lekciya_19_biomehanika_krovoobrashcheniya
реклама

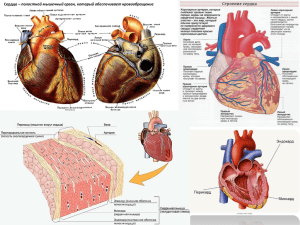
Лекция № 18 БИОМЕХАНИКА КРОВООБРАЩЕНИЯ Кровообращение происходит в сердечно-сосудистой системе. С позиций механики, сердце является комбинированным многокамерным насосом, нагнетающим кровь в большой и малый круги кровообращения, образованные сосудами разных типов (артериями, капиллярами, венами). Элементы биомеханики сердца. Механическая работа, совершаемая сердцем, развивается за счет сократительной деятельности миокарда. Вслед за распространением возбуждения происходит сокращение миокардиальных волокон. Функциональным элементом миокарда служит мышечное волокно, образованное цепочкой кардиомиоцитов, соединенных «конец в конец» и заключенных в общую сарколемму. В такие цепочки объединяются от 2 до 15 кардиомиоцитов, каждый из которых имеет длину от 100 до 200 мкм. Общая сарколемма имеет выраженный гликокаликс толщиной примерно 50 нм. Мышечные волокна собираются в пучки, отделенные друг от друга тонкими прослойками соединительной ткани. Пучки группируются в тяжи, взаимодействующие между собой посредством сложных нерегулярных контактов. Сократительной способностью обладает 99% всех кардиомиоцитов. Их называют сократительными (типичными, рабочими). В этих клетках миофибриллы упорядочены (направлены вдоль их длинной оси и параллельны друг другу), хорошо развиты саркоплазматическая сеть (СПС) и система Т-трубочек, много митохондрий. Толщина сократительных кардиомиоцитов у человека составляет 15−25 мкм. Поверхность каждого из них примерно 8,5 · 10-5 см2, а объем − 1,94 · 10-10 л, причем 34% объема приходится на митохондрии. В проводящих (атипичных, узловых, специализированных) кардиомиоцитах, на долю которых приходится всего 1% миокардиальных клеток, сократительный аппарат развит слабо. Способность сократительных кардиомиоцитов активно укорачиваться и развивать напряжение обеспечивает механическую (насосную) функцию сердца. Возбуждающий сигнал (потенциал действия) поступает без декремента к сократительным кардиомиоцитам от пейсмекера по миокардиальному синцитию. Потенциал действия на мембране Т-трубочек инициирует повышение концентрации Са2+ в миоплазме подле миофибрилл. Са2+ поступает к миофибриллам миоцита скелетной мускулатуры исключительно из цистерн СПС. В отличие от этого, в кардиомиоцитах СПС развита слабо, и ключевым событием в электромеханическом сопряжении является вход Са2+ в миоплазму из интерстиция через потенциалзависимые каналы в сарколемме. Кальциевый (медленный входящий) ток (Isi) течет через сарколемму в течение существования «плато» на потенциале действия. Чем оно продолжительнее, тем больше Са2+ входит в миоплазму. Входящий кальциевый ток выполняет триггерную функцию по отношению к потенциалзависимым кальциевым каналам в мембране СПС. При небольшом повышении концентрации Са2+ в миоплазме деполяризация мембраны T-трубочек открывает кальциевые каналы в мембране СПС кардиомиоцитов млекопитающих (рис. 75). Поток Са2+ из СПС в миоплазму на 2 порядка интенсивнее потока через сарколемму, но кальциевый ток из СПС в саркоплазму не пойдет без Isi В этом состоит важное отличие электромеханического сопряжения от аналогичного процесса в миоците скелетных мышц. 1 Миофибриллы Как и в миоцитах скелетных мышц, Рис. 75. Схема электромеханического сопряжения в кардиомиоците: СПР − саркоплазматический ретикулум (в тексте СПС) повышение концентрации Са2+ от 10-7 − 10-8 М до (0,2−1,5)·10-6 М устраняет тропомиозиновую блокаду сокращения, «расчехляя» активные центры для миозина на актиновых нитях. Однако в кардиомиоцитах нет тропонина, и процесс их сокращения происходит без его участия. Максимальное сокращение кардиомиоцита наблюдается при достижении концентрации Са2+ подле миофибрилл около 5 10-6 М. Однако в условиях покоя напряжение миокарда не превышает 70% от максимально возможного. После закрытия кальциевых каналов в сарколемме (в этот момент обрывается «плато» на потенциале действия) прекращается Isi а избыток Са2+ удаляется из миоплазмы. В кардиомиоцитах выведение ионов кальция осуществляется двумя путями. Во-первых, кальциевый насос, сосредоточенный в мембране СПС, обеспечивает их активный транспорт в цистерны СПС − так же, как в миоцитах скелетной мускулатуры. Во-вторых, кардиомиоцитам присущ своеобразный механизм выведения кальция в интерстиций − посредством кальций-натриевого обменника, находящегося в сарколемме. Эта система пассивного транспорта осуществляет во время диастолы обмен одного иона Са2+, который выходит из миоплазмы, на 2 иона Na+, входящих в нее из интерстиция. С работой Na-Ca-обменника связан лечебный эффект сердечных гликозидов, которые