УЧЕБНИКИ И УЧЕБНЫЕ ПОСОБИЯ
ДЛЯ СТУДЕНТОВ ВЫСШИХ УЧЕБНЫХ ЗАВЕДЕНИЙ
В.Ф.ВРАКИН М.В.СИДОРОВА
МОРФОЛОГИЯ
СЕЛЬСКОХОЗЯЙСТВЕННЫХ
ЖИВОТНЫХ
(АНАТОМИЯ С ОСНОВАМИ ЦИТОЛОГИИ,
ЭМБРИОЛОГИИ И ГИСТОЛОГИИ)
Допущено Главным управлением высших учебных заведений при Государственной комиссии Совета Министров СССР по продовольствию и
закупкам в качестве учебника для студентов высших учебных заведений
по специальности «Зоотехния»
МОСКВА ВО АГРОПРОМИЗДАТ- 1991
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ББК 45.2 В 81
УДК 636:611(075.8)
Редактор Н. И. Емельянова
Рецензенты: профессор Г. М. Удовин (Оренбургский сельскохозяйственный институт), профессор А. П. Елисеев и доцент Ю. А. Селиверстов (Санкт-Петербургский государственный аграрный университет)
Вракин В. Ф., Сидорова М. В.
Морфология сельскохозяйственных животных; Анатомия с основами
цитологии, эмбриологии и гистологии. — М: Агропромиздат, 1991. — 528 с,
[4] л. ил.: ил. (Учебники и учеб. пособия для студентов высш. учеб. заведений).
ISBN 5—10—000675—7
В книге приведены сведения по биологии клетки, основам эмбриологии,
гистологии, морфологии органов и систем: опорно-двигательного аппарата,
кожного покрова, пищеварительной, дыхательной, мочеполовой, сердечнососудистой и нервной систем, желез внутренней секреции, органов чувств.
Представлено научное обоснование изменений анатомической и гистологической структуры органов и их систем в зависимости от кормления и содержания, а также даны технологические приемы в промышленном животноводстве.
Для студентов по специальности «Зоотехния».
3705010000—332
В-261-91 ББК 45.2
035(01)—91
ISBN 5—10—000675—7
© В. Ф. Вракин, М. В. Сидорова, 1991
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Понятие о морфологии, ее предмет и методы изучения.
Морфология (греч. morphe — форма, logos — учение) —наука о форме и
строении организмов, является разделом биологии (греч. bios — жизнь) —
науки о жизни во всех ее проявлениях. Морфология животных тесно связана
с зоологией, физиологией и экологией. Предметом изучения морфологии животных как науки является внешняя форма животных организмов, их органов, топография, внешнее и внутреннее строение органов и систем органов,
их тканевая организация и, наконец, строение клеток и неклеточных структур, входящих в состав органов и организма в целом. Задача морфологии —
познание статики и динамики формы на всех ее уровнях.
Морфология имеет несколько направлений: функциональная морфология — исследует строение органов в процессе их функционирования и изменения в структуре органов с изменением их функциональной активности;
экологическая (греч. oikos — родина, место обитания) морфология — изменения в строении органов под влиянием внешних условий; возрастная морфология — преобразования в строении органов в процессе индивидуального
развития особи; эволюционная морфология — изменения в строении органов
и их систем в ряду животных форм по мере усложнения их организации в
процессе исторического развития (в эволюции).
Термин «морфология» был предложен в 1817 г. великим немецким поэтом и естествоиспытателем Иоганном Вольфгангом Гёте, которую он определил как «учение о форме, образовании и преобразовании органических существ».
Учение о внешней форме органов и их внутреннем строении, видимым
невооруженным глазом, называется анатомия (греч. anatome — рассечение,
разрез). Областью анатомии, изучающей органы в их естественной морфологической и функциональной связи в виде систем органов, является описательная, или системная, анатомия. Исследованием внешней формы тела,
соотношением и пропорцией его частей в статике и динамике занимается в
основном пластическая анатомия. Топографическая анатомия описывает
расположение и пространственные взаимоотношения органов в различных
участках, частях и областях тела. Различия в форме, строении и расположении одноименных органов и систем у разных пород, видов, семейств, классов
животных изучает сравнительная анатомия. С изобретением микроскопа
стали развиваться такие науки, как микроскопическая анатомия (греч. micros
— малый, scopeo — вижу) или частная гистология — наука о строении деталей органов, невидимых невооруженным глазом, общая гистология (греч. histos—ткань)—наука о тканях живого организма, эмбриология (греч. embryon
— зародыш) —наука о развитии зародыша и плода, цитология (греч. cytos —
сосуд, клетка) — наука о строении и жизнедеятельности клетки.
Объектом изучения морфологии сельскохозяйственных животных являются все виды домашних млекопитающих, птиц и рыб, используемых в сельском хозяйстве. Наиболее изучены лошадь, свинья, рогатый скот, курица.
Современная морфология исследует не только строение организма взрослого
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
животного, но и изменения строения в процессе их роста, развития, селекции,
под влиянием условий кормления и содержания.
