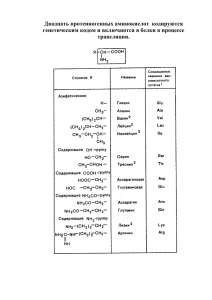
Тема лекции: Аминокислоты План 1. 2. 3. 4. Строение аминокислот Классификация аминокислот Свойства аминокислот Методы разделения и определения аминокислот 1.Строение аминокислот Первой аминокислотой, которая была выделена Бракконо в 1820 г из желатина, был глицин. Аминокислоты являются основными структурными компонентами белков. В настоящее время в природе обнаружено по разным источникам от 200 до 400 аминокислот, но в составе белков постоянно присутствует только 20 основных аминокислот. Остальные аминокислоты встречаются в живых организмах в свободном виде или в составе других соединений. Получить из белков аминокислоты можно путем кислотного, щелочного или ферментативного гидролиза. Общая формула аминокислоты Все аминокислоты за исключением пролина содержат карбоксильную группы, и отличаются друг от друга радикалом. амино- и 2.Классификация аминокислот. В зависимости от строения и свойств аминокислоты классифицируют на α-, β- и т.д. аминокислоты (по положению аминогруппы по отношению к α-углеродному атому) Все протеиногенные аминокислоты являются α-аминокислотами. протеиногенные (входят в состав белков), непротеиногенные (не входят в состав белков) К протеиногенным аминокислота относят 20 аминокислот: валин, лейцин, изолейцин, триптофан, лизин, метионин, треонин, фенилаланин, тирозин, гистидин, серин, глутаминовая кислота, глутамин, аспарагиновая кислота, аспарагин, пролин, цистеин, аргинин, глицин, аланин. по характеру бокового радикала: алифатические (ала, вал, лей, про, гли, иле), ароматические (фен, тир, три) серусодержащие (мет, цис), гетероциклические (три, про), содержащие в радикале карбоксильную группу (аспарагиновая кислота, глутаминовая кислота), содержащие в радикале аминогруппы (лизин, аргинин, гистидин, глутамин, аспарагин), содержащие в радикале гидроксогруппы (серин, тирозин, треонин). по кислотно-основным свойствам (кислые, основные, нейтральные). Кислыми аминокислотами являются глутаминовая и аспарагиновая кислоты. Основными аминокислотами являются лизин, аргинин, гистидин. по абсолютной конфигурации (L- и D-стереоизомеры) По абсолютной конфигурации выделяют L- и D-формы. Различия между изомерами связаны с взаимным расположением четырех замещающих групп, находящихся в вершинах воображаемого тетраэдра, центром которого является атом углерода в α-положении. Имеется только два возможных расположения вокруг него. На рисунке представлены две возможные конформации тетраэдра. L- и D- формы аланина В живых организмах в составе белков обнаружены только L-аминокислоты. по оптической активности на право- и левовращающие. Наличие в аминокислоте ассиметричного α-углеродного атома (хирального центра) делает возможным только два расположения химических групп вокруг него. Это приводит к особому отличию веществ друг от друга, а именно изменению направления вращения плоскости поляризации поляризованного света, проходящего через раствор. Величину угла поворота измеряют с помощью поляриметра. В соответствии с углом поворота выделяют правовращающие (+) и левовращающие(-) изомеры. Деление на L- и D- формы не соответствует делению на право- и левовращающие. Для одних аминокислот L-формы (или D- формы) являются правовращающими, для других – левовращающими. Например, L-аланин – правовращающий, а L-фенилаланин левовращающий. При смешивании L- и Dформ одной аминокислоты образуется рацемическая смесь, не обладающая оптической активностью. L- и D- формы аминокислот могут самопроизвольно неферментативно превращаться в рацемическую смесь, что может быть использовано для определения возраста людей, животных, ископаемых остатков. Известно, что L-аспарагиновая кислота содержится в дентине (белок твердой эмали зубов). Скорость рацемизации этой аминокислоты составляет 0,01% в год. У детей в зубах содержится только L- аспарагиновая кислота. Если определить содержание D-аспарагиновой кислоты, то можно определить возраст. по способности синтезироваться в организме (незаменимые и заменимые). В организме не могут синтезироваться 8 аминокислот: валин, лейцин, изолейцин, триптофан, лизин, метионин, треонин, фенилаланин. Остальные аминокислоты относятся к заменимым. 3.Свойства аминокислот. Амфотерность. Все аминокислоты являются амфотерными соединениями, поскольку способны проявлять свойства как кислот, так и оснований за счет наличия карбоксильных и амногрупп. В водных растворах аминокислоты проявляют свойства и кислоты и основания – по карбоксильной группе отдают протон и становятся отрицательно зараженными (свойства кислоты), а по аминогруппе присоединяют протон и становятся положительно заряженными (свойства основания). При определенном значении рН аминокислоты могут иметь нулевой заряд. Это состояние называется изоэлектрическим, а значение рН – изоэлектрической точкой (ИЭТ). Значение ИЭТ зависит от строения радикала аминокислоты. Для большинства аминокислот ИЭТ располагается в кислой среде и близкой к нейтральной. Основные аминокислоты имеют ИЭТ в щелочной среде. Если рН ниже ИЭТ, то аминокислота заряжается положительно, а если выше – то отрицательно. р1= ½(2.34+9,69)=6,01 р1= ½ (2,09+3,86)=2,77 Аминокислоты способны соединяться с образованием пептидной связи. Пептидные связи образуются за счет отщепления гидроксогруппы от карбоксильной группы одной аминокислоты и атома водорода аминогруппы другой аминокислоты Свойства пептидной связи: 1. За счет сопряжения π-электронов двойной связи и свободной пары электронов атома азота пептидная связь имеет характер частично двойной связи. За счет этого длина пептидной связи короче, чем одинарной, и длиннее двойной связи 2. Атомы, входящие в пептидную группу, располагаются в одной плоскости. Вращение вокруг пептидной связи невозможно. Пептидная цепь может вращаться только вокруг α – углеродных атомов. 3.Атомы водорода и кислорода пептидной группы располагаются в трансположении по отношению к пептидной связи. Радикалы также располагаются по обе стороны цепи 4.Атомы водорода и кислорода могут образовывать водородные связи с другими пептидными группами 5. Методы разделения и определения аминокислот Кислотно-основные свойства аминокислот используются для их разделения с помощью метода ионообменной хроматографии. Для ИОХ используются специальные смолы, имеющие прочно связанные заряженные группы. Смолы называют катионообменными, если они содержат анионные группы, и анионообменными, если содержат катионные группы. Для разделения аминокислот чаще всего используют катионообменные смолы. Этими смолами заполняют колонки, пропускают буферный раствор, содержащий положительно заряженные ионы натрия. Ионы натрия связываются с анионными группами смолы. Потом на колонку наносят кислый раствор смеси аминокислот. Положительно заряженные аминокислоты будут вытеснять ионы натрия и связываться с частицами смолы. Прочность связывания аминокислот с анионными группами смолы зависит от заряда аминокислот. Прочнее всего связываются лизин, аргинин, гистидин. Аспарагиновая и глутаминовая кислоты связываются слабее всего. Другие аминокислоты имеют промежуточную прочность связи. Элюцию (вымывание) аминокислот ведут с помощью буферного раствора с возрастающим значением рН. Скорость прохождения аминокислот через колонку будет различна. Менее прочно связанные аминокислоты будут быстро вымываться (асп, глу), потом будут выходить промежуточные аминокислоты и медленнее всего будут вымываться лиз, арг, гис. Элюат собирают небольшими порциями и в нем определяют аминокислоты. Этот процесс в настоящее время автоматизирован. Прибор называется аминокислотный анализатор. Количественное определение аминокислот ведут по реакции с нингидрином, с которым аминокислоты дают фиолетовую окраску, и только пролин - желтую Тема лекции: Белки План 1. 2. 3. Строение и структура белков Свойства белков Классификация белков, характеристика отдельных представителей 1.Строение белков. Белки – высокомолекулярные соединения, состоящие из аминокислот, соединенных пептидными связями. Содержание белков в живых организмах различается. Так, в организме человека воды содержится до 60%, 40% - сухое вещество, из которого 20% приходится на долю белков, 10% - жиры и жироподобные вещества, 1% углеводов, 5% минеральные вещества, 4% - все прочие соединения. В растительных организмах белков значительно меньше, за исключением бобовых культур. Количество аминокислот в составе белков может сильно варьировать от нескольких десятков до нескольких сот, следовательно, молекулярная масса белков также различна. Белки с молекулярной массой до 10000Да называют пептидами, с большей – белками. Важное значение для свойств и функций белков имеет последовательность аминокислот, которую называют первичной структурой. Предположение о том, что аминокислоты в белках соединены пептидными связями, впервые было высказано в 1888 году Данилевским, но доказано в 1904 г Эмилем Фишером. Для определения первичной структуры используются разные методы. 1.Метод Сенгера 2.Гидразиновый метод 3.Метод Эдмана 4.Использование ферментов Основные этапы определения первичной структуры белков 1. 2. 3. 4. 5. 6. 7. 