03 Red-Ox
реклама

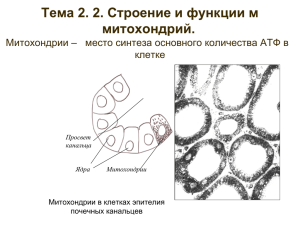
Компартменты растительной клетки Наружная мембрана (плазмалема) Клеточная стенка (апопласт) ЭПР Пластида Ядро Вакуоль Цитозоль (симпласт) Пероксисома Мембрана вакуоли (тонопласт) Митохондрия Плазмодесмы Поток электронов в растительной клетке Наружная мембрана (плазмалема) Клеточная стенка (апопласт) ЭПР Ядро Пероксисома Вакуоль Цитозоль (симпласт) ē Н2О ē НО 2 О2 ФC II ФC II О2 ЭТЦ ЭТЦ Пластида Митохондрия Плазмодесмы ē НО 2 ФC II Пластида ФC I Цит b/f МАЛИК-ЭНЗИМ: NADP+зависимая МДГ декарб. Порины Гликолиз ФЕП-карбоксилаза NADPH Оксалоацетат (С4) Оксалоацетат (С4) Порины ЭТЦ Н2О О2 Митохондрия АТФ NADP+зависимая МДГ декарб. Малат (С4) СО2 Гликолиз Пируват (С ) 3 Малат NAD+зависимая МДГ декарб. Гликолиз NADH АДФ+Ф NADP+ Пируват- Оксалоацетат NADH карбоксилаза NAD+ Цитозоль (симпласт) NAD+зависимая МДГ декарб. Малат (С4) СО2 NAD+ Пируват (С3) ē Н2О ФC II Цит b/f ФC I АДФ АТФ Пластида Гликолиз, 3-ФГК (С3) Цикл Кальвина Ф 1,3-ДФГК (С3) 3-ФГК (С3) Гликолиз АТФ 3-ФГА(С3) Ф Ф Порины Порины NADP+ NADPH Ф 3-ФГА (С3) 1,3-ДФГК (С3) АДФ NADH NAD+ ЭТЦ Н2О О2 Митохондрия Цитозоль (симпласт) Глюкоза (С6) 2 Пируват (С3) 2АТФ 2НАДН Сахароза или крахмал Гликолиз АТФ АДФ Глюкоза гексокиназа АТФ фруктозо-6-Ф Глюкозо-6-Ф АДФ фосфофруктокиназа Фруктозо-1, 6-Ф 3-ФГА 2НАДН ДГАФ НАД+ пируваткиназа 3-ФГК 1,3-ФГК АДФ АТФ 2-ФГК Субстратное фосфорилирование ФЕП АДФ ПВК АТФ Протекание гликолиза в клетках растений имеет свои особенности. Локализация: цитозоль и пластиды Обратимость гликолиза Регуляция гликолиза Глюконеогенез Гликолиз Глюкоза Пируват Глюконеогенез (Прорастающие семена) Синтез сахаров при обращении гликолиза называется глюконеогенезом. Как растения «обходят» необратимые реакции гликолиза, катализируемые гексокиназой, фосфофруктокиназой, пируваткиназой Особенности киназных реакций гликолиза цитозоль пластиды гексокиназа СН2О-(Р) СН2ОН АТФ О О АДФ ОН ОН ОН ОН глюкоза ОН ОН ОН ОН Глюкозо-6-фосфат СН2О-(Р) О СН2ОН АТФ (Р)-О--СН2 О АДФ Н2 С-О-(Р) ОН ОН ОН ОН ОН фруктозо-6-фосфат АТФ - фосфофруктокиназа ФФ - фосфофруктокиназа СН2О Р СНОН НС =О 3-фосфоглицериновый альдегид ОН фруктозо-1.6-бисфосфат альдолаза СН2ОН СО СН2О Р фосфодиоксиацетон Гликолиз Фн НАДН СН2О Р СНОН О=С -О ~ Р НАД+ фосфоглицерат киназа АДФ АТФ 1.3-фосфосфоглицерат СН2О Р СНОН НС =О 3-фосфоглицериновый альдегид СН2ОН СО СН2О Р фосфодиоксиацетон СН2О Р НСОН С ОО- СН2ОН НС-О- Р С ОО- 3-фосфоглицерат 2-фосфоглицерат Н2О енолаза СН2 С-О~ Р С ОО- фосфоенолпируват пируваткиназа АДФ АТФ СН3 С=О С ОО- пируват Глюкоза + 2НАД+ + 2 Фн + 2 АДФ = 2 пирувата + 2 НАДН + 2Н+ +2 АТФ Активация спиртового брожения наблюдается при аноксии НАДН НАД+ АЦЕТАЛЬДЕГИД ЭТАНОЛ АЛКОГОЛЬДЕГИДРОГЕНАЗА СО2 ФЕП ПИРУВАТДЕКАРБОКСИЛАЗ А ПИРУВАТ НАДН НАД+ ЛАКТАТ ФЕП-КАРБОКСИЛАЗА НСО3 ЛАКТАТДЕГИДРОГЕНАЗА – НАДН ОКСАЛОАЦЕТАТ НАД+ МАЛАТ Растения проявляют разную устойчивость к этанолу. Обращение реакции, катализируемой пируваткиназой СН3 С=О С ОО- пируваткиназа СН2 С-О~ Р С ОО- фосфоенолпируват АДФ пируват АТФ СО2 АТФ АТФ ФЕП-КАРБОКСИКИНАЗА СО2 пируваткарбоксилаза АДФ СОО С=О С Н2 СОО- НАД-МДГ оксалоацетат НАДН цитозоль НАД+ СОО НОСН С Н2 СОО- малат НАД-МДГ НАДН НАД+ СОО С=О С Н2 СОО- оксалоацетат митохондрии Регуляция гликолиза Животные Пластидный гликолиз Растения Цитозоль фруктозо-6-Ф фруктозо-6-Ф АТФ АТФ-ФФ-киназа ФФ АДФ Фруктозо-1, 6-Ф ФЕП Фруктозо-1, 6-Ф ФЕП пируваткиназа ПВК ПВК Фруктозо 2,6-бисфосфат – регуляторная молекула у всех эукариот Ключевое место регуляции гликолиза: отношение С6 / С3 киназы фруктозо-6-фосфат фруктозо-1.6–бисфосфат ФГА , ФГК фосфатаза Фн фруктозо-6-фосфат-2-киназа фруктозо-6-фосфат фруктозо-2.6–бисфосфат фруктозо-2,6-бисфосфат-2-фосфатаза АДФ Фр-2,6-бф Фн Фр-2,6-бф АТФ-ФФК ФФн-ФФК Фр-6-ф фосфатаза АТФ Цитрат Фр-1.6-бф ФЕП ФЕП,ФГК Окислительный пентозофосфатный шунт Глюкоза НАДФ НАДФН Глюкозо-6-Ф С3; 4; 5; 6; 7 3-ФГА 1,3-ФГК 5-Рибулезофосфат СО2 Фруктозо-1,6-БФ ФДА НАДФН 6-Фософглюконат Фруктозо-6-Ф Гликолиз НАДФ ПВ Митохондрия Первые две реакции пентозофосфатного цикла необратимы и сопровождаются образованием НАДФН и рибулозо-5-фосфата. СН2О(Р) 6НАДФ+ О 6х ОН ОН ОН ОН Глюкозо-6-фосфат 6НАДФН 6Н2О 6х COOH CНOH НOCH CНOH CНOH CH2O P 6-фосфоглюконовая кислота 6НАДФ+ 6НАДФН 6СО2 6х CH2ОН C =O CHOH CHOH CH2O P Рибулозо-5-фосфат CH2ОН 4 С5 C =O НОCH CHOH CH2O P Ксилулозо5-фосфат НС =О С3 НC = О 5 С5 РНК, ДНК, АТФ, NTP, НАД, КоА и др. 4 изомеризация ксилоза, ксилогликаны клеточной стенки C НOН CHOH CHOH CH2O P Рибозо--5фосфат СН2О Р НС =О 7 C НОН С4 СНОН CH2O P Эритрозо-4фосфат НС =О С3 C НОН CH2O P 3-фосфоглицериновый альдегид СН2ОН C=О С6 НОCH CHOH СНОН CH2O P Фруктозо-6фосфат C=О НОCH CHOH СНОН CH2O P Н О 2 Фруктозо-1.611 дифосфат Фн СН2ОН C=О С6 5х C=О CH2OН 3-фосфо -диоксиацетон 10 C=О СН2ОН C =O НОCH CHOH CH2O P ксилулозо-5фосфат СН2О Р 8 СН2ОН С7 НОCH CHOH СНОН СНОН CH2O P Седогептулозо-7фосфат 6 С5 9 C НОН CH2O P 3-фосфоглицериновый альдегид НОCH CHOH СНОН CH2O P Фруктозо-6фосфат СН2ОН C=О НОCH CHOH СНОН CH2O P Фруктозо-6фосфат шикиматный путь: синтез фенольных соединений (тирозин, триптофан, лигнин и др.) Растительные митохондрии имеют разнообразный размер и форму крахмал сахароза глюкоза гликолиз 2АТФ 2НАДН ЦИТОЗОЛЬ 2 х пируват