Развитие науки во многом зависит от методов исследования. Так, в анатомии основным методом, сохранившимся по настоящее время и отраженным в ее названии, считается рассечение, препарирование трупов с помощью
различных инструментов. На живых объектах многие детали строения изучаются с помощью осмотра, ощупывания (пальпации), измерения. Разработка
метода наливки сосудов привела к появлению коррозионных методов исследования. Предложение Н. И. Пирогова изучать взаиморасположение органов
и их частей на замороженных трупах способствовало накоплению сведений
по топографической анатомии. Рентгенография дала возможность изучать
строение скелета и полостных органов (при наполнении контрастирующим
веществом) на живом животном.
Первоначальными методами гистологии и эмбриологии были раздавливание, препаровка, мацерация и расщипывание иглами. Усовершенствование
микроскопической техники, применение красителей и других веществ, дающих цветные продукты химических реакций, привело к бурному развитию
этих наук. Сейчас они располагают богатым арсеналом методов исследования. Микроскопический, или светооптический, фазово-контрастный, люминесцентный, поляризационный методы применяют для изучения тонкого
строения тканей, органов, клеток на окрашенных и неокрашенных препаратах с помощью светового микроскопа в проходящем, отраженном или смешанном свете; ультрамикроскопический— для субмикроскопического строения тканей, органов и клеток с помощью электронного микроскопа; гистохимический, дифференциальное центрифугирование, радиоавтографический—
для функциональной активности структур с помощью модифицированных
(специфических, приспособленных) химических методов; культура тканей,
цейтраферная микрокиносъемка, витальная окраска, пересадка органов и
тканей — для прижизненных исследований структур. Существует макромикроскопический, или субмакроскопический, метод исследований, находящийся на стыке анатомии и гистологии, когда объект исследования слишком
велик для сильных увеличений и трудноразличим невооружейным глазом. При этом методе пользуются лупами или микроскопом с небольшим увеличением, в том числе стереоскопическим, что позволяет видеть мелкие структуры объемными.
Краткие сведения из истории морфологии. Зачатки анатомических
сведений возникли в глубокой древности: среди наскальных древнепалеолитических рисунков есть изображения внутренних органов и позвоночного столба. Цивилизация Древнего Востока сделала значительный шаг
вперед в познании природы, пытаясь объяснить окружающий мир. В египетском папирусе Эберса (XVI в. до н. э.) содержится довольно подробная анатомическая терминология. Накопленные знания оказали воздействие на науку античной Греции и Рима. Основоположником анатомии и физиологии в
Древней Греции считают врача и философа Алкмеона Кротонского (конец VI
— начало V в. до н. э.). Он первый начал анатомировать трупы животных для
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
научных целей и сделал ряд важных выводов о роли органов. В это время закладываются основы материалистической философии (Анаксагор, Демокрит). Современником и последователем Демокрита был величайший врач античности Гиппократ (460—377 гг. до н. э.). Вследствие запрета вскрывать человеческие трупы Гиппократ и его ученики изучали строение тела и органов
на животных. Лучше всего ими были изучены скелет, сердце, внутренние органы. В IV в. до н. э. Аристотель охватил весь круг биологических знаний того времени и по праву считается основателем зоологии, сравнительной анатомии и эмбриологии. Однако дуалистические (двойственные) философские
воззрения Аристотеля, противопоставление «пассивной» материи активному
«перводвигателю» в дальнейшем сыграли отрицательную роль в развитии
биологии.
В III—I вв. до н. э. центр науки и культуры перемещается в Александрию. Там работали знаменитые врачи Герофил и Эразистрат, которые далеко
продвинулись в изучении нервной системы.
После завоевания Александрии римлянами центр культуры переместился в Рим, где в I в. н. э. работал великий врач, анатом и физиолог Клавдий
Гален (130—200). Он изучал анатомию овец, быков, собак, свиней, медведей,
обезьян и других млекопитающих. Подметив сходство в строении человека и
обезьяны, он много внимания уделил изучению ее нервной системы, мышц,
костей и суставов, что потом было принято за описание человеческого тела.
Его авторитет в науке был столь велик, что считался непререкаемым на протяжении 1500 лет, вплоть до эпохи Возрождения, что в конце концов стало
являться тормозом, так как вместе с правильными воззрениями удерживались, канонизировались и ошибочные взгляды Галена — он утверждал,
что по дыхательным путям в сердце поступает воздух, а кровь переходит из
одного желудочка в другой через отверстие в перегородке.
Новый очаг культуры возник в средние века в государствах Передней и
Средней Азии. Особенно большой вклад внес таджикский ученый Ибн-Сина
(Авиценна, 980—1037). Его многотомный «Канон врачебной науки» содержит массу сведений анатомического и физиологического характера.