8. Получение чистого препарата белка. Раскрытие дисульфидных мостиков Определение концевых аминокислот с помощью ферментов Гидролиз белка ферментами для получения более коротких пептидов Гидролиз пептидов до аминокислот и их определение Определение концевых аминокислот у каждого пептида Длинные пептиды расшифровывают с помощью метода Эдмана Сопоставление полученных результатов и реконструкция полипептидной цепи Уровни структурной организации белков Первичная структура – это линейная цепь аминокислот, связанных между собой пептидной связью. • Вторичная структура – это способ укладки элементов первичной структуры в пространстве с образованием простейших структур: α-спираль, β-конформации, неупорядоченный клубок. На виток спирали приходится 3,6 аминокислотных остатка. Спираль у природных белков закручена вправо. α-спираль Поллинг и Кори предложили модель -спирали. Конфигурация имеет винтовую симметрию. Витки спирали регулярны. Спираль образована за счет внутримолекулярных водородных связей, обусловленных взаимодействием функциональных групп, входящих в состав пептидной связи. Спираль стабилизируется за счет внутрицепочечных водородных связей СО и NH группы 4-го за ним аминокислотного остатка. При расчете денной модели было показано, что расстояние между спиралями равно 0,54 нм. Ограничения на существование спирали. на участке не может быть остатков аминокислот, имеющих большие углеводородные радикалы; не может быть много заряженных аминокислот; не может быть остатков пролина. Примером может являться -керотин (основные аминокислоты являются глицин, аланин, цистеин). Цистеин участвует в формировании цистина за счет образования -SS- связей. β-конформации Была изучена на фиброине. Поллинг и Кори показали модель складчатости. -конформация стабилизируется за счет образования межцепочечных водородных связей. Расстояние между соседними полипептидными связями 0,7 нм. полипептидные цепи при конформации укладываются либо параллельно, либо антипараллельно. Ограничения те же, что и при -спирали. Β состав фиброина шелка, паутины входят β- кератины, у которых в основном антипараллельная ориентация, а α- спирали белков волос расположены в одном направлении. Третичная структура белка Третичная структура белка Это способ укладки доменов и элементов вторичной структуры в пространстве. образуется за счет межмолекулярных взаимодействий отдельных аминокислотных остатков. Структура стабилизируется за счет: водородные связи формируются между аминокислотными остатками имеющими полярные группы(ОН, NH2,…); вандервальсовы связи или гидрофобные взаимодействия образуются между ароматическими аминокислотами и аминокислотами, имеющими большое количество углеводородных радикалов; цистеиновые мостики или S-S связи образуются за счет димеризации 2х молекул цистеина в молекулу цистина; электростатические взаимодействия образующиеся за счет взаимодействия заряженных аминокислот. Четвертичная структура белка Характерна для олигомерных белков (состоят из нескольких полипептидных цепей). это способ укладки отдельных полипептидных цепей относительно друг друга. Структура стабилизируется за счет: гидрофобные взаимодействия электростатические взаимодействия образование ковалентных связей водородные связи ван-дер-ваальсовы силы. Пример: гемоглобин(22), т.е. 2 -субъединицы и 2 -субъединицы Четвертичная структура белка Если все субъединицы одинаковые, то это гомоолигомерные белки. Если разные гетероолигомерные белки. Гемоглобин гетеротетромер. Схематичное изображение четвертичной структуры гемоглобина Гемоглобин одновременно может переносить 4 молекулы кислорода. Присоединение очередной молекулы кислорода к гемоглобину облегчает присоединение следующей. 2. Свойства белков: Белки проявляют кислотно-основные свойства, являются буферами, заряжены и могут перемещаться в электрическом поле, имеют ИЭТ, способны к высаливанию и денатурации, способны к набуханию, растворимы в воде и других растворителях. Имея высокую молекулярную массу, растворы белков вязкие, имеют незначительную диффузию, низкое осмотическое давление, обладают свойствами коллоидных расторов, могут рассеивать свет и не проходят через полупроницаемые мембраны. Белки способны поглощать свет в разных участках спектра. В видимой области свет поглощают окрашенные белки, благодаря чему их можно идентифицировать. В ультрафиолете белки поглощают свет при 280 нм за счет содержания в них ароматических аминокислот. Белки, как и аминокислоты, являются оптически активными соединениями. В изоэлектрической точке белки имеют нулевой заряд. При рН выше ИЭТ белки становятся отрицательно заряженными, а при рН ниже ИЭТ – положительно заряженными. В формировании пространственной структуры белка важную роль выполняет вода как полярный растворитель. Благодаря ей гидрофобные радикалы аминокислот в основном ориентированы внутрь белковой глобулы, а полярные и заряженные располагаются на поверхности. Таким образом, гидратная оболочка стабилизирует белковую структуру. Одним из методов очистки, разделения и оценки гомогенности белков является электрофорез, основанный на способности белков перемещаться с разной скоростью в электрическом поле в зависимости от заряда, молекулярной массы и формы белковой молекулы. Нанесенный на линию старта суммарный белок при электрофорезе разделяется на фракции, которые можно идентифицировать. Денатурация белков – это нарушение пространственной структуры белков, кроме первичной. Денатурация происходит при нагревании, при механическом воздействии, действии кислот, щелочей, ряда солей и др. Денатурация может быть обратимой, когда структура восстанавливается. Структура белка, которую он имеет в физиологических условиях, называется нативной. 3.Классификация белков Классификация белков по составу Простые белки (неконъюгированные). Состоят только из аминокислот. -керотин, фиброин, коллаген, эластин Сложные белки (конъюгированные). Содержат небелковые группы: гем, ионы металлов, нуклеиновые кислоты, липиды, сахара, флавины. Классификация по простетическим группам гемапротеины (простетическая группа гем). Гемоглобин, цитохром.; металлопротеины (простетическая группа металл(Mg, Mn, Fe)). Ферритин, алкогольдегидрогеназа; липопротеины (простетическая группа липид). липопротеин крови; гликопротеины (простетическая группа углеводная часть). Рецепторы, -глобулины; фосфопротеины (простетическая группа фосфатная группа).казеин; нуклеопротеины (простетическая группа нуклеиновый кислотный остаток). Рибонуклеопротеины; флавопротеины (простетическая группа флавин). Сукцинатдегидрогеназа. Состав нуклеопротеинов – нуклеосом, состоящих их ДНК и белков гистонов. Важная роль этих белков – участие в хранении и передаче наследственной информации. В составе фосфопротеинов обнаружены остатки фосфорной кислоты, которые в основном присоединены к белку через гидроксо-группы серина, треонина, тирозина. Особенно богат остатками фосфорной кислоты казеин – белок молока, с которым в организм поступает важный для формирования костной ткани фосфор. Липопротеины имеют сложный состав. Липидная часть может быть представлена триглицеридами, полициклическими спиртами (холестеролом и его эфирами). Клеточная мембрана образована фосфолипидами, состоящими из жирных кислот, глицерина, фосфорной кислоты, азотистых оснований. Белки в составе мембраны в основном выполняют транспортные функции, как и липопротеины крови. Гликопротеины включают в свой состав углеводный компонент – глюкозу, галактозу и другие. Как видно из рисунка, они аминированы и по аминогруппе ацетилированы. К белку присоединяются по гидроксо- и амино-группам аминокислот. Гликопротеины обнаружены в покровных тканях, в слизях, где они выполняют в основном защитные функции. Обнаружены они и в составе нервных тканей и рецепторов. Гем Витамин В2 (рибофлавин) Хромопротеины представлены тремя группами окрашенных белков: Гемопротеины - они в своем составе содержат гем-группы. Транспорт кислорода выполняют гемоглобин, миоглобин, легоглобин (в основном красно-коричневого цвета). Цитохромы осуществляют перенос электронов в дыхательных цепях, в фотосинтезе, участвуют в обезвреживании токсичных веществ. Каталаза обезвреживает пероксид водорода. Флавопротеины (желтые) – они содержат в составе витамин В2 и участвуют в окислительно- восстановительных процессах. Родопсин- белок сетчатки глаза, содержит витамин А (ретинол), участвует в зрительном акте. В состав белков часто входят металлы, которые участвуют в стабилизации структуры белка, непосредственно принимают участие в каталитическом процессе и т.д. Их выделяют в группу металлопротеинов, например, белок ферритин, который содержит много железа, необходимого для синтеза других железосодержащих белков, таких как гемоглобин крови. Классификация по пространственной структуре Фибриллярные. Для них характерна линейная структура. Плохо растворимы в воде, обычно в их состав входят большое количество гидрофобных аминокислот. Глобулярные. Для них характерна сложная пространственная трехмерная структура. Имеют гидрофобное ядро и гидрофильную поверхность. Как правило, хорошо растворимы в воде. Нпр., яичный альбумин. Глобулярные белки имеют форму глобулы, отношение длины осей у них менее 10. У вытянутых белков – форма фибриллы, отношение длины осей у них более 10. Классификация белков по функциям. 