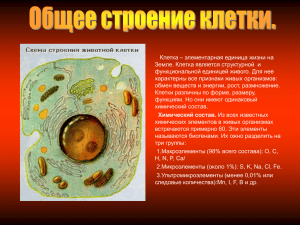
Глава 1. ЦИТОЛОГИЯ Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами. Рис.1. Объемная реконструкция клетки. 1.Ядро. 2.Ядрышко. 3.Кариолемма. 4.Ядерная пора. 5.Клеточный центр. 6.Комплекс Гольджи. 7.Эндоплазматическая сеть (ЭПС). 8.Митохондрия. 9.Плазмолемма. 10. Гликокаликс. 11.Лизосомы 12.Пероксисомы. Клетка представляет наименьшую структурнофункциональную единицу живых организмов, способную к самообновлению, саморегуляции и самовоспроизведению. Компоненты клетки распределены в ней упорядоченным образом. Выполнение этого условия обеспечивают мембраны. Мембранные структуры обособляют различные компартменты внутри клетки. Клетка – комплекс компартментов, которые делят её на функциональные зоны. Между структурными элементами клетки происходит взаимодействие благодаря движению гиалоплазмы (циклоз). Составными частями клетки являются её оболочка (плазмолемма), цитоплазма и ядро. Оболочка клетки (9) обеспечивает взаимодействие с другими клетками, с межклеточной жидкостью или другой окружающей средой. Плазмолемма (9) выполняет барьерную, транспортную, рецепторную функции. Толщина плазмолеммы около 10 нм. Она состоит из билипидного слоя в который погружены молекулы белка, которые выполняют транспортную функцию (белки – переносчики), ферментативную, являются циторецепторами. С молекулами плазмолеммы связан гликокаликс (10). Это цепочки полисахаридов, гликолипидов, гликопротеинов. Молекулы гликокаликса участвуют в сцеплении (контактах) клеток и в связывании сигнальных молекул, регулирующих деятельность клетки (гормоны, медиаторы и другие). Основной объем клетки занят гиалоплазмой (от греч. hyalos-стекло). Это коллоид из воды, ионов и различных молекул органических веществ. В гиалоплазме находятся органеллы и включения. Органеллы – это структурные элементы, выполняющие необходимые для клетки функции. Органеллы, присущие всем клеткам, относят к органеллам общего назначения. Среди них различают органеллы мембранные и немембранные. В приведенном рисунке – схеме отражены мембранные органеллы: гладкая и гранулярная ЭПС (7), комплекс Гольджи (6), лизосомы (11), митохондрии (8), пероксисомы (12). Каждая из указанных органелл имеет на своих мембранах ферменты, обеспечивающие специфику их деятельности. Немембранные органеллы: клеточный центр, рибосомы, элементы цитоскелета (микротрубочки и промежуточные филаменты), а также актиновые и миозиновые микрофиламенты. Клеточный центр (5) имеет две центриоли (диплосомы), расположенные под углом друг к другу. Каждая центриоль – цилиндр, стенка которого окружена 9 триплетами микротрубочек длиной около 0,5 мкм. Выделяют дочернюю и материнскую центриоль. Вокруг последней формируется центросфера из бесструктурного тонковолокнистого матрикса в который входят концы микротрубочек. Собранные микротрубочки участвуют в структурах цитоскелета. Центриоли в делящихся клетках формируют веретено деления. Рибосомы обеспечивают синтез белков, состоят из двух субъединиц – большой и малой. Каждая субъединица – комплекс рибосомальной РНК с белками. Большая субъединица содержит 3 молекулы рРНК, малая – одну. Число рибосом в клетке может достигать нескольких миллионов. Те рибосомы, которые не связаны с мембранами ЭПС (свободные рибосомы) синтезируют белки для жизни самой клетки. Рибосомы гранулярной ЭПС (7) синтезируют белки для выведения из клетки. В ядрышке образуются рибосомы, которые через ядерные поры (4) мигрируют в цитоплазму клеток. Ядро составная часть клетки, содержит кариолемму (3), ядрышко (2), кариоплазму. Размеры ядер могут варьировать. В ядре молекулы ДНК ассоциированы с белками – гистонами. Каждая хромосома образована такими молекулами. Хромосомы – важнейшие структуры, обеспечивающие через синтез ферментов метаболизм клеток. Назовите структуры, обозначенные цифрами и расскажите об их функциях. Рис. 2. Схема строения плазмолеммы. 1.Гидрофильные головки фосфолипидов плазмолеммы. 2.Гидрофобные хвосты фосфолипидов. 3.Микротрубочка. 4.Микрофиламент (промежуточный). 5.Интегральные белки. 6.Поверхностный белок. 7.Гликокаликс. 8.Наружный слой липидной мембраны 9.Внутренний слой липидной мембраны. 10.Полуинтегральные белки. Клеточная мембрана (цитолемма, плазмолемма) отграничивает клетку от внешней среды, осуществляет ее взаимодействие с внешней средой, в том числе и с другими клетками, обеспечивает избирательную проницаемость веществ, участвует в поддержании гомеостаза. Согласно современным представлениям, клеточная мембрана, как и остальные мембраны клетки, состоит из двух слоев липидов со встроенными в них белками. Липиды составляют от 25 до 40% массы. Это, в основном, фосфолипиды, сфинголипиды и холестерин. Их полярная часть (головка) является гидрофильной (1) и располагается поверхностно. Гидрофобная же часть (2) погружена внутрь мембраны. Белки составляют около 60% массы и обеспечивают специализированные функции мембраны. Это глобулярные белки. Они формируют скопления в плазмолемме. По степени погружения в мембрану белки могут располагаться поверхностно (с наружной или внутренней поверхности) (6), пронизывать всю толщу мембраны – интегральные белки (5); или глубоко погружаться, но не достигать одной из сторон – полуинтегральные белки (10). Интегральные белки играют роль в осуществлении трансмембранных процессов. С учетом разнообразия функций белки можно весьма условно разделить на группы: рецепторные белки - лежат поверхностно, взаимодействуют с внешними сигнальными молекулами; белки-переносчики - осуществляют трансмембранный перенос; адгезивные белки - обеспечивают прикрепление клетки к другим клеткам или к элементам межклеточного вещества; ферменты; структурные белки, в том числе и осуществляющие опорно-каркасные функции (они чаще всего связаны с микротрубочками, микрофиламентами и филаментами с внутренней стороны мембраны и с адгезивными белками с внешней стороны). Углеводы содержатся на внешней поверхности клеточной стенки и участвуют в формировании гликокаликса (7). Это, в основном, олигосахариды, связанные с пептидами (гликопротеины) или липидами (гликолипиды). Гликопротеины имеют высокую видовую, индивидуальную и тканевую специфичность. В подмембранные комплексы входят элементы цитоскелета (3,4). Эти элементы могут участвовать в формировании плотных и десмосомальных межклеточных контактов. Транспортные процессы через мембрану (трансмембранный перенос): 1.Пассивная диффузия по градиенту концентрации. Осуществляется в обе стороны в зависимости от парциального давления (концентрации) того или иного вещества. Не требует затрат энергии. 2. Облегченная диффузия. Отличается от пассивной тем, что возможность пассивной диффузии по градиенту концентрации может осуществляться только в одном направлении (либо в клетку, либо из нее). Облегченная диффузия осуществляется через специальные белки-переносчики, являющиеся интегральными, высокоспециализированными белками, осуществляющими перенос строго определенных веществ, зависящий от функциональной активности клетки. Они могут быть «открытыми», формируя «каналы», через которые проникает вещество, или «закрытыми». 3. Активный транспорт через клетку осуществляется с затратами энергии, против градиента концентрации, высокоспецифичен и нередко сопряжен с транспортом двух веществ в противоположном направлении. Так в клетку попадают ионы калия и одновременно выводятся ионы натрия. Высокомолекулярные вещества, макромолекулярные комплексы могут транспортироваться в клетку или выводиться из нее путем эндоцитоза или экзоцитоза. 2 Назовите структуры, обозначенные цифрами. Укажите функции этих структур. Рис. 3. Ядро и цитоплазма клетки. 1.Ядрышко. 2.Гетерохроматин. 3.Эухроматин. 4.Ядерная оболочка. 5.Ядерная пора. 6.Митохондрия. 7.Лизосома. 8.Гранулярная ЭПС. 9.Комплекс Гольджи. 10.Кариоплазма. Ядро состоит из кариоплазмы (10) и кариолеммы (4). В кариоплазме, представляющей коллоид в виде геля, можно выделить ядрышко (1), матрикс, хроматин (2,3). В ядре имеется от одного до нескольких ядрышек. Они могут иметь различные размеры, форму, плотность и область распределения в зависимости от функциональной активности клетки. Основная функция ядрышка – синтез рибосомальной РНК (рРНК) и субъединиц рибосом. Хроматин ядра – комплекс дезоксирибонуклеиновой кислоты с белками, где ДНК находится в различной степени спирализации. По электронной и светооптической плотности выделяют электронноплотный, грубо окрашенный гетерохроматин (2) и более нежно окрашенный, менее электронно-плотный эухроматин (3). Гетерохроматин - зона сильно конденсированной ДНК. При электронной микроскопии формирует темные глыбки неправильной формы. В гетерохроматине ДНК, в основном, связана с гистоновыми белками. Гетерохроматин представляет собой плотно упакованные скопления нуклеосом. В зависимости от локализации подразделяется на пристеночный, матричный и перинуклеарный. Пристеночный гетерохроматин прилежит к внутренней поверхности ядерной оболочки, матричный – распределен в матриксе кариоплазмы. Как вариант матричного – перинуклеарный гетерохроматин, примыкает к ядрышку. Эухроматин – это область слабо конденсированной ДНК. С нуклеиновыми кислотами в эухроматине связаны, в основном, негистоновые белки (проявляют менее выраженные основные свойства, более разнообразны по химическому составу). Ядерная оболочка (кариолемма) (4) отграничивает содержимое ядра от цитоплазмы. Представлена двумя мембранами и перинуклеарным пространством между ними (перинуклеарная цистерна). В ней имеются ядерные поры – области слияния наружной и внутренней мембран (5). Ядерная пора (5) обеспечивает избирательный транспорт макромолекул. Через нее свободно диффундируют по градиенту концентрации молекулы воды, растворенные в ней газы, неорганические ионы, низкомолекулярные органические вещества. Содержимое ядра отличается от цитоплазмы по составу органических веществ высокого молекулярного веса (ферменты, макромолекулярные соединения); по составу низкомолекулярных веществ, приближено к матриксу цитоплазмы. На внутренней и наружной поверхности комплекса ядерной поры имеются высокоспецифичные рецепторы, обеспечивающие транспорт из ядра субъединиц рибосом, информационной РНК (иРНК), транспортной РНК (тРНК) и некоторых других веществ. В ядро же избирательно транспортируются ферменты, белки ламин, гистоны. Транспорт макромолекул происходит активно, а белки комплекса ядерной поры обладают АТФ-азной активностью. Наружная ядерная мембрана по набору рецепторов и составу аналогична гранулярной или агранулярной ЭПС. Внутренняя ядерная мембрана участвует в формировании ядерной пластинки. Ядерная пластинка структурирует ядерную оболочку и перинуклеарный хроматин. Содержит белки – ламины, которые прикрепляются к белкам внутренней мембраны, выполняющим опорно-каркасные функции. Ламины образуют фибриллярные структуры, аналогичные промежуточным филаментам цитоплазмы. К внутренней поверхности ядерной оболочки нередко прикрепляется гетерохроматин (пристеночный). Перинуклеарная цистерна (пространство) характеризуется низкой электронной плотностью. Состав ферментов аналогичен содержимому гранулярной ЭПС. Цитоплазма состоит из матрикса (гиалоплазма), органелл и включений. Гиалоплазма может иметь свойства золя или геля. Органеллы – постоянно присутствующие структурные элементы цитоплазмы. Из органелл представленных на рисунке: митохондрии (6), лизосомы (7), гр. ЭПС (8), комплекс Гольджи (9). 3 Назовите структуры, обозначенные цифрами. Расскажите о последовательности процессов в ходе взаимодействия гранулярной ЭПС и комплекса Гольджи. Рис. 6. Схема взаимодействия гранулярной эндоплазматической сети и комплекса Гольджи. 1.Гранулярная эндоплазматическая сеть (ЭПС). 2.Гладкая эндоплазматическая сеть (ЭПС). 3.Комплекс Гольджи. 4.Диктиосомы. 5.Транспортные пузырьки (от ЭПС). 6.Лизосома. До недавнего времени не были раскрыты механизмы взаимодействия эндоплазматической сети и комплекса Гольджи. В настоящее время доказано наличие конвейера в воспроизводстве веществ в клетке. Первым блоком этого конвейера является эндоплазматическая сеть, а вторым комплекс Гольджи. Эти блоки взаимосвязаны. Эндоплазматическая сеть представлена трубочками, цистернами, мембранами, пузырьками. Большинство веществ синтезируется на наружной поверхности мембран. Затем вещества переносятся внутрь каналов эндоплазматической сети. На конце каналов эндоплазматической сети вещества накапливаются, а затем отделяются в виде транспортных пузырьков (5). Если образуются вещества для обновления мембран, то сборка мембран происходит в комплексе Гольджи, там же образуются лизосомы (6). Комплекс Гольджи является местом конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном, в эндоплазматической сети. При электронной микроскопии комплекс Гольджи состоит из скоплений плоских цистерн (диктиосом)(4) и пузырьков. Наиболее широкие уплощенные цистерны обращены в сторону эндоплазматической сети. Транспортные пузырьки от эндоплазматической сети переходят к цистернам (цис-полюс) комплекса Гольджи и там происходит их дальнейшая трансформация: образуются комплексы белков с углеводами и липидами. Образованные вещества переходят из одних цистерн в другие. На белковых концах цистерн (транс-полюс) образуются выпячивания, которые отщепляются в виде пузырьков и направляются в гиалоплазму или к плазмолемме для выведения, а в комплекс Гольджи поступают новые порции продуктов, синтезированных в гранулярной эндоплазматической сети. 4 Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами. Рис. 7. Фрагмент клетки. Митохондрии. 1.Крупная митохондрия с трубчатыми кристами. 2.Деление митохондрии путем почкования. 3.Митохондрии овальной формы с трабекулярными кристами. 4.Ядро. 5.Гранулярная эндоплазматическая сеть. 6.Трубчатая криста. 7.Трабекулярная криста. 8.Наружная мембрана. 9.Внутренняя мембрана. 10.Перимитохондриальное пространство. 11.Матрикс митохондрии. Митохондрии имеют средние поперечные размеры 0,5 – 3 мкм. В зависимости от размеров можно выделить мелкие, средние, крупные, гигантские органеллы. По форме они могут быть округлые, овальные (3) и нитевидные. Митохондрии состоят из матрикса (11); внутренней мембраны (9), образующей кристы (6,7) (впячивания в матрикс); перимитохондриального пространства (10) и наружной мембраны (8). Форма крист может быть в виде пластинок (7)(трабекулярные) и трубочек (6) (мультивезикулярные). Матрикс (11) митохондрии гомогенный, иногда мелкозернистый, различной электронной плотности. В матриксе содержатся ферменты, одноцепочечная циклическая ДНК, митохондриальная РНК, митохондриальные рибосомы, высокая концентрация ионов Са2+. ДНК митохондрий близка к ДНК прокариот. По строению митохондриальные рибосомы близки к рибосомам прокариотических организмов, но меньшего размера. Внутренняя мембрана митохондрии (9) обладает избирательной проницаемостью для электронов, образующихся при аэробном фосфорилировании и слабопроницаема для других веществ. К ее внутренней поверхности прикрепляются тесно прилежащие друг к другу ферменты цепи окислительного фосфорилирования. Наружная мембрана митохондрии (8) более проницаема для низкомолекулярных веществ. На поверхности наружной мембраны имеются специфические рецепторные комплексы, через которые белки из матрикса могут транспортироваться в перимитохондриальное пространство, или, за счет временного слияния с внутренней мембраной, попасть в матрикс митохондрии. В митохондриях имеются два автономных компартмента. Перимитохондриальное пространство (внешний компартмент) формируется за счет проникновения белковых комплексов гиалоплазмы, и внутренний компартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК. Во внутреннем компартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са 2+ в сравнении с гиалоплазмой. Ферментная активность внешнего и внутреннего компартментов, состав белков также сильно различаются. Внутренний компартмент нередко имеет более высокую электронную плотность, чем внешний. Специфическими маркерами митохондрии являются ферменты цитохромоксидаза и сукцинатдегидрогеназа. Некоторые авторы считают, что в клетках не множество, а одна гигантская митохондрия, которая многократно попадает в срез препарата, однако большинство исследователей указывает на ошибочность данной концепции, во всяком случае, для большинства клеток организма человека. Новые митохондрии образуются путем деления, способом перетяжки или почкования (2). В последнем случае образуется протомитохондрия, постепенно увеличивающаяся в размерах. При размножении в каждой из вновь образующихся митохондрий имеется свой собственный геном. Старые митохондрии разрушаются путем аутолиза (самопереваривания клеткой с помощью лизосом) с образованием аутолизосом. Из аутолизосомы образуется остаточное тельце. При полном переваривании содержимое остаточного тельца выводится путем экзоцитоза. При неполном переваривании остатки митохондрий могут накапливаться в клетке в виде слоистых телец или гранул с липофусцином. Часть митохондрий может обызвествляться с образованием нерастворимых солей кальция (кальцинатов). 5 Назовите структуры, обозначенные цифрами, укажите их функции. Рис. Клеточный центр. 1.Дочерняя центриоль. 2. Тонковолокнистый матрикс. 3. Триплет из микротрубочек 4. Микротрубочки центросферы. 5. Материнская центриоль. 6.Ручки с динеинами. Клеточный центр образован двумя перпендикулярно расположенными центриолями (1) и центросферой. Центриоли имеют цилиндрическую форму. Их поперечный диаметр составляет около 150 нм, а длина – 500 нм. Выделяют дочернюю и материнскую центриоли (5). Вокруг материнской центриоли формируется центросфера (4) из бесструктурного тонковолокнистого матрикса (2), в который входят концы микротрубочек. Центриоли составлены триплетами микротрубочек (3). Соседние триплеты взаимодействуют между собой с помощью ручек, образованных молекулами белка динеина (6). Динеин обладает АТФазной активностью и способен вызвать смещение (скольжение) одного триплета микротрубочек по отношению к другому, обеспечивая движение. Клеточный центр в интерфазной клетке организует ее компартменты, участвует в циклозе. При митотическом и мейотическом делении в животной клетке центриоли обеспечивают равномерное расхождение генетического и цитоплазматического материала делящейся клетки, участвуют в формировании веретена деления. Во время деления центриоли расходятся к полюсам, формируя новые клеточные центры клеток. Центриоли способны индуцировать синтез новых центриолей, до нескольких десятков или даже сотен на одну клетку. Вновь образуемые центриоли перемещаются на периферию и служат основой для формирования ресничек, формируя в них базальное тельце. 6 Назовите структуры, обозначенные цифрами, объясните функции этих структур. Рис. 11. Специальные структуры поверхности клетки. Реснички в эпителиоцитах. 1. Ресничка. 2.Аксонема. 3.Базальное тельце. 4.Десмосома. 5.Базальная мембрана. 6. Плазмолемма. Реснички относят к специализированным органоидам клетки. Они присутствуют в эпителии воздухоносных путей, маточных труб и обладают подвижностью. Ресничка (1) представляет собой выпячивание цитоплазмы, окруженное клеточной мембраной (6) диаметром 300 нм. В основании ресничек мелкие гранулы – базальные тельца (3), которые погружены в цитоплазму. К базальному тельцу могут прикрепляться спутники от которых отходят микротрубочки. Внутри реснички находится аксонема («осевая нить») (2), выступающая в просвет или полость органа, выстланного реснитчатым эпителием. Аксонема состоит из микротрубочек. Микротрубочки собраны в комплексы в виде дуплетов: по 9 пар на периферии и одна пара в центре. Каждая из двух первых микротрубочек девяти триплетов базального тельца является основой для дуплетов микротрубочек цитоплазматического выроста. Таким образом, две микротрубочки триплетов базального тельца являются микротрубочками дуплетов аксонемы. Соседние дуплеты соединяются друг с другом с помощью «ручек» из белка динеина, который обладает АТФ-азной активностью. В присутствии ионов кальция динеины сокращаются, потребляя при этом энергию, и микротрубочки скользят относительно друг друга. Все микротрубочки достигают своим дистальным отрицательным полюсом гомогенный матрикс аксонемы. Таким образом, базальное тельце и аксонема структурно связаны и составляют единое целое. Основной белок ресничек – тубулин к сокращению не способен. Смещения дуплетов микротрубочек относительно друг друга вызывают изгиб всей реснички. Движение микроресничек обеспечивает белок динеин. Реснички образуются за счет центриолей: от материнской синтезируется дочерняя. Вновь синтезированная органелла смещается к периферии. Процесс многократно повторяется и под мембраной образуется много базальных телец, которые являются основой для образования ресничек. Движения ресничек зависят от содержания внутриклеточного кальция, АТФ, ионного состава межклеточного вещества и т.д. 7 Назовите структуры, обозначенные цифрами, укажите функции этих структур. Рис. 12. Специальные структуры поверхности клетки: микроворсинки в эпителиоцитах 1.Гликокаликс. 2.Микроворсинки. 3.Межклеточный контакт в виде «замка». 4.Десмосома. 5. Плотный контакт Микроворсинки (2) видны под световым микроскопом как тонкие выросты клетки. За счет микроворсинок увеличивается площадь взаимодействия клетки с внешней средой. Микроворсинки состоят из клеточной мембраны, гиалоплазмы и тонких микрофиламентов (актиновых). Последние располагаются параллельно поверхности мембраны в виде упорядоченных пучков. Положительный полюс направлен к периферии и связывается с электронноплотным веществом «верхушки» микроворсинки. В основании микроворсинки актиновые филаменты вплетаются в сеть подобных структур, которые образуют кутикулу. С мембранами тонкие (актиновые) филаменты взаимодействуют с помощью минимиозина и виллина. Взаимодействие с минимиозином способствует сокращению микроворсинки. Микроворсинки являются неотьемлемой частью эпителиоцитов тонкой и толстой кишки, канальцев почки. В тонкой кишке ферменты, прикрепленные к гликокаликсу (1) и находящиеся в толще билипидного слоя мембраны микроворсинок, обеспечивают всасывание, переваривание веществ на поверхности клеток. Большое количество микроворсинок в клетке создает между ними узкие щели. В этих пространствах действуют капиллярные силы, способствующие присасыванию жидкости. Величина и размеры микроворсинок непостоянны. В почках микроворсинки клеток канальцев нефрона всасывают воду и электролиты, последние переходят в кровь. 8 Назовите элементы микротрубочки. Назовите структуры, в которых есть микротрубочки. Опишите варианты функций микротрубочек. Рис. 13. Схема строения микротрубочки. 1. Димер микротрубочки. 2. Просвет микротрубочки. 3. Ассоциированные белки. Микротрубочки представляют собой полые цилиндры диаметром 20 – 30 нм, внутренний просвет имеет ширину 15 нм (2). Являются элементами цитоскелета. Микротрубочки эволюционно древние образования клеток. Они участвуют в делении клеток, входят в состав центриолей, микроресничек, цитоскелета. В клетках микротрубочки обеспечивают внутриклеточный транспорт, в нейронах участвуют в аксотоке. Микротрубочки могут быть разбросаны в цитоплазме, либо собраны в виде пучков, например, в составе митотического веретена, а могут сливаясь друг с другом формировать пары ( в жгутиках), триплеты (в центриолях). Микротрубочки состоят из глобулярных белков – тубулинов, на поперечном срезе их 13. Тубулины подразделяются на альфа и бета тубулины. Альфа тубулины соединяются в виде цепочки, которая формирует спираль. Одновременно, в виде параллельной спирали полимеризуются бета-тубулины, т.е. образуются димеры тубулина (1). Таким образом, тубулины могут быть в двух формах – глобулярной (диспергированной в матриксе) и фибриллярной – в виде микротрубочек. С поверхностью микротрубочки могут быть связаны ассоциированные белки (3). Если эти белки сократительные (динеин), то их сокращение приводит к изменению положения и формы микротрубочки. Микротрубочки имеют полярность. У них есть положительный (+) и отрицательный (-) концы. Со стороны (-) полюса, лежащего ближе к клеточному центру, тубулины полимеризуются медленнее и легко распадаются до глобулярных частиц. Со стороны (+) конца легче осуществляется полимеризация и он направлен на периферию клетки. Воздействие на микротрубочки может привести к нарушению их функций и реконструкции, к их разрушению. Это используют в лечении больных со злокачественными опухолями, при этом нарушается деление злокачественных клеток. 9 Дайте название процессу и его фазам. Опишите изменения, происходящие на каждой из приведенных схем. Рис. 14. Митоз. 1. Клетка в интерфазе. 2, 3 Профаза. 4, 5. Метафаза. 6, 7.Анафаза. 8.Телофаза. 9.Дочерние клетки. В предмитотической фазе (1) в клетке удваивается число хромосом. В профазе (2,3) прекращается транскрипция, начинается спирализация хромосом (стадия рыхлого клубка), исчезает ядрышко. Распадается оболочка ядра. К концу профазы видны хромосомы, каждая из которых состоит из двух хроматид. Хроматиды сплетены и не видны отдельно. Характерной чертой профазы является формирование веретена деления. К каждому полюсу отходят по две центриоли и от них образуются микротрубочки. Формирование микротрубочек обеспечивает полимеризация белков тубулинов. С микротрубочками связываются хромосомы. Метафаза (4, 5) занимает 20-30 минут. В этот период завершено образование веретена деления и хромосомы распределяются в области экватора. К концу метафазы сестринские хроматиды разделяются. В анафазе (6,7) сестринские хроматиды становятся самостоятельными хромосомами и расходятся к полюсам формирующихся клеток. Это происходит при участии микротрубочек. Телофаза (8) является окончанием митоза. Ранняя телофаза – это завершение расхождения хромосом. В поздней телофазе начинается формирование новых ядер, обособление генетического материала. Завершается поздняя телефаза разделением исходной клетки на две дочерние (9) (цитокинез или цитотомия). Хромосомы начинают транскрибировать РНК. К концу телофазы ядрышко полностью сформировано. 10 Назовите структуры, обозначенные цифрами. Укажите функции, выполняемые этими структурами. Рис. 15. Фрагмент клетки. Процессы синтеза, накопления и выделения секрета. 1.Гранулярная ЭПС. 2.Транспортные пузырьки. 3.Цис-компартмент комплекса Гольджи. 4.Промежуточный компартмент комплекса Гольджи. 5.Транс-компартмент комплекса Гольджи 6.Секреторные включения. 7.Секреция (экзоцитоз). 8.Митохондрия. 9.Агранулярная ЭПС. 10.Клеточный центр. 11.Ядро. Местом конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки (в основном, в эндоплазматической сети) является комплекс Гольджи. Под электронным микросокпом органелла представляет собой скопления плоских цистерн, которые называются диктиосомы. Таких скоплений в клетке может быть несколько. Цистерны плотно прилежат друг к другу. В центральной части просвет между ними невелик, а на периферии цистерны образуют расширения – ампулы, ширина которых непостоянна. В каждой стопке около десяти цистерн. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул). Их больше по краям комплекса Гольджи. Одни авторы полагают, что цистерны взаимодействуют между собой за счет транспортных пузырьков (2), другие – что цистерны имеют участки слияния. Комплекс Гольджи поляризован, т.е. качественно не однороден с разных сторон и состоит в свою очередь из нескольких компартментов. Он имеет незрелую цисповерхность, лежащую ближе к ядру (11), и зрелую транс-поверхность, обращенную к наружной мембране клетки. Таким образом, органелла состоит из нескольких компартментов, которые, выполняя специфические функции, функционально взаимосвязаны между собой. Цис-компартмент (3) обычно обращен к клеточному центру (10). Его внешняя поверхность обычно имеет выпуклую форму. С его цистернами сливаются микровезикулы (транспортные пиноцитозные пузырьки), направляющиеся из эндоплазматической сети. Промежуточный компартмент (4) составлен несколькими цистернами в центре органеллы. Осуществляет гликозилирование (присоединяет углеводные цепочки), фосфорилирование (присоединение фосфорных остатков), карбоксилирование (присоединение карбоксильных групп), синтез гликолипидов и липопротеинов. В нем, как и в цис-компартменте, происходит формирование третичных и четвертичных белковых комплексов. Транс-компартмент (5) располагается ближе к периферии клетки. Внешняя поверхность обычно вогнутая. Частично он переходит в транс-сеть – систему везикул, вакуолей и канальцев. Отдельные диктиосомы могут быть связаны друг с другом системой пузырьков и цистерн, примыкающих к дистальному концу скопления плоских мешков, так что образуется рыхлая трехмерная сеть, выявляемая в световом и электронном микроскопах (транс-сеть комплекса Гольджи). В структурах транс-компартмента происходит сортировка (сегрегация) белков и других веществ, образование секреторных гранул, предшественников первичных лизосом и пузырьков спонтанной секреции. Секреторные пузырьки и прелизосомы являются окаймленными и окружены белком – клатрином. Окаймленные пузырьки отщепляются от транс-сети и их перемещение является гормонозависимым. Процесс транспортировки окаймленных пузырьков находится под влиянием микротрубочек. Белковые комплексы вокруг пузырьков распадаются после отщепления пузырька от транс-сети и вновь формируются в момент секреции. Пузырьки спонтанной секреции не окружены клатрином, их формирование происходит непрерывно и они, направляясь к клеточной мембране, сливаются с ней, обеспечивая ее восстановление. Таким образом, в комплексе Гольджи происходит формирование сложных биополимерных комплексов и, в частности, гликозилирование белков и липидов, в результате чего образуются гликопротеины, гликолипиды. В органелле формируются третичные и четвертичные структуры белков, частично происходит созревание секрета, его конденсация и упаковка. 11 . Назовите структуры, обозначенные цифрами. Опишите сущность процессов фагоцитоза и пиноцитоза. Рис. 18. Фагоцитоз и пиноцитоз. 1.Макромолекулярная структура. 2.Начало фагоцитоза. 3.Фагосома. 4.Фаголизосома (вторичная лизосома). 5.Гранулярная ЭПС. 6.Митохондрия. 7.Первичная лизосома. 8.Эндосома. 9. Аппарат Гольджи. 10.Фрагмент ядра. 11.Плазмолемма. 12.