СО2 Цикл трикарбоновых кислот АТФ 2СО2 4НАДН ФАДН2 e- Электрон-транспортная цепь АДФ + Фн ~+ МИТОХОНДРИЯ АТФ Пируватдегидрогеназный комплекс SH-СоА СО2 пируват О СН3 С S СоА НАДН Ацетил СоА Ацетил-S- СоА НАД+ • Е1 – пируватдегидрогеназа • Е2 –дигидролипоилацетил трансфераза • Е3 –дигидролипоил дегидрогеназа • Тиаминпирофосфат • Липоевая кислота НАД+, ФАД, СоА Ацетил-кофермент А (Acet-CoA) – восстановленный углеводный фрагмент, который окисляется в митохондриях для восстановления НАДН Кофермент А - универсальный переносчик и “активатор” ацильных (жирнокислотных) групп O || СоА-SH + CH3СOOH ↔ CoA-S~C-CH3 + Н2О. H3C~ || O H H O H H O H CH3 H | | || | | || | | | S-C-C-N-C-C-C-N-C-C– C – C — АДФ –3’Ф | | | | | | | | | H H H H H H OH CH3 H ¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯ пантетеин: пантоевая к-та + β-аланин + цистеамин цитратсинтаза цитрат оксалоацетат МДГ НАДН НАД+ Н2О НАД+ фумараза НАДН -кетоглутарат СoA-SH СО2 ФАДН2 сукцинатдегидрогеназа НАД-ИЦДГ СО2 фумарат фумарат ФАД аконитаза изоцитрат малат Н2О Н2О Н2О АДФ НАД+ НАДН -кетоглутарат дегидрогеназны комплекс Сукцинил СоА Сукцинат АТФ СoA-SH сукцинилтиокиназа С4-фотосинтез, Метаболизм азота CoASH НАДН Гликолиз, глюконеогенез С4-фотосинтез НАД+ НАД-МДГ Н2О фумараза СООц итратсинтаза СН2 С=О НО С-СООСН2 СН2 Н2О СОО оксалоацетат СООцитрат Н2О аконитаза аконитаза С ОО НО СН СН2 СООмалат СОО НСОН СН-СОО СН2 СООизоцитрат СО2 С ОО СН СН СООфумарат СОО- сукцинатдег идрогеназа ФАДН2 С ОО СН2 СН2 Сукцинил СоА-синтетаза СОО Сукцининат ФАД Глиоксилатный цикл Н2О СОО- изоцитратдегидрогеназа СО НАД+ СН2 СН2 НАДН СОО-кетоглутарат О CoASH С SCoA СН2 СН2 НАД+ СООСукцинил СоА НАДН СО2 -кетоглутаратдегидрогеназный комплекс АТФ АДФ CoASH Метаболизм азота Митохондрия КоASH С3ПВ НАД Цикл Кребса (1953) (ди- и три карбоновых кислот) СО2 9 реакций Ацетил КоА 1 НАДН С2 С4ЩУК НАД С4 малат НАДН ФАДН2 АТФ НАДН НАДН С4 фумаровая ФАД С4 янтарная (сукцинат) 2* 1мол АТФ 3 мол НАДН +1 мол НАДН 1 мол ФАДН2 2 лимонная С6 3 изолимонная С6 НАД СО2 2-оксоглутаровая (кетоглутаровая) С5 НАД АДФ СО2 Сукцинил КоА С4 ~тиоэфирная связь КоASH Субстратное 2*3СО2 фосфорилирование Реакции обращения реакци ФЕП -----ПВК Особенностью растительных Мх является присутствие малик-энзима малатдегидрогеназа декарбоксилирующая НАД-зависимый малик-энзим (цитозоль, Мх) НАДФ-зависимый малик-энзим (Хл) глюкоза Цитозоль гликолиз НСО3- пируваткиназа ФЭП АДФ гликолиз ФЕПК АТФ Фн оксалоацетат пируват НАДН МДГ Транспорт в митохондрии НАД+ малат НАД+ НАДН Транспорт в митохондрии пируват малат малик-энзим НАД+ МДГ СО2 ПДК СО2 НАД+ НАДН НАДН оксалоацетат ацетилСоА