Лекция 2. Экзон.
реклама

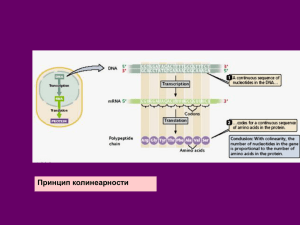
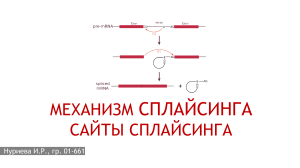
Лекция 2. Экзон [от английского ex(pressi)on — выражение, выразительность], участок гена (ДНК) эукариот, несущий генетическую информацию, кодирующую синтез продукта гена (белка). Соответствующие экзону участки ДНК, в отличие от интронов, полностью представлены в молекуле информационной РНК, кодирующей первичную структуру белка. По мнению некоторых исследователей экзоны соответствуют доменам (структурно автономным областям) в белке и являются первичными генетическими единицами, перекомбинация которых приводит к возникновению в ходе эволюции новых генов и соответственно новых белков. Экзоны чередуются в структуре гена с другими фрагментами — интронами. При альтернативном сплайсинге некоторые экзоны удаляются из зрелой РНК. Зрелая РНК может образоваться в результате: удаления интронов из незрелой мРНК в процессе цис-сплайсинга объединения и лигирования двух или более незрелых мРНК в процессе транс-сплайсинга. Зрелая РНК может кодировать полипептид (мРНК) или выполнять некодирующие функции (входить в состав рибосомы, рРНК или участвовать в трансляции в случае тРНК). В зависимости от контекста, экзон может соответствовать последовательности нуклеотидов и ДНК, и транскрипта РНК. Интрон — участок ДНК, который является частью гена, но не содержит информации о последовательности аминокислот белка. Схема нуклеотидной последовательности пре-мРНК гена CDK4 человека. Большую часть последовательности занимают интроны (показаны серым цветом) После транскрипции последовательности нуклеотидов, соответствующие интронам, вырезаются из незрелой мРНК (пре-мРНК) в процессе сплайсинга. Интроны характерны для генов эукариот. Интроны также найдены в генах, кодирующих рибосомальные РНК (рРНК), транспортные РНК (тРНК) и некоторые белки прокариот, эти интроны вырезаются на уровне РНК за счёт автосплайсинга. Число и длина интронов очень различны в разных видах и среди разных генов одного организма. Например, геном дрожжей Saccharomyces cerevisiae содержит в целом 293 интрона, в то время как в человеческом геноме можно насчитать свыше 300 тысяч интронов[1]. Обычно интроны длиннее экзонов. Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании матричной, или информационной, РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК)[1] и других некодирующих РНК. Автосплайсинг РНК тетрахимены обладает рибозимной активностью и может сплайсировать сама себя, замыкаясь в кольцо и одним своим концом вырезая интроны из другого. Сплайсинг белков, или белковый сплайсинг — внутримолекулярный автокаталитический процесс, происходящий в некоторых белках, при котором внутренняя часть белка (под названием интеин) выщепляется из белка-предшественника с последующим лигированием оставшихся частей. На месте сплайсинга в белке-предшественнике находится цистеин или серин, то есть аминокислота с нуклеофильной боковой группой. Известные в настоящее время реакции сплайсинга не требуют экзогенных кофакторов и источников энергии (например, АТФ или ГТФ). До открытия сплайсинга белков словом «сплайсинг» обозначали сплайсинг пре-мРНК.