Начало нового этапа в развитии анатомии приходится на эпоху Возрождения (XV—XVII вв.), когда в рамках феодального общества начинается вызревание капиталистических общественных отношений. Быстрый рост производительных сил, разностороннее использование природы, географические
открытия, новые рынки и расширение торговли — все это изменило экономику Европы и привело к перевороту в общественной жизни. Это была эпоха
культурного подъема, бурного развития науки, философии, искусства. Был
сломлен многовековой запрет на анатомирование человеческих трупов, и
единая до тех пор анатомия стала разделяться на медицинскую и ветеринарную. В XVI—XVII вв. наибольшие успехи делали анатомия человека, так как
она была тесно связана с практической медициной. Великий итальянский
анатом Андеас Везалий (1514—1564) пошатнул традиционную веру в неколебимость авторитетов Аристотеля и Галена. Вскрывая трупы людей, он разработал и довел до совершенства технику препаровки, получил массу новых
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
фактов о строении организма, пересмотрел анатомическую терминологию и
отразил все это в фундаментальном труде «Семь книг о строении человеческого тела» (1543).
В эпоху Возрождения интенсивно развивается и животноводство, стимулируя развитие биологии, в том числе анатомии животных. Преимущественное влияние уделялось лошади как основному средству передвижения,
тягловой силе в сельском и городском хозяйстве, в военном деле. Еще в XIII
в. в книге Дж. Руффо по коневодству есть сведения об экстерьере и анатомии
лошади. В XVI в. анатомию лошади изучали Леонардо да Винчи, К. Руини
(1598). В XVII в. Марко Аурелио Северино (1580—1656) написал «Демокритову зоотомию», труд, в котором он сравнивает строение мускулатуры, органов пищеварения и размножения у человека и животных, в том числе лошади. Во Франции в 1761 г. в Лионе и в 1765 г. в Альфорте К. Буржелем
(1712—1779) открыты ветеринарные школы. Им же написаны первые учебные руководства по ветеринарии, в том числе и по анатомии лошади (1766,
1769). Ф. Лафосс (1739—1820) в «Курсе гиппиатрии» также уделил внимание
анатомии лошади. Более полное руководство «Анатомия млекопитающих
животных» было написано Иозефом (1788). Практически ни один анатом не
обошел своим вниманием животных, изучая для тех или иных целей строение их органов, часто с точки зрения сравнения их с органами человека или
другого животного. Элементы сравнительной анатомии прослеживаются в
трудах анатомов, начиная с Гиппократа и Аристотеля, но только в XVII в.
можно говорить о зарождении сравнительной анатомии как раздела науки.
Сам термин «сравнительная анатомия» в его современном понимании был
введен Томасом Виллисом в «Анатомии мозга» (1666).
Сведения из эмбриологии также восходят ко времени античности, накапливаются в течение всего времени, но вплоть до XVIII в. содержат много натяжек, домыслов и ошибок и не сведены в единую систему, чтобы можно
было говорить об эмбриологии как науке. Вопросы зарождения и развития
волновали умы анатомов. Еще Гарвей выдвинул тезис: «Все живое из яйца»
(«О зарождении животных», 1651). В XVII в. к разгадке этой тайны ближе
других подошли Стеной, обнаруживший фолликулы в яичниках у акул, и Ренье де Грааф, описавший фолликулы в яичниках млекопитающих.
С изобретением микроскопа границы морфологических исследований
значительно расширились. Приоритет в изобретении микроскопа по разным
источникам принадлежит Г. Галилею (1609 г., Испания), Дребблю (1617—
1619 гг., Франция), отцу и сыну Янсенам (Голландия). В дальнейшем микроскоп был усовершенствован А. Левенгуком (1632—1723) и позволял увеличивать объект до 200 раз. Он описал сперматозоиды как мужские оплодотворяющие элементы, видел клетки, назвал их пузырьками, но не придал им
значения, открыл инфузории, саркодовые, бактерии и объединил их под общим названием анималькули — зверьки. Английский ученый Роберт Гук
(1635—1703) наблюдал строение растительных тканей и дал современное название их структурам «клетка», хотя и трактовал клетки как ячейки, пустоты
между волокнами («Микрография или некоторые физиологические описания
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мельчайших телец при помощи увеличительных стекол», 1665). Английский
натуралист Неэмия Грю (1641—1712) при описании растений («Анатомия
растений», 1682) установил понятие «ткань» как структуру.
Таким образом, к концу XVII в. морфология оформилась в общих чертах
как описательная наука о строении организмов и в ее недрах вызрели физиология, эмбриология и гистология, хотя еще и не оформились в самостоятельные науки. На протяжении XVIII—XIX вв. происходит дальнейшее
накопление фактов и поступательное развитие морфологии в связи с потребностями развивающихся производительных сил и совершенствованием приборов и методов исследования.
В развитии морфологической науки большую роль сыграли труды Ж.
Кювье (1769—1832)—основателя палеонтологии, одного из ведущих анатомов своего времени и Этьена Жоффруа Сент-Илера (1772—1844) — видного
французского анатома, много сделавшего для развития сравнительной анатомии, одного из основоположников теории «типа». Успехи микроскопической техники резко увеличили разрешающую способность прибора и
привели к бурному накоплению фактического материала в гистологии. Термин «гистология» ввел немецкий анатом Карл Майер в 1819 г.
