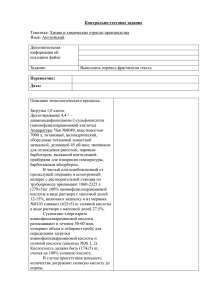
Новиков Д.А. Выделение и очистка продуктов биотехнологии КУРС ЛЕКЦИЙ Минск БГУ 2014 УДК 631.147(075.8) ББК 28.072 я 7 Б63 А в т о р ы - с о с т а в и т е л и: Д. А. Новиков Рецензенты: Кафедра биохимии и биофизики Международного государственного экологическогоуниверситета имени А.Д. Сахарова (заведующий кафедрой кандидат биологических наук, доцент С.Б. Бокуть); Доцент кафедрыбиотехнологии и биоэкологи Белорусского государственного технологического университета кандидат биологических наук, доцент Н.А. Белясова Выделение и очистка продуктов биотехнолгии. Б63 Методическое пособие / авт.: Д.А. Новиков – Минск.: БГУ, 2014. – 256 с. В Курсе лекций приводятся основные вопросы, касающиеся характеристики основных биотехнологических процессов, используемых для производства широкого спектра биологически активных субстанций, а также основные методы выделения очистки и фракционирования продуктов биотехнологии. Уделено внимание созданию нормативно-технической документации на продукты биотехнологии; основным продуктам биотехнологического производства, их характеристике, методам контроля и областям применения. Предназначено для студентов биологического факультета БГУ специальности 1-31 01 01 ―Биология‖ направление 1-31 01 01-03 Биотехнология. УДК 631.147(075.8) ББК 28.072 я 7 © БГУ, 2014 2 Оглавление Предисловие ГЛАВА 1. Биоткхнологические процессы в народном хозяйстве. 1.1. Общая характеристика биотехнологических процессов 1.2. Классификация продуктов биотехнологических производств 1.3 Общая схема биотехнологического производства 1.4. Лабораторный регламент ГЛАВА 2 Основные принципы получения белков 2.1 Использование микроорганизмов для производства белка 2.1.1 Использование дрожжей для производства белка 2.1.2 Использование бактерий 2.1.3 Использование водорослей 2.1.4 Использование грибов 2.2. Методы очистки белков 2.2.1 Приготовление экстракта 2.2.2. Разрушение клеток и экстракция 2.2.3. Оптимизация и осветление экстракта 2.2.4. Методы, используемые при очистке белков и ферментов, ассоциированных с частицами ГЛАВА 3. Производство аминокислот 3.1. Биотехнология синтеза аминокислот и их очистка. 3.2. Получение аминокислот с помощью иммобилизованных клеток и ферментов. 3.3. Получение оптических изомеров аминокислотпутем применения ацилаз микроорганизмов. ГЛАВА 4. Производство витаминов 4.1. Общая характеристика витаминов 4.2. Получение водорастворимых витаминов 4.3. Получение жирорастворимых витаминов ГЛАВА 5. Производство органических кислот ГЛАВА 6. Источники получения липидов и основные способы их выделения. 6.1. Промышленное получение липидов. 6.2. Практическое использование липидов. ГЛАВА 7. Получение нуклеотидов и нуклеиновых кислот. ГЛАВА 8. Получение сахаров и полисахаридов. 8.1. Полисахариды цитоплазмы и мембранных структур. 8.2. Полисахариды клеточных стенок. 8.3. Внеклеточные полисахариды. 8.4. Использование полисахаридов. 8.5. Промышленное получение полисахаридов 5 7 7 11 12 13 15 15 17 18 18 19 20 20 23 29 33 36 39 42 43 44 44 46 52 55 67 68 70 71 75 76 77 80 83 3 микроорганизмов. ГЛАВА 9. Получение антибиотиков 9.1. Принципы получения антибиотиков. 9.2. Применение антибиотиков. ГЛАВА 10. Получение меланинов. ГЛАВА 11. Основные принципы получения алкалоидов. ГЛАВА 12. Получение продуктов брожения 12.1.Молочнокислое брожение 12.2. Получение продуктов пропионовокислого брожения (витамин В12) 12.3. Ацетонобутиловое брожение 12.4. Спиртовое брожение. ГЛАВА 13. Выделение и очистка гормональных препаратов 13.1. Инсулин. 13.2. Препараты щитовидной железы 13.3. Препараты гипофиза 13.4. Гормон роста 13.5. Получение промышленно важных стероидов ГЛАВА 14. Биотехнология получения ферментов 14.1 Область применения и источники ферментов 14.2 Выбор штамма и условий культивирования 14.3 Технология культивирования микроорганизмов– продуцентов ферментов и выделение ферментов 14.4. Выделение и стабилизация ферментов. 14. 5.Применение ферментов микроорганизмов ГЛАВА 15. Получение пищквых добавок. ГЛАВА 16. Получение лигнина ГЛАВА 17. Получение антисывороток 17.1. Иммуногенность антигена 17.2. Получение иммунных антисывороток 17.3. Выделение и очистка антител. ГЛАВА 18 Использование продуктов биотехнологии для защиты окружающей среды. 18.1 Получение биогаза 18.2 Производство биоэтанола 18.3 Очистка сточных вод Литература 87 91 93 114 117 120 125 125 128 137 141 153 154 158 159 160 162 163 166 167 168 174 192 198 209 213 213 216 219 225 225 228 229 232 4 Прндисловие. Биотехнология как наука является важнейшим разделом современной биологии, которая, как и физика, стала в конце XX в. одним из ведущих приоритетов в мировой науке и экономике. Всплеск исследований по биотехнологии в мировой науке произошел в 80-х годах, когда новые методологические и методические подходы обеспечили переход к эффективному их использованию в науке и практике и возникла реальная возможность извлечь из этого максимальный экономический эффект. По прогнозам, уже в начале 21 века биотехнологические товары будут составлять четверть всей мировой продукции. В нашей стране значительное расширение научно-исследовательских работ и внедрение их результатов в производство также было достигнуто в 80-е годы. В этот период в стране была разработана и активно осуществлялась первая общенациональная программа по биотехнологии, были созданы межведомственные биотехнологические центры, подготовлены квалифицированные кадры специалистов-биотехнологов, организованы биотехнологические лаборатории и кафедры в научноисследовательских учреждениях и вузах. Однако в дальнейшем внимание к проблемам биотехнологии в стране ослабло, а их финансирование сокращено. В результате развитие биотехнологических исследований и их практическое использование в Беларуси замедлилось, что привело к отставанию от мирового уровня, особенно в области генетической инженерии. Что касается более современных биотехнологических процессов, то они основаны на методах рекомбинантных ДНК, а также на использовании иммобилизованных ферментов, клеток или клеточных органелл. Современная биотехнология — это наука о генно-инженерных и клеточных методах и технологиях создания и использования генетически трансформированных биологических объектов для интенсификации производства или получения новых видов продуктов различного назначения. Среди основных направлений современной биотехнологии особое место занимает раздел, связанный с практическим применением разнообразных продуктов биотехнологических процессов в отрослях народного хозяйства, медицине и ветиринарии, косметической и пищевой промышленности, защите окружающей среды и др. В настоящем курсе лекций рассматривеются общие принципы осуществления биотехнологических процессов, происходит знакомство с основными объектами биотехнологических процессов и сферами применения продуктов биотехнологии, основных способов промышленного получения очищенных биологически активных субстанций биотехнологического происхождения, рядом крупномасштабных промышленных биотехнологических производств, использующих микроорганизмы. 5 Микробиологическая промышленность в настоящее время использует тысячи штаммов различных микроорганизмов. В большинстве случаев они улучшены путем индуцированного мутагенеза и последующей селекции. Это позволяет вести широкомасштабный синтез различных веществ. Некоторые белки и вторичные метаболиты могут быть получены только путем культивирования клеток эукариот. Растительные клетки могут служить источником ряда соединений - атропин, никотин, алкалоиды, сапонины и др. Клетки животных и человека также продуцируют ряд биологически активным соединений. Например, клетки гипофиза липотропин, стимулятор расщепления жиров, и соматотропин - гормон, регулирующий рост. Созданы перевиваемые культуры клеток животных, продуцирующие моноклональные антитела, широко применяемые для диагностики заболеваний. В биохимии, микробиологии, цитологии несомненный интерес вызывают методы иммобилизации как ферментов, так и целых клеток микроорганизмов, растений и животных. В ветеринарии широко используются такие биотехнологические методы, как культура клеток и зародышей, овогенез in vitro, искусственное оплодотворение. Все это свидетельствует о том, что биотехнология станет источником не только новых продуктов питания и медицинских препаратов, но и получения энергии и новых химических веществ, а также организмов с заданными свойствами. В данном пособии рассматриваются основные принципы получения, очистки биологически активных соединений таких как белки, аминокислоты, витамины, органические кислоты, липиды, нуклеотиды и нуклеиновые кислоты, сахара и полисахариды, антибиотики, алкалоиды, различные продукты брожения, гормональные препараты, ферменты, пищевые добавки, антисыворотки, и др., а также основные направления их практического использования. 6 ГЛАВА 1. Биоткхнологические процессы в народном хозяйстве. 1.1. Общая характеристика биотехнологических процессов Биотехнология – это использование культур клеток бактерий, дрожжей, животных и растений, метаболизм и биосинтетические возможности которых обеспечивают выработку специфических веществ. Или более краткое определение, биотехнология – это производство с помощью объектов живой природы, или технология живого. Биотехнология возникла на стыке нескольких биотехнологических наук, таких как генетика, бактериология, вирусология, молекулярная биология, микробиология, биохимия, растениеводство. Важную роль сыграло открытие способов модификации ДНК и ее переноса из одних организмов в другие. биохимическая технология технология пищевой промышленности химическая технология БИОТЕХНОЛОГИЯ механическая технология электроника микробиология биохимия генетика Рисунок 1 – Связь биотехнологии с другими науками Исторически биотехнология возникла на основе традиционных микробиологических производств, таких как производство хлеба, сыра, вина, пива, молочных продуктов. Эти технологии до сих пор имеют большую значимость и постоянно развиваются. Биотехнологические процессы осуществляются за счет использования бактерий, дрожжей, плесневых грибов, водорослей, культур клеток и тканей растений и животных. В этих процессах используются особенности метаболизма и биосинтетические возможности клеток. Целью процесса может быть наработка клеточной биомассы или продуктов жизнедеятельности клеток – метаболитов. В настоящее время биотехнология включает в себя: промышленную микробиологию, прикладную микробиологию, генетическую инженерию, клеточную инженерию. Основные направления промышленной микробиологии представлены в таблице 1. Потребность в биотехнологии обусловлена дефицитом продовольствия, энергии, минеральных ресурсов и необходимостью улучшения состояния здравоохранения и охраны окружающей среды. 7 Таблица 1 – Основные направления биотехнологии в различных отраслях Отрасль Область применения Сельское Производство белково-витаминных концентратов. хозяйство Селекция, клонирование и генетическая инженерия животных и растений. Использование антибиотиков для лечения животных и птиц. Производство вакцин. Производство биоинсектицидов. Применение гормонов и других стимуляторов роста Производство Производство органических кислот. химических веПолучение витаминов, антибиотиков и др. ществ и Использование ферментов в составе СМС соединений Контроль за Улучшение методов тестирования и мониторинга состоянием загрязнений окружающей среды. окружающей Использование микроорганизмов для переработки среды сельскохозяйственных, бытовых и промышленных отходов Медицина Использование ферментов в диагностике. Использование микроорганизмов при создании и модификации сложных лекарственных средств. Синтез новых антибиотиков, гормонов и интерферонов. Применение в медицинской практике ферментов и штаммов микроорганизмов Энергетика Производство биогаза. Производство этанола МатериалоВыщелачивание руд. ведение Изучение и контроль биоразложения Пищевая Создание новых методов переработки и хранения промышленность пищевых продуктов. Применение пищевых добавок, полученных с помощью микроорганизмов. Использование белка одноклеточных. Применение ферментов. Совершенствование спиртового и молочнокислого брожения Биоиндустрия включает отрасли, в которых биотехнология может заменить широко используемые традиционные методы, и отрасли, в которых она всегда играла ведущую роль (Таблица 1). 8 Транснациональные корпорации инвестируют следующие биотехнологические отрасли: горнодобывающую, нефтехимическую, фармацевтическую. Быстрая отдача происходит в следующих биотехнологических отраслях: 1) совершенствование сбраживания; 2) производство биогаза; 3) производство безопасных и недорогих вакцин; 4) биоэнергетика; 5) улучшение техники компостирования; 6) гидролиз целлюлозы; 7) повышение уровня фиксации азота с помощью симбионтов. Из более чем 100 тыс. известных микроорганизмов в промышленности применяются всего несколько сотен видов, так как промышленный штамм должен отвечать ряду строгих требований: 1) расти на дешевых субстратах; 2) обладать высокой скоростью роста или давать высокий выход продукта за короткое время; 3) проявлять синтетическую активность в сторону желаемого продукта; образование побочных продуктов должно быть низким; 4) быть стабильным в отношении продуктивности и к требованиям условий культивирования; 5) быть устойчивым к фаговым и другим типам инфекций; 6) быть безвредным для людей и окружающей среды; 7) желательны термофильные, ацидофильные (или алкофильные) штаммы, поскольку с ними легче поддерживать стерильность в производстве; 8) интерес представляют анаэробные штаммы, так как аэробные создают трудности при культивировании – требуют аэрирования; 9) образуемый продукт должен иметь экономическую ценность и легко выделяться. На практике применяются штаммы четырех групп микроорганизмов: – дрожжи; – мицелиальные грибы (плесени); – бактерии; – аскомицеты. Термин «дрожжи» в строгом смысле не имеет таксономического значения. Это одноклеточные эукариоты, относящиеся к трем классам: Ascomycetes, Basidiomycetes, Deuteromycetes. К аскомицетам относят, прежде всего, Saccharomyces cerevisiae, определенные штаммы которого используются в пивоварении, виноделии, производстве хлеба, этилового спирта. Аскомицеты Saccharomyces lipolytica деградируют углеводороды нефти и употребляются для получения белковой массы. 9 Дейтеромицет Candida utilis используют как источник белка и витаминов и выращивают на непищевом сырье: сульфитных щелоках, гидролизатах древесины и жидких углеводородах. Дейтеромицет Trichosporon cutaneum окисляет многие органические соединения, в том числе токсичные (например, фенол), и используется при переработке стоков. Мицелиальные грибы используют: – в получении органических кислот: лимонной (Aspergillus niger), глюконовой (Aspergillus niger), итаконовой (Aspergillus terreus), фурмаровой (Rhizopus chrysogenum); – в получении антибиотиков (пенициллина и цефаллоспорина); – в производстве специальных видов сыров: камамбера (Penicillium camamberti), рокфора (Penicillium roqueforti); – вызывают гидролиз в твѐрдых средах: в рисовом крахмале при получении сакэ, в соевых бобах при получении темпеха, мисо. Полезные бактерии относятся к эубактериям. Промышленное применение с давних времен имеют молочнокислые бактерии родов Lactobacillus, Leuconostoc, Lactococcus. Уксуснокисные бактерии родов Acetobater, Gluconobacter превращают этанол в уксусную кислоту. Бактерии рода Bacillus используются для производства вредных для насекомых токсинов, а также для синтеза антибиотиков и аминокислот. Бактерии рода Corynebacterium используются для производства аминокислот. Из актиномицетов наиболее представительными являются рода Streptomyces и Micromonospora, используемые в качестве продуцентов антибиотиков. При росте на твердых средах актиномицеты образуют тонкий мицелий с воздушными гифами, которые дифференцируются в цепочки конидиоспор. В настоящее время с помощью микроорганизмов синтезируют следующие соединения: – алкалоиды, – аминокислоты, – антибиотики, – антиметаболиты, – антиоксиданты, – белки, – витамины, – гербициды, – ингибиторы ферментов, – инсектициды, – ионофоры, – коферменты, – липиды, 10 – нуклеиновые кислоты, – нуклеотиды и нуклеозиды, – окислители, – органические кислоты, – пигменты, – поверхностно-активные вещества, – полисахариды, – противоглистные агенты, – противоопухолевые агенты, – растворители, – ростовые гормоны растений, – сахара, – стерины и превращенные вещества, – факторы транспорта железа, – фармакологические вещества, – ферменты, – эмульгаторы. 1.2. Классификация продуктов биотехнологических производств Биотехнологические производства основаны на использовании жизнедеятельности микроорганизмов. Чтобы управлять микробиологическим процессом, необходимо знать физиологию применяемых культур микроорганизмов. Это позволит контролировать процессы, протекающие в клетке, условия культивирования и влияние основных факторов окружающей среды на направленный биосинтез. Продуктами биотехнологических производств являются природные макромолекулы – белки, ферменты, полисахариды, полиэфиры, выделенные из клеток микроорганизмов, тканей и органов растений и животных. Рисунок 2 – Динамика изменения биомассы и образования первичных (А) и вторичных (Б) метаболитов в процессе роста организма, 11 где 1 – биомасса; 2 – продукт По отношению к процессам роста низкомолекулярные продукты метаболизма живых клеток делятся на первичные и вторичные метаболиты. Первичные метаболиты – это низкомолекулярные соединения (молекулярная масса менее 1500 Да), необходимые для роста микроорганизмов. Одни из них являются строительными блоками макромолекул, другие участвуют в синтезе коферментов. Среди наиболее важных для промышленности метаболитов можно выделить аминокислоты, органические кислоты, пуриновые и пиримидиновые нуклеотиды, растворители и витамины. Вторичные метаболиты – это низкомолекулярные соединения, не требующиеся для роста в чистой культуре. Ко вторичным метаболитам относятся антибиотики, алкалоиды, гормоны роста растений и токсины. 1.3 Общая схема биотехнологического производства Процессы биотехнологических производств разнообразны, но все они имеют пять основных стадий: 1) приготовление питательной среды; 2) подготовка посевного материала; 3) культивирование микроорганизмов; 4) выделение целевого продукта; 5) очистка целевого продукта. Принципиальная биотехнологическая схема производства продуктов микробного синтеза показана на рисунке 3. Приготовление питательных сред. Среда должна отвечать двум основным требованиям. Во-первых, она должна быть полноценной для питания и недорогой. Углерод и азот в усвояемой форме требуются для биосинтеза белка; фосфор необходим для синтеза ДНК и АТФ; микроэлементы требуются для образования ферментов, также для нормальной жизнедеятельности нужны факторы роста и витамины. Вовторых, среда должна быть стерильной, что достигается температурной, ультрафиолетовой, ультразвуковой и другими видами обработки. Культивирование (ферментация) представляет собой совокупность последовательных операций от внесения в заранее приготовленную питательную среду посевного материала до завершения процессов роста и биосинтеза вследствие исчерпания питательных веществ среды. Существует два основных типа ферментаций: получение биомассы микроорганизмов и получение метаболитов. 12 пробирка получение культуры в микробиологической лаборатории выращивание микроорганизмов в малом инокуляторе выращивание микроорганизмов в большом инокуляторе Получение посевного материала (инокулята) проводится по следующей схеме: Качество полученного посевного материала контролируют путем микроскопирования. 1.4. Лабораторный регламент При изучении продуцента и образуемого им биологически активного вещества в лабораторных условиях подготавливается так называемый лабораторный регламент. Лабораторный регламент - это технологический документ, завершающий лабораторные исследования по выработке метода получения антибиотика. Он служит основой для разработки промышленного регламента. Задачей лабораторного регламента является определение оптимального метода промышленного производства антибиотического вещества. В соответствии с «Отраслевым стандартом промышленного регламента производства химико-фармацевтического препарата» ОСТ 59.01.002.40-85 лабораторный регламент получения антибиотика должен включать следующие разделы. 1. Характеристика антибиотика. Сюда входит название антибиотика, основное назначение, краткое описание свойств препарата, описание организма, образующего антибиотик, методы определения биологической активности, условия хранения. 2. Технологическая схема производства. Отражает последовательность работ по производству антибиотика с подразделением на стадии. Технологическая схема служит основой будущей технологии промышленного получения препарата. 3. Сырье и материалы. Сообщаются требования, предъявляемые к качеству сырья и материалам, которые используются при получении антибиотика с целью его максимальных выходов и обеспечения повторяемости результатов. При этом необходимо ориентироваться на сырье и материалы, выпускаемые отечественной промышленностью. 4. Аппаратурная схема производства антибиотика. Представляют схему процесса получения антибиотика с указанием аппаратов и приборов, 13 их конструкции, размера и других характеристик, которые могут иметь значение при производстве антибиотика. 5. Изложение технологического процесса. Отражает описание процесса получения антибиотика на основе завершенных экспериментальных результатов, выполненных в лабораторных условиях. Процесс включается в регламент в том случае, если удается получить воспроизводимые результаты как по качеству антибиотика, так и по его выходам. Технологический процесс описывают по стадиям. Подробно указываются объемы, концентрации веществ, входящих в среду, рН среды, степень аэрации, растворители, пеногасители, условия перемешивания, продолжительность процесса развития продуцента, температура и другие показатели. 6. Отходы производства, технологические и вентиляционные выбросы в атмосферу, их использование и обезвреживание. Приводится перечень возможных отходов и выбросов в атмосферу, наличие в отходах ценных веществ и рекомендации к их использованию, наличие вредных с точки зрения загрязнения окружающей среды веществ и способы их обезвреживания. 7. Контроль производства. Указываются особые требования к оборудованию (герметичность ферментера и всех коммуникаций, исправность и надежность работы мешалки и др.). Анализ качества сырья, соответствующего определенным стандартам. Режимы стерилизации сред и отдельных веществ, воздуха. Методы анализа за ходом процесса биосинтеза антибиотика и готовой продукции. 8. Техника безопасности, пожарная безопасность и производственная санитария. Приводится перечень веществ, способных воспламеняться и взрываться. Все вещества, применяемые в процессе получения антибиотика, должны быть изучены с позиций техники безопасности, пожарной опасности и производственной санитарии. 9. Перечень производственных инструкций. Приводятся все инструкции, которые должны быть разработаны на основе лабораторного регламента. 10. Технико-экономические нормативы. Выходы конечного продукта и промежуточных продуктов; удельные нормы расхода сырья и материалов, удельные нормы расхода технологических энергозатрат (пара, воды, электроэнергии, сжатого воздуха). 11. «Информационные материалы». В этом разделе должны быть указаны биологические и физико-химические свойства вещества, степень очистки. Фармакологические свойства (преимущества и особенности), сравнение с показателями идентичных зарубежных препаратов, сведения о патентной чистоте антибиотика и принятого метода его получения с перечислением охраняющих авторских свидетельств (патентов), сведения о вредности применяемых при получении препарата веществ и мерах предосторожности при работе с ними. 14 ГЛАВА 2 Основные принципы получения белков 2.1 Использование микроорганизмов для производства белка В соответствии с нормами питания человек должен ежедневно получать с пищей от 60 до 120 г полноценного белка. Для поддержания жизненных функций организма, построения клеток и тканей необходим постоянный синтез различных белковых соединений. Если растения и большинство микроорганизмов способны синтезировать все аминокислоты из углекислого газа, воды, аммиака и минеральных солей, то человек и животные не могут синтезировать некоторые аминокислоты (валин, лейцин, изолейцин, лизин, метионин, треонин, триптофан и фенилаланин). Эти аминокислоты называются незаменимыми. Они должны поступать с пищей. Их недостаток вызывает тяжелые заболевания человека и понижает продуктивность сельскохозяйственных животных. В настоящее время мировой дефицит белка составляет около 15 млн.т. Наиболее перспективен микробиологический синтез. Если для крупного рогатого скота требуется 2 месяца для удвоения белковой массы, для свиней – 1,5 месяца, для цыплят – 1 месяц, то для бактерий и дрожжей – от 1 до 6 часов. Мировое производство пищевых белковых продуктов за счет микробного синтеза составляет более 15 тыс. т в год. Рассмотрим пример: время удвоения кишечной палочки составляет 20 мин, тогда через 20 мин из одной клетки образуется две дочерних, через 40 мин – четыре «внучки», через 60 мин – восемь «правнучек», через 80 мин – 16 «праправнучек». Через 10 ч 40 мин из одной бактерии будет образовано свыше 6 млрд. бактерий, что соответствует населению Земли, а через 44 ч из одной бактерии массой 1•10-12 г образуется биомасса в количестве 6•1024 г, что соответствует массе Земли. Использование различных микроорганизмов в качестве источников белка и витаминов обусловлено следующими факторами: а) возможностью использования для культивирования микроорганизмов разнообразных химических соединений, в том числе отходов производств; б) относительно несложной технологией производства микроорганизмов, которое может осуществляться круглогодично; возможностью его автоматизации; в) высоким содержанием белка (до 60-70%) и витаминов, а также углеводов, липидов в микробиальных препаратах; г) повышенным содержанием незаменимых аминокислот по сравнению с растительными белками; д) возможностью направленного генетического влияния на химический состав микроорганизмов в целях совершенствования белковой и витаминной ценности продукта. 15 Для промышленного производства пищевых продуктов на основе микроорганизмов необходимы тщательные медико-биологические исследования. Такие продукты должны пройти всестороннюю проверку для выявления канцерогенного, мутагенного, эмбриотропного действия на организм человека и животных. Токсикологические исследования, усвояемость продуктов микробного синтеза – основные критерии целесообразности технологии их производства. Для получения белков используются дрожжи, бактерии, водоросли и мицелиальные грибы. Преимуществом дрожжей перед другими микроорганизмами является их технологичность: устойчивость к инфекциям, легкость отделения от среды благодаря крупным размерам клеток. Они способны накапливать до 60 % белка, богатого лизином, треонином, валином и лейцином (этих аминокислот мало в растительных кормах). Массовая доля нуклеиновых кислот составляет до 10 %, что вредно действует на организм. В результате их гидролиза образуется много пуриновых оснований, превращающихся затем в мочевую кислоту и ее соли, которые являются причиной мочекаменной болезни, остеохондроза и других заболеваний. Оптимальная норма добавок дрожжевой массы в корм сельскохозяйственных животных составляет от 5 до 10 % от сухих веществ. Дрожжи применяются для пищевых и кормовых целей. Преимуществами бактерий является высокая скорость роста и способность синтезировать до 80 % белка. Полученный белок содержит много дефицитных аминокислот: метионина и цистеина. Недостатками являются маленькие размеры клеток и низкая их концентрация в культуральной среде, что затрудняет процесс выделения. В некоторых бактериальных липидах могут содержаться токсины. Массовая доля нуклеиновых кислот до 16 %. Используются только для кормовых целей. Преимуществами водорослей являются высокое содержание полноценного по аминокислотному составу белка, накапливающегося в количестве 65 %, легкое выделение водорослей из культуральной среды, низкое содержание нуклеиновых кислот – 4% (для сравнения – у высших растений 1-2 %). Водоросли используются для пищевых и кормовых целей. Мицелиальные грибы традиционно используются в качестве пищевого продукта в странах Африки, в Индии, Индонезии, Китае и др. Накапливают до 50% белка, по аминокислотному составу приближающегося к белку животного происхождения, богаты витаминами группы В. Клеточные стенки тонкие и легко перевариваются в желудочно-кишечном тракте животных. Массовая доля нуклеиновых кислот составляет 2,5%. С 1985 г микробиальный белок используется в пищевой промышленности для изготовления различных продуктов и полуфабрикатов. В производстве пищевых продуктов рассматриваются три основные формы использования микробного белка: 1) цельная масса (без разрушения клеточных стенок); 16 2) частично очищенная биомасса (предусматривается разрушение клеточных стенок и удаление нежелательных компонентов); 3) выделенные из биомассы белки (изоляты). ВОЗ (Всемирная организация здравоохранения) сделала заключение, что белок микроорганизмов можно использовать в продуктах питания, но допустимое количество нуклеиновых кислот, вводимых вместе с белком в диету взрослого человека не должно превышать 2 г в сутки. Введение микробиального белка не вызывает отрицательных последствий, но встречается проявление аллергических реакций, желудочные заболевания и т.д. 2.1.1 Использование дрожжей для производства белка В конце 19 века в Германии была разработана технология производства хлебопекарных дрожжей, во время первой мировой войны дрожжи стали использоваться в качестве пищевой добавки в производстве супов и колбас, а также начала развиваться технология производства кормовых дрожжей. До сих пор культивирование пивных дрожжей Saccharomyces serevisiae (carlsbergensis) остается важным резервом пищевого белка и витаминов. Организм человека усваивает свыше 90% всех питательных веществ, содержащихся в них. В составе этих дрожжей обнаружено 14 витаминов, особенно они богаты витаминами группы В. При переработке биомассы в пищевой белок ее тщательно очищают. Сначала разрушают стенки дрожжевых клеток путем механической, щелочной, кислотной или ферментативной обработки с последующей экстракцией гомогенной дрожжевой массы подходящим органическим растворителем. Затем щелочным раствором растворяют белки, и белковый раствор отделяют от клеточной массы диализом. Очищенные от низкомолекулярных примесей белки осаждают и используют в качестве белковых добавок в различные пищевые продукты – сосиски, колбасы, паштеты, мясные начинки. Также сухой белок можно текстурировать. Некоторые дрожжевые клетки (родов Candida, Rhodotorula, Torulopsis, Trichosporon) в качестве источника углерода для роста способны использовать неразветвленные углеводороды с числом от 10 до 30 углеродных атомов в молекуле. В основном они представлены жидкими фракциями углеводородов нефти с температурой кипения от 200 до 320 ºС. Первоначально проект возник из необходимости утилизировать парафины, остающиеся в количестве от 10 до 15 % после очистки газойля. В питательную среду добавляют макро- и микроэлементы, витамины и аминокислоты. В России завод по производству кормовых дрожжей на парафинах нефти был построен в 1971г. (его продуктивность составила около 1 млн. т в год). Высушенная белковая масса гранулируется и используется как белково-витаминный концентрат в кормопроизводстве. Хорошим субстратом для выращивания кормовых дрожжей родов Torula, Kluyveromyces является молочная сыворотка. В 1 т молочной 17 сыворотки содержится около 10 кг белка и 50 кг лактозы. Методом ультрафильтрации белки отделяют, а раствор лактозы используют для культивирования дрожжей. В качестве источников углерода дрожжевые клетки могут использовать и низшие спирты – метанол и этанол, получаемые из природного газа или растительных отходов. При этом дрожжевая масса содержит больше белков (56-62 % от сухой массы) и меньше вредных примесей (производных бензола, D-аминокислот, аномальных липидов, токсинов, канцерогенов), чем кормовые дрожжи, выращенные на парафинах нефти. Для выращивания дрожжей на гидролизатах растительного сырья используются Candida arborea и Candida utilis, они применяются для пищевых целей и используются в качестве белковых добавок к различным продуктам. Например, в США на основе Candida utilis производят торутеин, который добавляют в продукты питания, после чего они считаются диетическими с высоким содержанием протеина. 2.1.2 Использование бактерий Известно более 30 видов бактерий, которые могут быть применены в качестве источников полноценного кормового белка. Источником углерода при культивировании бактерий могут служить природный и попутный газы, водород, а также спирты – метанол, этанол, пропанол. Чаще всего на газовых питательных средах выращиваются бактерии рода Methylococcus, способные утилизировать от 85 до 90 % метана в специальных ферментерах. Однако производство кормового белка на газовых средах достаточно дорого. Более широко применяется технология выращивания бактерий на метаноле, который легко получают путем окисления метана. Чаще всего используют бактерии родов Methylomonas, Methylophilus, Pseudomonas. Концерном ICI выпускается кормовой препарат прутин. В России – меприн. В этом препарате содержится до 74 % белков (от сухого вещества), до 5 % липидов, 10 % минеральных веществ, от 10до 13 % нуклеиновых кислот. К числу бактерий с высокой интенсивностью синтеза белков следует отнести водородокисляющие бактерии, способные накапливать до 80 % белка (в расчете на сухое вещество). Для их культивирования в газовой среде должно содержаться от 70 до 80 % водорода, от 20 до 30 % кислорода, от 3 до 5 % углекислого газа. Производство может быть организовано вблизи химических предприятий. 2.1.3 Использование водорослей Уже в 1521 г, после завоевания Мексики, испанец Бернал Диаз дель Кастильо сообщал, что ацтеки употребляют в пищу диковинные пирожки, похожие на сыр. На озере Чад (Африка) туземцы племени канембу употребляют в пищу клубки сине-зеленых водорослей. 18 Для получения кормового белка используют одноклеточные водоросли Chlorella и Scenedesmus, сине-зеленые водоросли (цианобактерии) Spirulina (Spirullina platensis, Spirullina getleri), способные синтезировать белки из углекислого газа, воды и минеральных веществ за счет энергии солнечного света. Для своего развития водоросли нуждаются в определенных режимах освещения и температуры и в больших объемах воды. Обычно их выращивают в естественных условиях южных регионов и бассейнах открытого типа (Мексика, Чад, Нигерия, Камерун, Италия, Япония, Израиль, Узбекистан и др.). Водоросли хлорелла и сценедесмус нуждаются в нейтральной среде, их клетки имеют достаточно плотную целлюлозную стенку, в результате чего хуже перевариваются в организме животных, чем спирулина, которую выращивают в щелочных озерах. С 1 га водной поверхности можно получать до 70 т сухой биомассы в год, что превышает выход биомассы при возделывании пшеницы, риса, сои, кукурузы. Содержание белков в клетках хлореллы и сценедесмуса составляет около 55 % (в пересчете на сухое вещество), а в клетках спирулины – 65 %. Водоросли хорошо сбалансированы по аминокислотному составу (кроме метиотина), в них содержится довольно много полиненасыщенных жирных кислот и β-каротина. При скармливании спирулины животным не обнаружено аномалий и патологических эффектов, обеспечивается норма скорости роста. Белковая масса из клеток водорослей поступает в продажу в виде суспензии, сухого порошка или пастообразного препарата. Процесс отделения клеток водорослей от массы воды – наиболее трудоемкая стадия. 2.1.4 Использование грибов Грибы рода Rhizopus sp. используют для твердофазной ферментизации соевых бобов. Через три дня мицелий гриба разрастается и связывает бобы в корж, содержащий до 40 % белка. В Индонезии такой корж жарят и используют в супах как заменитель мяса. Такой продукт называют «темпех». Подобным образом в странах Африки и Востока ферментизируют различные зернобобовые культуры. Преимуществом твердофазной ферментизации является снижение энергетических затрат, недостатком – низкая продуктивность. Различные мицелиальные грибы выращивают на крахмальных (зерновых) материалах, кожуре цитрусовых, соломе, отрубях, шелухе. Используют их либо для обогащения белками кормов, либо из полученной биомассы выделяют ферменты. Для получения пищевого продукта микопротеина культивируют гриб Fusarium graminearum. Его выращивают на дешевом глюкозном сиропе, полученном путем гидролиза пшеничного или кукурузного крахмала. Микопротеин хорошо переваривается. Ему придают консистенцию и аромат мяса, ветчины, кур, рыбы. Продукт долго сохраняет аромат и не дает усушки 19 в процессе кулинарной Великобритании. подготовки. Промышленно выпускается в 2.2. Методы очистки белков 2.2.1 Приготовление экстракта Исходный материал. При выборе организмов того или иного вида, используемых для выделения белков и ферментов, руководствуются преимущественно легкостью их выращивания. Из животных наиболее широко используются крыса (особенно при работе с печенью) и кролик (при работе со скелетными мышцами). Если работа проводится с органами, размеры которые слишком малы у лабораторных животных (сердце, мозг, почки, тимус), то используется мясной скот, в основном быки и свиньи. Кроме того, не следует забывать, что человек - это тоже животное, и именно ради него приносят в жертву такое множество лабораторных животных; при этом исходят из предположения, что то, что справедливо для крысы, вероятно, справедливо и для человека. Изучение ферментов тканей человека ограниченно по понятным причинам. С другой стороны, человеческая кровь вполне доступна, и существует множество методов выделения ферментов из эритроцитов и других клеток крови. Плазма крови бедна ферментами, но содержит значительные количества неферментных белков. В последнее время изучение ферментов из различных видов стало более обычным. Частично это обусловлено уяснением того, что многие важные особенности метаболического контроля, локализации ферментов и другие их характеристики значительно различаются даже в пределах типа позвоночных. Белкам и ферментам беспозвоночных уделяют довольно мало внимания в основном из-за того, что в большинстве своем беспозвоночные животные очень малы и для получения необходимого количества исходного материала приходится использовать много особей. Если нужный орган не извлекают из каждой особи (трудная задача при работе с насекомыми), то очистка гомогената, приготовленного из целого тела, осложняется тем, что в нем содержится большое количество пищеварительного сока, с протеолитическими ферментами, которые также высвобождаются в экстракт. Большая часть хорошо известных ферментов беспозвоночных получена из крупных ракообразных, таких, как речной рак и краб. Долгое время биохимия растений была очень слабо связана с биохимией животных, главным образом из-за того, что последняя имеет медицинскую направленность, а также потому, что при работе с растениями перед энзимологом встают определенные трудности. Качественные различия, связанные с сезонностью, могут быть преодолены, если выращивать растения в специальных камерах; в противном случае приходится мириться вариабельностью материала, выращенного под открытым небом. В силу очевидных причин из огромного множества видов для изучения используются в основном те, которые имеют экономическое значение, но, с 20 точки зрения биохимика, это не всегда наилучший выбор. Тем не менее работают преимущественно на одном или двух видах растений вследствие того, что из листьев можно без особого труда приготовить экстракты, них легко выделить хлоропласты. Наиболее часто используются шпинат Spinacia oleracea и в какой-то мере родственная ему свекла Beta vulgaris, которые хорошо растут в различных климатических условиях. Растительные клетки имеют множество компартментов (отсеков); в большинстве случаев основной объем клетки занимает вакуоль, заполненная весьма кислыми растворами, протеазами и целым рядом других, вредных для цитоплазмы веществ. Значительная часть цитоплазмы приходится на хлоропласты, гранулы крахмала и другие органеллы; в клетке имеется также объемистая клеточная стенка (целлюлоза). Цитоплазма (которая вместе с хлоропластами содержит большую часть ферментов) часто может составлять не более 1-2% всего объема растительной клетки. Следовательно, растительные экстракты содержат очень мало белка, даже если при их приготовлении добавляют минимальное количество жидкости. При работе с микроорганизмами возникают другие проблемы. Животный материал можно получать на местной скотобойне или из институтского вивария, растительное сырье можно приобрести на рынке, а микроорганизмы (за исключением дрожжей) необходимо специально выращивать в определенных условиях и в довольно больших количествах. Для каждой из таких групп микроорганизмов, как водоросли, грибы, дрожжи и бактерии, требуются особые условия выращивания, сбора клеток и экстрагирования. Особой проблемой, требующей тщательного изучения, является значительное изменение ферментного состава в различные фазы роста клеток. Желательно производить сбор бактериальных клеток и других одноклеточных микроорганизмов в течение логарифмической фазы роста, хотя активность изучаемого фермента может и не быть максимальной в этот период. Следует проводить предварительные исследования организма с целью определения физиологического состояния, при котором содержание нужного фермента наибольшее. Saccharomyces cerevisiae, пекарские дрожжи, доступны в больших количествах, так что если их можно использовать, то проблема сырья решена. Прессованные дрожжи, поступающие в продажу, в конце фазы роста состоят практически на 100% из дрожжевых клеток. Выращивание дрожжей производят на различных питательных веществах. Основным источником углерода для них служит меласса или какой-либо другой источник сахарозы. При 0°С дрожжи остаются жизнеспособными (и, следовательно, белки сохраняют нативность) в течение не скольких недель, а при хранении в замороженном виде их можно использовать и через несколько месяцев. В хлебопекарной промышленности и у биохимиков большой популярностью пользуются сухие дрожжи, жизнеспособность которых восстанавливается при добавлении к ним воды. Они служат очень удобным источником многих ферментов, хотя в процессе сушки активность некоторых, наиболее 21 чувствительных, ферментов может несколько снижаться. При хранении в вакууме или в атмосфере азота при 0-4 °С их можно использовать даже через несколько лет. Очень важно убедиться в том, что необходимый белок или фермент, присутствующий в исходном материале, экстрагируется из него полностью, для того чтобы можно было достоверно определить его содержание в ткани. При неполном разрушении клеток можно недооценить данный объект как источник ферментов из-за того, что значительная часть ферментативной активности не выходит в раствор, а удаляется вместе с осадком во время получения экстракта. Если фермент выделяют из бактериального материала, то экспериментатор должен учитывать возможность отбора штаммов с аномально высоким уровнем ферментативной активности. Это можно сделать либо путем обычного отбора мутантов, либо используя более сложную технику клонирования, посредством которой множественные копии гена фермента включаются в бактериальный геном с помощью плазмид. Экспрессия гена может быть еще более усилена за счет его слияния с сильным промотором. Эта работа занимает много времени, и выполнить ее может только опытный специалист; не стоит даже пытаться применять эти методы, если на выделение фермента отведено мало времени. Свежесть сырья и его хранение. Обычно чем быстрее используется сырье, тем лучше, т. е. препарат получается более соответствующим нормальному физиологическому состоянию, так как, если ткань некоторое время была «мертва», начинаются естественные процессы разложения. Однако бывают случаи, когда абсолютная свежесть может быть помехой. Например, в скелетных мышцах содержится большое количество АТР. В присутствии фосфокреатина и гликогена высокая концентрация АТР сохраняется в течение нескольких часов и после смерти. Структурные белки миозин и актин в норме нерастворимы при ионной силе ниже 0,25, но в присутствии АТР разорванные миофибриллы набухают, и часть белков переходит в раствор. Даже при физиологической ионной силе (~0,16) некоторое количество миозина может перейти в растворимое состояние. Впоследствии это может сказаться на качестве ферментных препаратов, так как миозин осаждается и смешивается с фракциями, в которых он не должен присутствовать. Примером особого рода служит получение препарата АМРдезаминазы. При экстракции совсем свежих мышц буфером с рекомендуемой ионной силой 0,26 в раствор переходит много актина и миозина, что мешает следующему этапу - адсорбции на фосфоцеллюлозе. Препарат лучшего качества можно получить, если мышцам сначала дать возможность перейти в состояние окоченения, что сопровождается потерей АТР. Однако это редко встречающийся случай. Обычно все же следует работать на свежем материале. Возможность получить свежий материал не всегда совпадает с возможностью использовать его. Как полученный материал, так и экстракт из него следует хранить замороженными. В процессе замораживания 22 происходит множество событий. Прежде всего, замерзает свободная вода, и образуются кристаллы льда. Кристаллы льда разрушают мембраны и органеллы, но не повреждают непосредственно белки и ферменты. По мере приближения температуры к точке эвтектики для различных солей, присутствующих в растворе, эти соли кристаллизуются. Наименее растворимая соль из пары, входящей в состав буфера, первой вы падает из раствора, вследствие чего значение рН может значительно измениться еще до того, как раствор полностью замерзает. Если хранение происходит при температуре от -15 до -25оС (обычная температура для испарителей в бытовых холодильниках и простых морозильников), то значение рН оставшегося незамороженным концентрированного раствора будет отличаться от значения рН исходного раствора. В таком концентрированном белковом растворе протеазы, освободившиеся из лизосом в процессе замораживания, могут начать функционировать (хотя и медленно при такой низкой температуре), и хранение материала в течение нескольких недель может сильно ухудшить его качество. Таким образом, при замораживании следует очень быстро снижать температуру до -25 °С и хранить материал, если возможно, даже при более низкой температуре. Коммерческие морозильники дают температуру до -80 °С. Иногда лучше замораживать экстракт, так как его состав можно менять в целях достижения оптимальных условий хранения. После получения экстракта к нему можно добавить ингибиторы протеолитических ферментов и довести его рН подходящим буфером до значения, при котором биомолекулы стабильны. Кроме того, экстракт можно получить из большого количества материала, так что образцы, взятые впоследствии из морозильника, будут различаться лишь временем хранения, различия в их свойствах в этом случае не будут обусловлены биологическими различиями замороженного материала. 2.2.2. Разрушение клеток и экстракция Большая часть белков и ферментов, изученных в ранний период истории белковой химии, была выделена из внеклеточных жидкостей. Причина этого, заключается не только в легкости получения материала, но и в том, что внеклеточные белки в основном более стабильны часто благодаря наличию в них дисульфидных мостиков, а также небольшим разменам молекул. Первоначально исследования структуры белков, естественно, сосредоточивались на этих небольших белках. Так, лизоцим, рибонуклеаза и химотрипсин были самыми первыми детально изученными белками, и все они получены из внеклеточных источников. Однако большинство ферментов локализовано внутри клеток. Часто такие белки менее стабильны - у них обычно отсутствуют дисульфидные связи вследствие того, что внутриклеточная среда обладает восстанавливающими свойствами. Цель данного раздела - дать описание методов разрушения клеток и выделения ферментов в водный «экстракт», что является первым этапом очистки 23 ферментов. Существует множество методов разрушения клеток, что обусловлено разнообразием типов клеток. Большинство клеток имеют присущие только им особенности, которые следует учитывать при их разрушении. Животные ткани широко варьируют по своей прочности: наряду с легко разрушаемыми эритроцитами используется и жесткий коллагенсодержащий материал, встречающийся в кровеносных сосудах и других тканях, содержащих гладкие мышцы. Растительные клетки обычно труднее разрушаются, чем животные, из-за наличия у них целлюлозной оболочки. Среди бактерий есть и довольно хрупкие организмы, которые могут быть разрушены пищеварительными ферментами или просто под действием осмотического шока, и более устойчивые, с толстой клеточной стенкой; для разрушения последних необходимо сильное механическое воздействие, которое, однако, не должно быть чрезмерным, так как при этом могут инактивироваться лабильные ферменты. В табл. 2.1 приводится перечень используемых методик. Эти методы служат для разрушения клеток и выделения ферментов в раствор. Если фермент локализован в органеллах, метод может быть использован только при условии, что органеллы также разрушаются. С другой стороны, может понадобиться предварительное выделение самих органелл, что позволит избавиться от загрязнения цитоплазматическими фермента ми перед экстракцией фермента из органелл. Для этого требуется менее сильное воздействие. Предварительное растворение поверхностных коллагеновых и целлюлозных структур препаратами гидролитических ферментов позволяет в дальнейшем разрушать клетки более мягкими способами, сохраняя целостность органелл. Примером подобной методики служит получение препаратов митохондрий из таких тканей, как скелетная или сердечная мышцы, под действием протеолитических ферментов. Однако эта методика применима только для получения препаратов в небольших масштабах и больше подходит для метаболических исследований органелл, чем для очистки ферментов. Выход очищенных органелл может быть очень низким, поэтому во многих случаях лучше сначала разрушить всю ткань, а затем уже приступить к выделению нужного фермента из сложной смеси белков в экстракте. Таким образом, ферменты митохондрий и хлоропластов часто получают из тканевого гомогената, а не из выделенных органелл. Наконец, фермент может быть нерастворим в буфере, используемом для экстракции. Таблица 2.1. Способы разрушения клеток Способ Лизис клеток Разрушение Пример Принцип Мягкое воздействие Эритроциты Осмотическое разрушение клеточной мембраны Обработка Разрушение клеточной стенки 24 под действием ферментов Химическая солюбилизация и автолиз Гомогенизация вручную Размельчение (растирание) Лопастной гомогенизатор (типа Уоринга) Растирание с абразивом (например, с песком или окисью алюминия) Пресс Френча Ультразвук Шаровая мельница Гомогенизатор МэнтонаГаулина бактерий лизоцимом Экстракция дрожжей толуолом приводит к осмотическому разрыву клеточной мембраны Клеточная стенка (мембрана) частично растворяется под действием химических веществ, освободившиеся при этом литические ферменты завершают процесс Ткань печени Клетки продавливают через узкий зазор, что приводит к разрушению клеточной мембраны Мышечная ткань и Клетки разрушаются в процессе др. размельчения ткани под действием силы сдвига Воздействие средней силы Мышечная ткань, Происходит механическое разрушение большинство крупных клеток и отделение друг от животных тканей, друга мелких растительные ткани Растительные Разрушение клеточных стенок ткани, бактерии происходит благодаря наличию на частицах абразива микрошероховатостей Сильное воздействие Бактерии, Клетки продавливают через маленькое растительные отверстие под очень большим клетки давлением; они разрушаются под действием силы сдвига Суспензии клеток Ультразвуковые волны создают высокий локальный градиент давления; в результате клетки разрушаются под действием напряжения сдвига и кавитации Суспензии клеток Разрушение клеточной стенки происходит под действием быстрой вибрации стеклянных шариков Суспензии клеток Действует так же, как и пресс Френча, но позволяет обрабатывать большие количества материала После разрушения клеток и осаждения нерастворимого материала центрифугированием получают «экстракт». Смесь до центрифугирования 25 обычно называют гомогенатом. После центрифугирования в жидкой фазе должна находиться по возможности большая часть фермента, присутствовавшего в исходном материале. Часть жидкости удерживается осадком, и общие потери будут пропорциональны отношению объема осадка к объему жидкой фазы. При получении экстракта потери за счет удерживания жидкости осадком не должны составлять более 20%, если только сырье не очень легко доступно, и высокая концентрация фермента при малом объеме экстракта не является более важным условием, чем полное выделение фермента из гомогената. Если объем жидкости, удерживаемой осадком, составляет половину объема осадка, то отсюда можно подсчитать, что для выделения 80% фермента необходимо, чтобы объем надосадочной жидкости в два раза превышал объем осадка. Большинство животных тканей содержит значительное количество нерастворимого клеточного материала, который связывает много воды. При работе с ними объем осадка получается практически равным исходному объему ткани (в некоторой мере это зависит от условий центрифугирования). Таким образом, когда получают гомогенат, скажем, из печени, необходимо добавить, по крайней мере, два объема подходящего экстрагирующего буфера. Чем больше буфера будет добавлено, тем большая часть растворимой фракции экстрагируется, но экстракт будет более разбавленным; его большой объем может создать трудности в дальнейшей работе. Для приготовления гомогенатов из печени, сердца и скелетных мышц обычно добавляют 2,5 объема экстрагирующего буфера. Растительные ткани значительно отличаются от животных. Как указывалось в предыдущем разделе, внутриклеточное содержимое составляет лишь малую часть объема растительной ткани. Наличие больших вакуолей (рассматриваемых здесь как внеклеточные образования) и межклетников служит причиной высвобождения большого количества жидкости при разрушении ткани. В этом случае практически не требуется добавления экстрагирующей жидкости. Объем осадка после центрифугирования может составлять только 20-40% объема исходной растительной ткани. Тем не менее, использование дополнительного экстрагирующего раствора может быть очень важно для контролирования нежелательных процессов, происходящих во время гомогенизации. Такими процессами могут быть подкисление среды или окисление нестойких соединений. Особую проблему представляют растения, содержащие фенольные соединения, которые окисляются в основном под действием эндогенных фенолоксидаз с образованием темных пигментов. Эти пигменты ковалентно соединяются с белками и инактивируют многие ферменты. Пригодны два способа решения этой проблемы. Во-первых, добавление какого-либо тиолового соединения, например β-меркаптоэтанола, сводящего до минимума действие фенолоксидаз. Во-вторых, часто оказывается полезным добавление порошкообразного поливинилпирролидона, адсорбирующего фенольные соединения. По объему осадка, получаемому после центрифугирования, 26 микроорганизмы больше похожи на животные ткани. Дрожжи и сходные с ними грибы с толстой клеточной стенкой дают осадок, объем которого практически равен исходному объему клеточного материала. То же самое характерно и для бактерий. Если в процессе экстракции действуют значительные силы сдвига, в раствор может перейти большое количество материала клеточной стенки. Обломки клеток легко отцентрифугировать. Это особенно относится к крупным обломкам животных и растительных тканей. Однако после центрифугирования, например, при 10 000 g в течение 15 мин в суспензии остаются внутриклеточные частицы. Мутность полученного раствора не имеет существенного значения, так как ее можно устранить на этапах фракционирования. Использование более мощных устройств для разрушения клеток, таких, как шаровая мельница, дает возможность разрушать клеточные стенки до мельчайших частиц относительно низкой плотности, обусловленной тем, что в них обычно содержатся липиды. Такие частицы трудно осаждаются при доступных для больших объемов величинах g, и экстракт может остаться очень мутным. Методы осветления экстракта описаны ниже. Далее в общих чертах описываются процедуры получения экстрактов из различных тканей. Ткани животных. Ткань разрезают на кусочки, удаляют, насколько это воз можно, соединительную ткань и жир. Разрезанную ткань помещают в лопастной гомогенизатор, добавляют 2-3 объема холодного экстрагирующего буфера в расчете на 1 г ткани. Смесь гомогенизируют в течение 30 с; гомогенизацию повторяют, если остались куски неразрушенной ткани. При доведении рН до нужного значения перемешивают гомогенат 10-15 мин, а затем переносят его в центрифужные пробирки. Центрифугируют при 5000— 10 000 g в течение 60 мин (2∙105 - 3∙105 g∙мин). Экстракт сливают через фильтр «Miracloth», марлю или стеклянную вату для удаления частиц жира. Эритроциты. Экстракт можно легко получить из осажденных центрифугированием эритроцитов после промывания их изотоническим раствором NaCl (0,9%, 0,15 М) и повторного центрифугирования. Клетки разрушают осмотическим шоком в воде (2 объема воды на 1 объем отцентрифугированных клеток). Однако около 90% белка, переходящего в раствор, составляет гемоглобин, и если вы занимаетесь очисткой другого белка, то очень полезно применить метод селективного удаления гемоглобина. Для денатурации гемоглобина успешно применяется смесь этанол - хлороформ. Мягкие растительные ткани. К растительной ткани добавляют только 0,5-1 объем холодного буфера, содержащего 20-30 mМ меркаптоэтанола, и гомогенизируют в течение 30 с. Вместо этого можно пропустить через бытовую соковыжималку, предварительно промытую буфером. Гомогенат следует отцентрифугировать как можно скорее, чтобы уменьшить его потемнение в результате окисления (2∙105 - 3∙105 g∙мин). 27 Жидкость осторожно сливают с поверхности осадка. Для адсорбции фенолов полезно добавить порошкообразный поливинилпирролидон. Дрожжи. 1. Полностью разрушить клетки можно с помощью гомогенизатора Мэнтона-Гаулина (Gaulin Corp. Mass.) или мельницы «Vibrogen Cell Mill» (Bühler, Tübingen), используя около двух объемов буфера на 1 г сырой массы. 2. Автолиз толуолом. Применяют различные методы с использованием толуола [8—110]; однако не все они пригодны для работы с коммерческими дрожжами. В основе этих методов лежит обработка дрожжей толуолом, обычно при температуре 35-40 °С. Через 20—30 мин дрожжи «разжижаются» вследствие экстракции компонентов клеточной стенки. После этого к дрожжам добавляют буфер и перемешивают их в течение нескольких часов или оставляют на ночь на холоде. Так как метод автолитический и структуры клеточной стенки разрушаются под действием ферментов, во время обработки может произойти деградация некоторых клеточных ферментов. Вместо толуола можно использовать этилацетат, но он не годится для работы со штаммами дрожжей, имеющими более прочные клеточные стенки. 3. Цитолиз аммиаком. Этот метод более всего подходит для работы с сухими дрожжами; он прост, но некоторые ферменты, не обладающие устойчивостью при рН 10, могут быть полностью утрачены. «Активные сухие» дрожжи размешивают в 0,5 М растворе NH4OH (2 объема раствора на 1 г сухого веса) в течение 16-20 ч при комнатной температуре. Введение небольшого количества толуола (5-10% от общего объема) иногда способствует растворению белков. Затем добавляют 1-2 объема воды и необходимое количество уксусной кислоты, чтобы снизить рН до нужного значения. После этого центрифугированием удаляют обломки разрушенных клеток. Бактерии. Бактериальные клетки можно разрушить ультразвуком, с помощью шаровой мельницы или пресса Френча, хотя не все эти способы удобны для приготовления больших объемов экстракта. В случае небольших объемов клетки можно растирать с окисью алюминия. Грамположительные бактерии обычно чувствительны к лизоциму. Для высвобождения компонентов цитоплазмы бывает достаточно 15-минутного перемешивания суспензии клеток в буфере, содержащем 0,2 мг/мл лизоцима яичного белка, при 37°С. Обработка полученного экстракта дезоксирибонуклеазой I (10 мкг/мл) улучшает его качество, так как при этом вязкость жидкости уменьшается. Грамотрицательные бактерии, если их предварительно не обрабатывать, менее чувствительны к лизоциму. Недавно описан способ комбинированной обработки клеток неионным детергентом, осмотическим шоком и лизоцимом. Модификацию этого метода используют при экстракции грамотрицательных микроорганизмов Zymomonas mobilis и Photobacterium phosphoreum. При этом достигается исчерпывающая экстракция ферментов, хотя полного диспергирования клеточной стенки не 28 происходит. К 10 г сухой клеточной массы, добавляют 0,1 мл 10%-ного (по объему) раствора тритона Х-100, 10 мкл меркаптоэтанола и 2,5 мл глицерина. Клетки диспергируют и энергично размешивают в течение 30 мин. Затем быстро добавляют 30 мл экстрагирующего буфера (например, 20 мМ КРi, рН 7,0+1 мМ ЭДТА), содержащего 0,2 мг/мл лизоцима и 10 мкг/мл дезоксирибонуклеазы, и продолжают перемешивание еще 30 мин. К смеси добавляют 5 мг фенилметилсульфонилфторида, растворенного в 0,5 мл ацетона, и 0,1 мг пепстатина А, после чего суспензию центрифугируют (15000 g в течение 20 мин). Здесь приведено лишь несколько примеров процедур, используемых для получения экстрактов из различных источников. На практике чаще пользуются опубликованной методикой, но следует помнить, что применяемое сырье так же, как и оборудование, может значительно отличаться от описанных в методике, поэтому иногда возникает необходимость приспособить метод к определенным условиям. Таким образом, в основе методов экстрагирования лежит размельчение материала в соответствующем буфере с последующим центрифугированием смеси для удаления нерастворимых остатков. 2.2.3. Оптимизация и осветление экстракта Часто поведение белков при фракционировании зависит не только от их особенностей, но и от состава раствора, в который входят также и другие белки. Неудачи могут быть связаны с неоптимальными условиями гомогенизации (низким давлением в ячейке пресса, износом шариков в шаровой мельнице, затуплением лопастей гомогенизатора), что приводит к неполному разрушению клеток. В результате экстракт может быть более разбавленным, и состав его будет другим, так как при неполном разрушении клеток не все компоненты переходят в раствор. Для оценки данного экстракта важно точно знать содержание фермента в одном грамме исходного материала. Пожалуй, лучше всего сначала получить экстракт из небольшого количества исходного материала, используя большой объем экстрагирующего буфера (скажем, 10 мл/г), и последовательно определять активность фермента через разные промежутки времени. С увеличением времени обработки активность фермента, вероятно, будет постепенно снижаться, так как сама обработка приводит к денатурации белков либо в результате нагревания, либо под действием сил сдвига в ходе измельчения материала. Часто бывает необходимо охлаждать систему в процессе разрушения клеток. Для этого применяют охлажденный буфер, и короткие периоды обработки материала чередуют с периодами охлаждения. Как упоминалось в предыдущем разделе, очень важно учитывать объем экстрагирующего буфера на грамм материала. При использовании больших количеств исходного материала приходится идти на компромисс между желанием добиться максимальной экстракции и стремлением получить 29 минимальный объем экстракта. Для определенных целей экстрагирующий буфер должен содержать вещества типа β-меркаптоэтанола. Для экстракции можно использовать и воду, но она экстрагирует не все белки. Было бы замечательно, если интересующий нас фермент полностью экстрагировался бы водой. Клетки содержат разные соли и много нерастворимого, несущего заряд материала белки, фосфолипиды, нуклеиновые кислоты. Ионная сила в цитоплазме типичной клетки колеблется в пределах 0,15-0,2 М. В этих условиях цитоплазматические белки «растворимы» в том смысле, что они могут перемещаться в клетке. При гомогенизации клеток с 2-3 объемами воды ионная сила гомогената может снизиться до 0,05 и ниже. В этих условиях заряженные частицы в растворе могут действовать в качестве ионообменников и адсорбировать белки, особенно основные. Если гомогенат готовится на разбавленном буфере, то чем больше добавляется буфера, тем большая часть белка переходят в не растворимую фракцию. Не всегда учитываются ионообменные адсорбционные свойства (в отличие от физиологически важных специфических взаимодействий) клеточных компонентов. Для того чтобы быть уверенным, что все растворимое содержимое клетки перешло в экстракт, следует использовать буфер с ионной силой, близкой к физиологической, и, конечно, с соответствующим значением рН. Если при этом экстрагируется слишком много нежелательных компонентов, лучше пойти на компромисс. Обычно используют следующие буферы: 20-50 мМ фосфат, рН 7-7,5, 0,1 М трис-HCl, рН 7,5 и 0,1 М КСl с небольшим содержанием в нем буфера; для выделения органелл применяют изоосмотические буферные смеси, содержащие наряду с солями и буферными мМ), β-меркаптоэтанол ионами сахарозу, маннит или сорбит. Буфер может содержать также ЭДТА (1-5 или цистеин (5-20 мМ) и специфические стабилизирующие агенты для определенных белков, например Zn2+ для цинкосодержащих белков или пиридоксальфосфат для ферментов, использующих его в качестве кофактора. После разрушения клеток в соответствующем буфере желательно проверить рН гомогената. Хотя после гомогенизации действительно свежего материала в буфере гомотенат имеет тот же рН, что и буфер, это значение может снизиться вследствие метаболических процессов, ведущих к закислению среды. Хорошим примером могут служить скелетные мышцы, в которых гликоген быстро превращается в молочную кислоту, и рН гомогената может упасть с 7,0 до 6,0 за 30-60 мин (включая время центрифугирования). В таких случаях перед центрифугированием значение рН можно поднять выше нужной величины (например, 1 М трис-буфером). Кроме того, добавление ингибитора гликолиза может остановить процесс. Для этой цели иногда используют фторид (10-30 мМ). Эти замечания относятся к улучшению условий экстракции всех растворимых компонентов. При экстрагировании какого-то одного фермента или белка лучше использовать другие условия, при которых не происходит 30 полной солюбилизации всех компонентов, тогда как нужный объект экстрагируется полностью. Часто экстракт получается мутным. После центрифугирования на его поверхности могут плавать частицы жира. Жир можно удалить грубой фильтрацией через стекловату или плотную фильтровальную ткань. Нерастворимые частицы суспензии содержат органеллы и фрагменты мембран, которые полностью осаждаются только при 100 000 g - процедура очень неудобная для больших объемов экстракта. Фильтрация в таких случаях обычно бесполезна, так как если фильтр достаточно плотен и задерживает частицы, то он быстро засоряется. Однако, если мутность незначительна, но нужен прозрачный экстракт - на пример для пропускания через адсорбционную колонку, фильтрацию можно провести с помощью целита. Различные типы экстрактов. При работе с некоторыми животными тканями частицы составляют незначительную часть экстракта и ими можно пренебречь. Они агрегируют при первом же фракционировании сульфатом аммония и затем удаляются. С другой стороны, есть животные ткани, содержащие больше жиров и мембранных структур; при экстрагировании таких тканей образуются суспензии. Подкисление среды до рН 6,0-5,0 обычно приводит к агрегации материала, который можно затем отцентрифугировать при относительно низкой скорости. Рибосомы и другие нуклеопротеиды обычно удаляются при подкислении, которое можно рассматривать как одну из форм изоэлектрического осаждения (разд. 3.2); фосфатные группы протонируются и нейтрализуют или по край ней мере снижают заряд частиц суспензии. Этот способ дает очень хорошие результаты при условии, что нужный белок а) не осаждается изоэлектрически при данном значении рН, б) не адсорбируется на образовавшемся осадке или в) остается стабильным, при нефизиологических значениях рН. Экстракт охлаждают и снижают рН соответствующей кислотой (например, 1 М уксусной). После перемешивания в течение 10-20 мин осадок отделяют центрифугированием, а рН надосадочной жидкости доводят, если необходимо, до нужного значения перед тем, как приступить к первому этапу фракционирования. Растительные ткани отличаются, прежде всего, тем, что реакция среды в них более кислая и содержащиеся в них частицы, такие, как фрагменты хлоропластов, агрегируют значительно труднее. К тому же растительные экстракты содержат сравнительно мало корпускулярного материала, если не считать грубых частиц типа зерен крахмала, которые легко осаждаются после приготовления экстракта. При приготовлении экстрактов из микроорганизмов возникает целый ряд проблем. Во-первых, из быстропролиферирующих клеток экстрагируется много нуклеиновых кислот. Во-вторых, в процессе разрушения клеток материал клеточной стенки либо сильно диспергируется, в результате чего раствор становится мутным, либо частично растворяется, так что в растворе наряду с белками и нуклеиновыми кислотами оказывается много 31 камедеподобных полисахаридов. Это создает определенные трудности на первых этапах фракционирования. Экстракты, приготовленные из бактериального материала при помощи пресса Френча, под воздействием ультразвука или при обработке лизоцимом, получаются вязкими из-за присутствия в них ДНК и содержат значительные количества рибосомного материала. Все нуклеиновые кислоты осаждаются в результате их агрегации с макромолекулярными поликатионами. Хорошими реагентами для этого служат протамин лососевых рыб и клупеин молок сельди, поскольку связывание с ДНК - естественная функция этих веществ. С обычно используемым протаминсульфатом следует быть весьма осторожным: он обладает сильными кислотными свойствами, и перед употреблением его необходимо растворить в воде и нейтрализовать. Следует установить, какое количество протаминсульфата потребуется; оно может достигать 5 мг на 1 г обрабатываемого материала. Протаминсульфат осаждает ДНК, рибосомные нуклеиновые кислоты, мРНК, тРНК и другие нуклеиновые кислоты. На образовавшемся осадке могут также адсорбироваться некоторые белки. Адсорбцию на протаминовом осадке используют даже как этап в процессе очистки ферментов. Если применение протамина окажется невозможным или нежелательным, вязкость раствора ДНК можно снизить, используя при получении экстракта ДНКазу. Рибосомы можно осаждать стрептомицином. Хотя стрептомицин хуже осаждает нуклеиновые кислоты, чем протамин, при его использовании теряется меньше белка в результате адсорбции на образующемся осадке. После удаления нуклеиновых кислот в растворе могут остаться капсульные камедеподобные углеводы. Это создает трудности при использовании обычных методов осаждения белков. В присутствии этих веществ невозможно применение сульфата аммония и других методов фракционного осаждения. Лучше полностью осадить белок в надежде, что камедеподобные вещества останутся в растворе. Практически весь белок можно осадить при 80%-ном насыщении сульфатом аммония или 55%-ной концентрации ацетона. В случае использования сульфата аммония для осаждения белка может понадобиться скоростное центрифугирование. Но даже после этого растворенный белковый осадок содержит вещества, снижающие воспроизводимость результатов при работе на колонке или при использовании других методов фракционирования. Недавно найдены два пути преодоления этих трудностей. Первый путь - это «амфифильная» адсорбция на геле агарозы, при которой все белки адсорбируются на гранулах агарозы в растворе сульфата аммония 70-80% -ного насыщения, тогда как небелковый материал вымывается. При экстракции гранул агарозы буфером белок переходит в раствор, который далее фракционируют обычным способом. Второй путь - использование соответствующего метода экстракции. При обработке клеток лизоцимом обычно не образуется слишком много капсульных камедеподобных веществ - лизоцим растворяет их, но не разрушает все бактериальные клетки. Для получения достаточно 32 прозрачного экстракта из бактерий успешным оказалось использование лизоцима вместе с неионным детергентом. После удаления нуклеиновых кислот такой экстракт можно непосредственно использовать для фракционирования. Подобные проблемы возникают при работе не со всеми бактериями. Опубликовано множество работ по выделению ферментов из бактериального материала, в которых успешно проводится обычное фракционирование вслед за обработкой экстракта протамином или стрептомицином. Приготовление экстрактов из дрожжей связано с несколькими проблемами. Главная из них состоит в том, что клеточные стенки у грибов очень толстые и их трудно разрушить. Поэтому для получения экстракта требуются более эффективные методы. Если экстракт нужно получить быстро, единственный надежный способ - это применение вибрационной шаровой мельницы или гомогенизатора Мэнтона - Гаулина. При использовании этих методов образуется большое количество частиц низкой плотности, которые не полностью удаляются центрифугированием. Мутность может снизить эффективность осаждения белка сульфатом аммония. Однако ее можно устранить путем агрегации частиц в органическом растворителе (ацетон, 20-25% по объему при 0°С). В прозрачном растворе остается большинство ферментов, которые осаждаются при повышении концентрации ацетона. Прозрачные экстракты получают также, используя автолиз толуолом, но в этих условиях может происходить протеолитическое расщепление белков. Для выделения очень стабильного фермента фосфоглюкомутазы применяют цитолиз под действием концентрированного аммиака. Эта процедура разработана и широко применяется при работе с «активными сухими» дрожжами. Ее используют для получения прозрачных экстрактов, содержащих большую часть гликолитических ферментов. Некоторые ферменты полностью разрушаются при высоких значениях рН. При использовании этого метода протеолиз не представляет серьезной проблемы, так как активность протеаз достаточно низка при рН 9,5-10. Эффективность экстракции может значительно варьировать в зависимости от партии дрожжей. 