Остаточное тельце. Существует несколько способов эндоцитоза (от греч. endon – внутри, kytos – клетка: пиноцитоз (от греч. pino – пью) и фагоцитоз (от греч. phagos – пожирающий). При пиноцитозе клетка захватывает жидкие коллоидные частицы, а при фагоцитозе – плотные частицы (макромолекулярные комплексы, части клеток, бактерии и др.). В этих процессах активную роль играют плазмолемма (11) и гликокаликс. При пиноцитозе и фагоцитозе вбираемые клеткой частицы взаимодействуют с плазмолеммой и окружаются ею (2). Жидкие частицы окаймляются дополнительно белком клатрином (окаймленные пузырьки) (12). В последующем и при пиноцитозе и при фагоцитозе происходит взаимодействие захваченных клеткой веществ с лизосомами. Фагоцитоз характерен для клеток макрофагов, которые находятся в рыхлой соединительной ткани в каждом органе, нейтрофилов и др. При фагоцитозе бактерий, частей клеток специализированными клетками идет взаимодействие фагоцитируемой частицы с рецепторами на поверхности клетки и активация фагоцитоза с изменением внутриклеточного содержания кальция. Это ведет к изменению полимеризации тонких микрофиламентов и микротрубочек, вызывает формирование выпячивания цитолеммы (псевдоподии) с погружением крупной частицы внутрь клетки (образование фагосомы). В дальнейшем эндоцитозные пузырьки (эндосомы) (8) могут сливаться друг с другом и внутри пузырьков, кроме поглощенных веществ обнаруживаются гидролитические ферменты, которые поступают из лизосом. Ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиалоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению. 12 Назовите структуру, приведенную на фотограмме. Укажите детали, обозначенные цифрами и их значение. Рис. 19. Структура десмосомы. 1. Плазмолеммы контактирующих клеток 2. Пятно десмосомы 3. Промежуточные филаменты 4. Десмоглеины в межклеточном пространстве. При развитии человека в ходе гисто– и органогенеза происходит интеграция клеток по тканевому и дифферонному принципу. Одним из элементов такой интеграции является межклеточное взаимодействие в форме межклеточных контактов. Разнообразие межклеточных соединений отражает многофункциональность клеток. Важную роль играет обмен ионами между клетками. Эта функция обеспечивается щелевидными контактами (нексусами). Подобный вид взаимодействия возможен при надежном механическом сцеплении клеток. Сцепления формируют межклеточные соединения называемые десмосомы. Десмосома представляет собой сложное макромолекулярное образование. Это локальный участок в форме диска, в котором мембраны соседних клеток (1) располагаются на расстоянии 30 – 40 нм. На внутренней поверхности мембран обеих взаимодействующих клеток имеются участки с высокой электронной плотностью – пятно десмосомы (2) с белками десмоплакинами, в которые входят промежуточные филаменты (3) и микротрубочки. Главными связующими элементами соседних клеток являются интегральные белки десмоглеины, которые, пронизывая плазмолеммы клеток, сцеплены в межклеточном пространстве (4). Чаще всего десмосомы можно встретить в эпителиях, но они имеются и в других тканях. 13 Глава 2. ТКАНИ Фрагмент какой ткани изображен на рисунке. Назовите структуры, обозначенные цифрами. Рис. 1. Десмосомы в шиповатом слое клеток эпидермиса кожи. 1.Ядра кератиноцитов шиповатого слоя. 2. Цитоплазма кератиноцита. 3.Десмосомы. 4. Простые контакты. 5.Пластинки прикрепления. 6.Межклеточная щель в области десмосомы. 7.Электронноплотная полоска по средней линии щели десмосомы. 8. Цитокератиновые тонофиламенты. 9. Митохондрии. 10. Комплекс Гольджи. 11. Гранулярная ЭПС. 12. Рибосомы. 13. Пучки тонофиламентов в цитоплазме. Эпидермис кожи является многослойным плоским ороговевающим эпителием, основным клеточным типом которого являются клетки кератиноциты. От базальной мембраны эпидермиса до его внешней поверхности (где находятся роговые чешуйки) располагаются колонки кератиноцитов единого происхождения, но на разных стадиях созревания. Толщина эпидермиса может быть различна. На данном рисунке кератиноциты шиповатого слоя эпидермиса кожи: фрагменты их ядер (1) и цитоплазмы (2). Между клетками два вида соединений: десмосомы (3) и простые контакты (4). При фиксации ткани клетки сморщиваются, цитолеммы соседних клеток расходятся и между ними находится свободное пространство, а в области десмосом клетки остаются тесно связанными. Таким образом, клетка приобретает «шиповатый» вид. Десмосома (пятно слипания) – сложный кнопковидный межклеточный контакт, скрепляющий клетки друг с другом. На внутренней поверхности мембран взаимодействующих клеток имеются участки с высокой электронной плотностью за счёт скопления трансмембранных (десмоглеин и десмоколлин) и цитоплазматических (плакоглобин и десмоплакин) белков. Эти участки, диаметром до 0,5 мкм, толщиной 15 нм, имеют дисковидную форму, и называются пластинками прикрепления (5). В десмосоме между цитолеммами клеток находится щель (6), шириной 25 нм, соответствующая их гликокаликсам. Электронноплотная полоска по средней линии щели (7) соответствует линии, по которой переплетаются белковые нити: внемембранные части трансмембранных Са2+-связывающих белков семейства кадгеринов десмоглеина и десмоколлина, отходящие от пластинок прикрепления, связывая клетки в единую систему. Т.е., белки пронизывая плазмолеммы контактирующих клеток сцеплены в межклеточном пространстве. С цитоплазматической стороны пластинки в десмосоме прикрепляются промежуточные филаменты (они толще микрофиламентов, но тоньше микротрубочек). В клетках находятся цитокератиновые тонофиламенты (8), которые состоят из предшественников кератина. Каждый тонофиламент подходит к пластинке, прошивает её и поворачивает обратно, не проходя через цитолемму. В цитоплазме кератиноцита шиповатого слоя эпидермиса развиты органеллы общего значения: митохондрии (9), комплекс Гольджи (10), грЭПС (11), рибосомы (12), необходимые для синтеза белков, участвующих в ороговении, а также в образовании десмосом; в ядре преобладает эухроматин, что говорит о синтетической активности клеток, т.е. локализация данного участка – нижние слои шиповатого слоя. Чем выше от базального слоя, тем ниже синтетическая и митотическая активность кератиноцитов. В цитоплазме шиповатых клеток накапливаются пластинчатые гранулы (кератиносомы), содержимое которых выделяется в межклеточное пространство, обеспечивая водонепроницаемость кожи. Тонофиламенты являются элементами цитоскелета (13), это - органеллы специального назначения в кератиноцитах, поскольку именно шиповатый слой отвечает за механическую прочность эпидермиса (в котором также выделяют базальный, зернистый, блестящий, не везде, роговой слои). В базальном слое или в основании шиповатого находятся эпидермальные макрофаги, которые выделяя тканевые гормоны, влияют на процессы созревания кератиноцитов. Кроме того, клетки базального слоя являются матричными (стволовыми). В базальной части шиповатого слоя клетки сохраняют способность к делению. 14 Фрагмент какой ткани на схеме? Фрагмент какого органа на схеме? Аргументируйте вывод. Рис. 2. Переходный эпителий мочевого пузыря. 1.Базальная мембрана. 2.Базальный слой эпителия. 3.Промежуточный слой эпителия. 4.Поверхностный слой эпителия. 5.Пластинки плазмолеммы. 6.Дисковидные пузырьки. 7.Микрофиламенты. 8.Подлежащая соединительная ткань. Переходный эпителий – особый вид многослойного эпителия, который выстилает большую часть мочевыводящих путей (почечные чашечки, лоханки, мочеточники, мочевой пузырь). Форма клеток и толщина этого эпителия зависят от степени растяжения органа. При наполненном мочевом пузыре эпителий более тонкий, клетки поверхностного слоя уплощены. При сокращении мышечной оболочки мочевого пузыря толщина эпителия увеличивается. Часть клеток промежуточного слоя при этом имеет грушевидную форму (клетки «выталкиваются» к поверхностному слою). В спавшемся состоянии в слизистой мочевого пузыря имеются многочисленные складки. Они отсутствуют в области дна мочевого пузыря в месте впадения мочеточников. В этом участке слизистая оболочка, состоящая из переходного эпителия и собственной пластинки слизистой лишена подслизистой основы и плотно срастается с мышечной оболочкой. Переходный эпителий образован тремя слоями клеток: базальным (2), промежуточным (3) и поверхностным (4). Базальный (2) слой образован неправильной кубической формой клетками, основанием прилежащими к базальной мембране. Базальная мембрана (1) образуется как самими эпителиальными клетками, так и за счет клеток подлежащей рыхлой волокнистой соединительной ткани. Состоит из светлой пластинки включающей аморфное вещество с большим содержанием ионов кальция, мало белков. Темная пластинка, наоборот, богата белками. В ее аморфном матриксе- фибриллярные структуры, в частности, коллаген IV типа; сложные белки –гликопротеины, которые индуцируют пролиферацию и дифференцировку эпителиоцитов, а с помощью фибронектина и ламинина, выполняющих роль адгезивного субстрата, эпителиоциты прикрепляются к базальной мембране; углеводы – гликозаминогликаны (придают базальной мембране упругость и создают отрицательный заряд, от которого зависит проницаемость веществ через базальную мембрану). В базальном слое переходного эпителия находятся стволовые клетки. Они способны делиться в течение всей жизни. Вновь образованные клетки дифференцируются в эпителиоциты. Промежуточный слой (3) состоит из полигональной формы клеток. Поверхностный слой (4) образован крупными округлыми одноядерными полиплоидными или двуядерными клетками (фасеточными клетками), которые в наибольшей степени изменяют свою форму при растяжении эпителия – от округлой до плоской. Этому способствует формирование в апикальной части цитоплазмы этих клеток в состоянии покоя инвагинаций плазмолеммы и дисковидных пузырьков – резервов плазмолеммы, которые встраиваются в нее по мере растяжения клетки. Формированию инвагинаций плазмолеммы способствуют многочисленные микрофиламенты (7), которые прикрепляются к особым участкам – пластинкам плазмолеммы (5). Пластинки плазмолеммы – утолщенные, малопроницаемые для воды участки полигональной формы. Они содержат скопления внутримембранных белковых частиц, каждая из которых образована шестью субъединицами. Пластинки разделены более гибкими участками плазмолеммы, не содержащими белковых частиц, которые способствуют образованию складок. Наличие указанных пластинок и плотных соединений между латеральными поверхностями клеток обеспечивает непроницаемость переходного эпителия для воды. Это свойство имеет важное функциональное значение: гипертоническая моча не разводится жидкостью из кровеносных сосудов подлежащей соединительной ткани. Переходный эпителий имеет высокую способность к регенерации. В пожилом и старческом возрасте эта способность снижена. 15 Дайте название процессу на схеме. Фрагмент какой железы представлен на схеме? Назовите типы клеток и структуры, обозначенные цифрами. Рис. 3. Сальная железа кожи. Голокриновый тип секреции. 1.Базальные клетки. 2.Базальная клетка в состоянии митоза. 3.Клетки, накапливающие секрет. 4.Разрушающиеся клетки. 5. Секрет. 6.Рыхлая соединительная ткань. Сальная железа является простой альвеолярной разветвленной железой. Между альвеолами располагается рыхлая волокнистая соединительная ткань (6). Клетки концевых секреторных отделов – себоциты. Базальные клетки (1) располагаются на базальной мембране. Они имеют кубическую или уплощенную форму. В них много гликогена, митохондрий, свободных рибосом. Слабо развиты комплекс Гольджи, гранулярная и агранулярная ЭПС. Базальные себоциты, которые выстилают верхнюю часть альвеолы не принимают участия в продукции кожного сала, являясь камбием. Дифференцируясь клетки увеличиваются в обьеме. В них хорошо развиты гранулярная и агранулярная ЭПС, комплекс Гольджи, много липидных капель. При дальнейшей дифференцировке обьем клеток значительно увеличивается (3). Они заполнены липидами. Между липидными гранулами – лизосомы, митохондрии. В клетках, накопивших секрет (3) ядра клеток становятся пикнотическими или совсем не видны, поскольку в клетке большое количество жира (агрегаты из жировых капель). В клетках увеличивается количество гидролитических ферментов в лизосомах. Начинается разрушение клеток (4), а секрет-кожное сало (состоит из триглицеридов) попадает в выводной проток. Такой тип секреции назывется голокриновым. Этот термин означает, что в процессе секреции происходит разрушение и гибель всей клетки. Голокриновые железы встречаются редко. Единственным широко распространенным примером желез такого типа являются сальные железы кожи. Тестостерон увеличивает интенсивность митоза в себоцитах. Появляется много зрелых клеток и отток кожного сала затруднен (угри). Тиреоидные гормоны усиливают функцию себоцитов. Эстрогены угнетают функцию сальных желез, а прогестерон ее стимулирует (у женщин). 16 Дайте название процессу на схеме. Фрагмент какой железы представлен на схеме? Назовите типы клеток и структуры, обозначенные цифрами. Рис. 4. Молочная железа. Апокриновый тип секреции. 1.Секреторные клетки концевого отдела (лактоциты). 2.Секрет в просвете концевого отдела. 3.Миоэпителиальная клетка. 4.Рыхлая волокнистая соединительная ткань. 5.Гемокапилляр. Молочная железа является альвеолярно-трубчатой и состоит из долек количество которых колеблется от 15 до 20. Между дольками плотная соединительная ткань. В лактирующей молочной железе дольки состоят из альвеол, их клетки - лактоциты (1) располагаются на базальной мембране. В них хорошо развита гранулярная ЭПС, комплекс Гольджи. Компоненты, вырабатывемые лактоцитами выделяются в просвет концевого отдела (2), образуют молоко. Так, в гранулярной ЭПС синтезируется белок молока казеин. Упаковывается в комплексе Гольджи в виде секреторных гранул, которые чаще всего выделяются в межклеточные канальцы по мерокриновому типу секреции. При мерокриновом типе секреции наиболее часто происходит экзоцитоз содержимого секреторных гранул путем слияния мембраны их гранул с плазмолеммой и выделения синтезированного продукта за пределы клетки. Встроенная в плазмолемму мембрана секреторных гранул затем отделяется из нее в цитоплазму механизмом эндоцитоза и возвращается в комплекс Гольджи для повторного использования. Таким образом, выведение секрета происходит без нарушения структуры клетки. Образующиеся липиды в виде крупных капель в апикальной части клетки выделяются вместе с окружающей цитоплазмой в просвет железы. Т.о., при апокриновом типе секреции в секрет отделяется часть апикальной цитоплазмы. Поверх базальной мембраны находятся отросчатые миоэпителиальные клетки (3), охватывая лактоциты они способствуют выведению молока в выводные протоки. 17 Фрагмент какой ткани на рисунке? Назовите структуры, обозначенные цифрами. Их строение, функция? Рис.5. Эпителиальные ткани. Многослойный плоский, ороговевающий эпителий. Эпидермис (кожа). I.Базальный слой. II. Шиповатый слой. III.Зернистый слой. IV. Роговые чешуйки. 1.Базальная мембрана.2.Кератиноцит. 3.Меланоцит. 4.Клетка Меркеля. 5.Чувствительное нервное окончание. 6.Миелиновое нервное волокно. В эпидермисе постоянно происходит обновление и дифференцировка клеток. Базальный слой эпидермиса (I). Кератиноциты (2) в базальном слое имеют призматическую форму, богатое хроматином ядро, базофильную цитоплазму. Размножаются путем митотического деления. От слоя к слою в них увеличивается содержание кератиновых тонофиламентов. Продвигаясь к верхним слоям, эти клетки превращаются в роговые чешуйки. Меланоциты (3) – имеют нейральное происхождение. Содержат мембранные органеллы – меланосомы с пигментом меланином. Пигмент синтезируется из аминокислоты тирозина и ДОФА - оксидазы. Меланосомы находящиеся в отростках могут покидать клетки, поступая в межклеточное пространство, а оттуда в другие клетки, в частности, в кератиноциты. Осязательные клетки Меркеля (4) находятся в сенсорных областях кожи. Имеют округлую форму, вытянутое ядро, отростки. К клеткам подходят дендриты чувствительных нейронов (5,6). В цитоплазме содержаться гранулы с гормоноподобными веществами: энкефалином, бомбезином, ВИП. Последние выделяются при тактильном раздражении клеток и путем диффузии попадают в межклеточное пространство. Оказывают влияние на регенерацию эпидермиса, проницаемость кровеносных капилляров. С соседними кератиноцитами соединяются с помощью десмосом. Относят к механорецепторам. Внутриэпидермальные макрофаги (на рисунке не изображены) - образуются из моноцитов крови, имеют отростки доходящие до зернистого слоя. Эти клетки захватывают антигены, которые попадают в эпидермис. Осуществляют их процессинг и транспорт в лимфатические узлы, где представляют лимфоцитам вызывая иммунную реакцию. Шиповатый слой (II) – находятся кератиноциты, клетки Лангерганса (внутриэпидермальные макрофаги). Кератиноциты полигональной формы. Клетки имеют короткие отростки – шипики, между которыми десмосомы. В клетках увеличивается количество кератиновых тонофибрилл, которые концентрически располагаются вокруг ядра. Появляются кератиносомы (плотные гранулы окруженные мембраной). В них синтезируются липиды - холестеринсульфат, церамиды и др., которые связывают клетки друг с другом. Зернистый слой (III). Клетки этого слоя имеют уплощенную форму, располагаясь в 3-4 слоя. Синтезируют белок филагрин формирующий гранулы к поверхности которых прилежат кератиновые тонофибриллы, распадающиеся органеллы клетки. В результате образуются кератогиалиновые гранулы, которые обуславливают зернистость. Продолжается синтез липидов, которые выделяются в межклеточное вещество, обеспечивая водонепроницаемость и осуществляя барьерную функцию. Под плазмолеммой накапливается синтезирующийся белок кератолинин, который «утолщает» оболочку. Роговой слой (IV) - состоит из закончивших дифференцировку кератиноцитов, (роговые чешуйки). Внутри нерастворимые в воде тонофибриллы (мягкий кератин), отсутствуют органеллы. Таким образом, от базального к шиповатому слоям кератиновые тонофибриллы полностью заполняют всю клетку, исчезают ядро и органеллы. Под плазмолеммой белок кератолинин, который синтезируют клетки и за счет которого формируется толстая оболочка. Специальные органеллы кератиносомы синтезируют липиды, фиксирующие клетки и обеспечивают водонепроницаемость. 18 Клетки какой ткани изображены на рисунке? Назовите структуры обозначенные цифрами. Рис. 6. Однослойный цилиндрический эпителий с бокаловидными клетками (тонкая кишка). 1.Цилиндрический эпителиоцит. 2.Ядро. 3.Комплекс Гольджи. 4.Гранулярная ЭПС. 5.Микроворсинки. 6.Бокаловидный экзокриноцит. 7.Ядро. 8.Гранулярная ЭПС. 9.Комплекс Гольджи. 10.Секреторные включения (вакуоли). 11.Базальная мембрана. Однослойный цилиндрический (призматический) эпителий, имеющий энтодермальное происхождение находится в желудке, тонкой и толстой кишке, желчном пузыре, выстилает выводные протоки желез, присутствует в почках (извитые канальцы) и т.д. Клетки располагаются на базальной мембране (10). Цилиндрические эпителиоциты (1) выстилают поверхность ворсинок и крипт в тонкой кишке. На апикальной поверхности клеток хорошо развиты микроворсинки (5), за счет которых увеличивается поверхность всасывания, а также здесь находится множество пищеварительных ферментов принимающих участие в пристеночном пищеварении. Клетки имеют хорошо развитую гранулярную (4) и агранулярную ЭПС, комплекс Гольджи (3), много митохондрий. Это короткоживущие клетки (3 суток). Регенерация осуществляется за счет стволовых клеток. Бокаловидные экзокриноциты (6) являются одноклеточными слизистыми железами. Располагаются поодиночке среди цилиндрических эпителиоцитов. Функционируют циклически: накапливают и выделяют секрет. Последний находится в секреторных вакуолях (10). При накоплении секрета ядро смещается к базальной поверхности клетки. В клетках хорошо развиты агранулярная и гранулярная ЭПС (8), комплекс Гольджи (9). Секрет содержит углеводно-протеидные комплексы. Количество бокаловидных клеток увеличивается по направлению к прямой кишке. 19 Назовите тип клетки. Аргументируйте вывод. Назовите структуры, обозначенные цифрами. Рис.7. Фибробласт рыхлой волокнистой соединительной ткани. 1.Отростки фибробласта. 2.Гранулярная ЭПС. 3.Комплекс Гольджи. 4.Ядро. 5.Митохондрии. 6.Коллагеновое волокно. 7.Эластическое волокно. Фибробласты – основной тип клеток рыхлой волокнистой соединительной ткани. Источником развития фибробластов в эмбриогенезе является мезенхима. Стволовая клетка →полустволовая клетка-предшественница →малоспециализированный фибробласт →фибробласт →фиброцит. Функция фибробластов заключается в продукции всех компонентов межклеточного вещества (коллагеновых (6), эластических (7), ретикулярных волокон и аморфной субстанции). Фибробласты осуществляют не только синтез, но также перестройку и частичное разрушение межклеточного вещества. Морфология этих клеток тесно связана с их синтетической активностью. Зрелый фибробласт – крупная отростчатая (1) клетка со светлым ядром (4), содержащим 1-2 ядрышка. Цитоплазма содержит органеллы хорошо развитого синтетического аппарата – гранулярную эндоплазматическую сеть (2), цистерны которой часто растянуты, комплекс Гольджи (3). В цитоплазме располагаются также лизосомы и митохондрии (5). Хорошо выражены все элементы цитоскелета, благодаря которым фибробласт обладает подвижностью, способностью изменять свою форму и обратимо прикрепляться к другим клеткам и волокнам. При старении фибробласты превращаются в малоактивную форму – фиброциты. 20 К какой ткани относится клетка на схеме? Назовите тип клетки и структуры, обозначенные цифрами. Рис. 8. Макрофаг (гистиоцит) рыхлой волокнистой соединительной ткани. 1.Отростки макрофага. 2.Фагоцитоз. 3.Пиноцитоз. 4.Фаголизосома. 5.Лизосома. 6.Гранулярная ЭПС. 7.Комплекс Гольджи. 8.Ядро. 9.Митохондрия. 10.Межклеточное вещество рыхлой волокнистой соединительной ткани. Макрофаги образуются из моноцитов крови после их миграции в соединительную ткань из кровеносных сосудов. Различают свободные макрофаги – это макрофаги рыхлой волокнистой соединительной ткани (РВСТ), серозных полостей, альвеолярные макрофаги, макрофаги воспалительных экссудатов. Фиксированные – в селезенке, лимфоузлах, внутриэпидермальные макрофаги, макрофаги ворсин плаценты, микроглия ЦНС. Ядра макрофагов (8) небольшого размера, овальной или бобовидной формы. В соединительной ткани макрофаги могут находиться как в покоящемся, так и в активном состоянии (блуждающие макрофаги). Покоящиеся макрофаги имеют уплощенную форму, плотное ядро и небольшое количество органелл. Неактивные макрофаги обычно прикреплены к коллагеновым волокнам. Блуждающие макрофаги, напротив, высоко подвижны, поверхность их неровная, с многочисленными выростами – псевдоподиями, микроворсинками. При электронной микроскопии в активных макрофагах выявляются множество лизосом (5), фагоцитированные частицы, фаголизосомы (4), митохондрии (9), гранулярная (6) и агранулярная ЭПС, включения гликогена, элементы цитоскелета. На поверхности цитолеммы макрофаги несут рецепторы для медиаторов иммунной системы, нейромедиаторов, гормонов, молекулы адгезии, позволяющие им мигрировать, взаимодействовать с другими клетками и межклеточным веществом. Функции макрофагов многообразны: 1) Распознавание, поглощение и расщепление с помощью ферментов микроорганизмов и других антигенов, погибших клеток, компонентов межклеточного вещества. 2) Антигенпредставляющая: переработка антигенов и передача информации об антигенах Т-лимфоцитам, благодаря этой функции макрофаги участвуют в запуске иммунных реакций. 3) Секреция веществ, регулирующих функции других клеток РВСТ, иммунокомпетентных клеток, стимулирующих регенерацию, противовирусных (интерферон) и антибактериальных (лизоцим) факторов. 21 Назовите тип клетки и вариант микроскопии. Аргументируйте свои выводы. Рис. 12. Тучная клетка (лаброцит, тканевой базофил) рыхлой волокнистой соединительной ткани в момент дегрануляции. Сканирующая электронная микроскопия. 1.Гранулы тучной клетки. Тучные клетки – постоянный компонент рыхлой волокнистой соединительной ткани. Обнаруживаются всюду, где есть рыхлая волокнистая соединительная ткань. Относятся к потомкам стволовой клетки крови. Тучные клетки располагаются преимущественно около мелких сосудов. Они многочисленны в дерме, собственной пластинке слизистых оболочек, строме молочной железы и тимуса. Их количество возрастает в активно функционирующих органах: в лактирующей молочной железе, в матке при беременности, в щитовидной железе при гиперфункции, в очагах воспаления, в опухолях и по периферии заживающих ран. Тучные клетки имеют округлую форму (диаметр 20-30 мкм), поверхность с многочисленными выростами. Однако, они могут иметь неправильную форму, овальную, иногда с отростками, что связано с их способностью к амебоидным движениям. Ядро небольших размеров, округлое с умеренным содержанием гетерохроматина. Цитоплазма содержит умеренно развитые органеллы и многочисленные гранулы, величина и состав которых вариабельны. Гранулы тучных клеток сходны по составу с гранулами базофилов, имеют диаметр до 1 мкм. Они содержат гепарин, гистамин, дофамин, хондроитинсульфаты, гиалуроновую кислоту, хемотаксические факторы эозинофилов и нейтрофилов, ферменты (протеазы, гидролазы, катепсин G). Малые дозы этих биологически активных веществ секретируются клеткой постоянно, регулируя тем самым тонус и проницаемость сосудов. Активация и быстрая массивная дегрануляция тучных клеток наступает после связывания их рецепторов с IgЕ (аллергическая реакция), белками комплемента, цитокинами, нейропептидами (вещество Р, соматостатин), протеиназами. Кроме выброса содержимого гранул тучные клетки выделяют эйкозаноиды (простагландины, лейкотриены), фактор активации тромбоцитов, цитокины (ИЛ-1, -2, -3, -4, -5, -6 и др.). Результатом дегрануляции тучных клеток являются разнообразные реакции, связанные со спазмом гладких мышц, расширением сосудов, повышением их проницаемости, хемотаксисом нейтрофилов, эозинофилов, макрофагов, фибробластов. Выделение различных ферментов вызывает разрушение компонентов межклеточного вещества, нередко повреждение тканей. Вместе с тем, некоторые вещества, вырабатываемые тучными клетками, стимулируют репаративные процессы. Количество тучных клеток вариабельно и изменяется в зависимости от физиологического состояния. Клинические проявления дегрануляции тучных клеток включают бронхоспазм, острый ринит, отеки, кожный зуд, диарею, падение кровяного давления. Вещества, блокирующие дегрануляцию тучных клеток, нашли широкое применение для лечения и профилактики аллергических заболеваний. В цитоплазме тучных клеток содержаться липазы, АТФ-аза, щелочная фосфатаза, цитохромоксидаза, гистидиндекарбоксилаза (является маркером) и другие ферменты. Метод сканирующей электронной микроскопии позволяет изучить поверхность клетки в трехмерном пространстве. При данном виде электронной микроскопии пучок электронов пробегает по поверхности обьекта. Полученная информация передается на электронно-лучевую трубку. Изображение получают в отраженных или вторичных электронах. Фиксированный и высушенный обьект при данном методе покрывают тонким слоем испаренного металла (золото). Отражаясь от него электроны попадают в приемное устройство, которое передает сигнал на электронно-лучевую трубку. За счет огромной глубины фокуса сканирующего электронного микроскопа (она больше, чем у просвечивающего) получается почти трехмерное изображение. 22 Назовите тип клетки. Аргументируйте вывод. Назовите структуры, обозначенные цифрами? Рис. 13. Плазматическая клетка (плазмоцит). 1.Цитолемма. 2.Ядро. 3.Хроматин. 4.Комплекс Гольджи. 5.Митохондрии. 6.Гранулярная эндоплазматическая сеть. 7.Секреторные вакуоли. Плазматические клетки в процессе дифференцировки развиваются из Влимфоцитов. Эти клетки обеспечивают выработку антител – гаммаглобулинов при появлении в организме антигена, т.е. участвуют в гуморальном иммунитете. Плазматические клетки образуются в лимфоидных органах. Из стимулированных В-лимфоцитов образуются В-лимфобласты, которые размножаются и часть из них приобретает способность к синтезу антител и становится плазмобластами, которые затем превращаются в плазмоциты. Плазмобласты-крупные клетки с большим количеством рибосом и небольшим числом уплощенных цистерн гранулярной эндоплазматической сети. Ядро содержит деконденсированный хроматин. Плазмоцит имеет меньшие размеры. Величина плазмоцитов от 7 до 10 мкм. Форма округлая или овальная. Ядро (2) лежит эксцентрично, хроматин (3) более компактный, расположен группами около ядерной оболочки (имеет вид колеса со спицами). Около ядра видна зона более светлой цитоплазмы, в которой расположен комплекс Гольджи (4). Большое количество концентрически расположенных узких канальцев гранулярной эндоплазматической сети (6). Цитоплазма резко базофильна (практически вся заполнена массой крупных цистерн гранулярной эндоплазматической сети, в которой синтезируются белки). Базофилия отсутствует только в небольшой светлой зоне цитоплазмы около ядра, где расположен аппарат Гольджи. Плазмоциты встречаются в рыхлой соединительной ткани собственного слоя слизистых оболочек полых органов, сальнике, лимфатических узлах, селезенке, красном костном мозге. Количество плазмоцитов увеличивается при различных инфекционно-аллергических реакциях, воспалительных заболеваниях. 23 Назовите клетку, аргументируя вывод. Укажите тип электронной микроскопии. Рис. 15. Плазматическая клетка (плазмоцит). Клетка округлой формы с колесовидным ядром, цитоплазма заполнена многочисленными цистернами гранулярной ЭПС. Трансмиссионная электронная микроскопия. Ув. 17000. К какой ткани относится клетка на схеме? Назовите тип клетки и структуры, обозначенные цифрами. Рис. 16. Хондроцит. 1.Ядро. 2.Гранулярная ЭПС. 3.Комплекс Гольджи. 4.Митохондрии. 5.Липидные капли. 6.Гранулы гликогена. 7.Лакуна. 8.Маткрикс хряща. Хондроциты – это клетки хрящевой ткани. Вырабатывают межклеточное вещество. Они имеют овальную или сферическую форму и лежат в полостях (лакунах) (7). В глубоких отделах хряща хондроциты могут располагаться группами в пределах одной лакуны, формируя путем деления изогенные группы (до 8-12 клеток). Под электронным микроскопом на их поверхности выявляются микроворсинки. Ядро (1) круглое или овальное, светлое (преобладает эухроматин), с одним или несколькими ядрышками. Цитоплазма содержит многочисленные цистерны гранулярной ЭПС (2), комплекс Гольджи (3), гранулы гликогена (6) и липидные капли (5), митохондрии (4). В зависимости от степени дифференцировки и функциональной активности выделяют три типа хондроцитов. Хондроциты I типа преобладают в молодом развивающемся хряще, характеризуются высоким ядерноцитоплазматическим отношением, развитым комплексом Гольджи, наличием митохондрий и рибосом в цитоплазме. Эти клетки делятся, формируя изогенные группы. Хондроциты II типа отличаются низким уровнем ядерно-цитоплазматических отношений, интенсивным развитием гранулярной ЭПС, комплекса Гольджи, которые обеспечивают образование и секрецию межклеточного вещества. Хондроциты III типа имеют самый низкий индекс ядерно-цитоплазматических отношений, сильно развитую гранулярную ЭПС, сохраняют способность к синтезу компонентов межклеточного вещества, но снижают продукцию гликозаминогликанов. 