Сравнительная анатомия достигла такого развития, что ее данные позволили сделать обобщающие выводы о том, что тело животных построено по
единому плану и обосновать так называемую теорию «типа» (А. МильнЭдвардс) и учение о гомологии — сходстве органов, имеющих общее происхождение (Р. Оуэн). Однако типы рассматривались как статичные «планы
строения», исторически не связанные между собой, а гомологичные сегменты сводились к позвонку, из которого выводились все детали скелета. Широко обсуждались вопросы происхождения головы (И. В. Гёте и А. Окен) и конечностей у позвоночных животных (К. Гегенбаур). Создавались различные
искусственные классификации живых существ (К. Линней, Ш. Бонне и др.).
В эмбриологии господствовала теория преформизма — теория предобразования, по которой все зародыши всех будущих поколений вложены как
матрешки в исходный организм и при развитии лишь увеличиваются в размерах (Ш. Бонне, Эразм Дарвин и др.).
С развитием общественной и философской мысли в XIX в. обнаруживалось все большее несоответствие между господствующей в морфологии телеологической антиэволюционной концепцией об изначальном целесообразном назначении органа и накапливающимися фактами. В науке все шире
распространялись эволюционные идеи (Э. Ж. Сент-Илер, Ж. Б. Ламарк, К. Ф.
Вольф, К. Ф. Рулье). Переворот был совершен книгой Ч. Дарвина «Происхождение видов путем естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь», вышедшей в 1859 г. Широкое распространение эволюционных идей Дарвина вдохнуло новые силы в морфологическую науку. Наиболее подготовленной для восприятия дарвиновского
учения оказалась русская наука (В. О. и А. О. Ковалевские, И. И. Мечников,
А. Дорн, Н. Клейненберг и др.).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Большинство теорий не только обрело новое звучание, но и наполнилось
новым содержанием. Типы стали восприниматься как выражение главных
направлений эволюционного процесса (Н. А. Холодковский, 1909), исторически связанных между собой. «Проблема головы» получила свое решение в
теории метамерии головы. Плодотворной оказалась гипотеза происхождения
пятипалой конечности из плавника, выдвинутая К. Гегенбауром и развитая
впоследствии А. И. Северцовым. В эмбриологии теория эпигенеза — теория
развития зародыша из недифференцированной закладки, сторонниками которой являлись К- Ф. Вольф, X. И. Пандер, К. М. Бэр, благодаря импульсу,
данному ей эволюционным учением, получила дальнейшее развитие в трудах
А. О. Ковалевского, И. И. Мечникова, О. и Р. Гертвигов, Н. Ф. Кащенко и
других и преобразовалась в эволюционную сравнительную эмбриологию.
Эволюционный принцип, привнесенный в сравнительную морфологию,
позволил переосмыслить накопленные ею факты и заложить фундамент филогенетической морфологии. Одним из первых морфологов, который, опираясь на учение Дарвина, пришел к выводу, что система животного царства
должна строиться на генеалогическом принципе (принципе происхождения),
был Э. Геккель (1834—1919). Он внес в морфологию много новых терминов,
укрепившихся в ней: онтогенез — индивидуальное развитие, филогенез —
историческое развитие вида и др. Филогенетическое направление в морфологии оказалось чрезвычайно плодотворным, способным объяснить причины
конкретных изменений формы и функции. Развиваясь в трудах В. О. Ковалевского (1842—1883), А. Дорна (1840—1909), Н. Клейненберга (1842—
1897) и др., оно превратилось в самостоятельное направление в морфологии
— филогенетику или эволюционную морфологию.
Одним из важных вкладов в биологические науки XIX в. стала клеточная теория, подготовленная работами многих микроскопистов (Р. Броун, Б.
Дюмортье, М. Шлейден) и особенно знаменитого чешского гистолога Яна
Эвангелисты Пуркине (1787—1869) и его школой (Г. Валентин, 1810—1883.
и др.) и сформулированная в 1838—1839 гг. Теодором Шванном (1810—
1882) в труде «Микроскопические исследования о соответствии в структуре
и росте животных и растений».
Развитие морфологии в России, как и на Западе, совпадало с развитием
образования, организацией университетов и медицинских школ. В 1658 г.
Епифаний Славинецкий перевел трактат А. Везалия о строении человеческого тела («Эпитоме»). Значительный шаг в развитии просвещения и науки был
сделан во времена Петра I (1672—1725). В 1699 г. в Москве был организован
курс лекций по анатомии, в 1707 г. открыта первая госпитальная школа в
Москве, а затем Петербурге, Кронштадте и других городах, в 1724 г. — Петербургская академия, в 1755 г. — Московский, в 1819 г. — Петербургский
университеты. В России в 1733 г. на 30 лет раньше, чем на Западе, открыта
первая ветеринарная школа, где стали готовить специалистов для работы на
конных заводах и в армии. В конце XVIII в. в России курс ветеринарной анатомии преподавался в Московском, Харьковском и Казанском университетах.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Первая отечественная анатомическая школа в России основана П. А. Загорским (1764—1846) при Петербургской медико-хирургической академии
(ныне Военно-медицинская академия им. С. М. Кирова). Его ученик И. Д.
Книгин (Булгаков) в 1808 г. организовал кафедру анатомии домашних животных при ветеринарном отделении академии. В 1811 г. организована кафедра домашних животных в Московской медико-хирургической академии.