1. Каталитическая. Это белки-ферменты, которые катализируют реакции в клетках. К ним относятся пепсин, трипсин, каталаза, липаза, амилаза, уреаза и множество других 2. Строительная (пластическая). Эту функцию выполняют многие белки, входящие в состав тканей организма: мышечные – актин, миозин, альбумины, глобулины; белки костей, кожи и множество других. 3. Защитная. Это белки иммунной системы – иммуноглобулины, белки покровных тканей насекомых – гликопротеины, муцины, входящие в состав слизистых тканей. 4. Транспортная. Эту функцию выполняют гемоглобины, переносящие кислород, липопротеины, которые переносят липиды. 5. Сократительные белки. Это белки мышечной ткани актин, миозин и другие. 6. Регуляторные белки. К ним относят гормоны пептидной природы – инсулин, глюкагон, соматотропный гормон, рецепторные белки. 7. Светочувствительные белки. Это белок сетчатки глаза родопсин. Кроме указанных классификаций можно рассматривать классификацию по происхождению, по аминокислотному составу, по растворимости. Лекция 3. Ферменты План 1.Сравнение биологических и небиологических катализаторов 2.Строение ферментов. Активный и аллостерический центры ферментов 3.Свойства ферментов 4.Механизм действия ферментов 5.Регуляция активности ферментов 6.Изоферменты. Многоферментные комплексы 7.Номенклатура и классификация ферментов 1. Сравнение биологических и небиологических катализаторов Ферментами, или энзимами, называются белки, обладающие каталитической активностью. К настоящему времени установлено, что практически все энзимы являются белками. На сегодняшний день известно около 20 тысяч ферментов. Сотни ферментов удалось выделить из живых клеток, очистить и получить в кристаллическом виде. Некоторые из них синтезированы в лабораторных условиях из соответствующих аминокислот (например, инсулин, рибонукеаза). Ферментативные процессы использовались человеком еще с глубокой древности. Получение вина и уксуса, изготовление сыра и хлеба, выделка шкур и кож известны с античных времен. Пастер показал, что сбраживание сахаров происходит под действием живых организмов, но в 1897 году братья Бюхнеры доказали, что сок, выделенный из клеток дрожжей, также способен сбраживать сахар с образованием спирта и углекислого газа, тем самым было доказано, что ферменты могут катализировать реакции вне клетки. В 1926 году Самнер выделил в кристаллическом виде фермент уреазу. В 1930–36 годах Нортроп и Кунитц выделили в кристаллическом виде пепсин, трипсин и химотрипсин. Для всех выделенных ферментов была доказана белковая природа. Все катализаторы ускоряют лишь термодинамически возможные реакции, не изменяют направление реакции Все катализаторы понижают энергетический барьер реакции, но ферменты понижают его более значительно. Так, энергетический барьер реакции разложения пероксида водорода без катализатора составляет 75,6 кдж/моль, в присутствии платины – 49,14 кдж/моль, а каталаза понижает этот барьер до 5,46 кдж/моль. Все катализаторы не смещают равновесие реакции, а лишь способствуют его быстрейшему достижению Все катализаторы ускоряют как прямую, так и обратную реакцию, если она обратима Ферменты по сравнению с другими катализаторами проявляют более высокую активность и специфичность действия. Так, каталаза за 1 минуту способна разложить 5. 106 молекул пероксида водорода При ферментативных реакциях практически не образуется побочных продуктов Все ферменты в отличие от других катализаторов имеют белковую природу Скорость ферментативных реакций, как и с участием других катализаторов, зависит от рН, температуры, активаторов, ингибиторов, но температурный и рН оптимум лежит в интервале, где не происходит денатурации белка Ферментативный запрограммирован процесс пространственно организован и 2.Строение ферментов Ферменты бывают простые и сложные. Простые состоят только из белковой части, а сложные содержат добавочную группу в виде кофермента (легко отделяется от белка) или простетической группы (прочно связана с белком) В настоящее время употребляются следующие названия составных частей ферментов Фермент в целом Белковая часть Добавочная группа холофермент Апофермент Простетическая группа Кофермент Апофермент каталитически не активен. Незначительной активностью обладает кофермент. Только в результате соединения кофермента с апоферментом возникает высокая каталитическая активность у фермента-протеида, так как в этом случае образуется активный центр фермента. В ферменте выделяют активный центр, куда присоединяется субстрат (это вещество, преобразование которого ускоряет фермент), и где идет сам каталитический процесс У некоторых ферментов есть аллостерический центр, который находится в другом месте фермента. Такие ферменты являются регуляторными. Центры образованы радикалами аминокислот с участием добавочных групп 3.Свойства ферментов Влияние рН Большинство ферментов максимальную активность проявляет вблизи нейтральных значений. Ряд ферментов (пепсин) активны в сильно кислой среде а такие, как аргиназа, в щелочной Оптимум рН для ферментов: пепсин-1,5-2,5, амилаза солода -4,7-5,2, трипсин – 7,8, каталаза – 7,6, аргиназа- 9,5-9,9, панкреатическая амилаза – 6,4-7,2. Влияние температуры Скорость ферментативной реакции возрастает в 2-3 раза при повышении температуры на каждые 10 градусов (правило Вант-Гоффа), но поскольку ферменты являются белками, при температуре выше 50-60 °С начинается процесс денатурации, и активность падает Влияние активаторов и ингибиторов Активаторы ускоряют реакции. Например, ионы кальция и хлора ускоряют гидролиз крахмала амилазой Ингибиторы, наоборот, замедляют катализ. Ингибиторы бывают обратимые, при удалении которых процесс восстанавливается, и необратимые, полностью ингибирующие фермент. Необратимый ингибитор – диизопропилфторфосфат, который необратимо блокирует серин в активном центре холинэстеразы. Аспирин может необратимо блокировать циклооксигеназу, ключевой фермент синтеза простагландинов В свою очередь обратимые ингибиторы делятся на конкурентные и неконкурентные Пример конкурентного ингибирования. Субстрат (сукцинат) и ингибитор (малонат) конкурируют за активный центр сукцинатдегидрогеназы. Увеличение концентрации субстрата понижает ингибирование. Конкурируют за активный центр алкогольдегидрогеназы этанол и метанол Сульфаниламид может конкурентно блокировать синтез фолиевой кислоты, что бывает важно при лечении некоторых бактериальных инфекций Неконкурентный ингибитор действует не на активный центр, а на другой участок фермента, но увеличение концентрации субстрата не понижает ингибирование. Примером неконкурентного ингибирования является действие цианидов, которые блокируют железо в составе цитохромов, тяжелых металлов. 4.Специфичность ферментов - важнейшее свойство, определяющее способность фермента «узнавать» свой субстрат. Различают несколько типов специфичности. Абсолютная спцифичность характерна для ферментов, у которых только один субстрат. Например, уреаза катализирует только гидролиз мочевины Групповая специфичность характерна для ферментов, способных узнавать тип химической связи. К ним относят липазы, катализирующие гидролиз сложноэфирных связей в составе триглицеридов, амилазы, катализирующие гидролиз гликозидных связей в составе полисахаридов. Специфичность в отношении изомеров проявляют изомеразы, которые превращают вещества в их изомеры. Они «узнают» оптические изомеры, геометрические изомеры и другие. Аспартаза катализирует только преобразование фумаровой кислоты, а не ее геометрического изомера малеиновой кислоты Активность ферментов и способы ее определения и выражения. Активность ферментов – это скорость преобразования субстрата при определенных условиях. Для определения активности ферментов используют разные методы, которые позволяют определить количество преобразованного субстрата или образованного продукта в единицу времени при данных условиях: спектрофотометрические, титриметрические, газометрические, вискозиметрические и другие. Например, активность каталазы можно определить газометрическим методом по количеству выделившегося кислорода в единицу времени, а активность амилазы по изменению вязкости крахмала в единицу времени. Активность ферментов выражают в международных единицах (МЕ). 1 МЕ соответствует такому количеству фермента, которое катализирует превращение 1 мкмоль субстрата за 1 минуту при оптимальных условиях. Активность ферментов выражают также в каталах. 1 катал соответствует такому количеству фермента, которое превращает 1 моль субстрата за 1 секунду при оптимальных условиях. Под удельной активностью понимают количество превращенного субстрата (мкмоль) за 1 минуту на один мг белка. Чем выше удельная активность, тем чище белок. Скорость реакции зависит от концентрации субстрата. При определенной концентрации субстрата достигается максимальная скорость реакции, которая может понизиться за счет ингибирующего действия продуктов реакции и других причин. Скорость реакции линейно зависит от концентрации фермента 4. Механизм действия ферментов Механизм действия ферментов основан на образовании промежуточных фермент-субстратных комплексов. Реакция обычно идет в несколько стадий Согласно Э.Фишеру, структура активного центра присоединении субстрата (ключ-замок) не меняется при По теории Кошланда (индуцированное соответствие) активный центр фермента изменяется при присоединении субстрата (рука-перчатка). Выделяют несколько стадий в ферментативном катализе: 1.