24 Назовите тип клетки? Аргументируйте вывод? Назовите структуры, обозначенные цифрами? Рис. 17. Остеобласт, дифференцирующийся в остеоцит. 1.Минерализованное вещество кости. 2.Зона минерализации. 3.Остеобласт. 4.Отростки остеобласта. 5.Капилляр. Остеобласты – это молодые клетки, создающие костную ткань. Образуются из малодифференцированных клеток мезенхимы (остеогенные клетки костного мозга). В сформировавшейся кости они встречаются только в глубоких слоях надкостницы, эндосте, вокруг внутрикостных сосудов, в местах регенерации костной ткани после ее травмы. Эти клетки мигрируют в участки будущей кости или зоны её перестройки, размножаются, трансформируются вначале в клетки-предшественники, а затем в остеобласты. Клетки имеют чаще кубическую форму, неровную поверхность, короткие отростки (4), цитоплазма базофильна. Ядро округлой или овальной формы, может располагаться эксцентрично. Ультраструктура остеобласта типична для секреторной клетки. Хорошо развита гранулярная ЭПС, расположенная у основания, комплекс Гольджи локализуется над ядром. Клетки содержат митохондрии, липидные включения. При помощи коротких отростков, содержащих актиновые микрофиламенты, остеобласты устанавливают контакты с соседними остеобластами. Основная функция остеобластов заключается в синтезе и секреции органического матрикса кости вокруг своих отростков. В клетках высока активность щелочной фосфатазы, необходимая для минерализации матрикса. Остеобласты выделяют так называемые матричные пузырьки, содержащие липиды, кальций, щелочную фосфатазу. Остеобласты обладают способностью к продукции белка, образуют преимущественно коллаген 1 типа, а также гликопротеины матрикса (остеонектин, остеокальцин) и протеогликаны. Принимают участие в процессе минерализации костной ткани (1). Остеобласты, выделяя фермент щелочную фосфатазу способствуют дефосфорилированию межклеточного субстрата, повышению концентрации фосфатных ионов. Этот процесс связан с выведением из цитоплазмы остеобластов мелких матричных пузырьков с высоким содержанием фосфата кальция и щелочной фосфатазы. Остеобласты, как правило, окружает остеоид – неминерализованный костный матрикс. До превращения в остеоциты остеобласты окружают себя со всех сторон матриксом. 25 К какой ткани относится клетка на схеме? Назовите тип клетки и структуры, обозначенные цифрами. Рис. 18. Остеокласт, разрушающий костную ткань. 1.Гофрированный край. 2.Зона резорбции. 3.Матрикс костной ткани. 4.Лизосомы. 5.Светлые зоны. 6.Комплекс Гольджи. 7.Ядра. 8.Гранулярная ЭПС. 9.Митохондрии. Остеокласты – многоядерные гигантские клетки (симпласты), образующиеся вследствие слияния моноцитов. Родоначальником последних является стволовая клетка крови из которой образуются моноциты (6 класс). Они через стенку капилляра попадают в костную ткань, где и преобразуются в остеокласты. Остеокласты в костной ткани находятся в местах ее регенерации, в периваскулярных пространствах остеонов.Они располагаются на поверхности костных перекладин. Остеокласты подвижны и осуществляют разрушение (резорбцию) костной ткани, обызвествленного хряща. Так как резорбция кости сопровождается высвобождением кальция, эти клетки играют важнейшую роль в поддержании кальциевого гомеостаза. Остеокласты располагаются в образованных ими углублениях на поверхности костной ткани (резорбционных лакунах), достигают размеров 20-100 мкм, содержат до 20-50 ядер (7). Цитоплазма ацидофильная, с высоким содержанием лизосом (4), митохондрий (9), диктиосом комплекса Гольджи (6). В активном остеокласте край, прилежащий к кости, образует многочисленные складки плазмолеммы (гофрированный край) (1). Здесь осуществляется синтез и секреция гидролитических ферментов. Над гофрированным краем многочисленные пузырьки и вакуоли. По сторонам гофрированного края имеются светлые зоны – участки плотного прикрепления клетки к кости. Остеокласт плотно прилегает к костному веществу и в области гофрированной зоны создается герметическое пространство. В зоне плотного прилегания светлая цитоплазма, мало органелл. Ядра и органоиды сосредоточены в удаленной от кости части остеокласта (базальной зоне). Разрушение костной ткани остеокластом включает несколько этапов: 1) Прикрепление остеокласта к поверхности кости обеспечивается взаимодействием рецепторов плазмолеммы остеокласта с белками костного матрикса (остеопонтином, витронектином) и перестройкой цитоскелета в области светлых зон (5), которые герметизируют участок резорбции (2) (лакуну). 2) Закисление содержимого лакун осуществляется благодаря действию протонных насосов, накачивающих ионы Н + в лакуну, и экзоцитозу пузырьков с кислым содержимым. 3) Растворение минеральных компонентов матрикса кислым содержимым лакун. 4) Разрушение органических компонентов матрикса протеолитическими ферментами лизосом, секретируемых в лакуну. 5) Удаление продуктов разрушения костной ткани осуществляется путем везикулярного транспорта через цитоплазму остеокласта или разгерметизации лакуны. Гормон щитовидной железы кальцитонин и женские половые гормоны угнетают деятельность остеокластов, гормон околощитовидных желез паратгормон активирует их. Костная ткань на протяжении всей жизни под влиянием внешних и внутренних факторов подвергается перестройке. В ней участвуют как клетки созидатели -остеобласты, так и клетки разрушители - остеокласты. Остеокласты, разрушая костную ткань, образуют в ней полости. Остеобласты вокруг сосудов в этих полостях образуют костное вещество имеющее вид пластинок цилиндрической формы (остеоны). При усилении нагрузки на костную ткань активность остеобластов повышается. При этом будет увеличиваться количество остеонов. При уменьшении нагрузки на костную ткань - повышается активность остеокластов, которые разрушая костную ткань, уменьшают ее прочность. По литературным данным активность остеокластов особенно высока в условиях невесомости. 26 Назовите тип клетки? Аргументируйте вывод. Назовите структуры, обозначенные цифрами? Рис. 19. Остеоцит. 1.Ядро. 2.Гранулярная эндоплазматическая сеть. 3.Комплекс Гольджи. 4.Митохондрии. 5.Стенка лакуны. 6.Костный каналец. 7.Отросток остеоцита. 8.Межклеточное вещество костной ткани. Остеоциты образуются из остеобластов. Это зрелые и уже не делящиеся клетки, расположенные в костных полостях. Они имеют отростчатую форму, компактное относительно крупное ядро, слабо базофильную цитоплазму. В клетке присутствуют цистерны гранулярной эндоплазматической сети (2), свободные рибосомы, комплекс Гольджи (3), округлые митохондрии (4) и лизосомы. Тонкие отростки (7) остеоцитов расположены в канальцах (6), отходящих в разные стороны от костных полостей. Отростки соседних остеоцитов, соприкасающиеся боковыми поверхностями, формируют щелевые контакты. Канальцы костных полостей заполнены тканевой жидкостью, анастомозируют между собой и с периваскулярными пространствами сосудов, заходящих внутрь кости. Остеоциты поддерживают структурную целостность минерализованного матрикса, участвуют в регуляции обмена кальция в организме. Это функция остеоцитов находится под контролем уровня кальция плазмы крови и различных гормонов. 27 Фрагмент какой ткани на фотограмме? Аргументируйте вывод. Назовите структуры, обозначенные цифрами. Рис. 20. Белая жировая ткань. Адипоцит белой жировой ткани. 1.Жировая капля. 2.Уплощенное ядро. 3.Узкий ободок цитоплазмы. 4.Кровеносный капилляр. 5.Адвентициальная клетка. 6.Ретикулярные волокна. Белая жировая ткань является преобладающим видом жировой ткани у человека. В эмбриогенезе развивается из мезенхимы, после рождения источником развития жировых клеток являются малодифференцированные фибробласты. Белая жировая ткань находится в подкожной жировой клетчатке, сальнике, межмышечно, в стенках внутренних органов. Белая жировая ткань состоит из скоплений жировых клеток - адипоцитов, разделенных тонкими прослойками рыхлой волокнистой соединительной ткани, несущими кровеносные сосуды и нервы. Кровеносные капилляры (4) и нервные волокна проникают и между адипоцитами. Адипоциты (липоциты) – крупные (диаметром 25-250 мкм) клетки сферической формы. Цитоплазма адипоцита содержит одну крупную жировую каплю (1), занимающую до 90-95% объема клетки, липиды в жировых клетках постоянно обновляются. Остальная часть цитоплазмы образует тонкий ободок (3), окружающий жировую каплю. Цитоплазма содержит агранулярную ЭПС, пиноцитозные пузырьки, комплекс Гольджи, митохондрии, промежуточные филаменты, а также уплощенное ядро (2), содержащее умеренно конденсированный хроматин. Каждый адипоцит снаружи окружен базальной мембраной, в которую вплетаются ретикулярные волокна (6). Функции белой жировой ткани: трофическая (депо жиров и жирорастворимых витаминов), энергетическая (при расщеплении жира образуется большое количество энергии), термоизолирующая, защитно-механическая, эндокринная (вырабатываются половые гормоны - эстрогены, гормон, регулирующий потребление пищи – лептин). 28 Фрагмент какой ткани на рисунке? Аргументируйте вывод. Назовите структуры, обозначенные цифрами. Рис. 21. Бурая жировая ткань. Адипоцит бурой жировой ткани. 1.Множественные жировые капли в цитоплазме адипоцита. 2.Ядро. 3.Митохондрии. 4.Комплекс Гольджи.5.Кровеносные капилляры. 6.Ретикулярные волокна. Бурая жировая ткань содержится у человека в небольшом количестве и сосредоточена в определенных участках тела (между лопаток, за грудиной, вдоль позвоночника, на задней поверхности шеи, под кожей, между мышцами). Она хорошо выражена у плодов человека и новорожденных, составляя 2-5% массы тела. Бурая жировая ткань образована дольками, состоящими из адипоцитов бурой жировой ткани, разделенных тонкими прослойками рыхлой волокнистой соединительной ткани. Между адипоцитами располагаются многочисленные кровеносные капилляры (5) и симпатические нервные волокна. Бурый цвет ткани связан как с ее обильным кровоснабжением, так и с наличием железосодержащих пигментов в митохондриях адипоцитов. Адипоциты бурой жировой ткани в диаметре до 60 мкм. Округлое ядро (2) располагается в центре клетки. Цитоплазма содержит множественные жировые капли (1) различных размеров и многочисленные митохондрии (3), слабо развита ЭПС и комплекс Гольджи (4). Ведущая функция бурой жировой ткани – термогенез – обеспечивается особенностями митохондрий. В митохондриях адипоцитов бурой жировой ткани выявлен особый белок – термогенин. Он разобщает процессы окисления и фосфорилирования, поэтому энергия окисления не накапливается в форме макроэргических связей АТФ, а рассеивается в виде тепловой энергии. Обильное кровоснабжение обеспечивает быстрое отведение вырабатываемого тепла. При понижении температуры окружающей среды активность окислительных процессов увеличивается. 29 Что представлено на электронной микрофотографии? Назовите тип электронной микроскопии. Рис. 23. Ретикулярные и коллагеновые волокна. На снимке соединительная ткань щитовидной железы. Фолликулы железы окружены плотным сплетением тонких ретикулярных волокон. Коллагеновые волокна выглядят как толстые мощные пучки. Сканирующая электронная микроскопия. Ув. 2500. Какие волокна изображены на рисунке? Как происходит их образование? Рис. 24. Коллагеновые волокна рыхлой волокнистой соединительной ткани. 1.Протофибриллы. 2.Микрофибриллы. 3.Фибриллы. 4.Коллагеновое волокно. Коллагеновые волокна являются волокнистым компонентом межклеточного вещества. Они формируют рыхло распределенную сеть и обеспечивают механическую прочность (на коллагеновой нити диаметром 1 мм можно подвесить груз, масса которого 70 кг). Состоят из белка коллагена. В процессе образования коллагеновых волокон выделяют два этапа: внутриклеточный и внеклеточный. В рыхлой волокнистой соединительной ткани коллагеновые волокна образуют фибробласты. На их гранулярной ЭПС синтезируются молекулы тропоколлагена представляющие три полипептидных альфа-цепи из аминокислот, которые свиваются в клетке, образуя проколлаген. Покидая клетку и соединяясь первоначально образуются протофибриллы (1). Пять - шесть протофибрилл соединяясь боковыми поверхностями образуют микрофибриллы (2), а они, также соединяясь боковыми поверхностями формируют коллагеновые волокна (4) диаметром от 1 до 10 мкм. Количество фибрилл может равняться нескольким десяткам в зависимости от топографии коллагеновых волокон. В параллельных рядах молекулы коллагена смещены относительно друг друга, поэтому под электронным микроскопом по ходу волокна чередуются темные и светлые полосы. В зависимости от молекулярной организации выделяют 14 типов коллагена. Наиболее часто встречаются первые четыре типа. В частности, коллаген I типа присутствует в костной ткани, в РВСТ; коллаген II типа в гиалиновом и волокнистом хряще; коллаген III типа в ретикулярных волокнах; коллаген IV типа в составе базальных мембран и т.д. 30 К какой ткани относятся клетки, изображенные на рисунке? В каких органах встречается эта ткань, какие функции она выполняет? Рис. 25. Ретикулярная ткань (ретикулярные клетки и ретикулярные волокна). Строма кроветворных органов. 1.Ретикулярные клетки. 2.Ретикулярные волокна. 3.Лимфоцит. 4.Макрофаг. Ретикулярная ткань – разновидность соединительной ткани со специальными свойствами. Образует строму органов кроветворения (за исключением тимуса). Состоит из отростчатых ретикулярных клеток (1) и связанных с ними ретикулярных волокон (2), образующих трехмерную сеть и вплетающихся в отростки фибробластоподобных ретикулоцитов, а также основного межклеточного вещества. Ретикулярная ткань создает микроокружение для развивающихся клеток крови. Ретикулярные клетки (1) вырабатывают элементы межклеточного вещества (преколлаген, гликозаминогликаны, проэластин) и ростовые факторы, регулирующие развитие гемопоэтических клеток (3,4). Ретикулярные волокна – продукт синтеза ретикулярных клеток и состоят из коллагена III типа. Они обнаруживаются при импрегнации солями серебра, поэтому называются еще аргирофильными. По растяжимости занимают промежуточное положение между коллагеновыми и эластическими волокнами, выполняют опорную функцию. 31 Что представлено на рисунке? Назовите структуры, обозначенные цифрами. Рис. 26.Схема строения тонких и толстых миофиламентов. 1.Толстый миофиламент. 2.Молекулы миозина. 3.Головки миозина. 4.Тонкий миофиламент. 5.Молекулы актина. 6.Тропомиозин. 7.Тропониновый комплекс. Сократительный аппарат мышечного волокна представлен миофибриллами – специальными органеллами, имеющими вид нитей диаметром 1-2 мкм. Они располагаются продольно на всю длину миосимпласта. Миофибриллы состоят из тонких (4) актиновых (диаметр 5 нм) и толстых (1) миозиновых (диаметр 12 нм) филаментов. В состав актиновых филаментов входит белок актин (5), а также белки тропонин и тропомиозин. В отличие от обычных актиновых нитей тонкие миофиламенты не подвергаются постоянному распаду и полимеризации. Это обьясняется присоединением к их актиновым цепочкам стабилизирующего белка - тропомиозина. Молекулы актина (5) имеют глобулярное строение и, соединяясь вместе, образуют длинные цепи. В актиновых филаментах таких цепей две, они образуют двойную спираль. В бороздках между цепями актина лежат молекулы тропомиозина (6). К молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина. Тропониновый комплекс (7) состоит из трех глобулярных субъединиц: тропонин T, тропонин C и тропонин I. Тропонин T осуществляет прикрепление тропонинового комплекса к тропомиозину. Тропонин C связывается с ионами кальция. Тропонин I препятствует взаимодействию миозиновых головок с актином. В покое молекулы актина связаны с тропомиозином и тропонинами таким образом, что актин блокирован и не может взаимодействовать с миозином. Толстые филаменты образованы упорядочено упакованными молекулами фибриллярного белка миозина. Каждая молекула миозина состоит из двух частей: головки и хвоста и может изменять пространственную ориентацию за счет шарнирных участков. Головки миозина проявляют ферментативную активность и способны расщеплять АТФ энергия которой идет на сокращение. Хвостовыми частями молекулы миозина соединяются в пучки и формируют толстый миофиламент (1). Головки миозина (3) расположены снаружи. В покое при низкой концентрации ионов Са2+ миозиновые головки не могут взаимодействовать с участками связывания на молекуле актина, потому что они прикрыты тропонин-тропомиозиновым комплексом. При возбуждении повышается концентрации ионов Са2+ в области миофиламентов. Взаимодействие актиновых и миозиновых филаментов при сокращении запускается ионами Са2+. Субъединица тропонина C тонкого миофиламента, связываясь с Са2+, к которому имеет высокую степень сродства, подвергается пространственному изменению. Это меняет форму тропонинового комплекса; происходит перестройка пространственной конфигурации тропомиозина и изменяется форма актиновой нити. Молекулы актина могут взаимодействовать с головками миозина. Миозиновые головки соединяются с ближайшими молекулами актина (с их миозинсвязывающими участками). При этом происходит сокращение миозинов в зонах шарнирных участков и толстая нить продвигается вперед в направлении центральной части изотропного диска. Следующим этапом является разрушение АТФ легкими цепями меромиозина (они присоединены к миозиновым головкам). При этом «разрывается» связь миозина с актином. Шарнирный участок «выпрямляется», т.е. принимает исходное положение. Но миозиновые головки располагающиеся впереди вновь связываются с последующими молекулами актина – и вновь движение вперед. Миозиновые головки «шагают» по актину. Расслабление после мышечного сокращения происходит в результате снижения концентрации Са2+, тропонин С высвобождается, актиновый филамент восстанавливает свою структуру. Миозинсвязывающие участки актина «закрываются» и взаимодействие актина и миозина невозможно. Миофибриллы занимают исходное положение и происходит расслабление мышечного волокна 32 Фрагмент какой ткани на схеме? Аргументируйте выводы. Назовите структуры, обозначенные цифрами, буквами? Рис. 27. Фрагмент мышечного волокна скелетной мышечной ткани. 1.Т-трубочка. 2.Терминальная цистерна саркоплазматической сети; А - Анизотропный диск; I – Изотропный диск; Z – линия Z; Н - Полоска H. 3.Митохондрии. 4.Ядро. 5.Базальная мембрана. 6.Плазмолемма. 7.Агранулярная ЭПС. 8.Миофибриллы. Структурно – функциональная единица скелетной мышцы – мышечное волокно. Это симпласт и миосателитоциты. Мышечное волокно имеет форму цилиндра с заостренными концами. Оболочка волокна (сарколемма) состоит из плазмолеммы симпласта и базальной мембраны. Между базальной мембраной и плазмолеммой расположены овальной формы ядра клеток – сателлитов. Ядра мышечного волокна (4) лежат под плазмолеммой в саркоплазме. В саркоплазме симпласта расположены: сократительный аппарат - миофибриллы (8); саркоплазматическая сеть (депо кальция) (7); энергетические станции – митохондрии (3); включения (гранулы гликогена). От поверхности мышечного волокна к расширенным участкам саркоплазматического ретикулума направляются трубковидные выпячивания плазмолеммы – поперечные трубочки (Т-трубочки) (1). Сократительный аппарат – миофбриллы придают мышечному волокну поперечную исчерченность, обусловленную регулярным чередованием в миофибриллах светлых и темных участков (дисков) изотропных (I) и анизотропных (А): светлых и темных. Разное светопреломление дисков обусловлено тем, что по длине саркомера упорядочено, расположены тонкие и толстые нити. Каждый светлый диск пересекает Z-линия. Участок миофибриллы между двумя Z-линиями определяется как саркомер – структурно–функциональная единица миофибриллы. Саркомер образуют расположенные параллельно друг другу тонкие (актиновые) и толстые (миозиновые) нити. Изотропный диск содержит только тонкие нити. Один конец тонкой нити прикрепляется к Z-линии, а другой конец направлен к середине саркомера. Толстые нити занимают центральную часть саркомера (Адиск). Тонкие нити частично входят между толстыми. Содержащий только толстые нити участок саркомера Н-зона, в середине её проходит М-линия. Саркомер содержит один А-диск (темный) и две половины изотропного диска (светлого). Формула саркомера: Z1 + 1/2 I1 + А + 1/2 I2 + Z2. Миозин – гексамер (две тяжелые цепи и четыре легкие). Тяжелые цепи – две спирально закрученные полипептидные нити, несущие на своих концах глобулярные головки. В области головок с тяжелыми цепями ассоциированы легкие цепи. Тонкая нить состоит из актина, тропомиозина и тропонинов. Тонкая нить - это две спирально скрученные цепочки F-актина. Полярные молекулы тропомиозина укладываются конец в конец в желобке между двумя спирально закрученными цепочками F-актина. Тропонин препятствует взаимодействию актина с миозином. При поступлении нервного импульса волна деполяризации доходит до цистерн саркоплазматического ретикулума, из них выделяются ионы кальция и концентрация кальция в саркоплазме резко возрастает. Кальций диффундирует к тонким нитям саркомера, где связывается с тропонином и миозиновыми головками. Происходит взаимодействие миозиновых головок с актином (актомиозиновые «мостики»). Это приводит к тому, что миозиновые головки «шагают по актину», образуя в ходе перемещения новые связи актина и миозина, сближая две Z-линии. Уменьшаются светлые диски. При расслаблении концентрация Са 2+ в саркоплазме становится низкой – Са 2+ - тропомиозин закрывает миозинсвязывающие участки тонких нитей и препятствует их взаимодействию с миозином. 33 Клетки какой ткани изображены на рисунке? Назовите структуры обозначенные цифрами. Рис. 28. Гладкий миоцит. 1.Ядро гладкого миоцита. 2.Плазмолемма. 3.Базальная мембрана. 4.Кавеола. 5.Гранулярная ЭПС. 6.Комплекс Гольджи. 7.Миофибриллы. 8.Двигательное нервное окончание. 9.Кровеносный сосуд. Структурно-функциональной единицей гладкой мышечной ткани является гладкий миоцит (гладкая мышечная клетка), имеющий мезенхимное происхождение. Гладкая мышечная ткань находится в стенке кровеносных сосудов, образует мышечные оболочки внутренних органов. Гладкие миоциты имеют веретенообразную форму, палочковидное ядро (1), снаружи базальная мембрана (3), а под ней – плазмолемма (2). Гранулярная ЭПС (5) и комплекс Гольджи (6) развиты слабо, много митохондрий, рибосомы расположены свободно. Акто-миозиновые комплексы (миофибриллы) (7) существуют только в момент сокращения. Актиновые нити расположены косо по отношению к длинной оси клетки, прикрепляются к плотным тельцам, которые являются сшивающими белками. Мономеры миозина располагаются рядом с актиновыми нитями. Ионы кальция при поступлении потенциала действия содержатся в кавеолах (покрытые мембраной пузырьки) (4). В гладких миоцитах нет L-трубочек. При поступлении потенциала действия и выделении медиатора изменяется состояние плазмолеммы. Через мембрану клетки в ее впячивания (кавеолы) поступают ионы кальция. Увеличивается содержание ионов кальция в цитоплазме. Происходит «сборка» миофибрилл, т.е. взаимодействие актина и миозина. Сближаются плотные тельца. Усилие передается на плазмолемму. Гладкий миоцит укорачивается. После прекращения потенциала действия ионы кальция накапливаются в кавеолах, миозин теряет сродство к актину. Т.е. происходит «разборка» миофибрилл. 34 Клетки какой ткани изображены на рисунке? Назовите структуры обозначенные цифрами.. Рис. 29. Поперечнополосатая сердечная мышечная ткань. Типичный кардиомиоцит (рабочий, сократительный). 1Ядро. 2.Комплекс Гольджи. 3.Гранулы гликогена. 4.Митохондрии. 5.Десмосомы. 6.Вставочный диск. 7.Миофибриллы. 8.Гемокапилляр. 9.Эндотелиоцит. 10.Перицит. 11.Эритроцит. Типичный кардиомиоцит – структурная единица сердечной мышцы (миокард). Клетки образуют так называемые «мышечные волокна», которые в отличие от мышечных волокон скелетной мышечной ткани, являющихся миосимпластом, представляют цепочки из клеток кардиомиоцитов, соединяющиеся друг с другом с помощью вставочных дисков (6). Кардиомиоциты обеспечивают сокращения сердечной мышцы. Клетки удлиненной формы с центрально расположенным ядром (1) (может быть два ядра). Вблизи ядра комплекс Гольджи (2) и гранулы гликогена (3). Сократительный аппарат представлен миофибриллами (7), между которыми лежат многочисленные митохондрии (4). Вблизи мембран Т-трубочек эндоплазматическая сеть образует терминальные расширения цистерн. В состав вставочных дисков входят десмосомы (5), плотные и щелевидные контакты (нексусы). Вставочные диски состоят из поперечных и продольных участков. Поперечные участки содержат преимущественно десмосомы и относительно небольшое количество щелевых контактов. Продольные участки имеют хорошо выраженные щелевые контакты. Десмосомы обеспечивают механическое сцепление, которые препятствуют расхождению кардиомиоцитов. Щелевые контакты способствуют передаче сокращения от одного кардиомиоцита к другому. Среди кардиомиоцитов видны гемокапилляры (8). 35 Фрагмент какой ткани на рисунке? Аргументируйте вывод. Назовите структуры, обозначенные цифрами? Рис. 34. Миелиновые и безмиелиновые волокна нервной ткани. 1.Миелиновое волокно: 2.Осевой цилиндр. 3.Слои миелина. 4.Нейролеммоцит. 5.Митохондрии. 6.Гранулярная ЭПС. 7.Безмиелиновое волокно: 8.Ядро леммоцита. 9.Осевые цилиндры. 10.Мезаксон. 11.Митохондрии. 12.Гранулярная ЭПС. 13.Фибробласт. Нервные волокна - это отростки нервных клеток, покрытые клетками олигодендроглии, которые называются нейролеммоцитами (шванновскими клетками) (4). Отросток нервной клетки в составе волокна называют осевым цилиндром (2). Оболочки нервных волокон в различных отделах нервной системы отличаются друг от друга по своему строению, на основании чего волокна делятся на две группы: миелиновые и безмиелиновые. Безмиелиновые волокна (7) находятся, в основном, в составе вегетативной нервной системы (например, в составе нервов, иннервирующих внутренние органы). Клетки олигодендроглии оболочек этих волокон располагаются плотно и образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В таких тяжах располагается не один, а несколько осевых цилиндров (9), принадлежащих различным нейронам. Нервные волокна, содержащие несколько осевых цилиндров (9), называются волокнами кабельного типа. При образовании безмиелиновых нервных волокон, осевые цилиндры (9) погружаются в нейролеммоцит, прогибая его оболочку, образуя складку-мезаксон (10). Оболочки нейролеммоцитов очень тонкие, поэтому мезаксон, границы клеток под микроскопом не видны. Миелиновые нервные волокна (1) встречаются как в центральной, так и в периферической нервной системе. Они толще, чем безмиелиновые. Миелиновые нервные волокна также состоят из осевого цилиндра (2), покрытого оболочкой из леммоцитов, но осевые цилиндры здесь толще, и оболочка построена сложнее. В миелиновом волокне различают два слоя оболочек: внутренний, более толстый – миелиновый слой (3) и наружный тонкий, состоящий из цитоплазмы и ядер нейролеммоцитов. По ходу волокон встречаются участки, где нет миелина-узловые перехваты, они соответствуют границам смежных леммоцитов. Отрезок волокна, заключенный между смежными перехватами называется межузловым сегментом. На определенном расстоянии по ходу волокна располагаются светлые линии - насечки миелина. При развитии миелинового волокна осевой цилиндр, погружаясь в нейролеммоцит, прогибает его оболочку, образуя глубокую складку формируя мезаксон. При развитии мезаксон удлиняется и концентрически наслаивается на осевой цилиндр, образуя вокруг него плотную слоистую зону - миелиновый слой (3). Наружным слоем называется периферическая зона нервного волокна, содержащая оттесненную цитоплазму нейролеммоцита и их ядра. Снаружи миелиновое волокно покрыто базальной мембраной. Осевой цилиндр нервных волокон состоит из нейроплазмы – цитоплазмы нервной клетки, содержащий продольно ориентированные нейрофиламенты и нейротубулы. В нейроплазме осевого цилиндра находятся митохондрии. С поверхности осевой цилиндр покрыт аксолеммой, обеспечивающий проведение нервного импульса. Скорость передачи импульса миелинового волокна больше, чем безмиелинового. Тонкие волокна, которые бедны миелином и безмиелиновые проводят нервный импульс со скоростью 1-2 м/с, а толстые миелиновые 5-120 м/с. В безмиелиновом волокне волна деполяризации мембраны идет по всей плазмолемме, не прерываясь, а в миелиновом волокне возникает только в области перехвата. Таким образом, для миелиновых волокон характерно сальтаторное (скачкообразное) проведение возбуждения. В пределах межузлового сегмента импульс движется как электрический ток, скорость которого выше чем прохождение волны деполяризации. Между нервными волокнами в нерве находится рыхлая волокнистая соединительная ткань (эндоневрий). На фотограмме клетка этой ткани - фибробласт (13). 36 Фрагмент какой ткани на рисунке? Назовите структуры, обозначенные цифрами. Рис. 35. Схема строения миелинового нервного волокна 1.Осевой цилиндр. 2 . Контакты нейролеммоциов в области перехвата Ранвье. 3.Миелиновый слой. 4. Насечки миелина. 5. Цитоплазма нейролеммоцита. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Диаметр поперечного сечения их колеблется от 2 до 20 мкм. Они состоят из осевого цилиндра (1), «одетого» оболочкой из нейролеммоцитов (шванновских клеток). Каждая шванновская клетка миелинизирует небольшой сегмент. Диаметр осевых цилиндров волокон этого типа значительно толще, а оболочка сложнее. В сформированном миелиновом волокне принято различать два слоя оболочки: внутренний, более толстый, — миелиновый слой и наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы. Миелиновый слой (3) содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии— насечки миелина (4), или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя— перехваты Ранвье (2). В процессе развития - аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. На электронных микрофотографиях видны главные плотные и интрапериодальные линии. Первые образуются от слияния цитоплазматических поверхностей плазмолеммы нейролеммоцита (или олигодендроглиоцита в центральной нервной системе), вторые — от контакта экстрацеллюлярных поверхностей соседних слоев плазмолеммы нейролеммоцита. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Аксолемма (оболочка аксона) обладает в области перехвата значительной электронной плотностью. Наличие большого числа митохондрий в этой области свидетельствует о высокой метаболической активности аксолеммы. Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (4) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита (5), т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана. Миелиновые волокна центральной нервной системы отличаются тем, что в них миелиновый слой формирует один из отростков олигодендроглиоцита. Остальные его отростки участвуют в образовании миелинового слоя других миелиновых волокон Миелиновые волокна центральной нервной системы не имеют насечек миелина, а нервные волокна не окружены базальными мембранами. Миелин в центральной нервной системе содержит миелиновый щелочной белок и протеолипидный белок. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые — со скоростью 5—120 м/с. В безмиелиновом волокне волна деполяризации мембраны идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехвата. Таким образом, для миелиновых волокон характерноскачкообразное (сальтаторное) проведение возбуждения. 37 Назовите тип клетки? Аргументируйте вывод. Назовите структуры, обозначенные цифрами? Рис. 36. Ультраструктура нейрона. 1.Плазмолемма 2. Ядро 3.Гранулярная эндоплазматическая сеть 4.Аппарат Гольджи 5.Лизосомы 6.Митохондрии 7.Элементы цитоскелета 8. Аксосоматический синапс 9. Кровеносный капилляр 10. Отростки нейронов. Нейроны, являются морфологически и функционально самостоятельной единицей, но с помощью своих отростков (10) осуществляет синаптический контакт с другими нейронами, образуя рефлекторные дуги. По количеству отростков различают: униполярные, имеющие только один аксон (у человека обычно не встречается); биполярные, имеющие один аксон и дендрит; мультиполярные, имеющие один аксон и много дендритов. Большинство нейронов человека содержит одно округлое ядро (2). При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляются в виде базофильных глыбок хроматофильная субстанция. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка состоит из цистерн гранулярной эндоплазматической сети (3), свободных рибосом и полисом, которые синтезируют белки цитозоля и интегральные белки плазмолеммы. В нейронах хорошо развит аппарат Гольджи (4), расположенный вокруг ядра, митохондрии (6), присутствуют лизосомы (5). Из элементов цитоскелета (7) в цитоплазме нейронов присутствуют нейрофиламенты и нейротубулы. Нейрофиламенты и нейротубулы участвуют в поддержании формы клеток, росте отростков и аксональном транспорте. Межнейрональные синапсы подразделяются на электрические и химические. К последним относится аксосоматический синапс (аксон одного нейрона контактирует с телом другого нейрона). 38 Что представлено на схеме? Назовите структуры, обозначенные цифрами. Рис. 37. Химический синапс. 1.Пресинаптическая часть. 2.Постсинаптическая часть. 3.Синаптические пузырьки. 4.Митохондрии. 5.Микротрубочки, микрофиламенты. 6.Пресинаптическая мембрана. 7.Постсинаптическая мембрана с постсинаптическим уплотнением. Химический синапс состоит из трех компонентов: пресинаптической части (1), постсинаптической части (2) и синаптической щели. Пресинаптическая часть образуется терминалью аксона. В ней содержатся митохондрии (4), агранулярная ЭПС, микрофиламенты, микротрубочки (5) и синаптические пузырьки (3) диаметром 20-65 нм, в которых находится нейромедиатор. Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. Нейромедиаторы вырабатываются в теле нейрона и механизмом быстрого транспорта переносятся в окончания аксона. На внутренней стороне пресинаптической мембраны имеется пресинаптическое уплотнение, образованное белковой сетью. Синаптическая щель шириной 20-30 нм содержит элементы гликокаликса, которые обеспечивают адгезию и направленную диффузию медиатора. Постсинаптическая часть представлена постсинаптической мембраной (7), содержащей интегральные белки – рецепторы, связывающиеся с нейромедиатором. Мембрана утолщена. В зависимости от того, является ли постсинаптической частью тело нейрона, дендрит или аксон, синапсы подразделяют на аксо-соматические, аксо-дендритические и аксо-аксональные. Под действием нервного импульса происходит открытие кальциевых каналов пресинаптической мембраны, Са 2+ устремляется в аксон, мембраны синаптических пузырьков в присутствии Са 2+ сливаются с пресинаптической мембраной, и содержащийся в них медиатор выделяется в синаптическую щель. Связываясь с рецепторами постсинаптической мембраны, медиатор вызывает ее деполяризацию и возникновение нервного импульса, или ее гиперполяризацию, обусловливая торможение. После экзоцитоза медиатора, большая часть его захватывается пресинаптической частью и используется повторно, поглощается окружающими глиальными клетками, некоторые медиаторы (например, ацетилхолин) расщепляются ферментами. Химический синапс обеспечивает передачу нервного импульса в одном направлении. 39 Рис. 38. Форменные элементы крови. 1.Эритроцит. 2.Сегментоядерный нейтрофил. 3.Эозинофил. 4.Базофил. 5.Лимфоцит. 6.Моноцит. 7.Тромбоцит. Эритроциты (1) – самые многочисленные клетки крови, утратившие в процессе дифференцировки ядро и практически все органеллы. Большинство эритроцитов (7585%) имеют форму двояковогнутого диска. Поддержание формы эритроцитов обеспечивается осмотическим равновесием (работой ионных насосов плазмолеммы), элементами цитоскелета. Функции эритроцитов: перенос газов (преимущественно кислорода) с помощью гемоглобина, перенос других веществ на своей поверхности (гормонов, иммуноглобулинов и др.). Нейтрофилы (2) – клетки с разной степенью дифференцировки, отличаются в мазках по форме ядра. Юные нейтрофилы имеют бобовидную форму ядра (составляют 0,5%). Палочкоядерные нейтрофилы имеют ядро в виде палочки или подковы (3,5%) . Сегментоядерные нейтрофилы (2) (60-65%) имеют дольчатое ядро, состоящее из 2-5 сегментов. Зрелые нейтрофилы содержат немного митохондрий, значительное количество гранул гликогена, хорошо развитый цитоскелет. Эозинофилы (3) составляют (1-5%), содержат палочковидное ядро или сегментированное (два сегмента), в клетках хорошо развита гранулярная ЭПС, большое количество рибосом, гранул гликогена. В цитоплазме гранулы двух типов: неспецифические (азурофильные), аналогичны лизосомам и специфические - содержат кристаллоид, образованный белками (обуславливает эозинофилию). Функции: уничтожение микроорганизмов, паразитов (гельминтов). Способны к фагоцитозу. Базофилы (4) составляют 0,1% от общего числа лейкоцитов циркулирующей крови. Чаще имеют трехдольное ядро. Содержат все виды органелл, свободные рибосомы и гликоген. Цитоплазма базофилов содержит специфические крупные метахроматические гранулы, которые содержат протеогликан, гистамин, пероксидазу; азурофильные гранулы - аналогичны лизосомам. При действии аллергена происходит быстрый экзоцитоз содержимого гранул с большим количеством биологически активных веществ, которые привлекают другие клетки к защитной реакции организма. Лимфоциты (5) составляют 20-45% от общего числа лейкоцитов, циркулирующих в крови. Эти клетки играют центральную роль в иммунологических реакциях. Способны выходить из крови в ткань, затем снова возвращаться в кровь через лимфу. В крови находится лишь 2% лимфоцитов, 98% рассредоточено по другим органам и тканям. Лимфоциты имеют округлую форму с крупными ядром, занимающим до 20% обьема клетки. Цитоплазма слабо базофильна. Все органеллы содержатся в небольшом количестве, кроме цитоскелета, который хорошо развит. По размерам лимфоциты делят: малые (6-7 мкм) их 80-90% от общего количества лимфоцитов крови; средние (8-9 мкм.) -10% и большие (10-18 мкм) - в норме в крови отсутствуют. Более существенна функциональная классификация. Согласно ей лимфоциты подразделяются на Т и В клетки. Они различаются местом дифференцировки. Т-лимфоциты дифференцируются в тимусе. В-лимфоциты в красном костном мозге. Обеспечивают различные типы иммунитета: Т-лимфоциты преимущественно клеточный, а В-лимфоциты гуморальный иммунитет. Функционально Т- и В-лимфоциты делят на субпопуляции. Среди Т-лимфоцитов выделяют Тхелперы - активируют эффекторные клетки; Т-киллеры – эффекторные цитотоксические клетки; Т-супрессоры – подавляют иммунный ответ, Т-памяти. В-лимфоциты дифференцируются в плазматические клетки, вырабатывающие иммуноглобулины (антитела) и клетки памяти, несущие информацию о встрече с антигеном. Т-лимфоциты составляют 70-80% в крови, Влимфоциты - 10-20%. Моноциты (6) - самые крупные лейкоциты (15 мкм) их 2-9% всех лейкоцитов циркулирующей крови. Образуются в красном костном мозге, а затем выходят в кровоток. Это незрелые клетки, находящиеся на пути из костного мозга в ткани. В тканях моноциты дифференцируются в подвижные макрофаги, их совокупность - система мононуклеарных фагоцитов. Тромбоциты (7) - являются фрагментами цитоплазмы мегакариоцитов красного костного мозга, поэтому правильнее называть их кровяные пластинки. В мазке крови агрегируются поэтому выявляются в виде скоплений. Кровяные пластинки – это овальные, двояковогнутые тельца с небольшими отростками. Внутреннее содержимое состоит из двух частей. Центральная часть – грануломер содержит азурофильные зерна, наружная –гиаломер имеет гомогенную структуру и бледно – голубую окраску. Плазмолемма покрыта слоем гликокаликса. Он состоит из множества рецепторов, которые обусловливают прикрепление тромбоцита к эндотелию и склеивание тромбоцитов друг с другом (агрегация). Гиаломер представляет собой однородную тонкозернистую структуру с микротрубочками и филаментами по периферии. Микротрубочки формируют краевое кольцо – жесткий каркас тромбоцита. В грануломере содержатся гранулы гликогена, единичные рибосомы и гранулы нескольких типов: 1) азурофильные гранулы содержат вещества, участвующие в свертывании крови (фибронектин, фибриноген);2) гранулы с плотным матриксом содержат АТФ, ионы кальция, магния, гистамин, серотонин; 3) гранулы, содержащие гидролитические ферменты, соответствуют лизосомам. Функции тромбоцитов: 1) восстанавление сосудистой стенки при повреждении; 2) свертывание крови; 3) участвуют в иммунных реакциях (вырабатывают факторы хемотаксиса клеток в иммунной системе). Количество тромбоцитов 200-400×109 (л). 40 Что представлено на схеме? Назовите структуры, обозначенные цифрами. Рис. 41. Ретикулоциты в мазке крови (окраска крезилвиолетом). 1.Эритроцит. 2.Ретикулоцит. 3.Базофильная зернистость. Эритроциты в организме ежедневно заменяются новыми. Ретикулоциты-это молодые эритроциты. В кровотоке в норме присутствует около 1% молодых эритроцитов, сохранивших в цитоплазме небольшое количество рибосом, обеспечивавших на более ранних стадиях развития синтез гемоглобина. Созревание ретикулоцитов происходит в течение 24-48 часов. При специальном окрашивании мазка крови бриллианткрезиловым синим рибосомы выявляются в виде базофильной зернистости, поэтому такие эритроциты назвали ретикулоцитами. Содержание ретикулоцитов может повышаться как вследствие абсолютного увеличения количества ретикулоцитов в крови, так и сокращения массы циркулирующих эритроцитов (анемия). Если причиной анемии являются кровопотеря или разрушение эритроцитов, то возрастает секреция эритропоэтина и относительное количество ретикулоцитов поднимается выше нормального уровня (1%), а абсолютное число ретикулоцитов превышает величину 100 000 в мкл. Отсутствие ретикулоцитоза при анемии указывает на нарушение продукции эритроцитов в костном мозге из-за недостаточности питания или заболеваний костного мозга. 41 Назовите клетку, изображенную на электронной микрофотографии. Укажите признаки, характерные для данного вида клеток. Определите тип электронной микроскопии. Рис. 43. Эозинофильный гранулоцит (эозинофил) крови. Видны характерные гранулы, содержащие кристаллоид и сегментированное ядро (два сегмента). Трансмиссионная электронная микроскопия. Ув. 16400. Назовите клетку, аргументируя вывод. Назовите структуры, обозначенные цифрами. Рис. 44. Базофильный гранулоцит (базофил). 1.Базофильные гранулы. 2.Азурофильные гранулы. 3.Ядро. 4.Гранулярная эндоплазматическая сеть. 5.Комплекс Гольджи. 6.Митохондрии. Базофилы – самая малочисленная группа гранулоцитов, их содержание в крови составляет 0,5-1,0 % от общего числа лейкоцитов. В крови базофилы циркулируют до 1 суток, а затем перемещаются в ткани. Строение и функции базофилов схожи с таковыми тучных клеток рыхлой волокнистой соединительной ткани. Размеры базофилов на мазках составляют 912 мкм. Ядро (3) клеток дольчатое (содержат 2-3 сегмента) или S – образное, относительно плотное, но с меньшим содержанием гетерохроматина, чем у нейтрофилов и эозинофилов. Ядра нередко трудно различимы, так как маскируются цитоплазматическими гранулами. В цитоплазме базофильных гранулоцитов под электронным микроскопом выявляются митохондрии (6), элементы цитоскелета, сравнительно слабо развитый синтетический аппарат и гранулы двух типов – специфические (базофильные) (1) и неспецифические (азурофильные) (2), представляют собой лизосомы). Специфические (1) (базофильные) гранулы – крупные (диаметром 0,5-2,0 мкм), сферической формы, хорошо видны в световой микроскоп, окрашиваются основными красителями. Гранулы окружены мембраной, более зрелые гранулы обладают большей плотностью. При дегрануляции содержимое выделяется путем слияния мембраны гранул с плазмолеммой базофила или часть гранул выстраивается в цепочки, где они сливаются друг с другом, а далее их содержимое выделяется из клетки. Дегрануляция активированных базофилов происходит в присутствии ионов кальция. Выделение содержимого гранул может происходить в виде медленной секреции или массивной дегрануляции. Последнее обусловливает участие базофилов в аллергических реакциях. Содержимое базофильных гранул: гистамин (расширяет сосуды, увеличивает их проницаемость), гепарин (антикоагулянт), хондроитинсульфат, ферменты (протеазы, пероксидаза), хемотаксические факторы эозинофилов и нейтрофилов. Выделение биологически активных веществ из гранул (дегрануляция) происходит в ответ на связывание рецепторов базофилов с иммуноглобулинами класса Е, компонентами комплемента, бактериальными продуктами, цитокинами. Снижение содержания базофилов в крови чаще всего происходит вместе со снижением количества эритроцитов (опухоли, инфекции, воспалительные заболевания и др.). 42 Глава 3. ОРГАНЫ СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ Определите тканевую и органную принадлежность клеток и опишите их. Назовите обозначенные структуры. Рис. 1. Кардиомиоциты проводящей системы сердца. А – Р-клетки (пейсмекеры) – водители ритма (I тип); Б – Переходные (промежуточные) клетки (II тип); В – клетки пучка Гиса и волокон Пуркинье (III тип); 1 – миофибриллы, 2 – ядра, 3 – митохондрии, 4 – гликоген. Кардиомиоциты проводящей системы сердца (атипичные кардиомиоциты) обеспечивают его функцию автоматизма – способность вырабатывать электрические импульсы при отсутствии внешних раздражений. Атипичные кардиомиоциты генерируют биопотенциалы, проводят их и передают на сократительные кардиомиоциты. В их цитоплазме содержится мало неупорядоченно расположенных миофибрилл (1), поэтому отсутствует поперечная исчерченность; с помощью десмосом и щелевидных контактов формируют волокна. Выделяют три типа клеток. Клетки I типа (А) (пейсмекеры, от англ. pace – темп, maker – создатель) составляют основу синуснопредсердного (синоатриального) узла – водителя ритма, расположенного в стенке правого предсердия между верхней полой веной и правым ушком, а также в небольшом количестве находятся в атриовентрикулярном узле. Несколько клеток заключены в единую базальную мембрану, к которой подходит много нервных окончаний (парасимпатическая система замедляет сердечные сокращения, симпатическая – ускоряет). Эти клетки меньше рабочих кардиомиоцитов, светлые, с небольшим содержанием миофибрилл (1) и крупными ядрами (2), с малым количеством органелл общего назначения (митохондрии – 3) и гликогена (4). Клетки синоатриального узла способны спонтанно генерировать биопотенциалы с частотой 60-90 импульсов в минуту в норме и передавать их на переходные клетки (II типа). Клетки II типа (переходные) (Б) передают импульсы от клеток I типа на клетки III типа. Их локализации – преимущественно предсердно-желудочковый (атриовентрикулярный) узел, находящийся в стенке между предсердиями над правым предсердно-желудочковым клапаном. Эти клетки чуть больше клеток I типа, имеют крупное ядро (2), содержат небольшое количество миофибрилл (1), митохондрий (3) и гликогена (4). Способны генерировать импульсы (при выключении синоатриального узла) с частотой 40-60 в минуту. Клетки III типа (В) - самые крупные клетки миокарда, находятся в предсердно-желудочковом пучке Гиса, идущего от предсердно-желудочкового узла в межжелудочковой перегородке и разделяющегося на правую и левую ножки, которые в миокарде желудочков ветвятся и называются волокона Пуркинье. При выключении вышестоящего атриовентрикулярного узла генерируют импульсы с частотой 20-40 в минуту. В клетках III типа содержится редкая неупорядоченная сеть миофибрилл (1), имеющих спиралевидный ход; многочисленные мелкие митохондрии (3), большое количество гликогена (4), т.е. активно проходят как аэробные так и анаэробные процессы. От клеток пучка и волокон Пуркинье биопотенциалы передаются на сократительные кардиомиоциты. Цитолемма пейсмекерных клеток обладает особыми свойствами: в то время как в диастолу трансмембранный потенциал сократительных кардиомиоцитов не меняется, оставаясь на уровне -90 mV, в клетках проводящей системы происходит медленная спонтанная диастолическая деполяризация (самопроизвольное увеличением проницаемости для ионов Nа+, медленно входящих в клетки). В результате этого разность потенциалов между наружной и внутренней поверхностью мембраны постепенно уменьшается. Как только разность достигает критического уровня (примерно —60 mV), проницаемость мембраны для ионов Na+ резко возрастает, что приводит к возникновению быстрой лавинообразной деполяризации клетки — ее возбуждению, которая является импульсом к возбуждению других клеток миокарда. На ЭКГ отражается сокращение только рабочих кардиомиоцитов. 43 Фрагмент какого органа изображен на рисунке? Какие ткани, клетки его образуют? Рис. 2. Фрагмент миокарда (мышечной оболочки сердца). Рабочие кардиомиоциты (типичные, сократительные). 1.Сократительный кардиомиоцит. 2.Анастомоз между соседними кардиомиоцитами. 3.Капилляр. 4.Рыхлая волокнистая соединительная ткань. 5.Вставочный диск. 6.Базальная мембрана. 7.Ядра кардиомиоцитов. 8. Комплекс Гольджи. 9.Гранулярная ЭПС. 10.Миофибриллы. 11.Митохондрии. Миокард - мышечная оболочка сердца. Состоит из тесно связанных между собой мышечных клеток – кардиомиоцитов (1). Клетки имеют удлиненную форму, близкую к цилиндрической, и соединяются друг с другом в цепочки, образуя функциональные волокна. Их поверхности покрыты базальной мембраной (6). Кардиомиоцит имеет обычно одно, реже два центрально расположенных овальных ядра (7), оксифильную цитоплазму. У полюсов ядра сосредоточены немногочисленные органеллы общего назначения (8,9), за исключением агранулярной эндоплазматической сети и митохондрий. Основную часть клетки занимают миофибриллы (10) – органеллы специального назначения, обеспечивающие сокращение. По строению они аналогичны миофибриллам скелетной мышечной ткани. Между ними располагаются многочисленные митохондрии (11) и цистерны гладкой эндоплазматической сети. В области соединения клеток образуются вставочные диски (5) – сложные межклеточные контакты, сочетающие плотные контакты (десмосомы) для прочности соединения кардиомиоцитов, интердигитации и щелевидные (нексусы) для передачи нервного импульса от клетки к клетке по волокну. При световой микроскопии межклеточные соединения представляют собой вид темных полос. Кардиомиоциты образуют анастомозы (2) формируя, таким образом, пространственную сеть. Благодаря анастомозам миокард представляеи собой единое целое. Между мышечными элементами располагаются прослойки рыхлой соединительной ткани (4), сосуды (3), нервы. Сердечная мышца прикрепляется к скелету сердца (фиброзные кольца вокруг атриовентрикулярных клапанов, клапанов аорты, легочной артерии). 44 Какой вид сосуда изображен на рисунке? Какими слоями и тканями образована его стенка. Где встречаются такие сосуды? Назовите структуры, обозначенные на схеме. Рис. 3. Гемокапилляр соматического типа. 1.Эндотелиоцит. 2.Непрерывная базальная мембрана. 3.Перицит. 4.Адвентициальная клетка. 5.Эритроцит в просвете капилляра. Кровеносные капилляры самые многочисленные и самые тонкие сосуды в организме. Основная функция – обменные процессы между кровью и тканями, которая обеспечивается тонкостью стенок капилляров, огромной площадью соприкосновения их с тканями, медленным кровотоком и низким кровяным давлением. Соматические капилляры – наиболее распространенный в организме тип гемокапилляров, они встречаются в ЦНС, сердечной и скелетных мышцах, легких. Их диаметр 4,5-8 мкм. Они участвуют в образовании гистогематических барьеров, которые препятствуют проникновению антигенов и токсинов из крови в ткани. Стенка соматического капилляра состоит из трех слоев: 1.Эндотелиальный слой представлен одним пластом эндотелиальных клеток полигональной формы (1), лежащих на сплошной базальной мембране (2). Их размеры варьируют от 5 до 175 мкм. Наименьшая толщина составляет 200 нм, а в ядросодержащей зоне, которая выпячивается в просвет капилляра, достигает 1-2 мкм. Эндотелиоциты соединяются друг с другом с помощью плотных, десмосомальных, интердигитирующих и черепичных контактов. Эндотелий выполняет транспортную (обмен веществ между кровью и тканями), атромбогенную (синтез веществ, ингибирующих агрегацию тромбоцитов), барьерную функции, обеспечивает регуляцию сосудистого тонуса (вырабатывает факторы регуляции расслабления и сокращения гладких миоцитов), участвует в образовании базальной мембраны. Базальная мембрана – это тонкофибриллярная, полупроницаемая пластина толщиной 30-35 нм. Состоит из коллагена IV и V типов, гликопротеинов, фибронектина, ламинина и сульфатсодержащих протеогликанов. Выполняет опорную, разграничительную, барьерную функции. 2.Перициты (3) – соединительнотканные клетки отростчатой формы, окружающие капилляры в виде корзинки. Располагаются в расщеплениях базальной мембраны, на которой располагается эндотелий. В капиллярах соматического типа образуют хорошо выраженный средний слой. 3.Адвентициальный слой состоит из малодифференцированных клеток (адвентициальных), окруженных аморфным веществом и тонкими коллагеновыми волокнами. Адвентициальные клетки (4) являются предшественниками фибробластов, адипоцитов и остеобластов. 45 Какой вид сосуда изображен на рисунке? Какими слоями и тканями образована его стенка. Где встречаются такие сосуды? Назовите структуры, обозначенные на схеме. Рис. 4. Гемокапилляр фенестрированного (висцерального) типа. 1.Эритроцит в просвете капилляра. 2.Эндотелиоцит. 3.Перицит. 4.Базальная мембрана. 5.Фенестры. Кровеносные капилляры самые тонкостенные сосуды микроциркуляторного русла. Основная функция – транспорт веществ из крови в ткани и из тканей в кровь. Для всех капилляров характерен общий план строения. Капилляры висцерального типа имеют диаметр от 8 до 12 мкм. Характеризуются непрерывным слоем эндотелиоцитов (2) с фенестрами (5), лежащих на истонченной базальной мембране (4) и сниженным (по сравнению с соматическими капиллярами) количеством перицитов (3) и адвентициальных клеток. Фенестры – это сильно истонченные участки эндотелия (отверстия, затянутые клеточной мембраной), увеличивающие проницаемость для макромолекул (например, гормонов в эндокринных железах; питательных веществ, всасывающихся в тонкой кишке). Проницаемость увеличивается про воздействии гиалуронидазы, которая разрушает гиалуроновую кислоту (при этом увеличивается обмен веществ), а также повышается при воздействии гистамина. Такие капилляры встречаются в эндокринных органах, слизистой оболочке тонкой кишки, почках. 46 Какой вид сосуда изображен на рисунке? Какими слоями и тканями образована его стенка. Где встречаются такие сосуды? Назовите структуры, обозначенные на схеме. Рис. 5. Гемокапилляр перфорированного (синусоидного) типа. 1.Эндотелиоцит. 2.Поры (щели). 3.Прерывистая базальная мембрана. 4.Эритроцит. Синусоидные капилляры отличаются от других типов гемокапилляров большим диаметром, который составляет более 12 мкм (например, в красном костном мозге встречаются капилляры с диаметром около 200-500 мкм). Для них характерен эндотелий со сквозными отверстиями (порами), которые облегчают проникновение зрелых клеточных элементов крови из кроветворных органов в кровеносное русло, а также сильно истонченная, прерывистая базальная мембрана и полное отсутствие перицитов и адвентициальных клеток. Базальная мембрана иногда также может отсутствовать. Проницаемость для зрелых клеток крови регулируется элементами стромы органов (например, в красном костном мозге ретикулярными клетками). Такие капилляры встречаются в кроветворных органах (красном костном мозге, селезенке), печени. 47 Какой сосуд изображен на рисунке? Какие клетки его образуют? Рис. 8. Лимфатический капилляр. 1.Полость капилляра. 2.Эндотелиоцит. 3.Стропные (фиксирующие) филаменты. 4.Лимфоцит. Лимфатические сосуды делят на лимфатические капилляры, выносящие интраорганные и экстраорганные лимфатические сосуды, крупные лимфатические стволы, которые несут лимфу в венозную систему (правый и левый венозные углы, образованные внутренними яремными и подключичными венами). Лимфатические капилляры располагаются рядом с кровеносными капиллярами. Лимфатические капилляры – замкнутые с одного конца начальные отделы лимфатической системы. Они, анастомозируя друг с другом, пронизывают органы (лимфатические капилляры не обнаружены в головном мозге, селезенке, плаценте, костном мозге, склере и хрусталике глазного яблока, в покровных эпителиях и хрящевых тканях). Их стенка образована крупными ( в сравнении с кровеносными капиллярами в 4 раза крупнее) эндотелиальными клетками (2). В клетках комплекс Гольджи, гранулярная ЭПС, митохондрии, лизосомы, вакуоли, везикулы. Эндотелитоциты характеризуются высокой проницаемостью и реактивностью на внешние воздействия и чужеродные агенты. Они обладают большой растяжимостью. Просвет лимфатического капилляра может увеличиваться в три раза без повреждения стенок. На наружной поверхности эндотелиальных клеток имеются якорные или стропные микрофиламенты (3). Они прикреплены к плазмолемме и прикрепляют ее, в свою очередь, к окружающим соединительнотканным структурам. В отличие от гемокапилляров перициты отсутствуют, а базальная мембрана значительно истончена и прерывиста. В лимфатических капиллярах образуется периферическая лимфа. Сюда мигрируют лимфоциты (4) из тканей, участвующие в рециркуляции. Могут поступать раковые клетки, которые далее попадают в кровеносную систему и разносятся по всему организму. Таким образом, из кровеносных капилляров в межклеточное вещество поступают необходимые компоненты основного вещества, откуда далее уже в лимфатические капилляры поступают продукты метаболизма. Лимфа образована компонентами межтканевой жидкости и состоит из лимфоплазмы (вода, органические веществыа, минеральные соли) и форменных элементов среди которых 98% составляют лимфоциты, а 2% приходится на остальные форменные элементы. Лимфа обновляет межклеточное вещество, очищает его от вредных продуктов. 48 ГЛАВА 4. ОРГАНЫ КРОВЕТВОРЕНИЯ Фрагмент какого органа изображен на рисунке? Какие клетки Вы узнаете? Рис. 1. Фрагмент красного костного мозга. Синусоидный капилляр. Мегакариоцит, другие кроветворные клетки, клетки стромы. Образование тромбоцитов. 1.Синусоидный (перфорированный) капилляр. 2.Мегакариоцит. 3.Отделяющиеся кровяные пластинки. 4.Дольчатое (многолопастное) полиплоидное ядро мегакариоцита. 5.Ретикулярная клетка стромы. Красный костный мозг – центральный орган кроветворения (гемопоэза), в котором образуются все форменные элементы крови (кроме Т-лимфоцитов). В красном костном мозге, как и в любой гемопоэтической ткани имеются следующие компоненты: гемопоэтические клетки (в данном случае-мегакариоцит), стромальный компонент –ретикулярная ткань, клетки макрофагического ряда, специфические сосуды. В красном костном мозге происходит антигеннезависимая дифференцировка В-лимфоцитов. Мегакариоцит (2) - дифференцированная клетка тромбоцитопоэза (процесс образования кровяных пластинок). Зрелый, активный мегакариоцит – крупная клетка диаметром до 100 мкм, располагается в тесном контакте с синусоидным капилляром (1), так что периферическая часть его цитоплазмы проникает в просвет сосуда. Содержит очень крупное, дольчатое полиплоидное ядро (до 64 n) (4). В цитоплазме накапливаются линейно расположенные микровезикулы и многочисленные азурофильные гранулы, содержащие факторы свертывания и другие биологически активные вещества необходимые для функционирования тромбоцитов. Из микровезикул формируются демаркационные мембраны, разделяющие цитоплазму мегакариоцита на фрагменты 1-3 мкм и содержащие по 1-3 гранулы (будущие кровяные пластинки). Процесс демаркации активно происходит в наружной части цитоплазмы и приводит к отделению тромбоцитов (3) от мегакариоцита. После завершения процесса демаркации остается резидуальный мегакариоцит – клетка, содержащая дольчатое ядро, окруженное узким ободком цитоплазмы. 49 Фрагмент какого органа изображен на рисунке? Какие структуры и клетки Вы узнаете? Рис. 2. Фрагмент красной пульпы селезенки. Синус селезенки. 1.Просвет синуса селезенки. 2.Макрофаг, фагоцитирующий эритроцит. 3.Эритроциты. 4.Эндотелиоциты стенки синуса. 5.