Создаются отечественные учебники по ветеринарной анатомии. Доктор медицины прозектор Московского университета И. С. Андреевский (1751 —
1809) издал в 1804 г. «Краткое начертание анатомии домашних животных
для употребления учащихся медицины, ветеринарии в Императорском Московском университете». В 1837 г. А. И. Кикин (1810—1852), руководитель
кафедры анатомии Московского университета, выпустил «Краткую зоотомию или руководство к познанию строения тела домашних животных» —
двухтомное руководство, отражающее уровень знаний по анатомии того времени в сравнительном и функциональном аспектах. Профессор Петербургской медико-хирургической академии В. И. Всеволодов (1790—1863) издал в
1846— 1847 гг. двухтомную «Анатомию домашних животных, преимущественно млекопитающих», в которой изложены сведения о скелете, мышцах и
внутренностях. Его ученик и преемник А. О. Стржедзиньский (1823—1882)
написал «Анатомию домашних животных (млекопитающих и птиц)». В 1873
г. он организовал кафедру анатомии в Казанском ветеринарном институте. А.
С. Измайлов (1833—1901), заведующий кафедрой анатомии Петербургской
медико-хирургической академии издал в 1879 г. «Анатомию домашних животных». После него кафедрой заведовал Э. К. Брандт (1839—1891)—автор
учебника «Анатомия домашних животных», вышедшего в двух изданиях.
Профессор Петровской сельскохозяйственной академии (ныне Московская сельскохозяйственная академия им. К- А. Тимирязева) Н. П. Чирвинский
(1848—1920) экспериментально доказал зависимость развития и строения
организма (скелета, органов пищеварения) от режима питания и других
внешних факторов, положив этим начало экспериментальной и экологической ветеринарной морфологии.
Первая русская гистологическая школа была основана А. И. Бабухиным
при Московском университете, где в 1869 г. он возглавлял кафедру гистология. Русские гистологи большое внимание уделяли изучению нервной системы. Морфология XX в. характеризуется углублением тех проблем, которые
разрабатывались в эпоху утверждения принципов дарвинизма, и появлением новых направлений, поставивших новые проблемы. Существенные успехи связаны с деятельностью русских и советских исследователей. Особенно велико значение их работ для развития эволюционной морфологии (А. Н.
Северное, В. М. Шимкевич, Н. А. Холодковский, П. П. Иванов, П. Г. Светлов, Г. А. Шмидт и др.), эволюционной гистологии (В. Н. Беклемишев, А. А.
За-варзин, Н. Г. Хлопин и их ученики), эмбриологии (Д. П. Филатов), закономерностей роста (И. И. Шмальгаузен), регенерации (М. А. Воронцова), культивирования тканей (Ф. М. Лазаренко) и постэмбрионального развития (М.
М. Завадовский). Обосновав морфологические закономерности эволюции, А.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Н. Северцов создал морфобиологическую теорию эволюции и теорию филэмбриогенеза, объясняющие пути эволюционного процесса. Творческое
применение принципов эволюции в гистологии позволило создать А. А. Заварзину теорию параллелизма гистологических структур (1934—1945), а Н.
Г. Хлопину — теорию дивергентной эволюции тканей (1946).
При изучении закономерностей роста и развития ученые на протяжении
веков сталкивались с проблемой взаимосвязи формы и функции. Основоположником функциональной морфологии в России по праву считают П. Ф.
Лесгафта (1837—1909), идеи которого отражены в фундаментальном труде
«Основы теоретической анатомии» (1892). Ф. Энгельс, стоя на позициях диалектико-материалистической философии, определил их единство: «Вся органическая природа является одним сплошным доказательством тождества или
неразрывности формы и содержания. Морфологические и физиологические
явления, форма и функция обусловливают взаимно друг друга» («Диалектика
природы»). Именно так понимали А. Н. Северцов, И. И. Шмальгаузен и их
ученики и последователи взаимосвязь формы и функции и говорили о морфофизиологических преобразованиях организма.
Изучение объектов биологии на стыке морфологии и физиологии привело к появлению таких направлений, как сравнительная физиология, топография функций, экспериментальная морфология (механика и физиология развития), функциональная анатомия, гистофизиология. С изобретением Руска
электронного микроскопа (в 1924) стало ясно, что форма и функция сливаются воедино на молекулярном уровне изучения строения клетки и ее жизнедеятельности. В. гистологии кроме описательного и экспериментального (Б.
В. Кед-ровский, П. А. Ковальский и др.) подходов сформировались эволюционный (А. А. Заварзин, Н. Г. Хлопин), экологический (Н. Л. Гербильский),
генетический (С. Г. и М. С. Навашины, А. А. Прокофьева-Бельговская). При
изучении эволюции тканей гистологи основывались на эмбриологическом
принципе (В. М. Шимкевич, В. П. Михайлов) и гистогенетических корреляциях (И. И. Шмальгаузен, А. Н. Студитский). Столь разнообразные углубленные и комплексные подходы позволили решить ряд кардинальных проблем
гистологии и цитологии: механизмы самовоспроизведения, передачи энергии
в сопряженных реакциях и запасание ее в макроэргических соединениях;
развитие по определенной программе в онтогенезе; матричный принцип хранения и передачи наследственной информации.