2.2.4. Методы, используемые при очистке белков и ферментов, ассоциированных с частицами Ряд специфических методов широко используются для выделения белков и ферментов, структурно связанных с нерастворимыми компонентами клетки, такими, как митохондрии, хлоропласты, плазмалемма, эндоплазматический ретикулум и ядерные мембраны. Эти методы не применяются к водорастворимым белкам, заключенным внутри органелл, например к белкам митохондриального матрикса, а лишь к молекулам, ковалентно связанным или прочно ассоциированным с частицами. Существенной степени очистки по сравнению с исходным материалом 33 можно добиться, многократно промывая гомогенат буфером, в котором данный белок не растворяется. После этого возможны два подхода к решению проблемы. Можно фракционировать осадок методом дифференциального центрифугирования (седиментация или градиент плотности), с помощью электрофореза или «молекулярных сит». Другой способ заключается в солюбилизации белков (субфракционирование, конечно, может предшествовать солюбилизации). Не все ферменты способны существовать в солюбилизированном состоянии, будучи изолированы от их нормального клеточного окружения; поэтому успех очистки таких соединений зависит от того, насколько полно можно отделить фрагменты частиц, содержащие фермент, от прочего корпускулярного материала. Белок, связанный с частицами, может быть солюбилизирован различными путями. Во многих случаях солюбилизированные белки ведут себя в дальнейшем как все водорастворимые белки, и их фракционирование может быть проведено с помощью процедур, описанных в гл. 3-5. С другой стороны, иногда возникает необходимость солюбилизировать вещество, используя детергенты. В этом случае детергент, связанный с белком, замещает липидсодержащую мембрану. Если впоследствии детергент удаляют, белок может денатурировать или, по крайней мере, агрегировать и выпадать из раствора. Большая часть подробно изученных мембраносвязанных белков и ферментов относится к первой категории: их можно выделить с помощью физических, химических или энзиматических методов; полученный раствор фракционируют далее обычным способом. Много таких ферментов содержится в митохондриях, в том числе ферменты, ассоциированные с системой переноса электронов. Для их выделения используют следующие методы: а) Разрушение митохондрий для выделения белков матрикса с последующим выделением мембранных белков из митохондриальных фрагментов с помощью 1) ультразвука, особенно при высоких значениях рН и повышенной температуре, 2) щелочи (рН 8-11) в присутствии или отсутствие агентов, образующих хелаты металлов (ЭДТА), 3) солюбилизации под действием детергентов, 4) органических растворителей (например, получение ацетонового порошка, экстракция н-бутанолом, этанолом и т.д.), 5) фосфолипазы А для расщепления липидов мембраны. б) Классический способ, до сих пор находящий применение и заключающийся в экстракции липидов и получении ацетонового порошка. Суспензию частиц обрабатывают несколькими объемами ацетона при очень низкой температуре (например, -20°С или ниже). Полученный таким образом обезвоженный порошок может храниться долгое время. Белки растворяют, диспергируя их в водном буферном растворе. Большинство мембраносвязанных белков можно экстрагировать из мембран в присутствии детергентов, в состав которых входят липофильные 34 цепи, взаимодействующие с гидрофобными поверхностями белка и вытесняющие его из комплекса с нормальной мембраной. Из детергентов, используемых для этой цели, наиболее широко применяются дезоксихолат натрия и тритон. Тритон - это торговое название целой серии в большинстве своем неионных детергентов на основе полиэтиленгликоля. Наиболее часто в различных целях используют тритон Х-100, хотя детергенты других типов также находят определенное применение. Особенно полезным является недавно описанное цвитерионное производное холевой кислоты. Детергенты способны вытеснять белок, прочно связанный с мембраной гидрофобными взаимодействиями, благодаря тому, что они, во-первых, растворяют мембрану и, во вторых, замещают компоненты мембраны алифатическими или ароматическими цепями, которые составляют липофильную часть детергента. Если белок солюбилизирован и установлено, что его целостность не нарушена, можно провести фракционирование. В большинстве случаев это не представляет особой проблемы. Необходимо сохранить биологическую активность белка; это зависит от выбора подходящего детергента. Неионные детергенты, такие, как тритон Х-100, очень мягки по своему действию, и большинство белков, как связанных с мембранами, так и свободных, выдерживают концентрации тритона Х-100 до 1-3% (вес/об.). Напротив, некоторые анионные детергенты (например, додецилсульфат) обладают наряду с солюбилизирующими свойствами очень сильным денатурирующим действием. Они, по-видимому, не найдут широкого применения при выделении ферментов. Липопротеины содержат ковалентно- или нековалентно- (но прочно) связанный липид, который в свою очередь погружен в мембрану и служит связующим звеном между белком и частицей. Экстракция в присутствии детергента может привести к удалению липида и замещению его детергентом. Это может быть причиной потери ферментативной активности, которая восстанавливается при добавлении природного липида - обычно фосфатида. С такими белками особенно трудно работать, так как они очень чувствительны к соотношению в растворе детергента и липида. Удаление детергента неизменно ведет к агрегации и чаще всего к денатурации. Избыток детергента может мешать фракционированию. На пример, высаливание сульфатом аммония приводит к появлению на поверхности раствора слоя тритона Х-100, в котором часто содержатся нужные белки. Однако эффективного разделения при этом не происходит. Можно провести колоночную хроматографию или отделить белки с помощью гельфильтрации, но не исключено, что мицеллы детергента будут двигаться в той же зоне, что и белок, и, следовательно, окажутся в одной фракции. Ионообменная хроматография успешно осуществляется в присутствии неионных детергентов. Действительно, тритон Х-100 в концентрации до 1 % оказывает незначительное влияние на ионообменные свойства нормальных водорастворимых белков. Но солюбилизированные белки мембран могут 35 находиться только в составе детергентных мицелл, что существенно влияет на процесс ионного обмена. Если исследуемый белок удается адсорбировать на ионообменнике, то избыток детергента свободно проходит через колонку. Это позволяет элюировать свободный (относительно) от детергента белок. С другой стороны, если полное удаление детергента приводит к денатурации белка, то, чтобы предотвратить это, в буфер вносят небольшое количество детергента (<0,1%). Собранная фракция будет, конечно, тоже содержать некоторое количество детергента. Тем не менее, так как обычно из смеси белков выделяют какой-то определенный фермент, присутствие в конечном препарате незначительной концентрации чистого детергента, не загрязненного жирами, не принесет большого вреда. Белки, связанные с мембранами, очевидно, отличаются по растворимости от типичных цитоплазм этических ферментов, и для их очистки иногда применяются (и вполне успешно) некоторые необычные эмпирически найденные методы. ГЛАВА 3. Производство аминокислот В промышленности аминокислоты получают: 1) гидролизом природного белоксодержащего сырья; 2) химическим синтезом; 3) микробиологическим синтезом; 4) биотрансформацией предшественников аминокислот с помощью микроорганизмов или выделенных из них ферментов (химикомикробиологический метод). Для гидролиза могут быть использованы отходы мясоперерабатывающей промышленности (отходы обработки животного сырья, кровь и т.д.), яичный белок, казеин молока, клейковина пшеницы, соевый шрот и т.д. При гидролизе белоксодержащее сырье нагревают с растворами кислот и щелочей, при температуре от 100 до 105 ºС в течение 20…48 часов. При этом аминокислоты переходят в гидролизат, и для выделения отдельных аминокислот необходима сложная многостадийная очистка. Кроме того, само сырье считается дефицитным и дорогим, поэтому аминокислоты имеют высокую себестоимость. Кроме того, может разрушиться часть аминокислот, таких как триптофан, цистеин, метионин, тирозин, а также происходит рацемизация. Химический синтез аминокислот достаточно эффективен, однако его недостатком является то, что в процессе синтеза образуется смесь из биологически активной L-формы и D-изомера аминокислоты. D-форма является балластом, так как не усваивается животными и человеком, а некоторые D-формы аминокислот обладают токсическими свойствами. 36 Разделение изомеров – дорогая и трудоемкая процедура. Синтетически производится незаменимая аминокислота метионин. приготовление питательной среды подготовка посевного материала продуцент культивирование разделение клетки биомасса живых клеток культуральная жидкость биомасса убитых клеток дезинтеграция убитых клеток выделение и очистка метаболитов концентрирование стабилизация продукта обезвоживание сухой продукт жидкий продукт хранение применение Рисунок 3 – Принципиальная биотехнологическая схема производства продуктов микробного синтеза 37 Наиболее перспективен и экономически выгоден микробиологический синтез аминокислот; 60 % высокоочищенных препаратов аминокислот получают именно этим способом. Преимущество его состоит в возможности получения L-аминокислот на основе возобновляемого сырья. В последние годы широко используется биотрансформация предшественников аминокислот, полученных химическим синтезом с помощью клеток микроорганизмов или иммобилизированных ферментов. Среди продуцентов аминокислот используются дрожжи (30 %), актиномицеты (30 %), бактерии (20 %). Brevibacterium flavum и Corynebacterium glutamicum более трети сахаров превращают в лизин. Для селекции продуцентов используются микроорганизмы, относящиеся к родам Micrococcus, Brevibacterium, Corynebacterium, Arthrobacter. Глутаминовая кислота – первая аминокислота, полученная микробным синтезом. Глутаминовая кислота относится к заменимым кислотам, обладает приятными органолептическими свойствами и находит самое широкое применение. Ее продуцентами являются бактерии Corinebacterium glutamicum, Brevibacterium flavum и др. Лизин образуют многие микроорганизмы: бактерии, актиномицеты, сине-зеленые водоросли, некоторые виды микроскопических грибов. В нашей стране в качестве продуцентов лизина используют бактерии родов Corinebacterium (С. glutamicum), Micrococcus, Brevibacterium. Триптофан образуют микроорганизмы бактериального и грибного происхождения: родов Micrococcus sp., Candida utilis, Bacillus subtilis. Основными потребителями аминокислот являются сельское хозяйство и пищевая промышленность. Аминокислоты, чаще всего лизин, используют в качестве обогатителей кормов и пищевых продуктов растительного происхождения для повышения их питательной ценности и для сбалансирования пищи по незаменимым аминокислотам. Использование 1 т лизина в комбикормовой промышленности позволяет экономить от 40 до 50 т фуражного зерна. Некоторые аминокислоты используют в качестве приправ, так как они обладают определенными вкусовыми свойствами и могут сообщать продукту приятные аромат и вкус. Большое распространение имеет глутаминовая кислота и ее натриевая соль (глутамат натрия), которая является эффективным усилителем вкуса мясных и овощных блюд. Данную аминокислоту добавляют во многие продукты при консервировании, замораживании и длительном хранении. Для улучшения органолептических показателей мясных продуктов, придания им специфического приятного вкуса и аромата используют цистин, лизин, гистидин. Цистеин и цистин с глутаматом натрия создают имитацию запаха и вкуса мяса, что используется при приготовлении приправ. 38 Многие аминокислоты: лизин, аланин, пролин, валин и другие могут снимать неприятные запахи и используются в качестве дезодорантов пищевых продуктов. Аминокислоты обладают оригинальным вкусом и участвуют в образовании вкусовых особенностей пищевых продуктов. Например, аспарагиновая и глутаминовая кислоты, кислые на вкус, в нейтральных растворах имеют очень приятный оригинальный вкус, глицин обладает характерным вкусом «освежающей» сладости, которая по интенсивности близка к сахарозе. Особый интерес представляет подсластитель аспартам, молекулу которого образуют две аминокислоты – фенилаланин и аспарагиновая кислота. Эти аминокислоты синтезируются микробиологическим путем, а аспартам из этих мономеров – с помощью ферментов. Сладость аспартама в 200 раз превышает сладость сахарозы. 3.1. Биотехнология синтеза аминокислот и их очистка. В последние годы широкое применение в народном хозяйстве и медицине находят различные аминокислоты. Особое значение они имеют для сбалансирования белкового питания. Некоторые пищевые и кормовые продукты не содержат в своем составе необходимых количеств незаменимых аминокислот, в частности лизина. К таким продуктам относятся пшеница, кукуруза, овес, рис и ряд других. Для ликвидации возможного дисбаланса аминокислоты используют в чистом виде или вводят в состав комбинированных кормов, выпускаемых промышленностью. Поэтому основной сферой применения аминокислот следует считать создание рационов, позволяющих понизить содержание растительных белков в кормах. Показано, что искусственные смеси аминокислот позволяют экономить расход естественных кормов. Кроме добавок к кормам сельскохозяйственных животных аминокислоты используются в пищевой промышленности. Применяются они и при изготовлении ряда полимерных материалов, например синтетической кожи, некоторых специальных волокон, пленок для упаковки пищевых продуктов. Ряд аминокислот или их производных обладают пестицидным действием. Метионин и γаминомасляная кислота широко применяются как лекарственные средства. Удельный вес применения аминокислот в различных отраслях хозяйства может быть продемонстрирован на примере Японии, где на долю пищевой промышленности приходится 65% всех производимых в стране аминокислот, на животноводство -18, для медицинских целей - 15 и на прочие нужды - 2 %. Мировой уровень производства аминокислот достигает в настоящее время нескольких миллионов тонн в год. В наибольших количествах в мире вырабатываются L-глутаминовая кислота, L-лизин, DL-метионин, Lаспарагиновая кислота, глицин. Основными способами получения аминокислот являются следующие: экстракция из белковых гидролизатов растительного сырья, химический синтез, микробиологический синтез 39 растущими клетками, при использовании иммобилизованных микробных клеток или ферментов, выделенных из микроорганизмов. Микробиологический синтез - в настоящее время весьма перспективный и экономически выгодный способ получения многих аминокислот. В процессе культивирования Продуцентов аминокислот непосредственно синтезируются L-аминокислоты. Одна из важных задач микробиологического синтеза аминокислот - получение высокоактивных штаммов - продуцентов, в частности, с использованием методов генной инженерии. Именно таким способом в СССР получен высокоактивный штамм - продуцент L-треонина. Кроме микробиологического синтеза аминокислоты можно получать, как указано выше, путем гидролиза природного белок-содержащего животного и растительного сырья. Это наиболее старый способ. Основным его недостатком является нерациональное использование сырья, которое с большой пользой может применяться в качестве белковых кормов или пищевых продуктов. Например, в странах юго-восточной Азии моноглутамат натрия получают из соевого шрота (обезжиренная соевая мука). В США описан способ получения аминокислот из клейковины пшеницы и кукурузного глютена, остающегося после отмывки крахмала. В равной мере, очевидно, могут быть названы и другие белки, однако их использование для получения аминокислот экономически невыгодно. Химический синтез аминокислот достаточно эффективен, позволяет получить соединения любой структуры и организовать непрерывное производство при высокой автоматизации. В нем в основном используется непищевое сырье, достигается высокая концентрация продукта. Однако, как правило, процесс этот многостадийный и требует сложной аппаратуры. Главный недостаток химического синтеза - получение рацемической формы аминокислот. Пока не разработаны достаточно эффективные и дешевые пути разделения соединений на оптические изомеры. Химический синтез рентабелен для получения только тех аминокислот, которые могут быть использованы в виде рацемического продукта. Наиболее хорошо разработан химический синтез LD-метионина, главным потребителем которого является птицеводство. L- и D-изомеры метионина усваиваются организмами одинаково хорошо. В последние годы имеются успехи в области асимметрического синтеза аминокислот, позволяющие избежать оптического разделения рацемических аминокислот. Новый подход к этой проблеме стал возможен благодаря открытию гомогенного каталитического гидрирования олефинов с помощью комплексов родия с фосфиновыми лигандами и разработке путей синтеза хиральных фосфинов. Применение комплексов родия с хиральными фосфинами в качестве гомогенных катализаторов для гидрирования Nациламиноакриловых кислот позволило осуществить асимметрический синтез α-аминокислот с высокой степенью стереоспецифичности и хорошими выходами. 40 Использование для пищевых, кормовых и медицинских целей аминокислот, полученных химическим синтезом, ставит еще одну существенную технологическую проблему - полное освобождение готового продукта от возможных токсических полупродуктов синтеза. В последние годы прочные позиции начинает занимать комбинированный химико-микробиологический метод синтеза, при котором исходное соединение получают в результате химических реакций, а конечная стадия осуществляется за счет активности ферментных систем соответствующих штаммов микроорганизмов. Микробиологический метод синтеза аминокислот основан на способности многих микроорганизмов накапливать в среде значительные количества таких продуктов. Среди микроорганизмов, получивших оценку как потенциальные продуценты глутаминовой кислоты, обнаружено много бактерий, ряд дрожжей и других грибов. Большинство обследованных штаммов микроорганизмов независимо от их систематического положения преимущественно накапливают α-аланин и глутаминовую кислоту. Значительно меньше штаммов и в меньшем количестве выделяют аспарагиновую кислоту, лейцин, валин, изолейцин, лизин. Строгой корреляции между видовой принадлежностью микроорганизмов и способностью их накапливать аминокислоты нет. Несмотря на широкое распространение микроорганизмов, накапливающих аминокислоты в процессе роста, продуцентов, обеспечивающих экономически выгодные выходы этих продуктов, не так много. Получают их обычно путем применения различных мутагенных факторов. Продуцент должен аккумулировать преимущественно одну аминокислоту. Одновременное присутствие нескольких аминокислот, особенно если они близки по своим физико-химическим свойствам, затрудняет их выделение и очистку. Ауксотрофные мутанты микроорганизмов, лишенные в результате действия мутагенов, ряда ферментных систем, признаны наиболее ценными продуцентами. Блокада у таких мутантов соответствующих реакций в цепи обмена веществ приводит к сверхсинтезу одного из метаболитов. Наиболее распространенные продуценты аминокислот грамположительные бесспоровые бактерии, относимые к родам Corynebacterium, Micrococcus, Arthrobacter, Brevibacterium и некоторым другим, но точное таксономическое положение большинства из них определить трудно, так как содержащаяся в публикациях информация явно недостаточна для этого. Одним из наиболее важных научных положений микробиологического синтеза аминокислот считается вопрос об их происхождении: находящиеся в среде аминокислоты - продукты ферментативного распада белков в результате автолитического процесса или они результат синтеза из других соединений. При использовании синтетических сред для культивирования 41 продуцентов достаточно определенно показано, что аминокислоты, обнаруживаемые в среде, представляют собой продукты синтеза de novo. Ферментативные реакции синтеза аминокислот протекают внутри клеток. Первоначально аминокислоты накапливаются внутри клеток в виде так называемых свободных аминокислот. На ранних этапах роста культуры свободные аминокислоты включаются в конструктивный обмен микроорганизма. Активное накопление аминокислот в среде в периодической культуре происходит обычно с середины экспоненциальной фазы ее роста, достигая максимума к концу. 3.2. Получение аминокислот с помощью иммобилизованных клеток и ферментов. В последние годы внимание исследователей привлекают методы получения аминокислот с использованием иммобилизованных ферментов. Способ имеет ряд преимуществ, в частности, конечный продукт отличается высокой концентрацией и чистотой, нет опасности заражения в ходе реакции посторонними микроорганизмами, в результате синтеза образуются только природные изомеры, имеется возможность осуществления непрерывных технологических процессов. Микроорганизмы являются основными источниками ферментов, переводимых в иммобилизованную форму. Имея в виду преимущества иммобилизованных ферментов, необходимо учитывать, что они всегда будут дороже растворимых, но их внедрение экономически оправдано при удовлетворении даже одного из приводимых ниже условий: повышение стабильности фермента, обеспечивающее его многократное применение и тем самым сокращение расходов на препарат; улучшение качества продукта благодаря отсутствию в нем следов фермента и предотвращению нежелательных побочных реакций. Из уже перечисленных способов получения аминокислот можно обратиться к превращению DL-α-амино-ε-капролактама в лизин. Ферменты, участвующие в реакциях гидролиза и рацемизации, могут быть иммобилизованы на ионообменных полисахаридах путем ковалентного связывания. Одно из серьезных технологических затруднений при осуществлении ферментативных реакций на носителях - возможность регенерации кофакторов (если реакция идет при их участии). Наиболее широкое распространение имеет ферментативный метод получения аспарагиновой кислоты из фумаровой и аммония благодаря активности аспартазы, катализирующей эту реакцию: Поскольку аспартаза, заключенная в полиакриламидный гели, относительно быстро теряет исходную активность, иммобилизации подвергают микробные клетки, обладающие аспартазной активностью. Хорошим продуцентом аспартазы признаны некоторые штаммы Escherichia coli, клетки которой фиксируются в полиакриламидном геле. При оптимальных режимах выход аспарагиновой кислоты при таком способе 42 достигает 12000-16000 мкМ/ч∙г сухих клеток. Отмечено существенное повышение активности аспартазы клеток Е. coli после их иммобилизации. Ферментативные методы используются также при синтезе L-аланина декарбоксилированием L-аспарагиновой кислоты с помощью Pseudomonas dacunhae. 3.3. Получение оптических изомеров аминокислотпутем применения ацилаз микроорганизмов. Одним из способов разделения рацематов аминокислот на L- и Dизомеры является ферментативный путь с использованием микроорганизмов, обладающих специфической L-ацилазной активностью. Ацилазы разрывают пептидную связь у ацилпроизводных аминокислот или пептидов, в результате чего образуются соответствующие свободные аминокислоты и пептиды, а также органическая кислота: При культивировании микроорганизмов, содержащих ацилазы, необходимо соблюдать ряд условий. В частности, вводить в среды вещества, близкие по природе к субстрату действия ацилаз, например, в случае превращения DL-лизина - ε-ацетил-L-лизин. Ацилазы грибов находятся внутри клеток. Поэтому их клетки необходимо предварительно подвергать дезинтеграции, например, применяя ультразвук. Основой использования микробных ацилаз для гидролиза ацилпроизводных аминокислот является специфичность их действия относительно оптической конфигурации и структуры субстрата. Соответственно характеру асимметрического деацетилирования субстрата различают L- и D-ацилазы, причем у микроорганизмов наиболее часто обнаруживаются L-ацилазы. В зависимости от характера отщепляемого ацильного радикала различают ацетил-, бензоил-, хлорацетил-, сукцинил- и другие производные аминокислот. Поскольку наиболее распространенными являются ацетил- и бензоилацилазы, именно эти производные аминокислот предпочтительно использовать в реакциях. Необходимо иметь в виду специфичность ацилазы к аминокислотному остатку. Указанные особенности ацилаз позволяют использовать их для выделения нужной аминокислоты из смеси многих ацилпроизводных: Остающиеся ацил-рацемат и ацил-D-аминокислота легко отделяются от оптически активной аминокислоты вследствие различий в физикохимических свойствах, например растворимости в органических растворителях и воде, способности сорбироваться на ионитах. Использование D-изомеров теоретически возможно, если иметь в виду дальнейшее применение рацемаз или химических способов. Например, некоторые N-ацетил-D-аминокислоты могут легко превращаться в N-ацетилDL-аминокислоты путем нагревания в присутствии уксусного ангидрида. Процесс рацемизации может быть реализован также путем применения иммобилизованных ферментов. 43 ГЛАВА 4. Производство витаминов 4.1. Общая характеристика витаминов Основоположник учения о витаминах русский врач Н. И. Лунин установил (1880 г.), что при кормлении белых мышей только искусственным молоком, состоящим из казеина, жира, молочного сахара и солей, животные погибают. Следовательно, в натуральном молоке содержатся и другие вещества, незаменимые для питания. В 1912 г. польский врач К. Функ, предложивший само название «витамины», обобщил накопленные к тому времени экспериментальные и клинические данные и пришѐл к выводу, что такие заболевания, как цинга, рахит, пеллагра, бери-бери — болезни пищевой недостаточности, или авитаминозы. С этого времени наука о витаминах (витаминология) начала интенсивно развиваться, что объясняется значением витаминов не только для борьбы со многими заболеваниями, но и для познания сущности ряда жизненных явлений. Метод обнаружения витаминов, примененный Луниным (содержание животных на специальной диете — вызывание экспериментальных авитаминозов), был положен в основу исследований. Было выяснено, что не все животные нуждаются в полном комплексе витаминов. Отдельные виды животных могут самостоятельно синтезировать те или иные витамины. В то же время многие плесневые и дрожжевые грибы и различные бактерии развиваются на искусственных питательных средах только при добавлении к этим средам вытяжек из растительных или животных тканей, содержащих витамины. Таким образом, витамины необходимы для всех живых организмов. Изучение витаминов не ограничивается обнаружением их в естественных продуктах с помощью биологических тестов и другими методами. Из этих продуктов получают активные препараты витаминов, изучают их строение и, наконец, получают синтетически. Исследована химическая природа всех известных витаминов. Оказалось, что многие из них встречаются группами по 3—5 и более родственных соединений, различающихся деталями строения и степенью физиологической активности. Было синтезировано большое число искусственных аналогов витаминов с целью выяснения роли функциональных групп. Это способствовало пониманию действия витаминов. Так, некоторые производные витаминов с замещенными функциональными группами оказывают на организм противоположное действие, по сравнению с витаминами, вступая с ними в конкурентные отношения за связь со специфическими белками при образовании ферментов или с субстратами воздействия последних. Витамины имеют буквенные обозначения, химические названия или названия, характеризующие их по физиологическому действию. В 1956г. принята единая классификация витаминов, которая стала общеупотребительной. Наличие химически чистых витаминов дало возможность подойти к выяснению их роли в обмене веществ организма. Витамины либо входят в состав ферментов, либо являются компонентами ферментативных реакций. 44 При отсутствии витаминов в организме нарушается деятельность ферментных систем, в которых они участвуют, а следовательно, — и обмен веществ. Известно несколько сот ферментов, в состав которых входят витамины, и огромное количество катализируемых ими реакций. Многие витамины — преимущественно участники процессов распада пищевых веществ и освобождения заключѐнной в них энергии (витамины B1, В2, PP и др.). Участвуют они и в процессах синтеза: B6 и В12 — в синтезе аминокислот и белковом обмене, В3 (пантотеновая кислота) — в синтезе жирных кислот и обмене жиров, Вс (фолиевая кислота) — в синтезе пуриновых и пиримидиновых оснований и многих физиологически важных соединений — ацетилхолина, глутатиона, стероидов и др. Таким образом, витамины имеют огромное физиологическое значение. Выяснение физиологической роли витаминов позволило использовать их для витаминизации продуктов питания, в лечебной практике и в животноводстве. Особенно широко стали применяться витамины после освоения их промышленного синтеза. Витамины не образуются у гетеротрофов. Способностью к синтезу витаминов обладают лишь автотрофные организмы. Микробиологическим способом можно получить практически все известные витамины. Однако экономически более целесообразно получать витамины выделением из природных источников или с помощью химического синтеза. С помощью микроорганизмов целесообразно получать сложные по строению витамины: β-каротин (провитамин А), В2, В12 и предшественники витамина D. Витамин В12 (цианокобаламин). В тканях животных концентрация витамина очень низкая (в печени быка 1 мг/кг) для того, чтобы использовать этот источник для промышленных целей. Химический синтез очень сложен. Синтезировать витамин В12 способны уксуснокислые бактерии, грибы и пропионовокислые бактерии. Наибольшее промышленное значение имеют Рго-pionibacterium и Pseudomonas (P. denitrificans). Концентрат витамина B12 предназначен для обогащения кормов животных. Для обогащения кисломолочных продуктов витамином B12 используют пропионовокислые бактерии, как в чистом виде, так и в виде концентрата, приготовленного на молочной сыворотке. Витамин В2 (рибофлавин) можно в небольших количествах выделять из природного сырья. В наибольшем количестве он содержится в моркови и печени трески. Наиболее активными продуцентами витамина B2 являются дрожжеподобные грибы рода Eremothecium ashbyii, входящие в класс аскомицетов, а также бактерии Bacillus subtilis. Витамином В2 обогащают некоторые сорта белого хлеба, его используют для окраски пищевых продуктов в оранжево-желтый цвет. Каротиноиды – это предшественники витамина А, среди которых наиболее активен β-каротин. В организме человека каротиноиды не 45 синтезируются, поэтому должны поступать извне. В печени каротин превращается в витамин А. Продуцентами каротиноидов могут быть грибы и дрожжи. В промышленности β-каротин чаще всего получают с помощью микроскопического гриба рода Blakeslea trispora. β-Каротин используют при изготовлении пищевых продуктов как краситель. Его применяют при изготовлении колбас с целью замены нитрита натрия и обеспечения высокой интенсивности и устойчивости цвета. Используют при производстве леденцов, пищевых паст, кексов и других кондитерских изделий. Во многих странах β-каротин применяют для подкрашивания сливочного масла. Кроме того, он обладает антиокислительными свойствами, которые используются для продления срока хранения продукта. Витамин D2 промышленно синтезируют с помощью дрожжей Saccharomyces serevisiae. Витамин используется для лечения и профилактики рахита человека и животных. Перспективно микробиологическое получение биотина, используемого в рационе кур и свиней. В настоящее время на Западе в большую часть комбикормов для свиней включают биотин, получаемый путем химического синтеза. В результате химического синтеза образуется рацемическая смесь, а биологической активностью обладает лишь D-форма витамина, которую синтезируют микроорганизмы. В мире существует 40 крупных промышленных производителей витаминов; 18 из них в США, 8 - в Японии, 14 - в Западной Европе Ведущее место в производстве витаминов занимает швейцарский концерн Hoffman La Roche, выпускающий 50-70% всех витаминов. 4.2. Получение водорастворимых витаминов Витамины получают главным образом синтетически и лишь в некоторых случаях отдельные стадии в цепи синтеза выполняются биологическими способами. Производство концентратов витаминов из продуктов растительного или животного происхождения почти полностью потеряло своѐ значение. Получение витаминов относится к тонкому органическому многостадийному синтезу. Химическими методами синтезируют следующие водорастворимые витамины: B1, B2, В3, B6, Вс, С, PP, а В12 — ферментативными методами микробиологического синтеза. Ферментацией пользуются также на одной из стадий синтеза витамина С. Этот витамин в виде индивидуального кристаллического вещества высокой степени чистоты образуется при восстановлении D-глюкозы в D-copбит. Последний ферментативно окисляют в L-copбозу, которую после ряда операций превращают в витамин С (I). 46 Витамин В1. Производство витамина B1 (тиамина, VI) основано на конденсации 2-метил-4-амино-5-хлор (бром) метилпиримидина с 4-метил-5b-оксиэтилтиазолом. Кофермент витамина B1 — кокарбоксилаза (VII), или дифосфорный эфир тиамина, применяемый для лечения заболеваний сердца, получают фосфорилированием тиамина с последующей очисткой на ионообменных смолах и кристаллизацией. Витамин В2 (рибофлавин, VIII) образуется при культивировании Eremothecium ashbyii и других микроорганизмов без выделения в виде сухой биомассы (с использованием только для кормления с.-х. животных), а синтетический рибофлавин (применяемый в медицине) получают в виде кристаллического продукта деструктивным окислением D-глюкозы (из кукурузного крахмала) в D-apaбоновую кислоту и рядом других операций превращают в конечный продукт — жѐлто-оранжевые кристаллы высокой степени чистоты. Важное производное рибофлавина — его кофермент рибофлавин-5'-фосфат натрия (IX, R = Na), применяемый для инъекций, получают фосфорилированием рибофлавина, а другой кофермент — ФАД (IX, R — остаток аденозин-5'-фосфата) получают конденсацией рибофлавина-фосфата и аденозин-5'-фосфата. Витамин В2 содержится в клетках различных микроорганизмов, будучи коферментом в составе флавопротеинов (прежде всего 47 соответствующих ферментов из класса оксидоредуктаз - ФМН, ФАД). Поэтому в качестве продуцентов рибофлавина (флавопротеинов) могут быть бактерии, дрожжи и нитчатые грибы. Однако наиболее заманчивыми являются те штаммы, которые образуют на жидких средах 0,5 г и более рибофлавина в 1 л среды. К подобным организмам относятся Ashbyii gossypii, Eremothecium ashbyii и Candida guilliermondii. Учитывая изменчивость активных продуцентов названных видов по способности синтезировать витамин В2, необходим систематический отбор культур в процессе их эксплуатации на производстве. Обычно активные продуценты первых двух видов формируют яркооранжевые колонии на агаризованных средах. Методами генной инженерии удалось получить штамм сенной палочки, образующий около 6 г рибофлавина в 1 л среды, включающей мелассу, белково-витаминный концентрат и его гидролизат. Высокий выход рибофлавина у Е.ashbyii коррелирует с азотом пуринов и другими азотистыми источниками, содержание которых должно быть достаточным. В качестве источников углерода применяют глюкозу или сахарозу, практикуют использование дрожжевого и кукурузного экстрактов, соевой муки, масла (жира). Ферментационная среда обычно включает кукурузную и соевую муку, сахарозу, кукурузный экстракт, калия дигидрофосфат, кальция карбонат, натрия хлорид и ненасыщенный жир. Обычно ферментацию проводят в течение 5 суток при рН 5,5- 7,7. После использования сахарозы (примерно через 30 часов) начинает заметно накапливаться витамин В2, вначале - в мицелии, а затем - в культуральной жидкости. Всю биомассу можно подвергнуть высушиванию и полученный сухой продукт с остаточной влажностью 8%, содержащий 1,5-2,5% рибофлавина, 20% белка, тиамин, никотиновую кислоту, пиридоксин, цианкобаламин, микроэлементы и другие вещества, рекомендуют для кормления животных. В случае высоких выходных показателей по рибофлавину, витамин можно выделять в индивидуальном состоянии и, наряду с синтетическим рибофлавином, использовать в медицине. Для Candida guillierniondii важно регулировать содержание железа в питательной среде; оптимальные концентрации колеблются, в среднем, от 0,005 до 0,05 мкг/мл. При этом определенные штаммы дрожжей могут образовывать за 5-7 дней до 0,5 г/л и более витамина. Однако для целей промышленного производства рибофлавина предпочитают использовать более продуктивные виды и штаммы грибов - E.ashbyii и Ashbyii gossypii. Витамин B6 (пиридоксин, X, а) синтезируют, конденсируя метоксиацетил-ацетон с циануксусным эфиром в присутствии аммиака в 2метил-4-метоксиметил-5-циан-6-оксипиридин, который подвергают нитрованию, затем рядом операций превращают в пиридоксин. Известен также и другой способ получения пиридоксина — через 4-метил-5пропоксиоксазол диеновым синтезом с формалем бутен-2-диола-1,4. Другими формами B6 являются пиридоксол (X, б) и пиридоксамин (X, в). 48 Витамин Вс (фолиевую кислоту, XI) синтезируют одностадийной конденсацией 2,4,5-триамино-6-оксипиримидина, 1,1,3-трихлорацетона и nаминобензоил-L-глутаминовой кислоты. Витамин PP (никотиновую кислоту, XII) получают окислением bпиколина (выделяемого из каменноугольного дѐгтя), ресурсы которого ограниченны, а также окислением хинолина или 2-метил-5-этилпиридина. Для медицинских целей пользуются, кроме никотиновой кислоты, никотинамидом (XIII). Витамин B3, оптически активная D-пантотеновая кислота для медицинских целей применяется в виде кальциевой соли. HOCH2C (CH3)2CH (OH) CONH (CH2)2COOH 49 Для нужд животноводства нет необходимости в разделении на промежуточных ступенях синтеза рацемата пантолактона на оптические антиподы. Синтез рацемического пантотената кальция состоит в альдольной конденсации изобутираля и формальдегида с последующим превращением в пантолактон, затем в его конденсации с b-аланином, приводящей к образованию конечного продукта. Витамин В12 (цианкобаламин) получают только микробиологическим синтезом. Его продуцентами являются прокариоты и, прежде всего, пропионовые бактерии, которые и в естественных условиях образуют этот витамин. Мутанты Propionibacterium shermanii M-82 и Pseudomonas denitrificans M-2436 продуцируют на жидкой среде до 58-59 мг/л цианкобаламина. Учитывая важную функцию витамина в организме человека (он является противоанемическим фактором), его мировое производство достигло 10 т в год, из которых 6,5 т расходуют на медицинские нужды, а 3,5 т - в животноводстве. Отечественное производство цианкобаламина базируется на использовании культуры P.freudenreichii var. shermanii, культивируемой в периодическом режиме без доступа кислорода. Ферментационная среда обычно содержит глюкозу, кукурузный экстракт, соли аммония и кобальта, рН около 7,0 поддерживают добавлением NH4OH; продолжительность ферментации 6 суток; через 3 суток в среду добавляют 5,6диметилбензимидазол - предшественник витамина Б12 и продолжают ферментацию еще 3 суток. Цианкобаламин накапливается в клетках бактерий, поэтому операции по выделению витамина заключаются в следующем: сепарирование клеток, экстрагирование водой при рН 4,5-5,0 и температуре 85-90?С, в присутствии стабилизатора (0,25% раствор натрия нитрита), Экстракция протекает в течение часа, после чего водный раствор охлаждают, нейтрализуют раствором едкого натра, добавляют коагулянты белка - хлорид железа трехвалентного и алюминия сульфат с последующим фильтрованием. Фильтрат упаривают и дополнительно очищают, используя методы ионного обмена и хроматографии, после чего проводят кристаллизацию витамина при 3-40 С из в одноацетонового раствора. Кристаллический цианкобаламин можно получать с помощью резорцина или фенола, образующих с ним аддукты, которые сравнительно легко разлагаются на составляющие компоненты. При реализации данного биотехнологического процесса не забывать о высокой светочувствительности витамина В12, поэтому все операции необходимо проводить в затемненных условиях (или при красном свете). На ацетонобутиловой и спиртовой бардах с добавлением солей кобальта и метанола в нашей стране получают кормовой препарат КМБ 12 - концентрат, содержащий витамин В12 и другие ростовые вещества. 50 Витамин С (аскорбиновая кислота) - это противоцинготный витамин, имеющийся у всех высших растений и животных; только человек и бактерии не синтезируют ее, но людям она необходима, а бактерии не нуждаются в ней. И, тем не менее, определенные виды уксуснокислых бактерий причастны к биосинтезу полупродукта этой кислоты - L-сорбозы. Таким образом, весь процесс получения аскорбиновой кислоты является смешанным, то есть химико-ферментативным. Биологическая стадия процесса катализируется мембраносвязанной полиолдегидрогеназой, а последняя (химическая) включает последовательно следующие этапы: конденсация сорбозы с диадетоном и получение диацетон - L-сорбозы, окисление диацетон -L-сорбозы до диацетон-2-кето-гулоновой кислоты, подвергаемой затем гидролизу с получением 2-кето-1-гулоновой кислоты; последнюю подвергают энолизации с последующей трасформацией в L-аскорбиновую кислоту. Ферментацию Gluconobakter oxydans проводят на средах, содержащих сорбит (20%), кукурузный или дрожжевой экстракт, при интенсивной аэрации (8-10 г О2/л/ч). Выход L-сорбозы может достичь 98% за одни-двое суток. При достижении культурой log-фазы можно дополнительно внести в среду сорбит, доводя его концентрацию до 25%. Также установлено, что G.oxydans может окислять и более высокие концентрации полиспирта (3050%), создаваемые на последних стадиях процесса. Это происходит благодаря полиолдегидрогеназы, содержащейся в клеточной биомассе. Ферментацию бактерий проводят в периодическом или непрерывном режиме. Принципиально доказана возможность получения L-сорбозы из сорбита с помощью иммобилизованных клеток в ПААГ. 51 4.3. Получение жирорастворимых витаминов Получение и применение эргостерина В промышленности эргостерин получают, используя дрожжи Sacch. cerevisiae, Sacch. carlsbergensis, а также мицелиальные грибы. Засев производят большим количеством инокулята. Культивирование ведут при высокой температуре и сильной аэрации в среде, содержащей большой избыток источников углерода по отношению к источникам азота. Дрожжи, а также грибы рода Aspergillus и Penicillium используют для получения кристаллического витамина D2 или концентрата. В качестве концентрата в животноводстве применяют облученные сухие дрожжи. Максимум поглощения эргостерина отмечен при 280 нм. Именно это излучение возбуждает отдельные связи колец А и В в молекуле эргостерина и вызывает его превращение в витамин D2. Облучение производят ультрафиолетовыми лампами с длиной волны 280-300 нм (сухие дрожжи) или в тонком слое 5 %-ной суспензии дрожжей. При более коротковолновом и длинноволновом излучении повышается выход других соединений стериновой природы. На выход витамина D2 (и образование других соединений) оказывают влияние длительность облучения, температура, наличие примесей. Поэтому облучение эргостерина, используемого в качестве пищевых добавок, производят с большой осторожностью. Промышленность СССР выпускает препарат под названием «Кормовые гидролизные дрожжи, обогащенные витамином D2». В 1 г абсолютно сухих дрожжей содержится 5000 ИЕ витамина D2, не менее 46 % сырого белка и незаменимые аминокислоты, в том числе лизин, метионин, триптофан. Для получения кристаллического витамина D2 дрожжи или мицелий грибов подвергают гидролизу раствором соляной кислоты при 110 °С. Гидролизованную массу обрабатывают спиртом при 75-78 °С и после охлаждения до 10-15 °С фильтруют. Фильтрат упаривают до содержания в нем 50 % сухих веществ и используют как концентрат витаминов группы В. Витамин D2 получают из массы, оставшейся после фильтрации. Массу промывают, сушат, размельчают и дважды обрабатывают при 78 °С трехкратным объемом спирта. Спиртовые экстракты сгущают до 70 %-ного содержания сухих веществ. Таким образом получают липидный концентрат. Его омыляют раствором NaOH, а стерины остаются в неомыленной фракции. Кристаллы эргостерина выпадают из раствора при 0°. Очистку кристаллов проводят путем перекристаллизации, последовательным промыванием 69 %-ным спиртом, смесью спирта и бензола (80:20) и повторной перекристаллизацией. Полученные кристаллы эргостерина сушат, растворяют в эфире, облучают, после чего эфир отгоняют, а раствор витамина концентрируют и кристаллизуют. 52 Для получения масляного концентрата раствор витамина после фильтрации разбавляют маслом до стандартного уровня. Источником получения эргостерина может служить мицелий грибов, остающийся как отход антибиотической промышленности и производства лимонной кислоты. В Советском Союзе планируется промышленное получение эргостерина из липидной фракции, использующей н-алканы. Сухую массу дрожжей для извлечения остаточных углеводородов экстрагируют петролейным эфиром. Получаемая при этом липидная фракция (микробный жир) является побочным продуктом микробиологической промышленности. Из микробного жира выделяют эргостерин, убихинон-9 и другие жирорастворимые соединения. Обогащенные эргостерином, облученные ультрафиолетовым излучением дрожжи используют в животноводстве как кормовую добавку. Эргостерин - исходный продукт для получения некоторых стероидных гормонов, лечебных и пищевых препаратов. Количество производимого пока эргостерина недостаточно для нужд народного хозяйства и внедрение новых производственных мощностей - задача ближайшего будущего. Получение каратиноидов. Каротиноиды получают с помощью химического синтеза и путем выделения из природных источников растений и микроорганизмов. Химическим путем получают β-каротин, витамин А, β-апо-8-каротиналь, этиловый эфир β-апо-8-каротиновой кислоты, кантоксантин и ряд других каротиноидов, синтез которых осуществляется в заводских масштабах. Традиционными источниками получения каротиноидов служат также некоторые растения (морковь, тыква, трава, шиповник, облепиха и др.). Наряду с этим все шире в тех же целях используют мицелиальные грибы и дрожжи. Как продуценты каротиноидов представляют также интерес бактерии и водоросли. Перспективными в данном отношении являются некоторые фототрофные бактерии, у которых в зависимости от интенсивности света можно регулировать выход каротиноидов. Биомассу пурпурных бактерий, богатую каротиноидами, в Японии используют в качестве добавок в рацион кур, что способствует более интенсивному окрашиванию желтка. Каротиноиды могут быть получены также в значительном количестве из некоторых водорослей (например, Spongiococcus excentricum, Chlorella sp.). Среди хемотрофов для получения каротиноидов используют дрожжи Rhodotorula gracilis, R. rubra, Rhodosporidium diobovatum, а также актиномицеты (Act. chrestomycetes var. aurantioideus, Act. chrysomallus var. carotinoides), микобактерии (Mycobacterium phlei, M. carotenum), грибы (Mucoraceae, Dacrymycetaceae и др.). Интерес представляют некоторые штаммы Flavobacterium, синтезирующие пигмент зеаксантин, который пока еще не может быть получен с помощью химического синтеза. 53 Продуцентами β-каротина, широко применяемыми для промышленного получения этого пигмента, являются гетероталличные мукоровые грибы Blakeslea trispora и Choanephora conjuncta. При совместном культивировании разнополых штаммов этих грибов на специально подобранных средах выход каротина составляет около 3-4 г/л среды. Для получения β-каротина с помощью В. trispora используют сложные по составу среды, например кукурузно-соевую, содержащую растительные масла, керосин, поверхностно-активные вещества и некоторые специальные стимуляторы. В последние годы в целях экономии для получения β-каротина начинают применять вторичные продукты отхода - кукурузный экстракт и гидрол. В качестве стимуляторов синтеза каротина используют β-ионон, который можно заменить более дешевой цитрусовой пульпой и цитрусовой мелассой. Как заменители β-ионона используют также изопреновые димеры или тримеры, а также циклогексан, циклогексанон и их 1 триметилпроизводные, среди которых наиболее эффективен 2,6,6 -триметил1-ацетилциклогексан (ТАЦ). Активаторами каротиногенеза у В. trispora могут быть также α-пирролидон, сукцинимид, нембутал и изониазид. Добавление этих активаторов, особенно последнего, на фоне действия βионона или ТАЦ позволяет значительно увеличить выход каротиноидов. Стимуляторы добавляются к культуре продуцента после окончания периода интенсивного роста биомассы. Процесс получения β-каротина при использовании гриба В. trispora многостадиен. Согласно одному из способов сначала выращивают отдельно (+)- и (-)-штаммы гриба. Следующая стадия - совместное выращивание разнополых штаммов в ферментере при 26 °С и достаточно интенсивной аэрации. Третья стадия выращивания - внесение в большой ферментер смешанной культуры В. trispora и инкубация в течение 6-7 сут. при той же температуре и аэрации. Используя соответствующие стимуляторы, можно не только значительно увеличить выход β-каротина, но и изменить состав каротиноидов у В. trispora. Под влиянием некоторых производных пиридина (2-аминопиридина, 4-аминопиридина) вместо β-каротина преобладающим пигментом становится ликопин, выход которого может составлять более 60 % от всех каротиноидов, синтезированных В. trispora. При добавлении 4аминопиридина наряду с ликопином образуется γ-каротин, причем оба каротиноида синтезируются почти в равных количествах. Таким образом, использование В. trispora интересно в практическом отношении не только потому, что это один из самых активных продуцентов β-каротина, но и потому, что с помощью этого организма можно получать и другие каротиноиды (γ-каротин, ликопин, α-каротин, β-зеакаротин), также имеющие практическое применение. Проводятся исследования, направленные на дальнейшее удешевление стоимости каротиноидов, получаемых микробиологическим способом. Показано, например, что синтез каротиноидов у В. trispora можно увеличить 54 почти в семь раз, если источником углерода в среде будет целлобиоза. Для удешевления производства с этой целью можно использовать отходы, остающиеся при производстве целлюлозных материалов. На таких средах В. trispora синтезирует кроме β-каротина еще и такой практически важный фермент, как β-глюкозидаза. Получение одновременно двух ценных продуктов значительно удешевляет производство β-каротина. Для практического использования предложен также высокопродуктивный мутант дрожжей Rhodosporidium diobovatum. На основе данного штамма получен каротинсодержащий белковый препарат. Разработан также метод получения каротинсодержащего препарата с помощью высокоактивного мутанта Муcоbacterium rubrum; препарат содержит α-, β- и γ-каротины, ликопин и ксантофиллы (лютеин, торулин и др.). Для получения ксантофиллов используют гриб Dacrymyces deliquescens, культивируемый на среде, содержащей глюкозу, глицерин и кукурузный экстракт при интенсивном освещении. Выход ксантофиллов в этом случае может составлять около 40 мг на 1 л среды. ГЛАВА 5. Производство органических кислот Органические кислоты широко используют в пищевой и фармацевтической промышленности, в технике и в качестве химического сырья. Отдельные органические кислоты (лимонную, яблочную) можно получать экстракцией из природного растительного сырья; другие (уксусную, молочную) - в процессах органического синтеза. Более 50 органических кислот могут быть получены на основе микробиологического синтеза. Биотехнологические методы их получения к настоящему времени детально разработаны. Более тогчэ, принято считать, что органические кислоты, полученные в результате микробиологического синтеза, для использования человеком предпочтительнее в сравнение с синтетическими кислотами. Для технических нужд органические, кислоты получают химическим путем; применяемые в пищевой и фармацевтической промышленности - в различных биотехнологических процессах. Это производства лимонной, молочной, уксусуной, итаконовой, пропионовой и глюконовой органических кислот; (молочная и уксусные кислоты производятся также и химическим путем). Органические кислоты в системе микробного метаболизма являются продуктами деградации источника энергии и углерода. Так, лимонная, изолимонная, кетоглутаровая, янтарная, фумаровая и яблочная кислоты интермедиа™ цикла трикарбоновых кислот у большинства аэробных микроорганизмов. Глюконовая, кетоглюконовая и винная кислоты промежуточные продукты прямого окисления глюкозы (без фосфорилирования) некоторых аэробных бактерий и грибов. Молочная, масляная и пропионовая кислоты являются конечными продуктами метаболизма углеводов у анаэробных бактерий. Уксусная кислота - продукт 55 окисления этанола; а алифатические моно и дикарбоновые кислоты промежуточные продукты окисления нормальных алканов. Таким образом, возможности микроорганизмов для получения на основе их метаболизма органических кислот велики. Для сверхсинтеза отдельных кислот нужны селективные, строго определенные условия. При сбалансированном росте микроорганизмов на полноценной среде накопления органических кислот не происходит, так как являясь промежуточными продуктами в^системе микробного метаболизма, органические кислоты - исходный материал для синтеза других макромолекул. Время максимальной скорости образования в клетке органических кислот, как и многих других метаболитов, не совпадает во времени со скоростью размножения клеток и накоплением биомассы. Сверхсинтез органических кислот наблюдается при торможении скорости роста продуцента и блокировании процессов биосинтеза, требующих участия кислот в качестве субстрата, то есть при нарушении процессов диссимиляции имеющегося эндогенного субстрата и процессов синтеза основных (азотсодержащих) компонентов клетки. Такими условиями, как правило, является полное или избыточное содержание в среде источника углерода и энергии и дефицит биогенных элементов, ограничивающих рост клеток. Большинство органических кислот получают, лимитируя рост клетокпродуцентов дефицитом азота или фосфора при избытке углеродсодержащего субстрата. Поэтому микробиологические процессы получения органических кислот - двухфазные. На первом этапе происходит так называемый сбалансированный рост при максимальном накоплении биомассы и потреблении углеродного и энергетического субстрата, а также лимитирующего биогена; на втором - происходит замедление скорости роста клеток. В результате этого прирост биомассы прекращается и начинается интенсивное кислотообразование. Длительность фазы интенсивного кислотообразования определяется наличием углеродсодержащего субстрата в среде. Важным условием кислотообразование большинства органических кислот (за исключением молочной) является хороший режим аэрации, а также величина рН среды. Способность продуцировать ту или иную кислоту - широко распространенное среди микроорганизмов свойство. В качестве производственных культур используют специально подобранные штаммы, продуцирующие целевую кислоту в виде монопродукта с высокими выходами и эффективным усвоением углеродного субстрата. При многих производствах органических кислот экономический коэффициент по углероду достигает 90 % и выше. В качестве продуцентов используют бактериальные, дрожжевые и грибные культуры (Lactobacillus, Arthrobacter, Alcaligenes, Candida, Aspergillus, Penicillium, Trichoderma). Способы ферментации в микробиологических процессах производства органических кислот - разнообразны. Среди них - поверхностные жидко и твердофазные процессы, а также глубинные, включая проточные культуры. В последние годы разработаны принципиально новые и эффективные биотехнологии с 56 использованием иммобилизованных целых клеток и ферментов. Также разнообразны и субстраты, используемые в производстве органических кислот. Применяемые в начале века глюкоза и сахароза со временем стали заменять более доступными комплексными средами (мелассой, гидролизным крахмалом); в 60е годы были разработаны новые процессы получения органических кислот на жидких парафинах нефти. В настоящее время биотехнологическими способами получают в промышленных масштабах ряд органических кислот. Из них лимонную, глюконовую, кетоглюконовую и итаконовую кислоты получают лишь микробиологическим способом, молочную, салициловую и уксусную – как химическим, так и микробиологическим, яблочную – химическим и энзиматическим путем. Уксусную кислоту продуцируют Aсеtobacter и Gluconobacter, лимонную – Aspergillus niger, Aspergillus wentii, молочную – Lactobacillus delbrueckii. В качестве сырья для получения пищевого уксуса используют виноградное вино, пивное сусло, мед, соки различных фруктов и ягод после спиртового брожения или водный раствор этилового спирта для получения белого уксуса. Кроме спирта среда содержит уксусную кислоту и минеральные соли, в состав которых входят азот, фосфор, сера, марганец, калий. Иногда добавляют источники витаминов в виде различных экстрактов. Спирт служит источником углерода и энергии для бактерий. Уксусная кислота стала первым микробиологическим продуктом, полученным с помощью иммобилизованных клеток. Этот способ может быть непрерывным и периодическим. В течение длительного времени применяется адсорбирование уксуснокислых бактерий на древесной стружке, древесном угле, коксе и других субстратах, Пропуская раствор этанола через генераторы с иммобилизованными бактериями, получают 10-15%-ный раствор уксусной кислоты. При этом из 100 л безводного спирта теоретически должно быть получено 103 л уксусной кислоты. На практике выход уксуса из 100 л этанола редко превышает 90 л, что связано с переокислением и неполным окислением этанола бактериями, а также с его испарением. Уксус, полученный при брожении, имеет приятные аромат и вкус, которые обусловливают побочные продукты брожения: сложные эфиры (этилацетат и другие), высшие спирты, органические кислоты. В столовом уксусе содержится от 5 до 9 % уксусной кислоты. Уксус с концентрацией кислоты от 20 до 30 % получают путем вымораживания исходного раствора. Путем перегонки получают 70-80%-ную уксусную кислоту, называемую уксусной эссенцией. Ледяная уксусная кислота содержит от 98,0 до 99,8 % кислоты. Уксусную кислоту или уксус широко используют в пищевой промышленности. Уксус, полученный микробиологическим путем (пищевая уксусная кислота, столовый уксус), различается по сортам в зависимости от 57 характера сбраживаемого субстрата. Известен яблочный, виноградный, грушевый и другие сорта уксуса. Уксус также применяют для растворения органических красителей, при получении медикаментов, пластмасс и т.д. Лимонная кислота широко распространена в природе, относительно много ее содержится в некоторых ягодах, фруктах, особенно в цитрусовых (в лимоне от 5 до 10 %), в листьях и стеблях некоторых растений. Ранее лимонную кислоту выделяли в виде лимоннокислого кальция из продуктов переработки листьев хлопчатника, стеблей махорки, хвои ели и в значительных количествах из плодов лимонов. Однако это производство является крайне дорогим и небольшим по объему. Поэтому лимонная кислота была дефицитным и дорогим продуктом. В настоящее время лимонная кислота по объему производства является одним из главных продуктов микробного синтеза, ее общий выпуск в разных странах достигает 400 тыс. тонн в год. Для получения лимонной кислоты используют микроскопические грибы родов Aspergillus, Penicillium, Mucor, Ustina и др. В настоящее время основными продуцентами лимонной кислоты являются различные штаммы гриба Aspergillus niger. Сверхсинтез лимонной кислоты происходит при лимитировании роста грибов-продуцентов минеральными компонентами среды и одновременном избыточном содержании источника углерода. В условиях лимитирования роста гриба недостатком железа и марганца после полного поглощения из среды дефицитного элемента он прекращает расти, однако продолжает потреблять имеющийся в среде источник углерода. При этом в клетках гриба начинает накапливаться лимонная кислота, которая в дальнейшем выделяется в среду. Посевной материал в виде спор (конидий) выращивают на мелассной среде поверхностным или глубинным способом. Лимонную кислоту выделяют из культуральной жидкости в виде плохо растворимой соли – цитрата кальция, которая образуется при добавлении мела. Перевод лимонной кислоты в свободное состояние достигается при добавлении строго определенного количества серной кислоты: Цитрат кальция + Н2SO4 = Цитрат + Гипс. Гипс удаляют фильтрованием. Раствор лимонной кислоты осветляют активным углем, упаривают, кристаллизуют. Лимонная кислота используется в кондитерской промышленности для подкисления карамели, пастилы, вафель, так как она хорошо подчеркивает фруктовый вкус. Данную органическую кислоту в целях подкисления добавляют в мороженое, пищевые концентраты, маргарин, некоторые сорта колбас и сыра. Лимонную кислоту применяют для торможения образования меланоидинов в сгущенном молоке с сахаром, раствором ее промывают и 58 дезодорируют жировое сырье, обрабатывают перед холодным хранением свежее мясо, рыбу, фрукты с целью стабилизации их цвета, вкуса и запаха. Соли лимонной кислоты используют для изготовления шампуней и других моющих средств, так как они стимулируют вспенивание и обеспечивают механическую устойчивость пен. Молочная кислота с 1881 г. производится промышленным способом с помощью молочнокислых бактерий. Для промышленного изготовления молочной кислоты пригодны только гомоферментативные молочнокислые бактерии, образующие до 98 % молочной кислоты. Применяются штаммы Lactobacillus delbrueckii, Lactobacillus bulgaricus. Молочнокислые бактерии преобразуют в молочную кислоту самые разные углеводы, поэтому для промышленного получения этой кислоты используют мелассу, молочную сыворотку, глюкозу, мальтозу, сахарозу, лактозу, осахаренный крахмал и пр. Молочную кислоту в промышленных условиях получают методом анаэробной глубинной ферментации. Во время ферментации рН среды поддерживают, добавляя мел. Через 6-7 суток культивирования в среде остается от 0,5 до 0,1 % сахаров и от 11 до 14 % лактата кальция. Из 100 г сахаров получают от 80 до 90 г лактата кальция. Осадок мела и коллоиды отделяют фильтрацией. Фильтрат упаривают, охлаждают и кристаллизуют. Кристаллы лактата отделяют центрифугированием. Молочную кислоту из лактата получают разложением серной кислотой. Реакция идет при температуре от 60 до 70 °С в соответствии с уравнением: Ca(C3H5O3)2 + H2SO4 2C3H6O3 + CaSO4 Молочную кислоту обрабатывают активированным углем, фильтруют и фасуют. Конечный продукт – в виде жидкого концентрата молочной кислоты. Молочную кислоту применяют для приготовления джемов, в которых она способствует хорошей консистенции. Молочная кислота как регулятор рН, улучшитель вкуса применяется в производстве многих сыров, квашении капусты, в сухом концентрате кваса. В хлебобулочном производстве молочная кислота и лактаты увеличивают объем мякиша и улучшают корку хлеба при использовании муки низкого качества. Способность лактатов удерживать влагу применяют в производстве колбас, сыров, детского питания. Молочную кислоту также используют для ускорения получения молочно-белкового сгустка при производстве творога. Получение лимонной кислоты. Лимонная кислота - трехосновная оксикислота, широко распространенная в плодах и ягодах. Она широко применяется в пищевой промышленности при производстве кондитерских изделий и напитков, в фармацевтической, химической и текстильной 59 промышленности. Лимонная кислота была идентифицирована в качестве продукта метаболизма плесневых грибов в 1893 г. Вемером. В настоящее время это кислота по объемам производства (свыше 350 тыс.т/г) занимает первое место среди всех органических кислот. У микроорганизмов синтез лимонной кислоты реализуется в цикле дикарбоновых кислот и осуществляется в результате конденсации кислоты с четырьмя атомами углерода и двумя карбоксильными группами и кислоты с одной карбоксильной группой. Образуемая в результате гликолиза пировиноградная кислота связывается с углекислотой; синтезируемая при этом щавелевоуксусная кислота реагирует с уксусной кислотой с образованием лимонной кислоты, то есть образование лимонной кислоты включает реакции гликолиза и ряд реакций цикла Кребса. При каждом обороте цикла молекула щавелевоуксусной кислоты взаимодействует с уксусной, образуя лимонную кислоту. Производство лимонной кислоты методом ферментации плесневых грибов принадлежит к числу давних биотехнологических процессов. Первое производство было реализовано в конце XIX века. Совершенствование процесса получения лимонной кислоты тесно связано с разработкой многих фундаментальных аспектов микробиологии (борьбой с микробным загрязнением производственной культуры, оптимизацией состава питательных сред, селекцией высокопродуктивных штаммов и др.). В промышленном производстве лимонной кислоты в качестве продуцента в основном используют Aspergillus niger, но также применяют и A. wentii. Процесс ферментации достаточно сложен, так как лимонная кислота, является продуктом первичного метаболизма грибов, и даже незначительное выделение данного продукта в окружающую среду свидетельствует о выраженном дисбалансе клеточного метаболизма. Рост продуцента и синтез кислоты обычно регулируют составом среды (сахара, Р, Mn, Sе, Zn). Сверхсинтез лимонной кислоты реализуется при больших концентрациях Сахаров в среде (14-24 %) и является ответной реакцией продуцента на дефицит фосфора, а также других металлов, хотя их роль до конца не ясна. Это, видимо, и подавление анаболизма, и влияние на свойства поверхности и морфологию гиф. Оптимум рН на стадии кислотообразования составляет 1,7-2,0. В более щелочной среде процесс сдвигается в сторону накопления щавелевой и глюконовой кислот. В качестве основы среды обычно используют глюкозный сироп, гидролизаты крахмала или мелассу. Последнюю предварительно разбавляют до требуемого уровня Сахаров и обрабатывают с целью снижения содержания металлов. Источником азота служат соли аммония (0,2 %); концентрация фосфатов (0,01-0,2 %). В качестве пеногасителей используют природные масла с высоким содержанием жирных кислот. Очень существенное значение имеет уровень аэрации культуры. В производстве лимонной кислоты применяют несколько вариантов процесса. Поверхностный способ реализуется на твердой сыпучей среде и в 60 жидкой фазе. При жидкофазной поверхностной ферментации питательную среду разливают в кюветы слоем от 8 до 18 см. Кюветы размещают на стеллажах в предварительно простерилизованной парами формалина бродильной камере. Через специальные воздуховоды с током стерильного воздуха поверхность среды засевают исходной музейной культурой. В качестве посевного материала используют предварительно полученные также в условиях поверхностной культуры и высушенные споры (конидии) из расчета 50-75 мг конидий на 1 м2 площади кювет. Известно несколько вариантов процесса: бессменный, бессменный с доливами и метод пленок. При бессменном режиме процесс осуществляется на одной среде от момента засева спор до завершения стадии кислотообразования. При использовании метода пленок через 7 суток после завершения кислотообразования сброженный раствор мелассы сливают из кювет, мицелий промывают стерильной водой; и в кюветы заливают новую среду. Бессменный способ с доливом характеризуется дробными добавками мелассы под пленку гриба на стадии кислотообразования (30-35 % от исходного объема), так называемый режим с подпиткой субстратом. Это позволяет повысить выход лимонной кислоты на 15-20 % с единицы поверхности при сокращении затрат Сахаров на 10-15 % по сравнению с другими методами. В ходе стадии ферментации на первом этапе (первые 2436 ч) происходит интенсивный рост мицелия. Температура среды в этот период стабилизируется на уровне 32-34°С, интенсивность аэрации составляет 3-4 м3 воздуха в ч/м мицелия. В период активного кислотообразования подачу воздуха увеличивают в 5-6 раз. В результате более интенсивного термогенеза температуру снижают до 30-32°С. По мере снижения процесса, кислотообразования режим аэрации становится менее интенсивным. Контроль процесса ведут по показателям титруемой кислотности среды. Процесс считают завершенным при остаточной концентрации Сахаров около 1-2 % и уровне титруемой кислотности 12-20%. Содержание лимонной кислоты от уровня всех кислот достигает 94-98 %. Сброженный раствор сливают в сборник и направляют на обработку; промытый мицелий используют в кормопроизводстве. Твердофазная ферментация имеет много общего с поверхностножидкофазным процессом. Разработанный в Японии процесс Коджи предусматривает использование в качестве среды пористого материла (багасса, картофель, пульпа сахарной свеклы, пшеничные отруби). Материал предварительно стерилизуют, после охлаждения инокулируют суспензией спор. Ферментация происходит в лотках при 25-30°С в течение 6-7 дней. Образованную лимонную кислоту экстрагируют водой. В Японии 20% общего объема производства лимонной кислоты получают методом Коджи. Начиная с 1950 г., промышленные процессы получения лимонной кислоты стали переводить в условия глубинной культуры. Стабильный процесс возможен при его организации в две стадии: рост мицелия на полной среде в ходе первой стадии и на второй (при отсутствии фосфора в среде)61 образование лимонной кислоты. Глубинная ферментация проводится в аппаратах емкостью 50 м с заполнением на 70-75 %. В качестве посевного материала используют мицелий, подрощенный также в условиях глубинной культуры. В производственном аппарате, куда подрощенный мицелий передается по стерильной посевной линии, питательная среда содержит 12-15 % Сахаров. Ферментацию проводят при 31-32°С при непрерывном перемешивании. В ходе процесса кислотообразования (5-7 суток) реализуют интенсивный режим аэрации (до 800-1000 мЗ/ч) с дробным добавлением Сахаров, 2-3 подкормки. Выход лимонной кислоты составляет от 5 до 12 %, остаточная концентрация Сахаров - 0.2-1.5 %, доля цитрата - 80-98 % от суммы всех органических кислот. В 60е годы начали разрабатывать процессы получения лимонной кислоты на основе жидких углеводородов (С9-С30) с использованием в качестве продуцентов дрожжей (Candida) и бактерий (Brevibacterium, Corynebacterium, Arthrobacter), а также с применением метода проточных культур. Эти технологии, пока не реализованные в промышленных масштабах, обещают в будущем определенные технологические перспективы. Готовый продукт - высокоочищенную кристаллическую лимонную кислоту получают в ходе постферментационной стадии. В сброженных растворах содержатся, помимо целевой кислоты, также глюконовая и щавелевая кислоты, остатки несброженных Сахаров и минеральные соли. Для выделения лимонной кислоты из данного раствора ее связывают гидроокисью кальция с образованием трудно растворимого цитрата кальция: 2 С6Н807 + 3 Са(ОН)2 = Са3(С6Н507)2 + 6 Н20. Одновременно образуются кальциевые соли глюконовой и щавелевой кислот, глюкоыат кальция СаССбНпСЬЪ и оксалат кальция СаС204. Кальциевые соли лимонной и щавелевой кислот выпадают в осадок, а глюконат кальция и основная часть органических и минеральных компонентов мелассы остаются в растворе. Осадок отделяется на вакуумфильтре, промывается и высушивается. Далее для перевода лимонной кислоты в свободное состояние и освобождения от оксалата кальция осадок обрабатывают серной кислотой с последующей фильтрацией. Раствор лимонной кислоты фильтруют, концентрируют вакуум-выпаркой и затем подвергают кристаллизации при медленном охлаждении до 8-10°С. Полученные кристаллы отделяют в центрифуге от маточника и высушивают в пневматических сушилках при 30-35°С. Готовый продукт содержит не менее 99,5 % лимонной кислоты (в пересчете на моногидрат), зольность - не выше 0,1-0,35%. Получение молочной кислоты. Молочная кислота - органическая одноосновная кислота, образуемая в результате анаэробного превращения углеводов молочнокислыми бактериями. В 1847 г. С. Блодно доказал, что данная кислота является продуктом брожения, а Л. Пастер установил, что этот процесс вызывают бактерии. Образование молочной кислоты из глюкозы 62 возможно несколькими путями. При сбраживании гомоферментными молочнокислыми бактериями: C3H1203 -> 2 СН2ОН2СНОНСНО (глицеральдегид) -» 2 СН3СОСНО (метилглиоксаль) + 2 Н20, СНзСОСНО (метилглиоксаль) + Н20 - СНЗСНОНСООН (молочная кислота). Второй путь, гетероферментный, включает распад глюкозы до пировиноградной кислоты и восстановление последней до молочной кислоты: С6Н,206 -> СНзСОСООН + Н2 -> СНзСНОНСООН. Для промышленного получения молочной кислоты используют гомоферментные молочнокислые бактерии. У гомоферментных молочнокислых бактерий только 3 % субстрата превращается в клеточный материал: а остальной - трансформируется в молочную кислоту, выход которой достигает до 1.5 %. Теоретически из 1 моля глюкозы должно образоваться 2 моля лактата. На практике эта величина несколько ниже, 1.8 моля, то есть выход продукта от субстрата достигает 90 %. Применяют молочную кислоту в пищевой промышленности для получения напитков, мармеладов, в процессах консервирования, а также в кормопроизводстве. Соли молочной кислоты используют в фармацевтике. Промышленное производство молочной кислоты начато в конце XIX века с участием молочнокислых бактерий Lactobacillus delbrueckii, L. leichmannii, L. bulgaricus. Молочнокислое брожение протекает в анаэробных условиях, однако лактобациллы относятся к факультативным анаэробам, поэтому при ферментации воздух полностью не удаляют из ферментеров. В качестве сырья используют сахарную и тростниковую мелассу игидролизаты крахмала, при этом концентрация Сахаров в исходной среде в зависимости от характера брожения составляет примерно от 5 до 20 %. Используют восстановленные формы азота, сульфаты или фосфаты аммония, а также солод и кукурузный экстракт в качестве источника факторов роста. Возможно использование сульфитного щелока с участием бактерий L. delbrueckii. Ферментацию проводят в глубинной культуре при рН 6,3-6,5 и строго постоянной температуре 50°С. Длительность процесса составляет до 7-11 суток. В ходе процесса брожения для коррекции изменяющегося рН в культуру вносят мел, 3-4 раза в течение суток. Конечная концентрация образующегося лактата кальция составляет 10-15 %, остаточная концентрация сахаров – 0,5-0,7 %. На стадии получения готового продукта культуральную среду нагревают до 80-90°С, затем нейтрализуют гашеной известью до слабощелочной реакции. После отстаивания в течение 3-5 ч взвешенные частицы декантируют. После этого раствор лактата кальция подают на фильтр-пресс. Фильтрат упаривают до концентрации 27-30 %, охлаждают до 25-30°С и подвергают кристаллизации. Промытый лактат кальция отделяют центрифугированием и подвергают расщеплению серной кислотой при 60-70°С. Сырую молочную кислоту 18-20 % концентрации 63 упаривают в несколько этапов в вакуум-выпарных аппаратах до 70 % концентрации. Отфильтрованную кислоту после фильтр-пресса подают на розлив с внесением небольших количеств мела, при этом около 10 % кислоты превращается в кристаллический лактат, который связывает молочную кислоту. Получение уксусной кислоты. Уксусная кислота - широко используется в пищевой, химической, микробиологической промышленности, в медицине. Получение уксусной кислоты из спиртосодержащих жидкостей было известно более 10 тыс. лет назад. В те времена древние греки и римляне использовали уксус в качестве освежающего напитка и получали, главным образом, оставляя вино открытым. В больших масштабах уксус долго получали в плоских открытых бочках, в которых пленка бактерий плавала на поверхности. В XIX веке поверхностные процессы стали заменять более эффективными. Так, был разработан процесс в струйном генераторе. В середине XX века появились глубинные процессы ферментации. Усовершенствованный генератор Фрингса используется в настоящее время. Уксуснокислое брожение основано на способности уксуснокислых бактерий окислять спирт кислородом воздуха с участием алкогольдегидрогеназы в уксусную кислоту: СН3СН2ОН + 02 -> СНзСООН + Н20, при этом из 1 моля этанола образуется моль уксусной кислоты, а из 1 л 12об.