цитрат Цикл трикарбоновых кислот Митохондрия Конверсия жиров в углеводы включает три этапа: Распад жиров и -окисление жирных кислот Глиоксилатный цикл в глиоксисомах Глюконеогенез • В клетках животных жирные кислоты подвергаются -окислению в митохондриях • В клетках растений жирные кислоты подвергаются -окислению в глиоксисомах, которые являются разновидностью пероксисом. Глиоксисомы впервые были выделены Биверсом в 1967 г. (Beevers) О || Н2С – С – С - (СН2)n -СН3 О || НС – С – С - (СН2)n -СН3 О || Н2С – С – С - (СН2)n -СН3 олеосома жиры глицерин липаза жирные кислоты Ацил-СоА Ацил-СоА глиоксисома nНАД nO2 -окисление + nНАДH nH2O2 Ацетил-СоА Ацетил-СоА Глиоксилатный цикл Глиоксилатный цикл сукцинат сукцинат сукцинат митохондрия глюкоза ЦТК малат глюконеогенез ФЕП-карбоксикиназа АТФ СО2 малатдегидрогеназа оксалоацетат фосфоенолпируват АД Ф малат НАД+ НАДН Глиоксилатный цикл 1цитратсинтаза Н3ССОS-CoA ацетилСоА СОО СН2 С=О СООоксалоацетат CoA Н2О Н2О Н2О аконитаза СОО СН2 НС СОО СНОН СООизоцитрат изоцитратлиаза МДГ НАДН СОО СН2 НОС СОО СН2 СООцитрат НАД+ СОО СН2 СНОН СООмалат НС = О СООглиоксилат Н2О Н3ССОS-CoA ацетилСоА CoA малатсинтаза СОО СН2 СН2 СОО- сукцинат Между митохондриями и цитозолем существует обмен восстановительными эквивалентами и метаболитами ЦТК. В митохондриях пересекается обмен углеводов, белков и жиров. Глюкоза Гликолиз липиды пируват аланин Жирные кислоты Ацетил СоА аспартат цитрат оксалоацетат ЦТК другие Изопреноидные соединения - кетоглутарат другие аминокислоты Сукцинил СоА аминокислоты порфирины белки цитохромы фитохромы глутамат белки хлорофиллы Результат: энергетическая система клетки - дыхание Обмен метаболитами между матриксом и цитозолем осуществляется с помощью белков-транспортеров цитозоль АТФ4- ОН- ОНАДФ3- НРО42- Пируват- НРО42- ацетилСоА Синтез аминокислот в хлоропластах оксалоацетат 2малат 2цитрат 3-/Н+ НАД+ -кетоглутарат2- ЦТК Цитрат3- малат 2- СО2 НАДН -кетоглутарат 2- -кетоглутарат 2- НАДН НАД+ малат оксалоацетат Синтез аминокислот в хлоропластах малат оксалоацетат НАДН НАД+ Мх-ЭТЦ основные компоненты 4Н+ 2Н+ 4Н+ e- Q e- НАДН ротенон Цит с III I НАД Сукцинат Межмембранное пространство IV e- О2 АНТИМИЦИН А Н2О СО CN- матрикс У растений работают дополнительно 4 НАД(Ф)Н-редуктазы 4Н+ НАДН НАДФН ДГex e- ДГex I ДГin НАДН ротенон Цит с III II e- nН+ 2Н+ 4Н+ Q IV e- ДГin НАДН НАДФНСукцинат АТФ синтаза О2 АНТИМИЦИН А Н2О СО CN- АДФ АТФ ОЛИГОМИЦИН Мх-ЭТЦ у растений 4Н+ НАДН НАДФН ДГex e- ДГex I Q НАДН ротенон АО ДГin НАДН НАДФНСукцинат IV e- eДГin Цит с III II e- nН+ 2Н+ 4Н+ АТФ синтаза О2 Н2О SHAM О2 АНТИМИЦИН А Н2О СО CN- АДФ АТФ ОЛИГОМИЦИН