Ретикулярные клетки стромы селезенки. Селезенка – периферический орган кроветворения и иммунной защиты. Участвует в формировании гуморального и клеточного иммунитета. В ней происходит лимфопоэз, элиминация отживших и поврежденных эритроцитов и тромбоцитов, депонирование крови и др. Снаружи селезенка покрыта брюшиной и капсулой из плотной соединительной ткани. От последней вглубь органа отходят трабекулы. В селезенке выделяют белую и красную пульпу. Красная пульпа внешне отличается от белой: в ней меньше лимфойдных элементов. Она образована ретикулярной тканью (5), образующей ее строму, многочисленными кровеносными сосудами (1), главным образом синусоидного типа и клеточными элементами крови (3), придающими красный цвет. Синусы являются началом венозной системы селезенки. Их диаметр колеблется от 12 до 40 мкм в зависимости от кровенаполнения. Стенка синусов образована эндотелиоцитами (4), которые располагаются на прерывистой базальной мембране. Перициты отсутствуют. Венозные синусы вследствие высокого содержания клеток крови на микропрепаратах плохо различимы. При растяжении синусов в результате кровенаполнения, между эндотелиальными клетками образуются щели, через которые кровь может проходить в ретикулярную строму (3). В красной пульпе находятся макрофаги селезенки (2), которые поглощают старые и поврежденные эритроциты. Гемоглобин, поглощенных макрофагами эритроцитов расщепляется на билирубин и содержащий железо трансферрин. Билирубин транспортируется в печень, где войдет в состав желчи. Трансферрин захватывается из кровотока макрофагами красного костного мозга и вновь участвует в образовании гемоглобина развивающихся там эритроцитов. 50 Фрагмент какого органа изображен на рисунке? Какие клетки Вы узнаете? К каким тканям они относятся? Рис. 3. Синус лимфатического узла. Ретикулярные клетки ретикулярной ткани, лимфоциты на разных стадиях развития (лимфоидная ткань). 1.Ретикулярные клетки. 2.Лимфоциты. 3.Плазмоцит. 4.Просвет синуса лимфатического узла. 5.Макрофаг. 6.Ретикулоэндотелиальная клетка. Лимфатические узлы – периферические органы кроветворения, играют роль в развитии иммунных реакций. Располагаются по ходу лимфатических сосудов. Пространства в лимфатических узлах ограниченные капсулой и трабекулами с одной стороны и узелками коркового вещества и мозговыми тяжами с другой стороны называются синусами (4). Они обеспечивают медленный ток лимфы по узлу, при этом очищают ее от антигенов, обогащение лимфоцитами и их продуктами (антителами, интерлейкинами и др.), таким образом, выполняют роль защитных фильтров. Ток лимфы осуществляется последовательно. Различают краевой синус (располагающийся между капсулой и лимфоидными узелками, который является продолжением приносящих лимфатических сосудов), вокругузелковые, мозговые и воротный (является началом выносящего лимфатического сосуда). Синусы выстланы ретикулоэндотелиальными клетками (6). На поверхности, обращенной к капсуле и трабекулам, они лежат на базальной мембране, а на поверхности, которая обращена к лимфоидным узелкам и мозговым тяжам, располагаются на сети ретикулярных волокон. Между клетками обнаруживаются щели, через которые в просвет синуса проникают лимфоциты. Среди ретикулоэндотелиальных клеток различают береговые клетки являющиеся макрофагами. Они фагоцитируют антигены, которые находятся в протекающей через лимфатический узел лимфе. Среди свободных клеточных элементов в синусах при различных состояниях организма встречаются лимфоциты (2), плазмоциты (3), свободные макрофаги (5), единичные зернистые лейкоциты. Отросчатые ретикулярные клетки замедляют ток лимфы. 51 ГЛАВА 5. ОРГАНЫ НЕРВНОЙ СИСТЕМЫ Что представлено на схеме? Назовите структуры обозначенные цифрами? Аргументируйте выводы. Рис. 1. Моторная бляшка (нервно-мышечное окончание). 1.Цитоплазма нейролеммоцита. 2. Митохондрия. 3.Плазмолемма нейролеммоцита. 4.Осевой цилиндр нервного волокна 5.Аксолемма 6.Постсинаптическая мембрана (сарколемма) 7.Митохондрии в аксоплазме. 8.Синаптическая щель. 9.Митохондрии мышечного волокна.10.Пресинаптические пузырьки 11.Пресинаптическая мембрана (аксолемма). 12.Сарколемма. 13.Ядра мышечного волокна. 14.Миофибриллы. Моторные бляшки представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Нервные терминали покрывают нейролеммоциты. Они контактируют и с мышечными волокнами, где уже являются глиальными клетками. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна (4) и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой плазмолемму и базальную мембрану мышечного волокна. Мембрана мышечного волокна (6) при этом образует складки. В месте формирования синапса в мышечном волокне много митохондрий (9). Плазмолеммы терминальных ветвей аксона и мышечного волокна разделены синаптической щелью (8), заполненной аморфным веществом, богатым гликопротеидами. Терминальные ветви нервного волокна в нервно-мышечном окончании характеризуются обилием митохондрий (7) и пресинаптическими пузырьками (10), содержащими характерный для этого вида окончаний медиатор – ацетилхолин. Импульс движется по плазмолемме аксона (пресинаптическая мембрана). Это приводит к поступлению в терминали ионов кальция. При этом синаптические пузырьки с ацетилхолином подходят к плазмолемме аксона и из пузырьков ацетилхолин изливается в синаптическую щель. Ацетилхолин захватывается рецепторами постсинаптической мембраны в результате чего повышается ее проницаемость. Ионы натрия с наружной поверхности плазмолеммы мышечного волокна переходят на ее внутреннюю поверхность, а ионы калия на наружную поверхность- это и есть волна деполяризации или потенциал действия. Локальная деполяризация постсинаптической мембраны приводит к генерации потенциала действия, который быстро распространяется по плазмолемме мышечного волокна, включая Т-трубочки. Запускается механизм мышечного сокращения. Взаимодействие ацетилхолина и рецепторов постсинаптической мембраны кратковременно. Ацетилхолинэстераза постсинаптической мембраны разрушает ацетилхолин и переход импульса через синаптическую щель прекращается. Нарушения нервно-мышечной передачи приводят к нарушению мышечного сокращения. В частности, нейротоксины при ботулизме блокируют секрецию ацетилхолина. Аналогичным образом действуют нейротоксины при столбняке. Фосфорорганические соединения инактивируют ацетилхолинэстеразу. Ряд заболеваний проявляется мышечной слабостью. Например, myasthenia gravis (псевдопаралитическая миастения), аутоиммунное заболевание, обусловленное образованием антител против ацетилхолиновых рецепторов. В результате количество участков постсинаптической мембраны доступных для инициирования деполяризации сарколеммы уменьшается. Первыми поражаются активные группы мышц - лица, глаз, языка. Затем вовлекаются и другие группы мышц. Поражение дыхательной мускулатуры приводит к развитию дыхательной недостаточности и смерти больного. Препараты, нарушающие нервно-мышечную передачу используют в анестезиологии (миорелаксанты). Сюда относят кураре и курареподобные препараты, которые препятствуют воздействию ацетилхолина на постсинаптическую мембрану; могут оказывать миопаралитический эффект характеризующийся стойкой деполяризацией постсинаптической мембраны. 52 Фрагмент какого органа на рисунке? Назовите структуры, обозначенные цифрами. Рис. 2. Нервный ствол в поперечном разрезе. 1.Миелиновое нервное волокно. 2.Безмиелиновое нервное волокно. 3.Осевой цилиндр. 4. Миелиновая оболочка. 5. Ядро нейолеммоцита. 6. Мезаксон. 7. Эндоневрий. Ув. 14400. Нервный ствол состоит из миелиновых (1) и безмиелиновых (2) нервных волокон и соединительнотканных оболочек. Центральная часть миелиновых нервных волокон занята осевым цилиндром (3). Миелиновая оболочка (4) окружает осевой цилиндр в виде темного ободка. Нервные волокна собираются в пучки разного диаметра, окружённые периневрием; между нервными волокнами прослойки соединительной ткани — эндоневрий (8). Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты (5) оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить на смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон (7) , на которой как бы подвешен осевой цилиндр. Оболочки нейролеммоцитов очень тонкие, поэтому ни мезаксона, ни границ этих клеток под световым микроскопом нельзя рассмотреть, и оболочка безмиелиновых волокон в этих условиях выявляется как однородный тяж цитоплазмы, «одевающий» осевые цилиндры. 53 ГЛАВА 6. ОРГАНЫ ЭНДОКРИННОЙ СИСТЕМЫ К какому органу относятся клетки? Аргументируйте вывод. Назовите структуры, обозначенные цифрами? Рис. 1. Клетки передней доли гипофиза. 1.Хромофобные клетки. 2.Тиротропоциты. 3.Соматотропоцит. 4.Пролактотропоцит. 5.Фолликулярно-звездчатые клетки. 6.Гонадотропоциты. 7.Кортикотропоциты. 8.Расширенный капилляр фенестрированного типа. Клетки аденогипофиза называются аденоцитами и располагаются в виде тяжей. Выделяют два вида клеток: хромофобные (1) и хромофильные. Хромофобные (1) клетки преобладают, поэтому их называют еще главными. Цитоплазма слабо окрашивается красителями. Хромофильные аденоциты располагаются по периферии тяжей и содержат в цитоплазме гранулы секрета. Гранулы интенсивно окрашиваются красителями и делятся на оксифильные и базофильные. Хромофильные аденоциты имеют хорошо развитые органеллы белкового синтеза (гранулярная ЭПС), секреции (комплекс Гольджи) и секреторные гранулы. В клетках вырабатывающих гормоны гликопротеидной природы, секреторные гранулы меньшего диаметра и менее развиты гранулярная ЭПС и комплекс Гольджи. Во всех аденоцитах развиты митохондриальный аппарат и компоненты цитоскелета. Каждый аденоцит тесно контактирует с гемокапилляром висцерального или синусоидного типа. Оксифильные эндокриноциты (аденоциты) – клетки округлой или овальной формы, с оксифильными гранулами, хорошо развитой гранулярной ЭПС и комплексом Гольджи. Делятся на две группы: соматотропоциты (3) – вырабатывают гормон роста стимулирующий деление клеток в организме и его рост. Лактотропоциты (4) вырабатывают лактотропный гормон (синонимы пролактин, маммотропин). Этот гормон усиливает рост молочных желез и секрецию молока во время беременности, а также способствует образованию в яичнике желтого тела. Базофильные эндокриноциты (аденоциты) содержат в цитоплазме базофильные гранулы. Подразделяются на два вида: тиротропоциты и гонадотропоциты. Гормоны базофильных аденоцитов имеют гликопротеиновую природу. Тиротропоциты (2) – клетки треугольной формы с большим количеством мелких базофильных гранул. Они вырабатывают тиротропный гормон. Этот гормон стимулирует выработку щитовидной железой тиреоидных гормонов. Гонадотропоциты (6) имеют круглую форму и лежащее эксцентрично ядро. В цитоплазме базофильные гранулы отсутствуют в области светлого участка вблизи ядра. Это место локализации аппарата Гольджи. Гонадотропоциты подразделяются на два вида: фоллитропоциты вырабатывают гормон фоллитропин (ФСГ), стимулирующий процессы овогенеза и синтез эстрогенов в женском организме. В мужском организме ФСГ активирует сперматогенез. Лютропоциты вырабатывают гормон лютропин, который в женском организме стимулирует развитие желтого тела и секрецию им прогестерона. В мужском организме ЛГ стимулирует выработку мужских половых гормонов клетками Лейдига. Есть клетки, которые занимают промежуточное положение между базофилами и оксифилами- кортикотропоциты (7) (адренокортикоциты). Они расположены в центре передней доли гипофиза. Гранулы клеток имеют вид пузырьков, содержащих в центре плотную часть. Клетки вырабатывают адренокортикотропный гормон, стимулирующий секрецию гормонов пучковой и сетчатой зонами коры надпочечников. Хромофобные (1) клетки расположены в центре трабекул. Их цитоплазма плохо воспринимает красители. К ним относят: 1.Незрелые, малодифференцированные клетки (выполняют регенераторную функцию). 2.Выделившие секрет и потому не окрашивающиеся в данный момент хромофильные клетки. 3.Фолликулярно-звездчатые клетки небольших размеров, имеющие длинные отростки, при помощи которых они соединяются друг с другом. Группа клеток, которые апикальными поверхностями обращены друг к другу, образует псевдофолликулы заполненные коллоидом. 54 Рис. 2. Фрагмент фолликула щитовидной железы. 1.Коллоид. 2.Тироцит. 3.С-клетка. 4.Микроворсинки тироцитов. 5.Ядро тироцита. 6.Ядро С-клетки. 7.Кровеносный капилляр. 8.Аксовазальный синапс. Щитовидная железа - паренхиматозный орган. Доли щитовидной железы покрыты капсулой из плотной волокнистой соединительной ткани, от которой внутрь отходят трабекулы с сосудами и нервами. Паренхиму образуют эпителиальные клетки, которые образуют шары – фолликулы. Фолликул является структурно-функциональной единицей щитовидной железы. Стенка фолликула образована двумя видами клеток, основными являются тироциты (2), которые лежат на базальной мембране. Тироциты имеют ядро (5), хорошо развитые органеллы белкового синтеза: гранулярная ЭПС, аппарат Гольджи, митохондрии. Апикальная поверхность тироцитов имеет микроворсинки (4), базальная – складки цитолеммы. В зависимости от функции форма тироцитов изменяется. В норме она кубическая, при гиперфункции – цилиндрическая, а при гипофункции - уплощенная. В просвете фолликула – коллоид (1), который является продуктом секреции тироцитов. Тироциты синтезируют и выделяют гормоны: трийодтиронин и тетрайодтиронин. Гормоны связаны с белками и накапливаются в виде коллоида. Латеральные поверхности тироцитов соединены комплексом межклеточных контактов. Исходные вещества для образования гормонов тироциты получают из крови: аминокислоты, основной из которых является тирозин, и ионы йода. Аминокислоты накапливаются в гр. ЭПС, где из них синтезируется полипептидная цепь, которая далее поступает в комплекс Гольджи. Здесь к ней присоединяются углеводные компоненты и образуется тироглобулин. Секреторные пузырьки с тироглобулином отделяются от комплекса Гольджи и поступают в полость фолликула. Ионы йода в клетке преобразуются в атомарный йод. Далее происходит процесс присоединения атомов йода к аминокислоте тирозину (находится в составе тироглобулина) с образованием трийодтиронина и тетрайодтиронина. Присоединение йода к тироглобулину происходит на поверхности тироцита. Образовавшиеся гормоны депонируются в полости фолликула в виде коллоида. При необходимости тироцит с помощью микроворсинок (4) захватывает коллоид. Ферменты лизосом отщепляют от тироглобулина молекулы тиреоидных гормонов, которые из тироцита через базальную мембрану поступают в кровь (7). Таким образом, выделяют три фазы секреторного цикла: 1.Биосинтез тироглобулина 2.Выделение тироглобулина в полость фолликула. Йодирование органической основы гормонов и депонирование тироглобулина в фолликуле. При необходимости тиреоглобулин захватывается микроворсинками и поступает в клетку. 3.Выведение гормонов из клетки в кровь. Гормоны щитовидной железы – тетрайодтиронин (тироксин) и трийодтиронин регулируют основной обмен, а также процессы развития, роста и дифференцировки тканей. Мишенями тиреоидных гормонов являются практически все клетки организма. Гормоны проникают в клетку, затем в ядро и воздействуют на генетический аппарат. Важную роль гормоны играют в дифференцировке нервной системы в период внутриутробного развития. С – клетки (3) составляют около 0,1% от общего числа паренхиматозных клеток. Располагаются отдельно или группами в стенке фолликула между тироцитами и базальной мембраной. Вырабатывают гормон кальцитонин, который накапливается в плотных гранулах и путем экзоцитоза выводится из клетки. Кальцитонин понижает уровень кальция в крови, поскольку стимулирует клетки костной ткани - остеобласты. При этом кальций откладывается в костях. Кальцитонин стимулирует экскрецию кальция почками. Связываясь с рецепторами на цитолемме остеокластов, ингибирует резорбцию кости. 55 Рис. 3. Пучковая зона коры надпочечников. 1.Светлый кортикоцит с липидными включениями. 2.Темный кортикоцит. 3.Ядро. 4.Митохондрии с везикулярными кристами. 5.Коплекс Гольджи 6.Гранулярная ЭПС. 7.Кровеносный капилляр. Наиболее выраженная зона коры (занимает 75% её объема). Образована идущими параллельно друг другу (в виде пучков) тяжами эндокринных клеток (1, 2), с оксифильной цитоплазмой, крупными размерами. Между тяжами в прослойках рыхлой соединительной ткани лежат синусойдные гемокапилляры (7). Различают два вида эндокриноцитов (кортикоцитов) этой зоны: темные (2) и светлые (1). Это один тип клеток, находящихся в разных функциональных состояниях. Цитоплазма клеток пучковой зоны выглядит вакуолизированной, что связано с большим содержанием липидных включений с холестерином, вымывающихся при обработке препаратов. Поэтому клетки имеют вид губки, и их называют «спонгиоциты». Для эндокриноцитов пучковой зоны характерно наличие митохондрий (4) особого строения: они имеют крупные и гигантские размеры, сильно развиты кристы, в виде извитых и ветвящихся трубок (везикулярные кристы). Митохондрии участвуют в одном из этапов биосинтеза гормонов. Развиты свободные рибосомы, комплекс Гольджи (5) и особенно гладкая ЭПС (основное место биосинтеза стероидных гормонов). Темные эндокриноциты имеют уплотненную цитоплазму, содержат мало липидных включений, но много рибонуклеопротеидов. В клетках выявляется аскорбиновая кислота, необходимая для биосинтеза стероидных гормонов. Полагают, что в темных клетках осуществляется синтез специфических белков-ферментов, которые в дальнейшем участвуют в образовании кортикостероидов, о чем свидетельствует обильное содержание в цитоплазме темных клеток рибосом. По мере выработки стероидов и их накопления, цитоплазма становится светлой. В пучковой зоне коры надпочечников вырабатываются глюкокортикоидные гормоны, в основном кортизол и кортизон. Эти гормоны влияют на метаболизм углеводов, белков и липидов, в частности, повышают уровень глюкозы в крови, за счет синтеза её из продуктов распада жиров и белков. Большие дозы глюкокортикоидов вызывают гибель лимфоцитов и эозинофилов. Синтез и секреция глюкокортикоидов резко возрастает при стрессе, этот процесс активируется гормонами гипофиза. 56 К какому органу относятся клетки? Аргументируйте выводы. Назовите структуры обозначенные цифрами? Рис. 4. Хромаффинные клетки мозгового вещества надпочечников. 1.Светлые клетки. 2.Темные клетки. 3.Гемокапилляр. 4.Гранулы секрета темных клеток. 5.Гранулы секрета светлых клеток. Мозговое вещество надпочечников продуцирует катехоламины – гормон адреналин и нейромедиатор норадреналин, которые вырабатываются при стрессе и мобилизуют все ресурсы организма в сложных ситуациях для сохранения работоспособности, а иногда и жизни. Хромаффиноциты хорошо окрашиваются солями хрома. Эти клетки делятся на два вида: 1) крупные светлые клетки – синтезируют гормон адреналин ( А-клетки) (1), содержат в цитоплазме умеренно электроноплотные гранулы (5); 2) темные мелкие хромаффиноциты (НА – клетки) (2), содержащие большое число плотных гранул, окруженных по периферии светлым ободком (4), они секретируют норадреналин. А и НА клетки на обычных срезах идентифицировать трудно, они хорошо различаются только при обработке солями хрома. Кроме катехоламинов секреторнве гранулы хромаффиноцитов содержат белки и полипептиды: хромогранины, энкефалины, иммуномодуляторы, а также АТФ и липиды. Как полагают, хромогранины обеспечивают осмотическую стабилизацию секреторных гранул клеток. 57 ГЛАВА 7. ОРГАНЫ ЧУВСТВ Фрагмент какой оболочки глазного яблока изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 1. Нейронный состав и глиоциты сетчатки. I – фоторецепторные нейроны. II – ассоциативные нейроны III – ганглионарные нейроны. 1. Колбочковый нейрон. 2.Палочковый нейрон. 3.Горизонтальный нейрон. 4.Амакринный нейрон. 5.Глиоцит (клетка Мюллера). 6.Внутренняя глиальная пограничная мембрана. 7.Отростки пигментоцитов. 8.Зрительный нерв. В сетчатой оболочке различают наружный пигментный слой и внутренний светочувствительный. Сетчатая оболочка состоит из трех типов нейронов: 1-фоторецепторы; 2-ассоциативны нейроны; 3-ганглионарные нейроны. Фоторецепторные нейроны (I). Для образования потенциала действия световой луч должен, пройдя через толщу сетчатой оболочки, достигнуть наружных сегментов палочек и колбочек (I., 1., 2.), где и происходит образование потенциала действия. Потенциал действия следует в противоположном направлении. Передача осуществляется через синапсы между фоторецепторными (I) и ассоциативными нейронами (II). Ассоциативные нейроны (II). К ним относят биполярные нейроны, которые своими дендритами образуют синапсы с аксонами фоторецепторных нейронов. Горизонтальные нейроны (3) располагаются в один или два ряда. Соседние клетки связаны щелевидными контактами. Их отростки обьединяют между собой фоторецепторы и задерживают сигнал в слое палочек и колбочек, обеспечивая время для процессов аккомодации. Амакринные нейроны (4) связывают аксоны биполярных нейронов друг с другом, что повышает контрастность изображения. Ганглионарные нейроны (III). Это крупные по размерам клетки. Имеют большой диаметр аксонов. По морфологическим и функциональным свойствам выделяют 18 типов. Ассоциативные нейроны образуют синапсы с ганглионарными нейронами (III). Миелинизированные аксоны последних составляют зрительный нерв (8). Его волокна образуют перекрест, а далее в составе зрительного тракта перекрещенные и неперекрещенные нервные волокна достигают латеральных коленчатых тел или верхних бугорков четверохолмия среднего мозга (подкорковые зрительные центры). От латеральных коленчатых тел потенциал действия передается в корковый конец зрительного анализатора. Таким образом, световой луч и потенциал действия следуют в противоположных направлениях. В составе нейроглии клетки Мюллера (5), которые проходят через всю толщу сетчатки, принимают участие в образовании наружной глиальной пограничной мембраны. Внутренняя пограничная мембрана образована окончаниями отростков клеток Мюллера и их базальными мембранами. Клетки Мюллера регулируют ионный гомеостаз, выполняют опорную функцию. Астроциты принимают участие в образовании гематоретинального барьера за счет того, что их отростки охватывают капилляры. Клетки микроглии находятся во всех слоях сетчатки и выполняют фагоцитарную функцию. Пигментоциты, отростки которых изображены на рисунке (7), располагаются кнаружи от сетчатой оболочки (пигментная часть сетчатой оболочки). Их основания расположены на базальной мембране входящей в состав сосудистой оболочки. На апикальной поверхности имеются отростки, которые контактируют с наружными сегментами фоторецепторных нейронов (I). Наличие меланосом (пигментных гранул) обуславливает поглощение 90% света попадающего в глаз. На свету меланосомы перемещаются в отростки. В темноте меланосомы из отростков перемещаются в тело пигментной клетки. Пигментоциты осуществляют транспорт метаболитов, солей, кислорода и др. из сосудистой оболочки к фоторецепторам; фагоцитарная функция ( убирают отработанные диски наружных сегментов палочек), регулируют ионный гомеостаз. 58 Назовите тип клетки. Аргументируйте вывод. Назовите структуры, обозначенные цифрами. Рис. 2. Палочковая (нейросенсорная) клетка. I –наружный дистальный сегмент. II – связующий сегмент (ресничка). III – внутренний проксимальный сегмент. IV – тело клетки. V – аксон. 1.Замкнутые мембранные диски. 2.Цитоплазма. 3.Микротрубочки. 4.Митохондрии. 5.ЭПС. 6.Ядро. 7.Синапс. Палочковая клетка-рецептор ночного зрения. Это длинная клетка, цилиндрической формы.Наружный сегмент (I) цилиндрической формы, и представлен стопкой пластинок (дисков), которые отделены от плазматической мембраны. В мембранных дисках (1) находится зрительный пигмент родопсин. Он разлагается под влиянием света, вызывая изменение ионной проницаемости мембран и возникновения потенциала действия. Ресинтез родопсина происходит в темноте при наличии витамина А с участием пигментных клеток. Диски в проксимальном отделе постоянно обновляются и смещаются в дистальные отделы, где фагоцитируются пигментными клетками. Наружный (I) и внутренний (II) сегменты палочек одинаковы по диаметру. Во внутреннем сегменте много митохондрий (4), развита гранулярная и агранулярная ЭПС (5). Здесь осуществляется синтез белка. Тело клетки (IV) расположено проксимальнее внутреннего сегмента и переходит в отросток – аксон (V), который образует синапс (7) с дендритом биполярного нейрона. 59 Назовите тип клетки. Аргументируйте вывод. Назовите структуры, обозначенные цифрами. Рис. 3. Колбочковая (нейросенсорная) клетка. I –наружный дистальный сегмент. II – связующий сегмент (ресничка). III – внутренний проксимальный сегмент. IV – тело клетки. V – аксон. 1.Незамкнутые мембранные диски. 2.Цитоплазма. 3.Микротрубочки. 4.Липидная капля. 5.Митохондрии. 6.ЭПС. 7.Ядро. 8.Синапс. Колбочковая клетка обеспечивает дневное и цветное зрение. Ее наружный сегмент (I) конической формы. В наружном сегменте диски не замкнуты и внутридисковое пространство сообщается с внеклеточной средой. Диски (1) образованы складками цитолеммы и содержат зрительный пигмент йодопсин. Существует три функциональных типа колбочек, каждый из которых содержит только один из зрительных пигментов (красный, синий, зеленый) и состоит из опсина, ковалентно связанного с хромофором. Их различия определяются структурой опсиновой молекулы с максимальной чувствительностью в длинноволновой, средневолновой и коротковолновой части спектра. Отсутствие колбочек определенных функциональных типов обусловлено изменениями гена в Х-хромосоме и вызывает цветовую слепоту (дальтонизм). Внутренний сегмент (III) по диаметру больше наружного. Во внутреннем сегменте имеется эллипсоид, который состоит из липидного тела, функция которого заключается в расщеплении луча света на составные части спектра, вокруг скопление митохондрий. Тело клетки (IV) расположено проксимальнее внутреннего сегмента и переходит в отросток – аксон (V). Аксоны образуют синапсы (8) с дендритами биполярных нейронов. Регенерация палочек и колбочек. У основания наружных сегментов палочек и колбочек разрастается цитолемма, которая впячивается внутрь наружного сегмента, образуя диски и полудиски. Старые диски фагоцитируют пигментные клетки. 60 Фрагмент какого органа изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 4. Обонятельный эпителий полости носа. 1.Обонятельная нейросенсорная клетка. 2.Дендрит. 3.Луковица дендрита. 4.Ресничка. 5.Аксон. 6.Поддерживающий эпителиоцит. 7.Базальный эпителиоцит. 8.Кровеносный капилляр. 9.Базальная мембрана. 10.Фрагмент концевого отдела железы. Полость носа относят к внелегочным воздухоносным путям, где воздух очищается, увлажняется, согревается. Кроме того, в верхнем носовом ходе слизистая оболочка специализирована на выполнение обонятельной функции. Обонятельный эпителий верхнего носового хода является многорядным. Здесь находятся нейросенсорные рецепторные клетки (1). Их периферический отросток дендрит (2) заканчивается на поверхности обонятельного эпителия утолщением – обонятельной булавой (3) или луковицей на поверхности которой подвижные реснички (4) с находящимися на их цитолемме рецепторными белками. При оседании пахучих веществ (молекул) возникает потенциал действия. В теле нейросенсорной клетки много митохондрий, цистерны ЭПС, комплекс Гольджи, лизосомы. Аксон (5) покидает полость носа через lamina cribrosa и следует к обонятельной луковице. Обонятельных клеток насчитывается порядка 6 млн. Они обновляются каждые 25-30 дней. Их функция заключается в регистрации первичных запахов (образуют потенциал действия). Поддерживающие эпителиоциты (6) – расположены на достаточно толстой базальной мембране (9). Имеют призматическую форму с ядром почти в центре клетки и органеллами общего назначения. Располагаются эти клетки между нейросенсорными клетками изолируя и поддерживая их. Выделяют жидкий секрет где растворяются пахучие вещества. Базальные эпителиоциты (7) имеют треугольную форму. Их основание располагается на базальной мембране. Это малодифференцированные клетки за счет которых обновляются нейросенсорные клетки. Под базальной мембраной расположена рыхлая волокнистая соединительная ткань с находящимися обонятельными железами (10), имеющими трубчатое строение, в жидком секрете которых и растворяются пахучие вещества. Подэпителиальный слой богато васкуляризирован (8), что способствует согреванию воздуха в полости носа. При изучении окрашенных микропрепаратов по расположению ядер можно предположить к какому типу клеток они относятся, в частности, ядра базальных эпителиоцитов расположены вблизи базальной мембраны, ядра нейросенсорных клеток располагаются центрально (в многорядном эпителии), а ядра поддерживающих эпителиоцитов - ближе к апикальной поверхности клеток. Механизм обоняния: на цитолемме подвижных ресничек булавы имеются рецепторные белки, которые захватывают молекулы пахучих веществ. Это приводит к изменению проницаемости цитолеммы и возникновению импульса. Белки чувствительны к молекулам специфических ароматических групп. При их связывании с рецептором (нейросенсорной клеткой), рецептор может являться одновременно и ионным каналом, который открывается под влиянием лиганда (молекулы, специфически связывающейся с рецептором). Либо под влиянием лиганда рецептор активирует аденилатциклазу, вызывая образование цАМФ, которая способствует открытию ионного канала, что и в том и в другом случаях приводит к поступлению ионов в нейросенсорную клетку. Происходит деполяризация ее плазмолеммы и возбуждение, которое по аксону достигает обонятельной луковицы с находящимися в них митральными клетками, аксоны которых следуют в древнюю кору (гиппокамп) и гиппокампову извилину, которая является корковым центром обонятельного анализатора. 61 Фрагмент какого органа изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 5. Фрагмент спирального (Кортиева) органа. 1.Фаланговый эпителиоцит. 2.Фаланга. 3.Наружный волосковый сенсорный эпителиоцит. 4.Стереоцилии. 5.Нервное волокно. 6.Афферентное нервное окончание. 7.Эфферентное нервное окончание. 8.