% спирта получается 12,4 весовых % уксусной кислоты. Данный процесс могут реализовать многие бактерии, но в промышленных технологиях для получения уксуса используют уксуснокислые бактерии рода Acetobacter, интерес представляют также бактерии Gluconobacter. Большую часть уксуса получают, используя разведенный спирт. В настоящее время процесс реализуют как поверхностным, так и глубинным способом. Поверхностный режим протекает в струйных генераторах, наполненных древесной стружкой, объемом до 60 м . Исходный питательный раствор с бактериями распыляют по поверхности стружек, ион стекает, собираясь в нижней части аппарата. После этого жидкость собирают и вновь закачивают в верхнюю часть аппарата. Процедуру повторяют 3-4 раза, в результате в течение Зх дней до 90 % спирта трансформируется в ацетат. Этот старый способ протекает более эффективно и равномерно в генераторах Фрингса с автоматическим поддержанием температуры и принудительной подачей воздуха. По такой технологии производят до 400 млн л уксусной кислоты в год. Современные промышленные процессы получения уксуса реализуют'в глубинной культуре в специальных аэрационных аппаратах с термостабилизацией и механической системой пеногашения. Скорость аэрации составляет 3,4 м3/ч., вращение ротора - 1500 об./мин., температура 30°С. Исходная инокулируемая смесь содержит этанол и уксусную кислоту, соответственно, около 5 и 7 %; конечная концентрация уксуса через 1,5 суток составляет 1213 %. Процесс - полупроточный, отливнодоливной. Каждые 30-35 часов до 64 60 % культуры заменяют на свежее сусло. При глубинной ферментации выход продукта на 1 м3 в 10 раз выше по сравнению с поверхностной ферментацией. К началу 90-х гг. таким способом производили до 715 млн. литров 10 % уксусной кислоты в год. Разработан и реализован эффективный непрерывный способ получения уксусной кислоты в батарее последовательно работающих ферментеров (обычно 5 аппаратов). Температура культивирования составляет 28°С для Acetobacter и 35°С при использовании в качестве продуцента культуры Bact. schutzenbachii. Наилучшим сырьем для процесса является этиловый спирт, полученный из зерно-картофельного сырья, при его концентрации около 10 %. Оптимум рН для развития бактерий - около 3. При увеличении содержания уксусной кислоты в культуре свыше 8 % рост бактерий замедляется, при 12-14 % прекращается. Поэтому процесс проводят в батарее последовательно соединенных аппаратов. Первый выполняет роль инокулятора, поэтому в него непрерывно подают свежую среду и поддерживают условия, оптимальные для быстрого образования биомассы бактерий. Культура из первого аппарата поступает во второй аппарат и далее в последующие, при этом транспортировка культуральной жидкости осуществляется воздухом. В каждом аппарате условия ферментации стабилизируются в соответствии с требованиями течения хода ферментации, при постепенном понижении температура среды от 28°С в первом аппарате до 25°С - в последнем. Режим аэрации также изменяется, от 0,4 до 0,15мЗ/мЗ мин. Концентрация спирта со второго по четвертый аппарат стабилизируется на требуемом уровне подачей в них среды с 40 % этанолом. Из последнего аппарата выводится культуральная жидкость с содержанием ацетата не ниже 9,0 и не выше 9,3 %. Выход кислоты составляет до 90 кг из 100 л безводного спирта. На постферментационной стадии после отделения бактериальной биомассы раствор уксуса фильтруют, освобождая от окрашенных и взвешенных частиц, и далее подвергают пастеризации. Для повышения концентрации исходные растворы вымораживают до 20-30 %. Дальнейшее концентрирование до получения ледяной уксусной кислоты (98,0 – 99,8 %),проводят методом перегонки. Получение пропионовой кислоты. Пропионовая кислота синтезируется грамположительными пропионовокислыми бактериями (Propionibacterium), используется в химикофармацевтической промышленности, при получении косметических средств, в качестве фунгицида для сохранения зерна. Химизм образования пропионовой кислоты заключается в следующем: пировиноградная кислота при участии биотина и углекислоты карбоксилируется в щавелевоуксусную, которая через яблочную и фумаровую кислоты восстанавливается до янтарной кислоты. Янтарная кислота при участии АТФ и КоА превращается в сукцинил-КоА, последний под воздействием метилмалонилКоА-изомеразы и при участии кофермента В И превращаетсяв метилмалонил-КоА. В результате карбоксилирования метилмалонил-КоА расщепляется с образованием 65 свободного КоА и пропионовой кислоты. Среди промышленных штаммов продуцентов - бактерии Pr. Arabinosum, Pr. shermanii, Pr. rubrum и др. В качестве субстрата брожения бактерии используют различные сахара (лактозу, глюкозу, мальтозу, сахарозу, органические кислоты - яблочную и молочную). Получают пропионовую кислоту в глубиной аэробной культуре на средах, содержащих (%): сахара 2, органический азот 0.4 (источник дрожжевой экстракт), соли молочной кислоты. Процесс реализуется за 12 суток при 30°С и рН 6,8-7,2; при этомсвыше 70 % Сахаров трансформируется в органические кислоты, на образование углекислоты расходуется менее 20 % углеродного субстрата. Получение итаконовой кислоты. Итаконовая кислота - ненасыщенная двухосновная кислота; ее образование плесневыми грибами открыл в 1931 г. Киношита. Данная кислота - важный промежуточный продукт для получения полимеров. Итаконовая кислота образует сополимеры с эфирами и другими мономерами, поэтому используется при производстве синтетических волокон и смол, ряда адгезивных средств, ПАВ, красителей и других сложных органических соединений. Синтез итаконовой кислоты связан с реакциями цикла Кребса; ее исходным продуктом является цисаконитовая кислота, которая при декарбоксилировании в результате перемещения электронов и переходадвойной связи из положения 2,3 в положение 3,4 превращается в итакановую кислоту. Получение итаконовой кислоты осуществляют поверхностным и глубинным методами ферментации. В качестве продуцентов используют отселектированиые грибные штаммы (Aspergillus itaconicus, Asp. terreus).Процесс аналогичен процессам получения лимонной кислоты. Среды содержат высокие концентрации Сахаров, обычно используют мелассу, при дефиците фосфора и железа. Особенностью процесса получения данно й кислоты является высокая потребность продуцента в солях цинка, магния и меди. При поверхностной ферментации в течение 10-12 суток образуется около 60 % продукта в пересчете на сахар, доля целевой кислоты в смеси (синтезируются также янтарная, щавелевая и фумаровая кислоты) -свыше 90 %. Содержание итаконовой кислоты достигает 15-20 %, остаточная концентрация Сахаров не превышает 0,6 %. В отличие от лимонной, итаконовая кислота - токсичный продукт, при ее концентрации около7 % рост продуцента угнетается, и скорость продукции кислоты снижается. Токсичность итаконовой кислоты нейтрализуют дробными добавками гидроксиаммония, рН среды при этом стабилизируется на уровне 3,5-3,8. При глубинной ферментации конечная концентрация итаконовой кислоты ниже, 4-6 %. Товарный продукт - кристаллическая итаконовая кислота92 % содержания, остальное - влага (3-6 %) и другие кислоты (1-3 %). Получение глюконовой кислоты Глюконовая кислота - одноосновная пентокислота, получаемая при ферментативном окислении глюкозы с 66 участием глюкозооксидазы. Глюконовая кислота имеет много областей применения. Комплексообразователь с металлами - глюконат натрия, применяют при производстве моющих средств; кальций, железо и калийные соли глюконовой кислоты широко используют в медицине и пищевой промышленности. Продуценты глюконовой кислоты - грибы (Penicillium, Aspergillus). Ферментацию в промышленных масштабах осуществляют поверхностыми глубинным способами; используют среды с высоким (до 30-35 %) содержанием глюкозы, в составе сред - сульфат магния, фосфат калия, источник азота, а также углекислый кальций. Процесс завершается при остаточной концентрации сахара около 1 %. Готовый продукт кристаллические соли - глюконаты. Получение фумаровой кислоты Фумаровая кислота используется при производстве синтетических смол, красок, лаков. Смолы фумаровой кислоты применяют для производства печатных красок. Магниевые и натриевые соли фумаровой кислоты используют в медицине. Фумаровая кислота - метаболит цикла трикарбоновых кислот и присутствует во всех живых клетках, однако редко экскретируется в среду. Продуцентом данной кислоты являются различные грибы (Penicillium, Aspergillus, Rhizopus), последние наиболее активны. Среды для получения фумаровой кислоты содержат глюкозу в концентрации 5-10 %, лимитирующий фактор- азот, цинк. Ферментация реализуется в условиях интенсивной аэрации поверхностным или глубинным способом. При этом в ходе ферментации проводят нейтрализацию среды углекислым кальцием или раствором щелочи. Максимальный выход кислоты - 58 % от потребленной глюкозы. Биотехнологические методы получения, органических кислот совершенствуются. Недавно в Японии разработан способ получения 2кетоглюконовой кислоты на основе биосинтеза бактерий Pseudomonas,Bbixofl кислоты достигает 90 % от использованного сахара. Разработана технология получения щавелевой кислоты на средах с сахарами на основе грибов А. ozyzae. На основе селектированных штаммов дрожжей (Candidalipolytica) созданы технологии получения лимонной и изолимонной кислот. Специально отселектированные штаммы дрожжей рода Candida синтезируют на средах с нормальными парафинами фумаровую, яблочную,янтарную кислоты. Процесс на данном сырье постоянного состава более стабилен, чем на комплексных природных средах на основе мелассы; также упрощается стадия выделения и очистки готового продукта. ГЛАВА 6. Источники получения липидов и основные способы их выделения. Липиды - большая группа природных веществ, разнообразных по химической структуре и физико-химическим свойствам. Имеется несколько трактовок понятия липиды и различных схем их классификации, основанных 67 на свойствах этих веществ. Общее свойство липидных соединений способность растворяться в эфире, хлороформе и других органических растворителях (но не в воде). Липиды по строению можно подразделить на две большие группы: 1. Простые липиды, или нейтральные жиры, представленные у большинства организмов ацилглицеринами, т. е. глицериновыми эфирами жирных кислот (свободные жирные кислоты встречаются в клетках лишь как минорный компонент). 2. Сложные липиды, к которым относятся липиды, содержащие фосфорную кислоту в моно- или диэфирной связи, - это фосфолипиды, в число которых входят глицерофосфолипиды и сфинголипиды. К сложным липидам относятся соединения, связанные гликозидной связью с одним или несколькими остатками моносахаридов, или гликолипиды, а также соединения стероидной и изопреноидной природы, в том числе каротиноиды. До 20-х годов нашего столетия липиды, особенно нейтральные, рассматривались лишь как запасной материал, который возможно без особого ущерба для жизнедеятельности организма заменить другими, равными по калорийности веществами. Первое доказательство того, что липиды содержат физиологически необходимые для высших животных соединения, получено в 1926 г. голландскими исследователями Ивансом и Буром. Несколько позднее было установлено, что этими соединениями являются полиненасыщенные жирные кислоты (линолевая, линоленовая и арахидоновая) - физиологически необходимые для большинства живых организмов (витамин F). В дальнейшем было установлено, что и в клетках микроорганизмов липиды выполняют самые различные биологические функции. Они входят в состав таких ответственных структур, как клеточная мембрана, митохондрии, хлоропласты и другие органеллы. Липопротеиновые комплексы играют важную роль в процессах метаболизма. С ними в значительной мере связаны активный перенос различных веществ через пограничные мембраны и распределение этих веществ внутри клетки. С составом липидов во многом связаны такие свойства организмов, как термотолерантность и термофильность, психрофильность, кислотоустойчивость, вирулентность, устойчивость к ионизирующей радиации и другие признаки. Кроме того, липиды могут выполнять функцию запасных продуктов. К таковым относятся поли-β-гидроксимасляная кислота, образуемая многими бактериями, и ацилглицерины, в частности триацилглицерин, накапливаемый в больших количествах некоторыми дрожжами и другими представителями грибов. 6.1. Промышленное получение липидов. Вопросам промышленного получения липидов с помощью микроорганизмов уделяется пристальное внимание как в нашей стране, так и за рубежом. Микроорганизмы можно использовать для получения 68 фосфолипидов, гликолипидов, незаменимых жирных кислот и препаратов на их основе, необходимых для использования в медицинской практике, сельском хозяйстве, пищевой и других отраслях промышленности. Ряд дрожжей и мицелиальных грибов рассматривается как потенциальные продуценты липидов, в том числе липидов - аналогов некоторым типам растительных масел. Мировая практика пока не имеет производств с целевым назначением получать микробные липиды. Однако изменение конъюнктуры на мировом рынке не исключает целесообразности организации получения липидов путем микробиологического синтеза. В настоящее время в небольших объемах получают липиды только с помощью дрожжей, причем липиды являются побочным продуктом основного производства (при получении белково-витаминных концентратов на углеводородах нефти). Получение липидов из мицелиальных грибов, а также бактерий, водорослей и простейших пока не вышло за рамки лабораторных исследований. Одной из причин медленного решения вопросов получения бактериальных липидов следует признать наличие в их составе соединений, токсичных для макроорганизма. С помощью дрожжей возможно получение липидов на различных субстратах: гидролизатах растительного сырья, сульфитных щелоках, углеводородах нефти и др. Эффективность производства дрожжевого жира во многом связана с количеством основого сырья, необходимого для получения определенной единицы массы дрожжей, и его стоимостью. Кроме того, сырье, на базе которого готовится питательный субстрат для выращивания дрожжей, должно обеспечивать получение липидов, отвечающих требованиям, предъявляемым промышленностью, перерабатывающей липиды в различные продукты. Наиболее отработаны технологические схемы получения липидов с помощью дрожжей на гидролизатах верхового торфа малой степени разложения и углеводородах нефти. Эти схемы различаются тем, что при получении липидов на гидролизатах торфа дрожжевой жир является основным продуктом, а при использовании углеводородов дрожжевой жир побочный продукт, появляющийся в результате очистки дрожжевой биомассы от остаточных углеводородов. В связи с этим и фракционный состав получаемых этими путями липидов весьма различен: доминирующая фракция углеводородных дрожжей - фосфолипиды, основная фракция при получении липидов на гидролизатах торфа - триацилглицерины. В нашей стране процесс получения дрожжевых липидов в условиях специализированной установки осуществлен на Кстовском опытнопромышленном заводе белково-витаминных концентратов, вырабатывающем сотни тонн этого продукта биосинтеза. В ближайшие годы планируется ввод в строй нескольких установок по получению липидов из дрожжей способом, аналогичным кстовскому. Процесс получения липидов на гидролизатах верхового торфа малой степени разложения включает несколько основных операций: получение 69 гидролизата торфа, отдувка фурфурола и нейтрализация гидролизата до рН 5,5-6,0, введение в гидролизат минеральных источников питания, выращивание дрожжей - продуцентов липидов, отделение биомассы и экстракция из нее липидов. Следовательно, весь процесс аналогичен процессу получения кормовых дрожжей, за исключением дополнительных операций, связанных с извлечением липидов. Система растворителей, применяемая для этой цели, идентична используемым в масло-жировой промышленности. Оставшаяся после экстракции липидов биомасса «биошрот» может быть использована в кормлении сельскохозяйственных животных. Продуцентами липидов на гидролизатах торфа являются выделенные в институте микробиологии АН БССР штаммы Lipomyces lipoferus, биомасса которых содержит до 40 % липидов и более. Из одной тонны абсолютно сухого торфа можно получить 50-70 кг дрожжевых липидов, содержащих до 70-75 % триацилглицеринов. Кроме гидролизатов торфа для культивирования липидообразующих дрожжей и получения липидов по указанной выше схеме могут быть использованы другие гидролизные среды, например, гидролизаты древесины, или смешанные субстраты древесины и торфа. 6.2. Практическое использование липидов. Многочисленными экспериментами показано, что дрожжевые липиды и продукты их переработки могут использоваться в самых различных отраслях народного хозяйства: в текстильной, керамической, кожевенной, металлообрабатывающей (прокат стального листа, протяжка проволоки, лужение жести) промышленностях. Дрожжевые липиды могут быть использованы также при производстве каучука, резины, фармацевтических препаратов, косметики, мыла, олиф, в процессах флотации руд и др. Наконец, как показали эксперименты, дрожжевые липиды могут найти большое применение в кормлении сельскохозяйственных животных и птиц. В этом случае из схемы производства липидов исключается процесс их экстракции из клеток - для кормовых целей используется богатая жиром биомасса микроорганизмов. После второй мировой войны значительное число работ было направлено на изыскание возможности получения микробных липидов для пищевых целей. Шведский исследователь Лундин показал, что богатый физиологически необходимыми жирными кислотами дрожжевой жир (Rhodotocula gracilis) может с успехом использоваться кроме технических и на пищевые нужды. Рацион из 25 г жировых дрожжей может обеспечить организм человека 10 г липидов, 6 г белка и многими другими необходимыми веществами, что на 20 % удовлетворяет дневную потребность в этих соединениях. Производство микробного жира для пищевых целей уже имело место в Германии во время первой мировой войны. В качестве питательной среды 70 использовали мелассу или другие сахарсодержащие субстраты, продуцентом служил дрожжеподобный гриб Endomycopsis vernalis. В пищу использовали богатую жиром биомассу, из которой готовили пасту, известную под названием «Эвернал» или «Мицета». Комбинируя питательные среды, а также подбирая продуцент и условия его культивирования, можно получать липиды, по составу отвечающие требованиям различных отраслей промышленности и сельского хозяйства. Например, при кормлении птиц предпочтение отдается липидам, содержащим до 65-70% ненасыщенных жирных кислот. Микробные липиды, содержащие значительное количество жирных кислот с двумя двойными связями, возможно использовать для приготовления лаков и красок, а также для приготовления медицинских препаратов, способствующих предотвращению атеросклероза и тромбоза. Липиды с преобладанием насыщенных жирных кислот можно употреблять на производство технических смазок. В первых случаях таким требованиям отвечают липиды мицелиальных грибов и дрожжей Lipomyces lipoferus, а во втором - липиды Candida humicola, выращенных на гидролизате древесины. Резюмируя сказанное, следует отметить, что состав липидов (а отсюда и область их возможного использования) в большой мере обусловлен систематическим положением организма-продуцента. В то же время соотношение отдельных компонентов в составе липидов определяется спецификой используемого сырья и физико-химическими условиями культивирования. Эти закономерности липидогенеза весьма существенны при организации промышленного производства микробного жира, так как в конкретных условиях позволяют получать продукт строго определенного состава и свойств. Такой управляемый микробный синтез может удовлетворить требованиям, предъявляемым к липидам различными отраслями народного хозяйства. ГЛАВА 7. Получение нуклеотидов и нуклеиновых кислот. Микробиологический синтез нуклеотидов и их производных имеет пока довольно ограниченные масштабы. Сферы применения этих соединений также пока невелики, за исключением нуклеотидов, используемых как вкусовые пищевые добавки, в особенности при производстве искусственной пищи. Нуклеотиды и их производные могут иметь применение и как лечебные препараты. Широко используются они в лабораторной биохимической практике. Характерная особенность большинства способов получения нуклеотидов - необходимость внесения метаболического предшественника в среду для культивирования микроорганизмов или в реакционную смесь. Получение аденозинтрифосфата (АДФ) и аденозинтрифосфата (АТФ) основано на осуществлении культурой микроорганизма реакции фосфорилирования аденина или адениловой кислоты (5'-АМФ). 71 В случае использования Brevibacterium ammoniagenes АТСС 6872 эту бактерию выращивают на среде, содержащей высокую концентрацию глюкозы, мочевину, дрожжевой экстракт, соли фосфора, магния, кальция и биотин. По ходу культивирования в среду вносят аденозан, который подвергается ферментативному фосфорилированию с образованием адениловой кислоты, АДФ и АТФ. При аналогичных условиях культивирования, но при внесении гуанозина можно получить гуаниловую кислоту, ГДФ и ГТФ. Фосфорилирующим агентом, по-видимому, выступает АТФ. Накопить АТФ в сфере реакции без передачи на акцепторную молекулу внесенного из вне аденозина не удается. Другим источником ферментных систем, осуществляющих фосфорилирование 5'-АМФ, могут служить пекарские дрожжи (Saccharomyces cerevisiae), взятые в виде гомогената или ацетонового порошка. Реакционная смесь содержит 5'-АМФ, глюкозу, фосфатный буфер (рН 7,0) и дрожжи, подготовленные, как сказано выше (рис. 17.1). Вслед за истощением глюкозы при минимальном количестве неорганического фосфата отмечается максимум накопления АТФ. После достижения максимума АТФ наблюдается его распад до АДФ и АМФ при одновременном повышении уровня неорганического фосфата. На выход продукта существенное влияние оказывает концентрация фосфата, присутствующего в виде фосфатного буфера. Отклонение от оптимальной величины может привести к ряду нежелательных реакций: вызвать образование инозиновой кислоты, инозина и гипоксантина. Вероятно, вносимый извне аденозин необходим для осуществления «затравочной» реакции. В равной мере это относится и к 5'-АМФ. Отсюда очевидна необходимость иметь в сфере реакции высокую концентрацию глюкозы, и сбалансированное количество фосфата. По несколько иному пути происходит синтез АТФ при введении в среду аденина (вместо аденозина). В этом случае ключевой реакцией является N-рибозидация аденина. Процесс обычно проводят как типичную аэробную ферментацию с добавлением предшественника. Продуцентами служат Brevibacterium ammoniagenes или Corynebacterium sp. В этих случаях также необходима среда с высокой концентрацией глюкозы. На первом 1 этапе синтеза происходит образование АМФ из внесенного в конце экспоненциальной фазы роста культуры аденина и синтезированного клеткой фосфорибозилпирофосфата (ФРФФ). Реакцию осуществляет аденинфосфорибозилтрансфераза: Дальнейший процесс происходит так же, как при наличии АМФ ФРФФ в клетках образуется в результате реакции, происходящей между рибозо-5фосфатом (Р-5-Ф) и АТФ при посредстве АТФ: D-рибоза-5-фосфат пирофосфаттрансферазы. АТФ и Р-5-Ф образуются в результате катаболизма глюкозы. Р-5-Ф в ходе функционирования ГМФ-пути, а АТФ - как продукт гликолиза. 72 Микробиологический синтез никотинамиддинуклеотида (НАД) происходит путем культивирования Brevibacterium ammoniagenes АТСС 6872 с предшественником. Выращивание продуцента рекомендуется проводить на средах, содержащих глюкозу, мочевину, дрожжевой экстракт, фосфаты, соли магния, кальция и биотин. Предшественниками НАД в среде могут быть никотиновая кислота и никотинамид. В условиях опыта предшественники обычно вводят на вторые сутки роста культуры (рис. 17.2). К пятым суткам в среде накапливается НАД, и культура вступает в фазу стационарного роста. Помимо НАД, в среде содержится промежуточный продукт его синтеза мононуклеотид никотиновой кислоты. Никотиновая кислота включается непосредственно в синтез, и если в среду вводят ее амид, происходит ферментативное дезаминирование с образованием кислоты. Ниже дана схема синтеза НАД, где фосфорибозилпирофосфат обозначен буквами ФРФФ, а остатки фосфорной кислоты в нуклеотидах - Ф (рис. ). На этой же схеме показана также реакция, в результате которой образуется никотинамиддинуклеотидфосфат (НАДФ). Реакцию осуществляет НАДкиназа, передающая фосфорный остаток от АТФ на НАД. Для получения НАДФ предложена также система, использующая в качестве ферментного препарата экстракт из клеток Proteus mirabilis IFO 3849, а компонентами служат НАД, п-нитрофенилфосфат, никотинамид, сульфат цинка, ацетатный буфер (рН 4,0). Однако в данном случае, помимо НАДФ, происходит синтез НАД-дифосфата и некоторых НАД-аналогов. Инозиновая кислота - метаболический предшественник важнейших пуриновых нуклеотидов - адениловой и гуаниловой кислот. Поэтому накопление инозиновой кислоты может происходить, если продуцент имеет блок в ферментных системах, осуществляющих дальнейшие ее превращения. Очевидно, что не следует лишать структуру столь необходимых и важных метаболитов (адениловой и гуаниловой кислот). Введение их в среду для восполнения дефицита возможно, но довольно дорого. Поэтому предпочитают вводить в сферу реакции нуклеозид - инозин. Для получения 5'-инозиновой кислоты (5'-ИМФ) необходимо иметь источник фермента, фосфорилирующего инозин в 5'-положении рибозного остатка. Источниками фосфорилирующего фермента - нуклеозиддифосфаттрансферазы - могут служить многие микроорганизмы, относящиеся к родам Flavobacterium, Serratia, Staphylococcus, Pseudomonas. Донорами фосфора в этой реакции могут быть нуклеотиды, а также неприродные соединения - nнитрофенилфосфат (n-НФФ), бензилфосфат, фенилфосфат. В СССР в качестве источника фосфорилирующего фермента предложен штамм Pseudomonas. Культуру предварительно выращивают на среде, содержащей глюкозу, пептон, дрожжевой экстракт. В качестве ферментного препарата, осуществляющего реакцию, используют живые клетки, высушенные под вакуумом или ацетоном, а также замороженные клетки, поскольку локализация фермента внутриклеточная. Из названных выше доноров фосфата лучшим считается п-нитрофенилфосфат. Реакция 73 происходит в ацетатном буфере, при рН 4,0, в присутствии ионов меди и цинка, заметно усиливающих выход продукта. Наиболее важными факторами, определяющими активность реакции фосфорилирования инозина, являются концентрации донора (фосфора), акцептора (инозина) и количества клеток. При оптимальных соотношениях пНФФ и инозина клетки Ps. trifolii могут фосфорилировать до 90% инозина в реакционной смеси. Выход инозиновой кислоты можно увеличить, если к реакционной смеси добавить поверхностно-активные вещества, в частности различные твины. Концентрации добавок подбирают опытным путем. Как и во всех других случаях использования в качестве ферментных препаратов клеток микроорганизмов, возникает необходимость оценить активность и условия ее проявления у ферментов, разрушающих целевой продукт синтеза. Наиболее нежелательно для синтеза инозиновой кислоты наличие активных фосфомоноэстеразы и 5'-нуклеотидазы. Если невозможно подавить их активность ингибиторами, то следует выбрать условия, при которых она будет минимальной. В первую очередь к условиям, наиболее легко регулируемым, относятся температура и рН среды. Инозиновую кислоту выделяют в виде бариевой соли и путем ионообменной хроматографии. Синтез гуанозинполифосфатов. Среди продуктов обмена веществ нуклеотидной природы у микроорганизмов обнаружены гуанозинполифосфаты. Преимущественно это тетра- или пентафосфаты. Они выполняют функции регуляторов в многочисленных биохимических реакциях, поэтому на их получение обращено внимание многих специалистов. Накопление гуанозинполифосфатов обнаружено в культуре Brevibacterium ammoniagenes KV 13510 в присутствии в реакционной смеси гуанозин-5'-монофосфата (Г-5'-Ф) или ксантозин-5'-монофосфата (Кс-5'-Ф). Штамм представляет собой мутант, требующий для выращивания присутствия в среде дорогостоящих для промышленного производства компонентов: дрожжевой экстракт, биотин, тиамин, пантотеновую кислоту и др. Ксантозин-5'-монофосфат метаболический предшественник гуаниловой кислоты, образующейся после амидирования пуринового основания глутаминов в присутствии АТФ. Собственно процесс ферментативного превращения метаболических предшественников (Г-5'-Ф или Кс-5'-Ф) в нуклеозидполифосфат происходит в среде, содержащей глюкозу, а также высокий уровень фосфатов и магния. Помимо полифосфатов в среде накапливается 5'-ГМФ, 5'-ГДФ, 5'-ГТФ, однако гуанозин полифосфаты образуются в более узком интервале рН и при ином температурном оптимуме. Фермент АТФ нуклеотид пирофосфаттрансфераза, осуществляющий перенос пирофосфатных остатков из АТФ на акцепторные молекулы, выделен из Streptomyces adephospholyticus А-4668. Продуктами реакции 74 могут быть нуклеозид-3'-дифосфат-5'-моно(ди- и три-) фосфаты, такие, как pppGpp, ppGpp, pGpp, а также производные аденина и инозина — рррАрр, ррАрр, pppIpp. ГЛАВА 8. Получение сахаров и полисахаридов. Полисахариды (или гликаны) - полимеры, построенные не менее чем из 11 моносахаридных единиц. Они могут состоять из одного или нескольких типов моносахаров. Соответственно различают гомополисахариды и гетерополисахариды. Полисахариды - обязательные компоненты всех организмов, составляют большую часть углеводов, встречающихся в природе, преобладающую долю в биомассе растений, а, следовательно, и основную массу органического вещества на Земле. Полисахариды встречаются в виде самостоятельных полимеров, а также в комплексах с нуклеиновыми кислотами, белками, липидами, фосфатом. Разнообразны они по мономерному составу и структуре. Особым разнообразием отличаются полисахариды микроорганизмов. Некоторые из них близки или идентичны полисахаридам растений и животных. Но подавляющее большинство микробных полисахаридов имеет уникальную структуру, специфическую для вида или для серологической группы вида. В микробных гликанах часто обнаруживаются ранее неизвестные моносахара, которые не встречаются ни у животных, ни у растений. О том, что слизь, образуемая многими микроорганизмами, может иметь углеводную природу, знали еще во времена Пастера. Однако особое внимание микробным полисахаридам стали уделять лишь с начала 20-х годов нашего столетия, когда узнали, что вещества, определяющие серологическую специфичность пневмококков, являются полисахаридами. В настоящее время исследование микробных полисахаридов приобрело особое значение в связи с открывшейся возможностью широкого применения их в медицине и ряде областей народного хозяйства. Полисахариды микроорганизмов в соответствии с локализацией делятся на внутриклеточные и внеклеточные. К внутриклеточным относят обычно полисахариды цитоплазмы, мембран и клеточных стенок, а к внеклеточным - полисахариды капсул, чехлов (пристеночные структуры) и свободной слизи, не прилегающей к клеточной стенке. Иногда к внеклеточным относят также полисахариды, локализованные снаружи от цитоплазматической мембраны. В этом случае в группу внеклеточных попадают и полисахариды клеточных стенок. У ряда микроорганизмов действительно трудно различить границу между капсулой и клеточной стенкой. Нередко по локализации выделяют три группы полисахариддов: внутриклеточные (цитоплазмы, мембран, периплазмы), полисахариды клеточных стенок и внеклеточные (капсул, чехлов и свободной слизи). 75 Термин «экзогликаны» применяют в основном к полисахаридам свободной слизи. Иногда экзогликанами называют также капсульные полисахариды. Микробные полисахариды объединяют в группы и по функциям: резервные, участвующие в активном транспорте, опорные, участвующие во взаимодействии между клетками, защитные и др. Некоторые исследователи классифицируют полисахариды, учитывая их топологию и функции. В соответствии с этим клеточные полисахариды подразделяются на две группы. Одна включает резервно-энергетические и модификаторы (внутриклеточные), вторая - структурные и структурнометаболические (в клеточной стенке). К внеклеточным относятся выделяющиеся при гиперпродукции структурно-метаболические гликаны и собственно экзогликаны. 8.1. Полисахариды цитоплазмы и мембранных структур. Полисахариды цитоплазмы обнаруживаются в двух формах: они могут быть диспергированы в ней или объединены в гранулы. Обычно в цитоплазме бактерий содержится 20-30% полисахаридов, а в условиях, способствующих их накоплению, до 50-60% от массы сухих клеток. Чаще всего в цитоплазме микроорганизмов обнаруживают гомогликаны, из которых особенно распространены глюканы типа гликогена. Их выделяли из цитоплазмы многих прокариотных и эукариотных микроорганизмов: представителей разных родов бактерий Agrobacteriutn, Arthrobacter, Bacillus, Clostridium, Escherichia, Mycobacterium, Nostoc, Rhodopseudomonas, Rhodospirillum, Streptococcus, а также дрожжей, мицелиальных грибов, ресничных и жгутиковых простейших, некоторых водорослей. Кроме гликогеноподобных полисахаридов в цитоплазме ряда микроорганизмов найдены крахмал, маннаны, леваны, арабаны и ксиланы. Гликоген и другие гомогликаны цитоплазмы могут образовывать комплексы с ДНК, РНК, белками, липидами, фосфатом. Гетерополисахариды обнаруживаются в цитоплазме реже. Однако у представителей Streptomyces и Mycobacterium они оказываются преобладающими. Функции полисахаридов цитоплазмы до конца не выяснены. До недавнего времени считали, что основное или даже единственное их назначение - быть резервным источником углерода и энергии для клетки. Они расходуются, например, при созревании эндоспор у бактерий родов Bacillus и Clostridium. Но теперь ясно, что полисахариды цитоплазмы могут выполнять ряд других важных функций. Предполагается, что комплексы гомогликанов с другими компонентами цитоплазмы участвуют в механизмах клеточной регуляции, контролирующих синтез различных веществ, рост и деление клеток. Так, гликогенрибосомные комплексы, возможно, контролируют синтез белков. Гликоген может оказывать радиозащитное действие на связанные с ним молекулы нуклеиновых кислот. 76 В мембранах микроорганизмов обнаруживается в среднем от 2 до 5%, иногда (у Micrococcus luteus) до 15-20% углеводов от массы мембраны. Возможно, в некоторых случаях эти углеводы полностью или частично представляют собой остаточный материал цитоплазмы или клеточных стенок. Тем не менее достоверно показано, что в мембранах грамположительных и некоторых грамотрицательных бактерий содержатся гликолипиды и гликопротеины. Все грамположительные эубактерии, за исключением микрококков и некоторых стрептококков, а также дрожжи и мицелиальные грибы содержат в области цитоплазматической мембраны тейхоевые кислоты (1-2% от сухой биомассы). Тейхоевые кислоты относят к кислым полисахаридам необычного строения. При их гидролизе наряду с моносахаридами образуются вещества, относящиеся к другим классам соединений. Разнообразие тейхоевых кислот определяется в основном числом присутствующих в них остатков сахаров и наличием связей различных типов. Мембранные тейхоевые кислоты - это всегда глицерофосфатные полимеры, часто связанные с гликолипидами и фосфолипидами (липотейхоевые кислоты). У некоторых микроорганизмов выявляются только липотейхоевые кислоты, а свободных тейхоевых кислот нет. В мембранах грамотрицательных бактерий тейхоевые кислоты не обнаружены. Мембранные гликолипиды участвуют в биосинтезе полисахаридов и транспорте сахаров. Тейхоевые кислоты, видимо, регулируют ионный обмен (связывают двухвалентные катионы, что необходимо для нормального функционирования ферментов, локализованных в мембране), действуют на связывание аминокислот с тРНК, осуществляют связь между мембраной и клеточной стенкой, проявляют антигенную активность. 8.2. Полисахариды клеточных стенок. У грамположительных эубактерии полисахариды составляют от 30 до 60% сухой массы клеточной стенки. Значительная их часть входит в состав муреинового комплекса, количество которого у грамположительных эубактерии достигает 50-90% веществ клеточной стенки. Линейные полисахаридные цепи муреина построены из повторяющихся β-1,4связанных единиц N-ацетилглюкозамина и N-ацетилмурамовой кислоты. Мурамовая кислота - производное глюкозамина, содержащее D-молочную кислоту. Клеточные стенки некоторых архебактерий, дающих положительную окраску по Граму, содержат псевдомуреин, гликановая часть которого состоит из N-ацетилглюкозамина, N-ацетилгалактозамина и Nацетилталозаминуроновой кислоты. Мурамовая кислота в псевдомуреине не найдена. У ряда грамположительных архебактерий клеточная стенка построена только из кислого гетерополисахарида, в состав которого входят галактозамин, нейтральные сахара и уроновые кислоты. В клеточных стенках подавляющего большинства грамположительных эубактерий, за 77 исключением микобактерий и коринебактерий, содержатся тейхоевые кислоты. Их количество обычно достигает 50-90% от массы клеточной стенки, у стрептомицетов оно колеблется от 4 до 50%. Как правило, у микроорганизмов обнаруживается либо глицеринтейхоевая, либо рибиттейхоевая кислота. Однако у Streptococcus faecalis и у одного штамма Streptomyces sp. найдены тейхоевые кислоты обоих типов. Другие полисахариды, содержащиеся в клеточных стенках грамположительных бактерий, отличаются большим разнообразием; чаще всего это гетерогликаны. В их составе обнаруживаются нейтральные моносахара, аминосахара, уроновые кислоты, ацетильные группы, остатки фосфорной кислоты. Клеточные стенки грамотрицательных бактерий содержат от 1 до 50% полисахаридов. Среди них полисахариды муреинового комплекса не занимают доминирующего положения, так как его количество составляет в среднем всего около 5% веществ клеточной стенки. Тейхоевые кислоты обнаруживаются только у отдельных представителей грамотрицательных бактерий. Особенно характерны для клеточных стенок грамотрицательных бактерий липополисахариды (ЛПС) биологически активные вещества, участвующие в формировании наружной мембраны. Число различных липополисахаридов велико и, несмотря на интенсивное изучение, строение и состав многих из них известны не до конца. В полисахаридной части комплекса различают базисную структуру и О-специфические боковые цепи. Моносахаридный состав, варьирование связей и структура их в основном и определяют биологическую активность липополисахаридов, выполняющих функцию соматических антигенов. Углеводный компонент липополисахарида - это обычно гетерополисахарид. У различных бактерий в его составе обнаружены нейтральные сахара, аминосахара, уроновые кислоты, метальные и ацетильные группы и, что особенно характерно, 3,6дидезоксипроизводные cахаров, которые в других природных объектах встречаются редко. Из них наиболее распространены абеквоза (3,6дидезокси-D-галактоза), колитоза (3,6-дидезокси-L-галактоза), тивелоза (3,6дидезокси-D-манноза), аскарилоза (3,6-дидезокси-L-манноза) и паратоза (3,6дидезокси-D-глюкоза). Эти сахара часто определяют серологическую специфичность бактерий. В О-специфической боковой цепи липополисахарида одного из видов рода Pasteurella обнаружена 6-дезокси-Dманногептоза - первая 6-дезоксигептоза, найденная в природе. Моносахариды особого типа - гликолактиловые кислоты - выявлены в составе О-антигенных полисахаридов представителей рода Shigella. Полисахаридные компоненты ЛПС ряда бактерий отличаются сложностью. Они могут содержать до 6 и более различных незамещенных и замещенных моносахаров. У энтеробактерий молекулы ЛПС могут образовывать комплексы с пептидогликаном, кислыми капсульными гликанами и другими гетерополисахаридами клетки. 78 Полисахариды - главные компоненты клеточной стенки дрожжей и мицелиальных грибов. Они могут составлять до 90% массы клеточной стенки (Saccharomyces cerevisiae). У дрожжей часто обнаруживаются гомогликаны: глюканы, маннаны, хитин (полимер N-ацетилглюкозамина). В дрожжевых маннанах нередко содержатся остатки фосфорной кислоты и (или) метильные группы, а в глюканах - ацетильные и аминогруппы. У многих дрожжей в клеточной стенке обнаруживаются гетерополисахариды галактоманнаны и глюкоманнаны, а также кислые гликаны, построенные из 3-4 мономеров. По-видимому, в клеточных стенках дрожжей присутствует несколько полисахаридов. Гликаны клеточных стенок дрожжей могут быть связаны с пептидами. Основным полисахаридным компонентом клеточных стенок большинства исследованных мицелиальных грибов является хитин. Обнаруживаются также неацетилированный или частично ацетилированный полимер глюкозамина - хитозан, глюканы (иногда целлюлоза), галактаны и различные гетерополисахариды, включающие незамещенные и замещенные сахара, уроновые кислоты. Структурные микрофибриллы клеточных стенок большинства микроформ водорослей состоят из целлюлозы, а у отдельных представителей - из других гомополисахаридов, часто из ксиланов и маннанов. Количество их может достигать 50-80% массы сухой клеточной стенки. Полисахариды матрикса представлены в основном гетерогликанами. Обнаруживаются и сульфатированные полисахариды. Полисахариды клеточных стенок выполняют разнообразные функции. Многие из них определяют механическую прочность клеточных стенок. Поэтому их часто называют «скелетными». Липополисахариды, тейхоевые кислоты, а также гетерополисахариды ряда грамположительных бактерий ответственны за антигенную активность клеток. ЛПС значительного числа грамотрицательных бактерий - токсины. ЛПС энтеробактерий защищают клетки от ингибирующего действия длинноцепочечных жирных кислот, позволяя этим бактериям выживать в кишечнике животных. С наличием Оспецифических боковых цепей ЛПС связана способность шигелл прикрепляться к надмембранному покрову эпителиальных клеток. Многие полисахариды определяют устойчивость бактерий к литическим ферментам и фагам. Полианионные полисахариды способствуют транспорту из клетки заряженных метаболитов и веществ, поступающих в нее из окружающей среды. Кроме того, такие полисахариды сообщают клетке отрицательный заряд, в результате чего происходит взаимное отталкивание клеток, диспергирование их в среде. Потеря О-боковых цепей ЛПС снижает гидрофильность клеток и приводит к их спонтанной агглютинации. Полисахариды клеточных стенок микроорганизмов, растущих на средах с налканами, обычно являются хорошими эмульгаторами и тем самым способствуют проникновению углеводородов в клетку. 79 8.3. Внеклеточные полисахариды. Внеклеточные полисахариды, как уже отмечалось, обнаруживаются в виде капсул и чехлов, прилегающих к клеточным стенкам, а также свободной слизи. Капсулы, имеющие толщину менее 0,2 мкм, не различимые в световом микроскопе, но хорошо видимые в электронный микроскоп, называют микрокапсулами. Микрокапсулы обычно связаны с клеточной стенкой прочнее, чем капсулы. У многих микроорганизмов капсулы имеют определенную структуру и четко отграничены от слизи. У некоторых бактерий капсульный материал рыхлый, бесструктурный, легко отторгается от клеток, поэтому границу между капсулой и свободной слизью в этом случае определить трудно. Такие аморфные капсулы называют слизистыми слоями. Чехлы в отличие от капсул имеют сложную структуру. В них нередко различают несколько слоев с разным строением. Количество внеклеточных полисахаридов может во много раз превышать биомассу клеток. Капсулы, чехлы и слизь не всегда состоят только из полисахаридов. Они могут кроме гликанов включать белки, полипептиды, нуклеиновые кислоты, липиды, образующие или не образующие комплексы с полисахаридами. Как правило, чехлы имеют более сложный химический состав. Так, у Sphaerotilus natans они содержат глюкозу, глюкозамин, белок, липид и фосфат. Чехлы бактерий, окисляющих восстановленные соединения металлов, часто содержат включения их окислов. Иногда неуглеводные полимеры - единственные компоненты капсул и слизи. Так, капсулы некоторых видов рода Bacillus построены из полипептида, слизь некоторых штаммов Pseudomonas aeruginosa состоит из ДНК. Внеклеточные полисахариды, капсульные или свободные, или те и другие, образуют многие микроорганизмы. Пожалуй, нет такой группы микроорганизмов, представители которой не обладали бы этой способностью. Однако синтез внеклеточных полисахаридов - не обязательная функция клетки и проявляется она лишь в определенных условиях. Встречаются микроорганизмы, у которых никогда не удавалось наблюдать ни капсул, ни слизи. Внеклеточные полисахариды микроорганизмов чрезвычайно разнообразны по составу и строению. К настоящему времени исследован состав около 200 экзогликанов, установлены первичная структура и детали строения многих из них. В составе внеклеточных полисахаридов различных микроорганизмов обнаружено более 20 моносахаров и их производных. Наиболее часто встречаются гексозы: глюкоза, галактоза, манноза и 6дезоксигексозы: фукоза и рамноза. Реже выявляются пентозы: арабиноза, ксилоза, рибоза. Распространены уроновые кислоты: галактуроновая, маннуроновая и особенно глюкуроновая. Многие содержат аминосахара: глюкозамин, галактозамин и маннозамин. Часто в экзогликанах присутствуют неуглеводные заместители - пируват и ацетат, встречаются также сукцинат и глицерин. Для ряда внеклеточных гликанов характерно наличие редких мономеров: 2,6- или 3,6-дидезоксисахаров, у некоторых 80 найдены ранее неизвестные моносахариды, например гликолактиловые кислоты (у сапротрофных микобактерий). Иногда обнаруживаются тейхоевые кислоты, фосфатные и сульфатные ионы. Внеклеточные полисахариды большинства видов бактерий - кислые гетерогликаны разнообразного состава, построенные из 2-5, иногда 6-7 мономеров, линейные и разветвленные, имеющие регулярную структуру из повторяющихся олигосахаридных звеньев. Так, например, Xanthomonas campestris синтезирует полианионит ксантан, включающий глюкозу, маннозу, глюкуроновую кислоту и О-ацетильную группу и пируват. Некоторые бактерии образуют нейтральные гетерополисахариды. Весьма распространены у микроорганизмов различные гомополисахариды, особенно глюканы, из которых наиболее известны декстраны (группа более или менее близких по строению нейтральных глюканов). Они могут содержать до 200000 остатков глюкозы, бывают линейными и разветвленными. Линейная (основная) цепь построена при участии α-1,6-связей, ветвление обусловлено α-1,2-, α-1,3- и α-1,4-связями. Молекулярная масса декстранов колеблется от 12 до 600 млн. Наиболее активные продуценты декстранов - представители молочнокислых бактерий Leuconostoc mesenteroides и L. dextranicum. Декстраны синтезируются также некоторыми видами Streptococcus (Str. sanguis, Str. mutans), Brevibacterium, Lactobacillus. Практически каждый продуцент синтезирует свой, несколько отличный от других видов декстран. Некоторые штаммы образуют одновременно два различных по структуре декстрана. Внеклеточную целлюлозу - полисахарид, распространенный в растениях, из бактерий синтезируют некоторые представители Pseudomonas, Zooglea, Azotobacter, подавляющее число видов Rhizobium и Agrobacterium, Acetobacter xylinum. β-(1→3)-Глюкан, называемый курдланом, образуют Alcaligenes faecalis var. myxogenes и виды Rhizobium. Сильно разветвленный α-(1→4)-глюкан с боковыми цепочками, присоединенными α-(1→6)-связями, - полимер типа гликогена, резервного полисахарида животных, многих бактерий и дрожжей - можно обнаружить в культуральной жидкости Neisseria perflava. Нигеран - α-глюкан с чередующимися (1→4)- и (1→3)связями - образует гриб Aspergillus niger. Разветвленные полимеры фруктозы - леваны с (2→6)-связями синтезируют уксуснокислые бактерии Gluconobacter охуdans, Acetobacter aceti, некоторые виды Pseudomonas, Erwinia (подгруппы «herbicola»), Aeromonas, Bacillus. Фруктаны типа инулина (резервный полисахарид растений семейства сложноцветных) с (2→1)-связями в основной цепи и ответвлениями в положении С6 образуют штаммы Str. mutans. Маннаны обнаружены в культурах некоторых видов Bacillus и Corynebacterium, а также у многих дрожжей. Чаще всего микроорганизмы, способные к образованию внеклеточных полисахаридов, синтезируют капсулы и свободную слизь. Мономерный состав слизи и капсул в большинстве случаев одинаковый. 81 Но не всегда можно определить, какие именно полисахариды свойственны той или иной группе микроорганизмов. В ряде случаев филогенетически близкие бактерии синтезируют внеклеточные гликаны, сходные по составу и строению. К ним относятся, например, бактериальный альгинат Pseudomonas aeruginosa и Azotobacier vinelandii, курдлан Rhizobium и A. faecalis var. myxogenes, кислые гетерогликаны Corynebacterium и Arthrobacter, декстраны Streptococcus и Lactobacillus и др. Однако нередко микроорганизмы, далеко отстоящие в таксономическом отношении, образуют гликаны сходного состава или одинаковые. Обычно в этом случае полимеры проявляют и функциональное сходство. Так, очень близки по coставу экзогликаны различных фитопатогенных бактерий и возбудителей менингита. У ряда бактерий цементирующим материалом при образовании клеточных агрегатов служит внеклеточная целлюлоза. Известно, что не только разные виды одного рода микроорганизмов, но часто и разные штаммы одного вида синтезируют неодинаковые экзополисахариды. Так, Е. coli имеет около 70 серотипов, различных по составу и структуре капсульных полисахаридов и, соответственно, по иммунохимическим свойствам. Экзогликаны эффективных и неэффективных штаммов клубеньковых бактерий различаются по мономерному составу, а разных штаммов дрожжей рода Lipomyces - по соотношению моносахаров. Неодинаковыми по моносахаридному составу могут быть внеклеточные гликаны М-, S- и R-форм бактерий. Отмечены случаи, когда в культуральной жидкости одного микроорганизма накапливается несколько различных гликанов. Например, L. mesenteroides образует декстран и леван, у Serratia marcescens обнаружено три экзополисахарида: кислый глюкорамнан, содержащий глюкуроновую кислоту, рамноглюкан и гептоглюкан. Гриб Aupeobasidium (Pullularia) pullulans образует два гомоглюкана: аубазидан - разветвленный полимер с α(1→4), β-(1→3) и β-(1→6)-связями, причем в боковой цепи может быть от одного до четырех остатков глюкозы на одно звено триозы, и пуллулан линейный глюкан, состоящий из мальтотриозных и мальтотетраозных фрагментов, соединенных α-(1→6)-связями. Внеклеточные полисахариды не являются жизненно необходимыми для микроорганизмов. В природе есть виды, никогда их не образующие. Экспериментально показано, что клетки, лишенные капсул, столь же жизнеспособны, как и капсулированные. Тем не менее внеклеточные полисахариды выполняют определенные функции, способствующие поддержанию условий, благоприятных для жизнедеятельности продуцента. Одни из них - универсальны для всех полисахаридов, поскольку определяются общими для этих веществ свойствами, другие - специфичны для определенного полисахарида, что обусловлено особенностями состава и строения данного полимера. Внеклеточные полисахариды предохраняют клетки от высушивания, от губительного действия ультрафиолетовых лучей и различных химических 82 агентов, в том числе тяжелых металлов и лекарственных препаратов. Замечено, что капсулированные бактерии устойчивее к химиотерапевтическим средствам, чем бескапсульные варианты. Располагаясь поверхностно, капсулы защищают клетки от бактериофагов и поглощения их простейшими, предупреждают денатурацию клеточного белка. Многие внеклеточные гликаны биологически активны и определяют иммунологические свойства и вирулентность штаммов. Как правило, чем толще капсула, тем выше вирулентность и патогенность бактерий. Некоторые патогенные бактерии, лишенные капсул, становятся авирулентными. К-антигены являются агрессинами, подавляющими фагоцитоз бактерий и тем создающими условия для их размножения. Одно из характерных проявлений биологической активности полисахаридов - способность модифицировать ферменты различных организмов, в результате чего стимулируется или снижается их активность. Экзополисахариды фитопатогенных бактерий являются фитотоксинами, участвующими в специфическом взаимодействии бактерий с растительной тканью. К ним относится, например, ксантан. В некоторых случаях внеклеточные гликаны служат резервным источником углерода и энергии, а азотсодержащие полисахариды источником азота для продуцента. Полианионные полисахариды концентрируют катионы из окружающей среды и способствуют транспорту их в клетку. Полисахаридно-липидные комплексы микроорганизмов, растущих на средах с н-алканами: псевдомонад, нокардий, коринебактерий, дрожжей - хорошие эмульгаторы. Они снижают поверхностное натяжение, увеличивают поверхность соприкосновения углеводорода и воды, образуют мицеллы и тем самым способствуют проникновению углеводородов в клетку. Нейтральные внеклеточные полисахариды поддерживают целостность нитчатых форм и различных скоплений клеток. Полисахариды, несущие определенный заряд, напротив, способствуют диспергированию клеток. Экзогликаны некоторых бактерий ответственны за прикрепление клеток к поверхности. Так, Str. mutans, вызывающий кариес зубов, прикрепляется к зубной эмали с помощью декстрана. 8.4. Использование полисахаридов. В настоящее время полисахариды микроорганизмов достаточно широко используются в практике (табл. 20.2). Они находят применение в самых различных сферах человеческой деятельности: в медицине, фармацевтической, пищевой, химической и текстильной промышленности, в гидрометаллургии, при добыче нефти и в ряде других областей народного хозяйства. При этом внимание исследователей и практиков привлекают и внутриклеточные и внеклеточные гликаны, однако в технико-экономическом плане предпочтительнее последние - масштаб их производства и применения значительно шире. 83 Возможность и перспективность использования полисахаридов в медицине в значительной мере определяется их биологической активностью. Многие микробные полисахариды обладают лечебным и профилактическим действием: повышают устойчивость организма к бактериальным и вирусным инфекциям, обладают противоопухолевой активностью, способствуют заживлению ран и регенерации тканей, благоприятно влияют на течение и исход воспалительных процессов, устраняют болевой синдром, снижают побочное действие лекарственных препаратов и рентгенотерапии. Лечебное и защитное действие полисахаридов определяется прежде всего их способностью повышать неспецифическую иммунобиологическую реактивность организма, влиять на различные защитные реакции, поддерживающие постоянство его внутренней среды. Преимущества многих полисахаридных препаратов перед другими средствами, повышающими неспецифическую резистентность организма, определяются тем, что они свободны от примесей, оказывающих нежелательное действие на организм. Некоторые микробные полисахариды уже нашли применение в лечебной практике различных клиник мира. В нашей стране для лечения последствий травм и нарушений проводимости нервной системы, для предупреждения образования грубых послеожоговых или посттравматических рубцов успешно применяли пирогеналь - препарат, выделяемый из клеток Salmonella typhi и Pseudomanas aeruginosa. В ФРГ и США с этой же целью использовали липополисахариды, изолированные из различных патогенных бактерий. Бактериальные ЛПС обладают также и противолучевой активностью. В клиниках Советского Союза уже более 20 лет применяют продигиозан - гетерополисахаридный комплекс с липидами, выделенный из клеток Serratia marcescens, и зимозан - препарат из оболочек клеток Sacch. cerevisiae, состоящий из глюкана, глюкоманнана и минорных количеств тейхоевых кислот. Эти препараты нормализуют ряд сдвигов в иммунобиологических реакциях, оказывают положительное действие при лечении опухолей, ряда инфекционных и неинфекционных заболеваний. Перспективны в качестве противоопухолевых агентов ЛПС ряда грамотрицательных бактерий, внутриклеточный глюкан парамилон (астазиан) бесцветных фитофлагеллят Astasia longa, внеклеточные полисахариды различных дрожжей родов Lipomyces, Cryptococcus, Bullera и др., бактерий родов Alcaligenes и Agrobacterium. Перечисленные соединения рекомендованы для клинических испытаний. Противовирусную активность проявляет продигиозан. Модифицированный (сульфатированный) полярный маннан - внеклеточный полисахарид Rhodotorula rubra - перспективен как средство профилактики и лечения атеросклероза. Полисахариды, обладающие антигенной специфичностью, начинают использоваться в медицинской практике в качестве диагностических средств. К ним относятся, например, полисахаридные препараты патогенных и условно патогенных видов дрожжей рода Candida, облегчающие диагностику 84 заболеваний кандидозной природы. Показана возможность использования модифицированных ЛПС-антигенов сальмонелл в диагностике сальмонеллезов. Очищенные специфические полисахариды менингококков групп А и С (полимеры N-ацетил, О-ацетилманнозаминфосфата и N-ацетил, Оацетилнейраминовой кислоты соответственно) используются для получения менингококковых вакцин. Микробные полисахариды могут быть основой для создания искусственных вакцин. Достигается это изменением их конфигурации или конъюгацией с синтетическими полиэлектролитами. Нейтральные декстраны с молекулярным весом около 75 000, продуцируемые L. mesenteroides, широко применяются у нас в стране и за рубежом в качестве заменителей плазмы крови. Перспективны как плазмозаменители пуллулан, а также леваны, синтезируемые G. oxydans и Вас. polymyxa. Декстраны определенного строения, как и многие другие полисахариды, способны стимулировать защитные реакции организма. В клиниках они применяются в комплексе с другими препаратами для лечения различных заболеваний брюшной полости. Сульфаты декстрана обладают антикоагулирующим действием, заменяют гепарин и могут применяться как антитромбогенное средство. В качестве антикоагулянта перспективен также хитин. Широкое применение микробных полисахаридов в фармацевтической, парфюмерной, пищевой и других отраслях промышленности определяется их свойствами: вязкостью, реологическими характеристиками, способностью к набуханию, взаимодействием с определенными структурами. В фармацевтике они используются в качестве основы для изготовления лекарственных форм: как мягчители, эмульгаторы и стабилизаторы суспензий, как склеивающие агенты и разрыхлители в мазях, пилюлях, таблетках. Они обеспечивают длительную устойчивость лекарственных препаратов, стабилизируют и пролонгируют их действие. На базе некоторых микробных полисахаридов (аубазидан, декстран) созданы стабильные в течение нескольких лет лекарственные препараты: бутадиона, серы, сульфаниламидов, суспензии сульфата бария для рентгеноскопии и др. Макромолекулярные конъюгаты модифицированных декстранов с ферментами (стрептокиназой, трипсином, фибринолизином) пролонгируют активность ферментов и снижают их аллергизирующее действие. Микробные полисахариды применяются как гельобразующие агенты при изготовлении косметических изделий, для создания гидрофильного буфера в кремах, в качестве набухающих веществ при производстве кремов, шампуней, лосьонов. Некоторые гликаны можно использовать вместо применяемой в настоящее время натрий-карбоксицеллюлозы в качестве связующего и биологически активного компонента в зубных пастах. В пищевой промышленности полисахариды микроорганизмов используются в виде пленок - покрытий продуктов, например сыров, для защиты их от высыхания и плесневения, в качестве стабилизаторов 85 мороженого, фруктовых соков, приправ к салатам, загустителей сиропов, джемов, подливок, желе и других кулинарных изделий. Особенно перспективным в этом плане считается ксантан. Слизеобразующие штаммы Streptococcus lactis применяют в Швейцарии при производстве густых кефиров, сметан и некоторых мягких сыров. Экзополисахариды дрожжей родов Saccharomyces и Ctreptococcus, бактерий родов Azotobacter и Arthrobacter могут использоваться для улучшения качества хлеба. Добавление их к муке при выпечке хлеба повышает газоудерживающую способность теста, улучшает его, реологические свойства. Хлеб, выпеченный из такого теста, отличается высоким удельным объемом, хорошей пористостью, медленнее черствеет. Как гельобразующие агенты экзогликаны применяются при производстве ядерного топлива, фотографических и рентгеновских пленок, как заменители альгиновой кислоты водорослей в пищевой, текстильной, фармацевтической и бумажной промышленности (полиурониды Azotobacter, P. aeruginosa и ряда других микроорганизмов), они могут заменять агар (гетерополисахариды Вас. subtilis и Ps. elodea, состоящие соответственно из глюкозы, галактозы, фукозы, глюкуроновой кислоты и глюкозы, рамнозы, глюкуроновой кислоты и О-ацетильных групп). Анионные полисахариды (ксантан, занфло - внеклеточный гетерогликан Erwinia tahitica, состоящий из глюкозы, галактозы, фукозы, уроновой кислоты и ацетильных групп, и др.) стабилизируют и предохраняют от высыхания катионные водные эмалевые краски. Некоторые гликаны, например гетерополисахарид Corynebacterium equi var. mucilagenosus, обладают высокой вязкостью и могут заменять дорогие клеящие средства. Сульфаты ксантана используются как загустители клеев. С другой стороны, способность ряда полисахаридов к образованию поверхностных пленок позволяет использовать их в качестве антисклеивающих веществ, например при освобождении слепков из отливочных форм. Декстран рекомендуется применять и в качестве смазочного средства. Полисахариды, водные растворы которых отличаются особой стабильностью при резких изменениях температуры и в условиях агрессивной среды, используются в нефтяной и газодобывающей промышленности как стабилизаторы и структурообразователи промывных жидкостей, предназначенных для бурения нефтяных и газовых скважин, и обеспечивают более полное извлечение нефти из нефтеносных пластов. Уже около 15 лет назад более половины нефти в США добывали с помощью полисахаридов, главным образом ксантана. Промышленные испытания проходит склероглюкан - капсульный линейный нейтральный глюкан, образуемый несовершенными грибами, преимущественно рода Sclerotium. В качестве стабилизатора буровых глинистых суспензий перспективен линейный внеклеточный гетерогликан облигатно-метилотрофных бактерий Methylobacillus methylophilus, состоящий из глюкозы, галактозы, маннозы, 86 рамнозы и глюкуроновой кислоты. Применение полисахаридов в нефтедобывающей промышленности является очень перспективным в техническом отношении. Полисахариды ряда микроорганизмов (пуллулан A. pullulans, гетерополисахарид бактерий рода Methylomonas и др.) являются флоккулирующими агентами и применяются в гидрометаллургии для получения металлических компонентов в виде гелей, включающих нерастворимые осадки. Процесс реализован при очистке, разделении и концентрации металлов из растворов их солей или смесей солей. На основе декстранов получают сефадексы, широко применяемые в лабораторной практике для гельфильтрации. Полианионные гликаны, например ксантан, хитин, рекомендуется использовать для очистки воды от тяжелых металлов, а также при промышленном синтезе полимеров для извлечения их из органических растворителей. Хитин может найти применение и для очистки сточных вод. В качестве носителя иммобилизованной α-амилазы используют аубазидан. Перспективны для иммобилизации ферментов курдлан и хитин. Микробные леваны - источники получения чистого препарата фруктозы, хитин - D-глюкозамина и N-ацетил-D-глюкозамина - соединений, используемых в химическом синтезе, из маннанов дрожжей можно получать маннозу. Полисахариды некоторых бактерий, например леван Вас. роlymyxa, оказались полезными в сельском хозяйстве При внесении в почву они повышают выживаемость семян культурных растений, способствуя сохранению в них влаги. Альгинатными пленками покрывают корни и семена растений для предохранения их от высыхания во время хранения и перевозок. Возможности практического применения полисахаридов микроорганизмов полностью еще не раскрыты. Всестороннее изучение гликанов в этом плане открывает новые перспективы и, несомненно, приведет к расширению соответствующей области микробиологической промышленности. 8.5. Промышленное получение полисахаридов микроорганизмов. Расширение спектра микробных полисахаридов, имеющих практическое значение, обусловило успехи в области организации их производства. В настоящее время микробиологическая промышленность многих стран выпускает ряд ценных экзогликанов: декстран (СССР и другие страны), ксантан (США, Франция), пуллулан (Япония), склероглюкан или «политран» (США), занфло (США), курдлан (Япония). Уже решены или решаются вопросы внедрения в производство ряда других полисахаридов, детально изученных в лабораторных условиях, проверенных на практике и производимых в полупромышленном масштабе. Производство различных полисахаридов не универсально. Для каждого гликана оно имеет свои 87 особенности, определяемые физиологией продуцента, локализацией и физико-химическими свойствами полимера, областью его применения. Получение экзополисахаридов имеет преимущества перед получением внутриклеточных, так как экзогликаны образуются, как правило, в значительно большем количестве, легче отделяются от биомассы и очищаются от примесей. Однако при производстве экзогликанов имеются свои технологические трудности. Накопление полисахарида в среде приводит к ограничению доступа кислорода к клеткам. У аэробных микроорганизмов это снижает энергетический баланс и тормозит синтез полисахарида. Повышенная вязкость среды делает невозможным отделение полисахарида от клеток продуцента из нативной культуральной жидкости. Ее приходится разбавлять в десятки раз, а после удаления клеток концентрировать до первоначального или меньшего объема. Решение этих проблем связано с дополнительными затратами. Приведем основные этапы производства наиболее широко применяемых сейчас полисахаридов - декстрана и ксантана. Плазмозаменители из декстранов выпускают под названиями: клинический декстран, полиглюкин, синкол, макродекс, плазмодекс, хемодекс и др. Для получения декстранов используют штаммы Leuconostoc mesenteroides. Ферментацию ведут на среде с 10-30 % сахарозы, декстраном «затравкой», дрожжевым экстрактом, минеральными солями. Создают условия, способствующие синтезу той формы декстрана, которая используется в качестве плазмозаменителя: линейного глюкана, имеющего более 90% α-1,6-связей, с молекулярной массой 60-80 тыс. Для этого ограничивают содержание в среде магния, стимулирующего синтез разветвленных декстранов, вносят «затравку» в виде декстрана, имеющего молекулярную массу 20-30 тыс. Такой акцептор обеспечивает преимущественное образование необходимого полимера. Наивысшей биологической активностью обладают декстраны, содержащие менее 70% α-1,6-гликозидных связей, т.е. более разветвленные. Синтезу биологически активных декстранов способствует, кроме магния, замена сахарозы мелассой. Оптимальное значение рН для роста продуцента лежит в пределах 6,5-8,0, а для накопления декстрансахаразы - около 7,0. Обычно значение рН среды задают в интервале 7,0-8,0. Бактерии расщепляют сахарозу на глюкозу и фруктозу. Фруктоза сбраживается по типу гетероферментативного молочно-кислого брожения с образованием молочной и уксусной кислот, маннита и СО2. Глюкоза полимеризуется в декстран. Процесс идет быстро и продукт можно выделить уже через 24 ч. Декстран выделяют из культуральной жидкости, например, метанолом. Можно, используя определенные приемы, осаждать фракции клинического декстрана с молекулярной массой 60-80 тыс. даже из смеси декстранов разной молекулярной массы. Можно осадить весь продукт, растворить его в воде и изолировать требуемый декстран фракционированием. При 88 необходимости выделенный декстран деполимеризуют (ферментативно, термической обработкой или ультразвуком). Для очистки декстран неоднократно растворяют в воде, переосаждают метанолом и фракционируют. Поскольку декстрансахараза в значительной степени выделяется в среду и синтез полимера идет вне клетки, декстраны получают и ферментативным путем. Для этого продуцент выращивается в условиях, обеспечивающих наиболее высокую активность внеклеточного ферментного комплекса. В период максимальной активности декстрансахаразы культуральную жидкость отделяют от клеток и консервируют, снижая значение рН до 5,0-5,2. При такой кислотности и температуре около 15 °С декстрансахараза, содержащаяся в культуральной жидкости, сохраняет активность не менее месяца. В СССР разработана технология получения частично очищенной декстрансахаразы. Ферментационная среда должна содержать сахарозу и декстран-«затравку». Процесс синтеза продолжается около 8 ч. Ферментативный способ удобнее микробиологического, так как он поддается более надежному контролю и регулированию, позволяет одним только варьированием исходных концентраций сахарозы и фермента, а также температуры процесса сразу получать декстран необходимой молекулярной массы. Это значительно упрощает и удешевляет последующие технологические операции. Широкое применение в промышленности может найти использование иммобилизованных декстрансахараз. В нашей стране в 1983 г. выпущена первая промышленная серия конъюгатов модифицированного декстрана со стрептокиназой «стрептодеказа» - пролонгированная с помощью декстрана форма стрептокиназы. Ксантан, продуцируемый Xanthomonas campestris, выпускают под названиями: биополимер Хс, келцан, ксантан, келтрол. Бактерии культивируют на среде, содержащей 1-5 % углеводов (кукурузный крахмал, сахар-сырец, меласса и др.), органическое соединение азота, двузамещенный фосфорнокислый калий и микроэлементы, рН среды 6,5-7,2. Инкубацию проводят в аэробных условиях, при 28 °С, в течение 72 ч. Для улучшения свойств полисахарида к среде во время ферментации добавляют формальдегид. Добавка позволяет получать гликан с повышенной устойчивостью к различным неблагоприятным факторам, в том числе к температуре и засолению. Полимер используют в виде раствора вязкой культуральной жидкости или в виде порошка, высушенного в струе горячего воздуха. В последнем случае полисахарид отделяют от клеток центрифугированием и очищают осаждением этанолом, метанолом или ацетоном в присутствии электролита. Интенсивные поиски продуцентов полисахаридов типа ксантана ведутся в различных странах. Активные продуценты среди бактерий рода Xanthomonas найдены в Советском Союзе. Разрабатывается технология получения отечественного ксантана в промышленном масштабе. 89 Нередко нативные полисахариды не обладают желаемыми качествами. Они могут быть, например, недостаточно активны или, будучи высокоэффективными, плохо растворимы или токсичны, что препятствует их применению, и т.д. Чтобы улучшить действие полисахаридных препаратов, устранить или снизить нежелательные явления, т.е. получить препарат с нужными свойствами, выделенные полисахариды иногда подвергают химической модификации. Так, декстран сульфатируют, чтобы придать ему антикоагулирующую активность. Обработка нативного гликана A. faecalis var. myxogenes солюбилизирующими агентами позволяет получить производные, образующие гели без предварительного нагревания. Растворимость глюкана A. pullulans и маннана R. rubra удается повысить карбоксиметилированием. Снижение токсичности противоопухолевого препарата белково-липо-полисахаридного комплекса из культуральной жидкости Serratia piscatorum достигается обработкой его щелочью. Получение производных полисахаридов связано с дополнительными технологическими операциями. Все более широкое применение для производства экзогликанов находит метод непрерывного культивирования продуцентов. С его помощью в ряде стран уже получают многие перспективные в практическом отношении полисахариды (ксантан, курдлан и др.). Этот способ весьма эффективен и экономичен, поскольку позволяет длительно получать продукт в период его максимального накопления и наиболее полно использовать субстрат. Подсчитано, что процент конверсии углеродного субстрата в экзополисахариды при проточном культивировании в 2-3 раза выше, чем при периодическом. Обычно в качестве лимитирующего фактора в хемостате используют азот, а также фосфор и серу. Это обеспечивает интенсивное накопление экзогликанов. Благодаря правильному подбору условий ксантан получают при непрерывном культивировании продуцента в течение многих сотен часов. В настоящее время разрабатывается производство ряда других практически ценных полисахаридов. У нас в стране на опытной установке Красноярского завода медпрепаратов получают аубазидан и суспензии сульфата бария на аубазидане. Создается опытно-промышленная установка для наработки маннана, развиваются работы по получению продигиозана и группоспецифических полисахаридов менингококков (вакцин). Перед микробиологическим производством полисахаридов стоит ряд задач. Одна из важнейших - замена дорогостоящих сахаров - традиционного источника углерода для получения многих полисахаридов, более дешевым сырьем. В связи с этим ведутся поиски микроорганизмов, растущих и образующих полисахариды при использовании углеводородов, этанола, метанола. Спирты как источники углерода несомненно удобнее углеводородов, так как они хорошо растворимы в воде, что значительно упрощает технологию выделения и очистки продукта. Использование для культивирования микроорганизмов возможно более простых по составу сред 90 - синтетических или диализованных, необходимо при получении высокоочищенных антигенов. Дешевый субстрат - гидролизат торфа предлагается использовать для производства пуллулана. Необходимы поиск новых продуцентов полисахаридов с полезными свойствами, особенно внеклеточных, и селекционно-генетические исследования перспективных микроорганизмов с целью получения вариантов с повышенной продуктивностью гликанов. Метод экспериментальной селекции активных штаммов нередко оказывается более быстрым и экономичным в сравнении с поиском таковых в природных условиях. Так, в результате действия низкой температуры с последующим облучением быстрыми нейтронами удалось отобрать варианты X. campestris, обладающие повышенным синтезом ксантана. Поскольку микроорганизмы являются поистине неиссякаемыми источниками полисахаридов, можно не сомневаться, что среди этих полимеров будут обнаруживаться вещества, оригинальные в химическом отношении и представляющие большую ценность для практики. ГЛАВА 9. Получение антибиотиков Принципы получения вторичных метаболитов основаны на особенностях их образования клетками микроорганизмов. Биосинтез вторичных метаболитов фазоспецифичен и происходит после завершения стадии роста, в идиофазе, благодаря чему их и называют идиолитами. Антибиотики – самый большой класс фармацевтических соединений, синтез которых осуществляется микробными клетками. К этому же классу относятся противогрибковые агенты, противоопухолевые лекарства и алкалоиды. В мире ежегодно производится антибиотиков на 20 млрд. долларов. Еще более широко, чем в медицине, они используются в растениеводстве, животноводстве, ветеринарии, пищевой промышленности. В 1928 г. английский микробиолог Флеминг установил способность зеленой плесени Penicillum notanum вызывать гибель микроорганизмов, а во время 2-ой Мировой войны был начат промышленный выпуск антибиотиков. Следует отметить, что в 1871 г. лечебные свойства плесени были описаны русским дерматологом А.