Особенности ЭТЦ-дыхания растений: Альтернативная оксидаза (цианид-резистентное дыхание) AO относится к семейству di-iron белков R-2 типа Обнаружена у E. coli, Paracoccus denitrificans, простейших Trypanosoma, водорослей и всех семенных растений. Впервые белок АО был получен из мтх Sauromatum guttatum Цианидрезистентное дыхание Термогенез ароидных растений Arum maculatum, Symplocarpus foetidis, Sauromatum guttatum, Philodendron selloum. Альтернативная оксидаза один полипептид ( М.В.~ 32 кДа) один ядерный ген АОХ1 В мембране Мх: мономерная или димерная форма Впервые белок АО был получен из мтх Sauromatum guttatum 2QH2 +O2 → 2Q + 2H2O • Термогенез в цветках некоторых растений связан с терморегуляцией: • При понижении температуры воздуха температура тканей цветка остается постоянной , а скорость поглощения кислорода возрастает. При повышении температуры воздуха скорость дыхания падает • t воздуха t цветка • Symplocarpus foetidis +3 -+240 +16 -+ 260 • Philodendron selloum +4 -+390 +39 -+ 440 • Существуют точки, (switchig temperature) за пределами которых «терморегуляция» не действует Sauromatum guttatum Symplocarpus foetidus Helicodeceros muscivorus OH Fe3+ Q QH2 Fe3+ Q + 2H2O OH диферри Fe2+ Fe2+ Fe3+-O – O -Fe3+ диферро QH2 Перокси-ферри Fe2+ Fe2+ O2 QH2 QH2 Гипотеза свехпотока через АОХ Особенности ЭТЦ-дыхания растений: Альтернативная оксидаза (цианид-резистентное дыхание) прямо или косвенно восстанавливает дисульфидную связь Аллостерически активирует АО пируват НАДФН ЦТК цитрат Регулирование синтеза и активности альтернативной оксидазы Пируват ЦТК НАДН I Q/ Q2 III IV O2 AO O2 Активация альтернативной оксидазы Инактивация Red-Ox! Активация АКТИВАЦИЯ СИНТЕЗА Альтернативной Оксидазы DE NOVO ПРИ ДЕЙСТВИИ СТРЕССОВ НИЗКОТЕМПЕРАТУРНЫЙ СТРЕСС ВНЕДРЕНИЕ ПАТОГЕНА ПОРАНЕНИЕ ЗАСОЛЕНИЕ ДЕФИЦИТ ФОСФОРА, АЗОТА ПРИ ВОДНОМ ДЕФИЦИТЕ ЗАСУХА ПРИ ЗАСОЛЕНИИ ПРИ ДЕЙСТВИИ ТЯЖЕЛЫХ МЕТАЛЛОВ И ГЕРБИЦИДОВ I Q.- O2.- H2O2 Q/QH2 •Интенсивность АД •зависит от содержания белка АО в мембране, •т.е. от интенсивности экспресии гена Аох1. III Q.- AOX1 AO СИНТЕЗ IV O2 O2 Н2O Н2O КЛЕТКИ ТАБАКА АНТИМИЦИН А КСN САЛИЦИЛОВАЯ К-ТА НАКОПЛЕНИЕ Н2О2 НАКОПЛЕНИЕ Н2О2 СИНТЕЗ АО +SHAM СИНТЕЗ АО УСТОЙЧИВОСТЬ К ВТМ УСТОЙЧИВОСТЬ К ВТМ НЕТ УСТОЙЧИВОСТИ Множество кольцевых молекул митохондриальной ДНК растений – результат гомологичных рекомбинаций по повторам. Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы Линейная ДНК митохондрий хламидомонады Сопоставление хлоропластного и митохондриального геномов риса. Варианты редактирования хлоропластных и митохондриальных РНК растений Гены митохондрий 1. Синтез белка. - 3 гена рРНК (оперон rrn) - 10 генов белков пластидных рибосом (rpl/rps) - 16 генов тРНК (trn) – не хватает! – импорт! 