являются специфическими ингибиторами K-Na-АТФазы. При блокаде ими калий-натриевого насоса в плазмолемме кардиомиоцитов активный транспорт (откачивание) натрия из миоплазмы тормозится, что приводит к повышению там концентрации Na+ . В результате понижается градиент ионов натрия на сарколемме, что ослабляет их поток в кардиомиоцит через Na-Ca-обменник, который начинает работать слабее, уменьшая выведение ионов кальция из миоплазмы. Их концентрация подле миофибрилл возрастает, усиливая сокращения кардиомиоцита. Так достигается преодоление сердечной недостаточности. Однако применять сердечные гликозиды следует очень осторожно, поскольку передозировка приводит к противоположному эффекту, вплоть до остановки сердца. При нарушении электромеханического сопряжения в сердце наблюдается явление электромеханической диссоциации − разобщения электрической активности и сокращений миокарда вследствие ослабления поступления Са2+ в миоплазму из интерстиция. Это происходит при понижении концентрации ионов кальция в интерстиции (например, вследствие недостатка их в пище и воде), при блокаде кальциевых каналов в сарколемме кардиомиоцитов (солями Со2+, Cd2+, Ni2+, La3+ и некоторых других элементов). В таких случаях бывает внезапная остановка сердца. Особенности сократимости миокарда: 1.Зависимость силы сокращения миокарда от силы стимула подчиняется закону 2 «все или ничего». Этот закон, характерный для отдельных клеток, свойствен всему миокарду, поскольку он представляет собой синцитий. 2.В зависимости силы сердечных сокращений от частоты стимула наблюдается феномен «лестницы Боудича» − нарастание силы сердечных сокращений по мере повышения частоты стимуляции. Такой (хроноинотропный) эффект объясняется тем, что при частоте стимуляции не весь Са2+, вошедший в миоплазму во время систолы, удаляется в диастолу. Его концентрация подле миофибрилл поддерживается на повышенном уровне, что и обусловливает усиление сокращений. 3.Сила сокращений желудочков сердца прямо пропорциональна исходной длине мышечных волокон − иными словами, чем больше расслабление миокарда в диастолу, тем сильнее сокращение в систолу. Эту зависимость называют основным законом сердца Франка-Старлинга. Он был установлен О. Франком в 1895 г. Зависимость «скорость−сила»; чем больше нагрузка на сердце, тем оно медленнее сокращается. Такая зависимость следует из уравнения А. Хилла, которому миокард подчиняется так же, как скелетная мышца. Вследствие ритмического характера автоматизма сердцу свойственна циклическая сократительная деятельность. Каждый цикл сердечных сокращений состоит из ряда механических процессов, идущих в строгой последовательности. Прежде чем характеризовать эти процессы, отметим, что левый желудочек нагнетает кровь в большой, а правый − в малый круги кровообращения. Кроме того, левое предсердие совершает работу по наполнению левого желудочка и тем самым способствует эффективности его нагнетательной функции. То же делает правое предсердие по отношению к правому желудочку. Поэтому принято условно говорить о левом сердце и правом сердце, как о двух насосах, соединенных друг с другом последовательно (выход одного является входом другого). Оба они перекачивают примерно одинаковые объемы крови за каждый цикл. При средней частоте пульса (75 мин-1) полное время сердечного цикла составляет 0,8 с. Сначала сокращаются предсердия. Благодаря атриовентрикулярной задержке их сокращение (систола предсердий) при расслабленных желудочках продолжается 0,1 с. Затем начинают сокращаться желудочки. В желудочковой систоле выделяют ряд стадий (их называют фазами). Первая из них − фаза асинхронного сокращения длится 0,05 с. В течение ее нет полной синфазности в возбуждении миокардиальных волокон. Из-за неодновременности сокращений отдельных из них желудочки развивают слабое усилие, но к концу фазы асинхронного сокращения степень синфазности нарастает, многие волокна вовлекаются в сокращение, вследствие чего достигается достаточная сила, чтобы создать внутри желудочков большее кровяное давление (КД), чем в предсердиях (около 5 мм рт. ст., что эквивалентно 0,66 кПа). В этот момент КД в магистральных артериальных стволах, куда кровь затем должна быть выброшена желудочками, значительно выше: в аорте − примерно 60 мм рт. ст. (8 кПа), в легочном артериальном стволе − 15 мм рт. ст. (2 кПа). Каждый из желудочков отделен от своего предсердия створчатым, а от соответствующего магистрального ствола − полулунными клапанами. Клапаны открываются и закрываются в зависимости от соотношения КД в полостях, которые ими разделяются. Створчатые клапаны закрыты тогда, когда КД внутри желудочков выше, чем в предсердиях. Полулунные