В ветеринарной анатомии выдающейся личностью был профессор Леонид Аполлонович Третьяков (1856—1922)—заведующий кафедрой анатомии
Казанского ветеринарного института, который сделал ее центром по подготовке научных кадров.
Из казанской школы ветеринарных анатомов вышли также видные ученые и создатели своих анатомических школ, как А. Ф. Климов (1878— 1940),
Д. М. Автократов (1868—1953), Н. А. Васнецов (1901—1961), А. И. Акаевский (1895—1982). Их труды и труды их учеников и последователей направлены в основном на изучение нервной, сердечно-сосудистой систем и аппаwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рата движения в сравнительно-анатомическом, возрастном, функциональном
и экологическом аспектах, а также в условиях эксперимента.
Наиболее многочисленной была школа А. Ф. Климова: профессора А. И.
Акаевский, Н. И. Акаевский, В. В. Александровский, А. В. Арсеньев, Б. В.
Богородский, П. А. Глаголев, Д. И, Заверткин, С. Б. Иванов, И. Ф. Мокрое, О.
Е. Пахоменко, П. П. Преображенский, С. М. Смиренский, И. А. Спирюхов.
Изученные этой школой особенности внутренней структуры мышц млекопитающих в связи с функцией и развитием получили всеобщее признание,
имеют теоретическое и практическое значение. Под руководством П. А. Глаголева велись комплексные работы по выяснению морфохимических особенностей мускулатуры и скелета сельскохозяйственных животных в возрастном, экологическом аспектах и в зависимости от принадлежности к тому
или иному морфофункциональному типу.
Морфологи большое внимание уделяют созданию учебников и учебных
руководств по анатомии, гистологии и эмбриологии для ветеринарных, зооин-женерных и агрономических факультетов вузов и техникумов: учебники
Д. М. Автократова, А. Ф. Климова, А. И. Акаевского, Ю. Т. Техвера, И. Ф.
Иванова и П. А. Ковальского, П. А. Глаголева и В. И. Ипполитовой, Г. С.
Крок, Г, Г. Тинякова прошли проверку временем и неоднократно переиздавались.
Современное промышленное животноводство с его специфическими
чертами: концентрация животных на ограниченной территории, высокий
уровень механизации, поточный характер производства, стабилизация сбалансированного кормления, микроклимата, условий содержания, высокая интенсивность эксплуатации животного приводят к несоответствию между
биологией животного и условиями новой технологической среды. В результате наблюдается частое нарушение воспроизводительных функций, сокращение сроков жизни, нарушения обмена веществ, рождение слабого потомства, заболевания конечностей, молочной железы, что отрицательно влияет
на внедрение индустриальных технологий, снижает их эффективность. В
этом свете перед животноводами стоит основная задача — создание таких
животных, которые сочетали бы в себе высокий генетический потенциал
продуктивности со способностью к его реализации в условиях промышленных технологий. Для морфологов это выливается в определение видовой, породной, возрастной, типовой нормы строения, в расшифровку (совместно с
физиологами, экологами, биохимиками, генетиками) сложных морфофункциональных комплексов, обусловливающих определенные продуктивные
признаки, в выявление структур и качеств, которые определяют уровень развития полезных признаков в конкретных условиях среды, особенно «в отношении таких свойств домашних животных, как молочность, мясность, рабочие качества, шерстность — признаков, находящихся во взаимодействии со
множеством свойств, со всей конституцией породы» (Н. И. Вавилов),
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Раздел первый.
ОСНОВЫ цитологии
Глава 1. ПОНЯТИЕ О КЛЕТКЕ, КЛЕТОЧНАЯ ТЕОРИЯ
Клетка (греч. — cytos, лат.— cellula)—элемент или участок протоплазмы
(protos — первый, первичный, plasma — нечто оформленное), отграниченный
оболочкой (плазмолеммой). Это основная форма организации живой материи, является целостной живой системой. Состоит она из ядра, цитоплазмы и
плазмолем-мы (цитолеммы), взаимодействие которых определяет ее жизненность, проявляющуюся в обмене веществ, росте, раздражимости, сократимости и размножении. Клетка — высокоорганизованная структура, длительность жизни или жизненный цикл которой определяется многими факторами и зависит от того, какой ткани она принадлежит: например, клетки крови, покровных эпителиев живут от нескольких часов до нескольких дней, а
нервные клетки могут жить в течение всей жизни индивидуума. Жизнь молодой малодифференцированной клетки часто завершается не гибелью, а делением с образованием двух дочерних клеток, и тогда говорят о митотическом
цикле. В процессе развития большинство клеток организма приобретает специализацию — дифференцируется и выполняет строго определенную функцию (вырабатывает тот или иной секрет, всасывает питательные вещества,
переносит кислород и др.). Дифференцированные клетки, как правило, теряют способность к размножению или она резко снижается. Пополнение клеток
осуществляется с помощью стволовых или камбиальных, обнаруживаемых в
большинстве тканей. Это малодифференцированные клетки, функцией которых и является размножение. Дифференцированные клетки отличаются друг
от друга формой, величиной, внутренним строением, химическим составом,
направленностью обмена веществ, выполняемыми функциями.
