потомков Сколько хиазм может образоваться в биваленте? Х-хромосома, унивалент, 0 хиазм 4 1 2 3 В генетике мы наблюдаем кроссинговер только по смене маркеров АВС (генов) А В С а b Видим c не видим Если гены (маркеры) расположены далеко (В и С), между ними может произойти 2 обмена, а мы их не заметим. Число кроссинговеров, который мы заметим, будет меньше, чем реально их произошло. В хромосоме на малых расстояниях действует интерференция обменов: обмены не могут происходить близко, один обмен запрещает другой обмен поблизости. Если маркеры А и В расположены близко, мы видим все обмены, которые произошли. Интерференцию на определенном участке хромосомы можно оценить по произошедшим двойным обменам А 0.3 В 0.2 С Пусть генетическое расстояние между АВ – 30%, ВС20%, двойные обмены – 1% С= количество реально произошедших двойных обменов на участке А-С к теоретически возможным. Теоретическая вероятность двойного обмена рассчитывается как вероятность одновременных обменов на участке АВ и ВС. Тогда С=0.01/(0.3х0.2) = 0.16 Это коэффициент Коинциденции, он оценивает интерференцию C= rf double/ (rf AB x rf BC) Функция Холдейна rf = 1/2(1-e-2d) d = -1/2 ln(1-2rf) rf- картирующая функция, частота видимых кроссоверов, d – реальное расстояние по карте. Расстояния меньше 10 сМ (в сумме) можно просто суммировать Картирующие функции: 1 – А.Стертеванта (rf=d), 2 – функция Дж.Холдейна Функция Холдейна rf = 1/2(1-e-2d) d = -1/2 ln(1-2rf) rf- картирующая функция, частота видимых кроссоверов, d – реальное расстояние по карте. Расстояния меньше 10 сМ (в сумме) можно просто суммировать Картирующие функции: 1 – А.Стертеванта (rf=d), 2 – функция Дж.Холдейна Теперь мы знаем, что кроссинговер происходит на стадии 4-х хроматид, каждый гомолог состоит из 2-х идентичных хроматид: В кроссинговере в одной точке участвует только 2 их 4-х хроматид, по одной от каждого гомолога Каждая клетка после мейоза дает 4 клетки с одной из 4-х хроматид. Из них 2 – некроссоверные, 2 кроссоверные. Или все 4 некроссоверные, если обмен произошел вне маркеров АВС. Поэтому кроссоверов может быть не больше 50% (<=50%) Цитологическое доказательство кроссинговера Эксперимент К. Штерна (1930) Хромосомы дрозофилы маркированы цитологически: Одна Х-хромосома несет кусочек Y, другая короче из-за транслокации участка на 4 хромосому; генетически : cr- глаза цвета гвоздики, cr+ - дикий тип, В – узкие глаза, В+ нормальные Некроссоверы кроссоверы Цитологическое доказательство кроссинговера Эксперимент Х. Крейтон и Б. МакКлинток на кукурузе Маркеры Цитологические: хромосома имеет блок гетерохроматина и более длинная из-за транслоцированного куска другой хромосомы Генетические: с, - бесцветный, с+ - окрашенный эндосперм, wx – восковой, wx+ - нормальный эндосперм некроссоверы кроссоверы