2. Дыхание - 9 генов белков НАД Н дегидрогеназы (nad) - ген апоцитохрома b (cob); - 5 генов белков биосинтеза цитохрома с (ccb) - 3 гена субъединиц цитохромоксидазы (гены сох). - 3 гена субъединиц сукцинатдегидрогеназы (sdh) у печеночников - 4 гена АТФ-синтазы (atp) Всего: около 50 генов (у печеночных мхов – более 100) , из них около 20 - «рабочих» и около 30 - «домашнего хозяйства». • Ядерный генов контролирует : • • • • • некоторые т РНК ДНК – полимеразу РНК – полимеразу Некоторые рибосомальные белки Синтез белков, входящие в состав комплексов электрон – транспортной цепи дыхания • Синтез ферментов ЦТК • Fzo – это гигантские ГТФ-азы, расположенные во внешней мембране митохондрий • Fzo (fuzzy onion) Цитоплазматическая мужская стерильность (ЦМС) Ядерный ген «Техасская» - Т-ЦМС Restorer of Fertility = rf1rf1 = Rf rf2rf2 Т-мх стерильность Т-мх Rf1Rf1 Rf2Rf2 восстановление фертильности «USDA»-тип - S-ЦМС Ядерный ген Rf3 «Молдавская» - M-ЦМС «Парагвайская» (Charrua-тип) - С-ЦМС Ядерный ген Rf4 АТФ-синтаза: ключевой фермент «Техасской» ЦМС 13 kDa-белок появляется в митохондриях как результат незаконной рекомбинации c геном atp6 и встраивается в АТФ-синтазу, вызывая ЦМС АТФ-синтаза: ключевой фермент ЦМС у петунии У петунии незаконная рекомбинация происходит с участием гена atp9. Изменения в АТФ-синтазном комплексе вызывают ЦМС Цитоплазматическая мужская стерильность (ЦМС) «USDA»-тип - S-ЦМС Ядерный ген Rf3 «Молдавская» - Т-ЦМС При S-ЦМС(=М-ЦМС) появляются линейные фрагменты в митохондриальном геноме: S1 и S2 – эписомы. Возможно, более крупные кольцевые молекулы мт-ДНК за счет рекомбинации могут линеаризоваться. Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль «USDA»-тип - S-ЦМС Ядерный ген Rf3 «Молдавская» - M-ЦМС S-мх S-мх S-мх rf3rf3 Стерильность Rf3rf3 Полное (!) восстановление фертильности Rf3Rf3 Восстановление фертильности Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль «Техасская» - Т-ЦМС Ядерные гены Rf1 и Rf2 T-мх Т-мх Т-мх rf1rf1 Rf1rf1 Rf1Rf1 Стерильность Половина (!) пыльцевых зерен Rf2-ген – фертильна доминантная гомозигота Восстановление фертильности ē Н2О ФC II Цит b/f ФC I АДФ АТФ Пластида NADP+ NADPH Гликолиз, 3-ФГК (С3) Цикл Кальвина Ф 1,3-ДФГК (С3) Ф Ф Порины 3-ФГК (С3) Гликолиз АТФ 3-ФГА(С3) Ф 1,3-ДФГК (С3) АДФ NADH 3-ФГА (С3) NAD+ Цитозоль (симпласт) 4Н+ НАДН НАДФН ДГex e- ДГex I Q НАДН ротенон АО ДГin НАДН НАДФНСукцинат IV e- eДГin Цит с III II e- nН+ 2Н+ 4Н+ АТФ синтаза О2 Н2О SHAM О2 АНТИМИЦИН А Н2О СО CN- АДФ АТФ ОЛИГОМИЦИН