В сложном многоклеточном организме кроме клеток существуют и неклеточные образования, однако это или производные клеток, или продукты
их деятельности. Наиболее распространенный продукт деятельности клеток
— межклеточное вещество, которое существует в виде волокон и аморфного — основного вещества. Производные клеток — синцитии и симпласты.
Симпласты — это крупные образования со множеством ядер, не разделенные на отдельные клеточные территории. Симпластами считаются мышечные волокна, один из слоев плаценты. Синцитии, или соклетия, —
образования, состоящие из клеток, соединенных между собой цитоплазматическими мостиками. Встречаются они при развитии сперматогенного эпителия. Изучением развития, строения, размножения и функционирования клетки занимается наука цитология.
В организме клетки объединены в ткани и органы —сложные, целостные системы, связанные межклеточными взаимодействиями и подчиненные
нейрогуморальной регуляции со стороны нервной, кровеносной и эндокринной систем. Поэтому организм является единой системой, качественно отличимой от суммы клеток, его составляющих.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Клеточная теория. Мысль о существовании элементарных единиц, из
которых состоят растения, животные и человек, появилась еще в глубокой
древности. В разные эпохи эти единицы трактовались по-разному (у Демокрита — это атомы; у Аристотеля — однородные и неоднородные части тела;
у Гиппократа и Галена — четыре первичные жидкости: кровь, слизь, черная
и желтая желчь; у Окена — органические кристаллы или инфузории и т. д.).
Однако это были умозрительные заключения, и только с изобретением микроскопа естествоиспытатели воочию убедились в существовании элементарных единиц, образующих живые тела.
Впервые клетки обнаружил английский ученый Роберт Гук (1635—
1703) при изучении среза пробки с помощью сконструированного им микроскопа, который увеличивал объект в 100 раз, и описал это в сочинении
«Микрография, или некоторые физиологические описания мельчайших телец, осуществленные посредством увеличительных стекол», вышедшем в
1665 г. Он же дал и названия обнаруженным им структурам — клетки, так
как трактовал их как пустоты, поры между растительными волокнами. Эту
дату можно считать временем рождения цитологии. Современники Гука М.
Мальпиги, Н. Грю, А. Левенгук подтвердили наличие структур, подобных
клеткам, но каждый из них называл их по своему: «пузырьки», «мешочки».
На протяжении XVII—XVIII вв. в цитологии происходит накопление
материала, часто разрозненного, противоречивого, с ошибочным трактованием фактов. Но время и опыт отбирают ценное, отбрасывая ошибочное и постепенно вырисовывалось истинное строение элементарных единиц. В конце
XVIII — начале XIX в. появляются попытки объяснения и обобщения накопленного материала. Сравнение тонкой структуры растений и животных наводило на мысль об их схожести (К. Вольф, Лоренц, Окен и др.). Идеи общности микроскопической структуры растений и животных витали в воздухе. В
1805 г. Г. Тревиранус, в 1807 г. Г. Линк показали, что растительные клетки
это не пустоты, а самостоятельные замкнутые образования. В 1831 г. Р. Броун доказал, что ядро является обязательной составной частью растительной
клетки, а в 1834 г. о том же заявили Я. Пуркинё и Г. Валентин применительно к животной клетке. Особенно большой вклад в учение о клетке внесли две
научные школы: И. Мюллера (1801—1858) в Берлине и Я. Пуркине (1787—
1869) в Бреславле. Ученик И. Мюллера Теодор Шванн (1810—1882) блестяще сопоставил литературные данные и собственные наблюдения, результатом чего явилась книга «Микроскопические исследования о ^соответствии в
структуре и росте животных и растений» (1839), в которой он доказывал, что
клетка — всеобщая элементарная единица, присущая обоим царствам организмов (животным и растительным), а процесс клеткообразования — это
универсальный принцип развития. Наблюдения Шванна были подчинены
общей идее, что дало возможность представить их в виде биологической теории, содержащей три главных обобщения: теорию образования клеток, доказательства клеточного строения всех органов и частей организма, распространение этих двух принципов на рост и развитие животных и растений.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Клеточная теория имела «революционизирующее» (Энгельс) влияние на
развитие биологии середины XIX в., обосновав идею единства живой природы, показав морфологическую основу этого единства. В числе других факторов она позволила Ч. Дарвину сделать допущение, что все животные и растения происходят от общего корня. Распространенная Р. Вирховом на область
патологии, она стала основной теоретической базой для понимания причин
болезней. Клеточная теория Шванна, несмотря на глубоко прогрессивный
характер, не была лишена ошибок, за которые неоднократно подвергалась
критике. Так он считал, что клетка — это автономная элементарная единица,
а организм — лишь сумма клеток.