Г. Полотебновым. К антибиотикам относятся низкомолекулярные эффекторы изначально природного происхождения, способные подавлять рост живых клеток. В настоящее время описано 12000 антибиотичных препаратов, из которых в клинике применяется около 200. Около 97 % известных антибиотиков токсичны, поэтому в практике не используются. В химическом отношении они представляют собой сборную группу органических веществ. По типу действия антибиотики делят на бактерицидные, вызывающие гибель микроорганизмов и бактериостатические, нарушающие способность микроорганизмов делиться. По спектру действия различают антибиотики 91 узкого и широкого действия. К последним относят тетрациклины, макролиды, амилогликозиды, которые назначают в случае неидентифицируемых возбудителей болезней, при длительном применении они вызывают дисбактериоз. Особенностью антибиотиков является специфичность их действия. Эти эффекторы подавляют один или несколько процессов лишь у некоторых микроорганизмов. В зависимости от специфики действия антибиотиков на молекулярном уровне различают следующие группы соединений, вызывающие у бактерий: 1) нарушение биосинтеза пептидогликанов клеточной стенки; 2) нарушение отдельных этапов процесса трансляции (трансляция – реализация генетической информации); 3) повреждение цитоплазматической мембраны; 4) нарушение биосинтеза нуклеиновых кислот; 5) нарушение энергетического обмена. Антибиотики широко используются для расшифровки механизмов биосинтеза белка, нуклеиновых кислот и структуры клеточных стенок бактерий. Изыскание новых антибиотиков обусловлено и потребностями практики, и накоплением резистентных форм микроорганизмов по отношению ко многим антибиотикам. Устойчивость микроорганизмам придает фермент лактамаза, превращающий пенициллин в пенициллиновую кислоту. Поэтому новые аналоги антибиотиков получают при использовании природных ингибиторов β-лактамаз – клавулановой и оливановой кислот. Химические методы получения антибиотиков очень сложны и не могут конкурировать с их биосинтезом методами биотехнологии. Существует несколько способов получения как природных, так и полусинтетических антибиотиков: 1) ферментация микроорганизма-продуцента с подходящим предшественником, что индуцирует синтез антибиотиков в идиофазе; 2) использование для биосинтеза блокированных мутантов. У таких мутантов блокирован синтез нужного антибиотика. Используя низкую субстратную специфичность ферментов вторичного метаболизма и вводя аналоги предшественников антибиотика, их переводят в аналоги самого антибиотика. Этот процесс называется биосинтез, или мутасинтез: а) предполагается последовательность реакций, ведущая к синтезу антибиотика фермент A B С D E антибиотик ; б) отсутствие синтеза антибиотика у «блокированного» мутанта фермент C D; А В блокированное звено метаболизма в) синтез модифицированного антибиотика после введения аналога предшественника (D*) фермент A B ...D* E * – модифицированный антибиотик. 92 Антибиотики продуцируются плесневыми грибами, актинамицетами, эубактериями и другими микроорганизмами. Шесть родов филаментозных грибов производят около 1000 различных антибиотиков, в том числе пенициллин и цефаллоспорин, а три рода актиномицетов – 3000 антибиотиков. Среди актиномицетов наибольший вклад вносит род Streptomyces, один из видов которого – S. griseus синтезирует более 50 антибиотиков. Процесс культивирования идиолитов проходит две фазы (двухступенчатое культивирование). На первой фазе происходит накопление достаточного количества биомассы, которая выращивается на подходящей для роста микроорганизма среде. Эта фаза должна быть быстрой, а питательная среда дешѐвой. На второй фазе осуществляется запуск и активный синтез антибиотика. На этой фазе ферментацию ведут на продуктивной среде. Образование антибиотиков регулируется условиями культивирования микроорганизмов. Поэтому оптимизация питательной среды является главным фактором в повышении выхода продукта. Большинство антибиотиков получают при глубинной аэробной ферментации периодического действия в асептических условиях. 9.1. Принципы получения антибиотиков. Широкое применение антибиотиков в медицине, сельском хозяйстве и других отраслях народного хозяйства поставило задачу получения этих биологически активных веществ в массовых масштабах. Решение этой задачи стало возможным благодаря созданию мощной антибиотической промышленности. В основе промышленного производства антибиотиков лежит ряд последовательных этапов: получение высокопродуктивных штаммовпродуцентов, разработка наиболее благоприятных условий культивирования продуцента антибиотика с максимальным биосинтезом этого вещества, подбор и внедрение в практику соответствующих методов выделения и очистки антибиотика, создание готовых препаратов и контроль их качества. Каждый из этих этапов должен обеспечиваться соответствующими специалистами (генетиками, микробиологами, технологами и др.). Производство антибиотиков представляет ныне мощную, хорошо развитую отрасль, входящую в фармацевтическую (в нашей стране в медицинскую и микробиологическую) промышленность. Она занимает одно из ведущих мест в производстве лекарственных препаратов. Особенно широко она развита в США, Англии, Японии, Франции, Италии и других странах. Например, в США ежегодно выпускается антибиотиков и их производных на сотни миллионов долларов. По общему производству антибиотиков Советский Союз занимает ведущее место в мире. По данным Лове и Эландера, только 5-лактамные антибиотики (пенициллин и цефалоспорин) составляют половину общего объема производимых антибиотиков. В 1979 г., отмечают названные авторы, путем биосинтеза 93 только пенициллина было получено около 14800 т на общую сумму 240 млн. долларов. Промышленный способ получения антибиотиков - сложный многоступенчатый процесс, включающий ряд технологических стадий. Ниже мы рассмотрим из них основные. Большое значение для получения разнообразных модификаций уже известных антибиотических веществ, обладающих более ценными свойствами по сравнению с исходными препаратами, имеет так называемый полусинтетический способ производства антибиотиков. Этой проблеме стали уделять заметное внимание с начала 60-х годов. В основе полусинтетического способа получения антибиотиков лежит следующий принцип. В результате биосинтеза получают исходные антибиотики (пенициллин, цефалоспорин, тетрациклины, рифамицин), которые затем подвергаются химической модификации. В ряде случаев основным исходным полуфабрикатом служит не вся молекула антибиотика, а лишь ее основное ядро. Так, при получении полусинтетических пенициллинов и цефалоспоринов используют соответственно 6аминопенициллановую кислоту и 7-аминоцефалоспорановую и 7аминодезацетоксицефалоспорановую кислоты, которые подвергаются затем химическому воздействию. 6-аминопенициллановую кислоту (6-АПК) получают тремя способами: 1) в результате развития продуцента пенициллина в среде, не содержащей предшественника; 2) химической обработкой (гидролизом) бензилпенициллина и 3) ферментативным гидролизом бензилпенициллина. На возможность получения 6-АПК в результате изменения условий культивирования продуцента пенициллина впервые указал Като в 1953 г. Бетчел и др. в 1959 г. установили, что пенициллинообразующие штаммы гриба способны при развитии в среде, не содержащей предшественника, образовывать 6-АПК. Однако практического, тем более промышленного значения такое получение 6-аминопенициллановой кислоты не имеет. Связано это с тем, что, во-первых, при развитии продуцента пенициллина в средах без предшественника образуется относительно небольшое количество 6-АПК. Во-вторых, довольно трудно выделять из культуральной, жидкости эту кислоту и очищать ее от сопутствующих веществ. Более эффективным способом получения 6-АПК является химический метод, в основе которого лежит обработка бензилпенициллина пятихлористым фосфором с получением легкогидролизуемого соединения, из последнего затем и получают 6-АПК. В данном случае выход 6-АПК составляет до 95%. Наиболее рациональным способом получения 6-аминопенициллановой кислоты следует назвать ферментативный гидролиз молекулы пенициллина. Фермент пенициллинацилаза осуществляет гидролиз молекулы бензилпенициллина с образованием 6-АПК и фенилуксусной кислоты: 94 6-Аминопенициллановая кислота практически лишена антибиотической активности. Пенициллинацилазу способны образовывать различные виды микроорганизмов, в том числе и грибы, продуцирующие пенициллин. Фермент, образуемый бактериями и проактиномицетами, осуществляет гидролиз бензилпенициллина более быстро, чем другие типы пенициллинов. В настоящее время получение 6-аминопенициллановой кислоты проводят с помощью иммобилизованной пенициллинацилазы, что обеспечивает большой выход продукта и его высокую степень чистоты. Важнейшим этапом после ферментации антибиотиков является их выделение и очистка из культуральной жидкости. В культуральной жидкости наряду с антибиотическим веществом, как правило, содержится огромное количество посторонних примесей, очень часто близких по своим химическим и физико-химическим свойствам к антибиотику. Примеси, сопутствующие антибиотическому веществу, представляют собой вещества минерального или органического характера, являются или продуктами биосинтеза, или компонентами питательной среды, а также веществами, добавляемыми в культуральную жидкость для ее предварительной обработки. Концентрация этих веществ часто достигает нескольких процентов и превышает концентрацию антибиотиков в десятки и сотни, а иногда и тысячи раз. Процесс выделения и очистки антибиотиков представляет собой сложный технологический процесс. Малая стабильность многих антибиотиков и возможность потери их активности при химических превращениях привели к преимущественному использованию для выделения и очистки антибиотиков физико-химических приемов разделения веществ, включая сорбцию, экстракцию и кристаллизацию, т.е. таких процессов, которые не сопровождались резким химическим воздействием на молекулу антибиотика. Для разделения, выделения и очистки антибиотиков применяются как равновесные, так и кинетические методы. Однако для производственных задач равновесные методы оказались более экономически выгодными и эффективными. Наибольшее значение приобрели здесь сорбционные и экстракционные методы. Равновесные методы могут быть одностадийными и многостадийными. При промышленном применении, а также при изучении свойств антибиотиков имеют большое значение многостадийные процессы, позволяющие значительно улучшать степень чистоты выделяемого препарата. Предварительная обработка и фильтрация культуральных жидкостей антибиотиков. Предварительная обработка культуральной жидкости и удаление мицелиальной массы являются первой стадией перед выделением и очисткой. Собственно, уже на этой стадии начинается частичная очистка культуральной жидкости от примесей. В зависимости от 95 свойств антибиотика и методов его выделения и очистки выбирается способ предварительной обработки культуральной жидкости. Основной задачей предварительной обработки культуральной жидкости является получение нативного раствора (а в случае нахождения антибиотика в мицелии мицелиальной массы) с наибольшей степенью чистоты, с наименьшими потерями, позволяющими обеспечить успешное проведение дальнейших операций выделения и химической очистки антибиотика. Большинство антибиотиков выделяются и очищаются из нативного раствора тремя методами: 1) экстракционным, 2) ионообменным, 3) осаждением нерастворимого соединения. При экстракционном методе извлечения антибиотиков из жидкости (пенициллин, эритромицин, новобиоцин) нативный раствор при предварительной обработке должен быть максимально освобожден от примесей, способных образовывать стойкие эмульсии с органическим растворителем. Белковые примеси, как правило, вызывающие образование стойких эмульсий, удаляются или вместе с мицелием с помощью различных химических способов или нагревом жидкости, или тем и другим вместе. Если мицелиальная масса удаляется легко без предварительной обработки, то к нативному раствору добавляют дезэмульгаторы, удерживающие белковые вещества в растворенном состоянии в условиях экстракции. При выделении и очистке пенициллина в качестве дезэмульгатора используется «контакт Петрова» (керосиновый контакт, катексол, ультравет, цетазол). При выделении и очистке эритромицина необходимо удалять из нативного раствора ионы кальция, которые могут при экстракции способствовать выпадению антибиотика в осадок. В случае применения ионообменного метода выделения антибиотика из нативного раствора последний должен быть максимально освобожден от конкурирующих ионов в случае сорбции на катионитах ионов кальция, магния, железа. Для удаления кальция применяется щавелевая кислота, для удаления магния - фосфаты, для удаления железа - желтая кровяная соль. Если антибиотик выделяется из нативного раствора с помощью осаждения, то из нативного раствора желательно удалить все примеси, способные в этих условиях перейти в осадок. Одной из основных задач предварительной обработки культуральной жидкости является коагуляция частиц, находящихся во взвешенном состоянии. Особенно важность этой задачи проявляется при коагуляции и фильтрации культуральной жидкости актиномицетного происхождения или бактериального. Отделение мицелиальной массы от нативного раствора в большинстве случаев связано со значительными трудностями. Это объясняется спецификой осадка, который обычно имеет аморфный, слизистый, бесструктурный характер и быстро забивает поры фильтрующего материала. Большое влияние на процесс фильтрации оказывают качество сырья и сырьевой состав питательной среды. Например, применение соевой муки, жмыхов в составе среды ухудшает фильтрацию жидкости. Применение 96 гидрола вместо глюкозы как источника углевода снижает скорость фильтрации (производство стрептомицина). Неполное потребление питательных веществ, применение жировых пеногасителей на последних этапах ферментации также приводят к ухудшению фильтрации. Для улучшения процесса фильтрации очень важно вовремя прекращать процесс ферментации. Прекращать ферментацию желательно при полном потреблении углеводов, но до наступления разрушения микробной клетки, так как процесс фильтрации автолизированной культуры обычно идет плохо. Кроме того, увеличение длительности ферментации ухудшает качество нативного раствора, увеличивает его пигментацию, содержание белковых примесей. Для коагуляции культуральные жидкости антибиотиков специально обрабатываются. В зависимости от свойства антибиотика, происхождения мицелиальной массы и метода выделения и очистки антибиотика культуральная жидкость для улучшения фильтруемости обрабатывается: 1) кислотной коагуляцией; 2) введением в жидкость электролитов; 3) тепловой коагуляцией; 4) применением наполнителей; 5) образованием наполнителя непосредственно в жидкости. Иногда используется сочетание двух методов. Кислотная и тепловая коагуляция используется в том случае, если антибиотики устойчивы при изменении рН раствора и температуры. Нагревание жидкости увеличивает скорость фильтрации вследствие свертывания и коагуляции белков при высокой температуре, а также благодаря значительному уменьшению вязкости фильтрата. С другой стороны, тепловая коагуляция увеличивает пигментацию нативного раствора и тем самым может ухудшить качество готового продукта. Для улучшения фильтрации часто применяются наполнители такие, как диатомит или инфузорная земля, перлит и т, д. Хорошим методом коагуляции культуральных жидкостей антибиотиков является метод образования наполнителя непосредственно в жидкости при добавлении реагентов, образующих в ней нерастворимый осадок. Выпадающий в жидкости осадок предотвращает слипание частиц мицелия, способствует образованию гранул, благодаря чему мицелий приобретает комковатую структуру и образует при фильтрации хорошо проницаемый слой. Эффективность этого метода высока и при правильной выборе условий позволяет увеличить скорость фильтрации в 3 - 10 раз (рис. 18). Исключительно большое влияние на эффективность коагуляции культуральной жидкости оказывают гидродинамические условия процесса. Культуральные жидкости, обработанные при разных гидродинамических режимах, отличаются по внешнему виду. Обработка при слабом перемешивании позволяет получать осадки в виде маленьких комочков, хорошо фильтруемые. С усилением перемешивания коагуляция становится менее отчетливой, жидкость отстаивается медленнее. Осадки после фильтрации получаются более влажными, липкими. Гидродинамические условия при обработке культуральных жидкостей оказывают большое 97 влияяние на фильтруемость их. Так, например, при увеличении скорости фильтрации культуральной жидкости от 1 до 2 м/сек фильтруемость суспензии и степень ее дисперсности практически одинаковы. Дальнейшая интенсификация гидродинамического режима (до ω = 10 м/сек) приводит к ухудшению фильтрационных характеристик суспензии. Следовательно, при фильтрации культуральных жидкостей имеет большое значение не размер кристаллов, а величина гранул, которые образуются при агрегации частиц коагулянта-наполнителя и мицелия. Большое значение для фильтрации коагулированного раствора имеет продолжительность выдержки после обработки. Уменьшение выдержки ниже определенного предела вызывает снижение скорости фильтрации и может в некоторых случаях привести к опалесценции фильтрата. Движущей силой процесса фильтрации является разность давлений по обе стороны слоя осадка, а одной из важнейших характеристик его - скорость фильтрации, т.е. количество фильтрата, получаемого с единицы поверхности в единицу времени. Скорость фильтрации зависит от давления, толщины слоя осадка, структуры и характера осажденного вещества, вязкости жидкой фазы суспензии и других факторов. Экстракционные процессы. Экстракция широко применяется для выделения и очистки многих химических соединений, в том числе и антибиотиков (и, особенно, когда они - или продукты их соединения с переносчиками не ионизированы в водной фазе). Несмотря на существенный недостаток экстракционных процессов, а именно использование вредных, взрывоопасных органических растворителей, все же она находит широкое применение в промышленности. Во-первых, экстракционные процессы по времени протекают значительно быстрее, чем ионообменные, коэффициенты распределения для некоторых систем очень велики, и это позволяет резко сокращать объемы перерабатываемых растворов. Аппаратурно этот процесс очень легко осуществить непрерывным способом. Особенно интересна экстракция с переносчиком; часто этот вид экстракции называют жидким ионным обменом. Экстракция с переносчиком. Перенос вещества осуществляется е образованием комплексного соединения с гидрофобным переносчиком. Перенос осуществляется не только за счет переносчика, но и за счет подавления полярных групп в молекуле антибиотика переносчиком. В качестве переносчиков используются (олеиновая кислота, ундециленовая кислота, из оснований - цетазол (цетилпиридиний бромид). Кислота (например, олеиновая) взаимодействует с основанием стрептомицина, образуя соль с большей энергией сольватации в органической фазе, и вещество переходит в органическую фазу. Переносчик может находиться как в водной, так и в органической фазе. Случай, когда переносчик находится в водной фазе, экстракцию можно представить следующим образом: А+В ↔ (АВ) р-р водный ↔ (АВ) органическая фаза (1) Переносчик находится в органической фазе: 98 А+ + В-С+ ↔ А+В- + С+ (2) Компонент В неспособен находиться в водной фазе. В этом случае мы имеем уравнение ионного обмена, и к нему можно применить все законы ионного обмена. Таким образом, если перенос осуществляется с помощью неионизированного переносчика, то используется уравнение (2), а если переносчик ионизирован, то уравнение (1). Наиболее ярким представителем антибиотиков, где до сих пор успешно применяется экстракционный метод выделения и очистки антибиотиков, является пенициллин. После предварительной обработки культуральной жидкости и отделения осадка она направляется на экстракцию, которая осуществляется с помощью бутилацетата при рН 2; коэффициент распределения при этих условиях может достигать величины 30. Добавляемая при этом серная кислота доводит рН раствора до 2 и этим самым подавляет степень диссоциации пенициллина в водной фазе, превращая его в недиссоциированную пенициллиновую кислоту, которая легко переходит в органическую фазу. Затем бутилацетатный экстракт обрабатывается слабым раствором щелочи и пенициллин в виде соли снова переходит в водную фазу. Экстракция повторяется еще раз для более полной очистки и концентрирования антибиотика. Представителем антибиотиков, которые выделяются и очищаются с помощью переносчиков, являются антибиотики тетрациклиновой группы (тетрациклин, окситетрациклин). Наличие в группе этих антибиотиков одной основной и двух кислых групп позволяет использовать в качестве переносчиков этих соединений вещества основного и кислого характера. В качестве кислых переносчиков можно использовать сульфокислоты, фенолы, жирные кислоты, а в качестве жидких анионитов - четвертичные аммониевые основания с длинной углеводородной цепью (С10 - С30). Схематически химические реакции при этом можно представить следующим образом: R • СОО-Na+ + окситетрациклин + ↔ РСОО • окситетрациклин+ + Nа+. R • N+Hal- + окситетрациклин - ↔ RN окситетрациклин + Hal-. Проведенные исследования (С.И. Каплан и др., 1962) говорят о том, что максимальная степень экстракции (95-97%) достигается при применении 5% цетазола в растворителе, взятом в количестве 15% от объема нативного раствора при рН водной фазы 9,5-10,2. Весьма существенным фактором, определяющим процесс извлечения антибиотика, является длительность контактирования двух жидких фаз. Исследования тех же авторов показали, что при оптимальном значении рН равновесие в системе достигается уже В течение первой минуты перемешивания. Важным моментом является последующая реэкстракция окситетрациклина из органической фазы. Использование для этой цели соляной, фосфорной, лимонной кислот хотя и позволяло получать растворы с концентрацией антибиотика 40 000—50 000 ЕД/мл, но эти растворы имеют интенсивную темную окраску. Реэкстракция 99 окситетрациклина 4-5% раствором щавелевой кислоты позволяет получить растворы с концентрацией антибиотика 35 000-45 000 ЕД/ мл со светлой окраской реэкстракта. Вероятно, это явление можно объяснить различной степенью диссоциации окрашенных примесей и свойством щавелевой кислоты взаимодействовать с ними. Аппаратура экстракции антибиотиков. Современное производство антибиотиков развивается в условиях усилившегося внедрения нового прогрессивного технологического метода выделения - очистки ионообменного. В связи с этим в аппаратурном оформлении экстракционного метода особенно актуален рациональный выбор конструкций аппаратов и режима их работы с применением экономических показателей. В химической промышленности используется много принципов экстракции и типов аппаратов, пригодных для экстракции антибиотиков. Если в прежние годы выбор экстракторов производился интуитивно, то в последнее десятилетие благодаря работам Пратта, Г.П. Питерских и Е.Р. Валашека, Д.Е. Шкоропада и И.В. Лысковцева выбор экстракторов может быть сделан довольно строго путем рациональной оценки достоинств и недостатков каждого типа экстракторов. Своеобразие физико-химических свойств антибиотиков - пенициллина, тетрациклина, эритромицина, бацитрацина и их нативных растворов позволяет выбирать лишь исключительно строгие режимы их экстракции. Описание конструкций экстракторов. Процесс экстракции слагается из двух элементов, проводимых либо в одном, либо в разных аппаратах: 1) эмульгирования одной жидкости в другой и их взаимного движения, при которых происходит экстракция, и 2) разделения жидкостей. Современные центробежные жидкостные экстракторы непрерывного действия по способу их работы делят на прямоточные и противоточные. В прямоточных экстракторах раствор и экстрагент, подводимые непрерывными потоками, смешиваются в отдельном аппарате смесителе или в смесительном устройстве экстрактора-сепаратора, затем транспортируются к сепаратору или сепараторному барабану экстрактора-сепаратора. В противоточных экстракторах-сепараторах экстрагент и раствор движутся противоточно в роторе аппарата. При раздельном непрерывном экстрагировании и сепарировании в качестве прямоточных смесителей-экстракторов применяют: 1) аппарат емкостью 10—20 л с мешалкой или центробежный насос; 2) трубчатый экстрактор, известный под названием трубы Питерских, длиной порядка 10 м, в котором создан развитый турбулентный режим движения жидкости; 3) струйный смеситель типа инжектора. Смеситель с мешалкой наименее эффективен, поскольку, в отличие от двух других экстракторов, длительность пребывания частиц жидкости в нем 100 различна. Кроме того, концентрация антибиотика в растворе из-за работы мешалки выравнивается во всем объеме и не отличается от концентрации удаляемого раствора. Это уменьшает движущую силу процесса (Г.П. Питерских и Е.Р. Валашек). Для разделения полученных эмульсий применяют тарельчатые сепараторы антибиотических жидкостей САЖ-3М (СССР), снабженные сдвоенными центробежными насосами с напором 1 ат. Производительность сепараторов 2,5 м3/час. За рубежом применяют сепараторы PSBS (ГДР), «Де-Лаваль» (Швеция) и др. При меньших объемах производства используют сверхцентрифуги СГС-100. Сорбционные процессы. Сорбционные методы выделения и очистки антибиотиков находят самое широкое распространение в промышленности. Они обладают целым рядом преимуществ по сравнению с другими методами и поэтому являются исключительно перспективными. Первые сорбционные методы выделения и очистки антибиотиков были основаны на применении молекулярных сорбентов (активированного угля, окиси алюминия и т.д.). Молекулярные сорбенты, такие как активированный уголь, обладают универсальной сорбционной способностью, т.е. одинаково хорошо сорбируют выделяемое вещество и целый ряд других примесей. Исключительно большие возможности синтеза ионообменных сорбентов, сорбентов с различной избирательностью и особенно со специфической избирательностью по отношению к отдельным антибиотикам быстро выдвинули их на первый план. Молекулярные же сорбенты в настоящее время находят применение на последних стадиях доочистки и удаления пигментных примесей. Ионообменные сорбенты принадлежат к классу высокомолекулярных соединений, макромолекулы которых имеют сетчатую или пространственную структуру и в большинстве случаев представляют собой аморфные вещества. Отдельные атомы этих гигантских молекул соединены друг с другом ковалентными связями. Такая структура ионообменных сорбентов и связанное с этим отсутствие растворимости в известной степени нарушают общепринятое представление об электролитах и ионообменных процессах между ними. Диссоциация растворимых в воде кислот или оснований вызывает изменение концентрации водородных ионов. Ионообменные сорбенты также содержат кислотные и основные группы, но их погружение в воду не вызывает изменения в ней концентрации водородных ионов. Катиониты представляют собой особый класс солей и кислот, которые характеризуются многовалентностью и, обладая громоздкой структурой, практически лишены подвижности. Катион, входящий в состав ионита, подвижен, и только сила электростатического притяжения препятствует ему отделяться в растворитель, поэтому вокруг адсорбента создается ионная атмосфера. Катионы в ионной атмосфере неравноценны по силе их электростатического сцепления с анионом адсорбента, поэтому процесс ионного обмена представляет собою многоступенчатую реакцию. 101 Ионы водорода или гидроксильные группы ионита свободно диффундируют в фазе сорбента. Вся остальная часть этого нерастворимого электролита, отдельные атомы и группы которого соединены между собою ковалентными связями, лишены подвижности, представляя собой гигантский анион (в случае катионита) или катион (в случае анионита). Таким образом, процесс ионного обмена можно рассматривать как взаимодействие двух электролитов, один из которых (сорбент) содержит практически неподвижный анионный (или катионный) комплекс. Процесс ионного обмена слагается из диффузии ионов растворенного электролита к поверхности зерна сорбента, диффузии ионов растворенного электролита внутрь сорбента, вытеснения подвижного иона из сорбента и диффузии вытесненного подвижного иона из фазы сорбента в раствор. Классификация сорбентов. Иониты в зависимости от наличия в них ионогенных групп можно разделить на два класса: 1) ионообменные сорбенты, содержащие в своей структуре кислотные группы - катиониты (нерастворимые кислоты); 2) ионообменные сорбенты, содержащие в своей структуре основные группы - аниониты (нерастворимые основания). Разграничение ионитов на кислоты и основания не исключает возможности существования ионитов амфолитов, ионогенные группы которых могут вести себя как кислоты или как основания, в зависимости от рН среды. Существуют также иониты, содержащие одновременно кислотные и основные группы. Все применяемые для сорбции антибиотиков материалы могут быть отнесены к одному из следующих классов: к молекулярным сорбентам, минеральным ионитам или к ионообменным смолам. Среди последних наибольшее значение имеют карбоксильные смолы и сульфокатиониты, а также аниониты различной степени основности. Самыми распространенными молекулярными сорбентами, применяемыми в производстве антибиотиков, являются активированный уголь и окись алюминия. Полимерные смоляные сорбенты могут быть синтезированы на основе процессов полимеризации или поликонденсации. Первые из них обычно обладают большей механической прочностью. Тем не менее ряд полимерных сорбентов, полученных путем поликонденсации, также с успехом применяются как в лабораторной практике, так и в промышленности. Основные требования, предъявляемые к ионитам. Обычно качество ионитов отражают условной оценкой сорбционных свойств ионита (полная емкость сорбента, емкость сорбента при различных рН среды, скорость установления сорбционного равновесия, объем десорбирующего раствора и полнота десорбции); физических свойств ионита в определенных условиях (набухаемость и прочность зерен, теплостойкость и химическая стойкость и т.д.). Весьма желательно, чтобы ионит, применяемый для извлечения ценных ионов из раствора или для очистки того или иного вещества, обладал максимальной емкостью. Для выполнения этого условия необходимо, чтобы в синтезируемом сорбенте содержалось возможно большее число 102 ионогенных групп на единицу веса, полностью ионизированных в данных конкретных условиях сорбции. Следует при этом помнить, что ионизация оксифенильных групп становится заметной при рН выше 9, ионизация карбоксильных групп при рН выше 5, сульфогруппы полностью ионизированы в кислой среде, амино- и иминогруппы вступают в реакцию ионного обмена в кислой среде, четвертичные аммониевые основания проявляют свойства ионообменного сорбента в нейтральных и даже слабощелочных средах. Иониты, применяемые для хроматографического анализа, должны содержать однотипные кислотные или основные группы. В этом случае легко достигнуть четкого разделения смеси соответствующих антибиотиков или других биологически активных веществ. При выборе ионитов необходимо также учитывать относительную прочность связи подвижных ионов с ионитом с тем, чтобы десорбция их осуществлялась столь же легко, как и сорбция. Чем больше энергия ионной связи между подвижными и неподвижными ионами сорбента, тем труднее сместить равновесие в период десорбции. Для набухающих ионитов большое значение приобретает степень набухания его в сорбируемых растворах. С увеличением набухаемости возрастает доступность ионогенных групп, что в свою очередь увеличивает емкость сорбента и скорость установления сорбционного равновесия. С величиной набухаемости тесно связана прочность зерен сорбента. С повышением набухаемости она уменьшается. В условиях резкого изменения степени набухания внутренние напряжения, возникающие в зернах, вызывают их разрушение. Механическую прочность ионита D определяют как отношение объема ионита после отсева пыли V2 к объему до встряхивания (в %) D=V2 / V1 * 100 Аппаратура ионообменной сорбции антибиотиков. Аппаратура для сорбционных ионообменных процессов, несомненно, более проста по устройству по сравнению с экстракционной и доступнее по стоимости. Конструкция ионообменного фильтра, называемого также колонной, для сорбции в динамических условиях должна обеспечивать: 1) постоянное нахождение смолы под слоем раствора; 2) минимальный унос мелких зерен смолы с уходящим фильтратом; 3) минимальную слеживаемость слоя смолы; 4) отсутствие мертвых пространств в сорбенте; 5) незначительное разбавление водой обрабатываемых растворов. Конструкция, размеры колонн и гидродинамический режим ее работы зависят от механических свойств смолы, ее сорбционной емкости и стоимости. Иониты, применяемые для умягчения и обессоливания воды, например сульфоугли, - это обычно твердые частицы, устойчивые к истиранию, создающие низкое сопротивление потоку жидкости в насадке. Высокая прочность смолы позволяет применить в качестве фильтров обычные емкостные аппараты с эллиптическими крышками и днищем, 103 снабженные поддерживающими устройствами, распределителями и сифонами. Такие фильтры благодаря малой емкости смолы и большой производительности имеют диаметр и высоту слоя смолы порядка 1,5 м. Вода в эти фильтры подается сверху, снизу же - лишь для периодического взрыхления слоя ионита после регенерации. Для целей деминерализации элюата стрептомицина и других антибиотиков применяют ионообменные фильтры аналогичной конструкции, хотя и меньшего размера, поскольку применяемые для этой цели недорогие смолы, например СБС-1 и ЭДЭ-10, обладают такими же хорошими механическими свойствами. Для сорбции антибиотиков применяются более дорогостоящие ионообменные смолы, например катиониты КБ-4П-2, КБ-2. Они являются слабосшитыми крупнопористыми ионообменными смолами, обладающими сравнительно небольшой механической прочностью. В набухшем состоянии они, находясь в толстом слое, деформируются под действием силы тяжести и давления нисходящего потока раствора. При этом порозность насадки становится значительно меньше порозности шарообразных недеформируемых частиц (равной 0,4), вследствие чего увеличивается гидравлическое сопротивление для потока раствора и уменьшается площадь контакта зерен смолы с раствором. В связи с этим отечественные предприятия в последние годы перешли к использованию восходящего потока раствора. Раствор подается с линейными скоростями (отнесенными к полному сечению аппарата), немного большими критической, при которой сопротивление слоя становится равным погруженному весу слоя, и зерна переходят во взвешенное состояние. Сорбция в таком псевдоожиженном слое протекает быстрее, увеличивается сорбционная емкость смолы и чистота элюатов. Критическая скорость нативного раствора для слоя сферических частиц одинакового диаметра определяется по уравнению, дающему погрешность ±25%: Re кр = 0,0007 * Аr. Для полидисперсного слоя вычисляется эквивалентный диаметр зерен. Критическая скорость имеет порядок десятых долей мм/сек. Для предупреждения уноса мелких частиц смолы применяется конструкция фильтра в виде колонны с диаметром 0,4—0,7 м, с расширенной открытой верхней частью, изготовленной из органического стекла. В верхней части колонны скорость раствора уменьшается до величин значительно меньших критической. Унесенная потоком смола улавливается в желобах и возвращается в рабочую часть колонны. Восходящий поток применяется также при регенерации, когда переход смолы в другую форму, например из водородной формы катионитов в натриевую, сопровождается набуханием смолы и увеличением ее объема раза в два. При подаче раствора сверху такое набухание смолы может разрушить колонну. Восходящий поток полезен также при промывке и взрыхлении слоя 104 смолы. Десорбция антибиотиков, сопровождаемая сжатием смолы, производится нисходящим потоком для увеличения концентрации элюата. Любой ионитовый фильтр имеет дренажный или опорный слой, нижний и верхний распределитель и устройство (сифон) для поддержания уровня раствора на 0,1—0,15 м выше уровня смолы. Дренажный слой служит для поддержания слоя смолы. Для фильтров с диаметром более 1,5 ж он состоит из слоя гравия, антрацита или другого инертного материала. При меньшем диаметре фильтров используют пористые пластинки или конические колпачки из пластмассы со щелями, имеющими поперечник, равный мелкой фракции зерен смолы, укрепленные в дырчатом ложном днище. Фильтрующую ткань используют реже. Такой дренажный слой служит и распределителем. Распределитель в виде дырчатой трубы, установленный непосредственно над слоем смолы (с учетом набухания) служит для равномерного распределения потока жидкости по сечению фильтра. Неравномерное распределение сильно ухудшает эффективность сорбционного процесса, ведет к возникновению мертвых зон, не омываемых растворов. Ионообменные фильтры изготовляют обычно из углеродистой стали. Для защиты от коррозии, вызываемой растворами кислот и щелочей, применяется гуммирование или стеклянная футеровка корпуса аппарата и трубопроводов. Такая защита эффективна благодаря отсутствию органических растворителей и повышенных температур растворов. Колонны с диаметром меньше 0,3 м целесообразно изготовить из винипласта, органического или обычного стекла. Прозрачные корпуса колонн очень удобны в работе. Трубопроводы из винипласта имеют недостаточную прочность, поэтому при длительной работе их следует избегать. Ионообменная аппаратура и технология развиваются по пути ускорения процесса сорбции, автоматизации приготовления растворов для регенерации, автоматизации контроля концентрации раствора, вытекающего из колонны, перевода процесса ионного обмена на непрерывный. Кристаллизация. Процесс кристаллизации в производстве антибиотиков, как правило, является завершающим этапом и поэтому требует тщательного исследования. Качество выпускаемого препарата в целом зависит от того, каким образом прошло выделение и химическая очистка его из культуральной жидкости, и, в частности, зависит от правильного соблюдения условий кристаллизации на завершающей стадии. В настоящее время требования к качеству выпускаемых препаратов повышаются, и это заставляет еще глубже вникать в процессы кристаллизации и искать пути и возможности их использования для получения препаратов в готовом виде. Твердые тела могут быть кристаллическими и аморфными; кристаллическое состояние отличается от аморфного расположением молекул, атомов или ионов в определенном и строгом порядке. Рентгеновский анализ показал, что многие вещества, 105 которые когда-то считались аморфными, имеют правильное расположение молекул, но термин «кристаллический» чаще всего применяется для обозначения высокой степени внутренней упорядоченности, приводящей к образованию определенных наружных граней кристалла. В газах и жидкостях движение молекул свободно и беспорядочно, поэтому физические свойства этих веществ одинаковы по всем направлениям, большинство же кристаллов обладает различными механическими, электрическими и магнитными свойствами в разных направлениях. Различные вещества могут при кристаллизации давать почти одинаковые кристаллические формы, такие вещества называются изоморфными. Изоморфные вещества часто аналогичны по химическому составу и имеют одинаковые химические формулы. Вещества, способные кристаллизоваться в различные, но химически идентичные формы, называются полиморфными. Основным условием процесса кристаллизации является получение пересыщенного раствора; находящийся в равновесии с твердой фазой считается насыщенным этой твердой фазой. Из насыщенного раствора сравнительно легко можно получить раствор, содержащий больший процент растворенной твердой фазы. Такой раствор называется пересыщенным. Если раствор, охлаждается без потери растворителя, то самопроизвольной кристаллизации не произойдет до тех пор, пока не будут достигнуты необходимые условия. В этой точке кристаллизация может происходить спонтанно, либо ее можно вызвать затравкой, перемешиванием или при помощи механического удара. Для начала кристаллизации может потребоваться дальнейшее охлаждение до некоторой новойточки, особенно если вещество обладает хорошей растворимостью. Несмотря на то, что склонность к кристаллизации увеличивается после того, как пройдена лабильная зона, все же иногда кристаллизация не происходит из-за увеличения вязкости раствора и перехода в стеклообразное состояние. Пересыщение раствора может достигаться при удалении из него некоторого количества растворителя испарением. Переход через кривую пересыщения в лабильную зону происходит редко, так как поверхность, от которой идет испарение, обычно в большей степени пересыщена, чем основная масса раствора. Кристаллы, образующиеся на этой поверхности, в конце концов попадают в раствор и затравляют его, прежде чем в основной массе раствора достигаются нужные условия. На практике чаще всего применяют сочетание охлаждения и испарения. Кристаллизация и растворение. Обычно процессы кристаллизации и растворения считают равновеликими и взаимносвязанными, и если бы они были по природе диффузионными, то тогда скорость кристаллизации должна равняться скорости растворения при данной температуре и при одинаковых движущих силах, т.е. при одинаковых отклонениях от равновесных насыщенных состояний; все грани кристалла должны расти и растворяться с 106 одинаковой скоростью. Такие условия редко достигаются на практике. Кристаллы обычно растворяются быстрее, чем растут, и различные грани обычно растут или растворяются с различными скоростями, хотя это не всегда наблюдается. Некоторые авторы считают, что кристаллы растворяются быстрее, так как открытые твердые поверхности неодинаковы в каждом случае. Когда кристалл растет, грани его плоские, а когда он растворяется, грани его обычно испещрены ямками, что приводит к увеличению площади контакта между твердой и жидкой фазами. Растворимость кристаллов зависит от их размеров. Растворимость весьма малых кристаллов значительно повышается, и при слишком малых размерах кристаллов мелкие кристаллы будут исчезать из системы, а крупные расти. Это явление вызывает некоторые затруднения в начальной фазе кристаллизации. Первые кристаллы (зародыши) не могут образовываться изза весьма малых размеров, имеют высокую растворимость. Величина кристаллов зависит от поверхностного натяжения раствора, которое может изменяться в зависимости от содержания присутствующих примесей. Чем больше поверхностное натяжение, тем сильнее его действие в направлении уменьшения общей межфазной поверхности на границе твердое вещество жидкость. Это условие ведет к образованию кристаллов больших размеров, так как их удельная поверхность (на единицу веса) меньше удельной поверхности мелких кристаллов. Следует заметить, что крупные кристаллы не могут обладать большой чистотой, вследствие агломерации содержат включения маточного раствора, в то время как мелкие кристаллы отличаются однородностью, но большая поверхность затрудняет их промывку. Рост кристаллов. Как только в пересыщенной или переохлажденной системе образовались устойчивые зародыши, т.е. частицы больше критического размера, они начинают расти, превращаясь в кристаллы видимого размера. Рост кристаллов зависит от очень многих причин. Остановимся на некоторых из них. Целый ряд таких кривых для многих систем был получен Таmmаnn. Для всех изученных систем было установлено, что оптимальная температура зарождения центров кристаллизации была гораздо ниже той, которая требовалась для максимального роста кристаллов. При температуре ниже оптимальной вязкость увеличивалась до такой величины, которая препятствовала кристаллизации, в то время как выше оптимальной температуры зарождению центров кристаллизации препятствует интенсивное молекулярное движение жидкости. Влияние перемешивания. Скорость роста кристаллов при данной температуре в постоянных условиях пересыщения может значительно изменяться при перемешивании жидкости или вращении кристалла относительно жидкости. Скорость роста значительно увеличивается на первых стадиях, но вскоре достигаются такие условия, когда дальнейшее перемешивание не оказывает никакого влияния. Из целого ряда подобных 107 исследований был сделан вывод, что диффузия не является единственным и наиболее важным фактором, который следует рассматривать при изучении процесса кристаллизации. Следует также отметить, что в кристаллизаторе, в котором кристаллы поддерживаются во взвешенном состоянии путем перемешивания, большие кристаллы будут расти быстрее, чем маленькие, так как для первых окажутся благоприятными более высокие относительные скорости реакции между твердым веществом и раствором. Форма кристаллов. Форма кристаллов зависит от очень многих факторов: например, свойства растворителя, рН раствора, наличия примесей, степени пересыщения или переохлаждения, скорости охлаждения, температуры кристаллизации, интенсивности перемешивания и т. д. Быстрое охлаждение раствора часто вызывает предпочтительный рост кристалла в одном направлении, что приводит к образованию иголок. Наиболее частым случаем, вероятно, следует считать изменение формы кристалла в присутствии примесей в кристаллизирующемся растворе. Изменение формы под влиянием примесей представляет собой, в сущности, поверхностное явление: молекулы примесей или ионы притягиваются к различным граням кристалла и поглощаются на поверхности физически или химически. Это явление уменьшает площадь, пригодную для зарождения центров кристаллизации на поверхности для осаждения растворенного вещества, и рост на той грани замедляется. Кристаллизация тетрациклинов. Наличие в молекуле антибиотиков тетрациклиновой группы 2 кислых и одной основной группировок делает возможным получение их в виде основания, соли какой-либо кислоты, а способность к комплексообразованию с тяжелыми металлами позволяет осаждать их даже из нативных растворов в виде кальциевых солей. Растворимость этих соединений в воде различна и находится в зависимости от рН водной фазы. Наименьшую растворимость тетрациклин и окситетрациклин имеют при значениях рН от 4,0 до 7,0. Это свойство антибиотиков тетрациклиновой группы позволяет их осаждать из водных растворов в виде нерастворимых кристаллических соединений. Например, осаждение основания окситетрациклина производят следующим образом. Реэкстракт с предыдущей стадии обрабатывают активированным углем в количестве 1% для удаления остатков экстрагента и пигментных веществ. Уголь отделяют, а к прозрачному фильтрату при перемешивании добавляют 10% раствор едкого натра до получения устойчивого значения рН водной фазы 4,2-4,5. Выпавший кристаллический осадок основания окситетрациклина отфильтровывают от маточника, промывают водой и высушивают в вакууме при 50°. Высокая гидрофильность хлоргидрата окситетрациклина препятствует выделению его из водных растворов даже больших концентраций. Поэтому для получения кристаллического хлоргидрата окситетрациклина необходимо применять органический растворитель, в частности метиловый спирт. 108 Из литературных данных известно, что растворимость основания окситетрациклина в метаноле при 20°С составляет 7500 ЕД/мл. Известно также, что растворимость основания окситетрациклина в метаноле возрастает в присутствии хлористого кальция. Вероятно, это повышение растворимости основания окситетрациклина в присутствии хлористого кальция можно объяснить способностью антибиотиков тетрациклиновой группы к образованию комплексных соединений с тяжелыми металлами (типа хелатов). Отмечено, что растворимость основания окситетрациклина в метиловом спирте сильно возрастает с увеличением концентрации хлористого кальция. Однако с повышением концентрации окситетрациклина возрастает вязкость раствора, вследствие чего уменьшается скорость его фильтрации и ухудшаются условия кристаллизации. С другой стороны, растворимость гидрохлорида окситетрациклина в кислом метаноле уменьшается с повышением концентрации хлористого кальция. Сопоставление этих данных привело к выводу, что оптимальным условием для получения кристаллического гидрохлорида окситетрациклина является концентрация основания 100000-120000 ЕД/мл в 7-9% растворе хлористого кальция в метаноле. При подкислении такого раствора соляной кислотой происходит быстрая кристаллизация гидрохлорида, растворимость которого в метанольном растворе данного состава минимальна. Процесс кристаллизации осуществляется следующим образом. Техническое основание окситетрациклина растворяют в 8% метанольном растворе хлористого кальция до получения указанной выше концентрации. К раствору, обработанному активированным углем, при 3-5° прибавляют концентрированную соляную кислоту. Смесь перемешивают в течение 30 мин при этой же температуре, после чего выпавший кристаллический осадок гидрохлорида окситетрациклина отфильтровывают и на фильтре промывают охлажденным метанолом. Осадок сушат в вакууме при 35-38° и получают гидрохлорид окситетрациклина в виде кристаллов лимонно-желтого цвета с активностью 900 ЕД/мг. Аналогичным образом протекает и осуществляется процесс кристаллизации двух представителей антибиотиков тетрациклиновой группы (тетрациклина, хлортетрациклина) с небольшими деталями и изменениями в каждом частном случае. Следует указать также, что процесс кристаллизации как в случае получения основания, так и в случае получения гидрохлорида окситетрациклина является процессом кристаллизации с химической реакцией. Кристаллизация пенициллина. Исключительно высокая гидрофильность калиевой и натриевой солей пенициллина не позволяет проводить кристаллизацию их из водных растворов. Кристаллизация калиевой соли пенициллина осуществляется из бутилацетатного экстракта добавлением насыщенного водного раствора ацетата калия. При этом пенициллин, находясь в бутилацетате в виде пенициллиновой кислоты, реагирует с 109 ацетатом калия; в результате химической реакции образуется калиевая соль, которая вследствие малой растворимости в бутилацетате выпадет в осадок. В данном случае кристаллизация протекает с химической реакцией. Выпавшие кристаллы промываются бутанолом для удаления ацетата калия и высушиваются. Натриевая соль пенициллина кристаллизуется методом азеотропной отгонки растворителя. В вакуум-выпарной аппарат к водному раствору пенициллина добавляется бутанол, который с водой образует азеотропную смесь, состоящую из 68 частей бутанола и 32 частей воды. Отгонка азеотропной смеси бутанола с водой производится при 20° и остаточном давлении 10 мм рт. ст., при полном удалении воды из кубового остатка, при охлаждении кристаллизуется Nа-соль пенициллина, которая затем также промывается бутанолом и высушивается. Сушка антибиотиков. Вследствие нестабильности антибиотиков даже в слабо увлажненном состоянии их выпускают в хорошо высушенном состоянии - с остаточной влажностью 0,5-2,0%. В зависимости от агрегатного состояния антибиотика в конце очистки и его стабильности при повышении температуры применяют четыре метода сушки: 1) осажденные в виде кристаллов антибиотики, имеющие сравнительно высокую стабильность (например, тетрациклиновые), подвергаются сушке при атмосферном давлении и температуре до 90°С в камерных или пневматических сушилках; 2) осажденные малостабильные антибиотики (пенициллины) высушиваются в вакуумных сушилках при техническом вакууме и температуре около 40° С - шкафах, сушилках-венулет и др. Антибиотики, полученные при выделении-очистке в виде концентрата, т.е. 5-15% водного раствора (стрептомицин, антибиотики группы неомицина), весьма нестабильные в растворенном состоянии, подвергаются сушке двумя методами, исключающими инактивацию и ухудшение качества препаратов; 3) медленная сушка (в течение нескольких часов) при отрицательной температуре путем сублимации воды из замороженного раствора под средним или глубоким вакуумом; эту сушку называют молекулярной; 4) скоростная сушка (в течение долей секунды) при высоких температурах порядка 130° в виде аэрозоля, образованного из раствора в токе горячего воздуха; эту сушку называют распылительной. Молекулярная сушка. Метод сублимационной сушки впервые был открыт и запатентован в 1921 г. советским инженером Г. И. ЛаппаСтарженецким. Метод заключается в том, что влажный материал или раствор (бактериальная масса, сыворотка крови, раствор антибиотика, фруктовый сок, фрукты, мясо, рыба), замороженные до температуры минус 20-40° С сушатся в вакууме с остаточным давлением около 0,01 мм рт. ст., путем возгонки воды из кристаллов льда. 110 При этом благодаря вакууму средняя длина свободного пробега молекул воды в порах высушиваемого материала (≈ 5 мм) значительно больше поперечника капиллярных пор (10-4 - 10-2 мм) замороженного материала, по которым удаляются пары воды. В этих условиях молекулы воды движутся в порах в виде молекулярных пучков (эффузия). Поэтому сублимационный метод сушки, по предложению А. В. Лыкова, назван молекулярной сушкой. Технология молекулярной сушки заключается в следующем. Раствор антибиотика, чтобы не допустить его вспенивания в сублимационной камере и выброса в вакуумную систему, предварительно замораживают в камерах обычных холодильных установок до температуры -40° С. Перед замораживанием раствор стерилизуют фильтрацией и разливают во флаконы с помощью полуавтоматического дозатора по 3-5 мл в зависимости от клинической дозы антибиотика. Чем быстрее происходит замораживание, тем более мелкокристаллической и более гомогенной по концентрации антибиотика становится структура образующегося льда и тем быстрее идет последующая сушка благодаря более развитой поверхности сублимации. Удаление влаги в период сублимации происходит с постоянной скоростью и при сохранении формы замороженного материала или раствора. Поскольку сушка происходит с поверхности тела, то, чем мельче дозировка, тем быстрее заканчивается сушка. Оптимальный режим сутки соответствует такой температуре и вакууму, при которых скорость сублимации наибольшая. Это достигается сушкой при постоянной температуре, немного меньшей температуры плавления продукта - криогидратной точки. Плавление продукта не допустимо, так как вспенивание раствора вызывает выброс раствора из флакона или частичный вынос его на стенки флакона. Такие «вспененные флаконы» выбраковываются при контроле качества сушки. В период сублимации (5-8 ч) удаляется около 80% влаги при температуре замороженного продукта минус 35-40° С и остаточном давлении 10-30 мм рт. ст. В конце периода сублимации температура внутренних слоев материала повышается до 0° С, и начинается период испарения остаточной влаги. В этот период материал постепенно нагревается до температуры окружающей среды (температуры теплоносителя), а скорость сушки постепенно уменьшается до нуля. Стерильные растворы антибиотиков сушатся в открытых флаконах, помещенных в кассеты и прикрытых тканью Кассеты размещают на нескольких лодках прямоугольного или цилиндрического шкафа сублиматора 1. В полые полки шкафа, после создания в системе вакуума, подают вначале холодную, а затем подогретую воду с целью подвода теплоты, необходимой для сублимации. В конце сушки температура воды повышается до 80° С. Вакуум в шкафу создается двумя ступенями насосов. Форвакуумный (или газобалластный) по конструкции ратационно-масляный насос 4, включаемый в начале сушки, создает остаточное давление в несколько мм рт. ст. Затем пароструйный диффузионный насос снижает 111 остаточное давление до 10-50 мк. Для уменьшения нагрузки на насос и защиты его от конденсата водяного пара в схеме предусмотрен скребковый или, при больших мощностях установки, трубчатый конденсатор. В скребковом конденсаторе на цилиндрической поверхности, охлаждаемой хладоагентом до температур, более низких, чем в сублиматоре, конденсируются и замораживаются пары воды, образуя слой льда. Скребком лед непрерывно снимается в льдоприемник, откуда он периодически удаляется. Распылительная сушка. Сушка распылением является одним из наиболее современных и перспективных методов обезвоживания лекарственных растворов термолабильной природы и пищевых продуктов (молока, яиц). В принципе метод сушки заключается в том, что высушиваемый раствор распыляется с помощью форсунок, струи сжатого воздуха или быстро вращающегося диска до частиц размером 5-25 мк в токе протекающего через сушильную камеру нагретого до температуры порядка 160° С воздуха. Величина поверхности частиц порядка 0,5 млн. м2 на 1 м3 раствора обеспечивает сушку в течение долей секунды. Высушенный продукт в виде порошка отделяется от отработанного теплоносителя в специальных пылеотделителях. Киносъемка процесса сушки одиночных капель раствора стрептомицина, проведенная О.А. Кремневым и соавт. (1963), позволила выявить пять этапов сушки капель коллоидных растворов: а) прогрев капли; б) сушку с постоянной скоростью при температуре, равной температуре мокрого термометра (около 40° С); в) образование при влажности капли 200250% на ее поверхности корки из сухого стрептомицина, препятствующей выходу пара, и повышение температуры капли до температуры кипения (≈ 105о С); г) сушку капли с постоянной скоростью при температуре кипения; д) сушку с падающей скоростью, приближающейся к температуре окружающей среды. Такой характер кинетики сушки капель привел авторов к выводу о целесообразности разделения процесса обезвоживания высоковлажных растворов антибиотиков на два этапа с целью интенсификации и удешевления процесса: 1. Испарение от начальной влажности, равной 1600-2000%, до влажности коркообразования. Поскольку на этом этапе капля имеет низшую температуру, длительность ее пребывания в камере, можно выбирать произвольно. Теплоноситель целесообразно использовать в максимальной степени, т.е. удалять его с температурой на 3-5° С выше температуры раствора, а относительную влажность доводить до 90%. Скорость теплоносителя по сечению камеры для интенсификации процесса можно брать в 3-8 раз больше, чем в сушильной камере, а размеры камеры делать в 1,5-2 раза меньше размера факела распыленного материала. 2. Сушка от влажности коркообразования (200-250%) до конечной влажности 1,5-2% в течение минимального временя, поскольку испарение 112 термолабильного раствора происходит при температуре его кипения. Это достигается применением очень сухого воздуха (конечная относительная влажность 4-6%) и малым расходом высококонцентрированного раствора. Такие условия позволяют также увеличить скорость теплоносителя по сравнению с одноступенчатой сушилкой. Отработанный теплоноситель со ступени сушки с температурой 100-105° С подается на испарительную ступень и полностью используется. Применение такого двухступенчатого испарительно-сушильного метода обезвоживания стрептомицина позволило при внедрении на Киевском заводе медицинских препаратов по сравнению с одноступенчатой распылительной сушилкой датской фирмы «Niro atomizer» увеличить коэффициент использования тепла теплоносителя с 25 до 75%, повысить выход антибиотика (с учетом потерь на вакуум-выпарке, необходимой при одноступенчатом режиме) с 85 до 93% и снизить себестоимость процесса обезвоживания стрептомицина с 0,98 до 0,34 руб./кг стрептомицина (О.А. Кремнев и соавт., 1963). Воздух всасывается из атмосферы через тканевой фильтр грубой очистки вентилятором и нагнетается через фильтр Петрянова, паровой калорифер и электрокалориферы в испарительную ступень емкостью 7 м3. Воздух с температурой 165°С подается в камеру через газораспределительное устройство. Стерильный раствор стрептомицина подается на диск, вращающийся со скоростью 18 000 об/мин. Поток воздуха, проходя между потолком камеры и радиальным факелом распыляемого раствора, предупреждает оседание раствора на потолке камеры. Затем воздух поворачивает вниз. Испарение раствора уменьшает его объем в четыре раза. Раствор с влажностью 250-300% отделяется на 98-99% от воздуха в мокром циклоне. Воздух выбрасывается в атмосферу, а раствор немедленно распыляется на диске сушильной камеры емкостью 17 м3. Подача теплоносителя в сушильную камеру производится из аналогичной системы стерилизации и нагрева атмосферного воздуха. Порошок антибиотика отделяется от отработанного воздуха в циклоне и выгружается в герметичное устройство для стерильной фасовки антибиотика в контейнеры. Отработанный воздух, содержащий около 7% порошка антибиотика, направляется в испарительную камеру, где, благодаря малой влажности, используется как вторичный сушильный агент. Сушильные камеры изготовляются из полированной изнутри камеры нержавеющей стали и имеют форму цилиндра с конусным днищем. Производительность сушилки 200 л/час испаренной влаги. Система автоматических приборов регулирует температуру теплоносителя на входе в камеры и расход раствора. Сравнение молекулярной и распылительной сушки. С экономической точки зрения оба метода в настоящее время приблизительно равноценны. Молекулярная сушка требует в 2,7 раза больше производственной площади и в 2,1 раза больший расход электроэнергии. Трудовые затраты также выше. 113 Вместе с тем выход антибиотиков при молекулярной сушке (97-98%) выше, чем при распылительной сушке (92-94%). Если же учесть потери при фасовке порошка антибиотиков, то снижение выхода ниже 90%, наблюдающееся в практической работе предприятий, сводит на нет экономические преимущества распылительного метода. При более тщательной отработке режима недавно освоенной двухступенчатой распылительной сушилки и устранения конструктивных недостатков, ее экономические показатели станут несомненно выше показателей молекулярной сушки. Однако молекулярная сушка имеет преимущества в более важном, чем экономический, показателе для антибиотиков медицинского парентерального применения - качестве препаратов. Сушка при отрицательной температуре под вакуумом обеспечивает гарантию сохранения качества - растворимости, бесцветности, апирогенности, отсутствию опалесценции растворов и пр. Отсутствие последующей фасовки обеспечивает сохранение стерильности. При распылительной сушке 7% антибиотика, возвращаемые со второй ступени на первую, сушатся повторно. Этот и некоторые другие технические особенности процесса (например, полидисперсность распыла) снижают качество продукта. По мере освоения распылительной сушки качество сухого продукта будет повышаться. В производстве антибиотиков перспективны оба рассмотренных метода сушки растворов антибиотиков. 9.2. Применение антибиотиков. Антибиотические вещества находят применение в различных отраслях народного хозяйства, в научных исследованиях. Они широко используются в медицине, в сельском хозяйстве, в пищевой и консервной промышленности, как специфические ингибиторы в биологических исследованиях. Антибиотики в медицине. Открытие антибиотиков вызвало переворот в медицине. Многие антибиотические вещества оказались незаменимыми лечебными препаратами. Они нашли широкое применение при лечении многих инфекционных заболеваний, которые ранее считались неизлечимыми или сопровождались высоким летальным исходом. К числу таких заболеваний необходимо отнести некоторые формы туберкулеза и, прежде всего туберкулез менингитный, который до применения антибиотиков вызывал 100%-ный летальный исход, чуму, азиатскую холеру, брюшной тиф, бруцеллез, пневмонию, различные септические процессы и др. Некоторые антибиотики способны подавлять развитие злокачественных опухолей и проявлять активность в отношении ряда вирусов. К настоящему времени в медицинской практике нашло применение около ста антибиотиков. Поиски новых антибиотических веществ, получение ценных полусинтетических препаратов антибиотиков расширяют возможность их практического применения в медицине. 114 Антибиотики в сельском хозяйстве. Наряду с медицинским использованием антибиотики находят широкое применение в сельском хозяйстве. Прежде всего, антибиотики используются в качестве препаратов в ветеринарии для лечения различных заболеваний сельскохозяйственных животных. В этом случае они, как и в медицине, оказались весьма эффективными средствами. Антибиотические вещества находят все возрастающее применение в борьбе с фитопатогенными организмами - возбудителями заболеваний растений, наносящими ощутимый урон сельскохозяйственному производству. При выборе антибиотиков, используемых в растениеводстве, необходимо руководствоваться следующими основными требованиями к препарату: 1) антибиотик должен обладать специфической биологической активностью к возбудителю заболевания растений; 2) он должен легко проникать в ткани растений и проявлять внутри них биологическую активность; 3) лечебные дозы антибиотика должны быть безвредными для растения; 4) антибиотик, находясь на поверхности и внутри тканей растения, должен относительно длительное время проявлять биологическую активность, но должен также легко и быстро инактивироваться, попадая в почву. Одним из самых главных требований к антибиотикам, используемым в сельском хозяйстве, должно быть то, чтобы эти препараты не применялись в медицинской практике. Это принципиальное условие обеспечивает снижение уровня появления резистентных форм микроорганизмов, патогенных для человека. Для борьбы с фитопатогенными организмами могут применяться различные антибиотики. Гризеофульвин - образуется плесневыми грибами из рода Penicillum (P. urticae, P. nigricans, P. rustrichi). Строение антибиотика см. с. . Гризеофульвин применяют против фитопатогенных грибов и, прежде всего грибов, относящихся к роду Botrytis. Он активен в отношении возбудителя ржавчины, мучнистой росы, килы капусты. Трихотецин - продуцируется плесневым грибом Trichothecium roseum. Антибиотик подавляет развитие ряда фитопатогенных грибов, в том числе Botrytis cinerea, Helmintosporium. Попадая в почву, трихотецин инактивируется ферментом трихотециназой, образуемой почвенными грибами из рода Fusarium, Aspergillus, Penicillum. Касугамицин, продуцируемый Streptomyces kasugaensis, имеет следующее строение: 115 Активен против грибного заболевания риса пирикуляриоза, широко распространенного в Японии. Заболевание вызывается грибом Pericularia oryzae. Полиоксины - антибиотики, относящиеся к своеобразным соединениям пептидил-пиримидин-нуклеозидам, обладают противогрибковой активностью. Образуются культурой Streptomyces cacaoi и имеют следующее строение: Антибиотики подавляют рост фитопатогенных грибов из рода Alternaria, Cochliobalus, Pirularia. Валидамицин А, образуемый Streptomyces hygroscopicus var. limoneus, обладает биологической активностью против фитопатогенного гриба Rizoctonia solani - возбудителя заболевания риса. Известна суммарная формула валидамицина A: C20H35NO13∙H2O. Антибиотик легко разлагается почвенными микроорганизмами. Тетранастин антибиотическое вещество актиномицетного происхождения. Его продуцентом является Streptomyces aureus. Обладает специфической активностью против паразитарных паучков и клещей плодовых деревьев. Гербицидины А и В, синтезируемые Streptomyces saganoensis, подавляют развитие возбудителя заболевания риса, вызываемое Xanthomonas oryzae. Гербицидин А задерживает прорастание семян риса и китайской капусты, обладает избирательной гербицидной активностью против двудольных растений. Гербицидины А и В по химическому строению и биологической активности близки к тойокамицину, образуемому культурой Str. toyocaensis. Антибиотики в пищевой и консервной промышленности. При борьбе с микроорганизмами, вызывающими порчу продуктов питания, наряду с физическими и химическими методами применяются и антибиотики. Однако для этих целей не могут быть использованы антибиотики, применяемые в медицине. Это правило введено в нашей стране и ряде других государств и связано оно с предупреждением процесса возникновения и распространения устойчивых к антибиотикам форм микробов. Среди антибиотиков, применяемых в пищевой и консервной промышленности, можно назвать субтилин, низин и некоторые другие. Субтилин образуется культурой Bacillus subtilis и представляет собой полипептид. Активен в отношении грамположительных и грамотрицательных микроорганизмов, в том числе и кислотоустойчивых бацилл. Применяя субтилин при консервировании овощей, можно значительно смягчить их термическую обработку, что имеет большое значение для сохранения витаминов, вкусовых качеств и консистенции продукта. 116 Низин - высокомолекулярный пептид, а может быть даже низкомолекулярный белок, образуемый Streptococcus lactis. Низин не используется в медицинской практике. Его применяют при консервировании томатов, зеленого горошка, цветной капусты и других продуктов. Хорошие результаты получены при сохранении сыров. Антибиотик подавляет развитие ряда термофильных спорообразующих бактерий, не токсичен для человека. Применение антибиотиков в растениеводстве, пищевой и консервной промышленности должно происходить под постоянным и строгим контролем специалистов и соответствующих компетентных органов. ГЛАВА 10. Получение меланинов. Меланины (от греч. melas, родительный падеж melanos - черный), обычно черные или темно-коричневые пигменты животных, растений и микроорганизмов. У высших животных и человека меланины - основная группа пигментов. У животных меланины придают окраску шерсти, у птиц оперению, у человека ответственны за цвет глаз, волос, окраску кожи. Меланины - аморфные высокомолекулярные вещества. Обширную группу природных конденсированных фенолов составляют меланиновые пигменты, имеющиеся у всех живых организмов. Охарактеризовать их можно как класс соединений с низкой токсичностью, обладающих разнообразным биологическим действием. Эти свойства могут найти широкое применение в ряде областей профилактической и клинической медицины. Наибольшее применение и более глубоко изучены меланины млекопитающих и растений. Не растворимы в воде, минеральных кислотах, орг. растворителях; хорошо растворяются в щелочах, а затем выпадают в осадок при подкислении растворов, что используется для их выделения. Хим. разрушение меланинов происходит при нагревании выше 200°С, сплавлении со щелочью, окислении концентрированными растворами КМnО4 или Н2О2. По предшественникам меланинов в организме их разделяют на эумеланины, феомеланины и алломеланины. Эумеланины (черные) и феомеланины (желтые, красные и коричневые) распространены у животных, алломеланины (черные) - и в растениях, грибах, бактериях. Предшественник эумеланинов -тирозин; из него в организме получаются пигменты, содержащие С, Н, N и О. Предшественники феомеланинов - тирозин и цистеин; в организме превращается в серосодержащие пигменты. Предшественники алломеланинов - дифенолы (пирокатехин и др.); из них образуются меланины, не содержащие азота. В организмах меланины находятся в комплексе с белками. Многие свойства и биологические функции меланинов определяются их способностью функционировать в организме в виде системы фенолсемихинон-хинон. Механизм образования меланинов окончательно не выяснен. Полагают, что первые стадии биосинтеза меланинов являются 117 ферментативными и катализируются о-дифенол: кислород оксидоредуктазами (например, тирозиназой, полифенолоксидазой и др.), последующие стадии протекают спонтанно с участием свободных радикалов. Меланины могут быть получены автоокислением ДОФА (3,4дигидроксифенилаланина), 5,6-дигидроксииндола и др. дифенолов. Меланины характеризуются наличием в их структуре неспаренных электронов и обладают свойствами стабильных своб. радикалов. Эта особенность меланинов важна для выполнения ими защитные функций в организме. Меланины не только поглощают различные излучения, но и нейтрализуют и обезвреживают опасные для клеток своб. радикалы, образующиеся при действии ионизирующего излучения и некоторых хим. веществ на живые организмы. Меланины могут существовать в нескольких окислительновосстановительных состояниях. Обладают электрон-транспортными свойствами. У животных и человека процесс образования меланинов в организме происходит в специальных клетках - меланоцитах и регулируется гормональной системой, главным образом с помощью гормонов гипофиза (aи b- меланоцитстимулирующих гормонов). Некоторые физ. факторы (солнечные, УФ и рентгеновские лучи) и хим. реагенты стимулируют образование меланинов в организме. Повышенное содержание меланинов наблюдается также в некоторых злокачественных опухолях-меланомах. Получение меланина из грибов. Меланин из Aspergillus niger был получен согласно общепринятой методике. Культивирование продуцента проводили на минеральной среде, содержащей глюкозу, при 26ºС до появления сильной пигментации мицелия. Мицелий отделяли фильтрованием, промывали дистиллированной водой и замораживали при – 16 ºС. Мицелий подвергали щелочному гидролизу (0,1 М NaOH) с последующим осаждением концентрированной HCl. Полученный осадок центрифугировали при 7000 g в течение 30 мин, промывали дистиллированной водой, затем снова центрифугировали и диализовали против дистиллированной воды. Диализ проводили не менее 24ч и заканчивали при достижении диализуемого раствора pH 4,5. Диализированный препарат грибного меланина высушивали при 40 ºС. Такой же методикой были получены из базидиомицетов, как Inonotus obliquus (трутовик скошенный или чага), Phellinus robustus (трутовик ложного дуба) и Phellinus igniarius. Очищенные меланиновые пигменты могут быть использованы для создания фармакологических препаратов, обладающих разнообразными протекторными свойствами, к примеру, противовоспалительный и противоязвенный препарат «Бефунгин», получен из чаги берѐзовой – стерильной формы гриба Inonotus obliquus. Способ микробиологического получения меланина. Изобретение относится к области микробиологического синтеза органических соединений. При реализации способа проводят культивирование штамма дрожжей 118 Saccharomyces neoformans var. nigricans ГСП - ТБ 17 Д на углесодержащих средах. Процесс проводят при 27 - 35oС и рН культуральной жидкости 4,5 5,5 в условиях аэрации и перемешивании. Полученную биомассу отделяют от культуральной жидкости с последующим выделением целевого продукта. Способ позволяет сократить время синтеза и повысить выход меланина. Меланины представляют собой черные, коричневые и желтые пигменты. Молекулы меланинов представляют собой сложные комплексы, образованные полимерами производных тирозина и белков. Молекулярная масса меланинов зависит от способа его получения. Следующие примеры получения меланина: 1. Дрожжи штамма Saccharomyces neoformans var. nigricans ГСП-ТБ 17 Д выращивали в аппарате, конструкция которого обеспечивала аэрацию культуральной жидкости при перемешивании и термостатировании, объемом 5 дм3 на питательной среде, содержащей в качестве источника углерода отход спиртового производства, в количестве 4% по объему от объема культуральной жидкости, а также в качестве источников минерального питания сульфат аммония (6 г/дм3), однозамещенный фосфат калия (1,5 г/дм3), сульфат магния (0,5 г/дм3), а также сульфат марганца, сульфат цинка, сульфат железа (II) и сульфат меди по (0,05 г/дм3). А рН культуральной среды составило 5,5 при температуре 30oС. Режим аэрации составил 1 об/об*мин. При этом в течение 20 часов культивирования была получена суспензия, содержащая 3,9% биомассы. Биомассу обработали 0,5 N гидроксида натрия при давлении 0,5 атм в течение 30 мин. Клетки дрожжей осадили центрифугированием. Фугат подкислили 40%-ной соляной кислотой до рН 3,2. В результате вышеуказанных операций меланин выпал в осадок. Осадок центрифугировали при 5000 об/мин в течение 15 мин с последующей последовательной промывкой водой и этиловым спиртом. В результате было получено 4,5 г меланина, что составило выход 2,5% от субстрата и 3,1% от содержания биомассы. 