клапаны, напротив, открываются при высоком давлении в желудочках (при КД в левом желудочке выше, чем в аорте, и при КД в правом желудочке выше, чем в легочном артериальном стволе). Когда в конце фазы асинхронного сокращения внутрижелудочковое КД станет выше, чем внутрипредсердное, створчатые клапаны закрываются, после чего желудочки 3 продолжают сокращаться в условиях герметичности своих полостей, заполненных кровью. Следовательно, первая фаза систолы желудочков сменяется второй − фазой изометрического сокращения. Изометрический режим обусловлен тем, что сердечная мышца сокращается вокруг несжимаемого тела − крови, содержащейся в желудочках, из которых она не может выйти, так как обе пары отверстий перекрыты клапанами. Существование фазы изометрического сокращения является важным условием нормальной сердечной деятельности. Все усилие, развиваемое миокардом, идет на повышение его потенциальной энергии и, следовательно, на создание напряжения. В результате КД внутри желудочков стремительно нарастает: в левом желудочке − от 5 до 60 мм рт. ст., в правом − от 5 до 10 мм рт. ст. Это происходит примерно за 0,03 с − такова продолжительность фазы изометрического сокращения. Благодаря достижению внутри желудочков КД, большего, чем в магистральных артериальных стволах, открываются полулунные клапаны, и кровь устремляется из сердца в сосуды. Вначале КД в желудочках еще продолжает повышаться, несмотря на открытие клапанов. Этот режим сокращения называют изотоническим, что не совсем точно выражает его характер, поскольку при нем КД не удерживается на постоянном уровне. Например, в левом желудочке давление повышается примерно на такую же величину, как и в течение фазы изометрического сокращения (около 60 мм рт. ст.), но не за 0,03, а за 0,12 с. За это время кровь быстро вытекает из желудочков в сосудистое русло. Поэтому третья фаза сердечного цикла называется фазой быстрого изгнания. Затем еще 0,13 с кровь вытекает из желудочков в фазу медленного изгнания, причем в течение четвертой фазы скорость истечения крови неуклонно понижается, поскольку падает внутрижелудочковое давление. Так, КД в полости левого желудочка, достигшее к началу фазы медленного изгнания 120−130 мм рт. ст. (16,0−17,3 кПа), к концу ее становится меньше диастолического давления в аорте. После расслабления − диастолы наступает новое сокращение. Сердечный цикл повторяется. Сила, возникающая в сердечной мышце при ее сокращении, зависит от исходной длины миокардиальных волокон, поскольку по мере укорочения волокна увеличивается число миозиновых мостиков, участвующих в сокращении. Это общее свойство мышечной ткани. В миокарде оно находит выражение в законе Франка−Старлинга: сила сердечного сокращения прямо пропорциональна исходной длине миокардиальных волокон (перед началом систолы), а исходная длина зависит от степени растяжения их кровью, поступающей из предсердий. Следовательно, сила сокращения миокарда желудочков тем значительнее, чем больше крови поступает в них во время диастолы. От диастолического наполнения желудочков зависит основной показатель (параметр) сократительной функции сердца − систолический (ударный) объем (Vc). Так называют объем крови, выбрасываемый каждым из желудочков при одном сокращении. Систолические объемы левого и правого желудочков одинаковы. Для каждого из желудочков Vc составляет 70−100 мл в условиях покоя, причем изгоняется 60−70% крови, содержавшейся там в период диастолы. При физической работе и эмоциях Vc увеличивается в 2−3 раза. От систолического объема зависит работа сердца, но ее значение может быть разным при одной и той же величине Vc Работа, совершаемая сердцем, затрачивается, во-первых, на выталкивание крови в магистральные артериальные сосуды против сил давления и, во-вторых, на придание крови кинетической энергии. Первый компонент работы называется статическим (потенциальным), а второй − кинетическим. Статический компонент работы сердца вычисляется по формуле Аст = =Pcp Vc , (70) где Pсp − среднее давление крови в соответствующем магистральном сосуде 4 (аорте − для левого желудочка, легочном артериальном стволе − для правого желудочка). Поскольку изменение КД в артериях является сложной периодической функцией, среднее давление равно не полусумме максимального (систолического) и минимального (диастолического) давлений, а среднему из бесконечно малых изменений давления от максимального до минимального в течение одного сердечного цикла (рис. 76) и определяется по формуле: (71) где t = t 2- t1 − время сердечного цикла. Рис. 76. Кривая динамики кровяного давления в артерии:по оси абсцисс − время, по оси ординат – кровяное давление. На графике показан способ планиметрического определения среднего давления: определив площадь под кривой кровяного давления за время сердечного цикла (площадь фигуры АабвгД) при помощи планиметра, строим равновеликий ей прямоугольник АВСД, основание которого равно основанию фигуры АабвгД (тоесть АД = ∆t); тогда высота этого прямоугольника (ЕН) численно равна величине среднего давления. Рс − систолическое давление; РД − диастолическое давление; tс = АД – время сердечного цикла (∆t) В отличие от систолического и диастолического давлений, изменяющихся в довольно широких пределах, рср характеризуется постоянством. И.