сближение фермента и субстрата; 2.ориентация субстрата в активном центре фермента; 3.образование промежуточных комплексов; 4.образование комплекса ферментпродукт; 5. высвобождение продуктов реакции. 5.Регуляция активности ферментов осуществляется разными способами. 1.Повышение или понижение концентрации субстрата влияет на активность фермента Скорость реакций цикла Кребса зависит от концентрации щавелевоуксусной кислоты, являющейся катализатором цикла 2.Активность ферментативной реакции зависит от концентрации фермента, синтез которого в клетке может изменяться 3.Некоторые ферменты синтезируются в неактивной форме. Переход в активное состояние происходит за счет частичного протеолиза. Это характерно для пищеварительных ферментов, гидролизующих белки. Благодаря такой особенности не расщепляются собственные белки организма 4.Аллостерическая регуляция Продукт В стимулирует фермент Е2, а продукты С и Х аллостерически подавляют активность фермента Е2. Стимулятором данной реакции является АМФ, а АТФ подавляет активность этого фермента 5. Белок-белкововое взаимодействие Неактивная протеинкиназа состоит из 2 регуляторных (R) и 2 каталитических субъединиц (C). цАМФ может присоединяться к регуляторным субъединицам, высвобождая каталитические, в результате чего протеинкиназа переходит в активное состояние Активация аденилатциклазы. 6.Ковалентная модификация Ковалентная модификация осуществляется путем присоединения или отщеплении какой-либо химической группировки. Это может быть остаток фосфорной, уксусной кислоты, метильная группа. Присоединение или отщепление группы меняет активность фермента 6. Кинетика ферментативных реакций Общие принципы кинетики химических реакций применимы и к ферментативным реакциям. Однако у последних имеется одна отличительная особенность, не свойственная обычным реакциям, протекающим в отсутствии ферментов. Эта особенность – насыщение субстратом. V [S] Влияние концентрации субстрата на скорость ферментативных реакций. При низкой концентрации субстрата скорость реакции V возрастает пропорционально концентрации субстрата, то есть в отношении субстрата реакция имеет первый порядок. По мере увеличения концентрации субстрата скорость реакции возрастает все медленнее, пропорциональность нарушается. В этой зоне мы имеем реакцию дробного порядка. При дальнейшем увеличении концентрации субстрата скорость реакции становится постоянной и не зависит от концентрации субстрата. В этой области в отношении субстрата реакция приобретает нулевой порядок. Происходит как бы насыщение субстратом. При этих условиях фактором, лимитириующим скорость реакции, становится концентрация фермента. Исследование эффекта насыщения привело Л. Михаэдиса и М. Ментен в 1913 году к созданию общей теории действия ферментов и ферментативной кинетики. Михаэлис и Ментен предложили, что фермент E на первой стадии реагирует с субстратом S с образованием фермент-субстратного комплекса ES, который находится в равновесии со свободным ферментом и субстратом: E+S K+1 K-1 ES ферментсубстратный комплекс К + 1, К – 1 – константы скорости прямой и обратной реакций Тогда V1 = K + 1 [E] [S] V2 = K - 1 [ES] Если V1 = V2 , то K + 1[E] [S] = K - 1 [ES], т.е. [ES] K-1 = [E] [S] K+1 = Ks ОШИБКА в уравнении! КS – константа диссоциации фермент-субстратного комплекса, или субстратная константа. Так как концентрация фермент-субстратного комплекса изменяется вследствие перехода последнего в продукт реакции Р с регенерацией свободного фермента: K+2 ES E+P где К + 2 – константа скорости второй стадии реакции, то первоначальное выражение после дополнения будет иметь вид: E+S K+1 K+2 ES E+P K-1 (1) Отношение констант всех трех реакций называется константой Михаэлиса К М. KM = K-1+K+2 K+1 КМ всегда несколько выше по числовому значению, чем КS. Константа Михаэлиса более полно характеризует состояние фермент-субстратного комплекса. Так как последняя стадия реакции практически необратима, то есть E+P ES то ею можно пренебречь. Тогда общая концентрация фермента в системе [Et] равна [Et] = [E] + [ES] где [E] – концентрация свободного фермента, [ES] – концентрация ферментсубстратного комплекса. Отсюда скорость реакции V = K + 2 [ES] Это, по существу, уравнение скорости для реакции (1), однако им нельзя воспользоваться для расчетов, поскольку ни К + 2, ни [ES] не могут быть непосредственно измерены. Поэтому используют максимальную скорость: Vmax = K + 2 [Et] Ее можно определить, поскольку Vmax не может превышать той скорости, которая наблюдается при полном насыщении фермента субстратом. Используя указанные величины можно вывести уравнение Михаэлиса-Ментен: Vmax [S] V= KM + [S] т.е. скорость реакции пропорциональна концентрации субстрата в данный момент времени. Чем больше значение КМ, тем слабее связь между ферментом и субстратом. КМ характеризует сродство фермента к субстрату. Однако кинетическое поведение большинства ферментов значительно сложнее, чем это вытекает из идеализированной схемы, лежащей в основе уравнения МихаэлисаМентен. При выводе этого уравнения полагали, что существует только один ферментсубстратный комплекс. В действительности в большинстве ферментативных реакций образуется по меньшей мере два или три таких комплекса, возникающих в определенной последовательности: E+S ES EZ EP E+P Кроме того, в большинстве ферментативных реакций участвует более одного субстрата, и образуется два или большее число продуктов и т.д. В таких реакциях много промежуточных стадий. E I II P (продукты) Кривые, иллюстрирующие изменения внутренней энергии Е в процессе реакций, протекающих в одну стадию (I) и через несколько метастабильных промежуточных соединений (II). 7.Изоферменты. Многоферментные комплексы. Изоферменты Креатинкиназа представлена тремя формами, каждая состоит из 2 субъединиц - мышечной (М) и мозговой (В). Субъединицы незначительно отличаются по первичной структуре. Изоформы отличаются сродством к субстрату. КК-1 локализована в мозге, КК-2 – в миокарде, КК-3 – в скелетных мышцах Лактатдегидрогеназа представлена пятью формами, состоящими из четырех субъединиц. Н-сердечная, М- мышечная. ЛДГ-1 и ЛДГ-2 характерны для тканей с аэробным метаболизмом и превращают лактат в пируват. ЛДГ-4 и ЛДГ-5, наоборот, работают при анаэробных условиях, превращая пируват в лактат. ЛДГ-3 активна в тканях с промежуточным метаболизмом. Многоферментные комплексы. В таких комплексах ферменты собраны в ансамбли, реакции становится субстратом для другого фермента. относится пируватдегидрогеназный комплекс, синтетаза кислот. Такие пространственно организованные комплексы ферментативные реакции. причем продукт одной К таким комплексам высших карбоновых значительно ускоряют 8..Номенклатура и классификация ферментов Энзимология долгое время не располагала научной номенклатурой ферментов. Наименования ферментам давали: 1. По случайным признакам (тривиальная номенклатура); 2. По названию субстрата (рациональная); 3. По химическому составу фермента; 4. По типу катализируемой реакции и характеру субстрата. Тривиальные названия: пепсин (от греч. pepsis – пищеварение), трипсин (от греч. tripsis – разжижаю), папаин (от названия дынного дерева papaja, из сока которого он выделен) и т.д. Наибольшее распространение получила рациональная номенклатура. Здесь принято добавлять суффикс «аза» к части названия атакуемого субстрата; например, уреаза действует на мочевину (urea), аргиназа - на аргинин, рибонуклеаза – на РНК и т.д. В 1961 году Международным биохимическим съездом в Москве была принята единая научная номенклатура и классификация ферментов. Классификация ферментов основана на типе катализируемой реакции. Всего выделяют 6 классов ферментов. Каждый фермент имеет собственный шифр или кодовый номер, в котором первое число означает класс фермента, второе - подкласс, третье – подподкласс, а четвертое – номер фермента в подподклассе. 1 класс – оксидоредуктазы. Они катализируют окислительно- восстановительные реакции 2 класс – трансферазы. Они катализируют реакции переноса различных групп 3 класс – гидролазы. Они катализируют гидролиз веществ. 4 класс – лиазы. Они катализируют негидролитические реакции с высвобождением низкомолекулярных веществ или их присоединением по двойным связям. 5 класс – изомеразы. Они катализируют превращение веществ в их изомеры. 6 класс – синтетазы (лигазы). Они катализируют синтетические реакции с использованием энергии макроэргических соединений По научной номенклатуре названия ферментам даются следующим образом: Тема лекции : ВИТАМИНЫ Витамины - это низкомолекулярные органические вещества разнообразного строения. Объединены в одну группу по следующим признакам: 1. Витамины абсолютно необходимы организму и в очень небольших количествах. 