2. Дрожжи штамма Saccharomyces neoformans var. nighcans ГСП-ТБ 17 Д выращивали в аппарате, конструкция которого обеспечивала аэрацию культуральной жидкости при перемешивании и термостатировании, объемом 5 дм3 на питательной среде, содержащей в качестве источника углерода смесь мелассы с углеводородами в соотношении 10:1 в количестве 6% по объему от объема культуральной жидкости, а также в качестве источников минерального питания сульфат аммония (5,8 г/дм3), однозамещенный фосфат калия (1,6 г/дм3), сульфат магния (0,6 г/дм3), а также сульфат марганца, сульфат цинка, сульфат железа (II) и сульфат цинка по (0,045 г/дм3). рН культуральной среды составило 5,6 при 30oС. Режим аэрации составил 1,1 об/об*мин. При этом в течение 18 ч культивирования была получена суспензия, содержащая 3,5% биомассы. Биомассу обработали 0,5 N гидроксида калия при давлении 0,5 атм в течение 30 мин. Клетки дрожжей осадили центрифугированием при 5000 об/мин в течение 20 мин. Фугат содержал 70% меланина от а.с.в, 15% белков от а.с.в. и 6% липидов от а.с.в. 119 В результате выход продукта составил 13,8% от исходной биомассы, при этом чистого меланина получено 6,9% от содержания биомассы. 3. Дрожжи штамма Saccharomyces neoformans var. nigricans ГСП - ТБ 17 Д выращивали в аппарате, конструкция которого обеспечивала аэрацию культуральной жидкости при перемешивании и термостатировании, объемом 5 дм3 на питательной среде, содержащей в качестве источника углерода отход спиртового производства, в количестве 4% по объему от объема культуральной жидкости, а также в качестве источников минерального питания сульфат аммония (6 г/дм3), однозамещенный фосфат калия (1,5 г/дм3), сульфат магния (0,5 г/дм3), а также сульфат марганца, сульфат цинка, сульфат железа (II) и сульфат меди по (0,05 г/дм3). рН культуральной среды 5,5 при температуре 30oС. Режим аэрации составил 1 об/об*мин. При этом в течение 18 ч культивирования была получена суспензия, содержащая 3,3% биомассы. Биомассу обработали ультразвуковыми колебаниями частотой 50100 кГц в течение 5 мин. Клетки дрожжей осадили центрифугированием. Фугат пропустили через мембранный фильтр фирмы Millipore с размером пор 0,45 мкм. Полученный продукт содержал 98% меланина от а.с.в. 4. Для получения путем микробиологического синтеза меланина в качестве углеродсодержащего сырья использовали молочную сыворотку. Дрожжи штамма Saccharomyces neoformans var. nigricans ГСП-ТБ 17 Д выращивали в аппарате, конструкция которого обеспечивала аэрацию культуральной жидкости при перемешивании и термостатировании, объемом 10 дм3 на молочной сыворотке с содержанием сухих веществ 6%. В качестве источников минерального сырья использовали сульфат аммония (4 г/дм3), однозамещенный фосфат калия (1,0 г/дм3), сульфат меди (0,1 г/дм3), сульфат железа (II) (0,01 г/дм3). рН культуральной среды составило 5,5 при 30 oС. Режим аэрации составил 1 об/об•мин. При этом в течение 20 ч культивирования была получена суспензия, содержащая 2,6% (вес.) биомассы. Для извлечения меланина биомассу обработали в ультразвуковом дезинтеграторе при частоте колебаний 100 кГц в течение 30 мин, затем дезинтегрированные клетки дрожжей осадили центрифугированием. Фугат подкислили соляной кислотой до рН 3,0 и отделили центрифугированием осадок. Полученный продукт содержал 91% меланина от а.с.в. В результате проведенных экспериментальных работ установлено, что молекулярный вес получаемого меланина зависит от условия проведения эксперимента. ГЛАВА 11. Основные принципы получения алкалоидов. Алкалоиды - гетероциклические азотосодержащие вещества щелочного характера, обладающие сильным физиологическим действием. Во второй половине 18 века и в начале 19 века, при изучении химического состава растений, были выделены отдельно сложные производственные гетероциклов, получившие впоследствии объединяющее название 120 "алкалоиды". Сам термин был введен В. Мейснером в 1819 году: по-латыни alkali-щелочи, oides-подобный, то есть "подобные щелочам". Современное определение алкалоидам впервые было дано в 1910 году Э. Винтерштейном и Г. Триром. В определении были сформулированы основные условия, которым должен отвечать истинный алкалоид: 1. растительное происхождение; 2. сложная молекулярная структура соединения; 3. атом азота - часть гетероциклической системы; 4. проявление высокой фармакологической активности. На сегодняшний день классическое определение Винтерштейна -Трира несколько устарело, так как соединения, рассматриваемые большинством химиков и формаколагов как алкалоиды, не отвечают всем его требованиям. Это не удивительно, ведь известно свыше 5000 алкалоидов, строение установлено примерно для 3000. Обратим внимание на некоторые недостатки определения Винтерштейна - Тирра: на сегодняшний день известны многие вещества со структурой классических алкалоидов, но полученные из материалов нерастительного происхождения (бактерий, грибов, тканей животных). Определение "сложная молекулярная структура соединения" слишком расплывчато и не дает представления о положении в целом. Термин "высокая фармакологическая активность" - применен не слишком корректно, так как фармакологическая активность веществ во многом зависит от уровня доз, и проявляется, при достаточном содержании многими веществами, не имеющими отношения к алкалоидам. Учитывая все вышеперечисленное было предложено новое определение понятия "алкалоид", которое безусловно охватывает большее число соединений, исключая тем не менее такие классы природных азотсодержащих соединений, как аминокислоты, аминосахара, алифатические амины, белки и пептиды, нуклеотиды, нуклеиновые кислоты, витамины, птерины, порфирины. Новое определение было сформулировано У. Плетье: "Алкалоид - это циклическое органическое соединение, содержащее азот в отрицательной степени окисления и имеющее органическое распространение среди живых организмов". Определение, отвечает необходимым условиям и получило повсеместное признание. Достоинство определения алкалоидов, предложенного Плетье, состоит в том, что подтверждает отнесение к алкалоидам большинства тех соединений, которые, хотя традиционно считаются алкалоидами, не подходят под классическое определение Винтерштейна - Трира. Это, например, колхицин, пиперин, b -фенилэтиламины, рицинин, генцианин, буфотоксин. Способы выделения алкалоидов Главной проблемой при выделении алкалоидов является отделение вещества от "балластного" материала, составляющего главную массу растительного сырья. В случае с легколетучими алкалоидами, выделение производится путем отгонки с водяным паром, однако такие случаи редки. 121 Так как обычно алкалоиды находятся в растении в виде различных кислот, то необходимо сначала освободить их путем смачивания измельченного растения раствором щелочи. Нередко для извлечения алкалоидов прибегают к экстракции при помощи подходящих растворителей. Подобные методики выделения делятся на две группы: экстракция в виде солей и экстракция в виде свободных оснований. Экстракция в виде солей. В первом случае растительное сырье обрабатывается подходящим растворителем, к которому прибавляется небольшое количество какой-либо кислоты (уксусной, соляной, винной, лимонной и др.). Экстракция ведется обычно в конических экстракционных аппаратах (перколяторах), в которые загружается мелко размолотое сырье и заливается растворитель. После настаивания в течение нескольких часов раствор медленно выпускают через кран, имеющийся в нижней части перколятора, а сырье снова заливают свежим растворителем и продолжают так до полного извлечения, т.е. до того момента, когда в пробе жидкости, стекающей из перколятора, при помощи подходящих качественных реакций не удается больше открыть присутствие алкалоида. Еще более совершенной является непрерывная перколяция; при этом способе, по мере того как из крана перколятора медленно сливается раствор, в верхнюю его часть автоматически добавляется такое же количество свежего растворителя. При наличии аппаратуры целесообразно использование экстракцию в нескольких перколяторах по принципу противотока: раствор, вытекающий из первого перколятора, поступает на свежее сырье, находящееся в во втором перколяторе; из второго обогащенный раствор поступает на свежее сырье - в третьем перколяторе и т.д. Этим путем удается получить более концентрированные растворы алкалоида и обойтись с меньшим количеством растворителя. На производстве устанавливаются "экстракционные батареи", состоящие из 5-10 перколяторов. Соли алкалоидов обычно растворимы в воде, спиртах (метиловом и этиловом) и нерастворимы в эфире и углеводородах. Поэтому при извлечении алкалоидов в виде солей в качестве растворителя обычно применяется вода или спирт. Хотя экстракция алкалоидов идет в большинстве случаев легко и быстро, однако этот способ имеет недостаток, что спирт, а особенно вода, извлекает из растений наряду с алкалоидами большое количество так называемых "экстрактивных веществ" (белки, смолы, дубители, слизи и др.), присутствие которых часто сильно затрудняет обработку таких растворов. Экстракция алкалоидов в виде свободных оснований. При этом методе необходимо предварительно выделить алкалоиды, находящиеся в растении в виде солей, что достигается обработкой щелочью. Иногда для этого слегка влажный порошок растительного сырья тщательно смешивают с сухим основанием (окись магния или известь), а затем 122 подвергают экстракции. В других случаях растение смачивают и тщательно растирают с раствором щелочи (аммиак, сода, едкий натр) и затем подвергают экстракции в перколяторе. Так как свободные алкалоиды растворимы не только в воде и спирте, но и в большем числе органических растворителей, то выбор подходящего растворителя в этом случае гораздо богаче. Чаще всего для этой цели применяют бензол, дихлорэтан, реже пользуются эфиром, хлороформом, четыреххлористым углеводородом, петролейным эффиром и керосином. Сама экстракция ведется путем перколирования совершенно так же, как в случае экстракции в кислой среде. Выбор подходящей щелочи является очень важным моментом, так как многие алкалоиды очень чувствительны к действию сильных щелочей и могут при этом подвергаться нежелательным изменениям, также могут встретить случаи, когда алкалоид представляет собой настолько сильное основание, что для его выделения из солей недостаточно слабых оснований вроде аммиака. Предварительная экстракция. Для разделения смеси алкалоидов от балластных веществ применяется способ предварительной очистки сырья. Для этого сырье сначала обрабатывают какой-либо слабой кислотой (или солью, имеющей слабокислую реакцию) и подвергают экстракции бензолом или петролейным эфиром. Алкалоиды, будучи связаны в виде солей, в эти растворители не переходят, а растворитель извлекает только нейтральные или кислые экстрактивные вещества. После этой предварительной обработки растительный материал снова обрабатывают подходящей щелочью и вторично извлекают по тому же способу. Раствор алкалоидов при этом получается значительно более чистым, содержащим гораздо меньше посторонних веществ, и выделение из него чистых оснований значительно облегчается. Однако, вследствие громоздкости и большой затраты времени этот метод применяется только в исключительных случаях, там где приходится иметь дело с сырьем, особо богатым балластными веществами, или в случае очень чувствительных и легко изменяющихся алкалоидов. Экстракты, полученные тем или иным способом, содержат алкалоиды (и балластные вещества), или в виде солей, или уже в свободном виде. Сообразно с этим дальнейшая обработка их несколько отличается. Далее проводится обработка экстрактов. При этом для выделения алкалоидов из водных, кислых растворов их солей эти растворы подщелачиваются и алкалоиды отсасываются (если они труднорастворимы в воде и прямо выпадают в твердом виде) или же извлекаются подходящим растворителем (эфиром, хлороформом, бензолом, амиловым спиртом и др.), не смешивающимся с водой. Такую обработку часто проводят многократно. В случае спиртовых кислых растворов необходимо сначала удалить спирт, что делается путем отгонки на водяной бане; остающаяся после этого густая масса обрабатывается водой (или разбавленной кислотой), причем 123 часть смолистых веществ остается нерастворенной и отделяется путем фильтрации. Эти смолы часто адсорбируют значительное количество алкалоидов, так что приходиться обрабатывать их несколько раз горячей водой (или разбавленной кислотой) до полного выделения из них алкалоидов. В последнее время для выделения алкалоидов из водных или кислых диффузионных соков применяется более простой метод адсорбции. В качестве адсорбента обычно применяются угли и ионообменные адсорбенты: природные глины или искусственные смолы. Растворы свободных алкалоидов в не смешивающемся с водой растворителе, полученные путем щелочной экстракции растения, обычно значительно чище, чем водные и спиртовые экстракции, т.е. содержат меньше балластных веществ. Для получения из них алкалоидов эти растворы сначала взбалтываются с разбавленной кислотой (1-5 %), в которую переходят все алкалоиды. Последние, таким образом, сразу концентрируются в сравнительно небольшом объеме жидкости. Этот кислый раствор подвергается обычной очистке, подщелачивается и алкалоидная смесь или отсасывается или снова извлекается органическим растворителем. Разделение алкалоидов. Разделение алкалоидов на основании различных температур кипения. Этот метод используется в случае, когда алкалоиды, находящиеся в смеси, сильно отличаются один от другого по своей температуре кипения. Таким образом, возможно разделить их путем дробной перегонки. Методы, основанные на различии растворимости. Различие в растворимости алкалоидов и их солей в различных растворителях является основой наиболее часто применяемых методов их разделения и очистки. Уже при извлечении "суммы алкалоидов" из первичного кислого раствора, полученного при экстракции, можно, путем применения различных несмешивающихся органических растворителей, достигнуть частичного разделения смеси. Так, например, при взбалтывании подщелоченного раствором эфиром в последний переходит часть алкалоидов, тогда как часть остается в водном растворе и извлекается из него только применением другого растворителя, например, хлороформа или бензола. Такое частичное разделение алкалоидной смеси на две или более группы, применялось например, в случае алкалоидов кактуса - Anhalonium . Это разделение, конечно, никогда не бывает полным и представляет собой только грубую первую фракционировку. Разделение на основании различной "силы основности". Метод основан на том, что различные алкалоиды обладают различной "силой Основности". Если к смеси таких алкалоидов прибавить количество кислоты, недостаточное для нейтрализации всей массы, то в первую очередь с кислотой свяжутся наиболее сильные основания, тогда как более слабые останутся свободными. Наоборот, если к раствору смеси алкалоидов в 124 теоретическом количестве какой-либо кислоты прибавить количество щелочи, недостаточное для освобождения всей суммы алкалоидов, то в первую очередь разложатся соли наиболее слабых оснований, тогда как сильные останутся в связанном с кислотой виде. Разделение этим способом обычно бывает неполным, в особенности при сложных смесях, и в отдельных фракциях наблюдается обогащение одним из оснований. Для полного разделения и очистки этот метод комбинируют с другими способами, основанными на кристаллизации солей или свободных оснований. Разделение на основании различной адсорбционной способности (хроматография). Метод хроматографии состоит в том, что через колонку, наполненную адсорбентом, пропускается испытуемый раствор, содержащий несколько алкалоидов. После того как раствор полностью проникает в слой адсорбента, колонку промывают органическим растворителем или смесью нескольких растворителей и собирают отдельные фракции вытекающей из колонки жидкости. Дальнейшая обычная обработка отдельных фракций позволяет выделять индивидуальные соединения. Разделение путем получения производных Он используется в случаях, когда алкалоиды, находящиеся в смеси, отличаются одни от других такими химическими особенностями, которые позволяют произвести их разделение путем получения каких-либо подходящих производных. Этот метод разделения основывается на том, что один из алкалоидов вступает в реакцию с каким-либо реактивом, тогда как другой остается неизменным. Свойства образовавшегося таким образом производного первого алкалоида (растворимость и пр.) часто сильно отличаются от свойств исходного вещества и позволяют провести разделение его обычным методом кристаллизации. Основным условием при этом является то, что алкалоид должен легко обратно получаться из полученного производного и не должен претерпевать при этом никаких существенных изменений. ГЛАВА 12. Получение продуктов брожения 12.1.Молочнокислое брожение При молочнокислом брожении конечным продуктом является молочная кислота. С этим брожением люди знакомы издавна. Сквашивание молока, приготовление простокваши, кефира, квашение овощей — результаты молочнокислого сбраживания сахара молока или углеводов растений. Этот вид брожения осуществляется с помощью молочнокислых бактерий, которые подразделяются на две большие группы (в зависимости от характера брожения): гомоферментативные, образующие из сахара только молочную кислоту, и гетероферментативные, образующие, кроме молочной кислоты, спирт, уксусную кислоту, углекислый газ. 125 Гомоферментативное молочнокислое брожение вызывают бактерии рода Lactobacillus и стрептококки. Они могут сбраживать различные сахара с 6-ю (гексозы) или 5-ю (пентозы) углеродными атомами, некоторые кислоты. Однако круг сбраживаемых ими продуктов ограничен. У молочнокислых бактерий нет ферментативного аппарата для использования кислорода воздуха. Кислород для них или безразличен, или угнетает развитие. Молочнокислое брожение может быть описано уравнением: C6H12 O6→2CH3∙CHOH∙COOH+21.8∙104дж Гетероферментативное молочнокислое брожение — процесс более сложный, чем гомоферментативное: сбраживание углеводов приводит к образованию ряда соединений, накапливающихся в зависимости от условий процесса брожения. Одни бактерии образуют, помимо молочной кислоты, этиловый спирт и углекислоту, другие — уксусную кислоту; некоторые гетероферментативные молочнокислые бактерии могут образовывать различные спирты, глицерин, маннит. Гетероферментативное молочнокислое брожение вызывают бактерии рода Lactobacterium и рода Streptococcus. Химизм этих брожений изучен не так хорошо, как спиртового или гомоферментативного молочнокислого брожения. Гетероферментативные бактерии образуют молочную кислоту иным путем. Последняя стадия — восстановление пировиноградной кислоты до молочной — та же самая, что и в случае гомоферментативного брожения. Но сама пировиноградная кислота образуется при ином расщеплении глюкозы — гексозомонофосфатном. Выход энергии гораздо меньше, чем при спиртовом брожении. Гетероферментативные бактерии сбраживают ограниченное число веществ: некоторые гексозы (причем определенного строения), пентозы, сахароспирты и кислоты. Молочнокислое брожение широко используется при выработке молочных продуктов: простокваши, ацидофилина, творога, сметаны. При производстве кефира, кумыса наряду с молочнокислым брожением, вызываемым бактериями, имеет место и спиртовое брожение, вызываемое дрожжами. Молочнокислое брожение происходит на первом этапе изготовления сыра, затем молочнокислые бактерии сменяются пропионово кислыми. Молочнокислые бактерии нашли широкое применение при консервировании плодов и овощей, в силосовании кормов. Чистое молочнокислое брожение применяется для получения молочной кислоты в промышленных масштабах. Молочная кислота находит широкое применение в производстве кож, красильном деле, при выработке стиральных порошков, изготовлении пластмасс, в фармацевтической промышленности и во многих других 126 отраслях. Молочная кислота также нужна в кондитерской промышленности и для приготовления безалкогольных напитков. Типичное и нетипичное молочнокислое брожение. Возбудителями типичного молочнокислого брожения "являются следующие микроорганизмы: 1) Bact. Streptococcus lactis небольшая бесспоровая палочка, молодая культура имеет вид стрептококка; оптимальная температура для развития этой бактерии 30—35°; она сбраживает лактозу, глюкозу, галактозу и мальтозу; молочной кислоты накапливается в среде около 1 %; 2) Bact. Delbriicki представляет собой длинную бесспоровую палочку, развивающуюся при температуре 45—48°; используется для производственного получения молочной кислоты, количество которой достигает 2,2%; в присутствии нейтрализующих агентов (мел) можно накопить до 10% молочной кислоты; 3) Bact. bulga-ricum или болгарская палочка, представляющая неподвижную бесспоровую палочку, длиной от 10 до 20 ц, оптимальная темпе-| ратура для ее жизнедеятельности 40—48°. Эта бактерия накапливает наибольшее количество молочной кислоты, равное 3,2%. Молочнокислые бактерии могут сбраживать различные сахара в зависимости от наличия в них тех или иных ферментов. Нетипичным молочнокислым брожением называется такое, при котором наряду с молочной кислотой образуются и другие продукты, например этиловый спирт, уксусная и янтарная кислота, углекислота и водород. Количество получающихся продуктов колеблется. В среднем молочной кислоты образуется до 40% от сброженного сахара, янтарной кислоты около 20%, уксусной кислоты и этилового спирта по 10% и 20% составляют газы. Представителями бактерий, вызывающих нетипичное молочнокислое брожение, являются В. coif, В. pentoaceticum и др. Нетипичному молочнокислому брожению подвергаются также пентозы. При брожении пентоз образуются молочная и уксусная кислоты по уравнению: C5H10O5 = CH3∙CHOH∙COOH • CH3COOH + к кяп Пентоза Молочная кислота Уксусная кислота Встречаются также так называемые ложные бактерии молочнокислого брожения, они размножаются при температуре 25—35° и образуют наряду с молочной другие кислоты. Типичное и нетипичное молочнокислое брожение широко применяется в различных областях народного хозяйства. Первое используется для приготовления различных молочных продуктов (простокваши^ кефира, кумыса и др.), кислого черного хлеба, для получения молочной кислоты и консервирования фруктов и овощей. Брожение пентоз имеет значение при силосовании кормов, так как в сене, соломе и в некоторых растениях содержатся пен-тозаны, гидролизующиеся до пентоз, которые превращаются 127 в указанные выше кислоты, придающие предохраняющие его от загнивания. силосу кислый вкус и 12.2. Получение продуктов пропионовокислого брожения (витамин В12) Витамин В12, или цианкобаламин, - важное биологическое соединение, активный гематопоэтический фактор млекопитающих и ростовой фактор для многих видов микроорганизмов и животных. Начало истории витамина B12 положили исследования злокачественного малокровия (пернициозной анемии) - заболевания, поражающего в основном пожилых людей, но наблюдаемое иногда и у детей. До 1926 г. это заболевание было неизлечимым и обычно кончалось смертью. При этом заболевании в организме вырабатываются аномально большие, недоразвитые и нестойкие эритроциты, общее количество которых значительно снижалось (1x106-3xЮ6 мм~3 вместо 4,5x106-6x106 мм~3). Заболевание поражает также и другие быстрорастущие ткани, например слизистую оболочку желудка (в результате прекращается секреция НСl) и нервные ткани. При этом часто наблюдается демиелинизация центральной нервной системы с нарушением координации движений (атаксия) и психотическими расстройствами. Внутримышечное введение 3-6 мкг витамина B12 достаточно, чтобы вызвать ремиссию у пациента, больного перницитозной анемией. В 1926 г. Мино и Морфи обнаружили, что с пернициозной анемией можно справиться, если употреблять в пищу сырую или слегка поджаренную печень из расчета 1/4 кг в день. И только 22 годами позднее из печени крупного рогатого скота были выделены первые красные кабальтсодержащие кристаллы витамина B12. Позже выяснилось, что более богатым источником витамина могут служить жидкие ферментационные среды бактерий. Структура витамина B12 была впервые определена в 1956 году Дороти Ходжкин методом дифракции рентгеновских лучей. Это была самая крупная органическая молекула, структуру которой удалось определить методами рентгено-структурного анализа. Циклическая система витамина B12 подобна циклической системе порфиринов (рис. 3); она состоит из четырех пиррольных колец с атомом кобальта в центре. Нормальный уровень витамина B12 в крови составляет ~2-10~-10 M или немного выше, однако у вегетарианцев этот уровень может опускаться ниже половины этой величины. Недостаточность фолиевой кислоты также может вызывать, мегалобластическую анемию, а большой избыток фолиевои кислоты может в некоторой степени уменьшить анемию у пациентов, больных злокачественным малокровием. В тканях животных концентрация витамина очень низкая (в печени быка -1 мг/кг), чтобы использовать этот источник для промышленных целей. Активированный ил сточных вод содержит 4-10 мкг/кг витамина, но при этом требуется разделение большого числа различных форм (аналогов). 128 Химический синтез витамина В12 очень сложен. Больше 10 лет потребовалось сотрудникам двух больших лабораторий - Вудворта и Эшенмозера для осуществления химического синтеза витамина В12, включающего 70 стадий. Поэтому в настоящее время витамин В12 в промышленности получают исключительно биосинтетическим путем. Из 10 тонн ежегодно выпускаемого в мире витамина B12 3,5 т приходится на цианкобаламин (собственно витамин B12), 2 т на гидроксокобаламин, 1т - на коэнзим В12 и небольшое количество на метилкобаламин; эти формы в указанных количествах используют в медицине. Остальное количество витамина используется для животноводства. Витамин B12 применяют при лечении злокачественной анемии, цирроза печени, при нервных и психических расстройствах. Он широко используется в производстве кормов. В настоящее время большинство комбикормов для свиней и птиц обогащают витамином В12, особенно благоприятное действие на животных оказывает сочетание витамина B12 с малыми дозами антибиотиков, в частности биомицина. Витамин В12 воздействует на кроветворную функцию и на обмен белков, принимает участие в регуляции оптимального содержания в организме животного метионина, валина, треонина, лейцина, изолейцина. У птиц снижение содержания B12, в желтке яиц приводит к резкому падению выводимости потомству. При добавлении В12 в корма (10-15 мг/т) прирост поросят увеличивается на 10-15%, цыплят - до 20%, яйценоскость кур - со 180 до 208 яиц в год. При добавлении Bi2 к кормам можно заменить животный белок растительным. Отечественная микробиологическая промышленность выпускает витамин B12 двух марок: А-200 мг/кг, В - не менее 500 мг/кг препарата. Люди получают витамин В12 с пищей и не могут усваивать витамин, выделяемый бактериями кишок. Животные получают витамин В12 с кормом и утилизируют витамин, образуемый кишечной микрофлорой. Молекулярная структура. Витамин B12 - первое органометаллическое соединение, выделенное из биологической системы. Из неполимерных органических соединений имеет наиболее сложное строение, изображенное на рисунке. Молекула состоит из двух почти планарных циклических структур и линейного участка. Металл Со+3 связан с макроциклом, сильно напоминающим порфириновое ядро гема. Это тетрапиррольная структура, но имеющая ту особенность, что вместо метановых мостиков, связывающих 4 пиррольных кольца, кольца А и D непосредственно связаны. Вторая кольцевая структура - азотистое основание - 5,6-диметилбен-зимидазол (5,6 ДМБ}. 5,6 ДМБ соединен с первой кольцевой системой гетерогенной боковой цепью, состоящей из N-амино-2-пропанола (изопропанола), этерифицированного фосфатом 3-мононуклеотида, связанного с основанием 5,6 ДМБ Na-гликозидной связью. Структура витамина В12 не только очень сложная, но содержит некоторые необычные части: 1) корриновая структура ранее не была известна 129 в органической химии (до открытия витамина В12 в 1948 г. независимо Риксом и Смитом); 2) Na-гликозидная связь встречается в природе очень редко и обнаружена лишь в нескольких соединениях, содержащих рибозо-3фосфат; 3) 5,6 ДМБ тоже принадлежит к уникальным соединениям и встречается в природе только в составе кобаламинов. Рис. 3. Строение кобаламина. Атом кобальта имеет 6 координационных связей; 4 из них заняты пиррольными кольцами. Одна - N-3-5,6 ДМБ и последняя - верхним лигандом (У), природа которого может варьировать. В коммерческом витамине В12 (цианкобаламине) лиганд -CN-группа (артефакт процесса выделения). 130 In vivo чаще всего встречаются дезоксиаденозильная группа (Co-B12-I), метильная группа (метилкобаламии, СН3-B12-CoB-II) или оксогруппа (оксокобаламин). Кроме этих соединений, известных как кобаламины, есть другие корриноидные соединения с иным нуклеотид-аным основанием. Нижний лиганд (х) -5,6 ДМБ может быть заменен на аденин (псевдовитамин В12), на гуанин (фактор С), 2-метиладенин (фактор А) и др. Они могут проявлять активность для некоторых микроорганизмов, но неактивны для людей. Из всех витамин В12-подобных соединений только Со-В12-I и Со-В12-II (СНз-В12) активны на клеточном уровне и как кофакторы вовлекаются в катализ двух типов реакций. Аденозил B12 используется в реакциях, в которых имеет место перестройка углеродуглеродных связей. Метил В12 вовлекается в реакциях переноса метильных групп, например в синтезе метионина из гомоцистеина (Воробьева, 1982). Биосинтез. Путь биосинтеза витамина В12 включает три ступени образования: 1) порфиринового ядра; 2) корринового ядра; 3) кобаламинов. Путь до уропорфирина III хорошо изучен в отличие от последующих ступеней от УПГ III к кобириновой кислоте. В последние годы открыт ряд пигментированных интермедиатов метилкорриноидов. Источником СНз-групп служит S-аденозилметионин. Диметилкоррифирин идентичен сирогидрохлорину (сирогему микробной сульфитредуктазы), следовательно, этот интермедиат и возможно другие коррифирины имеют витаминнезависимую биологическую функцию. Образование кобаламина включает: амидирование семи карбоксильных групп корринового кольца, включение остатка изопропиламина (происходит из треонина), активацию кобипамида с участием GTP и с образованием GMP, включение нуклеотидного основания. Включение атома кобальта в систему происходит вскоре после образования кобириновой кислоты. У бактерии Prop. shermanii все известные корриноиды, амидированные больше, чем кобириновая кислота, находятся в 5'-аденозильной форме. Аэробные и аэротолерантные формы синтезируют 5,6 ДМБ из рибофлавина через FMN. Для синтеза требуется кислород. Анаэробы образуют 5,6 ДМБ другим путем, включающим глицин и метионин. 6-Аминолевулиновая кислота (6-АЛК) у аэробов образуется из глицина и сукцинил-СоА, а у анаэробов -из глутамата (как у растений). Биосинтез витамина В12 у бактерий происходит двумя независимыми путями, но оба приводят к образованию 5'-дезоксиаденозилкобинамидгуанозипдифосфата и а-рибазол-5'-фосфата, являющихся субстратами для конечной стадии синтеза. Из клеток мутанта Е. coli выделили 70S рибосомы, способные синтезировать витамин B12 в реакционной системе. Основным катализатором синтеза является белок 18L. Белки 5S RNA также 131 катализировали синтез витамина. Витамин В12, синтезированный при участии изолированных рибосом, был биологически активным. Гем оказывает ингибирующее действие на общие начальные пути синтеза корриноидов и порфиринов. Синтез витамина В12 регулируется на уровне монометилкоррифирина. Регуляция осуществляется с участием кобаламинов и аналогов, в которых 5,6 ДМБ замещен на аденин или метиладенин. Фактор В регулирующей активностью не обладает. На биосинтез витамина В12 пропионовыми бактериями стимулирующее действие оказывает ион NH4+; в его отсутствии клетки витамин не синтезируют. Глутамин и аспарагин могут заменить ион NH4+. Стимулятором синтеза B12 у микроорганизма Ps. de-nitrifleans выступает бетаин (триметилглицин). Продуценты. Витамин синтезируют многие бактерии. Дрожжи и мицелиальные грибы не образуют корриноиды. В организме человека и животных витамин B12 синтезируется исключительно бактериальной микрофлорой кишечника. Таблица 3. Образование витамина В12 различными штаммами Штамм Источник углерода Выход, мг/л Micromonospora Nocardia rugolaa Propionibacierium freudenreichii Propionibaderium Khermanii Propionibacterium vannielii sp. глюкоза 11,5 глюкоза-тростниковая меласса глюкоза 14 глюкоза 23-40 25 Pseudomonas denitr if глюкоза leans Streptomyces olivaceus Bacterium FM-O2T свеклосахарная меласса 25 Methanobacillus omelianskii глюкоза 35 Protaminobacter ruber лактоза 2,6 Corynebacterium метанол 8,8 Rhodopseudomon парафины 2,5 59 132 В табл. 3 перечислены микроорганизмы, которые в силу высокого уровня биосинтеза кобаламинов в разное время рассматривались как его потенциальные продуценты в промышленности. В настоящее время в промышленности используют в основном три штамма бактерий: Pseudomonas denitrificans, представителей рода Propioni-bacterium и метаногенные бактерии (смешанную культуру). Во всех случаях независимо от используемого штамма и условий культивирования в среду вводят ионы кобальта и часто 5,6 ДМБ. .Добавление таких предшественников корриноидов, как глицин, треонин, 6-АЛК и аминопропанол, может оказывать стимулирующее действие на витаминообразование. Получение витамина В12 с помощью пропионовокислых бактерий В настоящее время для получения витамина В12 используют следующие микроорганизмы Prop, freudenreichii ATCC 6207, Prop, shermanii ATCC 13673, Prop, shermanii BKM-103 и их варианты и мутанты. Наибольший интерес представляют штаммы, способные к самостоятельному синтезу 5,6 ДМБ. Поскольку синтез 5,6 ДМБ лучше происходит при доступе воздуха, осуществляют двустадийный процесс, в котором получают наиболее высокий выход продукта. На 1-й стадии культуру выращивают в анаэробных условиях до полной утилизации сахара. На 2-й стадии включают аэрацию, тем самым создавая условия для синтеза 5,6 ДМБ и превращения этиокобаламнна в дезоксикобаламин. Обе стадии осуществляют в двух разных ферментерах или в одном. Анаэробно выросшие клетки можно собрать путем центрифугирования и инкубировать густую суспензию на воздухе и, если нужно, в присутствии 5,6 ДМБ и цианида. Добавление ДМБ производят только во 2-й стадии ферментации (если бактерии не синтезируют его самостоятельно), поскольку в его присутствии образуются полные формы витамина, ингибирующие его синтез. Среда для ферментации обычно содержит глюкозу или инвертированную-мелассу (10-100 г/л), небольшие количества солей Fe, Mn и Mg, а также Со (10-100 мг/л), источники азота [(Nl-UhSCU]. В среду добавляют кукурузный экстракт (3070 г/л), содержащий молочную и пантотеновую кислоты, усиливающие рост бактерий. Пантотеновую кислоту, стимулирующую также синтез витамина, рекомендуют вносить в среду дополнительно. Бактерии-культивируют при 30?, поддерживая рН на уровне 6,5-7,0 путем введения (NH4)OH. Ферментацию производят в ферментерах на 500 л, содержащих 340 л среды, инокулированных 7 л посевного материала. В первые 80 ч культура растет под небольшим давлением N2 и слабым перемешиванием (без аэрации), в следующие 88 ч включают аэрацию (2 м3/ч) и перемешивание. Возможны некоторые вариации в культивировании. Витамин В12 сохраняется в клетках бактерий, поэтому проводят его экстракцию: 1) выделение витамина из клеток и превращение его в цианокобаламин; 133 2) выделение неочищенного продукта (80% чистоты), который можно использовать в животноводстве; 3) дальнейшую очистку до уровня 91-98% (для медицинских целей). Для экстракции витамина из клеток последние нагревают при 80?-120? в течение 10-30 мин при рН 6,1-8,5. Превращение в CN-кобаламин достигают, обрабатывая горячий раствор или клеточную суспензию цианидом или тиоционатом, часто в присутствии NaNO2 или хлорамина В. Корриноиды сорбируют на различных носителях: амберлите IRC-50, А12О3, активированном угле и элюируют водными спиртами или водно-фенольными смесями. Из водных растворов корриноиды экстрагируют фенолом или крезолом или смесью этих спиртов с бензином, бутанолом, углеродистым тетрахлоридом или хлороформом. При упаривании различных растворителей получают осадок или кристаллы витамина, которые растворяют в соответствующем растворителе до нужной концентрации. Выход витамина B12 при использовании пропиоиовокислых бактерий 25-40 мг/л. Но есть патентное сообщение (Франция) о достижении невероятно высокого выхода - 216 мг/л. Получение витамина В12 с помощью бактерии Pseudomonas denitrificans Ряд штаммов рода Pseudomonas образует в значительных количествах Bia, но чаще всего используют мутант Ps. denitriflcans, у которого в результате мутагенеза уровень витамина В12 удалось поднять с 0,6 мг/л (дикий штамм) до 60 мг/л. Бактерии культивируют с аэрацией и перемешиванием в периодических (или проточных) условиях в среде (а) следующего состава: свеклосахарная меласса - 100 г, дрожжевой экстракт - 2 г, (NHtb НРО4 - 5 г, MgS04 - 3 г, MnS04 - 200 мг, CoNO3 - 188 мг, 5,6 ДМБ 25 мг, ZnSO4 - 20 мг, Na2MoO3 - 5 мг, вода водопроводная - до 1 л, рН 7,4. Меласса богата бетаином и глутаминовой кислотой, оказывающими положительный эффект на выход витамина. Бетаин стимулирует синтез бАЛК и, возможно, также изменяет проницаемость мембраны. Культуру сохраняют в лиофилизированном состоянии, поддерживают в вышеприведенной среде. Пересев осуществляют в пробирку с плотной средой (б). Состав среды (б): свеклосахарная меласса - 60 г, пивные дрожжи 1 г, N-Z-амин - 1 г, (МН4НРО4 - 2 г, MgSO4 - 1 г, MnSO4 -- 200 мг, ZnS04 -20 мг, MoSO4 - 5 мг, агар - 25 г, вода водопроводная - до 1 л, рН 7,4. Инкубируют 4 дня при 28 С. Далее клетки переносят в 150 мл жидкой среды того же состава (но без агара), налитую в литровую колбу Эрленмейера. Инкубируют 3 дня при 28С на качалке. Содержимое колбы вносят в ферментер на 5 л, содержащий 3,3 л среды (см. выше), стерилизованный 75 мин при 120?. Инкубируют 90 ч при 29 С с перемешиванием (420 об/мин) и аэрацией (0,2 м3/ч). Чистый витамин В12 получают в результате проведения следующих последовательных операций: Получение витамина B12 с помощью метаногенных бактерий 134 В клетках метанобразующих бактерий витамин В12 присутствует от 4,1 нмоля/мг сухих клеток у Methanosarcina barkeri до 0,65 наномолей/мг сухих клеток у Metanobacterium formicum. Биосинтез кобаламинов архебактериями (изучали на М. barkei) сходен с биосинтезом корриноидов у анаэробных эубактерий. У метанотрофа Mtb. thermoautotrophicum большая часть клеточного кобамида локализована во фракции мембран и связана с мембранным белком. Предполагают, что содержащий кобамид интегральный мембранный белковый комплекс играет существенную роль в метаболизме этих бактерий при утилизации H2 + CO2, которая, видимо, сводится к переносу электронов. Корриноиды у метанобразующих бактерий участвуют также в катаболизме ацетата и метанола. Превращение метанола в метан у Mis. barkeri происходит через образование СНз-СоМ, в метилировании которого за счет метанола участвуют две метилтрансферазы, зависимые от кобамида. Корриноид, видимо, служит простетической группой фермента. Во Франции из ила сточных вод выделили мезофильные метаногенные бактерии и инкубировали их в ассоциации с другими бактериями в полупроточном режиме в среде, содержащей метанол (3-12 г/л), мелассу, кукурузный экстракт, NH4, Co, ортоксилидин и 5,6 ДМБ. Ферментацию ведут при 35 С в ферментере на 1000 м3 с ежесуточной заменой 10% бродящего субстрата свежей средой. Биомассу отделяют на сепараторах и высушивают на распылительной сушилке. Высушенный концентрат усредняют мелом до стандартной активности 1000 мкг/г препарата, который используют в таком виде как добавку к кормам. Сухой концентрат до усреднения содержит -3000 мкг/г витамина B12 что составляет 45-50% суммы корриноидов, фактор III - 10-15% и другие неполные корриноиды 40-50%. Термофильные штаммы метанотрофных бактерий родов Methanobacillus и Methanobacterium образуют 2 мг/л кобаламина при содержании в среде 8 г/л метанола. В России производство кормового препарата витамина B12 основано в основном на переработке барды (отхода ацетоно-бутилового или спиртового производства) биоценозом бактерий, осуществляющих термофильное метановое брожение сточных вод. Используют сложный консорциум анаэробных микроорганизмов, включающих углеводсбраживающие, аммонифицирующие, сульфатвосстанавливающие и метанобразующие бактерии. К, барде добавляют метанол - до 2%, СоСl2-6Н2О - 10 г/м3, мочевину - 300 г/м3 и сухие кормовые дрожжи - 230 г/м3. Дозировку обогатителей производят автоматически. Барду подают в нижнюю часть ферментатора метантенка (па 4-5 тыс. м3), в котором автоматически регулируют параметры процесса, обеспечивая контроль температуры (55-57?С), рН (7,5-8,0) и длительности брожения. Брожение ведут непрерывно, ежесуточно заменяя 20-25% бродящей жидкости на свежую барду. В качестве пеногасителя используют рыбий жир. 135 Для получения кормового препарата бражку выпаривают и сушат. Поскольку витамин В!2 неустойчив при тепловой обработке, особенно в щелочной среде, его стабилизируют. Для этого получаемую в процессе брожения жидкость перед выпариванием подкисляют до рН 5,0-5,3 и добавляют к ней сульфит натрия (0,1-0,25%). Содержание витамина В12 в исходной сброженной жидкости - 4,4 г/м3. Сгущение сброженной барды осуществляют на выпарных аппаратах (до 14-17% содержания сухих веществ), а сушку в распылительной сушилке. Концентрация витамина Bi2 в высушенном препарате - 500-600 мг/кг. Истинный витамин составляет 2025% от суммы корриноидов, фактор III - 35-40%, фактор В и другие - 40-45%. Получаемый препарат называют КМБ-12. Мезофильные и термофильные метаногенные бактерии, в том числе Metanobacterium thermoautotrophi-сит, Mb. thermoformicuin, Mb. bryantii, Metanosarcina barkeri, Ms. vacuolata, Ms. mazei, Methanococcus hatopilus, синтезируют исключительно фактор III. Истинный витамин B12 образуют неспорообразующие метилотрофы: Eubacterium limosum, близкий к нему Butyribacterium methylotrophicum и Acetobacter woodi. Путем создания искусственных биоценозов и подбора условий ферментации возможно целенаправленное регулирование процесса биосинтеза витамина В12. Новые разработки. Для удешевления производства витамина B12 и утилизации дешевого возобновляемого сырья изучалось образование корриноидов бактериями Prop, atidipropionici ATCC 25562 при росте на ксилозе как главной составной части гидролизатов гемицеллюлоз. Используя ксилозу, бактерии аккумулировали 0,35 мг корриноидов в одном литре среды без внесения солей Со. Для продукции корриноидов из ксилозы больше всего подходит UFR-реактор, работающий с ультрафильтрационной рециркуляцией клеток. Иммобилизованные клетки. В Японии сразнивали стабильность и продуктивность биокатализатора при включении клеток Propionibacterium sp. в гели каппа-каррагенана, Na-альгината, агара и форполимерные уретановые смолы. Оптимальной подложкой служит форполимер PU-9, полимерная матрица которой не снижала активности включенных в нее клеток. В оптимальных условиях ферментации 5 г иммобилизованных клеток вновь синтезировали и экскретировалн 900 мкг витамина за 18 дней повторной периодической ферментации, продемонстрировав возможность проведения многоступенчатого сложного синтеза (подобных примеров известно немного). Усовершенствование штаммов-продуцентов. В последние годы усовершенствование штаммов было достигнуто путем мутаций и селекции. Этим методом в 50 раз увеличена продуктивность по витамину у Ps. denitrificans. Для грамположительных бактерий Propionibac-terium, Bacillus, Streptotnyces применимо слияние протопластов, для грамотрицательных бактерий, например Pseudomonas, доступны конъюгативные плазмиды, как 136 Inc PI. Пока весомых практических результатов этими новыми и мощными методами не получено, но начало таким работам положено. Клонировали 11 генов, кодирующих ферменты биосинтеза витамина B12 у бактерии Вас. megaterium. Полагают, что в геноме содержится всего 20-30 таких генов. Поэтому DNA Вас. megaterium подвергли фрагментированию и крупные фрагменты встраивали в плазмиды, которыми далее трансформировали мутанты-ауксотрофы по В12. Такие мутанты приобрели способность синтезировать витамин В12. Метод может быть использован для получения штаммов-продуцентов в производственных масштабах. В бактерии Е. coll клонированы гены Prop, technicum, ответственные за синтез витамина В12. Бактерия Prop. technicum не содержит плазмид, поэтому из этого штамма выделили, очистили и частично разрушили ДНК, получив фрагменты 15-20 килобаз. Эти фрагменты включили в расщепленную плазмиду pBR 322 и полученной гибридной плазмидой трансформировали Е. соli. Новые трансформанты отличались от контрольного штамма в отношении морфологических и физиологических признаков. 12.3. Ацетонобутиловое брожение Ацетонобутиловое брожение представляет собой биохимически более сложный тип маслянокислого брожения, в котором образовавшиеся на первой стадии жирные кислоты превращаются в нейтральные конечные продукты — ацетон, бутиловый и изопропиловый спирты. Если в первой фазе брожения увеличивается кислотность, то во второй фазе кислотность не только не возрастает, но даже уменьшается. Таким образом, за счет превращения кислот в нейтральные продукты регулируется рН среды (кислотность). Ацетонобутиловое брожение углеводов вызывается Clostridium acetobutylicum и близкими к нему по физиологии видами. Cl. butylicum осуществляет брожение, очень сходное по типу с ацетонобутиловым, но отличается тем, что вместо ацетона в этом случае образуется изопропиловый спирт. Получение ацетона и бутанола. Получение ацетона и бутанола относится к числу важных бродильных производств. Впервые в промышленно масштабе они были осуществлены в Манчестере Вейсманом в ходе Первой Мировой Войны. Ацетон был необходим для производства кордита и как метательное взрывчатое вещество в тяжелой артиллерии. Ацетон низкого качества получали путем сухой перегонки древесины, но для упомянутых целей нужен был высококачественный растворитель. Бродильный процесс (ферментация) был основан на переработке крахмала, концентрация которого составляла до 3,8% (вес/объем), анаэробными спорообразующими бактериями Clostridium acetobutylicum. Превращению подвергалось до 30% субстрата, в результате получали смесь растворителей (60% бутанола, 30% ацетона, 5-10% этанола, изопропанола и мезитилоксида). Остальная часть субстрата в ходе процесса брожения 137 превращалась в водород и углекислый газ, как представлено на схеме (Рис. 4). Поскольку образовывались большие объемы газов, при крупномасштабном производстве перемешивания не требовалось, а главная сложность заключалась в гашении пены. В зависимости от штаммов отношение ацетон/спирт несколько варьировало. Многие микробы, разрушающие крахмал и способные образовывать растворители, могут также сбраживать мелассу при содержании сахара в среде до 6% (вес/объем). Фактором, определяющим количество использованного субстрата, оказалась чувствительность организмов, участвующих в процессе, к н-бутанолу (верхний предел – около 1,2% по объему) и ацетону (0,4%). Заражения бродильных емкостей аэробными бактериями обычно не происходило, и главной проблемой была инфекция бактериофагами. Впоследствии выяснилось, что участвующие в процессе микроорганизмы можно «иммунизировать» путем нескольких пересевов в присутствии бактериофага. Было установлено, что фаговая инфекция является штамм-специфичной. Рис. 4 Схема реакций ацетон-бутанольного брожения. 138 Растворители отделяли от среды отгонкой. В конце Первой Мировой Войны главную роль стало играть производство бутанола: он нашел применение при получении широкого круга веществ, включая мочевиноформальдегидные пластмассы, пластификаторы и тормозные жидкости. Побочный продукт, водород, стал использоваться в производстве синтетического метанола и для гидрогенизации пищевых масел; углекислый газ либо сжижали, либо превращали в сухой лед. Твердые вещества отходов содержали большое количество рибофлавина (витамина В2), и их можно было использовать как богатую белком добавку к кормам. После Второй Мировой Войны бродильное производство этих растворителей сильно сократилось, так как относительная стоимость нефтехимических продуктов по сравнению с полимерами сахаров уменьшилась. Производство н-бутанола путем ферментации продолжалось только в ЮАР. Однако в настоящее время получение бутанола с помощью ферментации становится все более выгодным. Главный недостаток существующих штаммов – низкая устойчивость к конечным продуктам и относительно низкий выход растворителей. В настоящее время ацетон и бутиловый спирт получают, сбраживая зерновые, мелассно-зерновые заторы или мелассу. Если среду готовят из зерна, например кукурузы, то сначала получают муку грубого помола, ее смешивают с водой из расчета 6-8 кг муки на 100 л воды. Затем затор варят 2 ч под давлением 0,2 МПа и стерилизуют. Охлажденную до 37-42 С° массу сбраживают в течение 2 сут, рН среды 5-7. В процессе брожения из глюкозы образуется смесь, содержащая 6 частей бутилового спирта, 1 часть этилового спирта и 3 части ацетона. Практически из 3 кг крахмала получается 1 кг органических растворителей. Разработаны методы получения ацетона и бутилового спирта из сульфитного щелока и гидролизатов древесины. В ацетоно-бутиловом брожении в первый период образуется уксусная и масляная кислоты, выделяется водород и углекислый газ. Затем масляная кислота восстанавливается до бутилового спирта. Ацетон образуется из продукта конденсации уксусной кислоты – ацетонуксусной кислоты при декарбоксилировании. Во время брожения в среде накапливается много рибофлавина, причем его количество тем больше, чем интенсивнее образуется ацетон. Получение рибофлавина. Разработаны способы получения кормового препарата рибофлавина из ацетоно-бутиловой барды после отделения органических растворителей. Для этого в качестве субстрата используют декантированную ацетонобутиловую барду, содержащую 2.0–2.5 % сухих веществ. Брожение проходит при 55–57°С в нестерильной культуре в две фазы: на первой образуются жирные кислоты и метан, на второй – метан, углекислота и витамин В2. Длительность процесса в одном аппарате составляет 2.5–3.5 суток, в двух последовательных – 2–2.5 суток. Концентрация витамина в бражке достигает 850 мкг/л. Параллельно в 139 значительных количествах, до 20 м3/м3 образуется газ (65 % метана и 30 % углекислоты). Бражка имеет слабощелочную реакцию. Для стабилизации витамина ее подкисляют соляной или фосфорной кислотой, затем в выпарном аппарате сгущают до 20 % содержания сухих веществ и высушивают в распылительной сушилке. Содержание В2 в сухом препарате – до 100 мкг/г. Аппараты для ацетонбутилового брожения. Поскольку ацетонбутиловое брожение это анаэробный процесс, для ферментации используются аппараты для анаэробных процессов. Аппараты для анаэробных процессов достаточно просты и применяются в процессах конверсии растительного сырья, в том числе растительных отходов, а также различных промышленных отходов. При метановом брожении для получения биогаза, а также в ряде других процессов (получение ацетона, шампанских вин) используют ферментационные аппараты (метанотенки). Эти аппараты имеют различную конструкцию (от простой выгребной ямы до сложных металлических конструкций или железобетонных сооружений) и объемы (от нескольких до сотен кубометров). Метановые установки оборудованы системой подачи сырья, системой теплообменах труб для стабилизации температуры, несложным перемешивающим устройством для гомогенного распределения сырья и биомассы продуцента, газовым колпаком и устройством переменного объема (газгольдер) для сбора образуемого биогаза. Использование ацетона. Технический ацетон получается кумольным методом в процессе синтеза фенола, методом ацетонобутилового брожения, а также в качестве побочного продукта в ряде производств. Ацетон находит применение в лакокрасочной промышленности, используется при производстве пластмасс, синтетического каучука и химических волокон, служит сырьем для синтеза многих других органических продуктов. Он используется в производстве уксусного ангидрида, ацетонциангидрина, дифенилолпропана, для обезжиривания поверхностей, как растворитель в различных отраслях промышленности. Перспективы использования низших спиртов (метанола, этанола), а также ацетона и других растворителей в качестве горючего для двигателей внутреннего сгорания вызвали в последние годы большой интерес к возможности их крупномасштабного получения в микробиологических процессах с использованием различного растительного сырья. Промышленные объекты получения продуктов ацетонбутилового брожения. При ацетонобутиловом брожении из 1 т картофеля можно получить 25 м3 водорода, 340 кг бутанола и 110 кг ацетона. Биоводород из биомассы можно получать путем бутилового или ацетонобутилового брожения сахарозы или крахмала. При этом из 1 т мелассы образуется 80 м3 водорода. Это означает, что с 1 га плантаций сахарной свеклы (мелассы) можно получить до 140 м3 водорода, или в 140 пересчете на всю произведенную в 2003 г. мелассу – 88 млн. м3. Дополнительно к водороду 1 т мелассы дает до 114 кг бутанола и до 36 кг ацетона, а весь годичный объем мелассы – 125000 т бутанола и 40000 т ацетона. В СССР до конца 70-х годов XX столетия в эксплуатации находилось 4 ацетонобутиловых завода: в городах Грозном, Нальчике, Талица (Свердловской области) и Ефремов (Тульской области). К концу 90-х годов остались Грозненский и Ефремовский заводы, причем Ефремовский завод можно восстановить. Ефремовский завод производил до 50 т растворителей (бутанол/ацетон/этанол = 13/4/1) и до 29 тыс. м3 водорода в сутки, или 15 тыс. т растворителей и до 8.7 млн. м3 водорода в год, а Грозненский завод – 74 т растворителей и 43 тыс. м3 водорода в сутки, или до 22 тыс. т растворителей и 12.9 млн. м3 водорода в год. К сожалению, весь образующийся водород в то время выпускался в атмосферу (углекислый газ шел на производство жидкой и твердой углекислоты). В конце 60-х годов XX столетия на Ефремовском ацетонобутиловом заводе (1967 г.) и Грозненском АБЗ (1969 г.) были введены в эксплуатацию два цеха по производству кормового витамина В2 методом термофильного метанового брожения жидких отходов этих производств – барды (3000 мэ/сутки). Кроме витамина В2 каждый цех производил в сутки до 30 тыс. м3 биогаза, который целиком направлялся на производство тепловой энергии для всего производственного цикла. Таким образом, в конце 60-х годов в СССР впервые в мире были созданы промышленные производства биотоплив из биомассы (биоводород, биометан, биобутанол, биоацетон и биоэтанол). В Республике Беларусь в настоящее время источником ацетона и бутанола является завод ОАО «БелХим», однако в республике есть источники мелассы и другого сырья, которое может быть использовано при микробном синтезе продуктов ацетонбутилового брожения. 12.4. Спиртовое брожение. Этиловый спирт (этанол) — бесцветней жидкость, легко испаряющаяся. Спирт, содержащий 4—5 % воды, называют ректификатом, а содержащий только доли процента воды — абсолютным спиртом. Такой спирт получают химической обработкой в присутствии водоотнимающих средств (например, свежепрокаленного СаО). Этиловый спирт — многотоннажный продукт химической промышленности. Получают его различными способами. Один из них — спиртовое брожение веществ, содержащих сахаристые вещества, в присутствии ферментов (например, зимазы — фермента дрожжей). Такой спирт называют пищевым или винным спиртом. Этиловый спирт можно получать из целлюлозы, которую предварительно гидролизуют. Образующуюся при этом глюкозу подвергают 141 в дальнейшем спиртовому брожению. Полученный спирт называют гидролизным. Как известно, для получения этилового спирта существуют и синтетические способы, такие, как сернокислотная или прямая гидратация этилена. Себестоимость спирта, полученного таким способом, намного дешевле, чем приготовленного из пищевых продуктов. Этиловый спирт широко используют в различных областях промышленности и прежде всего в химической. Из него получают синтетический каучук, уксусную кислоту, красители, эссенции, фотопленку, порох, пластмассы. Спирт является хорошим растворителем и антисептиком. Поэтому он находит применение в медицине, парфюмерии. В больших количествах этиловый спирт идет для получения спиртоводочных изделий. Этиловый спирт в больших количествах идѐт на производство синтетического каучука. Окислением спирта получают пищевую уксусную кислоту. Путѐм его дигидратации готовят диэтиловый (медицинский) эфир, с взаимодействием с хлороводородом получают хлорэтан, для местной анестозии. Спирт применяется при изготовлении многих лекарств. В парфюмерии он идѐт на изготовление духов и адеколонов. Этиловый спирт – наркотик. При приеме внутрь он вследствие высокой растворимости быстро всасывается в кровь и возбуждающе действует на организм. Под влиянием спиртного у человека ослабевает внимание, затормаживается реакция, нарушается координация, появляется развязность, грубость в поведении и т. д. Все это делает его неприятным и неприемлемым для общества. Но следствия употребления алкоголя могут быть и более глубокими. При частом потреблении появляется привыкание, пагубное пристрастие к нему и в конце концов тяжелое заболевание – алкоголизм. Спиртом поражаются слизистые оболочки желудочнокишечного тракта, что может вести к возникновению гастрита, язвенной болезни желудка, двенадцатиперстной кишки. Печень, где должно происходить разрушение спирта, не справляясь с нагрузкой, начинает перерождаться, в результате возникает цирроз. Проникая в головной мозг, спирт отравляюще действует на нервные клетки, что проявляется в нарушении сознания, речи, умственных способностей, в появлении психических расстройств и ведет к деградации личности. Особенно опасен алкоголь для молодых людей, так как в растущем организме интенсивно протекают процессы обмена веществ, а они особенно чувствительны к токсическому воздействию. Поэтому у молодежи быстрее, чем у взрослых, может появиться алкоголизм. Спирт опасен в любой концентрации (водка, настойки, вино, пиво и т.д.). Этиловый спирт, применяемый для технических целей, специально загрязняют дурно пахнущими веществами. Такой спирт называют денатуратом (для этого спирт подкрашивают, чтобы отличить его от чистого спирта). 142 Особенности технологии этилового спирта. До начала 30-х годов 20 века этиловый спирт получали исключительно сбраживанием углеводсодержащего сырья, и при обработки зерна (рожь, ячмень, кукуруза, овѐс, просо). В 30-е по 50-е годы было разработанно несколько способов синтеза этилового спирта из химического сырья. В США в 1976 г. было выработано около 800 тыс. тонн этонола, в т.ч. 550 тыс. тонн прямой гидротацией (остальное сбраживание пищевого сырья). В других странах (СССР, Франция и др.) этиловый спирт получают также двухстадийной гидраитацией этилена при 75-80 С. В ряде стран этиловый спирт получают также сбраживанием продуктов гидролиза растительных материалов. Очистку технических спиртов проводят различными способами. Пищевой спирт-сырец, обычно освобождают от примесей (сивушные масла и др.) рекитификацией. Слинтентичиский этиловый спирт очищают от этилового эфира, ацетальдегида и др. рекитификаций в присутствии щѐлочи и гидрированием в паровой фазе на никелевых католизаторах при 105 С. Спирт –рекитификат представляет собой асеотропную смесь 0 этилового спирта с водой (95,57% спирта t кипения 78,15 С.). Для многих целей требуется обезвоженый, абсолютный этиловый спирт. Для получения этилового спирта издавна пользуются различными сахаристыми веществами, например, виноградным сахаром, или глюкозой, которая путем "брожения", вызываемого действием ферментов (энзимов), вырабатываемых дрожжевыми грибками, превращается в этиловый спирт. Глюкоза в свободном виде содержится, например, в виноградном соке, при брожении которого получается виноградное вино с содержанием спирта от 8 до 16%. Исходным продуктом для получения спирта может служить полисахарид крахмал, содержащийся, например, в клубнях картофеля, зернах ржи, пшеницы, кукурузы. Для превращения в сахаристые вещества (глюкозу) крахмал предварительно подвергают гидролизу. Для этого муку или измельченный картофель заваривают горячей водой и по охлаждении добавляют солод – проросшие, а затем подсушенные и растертые с водой зерна ячменя. В солоде содержится диастаз (сложная смесь ферментов), действующий на процесс осахаривания крахмала каталитически. По оканчании осахаривания к полученной жидкости прибавляют дрожжи, под действием фермента которых образуется спирт. Его отгоняют, а затем очищают повторной перегонкой. В настоящее время осахариванию подвергают также другой полисахарид – целлюлозу (клетчатку), образующую главную массу древесины. Для этого целлюлозу подвергают гидролизу в присутствии кислот (например, древесные опилки при 150 -170(С обрабатывают 0,1 - 5% серной кислотой под давлением 0,7 - 1,5 МПа). Полученный таким образом продукт также содержит глюкозу и сбраживается на спирт при помощи 143 дрожжей. Из 5500 т сухих опилок (отходы лесопильного завода средней производительности за год) можно получить 790 т спирта (считая на 100%ный). Это дает возможность сэкономить около 3000 т зерна или 10000 т картофеля. В зависимости от исходного сырья и степени очистки спиртректификат делят на люкс, экстра, высшей очистки и 1 сорта. Питьевой спирт 95% - это смесь спирта-ректификата высшей очистки, умягченной воды, профильтрованная и выдержанная. Спирт-ректификат служит сырьѐм для производства водок, вин, ликеро- водочных изделий. Водка - крепкий алкогольный напиток, содержащий 40-56% спирта. Получают из спирта-ректификата, умягченной воды, фильтруют через активированный уголь. Добавляют: сахар-песок, питьевую соду, уксус, лимонную кислоту, мѐд, соль. Для производства этилового спирта из крахмалистого сырья на спиртзаводе используется зерно. Основными зерновыми культурами, поступающими в производство, являются рожь, пшеница, ячмень и кукуруза. Основными из показателей является условная крахмалистость, по которой ведѐтся учѐт выхода спирта. При поступлении зерна определяют показатель зерновой примеси, по наличию зерна другого вида и устанавливают, что идѐт на переработку, чистая культура или зерносмесь. Зерно на завод поступает железнодорожным транспортом в хопперах и автотранспортом. При поступлении зерна лаборатория завода производит отбор проб для определения качества зерна в соответствии с требованиями ГОСТ 13586-83 «Зерно. Правила приѐмки и методы отбора проб». При поступлении зерна железнодорожным транспортом выдаѐтся документ о качестве зерна. Для проверки соответствия качества зерна требованиям нормативной документации анализируют среднюю пробу от поступившей партии. При поставке зерна автотранспортом от колхозов допускается отметка о качестве в сопроводительном документе. Поступающее зерно взвешивается на автомобильных весах и отправляется на хранение в склад. С момента поступления зерна на склады завода, в течение всего периода его хранения, за качеством и состоянием каждой партии ведѐтся наблюдение. К числу показателей, по которым оценивают состояние зерна при хранении, относят: температуру, влажность, а в партиях солодового зерна дополнительно проверяют энергию и способность его прорастания. На производство зерно подаѐтся автотранспортом, взвешивается, очищается и поступает в зерновые силоса (V = 48 м3). Для получения спирта-ректификата в качестве сырья также используется спирт сырец, который поступает на завод посредством спиртовозов и хранится в спиртохранилище открытого типа. Спирт этиловый ректификованный вырабатывают на заводе путем сбраживания крахмало- и сахаросодержащего сырья специальными расами 144 дрожжей, перегонки полученной спиртовой бражки и ректификации на брагоректификационных установках. Спирт этиловый ректификованный вырабатывают также путем ректификации спирта-сырца на непрерывно-действующей ректификационной установке. Спирт этиловый ректификованный из пищевых видов сырья вырабатывают в виде спиртоводной смеси, концентрация спирта в которой, в зависимости от сорта, составляет 96,2...96,6% объемных. В производстве в соответствии с ГОСТ 5962-67 вырабатывают спирт этиловый ректификованный "Люкс", "Экстра", "Высшей очистки". Спирт "Супер-Люкс" является нововведением РУП "Кристалл" и вырабатывается по ТУ РБ 0179 7526.542-97. В спирте этиловом ректификованном кроме воды в микро количествах содержатся различные примеси (альдегиды, эфиры, высшие спирты и другие химические соединения), которые формируют у спирта свойственные ему вкус и аромат в зависимости от вида перерабатываемого сырья. По органалиптическим показателям спирт этиловый ректификованный должен соответствовать следующим показателям (ГОСТ 5962-67): - внешний вид - прозрачная жидкость без посторонних частиц; - цвет - бесцветная жидкость; - вкус и запах - характерные для каждого вида этилового спирта, выработанного из соответствующего сырья, без привкуса и запаха посторонних веществ. Качество воды, используемой для технологических целей, оказывает большое влияние на технологические процессы и качество готовой продукции. Наиболее важными показателями качества воды являются еѐ жѐсткость, окисляемость и бактериальная чистота. Вода не содержит различимых невооружѐнным взглядом водных организмов, а также посторонних вредных веществ, угнетающих дрожжи и ухудшающих качество готовой продукции. В зависимости от места применения в производстве к воде предъявляются различные требования. Для приготовления замеса используется техническая вода с температурой не более +50 С, рН не менее 5,0, жѐсткостью не выше 12 мг/ экв/л. Для нормального ведения процесса вода, поступающая на охлаждение в теплообменные аппараты и на вакуум-охлаждение имеет температуру не более +12 С. Для питания котлов используется оборотная вода из дефлегматоров с температурой 60 – 65 С. Чтобы предотвратить образование накипи, воду предварительно умягчают, с целью удаления ионов кальция и магния. Эта же вода используется для рассиропки спирта-сырца при его ректификации. Контроль ведѐт заводская лаборатория. Формалин (НСОН) представляет собой водно-метанальный раствор формальдегида и предназначается для использования в качестве антисептика. 145 Кислота серная (H2SO4) применяется для подкисления дрожжевого сусла. Содержание моногадрата не менее 92,5 %. Карбамид (H2N-CO-NH2, мочевина) используется на заводе в качестве азотосодержащего питания при выращивании дрожжей. Содержание азота , в пересчѐте на сухое вещество составляет 46,3 %. Хлорная известь (CaCl2) применяется для обеззараживания солодового зерна, а также для дезинфекции бродильных чанов, трубопроводов, различных ѐмкостей, полов и других загрязнѐнных мест. Хлорная известь представляет собой белый порошок с содержанием активного хлора 32 – 35 %. Побочные продукты при брагоректификации бражки (ЭАФ, сивушное масло). Фракция головная этилового спирта ТУ РБ 00966671-494-95 (изменение введено 10.09.99 г.) представляет собой смесь, состоящую из метилового спирта, уксусного альдегида, кислот, сложных эфиров карбоновых кислот и других примесей. Применяется для получения технического и денатурированного спиртов, а также для изготовления средств и препаратов различного назначения. Продукт производства, пройдя контрольный снаряд, отправляется на хранение в спиртохранилище, в специально оборудованные металлические резервуары. Сивушное масло ГОСТ 17071-91 представляет собой смесь амиловых спиртов, изобутилового, н-пропилового и этилового спирта, воды и незначительных количеств других органических соединений. Сивушное масло служит сырьѐм для получения технических спиртов применяемых в пищевой, парфюмерной, фармацевтической, лакокрасочной промышленности. В процессе перегонки бражки на спиртзаводе образуются промежуточные продукты: бражной дистиллят, эпюрат, не пастеризованный спирт, сивушный спирт, промывная вода с ЭРК, барда и лютерная вода. С бардой и лютерной водой выводятся нелетучая часть бражки. Летучие примеси выводятся с головной фракцией этилового спирта, с сивушным маслом и сивушным спиртом. Отделение подработки зерна предназначено для очистки зерна от минеральных, металических и органических примесей. Зерно, поступающее со склада, выгружается в приѐмный бункер, оборудованный защитной решѐткой, предназначенной для отделения крупных предметов от зерна. По спиральному шнеку зерно подаѐтся на норию и поступает в бункер-накопитель неочищеного зерна. Из бункеранакопителя зеоно поступает на воздушно ситовой сепаратор, где при помощи сита отделяются механические и органические примеси. Отходы поступают в специальный бункер. 146 С сепаратора зерно поступает в бункер очищеного зерна, а затем на весы-дозатор, которые предназначены для определения количества очищеного зерна, подаваемого на производство. С весов-дозатора зерно, порциями до 50 кг, по нории поступает в силоса. В процессе подработки зерна выделяется пыль, которая может образовать взрывоопасную смесь. Поэтому на всех шнеках, бункерах, нориях и воздушно-ситовом сепараторе установлена аспирация, состоящая из вентиляторов и циклонов, куда поступает пыль. Из силосов зерно поступает норией и подаѐтся в накопительный бункер производства. На заводе действует Мичуринская схема неприрывного разваривания крахмалистого сырья. Разваривание крахмалистого сырья в неприрывнодействующих агрегатах предусматривается в измельчѐнном виде. Очищеное зерно из накопительного бункера самотѐком идѐт на молотковые дробилки и измельчается в них. В измельчѐнном зерне допускается остаток крупки, на сите с диаметром 1 мм, 7 – 8%. Измельчѐнное зерно самотѐком непрерывно поступает на смесители. Туда же через ротаметры подаѐтся требующееся количество воды, с температурой 40 – 45 ºС. Для приготовления замеса на 1 кг размолотого зерна берѐтся 2,5 – 3,5 л воды, в зависимости от крахмальности исходного сырья. Концентрация сусла при правильной дозировке должна быть 16 – 18% по сахарометру. Зерновой замес плунжерным насосом через подогреватель типа ―труба в трубе‖, где он нагревается вторичным паром из выдерживателя, до темпетатуры 55 – 65 ºС, подаѐтся на контактную головку 1-й ступени, где нагревается до температуры 65 – 75 ºС. В контактной головке 2-й ступени зерновой замес нагревается до максимально-возможной температуры – 138 – 140 ºС. При прогреве до более высокой температуры сокращается количество подаваемого пара в варочную колонну 1-й ступени. Варочный аппарат состоит из одной колонны 1-й ступени и трѐх колонн 2-й ступени. Равномерность разваривания замеса достигается многосекционностью аппарата и специальной конструкцией колонны 1-й ступени. Время разваривания должно составлять 40 – 50 мин при температуре 138 – 140 ºС. Для кукурузы рекомендуется режим варки при температуре 144 – 150 ºС в течении 60 мин. В варочные колонны 2-й ступени пар не подаѐтся. Работа варочного аппарата проходит при одинаковом давлении в варочных колоннах 1-й и 2-й ступени и корпусе регулятора уровня, что обеспечивается соединением их уравнительной линией. Переток массы осуществляется только за счѐт разницы уровней в колоннах. Разваренная масса из последней колонны 2-й ступени, через регулятор уровня, выдувается в выдерживатель. В регуляторе уровня выдувной клапан расположен ниже рабочего уровня массы в сосуде не менее чем на 500 – 600 147 мм. Неконденсирующиеся газы и пары периодически, через 15 мин, в течении 15 сек, отводятся в паропровод через циркуляционный клапан (вентиль), расположенный в верхней части колонн далее в атмосферу. В выдерживателе поддерживается постоянное избыточное давление около 0,02 МПа, что соответствует температуре замеса 105 ºС. Разваренная масса находится в выдерживателе в течении 15 – 20 мин. Качество разваривания определяют визуально, не менее 4-х раз в смену. Отбор проб производят из пробника, установленного на выдувной трубе. Расчѐтное количество Амилосубтилина Гх взвешивают или отмеряют в литрах и вносят в расходную ѐмкость, содержащую воду с температурой 20 – 30 ºС. За 10 – 15 минут до подачи раствора в осахариватель, добавляют расчѐтное количество Глюкаваморина Гх из мерника, при постоянном перемешивании. Осахаривание – это гидролиз крахмала разваренной массы до сбраживаемых сахаров, под действием амилолитических ферментов, которые содержат осахаривающие материалы (ферментные препараты). Одновременно под действием протеолитических ферментов протекает гидролиз белков. При неприрывном осахаривании с вакуум-охлаждением для охлаждения массы под вакуумом используется специальная установка. Разваренную массу до поступления еѐ в осахариватель пропускают через испаритель, в котором температура массы снижается до 62 – 63 ºС, так как разряжение в нѐм составляет 0,08 – 0,081 МПа. Разряжение в испарителе поддерживается за счѐт резкого снижения температуры и конденсации выделившихся из разваренной массы паров. Вакуум в нѐм создаѐтся при помощи барометрических конденсаторов и суховоздушного вакуум-насоса. Сконденсировавшиеся пары вместе с холодной водой, подаваемой на барометрические конденсаторы, сливаются в сборник, откуда направляются в канализацию. Охлаждѐнная масса из испарителя поступает в осахариватель, где смешивается с ферментными препаратами, быстро охлаждается о температуры 57–58 ºС. Если еѐ температура не достигла требуемой, то производится дополнительное охлаждение с помощью водяной рубашки, которой снабжѐн осахариватель. Продолжительность осахаривания не менее 10 минут. Солодовое молочко дозируется по объему, его расход составляет 16-18% от объема разваренной массы. Раствор ферментного препарата дозируется в зависимости от их активности и крахмалистости осахаренной массы. Осахаренная масса центробежным насосом подаѐтся через теплообменники, где охлаждается до температуры складки 18 – 20 ºС, в бродильный чан. Основной целью спиртового брожения является сбраживание сахаров сусла дрожжами в спирт. Периодический способ брожения предусматривает проведение процесса от начала до конца в одном аппарате. При заполнении стерилизованных и охлаждѐнных бродильных чанов одновременно с суслом 148 вводится необходимое количество дрожжей. Расход дрожжей должен составлять 6 – 8 % от объѐма сбраживаемого сусла. Заполнение бродильного чана должно продолжаться не более 8-и часов. Более длительное заполнение приводит к ухудшению отбродов. Продолжительность брожения, считая от начала залива чана до начала перегонки зрелой бражки, составляет 72 часа. Температура складки при трѐхступенчатом периодическом брожениии должна быть 18 – 20 ºС. Во время главного брожения поддерживается температура 29 – 30 ºС, во время дображивания – 27 – 29 ºС. Снижение температуры при дображивании исключает голодание дрожжей и способствует предотвращению нарастания кислотности бражки. Температуру регулируют подачей холодной воды на водяную рубашку бродильных чанов и измеряют ртутным термометром в защитной гильзе, при этом бражку нельзя переохлаждать, так как это замедлит еѐ дображивание. Во время брожения бродильные чаны соединяют со спиртоловушкой для конденсации спиртовых паров, уносимых выделяющимся при брожении диоксидом углерода. Промывные воды из спиртоловушки отводят в сборник, откуда отправляют в приямок в брагоректификационное отделения. Концентрация спирта в промывной воде не должна превышать 7% об. Содержание спирта в промывной воде определяется стеклянным спиртометром. По окончанию брожения зрелую бражку из бродильных чанов направляют в брагоректификационное отделение. Бродильные чаны после освобождения от бражки моют с антисептиком и пропаривают паром до температуры 105 ºС. Во время сгонки бродильные чаны полностью освобождают при помощи плунжерных насосов. Нормативными показателями зрелой бражки являются: кислотность; количество несброженных углеводов; крепость. Нарастание кислотности в зрелой бражке при нормальных условиях производства не должно превышать 0,15 – 0,20 град. кислотности Содержание несброженных углеводов при отличной работе – до 0,25 г/100 мл.; при хорошей работе – от 0,25 до 0,35 г/100 мл.; - при удовлетворительной работе – до 0,45 г/100 мл. Крепость бражки при нормальных условиях производства должна быть 7,5 – 9,0 % об. Для оперативного контроля процесса брожения при переработке крахмалистого сырья определяют содержание сухих веществ в фильтрате бражки по сахарометру. Видимая концентрация зрелой бражки, в зависимости от вида сырья должна иметь следующие значения (в %): кукуруза -0,4; просо -0,2; пшеница 0,0; рожь +0,7; ячмень +0,7; овѐс +0,9. Целью процесса приготовления дрожжей является получение зрелых дрожжей, выращиваемых на специально приготовленном дрожжевом сусле, используемых для сбраживания сусла в бродильных чанах. Задачей процесса является такое ведение технологического процесса, которое обеспечивало бы максимальное накопление дрожжевых клеток при отсутствии развития посторонних микроорганизмов. 