П. Павлов относил его к гомеостатическим константам организма. Величина рср в большом круге кровообращения составляет приблизительно 100 мм рт. ст. (13,3 кПа). В малом круге рср = 15 мм рт. ст. (2 кПа), т. е. примерно в 6 раз меньше, чем в большом. Поскольку Vc обоих желудочков одинаков, а давление, против которого они совершают работу, имеет шестикратное различие, то и статический компонент работы левого желудочка приблизительно в 6 раз больше: А ст левого желудочка =13,3 103 Па 70 10-6 м3 ≈ 0,9 Дж; А ст правого желудочка =0,15 Дж. В этих расчетах Vc принят равным 70 мл. Кинетический компонент работы сердца определяется по формуле: (72) где ρ − плотность крови (примерно 10 кг · м ); υ − скорость кровотока в магистральном артериальном стволе (в среднем 0,7 м · с-1). Следовательно, Ak =1/2 · 103 кг · м3 · 70 · 10-6 м3 · 0,49 м2 · с-2 ≈ 0,02 Дж. В целом работа левого желудочка за одно сокращение в условиях покоя составляет около 1 Дж, а правого − менее 0,2 Дж, причем статический компонент доминирует, 3 -3 5 достигая 98% всей работы, тогда как на долю кинетического компонента приходится всего 2%. Средняя мощность миокарда поддерживается на уровне 1 Вт. При физической работе и эмоциях повышается КД в магистральных сосудах, увеличивается Vc , но в еще большей степени возрастает скорость кровотока. К тому же Ak зависит от квадрата этой скорости. Поэтому при физических и психических нагрузках вклад кинетического компонента в работу сердца становится весомее (до 30% всей работы), чем в покое. Например, при выполнении тяжелой физической работы тренированным человеком его ρср достигает 16 кПа, Vc − 200 мл, υ − 3 м · с 1 . Тогда работа левого желудочка достигает А = 16 · 103 · 200 · 10-6 + 1/2 103 · 200 · 10-6 · 9 = 3,2 + 0,9 = 4,1 Дж. Средняя мощность возрастает до 8,2 Вт. Приведенные числа не являются предельными. Биофизические закономерности движения крови по сосудам. Основные показатели гемодинамики. Движение крови по кровеносным сосудам изучает гемодинамика, являющаяся одним из разделов биомеханики. Принято выделять две группы показателей, интегрально характеризующих кровообращение. Их называют основными гемодинамическими показателями. Первую группу составляет скорость кровотока, вторую − кровяное давление (КД). Различают объемную и линейную скорости кровотока. Объемной скоростью (Q) называют объем жидкости (V), протекающей в единицу времени: (73) Применяются разные единицы измерения этой величины: мл · с , л · мин и др. Линейная скорость (υ) представляет собой путь (l), проходимый частицами крови в единицу времени: -1 -1 (74) Объемная и линейная скорости связаны простым отношением: (75) где S − площадь поперечного сечения потока жидкости. Для сплошного течения несжимаемой жидкости, выполняется условие неразрывности струи: через любое сечение струи в единицу времени протекают одинаковые объемы жидкости: Q = υ · S = const. Так формулируется в гидродинамике закон сохранения массы. Поскольку для кровообращения характерно сплошное течение, условие неразрывности струи выполняется и в гемодинамике, где принята его следующая формулировка: в любом сечении сердечно-сосудистой системы объемная скорость кровотока одинакова: Q = const. В покое она составляет примерно 5 л · мин-1. Словосочетание «любое сечение сердечно-сосудистой системы» требует пояснения. Одной из моделей круга кровообращения (как большого, так и малого) служит так называемая разветвленная сосудистая трубка (рис. 77, а). Каждое ее сечение представляет собой поперечный разрез всех кровеносных сосудов одного уровня ветвления. Например, в большом круге кровообращения первое сечение проходит через аорту, второе − через все артерии, на которые разветвляется аорта непосредственно, третье − через все ветви этих артерий и т. д. В одно сечение попадают все капилляры большого круга кровообращения. Площадь последнего 6 сечения большого круга (перед правым предсердием) равна сумме площадей поперечных разрезов верхней и нижней полых вен, которыми он заканчивается. Рис. 77. Схема разветвления сосудов в большом круге