2. Витамины не синтезируются в организме и должны поступать извне или синтезироваться микрофлорой кишечника. Витамины играют одинаковую роль во всех формах жизни, но высшие животные утратили способность к их синтезу. Например, аскорбиновая кислота (витамин ”С”) не синтезируется в организмах человека, обезьян и морской свинки, так как в процессе эволюции была утеряна ферментная система синтеза этого витамина из глюкозы. АВИТАМИНОЗ - это заболевание, которое развивается при полном отсутствии того или иного витамина в организме. В настоящее время авитаминозы обычно не встречаются, а бывают ГИПОВИТАМИНОЗЫ при недостатке витамина в организме. ПРИЧИНЫ РАЗВИТИЯ ГИПО- И АВИТАМИНОЗОВ Все причины можно разделить на внешние и внутренние. ВНЕШНИЕ причины гиповитаминозов: 1. Недостаточное содержание витамина в пище (при неправильной обработке пищи, при неправильном хранении пищевых продуктов) 2. Состав рациона питания (например, отсутствие в рационе овощей и фруктов) 3. Не учитывается потребность в том или ином витамине. Например, при белковой диете возрастает потребность в витамине “РР” (при обычном питании он может частично синтезироваться из триптофана). Если человек потребляет много белковой пищи, то может увеличиться потребность в витамине “В6“ и снизиться потребность в витамине РР. 4. Социальные причины: урбанизация населения, питание исключительно высокоочищенной и консервированной пищей; наличие антивитаминов в пище. Социальные причины развития авитаминозов существуют в мире. Например, в отдаленных районах Севера, в рационе людей мало овощей и фруктов. Урбанизация также имеет значение, т.к. в пищу потребляется много консервированных и рафинированнных продуктов. В крупных городах люди недостаточно обеспечены солнечным светом - поэтому может быть гиповитаминоз Д. ВНУТРЕННИЕ причины гиповитаминозов: 1. Физиологическая повышенная потребность в витаминах, например, в период беременности, при тяжелом физическом труде. 2. Длительные выздоровления; тяжелые инфекционные заболевания, а также период 3. Нарушение всасывания витаминов при некоторых заболеваниях ЖКТ, например, при желчнокаменной болезни нарушается всасывание жирорастворимых витаминов; 4. Дисбактериоз кишечника тоже имеет значение, так как некоторые витамины синтезируются полностью микрофлорой кишечника (это витамины В 3, Вc, В6, Н, В12 и К); 5. Генетические дефекты некоторых ферментативных систем. Например, витамин Д-резистентный рахит развивается у детей при недостатке ферментов, участвующих в образовании активной формы витамина Д (1,25-диоксихолекальциферола). КЛАССИФИКАЦИЯ ВИТАМИНОВ 1. Водорастворимые витамины. К этой группе относят витамины С, Р, В 1, В2, В3, ВC, В6, В12, РР, Н. 2. Жирорастворимые витамины: А, Д, Е, К. Большинство водорастворимых витаминов должно поступать регулярно с пищей, т.к. они быстро выводятся или разрушаются в организме. Жирорастворимые витамины могут депонироваться в организме. Кроме того, они плохо выводятся, поэтому иногда при избытке жирорастворимых витаминов наблюдаются ГИПЕРВИТАМИНОЗЫ - заболевания, связанные с интоксикацией организма высокими дозами жирорастворимых витаминов. Такие заболевания описаны для витаминов А и Д. Для большинства витаминов известно, что их производные входят в состав коферментов и простетических групп ферментов. Для некоторых витаминов (витамин С) точно известно, в каких реакциях они участвуют, но коферментная функция пока не открыта. ЖИРОРАСТВОРИМЫЕ ВИТАМИНЫ ВИТАМИН “А” ( ретинол, антиксерофтальмический) Наиболее ранний и специфический признак гиповитаминоза А - гемералопия ("куриная слепота") нарушение сумеречного зрения. Возникает из-за недостатка зрительного пигмента - родопсина. Родопсин содержит в качестве активной группы ретиналь (альдегид витамина А) - находится в палочках сетчатки. Эти клетки (палочки) воспринимают световые сигналы низкой интенсивности. РОДОПСИН = опсин (белок) + цис-ретиналь. При возбуждении родопсина светом, цис-ретиналь, в результате ферментативных перестроек внутри молекулы переходит в полностью-трансретиналь (на свету). Это приводит к конформационной перестройке всей молекулы родопсина. Родопсин диссоциирует на опсин и транс-ретиналь, что является пусковым механизмом, возбуждающим в окончаниях зрительного нерва импульс, который затем передается в мозг. В темноте, в результате ферментативных реакций транс-ретиналь вновь превращается в цис-ретиналь и, соединяясь с опсином, образует родопсин. Витамин А также влияет на процессы роста и развития покровного эпителия. Поэтому при авитаминозе наблюдается поражение кожи, слизистых оболочек и глаз, которое проявляется в патологическом ороговении кожи и слизистых. У больных развивается ксерофтальмия - сухость роговой оболочки глаза, т.к. происходит закупорка слезного канала в результате ороговения эпителия. Так как глаз перестает омываться слезой, которая обладает бактерицидным действием, развиваются конъюнктивиты, изъязвление и размягчение роговицы -кератомаляция. При авитаминозе А может быть также поражение слизистой ЖКТ, дыхательных и мочеполовых путей. Нарушается устойчивость всех тканей к инфекциям. При развитии авитаминоза в детстве - задержка роста. В настоящее время показано участие витамина А в защите мембран клеток от окислителей - т.е. витамин А обладает антиоксидантной функцией. Витамин А запасается в печени. Пищевые источники - печень морских рыб и млекопитающих, желток яиц, цельное молоко, рыбий жир. Овощи и фрукты красно-оранжевого цвета (томаты, морковь и др.) содержат много каротина - водорастворимого предшественника витамина А, имеющего в молекуле 2 иононовых кольца. В настоящее время, гиповитаминоз А наблюдается у людей с заболеваниями кишечника, поджелудочной железы, при нарушении желчевыделительной функции печени, то есть при заболеваниях, при которых нарушается всасывание жира. Высокие дозы витамина А могут приводить к токсическим эффектам. Характерные проявления гипервитаминоза - воспаление глаз, гиперкератоз, выпадение волос, диспептические явления. Суточная потребность в витамине А - 1-2.5 мг, в каротине - в 2 раза больше. ВИТАМИН Д (холекальциферол, антирахитный) Сам витамин Д не обладает витаминной активностью, но он служит предшественником 1,25дигидрокси-холекальциферола (1,25дигидроксивитамина Д3). Синтез активной формы протекает в два этапа - в печени присоединяется оксигруппа в положении 25, а затем в почках оксигруппа в положении 1. Из почек активный витамин Д 3 переносится в другие органы и ткани - главным образом в тонкий кишечник и в кости, где витамин Д участвует в регуляции обмена Са и Р. Недостаток витамина Д приводит к развитию нарушений фосфорно-кальциевого обмена и процессов окостенения. В результате у детей развивается рахит, связанный с недостатком Са и Р. Характерные признаки рахита - остеомаляция ("размягчение" костей запаздывание окостенения), запаздывание закрытия родничков, деформации грудной клетки, позвоночника, конечностей. У таких детей снижен мышечный тонус, наблюдается раздражительность, потливость, выпадение волос. У взрослых при недостатке витамина Д наблюдается остеопороз - разрежение костной ткани в результате вымывания солей кальция из скелета. Потребность в витамине Д повышается у беременных. При благоприятных условиях витамин Д может синтезироваться в организме человека из предшественника - 7-дегидрохолестерина под действием ультрафиолетовых лучей (фотохимическая реакция) в результате разрыва связи в кольце В. Пищевые источники - рыба, рыбий жир, печень, сливочное масло, желток яиц. Суточная доза витамина Д3 - 10-20 мкг. Высокие дозы витамина Д (выше 1,5 мг в сутки) крайне токсичны. При гипервитаминозе кроме интоксикации наблюдается отложение гидроксиапатита в некоторых внутренних органах (кальцификация почек, кровеносных сосудов). ВИТАМИН К (филлохинон). Витамин К необходим для нормального синтеза протромбина (фактор II) предшественника одного из белков системы свертывания - тромбина. Тромбин это фермент, который катализирует реакцию превращения фибриногена в фибрин - основу кровяного сгустка при активации системы светрывания крови. При недостатке витамина К синтезируется дефектная молекула протромбина и ряда других факторов свертывания крови. Причина - нарушение ферментативного карбоксилирования глутаминовой кислоты, необходимой для связывания Са2+ белками системы свертывания. Основное проявление недостаточности - нарушение свертывания крови, в результате которого происходят самопроизвольные паренхиматозные и капиллярные кровотечения. Авитаминоз, как правило связан с нарушением выделения желчи в ЖКТ (при желчнокаменной болезни). Пищевые источники - ягоды рябины, капуста, арахисовое масло и др. растительные масла. Витамин К также синтезируется микрофлорой кишечника, поэтому одна из причин гиповитаминозов при недостатке витамина в пище дизбактериоз кишечника (например, при антибиотикотерапии). Если больной страдает гиповитаминозом К, например, при некоторых видах желтух, то операции - даже удаление зуба - могут сопровождаться длительным кровотечением. Синтезирован водорастворимый аналог витамина К - викасол, который используют при лечении гиповитаминозов, связанных с нарушением всасывания витамина К из кишечника. Известны природные антивитамины К - например, ДИКУМАРИН, САЛИЦИЛОВАЯ кислота, которые применяют при лечении тромбозов, т.к. антивитамины К способны снижать количество протромбина в крови. Суточная потребность точно не установлена, т.