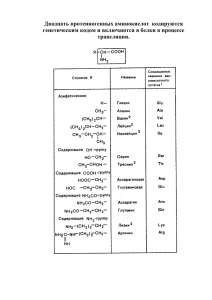
Лекция 4 Метаболизм аминокислот (часть 2) БИОСИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ Из 10 заменимых аминокислот 8 образуются из амфиболических метаболитов: Ala, Asp, Asn, Gly, Glu, Gln, Pro, Ser, а две (Суs и Туr) – из незаменимых аминокислот. Центральное место в биосинтезе аминокислот занимают глутаматдегидрогеназа, глутаминсинтаза и трансаминазы. Благодаря совместному действию этих ферментов катализируется включение неорганического иона аммония в α-аминогруппу аминокислот. Глутамат Восстановительное аминирование α-кетоглутарата катализируется глутаматдегидрогеназой. Данный фермент найден у всех организмов, однако наиболее важную роль он играет у животных. У растений и бактерий значительная часть глутамата синтезируется по другому механизму глутаматсинтетазой через промежуточное образование глутамина. Помимо того, что эта реакция приводит к образованию L-глутамата из амфиболического метаболита, α-кетоглутарата, она является ключевой стадией биосинтеза многих других аминокислот. Глутамин Биосинтез глутамина из глутамата катализируется глутаминсинтетазой. Рис. Структура глутаминсинтетазы. Наряду с предыдущей реакцией, образование глутамина является ключевым как в синтезе аминокислот, так и в утилизации токсичного соединения – аммиака. Данная реакция имеет как сходство с реакцией, катализируемой глутаматдегидрогеназой, так и отличия от нее. В обоих случаях «фиксируется» неорганический азот, который в одном случае включается в аминогруппу, а в другом — в амидную группу. Обе реакции сопряжены с сильно экзергоническими реакциями: в случае глутаматдегидрогеназы с окислением NAD(P)H, а в случае глутаминсинтетазы с гидролизом АТР. Данная реакция состоит из двух этапов: 1. Глутамат + ATP → γ-Глутамилфосфат + ADP γ-Глутамилфосфат + NH4+ → Глутамин + Рi + Н+ ----------------------------------------------------------------------Итог: Глутамат + ATP + NH4+ → Глутамин + ADP + Рi + Н+ 2. Рис. Синтез глутамина. Алании и аспартат L-аланин образуется из пирувата путем переаминирования с глутаматом, а L-аспартат – тем же путем из оксалоацетата. Ферменты аминотрансферазы (трансаминазы). Рис. Общий принцип функционирования аминотрансфераз Рис. Схема синтеза аланина. Рис. Схема синтеза аспартата. Перенос α-аминогруппы глутамата на амфиболические метаболиты иллюстрирует участие трансаминаз в процессах включения иона аммония в α-аминогруппы аминокислот. Дополнительно аланин может образовываться путем удаления β–карбоксигруппы аспартата (декарбоксилирования). Фермент аспартатдекарбоксилаза. Аспарагин Образование аспарагина из аспартата, катализируемое аспарагинсинтетазой, сходно с синтезом глутамина. Аспарагинсинтетаза млекопитающих в качестве источника азота использует не ион аммония, а глутамин и, следовательно, не «фиксирует» неорганического азота. Бактериальные же аспарагинсинтетазы используют ион аммония, следовательно, «фиксируют» неорганический азот. Как и в случае других реакций, сопровождающихся образованием РРi, последующий гидролиз РPi до Pi с участием неорганической пирофосфатазы обеспечивает энергетически благоприятные условия для протекания реакции. Особенность: донором азота в данной реакции выступает α-аминогруппа глутамата (а не амидная группа), поэтому образуется амид α-кетоглутарата – глутарамат. Важно! Выше рассмотренные реакции синтеза глутамата и аспартата носят обратимый характер (реакции дегидрогеназ и аминотрансфераз обратимы!), в то время как реакции синтеза глутамина и аспарагина (катализируются синтетазами) идут с высвобождением значительного количества энергии (распад ATP) и не обратимы. Серин Серин образуется из промежуточного продукта гликолиза – 3-фосфоглицерата. В первой реакции α-гидроксильная группа при участии NAD+ окисляется в кетогруппу (фермент 3-фосфоглицератдегидрогеназа). Далее в результате переаминирования образуется фосфосерин (фосфосерин-аминотрансфераза), который затем дефосфорилируется, образуя серин (фосфатаза). Также возможен вариант, когда сначала происходит дефосфорилирование 3фосфоглицерара до глицерата (фосфатаза), а затем уже идут реакции окисления (глицератдегидрогеназа) и переаминирования (серин-аминотрансфераза). Другим важным путем образования серина является его обратимый синтез из глицина. Донором метильной группы является N5,N10-метилен-тетрагидрофолат, переходящий в тетрагидрофолат. Гидроксильная группа серина поступает из воды. Реакция катализируется серин-гидроксиметилтрансферазой. А Б Рис. А – синтез серина из 3-фосфоглицерата; Б – реакция взаимопревращения серина и глицина. Рис. Структура тетрагидрофолата – одного из основных переносчиков одноуглеродных фрагментов в клетке (синим указаны атомы азота 5 и 10, к которым происходит присоединение одноуглеродных фрагментов – метильных, метиленовых, формильных и других групп). Глицин Синтез глицина в тканях млекопитающих осуществляется несколькими путями. В цитозоле печени содержится глицинтрансаминаза, катализирующая синтез глицина из глиоксилата, донором аминогруппы выступает глутамат (или аланин). В отличие от большинства реакций переаминирования равновесие этой реакций сильно смещено в направлении синтеза глицина. Рис. Синтез глицина из глиоксилата. Два важных дополнительных пути, функционирующие у млекопитающих, используют