кровообращения (модель разветвленной сосудистой трубки) – а и соотношение между суммарным поперечным сечением каждой генерации сосудов большого круга кровообращения (S) на разных уровнях (сплошная линия) и линейной скоростью кровотока (υ) в соответствующих сосудах (штриховая линия) – б. Уровни сечения: 1 – аорта; 2 – капиллярное русло; 3 – полые вены (верхняя и нижняя) Самым узким сечением в большом круге кровообращения обладает аорта (S около 4 см2 − см. рис. 77, б). Суммарная площадь сечения обеих полых вен немного больше. Самое обширное сечение сосудистой трубки приходится на уровень кровеносных капилляров (примерно 11 000 см2, из которых лишь через 3 000 см2 течет кровь, а остальные капилляры в покое находятся в спавшемся состоянии). Следовательно, в большом круге кровообращения площадь суммарного просвета капилляров, в которых есть кровоток, в 700−800 раз больше поперечного сечения аорты. С учетом условия неразрывности струи это означает, что линейная скорость кровотока в капиплярной сети в 700−800 раз меньше, чем в аорте, и составляет около 1 мм · с-1. В покое средняя υ в аорте лежит в пределах от 0,5 до 1 м · с-1, а при большой физической нагрузке может достигать 20 м · с-1. Ускорения в аорте также максимальны и колеблются от 1 до 8 g, где g − ускорение свободного падения (9,81 м · с-2). Средняя линейная скорость кровотока в полых венах 0,10− 0,16 м · с-1, но -1 может достигать 0,5 м · с . Время полного кругооборота крови равно 27 систолам сердца, т. е. при частоте сердечных сокращений около 75 мин-1 кругооборот крови занимает 20−23 с. Из них 4/5 затрачивается на движение в большом и 1/5 − в малом кругах кровообращения. В разных точках сечения сосуда кровь движется с неодинаковой скоростью. Она максимальна на продольной оси сосуда и постепенно падает до нуля у стенки (пристеночный слой текущей жидкости неподвижен). Между слоями движущейся по сосуду крови существует градиент скорости. Его принято называть скоростью сдвига: γ = dυ/dy · [с-1], (76) Из формулы Ньютона следует, что сила внутреннего трения (вязкость) текущей жидкости определяется скоростью сдвига: Fтр = η · s · γ. Коэффициент вязкости: η =Fтр /s · γ [Па · с]. У цельной крови η, измеренный на вискозиметре, составляет около 5 мПа · с (5 сП), что в 5 раз больше вязкости воды. При патологии η крови колеблется от 1,7 до 22,9 мПа · с. Между вязкостями крови и воды обнаружена не только количественная, но и качественная разница. В воде, как и других однородных жидкостях, η не зависит от 7 скорости сдвига, а значит, и от линейной скорости. Такие жидкости называют ньютоновскими. У крови же вязкость принимает разные значения при различных скоростях сдвига. Так, η = 5 мПа · с при сравнительно большой υ крови, присущей методике определения вязкости (вискозиметрии) и движению крови в аорте. При малых скоростях сдвига, свойственных более мелким сосудам, η оказывается заметно больше: в мелких артериях − 10 мПа · с, в капиллярах − до 800 мПа · с. Кровь вместе с другими жидкостями, вязкость которых зависит от скорости сдвига, относится к неньютоновским жидкостям. Они представляют собой неоднородные жидкие среды, в основном суспензии. В крови зависимость η от скорости сдвига связана с наличием форменных элементов (прежде всего, эритроцитов). При самых низких скоростях сдвига (γ порядка 1 с-1) большая часть эритроцитов объединяется в разделенные плазмой группы из нескольких клеток. Эти конгломераты образуют жесткие структуры, что придает крови некоторые свойства твердого тела. Когда же напряжение сдвига превосходит предел текучести такого твердого тела (это достигается при повышении v), конгломераты изгибаются и разрушаются, вследствие чего вязкость крови, движущейся достаточно быстро, начинает уменьшаться, причем тем в большей степени, чем выше скорость кровотока. Следовательно, различия вязкости крови при разных скоростях (и, значит, в разных сосудах) обусловлены обратимой агрегацией эритроцитов. Вязкость медленно текущей крови (ηк) можно приближенно рассчитать по формуле: (77) где η0 − вязкость кровяной плазмы (около 1,2 мПа · с); К1 − коэффициент ≥ 2,5; С − объем форменных элементов крови в 1 см3 (около 0,42−0,45 см3). Величина ηк («кажущейся вязкости») зависит прежде всего от свойств форменных элементов крови, 93% которых составляют эритроциты. Поэтому вязкость крови определяется главным образом свойствами эритроцитов, причем не только их агрегацией. При повышении жесткости эритроцитарной мембраны, например при атеросклерозе, вязкость крови также возрастает, что приводит к увеличению нагрузки на сердце. Вязкость крови неодинакова в широких и узких сосудах, причем влияние диаметра кровеносного