к. витамин синтезируется микрофлорой. Считают, что в сутки потребность около 1 мг. ВИТАМИН Е (токоферол, витамин размножения). Является антиоксидантом. При недостаточности витамина Е - дегенеративные изменения в печени, нарушение функций биологических мембран. Витамин Е предохраняет липиды клеточных мембран от окисления активными формами кислорода. Авитаминоз проявляется при очень длительном голодании или при стойком нарушении желчевыделительной функции печени (нарушение всасывания жиров). При этом наблюдаются шелушение кожи, мышечная слабость, стерильность - нарушением функции размножения. Поскольку витамин Е широко распространен в природе (растительные масла, семена пшеницы и др. злаков, сливочное масло), то авитаминоз встречается редко. Суточная потребность - около 10-30 мг. ВИТАМИН “С” (аскорбиновая кислота, антицинготный, антискорбутный) В 1932 г. впервые выделен из сока лимона, через два года искусственно синтезирован. Важное свойство - способность аскорбиновой кислоты легко окисляться. Биологическая роль витамина “С” (связана с его участием в окислительновосстановительных реакциях) 1. Витамин С, являясь сильным восстановителем, играет роль кофактора в реакциях окислительного гидроксилирования, что необходимо для окисления аминокислот пролина и лизина в оксипролин и в оксилизин в процессе биосинтеза коллагена. Коллаген может синтезироваться и без участия витамина С, но такой коллаген не является полноценным (не формирутся его нормальная структура). Поэтому при недостатке витамина С ткани, содержащие много коллагена, становятся непрочными, ломкими. В первую очередь нарушается структура стенок сосудов, повышается их проницаемость, наблюдаются кровоизлияния под кожу и под слизистые оболочки. 2. Участвует в синтезе стероидных гормонов надпочечников. 3. Необходим для всасывания железа. 4. Участвует в неспецифической иммунной защите организма. Авитаминоз “С” - цинга. Проявления цинги: болезненность, рыхлость и кровоточивость десен, расшатывание зубов, нарушение целостности капилляров подкожные кровоизлияния, отечность и болезненность суставов, нарушение заживления ран, анемия. Иногда цинга развивается у новорожденных на искусственном вскармливании пастеризованным молоком, в которое не добавлен витамин С. В основе всех изменений при цинге, за исключением анемии, лежит нарушение синтеза коллагена. Анемия связана с нарушением всасывания железа. В настоящее время цинга не распространена, но весной у многих людей наблюдается недостаток (гиповитаминоз) витамина “С”, что проявляется, например, повышенной утомляемостью, понижением иммунитета. Основные источники витамина “С”: свежие зеленые овощи и фрукты. Следует помнить, что витамин С легко разрушается при нагревании, особенно в щелочной среде в присутствии кислорода, ионов железа и меди. Хорошо сохраняется в кислой среде (в квашеной капусте, в клюкве, в ягодах черной смородины и плодах шиповника). При длительном хранении овощей и фруктов содержание в них витамина “С” уменьшается. Источником витамина С является также хвоя ели и сосны. Суточная потребность - около 100 мг в сутки. Лечебная доза - до 1-2 г в сутки. ВИТАМИН “Р” (рутин, витамин проницаемости) Биологическая роль - стабилизация основного вещества соединительной ткани, путем ингибирования фермента гиалуронидазы. При недостатке витамина Р у людей повышается проницаемость кровеносных сосудов, которое сопровождается кровоизлияниями и кровотечениями. Витамин Р усиливает действие витамина С (снижает потребность в нем) Пищевые источники: зеленые овощи и фрукты, кожура лимона. Суточная потребность - не установлена В И Т А М И Н Ы Г Р У П П Ы "B" ВИТАМИН B1 (тиамин, антиневритный) Производное вит.В1 - ТДФ (ТПФ) является коферментом пируватдегидрогеназного комплекса (фермента пируваткарбоксилазы), альфакетоглутаратдегидрогеназного комплекса и фермента транскетолазы (фермента альфа-тотаратдекарбоксилазы), а также входит в состав кофермента транскетолаз - ферментов неокислительного этапа ГМФ-пути.. При недостаточности вит.В1 может возникнуть болезнь "бери-бери", характерная для тех стран Востока, где основным продуктом питания служит очищенный рис и кукуруза. Для этого заболевания характерна мышечная слабость, нарушение моторики кишечника, потеря аппетита, истощение, периферический неврит (характерный признак - человеку больно вставать на стопу - больные ходят “на цыпочках”), спутанность сознания, нарушения работы сердечно-сосудистой системы. При "бери-бери" повышается содержание пирувата в крови. Пищевые источники витамина В1 - ржаной хлеб. В кукурузе, рисе, пшеничном хлебе витамин В1 практически отсутствует. Это объясняется тем, что в зерне ржи тиамин распределен по всему зерну, а в других злаках он содержится только в оболочке зерен. Суточная потребность - 1.5 мг/сутки. ВИТАМИН В2 (рибофлавин) Витамин В2 входит в состав флавинмононуклеотида (ФМН) и флавинадениндинуклеотида (ФАД) простетических групп флавиновых ферментов. Его биологическая функция в организме - участие в окислительно-восстановительных реакциях в составе флавопротеидов (ФП). Недостаточность этого витамина часто встречается в России. Особенно часто бывает у людей, которые не употребляют в пищу черный ржаной хлеб. Проявление гиповитаминоза: ангулярные дерматиты в углах рта (“заеда”), глаз. Часто это сопровождается кератитами (воспаление роговицы). В очень тяжелых случаях бывает анемия. Очень часто сочетаются сочетанные гиповитаминозы витаминов "В2" и "РР",так как эти витамины содержатся в одних и тех же продуктах. Пищевые источники: ржаной хлеб, молоко, печень, яйца, овощи желтого цвета, дрожжи. Суточная потребность: 2-4 мг/сутки. ФОЛИЕВАЯ КИСЛОТА (ВC) В составе 3 структурных единицы: птеридин, ПАБК (парааминобензойная кислота) и глутаминовая кислота. Часто ПАБК (парааминобензойную кислоту) тоже называют витамином. Но это неверно. ПАБК - это фактор роста для микроорганизмов, которые синтезируют фолиевую кислоту. Активный С1 извлекается из глицина или серина с помощью фермента, в небелковой части которого содержится витамин Вc - фолиевая кислота. Фолиевая кислота два раза восстанавливается в организме (к ней присоединяется водород). ТГФК является коферментом ферментов, переносящих одноуглеродные радикалы. Из метилен-ТГФК могут образовываться все другие формы активного С1: формилТГФК, метил-ТГФК, метен-ТГФК, оксиметил-ТГФК в результате реакций окисления или восстановления метилен- ТГФК. Фолиевая кислота в виде тетрагидрофолиевой кислоты является коферментом, участвующим в ферментативных реакциях, связанных с переносом активных одноуглеродных радикалов. Например: биосинтез пуриновых и пиримидиновых мононуклеотидов. При авитаминозе у человека наблюдается макроцитарная анемия, при которой нарушен синтез ДНК в клетках красного костного мозга, для больных характерна потеря веса. Пищевые источники: зеленые листья овощей, дрожжи, мясо, шпинат. Авитаминозы встречаются редко, так как потребность в этом витамине компенсируется за счет микрофлоры кишечника. При некоторых заболеваниях кишечника, когда возникают дисбактериозы, нарушается всасывание фолиевой кислоты. Суточная потребность: 0.2 - 0.4 мг. ВИТАМИН В6 (пиридоксин) В6 в форме пиридоксальфосфата является простетической группой трансаминаз и декарбоксилаз аминокислот. Он необходим и для некоторых реакций обмена аминокислот. Поэтому при авитаминозе В6 наблюдаются нарушения обмена аминокислот. В6 также участвует в реакциях синтеза гема гемоглобина (синтез d-аминолевулиновой кислоты). Поэтому при недостатке В6 у человека развивается анемия. Кроме анемии, наблюдаются дерматиты. Недостаток В6 может развиться у больных туберкулезом, потому что этих больных лечат препаратами, синтезированными на основе изониазида - это антагонисты витамина В6. Пищевые источники: ржаной хлеб, горох, картофель, мясо, печень, почки. Суточная потребность взрослого человека: 0.15-0.20 мг. ПАНТОТЕНОВАЯ КИСЛОТА (витамин В3) Молекула пантотеновой кислоты состоит из бета-аланина и 2,4-дигидроксидиметил-масляной кислоты. Формулу знать необязательно. Важность этого витамина в том, что он входит в состав HS-KoA (кофермента ацилирования). Строение КоА: а) тиоэтиламин б) пантотеновая кислота в) 3-фосфоаденозин5-дифосфат. HSКоА - кофермент ацилирования, то есть входит в состав ферментов, которые катализируют перенос ацильных остатков. Поэтому В3 участвует в бета-окислении жирных кислот, окислительном декарбоксилировании альфа-кетокислот, биосинтезе нейтрального жира, липоидов, стероидов, гема, ацетилхолина. При недостатке пантотеновой кислоты при дисбактериозе у человека развиваются дерматиты, в тяжелых случаях - изменения со стороны желез внутренней секреции, в том числе надпочечников. Также наблюдается депигментация волос, истощение. Пищевые источники: яичный желток, печень, дрожжи, мясо, молоко. Суточная потребность: 10мг/сут. ВИТАМИН В12 (кобаламин) (антианемический витамин) Имеет сложное строение, структура молекулы похожа на гем, но вместо железа - кобальт. В состав В12 входит также нуклеотидная структура, похожая на АМФ. Производное витамина В12 является коферментом. Этот витамин необходим для синтеза нуклеиновых кислот. Он обеспечивает переход оксирибонуклеотидов в дезоксирибонуклеотиды (РНК в ДНК). Недостаток этого витамина может привести к развитию злокачественной тромбоцитарной анемии, нарушениям функции центральной нервной системы. Как правило, встречается сочетанный недостаток витамина В12 и фолиевой кислоты. Анемия развивается не потому, что В12 мало поступает с пищей, а при отсутствии особого гликопротеина, который называется "внутренний фактор Кастла" и