149 Для приготовления дрожжей приготавливают сусло из различных зерновых культур. При переработке толстокожурного зерна (ячменя, овса, проса) оно обязательно должны подрабатываться или перерабатываться в смеси с бесплѐночным зерном, чтобы покровное брожение было полностью исключено. В спиртовом производстве процесс размножения дрожжей происходит при повышенной кислотности среды (рН 3,8 – 4,2), что угнетает развитие молочнокислых бактерий и создаѐт нормальные условия для развития дрожжевых клеток. Для подкисления дрожжевого сусла используется серная кислота. При переработке зернового сырья нормального качества, за исключением кукурузы, дрожжевое сусло готовиться без дополнительного питания. При переработке дефектного сырья, а также кукурузы, в дрожжевое сусло вносится дополнительное солодовое питание. Технологической инструкцией предусмотрены следующие нормы дополнительного солодового питания: - 0,8 кг/дал дрожжевого сусла - при переработке кукурузы; - 0,6 кг/дал дрожжевого сусла - при переработке остальных видов сырья. Технологический процесс выращивания дрожжей состоит из двух операций: приготовление дрожжевого сусла и сбраживание этого сусла с целью накопления в нѐм дрожжевых клеток, используемых в качестве засевных дрожжей для сбраживания сусла в бродильных чанах. Приготовление дрожжевого сусла. Дрожжанка приготовления дрожжевого сусла перед началом процесса должна быть тщательно промыта, продезинфицирована раствором хлорной извести в течении 25 – 30 минут, затем вновь промыта тѐплой водой и пропарена таким образом, чтобы выдержка еѐ при температуре 100 ºС была не менее 20 минут. В эту дрожжанку отбирают сусло из осахаривателя после задачи в него всего осахаривающего материала. Концентрация сусла должна быть в пределах 17 – 18 %. Данный отъѐм сусла выдерживают в течении 1-го часа, для полного осахаривания крахмала , а затем пастеризуют при температуре 80ºС, охлаждают до температуры 50 ºС и подкисляют серной кислотой из мерника, при тщательном перемешивании, до 0,7 – 0,8 град. кислотности, что соответствует рН 3,8 – 4,0. Серную кислоту для подкисления дрожжевого сусла используют в разбавленом виде с соблюдением всех правил по технике безопасности. После охлаждения дрожжевого сусла до температуры 30ºС в него центробежным насосом вносятся засевные дрожжи, в качестве которых используются зрелые дрожжи. Количество засевных дрожжей - 8 – 12 % от объѐма сусла. После чего сусло охлаждают до температуры складки – 18 – 20ºС. Сбраживание дрожжевого сусла. В процессе производства в качестве засевной культуры для каждой последующей дрожжанки используются зрелые дрожжи предыдущей. Процесс сбраживания дрожжевого сусла осуществляется в дрожжанках при 150 строгом контроле за температурой и чистотой брожения. Повышение температуры брожения выше 30ºС не допускается, так как это свидетельствует о развитии посторонней микрофлоры. Дрожжи считаются зрелыми по достижении видимого отброда, равного 1/3 первоначальной концентрации дрожжевого сусла. Требования к зрелым дрожжам: - дрожжевые клетки должны быть хорошо упитаными; - наличие мѐртвых клеток не должно превышать 3,0 %; не допускается наличие посторонних микроорганизмов в поле зрения микроскопа. Технологическая схема браго-ректификационной установки с элементами под малым вакуумом типа «БРУВАК-М» состоит из четырѐх колонн: бражной, эпюрационной, ректификационной и экстрактивноректификационной. Бражку из бродильного отделения насосом подают в подогреватель бражки ректификационной колонны, в котором еѐ нагревают до температуры 53 – 55 ºС. Затем бражку подают в подогреватель бражной колонны, где еѐ догревают до температуры 85 – 90 ºС паром бражного дистиллята, поступающим в верх бражной колонны. Подогретую бражку, из подогревателя бражной колонны, направляют в сепаратор бражки, в котором бражка освобождается от диоксида углерода. Из сепаратора бражки она направляется на тарелку питания бражной колонны, а диоксид углерода в смеси с другими газами и водно-спиртовыми парами попадает в конденсатор сепаратора бражки и далее на спиртоловушку бражной колонны, где они конденсируются и конденсат подаѐтся на 36-ю, питательную тарелку эпюрационной колонны. Неконденсирующиеся газы из спиртоловушки бражной колонны, через огнепреградитель и клапан дыхательный, отводятся в атмосферу. В бражной колонне из бражки выделяют спирт и другие летучие компоненты бражки. Барду отводят из куба колонны через бардорегулятор в сборник барды. Водно-спиртовой пар из верхней части бражной колонны направляют в межтрубное пространство подогревателя бражки бражной колонны. Бражной дистиллят из него и его водяной части поступает на 28-ю, питательную тарелку эпюрационной колонны. Часть бражного дистиллята из водяной части подогревателя бражки бражной колонны через ротаметр возращается на верхнюю тарелку бражной колонны для еѐ орошения. Бражной дистиллят из конденсатора бражной колонны и еѐ спиртоловушки поступает на 36-ю, питательную тарелку эпюрационной колонны. Бражной дистиллят из кипятильника эпюрационной колонны собирают в сборник, из которого под воздействием работы инжектора, подают на 23-ю питательную тарелку эпюрационной колонны. При этом уровень в сборнике поддерживают постоянным. Водно-спиртовой пар из эпюрационной колонны направляют для конденсации последовательно в еѐ дефлегматор, конденсатор и спиртоловушку. Флегму из указанных теплообменников возвращают на 151 верхнюю тарелку эпюрационной колонны. Из конденсатора эпюрационной колонны отбирают головную фракцию этилового спирта и через холодильник ЭАФ, фильтр, ротаметр, эпрюветку и спиртоизмеряющий аппарат. Отводят в спиртоприѐмное отделение. Эпюрат из куба эпюрационной колонны направляют на тарелку питания ректификационной колонны. Спиртовые пары из ректификационной колонны последовательно конденсируют в подогревателе бражки ректификационной колонны, еѐ дефлегматорах, конденсаторе и спиртоловушке. Из конденсатора ректификационной колонны отбирают непастеризованный спирт, расход которого регулируют ротаметром и направляют на верхнюю тарелку эпюрационной колонны. Избыток непастеризованного спирта возвращают на верхнюю тарелку ректификационной колонны. Ректификованный спирт отбирают с 71, 72, 73, 75, 77 и 79-й тарелок ректификационной колонны и через холодильник спирта, эпрюветку, и спиртоизмеряющие аппараты направляют в спиртопиѐмное отделение. Лютерную воду из ректификационной колонны отводят через гидрозатвор с паросепаратором в сборник лютерной воды, из которого насосом подают в напорный бак лютерной воды. Из данного сборника лютерная вода через ротаметр подаѐтся на верхнюю тарелку экстрактивноректификационной колонны и на промывку сивушного масла в насадочной колонке деконтатора. Из зоны 5, 7, 9, 11-ой нижних тарелок ректификационной колонны отбираются пары сивушного масла, которые инжектором поддают в куб экстрактивно-ректификационной колонны. Пары из неѐ направляют в дефлегматор и конденсатор экстрактивно-ректификационной колонны. Гетерогенный дистиллят из дефлегматора направляют в разделитель, нижний слой из которого возвращают на верхнюю тарелку экстрактивноректификационной колонны, а верхний слой, обагащѐнный высшими спиртами, через ротаматр, подают в насадочную колонку деконтатора, где его смешивают с лютерной водой. В деконтаторе сивушная фракция поднимается вверхнюю его часть, откуда отстоявшееся сивушное масло собирается в сборник сивушного масла. Кубовой остаток из куба экстрактивно-ректификационной колонны и подсивушный слой из деконтатора направляют на 12-ю тарелку бражной колонны или на 7,9-ю тарелки ректификационной колонны. Из зоны 19, 21, 23и 24-ой тарелок ректификационной колонны сивушные спирты с помощью инжектора подают на 36-ю тарелку эпюрационной колонны, а также периодически в количестве 0,5% сивушные спирты, через пробный холодильник, отводят на спиртоизмеряющий снаряд ЭАФ. Вакуум в эпюрационной и верху ректификационной колонн создаѐтся вакуум-насосом через барометрический конденсатор, для чего спиртоловушки эпюрационной и ректификационной колонн соеденены по 152 газовой линии с нижней частью барометрического конденсатора и далее с вакуум-насосом. Для улавливания спирта и других летучих веществ в барометрический конденсатор подают холодную воду, а водно-спиртовую жидкость из него направляют на 12-ую тарелку бражной колонны. Вакуум в верху колонн регулируют подачей охлаждающей воды из коллектора холодной воды на поверхности теплообмена теплообменного оборудования колонн. Горячую воду после теплообменного оборудования колонн отводят в сборник горячей воды. Конденсат пара из кипятильника ректификационной колонны возвращается в котельную. Для контроля отсутствия потерь спирта в барде и лютерной воде, установка оснащенна пробными холодильниками. Для предохранения колонн от деформации в следствии избыточного давления или глубокого вакуума, верх и низ бражной, низы ректификационной и экстрактивноректификационной колонн снабжены вакуум-прерывателями. При переработке спирта-сырца он из спиртохранилищща подаѐтся в сборник спирта-сырца, из которых насосом его подают в напорный сборник спирта-сырца. От туда спирт-сырец, через ротаметры и смеситель, где он смешивается с умягчѐнной водой, поступающей из напорного бака умягчѐнной воды через ротаметры, поступает в подогреватель спирта-сырца, где он подогревается до температуры 73 – 75 ˚С. Подогретая смесь спиртасырца с умягчѐнной водой подаѐтся на 23-ую и 28-ую тарелку эпюрационной колонны. Ректификованный спирт, полученный на браго-ректификационной установке и учтѐнный спиртоизмеряющими аппаратами отводится в накопительные ѐмкости. Схемой спиртоприѐмного отделения предусмотрена возможность отводить ректификованный спирт в отдельную ѐмкость для индивидуального учѐта и возможности контроля слива в любое время. Из накопительных ѐмкостей в установленном порядке отбирают пробы для химического и контроля качества ректификованного спирта. Все накопительные ѐмкости в верхней их части соединены чересными переливными устройствами, для предотвращения переполнения ѐмкостей ректификованным спиртом и его потерь от проливов. Для передачи на хранение ректификованный спирт из накопительных ѐмкостей самотѐком поступает в мерники, где производится измерение его объѐма и температуры, и насосом передаѐтся в спиртохранилище на хранение. ГЛАВА 13. Выделение и очистка гормональных препаратов Гормоны (от греч. hormao –приводить в движение, возбуждать) – биологически активные вещества разной химической природы, образующиеся специализированными клетками желез внутренней секреции, которые выделяются непосредственно в кровь, лимфу и регулируют обмен 153 веществ физиологические функции организма. Сейчас известно около 60 биологически активных секретов, которые продуцируются эндокринными железами и имеют гормональную активность. Гормональные препараты чаще классифицируются по химической структуре: 1 Гормоны белковой природы: простые инсулин, пролактин, гормон роста) и сложные (фолатропин, лютропин, тиротропин) белки. 2 Гормоны пептидной природы: глюкоген, кальцитонин, соматостатин, вазопрессин, окситоцин. 3 Гормоны – производные аминоспиртов: адреналин, норадреналин, окситоцин. 4 Гормоны липойдной природы (стеройдные гормоны): котикостеройды, андрогены, эстрогены, простагландины. 5 Парагормоны, тканевые гормоны: гастрин, секретин, гепарин. Получают гормоны эндокринных желез путем химического синтеза, методами генной инженерии и выделением из сырья животного происхождения. Кроме приведенных выше существует методика биотрансформации. Так один из штаммов хлебной плесени Rhizopus arrhizus используется на начальном этапе синтеза производного стероида, кортизона, путем гидроксилирования прогестерона по 11-му положению. Синтезированы инсулин и некоторые белковые гормоны гипофиза (кортикотропин, соматотропин) и т.д. Препараты стероидных гормонов, производных аминокислот и пепидов (окситоцин, вазопрессин) получают путем химического синтеза. Но получение полипептидных гормонов из-за многостадийности и сложности не рентабельно [2]. Далее будут приведены примеры получения некоторых гормонов из различных источников. 13.1. Инсулин. Выделение инсулина из животного сырья Инсулин – гормон поджелудочной железы, вырабатываемый клетками островков Лангерганса, играет важную роль в углеводном обмене. Молекула инсулина человека состоит из двух полипептидных цепей – А и В, соединенных двумя дисульфидными связями. А цепь состоит из 21, а В-цепь – из 30 аминокислотных остатков. В 1952 году был впервые выделен инсулин в кристаллическом виде. В настоящее время существует несколько методов выделения инсулина из поджелудочных желез рогатого скота и свиней. Получение инсулина состоит из ряда страдий: 1) Измельчение замороженных поджелудочных желез и экстракция кислым спиртовым раствором. 2) Осаждение балластных белков и освобождение от липидов. 3) Изоэлектрическое осаждение фракций инсулина (при pH=5,5) и осаждение спиртом, ацетоном, эфиром. 154 4) Очистка инсулина: осаждение солями, фракционирование методами хроматографии, гель-фильтрации. 5) Осаждение инсулина в виде кристаллов. 6) Переосаждение цинк-инсулина. Свежие или замороженные поджелудочные железы измельчают в мясорубке-волчанке и экстрагируют способом бисмацерации первый раз 8085 % этанолом в реакторе с мешалкой. Второй раз экстрагируют 57% этанолом, который подкислен ортофосфорной кислотой (хлороводородной или серной) до значения pH 2,8-3. Экстракцию проводят 1,5-4 часа при постоянном перемешивании. Подкисленный спирт способствует инактивации фермента трипсина, находящегося в поджелудочной железе, благодаря чему удается сохранить инсулин в неизменном состоянии. Но минском заводе эндокринных препаратов используют роторнопульсационный аппарат для экстракции, что дает интенсивность экстрагирования и сокращает время до 1.5ч. Полученные вытяжки объединяют, отставляют на холоде на 48 ч для освобождения от нежелательных белков, которые выпадают в осадок. Осадок отделяют центрифугированием и удаляют. Далее для выделения и очистки инсулина применяют ионообменную хроматографию. Осуществляют сорбцию инсулина из прозрачной жидкости на макропористом сульфокатионите КУ-33-30/100 при значении pH 3,0-3,3 в режиме псевдоожижения. Жир удаляют путем промывки катионита 65-67% этанолом, а балластные белки 0,3М раствором ацетатного буфера. Десорбцию инсулина осуществляют с помощью 0,01-0,05 раствора аммонийного буфера (pH 10) и немедленно подкисляют хлороводородной кислотой до pH 4,5 и добавляют ацетон. Выпавший осадок балластных веществ удаляют. Инсулин осаждают раствором цинка-ацитата (pH 6,2). Полученный цинк-инсулин очищают методом кристаллизации, после чего растворяют в воде, подкисленной до pH 2,8 лимонной кислотой. Раствор отстаивают 1ч. Выпавшие балластные вещества удаляют фильтрацией через кизельгур. Фильтрат смешивают с ацетоном, добавляют хлористый цинк и охлажденный до 0ºС фенол. Для медленной кристаллизации инсулина создают условия с последовательным изменением pH раствора. Раствор подщелачивают до pH 8,5,; оставляют на 2-3 минуты, изменяют pH до 6,8 и перемешивают 1 ч; при значении pH 6,5 перемешивают 2 ч; при pH 6,2 и 6,0 перемешивают 2 ч и отстаивают 20ч; при значении pH 5,8 перемешивают 2ч. И отстаивают 96 ч при температуре 5 °С. Выпавшие кристаллы инсулина отделяют центрифугированием, промывают на воронке Бюхнера последовательно холодной водой, ацетоном, эфиром. Досушивание проводят на воздухе, в вытяжном шкафу и эксикаторе. Датская компания «Ново индастри» производит человеческий инсулин методом, в основе которого лежит замена остатка аланина, в цепи В на остаток треонина. Это достигается путем ферментативного замещения с последующей хроматографической очисткой продукта, в результате чего 155 получается однокомпанентный инсулин человека, с 99% содержанием чистого препарата. Создание промышленного производства генно-инженерного инсулина. Первой попыткой создать производство в Российской Федерации можно считать пилотную установку ВНИИА, основанную на технологии, разработанной совместно с немецкой фирмой Genbiotech GmbH (Гейдельберг) в 1987—1989 гг. Процесс был основан на продуцировании генноинженерным штаммом Е. coli рекомбинантного белка (молекулярная масса около 17000 Да), содержащего лидерную аминокислотную последовательность, соединенную через аминокислотный остаток метионина с проинсулином человека. Исходная технологическая схема включала в себя следующие стадии: 1. Ферментация (выращивание штамма-продуцента в ферментере). 2. Отделение биомассы. 3. Разрушение клеток с выделением телец включения. 4. Очистка рекомбинантного белка последовательно ионообменной хроматографией на DEAE-сефарозе FF и гель-фильтрацией на сефадексе G25M. 5. Расщепление рекомбинантного белка бромцианом. 6. Окислительный сульфитолиз проинсулина. 7. Хроматографическая очистка гексасульфоната проинсулина последовательной гель-фильтрацией на Сефадексе G-50 F, ионообменной хроматографией на DEAE-сефарозе FF и гель-фильтрацией на сефадексе G25M. 8. Ренатурация проинсулина. 9. Хроматографическая очистка нативного проинсулина ионообменной хроматографией на DEAE-сефарозе FF. 10. Ферментативное расщепление проинсулина. 11. Хроматографическая очистка инсулина ионообменной хроматографией на S-сефарозе FF, ультрафильтрацией и гель-фильтрацией на сефадексе G-50 SG. 12. Лиофильная сушка Na-соли инсулина человека. Несмотря на очевидную громоздкость приведенной схемы технологического процесса на опытной установке ВННИА были выпущены опытные партии субстанции генноинженерного инсулина человека (ГИЧ) и зарегистрирована соответствующая ВФС (временная фармакопейная статья. Однако, с распадом Советского Союза и сопутствующими экономическими и организационными последствиями выпуск препарата во ВНИИА более не производился. В 1996 г. был издан Указ Президента РФ «О мерах государственной поддержки лиц, больных сахарным диабетом», в соответствии с которым Правительством РФ была принята Федеральная программа «Сахарный диабет». На основании этого, под эгидой РАО «Биопрепарат» в ИБХ РАН и ГНЦ прикладной микробиологии (г .Оболенск, МО) были возобновлены 156 широкомасштабные работы по созданию промышленной технологии и производства препаратов генноинженерного инсулина человека. В рамках совместных работ были созданы высокоэффективные штаммы продуценты препроинсулина человека и разработана лабораторная технология получения субстанции инсулина. К настоящему времени в ЗАО «Национальные биотехнологии» (правопреемник РАО «Биопрепарат») в г. Оболенске открыто пилотное производство субстанции генноинженерного инсулина человека, но возможность выпуска готовых лекарственных форм отсутствует. Технологический процесс производства субстанции ГИЧ в ИБХ РАН основан на следующей схеме: 1. Получение посевного материала штамма-продуцента 1.1. Оживление консервированной культуры. 1.2. Выращивание маточной культуры. 1.3. Выращивание инокулята. 2. Биосинтез гибридного белка 2.1. Выращивание продуцента в ферментере (ферментация). 2.2. Получение биомассы (сепарирование). 2.3. Дезинтеграция клеточной суспензии. 2.4. Выделение и отмывка телец включения (центрифугирование). 3. Выделение и очистка рекомбинантного белка 3.1. Солюбилизация телец включения и восстановление рекомбинантного белка. 3.2. Ренатурация рекомбинантного белка. 3.3. Хроматографическая очистка рекомбинантного белка. 4. Ферментативное расщепление рекомбинантного белка 5. Хроматографическая очистка инсулина 1 6. Хроматографическая очистка инсулина 2 7. Хроматографическая очистка инсулина 3 8. Получение кристаллического инсулина В основе процесса биосинтеза — использование штамма-продуцента Е. coli JM 109 с рекомбинантной плазмидой pPINS 07.Рекомбинантная плазмидная ДНК содержит искусственный ген, кодирующий гибридный полипептид, состоящий из одного IgG-связывающего домена белка А из Staphylococcus aureus, пептидного линкера His6GlySerArg и проинсулина человека. Плазмида (молекулярная масса 3,3 МДа, 5051 п. о.) содержит в качестве генетического маркера ген β- лактамазы, определяющий устойчивость трансформированных ею клеток бактерий к ампициллину. Экспрессия рекомбинантного белка штаммом-продуцентом индуцируется изопропил- β-D-тиогалактозидом (благодаря наличию в плазмиде tаcпромотора) и достигает 30% от суммарного клеточного белка. Клетки продуцента рекомбинантного препроинсулина человека отделяют сепарированием, дезинтегрируют при высоком давлении для сбора телец включения, многократно отмывают полученные тельца включения от 157 водорастворимых белков и иных клеточных компонентов. За 1 цикл получают 15—25 кг влажных телец включения, содержащих более 13% рекомбинантного белка. Полученные тельца включения солюбилизируют в буферном растворе, содержащем мочевину и дитиотреитол (ДТТ) для восстановления внутри- и межмолекулярных ди-сульфидных связей в рекомбинантном белке. Далее восстановленный рекомбинантный белок подвергают воздействию кислорода воздуха при сильном разбавлении, при этом белок окисляется с образованием дисульфидных связей, соответствующих нативной конформации инсулина (рефолдинг). Происходит ренатурация рекомбинантного белка. Ренатурированный рекомбинантный белок очищают ионообменной хроматографией и подвергают ферментативному расщеплению трипсином и карбокси-пептидазой Б в массовом соотношении 4000 : 2 : 1. После осаждения и предварительной обработки получают инсулин-сырец 86—88%ной чистоты. На первой стадии очистки гидрофобной хроматографией содержание инсулина достигает 96%, на второй стадии очистки ионообменной хроматографией проводится окончательное освобождение от иммуногенных, финишная гель-фильтрация проходит в стерильных условиях и позволяет довести качество препарата до фармакопейного. В завершении процесса получают кристаллический цинк-инсулин, который высушивают до необходимой степени влажности. Выход продукта составляет не менее 100 г с 1000 л культуральной жидкости штамма-продуцента. Преимущества процесса: — одностадийная очистка ренатурированного рекомбинантного белка; — совместное ферментативное расщепление рекомбинантного белка трипсином и карбоксипептида-зой Б; — эффективное использование хроматографии низкого давления для очистки инсулина. Требования к качеству субстанции инсулина человека изложены в соответствующих разделах Американской Фармакопеи (Ф. США) и Европейской Фармакопеи (ЕФ). Инсулин должен быть охарактеризован качественно и. Первый показатель определяется методом ВЭЖХ в сравнении со стандартом, сравнением хроматографических профилей инсулина и инсулина-стандарта, расщепленных специфическим ферментом на 4 фрагмента («метод пептидных карт») и определением биоидентичности (in vivo, на кроликах, по сравнительному содержанию глюкозы в крови). Количественное определение проводится также методом обращенно-фазовой ВЭЖХ. Количество гормона принято оценивать в международных единицах (ME), при этом 1 ME (IU) = 0,0347 мг инсулина. 13.2. Препараты щитовидной железы Тиреоидин. Гормональный препарат, получаемый из высушенных обезжиренных щитовидных желез убойного скота. Щитовидные железы извлекают немедленно после убоя от нормально развитых и здорвых 158 животных на бойнях или мясокомбинатах. Для производства препарата их замораживают при температуре от -8 до -12 ºС и доставляют в морозильных камерах для переработки. Перед переработкой отобранные железы размораживают, моют в воде, очищают от окружающих посторонних тканей: жира, соединительных тканей, мышц, крупных сосудов и т.д. Затем железы измельчают, и полученную кашу раскладывают на плоские эмалированные противени и высушивают в вакуум-сушильном шкафу при температуре не выше 40ºС. После чего материал обезжиривают в аппарате Сокслета органическими растворителями с низкой температурой кипения, хорошо извлекающими жиры. Остатки органических растворителей удаляют из сырья просушиванием в вакуум-сушилках при температуре не выше 40ºС. Сухую обезжиренную массу измельчают в фарфоровых шаровых мельницах. Препарат стандартизируют по содержанию органически связанного йода, которого должно быть 0,17-0,23%.При необходимости препарат разбавляют молочным сахаром. Действие данного препарата обусловлено наличием тироксина и трийодтиронина. Химически тироксин отличается от трийдодтиронина наличием дополнительного атома йода. Тиреоидин назначают при недостаточной функции щитовидной железы. 13.3. Препараты гипофиза Из передней доли гипофиза убойных животных получают для медицинского применения препараты: кортикотропин для инъекций, суспензию цинк- кортикотропина, тиротропин, адипозин. Кортикотропин. Адренокортикотропный гормон (АКТГ). Образуется в базальных клетках передней доли гипофиз. Это полипептидный гормон, состоящий из 39 аминокислот. Его активность определяется биологическим путем и выражается в единицах действия. Кортикотропин является физиологическим стимулятором коры надпочечников. Распространенный способ промышленного производства гормонов из гипофиза разработан во ВНИИ технологии кровезаменителей и гормональных препаратов. Заключается он в комплексной переработке сырья, когда последовательно выделяют отдельные гормоны из передней доли гипофиз. Из свежезамороженных передних долей гипофизов готовят фарш. Который экстрагируют подкисленным ацетоном (1% раствор HCl в 90% ацетоне). Кислая водно-ацетоновая вытяжка центрифугируется. В фильтре находиться АКТГ и лактогенный гормон, которые осаждаются ацетоном. Концентрация ацетона в смеси достигается 92%. Эта смесь отстаивается на холоде при температуре от -2 до -5 °С в течение 10-12 ч. Получают кислый ацетонированный осадок, который отделяют, промывают на нутч-фильтре охлажденным 98% ацетоном и высушивают на воздухе. Кислый ацетонированный порошок растворяют в воде, подкисленной уксусной кислотой, и постепенно добавляют раствор аммиака до значения pH 5,0. В изоэлектрической точке осаждается лактогенный гормон. Осадок отделяют 159 центрифугированием и используют для получения препарата лактина. После отделения лактогенного гормона к оставшемуся добавляют аммонийноацетатный буферный раствор (pH 5,0) и пропускают через колонку, наполненную катионитом КМ-сефадекс К-25. После завершения сорбции кортикотропина на ионообменной смоле проводят его десорбцию этанольным раствором аммонийно-ацететного буфера. Из элюэнта АКТГ осаждают этанолом. Осадок отделяют центрифугированием, промывают этанолом, ацетоном и высушивают на воздухе. Препарат стандартизуют. Из оставшегося от изначального сырья далее получают фоллукилостимулирующий, лютеинизирующий и тиреотропный гормоны. Разработан ряд методик подобного ступенчатого выделения лютеинизирующего, фолликулостимулирующего и тиреотропного гормонов из тканей после выделения гормона роста, но уже человеческого гипофиза. 13.4. Гормон роста Гормон роста был открыт в 1920-х гг., а получен в кристаллическом виде из гипофиза животных в 1944 г. учеными Лайем и Эвансом. В 1956 г. был выделен человеческий соматотропин, а в 1958 г. эндокринолог из Новоанглийского медицинского центра в Бостоне Морис Рабен впервые ввел его ребенку, который не рос из-за того, что его организм вообще не вырабатывал этого гормона. Лечение помогло, и ребенок стал расти. На тот момент единственным источником получения гормона роста был человеческий мозг – мозг трупов, так как гормон животного происхождения не эффективен для человека. Большей частью трупный материал поступал из Африки. Гормон извлекался из гипофиза, и, поскольку он разрушается при нагревании, на фармацевтических заводах его пастеризовали, а не стерилизовали. В 1980-х гг. сразу у троих детей, получавших гормон роста, развилось редкое вирусное заболевание – болезнь Крейцфельдта – Джейкоба (БКД). Оно характеризуется прогрессирующим слабоумием и потерей контроля над мышцами. В течение примерно 5 лет больной человек погибает. После обнаружения болезни у детей, получавших гормон роста, распространение лекарства было остановлено. В 1991 г. БКД развилась у семерых детей в США, а по всему миру насчитывалось 50 случаев заболевания, связанных с инъекциями гормона роста. И количество больных может продолжать расти, поскольку болезнь вызывается инфекционным агентом, который до появления симптомов может не давать о себе знать длительное время (до 15 лет). Поскольку от мозга трупов как источника гормона пришлось отказаться, возникла очень трудная проблема – получение синтетического гормона. Гормон роста – это крупнейший белок, производимый гипофизом и состоящий из 191 аминокислоты. В 1985 г. компания «Генентех», которая ранее успешно клонировала человеческий ген на инсулин, создала второе в истории лекарство на основе 160 рекомбинантной ДНК – соматотропин. Генно-инженерный гормон роста отличался от своего человеческого аналога одной только аминокислотой. И хотя это небольшое несоответствие никак не отразилось на эффективности препарата при его действии на человеческий организм, оно открыло дверь для конкурентов. На следующий год базирующаяся в Индианаполисе фармацевтическая компания «Эли Лилли» создала состоящий из 191 аминокислоты новый гормон роста (препарат гуматотроп), который был на 100 % идентичен (физически, химически и биологически) тому, который вырабатывается гипофизом человека. На сегодняшний день из-за монополии этих компаний лечение препаратом обходится пациенту от 14 до 30 тыс. долларов в год, что создает трудности при лечении, но экономического порядка. После выделения гормона железой он попадает в кровеносное русло, где немедленно связывается с альбуминами и другими специфическими переносчиками, только 5 % секретируемого гормона остается в крови в несвязанном состоянии. С кровотоком гормон доставляется ко всем органам и тканям и контактирует с ними. Однако его специфическое действие начинается после соединения с чувствительными только к этому гормону рецепторами. Известно, что рецепторы к гормонам, имеющим пептидную, белковую и аминокислотную структуры, расположены на поверхности клеток, а рецепторы к гормонам – дериватам жирных кислот и стероидам находятся внутри клетки (в цитоплазме или на мембране ядра). Специфическое избирательное восприятие гормонального сигнала объясняется высочайшим сродством и чувствительностью рецепторов и гормонов. Дефицит гормона в организме может иметь очень различную основу и ее необходимо всегда устанавливать. Например, недостаток инсулина может быть от того, что его мало вырабатывается в организме, он быстро расщепляется инсулиназой, вступает в новые соединения; кроме того, структура эфферентных систем не взаимодействует с ним. Каждая из перечисленных причин может быть обусловлена различными факторами. Так, слабая выработка инсулина может иметь в основе: а) истощение продуцирующих клеток; б) подавление функции продуцирующих клеток аллоксаном, дегидроаскорбиновои кислотой, нингидрином и др.; в) нарушение отдельных звеньев синтеза инсулина. Из этого ясно, что гормональные препараты эффективны только в определенных условиях. В соответствии со специфичностью фармакодинамики гормональных препаратов их наиболее часто применяют: для восполнения недостающего гормона в организме (при избытке гормона обычно используют фармакологические средства, противодействующие гормону или снижающие функцию гормонопроизводящей железы); для специфической фармакотерапии и фармакостимуляции при заболеваниях, не связанных с недостатком гормона в организме. Широко известно применение гормональных стероидных препаратов в спорте. Их обнаружение уже не является первостепенной задачей и от 161 контролирующих лабораторий требуют детекции малых количеств препаратов-аналогов. Активно применяются гормональные препаратыконтрацептивы. Гормональные препараты используются как в медицине так и животноводстве. Разрабатываются методики выращивания свиней с использованием регулярных инъекций инсулина для ускорения набора веса и стимуляция охоты у коров введением фоллюкулостимулируещего гормона. 13.5. Получение промышленно важных стероидов К стероидам относится большая группа биологически важных соединений, среди которых – половые гормоны, сердечные гликозиды, желчные кислоты, витамины, алкалоиды, регуляторы роста растений. В основе стероидов лежит скелет пергидроциклопентанофенантрена. Процессами биотрансформации называют реакции превращения исходных органических соединений (предшественников) в целевой продукт с помощью клеток живых организмов или ферментов, выделенных из них. Способность клеток микроорганизмов к высокоспецифичной биотрансформации используется в производстве стероидов. Использование абсолютной стереоспецифичности и субстратной специфичности ферментов клеток позволило разработать условия осуществления множества химических реакций для структурных перестроек стероидов. В результате были получены новые соединения с лучшими фармакологическими свойствами. Биотрансформация стероидов обычно заключается в селективном воздействии на одно из положений стероидного скелета. Значимость разработанной микробной трансформации определяется тем, что процессы гидроксилирования прогестерона и его производных лежат в основе промышленного синтеза многих ценных продуктов: противовоспалительных и противоопухолевых препаратов, транквилизаторов, анестезирующих средств, половых гормонов и др. Кроме биотрансформации, стероидные гормоны можно получать с помощью культур клеток растений. Например, культура клеток корня растения диоскореи дельтовидной (Dioscorea deltoidea) продуцирует фитостерин, диосгенин и его гликозидные производные (сапонины). Существенно, что способность к сверхсинтезу фуростаноловых гликозидов ряда штаммов диоскореи, например, штамма ДМ-ОГ, стабильно поддерживалась в течение 27 лет. Таким образом, культивирование клеток растений in vitro представляет собой новое решение проблемы промышленного получения вторичных метаболитов. В дальнейшем для производства стероидных гормонов прогнозируется применение иммобилизованных клеток, использование оптимального сочетания биологических и химических превращений, а также совершенствование технологии очистки получаемых соединений. 162 ГЛАВА 14. Биотехнология получения ферментов Источники получения ферментных препаратов. Ферменты присущи всем живым объектам и находятся практически во всех растениях, животных и микроорганизмах. Однако процесс биосинтеза ферментов в организме связан с обеспечением метаболизма клеток, и количество синтезируемых ферментов строго определяется жизненной потребностью организма; такие объекты не могут служить источником получения ферментных препаратов. Для этого пригодны только микроорганизмы, некоторые растения или отдельные органы растений и животных, способные накапливать значительное количество ферментов. Растительное сырье. Источником ферментов может быть проращенное зерно различных злаков (солод). Оно может либо использоваться непосредственно как технический ферментный препарат, либо служить исходным материалом для получения очищенных ферментных препаратов. В тропических и субтропических странах в качестве сырья для промышленного производства протеиназ используют латекс дынного дерева, латекс растений, относящихся к виду фикусовых, например, листья, побеги инжира, сок зеленой массы ананаса и др. Органы, и ткани животных. На всех мясоперерабатывающих комбинатах собирают сырье, содержащее ферменты, консервируют его и используют для получения ферментных препаратов. Таким сырьем являются поджелудочная железа, слизистые оболочки желудков и тонких кишок свиней, сычуги крупного рогатого скота, сычужки молочных телят и ягнят, семенники половозрелых животных. Поджелудочная железа содержит большое количество разнообразных ферментов: химотрипсин, коллагеназу, эластазу, трипсин, амилазу, липазу и др. Слизистая оболочка желудков свиней и сычугов крупного рогатого скота служит источником пепсина и липазы. Из сычужков молочных телят и ягнят получают реннин (сычужный фермент). Семенники половозрелого скота содержат фермент гиалуронидазу. Микроорганизмы. В специально созданных условиях микроорганизмы способны синтезировать огромное количество разнообразных ферментов. Они неприхотливы к составу питательной среды, легко переключаются с синтеза одного фермента на другой и имеют сравнительно короткий цикл роста (16-100 ч). Для промышленного получения ферментных препаратов используют как природные штаммы микроорганизмов, выделенные из естественных объектов, так и мутантные штаммы. Продуцентами ферментов могут быть различные микроорганизмы: бактерии, грибы, дрожжи, актиномицеты. Микроорганизмы могут синтезировать одновременно целый комплекс ферментов, но есть и такие, особенно среди мутантных штаммов, которые являются моноферментными и образуют в больших количествах только один фермент. Классификация и номенклатура ферментов и ферментных препаратов. По современной классификации все ферменты делятся на шесть 163 основных классов по типу катализируемой реакции: 1) оксидоредуктазы; 2) трансферазы; 3) гидролазы; 4) лиазы; 5) изомеразы и 6) лигазы (синтетазы). Большинство промышленно важных ферментов, потребность в которых определяется десятками тысяч тонн, относятся к третьему классу гидролазам. Подавляющее количество препаратов, выпускаемых различными фирмами мира, являются комплексными, содержащими помимо основного фермента еще значительное количество сопутствующих ферментов и белков. Поэтому в технологии ферментов препараты чаще классифицируют по основному компоненту в смеси ферментов, присутствующих в данном препарате: амилолитические, протеолитические, липолитические и т. д. В нашей стране существует определенная система названия ферментных препаратов, в которой учитываются основной фермент, источник получения и степень очистки. Наименование каждого препарата включает сокращенное название основного фермента, затем добавляется видовое название продуцента и заканчивается название препарата суффиксом «ин». Например, амилолитические препараты, получаемые из культур Aspergillus oryzae и Bacillus subtilis, называются соответственно амил-ориз-ин (амило-ризин) и амил-о-субтил-ин (амилосубтилин). Далее ставится индекс, в котором обозначены способ производства и степень очистки фермента от балластных веществ. При глубинном способе культивирования после названия ставится буква Г, а при поверхностном - П. Если это неочищенная культура продуцента, то далее следует буква х. Между буквами П, Г и х может стоять цифра, обозначающая степень чистоты препарата. Индекс 2 обозначает жидкий неочищенный концентрат исходной культуры; 3 - сухой ферментный препарат, полученный высушиванием распылением неочищенного раствора фермента (экстракт из поверхностной культуры или культуральной жидкости); 10 - сухие препараты, полученные осаждением ферментов органическими растворителями или методом высаливания. Индексы 15, 18, 20 обозначают препараты, частично освобожденные не только от балластных веществ, но и от сопутствующих ферментов. Номенклатура препаратов с индексом выше 20 не используется, так как в этих случаях речь идет о высокоочищенных и даже гомогенных ферментных препаратах, которые именуются согласно классической номенклатуре и классификации ферментов. Характеристика активности ферментных препаратов. Ферменты являются веществами белковой природы, поэтому в смеси с другими белками определить их количество невозможно. Наличие определенного фермента в данном препарате может быть установлено по результатам той реакции, которую катализирует фермент, т. е. по количеству образовавшихся продуктов реакции или уменьшению исходного субстрата. В количественном выражении условно активность фермента определяется по начальной скорости ферментативной реакции. Начальная скорость зависит от многих факторов, наиболее важные из них - температура, концентрация субстрата, рН реакционной смеси и время от начала реакции. Поэтому по предложению 164 Комиссии по ферментам Международного биохимического союза были приняты правила определения активностей препаратов и их выражения в единицах активности. Стандартная единица активности. Эта величина для любого фермента обозначает то количество его, которое катализирует превращение 1 мкмоль субстрата в 1 мин при заданных регламентированных условиях. На русском и немецком языках эта единица обозначается буквой Е, на английском, французском, итальянском и испанском - U. Часто количество субстрата нельзя выразить числом микромолей, так как точно не известна масса молекулы, например, при действии на белок, крахмал, пектин, целлюлозу. В этих случаях определяют микроэквивалент затронутых реакцией групп. Так, при гидролизе белка учитывают не число прогидролизованных молекул, а число образовавшихся свободных карбоксильных или аминных групп, т. е. число расщепленных пептидных связей; при гидролизе крахмала и полисахаридов число прогидролизованных глюкозидных связей и т. д. Комиссия по ферментам рекомендовала придерживаться определенных условий при установлении активности фермента: стараться вести определение при температуре 30 °С и определять активность по начальной скорости реакции, когда концентрация субстрата достаточна для насыщения фермента и соответствует кинетике реакции нулевого порядка. Концентрации субстрата, фермента и рН выбирают оптимальными для данного фермента. Если количество прореагировавшего субстрата очень мало или велико, допускается выражение результатов в миллиединицах (мЕ или мU) и килоединицах (кЕ и кU). Активность ферментных препаратов. Содержание фермента в данном препарате условно выражается в стандартных единицах активности фермента на 1 мл ферментного раствора или 1 г препарата. Активность ферментного препарата выражается в микромолях субстрата, прореагировавшего в присутствии 1 мл ферментного раствора или 1 г препарата в заданных условиях за 1 мин. Число микромолей и будет равно числу стандартных единиц. Если фермент гомогенен, то его удельная активность может быть выражена в стандартных единицах на 1 мг фермента: если же препарат содержит балласт в виде неактивного белка, его удельная активность выражается в стандартных единицах на 1 мг белка в ферментном препарате. Молекулярная активность представляет собой число миллимолей субстрата или эквивалентов затронутой реакцией групп, прореагировавших в течение 1 мин с 1 ммоль фермента при оптимальных концентрациях субстрата, или число стандартных единиц, содержащихся в 1 ммоль фермента. Если фермент содержит характерную простетическую группу или несколько каталитических центров, которые поддаются измерению, его активность можно выразить в величинах активности каталитического центра. 165 Такая активность будет соответствовать молекулярной активности, если молекула фермента имеет один активный центр; если же число каталитических центров n, то активность одного центра будет в n раз меньше молекулярной. Активность условного препарата. В технологии ферментов помимо общепринятых понятий об активности ферментных препаратов принято пользоваться понятием активности условного ферментного препарата. Это необходимо для оценки работы предприятия, сравнения его с другими аналогичными заводами, т. е. для сопоставления показателей по всем видам выпускаемой продукции. Для осуществления этого пересчета предполагают, что предприятие выпускает товарную продукцию в виде стандартного препарата с точно определенной активностью, измеряемой по основному ферменту в стандартных единицах в препарате на единицу массы препарата. Активность основного фермента в таком стандартном условном препарате устанавливается нормативами и называется активностью условного препарата. За 1 усл. т ферментного препарата принимается 1 т препарата со стандартной активностью. Для пересчета выработанной товарной продукции в условные тонны можно пользоваться формулой Qусл=QтовАф / Aусл , где Qусл - количество условного препарата, т; Qтов - количество товарного препарата, т; Аф - фактическая активность товарного препарата, ед./г; Aусл - активность условного препарата, ед./г. 14.1 Область применения и источники ферментов Производство ферментных препаратов занимает одно из ведущих мест в современной биотехнологии. Постоянно увеличивается объем их выпуска и расширяется сфера применения. Ферменты являются высокоактивными нетоксичными биокатализаторами белкового происхождения. Их преимуществом перед химическими катализаторами является действие при нормальном давлении, температуре от 20 до 70 ºС, рН от 4 до 9. Они имеют высокую субстратную специфичность, что позволяет в сложной смеси субстратов направленно воздействовать только на определенные соединения. Согласно принятой классификации и номенклатуре сейчас идентифицировано около 2000 ферментов. Промышленно выпускается около 250 наименований, причем 99 % общей суммы реализации ферментных препаратов приходится только на 18 ферментов. Наибольший удельный вес среди выпускаемых препаратов занимают протеиназы, широко используемые в синтетических моющих средствах, и амилазы для переработки крахмала. Эти два вида препаратов составляют 60 % общего объема выпуска ферментных препаратов за рубежом. Другими крупными отраслями – потребителями ферментов, являются – производство вин и соков 10 %; – производство спирта 8 %; 166 – сыроделие 5 %; – хлебопечение 5 %; – пивоварение 6 %; – прочие отрасли 6 %. В России, кроме того, ферменты внедряются в кормопроизводство. Особое место в общем объеме производства ферментов занимают высокоочищенные ферментные препараты. Их доля в общем объеме очень мала, так как технология сложна, требует больших материальных затрат и времени. Эти препараты очень важны для медицины, аналитических целей и научных исследований. Ферменты присущи всем живым существам, однако для их выделения используют только те природные объекты, в которых содержание используемого энзима составляет не менее 1 %. Источниками ферментов могут быть: 1) проросшее зерно различных злаков (солод) – для получения амилаз, латекс фикусовых, дынного дерева – для получения протеиназ; 2) отдельные ткани и органы животных (поджелудочная железа, слизистые оболочки желудков и тонких кишок, сырный сычуг крупного рогатого скота); 3) микроорганизмы. В специфических условиях микроорганизмы способны синтезировать огромное количество ферментов. Они легко переключаются с синтеза одного фермента на другой, имеют короткий цикл роста (от 16 до 100 ч). Для промышленного получения ферментов используют как естественные штаммы, так и полученные с помощью мутагенеза, селекции и индукции биосинтеза. Ферменты способны синтезировать бактерии, грибы, дрожжи, актиномицеты; микроорганизмы могут быть моно- или полиферментами. 14.2 Выбор штамма и условий культивирования Целесообразность применения микроорганизмов для производства ферментов заключается в следующем: 1) генетическими манипуляциями удаѐтся в тысячу и более раз увеличить уровень катаболитных и в несколько сот раз уровень биосинтетических ферментов; 2) энергетически оправдано выращивание микробных клеток в больших масштабах в связи с применением недорогих сред и быстрого роста микроорганизмов; 3) огромное разнообразие реакций, к которым способны микроорганизмы. Это особенно касается вторичного метаболизма; 4) микроорганизмы служат источником не только таких ферментов, которые встречаются у животных и растений, но и ряда уникальных ферментов, нигде более не обнаруженных. Например, целлюлоза, танназа, гидрогеназа и др. Среди микроорганизмов есть виды, развивающиеся при 167 экстремально высоких температурах (87 ºС), в связи с чем потенциально возможно создание термостабильных штаммов; 5) способность микроорганизмов адаптироваться к различным окружающим условиям, что позволяет переносить культуру на производство, где она растет на дешевых субстратах. Промышленное производство и применение ферментов основано на двух важных факторах: во-первых, ферменты образуются в живых клетках; во-вторых, они могут проявлять свое специфическое действие в среде независимо от живых клеток. При первоначальном выделении штамма исходят из того, что микроорганизмы адаптируются к утилизации субстрата, находящегося в изобилии в местах их обитания, и, следовательно, образуют ферменты, реагирующие с этим субстратом. Поэтому продуценты целлюлаз и лигнолитических ферментов выделяются из лесных почв, продуценты пектиназ – из фруктов и растений, деструкторы мочевой кислоты – из птичьего загона и т.д. Ферменты близкородственных штаммов имеют сходные свойства, а у дальнеродственных – могут сильно отличаться. Первая ступень производства ферментов состоит в селекции организма, образующего желаемый фермент в наибольшем количестве. При этом учитывают следующие общие требования к продуценту: 1) желательно образование внеклеточных ферментов, так как их легче выделить; 2) высокий выход фермента за короткое время; 3) очистка фермента от культуральной жидкости должна быть легкой; 4) штаммы не должны продуцировать антибиотики, токсичные вещества и не должны быть родственниками штаммов, образующих токсины. Чтобы вызвать сверхсинтез ферментов используют ряд методов, связанных с изменением окружающих условий роста и ведущих к изменению генетики организма. 14.3 Технология культивирования микроорганизмов – продуцентов ферментов и выделение ферментов Технологический процесс можно разбить на три стадии: 1) получение посевного материала; 2) получение производственной культуры методами поверхностного или глубинного культивирования; 3) выделение из готовой производственной культуры технических или очищенных ферментных препаратов. Поверхностный метод состоит в культивировании микроорганизмов на поверхности увлажненных стерилизованных отрубей, размещенных в кюветах. Инкубацию ведут в специальном термостатируемом цехе при постоянном контроле в нем температуры, влажности и подачи воздуха. 168 Глубинный метод более экономичен. Для его реализации применяются ферментеры из нержавеющей стали, снабженные устройствами для перемешивания и подачи в жидкую питательную среду стерильного воздуха. Наиболее прогрессивен проточный метод, который обеспечивает непрерывную подачу в ферментер питательной среды и посевного материала и непрерывный отбор продуктов жизнедеятельности и микробиальной массы. Достоинством метода является возможность длительное время поддерживать в автоматическом режиме рост культуры микроорганизмов (до 200 суток). Выделение и очистка фермента из культуры микроорганизмов – достаточно трудоемкая и дорогостоящая процедура, поэтому, если фермент можно использовать в виде неочищенного препарата, его не очищают. В промышленности широко используют коммерческие препараты, чистота которых составляет всего 0,1 % (то есть 99,9 % составляют примеси). К таким отраслям относят спиртовую, кожевенную, текстильную промышленность, сельское хозяйство, производство бытовой химии. Неочищенные препараты получают в мягком режиме высушивания культуры микроорганизмов вместе с остатками питательной среды. Такие препараты получают или из экстракта культуры продуцента, выращенного поверхностным способом, или из фильтрата культурной жидкости продуцента, выращенного глубинным способом. Для большинства отраслей пищевой промышленности, научных исследований и медицины требуются очищенные ферментные препараты. Источником выделения фермента может быть биомасса микроорганизмов, экстракт или фильтрат культуральной жидкости. Биомассу микроорганизмов необходимо тщательно измельчить, вплоть до разрушения субклеточных структур: лисозом, митохондрий, ядер и т.д. Затем препараты очищают осаждением органическими растворителями, солями, после чего диализом, аффинной хроматографией, переосаждением, гель-фильтрацией, сорбцией, кристаллизацией и пр. Так как ферменты имеют белковую природу, то особое значение придают соблюдению режимов, сохраняющих их активность при инактивации сопутствующих балластных белков. Очищенные ферменты хранят при низкой температуре (до минус 80 ºС). Для стабилизации ферментов в их препараты добавляют коферменты и субстраты. Ферменты широко используются в различных областях практической деятельности человека как биологические катализаторы. Источниками ферментов могут быть животные, растения и микроорганизмы. К настоящему времени установлено наличие более двух тысяч ферментов, а несколько сотен из них получены как индивидуальные вещества. Микроорганизмы в качестве продуцентов ферментов представляют особый интерес, поскольку их метаболизм, а, следовательно, и работа ферментных систем осуществляются с очень большой интенсивностью. Если характеризовать суммарную интенсивность ферментных реакций по поглощению молекулярного кислорода в виде коэффициента Qo2 (см3 O2, 169 потребленное одним мг сухой биомассы за один час), то для клеток печени он равен 2-5; почки – 10-20, дрожжевой клетки – 50-110; у бактерий рода Acetobacter - 1800, а у Azotobacter – 2000. Помимо высокой интенсивности метаболизма, следует помнить об очень большой скорости прироста биомассы микроорганизмов. Это позволяет в течение коротких промежутков времени (иногда за 24-72 ч) получать такие количества сырья для выделения ферментов, которые не могут сравниться с тем, что дают растения и животные. Важное свойство многих микроорганизмов - способность расти, используя различные дешевые субстраты, в том числе не имеющие пищевого значения (целлюлоза, углеводороды нефти, метан, метанол и др.). Некоторые субстраты, используемые микроорганизмами, токсичны для человека и животных. Содержание отдельных ферментов в клетках микроорганизмов может быть весьма высоким. Так, количество рибулезобисфосфаткарбоксилазы у фототрофных бактерий иногда достигает 40-60% от всех растворимых белков. Многие микроорганизмы образуют ферменты, которые в большом количестве выделяются в культуральную среду. Эти ферменты в основном принадлежат к гидролазам, расщепляющим белки, крахмал, целлюлозу, жиры и другие нерастворимые в воде вещества. Выделение ферментных препаратов из клеток микроорганизмов и особенно из культуральной среды после их выращивания проще и экономичнее, чем из растительных и животных тканей. Ряд ферментов обнаружены только у микроорганизмов. К таким ферментам относятся: танназа, расщепляющая дигаллат до галловой кислоты, рацемазы многих аминокислот, кератиназы, гидролизующие серусодержащие белки - кератины, входящие в состав волоса, перьев, рогов и копыт. Некоторые микроорганизмы обладают специфическими декарбоксилазами аминокислот, образуют пенициллиназу, расщепляющую пенициллин до пенициллиновой кислоты и воды. Такой фермент, как нитрогеназа, участвующая в образовании аммиака из молекулярного азота, обнаружена лишь у бактерий, способных к фиксации N2. Характерной особенностью некоторых бактерий является их способность окислять неорганические субстраты: аммиак, нитриты, сульфид и другие соединения серы, а также двухвалентное железо, что связано с наличием у них особых ферментов. Ряд бактерий и водорослей синтезируют гидрогеназы, катализирующие окисление и образование молекулярного водорода. Значительное число бактерий способны синтезировать ферменты, позволяющие им использовать метан, метанол, метилированные амины, окись углерода и другие одноуглеродные соединения в качестве субстратов для роста. 170 Очистка окружающей среды от ряда загрязняющих ее веществ возможна благодаря способности ферментов, образуемых микроорганизмами, разрушать компоненты пластмасс, пестициды и другие ядовитые соединения. Способность микроорганизмов развиваться в экстремальных условиях, т. е. при низких и высоких температурах, в отсутствие молекулярного кислорода в кислых или щелочных средах, при высоких концентрациях солей, определяется часто характером их ферментов. Психрофильные дрожжи Candida gelida имеют оптимум роста 15°С, при нагревании до 35°С они погибают: у них необратимо инактивируется только пируваткарбоксилаза. У облигатно психрофильного штамма Micrococcus cryophilus кратковременное нагревание до 25°С изменяет вторичную и третичную структуру глутамин- и пролин-тРНК-синтетаз. В результате клетки теряют способность соединять т-РНК с соответствующей аминокислотой. Некоторые псевдомонады обнаруживают активный рост при +3, +4°С, а их ферменты активны при -5, -10°С. Исключительное положение среди живых существ занимают термофильные бактерии, имеющие оптимум роста при 60-80°С и выше. Возможность развития этих бактерий при высоких температурах обусловливается термостабильностью белков. Например, экстремальный термофил Thertnus aquaticus обладает энолазой с оптимумом действия при 90°С. Гриб Malbranchea pulchella var. sulfurica, способный расти при 45°С, выделяет в среду термостабильную протеиназу, сохраняющую активность при 73°С. Некоторые мезофильные микроорганизмы также образуют ферменты с высокой термостабильностью, проявляющие максимальную активность при более высокой температуре, чем нужна для роста продуцента. Необычна форма жизни галофильных микроорганизмов, рост их наблюдается даже в насыщенном растворе NaCl. Ферменты таких микроорганизмов требуют для проявления максимальной активности высоких концентраций солей. Отношения микробных ферментов к кислотности и щелочности среды во многих случаях также уникальны. Амилолитическая активность Aspergillus niger обеспечивается двумя амилазами, из которых одна стабильна при очень низких значениях рН. Некоторые амилазы, напротив, требуют для проявления активности резко щелочных условий. Например, амилаза Bacillus sp. проявляла максимум активности при рН 10,5. Известны бактерии, развивающиеся в щелочной среде (рН 9,0 и выше) и образующие ферменты с высоким оптимумом рН. Большой интерес как продуценты ферментов представляют анаэробные микроорганизмы. Многие анаэробные бактерии превращают аминокислоты, пурины, пиримидины и другие субстраты путями, отличными от тех, которые известны для аэробных микроорганизмов, животных и растений. 171 В последние годы установлено, что среди строгих анаэробов имеются бактерии, способные расти в хемолитоавтотрофных условиях. При этом у них действуют системы ассимиляции углекислоты, не свойственные другим организмам, что обусловлено наличием особых ферментов. К числу таких автотрофов принадлежат некоторые метанобразующие, ацетатобразующие и сульфатредуцирующие бактерии. Внимание, проявляемое к анаэробам, объясняется возможностью получения в результате их деятельности этанола, бутанола, метана, ацетата и других полезных продуктов при переработке растительных остатков и другого дешевого сырья. Культивирование анаэробов исключает необходимость аэрирования и перемешивания питательной среды; снижается возможность инфицирования, вызываемого недостаточной стерильностью воздуха. Кроме того, использование анаэробных микроорганизмов позволяет проводить процесс в больших емкостях с высоким слоем среды. Свойством многих ферментов микробного происхождения является их индуцибельность. Так, синтез β-галактозидазы у Escherichia coli индуцируется лактозой и начинается менее чем через три минуты после ее внесения, а при удалении индуктора синтез так же быстро прекращается. Поэтому путем изменения условий культивирования можно добиться образования микроорганизмами больших количеств многих практически важных ферментов. Известно, что ферменты, катализирующие одну и ту же реакцию, но полученные из разных организмов, могут существенно различаться по свойствам. Примером этого может служить глутаминаза, осуществляющая дезамидирование глутамина с образованием глутаминовой кислоты. У глутаминаз, полученных из клеток Azotobacter agilis, оптимум рН = 6,0 - 7,0; из Clostridium welchii - 5,0; из Mycobacterium tuberculosis - 8,3. Хлориды ингибируют глутаминазу Pseudomonas sp., но не влияют на ее активность у Е. coli и активируют глутаминазу Cl. welchii. Аспарагиназы Е. coli и Alcaligenes euirophus различны по субстратной специфичности. Резюмируя сказанное, следует подчеркнуть, что микроорганизмы обладают очень высокой активностью ферментативных реакций и способны осуществлять многие процессы, отсутствующие у макроорганизмов, благодаря наличию специфических ферментов. Ферменты сходной функции у микро- и макроорганизмов могут быть различны по свойствам и у микроорганизмов иногда для проявления своей активности нуждаются в особых условиях. Поэтому изучение ферментов разных микроорганизмов - задача весьма важная. Получение активных продуцентов. Мутанты в большинстве случаев ауксотрофны по ряду соединений, так как в них произошли определенные нарушения обмена веществ, вызвавшие гипертрофию некоторых функций клетки. Обычно активные штаммы, выявленные из естественных источников, подвергают действию мутагенов несколько раз, т. е. осуществляют ступенчатую селекцию. В результате получают 172 высокопродуктивные штаммы. Часто эффективно комбинированное воздействие мутагенов химической и физической природы. Так, применение этиленимина и ультрафиолетового излучения в сочетании со ступенчатым отборов позволило получить очень активные штаммы Asp. awamori, используемые как продуценты амилолитического, протеолитического и других ферментных комплексов. Селекция производственно ценных штаммов ведется и в условиях производства. Непременным условием правильно поставленного промышленного процесса являются такие мероприятия по сохранению микроорганизмовпродуцентов. Существует ряд методов хранения производственно ценных штаммов, обеспечивающих их высокую биохимическую активность (см. гл 8). В музеях живых культур при заводских лабораториях культуры периодически пересеваются. Однако пересевы через короткие промежутки времени могут снизить активность микроорганизмов, поэтому после развития культуры на плотной среде ее заливают стерильным вазелиновым маслом. Во многих случаях лучшим способом хранения является лиофилизация культур. Питательные среды для культивирования микроорганизмов. Питательные среды по своему назначению разделяются на три основные группы: среды для поддержания штаммов-продуцентов в лаборатории и в музее, среды для получения посевного материала и среды, употребляемые в больших объемах в основном процессе производства. Если первая группа сред подбирается исходя из физиологических потребностей микроорганизмов, то две последние группы должны удовлетворять и требованиям производства, т. е. они должны быть достаточно дешевыми. Состав сред, обеспечивающих накопление того или иного фермента, может значительно отличаться от состава сред, применяемых для выделения и поддержания культуры продуцентов. Как уже было отмечено выше, некоторые ферменты микроорганизмов индуцибельны и требуют для своего синтеза присутствия в среде индуктора, причем индуктором не всегда может быть субстрат, на который фермент действует. Так, например, если для синтеза липазы Aps. niger благоприятно присутствие в среде растительного масла, а для синтеза рибонуклеазы нуклеиновых кислот, то для повышенного синтеза α-амилазы Вас. polymyxa недостаточно присутствия в среде растворимого крахмала, необходимо добавление гидролизата казеина. При замене органических соединений азота неорганическими фермент не синтезировался. С целью удешевления производства ферментов подбирают наиболее дешевые источники сырья, пригодные для культивирования микроорганизмов-продуцентов При промышленном получении гидролаз обычно используют различные отходы, содержащие специфические субстраты. Так, в производстве пектолитических ферментов применяют пектинсодержащее сырье, например свекловичный жом, для целлюлолитических - солому, отруби и другие целлюлозосодержащие субстраты. Но использование в 173 промышленности сред с такими нерастворимыми субстратами не всегда оказывается технологичным, а также затрудняет направленное ведение ферментации при глубинном культивировании продуцентов Применение сред с растворимым соединением углерода дает возможность устранить эти недостатки, сократить сроки ферментации и вести процесс на современном уровне. Перспективным сырьем для получения некоторых микробных экзоферментов (пектиназы, целлюлазы и др.) признана молочная сыворотка - дешевый побочный продукт производства сыра, творога, казеина Использование молочной сыворотки в качестве питательной среды позволяет также осуществить непрерывный способ получения ряда экзоферментов. Для получения целлюлазы при помощи термотолерантного штамма Asp. terreus подобрана дешевая питательная среда с пшеничной соломой, которая может быть использована в виде сечки и муки. При приготовлении сред для промышленного выращивания продуцентов ферментов может быть использована биомасса других микроорганизмов, подвергнутая гидролизу или экстракции. Разработан способ экстрагирования биомассы грибов-продуцентов пектиназы и последующего введения экстрактов в питательную среду, позволяющий полностью утилизировать всю получаемую биомассу, сократив на 9% количество экстракта свекловичного жома и интенсифицировав биосинтез пектолитических ферментов в 2,4 раза. Разработана также замкнутая технологическая схема получения пектолитических ферментов, позволяющая полностью утилизировать отходы микробиологического производства, путем возврата их в технологический процесс. Поиски новых питательных сред, удовлетворяющих современное производство ферментов, способных заменить исходное сырье (кормовые и пищевые продукты), актуальная проблема биотехнологии. 14.4. Выделение и стабилизация ферментов. Выбор метода выделения и очистки ферментов микроорганизмов различны и определяются в зависимости от локализации (в клетках фермент или в культуральной среде) и целями применения. Неочищенные ферментные препараты получают путем сушки и размельчения мицелия гриба вместе с твердым субстратом (отрубями, жомом и др.) или высушиванием продуцента вместе с культуральной жидкостью на распылительной сушилке. Высушенные препараты размалываются в порошок и в таком виде используются. Удельная ферментная активность таких препаратов невелика, но они дешевле и достаточно устойчивы при хранении. Таким способом получают амилазы, протеазиназы, целлюлазы для сельского хозяйства и некоторых отраслей промышленности. Получение сиропов производится при сгущении отделенной от биомассы микроорганизма культуральной жидкости, содержащей фермент. 174 В лабораторной практике и промышленности существуют различные способы концентрирования, однако лишь некоторые из них могут быть использованы для сгущения ферментных растворов. При промышленном производстве ферментов наряду с активностью важными показателями являются их термоустойчивость, рН и стабильность. Следует также учитывать, что увеличения концентрации неочищенных растворов ферментов может сопровождаться повышением содержания ингибирующих примесей до критического с одновременным разложением и удалением стабилизирующих продуктов. Но обычно уменьшение содержания воды при сгущении приводит к снижению скорости химических и биохимических процессов. Способов концентрирования известно несколько: 1) без изменения фаз (мембранные); 2) с изменением фаз (вымораживание, выпаривание). Концентрирование с помощью мембран основано на том, что состав жидкости может быть изменен при пропускании ее через мембрану с селективной проницаемостью. Идеальная мембрана должна быстро пропускать необходимое вещество из раствора, но быть барьером для всех других компонентов. Возможен и другой вариант мембраны, при котором необходимое вещество остается, а все другие компоненты пропускаются. На практике применяют мембраны обоих типов. Для сгущения термочувствительных жидкостей используется прием, основанный на избирательной кристаллизации воды в растворе при охлаждении ниже 0°. Сущность этого метода концентрирования заключается в том, что вода сгущаемого раствора превращается в кристаллы льда, увеличивая тем самым концентрацию растворимых веществ. Кристаллы льда удаляются центрифугированием или прессованием. Операция повторяется многократно до получения необходимой концентрации фермента. При таких условиях полностью исключаются потери термостабильных компонентов раствора (отсутствует термическое разрушение). Разработка новых способов концентрирования биологических жидкостей не умаляет значений такого общеизвестного приема, как вакуумвыпаривание. Обилие конструкций выпарных установок и уровень современного развития исследований в этой области открывают большие возможности использования процесса выпаривания. Физический смысл выпаривайся растворов - разделение их на две части: воду, удаляемую виде пара, и обогащенный растворимыми веществами концентрат. В ферментной промышленности широко распространен процесс концентрирования водных растворов под вакуумом. Так производят, например, сгущение водного экстракта из мицелия Asp. oryzae для получения препарата протеоризина. Большое количество ферментных препаратов производят с использованием способа вакуумконцентрирования. 175 Высушивание ферментных препаратов производится в вакууме при распылении или из замороженного состояния. После высушивания препарат в случай нестойкости необходимо смешивать со стабилизатором или с наполнителем (крахмал, декстрины, неорганические нейтральные соединения, тальк и др.). В некоторых случаях удается стабилизировать фермент добавлением в культуральную жидкость хелатирующих агентов. Так удалось стабилизировать щелочную протеазу Вас. subtilis добавлением в среду триполифосфата натрия, нитрилотриацетата натрия и этилендиаминтетраацетата. Используются ферменты и в виде гомогенных белков. Получение ферментов в чистом виде осуществляется различными способами в зависимости от их свойств и области применения. Обычная схема получения экзоферментов может быть следующей: Культуральная жидкость ↓ центрифугирование Надосадочная жидкость ↓ диализ Диализат ↓ выпаривание в вакууме I. Концентрат ↓ осаждение солями аммония Осадок ↓ диализ Водный раствор ↓ гельфильтрация Обессоленный раствор ↓ выпаривание II. Ферментный препарат Обессоленный раствор ↓ осаждение ацетоном Фильтрат ↓ фракционирование ацетоном Осадок ↓ Водный раствор ↓ гельфильтрация Раствор фермента ↓ высушивание в вакууме III. Чистый фермент В промышленности высокая степень очистки ферментов не всегда нужна. Очистка должна быть очень высокой в случае использования ферментов как терапевтических препаратов. Очистку и разделение ферментов, образующих комплексы, проводят, применяя ионообменные смолы. Ионообменный метод, например, позволяет разделить ферменты пектолитического комплекса грибов, очистить и выделить пектинэстеразы, полигалактуроназы и пектинтрасэлиминазы. Для использования фермента как терапевтического препарата применяется иногда микрокапсулирование. Фермент, заключенный в специальные капсулы, достигает органа или ткани, где он должен проявить свою активность, там капсула растворяется и фермент попадает в тканевую жидкость В настоящее время все шире используются так называемые иммобилизованные ферменты, т. е. ферменты, прочно связанные с 176 различными носителями. Помимо иммобилизации высоко очищенных ферментов, весьма перспективен способ иммобилизации целых клеток микроорганизмов, обладающих той или иной ферментной активностью. В этом случае фермент более стабилен. Схема очистки фермента от балластных веществ сводится к освобождению его от нерастворимых веществ, сопутствующих растворимых веществ и других ферментов. Процессы получения очищенных препаратов из поверхностных и глубинных культур несколько различны. Из поверхностных культур труднее получить высокоочищенные препараты из-за большого количества балластных веществ. Из глубинных культур получить очищенные препараты несколько легче, но при этом приходится вести выделение из разбавленных растворов, если выделение ферментов проводится из жидкой части культуры. Выделение осложняется, если фермент внутриклеточный, и тогда необходимо разрушать клетки микроорганизмов. Из схемы ясно, что экстракт из поверхностной культуры или фильтрат культуральной жидкости является исходным материалом для получения препаратов ферментов различной степени очистки. На первом этапе выделения отходом процесса является нерастворимая часть культуры биошрот, содержащий нерастворимые включения среды и биомассу продуцента. Далее в зависимости от свойств выделяемого фермента и сопутствующего ему балласта схема очистки и получения ферментного препарата может включать различные приемы и методы, такие как концентрирование, диализ, осаждение органическими растворителями, солями, гель-фильтрование, афинная хроматография, иммобилизация, сушка термолабильных материалов и т. д. Поэтому первоначально рассмотрим получение технических ферментных препаратов, а затем без особого акцента на поверхностный или глубинный способ культивирования более сложные способы очистки и выделения ферментов. 