Покровная мембрана. Спиральный орган расположен на базилярной пластинке в перепончатом лабиринте улитки. Образован рецепторными клетками и поддерживающими (опорными) клетками, которые подразделяют на наружные и внутренние. На рисунке изображен фрагмент Кортиева органа с рецепторными и поддерживающими клетками. Наружный волосковый сенсорный эпителиоцит (3) имеет цилиндрическую форму. Базальная поверхность располагается в чашеобразных углублениях наружных фаланговых эпителиоцитов (1). На апикальной поверхности стереоцилии (4), которые содержат плотно упакованные фибриллы, в составе которых актин и миозин. Поэтому стереоцилии выпрямляются после соприкосновения с покровной мембраной. На цитолемме стереоцилий имеются холинорецепторные белки и фермент ацетилхолинэстераза. Стереоцилии погружены в желеобразную покровную мембрану (8). Покровная мембрана нависает над всем спиральным органом. Представляет соединительнотканную пластинку, состоящую из радиально направленных коллагеновых волокон, погруженных в аморфный матрикс. Ее внутренний край прикреплен к спиральному гребешку, а наружный свободно нависает над клетками. Наружные фаланговые эпителиоциты (1) имеют призматическую форму. Ядро в базальной части клетки. На апикальной поверхности чашеобразное углубление, где находится базальная часть рецепторных клеток. Длинный отросток – фаланга (2) с находящимися там микротрубочками прилежит к апикальной части рецепторных клеток. В цитоплазму фаланговых клеток внедряются нервные волокна (5), которые образуют нервные окончания на базальной части волосковых сенсорных эпителиоцитов (6, 7). Звуковые волны, доходя до внутреннего уха, приводят в движение перилимфу барабанной лестницы, что вызывает перемещение базилярной мембраны. Во время колебательных движений холинорецепторы стереоцилий захватывают ацетилхолин, который находится в эндолимфе. Это приводит к изменению проницаемости цитолеммы, возникает потенциал действия. 62 Назовите фрагмент органа, типы клеток, структуры, обозначенные цифрами. Рис. 6. Фрагмент макулы мешочков (орган равновесия). 1.Грушевидный эпителиоцит (рецепторная клетка I типа). 2.Столбчатый эпителиоцит (рецепторная клетка II типа). 3.Киноцилия. 4.Стереоцилия. 5.Отолитовая мембрана с отолитами. 6.Нервные окончания. 7.Нервные волокна. 8.Поддерживающий эпителиоцит. 9.Соединительная ткань. Рецепторные клетки в маточке и мешочке расположены не повсеместно, а образуют скопления, которые называют пятна (макулы). Они выстланы эпителием, располагающимся на базальной мембране. Среди клеток выделяют рецепторные и опорные. Грушевидный эпителиоцит (1) имеет расширенную базальную часть к которой прилежат нервные окончания (6) в виде чаши. Столбчатые эпителиоциты (2) имеют призматическую форму. К их базальной части так же прилежат нервные окончания (6). На апикальной поверхности клеток неподвижные стереоцилии (4) и одна подвижная киноцилия (3). Поддерживающие эпителиоциты (8) имеют призматическую форму с многочисленными микроворсинками на апикальной поверхности. Отолитовая мембрана (5) представляет собой студенистое вещество (гликопротеины) покрывающее макулы. В нее погружены стереоцилии и киноцилии. На ее поверхности располагаются кристаллы карбоната кальция – отолиты. При потере равновесия смещается отолитовая мембрана и соответственно изменяется положение стереоцилий и киноцилий: при смещении стереоцилий в сторону киноцилии клетка возбуждается, а противоположный эффект вызывает торможение. 63 Рис. 7. Вкусовая почка. 1.Опорная (поддерживающая) «темная» клетка. 2. Опорная (поддерживающая) «светлая» клетка. 3.Сенсоэпителиальная (рецепторная) клетка. 4.Базальная малодифференцированная клетка. 5.Нервное волокно. 6.Вкусовая пора. 7. Периферические (перигеммальные) клетки. 8.Собственная пластинка слизистой оболочки языка. Вкусовые почки располагаются в многослойном эпителии сосочков языка, за исключением нитевидных сосочков. Вкусовая почка имеет эллипсоидную форму. Состоит из плотно прилежащих друг к другу клеток, количество которых колеблется от 40 до 60. Воспринимает вкусовые раздражения пищевые и непищевые. Источником развития клеток во вкусовых почках является многослойный эпителий языка, где происходит их дифференцировка под индуцирующим влиянием врастающих нервов (7, 9, 10 пар черепно-мозговых нервов). Среди них различают сенсоэпителиальныне клетки (3), располагающиеся на базальной мембране. Апикальная поверхность имеет микроворсинки между которыми электронноплотное вещество с высокой активностью фосфатаз, большим содержанием мукопротеидов, которые играют роль адсорбента, с встроенными в цитолемму рецепторными белками. Конформационные (пространственные) изменения рецепторных белков приводят к изменению проницаемости мембран сенсоэпителиальных клеток и генерации потенциалов действия, которые с сенсоэпителиальной клетки переходят на нервное окончание, контактирующее с базальным полюсом клетки. В базальной части располагается овальной формы ядро, и эта часть клетки образует синапсы с чувствительными нервными волокнами. Поддерживающие клетки имеют вытянутую форму с овальным ядром в центральной части клетки, комплекс Гольджи, гр.и агр. ЭПС, митохондрии. Они отделяют сенсоэпителиальные клетки друг от друга. Базальные малодифференцированные клетки (4) конической формы, располагаются на базальной мембране. Имеют небольшой обьем цитоплазмы вокруг ядра. Слабо развиты органеллы. Эти клетки способны к митотическому делению и служат источником обновления сенсоэпителиальных и поддерживающих клеток. Периферические (перигеммальные) клетки (7) располагаются на поверхности вкусовой луковицы. Имеют серповидную форму. Вероятно, они выполняют разграничительную функцию, отделяя вкусовую почку от эпителия сосочка. Орган вкуса у новорожденных хорошо развит. Количество вкусовых почек в грудном возрасте больше, чем у взрослого. С возрастом темпы обновления клеток замедляются, количество вкусовых почек уменьшается, появляются нарушения вкусовой чувствительности. Это может быть связано с дистрофическими изменениями в луковицах, разрушении рецепторов языка. В старческом возрасте снижается в большей степени чувствительность на сладкое, а наиболее устойчивым является восприятие соленого. Механизм вкусовой рецепции: при попадании пищи в ротовую полость, молекулы пищевых веществ захватываются рецепторными белками, каждый из которых воспринимает определенный вкус, изменяется потенциал сенсоэпителиальных клеток (изменяется проницаемость цитолеммы, т.е. изменяется состояние ионных каналов), меняется концентрация медиаторов внутри клетки. Через синапс потенциал действия передается на дендриты нейронов, тела которых находятся в ганглиях лицевого, блуждающего, языкоглоточного нервов – это 1 нейроны. Далее их аксоны следуют к дендритам 2-ых нейронов, клеточные тела которых заложены в ядре одиночного пути, а аксоны вторых нейронов несут потенциал действия к слюнным железам, мышцам языка, мимическим мышцам. Часть аксонов вторых нейронов направляется к подкорковому чувствительному центру – таламусу, где находятся 3 нейроны, аксоны которых достигают постцентральной извилины коры головного мозга (корковый конец вкусового анализатора). 64 ГЛАВА 8. ОРГАНЫ ПОЛОВОЙ СИСТЕМЫ Рис. 1. Извитой каналец яичка. А. Сперматогенный эпителий: Сустентоцит (клетка Сетоли): 1 – базальная мембрана, 2 – основание клетки, 3 - апикальная поверхность, 4 – ядро, 5 – ядрышко, 6 – межклеточные контакты; Предшественники сперматозоидов на разных стадия дифференцировки: 7 – светлая сперматогония типа А, 8 – тёмная сперматогония типа А, 9 – сперматогонии типа В, 10 – сперматоциты 1-го порядка, 11 – цитоплазматические мостики, 12 – сперматоцит 2-го порядка, 13 – ранние сперматиды, 14 – поздние сперматиды; Б - соединительнотканная оболочка канальца, В – слой миоидных клеток, Г – просвет канальца. В извитом канальце яичка происходит сперматогенез, или образование мужских половых клеток – сперматозоидов. Почти всю толщу стенки извитого канальца яичка составляет эпителий, окружённый базальной мембраной (1), который называется сперматогенный и включает два типа клеток: сустентоциты (клетки Сертоли) и предшественники сперматозоидов на разных стадиях дифференцировки. Сустентоциты, или поддерживающие клетки, основанием (2) лежат на базальной мембране, апикальной частью (3) обращены в просвет канальца (Г). Ядро клетки (4) чаще неправильной формы, располагается в базальной части, оно крупное, светлое, имеется ядрышко (5). Вдоль латеральных поверхностей в цитоплазматических карманах располагаются развивающиеся сперматозоиды, которые, созревая, перемещаются от базальной мембраны к просвету канальца. Сустентоциты формируют друг с другом специализированные межклеточные контакты (6), разделяющие сперматогенный эпителий на базальное и адлюминальное пространство. Клетки Сертоли, как и фолликулярные клетки яичника, выполняют барьерную роль между половыми клетками и внутренней средой организма. Принимают участие в образовании гематотестикулярного барьера. Вырабатывают жидкий секрет, который заполняет просвет извитых семенных канальцев. В базальном отделе эпителия, находясь на базальной мембране, располагаются сперматогонии. Их несколько типов: сперматогонии типа А по степени конденсации хроматина делят на светлые (7), находящиеся в непрерывно следующих друг за другом клеточных циклах, и тёмные (8), или резервные, редко вступающие в митоз. Сперматогонии типа В (9), образующиеся из светлых сперматогоний А, имеют округлые ядра и конденсированный хроматин, делятся митозом (стадия размножения сперматогенеза). Их дочерние клетки – сперматоциты 1-го порядка (10), связанные друг с другом цитоплазматическими мостиками (11), образуя синцитий, увеличиваются в размерах, проходя стадию роста, прекращают делиться митозом и вступают в мейоз, при этом перемещаются из базального в адлюминальное пространство канальца. Сперматоциты 2го порядка (12), образующиеся в стадию созревания в результате 1-го деления мейоза, меньше по размерам своих предшественников, располагаются над сперматоцитами 1-го порядка. У просвета канальца располагаются образованные в результате 2-го деления мейоза гаплоидные сперматиды: сначала в виде округлых клеток – ранние (13), на этой стадии разрываются цитоплазматические мостики между клетками, затем происходит формирование сперматозоида – поздняя сперматида (14). Помимо эпителия, в состав извитого канальца входит соединительнотканная оболочка (Б), представленная плотной волокнистой тканью, выполняющей защитную функцию. Снаружи от волокнистого слоя находятся миоидные клетки (В). Их сокращения приводят к продвижению сперматозоидов из извитых канальцев в сеть яичка. Также миоидные клетки активируют выработку андроген-связывающего белка клетками Сертоли. 65 Рис. 2. Эндокриноцит яичка (клетка Лейдига) и миоидная клетка. 1.Клетки Лейдига. 2. Интерстиций яичка. 3.Кровеносный капилляр. 4. Сперматогенный эпителий. 5.Базальная мембрана сперматогенного эпителия. 6. Волокнистый слой извитого канальца. 7.Миоидные клетки. 8.Базальная мембрана миоидных клеток. Клетка Лейдига: 9 – ядро клетки Лейдига, в нём а) – ядрышко, 10 – митохондрии везикулярного типа, 11 – агр. ЭПС, 12 – гр. ЭПС, 13 – лизосомы, 14 – липидные включения, 15 – кристаллы Рейнке; Миоидная клетка: 16 – ядро, 17 – миофибриллы, 18 – десмосомы, 19 – щелевидные соединения. Клетки Лейдига (1) располагаются в интерстиции яичка (2) поодиночке или группами около кровеносных капилляров (3). На рисунке, помимо интерстиция, представлен фрагмент стенки извитого канальца: базальные части клеток сперматогенного эпителия (4), расположенные на базальной мембране (5), волокнистый слой канальца (6) и миоидные клетки (7), окружённые базальной мембраной (8). Клетка Лейдига достаточно крупная по размерам, имеет округлую форму, светлое ядро (9), с одним ядрышком (а), оксифильную цитоплазму. Главный секреторный продукт - тестостерон, образуется из холестерина ферментными системами агр. ЭПС и митохондрий. Поэтому в цитоплазме содержится большое количество удлинённых митохондрий везикулярного типа (10), хорошо развита агр. ЭПС (11) и гр. ЭПС (12), много лизосом (13), липидных включений (14). Особенностью в строении клеток является наличие кристаллов Рейнке (15) – белковых включений правильной вытянутой геометрической формы с острыми углами, состоящие из филаментов. Функция кристаллов до конца не выяснена. Продуцируемые клетками Лейдига андрогены (тестостерон, дигидротестостерон, андростендион) необходимы для нормального сперматогенеза, они регулируют развитие и функцию добавочных желёз половой системы (предстательной, бульбоуретральных, семенных пузырьков), обеспечивают развитие вторичных половых признаков. Так, тестостерон стимулирует выработку андрогенсвязывающего белка (АСБ) в клетках Сертоли, поддерживающего высокие концентрации тестостерона в извитых канальцах. Тестостерон влияет на репрессию генов апоптоза в развивающихся половых клетках, напрямую стимулирует трансформацию сперматид в сперматозоиды. Образование тестостерона, как и сперматогенез, у мужчин происходит постоянно, начиная с постпубертатного периода, и не имеет цикличности, как у женщин. Это связано с отсутствием положительной обратной связи между количеством андрогенов и синтезом гонадотропинов. Активность клеток Лейдига регулируется ЛГ. Усиление выработки тестостерона через механизмы отрицательной обратной связи угнетает секрецию ЛГ передней долей гипофиза и гонадолиберина гипоталамусом. Помимо половых гормонов, клетки Лейдига вырабатывают некоторые производные проопиомеланокортина, оказывающие паракринное действие на клетки Сертоли; небольшие количества окситоцина, стимулирующего сокращения перитубулярных миоидных клеток (16), а также интерлейкин-1, действующего как фактор роста на сперматогонии типа В. В миоидных клетках (миофибробластах) ядра вытянутые, палочковидные (16), в цитоплазме находятся помимо органелл общего значения специализированные - миофибриллы (17), с участием которых происходят сокращения клеток, необходимые для проталкивания спермиев из извитого канальца в сеть яичка. Межклеточные соединения в миофибробластах представлены десмосомами (18) и щелевидными контактами (19). Миоидные клетки под действием тестостерона вырабатывают белковый паракринный фактор P-Mod-S (в переводе - белок, модулирующий сустентоциты), активирующий синтетическую деятельность сустентоцитов и повышающий их чувствительность к тестостерону. 66 Дайте характеристику клеток в соответствии со стадиями изображённого процесса. Назовите структуры, обозначенные цифрами. Рис. 3. Сперматогенный эпителий. Сустентоциты (Клетки Сертоли). 1. Базальная мембрана. 2. Апикальная поверхность сустентоцита. 3.Просвет канальца. 4.Ядро. 5.Ядрышко. 6.грЭПС. 7. Митохондрии. 8.Лизосомы. 9. Кристаллы (включения). 10.Контакты соседних клеток. Предшественники сперматозоидов на разных стадиях дифференциации. В базальном отделе эпителия: 11 – сперматогония А светлая, 12 – сперматогония А тёмная, 13 – сперматогония В; в адлюминальном отделе эпителия: 14 – сперматоциты 1-го порядка, 15 – цитоплазматические мостики между клетками, 16 – сперматоцит 2-го порядка, 17 – сперматида ранняя, 18 – сперматида поздняя. Сперматогенный эпителий выстилает извитые канальцы яичка и включает два типа клеток: сустентоциты (клетки Сертоли) и сперматозоиды с предшественниками на разных стадиях дифференцировки. Сустентоциты, или поддерживающие клетки, основанием лежат на базальной мембране (1), апикальной частью (2) обращены в просвет канальца (3). Ядро клетки (4) чаще неправильной формы, располагается в базальной части, оно крупное, светлое, имеется ядрышко (5). В цитоплазме хорошо развиты органеллы: грЭПС (6), митохондрии (7), лизосомы (8). В базальной части находятся включения – кристаллы (9), состоящие из филаментов (функция неясна). Сустентоциты обеспечивают развитие сперматозоидов, предшественники которых охвачены отростками сустентоцитов, находясь на разных уровнях - этажах. Кроме того, сустентоциты формируют друг с другом плотные контакты (10), разделяющие сперматогенный эпителий на базальное и адлюминальное пространство. Базальный отдел (нижний этаж) содержит сперматогонии, лежащие на базальной мембране, которые делятся митозом и подобны соматическим клеткам. Базальный отдел обменивается различными веществами с интерстицием яичка. В адлюминальном (околопросветном) отделе вступившие в мейоз предшественники сперматозоидов изолированы от системы крови и лимфы. Клетки Сертоли являются ведущими в создании гематотестикулярного барьера, предотвращающего иммунную реакцию на развивающиеся сперматозоиды. В базальном отделе эпителия располагаются сперматогонии типа А. Их по степени конденсации хроматина делят на светлые (11), находящиеся в непрерывно следующих друг за другом клеточных циклах, и тёмные (12), или резервные, редко вступающие в митоз. Сперматогонии типа В (13), образующиеся из светлых сперматогоний типа А, имеют округлые ядра и конденсированный хроматин, делятся митозом, т.е. происходит стадия размножения сперматогенеза. Их дочерние клетки – сперматоциты 1-го порядка (14), связанные друг с другом цитоплазматическими мостиками (15), образуя синцитий, увеличиваются в размерах, проходя стадию роста, прекращают делиться митозом и вступают в мейоз, при этом перемещаются из базального в адлюминальное пространство канальца. Сперматоциты 2-го порядка (16), образующиеся в стадию созревания в результате 1-го деления мейоза, меньше по размерам своих предшественников, располагаются над сперматоцитами 1-го порядка. У просвета канальца располагаются образованные в результате 2-го деления мейоза гаплоидные сперматиды: сначала в виде округлых клеток – ранние (17), на этой стадии разрываются цитоплазматические мостики между клетками, затем происходит формирование сперматозоида – поздняя сперматида (18). Сперматогенез начинается в период полового созревания и продолжается в течение всей жизни, протекает при пониженной по сравнению с телом температуре (в мошонке). Под действием ФСГ сустентоциты синтезируют: АСБ (андроген-связывающий белок), создающий в канальцах высокие концентрации необходимого для сперматогенеза тестостерона; ингибин и активин, которые по принципу обратной связи ингибируют и активируют соответственно выработку ФСГ; в эмбриональном периоде предшественники клеток Сертоли продуцируют мюллеров-ингибирующий фактор (МИФ). 67 Опишите клетку. Назовите структуры, обозначенные цифрами. Рис. 4. Сперматозоид. 1.Головка. 2.Ядро. 3.Акросома. 4.Шейка. 5.Центриоль. 6.Аксонема. 7.Периферические дуплеты микротрубочек аксонемы. 8.Центральный дуплет микротрубочек аксонемы 9. Дистальный (концевой) отдел хвоста. 10.Проксимальный (главный) отдел хвоста. 11. Промежуточная часть хвоста.12.Фибриллы. 13. Митохондрии. Сперматозоид – образующаяся в извитом канальце яичка мужская половая клетка, максимально приспособлен для выполнения своей функции: содержит ядро, отсутствует цитоплазма, имеются только необходимые для передвижения и проникновения в яйцеклетку органеллы: акросома, митохондрии, центриоли и жгутик. В сперматозоиде различают головку, шейку и хвост. Головка (1) длиной до 6 мкм и по экватору шириной 4 мкм, содержит ядро (2) с конденсированным хроматином и акросому (3), которая образуется из комплекса Гольджи и является специализированной лизосомой. В шейке (4) расположены центриоли (5). Хвост – самая длинная часть клетки до 50 мкм, в нём выделяют три части: промежуточную (которую также называют телом сперматозоида), проксимальный (главный) и дистальный (концевой) отделы. В основе хвоста в виде оси располагается аксонема (6), представленная микротрубочками в соответствии с формулой 9х2+2, т.е. 9 дуплетов микротрубочек (7) по периферии и 1 дуплет в центре (8). В дистальном отделе хвоста (9) других структур нет, кроме аксонемы, распадающейся в конце на отдельные микротрубочки. В главном отделе (10) и промежуточной части (11) хвоста вокруг аксонемы располагается 9 продольно ориентированных плотных одиночных элементов цитоскелета – фибрилл (12), формируя волокнистый футляр осевой нити. В промежуточной части вокруг фибрилл, окружающих аксонему, находятся митохондрии (13), образующие митохондриальную спираль. Сперматогенез (образование сперматозоидов), происходящий в извитых канальцах яичка, длится 60-70 дней, затем происходит дозревание клеток в протоке придатка в течение 10-15 суток. В эякуляте содержится около 200 млн сперматозоидов, из которых только около 1% (несколько сотен) достигает маточной трубы и участвует в процессе оплодотворения. Сперматозоиды передвигаются из влагалища в маточную трубу со скоростью 2-4 мм/мин за 7-10 часов. Движение сперматозоидов в женских половых путях осуществляется в результате положительного рео- и хемотаксиса, а также происходящей активации сперматозоидов – капацитации. Капацитация – процесс приобретения спермиями оплодотворяющей способности под влиянием секреторных продуктов женского полового тракта, в результате чего с плазмолеммы сперматозоидов удаляется гликопротеиновый слой, блокирующий активные центры рецепторных молекул, узнающие поверхность яйцеклетки и взаимодействующие с рецепторами прозрачной зоны (ПЗ). У капацитированных сперматозоидов изменяются движения жгутика с регулярных волнообразных на хлыстоподобные, что способствует более эффективному их перемещению; увеличивается чувствительность к ионам кальция. Капацитированный сперматозоид взаимодействует с рецепторами ПЗ, под действием активирующих субстанций овоцита ионы Са++ поступают в сперматозоид, и начинается акросомальная реакция. Передняя часть мембраны акросомы и плазматическая мембрана сперматозоида сливаются и разрываются, из акросомы выделяются ферменты, лизирующие оболочки яйцеклетки: акрозин (разрушает ПЗ овоцита), пенетраза (диссоциирует клетки лучистого венца), гиалуронидаза (расщепляет гиалуроновую кислоту), кислая фосфатаза (разрушает фосфохолин при прохождении спермиев через плазмолемму овоцита) и др. Ядро спереди становится прикрытым только задней частью мембраны акросомы, где находятся рецепторы, удерживающие сперматозоид около овоцита. При оплодотворении в яйцеклетку проникает ядро и центриоли сперматозоида, хвостик остаётся снаружи и отпадает. Сперматозоиды человека находятся в жизнеспособном состоянии в женском репродуктивном тракте от 24 до 48 часов. 68 Назовите и опишите клетки. Проанализируйте значение комплекса данных клеток. Назовите обозначенные структуры. Рис. 5. Овоцит с лучистым венцом. 1.Ядро овоцита, в нём а) – ядрышко, б) – ядерная оболочка. 2. Желточные (вителлиновые) включения в цитоплазме. 3.ЭПС. 4.Митохондрии. 5.Кортикальные гранулы. 6. Перивителлиновое пространство. 7. Прозрачная зона. 8.Ядра фолликулярных клеток. 9. Отростки фолликулярных клеток. Выросты на поверхности овоцита. На данной схеме представлена часть третичного фолликула яичника, когда яйцеклетка, или овоцит, окружён двумя слоями фолликулярных клеток – лучистым венцом, находится в составе яйценосного бугорка в конце фолликулярной стадии овариального цикла. Овоцит – это крупная до 125-150 мкм в диаметре клетка, с округлым ядром (1); визуализируется ядрышко (а) и ядерная оболочка (б), в ядре преобладает эухроматин, т.е. в клетке происходят процессы синтеза, и овоцит находится на стадии диплотены профазы 1-го деления мейоза, когда блок мейоза ещё не снят. В цитоплазме овоцита происходит накопление необходимых для последующих процессов оплодотворения, дробления и формирования бластоциты цитоплазматических органелл и веществ: желточные (вителлиновые) гранулы (2), расположенные равномерно в небольшом количестве, т.к. яйцеклетка человека – олиго- и изолецитальная; множество рибосом (видимые как точки); канальцы ЭПС (3); большое количество митохондрий (4); накапливаются белки, участвующие в формировании структур веретена деления. В периферической зоне цитоплазмы располагаются кортикальные гранулы (5) диаметром 200-600 нм, содержащие ферменты, которые после проникновения сперматозоида выделяются из клетки в перивителлиновое пространство (6) между прозрачной зоной и оволеммой (плазмолеммой овоцита) и изменяют конфигурацию белков прозрачной зоны, в первую очередь рецепторов сперматозоидов, что предотвращает полиспермию. Поверхность овоцита неровная, образует выросты, контактирующие с отростками фолликулярных клеток (9). Овоцит окружён защитной прозрачной зоной (7), состоящей из гликозаминогликанов. Через поры прозрачной зоны (ПЗ) могут проникать микромолекулы. Белковый состав ПЗ представлен гликопротеинами, из которых важное значение имеют белки - рецепторы для сперматозоидов. Фолликулоциты, как эпителиальные клетки, прилежат плотно друг к другу, но в лучистом венце находятся без базальной мембраны. Ядра клеток (8) округлые, с гетерохроматином по периферии. В цитоплазме хорошо развиты органеллы синтеза. Отростки фолликулярных клеток (9) пронизывают прозрачную зону, контактируя с выростами оволеммы, обеспечивая регуляцию мейоза, питание половой клетки. Третичный фолликул яичника представляет собой пузырёк до 2,5 см в диаметре, покрытый снаружи соединительнотканной капсулой – текой, состоящей из двух слоёв: наружный слой представлен плотной волокнистой тканью; внутренний - содержит РВСТ с кровеносными сосудами, нервами, а также клетки текоциты, которые вырабатывают тестостерон под действием ЛГ. Изнутри фолликул выстлан зернистым (гранулёзным) слоем - фолликулярными клетками, лежащими на базальной мембране, преобразующими тестостерон текоцитов в эстрогены под действием ФСГ. Овоцит окружён лучистым венцом (на данной схеме) и находится в яйценосном бугорке (кумулюсе). В полости фолликула находится жидкость, содержащая большое количество биологически активных веществ, в том числе эстрогенов. За 36 часов до овуляции происходит выброс ЛГ, под действием которого в овоците мейоз возобновляется и затем останавливается на стадии метафазы II, когда исчезает ядерная мембрана в ядре яйцеклетки. В таком состоянии, окружённый лучистым венцом, с полярным тельцем в перивителлиновым пространстве за 1-2 часа до овуляции овоцит отрывается от кумулюса и плавает в полости фолликула. После овуляции этот комплекс клеток попадает в маточную трубу. Мейоз завершается при оплодотворении. Зрелая (гаплоидная) женская половая клетка жизнеспособна 6-24 часа с момента овуляции. 69 Опишите клетки, определите их локализацию. Назовите структуры, обозначенные цифрами. Рис. 6. Эпителий слизистой оболочки маточной трубы. 1.Базальная мембрана. 2. Ядро реснитчатой клетки. 3.Реснички. 4.Ядро секреторной клетки. 5.Секреторные гранулы. 6. Межклеточные соединения. Однослойный однорядный призматический эпителий слизистой оболочки маточной трубы располагается на базальной мембране (1). Под эпителием собственная пластинка слизистой из рыхлой волокнистой соединительной ткани. Эпителий образован клетками двух видов: реснитчатыми и секреторными. Эпителий маточной трубы - целонефродермального типа, формируется из эпителия парамезонефрального (Мюллерова) протока. Слизистая оболочка образует глубокие складки. Овоцит достигает матки продвигаясь между складками в течение 5 суток. Реснитчатые клетки имеют округлое ядро (2), светлую цитоплазму, слабо развитые органеллы синтеза, множество митохондрий. На апикальной поверхности находятся реснички (3), которые начинаются от базальных телец, лежащих в поверхностном слое цитоплазмы клетки. Реснички мерцают с частотой около 22 биений в секунду и способствуют продвижению овоцита до и после оплодотворения в сторону матки. Секреторные эпителиоциты характеризуются крупным ядром (4), развитым синтетическим аппаратом. В апикальной части клеток, покрытой микроворсинками, накапливается в гранулах секрет (5), который затем апо- и мерокриновым способом выделяется в полость маточной трубы для питания овоцита, а также капацитации спермиев. Основными компонентами секрета являются преальбумины, трансферин, глобулин и липопротеиды, а также гликозаминогликаны, простагландины, утероглобин. Продукты химического распада гликозаминогликанов в просвете труб служат источником энергии, необходимой для поддержания функциональной активности и подвижности сперматозоидов. Эпителий слизистой оболочки маточной трубы изменяется под действием женских половых гормонов. На данном рисунке реснитчатые клетки высокие, реснички на их поверхности хорошо развиты, в секреторных эпителиоцитах много гранул, что говорит о насыщенности организма эстрогенами, наблюдаемой в конце фолликулярной стадии овариального цикла. Частота биения ресничек, также как и сократительная активность трубы усиливаются в периовуляторный период и угнетаются прогестероном в лютеиновую фазу цикла. Это связано с тем, что женская половая клетка по маточной трубе самостоятельно передвигаться не может. Движения овоцита с лучистым венцом осуществляются с участием биения ресничек клеток эпителия, складок слизистой оболочки, перистальтических сокращений мышечной оболочки стенки маточной трубы, всё это вызывает направленный в сторону матки ток жидкости. Клетки лучистого венца способствуют перемещению яйцеклетки. В предменструальный период (конец лютеиновой фазы цикла) реснички в реснитчатых клетках дегенерируют. В секреторных эпителиоцитах снижается образование веществ. Поскольку на рисунке реснитчатые клетки преобладают по количеству над секреторными, то локализация данного участка эпителия - воронка или ампулярный отделы маточной трубы, где количество реснитчатых клеток в эпителии максимально. Реснитчатые клетки убывают по направлению движения яйцеклетки – от воронки трубы к матке. И, наоборот, в эпителии перешейка и интерстициального (маточного) отдела маточной трубы, преобладают секреторные клетки, образующиеся продукты которых входят в состав трубной жидкости, необходимые для капацитации и энергообеспечения сперматозоидов, а также освобождения овоцита от фолликулярных клеток лучистого венца. Эпителиоциты слизистой оболочки маточной трубы, образуют различные межклеточные соединения в области латеральных поверхностей: простые и плотные соединения, интердигитации, десмосомы (6). 70 Глава 9. ЭМБРИОЛОГИЯ Назовите структуры, обозначенные цифрами. Опишите происходящий процесс. Рис. 1. Поперечный срез зародыша на стадии первичной полоски. 1.Первичная полоска. 2.Первичная эктодерма (эпибласт). 3.Первичная энтодерма (гипобласт). 4. Выселяющиеся клетки мезодермы. 5.Первичная бороздка. 