для образования глицина серин (см. синтез серина) и холин. Рис. Синтез глицина из холина. adoHcy – S-аденозилгомоцистеин; adoMet – S-аденозилметионин. В клетках печени хордовых существует еще один путь синтеза глицина, катализируемый ферментом глицинсинтетазой. Глицин образуется из неорганических соединений: CO2 и NH4+. Донором дополнительного одноуглеродного фрагмента выступает N5,N10-метилентетрагидрофолат, восстановление происходит за счет энергии NADH. Пролин У млекопитающих и некоторых других организмов пролин образуется из глутамата путем обращения реакций катаболизма пролина. Рис. Синтез пролина. Цистеин Цистеин, относящийся к заменимым аминокислотам, образуется из незаменимого метионина и заменимого серина. Сначала происходит превращение активация метионина с образованием S-аденозилметионина (см. лекцию 3), затем метилтрансфераза переносит метильную группу с S-аденозилметионина на какой-либо акцептор (тетрагидрофолат, диметилглицин, фосфатидилэтаноламин) и образуется S-аденозилгомоцистеин. Данное соединение распадается на аденозин и гомоцистеин. Далее гомоцистеин взаимодействует с серином. Получаемый в последней реакции α–кетобутират может подвергаться дальнейшему окислению с образованием пропионил-СоА. Тирозин Тирозин образуется из фенилаланина в реакции, катализируемой фенилаланингидроксилазой, поэтому фенилаланин относится к незаменимым аминокислотам, а тирозин – нет (при условии, что диета содержит достаточное количество фенилаланина). Реакция необратима, и поэтому тирозин не может заменить пищевой фенилаланин. Фенилаланингидроксилаза является оксигеназой со смешанной функцией, она имеется в печени млекопитающих и отсутствует в других тканях. В результате реакции один атом молекулярного кислорода включается в пара-положение фенилаланина (в виде гидроксильной группы), а другой восстанавливается, образуя воду. В качестве восстановителя (кофактора фермента) непосредственно в реакции участвует соединение тетрагидробиоптерин (воостановленная форма). В процессе реакции оно окисляется до дигидробиоптерина, восстановление которого в дальнейшем происходит за счет NADH. Биосинтез незаменимых аминокислот Ниже кратко описан биосинтез незаменимых аминокислот, осуществляемый в бактериях, грибах и растениях из глутамата, аспартата или других амфиболических метаболитов. В тканях млекопитающих эти реакции не происходят. Аргинин Аргинин – незаменимая пищевая аминокислота для человека в период его роста – может синтезироваться у крыс, но в таких количествах, которые не достаточны для нормального роста этих животных. В микроорганизмах идет биосинтез аргинина из глутамата с образованием Nацетилированных интермедиатов. Одним из них является N-ацетилглутамат-γполуальдегид, который служит также предшественником пролина у бактерий. У человека, а также у животных пролин образуется из глутамата. Аспартат является предшественником семейства аминокислот, в которое входят лизин, метионин, треонин и изолейцин. Метионин и треонин Аспартат сначала превращается (восстанавливается) в аспартат-β-полуальдегида, затем в гомосерин. Далее гомосерин может превратится в метионин (реакции обратные синтезу цистеина) или через гомосеринфосфат в треонин. Лизин Наиболее распространены два пути синтеза лизина: 1. Диаминопимелиновый путь (бактерии, низшие грибы, водоросли, высшие растения). При этом лизин образуется из аспартата, путем конденсации аспартат-βполуальдегида с пируватом. Образующийся при этом промежуточное соединение – диаминопимелат участвует также в образовании клеточной стенки бактерий. 2. Аминоадипиновый путь (некоторые низшие грибы, высшие грибы) Биосинтез лизина начинается с α-кетоглутарата и ацетил-СоА и проходит серию реакций, аналогичных реакциям цикла лимонной кислоты, катализируемых группой ферментов с несколько иной субстратной специфичностью. Лейцин, валин и изолейцин Исходными соединениями для синтеза лейцина является α– кетоизовалерьяновая кислота и ацетил-СоА, для валина – 2 молекулы пирувата, для изолейцина – треонин (то есть, изначально, аспартат). Следует отметить, что хотя лейцин, валин и изолейцин являются незаменимыми аминокислотами для человека, а также других высших животных, в тканях млекопитающих имеются трансаминазы, катализирующие обратимые взаимопревращения всех трех указанных аминокислот и соответствующих α-кетокислот. Этим объясняется способность соответствующих кетокислот заменять рассматриваемые аминокислоты в диете. Гистидин Гистидин подобно аргинину может быть назван полунезаменимым. Организм взрослого человека в течение некоторого периода может поддерживать азотистый баланс и в отсутствие гистидина. Растущий организм, однако, нуждается в гистидине. Процесс биосинтеза начинается с 5-фосфорибозил-1-пирофосфата (ФРПФ), который конденсируется с АТР с образованием N'-(5-фосфорибозил)-АТР. Эта реакция, следовательно, сходна с начальной стадией биосинтеза пуринов. Фенилаланин, триптофан. Исходными соединениями для синтеза фенилаланина, тирозина и триптофана у микроорганизмов и растений являются продукты углеводного обмена – эритрозо-4фосфат и фосфоенолпируват. В результате 7 реакций образуется соединение хоризмат, которое либо за счет нескольких превращений дает фенилаланин и тирозин, либо конденсируется с одной молекулой ФРПФ с дальнейшим првращением в триптофан.