177 Таблица 4. Принципиальная схема получения очищенных ферментных препаратов из культур микроорганизмов Стадия очистки Объем, Общее Глюкозамилазная активность Амилолитическая Трансглюк мл количество активность о-зидазная белка, мг общая, удельная, выход, степень общая, ед выход, % активност ь* ед ед/мг % очистк белка и Исходная культуральная 1200 13600 28500 2,1 100,0 1,0 9500 100 Глюкоза, жидкость изомальто за, паноза Отделение биомассы, концентрирование, отделение балласта 560 11100 25600 2,3 90,0 1,1 8500 89,00 То же Осаждение ацетоном, растворение в воде 350 2040 19800 9,7 69,5 4,6 1050 11,30 » Ультрафильтрация 55 1610 18200 11,3 64,0 5,4 860 9,10 » Хроматография на ДЭАЭ-целлюлозе 555 298 15000 50,4 52,5 245 30,0 0,35 Нет Ультрафильтрация 16 250 13450 54,0 47,2 25,7 27,5 0,29 » Гель-фильтрование через акрилекс П-100 100 140 11400 76,5 40,5 36,5 22,8 0,24 » Обессоливание, лиофилизация 0,1** 92 7150 77,0 25,0 37,0 14,3 0,15 » * Характеризуется хроматографически по наличию изосахаров. ** Выражено в г. Получение неочищенных ферментных препаратов Неочищенные ферментные препараты представляют собой культуру микроорганизма вместе с остатками питательной среды, высушенную при мягком режиме до влажности не более 8-12 %. Неочищенный ферментный препарат может быть получен на основе поверхностной или глубинной культуры. Глубинная культура может быть перед сушкой очищена от нерастворимой части (твердая взвесь среды и биомассы продуцента) или высушена вместе с ней. Получение сухой поверхностной культуры. Поверхностная культура микроорганизма имеет влажность от 35 до 58 %. Это низкостабильный продукт, который следует либо немедленно использовать в производстве, либо высушивать до равновесной влажности (10-12 %). Перед высушиванием культура, выгруженная из растильной камеры, измельчается до определенной величины частиц и далее поступает на высушивание. Для сушки культуры микроорганизма могут быть использованы ленточные, тоннельные, шахтные, барабанные, шкафные и вибрационные сушилки. Сравнительно часто для сушки культур микроорганизмов используют барабанные сушилки прямоточного типа. Влажная культура поступает в сушилку одновременно с теплоносителем, имеющим температуру 80-85°С. Такую высокую температуру допустимо применять потому, что высушиваемый материал содержит большое количество влаги, а при ее испарении частицы культуры почти не нагреваются, и активность ферментов сохраняется почти полностью. У большинства барабанных сушилок на внутренней поверхности есть насадка в виде лопаток или крестовины. Барабан вращается медленно с частотой от 3 до 8 мин -1. Высушиваемый материал с помощью лопаток поднимается, пересыпается и передвигается вдоль барабана. Происходит некоторая дифференциация частиц культуры по размерам - более крупные частицы падают почти вертикально вниз и вновь подхватываются лопатками, а мелкие подхватываются теплоносителем, траектория их падения несколько смещается, и они быстрее перемещаются по барабану. Поэтому высушенный в такой сушилке продукт имеет равномерную влажность по всей массе. Длительность пребывания высушиваемой частицы в сушилке 3-7 мин, скорость движения подаваемого теплоносителя 2-3 м/с, температура воздуха на входе 80-85°С, на выходе 60-65°С, температура высушенного материала 40°С. Потери активности в процессе сушки составляют 3-10% . Другой вид сушилок, используемых для сушки культур, - это паровые конвейерные сушилки, представляющие собой герметизированный ленточный конвейер. При сушке в таких установках потери активности больше, но они очень компактные и имеют большую производительность. Но все же необходимо отметить, что потери могут доходить до 10-20%. Это связано с тем, что нет дифференцирования частиц культуры по размерам, мелкие и крупные частицы находятся в сушилке одинаковое время, мелкие частицы сильно пересыхают, ферменты инактивируются быстрее, крупные 179 частицы немного не досушиваются, в них потери активности ниже. Но они могут возрастать из-за повышенной влажности при хранении. Для высушивания поверхностной культуры можно использовать самые различные конструкции сушилок, в которых длительность пребывания культуры сокращена до 5-8 мин при температуре продукта на выходе не выше 40-42°С, что позволяет свести до минимума потери активности. Готовую сухую культуру обычно упаковывают на специальной фасовочной машине по 25-40 кг, водонепроницаемые мешки зашивают на зашивочной машине и отправляют на склад готовой продукции. Очистка культуральной жидкости от твердых взвесей. Большинство продуцентов накапливает основную часть синтезируемых ими ферментов в питательной среде. При получении очищенных ферментных препаратов нерастворимую часть среды вместе с биомассой продуцента отделяют на фильтрах, центрифугах или сепараторах. Наиболее широко в микробиологической промышленности используют ячейковый барабанный вакуум-фильтр непрерывного действия с наружной поверхностью фильтрования. Эти фильтры имеют высокую степень механизации и позволяют осуществлять фильтрование различных суспензий с постоянной скоростью. Барабанные вакуум-фильтры представляют собой барабан, погруженный в емкость, в которую непрерывно подается культуральная жидкость. Поверхность барабана перфорирована и обтянута фильтрующей тканью (батист или другая синтетическая ткань аналогичного типа). Иногда при наличии в культуральной жидкости трудно отделяемых осадков с высокими удельными сопротивлениями в качестве фильтрующей поверхности используют намывной слой. Съем осадка на этих фильтрах производится специальным ножом. При каждом обороте барабана вместе с осадком удаляется часть намывного слоя и фильтрующая поверхность обновляется. Барабан вращается медленно с частотой 0,13-0,26 мин-1 и проходит последовательно зоны фильтрования, подсушивания, промывания осадка, подсушивания и отдувки. Барабан разделен на секции с помощью неподвижной распределительной головки, состоящей из нескольких камер, которые соединены соответственно с вакуум-приемниками фильтрата и промывных вод, а также с линией сжатого воздуха. Барабанные фильтры удобны для отделения не только биомассы продуцента, но и нерастворимых взвесей, которых сравнительно много в среде (выжимки, отруби, жмых, ростки и т. д.). К недостаткам фильтров этого типа можно отнести их сравнительно низкую производительность, громоздкость (отношение удельной поверхности фильтрования к объему фильтрата небольшая) и невозможность обеспечения асептических условий. В ферментной промышленности реже используют рамные фильтрпрессы периодического действия с ручной выгрузкой осадка. С их помощью можно получать прозрачные фильтраты, но эти фильтры работают периодически без регенерации фильтрующей поверхности. Так как шламовое 180 пространство ограничено, а слой осадка к концу фильтрования достигает значительной толщины, то скорость фильтрования падает, несмотря на повышение рабочего давления. При заполнении шламового пространства осадком фильтр-пресс отключают, разбирают и промывают или меняют фильтрующее полотно. Производительность фильтр-пресса много меньше, чем барабанного вакуум-фильтра, она лимитируется содержанием осадка в фильтруемой жидкости и объемом рамного пространства фильтр-пресса. Процесс фильтрования в рамном фильтре ведется под давлением 0,6-0,4 МПа. Культуральная жидкость через отверстия в стенке рамы поступает во внутреннюю полость фильтрующего элемента, взвесь задерживается на фильтрующих поверхностях, а фильтрат проходит через фильтрующую салфетку и стекает по канавкам в плитах в трубопровод. Обычно первые порции фильтрата бывают мутные и их повторно фильтруют. Недостатки фильтр-пресса в значительной степени устранены в конструкции с горизонтальными камерами типа ФПАКМ. Он состоит из ряда расположенных одна над другой горизонтальных фильтровальных плит, между которыми натянута фильтровальная ткань. Фильтровальные плиты размещены между верхней и нижней поддерживающими плитами, а фильтровальная ткань натянута на направляющие ролики. Цикл работы фильтр-пресса состоит из сжатия плит, фильтрования, промывания и обезвоживания осадка, раздвигания плит и разгрузки осадка одновременно с перемещением ткани и ее промыванием. Работа фильтр-пресса ФПАКМ полностью автоматизирована. Эти фильтры имеют развитую фильтрующую поверхность (на 8 м2 площади, занимаемой установкой, приходится до 25 м2 фильтрующей поверхности); осадок отжимается под давлением 0,8-1,5 МПа и имеет влажность не более 60-70%; сравнительно небольшие энергозатраты (0,8-1 кВт-ч на 1 м2 фильтрующей поверхности); удельная производительность его в 6-8 раз выше, чем у других фильтр-прессов (при концентрации твердой фазы в суспензии 4-7 г/л до 1000 л/(м2 • ч), потери активности не превышают 4-5%). Установки ФПАКМ выпускаются с площадью фильтрующей поверхности от 2,5 до 50 м2. Их применение для очистки ферментных растворов очень перспективно; особенно этот тип фильтров рекомендуется для фильтрования взвеси культуральной жидкости бактерий. К фильтрам, работающим под давлением, относятся различные конструкции листовых фильтров. Общим для фильтров этого типа является наличие плоских фильтровальных элементов с жестким каркасом. Осадок с фильтрующей поверхности может удаляться различными способами: сжатым воздухом, паром, вибрацией, под действием центробежной силы. Некоторые бактериальные культуры даже при использовании вспомогательных фильтрующих материалов фильтруются со скоростью ниже 30 л/(м2 • ч). В этих случаях для отделения биомассы и удаления взвеси широко применяются сепарирующие центробежные машины. В ферментной промышленности применяются сепараторы-кларификаторы типа ВСМ, 181 представляющие собой емкость, внутри которой располагается барабан. Внутри барабана находятся концентрические цилиндры-вставки. Очищаемый раствор по патрубку поступает в цилиндр с наименьшим радиусом, затем проходит вдоль установленных цилиндров, каждый раз меняя направление. Осветленная жидкость удаляется из барабана с помощью напорного диска под давлением, а осадок под действием центробежной силы отбрасывается к внутренней стенке цилиндрических вставок. Производительность сепараторов этого вида может достигать до 2000-5000 л/ч. При производстве ферментных препаратов используются различные типы и конструкции саморазгружающихся сепараторов. В нашей стране применяют сепараторы типов АСЭ-3, АСИ, АСЭ-Б с центробежной пульсирующей выгрузкой осадка, имеющие соответственно производительность 500, 1500 и 2000 л/ч, при диаметре барабана 600 мм и межтарелочном зазоре 0,5 мм. На предприятиях очень большой производительности используются так называемые сопловые сепараторы фирмы «Альфа-Лаваль» (Швеция) типов QX и FEUX производительностью от 80 до 200 м3/ч. Есть той же фирмы «Альфа-Лаваль» фактически обеспложивающие сопловые высокоскоростные сепараторы так называемые бактофуги типа D3187M (производительность 6 м3/ч) и АХ-213 (производительность до 36 м3/ч). Фактор разделения, например, на бактофуге АХ-213 равен 142 000, что позволяет получать фугат, почти полностью очищенный от микроорганизмов и тончайшей взвеси, что очень важно. Бактофуга закреплена на станине. На ней расположен вал с фрикционной муфтой и тормозом, червячная передача, вертикальный полый шпиндель ротора с питающим насосом. На полый вал насажен барабан с набором конических тарелок. Верхняя часть станины вместе с колпаком и устройством для отвода удаляемой жидкости заключена в охлаждающую рубашку, что обеспечивает низкие температуры при сепарировании - это чрезвычайно важно, т. к. повышение температуры при отделении осадка может привести к инактивации ферментов. Исходная суспензия попадает в ротор снизу через полый вал и под действием центробежной силы распределяется по тарелкам. Твердые частицы направляются к стенкам ротора и непрерывно выгружаются через сопла с небольшим количеством жидкости. Основная часть жидкости удаляется из бактофуги под давлением через верхний параксиальный выпуск. Удаление чистого фильтрата происходит непрерывно. Микробные клетки и другие взвешенные частицы с небольшим количеством жидкости собираются под крышкой над ротором и поступают вниз вдоль сборной крышки по впускной трубе в циклон, где деаэрируются. Загрязненный воздух направляется в верхнюю часть крышки ротора, где вновь смешивается с микробными клетками, выходящими через сопла, образуется замкнутый цикл. Концентрат микробных клеток, отделенный от воздуха, удаляется через нижний патрубок циклона. 182 Этот вид сепаратора обладает большими преимуществами: герметичность процесса, непрерывность загрузки суспензии и отбора фильтрата, очистка воздуха от продуцента или фермента, ведение процесса при низких температурах и т. д. Можно смело утверждать, что этот тип сепараторов будет успешно использоваться в ферментной промышленности в XXI-м веке. Но надо помнить, что эффективность отделения биомассы во многом зависит не только от типов используемых аппаратов, но и от состава среды, размеров отделяемых частиц, количества нерастворимой фракции, физико-химических характеристик фильтрующих материалов, температурных режимов. Для улучшения процесса фильтрования проводят предварительную химическую обработку культуральной жидкости. Для этого культуральную жидкость подщелачивают до рН 8-8,5 и вводят 0,1% ный раствор хлористого кальция, в результате образуется гель фосфата кальция, который способствует наиболее полному отделению осадка при наименьших потерях. Но предварительная химическая обработка не всегда дает хорошие результаты, поэтому для повышения эффективности процесса часто используют различные кизельгуры, например, диатомит и радиолит (Япония), микрозил (Франция), диатомит (Бельгия), кларгель (Великобритания) и т. д. Использование этих наполнителей может резко повысить скорость фильтрования, но вместе с этим увеличиваются потери активности на этой технологической стадии. Полученную биомассу продуцента вместе с нерастворимыми частицами среды (биошрот) при необходимости стерилизуют, высушивают и используют на корм животным. Фильтрат культуральной жидкости нестабилен, он не может храниться и должен немедленно направляться на дальнейшую обработку для получения очищенных ферментных препаратов. Экстрагирование ферментов из поверхностных культур. Все ферменты являются водорастворимыми белками, поэтому наилучшим экстрагентом для них является вода. Для извлечения ферментов из дрожжей или бактерий необходимо подвергнуть механическому или автолитическому разрушению их клеточные стенки, обладающие высоким диффузионным сопротивлением. Оболочки мицелиальных нитей имеют меньшее диффузионное сопротивление, чем оболочки бактериальных и дрожжевых клеток, поэтому дезинтеграции культуры грибов не требуется. Извлечение ферментов проводят как из влажных, так и из сухих поверхностных культур грибов. Сухая культура может храниться длительное время без потери активности ферментов, и из нее получают более концентрированные экстракты. Технологически это выгоднее, но при подсушивании культуры имеют место потери активности, и потому экстрагирование целесообразно вести из влажной культуры. При экстрагировании различные водорастворимые вещества извлекаются из культуры с неодинаковой скоростью, происходит их частичное фракционирование, удельная активность ферментов в экстракте повышается 183 в 3,5-4 раза по сравнению с исходной культурой в результате отделения большой части веществ (до 75%) с нерастворимым остатком - биошротом. На полноту экстрагирования ферментов из культур оказывают влияние многие факторы: температура, рН, длительность процесса, конструктивные особенности экстракционных аппаратов, природа извлекаемого фермента, количество отобранного экстракта с единицы массы загруженной в аппарат культуры и т. д. Одновременно с ферментами экстрагируются многие другие соединения, и часто скорость извлечения балластных веществ больше скорости экстрагирования из культуры целевого фермента. Поэтому рациональнее пойти на некоторые потери фермента и закончить экстрагирование на оптимальном значении отношения активности фермента в экстракте к сумме извлекаемых веществ. Этот вопрос решается экспериментально для каждого вида продуцента. Влиять на процесс экстрагирования с помощью такого фактора, как температура, практически невозможно, так как ферменты очень термолабильны и инактивируются даже при 35-40 °С. Кроме того, повышение температуры до 35-40 °С влечет за собой увеличение содержания сухого вещества в экстракте и уменьшение удельной ферментативной активности на 1 г сухого вещества, повышение опасности инфицирования экстрактов. Поэтому при проведении экстракции в заводских условиях стремятся подавить развитие микрофлоры путем максимального снижения температуры воды до 22-25 °С и применения антисептиков (формалин, бензол, толуол, хлороформ и др.). В большинстве случаев ферменты наиболее полно извлекаются при рН 5-7. Для получения концентрированных экстрактов при небольших потерях ферментов с биошротом необходимо применять специальные экстракционные установки. До недавнего времени для этого широко использовались диффузионные батареи. В них можно получить экстракт с содержанием сухого вещества от 7 до 14% в зависимости от вида культуры, среды и величины отбора экстракта. Диффузор представляет собой цилиндро-коническую емкость, снабженную рубашкой. В центральную цилиндрическую часть аппарата помещают сухую или влажную поверхностную культуру и фиксируют ее двумя сетками снизу и сверху. Экстрагент подают в диффузор № 1 снизу, обычно это вода с температурой 22-28 °С, и заполняют диффузор полностью до уровня сливной трубы. Подача воды прекращается на определенное время (в зависимости от вида фермента это может быть от 30 до 60 минут), затем вновь включают подачу воды и с помощью свежей воды вытесняют экстрагент из диффузора № 1 в диффузор № 2 до его полного заполнения, и вновь вся система останавливается на определенное время. Таким образом все повторяется до тех пор, пока в последнем, восьмом диффузоре экстрагент простоит еще τ8, т. е. если τ = 30 мин, то τобщ будет 4 часа (0,5 ч х 8). После этого отбирают готовый экстракт. Непрерывность работы 184 батареи обеспечивается наличием в системе 10 диффузоров. После прохождения первого цикла экстракции через 8 диффузоров в диффузоре № 1 уже прошла 8-ми кратная экстракция, там в культуре практически нет ферментов, поэтому этот (№ 1) диффузор отключается для разгрузки, вода подается на диффузор № 2, а в конце процесса диффузор № 9 заполняется свежей культурой и съем экстракта через 30 минут настаивания производят с диффузора № 9. К этому времени диффузор № 1 разгружен и вымыт, диффузор № 10 загружен свежей культурой, диффузор № 2 отключен на разгрузку, вода подается на диффузор № 3 и т. д. Фактически диффузионная батарея работает в непрерывном режиме: каждый диффузор периодически; находится в головном, промежуточном и хвостовом состоянии. Но эти установки для экстрагирования ферментов из поверхностной; культуры имеют сравнительно небольшую производительность, требуют больших затрат ручного труда и в них наблюдаются сравнительно большие потери активности. Поэтому постоянно ведутся исследования и поиск наиболее совершенной конструкции по экстракции ферментов из поверхностных культур микроорганизмов в непрерывном режиме и с минимальной затратой ручного труда. На ферментных предприятиях нашей страны успешно испытывался и определенное время с хорошими показателями работал диффузионный аппарат конструкции С. М. Гребенюка. Экстрактор состоит из горизонтального корпуса со шнеком и вертикального корпуса со шнеком. Загрузка свежей культуры осуществляется через дозатор; экстрагент подается через течку и поступает сначала в вертикальный корпус, затем переходит в горизонтальный корпус вплоть до ситового пояса, из которого и происходит отбор экстракта из грибной поверхностной культуры, т. е. поток экстрагента находится в состоянии противотока с культурой микроорганизма. В ферментной промышленности используются экстракторы роторного типа фирмы «Роунс Дауне», состоящие из неподвижного корпуса, внутри которого находится ротор, разделенный на 16-20 секторных отсеков, вращающийся вокруг вертикальной оси. Каждый отсек имеет ситчатое дно, на которое подается измельченная культура гриба. Ротор медленно вращается и последовательно проходит четыре участка, в каждом из которых культура смачивается водой или экстрагентом из предыдущих отсеков, вытяжка отсасывается вакуум-насосом и подается в следующий отсек для увлажнения свежей культуры, вновь отбирается вытяжка и передается в 3-й отсек и т. д. При завершении одного оборота ротора биошрот разгружается, и отсеки вновь загружаются свежей культурой. В настоящее время наблюдается тенденция к более широкому использованию пресс-диффузии. Она заключается в том, что культура после настаивания с водой отпрессовывается, затем снова настаивается при меньшей концентрации ферментов в получаемом экстрагенте, вновь прессуется и т. д. Вероятно, после удачного аппаратурного решения данного 185 принципа он найдет широкое применение в ферментной промышленности. Концентрирование ферментных растворов Экстракты из поверхностных культур микроорганизмов и фильтраты глубинной культуры являются нестабильными при хранении. Для получения готовых форм технических препаратов (П2х и Г2х) их необходимо сконцентрировать. Чаще всего для этих целей в технологии ферментных препаратов используются методы вакуум-выпаривания. Вакуум-выпаривание также применяется как один из этапов получения сухих технических или очищенных ферментных препаратов. Ферменты очень чувствительны к температуре выпаривания, поэтому основным условием концентрирования ферментных растворов является кратковременное ведение процесса при низких температурах кипения, чтобы выпариваемая жидкость не нагревалась выше определенной критической для данного фермента температуры, т. е. чтобы не наблюдалось инактивации фермента. Также следует учитывать, что чем чище раствор, чем меньше он содержит сопутствующих веществ, тем ферменты более чувствительны к воздействию высоких температур. В концентрированных экстрактах из поверхностных культур инактивация ферментов значительно меньше, так как в экстракте содержится очень большое количество защитных соединений, которые препятствуют инактивации ферментов. Зависимость стабильности ферментов экстракта от температуры после концентрирования вакуум-выпариванием близка к тому, как ведут себя ферментные растворы со стабилизатором при различных температурах кипения. При концентрировании фильтратов культуральной жидкости наблюдаются несколько большие потери, поэтому ферменты культуральной жидкости стабилизируют различными соединениями. В процессе концентрирования ферментных растворов происходят изменение растворимости многих соединений и выпадение их осадков, и суммарное содержание сухого вещества в концентрате снижается на 11-20%, изменяется рН концентрата. В осадок выпадают минеральные соли, некоторые органические вещества и продукты их распада, наблюдается потеря азота в результате уноса аммиака. При концентрировании культуральной жидкости различных микроорганизмов происходит значительное изменение минерального состава получаемого концентрата, например, при концентрировании культуральной жидкости В. mesentericus это четко видно. Наиболее резко снижается содержание кальция, меди и магния, заметно уменьшается содержание цинка и марганца. Такое изменение минерального состава культуральной жидкости сказывается на стабильности ферментов в процессе концентрирования. При сгущении культуральной жидкости до содержания сухого вещества 10% количество кальция снижается всего на 5%, а меди - на 75%. Известно, например, что медь оказывает на ферменты ингибирующее действие, а кальций - стабилизирующее. Поэтому на первых 186 стадиях концентрирования наблюдается повышение активности ферментов, особенно протеиназ. При более глубоком концентрировании вместе с резким снижением содержания кальция снижается активность ферментов. Большинство ферментов очень чувствительно к термической обработке и нуждается в мягких режимах концентрирования. При последующих исследованиях было установлено, что не только температура кипения концентрируемого раствора имеет большое значение, но и температура греющего пара (теплоносителя). Так, даже при очень низкой температуре кипения (25-30°С), происходит заметная инактивация ферментов (до 12%), если температура греющего пара равна 120°С. При температуре теплоносителя 90-100°С и температуре кипения 35-40°С потери активности не превышают 10%. И еще следует отметить, что чем выше температура теплоносителя, тем больше сухих веществ концентрируемой жидкости выпадает в осадок, особенно при высоких температурах кипения. Важно заметить, что в зависимости от вида продуцента культуральная жидкость имеет различный химический состав и содержит различный комплекс ферментов, поэтому тепловые режимы вакуум-выпаривания уточняются экспериментальным путем в каждом конкретном случае. Суммарные потери активности при вакуум-выпаривании в значительной степени зависят не только от режима концентрирования, но и от конструкции аппарата. Аппараты для стадии вакуум-выпаривания в последние года значительно усовершенствованы, в десятки раз сокращена длительности процесса, что привело к значительному уменьшению потерь активности ферментов, а также позволило несколько ужесточить температурные режимы концентрирования ферментных растворов. Помимо трубчатых вакуум-выпарных установок с различным расположением трубок (горизонтальным) вертикальным и наклонным), со встроенной и выносной поверхностью нагрева, с использованием принудительной циркуляции созданы новые конструкции пленочных выпарных аппаратов, ультрацентробежных вакуум-выпарных установок и пластинчатых испарителей. Особый интерес представляют ротационные пленочные выпарные аппараты, где упариваемая жидкость в виде пленки движется по внутренней стенке аппарата. Лопатки, смонтированные на вращающемся роторе, непрерывно направляют движение ее сверху вниз. Время прохождения жидкости через аппарат составляет несколько секунд. В настоящее время фирма «Альфа-Лаваль» изготовляет вакуум-выпарные центробежные аппараты типа «Центритерм». Они очень компактны, время контакта ферментного раствора с обогревающей поверхностью предельно сокращено (не более 1 с), потери не превышают 10%, производительность этих установок от 800 до 4800 л/ч. Создана центробежная вакуум-выпарная установка пленочного типа производительностью 800 л/ч по испаренной влаге. Время контакта культуральной жидкости с теплоносителем не более 1 с, температура греющего пара 60-80°С. Для увеличения производительности можно 187 монтировать установку из трех модулей, каждый из которых работает либо автономно, либо последовательно, либо первые два модуля работают параллельно и соединены с третьим модулем последовательно. Представляет интерес для ферментной промышленности центробежная пленочного типа вакуум-выпарная установка «Единство» (Югославия) производительностью до 200 л/ч и с температурой упаривания 30-40 °С. Хорошие технологические показатели имеют роторные выпарные аппараты фирмы «Люва» (Швейцария), имеющие производительность по испаренной влаге от 50 до 200 л/(м2 • ч). Французская фирма APV изготовляет пластинчатые вакуумвыпарные установки производительностью до 20 000 л/ч. Несмотря на наличие высокопроизводительных вакуум-выпарных аппаратов полностью устранить недостатки метода вакуум-выпаривания не удается (потери активности, выпадение осадков и т. д.), и этот метод все больше заменяется методом ультрафильтрации. Мембранные методы очистки и концентрирования ферментов. В зависимости от движущей силы процесса мембранные методы классифицируются на диффузионные - диализ (движущая сила - разность концентраций по обе стороны мембраны), электромембранные электродиализ (разность электрических потенциалов), баромембранные обратный осмос, ультрафильтрация, микрофильтрация (разность давлений). Все эти процессы применяются для переработки ферментных растворов, выбор их определяется целью переработки: для очистки от низкомолекулярных примесей при небольших производительностях - диализ, для обессоливания в интенсивных условиях - электродиализ, для глубокой очистки от примесей с одновременным концентрированием ультрафильтрация и т. д. Рассмотрим подробнее сущность и технические решения каждой группы процессов, что (дет возможность в дальнейшем выбрать оптимальный вариант для любой конкретной задачи. Диализ - это первый изученный и промышленно развитый мембранный процесс, поскольку для его осуществления не нужна сложная аппаратура и специальные мембраны. Сущность диализа в том, что если два раствора с различной концентрацией какого-либо компонента разделить мембраной, то начнется естественный процесс диффузии, достигающий равновесия при выравнивании концентраций этого компонента с обеих сторон мембраны. Интенсивность переноса вещества QВ через мембрану определяется коэффициентом диффузии этого вещества в материале мембраны DB, и пропорциональна разности концентраций ΔС. Эту зависимость можно записать в виде сравнения: QВ = DB • S • ΔС, где S - площадь мембраны. Соответственно, чем больше различие в величинах коэффициентов диффузии двух компонентов, находящихся в растворе, тем лучше они разделяются мембраной. Понятно, что белковые молекулы (высокомолекулярные вещества) и органические и неорганические низкомолекулярные молекулы и ионы сопутствующих компонентов (сахара, 188 аминокислоты, минеральные соли и т. п.) в силу огромных различий в коэффициентах диффузии практически полностью разделяются мембраной. В качестве диализных мембран используют обычно пленки из целлюлозы - целлофан, купрофан, а также из других синтетических полимеров. Процесс проводят либо по проточной схеме, когда исходный раствор ферментов постоянно прокачивают с одной стороны мембраны, а диализирующую жидкость (обычно воду) - с другой ее стороны, либо по полупроточной схеме, когда раствор ферментов помещают на определенное время в мешочки из диализной мембраны, которые постоянно омываются водой. Таким образом можно удалить основную массу сопутствующих низкомолекулярных примесей и повысить активность ферментных растворов в пересчете на сухое вещество в несколько раз. Процесс диализа применительно к очистке растворов ферментов имеет ряд существенных недостатков. Во-первых, при диализе возможна «потеря» фермента в результате вымывания ионов металлов, входящих в состав молекулы фермента, или стабилизирующих фермент соединений, или фрагментов самого фермента, например, простетической группы его. Вовторых, при диализе против обычной водопроводной воды может происходить потеря активности фермента в результате попадания из воды в раствор фермента ионов металлов - ингибиторов фермента. Следует также отметить, что в процессе диализа одновременно с очисткой происходит сильное разбавление ферментного раствора из-за проникновения воды под действием сил прямого осмоса в диализуемый раствор. Объем продиализованного раствора увеличивается примерно на 20-25%, а если учесть, что происходит активное удаление балластных веществ, то в результате диализа получают очень разбавленные ферментные растворы. Поэтому сейчас этот метод очистки ферментных растворов от балластных веществ в ферментной промышленности почти не используется. Этот метод иногда применяют в лабораторных исследованиях и при получении высокоочищенных ферментных препаратов. Электродиализ. Если в процессе очистки ферментов стоит задача удалить из раствора электролитные примеси, т. е. органические и минеральные ионы, иногда пользуются электродиализом. Сущность этого мембранного метода в том, что перенос ионов через мембрану интенсифицируют с помощью постоянного электрического поля, а мембраны изготавливают из специальных ионо-обменных материалов на основе синтетических полимеров. Электрический потенциал к аппарату подводится через два электрода, размещенных в соответствующих электродных камерах. Обе камеры отделены от рабочей обессоливающей камеры, куда подается исходный pacтвор, ионообменными мембранами, со стороны катода - анионообменной, со стороны анода - катионообменной. При работе аппарата катионы под действием постоянного электрического поля смещаются к аноду, встречают 189 на пути катионообменную мембрану, проходят через нее в электродную камеру и в виде слабого раствора щелочи выводятся из аппарата. Соответственно ведут себя и анионы, выходя из аппарата в виде слабого раствора кислоты. Обессоленный раствор ферментов (диализованный раствор) выводится из рабочей камеры. Электродиализный метод осуществляется всегда в непрерывном режиме и существенно более энергоемок, чем диализ. Применительно к обработке ферментных растворов он имеет те же недостатки. Кроме того, электродиализ нельзя применять при выделении ферментов, имеющих, например, четвертичную структуру, которая формируется с участием ионов металлов, а также при выделении металлоферментов, которые, как правило, теряют активность при электродиализе (α-амилазы, β-галактозидазы и др.). Часто баромембранные методы разделения жидких смесей относят к процессу обычной фильтрации, но они лишь внешне похожи благодаря тому, что движущей силой является разность давлений. В действительности с помощью полупроницаемых мембран разделяются истинные растворы, т. е. гомогенные системы, в то время как фильтрованием можно разделить лишь суспензии, т. е. твердую фазу отделить от жидкой. Вместе с тем, считая мембранные методы фильтрованием на молекулярном уровне, можно построить условный спектр фильтрации, разместив мембранные методы - обратный осмос, нанофильтрацию, ультрафильтрацию и микрофильтрацию - в некоторый ряд и дополнив его обычной механической фильтрацией по порядку увеличения размера и молекулярной массы задерживаемых частиц. Сегодня баромембранные методы получили широкое распространение в биотехнологической, пищевой, фармацевтической, химической промышленности. В частности, ни одно современное производство ферментов не может уже обойтись без ультрафильтрационной очистки и концентрирования продукта. Растворение вещества в растворителе возможно только тогда, когда они имеют сродство друг к другу, т.е. когда на уровне межмолекулярного взаимодействия происходит сольватация молекулами растворителя молекул или ионов растворяемого вещества. Когда речь идет о водных растворах, процесс называется гидратация. Поскольку молекула воды представляет собой крохотный диполь, ее энергия связи с частицей растворимого вещества тем больше, чем больший заряд несет эта частица на себе. Понятно, что чем больше заряд иона, тем больше количество молекул воды окажется связанным с ионом в виде многослойной гидратной оболочки. Именно образованием гидратных оболочек объясняется явление, которое называется прямой осмос. Если раствор любого вещества отделить полупроницаемой мембраной от объема чистого растворителя, то мы будем наблюдать односторонний перенос молекул растворителя (в данном случае воды) в раствор, где они достраивают гидратные оболочки. Чем выше 190 концентрация растворенного вещества слева, тем больше молекул воды должно пройти через мембрану в раствор. Количественно этот перенос выражается величиной осмотического давления (РО): РО = C R Е , где: С - массовая концентрация растворенного вещества; R - газовая постоянная; Т - абсолютная температура. Если осмотическое давление (РО) больше гидравлического (Рr), то происходит прямой осмос, если РО = Рr, то диффузия через мембрану прекращается. Если же теперь к раствору приложить рабочее давление, превышающее осмотическое, т. е. Рr > РО то начнется перенос молекул воды слева направо, т. е. будет происходить дегидратация раствора, концентрирование растворенного вещества и получение чистой воды в правой половине сосуда. Это механизм называется обратным осмосом. Обратный осмос по механизму близок к ультрафильтрации. Ультрафильтрация год от года все шире используется в технологии ферментных препаратов. Весьма убедительны данные, приводимые фирмой «Амикон корпорейшн» (США) о преимуществах очистки и концентрирования методом ультрафильтрации (см. табл. 5). Таблица 5 Метод разделения Центрифугирование Гель-фильтрация Сушка барабанная распылительная лиофильная Осаждение этиловым спиртом или солями Ультрафильтрация Затраты на Концентрация сухого вещества, удаление 1 м3 % воды, доллары исходная в концентрате 1-2 10-15 0,15-0,9 3-5 Разбавленная 6-30 30 100 7,5 10-20 100 15 10 100 60-90 1-2 Различная 1500 1-10 10-50 0,15-0,30 Действительно, из таблицы следует, что ультрафильтрация обладает способностью не инактивировать ферменты и требует минимальные энергозатраты. Скорость ультрафильтрации будет тем выше, чем больше разница между рабочим гидравлическом (Рr) давлением и осмотическим. Однако между процессами обратного осмоса и ультрафильтрации все же есть различия. Так, при обратном осмосе разделение низкомолекулярных веществ происходит при рабочем давлении до 0,7-14 МПа, так как 191 осмотическое давление РО в этих растворах велико. При обратном осмосе используются мембраны с очень маленькими порами (от 1 • 10~4 до 2 • 10~3 мкм). При ультрафильтрации происходит разделение высоко- и низкомолекулярных соединений, и целью этого процесса является получение концентрата высокомолекулярных соединений (например, ферментов). Рабочее давление в этом случае низкое (от 0,07 до 0,7 МПа), так как РО небольшое. Величина пор мембран значительно больше - от 3 • 10~3 до 150 • 10~3 мкм. Однако названные различия достаточно условны. Механизм процессов обратного осмоса и ультрафильтрации пока остается недостаточно ясным. Для математического описания процесса мембранного разделения служит модель движения вязкого потока через поры (уравнение Пуазейля) и модель диффузионного массопереноса (закон Фика). Принято считать, что если размер пор мембраны меньше 3 • 10~3 мкм (обратный осмос), то процесс подчиняется закону Фика, если же размер пор больше 3 • 10~3 мкм (ультрафильтрация), то процесс подчиняется уравнению Пуазейля. В этом и заключается принцип любого баромембранного процесса. Отличия между ними лишь в размерах пор используемой мембраны и в величинах приложенного к раствору давления. Поскольку осмотическое давление белковых растворов мало, для осуществления процесса достаточно 0,3-0,6 МПа, а размер пор мембраны составляет 10-50 нм. На основе этих знаний каждый может теперь дополнить диаграмму на рис. 1.55 новыми системами. 14. 5.Применение ферментов микроорганизмов Использование ферментов микроорганизмов в различных областях народного хозяйства весьма перспективно. В настоящее время ферментные препараты, полученные из микроорганизмов, применяются в различных областях промышленности, сельского хозяйства и медицины. Применение ферментов в пищевой промышленности. В пивоварении и винокурении для замены солода используют препараты грибных амилаз. Это удешевляет производство и сокращает расход зерна. Амилазы используются также для получения растворимого крахмала, декстрина и патоки. Продукты из овощей и фруктов, полученные с применением амилаз, содержат больше сахара и лучше усваиваются, особенно детьми. В хлебопечении амилазы ускоряют процесс созревания теста и улучшают качество хлеба. В кондитерской промышленности используется инвертаза (сахараза) дрожжей, превращающая сахарозу в глюкозу и фуктозу, она предупреждает кристаллизацию сахарозы при высоких концентрациях. Комплекс ферментов (цитаз) грибов, расщепляющих вещества стенок растительных клеток, используют для улучшения экстракции их содержимого (сока, эфирных масел, жиров, крахмала). 192 Пектиназы грибов применяют для осветления фруктовых и ягодных соков, для повышения выхода виноградного сока в виноделии, при производстве кофе. Применение пектиназ особенно эффективно при производстве сока из плодов и ягод, содержащих много пектина (черная смородина, крыжовник, слива). В СССР в промышленных масштабах выпускаются пектолитические препараты «Авоморин ППК», «Пектавоморин П10x». Грибная глюкоамилаза применяется в пивоваренной промышленности для удаления остатков декстринов из пива. Глюкозоизомераза используется для получения глюкозо-фруктозных сиропов, заменяющих сахарозу, что важно для улучшения рационов питания, поскольку использование в большом количестве сахарозы вредно для человека. Лактаза применяется для получения молока без лактозы. После такой обработки оно приобретает лучшие вкусовые качества. Кроме того, некоторая часть населения не может употреблять молоко из-за наличия в нем лактозы, которая вызывает аллергическую реакцию. С помощью лактазы получают также сахара (глюкозу, галактозу) из молочной сыворотки, содержащей большое количество лактозы. Большое значение имеет глюкозооксидаза грибов, так как она позволяет пищевым продуктам освобождаться от остатков глюкозы и молекулярного кислорода и этим повышает сроки их хранений. Глюкозооксидазу добавляют к яичному порошку, к майонезу, к пиву при его длительном хранении. С помощью этого фермента замедляется окисление аскорбиновой кислоты при обработке им овощей и фруктов. Применение ферментов облегчает получение глюконовой кислоты. Каталазу Asp. niger применяют в пищевой промышленности для удаления остатков пероксида водорода – стерилизующего агента при получении пищевых концентратов, стерильного молока, меланжа. Препараты целлюлазы используют для осахаривания картофельной мезги, выделения крахмала из картофеля и зерна, увеличения выхода агарагара из водорослей, для приготовления овощной пасты, удаления кожуры у цитрусовых. Используют их и для получения редуцирующих сахаров из растительных материалов. Такой способ производства сахаров может быть дешевле, чем при использовании в качестве исходного субстрата крахмала. Протеолитические ферменты микробного происхождения заменяют ренин в сыроделии для получения сгустка. Начинают их использовать для размягчения (тендеризации) мяса, ускорения созревания рыбы при посоле, в виноделии и пивоварении. Липазы находят применение в производстве цельного сухого молока, в сыроделии для ускорения созревания сыров и придания им специфического вкуса и аромата. Применение ферментов в текстильной промышленности В текстильной промышленности пектолитические ферменты микроорганизмов давно и широко применяются для переработки льносоломы 193 и получения из нее волокна. В настоящее время используется тепловая мочка льна на льнозаводах. Основными микроорганизмами, участвующими в процессе мочки, признаны анаэробы рода Clostridium. Процессы, протекающие во время мочки, приводят к разрушению пектиновых веществ льносоломы и высвобождением льноволокна. Для ускорения процесса (обычно идущего 2,5-3 сут.) и повышения качества волокна был получен ферментный препарат пектоклостридин ГЗх отфильтрованная и высушенная культуральная жидкость Clostridium sp. Препарат позволяет ускорить процесс мочки в 2-2,5 раза и получить волокно с высокой механической прочностью. Амилолитические препараты используются для удаления клея из тканей (расшлихтовка). Некоторые протеиназы, в частности протосубтилин, используются для обесклеивания шелка (удаления серицина) и высвобождения шелковых волокон, состоящих из фиброина. Для освобождения шелкового волокна от жира применяют препараты липаз. Другие области промышленного применения ферментов В кожевенной промышленности микробные протеиназы используют для обезволашивания шкур и мягчения кожи. Применение комплексного ферментного препарата, состоящего из протеазы и липазы, ускоряет процесс и позволяет получить высококачественную шерсть. Использование микробных ферментов при производстве моющих средств приобретает все большее распространение. Обычно в них добавляют ферменты Вас. subtilis обладающие протеолитической, амилолитической и липолитической активностями; препараты используются в комбинации с поверхностно-активными веществами. Моющие средства, содержащие ферменты, сокращают продолжительность стирки, повышают сохранность тканей, так как обработка ведется при 40-60°С (не выше). Применение ферментов в сельском хозяйстве Применение ферментов в сельском хозяйстве развивается в двух направлениях: 1) использование в рационах животных, 2) обработка кормов ферментами для повышения их усвояемости. При поверхностном способе культивирования Asp. oryzae у нас в стране получают препарат амилоризин - высушенная культура гриба, содержащая α-амилазу, декстриназу, мальтазу, глюкоамилазу и протеиназу. Препарат глюкаваморин - высушенная культура Asp. аwamory, выращенного на отрубях, содержит α-амилазу, декстриназу, мальтазу, глюкоамилазу, кислую протеиназу и гемицеллюлазу. Препарат амилосубтилин, содержит αамилазу протеазы, β-глююконазу и литические ферменты. Эти ферменты входят и в состав протосублитина ГЗх и ксилаваморина ГЗх, содержащих также гемицеллюлазу и пектиназу. Применение ферментов в медицине Микробные ферменты используются в различных областях медицины как терапевтические средства и при проведении клинических анализов. При лечении воспалительных процессов и ожогов применяются препараты 194 протеиназ, разрушающие некротизированные ткани и клетки, способствуя быстрому заживлению ран. При терапии злокачественных новообразований используют бактериальную L-аспарагиназу, превращающую L-аспарагин, необходимый лейкозным клеткам, в L-аспарагиновую кислоту, в результате чего рост опухоли значительно замедляется. Тромболитическими свойствами обладают протеиназы террилитин и стрептокиназа, имеющие микробное происхождение. В заместительной терапии, т. е. при нарушении синтеза некоторых ферментов в организме человека, применяют отдельные ферменты и комплексные ферментные препараты. Например, при нарушении функции поджелудочной железы употребляют комплексный препарат, содержащий протеиназу, амилазу и липазу. При потере способности к синтезу лактазы и глюкоамилазы также используют эти ферменты, полученные из микроорганизмов. При нарушении процессов пищеварения в некоторых случаях используют комплекс ферментов (α-амилаза, целлюлаза, липаза и протеиназа). Использование микробных ферментов в медицине весьма перспективно и, несомненно, будет расширяться. Трудности в применении ферментных препаратов для целей медицины состоят в необходимости очистки их от пирогенных веществ, токсинов и других примесей. Принимая во внимание белковую природу ферментов, необходимо проверять их на антигенное действие и аллергическую реакцию организма. Использование ферментов при проведении химических анализов. Применение ферментов в качестве химических реактивов составляет особую область аналитической и препаративной биохимии. В качестве примеров можно привести определение кокарбоксисилазы в крови при помощи протеиназы Asp. niger, расщепляющей ее с выделением тиамина. По количеству свободного тиамина рассчитывают количество фермента. Определение галактозы производят, используя галактозооксидазу гриба Polysporus ciracinatus. Фермент окисляет галактозу, переводя ее в соответствующий лактон с одновременным образованием пероксида водорода. При добавлении пероксидазы и орто-дианизидина появляется синяя окраска колириметрируемого раствора. Определение оротовой кислоты можно проводить с соответствующей дегидрогеназой оротовой кислоты из Clostridium оrоticum. Фермент катализирует восстановление оротовой кислоты НАДН2 с образованием дигидрооротата и НАД. Бактериальная гиалуронидаза расщепляет гиалуроновую кислоту количественно с образованием ненасыщенного дисахарида. При этом образуются N-ацетилглюкозаминовые группы, дающие при нагревании в 195 щелочи с пара-диметиламинобензальдегидом красную окраску (важно для колориметрирования). Бактериальные и грибные протеиназы используются для расщепления белков, причем расщепление может быть фрагментарным. Например, субтилизин при коротком воздействии на рибонуклеазу расщепляет только одну пептидную связь этого белка. Карбоксидопептидаза дрожжей постепенно расщепляет белки, отделяя одну за другой аминокислоты с карбоксильного или аминного конца. Определение L-аминокислот (L-лизин, L-аргинин, L-орнитин, Lтирозин и др.) можно проводить с помощью специфических декарбоксилаз аминокислот. Ферменты синтезируются некоторыми бактериями (например, Cl. welchii, Cl. septica) и эффективны при низком значении рН. Образовавшаяся при реакции углекислота может быть измерена количественно. Определение глутамина и аспарагина проводится путем их дезаминирования специфическими дезаминадами Pseudomonas sp. Освобождающийся аммиак определяют по цветной реакции с фенолятом в присутствии нитропруссида. Весьма перспективны такие ферменты микроорганизмов, как холестериноксидаза для определения холестерина в крови, алкогольоксидаза для обнаружения спирта и др. Ферменты используют также для изучения первичной структуры высокомолекулярных соединений: нуклеиновых кислот, белков, полисахаридов. Использование ферментов в органическом синтезе Перспективная область применения ферментов микроорганизмов использование их для синтеза различных практически важных соединений. Выше отмечалось, что с помощью пенициллинамидазы, образуемой некоторыми бактериями, можно получить 6-аминопенициллановую кислоту субстрат для синтеза полусинтетических пенициллинов. L-Аспарагиновую кислоту получают с помощью аспартазы Е. coli. При этом используют как препараты фермента, так и клетки с увеличенной проницаемостью в иммобилизованном состоянии. Аналогичным образом могут быть синтезированы и некоторые другие практически важные L-аминокислоты, в частности тирозин, 3,4диоксифенилаланин. Использование ацилаз аминокислот позволяет разделить их L- и Dизомеры. В Японии создано производство L-яблочной кислоты из фумаровой на основе использования бактериальной фумаразы. Эта кислота применяется как заменитель лимонной кислоты в пищевой и фармацевтической промышленности. Ряд реакций трансформации, ведущих к получению стероидных препаратов, основан практически на ферментативной активности не растущих клеток микроорганизмов. 196 С помощью ферментов микроорганизмов возможно и проведение многостадийных процессов. Например, иммобилизованные клетки дрожжей способны синтезировать глутатион из глюкозы и фосфата, что требует последовательного действия нескольких ферментов. Как уже отмечалось выше, особая область применения ферментов микроорганизмов - их использование в генетической инженерии. Даже эти немногочисленные примеры позволяют представить, что возможности использования ферментов микроорганизмов для получения практически важных соединений, а также для других целей весьма разнообразны. Очевидно, что в недалеком будущем количество и число ферментов микробного происхождения, выпускаемых промышленностью, будет существенно расширено. Производство ферментных препаратов занимает одно из ведущих мест в современной биотехнологии и относится к отраслям, объем продукции которых постоянно растет, а сфера применения неуклонно расширяется. Такое быстрое развитие связано с тем, что ферменты являются высокоактивными, нетоксичными биокатализаторами белкового происхождения, которые широко распространены в природе, без них невозможно осуществление многих биохимических процессов и жизнь в целом. Познание роли ферментов для всего живого на Земле послужило основой для становления и развития технологии ферментных препаратов как науки и для создания промышленного производства наиболее широко используемых ферментных препаратов. Применение этих препаратов помогло существенно изменить, интенсифицировать и усовершенствовать многие существующие технологии или даже создать принципиально новые высокоэффективные процессы. Применение ферментных препаратов различной степени очистки позволило не только улучшить показатели и выходы в различных биотехнологических процессах, но позволило усовершенствовать кормопроизводство, повысить усвояемость кормов, сделать более целенаправленным и эффективным действие синтетических моющих средств, улучшить качество косметических препаратов, создать целый арсенал специфических, чувствительных и точных аналитических методов, наладить производство лекарственных и профилактических средств для медицинской промышленности и т. д. Большим и неоспоримым достоинством ферментов перед химическими катализаторами является то, что они действуют при нормальном давлении, при температурах от 20 до 70 °С, рН в диапазоне от 4 до 9 и имеют в большинстве случаев исключительно высокую субстратную специфичность, что позволяет в сложной смеси биополимеров направленно воздействовать только на определенные соединения. Все это свидетельствует о том, что производство ферментных препаратов является одним из перспективных направлений в биотехнологии, которое будет и далее интенсивно развиваться и расширяться. 197 ГЛАВА 15. Получение пищквых добавок. Пищевые добавки — вещества, добавляемые в продукты питания для придания им желаемых свойств, например определѐнного аромата (ароматизаторы), цвета (красители), длительности хранения (консерванты), вкуса, консистенции. Подавляющее количество продукции, потребляемой горожанами, содержат в себе пищевые добавки, которые, в свою очередь обозначаются кодом с буквой «Е». Практически любой продукт, предлагаемый нам в супермаркетах – будь то сухарики, морожение, то ли это сок, то ли полуфабрикаты, или даже самая обычная буханка хлеба – на каждой упаковке практически всегда есть «E». Можно ли питаться такой пищей – решать нужно только самому покупателю, который естественно не проинформирован о возможной вредности определенной пищевой добавки, содержащейся в данном продукте. Но перед тем как купить что-либо в супермаркете нужно обладать специальной информацией, которая выведет Вас из дебрей закодированных обозначений «Е». Пищевые добавки - являются неотъемлемой частью практически всех продуктов быстрого приготовления и многих других видов не скоропортящихся и долго хранящихся продуктов. Благодаря пищевым добавкам можно намного улучшить вкус продукта, а так же продлить его срок годности. Но помимо всех положительных качеств пищевых добавок, они в основном вредны для нашего здоровья. Человек, в среднем, в течении одного года употребляет вместе с пищей около 5 килограммов различных пищевых добавок. Во многих странах некоторые пищевые добавки вовсе запрещены и являются так называемыми запрещѐнными пищевыми добавками. Запрещенные в Беларуси: Е103, Е106, Е111, Е121, Е123, Е126, Е130, Е181, Е216, Е217, Е240, Е924, Е924а. Существует мнение, будто бы пищевые добавки несут с собой только вред человеческому организму. Но не стоит забывать, что благодаря многим пищевым добавкам, пищевые продукты остаются свежими и полезными для употребления человеку. В некоторых случаях они просто незаменимы. Пищевые добавки, по некоторым расчетам, имеют около пятидесяти различных функций. Из которых можно выделить одиннадцать групп наиболее известных нам добавок. К этим добавкам относятся; питательные добавки или природные компоненты пищи, добавки сохраняющие свежесть продукта, добавки с помощью которых облегчается переработка или изготовление продуктов питания, а как же консерванты такие как приправы, красители, уплотнители или текстуранты, подсластители, наполнители, добавки благодаря которым можно снизить калорийность пищи, и другие. Питательные добавки одни из наиболее полезных человеческому организму. С их непомерной помощью в некоторых странах удалось полностью 198 ликвидировать болезни вызванные недостатком в какого либо вещества в пище. Например - зоб развивается благодаря недостатку иода в организме, цинга развивается благодаря недостатку витамина C, рахит в связи с нехваткой витамина D, кальция, фосфора. В развитых странах в пищевые продукты для улучшения их питательной ценности, добавляют такие микроэлементы пищи как жиры, углеводы, белки, а так же клетчатку. Все это делается для того что бы улучшить питательную ценность употребляемой пищи. Е 100 —Е 182 —красители; Е 200 — Е 299 — консерванты; Е 300 —Е 399 — антиокислители (антиоксиданты); Е 400 —Е 449 — стабилизаторы консистенции; Е 450 — Е 499 — эмульгаторы; Е 500 — Е 599 — регуляторы кислотности, разрыхлители; Е 600 — Е 699 — усилители вкуса и аромата; Е 700 — Е 800 — запасные индексы для другой возможной информации; Е 900 и далее — антифламинги, улучшители качества хлеба и т.д. Эти вещества делятся на натуральные и синтетические. Натуральные производятся только из естественного сырья – трав, специй, фруктов, овощей, мяса, птицы, дрожжей, древесной коры, грибков и даже из насекомых-вредителей. Однако различие между натуральными и синтетическими добавками довольно условно, так как касается не столько состава, сколько способа их производства. Натуральные добавки необязательно безопаснее искусственных. Часто они содержат больше химических примесей. Натуральные пищевые красители. Относительно дешевые синтетические красители нашли очень широкое применение в пищевой промышленности. Однако среди них практически нет абсолютно безвредных. Более того, многие импортные красители потенциально опасны как канцерогены и аллергены. К тому же они не всегда удачно имитируют цвет натурального продукта. Именно поэтому в последнее время все больше внимания уделяется природным окрашивающим веществам. Натуральные (природные) пищевые красители – это красящие вещества, выделенные физическими способами из растительных и животных источников. Иногда их подвергают химической модификации для улучшения технологических и потребительских свойств. Ряд красителей получают не только их выделением из природного сырья, но и синтетически. Сырьѐм для натуральных пищевых красителей могут быть ягоды, цветы, листья, корнеплоды и т д., в том числе, в виде отходов переработки растительного сырья на консервных и винодельческих заводах. Технология производства натуральных красителей постоянно совершенствуется, что позволяет увеличивать их стойкость и оптимизировать цену. Натуральные красители 199 обычно выделяют из природных источников в виде смесей различных по своей природе соединений, состав которых зависит от сырья и технологии получения, поэтому обеспечение постоянства их качества — очень сложная задача. Оттенки и стойкость одних и тех же натуральных пигментов в красителях разных производителей могут варьировать. Технологические свойства натуральных пигментов могут быть откорректированы и улучшены при помощи натуральных антиоксидантов и технологий суспендирования, эмульгирования и микрокапсулирования, применение которых позволяет значительно расширить сферу применения натуральных красителей. Производство натуральных красителей— одна из сложнейших областей пищевой промышленности, где достичь высокого уровня качества может лишь производитель, обладающий глубокой научной и высокоразвитой производственной базой. Содержание красящих веществ в растительном сырье зависит от климатических условий произрастания и времени сбора, но, в любом случае, оно относительно невелико (обычно, несколько процентов или доли процента). Количество других химических соединений – сахаристых, пектиновых, белковых веществ, органических кислот, минеральных солей и т.д. – может превышать содержание красящих в несколько раз. Эти вещества не представляют опасности для здоровья, а часто даже полезны для человека, но своим присутствием они снижают интенсивность окрашивания готового продукта. При производстве препаратов натуральных пищевых красителей от побочных веществ, в той или иной степени, избавляются. Современные технологии позволяют получать препараты натуральных пищевых красителей с заданными свойствами и стандартным содержанием основного красящего вещества. Потребители интуитивно связывают внешний вид и качество продукта в единое целое. Именно поэтому пищевые красители стали важнейшими ингредиентами, от которых зависит «товарный вид». Натуральные красители могут быть и водо-, и жирорастворимыми, что расширяет сферу их применения. Более того, многие природные красящие вещества являются диетологическими средствами. Существует большое количество природных красителей таких, как, например, красный сандал, кверцитрон, кармин, сепия, кампешевое дерево и т.д. Кампешевое дерево растет в Южной Америке, сандал - в Южной Азии, сепию добывают из каракатиц, кармин - из кошенили (крошечных насекомых). Растения главным образом применяются в высушенном виде. Сушка производится не на солнце и не в печи, а в тени, на открытом воздухе, на растянутых веревках или полотне. Красящие вещества, содержащиеся в растениях, очень редко находятся в чистом виде, и для получения их пользуются следующими приемами и способами: прежде всего происходит измельчение сырых материалов, состоящее в резке, крошке или толчении их; степень измельчения зависит от свойств и качеств самого материала, но во всяком случае оно должно быть равномерно и как можно мельче. 200 Измельченный материал подвергается выщелачиванию, состоящему в кипячении в дождевой, речной или перегнанной воде, причем для более полного извлечения красок к воде необходимо прибавить немного поташа или соды. По мере выкипания воды, прибавляют свежей и так поступают до полного выщелачивания материала, т.е. пока вода больше уже не будет окрашиваться. После этого отвар кипятят для выпаривания, т.е. для получения более густой краски. Дают отстояться и фильтруют через сито, на которое положен кусочек полотна. Коричневый краситель. Если отварить в воде сухую кожуру репчатого лука, то получается коричневый краситель разных оттенков - от почти желтого до темно-коричневого. Другой источник такого красителя - сухая кора жостера. Кстати, некоторые прменяют отвар кожуры репчатого лука для окраски пасхальных яиц (яйца варят в отваре). Колер карамельный (Е 150 a-d) получают путем термической обработки различных видов сахаристых веществ. Он имеет коричневую окраску, хорошо растворим в водной фазе, не чувствителен к воздействию света, температуры. За последние годы развитие рынка мороженого, йогуртов, творожных и кисломолочных продуктов привело к появлению множества новых оригинальных видов продукции. Варьировать оттенки конечной продукции можно также изменением дозировки красителя, либо использованием двух или нескольких красителей одновременно. Например, для получения лаймового цвета рекомендуется одновременно использовать меднохлорофиллиновый краситель и куркумин. Жирорастворимые натуральные красители адресованы в первую очередь производителям шоколадных глазурей, шоколада. Они имеют в своем составе жировую составляющую, поэтому их внесение в продукцию на жировой основе гораздо технологичнее, нежели порошков синтетических лаков (не требуются высокоскоростные миксеры и т.п.). Применение натуральных жирорастворимых красителей дает возможность производителю использовать натуральное сырье, получая широкий спектр оттенков готовой продукции. Итак, современное производство немыслимо без красителей. Но, в соответствии с тенденциями развития рынка, сейчас необходимы именно природные компоненты. Это подтверждают и маркетинговые исследования. Правда, объемы производства синтетических красителей пока в 3,5 раза выше, чем натуральных. Но зато темпы роста производства натуральных выше, чем синтетических. Красный краситель. Красный краситель можно получить из стебля зверобоя (отвар надо подкислить) или из корня подмаренника. Ольховую кору положите в воду на несколько дней, а потом приготовьте отвар. Красный краситель можно извлечь из корней конского щавеля, но в этом случае не забудьте прибавить к готовому отвару немного алюминиевых квасцов - иначе цвет будет тусклым. Красные оттенки 201 представлены различными антоцианинами Е 163, карминами Е 120, а также свекольным соком. Оттенки антоцианинов могут варьироваться от красных до красно-фиолетовых. Источниками их получения могут служить кожица винограда, черная морковь, ягоды бузины и т.д. Красители водорастворимы, свето- и термоустойчивы, оттенок зависит от рН. Красные красители флавоноидного типа (в частности, антоциановые), получают элюированием 50% уксусной кислотой или экстрагированием соляной кислотой и этилацетатом. Кармин получают экстракцией из кошенили - высушенных и растертых женских особей насекомых вида Coccus Sactic, обитающих на кактусах в Южной Америке, Африке. Пигмент кармин обладает хорошей устойчивостью к воздействию света и высокой температуры. Кармины могут быть представлены и водорастворимыми, и жиродисперсными формами, а также оттенками от красного до красно-фиолетового. Источником красителя свекольного сока (Е162) является свекла Beta vulgaris. Основные пигменты, содержащиеся в соке, - бетанин и валгаксантин. Свекольный сок придает продуктам окраску от розового до красно-синего оттенков. Главным недостатком свекольного сока является слабая термоустойчивость. Однако для молочной промышленности это не является ключевым моментом. Краситель может быть представлен либо в форме водорастворимой жидкости, либо порошка. Синий краситель. Из корней девясила (относится к лекарственным травам) можно получить синий краситель. Для этого корни надо сначала подержать в нашатырном спирте - водном растворе аммиака. Синий краситель можно добыть также из цветов живокости и корней птичьей гречишки. Зеленый краситель. Хлорофилл (Е140). Натуральный краситель, который изготавливается из зеленой массы растений (крапива, шпинат), придает продукту оливковые оттенки. Медные комплексы хлорофиллов, хлорофиллинов (Е141) применяют для придания продуктам яркозеленых оттенков. Краситель зеленого цвета извлекают из листьев трилистника (лекарственная трава). Не такой яркий, но тем не менее красивый серозеленый краситель можно получить из листьев и стеблей манжетки; перед приготовлением отвара их необходимо тщательно измельчить. Желтый краситель. Желтый краситель дают многие растения: дрок красильный, орешник (кора), ольховидная крушина (кора, листья, ягоды), подмаренник (цветы). Из плодов барбариса получается желтый краситель с лимонным оттенком. Фиолетовый краситель. Ягоды черники и ежевики, как нетрудно догадаться, содержат фиолетовый краситель. Он не очень стоек, но вполне может пригодиться для акварельных красок. Оранжевый краситель. Из стеблей и листьев чистотела удается извлечь краситель оранжевого цвета. Каротиноиды получают путем добавления к овощам эфирно-фцетоновой смеси и уксусной кислоты, а затем диэтилового эфира. 202 Черный краситель. Во-первых, для этого можно приготовить отвар из ягод и корней воронца. Но есть и другой способ, более простой: добавить железный купорос к одному из полученных ранее отваров. Почти все наши отвары содержат дубильные вещества типа танина, и в присутствии солей двухвалентного железа они становятся черными. Мясная и молочная промышленность. Естественный неброский цвет вареного мяса, к сожалению, не особо привлекает большинство потребителей продукции мясной индустрии. Поэтому в мясной промышленности широко применяются ингредиенты для восстановления коррекции цвета мясных продуктов. Мясная промышленность потребляет 13% (в денежном выражении, по состоянию на 2004 год) производимых во всем мире натуральных красителей. Идеальный пищевой краситель для мясной промышленности должен быть устойчивым, безвредным при рекомендуемых дозировках и условиях применения, не оказывать неблагоприятного влияния на свойства пищевого продукта, не реагировать с основными и сопутствующими компонентами окрашиваемого изделия, обладать высокой красящей способностью, быть простым в применении, а также иметь приемлемую цену аннато куркумин кошениль — источник кармина. Аннато. В мясной промышленности очень широко используется натуральный желто-оранжевый краситель аннато (E160b), относящийся к группе каротиноидов. Аннатто получают из семян орлеанового дерева Bixa orellana. Его родиной является тропическая Америка. Это дерево издавна культивируют в тропических странах для получения семян оранжевой окраски, из которых добывают натуральный краситель оранжевого цвета. Аннатто имеет в своем составе смесь каротиноидов, главным из которых является биксин. Биксин - жирорастворим, а его производное норбиксин -водорастворимый пигмент. Аннатто обладает хорошей термо- и светоустойчивостью. Для использования в продуктах с кислым рН разработаны специальные кислотоустойчивые формы красителя. Водорастворимая форма аннато (пигмент норбиксин) имеет способность образовывать прочные комплексы с белками. Благодаря этой способности аннато хорошо подходит для окраски оболочек и поверхностей копченных продуктов. Для получения рыжего или яркого красно-коричневого оттенка следует внести 50—100 г красителя на 10 л воды (дозировка указана для красителя с концентрацией пигмента около 1%). Подготовленную оболочку необходимо погрузить в приготовленный раствор на 2—3 ч и более в зависимости от желаемой интенсивности окрашивания. Краситель растворяют в холодной мягкой воде, и мясные изделия погружают в раствор на 2—5 минут до получения необходимой окраски. Для окрашивания копченостей этот краситель также иногда комбинируют с натуральным коричневым красителем карамель, причем из четырех ее видов в мясной промышленности используют только Е150с и Е150d. Наиболее подходящим для сочетания с аннато типом карамели и по оттенку, и с технологической 203 точки зрения является Е150с. Если необходимо придать копченостям несколько более яркий оранжевый оттенок, к аннато прибавляют немного кармина. Куркумин. Для получения золотистожелтого оттенка аннато иногда смешивают с натуральным красителем куркумином (турмериком) (Е100). Натуральный пигмент куркумин позволяет окрашивать пищевые продукты в желтый цвет. Куркумин (турмерик) (Е100) получают из корневища пряного растения куркумы Curcuma longa L. семейства имбирных, произрастающего в Индии. Куркумин жирорастворим по своей природе и диспергируется в воде при добавлении эмульгаторов. Он также применяется для коррекции оттенков смесей специй, приправ, для имитации желтизны куриного жира. Куркумин является сильным антиоксидантом, улучшает пищеварение, способствует нормализации кишечной микрофлоры, связывает свободные радикалы, помогает в борьбе с возрастными болезнями, предотвращает рак, является антибактериальным агентом, способствует детоксикации печени, предотвращает формирование катаракты и желчных камней, стимулируетработу желчного пузыря. Паприка. Одним из наиболее широко применяемых в пищевой промышленности натуральных красителей является экстракт паприки (Е160с). Паприка - краситель, полученный экстракцией из красного сладкого перца Capsicum annuum, произрастающего в Европе и Северной Америке. Паприка содержит каротиноидные пигменты капсантин, β-каротин и капсорубин. В зависимости от содержания пигмента краситель может быть от красного до оранжевого цвета. Он улучшает пищеварение, обладает антисептическими свойствами. Пигменты паприки жирорастворимы по своей природе, но на их основе получают также диспергируемые в воде красители. В зависимости от условий производства и состава вододисперсной паприки этот краситель может по-разному вести себя при смешивании с водой. Наиболее дешевые формы вододисперсной паприки диспергируются в воде при тщательном перемешивании; красите ли, легко смешиваемые с водой, имеют несколько более высокую цену. Наиболее востребован этот краситель в мясной отрасли и широко применяется при производстве смесей специй, приправ. Паприка используется практически во всех сферах пищевой промышленности. Хорошо очищенный краситель при соблюдении рекомендуемой дозировки не придает продуктам никаких ощутимых посторонних привкусов и запахов. Олеорезины паприки не проявляют заметной чувствительности к рН среды, однако без специальной защиты пигменты паприки не очень устойчивы к воздействию света и при высокотемпературной обработке. Для увеличения свето- и термостойкости паприки применяются натуральные антиоксиданты (токоферолы, экстракт розмарина и др.) В мясных полуфабрикатах водо- и жирорастворимые формы красителя (с содержанием 1,5% красящего пигмента) обычно применяются в количестве от 0,05 до 0,1%. Кармин. Во всем мире для придания мясным изделиям естественного 204 красного цвета наиболее часто применяется натуральный краситель животного происхождения — кармин (Е120), представляющий собой комплексное соединение карминовой кислоты с кальцием и алюминием. На основе этого пигмента могут производиться водо- и жирорастворимые красители, кроме того, он используется в виде лака — нерастворимого в воде и жирах комплексного соединения с кальцием и алюминием. Карминовый лак имеет более светлый оттенок красного и легко смешивается с продуктами на водной и жировой основе. Такая форма этого красителя имеет низкую миграционную способность. Кармин зарекомендовал себя как один из самых устойчивых пищевых красителей: придавая колбасным и деликатесным изделиям естественный сочный оттенок, он не проявляет заметной чувствительности к свету, окислению и температурной обработке. Кармин используется как при производстве сырокопченых и сыровяленых продуктов, так и при выработке колбасных изделий, подвергаемых термической обработке. Кармин в водорастворимой форме — единственный натуральный краситель, применяемый при инъецировании вареной ветчины и для окраски оболочек в разные оттенки красного цвета (часто в сочетании с аннато). Этот краситель позволяет получать стабильный цвет при производстве мясопродуктов, регулировать степень окрашивания продуктов в соответствии с их типом и пожеланиями потребителя, а также улучшать их товарный вид. Лютеин (Е161) получают экстрагированием из лепестков бархатцев Aztec Marigolds, эти цветы произрастают в Южной Америке. Краситель представляет собой вододисперсный экстракт, растворимый в жирах и масле. Он придает продукту различные оттенки желтого цвета от лимонного до золотистого. Лютеин менее чувствителен к окислению, чем другие красители каротиноидного типа, стабилен к нагреванию, светоустойчив. Консерванты. Наиболее используемыми консервантами являются: поваренная соль, этиловый спирт, уксусная, сернистая, сорбиновая, бензойная кислоты и некоторые их соли. Консерванты можно условно разделить на собственно консерванты и вещества, обладающие консервирующим действием. Действие первых направленно непосредственно на клетки микроорганизмов, вторые отрицательно влияют на микробы, в основном, за счет снижения pH среды, активности воды или концентрации кислорода. Соответственно, каждый консервант имеет свой спектр действия. Накопление консерванта в продукте может происходить не только при его внесении извне, но и в связи с химическими изменениями, происходящими в сырье в результате деятельности микроорганизмов. Квашение капусты, соление огурцов и других овощей основано на молочнокислом брожении сахара, в результате которого в продукте накапливается молочная кислота, являющаяся природным консервантом. По мере роста населения консерванты приобретают все большее значение, поскольку обеспечить 10%-й прирост запасов пищевых продуктов 205 с их помощью гораздо легче, нежели путем расширения сельскохозяйственного производства. Низин (Е 234) - антибиотик естественного происхождения, продукт жизнедеятельности молочнокислых бактерий. Он активно подавляет рост термоустойчивых граммположительных спорообразующих бактерий, предотвращает образование ботулиновых токсинов. Применяется в производстве сыров и других молочных продуктов, овощных, мясных и рыбных консервов, а также в виноделии, пивоварении и хлебопечении. Молочная кислота (Е 270) образуется в процессе молочнокислого брожения и, благодаря низкому значению рН, широко используется в качестве консерванта при изготовлении сыра и целого ряда других молочных продуктов. Муравьиная кислота и ее производные (Е 236-238) используются, в основном, при консервировании овощей и производстве безалкогольных напитков. Антиоксиданты. Добавки, сохраняющие свежесть, включают в первую очередь антиоксиданты. Их добавляют к маслам и к упаковочным материалам, чтобы предотвратить прогоркание. Используют также хелатирующие агенты и секвестранты. Они предотвращают взаимодействие между металлами и компонентами пищи, что сводит к минимуму обесцвечивание, а также утрату вкуса и аромата. Ряд веществ используется для того, чтобы предотвратить потемнение фруктов на поверхности разреза. Как и консервирующие вещества, антиоксиданты применяются для увеличения сроков хранения пищевых, главным образом жироемких, продуктов. В основе их действия лежит ингибирова-ние реакций окисления пищевых компонентов. Окисление происходит под влиянием кислорода воздуха, света, температуры, технологических факторов производства. В первую очередь окисляются липиды и их соединения, витамины и другие биологически важные нутриенты, что снижает пищевую ценность продукта. Конечные продукты окисления отрицательно влияют на органолептические свойства и могут быть токсичны для организма человека. Например, окисление липидных компонентов приводит к образованию гидропероксидов, которые, также окисляясь, дают такие токсичные соединения, как альдегиды, кетоны, отдельные жирные кислоты и многочисленные продукты их полимеризации. Натуральные антиокислители. К ним относят токоферолы (витамин Е), аскорбиновую кислоту (витамин С), флавоны (кверцетин), эфиры галловой кислоты, гваяковую кислоту и т.