сосуда на вязкость начинает сказываться при просвете менее 1 мм. В сосудах тоньше 0,5 мм η уменьшается прямо пропорционально укорочению диаметра (эффект Фареуса−Линдквиста), поскольку в них эритроциты выстраиваются вдоль центральной оси в цепочку наподобие змейки и окружены слоем плазмы, изолирующей «змейку» от сосудистой стенки. В этой бесклеточной краевой зоне вязкость невелика (около 2 мПа · с), что несколько понижает величину кажущейся вязкости крови в артериолах и капиллярах, но все-таки она достигает 800 мПа · с. От вязкости крови зависит характер ее течения по сосудам, которое может быть ламинарным или турбулентным. Если слои жидкости движутся параллельно друг другу, не смешиваясь между собой, то такое течение называют ламинарным. Когда же ток жидкости сопровождается перемешиванием слоев, обусловленным образованием вихрей в ней, то говорят о турбулентном (вихревом) движении. Переход одного вида течения в другой определяется числом Рейнольдса (Re): (78) 8 где ρ − плотность жидкости; v = η/ρ − кинематическая вязкость; d − диаметр сосуда; υ − линейная скорость движения жидкости. Существует критическое значение числа Рейнольдса (Reкp), которое служит граничным параметром перехода ламинарного течения в турбулентное. Если фактическое значение Re < Reкp, то жидкости свойственно ламинарное течение, тогда как при Re > Reкp в потоке возникают вихри, и движение приобретает турбулентный характер. Для однородной жидкости Reкp = 2300, для крови Reкp = 970 ± 80, но уже при Re > 400 возникают локальные завихрения в разветвлениях артерий и в области их крутых изгибов. Ламинарное течение крови создает меньшую нагрузку на сердце, поскольку при нем работа сердца (А) прямо пропорциональна объемной скорости кровотока (Q), тогда как при турбулентном движении крови между ними установлена почти квадратичная зависимость (А ~ Q1,8). В физиологических условиях кровоток в магистральных сосудах имеет преимущественно ламинарный характер. При тяжелой физической нагрузке он может стать турбулентным из-за повышения скорости. Переход к турбулентному течению крови в сосудистом русле сопровождает ряд заболеваний, отягощая их. Например, при анемии (малокровии) Re возрастает за счет понижения кинематической вязкости крови вследствие уменьшения количества эритроцитов и нарушения их свойств. Вязкость крови оказывает существенное влияние и на второй основной гемодинамический показатель − кровяное давление (КД). Это сила, с которой движущаяся кровь воздействует (давит) на единицу площади стенки кровеносного сосуда. Анализ факторов, определяющих КД, целесообразно проводить исходя из уравнения Пуазейля: (79) где Q − объемная скорость кровотока, r − радиус сосуда, l − его длина, η − вязкость крови, р − разность давлений на концах сосуда. Если кровь течет по нескольким одинаковым сосудам, включенным параллельно, то Q = п(πr 4/8ηl) р, (80) где n − число сосудов данного типа (калибра). Для анализа факторов, от которых зависит КД, разрешим уравнение Пуазейля относительно р: р = (8ηl/πr 4) · Q и, обозначив сомножитель 8ηl/πr 4 символом Rr, запишем: р = Rr · Q. При такой записи уравнение Пуазейля обнаруживает сходство с законом Ома: U = R · I, если признать аналогию потенциальных ( U и р), а также потоковых (I и Q) величин. Заметим, что между ними существует не формальное, а смысловое сходство. Тогда Rr аналогично электрическому сопротивлению (R). И действительно, величина Rr = 8ηl/πr 4отображает сопротивление сосудистого русла кровотоку, включая все факторы, от которых оно зависит. Поэтому Rr называют гемодинамическим сопротивлением (или общим периферическим сопротивлением сосудов − ОПСС). Его единица измерения в системе СИ: [Н ·м-5 · с = Па · с · м - 3 ]. В гемодинамике обычно используют другую единицу: [Па · с · мл-1]. В большом круге кровообращения ОПСС = 140 Па · с · мл-1, а в малом − на порядок меньше (около 11 Па · с · мл-1). Вклад в ОПСС большого круга различных его частей характеризуют следующие величины: аорта и крупные артерии − 19%, артериолы − 50%, капилляры − 25%, вены − 4%, артериовенозные анастомозы и сосуды-сфинктеры − 3%. Аналогия, существующая между законами Пуазейля и Ома, позволяет моделировать кровообращение при помощи электрических цепей. В электротехнике 9 хорошо разработаны методы расчета и экспериментального исследования сложных (разветвленных) электрических цепей. Применяя эти методы к изучению кровообращения на его электрических моделях, приходят к важным теоретическим и практическим выводам о закономерностях гемодинамики. Электрическое моделирование сердечно-сосудистой системы оказалось весьма плодотворным при создании аппаратов искусственного