вырабатывается в желудке. Фактор Кастла необходим для всасывания витамина В12. При удалении части желудка, гастритах уменьшается выработка фактора Кастла. Это единственный кишечника. витамин, который синтезируется только микрофлорой Это единственный водорастворимый витамин, который депонируется в организме (в печени). Суточная потребность: 2.5-5 мкг. ВИТАМИН РР (антипеллагрический) Химическое название: никотинамид. Входит в состав НАД и НАДФ, то есть входит в состав коферментов никотинамидных дегидрогеназ. Его роль - участие в окислительно-восстановительных реакциях. При недостатке РР развивается пеллагра. При пеллагре наблюдаются три “Д”: - дерматит - диарея - деменция (поражение центральной нервной системы) Источники РР: мясо, бобовые, орехи, рыба и вообще продукты, богатые белком. Витамин РР может частично синтезироваться из триптофана. Если человек съедает много белковой пищи, то потребность в этом витамине снижается. Из 60 гр. белка может синтезироваться 1 мг витамина РР. Суточная потребность: 15-25 мг/сутки. ВИТАМИН “Н” (БИОТИН) В составе молекуы биотина имеются имидазоловое и тиоэфирное кольца, к ним присоединен радикал - валериановая кислота. Витамин Н входит в состав ферментов карбоксилаз: Ацетил-КоА-карбоксилазы, пируваткарбоксилазы и других. Всасыванию биотина в кишечнике препятствует овидин - белок, содержащийся в сырых яйцах. При термической обработке яиц происходит днатурация овидина. При авитаминозе наблюдаются Синтезируется микрофлорой кишечника. дерматиты, поражения ногтей, анемия. АВИТАМИНОЗЫ, СВЯЗАННЫЕ С НЕДОСТАТКОМ ФОЛИЕВОЙ КИСЛОТЫ (Вc), ПАНТОТЕНОВОЙ КИСЛОТЫ (В3), БИОТИНА (Н), ПИРИДОКСИНА (В6), КОБАЛАМИНА (В12) ВСТРЕЧАЮТСЯ ОЧЕНЬ РЕДКО, ПОСКОЛЬКУ ЭТИ ВИТАМИНЫ, ТАК ЖЕ, КАК И ВИТАМИН К, СИНТЕЗИРУЮТСЯ МИКРОФЛОРОЙ КИШЕЧНИКА. АВИТАМИНОЗЫ НАБЛЮДАЮТСЯ ПРИ ДИСБАКТЕРИОЗЕ КИШЕЧНИКА, ПРИ НЕОБЫЧНОЙ ДИЕТЕ ИЛИ ПРИ НАРУШЕНИИ ВСАСЫВАНИЯ ИЗ КИШЕЧНИКА. Лекция Нуклеиновые кислоты. План 1.Химический состав нуклеиновых кислот 2.Нуклеозиды, нуклеотиды 3.Структура нуклеиновых кислот 4.Свойства нуклеиновых кислот. 5.Функции нуклеиновых кислот Нуклеиновые кислоты были выделены Мишером в 1868 году из ядер клеток гноя. Поскольку они были найдены в ядрах клеток, то и название получили «нуклеиновые кислоты». Состав нуклеиновых кислот, нуклеотидов и нуклеозидов приведен на схеме. Нуклеиновые кислоты → Нуклеотиды Нуклеотиды → Азотистые основания D-рибоза ( D-дезоксирибоза) Фосфорная кислота Нуклеозиды → Азотистые основания D-рибоза ( D-дезоксирибоза) Нуклеиновые кислоты находятся в биологическом материале в комплексе с белками. Обработка 10% раствором хлорида натрия высвобождает нуклеиновые кислоты. После удаления осадка нуклеиновые кислоты из раствора осаждают этанолом, ТХУ, хлороформом, фенолом. Дальнейшую очистку ведут хроматографией, электрофорезом. При полном кислотном гидролизе нуклеиновые кислоты распадаются на составные части – азотистые основания, сахара и фосфорную кислоту. Азотистые основания. Различают пуриновые и пиримидиновые основания В составе нуклеиновых кислот встречаются так называемые минорные основания. Они являются производными пурина и пиримидина и отличаются наличием других боковых радикалов. Особенно много минорных оснований в тРНК Сахара Фосфорная кислота Н3РО4 2.Нуклеозиды, нуклеотиды Нуклеозиды содержат в своем составе азотистое основание и остаток сахара Нуклеотиды содержат в своем составе азотистое основание, остаток сахара и фосфорной кислоты. Фосфорная кислота может присоединяться как по 5', так и по 3' углеродному атому сахара К нуклеотидам относят также АТФ, ГТФ, ЦТФ, ТТФ, УТФ. АТФ – универсальный источник энергии, который используется для выполнения мышечной работы, для синтеза и транспорта веществ, для передачи генетической информации. АТФ содержит по 5' углеродному атому три остатка фосфорной кислоты, два из которых присоединяются через макроэргические связи. При гидролизе АТФ высвобождается более 30 кдж/ моль энергии. Нуклеотид входит в состав кофермента А, участвующего в активации и транспорте карбоновых кислот В энергетическом обмене важную функцию выполняет кофермент НАД – никотинамидадениндинуклеотид, который может присоединять и отдавать два атома водорода На схеме приведено уравнение реакции восстановления пировиноградной кислоты до молочной кислоты, которая является заключительной реакцией молочно-кислого брожения Различают два типа нуклеиновых кислот ДНК и РНК. Они различаются по химическому составу, структуре и функциям. В состав ДНК входят тимин, аденин, гуанин, цитозин, дезоксирибоза, фосфорная кислота. В состав РНК входят урацил, аденин, гуанин, цитозин, рибоза, фосфорная кислота. Структура нуклеиновых кислот Первичная структура. Под первичной структурой нуклеиновых кислот понимают последовательное расположение нуклеотидов, соединенных между собой фосфодиэфирными связями. Сокращенно первичную структуру нуклеиновых кислот изображают как последовательность азотистых оснований, указывая нумерацию углеродных атомов, содержащих свободные гидроксогруппы 5' –ААТГГЦТАГЦЦГТТ- 3' - одна цепь ДНК 5'–ГГУЦЦАГУЦЦГААГ -3' - цепь РНК Вторичная структура нуклеиновых кислот ДНК Вторичная структура ДНК в виде двойной спирали была расшифрована в 1953 году Дж. Уотсоном и Ф.Криком, которые использовали результаты экспериментальных исследований М.Уилкинса, Р.Франклин и Э.Чаргаффа. Э.Чаргаффом были установлены закономерности, которые впоследствии получили название правила Чаргаффа. Согласно этим правилам 1.Количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина: А=Т, Г=Ц. 2.Сумма пуриновых оснований равна сумме пиримидиновых оснований А+Г=Ц+Т 3.Химический состав ДНК не меняется с возрастом и не зависит от среды 4. Химический состав ДНК одинаков в клетках одного вида организмов, а у разных видов разный. Молекула ДНК состоит из двух цепей, спирально закрученных относительно друг друга Цепи ДНК антипараллельны. Диаметр спирали 2 нм, расстояние между витками – 3,4 нм, на один виток спирали приходится 10 пар нуклеотидов. Цепи друг другу комплементарны: аденин комплементарен тимину, и между ними образуется две водородные связи, а гуанин – цитозину, между этими основаниями образуется три водородных связи. Комплементарные пары располагаются в плоскости практически перпендикулярно оси спирали. Важную функцию в стабилизации вторичной структуры ДНК выполняют «стекингвзаимодействия» гидрофобные взаимодействия, возникающие между расположенными друг над другом комплементарными парами азотистых оснований. Эти взаимодействия направлены примерно параллельно оси спирали. Существует несколько структурных форм, в которых может находиться ДНК. Каждая из них предназначена для выполнения определенных функций. Так, форма А предназначена для передачи информации от ДНК к РНК (транскрипция). Форма В связана с репликацией ДНК, форма С предназначена для хранения ДНК, Форма Z – левая спираль, а форма SBS используется при биосинтезе ДНК. Вторичная структура РНК РНК является одиночной цепью, и лишь на отдельных участках она может быть заспирализована сама на себя, образуя петли. В РНК комплементарными являются А и У, Г и Ц. Третичная структура нуклеиновых кислот ДНК – очень большая молекула. Общая длина ДНК в организме человека достигает 2 метров. Молекулярная масса ДНК составляет порядка 106-109Да. Поэтому ДНК плотно упакована, образуя суперспираль. Большую роль в формировании пространственной структуры ДНК играют белки гистоны. По мере упаковки в нуклеосомы, хроматин, хромосомы достигается уплотнение в 1,2.104 раз. Третичная структура РНК. Существует несколько видов РНК – матричная (м-РНК), транспортная (т-РНК), рибосомная (р-РНК), малая ядерная (мя-РНК). Первой была изучена пространственная структура ала- т-РНК в 1965 году Холли. Было показано, что пространственная структура т-РНК напоминает клеверный лист. Каждая петля получила свое название. 1 петля – дигидроуридиловая. 2 петля – антикодоновая. 3 петля – псевдоуридиловая. 4 – акцепторный конец. 5 – антикодон. Все другие т-РНК имеют схожую структуру. Третичная структура м-РНК может иметь вытянутую форму, форму клубка, спирали. р-РНК входит в структуру рибосомы. мя-РНК участвует в удалении интронов. Свойства нуклеиновых кислот ДНК белого цвета, волокнистого строения, плохо растворима в воде, хорошо растворяется в растворах солей щелочных металлов, в растворах других солей. Растворы ДНК имеют высокую вязкость, обладают двойным лучепреломлением, являются оптически активными соединениями. По своей химической природе нуклеиновые кислоты относятся к кислотам, поскольку способны отдавать протоны по остаткам фосфорной кислоты, следовательно, в щелочной среде заряжаются отрицательно и могут перемещаться в электрическом поле (электрофорез). При нагревании водородные связи в нуклеиновых кислотах разрушаются. Плавление, денатурация ДНК происходит в интервале от 80 до 90 градусов, вязкость раствора уменьшается и возрастает поглощение в УФ (260 нм). Наблюдается гиперхромный эффект. Чем больше пар Г-Ц, тем выше температура плавления. При охлаждении происходит отжиг, т.