6. Митозы в клетках эктодермы. Зародыш находится на стадии первичной полоски (1), соответствующей концу второй – началу третьей недели развития (смотри также рис. «Зародыш человека в плодном пузырьке»). У зародыша человека гаструляция начинается в конце 1-й недели развития, по окончании дробления и освобождения от прозрачной зоны, одновременно с имплантацией. На 7-е сутки развития зародыш имеет вид бластоцисты: пузырька с бластоцеломической полостью, стенка которого состоит из клеток трофобласта; у одного из полюсов пузырька изнутри – скопление клеток эмбриобласта. Гаструляция у человека протекает в две фазы. Первая фаза гаструляции продолжается всю 2-ю неделю. Материал внутренней клеточной массы бластоцисты (эмбриобласт) расщепляется способом деламинации на два листка, формируя зародышевый диск: эпибласт (первичную эктодерму), который даёт начало трём зародышевым листкам эмбриона и образует внезародышевую мезодерму; и гипобласт (первичную энтодерму), дифференцирующийся в энтодерму желточного мешка. Между клетками эпибласта образуется полость, разделяющая эпибласт на внезародышевую эктодерму амниона и зародышевый эпибласт. Вторая фаза гаструляции происходит на 3й неделе развития способом иммиграции и завершается формированием трёх зародышевых листков – эктодермы (2), энтодермы (3), мезодермы (4). В основе гаструляции лежит клеточная пролиферация и перемещение клеток зародышевого эпибласта сразу после освобождения бластоцисты от прозрачной зоны. Скапливающиеся на периферии зародышевого диска клетки передвигаются к каудальному концу зародыша, где встречаются и устремляются кпереди в виде клеточного потока (к краниальному концу). Удлиняющийся клеточный поток представляет материал первичной полоски – утолщение эпибласта по середине зародышевого диска. Второй поток клеток (более медленный) распространяется со стороны головного конца зародыша и движется по средней линии навстречу первому потоку. Место встречи двух потоков называется первичный (гензеновский) узелок – возвышение в переднем отделе первичной полоски. В центре первичного узелка появляется первичная ямка, а по средней линии первичной полоски – первичная бороздка (5), как продолжение первичной ямки. Иммиграция инициируется в области гензеновского узелка. Мигрирующие клетки второго потока подворачиваются вниз и через первичную ямку перемещаются в краниальном направлении. Это приводит к формированию головного отростка – зачатка хорды. Часть мигрирующих клеток первичного узелка и передней трети первичной полоски встраивается в материал головной кишки (прехордальной пластинки) и зародышевой энтодермы, оттесняя в стороны клетки гипобласта (3), при этом гензеновский узелок деформируется, а первичная полоска укорачивается. На данной схеме срез зародыша произведён через заднюю часть первичной полоски, соответствующую первому, более быстрому потоку клеток эпибласта, которые выселяются в пространство между эпибластом и гипобластом, точнее уже энтодермой, мигрируют в латеральном направлении и образуют зародышевую и внезародышевую мезодермы (4), а также часть клеток формирует энтодерму. Остальные клетки зародышевого эпибласта (2), не проходящие через первичную полоску, образуют эктодерму зародыша. Клетки эктодермы высокие, крупные, призматические, имеют вид многорядного призматического эпителия с многочисленными митозами (6). 71 Опишите строение зародыша, формирующихся внезародышевых органов; определите стадию развития. Назовите структуры, обозначенные цифрами. Рис. 2. Зародыш человека в плодном пузырьке (15 сутки развития). 1.Первичная полоска. 2.Первичный (гензеновский) узелок. 3.Первичная ямка. 4. Зародышевая эктодерма. 5.Зародышевая энтодерма. 6.Выселяющиеся клетки мезодермы. 7.Внезародышевая эктодерма. 8.Внезародышевая энтодерма. 9.Полость амниона. 10. Желточный мешок. 11.Внезародышевая мезодерма. 12.Амниотическая ножка. 13. Цитотрофобласт. 14.Базальная мембрана трофобласта. 15.Симпластотрофобласт, образующий первичные ворсины хориона. 16.Собственная пластинка эндометрия. 17. Лакуны с материнской кровью. 18. Бластоцеломическая полость. Зародыш находится на стадии первичной полоски (1), в середине которой – первичная бороздка. На зародышевом диске виден возвышающийся над плоскостью зародышевого диска первичный узелок (2), в котором углубление (3) – первичная ямка. На данном этапе, соответствующем концу второй – началу третьей недели развития, идёт вторая стадия гаструляции: зародыш состоит из зародышевой (первичной) экто- и энтодермы (4, 5), клетки первичной эктодермы мигрируют с образованием третьего зародышевого листка – мезодермы (6), зародышевой и внезародышевой. Таким образом, образуется трёхслойный зародыш, длина которого около 0,4 мм. Все остальные структуры плодного пузырька относятся к развивающимся внезародышевым органам. Сформированы выстланные внезародышевой эктодермой (7) и внезародышевой энтодермой (8) соответственно полость амниона (9) и желточный мешок (10). Снаружи к их стенкам подрастает внезародышевая мезодерма (11). Амнион существует до конца эмбрионального развития, он полностью окружает зародыш и является самой внутренней оболочкой плода. Внезародышевая эктодерма в процессе развития становится призматическим эпителием, который вырабатывает околоплодные воды, обеспечивающие оптимальную среду для развития зародыша и предохраняющие его от высыхания и механических воздействий. Желточный мешок у человека – рудиментарное образование, утратившее функцию накопления запасов питательных веществ. Внезародышевая энтодерма трансформируется в плоские эпителиоциты - особый тип кишечного эпителия. Внезародышевая мезодерма даёт начало мезенхиме, в которой до 7-8-й недели эмбриогенеза происходит кроветворение, а также образуются первичные половые клетки – гонобласты из мигрирующих сюда клеток первичной эктодермы. В дальнейшем: в каудальной части первичной кишки появится вырост – аллантоис, врастающий в амниотическую ножку (12), через него сосуды, образующиеся в аллантоисе и желточном мешке, связываются с ворсинками хориона, обеспечивая питание развивающегося зародыша. Амниотическая ножка (12), посредством которой зародыш связан с хорионом, постепенно трансформируется в пупочный канатик, в состав которого также входит редуцирующиеся к 3-му месяцу желточный мешок в виде желточного стебелька и аллантоис. Зародыш и внезародышевые органы: амнион, желточный мешок, аллантоис, мезодерма хориона, образуются из материала внутренней клеточной массы (эмбриобласта) бластоцисты, в то время как трофобласт, являющийся эпителием хориона, образуется из наружной клеточной массы. Внутренний слой трофобласта - клетки цитотрофобласта (13), располагаются на базальной мембране (14). Находясь снаружи, симпластотрофобласт образует выросты, или первичные ворсины хориона (15), которые проникают, разрушая, в соединительнотканную строму эндометрия матки (16), для взаимодействия с кровеносными сосудами матери (17). В плодном пузырьке, как следующего этапа развития бластоцисты, остаётся бластоцеломическая полость (18), которая впоследствии уменьшается и практически исчезает из-за слияния соединительнотканных пластинок хориона и аниона. Плодный пузырёк в конце второй недели достигает 10 мм в диаметре, находится в толще эндометрия матки. 72 Представлен фрагмент плаценты. Назовите обозначенные структуры и охарактеризуйте комплекс. Рис. 3. Гематоплацентарный барьер. 1 - эритроциты крови плода, 2 – просвет капилляра третичной ворсинки хориона, 3 – ядро эндотелиоцита капилляра, 4 – базальная мембрана капилляра, 5 – соединительнотканная строма ворсинки, 6 – базальная мембрана трофобласта, 7 – ядра клеток цитотрофобласта, 8 – ядра симпластотрофобласта, 9 – гр.ЭПС, 10 – комплекс Гольджи, 11 – митохондрии, 12 – секреторные гранулы, 13 – пиноцитозные пузырьки, 14 – микроворсинки симпластотрофобласта, 15 – лакуны с материнской кровью, 16 - эритроциты крови матери. На данном участке плаценты представлены компоненты гематоплацентарного (фетоплацентарного) барьера, который способствует избирательной проницаемости веществ из крови матери (16) в кровь плода (1) и в обратном направлении. Плацента – это комплекс структур материнского организма, представленный видоизменённой слизистой оболочкой матки (децидуальной оболочкой), и плодной оболочкой – хорионом. Гематоплацентарный барьер включает все структуры третичной ворсины хориона плода: эндотелий капилляра соматического типа (3), базальная мембрана капилляра (4), соединительнотканная строма ворсинки (5), базальная мембрана трофобласта (6), внутренний слой трофобласта - клетки цитотрофобласта (7), наружный слой трофобласта - симпластотрофобласт (синцитиотрофобласт) (8), который непосредственно контактирует с материнской кровью, находящейся в лакунах эндометрия матки (15). Ядра симпластотрофобласта овальные (8), с гетерохроматином по периферии. В базальной части симпластотрофобласта находятся хорошо развитые органеллы: агр.ЭПС и гр.ЭПС (9), комплекс Гольджи (10) митохондрии (11), в апикальной части – множество секреторных гранул (12), пиноцитозных пузырьков (13), на поверхности имеются микроворсинки (14), по строению и функциям подобные микроворсинкам столбчатых энтероцитов (в тонкой кишке). Щёточная каёмка принимает участие в транспорте веществ. Важной функцией клеток трофобласта в составе плаценты является иммунодепрессивная, в результате чего организм матери не отвергает плод. Это связано с продукцией хорионического гонадотропина (ХГч) и прогестерона, который является «натуральным супрессором», стимулируя выработку лимфоцитами матери медиаторного белка - прогестерон-индуцированного блокирующего фактора (ПИБФ), подавляющего активность клеточного иммунитета (угнетает Т-х1, активируют Т-х2). Другой механизм защиты плода от лимфоцитов матери связан с отсутствием антигенов главного комплекса гистосовместимости в клетках трофобласта человека. В трофобласте образуются гормоны, многие из которых регулируются по двуклеточной модели, что имитирует взаимосвязи между гипоталамусом и гипофизом. В цитотрофобласте вырабатываются гонадолиберин, соматолиберин, кортиколиберин, а под их действием в симпластотрофобласте образуются соответственно ХГч, плацентарный лактоген, АКТГ. ХГч активирует образование прогестерона по механизму аутокринной регуляции тем же симпластотрофобластом (после 9-й недели беременности, после чего жёлтое тело в яичнике угасает). В трофобласте вырабатываются эстрогены, необходимые, в первую очередь, для увеличения кровотока по сосудам плаценты. Плацентарный лактоген обладает соматотропным, лактогенным и лютеотропным действием. Выработка плацентарного кортиколиберина, стимулирующего выработку АКТГ плода, усиливается перед родами, что связано с их участием в определении срока родов. Также локально кортиколиберин усиливает плацентарный биосинтез простагландинов и сократимость матки. Гематоплацентарный барьер проницаем для некоторых веществ, которые переносятся путём диффузии и активного трансмембранного транспорта. Барьер проницаем для газов, электролитов, низкомолекулярных белков (альбуминов) и аминокислот, углеводов, витаминов, иммуноглобулинов G (обеспечивающих пассивный иммунитет плода), а также для алкоголя, никотина, наркотических веществ, лекарств, в том числе антибиотиков, большинства вирусов (особенно опасен вирус краснухи, вызывающий уродства у плода). 73 ГЛАВА 10. ОРГАНЫ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ Какая клетка изображена на рисунке? Назовите структуры, обозначенные цифрами. Рис. 1. Собственные железы желудка. Главный экзокриноцит. 1.Ядро. 2.Митохондрии.3.Комплекс Гольджи. 4.Гр. ЭПС. 5.Гранулы белкового секрета. 6.Микроворсинки.7.Кровеносный капилляр. Главные экзокриноциты находятся в области дна и тела собственных желез желудка. Это небольшие по размеру клетки имеют базофильную цитоплазму ( за счет высокого содержания РНК). В апикальной части короткие микроворсинки (6), секреторные гранулы (5). Клетки имеют хорошо выраженный комплекс Гольджи (3), гр. ЭПС (4). Главные экзокриноциты секретируют пепсиноген, который в присутствии соляной кислоты преобразуется в пепсин. 74 Фрагмент какого органа изображен? Назовите структуры, обозначенные цифрами. Рис. 2. Собственные железы желудка. Париетальный экзокриноцит. 1.Внутриклеточные канальцы. 2.Митохондрии. 3.Комплекс Гольджи. 4.Гр. ЭПС. 5.Межклеточные канальцы. Собственные железы желудка относятся к простым трубчатым железам, где различают дно, тело, перешеек, шейку. Дно желез расположено в мышечной пластинке слизистой оболочки желудка. Париетальные экзокриноциты являются самыми крупными по размерам в собственных железах. Располагаются поверх главных экзокриноцитов и мукоцитов. Имеют неправильную округлую форму. Находятся в области тела и шейки железы. Имеют оксифильную цитоплазму с 1 или 2 ядрами. Особенностью этих клеток является наличие в них внутриклеточных канальцев. Внутриклеточные канальцы (1) с микроворсинками, везикулами, трубочками транспортируют ионы водорода и хлора. Они переходят в межклеточные канальцы (5), которые находятся между главными экзокриноцитами и мукоцитами, а далее открываются в просвет железы. Функция этих клеток заключается в выработке ионов водорода и хлоридов из которых образуется соляная кислота. Процесс синтеза соляной кислоты энергозатратный, поэтому в клетке большое количество митохондрий (2). В присутствии соляной кислоты пепсиноген преобразуется в пепсин, соляная кислота разрушает связи в белках, уничтожает бактерии. Устанавливается оптимальная рН для протеолитического действия пепсина. Кроме того, эти клетки синтезируют и выделяют антианемический фактор (фактор Кастла), являющийся по химической природе мукопротеидом, он связывает витамин В12, способствуя его всасыванию в тонкой кишке. В базальной части париетальных клеток синтезируются и секретируются бикарбонаты. которые далее диффундируют в кровь сосудов находящихся в собственной пластинке слизистой. Далее попадают в базальную часть клеток эпителия, а они выделяют бикарбонаты в слизь. 75 Фрагмент какого органа изображен? Назовите структуры, обозначенные цифрами. Рис. 3. Собственные железы желудка. Главный экзокриноцит. Мукоцит (слизистая клетка). 1.Главный экзокриноцит. 2.Ядро. 3.Митохондрии. 4.Комплекс Гольджи. 5.Гр. ЭПС. 6.Гранулы белкового секрета. 7.Микроворсинки.8.Кровеносный капилляр. 9.Мукоцит. 10.Агр. ЭПС. 11.Митохондрии. 12.Гранулы слизистого секрета. Главные экзокриноциты (1) находятся в области дна и тела собственных желез желудка. Это небольшие по размеру клетки, имеют базофильную цитоплазму (высокое содержание РНК). В апикальной части - короткие микроворсинки (7), секреторные гранулы (6) диаметром 0,9 – 1мкм. Клетки имеют хорошо выраженный комплекс Гольджи (4), гр. ЭПС (5). Главные экзокриноциты секретируют пепсиноген, который в присутствии соляной кислоты преобразуется в активный пепсин. Между клетками имеются межклеточные канальцы. В грудном возрасте главные экзокриноциты продуцируют химозин, липазу, которые находятся в составе зимогенных гранул. У взрослых секреция этих компонентов минимальна, либо отсутствует совсем. Мукоциты - слизистые клетки (9). Располагаются в теле собственных желез. Имеют призматическую форму, светлую цитоплазму и микроворсинки (7) на апикальной поверхности. Ядро (2) расположено в базальной части клетки, содержит плотный хроматин. В цитоплазме комплекс Гольджи (4), агранулярная ЭПС (10), митохондрии (11). В апикальной части гранулы слизистого секрета (12). Помимо выработки слизи, эти клетки, будучи малодифференцированными, являются источником регенерации. Между клетками располагаются межклеточные канальцы. 76 Назовите фрагмент органа, тип клеток, структуры, обозначенные цифрами. Рис. 4. Столбчатые эпителиоциты ворсинки тонкой кишки. 1.Базальная мембрана. 2.Микроворсинки. 3.Гликокаликс. 4.Ядро. 5.Межклеточные канальцы. Столбчатые эпителиоциты (каемчатые клетки, энтероциты, клетки со щеточной каемкой) располагаются на базальной мембране (1). Это короткоживущие клетки. Имеют высокопризматическую форму, ядро (4) в базальной части клетки. На поверхности микроворсинок (2) высотой 1 мкм– гликокаликс (3) с ферментами – аминопептидазы, щелочная фосфатаза, гликозидазы, а так же гликозаминогликаны, гликопротеины и др. Ферменты синтезируются самими клетками. В микроворсинках пучки актиновых филаментов, которые идут в цитоплазму, а под микроворсинками терминальный слой богатый микрофиламентами. Актиновые филаменты вплетаются в этот терминальный слой. Поэтому в разные периоды пищеварения высота микроворсинок меняется: они могут быть выше или ниже. Апикальные и латеральные поверхности клеток связаны плотными изолирующими контактами, опоясывющими десмосомами. Это препятствует проникновению молекул химуса через эпителий. Формируется кишечный барьер, при нарушении которого может возникнуть пищевая аллергия. Ниже зоны опоясывающих десмосом контакты менее плотные, поэтому, в частности, жиры выходят в межклеточное пространство (5), откуда далее транспортируются через базальную мембрану в лимфатические капилляры собственной пластинки слизистой оболочки. В цитоплазме хорошо развит комплекс Гольджи, ЭПС, митохондрии, лизосомы. Таким образом, столбчатые эпителиоциты вырабатывают ферменты, за счет наличия микроворсинок осуществляют всасывательную функцию. Из продуктов расщепления образуются мономеры, которые всасываются клеткой. Эти клетки способны расщеплять белки, жиры, углеводы. Участвуют в пристеночном пищеварении. 77 Фрагмент какого органа на рисунке? Назовите структуры, обозначенные цифрами. Рис. 5. Дно крипты в тонкой кишке. 1.Клетка с ацидофильными гранулами (Панета). 2.Эндокриноцит. 3.Бокаловидный экзокриноцит. 4.Недифференцированная (бескаемчатая) клетка. 5.Тучная клетка в соединительной ткани между криптами. Крипты - углубления эпителия в собственную пластинку слизистой. В области дна крипт находятся клетки имеющие форму пирамиды с ацидофильными гранулами в апикальной части (1). В составе гранул белково-полисахаридный комплекс (ацидофилия объясняется наличием щелочного белка аргинина). Клетки вырабатывают бактерицидный фермент лизоцим; вырабатывают дипептидазы, расщепляющие дипептиды до аминокислот; вырабатывают секрет, нейтрализующий соляную кислоту. Эндокриноциты (2) также имеют форму пирамиды с зернистостью в базальной части клетки. Вырабатывают гормоны. Эндокриноциты контактируя с поступившим химусом активируются и начинают секретировать гормоны, которые далее поступают в кровеносную систему. В частности, вырабатывают мотилин, который стимулирует двигательную функцию ворсинок, вещество Р - регулирует кровоснабжение слизистой оболочки и др. Клетки относят к диффузной эндокринной системе. Недифференцированные клетки (4) (бескаемчатые) находятся в глубине крипт и являются источником обновления клеток эпителия в криптах и ворсинках. Бокаловидный экзокриноцит (3) - клетка с хорошо развитым комплексом Гольджи, агр. ЭПС. Слизистый секрет имеет полисахаридную природу, накапливается в апикальной поверхности клетки. Ядро смещено к базальной части клетки. Бокаловидные экзокриноциты выполняют защитную функцию (защищают от самопереваривания и механических воздействий). За 2-4 дня жизни этих клеток они совершают 2-3 цикла секреции. 78 Фрагмент какого органа на рисунке? Назовите структуры, обозначенные цифрами. Рис. 6. Фрагмент крипты в толстой кишке. 1.Бокаловидный экзокриноцит. 2.Столбчатый эпителиоцит. 3.Эндокриноцит. 4.Базальная мембрана. В эпителии крипт содержатся бокаловидные экзокриноциты (1), количество их увеличивается по направлению к прямой кишке. Вырабытываемая слизь облегчает продвижение каловых масс и защищает слизистую оболочку. Эндокриноциты (3) имеют форму пирамиды с зернистостью в базальной части клетки. Вырабатывают гормоны. Эндокриноциты, контактируя с поступившим химусом, активируются и начинают секретировать гормоны, которые далее поступают в кровеносную систему. В частности, вырабатывают мотилин, который стимулирует двигательную функцию ворсинок, вещество Р регулирует кровоснабжение слизистой оболочки и др. Столбчатые эпителиоциты (каемчатые клетки, энтероциты, клетки со щеточной каемкой) располагаются на базальной мембране (2). Это короткоживущие клетки. Имеют высокопризматическую форму, ядро в базальной части клетки. На поверхности микроворсинок высотой 1 мкм– гликокаликс с ферментами – аминопептидазы, щелочная фосфатаза, гликозидазы, а так же гликозаминогликаны, гликопротеины и др. Ферменты синтезируются самими клетками. В микроворсинках пучки актиновых филаментов, которые идут в цитоплазму, а под микроворсинками терминальный слой богат микрофиламентами. Актиновые филаменты вплетаются в этот терминальный слой. Поэтому в разные периоды пищеварения высота микроворсинок меняется. Апикальные и латеральные поверхности клеток связаны плотными изолирующими контактами, опоясывющими десмосомами. В цитоплазме хорошо развиты: комплекс Гольджи, ЭПС, митохондрии, лизосомы. Таким образом, столбчатые эпителиоциты сходны по строению с аналогичными клетками тонкой кишки, однако их щеточная каемка развита слабее. 79 Какая клетка изображена на рисунке? Назовите структуры, обозначенные цифрами. Рис. 9. Клетка диффузной эндокринной системы в тонкой кишке. 1.Эндокриноцит. 2.Базальная мембрана. 3.Капилляр. 4.Инкреторные включения. 5.Столбчатый эпителиоцит. Эндокриноциты (1) имеют форму пирамиды с зернистостью в базальной части клетки. Вырабатывают гормоны: секретин (Sклетки) который стимулирует выделение сока поджелудочной железы, G –клетки вырабатывают гастрин и др. Клетки относят к диффузной эндокринной системе, которая представлена диффузно рассеянными в органах и тканях эндокриноцитами. Столбчатые эпителиоциты (5) (каемчатые клетки, энтероциты, клетки со щеточной каемкой) располагаются на базальной мембране (2). Имеют высокопризматическую форму, ядро в базальной части клетки. На поверхности микроворсинок высотой 1 мкм– гликокаликс с ферментами – аминопептидазы, щелочная фосфатаза, гликозидазы, а так же гликозаминогликаны, гликопротеины и др. Ферменты синтезируются самими клетками. Апикальные и латеральные поверхности клеток связаны плотными изолирующими контактами, опоясывющими десмосомами. В цитоплазме хорошо развиты: комплекс Гольджи, ЭПС, митохондрии, лизосомы. Таким образом, столбчатые эпителиоциты вырабатывают ферменты, за счет наличия микроворсинок осуществляют всасывательную функцию. Из продуктов расщепления образуются мономеры, которые всасываются клеткой. Эти клетки способны расщеплять белки, жиры, углеводы. Участвуют в пристеночном пищеварении. 80 Фрагмент какого органа на рисунке? Назовите структуры, обозначенные цифрами. Рис. 10. Фрагмент дольки печени. 1.Капилляр. 2.Перикапиллярное пространство (Диссе). 3.Ямочная клетка (Pit cells). 4. Эндотелиоцит. 5.Эритроцит. 6.Печеночный эпителиоцит (гепатоцит). 7.Комплекс Гольджи. 8.Митохондрия. 9. Ядро. 10. Ядрышко. 11. Гранулярная ЭПС. 12. Секреторные включения. 13. Включения гликогена. 14.Желчный капилляр, 15.Десмосома. 16. Перисинусоидальный липоцит ( клетка Ито). 17. Звездчатый макрофаг ( клетка Купфера). Структурно-функциональной единицей печени является печеночная долька, имеющая форму призмы. Состоит из печеночных балок, а они, в свою очередь, образованы гепатоцитами (6). Гепатоциты имеют многоугольную форму, 1 или 2 ядра. Составляют 80% всех клеток печени, живут более 1 года. В печеночных балках гепатоциты располагаются в 2 ряда. Между собой клетки соединяются при помощи десмосом (15), плотных контактов, по типу «замка». Между рядами располагаются желчные капилляры (14), которые не имеют собственной стенки (ею являются билиарные поверхности гепатоцитов) и начинаются они слепо. Поверхность гепатоцита обращенная к синусоидному капилляру называется васкулярной. Васкулярной поверхностью гепатоцит выделяет в кровь белки, витамины, глюкозу, липидные комплексы. Васкулярная и билиарная поверхности гепатоцитов имеют микроворсинки. В норме желчь не поступает в кровь. Возможность попадания желчи в кровь создается при повреждении гепатоцитов (возникает паренхиматозная желтуха). Цитоплазма печеночных эпителиоцитов воспринимает кислые и основные красители. Клетки содержат много органелл. Хорошо развит комплекс Гольджи (7), где осуществляется биосинтез липопротеинов, гликопротеинов. Комплекс Гольджи может смещаться к той или иной поверхности гепатоцита в зависимости от того, что в данный момент синтезирует гепатоцит. Гранулярная ЭПС (11) располагается плотно, на ее поверхности синтезируется много белков, которые затем поступают в комплекс Гольджи. Агранулярная ЭПС отвечает за синтез гликогена, липидов. Много митохондрий (8), имеющих овальную форму, с малым количеством крист. Митохондрии обеспечивают энергетические процессы. Лизосомы располагаются вблизи ядра, принимают участие во внутриклеточном переваривании. Пероксисомы расщепляют эндогенные перекиси. Включения гликогена (13), жира относят к трофическим. Их количество связано с пищеварением. Между печеночными балками находятся синусоидные капилляры (1), стенка которых выстлана эндотелиоцитами (4). Между эндотелиальными клетками располагаются звездчатые макрофаги (17) – клетки Купфера. Их функция осуществляется за счет высокой фагоцитарной активности, наличия лизосомального аппарата. Они очищают кровь от антигенов, токсинов, микроорганизмов; фагоцитируют поврежденные эритроциты. Со стороны просвета капилляра с помощью псевдоподий прикрепляются ямочные клетки (3). В их цитоплазме содержатся гранулы с биологически активными веществами и пептидными гормонами. Ямочные клетки относят к натуральным киллерам. Они уничтожают поврежденные гепатоциты; обладают эндокринной функцией (стимулируют пролиферацию печеночных клеток), их относят к APUD системе. Перисинусоидальное пространство (перикапиллярное) (2), в норме заполнено жидкостью, богатой белками. Здесь находятся микроворсинки гепатоцитов, отростки звездчатых макрофагов. Перисинусоидальные липоциты (16) – отросчатые клетки со слабо развитыми органеллами. Вокруг ядра и в отростках липидные капли. Располагаются в перисинусоидальном пространстве между гепатоцитами. В норме эти клетки накапливают витамин А (в цитоплазме в виде мелких липидных капель), в патологических условиях вырабатывают коллаген, что может приводить к фиброзу печени. В этих клетках много рибосом, меньше митохондрий. 81 Фрагмент какого органа изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 11. Синусоидный капилляр в печеночной дольке. 1.Капилляр. 2.Перывистая базальная мембрана. 3.Эритроцит. 4.Эндотелиоцит. 5.Фрагмент печеночного эпителиоцита (гепатоцита). 6.Звездчатый макрофаг (клетка Купфера). 7.Перисинусоидальный липоцит (клетка Ито). 8.Пространство Диссе (периваскулярное). Между печеночными балками находятся синусоидные капилляры (1), стенка которых выстлана эндотелиоцитами (4). Между эндотелиальными клетками располагаются звездчатые макрофаги (6) – клетки Купфера образующиеся из моноцитов крови. Этих клеток больше на периферии печеночной дольки. Отростки этих клеток проникают в пространство Диссе (8). Их функция состоит в высокой фагоцитарной активности. Они очищают кровь от антигенов, токсинов, микроорганизмов, фагоцитируют поврежденные эритроциты, стимулируют регенерацию гепатоцитов. Перисинусоидальное пространство (перикапиллярное, пространство Диссе), в норме заполнено жидкостью, богатой белками. Здесь находятся микроворсинки гепатоцитов, отростки звездчатых макрофагов (6). Перисинусоидальные липоциты (клетки Ито) (7) – отросчатые клетки со слабо развитыми органеллами. Их отростки контактируют как с синусоидными капиллярами, так и с гепатоцитами. Вокруг ядра и в отростках липидные капли. Клетки располагаются в перисинусоидальном пространстве между гепатоцитами. В норме эти клетки накапливают витамин А ( в цитоплазме в виде мелких липидных капель) и другие жирорастворимые витамины (А, Д, Е, К). В патологических условиях вырабатывают коллаген, что может приводить к циррозу печени. В этих клетках много рибосом, меньше митохондрий. 82 Фрагмент каких клеток изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 12. Печеночный эпителиоцит. Включения в клетках печени. 1.Микроворсинки. 2.Секреторные белковые включения. 3.Митохондрия. 4.Включения гликогена. 5.Лизосома. 6.Липидные включения. 7.Гр.ЭПС. 8.Комплекс Гольджи. Печеночная долька состоит из печеночных балок, а они, в свою очередь, состоят из гепатоцитов (печеночных эпителиоцитов). Гепатоциты имеют многоугольную форму, 1 или 2 ядра. На апикальной поверхности микроворсинки (1).Между собой клетки соединяются при помощи десмосом, плотных контактов, по типу «замка». Цитоплазма печеночных эпителиоцитов воспринимает кислые и основные красители. Клетки содержат много органелл. Хорошо развит комплекс Гольджи (8), где осуществляется биосинтез липопротеинов, гликопротеинов. Комплекс Гольджи может смещаться к той или иной поверхности гепатоцита в зависимости от того, что в данный момент синтезирует гепатоцит. Гранулярная ЭПС (7) располагается плотно, на ее поверхности синтезируется много белков, которые затем поступают в комплекс Гольджи. Агранулярная ЭПС отвечает за синтез гликогена, липидов, осуществляет дезинтоксикационную функцию за счет ферментов, которые поступают из гранулярной ЭПС. Много митохондрий (3), имеющих овальную форму, с малым количеством крист. Митохондрии обеспечивают энергетические процессы. Лизосомы располагаются вблизи ядра, принимают участие во внутриклеточном переваривании, а также участвуют в защитных реакциях. Пероксисомы расщепляют эндогенные перекиси. Включения гликогена (4), жира относят к трофическим. Их количество связано с пищеварением. Липиды захватываются печенью из крови, а так же синтезируются самими гепатоцитами. Накапливаются в виде липидных включений (6). Помимо липидных включений в клетках присутствуют секреторные белковые включения (2). 83 Фрагмент какого органа изображен на рисунке? Назовите структуры, обозначенные цифрами. Рис. 13. Фрагмент поджелудочной железы. Ацинозно-инсулярная клетка. 1.Кровеносный капилляр. 2.Секреторные включения с гормонами. 3.Зимогенные гранулы. 4.Комплекс Гольджи. 5.Гранулярная ЭПС. 6.Митохондрии. 84 Структурно-функциональная единица какого органа пищеварительной системы изображена на рисунке? Каково строение и функция? Рис. 15. Поджелудочная железа. Панкреатический ацинус. 