кровообращения, в протезировании сердца и сосудов, а также во многих других работах. Анализ уравнения Пуазейля, разрешенного относительно р, свидетельствует, что КД зависит от объемной скорости кровотока и, следовательно, от массы циркулирующей крови и сократительной деятельности миокарда, определяющих эту скорость. Еще более выраженное влияние на динамику КД оказывает гемодинамическое сопротивление (ОПСС), прежде всего такой его компонент, как радиус сосуда, поскольку в формулу он входит в четвертой степени (r 4). Поэтому изменение радиуса на 20% (при Q = const) приводит к повышению КД в сосуде в (1,2)4 раз, т. е. более чем вдвое. Даже небольшие колебания просвета кровеносных сосудов сильно сказываются на кровообращении. Не случайно регуляция уровня КД в организме связана с нервными и гуморальными влияниями прежде всего на гладкомышечную оболочку кровеносных сосудов в целях активного изменения их просвета. Сюда же направлены основные фармакологические средства нормализации КД. В крупных и средних артериях КД неодинаково в систолу и диастолу). Принято различать систолическое (максимальное) и диастолическое (минимальное) КД, а также пульсовое давление крови, равное разности их и составляющее в большом круге примерно 40 мм рт. ст. (5,3 кПа), поскольку первое из них 110−130, а второе − 60−80 мм рт. ст. В малом круге кровообращения эти величины составляют соответственно 20−30 и около 10 мм, а пульсовое давление − 10−20 мм рт. ст. Кроме того, измеряют среднее КД. Названные виды кровяного давления составляют вторую группу основных гемодинамических показателей − показателей давления крови. Понятно, что между артериальным, капиллярным и венозным давлениями крови существует большая разница. В капиллярах большого круга КД падает (от 30 до 15 мм рт. ст. на протяжении всего 0,75 мм), а в малом круге капиллярное давление около 7 мм рт. ст. В венулах большого круга КД = 15−20 мм рт. ст., а в крупных венах (за пределами грудной полости) падает до 5−6 мм рт. ст. В предсердиях КД имеет отрицательное значение (на 2−4 мм рт. ст. ниже атмосферного). Волна давления, как установил Н.Е. Жуковский, распространяется по жидкости со скоростью звука (порядка 103 м · с-1). Следовательно, в самых удаленных от сердца артериях большого круга кровообращения уровни систолического и диастолического давлений достигаются мгновенно после того, как они устанавливаются в аорте. То же происходит и в малом круге относительно легочного артериального ствола. В биофизике принято выделять 2 группы основных гемодинамических показателей. Первая из них (скорость) служит потоковой характеристикой, а вторая (кровяное давление) − потенциальной. Они исчерпывающим образом характеризуют линейные процессы массопереноса, поскольку коэффициент связи между ними − постоянная величина. В гемодинамике Rr является функцией скорости, что свидетельствует о нелинейном характере движения крови по сосудам. Поэтому Rr следует считать третьим основным гемодинамическим показателем, так как он не определяется однозначно из первых двух. В биофизике принято различать четыре типа кровеносных сосудов: артерии эластического типа, артерии мышечного типа, капилляры, вены. Их функциональное 10 предназначение в системе кровообращения неодинаково. Артерии эластического типа (или амортизирующие кровеносные сосуды) поддерживают кровоток во время диастолы сердца и тем самым обеспечивают непрерывность движения крови в сосудистой системе. Артерии мышечного типа (или резистивные кровеносные сосуды) создают переменное сопротивление кровотоку и, следовательно, регулируют уровень КД в системе кровообращения, а также объемную скорость кровотока в каждом из органов. В капиллярах (обменных кровеносных сосудах) осуществляется обмен веществ между кровью и тканями, ради чего существует вся сердечно-сосудистая система. Поэтому капилляры зачастую называют центральным (главным) ее элементом. Их характерной особенностью является то, что сосудистая стенка образована одним слоем эндотелия, содержит тонкую базальную мембрану при отсутствии мышечной и адвентициальной оболочек. Диаметр капилляра от 5−7 до 10 мкм, а длина от 0,5 до 1,1 мм, но если все капилляры человека приложить друг к другу конец в конец, то общая длина составит 100 000 км. Ими можно опоясать Землю по экватору 2,5 раза. Общая площадь обмена через стенки капиллярного русла превышает 1000 м2. В1 мм3 тканей человека содержится в среднем 600 капилляров, но в разных тканях этот показатель неодинаков: в миокарде, головном мозге, почках и печени он достигает 3000, в скелетных мышцах − 300−400 (в тонических даже 100), а в костной и жировой тканях и того меньше. Линейная скорость капиллярного