е. ренатурация. Химически нуклеиновые кислоты достаточно инертны, тем не менее подвергаются дезаминированию, алкилированию, связываются с металлами, полиаминами. Молекулярная масса нуклеиновых кислот различна. Так, для ДНК она может достигать порядка 106-109Да, для м-РНК – 106 Да, для р-РНК – 106Да, для т-РНК – 17-35 тысяч Да Функции нуклеиновых кислот. ДНК выполняет функцию хранения и передачи последовательности ее нуклеотидов закодирована в белке. Каждая аминокислота кодируется подвергается репликации - удвоению, благодаря дочерние клетки из поколения в поколение. наследственной информации. В последовательность аминокислот триплетом нуклеотидов. ДНК чему информация передается в м-РНК при транскрипции снимает информацию с ДНК и служит впоследствии матрицей для синтеза белка рРНК образует структуру рибосомы, где происходит синтез белка. т-РНК является переносчиком аминокислот, когда идет синтез белка. мя-РНК принимает участие в удалении интронов из м-РНК после транскрипции. Тема лекции Липиды План 1.Общая характеристика липидов 2.Классификация липидов. Простые и сложные липиды 1.Общая характеристика липидов. Под липидами понимают вещества органического происхождения, плохо растворяющиеся в воде (гидрофобные), и хорошо растворяющиеся в органических растворителях – хлороформе, бензоле, эфире. В состав липидов могут входить спирты, высшие жирные кислоты, азотистые основания, углеводы, фосфорная кислота и др. Имея разнообразное строение, липиды способны выполнять многочисленные функции. -в химических связях липидов заключено много энергии, которая высвобождается при их распаде. Эта энергия расходуется на выполнение работы, синтез веществ, транспорт, передачу генетической информации и др. -липиды являются структурными компонентами клеточных мембран; -липиды выполняют защитные функции; - некоторые липиды могут участвовать в регуляции метаболизма; - липиды способны запасаться в организме. 2.Классификация липидов. Все липиды можно разделить на омыляемые, которые подвергаются гидролизу, и неомыляемые, не подвергающиеся гидролизу. Омыляемые липиды 2. Среди омыляемых липидов можно выделить простые и сложные липиды. Простые липиды, как правило, двухкомпонентные. К ним относят триглицериды, воска, стериды. К сложным липидам относят фосфолипиды, гликолипиды, сфинголипиды, орнитинолипиды. Триглицериды – сложные эфиры высших карбоновых кислот и трехатомного спирта глицерина. СН2ОН-СНОН-СН2ОН Глицерин СН3 –СН2 - СН2- СН2- СН2- СН2- СН2- СН2- СН2- СН2 -СН2 -СН2 -СН2 -СН2 -СН2 -СН2СН2 –СООН Стеариновая кислота СН3 –СН2 - СН2- СН2- СН2- СН2- СН2- СН2- СН2- СН2 -СН2 -СН2 -СН2 -СН2 -СН2 – СООН Пальмитиновая кислота СН3 –СН2 - СН2- СН2- СН2- СН2- СН2- СН2- СН = СН -СН2 -СН2 -СН2 -СН2 -СН2 – СН2 СН2 СООН Олеиновая кислота Триглицериды составляют основную часть жиров. В жире помимо триглицеридов обнаружены в небольших количествах другие вещества: белки, витамины минеральные элементы, вода и др. В составе жирных кислот чаще всего четное число углеродных атомов, хотя встречаются жирные кислоты с нечетным числом углеродных атомов. Существуют предельные, насыщенные жирные кислоты и непредельные, ненасыщенные кислоты. Число двойных связей в кислотах может быть различным: от одной до шести. К насыщенным кислотам относят пальмитиновую, стеариновую, а к ненасыщенным олеиновую, линолевую, линоленовую, арахидоновую. Последние три кислоты в организме человека не синтезируются и поэтому их относят к витаминам – витамин F. В состав триглицеридов могут входить как насыщенные, так и ненасыщенные кислоты. В составе твердых жиров больше насыщенных кислот, а в составе растительных масел больше ненасыщенных кислот. Как правило, в состав жиров входят цис-формы жирных кислот. Триглицериды Тристеарин Триглицериды подвергаются щелочному и ферментативному гидролизу. В результате ферментативного гидролиза триглицеридов образуются глицерин и свободные жирные кислоты. Гидролиз осуществляют ферменты липазы. В результате щелочного гидролиза образуются глицерин и соли жирных кислот, которые называются мылами. Калиевые соли обычно образуют жидкое мыло, а натриевые – твердое. Для характеристики жиров определяют константы жиров. Кислотным числом называют количество мг гидроксида калия, которое требуется для нейтрализации жирных кислот, содержащихся в 1 грамме жира. Кислотное число характеризует качество жира. Чем это число больше, тем хуже качество. Оно свидетельствует о том, что жир получен из незрелых семян, или долго хранился в неподходящих условиях Числом омыления называют количество мг гидроксида калия, которое требуется для нейтрализации свободных и связанных жирных кислот, содержащихся в 1 грамме жира. Это число характеризует количество связанных и свободных жирных кислот. Эфирным числом называют количество мг гидроксида калия, которое требуется для нейтрализации связанных жирных кислот, содержащихся в 1 грамме жира. Это число определяют по разности между числом омыления и кислотным числом. Эфирное число характеризует количество связанных жирных кислот э.ч. = ч.о - к.ч. Иодным числом называют количество г йода, которое необходимо для присоединения по двойным связям непредельных кислот, содержащихся в 100 г жира. Иодное число характеризует содержание в жире непредельных кислот. Основная функция триглицеридов – энергетическая и запасная. 1 г жира может дать 38 кдж энергии Воска. Воска – сложные эфиры высокомолекулярных спиртов и высших карбоновых кислот. Спирты могут быть одно- или двухатомными алифатического ряда, а также ароматическими. СН3- ( СН2)14 – СО-О-СН2 –(СН2)28 –СН3 - пальмитиновомирициловый эфир, который входит в состав пчелиного воска. К воскам относят ланолин (овечий воск), пчелиный воск, воска, покрывающие листья и плоды. Эти вещества обладают выраженными гидрофобными свойствами, благодаря чему их основная функция – защитная. Стериды. Стериды – сложные эфиры циклических спиртов и высших карбоновых кислот Сложные липиды. В состав сложных липидов входят помимо спиртов и карбоновых кислот другие вещества Фосфолипиды. В их состав входят глицерофосфолипиды, содержащие в качестве спирта глицерин, и глицеросфинголипиды, в которых спирт – сфингозин. В составе глицерофосфолипидов также обнаружены высшие карбоновые кислоты, азотистые основания (серин, холин, аминоэтиловый спирт) и фосфорная кислота. Наличие в структуре заряженных групп и гидрофобных радикалов жирных кислот позволяет глицерофосфолипидам участвовать в построении клеточных мембран, что является их важнейшей функцией. Щелочной гидролиз фосфатидилэтаноламина Глицеросфинголипиды в свой состав включают спирт сфингозин Глицеросфинголипид К неомыляемым липидам относят те, которые не подвергаются гидролизу. Среди них витамины, гормоны, холевые кислоты. холестерин и другие вещества ЛИПОПРОТЕИНЫ Липопротеины - это сферические частицы, в которых можно выделить гидрофобную сердцевину, состоящую из триглицеридов (ТРГ) и эфиров холестерина (ЭХС) и амфифильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки. Белки оболочки называются апобелками. Холестерин (ХС) обычно занимает промежуточное положение между оболочкой и сердцевиной. Компоненты частицы связаны слабыми типами связей и находятся в состоянии постоянной диффузии – способны перемещаться друг относительно друга. Основная роль липопротеинов – транспорт липидов, поэтому обнаружить их можно в биологических жидкостях. При изучении липидов плазмы крови оказалось, что их можно разделить на группы, так как они отличаются друг от друга по соотношению компонентов. У разных липопротеинов наблюдается различное соотношение липидов и белка в составе частицы, поэтому различна и плотность. Липопротеины разделяют по плотности методом ультрацентрифугирования, при этом они не осаждаются, а всплывают (флотируют). Мерой всплывания является константа флотации, обозначаемая Sf (сведберг флотации). В соответствии с этим показателем различают следующие группы липопротеинов: Липопротеины можно разделить и методом электрофореза. При классическом щелочном электрофорезе разные липопротеины ведут себя по-разному. При помещении липопротеинов в электрическое поле хиломикроны остаются на старте. ЛОНП и ЛПП можно обнаружить во фракции пре-глобулинов, ЛНП - во фракции -глобулинов, а ЛВП - -глобулинов: Определение липопротеинового спектра плазмы крови применяется в медицине для диагностики атеросклероза. Все эти липопротеины отличаются по своей функции. 1. Хиломикроны (ХМ) - образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном - в жировую ткань), а также транспорт экзогенного холестерина из кишечника в печень. 2. Липопротеины Очень Низкой Плотности (ЛОНП) - образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань. 3. Липопротеины Низкой Плотности (ЛНП) - образуются в кровеносном русле из ЛОНП через стадию образования Липопротеинов Промежуточной Плотности (ЛПП). Их роль: транспорт эндогенного холестерина в ткани. 4. Липопротеины Высокой Плотности (ЛВП) - образуются в печени, основная роль - транспорт холестерина из тканей в печень, то есть удаление холестерина из тканей, а дальше холестерин выводится с желчью. При определении содержания в крови липопротеинов различной плотности их обычно разделяют методом электрофореза. При этом ХМ остаются на старте, ЛОНП оказываются во фракции пре-глобулинов, ЛНП и ЛПП находят во фракции глобулинов, а ЛВП - 2-глобулинов. Если в крови повышено содержание -глобулинов (ЛНП) - это означает, что холестерин откладывается в тканях (развивается атеросклероз).