Дифференцировка клеток в онтогенезе.
реклама

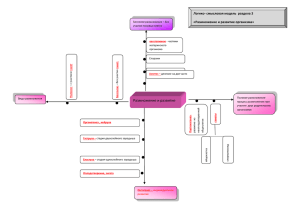
ДИФФЕРЕНЦИРОВКА КЛЕТОК В ОНТОГЕНЕЗЕ лекция 13 Основные понятия Дифференцировка: процесс развития специализированных клеточных типов из оплодотворённой яйцеклетки Детерминация: процесс определения судьбы данной части зародыша • недетерминированные части – судьба не определена • детерминированные части – определённая судьба Потенция части зародыша – то, что она может дать при любых условиях: • проспективные потенции – могут осуществиться в будущем при определённых условиях • проспективные значения – то, что данная часть даёт при нормальных условиях развития • тотипотентность – начало всем клеточным типам взрослого организма • мультипотентность – начало многим, но не всем клеточным типам • унипотентность – лишь одно направление развития (детерминированность) Дифференциальная активность генов Дифференцировка клеток в онтогенезе – результат последовательных реципрокных (взаимных) влияний цитоплазмы и меняющихся продуктов активности ядерных генов (дифференциальная экспрессия генов ) • наследственный материал соматических клеток способен оставаться полноценным не только в количественном, но и в функциональном отношении • взаимодействие цитоплазмы и ядра в жизнедеятельности клеток и развитии организма имеет решающее значение • чем старше стадия зародыша Опыт Дж. Гердона донора, из клеток которого брали ядро для пересадки, тем в меньшем проценте случаев развитие оказывалось полностью завершенным Способы детерминации • автономная обусловлена материнскими цитоплазматическими факторами, вырабатывающимися в период оогенеза под контролем генов материнского организма. Достигается в результате неравномерного распределения в ооците транскрипционных факторов (цитоплазматическая сегрегация) • зависимая устанавливается с помощью межклеточных взаимодействий, вследствие передачи сигнала от одних клеток и детектирования сигнала другими клетками эмбриона, побуждая их специфицироваться в определённом направлении Аристотель (384-322 до н.э.) Гиппократ (460-370 до н.э.) Эпигенез Преформизм Автономная детерминация (1): цитоплазматическая спецификация Цитоплазма зиготы Styela partita и её проспективное значение Мозаичная детерминация у оболочников (Reverbery & Minganti, 1946) Локализация АХЭ в зародышах оболочников (Whittaker, 1973) Автономная детерминация (2): цитоплазматическая локализация Изоляция ранних бластомеров моллюска Patella coerulea (Wilson, 1904) Полярная лопасть моллюсков Автономная детерминация (3): локализация детерминантов половых клеток Распределение половой (зародышевой) плазмы у аскариды (Бовери, 1910) Сегрегация линии половых клеток и диминуция хроматина у галлицы Зависимая детерминация (1) Опыт В. Ру (1888) Регуляционное развитие (эмбриональная регуляция): некоторая часть зародыша может дать целостный организм нормальной структуры. • эмбриональная регуляция возможна лишь при наличии мульти- (тоти-) потентности клеток зародыша • «проспективное значение каждого элемента системы есть функция его положения в целом» (закон Дриша). Плутеус из 1 клетки 4-х клеточного Нормальный зародыша плутеус Зависимая детерминация (2) Асимметрия в зародыше и яйце морского ежа (Гёрстадиус, 1928-1935) 1 – задержанная бластула (перманентная бластула); 2 – личинка (слегка аномальная; 3–6 – личинка (маленькая, но нормальная); 7 – задержанная бластула (аномальная бластула); 8 – личинка (почти нормальная) Модель «двойного градиента» (J. Runnstrom, ограничение потенций в ходе онтогенеза. 1929): Бластомеры остаются способными к регуляции до тех пор, пока не будут формироваться исключительно из анимальной или вегетативной цитоплазмы. Зависимая детерминация (2а) Обнаружение «анимализирующего» и «вегетализирующего» градиентов в развитии зародышей морского ежа Эмбриональная индукция Ранние опыты Г. Шпемана Детерминация: предназначение клеток к дифференцировке в специфический клеточный тип Равноценность ядер у зародышей тритона Асимметрия в яйце у амфибий зависимое развитие независимое развитие Детерминация клеток эктодермы на стадии ранней (А) и поздней (Б) гаструлы у тритона: 1 – презумптивная нейральная эктодерма; 2 – презумптивный эпидермис; 3 – нервная пластинка (НП); 4 – формирующийся эпидермис; 5 – формирующаяся вторичная НП Эмбриональная индукция (опыт Г. Шпемана и Г. Мангольд, 1924) • участок, взятый из спинной губы бластопора, способен направлять (переключать) развитие находящегося вокруг материала на определенный путь развития; • боковая и брюшная сторона Эмбриональная индукция: взаимодействие частей развивающегося зародыша А – пересадка спинной губы бластопора на место презумптивного эпидермиса брюшной стороны; Б – формирование комплекса осевых структур в области трансплантата; В – формирование второго зародыша, соединённого с первым гаструлы обладают более широкими потенциями к развитию, нежели их презумптивное проспективное направление; • достаточно точное строение новообразованных органов в месте пересадки указывает на эмбриональную регуляцию. Спинная губа бластопора – первичный эмбриональный (шпемановский) организатор. Эмбриональная индукция: особенности • головной индуктор • туловищный индуктор Компетенция – способность ткани отвечать специфическим образом на данный стимул (индуктор). Виды индукции: • первичная: формирование осевых органов, под действием дорсальной губы; • вторичная: действие глазного бокала на прилежащий покровный эпителий ; • третичная: изменения находящегося над хрусталиком покровного эпителия и трансформация его в прозрачный эпителий роговицы • гетерономная: одна часть зародыша индуцирует образование иного органа; • гомономная: индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам. Эмбриональная индукция: механизм (1) Ньюкоповская индукция: дорсальные вегетативные бластомеры оказывают индуцирующее действие на клетки краевой зоны (развитие в мезодермальном направлении). Центр Ньюкупа (ньюкуповский организатор): вегетативно-дорсальная область зародыша. Материал хордомезодермы не предетерминирован, а специфицируется в ходе дробления. Если на стадии средней бластулы сделать разрез в экваториальной области и повернуть анимальную половину зародыша на 180о, то хордомезодерма формируется из материала бывшей вентральной стороны, который после поворота занял дорсальное положение. Эмбриональная индукция: механизм (2) Дорсализация зародыша амфибий: А – момент оплодотворения; Б – поворот кортикальной цитоплазмы; В – накопление -катенина; Г – инициация экспрессии siamois Черные круги – Dsh, кортикальная цитоплазма анимального полюса – черный, вегетативного полюса – белый цвет. Ген goosecoid (gsc) Гены организатора шпорцевой лягушки Эмбриональная индукция: механизм (3) Дифференцировка нейральных производных На вентральной стороне зародыша создаётся максимальная концентрация белков семейства BMP (Bone Morphogenetic Proteins, морфогенетические белки костного мозга), в частности – ВМР-4. Нейрализация эктодермы – реализация «основного состояния» (ground state) системы (индукцией по умолчанию (default induction), поскольку формирование нервной трубки происходит в случае отсутствия вентрализирующего сигнала. Дополнительная литература по теме: • Gastrulation: From Cell to Embrio. Edited By Claudio D. Stern, University College London. Electronic resource: http:// www.gastrulation.org The web site provides supplementary data and movies to accompany the chapters in the book.