1 - панкреатический экзокриноцит на базальной мембране; 2 – зимогенная зона; 3 – гомогенная зона; 4 – центроацинозный эпителиоцит; 5 - вставочный проток; 6 – кровеносный капилляр Поджелудочная железа является смешанной: в ней осуществляется выработка панкреатического сока необходимого для пищеварения (экзокринная часть), а также присутствуют островки из эндокринных клеток (эндокринная часть). Экзокринная часть представлена ацинусами, где различают секреторный отдел и вставочный проток (5). В составе ацинуса может быть более десяти клеток называемых панкретатическими экзокриноцитами (ациноциты, экзокринные панкреатоциты), которые располагаются на базальной мембране. Эти клетки синтезируют ферменты входящие в состав панкреатического сока. Секреция и выделение пищеварительных ферментов стимулируется гормоном холецистокинином, который вырабатывают эндокринные клетки находящиеся в тонкой кишке. В панкреатическом экзокриноците (1) ядро располагается ближе к базальной поверхности. В клетке выделяют гомогенную и зимогенную зоны. В зимогенной зоне (2) находятся секреторные гранулы различной степени зрелости. В гомогенной зоне (3) присутствует гранулярная ЭПС, где осуществляется синтез ферментов панкреатического сока. Некоторые ферменты выделяются в форме проферментов (трипсин, эластаза и др.), а некоторые в форме активных ферментов (амилаза, липаза и др.). Выработка секрета осуществляется циклически и включает фазу поступления исходных веществ для образования панкреатического сока, далее их синтез, накопление секрета и выделение по мерокриновому типу (без нарушения целостности клеток). Выделившийся секрет попадает во вставочный проток (5). Клетки образующие начальный отдел вставочного протока называются центроацинозными эпителиоцитами (4). Они имеют уплощенную форму, овальное ядро, бедны органеллами. В соединительной ткани вокруг ацинусов кровеносные сосуды. На рисунке изображены капилляры прилежащие к базальной мембране ацинуса. 85 Какая клетка изображена на рисунке? Назовите структуры, обозначенные цифрами. Рис. 16. Дентинобласт (одонтобласт). 1.Гранулярная ЭПС. 2.Митохондрии. 3.Ядро. 4.Комплекс Гольджи. 5.Отросток (волокна Томса). 6.Дентин. 7. Дентинные канальцы.8.Десмосома. Клетки зубной пластинки имеют эктодермальное происхождение. В сформированном зубе дентинобласты находятся в периферическом слое пульпы, а в дентине находятся их отростки. Дентинобласты - клетки вытянутой формы с овальным ядром (3), располагающимся ближе к базальной поверхности. В цитоплазме гр. ЭПС (1), митохондрии (2), комплекс Гольджи (4). Клетки имеют отростки (5), в которых органелл меньше, чем в теле. Вырабатывают молекулы коллагена для образования коллагеновых волокон (коллаген I типа) и цементирующее вещество (фосфопротеины, протеогликаны, гликопротеины и др.). Элементы цитоскелета локализованы по длинной оси клетки. Первоначально образуется предентин, при этом удлиняются отростки, которые за счет образовавшегося предентина оказываются в канальцах (7). Канальцы придают исчерченность и направлены от границы между пульпой и дентином к дентинно-эмалевому и дентинно-цементному соединениям, параллельно друг другу. На поверхности плазмолеммы дентинобласты имеют рецепторы к ряду биологически активных веществ. Предентин (необызвествленный дентин) состоит из органических веществ. Минерализация дентина осуществляется при участии фермента щелочной фосфатазы, находящейся в цитоплазме дентинобластов, которая расщепляет глицерофосфат крови на фосфорную кислоту (которая в предентине далее соединяется с кальцием) и моносахара. Образовавшийся фосфорнокислый кальций, откладываясь в предентине, образует дентин (6). Т.о. дентин представляет кальцинированный материал на основе коллагена. Он плотнее костной ткани, более проницаем, чем эмаль. Минерализуется не весь предентин. В коронке зуба присутствуют интерглобулярные пространства, представленные неминерализованным дентином. Не подвергается минерализации предентин расположенный рядом с пульпой. С формирования дентина начинает формироваться корень зуба и осуществляется его прорезывание. Вторичный дентин (дентин раздражения) откладывается между первичным дентином и предентином. Предентин является местом постоянного роста дентина, который не прекращается и у взрослых. Дентинобласты сохраняются в пульпе взрослого человека в течение всей жизни, постоянно осуществляя дентинообразующую функцию. Дентинобласты связаны между собой межклеточными контактами, в частности, десмосомами (8), плотными и щелевидными контактами. 86 ГЛАВА 11. ОРГАНЫ ВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ Определите, фрагмент какого органа на схеме. Назовите структуры, обозначенные цифрами. Рис. 1. Почечное тельце с юкстагломерулярным аппаратом. 1.Приносящая артериола. 2.Выносящая артериола. 3.Полость проксимального извитого канальца. 4. Эндотелиальные клетки. 5. Базальная мембрана. 6.Мезангиальные клетки. 7. Полость эпителиальной капсулы клубочка. 8.Эпителий наружного листка капсулы клубочка. 9.Эпителий извитого канальца. 10.Подоциты. 11.Цитоподии подоцитов. 12. Клетки плотного пятна дистального канальца нефрона. 13.Юкстагломерулярные клетки. 14.Юкставаскулярные клетки. 15. Гранулы с ренином в цитоплазме юкстагломерулярных клеток. 16. Строма почки. Почечное тельце является начальной частью структурно-функциональной единицы почки – нефрона и состоит из сосудистого клубочка, покрытого эпителиальной капсулой. Тельце имеет два полюса: сосудистый (в области расположения приносящей (1) и выносящей (2) артериол) и мочевой (в участке отхождения проксимального извитого канальца (3)). В почечном тельце происходит процесс фильтрации плазмы крови с образованием первичной мочи. Сосудистый клубочек состоит из 50-100 капиллярных петель, на которые распадается приносящая артериола. Это первичная капиллярная сеть, или «чудесная»; выносящая артериола распадается с образованием вторичной капиллярной сети вокруг канальцев нефрона. Капилляры сосудистого клубочка выстланы клетками эндотелия (4), расположенными на базальной мембране (5). Между петлями капилляров находится мезангий, состоящий из межклеточного вещества (матрикса) и мезангиальных клеток (6), являющихся поддерживающими и фагоцитарными. Эпителиальная капсула клубочка (Шумлянского-Боумена) образована двумя листками – наружным (париетальным) и внутренним (висцеральным), между которыми находится полость, куда из просвета кровеносных капилляров фильтруется плазма крови с образованием первичной мочи. Полость капсулы (7) сообщается с просветом извитого канальца. Наружный листок капсулы представлен однослойным плоским эпителием (8), отграничивающим полость капсулы снаружи. В области мочевого полюса этот эпителий переходит в эпителий извитого канальца (9), а в области сосудистого полюса - в эпителий внутреннего листка (10). Клетки висцерального листка капсулы – подоциты (10), охватывают капилляры снаружи, их тела выступают в полость, а с помощью многочисленных отростков – цитоподий (11), подоциты прикрепляются к общей с эндотелиоцитами базальной мембране (5), участвуя в формировании фильтрационного барьера. В области сосудистого полюса почечного тельца находится юкстагломерулярный (околоклубочковый) аппарат, структуры которого: клетки плотного пятна дистального канальца нефрона (12), юкстагломерулярные (13) и юкставаскулярные клетки (14), регулируют артериальное давление посредством ренин-ангиотензиновой системы. Клетки плотного пятна обладают осморецепторной функцией: сигналы об изменениях концентрации Nа+ в просвете дистального канальца передаются ими юкстагломерулярным клеткам. Юкстагломерулярные клетки (миоидные эндокриноциты) – это видоизменённые гладкомышечные клетки средней оболочки приносящей артериолы клубочка в месте ее контакта с плотным пятном дистального канальца. В цитоплазме находятся гранулы (15), содержащие эндокринное вещество – ренин. Юкстагломерулярные клетки относятся к барорецепторам: при снижении артериального давления уменьшается растяжение стенки артериолы, приводящее к усилению секреции ренина и повышению артериального давления. Юкставаскулярные клетки (Гурмагтига) образуют скопление треугольной формы между приносящей, выносящей артериолами и плотным пятном, переходящее в мезангий. Точная функция не установлена. Возможно, эти клетки передают сигнал с клеток плотного пятна на сосуды, и при истощении юкстагломерулярных клеток в юкставаскулярных клетках может вырабатываться ренин. Вокруг почечного тельца находится соединительнотканная строма почки (16). 87 На схеме канальцы нефрона. Укажите, к каким отделам они принадлежат. Назовите обозначенные структуры. Рис. 2. Почка. Канальцы нефрона. А – проксимальный каналец, Б – тонкий каналец петли Генле, В – дистальный каналец; Г – просвет кровеносного капилляра; 1.Базальная мембрана. 2.Ядро. 3.Пиноцитозные пузырьки. 4.Митохондрии. 5.Микроворсинки. Проксимальный каналец (А). Стенка канальца образована однослойным кубическим эпителием, лежащим на базальной мембране (1). Для клеток характерно наличие крупного округлого ядра (2), множество пиноцитозных пузырьков (3), вакуолей и лизосом в цитоплазме. В базальной части клеток плазмолемма образует инвагинации, между которыми перпендикулярно базальной мембране располагается большое количество митохондрий (4), что создаёт на светооптическом уровне картину «базальной исчерченности». На апикальной поверхности клеток имеются многочисленные микроворсинки (5), образующие щёточную кайму. Проксимальный каналец начинается от мочевого полюса капсулы почечного тельца, имеет толщину 40-60 мкм, включает извитую часть и прямой нисходящий отдел (нисходящую толстую часть петли) и переходит в тонкую часть петли. Это самый протяжённый участок канальцев нефрона – до 14 мм. В проксимальном канальце происходит обратное всасывание (реабсорбция) более 80% объёма первичной мочи в окружающие каналец капилляры вторичной сети. Путём пиноцитоза происходит реабсорбция аминокислот и белков, которые сначала расщепляются в лизосомах. Из просвета канальца активно с помощью Na-K-АТФазы перекачиваются ионы Na+, сопровождающиеся пассивным переносом Cl- и воды. Здесь также всасывается глюкоза. В просвет проксимального канальца секретируется креатинин, лекарственные препараты и их метаболиты. Тонкий каналец петли Генле (Б). Стенка канальца образована однослойным плоским эпителием, лежащим на базальной мембране (1). Ядросодержащая часть клетки (2) выступает в просвет канальца. В цитоплазме – большое количество пиноцитозных пузырьков (3). На апикальной поверхности имеются немногочисленные микроворсинки (5). Тонкий отдел петли Генле располагается в мозговом веществе, его толщина - 13-15 мкм. В этом отделе нефрона происходит пассивный транспорт воды, что становится возможным благодаря осмотическому градиенту: из гипотонического содержимого просвета в гипертоническую среду окружающих канальцы кровеносные капилляры вторичной сети (Г). В результате моча становится концентрированной и гипертонической. В просвет тонкого канальца секретируется мочевина. Дистальный каналец (В). Стенка канальца представлена однослойным кубическим эпителием, лежащим на базальной мембране (1). В клетках находится по одному округлому ядру (2), в цитоплазме пиноцитозные пузырьки (3) и немногочисленные лизосомы. В базальной части клеток, также как в проксимальном канальце, находятся инвагинации плазмолеммы с большим количеством митохондрий (4). На апикальной поверхности имеются микроворсинки (5), не образуя щёточной каймы. Дистальный каналец имеет толщину 30-50 мкм, включает дистальный прямой каналец (восходящую толстую часть петли) и дистальный извитой каналец, который прилежит к сосудистому полюсу, где образует компонент юкстагломерулярного аппарата – плотное пятно, клетки которого регулируют выработку ренина. В дистальном канальце нефрона завершается обратное всасывание веществ, осуществляется транспорт электролитов (ионов Na+ и Cl-) из просвета, в то же время стенка канальца непроницаема для воды, которая остаётся в канальце. Это приводит к тому, что моча становится всё более гипотонической. В извитом отделе канальца в просвет транспортируются ионы Н+, K+ и NH+, придающие окончательной моче кислую реакцию и антимикробные свойства. На границе перехода дистального канальца в собирательные трубочки заканчивается нефрон. 88 Процесс реабсорбции регулируется антидиуретическим гормоном (вазопрессином) гипофиза и минералокортикоидами надпочечников. 89 Опишите и проанализируйте значение комплекса гистологических элементов. Назовите обозначенные структуры. Рис. 3. Связь подоцитов с капиллярами сосудистого клубочка почечного тельца. 1.Эритроциты. 2.Ядра эндотелиоцитов. 3.Фенестры в эндотелии. 4. Базальная мембрана. Подоцит: 5 – ядро, 6 – гр.ЭПС, 7 – митохондрии, 8 - комплекс Гольджи, 9 – элементы цитоскелета, 10 – первичные отростки, 11 – фильтрационные щели, 12 – цитоподии, 13 – интердигитации цитоподий соседних клеток; 14 – полость капсулы клубочка. Представлен фрагмент взаимодействия клеток внутреннего листка эпителиальной капсулы клубочка – подоцитов, с капиллярами сосудистого клубочка почечного тельца в нефроне почки. Подоциты охватывает со всех сторон капилляры. В просвете капилляра висцерального типа располагаются эритроциты (1), агрегация которых предотвращается отрицательно заряженной внутренней поверхностью капилляра. Эндотелиоциты, выстилающие капилляры, имеют крупные ядра (2), в ядросодержащей зоне клеток находятся немногочисленные органеллы. На остальном протяжении цитоплазма максимально (до 30% поверхности) истончена, образуя фенестры (3) – окошки диаметром до 100 нм, в результате плазма крови в этих участках контактирует непосредственно с базальной мембраной (4). Подоциты, или клетки внутреннего листка эпителиальной капсулы клубочка, - это крупные клетки, в них выделяют тело, где располагается ядро (5), органеллы синтеза (6), митохондрии (7), хорошо развитый комплекс Гольджи (8) и элементы цитоскелета (9): микротрубочки и филаменты, которых особенно много в отростках клеток. От тела отходят первичные отростки (10) – ножки (подии), которые ветвятся до тончайших нитей. Толщина отростков уменьшается по мере их ветвления, так что на поверхности капилляра образуется лабиринт из фильтрационных щелей (11), находящихся между многочисленными выростами – цитоподиями (12). Ножки подоцитов прикрепляются к базальной мембране, образуя интердигитации с цитоподиями соседних клеток (13). Между подоцитами и эндотелиоцитами находится общая для клеток толщиной до 300 нм базальная мембрана, формирующаяся обоими типами клеток и состоящая из трёх слоёв: наружный и внутренний слои светлые, средний слой более толстый и электронноплотный. Светлые слои содержат ламинин и гепаран сульфат, обладают анионными участками, отталкивающими отрицательно заряженные молекулы. Плотный слой содержит коллаген IV типа, формирующий филаменты в виде молекулярного сита с размерами пор 4 нм. Цитоплазма фенестрированных эндотелиоцитов капилляра клубочка, трёхслойная базальная мембрана и щелевые диафрагмы, закрывающие фильтрационные щели между цитоподиями подоцита, образуют фильтрационный барьер, через который вещества из просвета кровеносного капилляра фильтруются в полость капсулы клубочка (14). Через фильтрационный барьер проникают практически все компоненты плазмы крови: вода, хлориды, глюкоза, мочевина, мочевая кислота, креатинин, низкомолекулярные белки и др., образуя ультрафильтрат – первичную мочу, которой за сутки образуется около 150 л (из них около 148 л обратно всасывается в канальцах нефрона). Барьер непроницаем для форменных элементов крови и высокомолекулярных белков. Эпителиальная капсула Шумлянского-Боумена, как и канальцы нефрона, являются производными метанефрической мезодермы, окружающей метанефрический дивертикул, который даёт начало мочевыводящим путям. Начиная с 5-й недели внутриутробного развития медиальный конец каждого выделительного канальца метанефроса утолщается, в нём появляется углубление для врастающих веточек аорты, формирующих шаровидные скопления капилляров. Постепенно сосудистый клубочек охватывается медиальной частью канальца, которая становится двухслойной капсулой, и образуется почечное тельце. Первая функциональная деятельность почки – поступление в чашечки лоханки жидкости, не содержащей белок, отмечается на 11-12 неделе эмбрионального развития, что говорит об установившихся к этому времени связях подоцитов и эндотелиоцитов сосудистого клубочка в почечном тельце, начале формирования фильтрационного барьера. 90 Рис. 4. Юкстагломерулярный аппарат почки. 1.Эндотелий артериол. 2.Базальная мембрана эндотелия выносящей артериолы. 3. Эритроциты. 4.Сегментоядерный нейтрофил. 5.Гладкие миоциты выносящей артериолы. 6.Миоидные эндокриноциты (юкстагломерулярные клетки) приносящей артериолы. 7. Просветы капилляров сосудистого клубочка. 8.Клетки внутреннего листка капсулы клубочка (подоциты). 9.Эпителиоцит наружного листка капсулы клубочка. 10.Ядра мезангиальных клеток. Клетки плотного пятна: 11 – базальная мембрана, 12 ядра, 13 – митохондрии, 14 – интердигитации между соседними клетками, 15 микроворсинки; 16.Просвет дистального канальца нефрона. 17.Гранулы с ренином в цитоплазме юкстагломерулярных клеток. 18.Окончания аксонов симпатических нейронов. 19.Ядра юкставаскулярных клеток. Юкстагломерулярный (околоклубочковый) аппарат включает структуры, расположенные в области сосудистого полюса почечного тельца нефрона почки и участвует в регуляции кровяного давления посредством ренинангиотензиновой системы. Артериолы выстланы эндотелием (1), который располагается в выносящей артериоле на базальной мембране (2). В приносящей артериоле в области контакта с плотным пятном базальная мембрана прерывистая. В просвете видны эритроциты (3) и сегментоядерный нейтрофил (4). Средняя оболочка выносящей артериолы представлена гладкими миоцитами (5), приносящей – миоидными эндокриноцитами (6). Приносящая артериола даёт начало капиллярной сети (7) сосудистого клубочка почечного тельца, выстланной эпителиоцитами внутреннего листка капсулы клубочка (8), которые здесь переходят в эпителиоциты наружного листка (9). Между петлями капилляров находятся мезангиальные клетки, являющиеся поддерживающими и фагоцитарными (10). Непосредственно к юкстагломерулярному аппарату относятся: клетки плотного пятна, юкстагломерулярные и юкставаскулярные клетки. Плотное пятно образовано клетками дистального извитого канальца в участке между приносящей и выносящей артериолами клубочка. Состоит из 15-40 специализированных высоких эпителиоцитов, тесно контактирующих с приносящей артериолой и юкставаскулярными клетками, т.к. базальная мембрана в этой зоне тонкая (11). Ядра клеток округлые (12), в базальной части плазмолемма образует инвагинации, вдоль которых располагаются многочисленные митохондрии (13), придавая вид «базальной исчерченности». Между латеральными поверхностями клеток – интердигитации (14). На апикальных поверхностях клеток (15), обращённых в просвет дистального канальца (16), находятся микроворсинки. Клетки плотного пятна обладают осморецепторной функцией: сигналы об изменениях концентрации ионов Nа+ в крови и жидкости, омывающей плотное пятно, передаются ими юкстагломерулярным клеткам. Юкстагломерулярные клетки (миоидные эндокриноциты) – это видоизменённые гладкомышечные клетки средней оболочки приносящей и выносящей артериол клубочка (6). Органеллы в клетках развиты умеренно. В цитоплазме накапливаются гранулы округлой формы (17), содержащие протеолитический фермент - ренин. Эти клетки относятся к барорецепторам: при снижении артериального давления уменьшается растяжение стенки артериолы, что вызывает усиление ими секреции ренина. Клетки плотного пятна блокируют выработку ренина при избыточном содержании ионов Na+ в просвете дистального канальца. Медиатор норадреналин, выделяющийся из окончаний аксонов (18) симпатических нейронов, способствует выработке ренина. Юкставаскулярные клетки (Гурмагтига) образуют скопление треугольной формы (19) между приносящей, выносящей артериолами и плотным пятном, переходящее в мезангий. Полагают, что в юкставаскулярных клетках может вырабатываться ренин. 91 Рис. 5. Фильтрационный барьер в нефроне. 1.Просвет капилляра сосудистого клубочка. 2.Полость капсулы Шумлянского-Боумена. 3.Ядро эндотелиоцита. 4.Фенестры в цитоплазме эндотелиоцита. 5.Эритроциты в просвете капилляра. 6.Трёхслойная базальная мембрана. 7.Ядро подоцита. 8.Первичные отростки подоцита. 9.Фильтрационные щели. 10.Цитоподии эпителиоцитов внутреннего листка капсулы клубочка. 11.Контакты цитоподий соседних подоцитов. Фильтрационный барьер – совокупность структур почечного тельца (являющимся начальным отделом структурнофункциональной единицы почки – нефрона), находящихся между просветом капилляров сосудистого клубочка (1) и полостью эпителиальной капсулы клубочка (2). Через барьер фильтруются компоненты плазмы крови, образуя первичную мочу. Фильтрационный барьер почки состоит из трёх компонентов: эндотелиоцитов, базальной мембраны и подоцитов. Эндотелиоцит капилляра висцерального типа сосудистого клубочка имеет крупное ядро (3) в ядросодержащей зоне клетки, где также находятся немногочисленные органеллы, на остальном протяжении цитоплазма максимально (до 30% поверхности) истончена, образуя фенестры (4) – окошки диаметром до 100 нм, в результате плазма крови в этих участках контактирует непосредственно с базальной мембраной. Поверхность эндотелиоцита заряжена отрицательно, что отталкивает эритроциты (5) и предотвращает их агрегацию. Базальная мембрана (6) – общая для эндотелиоцита и подоцита, состоит из трёх слоёв: наружный и внутренний слои светлые, средний слой более толстый и электронноплотный. Светлые слои содержат ламинин и гепаран сульфат, обладают анионными участками, отталкивающими отрицательно заряженные молекулы, плотный слой содержит коллаген IV типа. Оба вида клеток участвуют в образовании базальной мембраны. Подоциты, или клетки внутреннего листка эпителиальной капсулы клубочка, - это крупные клетки, имеют тело, где располагается ядро (7) и органеллы, от тела отходят ветвящиеся первичные отростки (8), или подии. Толщина отростков уменьшается по мере их ветвления, так что на поверхности капилляра образуется лабиринт из фильтрационных щелей (9), находящихся между многочисленными выростами – цитоподиями (10). Ножки подоцитов прикрепляются к базальной мембране, образуя интердигитации с цитоподиями соседних клеток (11). Фильтрационные щели, имеющие ширину 30-40 нм, закрыты тонкими щелевидными диафрагмами, представляющими собой сеть из волокнистых структур с размерами ячеек сети до 10 нм. Молекулы, которые при фильтрации откладываются в щелях, фагоцитируются подоцитами. В области фенестр эндотелия фильтрационный барьер состоит только из базальной мембраны и щелевых диафрагм. Таким образом, фильтрационные щели являются главной частью барьера. Процесс фильтрации является первым этапом мочеобразования. При фильтрации не происходит расходования энергии, так как поток жидкости обеспечивается гидростатическим давлением равным 70-80 мм рт. ст., образующимся за счёт разницы диаметра приносящей и выносящей артериол. Через фильтрационный барьер проникают практически все компоненты плазмы крови: вода, аминокислоты, хлориды, глюкоза, мочевина, мочевая кислота, креатинин, низкомолекулярные белки и др., образуя ультрафильтрат – первичную мочу, которой за сутки образуется около 150 л (из них около 148 л реабсорбируется в канальцах нефрона). Барьер непроницаем для форменных элементов крови и высокомолекулярных белков. Полноценная фильтрация мочи осуществляется с 2-х лет жизни, когда дифференцируются элементы барьера (у новорожденных фильтрация снижена на 60-70% по сравнению с таковой у взрослых). Снижение уровня фильтрации начинается с третьего десятилетия и неизменно продолжается в дальнейшем. 92 ГЛАВА 12. ОРГАНЫ ДЫХАТЕЛЬНОЙ СИСТЕМЫ Рис. 1. Эпителий слизистой оболочки воздухоносных путей. 1.Реснитчатая клетка. 2.Реснички. 3.Бокаловидная клетка (Бокаловидный экзокриноцит). 4.Эндокринная клетка. 5.Базальная (камбиальная клетка). 6.Собственная пластинка слизистой оболочки. Эпителий в разных отделах дыхательной системы имеет разное строение: многослойный ороговевающий - в верхних отделах, многорядный и однослойный реснитчатый – в дистальных отделах. Реснитчатые клетки (1) имеют на апикальной поверхности реснички (2), которые колеблются против движения вдыхаемого воздуха, в основе ресничек – аксонема (дуплеты микротрубочек, связанные динеиновыми мостиками). Ядра реснитчатых клеток в многорядном эпителии образуют самый верхний ряд. Клетки имеют рецепторы, в частности холинорецепторы, рецепторы глюкокортикоидов. Они могут изменять частоту биения ресничек, которые обеспечивают продвижение слизи с находящимися в ней частицами пыли, инородными телами от бронхов к трахее, глотке и т.д. Реснитчатые клетки могут секретировать вазо- и бронхоконстрикторы. Оптимальная температура для колебания ресничек от 18 до 33 градусов. Бокаловидные клетки (бокаловидные экзокриноциты) (3). Ядра в многорядном эпителии образуют средний ряд. Это клетки с хорошо развитой агранулярной ЭПС, комплексом Гольджи. Секретируют муцины – слизь, которая обволакивает частицы пыли и способствует их выведению. Плазматические клетки, находящиеся в собственной пластинке слизистой, вырабатывают иммуноголобулины класса А, которые также содержатся в слизи. Кроме того, в составе слизи может содержаться бактерицидный фермент лизоцим, сиаловая, гиалуроновая кислоты, которые обладают бактерицидным действием, гликозаминогликаны. В норме одна бокаловидная клетка приходится на 5-6 реснитчатых. Эндокринные клетки имеют одиночное расположение (4). В клетках хорошо развит синтетический аппарат. В их цитоплазме ближе к базальной поверхности находятся секреторные гранулы. Клетки синтезируют биогенные амины и пептиды (норадреналин, бомбезин, дофамин и др.), которые влияют на сокращение гладких мышечных клеток, находящихся в стенке дыхательных путей. Базальные клетки (5) небольших размеров, прилежат к базальной мембране. Их апикальная часть не доходит до апикальной поверхности других клеток. Способны к митотическому делению, являются стволовыми клетками. Их ядра образуют нижний ряд в многорядном эпителии. Более широкое основание располагается на базальной мембране. Являются источником регенерации. За счет активности этих клеток эпителий бронхов обновляется в течение 2 – 3 месяцев. Помимо вышеперечисленных элементов в составе эпителия верхних дыхательных путей присутствуют антигенпредставляющие клетки (клетки Лангерганса). Их относят к макрофагам. Это отросчатые клетки с органеллами общего значения. Вырабатывают цитокины, стимулируют Т-лимфоциты. Захватывая антигены вызывают аллергическую реакцию. Участвуют в обоазовании иммунной системы дыхательных путей. Микроворсинчатые клетки (каемчатые, щеточные) встречаются в дистальных отделах воздухоносных путей. На апикальной поверхности микроворсинки, а базальная поверхность контактирует с дендритами чувствительных нейронов. Клетки рассматриваются как рецепторы общей чувствительности. В мелких бронхах и бронхиолах присутствуют секреторные клетки Клара. Они вырабатывают ферменты, которые препятствуют слипанию бронхиол, принимают участие в обезвреживании токсинов. Собственная пластинка слизистой (6) образована рыхлой волокнистой соединительной тканью с находящимися лимфатическими узелками, кровеносными капиллярами. 93 Фрагмент какого органа на рисунке? Назовите структуры, обозначенные цифрами. Их строение, функция? Рис. 2. Фрагмент стенки альвеолы в легком. Аэрогематический барьер. 1.Сурфактант. 2.Альвеолярный макрофаг. 3.Альвеолоцит I типа. 4.Кровеносный капилляр. 5.Базальная мембрана. 6.Альвеолоцит II типа. 7.Эндотелиоцит кровеносного капилляра. 8.Просвет альвеолы. Респираторный отдел в легких представлен ацинусами, включающими респираторные бронхиолы, альвеолярные ходы, альвеолярные мешочки. Альвеолы представляют собой незамкнутые пузырьки. Фрагмент стенки альвеолы изображен на рисунке. В альвеолах происходит газообмен между воздухом альвеол и кровью. Ацинусы отделяются друг от друга соединительнотканными прослойками. Альвеолы разделены соединительнотканными перегородками с проходящими в них сосудами. На внутренней поверхности альвеол выделяют два вида клеток, которые располагаются на базальной мембране: альвеолоциты I типа (3) и альвеолоциты II типа (6). Альвеолоциты I типа (респираторные альвеолоциты) имеют уплощенную форму, цитоплазматические выросты, обращенные в сторону цитоплазмы. К безъядерным участкам этих клеток, лежащих на базальной мембране (5) прилежит базальная мембрана кровеносного капилляра с располагающимися на ней эндотелиальными клетками (4). Поэтому альвеолоциты I типа принимают участие в образовании аэрогематического барьера. Альвеолоциты II типа (секреторные) составляют 5%. Имеют кубическую форму, в цитоплазме присутствуют органеллы характерные для секреторных клеток. Кроме того, в цитоплазме есть «слоистые тельца» - цитофосфолипосомы, основным компонентом которых являются фосфолипиды. Являясь секреторными клетками, альвеолоциты II типа синтезируют поверхносто активные вещества (ПАВ), фосфолипиды, белки. ПАВ предотвращают спадение альвеол во время выдоха. Макрофаги (2) отросчатой формы с овальным ядром, с хорошо развитым лизосомальным аппаратом. Могут располагаться в стенке альвеолы или на поверхности альвеолы. Из альвеолы могут передвигаться в интерстициальную ткань. Эти клетки фагоцитируют частицы пыли, микроорганизмы, разрушенные клетки, сурфактант. Снаружи к альвеолам прилежат кровеносные капилляры (4), сеть эластических и коллагеновых волокон, тучные клетки и фибробласты. Внутренняя поверхность альвеол (над клетками) выстлана сурфактантом (1) в состав которого входят глицерофосфолипиды, холестерол, белки. Для образования сурфактанта его компоненты поступают в альвеолоциты II типа из крови. Удаление сурфактанта из альвеол происходит через лимфатическую систему, через бронхи, а также альвеолярными макрофагами. Сурфактантный альвеолярный комплес включает: 1.Мембранный компонент (построен как клеточные мембраны). Состоит из фосфолипидов и белков синтезируемых альвеолоцитами II типа. 2.Гипофаза - это жидкий компонент. состящий из липопротеинов и гликопротеинов. Вырабатывается секреторными клетками Клара. 3.Резервный сурфактант. Комплекс препятствует слипанию внутренней поверхности альвеол при выдохе; формирует противоотечный барьер (жидкость из интерстициальной ткани не попадает в просвет альвеолы); препятствует проникновению попавших в альвеолы микроорганизмов в интерстициальную ткань. Однако, сурфактант проницаем для газов. До рождения вырабатывается избыточное количество сурфактанта, которое после рождения удаляется альвеолярными макрофагами (2). Таким образом, в состав аэро-гематического барьера входят: 1.Сурфактант (1). 2.Цитоплазма альвеолоцитов I типа (безьядерные участки) (3). 3.Слившиеся базальные мембраны (5) с располагающимися альвеолоцитами 1 типа и базальная мембрана кровеносного капилляра. 4. Цитоплазма эндотелиальных клеток капилляров (7). 94