кровотока порядка 1 мм · с-1 (от 0,05 до 1,93 мм · с-1) и, следовательно, каждая частица крови проходит капилляр от начала до конца примерно за 1 секунду. Эритроциты могут двигаться в нем как поодиночке, так и объединенными в группы, содержащие от 3−4 до 8−10 клеток. Прослойки плазмы внутри такой группы имеют размер вдоль оси капилляра от 0,5 до 1 мкм, а между группами − по 10−15 мкм. Упакованные в одну группу эритроциты вместе с находящейся между ними плазмой составляют единое целое в кинетическом отношении, т. е. не изменяют своих параметров при прохождении через капилляры. Основная масса эритроцитов подвергается в капиллярах продольной деформации. Деформированный эритроцит имеет форму колокола, обращенного выпуклым концом по направлению кровотока. При колоколообразной деформации эритроцита площадь его поверхности увеличивается примерно на 10 %. Эритроциты могут протиснуться через капилляры и меньшего диаметра, чем 7 мкм. При исследовании этого процесса на поликарбональных фильтрах было установлено, что эритроциты проникают через поры, диаметр которых 2,4 мкм. Если скорость капиллярного кровотока падает до 0,1−0,2 мм · с-1, то эритроциты образуют агрегаты по 20−50 клеток в виде монетных столбиков. В них эритроциты сохраняют форму двояковогнутых дисков, но утрачивают способность к пластической деформации, вследствие чего могут закупорить микрососуды. Их локальную закупорку нередко создают лейкоциты, даже не объединенные в агрегаты, поскольку они деформируются в гораздо меньшей степени, чем эритроциты, под действием как продольных, так и радиальных сил. В зависимости от ультраструктурной организации стенки различают три типа капилляров: с непрерывной стенкой, причем эндотелиальные клетки с диаметром пор в плазмолемме 4−5 нм уложены наподобие черепицы − проницаемость стенки таких капилляров наименьшая по сравнению с другими (капилляры с непрерывной стенкой присущи мышцам, легким, жировой ткани); с фенестрированной (окончатой) стенкой − диаметр «окон» в ней достигает 100 нм, что обеспечивает их лучшую проницаемость, например, в почечных капиллярных клубочках и в слизистой оболочке кишки; 11 с прерывистой стенкой − за счет крупных пространств (с линейными размерами более 10 мкм), например между эндотелиальными клетками в капиллярах костного мозга, печеночных и селезеночных синусах, где проницаемость капиллярной стенки наибольшая. Вены являются емкостными кровеносными сосудами. Благодаря выраженной пластичности они могут сильно деформироваться (растягиваться) и вмещать много крови без развития напряжения в сосудистой стенке. При повышении венозного КД на 10 мм рт. ст. вместимость венозного русла большого круга кровообращения увеличивается в 6 раз. Поэтому вены называют резервуаром крови с переменной (регулируемой) емкостью. Ими образованы кровяные депо организма, которые в покое содержат 40−50% всего объема крови (примерно 1,5 л в венозных синусах селезенки, до 1 л в венозных сплетениях кожи и т. д.). Во всех венах большого круга сосредоточено до 80% крови, протекающей по нему. Кровь течет по венам под действием двух сил: «vis a tergo» (действующая сзади) и «vis a fronte» (действующая спереди). Первым термином прежде всего обозначают силу, которая обусловлена механической энергией, сообщенной крови сердцем во время систолы и сохранившейся после кровотока по артериям и капиллярам. Остаточная энергия в венулах большого круга кровообращения составляет в покое не более 13% работы левого желудочка (13% от 0,92 Дж за 1 систолу). Это основной компонент vis a tergo. Дополнительными служат массирующее действие пульсирующих рядом с венами артерий и, в большей степени, сокращения скелетных мышц, сжимающие венозные сосуды и проталкивающие кровь. Такое проталкивание действует в одном направлении (к предсердиям) благодаря наличию в венах клапанов. Движению крови в венах конечностей способствует то обстоятельство, что они образуют с артериями сообщающиеся сосуды. Вторая сила обусловлена присасывающим действием на кровь грудной клетки («дыхательного насоса» и отчасти смещения в сторону желудочков предсердножелудочковой перегородки в определенную фазу сердечного цикла). КД в правом предсердии называется центральным венозным давлением (ЦВД). На выдохе оно повышается, а на вдохе понижается. При понижении ЦВД от 0 до - 4 мм рт. ст. приток венозной крови в правое предсердие возрастает на 20−30%. Однако дальнейшее снижение ЦВД ухудшает кровоток, поскольку это приводит к спадению вен. Пределы изменений ЦВД, при которых сердце работает устойчиво, лежат в диапазоне от 0,01−0,05 кПа до 0,1−0,12 кПа. Средняя величина ЦВД у здорового человека в покое от 0,04 до 1,12 кПа. 12