д. Пищевые стабилизаторы. Это особая группа добавок, применяемых в разных отраслях пищевой промышленности, главным назначением которых является формирование и сохранение консистенции, текстур, форм и потребительских качеств продуктов молочного, мясоперерабатывающего, хлебопекарного и кондитерского производств. 206 Обычно выделяют три главные группы пищевых стабилизаторов: пектины, каррагинаны и камеди. Все они являются производными натуральных веществ, хотя в последнее время объемы мирового производства продуктов питания потребовали и промышленного синтеза некоторых видов пищевых стабилизаторов. Пищевые стабилизаторы не представляют опасности для здоровья и являются очень важным подспорьем для наращивания мирового производства продуктов питания. Сырьем для них служат яблоки, плоды цитрусовых, пшеница, кукуруза, морские водоросли, смолы различных наземных растений и т.п. Отдельные виды стабилизаторов являются продуктами микробиологической промышленности. Пектин (Е440) — это натуральное желеобразующее вещество, содержащееся во фруктах и многих видах овощей. Пектин обычно получают в результате экстракции из цитрусовых или яблок. Особенность пектина как студнеобразователя — способность формировать гели в водных растворах только в присутствии определенного количества сахара и кислоты или ионов кальция. Применяются пектины в кондитерском, молочноперерабатывающем, мясоперерабатывающем производстве. К другой группе относятся камеди трех видов: гуаровая (Е412), ксантана (Е 415) и камедь рожкового дерева (Е 410). Камеди являются загустителями, стабилизаторами, гелеобразователями, средством для капсулирования. Широко используются в производстве плавленых сыров, мороженого и молочных продуктов, фруктовых и овощных консервов, сырокопченых колбас, соусов, кетчупов, майонезов, хлебобулочных изделий, рыбных консервов, низкожирных маргаринов и спредов. Камеди также применяются в связке с другими загустителями и гелеобразователями для регулировки процесса. Так например, гуаровая камедь применяется для производства сыра в сочетании с каррагинаном. Каррагинан (Е 407) – природный загуститель, получаемый при переработке красных морских водорослей класс Rhodophyceae. Этот класс водорослей произрастает практически по всей акватории Земли, на подводных скалах на глубине до трех метров. Каррагинаны также широко применяются в вышеперечисленных областях пищевой промышленности. Эмульгаторы. Вещества, способные образовывать и стабилизировать эмульсию. Они обеспечивают возможность создания и сохранения дисперсии двух или более несмешивающихся веществ. Впервые в качестве эмульгаторов стали использовать камеди, сапонины, лецитин и другие натуральные вещества. В настоящее время список эмульгаторов расширился главным образом за счет синтезированных препаратов. Эмульгирующая способность рассматриваемой группы веществ связана с их поверхностно-активными свойствами, поэтому термин «эмульгатор» можно рассматривать как синоним терминов «эмульгирующий агент» и «поверхностно-активное вещество» (ПАВ). Основная область применения эмульгаторов и стабилизаторов — 207 масложировая промышленность. Так, для приготовления жиров, используемых в хлебопечении и кондитерском производстве, разрешены эмульгаторы: Т-1 — моно- и диглицериды жирных кислот; Т-2 — продукт этерификации полиглицерина насыщенными жирными кислотами (С16 и С18). Их добавляют в количестве не более 2000 мг на 1 кг продукта. Для этих соединений ДСД составляет 125 мг на 1 кг массы тела человека. Наряду с основной функцией эмульгаторы используют для равномерного распределения в воде жирорастворимых веществ и соединений: ароматизаторов, эфирных масел, экстрактов пряностей и т.д. Усилители вкуса. Свежие овощи, мясо, рыба и другие продукты имеют яркий вкус и аромат за счет содержания в них нуклеотидов. В процессе хранения и промышленной переработки количество нуклеотидов уменьшается, что сопровождается потерей вкуса и аромата продукта. Глутамат натрия (Е 621 или натриевая соль глутаминовой аминокислоты) – пищевая добавка, предназначенная для усиления вкусовых ощущений за счет увеличения чувствительности рецепторов языка. Глутамат натрия не содержит питательных веществ, вместо этого он обманывает ваш мозг, в результате чего вы думаете что еда, которую вы едите, очень вкусная. Пряности. Во времена, когда не было постоянных торговых путей, обеспечивающих ввоз пряностей, пища европейцев была крайне однообразной и неудовлетворительной не только в количественном, но и в качественном отношении. В наше время в ходу свыше 2000 различных пряностей для любого мыслимого употребления. Природные пряности имеют очень сложный состав; в кофе, например, содержится свыше 1000 различных соединений (впрочем, обычно пряности далеко не столь сложны). Большинство пряностей, которыми мы пользуемся теперь, – это смеси, составленные из синтетических веществ. Текстуранты. Так называют различные добавки, предназначенные для улучшения текстуры пищевых продуктов. Соединения кальция делают консервированные томаты более плотными и крепкими. Фосфаты улучшают вкус консервированных груш, делая их более нежными. Пирофосфаты улучшают текстуру пудингов быстрого приготовления и молочных продуктов. Эмульгаторы придают стабильность водным и масляным эмульсиям в заправках для салата. Разнообразное применение находят вещества типа крахмала, придающие продуктам большую плотность. Разрыхлители обеспечивают соответствующую текстуру выпекаемых хлебобулочных и кондитерских изделий. Подсластители. Природные подсластители, такие, как сахар, известны людям на протяжении тысячелетий. Их всегда добывали в больших количествах. Однако забота о снижении калорийности пищи вынудила обратиться к 208 непищевым подсластителям. В США в настоящее время разрешены к применению пять таких веществ: сахарин, аспартам, ацесульфам, тауматин и глициризин. Рассматривается и возможность применения ряда веществ, разрешенных в других странах. Аспартам и ацесульфам приблизительно в 200 раз слаще сахарозы, и ведутся работы по созданию новых, более эффективных искусственных подсластителей. Наполнители. Эта тенденция к применению непищевых подсластителей заставила искать вещества, которые могли бы выполнять роль, традиционно выполняемую сахарами в напитках, джемах, желе и копченостях. Желатинированный крахмал люди употребляют на протяжении веков, теперь же получен ряд производных крахмала и целлюлозы. Используется полидекстроза – также одно из производных сахара. Способ получения пищевых добавок из растительного сырья, содержащих витамины, минеральные соли и растительные волокна и может найти широкое применение в пищевой и медицинской промышленности. Он заключается в следующем: подготовленное растительное сырье предварительно измельчают, затем его сушат в определенных параметрах до влажности не более 10%, после чего полученную массу охлаждают хладагентом до достижения температуры во всей массе от -80 до -165oС, с последующим ее измельчением до размеров частиц 50 - 600 мкм и готовый продукт затем упаковывают. Перед упаковкой продукт может быть таблетирован. Это позволяет получить биологически активную добавку из традиционно используемого пищевого сырья, обладающую высокой биологической активностью за счет наиболее полного сохранения биологически активных веществ и повышенной их усвояемости организмом. Товарная экспертиза пищевых добавок проводится на стадии изготовления и на всех этапах их товародвижения. Одним из таких этапов являются создание и анализ технологии подбора и внесения пищевой добавки или их комплекса в продукт с учетом особенностей химического состава, функциональных свойств пищевых добавок и сырья, характера действия, вида продукта, технологии, в отдельных случаях — упаковки и хранения. ГЛАВА 16. Получение лигнина Обычно присутствие лигнина в растительном мире связывается с наличием опорных и проводящих тканей (например, ксилемы). Лигнин является важным компонентом клеточных стенок этих тканей. Так, лигнин найден в таких растениях, как плауны, папоротники, голосеменные и покрытосеменные, но его нет в несосудистых растениях, таких, как грибы и водоросли. Мхи составляют исключение в том отношении, что, не имея клеток, подобных ксилеме, они, тем не менее, содержат лигниноподобные 209 соединения; правда, имеются некоторые сомнения в том, что это настоящие лигнины. Лигнин содержится не только в клеточных стенках проводящих тканей, но и в клеточных стенках сердцевины, корней, плодов, почек, коры и пробки. Лигнификация клеточной стенки происходит после отложения полисахаридных компонентов стенки и к концу ростового периода клетки. Распределение лигнина в стенке происходит неравномерно; обычно те слои, которые откладывались первыми, бывают наиболее богаты лигнином. Следовательно, срединная пластинка и первичная стенка подвергается наибольшей лигнификации, а вторичная – наименьшей. Лигнификация имеет двоякое значение. Она укрепляет клеточную стенку, образуя разветвлѐнную сеть по всему матриксу, закрепляя, таким образом, более прочно микрофибриллы целлюлозы. Кроме того, лигнин предохраняет микрофибриллы стенки от химических, физических, биологических воздействий. Важное свойство лигнина – устойчивость по отношению к микроорганизмам. Лишь немногие из них, и то сравнительно медленно, разрушают лигнин. Термин лигнин определяет группу близкородственных, высокомолекулярных полимеров, главным, если не единственным, строительным блоком которых является фенилпропановый остаток. Структурных вариантов фенилпропанового остатка известно мало, но существует много путей, по которым они соединяются вместе. Порядок в структуре полимера отсутствует. Процесс полимеризации происходит путѐм хаотичной конденсации свободных радикалов. По этой причине структура каждой молекулы лигнина может быть уникальной, и, следовательно, структуру лигнина записать невозможно. Однако можно нарисовать гипотетическую частичную структуру, которая отражает различные типы фенилпропановых строительных блоков и различные пути связывания их вместе. Лигнины однодольных и двудольных покрытосеменных и голосеменных растений структурно различаются между собой. В основе этого различия лежит неодинаковая структура фенилпропановых строительных блоков. Об этом свидетельствуют различные ароматические альдегиды, получающиеся при мягком окислении нитробензолом в щелочной среде лигнинов из указанных источников. Лигнин голосеменных растений даѐт преимущественно ванилин и немного n-гидроксибензальдегида. Лигнин двудольных растений даѐт главным образом ванилин и сиреневый альдегид и также немного n-гидроксибензальдегида; лигнин однодольных растений даѐт все три альдегида. Накапливается всѐ больше данных в пользу того, что лигнин ковалентно связан с полисахаридами матрикса клеточной стенки, однако природа этой связи пока не установлена. 210 Лигнин исключительно трудно экстрагировать из клеточной стенки. До сравнительно недавнего времени все методы экстрагирования включали использование очень агрессивных химических реагентов при повышенных температурах, например 40% HCl, смесь 3:1 36% HCl и 80% H3PO4, 2% H2SO4, а затем аммиачный CuO . Эти методики приводили к значительной модификации экстрагируемого лигнина. Более того, природа модификации зависела от используемого для экстракции реагента. Дополнительное осложнение вносила самоконденсация молекул лигнина, происходившая при повышенных температурах. Такие модифицированные лигнины обычно называли по имени использованного экстрагирующего реагента, например солянокислый лигнин, купроксамовый лигнин. Указанные трудности были в значительной мере преодолены, когда Браунс показал, что около 5% общего количества лигнина древесины можно экстрагировать, обрабатывая свежие опилки 95%-ным этанолом в течение нескольких дней при комнатной температуре. Такое сочетание нейтрального, относительно инертного растворителя и низкой температуры не вызывало заметных структурных изменений и поэтому позволяло получать немодифицированный лигнин. Экстрагированный таким образом лигнин называют лигнин Браунса или нативный лигнин. Выход нативного лигнина можно увеличить, если для экстрагирования по методу Браунса использовать древесину, поражѐнную бурой гнилью, вызываемой, например, грибом Poria vaillantii. Подобные грибы выделяют смесь ферментов, расщепляющих целлюлозу и другие полисахариды стенки, но не атакующих лигнина, который в результате становится более доступным экстрагирующему растворителю. Бурой гнилью эти грибы называются потому, что поражѐнная область древесины имеет бурый цвет. Это отличает их от грибов, вызывающих белую гниль, которые расщепляют все компоненты древесины, включая лигнин, и оставляют белые изгнившее области. Утилизация лигноцеллюлозной биомассы основана на процессе еѐ конверсии в сбраживаемые субстраты, которые в свою очередь служат сырьѐм для многих отраслей промышленности, производящих химические вещества, горючее и продукты питания. При этом необходимо эффективно использовать все компоненты биомассы: целлюлозу, гемицеллюлозу и лигнин. Лигнин защищает полиглюкан от действия ферментов, так что для успешного использования полимерной целлюлозы необходимо, прежде всего, удалить гемицеллюлозы, далее разрушить комплекс лигнина с целлюлозой, а затем – и кристаллическую структуру самой целлюлозы. Эти задачи могут быть решены с применением разнообразных химических, физических и микробиологических методов. Гемицеллюлозы легко удаляются путѐм растворения в слабой кислоте в ходе первичной обработки биомассы. При производстве глюкозы из древесины довольно успешно применяется гидролиз слабым раствором кислоты при высоких давлении и температуре. Этот способ нашѐл применение в ряде стран. Гораздо эффективнее гидролиз концентрированной 211 кислотой, но для осуществления этого процесса требуются большие капиталовложения. Особенно успешно применяется плавиковая кислота, причѐм безводную кислоту можно регенерировать. Лигноцеллюлозу можно также размалывать и затем подвергать радиационному облучению в больших дозах. Получаемый такими способами лигнин различается по своим свойствам. Очищенные лигнины используются при выработке клеев, смол, адгезивов, типографической краски, для диспергирования красителей; они применяются как адсорбенты, изоляторы, используются при добыче нефти, в асфальтовых смесях и при нанесении полимеров. После удаления лигнина экстракцией растворителями получают кристаллическую целлюлозу, которая идѐт на нужды бумажной промышленности. Рентабельность этого процесса определяется главным образом стоимостью регенерации растворителя. Примером одностадийного процесса, протекающего в мягких условиях, является экстракция фенолом при 100˚С, когда лигнин и гемицеллюлозы растворяются в феноле, а высвободившуюся после фильтрования целлюлозу используют для выделки бумажной массы. По мере остывания фенольного экстракта гемицеллюлоза концентрируется главным образом в водной фазе, отделяясь от масел и лигнина, которые можно использовать для производства новых порций фенола методом гидрокрекинга. Таким образом, в отношении фенола и энергетических потребностей – это самообеспечивающийся процесс. При ―паровом взрыве‖ лигноцеллюлоза претерпевает превращения, в результате которых она становится более чувствительной к ферментативному гидролизу. При этом увеличивается и выход реакционноспособных лигнинов, высвобождаемых под действием слабой щѐлочи. При разрушении замораживанием используется жидкий аммиак. В случае умеренно твѐрдой древесины метод автогидролиза на основе парового взрыва даѐт отходы, пригодные для дальнейшей переработки микробами; древесина хвойных и тропических пород, более богатая лигнином, плохо поддаѐтся такой обработке. Гидролизный лигнин – неоднородный продукт кислотной переработки древесины, который состоит из собственно лигнина (до 80%),остатков полии моносахаридов, органических кислот, смол, воска, азотистых веществ, зольных элементов и не отмытых при гидролизе древесины минеральных кислот. Научно-исследовательские работы по изысканию рациональных путей использования гидролизного лигнина проводятся практически с начала возникновения гидролизного производства, но реализация многих предложений из-за сложности и неоднородности состава лигнина, недостаточной технологической отработанности отдельных стадий и аппаратурного оформления так и не была осуществлена. Например, разработаны технологии использования гидролизного лигнина в качестве компонента фенолформальдегидных смол и резин, сорбента для очистки сточных вод, реализуется технология получения лечебного препарата ―Полифепан‖ и другие. И всѐ же переработке подвергается лишь около 2.7% 212 образующегося гидролизного лигнина. Существуют рекомендации по использованию гидролизного лигнина в качестве энергетического топлива, так как невысокая теплотворная способность гидролизного лигнина может быть эффективной мерой по его утилизации, но не использованием его потенциальных возможностей как природного органического сырья. Вернуть гидролизный лигнин в круговорот углерода в природе можно с помощью микробиологического компостирования. Существует достаточное количество рекомендаций для использования гидролизного лигнина и смеси его со щелочными добавками, например навозом в качестве удобрения. Наряду с этим имеются сведения об ухудшении структуры и механического состава почвы из-за цементирующей способности гидролизного лигнина. Очевидно, для получения полноценного удобрения простого смешивания гидролизного лигнина с органическими добавками недостаточно. Одно из направлений переработки технических лигнинов в органоминеральные удобрения связано с использованием микроорганизмов и их ферментов в качестве делигнифицирующих агентов, так как разрушение лигнина является одним из основных этапов компостирования. В силу сложного химического строения, лигнин чрезвычайно медленно разлагается. Разрушение лигнина осуществляют в основном базидиальные грибы, и в первую очередь грибы белой гнили. Биодеградация лигнина этими грибами – многоступенчатый процесс с участием ферментов лигнолитического комплекса. Проблема загрязнения окружающей среды выбросами целлюлознобумажной и лесохимической промышленности особенно актуальна в областях, где расположены гидролизные предприятия, с которых ежегодно на свалку вывозится огромное количество гидролизного лигнина. Такие крупнотоннажные отходы занимают значительные земельные участки и загрязняют прилегающие к предприятиям территории. Следовательно, основной задачей биотехнологии является не получение, а утилизация лигнина как отхода целлюлозно-бумажного производства. ГЛАВА 17. Получение антисывороток 17.1. Иммуногенность антигена Иммуногенность антигена - это способность в организме иммунизированного животного образования антител. Иммуногенность как биологическое свойство антигена является более сложным, чем антигенность. Антигенности того или иного вещества недостаточно, чтобы вызвать образование антител. В качестве примера можно привести гаптены, которые приобретают иммуногенность только после конъюгирования с соответствующим носителем. Иммуногенность веществ сильно зависит от их молекулярной массы: чем выше молекулярная масса, тем выше иммуногенность. Отсюда вытекает важное практическое следствие - сшивка биополимеров между собой и 213 другими белками повышает иммуногенность. Зависимость иммуногенности от молекулярной массы, по-видимому, определяется следующими причинами: во-первых, увеличение времени пребывания антигена в организме при возрастании его молекулярной массы; во-вторых, у высокомолекулярных антигеноа существенно возрастает способность взаимодействовать с макрофагами, в-третьих, с увеличением молекулярной массы в антигене увеличивается как общее количество антигенных детерминант, так и их разнообразие, что повышает эффективность взаимодействия] антигенов как с B-, так и с T-лимфоцитами. Плотность расположения и количество антигенных детерминант на поверхности антигенов также имеет важное значение: по мере увеличения этих показателей иммуногенность в начале растет, а затем начинает уменьшаться. Так, например, для динитрофенильной гаптеновой группы было показано, что из конъюгатов, содержащих 3, 16 и 28 групп на молекулу бычьего альбумина, максимальной антигенностью обладал конъюгат, содержащий 16 молекул гаптена. Одной из причин такого эффекта, повидимому, является сложность межклеточной кооперации. В частности, показано, что в иммунном ответе против антигенов, имеющих повторяющиеся антигенные детерминанты, участвуют только В-лимфоциты; такие антигены называются независимыми. Для этих антигенов, например полимеров. D-аминокислот, также характерно снижение скорости метаболизма в организме. Очень важным является понятие "чужеродность" иммуногена. Установлено, что чем более антиген отличается по своей структуре от гомологичного антигена иммунизируемого животного, тем выше его иммуногенность. Например, инсулины человека и многих видов животных имеют близкую первичную структуру и поэтому для них инсулин человека малоиммуногенен. Однако между инсулином человека и морской свинки имеются достаточные отличия, что позволяет использовать этих животных как продуцентов соответствующих антисывороток. Однако это правило нельзя считать абсолютным. Так, например, гормон тироксин имеет одинаковую структуру у всех животных, тем не менее, будучи конъюгированным с подходящим белком, он становится хорошим иммуногеном. В данном случае антигенная детерминанта состоит не только из гормона, но и "ножки" и части белковой глобулы, что в целом создает "чужеродную" структуру. Именно на этом принципе основано получение антител против различных низкомолекулярных физиологически активных веществ. "Чужеродность" зависит от генетических особенностей иммунизируемого животного, поэтому часто иммуногенность связывают с генетической чужеродностью антигена. Из "чужеродности" следует, что иммуногенность - это не абсолютное свойство антигена по отношению к данному виду животного, а иногда даже к индивидуальному организму. Необходимо иметь в виду, что иммунная система организма сама находится 214 под жестким генетическим контролем, который определяет как биологическую активность различных участников иммунного процесса, так и многообразие специфичностей рецепторов, а значит, и специфичностей антител. Именно видовая и индивидуальная вариабельность организмов требует внимательного выбора вида животного. Чем менее "чужеродный" антиген, тем большее количество животных следует брать для иммунизации. Так, например, для получения антисывороток против инсулина наиболее иммунореактивными являются морские свинки, при этом в среднем только одна из семи морских свинок дает удовлетворительную для целей анализа антисыворотку. Даже в случае получения антисывороток против достаточно "чужеродных" антигенов необходима большая группа животных, так как в этом случае нивелируются индивидуальные различия. Смесь антисывороток против данного антигена от разных животных одной группы называют пулом. Из лабораторных животных чаще всего берут для иммунизации кроликов, морских свинок или мышей в зависимости от количества имеющегося антигена, доступности животного и т.д. Возможность использования группы лабораторных животных позволяет решить проблему отбора из них наиболее иммунореактивных. Иммунизировать удобнее самцов, так как у них иммуногенный ответ менее подвержен влиянию гормональных циклов. Для получения антител против вирусов эффективными оказались куры, у которых антитела накапливаются в яйцах. Большие количества антисывороток получают иммунизацией крупных животных: козлов, баранов, ослов, лошадей. Для получения специфических антисывороток важное значение имеет гомогенность антигена. Это обусловлено тем, что примеси чужеродных антигенов могут обладать большей иммуногенностью, чем основной антиген, в результате чего, несмотря на небольшое количество примеси, против нее может образоваться достаточное количество антител. Так, например, вирусные антигены, выделенные из культуры ткани животных, содержат примесь тканевых антигенов, против которых вырабатываются антитела, дающие ложноположительные реакции в иммунохимическом анализе. Степень иммунного ответа также зависит от количества введенного антигена. При определенных концентрациях антигена, как высоких, так и низких, наступает торможение гуморального иммунного ответа, называемое толерантностью. Это обусловливает необходимость выбора оптимальной дозы в каждом конкретном случае, с учетом чистоты препарата и его иммуногенности. Доза иммуногена для одной инъекции кролику или морской свинке составляет в среднем 100-300 мкг на 2 кг массы. Доза, необходимая для крупных животных, не увеличивается пропорционально их массе. Так, для овец достаточна доза, равная 0,25-5 мг иммуногена на инъекцию, для осла - 0,5-10 мг. В случае использования в качестве иммуногена конъюгата гаптенноситель доза зависит от молекулярной массы конъюгата. 215 Способ введения антигена и периодичность введения влияют на иммунологическую активность антисывороток. Так как иммунный ответ формируется в организме постепенно, принято различать первичный ответ и вторичный ответ. Первичные и вторичные антисыворотки отличаются по составу антител и их специфичности. Обычно высокоактивные антисыворотки получают после нескольких циклов иммунизации. Однако очень длительные иммунизации могут привести к снижению специфичности из-за постепенного увеличения титра антител к примесным антигенам. В процессе иммунизации изменяется также аффинность и соотношение между различными фракциями антител. Такая вариабельность качества антисывороток по специфичности антител, их физико-химическим свойствам и концентрации является следствием популяционной природы иммунного ответа. В связи с этими обстоятельствами на практике необходимо вести непрерывный контроль за качеством получаемых антисывороток. 17.2. Получение иммунных антисывороток Адъюванты - это соединения, которые при введении в организм вызывают неспецифическое усиление иммунного ответа и тем самым повышают способность организма реагировать на любой иммуноген. Адъювантными свойствами обладают масла, липосомы, клетки бактерий, полимеры и др. Адъюванты, введенные в организм вместе с иммуногеном, выполняют две функции. Во-первых, они способствуют более медленному освобождению иммуногена из участков инъекции, что замедляет его поступление в кровоток, в результате чего увеличивается вероятность встречи иммуногена с иммунокомпетентными клетками, а также резко снижается его токсичность. Во-вторых, адъюванты вызывают сильное воспаление в месте введения иммуногена, при этом активируется фагоцитоз и стимулируется местная циркуляция лимфоцитов, происходит неспецифическая стимуляция иммунокомпетентных клеток. Для усиления такой неспецифической иммуностимуляции в состав адъюваитов дополнительно включают препарат бактериальных клеток рода Bacillus pertussium. В настоящее время для целей иммунизации широко применяется коммерческий препарат полного адъюванта Фрейнда, в состав которого входят смесь минеральных масел, эмульгатор и убитые микобактерии. Препарат адъюванта можно приготовить в лабораторных условиях, тщательно смешав три части минерального масла, одну часть безводного ланолина, четыре части 0,15 M К-фосфатного буфера и препарат микобактерии до конечной концентрации 10 мг/мл так, чтобы частички микобактерии равномерно распределились по всему объему. Адъювант смешивают с водным раствором иммуногена в отношении 2: 1 до образования нерасслаивающейся эмульсии, в которой водный раствор иммуногена находится в мицеллах. 216 Использование в иммунизации адъюванта снижает возможность появления толерантности, позволяет расширить диапазон вводимого иммуногена от 50 до 200 мкг на одну инъекцию. После введения адъюванта у животных часто образуются гранулемы, которые влияют на самочувствие животных, поэтому в течение иммунизации необходимо тщательно наблюдать за состоянием здоровья животного. В случае ухудшения самочувствия очередную инъекцию пропускают и лишь после выздоровления животного продолжают иммунизацию. Способы иммунизации. Введение иммуногена приводит к активации лимфоидных клеток, расположенных вблизи от места инъекции. Наиболее эффективно вводить иммуноген малыми порциями в большое количество точек. Введение иммуногена можно осуществлять различными способами. Внутрикожное введение. На очищенном от шерсти участке кожи животного делают острым скальпелем несколько царапин, а затем втирают в это место раствор иммуногена. Применение этого способа позволяет получать высокий иммунный ответ уже после однократного введения, в результате чего значительно сокращается расход иммуногена. Однако описанный способ трудоемок в исполнении и обычно вызывает сильные болевые ощущения у животного от обширных участков изъязвлений в местах введения иммуногена. Подкожная иммунизация. В точки, расположенные вдоль позвоночника животного, вводят 5-6 порций раствора иммуногена объемом приблизительно 2 мл. Внутримышечное введение. Одновременно часть иммуногена вводят в мышцу задних ног животного и небольшими порциями. Внутрибрюшинное введение. Этот способ используют для иммунизации мелких лабораторных животных, таких, как мыши или морские свинки. Прямое введение иммуногена в лимфатические узлы. Иммуноген инъецируют в лимфоидные узлы, расположенные в подколенной ямке задних ног кролика, что позволяет уменьшить его количество до 10-50 мкг, а объем вводимого раствора - до 25 мкл. Этот способ является сложным в техническом исполнении, поскольку включает в себя разрез кожи, поиск лимфоузла и введение иммуногена без его повреждения. Внутривенное введение. Этот способ обычно применяется для повторных инъекций, после которых проводят отбор крови у животного. Раствор иммуногена вводят непосредственно в кровоток. Адъювант в этом способе введения антигена не применяется, поскольку он оказывает токсический эффект и животное может погибнуть. Более редко используются такие способы иммунизации, как введение иммуногена в подушечки лап или в конъюнктиву глаза, которые вызывают сильные болевые ощущения у животного. Иммунизацию начинают введением животному иммуногена в смеси с полным адъювантом Фрейнда, а для повторных инъекций применяют неполный адъювант Фрейнда. Перед 217 отбором крови за 7 - 9 дней проводят 1-3 внутривенных инъекций для повышения уровня антител. Число инъекций и интервалы между ними чаще всего подбираются экспериментально для конкретного иммуногена. В процессе иммунизации у животных отбирают небольшие пробы крови для оценки количества антител. Максимальный уровень иммунного ответа на введение большинства растворимых антигенов достигается через 40-60 дней после первой инъекции. В том случае, когда иммуногеном являются клетки микроорганизмов, максимально высокий уровень антител наблюдается гораздо раньше. После окончания первого цикла иммунизации животному в течение 30 дней дают восстановить здоровье и проводят реиммунизацию, включающую 1-3 внутривенные инъекции. Ниже приводятся схемы иммунизации морских свинок и кроликов инсулином и конъюгатом тироксин - бычий сывороточный альбумин. Для получения антисывороток морских свинок к инсулину свиньи проводят многоточечные инъекции инсулина подкожно, внутримышечно и внутрибрюшинно по следующей схеме: 1, 8, 15-й день - 200 мкг инсулина в 0,5 мл физиологического раствора смешивают с 0,5 мл полного адъюваита Фрейнда и вводят животному. После 30 дней отдыха животному в течение 3 дней ежедневно вводят антиген путем многоточечных инъекций по схеме: 45, 46, 47-й день - 150 мкг инсулина в 0,7 мл физиологического раствора. Отбор крови проводят через 7-9 дней из сердца. Второй цикл иммунизации осуществляется после месячного отдыха животных по схемам 45, 46 и 47 дней. Для получения антисывороток с титром, удовлетворительным для проведения ИФА, часто требуется 4-5-кратное повторение циклов иммунизации. Отбор крови каждый раз проводится на 7 - 9-й день после последней инъекции. Иммунизацию кроликов конъюгатом тироксин - БСА проводят многоточечными инъекциями вдоль позвоночника и внутримышечно в область микроузлов задних лап по следующей схеме: 1-й день - 1-2 мг конъюгата. T4 - БСА в 0,7 мл физиологического раствора смешивают с 0,7 мл адъюванта Фрейнда и полученную эмульсию вводят животному; 31, 32, 33 или 45, 46, 47-й дни - внутривенно вводят 0,7-1 мл физиологического раствора, содержащего 1-2 мг T4 - БСА. Кровь берут из сердца на 7-9-й день после последней инъекции. Через месяц цикл внутривенной иммунизации, повторяют. Отбор крови и получение антисыворотки. Иммунизированное животное используется в качестве донора иммунной сыворотки в течение 5-7 мес, за это время удается провести 5-6 циклов иммунизации. Животных, прошедших несколько циклов иммунизации, называют гипериммунными. Кровь у животных отбирают из вены уха или непосредственно из сердца путем кардиальной пункции в объеме 50-70 мл у кролика и 5-10 мл у 218 морской свинки в стерильные пробирки, промытые стерильным буферным раствором. Кровь, лишенная клеточных элементов, называется плазмой. В плазме содержится фибриноген, приводящий к образованию во всем объеме пробирки сгустка фибрина, который осторожно удаляется центрифугированием при 1000-2000 об/мин в течение 15 мин. Плазма, лишенная фибрина, называется сывороткой. Для удаления белков системы комплемента сыворотку прогревают в течение 30 мин при 56°С, при этом антитела сохраняют свою активность. Обработку крови и получение сыворотки надо проводить с максимальной осторожностью, избегая разрушения эритроцитов. Наличие внутриклеточных белков и ферментов в сыворотке может приводить к появлению дополнительного фона в некоторых модификациях иммуноферментного анализа. Это замечание касается прежде всего использования антисывороток в гомогенных методах. Хранение антисывороток. Нативную иммунную сыворотку можно хранить 3-6 мес без потери иммунологической активности в замороженном состоянии при - 20°С, предварительно разливая ее во флаконы по 0,5-1 мл. Отмораживание-замораживание ведет к снижению иммунологической активности сыворотки, поэтому лучше замораживать ее небольшими порциями для одноразового употребления. Удобно хранить антисыворотку в лиофилизованном состоянии в ампулах под вакуумом. В сухом виде антисыворотка сохраняет иммунологическую активность при комнатной температуре в течение 1-2 лет. Размороженную или разведенную сухую сыворотку в растворе можно хранить при +4°С в течение недели, предварительно добавив в нее консервант - хлороформ, 0,1% азида натрия или 0,01% мертнолата. Следует иметь в виду, что консерванты могут быть ингибиторами ферментовмаркеров в иммуно-феремнтном анализе. 17.3. Выделение и очистка антител. Для увеличения относительного количества антител обычно используют г-глобулиновую или IgG-фракции иммунной сыворотки. Наиболее полное выделение г-глобулиновой фракции без снижения ее иммунологической активности достигается при осаждении сульфатом аммония, после чего осадок длительно диализуется с частой сменой буферных растворов. Другим широко распространенным способом является осаждение полиэтиленгликолем с Air=4000-6000. В 10% -иом растворе полиэтиленгликоля происходит агрегация всех белков с Мг> ~> 150 000, в результате чего осаждаются белки г-глобулиновой фракции, а белки меньшей молекулярной массы остаются в растворе. Этот метод позволяет очень быстро получить препарат г-глобулиновой фракции антисыворотки. Способ достаточно эффективный, если препарат не подвергается хранению при низких температурах. Длительное хранение полученного препарата с 219 сохранением иммунологической активности возможно после предварительного тщательного диализа. Антитела в большей части антигенов относятся к IgG-фракции иммунной сыворотки. С целью повышения чувствительности и специфичности анализа во многих случаях полезно использовать для сорбции на твердой фазе и для получения конъюгатов IgG-фракцию нативной сыворотки. Наиболее простой и доступный способ выделения IgG метод ионообменной хроматографии на ДЗАЭ-сефадексе или ДЗАЭцеллюлозе. Выделение IgG-фракции кроличьей аитисыворотки. 1. 10 мл аитисыворотки кролика разбавляют в 2 раза 0,15 M NaCl1 добавляют 6,26 г 2SO4, перемешивают и инкубируют 12-16 ч при 4°С. 2. Выпавший осадок удаляют цеитрифугироваиием, растворяют в 10 мл фосфатного буфера и затем диализуют против того же буфера в течение ночи при комнатной температуре. 3. После удаления осадка центрифугироваиием раствор наносят на колонку с ДЭАЭ-целлюлозой, уравновешенную фосфатным буфером. 4. IgG-фракцию определяют, измеряя оптическую плотность элюата при 280 им, концентрацию рассчитывают, используя коэффициент молярного поглощения е=1,5 г/л см-1. Для выделения IgG кролика используют 50 мМ фосфатный буфер, содержащий 20 мМ ЭДТА.8 г Na2HPO4 ∙2З20 н 7,4 г ЭДТА-Na растворить в 900 мл воды, довести рН до 8 IM NaOH и затем добавить воды до 1 л. Выделение антител методом иммуносорбции. Специфичность сыворотки проверяется в реакциях с используемым для иммунизации препаратом антигена и с набором близких ему по структуре химических соединений. В случае недостаточна очищенного антигена возможны неспецифические реакции, обусловленные наличием антител к применяемым антигенам. Перекрестные реакции с другими соединениями могут наблюдаться при наличии у них химических структур, близких к структуре антигенных детерминант иммуногена. Удаление неспецифических антител из антисыворотки обычно осуществляют с помощью метода адсорбции на соответствующем неспецифическом антигене. Адсорбция неспецифических антител в реакции преципитации с растворимой фракцией путем центрифугирования может привести к появлению в антисыворотке или растворимых комплексов AT-Ar, или избытка добавленного Ar, присутствие которых влияет на способность антисыворотки реагировать со сцецифическим антигеном в ИФА. Наиболее удобно для адсорбции антисыворотки использовать антиген, иммобилизованный на твердой фазе. Добавляя такой иммуносорбент в антисыворотку или пропуская антисыворотку через колонку с иммуносорбентом, можно быстро освободиться от перекрестно реагирующих антител. Метод аффинной хроматографии на иммуносорбентах используется для получения препаратов очищенных антител. Наиболее широкое 220 распространение лолучили иммуносорбенты на основе CNBrактивированной сефарозы. Выпускаемая в нашей стране CNBrактивированная агароза по своим основным параметрам не уступает зарубежному аналогу. Основные операции, используемые для получения иммобилизованных на CNBr-агарозе антигенов или антител, приведены в табл.6. Таблица 6. Ковалентное связывание с CNBr-активированной сефарозой Операция Условия 1. Взвесить требуемое 1 г высушенного препарата дает около количество CNBr-активирован 4,5 мл геля ион сефарозы 2. Промыть на стеклянном Использовать для промывки раствор 1 фильтре и дать набухать гелю мМ HCl, а затем для уравновешивания и набухания - боратный или гидрокарбонатный буфер 3. Растворить в буфере белок Использовать гидрокарбонатный или обратный буфер Гидрокарбонатный буфер: 0,1 M NaHCO3, содержащий 0,5 NaCl 4. Смешать белковый раствор с Раствор должен содержать 5-10 мг суспензией геля антигена 5. Блокировка оставшихся Два часа при комнатной температуре или активных групп ночь при +4 °С Гель промывается буфером, содержащим блокирующие соединения. Используются 1M этаиоламин или 0,2 M глицин, рН 8,0 6. Отмывка от несвязавшегося Используется буфер, в котором антигена осуществлялось ковалентное связывание, затем 0,1 M ацетатный буфер, рН 4, содержащий 0,5 M NaCl, после, чего гель опять уравновешивается боратным или гидрокарбонатным буфером Следует отметить, что при работе с иммуносорбентами на основе CNBr-активированной сефарозы нельзя использовать буфер, содержащий аминогруппы. Получение фрагментов антител. При разработке некоторых модификаций ИФА может возникнуть необходимость использования конъюгатов фермента не с целой молекулой иммуноглобулина, а с тем ее фрагментом, который специфически связывается с молекулой антигена - Fabфрагментом. Обусловлено это прежде всего тем, что Fc-фрагмент молекулы, 221 ответственный за эффекторные функции иммуноглобулина, обладает способностью неспецифически взаимодействовать с другими белками, сорбированными на носителях при проведении твердофазного ИФА. Эффект неспецифической сорбции определяется природой антигена, концентрацией сорбированного на носителе белка и рядом других факторов. Получение Fab-фрагментов. 1. Готовят раствор, содержащий 0,1-3 мг Р2-фрагмента в 0,45 мл 0,1 M Na-фосфатиом буфере, рН 6,0. ∙ 2. Добавляют 0,05 мл 0,1 M 2-меркаптоэтанола в том же буфере, содержащем 5 мЛ\ ЭДТА. 3. Смесь инкубируют при 37 0C в течение 1,5 ч. 4. Тестирование антисывороток. Антисыворотки, полученные даже от одного животного, значительно различаются по своей способности связывать антиген. Для сравнительной характеристики и оценки качества антисывороток проводят их тестирование, которое позволяет решить следующие две важные задачи: во-первых, произвести отбор именно тех сывороток, которые по своим свойствам удовлетворяют требованиям иммунохимического анализа, во-вторых, осуществить стандартизацию антисывороток при их промышленном производстве для иммуноферментных наборов. Первичный отбор антисывороток проводят на основании нахождения их титра, который представляет собой интегральный параметр, характеризующий взаимодействие с антигеном. Более детальные сведения о сыворотке получают, определяя аффинность и концентрацию антител. Следующий этап - это установление антигенной специфичности антител, т.е. возможности взаимодействовать со структурно сходными антигенами. Принципиальное различие этих этапов заключается в следующем: при определении титра исследуют связывание выбранной концентрации антигена при различных разведениях аитисыворотки. При определении аффинности и концентрации антител исследуют связывание антисыворотки с различными концентрациями меченого антигена. При изучении специфичности антисывороток в соответствующем разведении и при постоянной концентрации меченого антигена добавляют различные концентрации перекрестно реагирующего антигена. Во всех случаях используют концентрации антигена либо близкие к минимально детектируемым в данном виде анализа, либо в том диапазоне, который соответствует их концентрации в Образце. Определение титра аитисыворотки. Титр - это эффективная величина, характеризующая связывающую способность антител, зависящая от их концентрации и аффинности. Абсолютное значение титра также зависит от метода и условий проведения эксперимента и от начальной концентрации свободного антигена. 222 Количественно титр находят как предельное разведение сыворотки, при котором еще наблюдается положительный регистраруемый данным методом эффект взаимодействия антисыворотки с антигеном. Например, если титр устанавливают методом преципитации свободного антигена в геле и для первой сыворотки образование преципитата наблюдается при разведении в 2т раза, а для другой - в 2" раз, фп титр первой сыворотки равен 2т, а второй - 2", причем вторая антисыворотка менее активная, чем первая. Иногда оперируют понятием "50% -ный титр", подразумевая под этим, соответственно, разведение сыворотки, вызывающее 50% -ное связывание антигена. "Рабочим титром" называют то начальное разведение сыворотки, которое используют непосредственно в эксперименте (табл.7). Титр антисыворотки сильно зависит от концентрации антигена, используемого для тестирования и от способа его определения. Например, одна и та же сыворотка может иметь титр 100 в тесте иммунопреципитации и 100 000 в тесте иммуноферментного анализа. Поэтому при тестировании сыворотки и определении титра лучше всего применять тот же метод, что и при анализе, а концентрацию антигена выбирать близкую к минимальной в том диапазоне, который выбран для анализа. Кроме того, следует учитывать, что в иммуноферментном анализе используют как гомогенные, так и гетерогенные методы определения концентрации антигена, которые могут сильно отличаться по структуре образующихся иммунных комплексов. В частности, в твердофазных методах вероятность образования циклических комплексов антиген - антитело значительно меньше, чем в гомогенных, а следовательно, и наблюдаемая аффинность антител в обеих системах будет разная. Таблица 7. Схема тестирования сывороток Этап исследования Цель Титр Аффинность Специфичность Отбор высокоактивных иммунных аитисывороток, определение конечного и рабочего титра Оценка аффинности и концентрации фракции высокоаффиниых антител Определение специфичности антител При разработке методов ИФА обычно пользуются значением титра антисыворотки, определенным в непрямом методе, в котором на стенках лунок микроллаты первоначально сорбируют антиген и затем изучают связывание с ним иммунной сыворотки в последовательных разведениях. Связывание исследуемых антител с иммобилизованным антигеном регистрируют с помощью антивидового конъюгата к иммуноглобулиновой фракции сыворотки животного, используемого для иммунизации. 223 Результаты в этом случае обычно оценивают с помощью понятия "50% -ный титр". На рис. приведены кривые титрования антисывороток к инсулину, полученных иммунизацией морских свинок. Максимальный сигнал, регистрируемый по оптической плотности А продукта реакции окисления перекисью водорода 5-аминосалициловой кислоты, составляет ~1,4 оптических единиц. За титр сыворотки принимается ' такое разведение, при котором оптическая плотность, регистрируемая в ИФА, имеет значение, близкое к 0,7. Обычно такой подход используется в качестве 1-го этапа оценки качества полученных иммунных сывороток и позволяет отобрать высокоактивные иммунные сыворотки. Однако для более глубокой оценки полученных антител проводят определение их аффинности. При разработке методов ИФА ряда антигенов, присутствующих в биологических жидкостях в низких концентрациях порядка Ю-10-IO11M, должны быть использованы антисыворотки, имеющие не только высокий титр, но и достаточное содержание антител, обладающих высокой константой связывания. Аффинность антител. В основном в таких экспериментах применяются иммобилизованные антитела и антигены, меченные радиоактивной меткой. Титрование антисывороток к инсулину методом непрямого твердофазного ИФА: по оси абсцисс - разведение разбавленной в 100 раз сыворотки, по оси ординат - оптическая плотность при 430 им продукта пероксидазного окисления 5-аминосалициловой кислоты перекисью водорода. В качестве контроля - использована сыворотка крови неиммунизированного животного. При использовании ферментных меток химическая структура меченого антигена может существенно отличаться от немеченого. В этих случаях полученные значения констант связывания характеризуют, как правило, взаимодействие в данной конкретной системе, вследствие - чего иммунная сыворотка, обладающая высокой аффинностью в реакции с конъюгатом одной структуры, может иметь значительно более низкие значения константы связывания с конъюгатом, полученным другим способом. Если исследователь имеет дело с разработкой конкурентных методов ИФА, т.е. располагает набором иммунных сывороток и конъюгатом антигенфермент, то важным этапом создания тест-системы является выбор пары антитело - конъюгат, характеризующейся Необходимой константой взаимодействия. Для проведения скрининга у-глобулиновую фракцию каждой антисыворотки иммобилизуют на поверхности микропланшета и изучают ее связывание с имеющимся набором конъюгатов. Эффективное значение связи является величиной, обратной значению концентрации конъюгата, при которой связывается 50% активных центров на носителе. Получение такого конъюгата для широкого круга антигенов представляется непростой задачей. Более универсальным реагентом являются антитела, меченные ферментом, поэтому остановимся на более общем подходе к определению аффинности антител в сыворотке или асците, 224 основанном на твердофазном ИФА. Этот метод, предложенный Б. Фриге, позволяет определять константу связывания антител с антигеном при их взаимодействии в растворе. Постановка метода включает две основных стадии: 1) проведение реакции антиген - антитело в растворе; 2) определение концентрации антител после установления равновесия с помощью метода твердофазного ИФА. ГЛАВА 18 Использование продуктов биотехнологии для защиты окружающей среды. Экобиотехнология решает проблемы по охране окружающей среды, такие как переработка отходов, очистка воды, устранение загрязнений. В процессе круговорота загрязняющих веществ в экосистемах огромную роль играют микроорганизмы. Однако, многие из созданных человеком низкомолекулярных соединений (ядохимикаты, детергенты) и высокомолекулярных полимеров оказались устойчивыми и не разлагаются микроорганизмами, кроме того, они проявляют мутагенное, канцерогенное, тератогенное влияние, поэтому для их утилизации требуется разработка более совершенных технологий очистки. Обычно для утилизации отходов применяют комплексы микроорганизмов и специальные приборные устройства. Чужеродные вещества (ксенобиотики), попадая в организм человека и животных, претерпевают различную биотрансформацию: окисление, восстановление, гидролиз и другие превращения с участием ферментных систем. В воде и почве биотрансформация ксенобиотиков протекает под воздействием ферментов и микроорганизмов. Изучение реакций в почвах затруднено гетерогенностью среды и адсорбцией ксенобиотиков, микроорганизмов и ферментов на частицах и коллоидах почв. Многие ксенобиотики в биосфере достаточно устойчивы, например, ДДТ не исчезает из почвы 30 лет, альдрин и хлордан – 15 лет, диэльдрин – 25 лет, гептахлор – 14 лет. Некоторые вещества при распаде образуют еще более устойчивые и токсичные соединения. Одним из направлений экобиотехнологии является получение экологически чистой энергии. Экологически чистой считается энергия, получаемая путем преобразования солнечной энергии в электрическую с помощью солнечных коллекторов, а также энергия биогаза и микробного этанола. 18.1 Получение биогаза Биогаз – это смесь, состоящая из 65 % метана, 30 % углекислого газа, 1 % сероводорода и незначительных примесей азота, кислорода, водорода и угарного газа. Энергия, заключенная в 1 м3 биогаза, эквивалентна энергии 0,6 м3 природного газа или 0,74 л нефти, или 0,66 л дизельного топлива. В 225 основе получения биогаза лежит процесс метанового брожения, или биометаногенез – процесс превращения биомассы в энергию. Биометаногенез – сложный микробиологический процесс, в котором органическое вещество разлагается в анаэробных условиях до метана и диоксида углерода. Микробиологическому разложению поддаются практически все соединения природного происхождения, а также значительная часть ксенобиотиков органической природы. Для получения биогаза можно использовать отходы животноводства, отходы сельского хозяйства, испорченные продукты, стоки крахмалоперерабатывающих предприятий, жидкие отходы сахарных заводов, бытовые отходы, сточные воды городов. Процесс ведут при температуре от 30 до 60 ºС и значения рН от 6 до 8. Получение биогаза широко применяют в Индии, Китае, Японии. Чаще всего используют вторичные отходы (то есть отходы животноводства и сточные воды городов). Рисунок 7 – Схема устройства реактора для обработки сельскохозяйственных отходов Подача навоза, остатков растениеводства (субстрата) и отбор отработанных (стоков) осуществляют в нижней части реактора. Режим его работы может быть как периодический, так и полунепрерывный. Реактор обычно имеет две (или более) секции для разделения стадий процесса. Биогаз сгорает с образованием углекислого газа и воды, а в реакторе остается естественное удобрение – сапропель. Он содержит азот, фосфор, соли калия, необходимые для роста растений. Использование сапропеля более целесообразно, чем использование навоза, поскольку навоз перегружает почву. В анаэробном процессе биометаногенеза выделяют три последовательных стадии, в которых участвуют свыше 190 различных микроорганизмов. На первой стадии ферментативному гидролизу подвергаются сложные многоуглеродные соединения: белки, липиды, 226 полисахариды. На второй стадии идет образование ацетата, которое может протекать двумя путями: а) ацетогенные микроорганизмы усваивают водород, углекислый газ и некоторые одноуглеродные соединения с образованием ацетата; в) гомоацетатные микроорганизмы усваивают водород, углекислый газ и некоторые одноуглеродные соединения с образованием ацетата. На третьей стадии образуется метан. Он может синтезироваться через стадию восстановления углекислого газа с молекулярным водородом, а также из метильной группы ацетата. Некоторые метановые бактерии в качестве субстрата используют формиат, углекислый газ, метанол, метиламин и ароматические соединения. 4H2 + CO2 CH4 + 2H2O + CO CH4 + H2O 3H2 2H2O + CH4 CO 4HCOOH 4CH3OH CH3COOH CH4 3CH4 + + + CO2 3CO2 CO2 + + CO2 CH4 H2O В зависимости от температуры протекания процесса метановые бактерии разделяют на мезо- и термофильные. Оптимальная температура для мезофильных бактерий – от 30 до 40 ºС, для термофильных – от 50 до 60 ºС. В целом термофильный процесс метаногенеза идет интенсивнее мезофильного, причем субстрат обеззараживается от патогенной микрофлоры и гельминтов. Микрофлора для метаногенеза формируется в основном микрофлорой желудочно-кишечного тракта животных: Lactobacillus acidophilus, Eubacterium aerofaciens, Methanobacterium mobile, Methanosarcina sp., Methanobrevibacterium ruminantium. Метанобразующие бактерии от 90 до 95 % используемого углерода превращают в метан и лишь от 5 до 10 % углерода – в биомассу. Анаэробная биоконверсия органических отходов в метан – наиболее конкурентоспособная область биоэнергетики. Она позволяет получать из местного сырья биогаз как локальный источник энергии. Экологически чистые источники энергии не влияют отрицательно на окружающую среду. Современные источники энергии – ГЭС, ТЭС, АЭС – вызывают серьезные нарушения во внешней среде. ГЭС служат причиной затопления территорий, изменения ландшафта, гибели биоценозов. ТЭС загрязняют атмосферу, вызывают отчуждение земель. АЭС создают угрозу радиационного загрязнения. Сжигание нефти и газа вызывает повышение концентрации 227 углекислого газа, образование смога и, кроме того, уменьшение ресурсов нефти и газа. Основное преимущество биогаза состоит в том, что он является возобновляемым источником энергии. В природе в результате деятельности бактерий образуется ежегодно около 800 млн. тонн метана, примерно столько же добывается людьми. 18.2 Производство биоэтанола С 1975 г. производство пищевого этилового спирта остается постоянным, а производство топливного этанола увеличилось в 10 раз. Этанол может применяться как топливо самостоятельно или в смеси с бензином в количестве от 10 до 26 %, такую смесь в США называют газохол; или в смеси с дизельным топливом в количестве 3 %. Эти смеси могут быть использованы без изменений в конструкции двигателей внутреннего сгорания. В качестве источника энергии спирт используется в Бразилии, США, странах ЕС, то есть в энергодефицитных зонах. Кроме того, спирт широко применяется в химической промышленности в качестве растворителя, экстрагента, антифриза. Этанол преимущественно получают биотехнологическим путем сбраживания сахаров, содержащихся в растениях. В мировом производстве первое место занимает производство спирта из сахарного тростника (Бразилия, США). При переработке сахарного тростника его тщательно давят, сок концентрируют и подвергают брожению. На втором месте находится маниок (кассава) – крахмалистое растение, способное расти на скудных почвах. Считают, что бразильский вариант биотехнологического решения топливной проблемы – наилучший, однако получилось, что лучшие пахотные земли засевались сахарным тростником, при этом для одного автомобиля требуется примерно 13000 м2, в то время, как для одного человека 800 м2 в год. То есть один автомобиль отбирает пищу у 18 жителей. В то же время в Бразилии миллионы людей страдают от недоедания. Кроме того, стоки со спиртовых заводов загрязняют водоемы и нарушают экологическое равновесие. Кроме сахарного тростника и маниока для производства спирта используют злаки, особенно кукурузу, топинамбур, ананас, сахарную свеклу, сорго. При переработке крахмалосодержащего сырья необходимо его предварительное разваривание и обработка ферментами для превращения крахмала и других полисахаридов в усваиваемые микроорганизмами сахара. Для производства спирта можно также использовать мелассу – остаток производства сахара из сахарной свеклы и сыворотку, остающуюся после производства сыров. 228 Производство спирта из сахарного тростника экономически неоправданно (Бразилия), из кукурузы (США) – субсидируется государством, чтобы цена на этанол была ниже, чем на нефтепродукты. В других регионах себестоимость биоэтанола еще выше, поскольку выше себестоимость сырья. Снижение себестоимости этанола может быть достигнуто заменой сырья или кардинальным изменением технологии ферментации. Замена сырья заключается в том, что вместо зерна злаков для превращения в этанол используется биомасса целых растений, как травянистых, так и деревьев, а также твердые коммунальные отходы, то есть мусор. Лигноцеллюлоза (древесина) состоит из трех полимеров: целлюлозы, гемицеллюлозы и лигнина. Путем химического или ферментативного гидролиза эти полимеры расщепляются до мономеров с последующей ферментацией сахаров до этанола. Несмотря на неполный гидролиз (из-за сложности химического строения молекул полимеров) процесс экономически выгоден. Кроме того, найдены виды дрожжей, способные сбраживать в спирт не только гексозу (глюкозу), но и ксилозу; использование таких дрожжей приводит к более полному использованию сахаров, а, следовательно, повышается выход спирта, и снижается его себестоимость. Из гидролизатов древесины и сульфитных щелоков (отход в целлюлозно-бумажном производстве) в России получают технические спирты. 18.3 Очистка сточных вод Важнейшая проблема экологической биотехнологии – очистка сточных вод. Потребность в воде в связи с ростом городов, бурным развитием промышленности, интенсификацией сельского хозяйства огромна. Ежегодный расход воды на земном шаре по всем видам водоснабжения составляет от 3300 до 3500 км3, при этом в сельском хозяйстве – 70 % всего водопотребления. Для производств химической, целлюлозно-бумажной, энергетической промышленности, черной и цветной металлургии и бытовых нужд населения требуется также значительное количество воды. Большая часть этой воды после ее использования возвращается в реки и озера в виде сточных вод. На современном этапе выделяются следующие направления рационального расхода водных ресурсов: более полное использование и расширение воспроизводства ресурсов пресных вод; разработка новых биотехнологических процессов, позволяющих предотвратить загрязнение водоемов и свести к минимуму потребление свежей воды. Загрязнение поверхностных и подземных вод можно подразделить на несколько типов: механическое, сопровождающееся повышением содержания механических примесей и относящееся, в основном, к поверхностным видам загрязнений; химическое, обусловленное 229 присутствием в воде органических и неорганических веществ токсического и нетоксического действия; биологическое, связанное с наличием в воде разнообразных патогенных микроорганизмов, грибов и мелких водорослей; радиоактивное; тепловое. Основные источники загрязнения и засорения водоемов – недостаточно очищенные сточные воды промышленных и коммунальных предприятий, крупных животноводческих комплексов, отходы производства при разработке рудных ископаемых (воды шахт, рудников); сбросы водного и железнодорожного транспорта; пестициды и т.д. Загрязняющие вещества, попадая в природные водоемы, качественно изменяют их состав. Сточные воды содовых, сульфатных, азотно-туковых заводов, обогатительных фабрик свинцовых, цинковых, никелевых руд, содержащие кислоты, щелочи, ионы тяжелых металлов, меняют физические свойства воды (появление неприятных запахов, привкусов и т.д.). Сточные воды нефтеперерабатывающих, нефтехимических заводов, предприятий органического синтеза содержат различные нефтепродукты, аммиак, альдегиды, смолы, фенолы и другие вредные вещества. Вследствие окислительных процессов уменьшается содержание в воде кислорода, ухудшаются ее органические показатели. Нефть и нефтепродукты – основные загрязнители внутренних водоемов, вод и морей Мирового океана – создают разные формы загрязнения: плавающую на воде нефтяную пленку, осевшие на дно водоемов тяжелые фракции. Вода приобретает токсические свойства и представляет собой угрозу для всего живого: 12 г нефти делают непригодной для употребления одну тонну воды. Вредным загрязнителем промышленных вод является фенол, содержащийся в сточных водах многих нефтехимических предприятий. На жизнь населения водоемов пагубно влияют сточные воды целлюлозно-бумажной промышленности. Окисление древесной массы сопровождается поглощением значительного количества кислорода, что приводит к гибели икры, мальков и взрослых рыб. Сточные воды, имеющие повышенную радиоактивность (100 кюри на 1 л и более), подлежат захоронению в подземные бессточные бассейны и специальные резервуары. В значительной степени загрязняют водоемы моющие синтетические средства, широко используемые в быту, промышленности и сельском хозяйстве и парализующие жизнедеятельность бактерий. Пестициды, попадая в водоемы, накапливаются в планктоне, бентосе, рыбе и по цепочке питания попадают в организм человека, действуя отрицательно, как на отдельные органы, так и на организм в целом. Сточные воды, содержащие отходы кожевенной и целлюлозно-бумажной промышленности, сахарных и пивоваренных заводов, предприятий мясомолочной, консервной и кондитерской промышленности, служат причиной органических загрязнений водоемов. Нагретые сточные воды тепловых электростанций вызывают тепловое загрязнение, которое резко изменяет термический 230 режим, отрицательно влияет на флору и фауну водоемов. Возникают благоприятные условия для массового развития в водохранилищах синезеленых водорослей (так называемое «цветение воды»). Применение того или иного метода (механического, химического, физико-химического и биологического) в каждом конкретном случае определяется характером и степенью вредности примесей. Сущность этих методов состоит в том, что из сточных вод путем отстаивания и фильтрации удаляют механические примеси. Грубодисперсные частицы в зависимости от размеров улавливаются решетками, ситами, песколовками, навозоуловителями, нефтеловушками и т.д. Механическая очистка позволяет выделять из бытовых сточных вод до 60-75 % нерастворимых примесей, а из промышленных – до 95 %, многие из которых как ценные примеси используются в производстве. Химический метод. В сточные воды добавляют различные химические реагенты, которые вступают в реакцию с загрязнителями и осаждают их в виде нерастворимых осадков. Химическая очистка уменьшает количество нерастворимых примесей до 95 %, а растворимых – до 25 %. Физический метод. Эти методы используют для удаления тонкодисперсных и растворенных неорганических примесей, а также для разрушения органических и плохо окисляемых веществ. В арсенал этих методов входят электролиз, окисление, сорбция, экстракция, ионообменная хроматография, ультразвук, высокое давление и др. Биологический метод. Этот метод основан на использовании закономерностей биохимического и физиологического самоочищения рек и других водоемов. Для очистки сточных вод используют биофильтры, биологические пруды и аэротенки. В биофильтрах сточные воды пропускают через слой крупнозернистого материала, покрытого тонкой бактериальной пленкой, благодаря которой интенсивно протекают процессы биологического окисления. В биологических прудах в очистке сточных вод принимают участие все организмы, населяющие водоем. Аэротенки – огромные резервуары из железобетона, в которых очистка происходит с помощью активного ила из бактерий и микроскопических животных, которые бурно развиваются в этих сооружениях, чему способствуют органические вещества сточных вод и избыток кислорода, поступающего с потоком подаваемого воздуха. Бактерии, склеивающиеся в хлопья, выделяют в среду ферменты, разрушающие органические загрязнения. Ил с хлопьями оседает, отделяясь от очищенной воды. Инфузории, жгутиковые, амебы, коловратки и другие мельчайшие животные, пожирая бактерии, не слипшиеся в хлопья, тем самым омолаживают бактериальную массу ила. Сточные воды сначала подвергают механической, а после химической очистке для удаления болезнетворных бактерий путем хлорирования жидким хлором или хлорной известью. Для 231 дезинфекции используют также ультразвук, озонирование, электролиз и другие методы. Биологический метод дает существенные результаты при очистке коммунально-бытовых стоков, а также отходов предприятий нефтеперерабатывающей, целлюлозно-бумажной промышленности и производства искусственного волокна. ЛИТЕРАТУРА 1. Безбородов А.М. Биохимические основы микробиологического синтеза. – М.: Легкая и пищевая промышленность, 1984. 2. Бекер М.Е., Лиепинен Г.К., Райпулис Е.П. Биотехнология - М.: Агропромиздат, 1990. 3. Боярский Л.Г., Коршун В.П., Бикташев Р.У. и др. Ферментные препараты в кормлении животных. – М.: Россельхозиздат, 1985. 4. Булдаков А. – Пищевые добавки (справочник) – С.Пб., 1996. 5.Виестур У.Э., Шмите И.А., Жилевич А.В. Биотехнология: Биологические агенты, технология, аппаратура. – Рига: Занатне, 1987. 6. Грачева И.М., Иванова Л.А., Кантере В.М. Технология микробных белковых препаратов, аминокислот и биоэнергия. – М: Колос, 1992. 7. Грачева И.М., Кривова А.Ю. Технология ферментных препаратов. – 3-е изд. – М.: Изд-во «Элевар», 2000. 8. Елинов Н.П. Основы биотехнологии. – С.-Пб.: Наука, 1995. 9. Квеситадзе Г.И., Безбородов А.М. Введение в биотехнологию. – М.: Наука, 2002. 10. Кашкин П.Н. и др. Антибиотики - Л.: Медицина, 1970. 11. Нечаев А.П., Кочеткова А.А., Зайцев А.Н. Пищевые добавки – М.: ―Колос‖, ―Колос-Пресс‖, 2002. 12. Промышленная микробиология / Под ред. Егорова Н.С. - М.: ―Высшая школа‖, 1989. 13. Самарцев М.А., Беляков Н.В., Кестнер А.И. Применение иммобилизованных ферментов в промышленных процессах. – М.: ОНТИТЭИмикробиопром, 1984. 14. Р. Скоупс. Методы очистки белков - М.: Мир, 1985. 15. Технология переработки жиров. / Под ред. Арутюняна Н.С. – М., 1985. 16.Тютюнников Б.Н. Химия жиров – М.: ―Пищевая промышленность‖, 1966. 17. Биотехнология/ под ред. А.А. Бабаева. – М.: Наука, 1984. 18. Биотехнология: принципы и применение./Под ред. И.Хиггинса, Д.Беста и Дж. Джонса. - М.: Мир, 1988. 19. Варфоломеев С.Д., Калюжный С.В. Биотехнология. – М.: Высшая школа, 1990. 232 20. Глик Б., Пастернак Дж. Молекулярная технология. Принципы и применение. – И: мир, 2002. 21. Дудкин М.С., Громов В.С., Ведерников Н.А. и др. Гемицеллюлазы. – Рига: Занатне, 1991. 22. Елинов Н.П. Химическая микробиология – М.: ―Высшая школа‖, 1989. 23. Квеситадзе Г.И. Грибные и бактериальные амилазы. – Тбилиси: Мецниереба, 1984. 24. А. Сассон. Биотехнология: свершения и надежды. - М.: Мир, 1987. 25. Технология продуктов из гидробионтов. / Под ред. Сафроновой Т.М., Шендерюка В.И. – М.: ―Колос‖, 2001. 26. Технология спирта / Под ред. Яровенко В.Л. – М.: ―Колос‖, 2002. 27. Альбертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. Т. 1. М.: Мир, 1994. 28. Артамонов В.И. Занимательная физиология растений. М.: Агропромиздат, 1991. 336 с. 29. Безбородов А.М. Ферменты микроорганизмов и их применение // Биотехнология. М.: Наука, 1984. 30. Березин И.В., Клесов А.А., Швядас В.К. и др. Инженерная энзимология. М.: Высшая школа, 1987. 144 с. 31. Березин И.В., Клячко Н.Л., Левашев А.В. и др. Иммобилизованные ферменты. М.: Высшая школа, 1987. 160 с. 32. Биология наших дней. Вып. 2. – М.: Знание, 1987. 160 с. 33. Биотехнология. Принципы и применение /Хиггинс И., Бест Д., Джонс Дж. М.: Мир, 1988. 480 с. 34. Биотехнология сельскохозяйственных растений. М.: Агропромиздат, 1987. 301 с. 35. Биотехнология - сельскому хозяйству /Лобанок А.Г., Залашко М.В., Анисимова Н.И. и др. Минск: Урожай, 1988. 199 с. 36. Биотехнология растений: культура клеток. М.: Агропромиздат, 1989. 280 с. 37. Быков В.А., Крылов И.А., Манаков М.Н. и др. Микробиологическое производство биологически активных веществ и препаратов. М.: Высшая школа, 1987. 142 с. 38. Быков В.А., Манаков М.Н., Панфилов В.И. и др. Производство белковых веществ. М.: Высшая школа, 1987. 142 с. 39. Варфоломеев С.Д., Панцхава Е.С. Биотехнология преобразования солнечной энергии. Современное состояние, проблемы, перспективы // Биотехнология. М.: Наука, 1984. 40. Голубовская Э.К. Биологические основы очистки воды. М.: Высшая школа, 1978. 270 с. 41. Грачева И.М., Гаврилова Н.М., Иванова Л.А. Технология микробных белковых препаратов, аминокислот и жиров. М.: Пищевая промышленность, 1980. 448 с. 233 42. Егоров Н.С. Основы учения об антибиотиках. М.: Высшая школа, 1986. 448 с. 43. Иммобилизованные клетки и ферменты. Методы / Под ред. Дж. Вудворта. М.: Мир, 1988. 215 с. 44. Казанская Н.Ф., Ларионова Н.И., Торчилин В.П. Ферменты и белковые препараты в медицине // Биотехнология. М.: Наука, 1984. 45. Каравайко Г.И. Биогеотехнология металлов // Биотехнология. М.: Наука, 1984. 46. Кефели В.И., Дмитриева Г.А. Биотехнология: курс лекций. Пущино, 1989. 96 с. 47. Клесов А.А. Применение иммобилизованных ферментов в пищевой промышленности//Биотехнология. М.: Наука, 1984. 48. Мартинек К. Иммобилизованные ферменты // Биотехнология. М.: Наука, 1984. 49. Методы культивирования клеток. Л.: Наука, 1988. 313 с. 50. Печуркин Н.С., Брильков А.В., Марченкова Т.В. Популяционные аспекты биотехнологии. Новосибирск: Наука, 1990. 173 с. 51. Пирузян Л.А., Михайловский Е.М. Сапротрофная микрофлора в качестве продуцента биологически активных веществ для целей микробной сапротрофной фармакотерапии // Изв. АН Серия биологическая, 1992. № 6. С. 860 - 866. 52. Реннеберг Р., Реннеберг И. От пекарни до биофабрики. М.: Мир, 1991. 112 с. 53. Рычков Р.С., Попов В.Г. Биотехнология перспективы развития // Биотехнология. М.: Наука, 1984. 54. Сассон А. Биотехнология: свершения и надежды. М.: Мир, 1987. 411 с. 55. Скрябин Г.К., Кощеенко К.А. Иммобилизованные клетки микроорганизмов // Биотехнология. М.: Наука, 1984. 56. Тривен М. Иммобилизованные ферменты. М.: Мир, 1983. 213 с. 27. 57. Хотянович А.В. Методы культивирования азотфиксирующих бактерий, способы получения и применение препаратов на их основе (методические рекомендации). Л., 1991. 60 с. 58. Шлегель Г. Общая микробиология. М.: Мир, 1987. 566 с. Рекомендуемые Web-сайты для поиска и самостоятельного изучения рассматриваемых в курсе тем: www.antibiotic.ru www.biengi.ac.ru www.bioinforum.ru www.biolinks.net.ru www.bioscience.ru 234 www.bio.1september.ru www.chem.ac.ru www.chshb.ru www.chemistry.narod.ru www.edu.ru www.informnauka.ru www.markovsky.virtualate.ney www.medline.ru www.medlinks.ru www.mednovosti.ru www.membrana.ru www.molbio.ru www.nature.ru www.rusbiotech.ru www.scientific.ru www.sciteclibrary.com www.washingtonprifile.com 235