В конце XIX — первой половине XX в. вокруг клеточной теории развернулась оживленная дискуссия, в процессе которой происходило критическое
переосмысление ее основных положений. Подытоживая результаты данного
обсуждения, П. И. Лаврентьев писал: «Очищенная от метафизической шелухи, от персонификации клеток, от аналогии с государством, от сведения к
элементарным составным частям, теория клеточного строения растений и
животных остается и останется одним из величайших и плодотворнейших завоеваний биологии».
В современной клеточной теории отражено все лучшее, что было достигнуто учеными прошлого. Углублены и расширены представления о клетке
на основе последних достижений науки в свете материалистического мировоззрения и диалектического подхода к строению и развитию организма.
Биология клетки накопила богатый материал, позволяющий глубже разобраться в жизнедеятельности клетки, ее строении, развитии и значении. Основные положения современной клеточной теории можно свести к следующему.
1. Клетка лежит в основе строения всех многоклеточных организмов.
Клетки всех организмов, несмотря на их различия, имеют общие принципы
строения и образуются в результате деления.
2. Клетка — основная, но не единственная форма организации живой
материи. Наряду с ней существуют доклеточные формы (бактериофаги, вирусы), а у многоклеточных организмов — неклеточные живые образования
(волокна, межклеточное вещество и др.).
3. Клетка, обладающая большой сложностью строения, имеет длительную историю развития, свой филогенез. Она возникла на определенной ступени развития органической материи из более простых форм.
4. Клетка имеет индивидуальную историю развития, свой онтогенез, в
процессе которого клетка многоклеточного организма изменяется, развивается, приобретает новые качества. Онтогенез клетки подчинен онтогенезу организма.
5. Клетка — часть многоклеточного организма, и ее развитие, форма и
функция зависят от всего организма. Функция организма не является суммой
функций отдельных клеток. Это качественно новое явление.
6. Возникновение клеточного строения сыграло в эволюционном процессе очень важную роль, дало большие преимущества многоклеточному орwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ганизму, в связи с чем явилось главным направлением в эволюции как растений, так и животных: а) расчленение на клетки создало значительно большую
поверхность клеточных мембран, что, в свою очередь, коренным образом изменило ход и уровень обменных процессов, увеличило жизнедеятельность
организмов, б) привело к гораздо более глубокой структурной дифференцировке, чем у неклеточных организмов (например, у сифонофор). Благодаря
этому возросла специализация клеток, которая сильно повысила приспособляемость организмов к среде существования, в) Только клеточное строение
дало возможность развития крупных форм животных и растений. Увеличение размеров тела позволило освоить новые условия существования и обеспечило прогрессивную эволюцию органического мира, г) Клеточное строение облегчает обновление, замену изношенных и патологически измененных
частей тела.
Вопросы для самоконтроля. 1. Что такое клетка? Какое значение для
развития биологии имела клеточная теория? 3. В чем механистичность, ошибочность клеточной теории Шванна? 4. Перечислите и раскройте основные
положения современной клеточной теории.
Глава 2. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА И МОРФОЛОГИЯ
КЛЕТКИ
ХИМИЧЕСКИЙ СОСТАВ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА
ПРОТОПЛАЗМЫ
Элементарный состав протоплазмы. Протоплазма — содержимое живой
клетки, включая ее ядро и цитоплазму. В ее состав входят практически все
химические элементы, но распространение их не совпадает с распространением в неживой природе. В земной коре больше всего О, Si, Al, Na, Са, Fe,
Mg, Р (99 %). Основными элементами любой структуры живого вещества являются С, О, N и Н. Имеют немаловажное значение S, Р, К, Са, Na, CI, Fe, Си,
Mn, Zn, I, F. В организме данные элементы распространены неравномерно:
например, в костях много Са и Р, в щитовидной железе — I. В зависимости
от количества их делят на макроэлементы, микроэлементы и ультрамикроэлементы. Микро- и ультрамикроэлементы необходимы для жизни и деятельности клетки, как и макроэлементы, хотя и действуют в ничтожно малых
количествах (10-8—10~12%). Как правило, микроэлементы входят в состав
биологически активных веществ — гормонов, витаминов, ферментов, определяя их специфическую активность. Конечно, не все элементы имеются в
каждой клетке. Клетки отличаются как количеством, так и составом элементов, что во многом определяет особенности их структуры и характер функционирования.
Вещества, входящие в состав протоплазмы. Знание элементарного состава протоплазмы не объясняет нам тайн живого. Почему химические элементы, войдя в состав живого вещества, приобретают способность участвоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
вать в сложнейших биологических процессах? Дело в том, что в протоплазме
химические элементы образуют сложные высокомолекулярные вещества, которые строго упорядоченно взаимодействуют между собой. Изучая свойства
и характер взаимодействия этих веществ, то есть познавая структурную организацию протоплазмы, мы подходим к раскрытию тайн живого, тайн жизни.
В клетках химические элементы находятся в виде органических и неорганических веществ. Многие органические вещества протоплазмы — полимеры — это гигантские молекулы, состоящие из мономеров. Полимеры совмещают в себе свойства устойчивости и изменчивости, благодаря чему на
их основе возможна структурная организация клетки и пространственная организация химических реакций, протекающих в клетке. Приблизительный
состав протоплазмы известен. Ее вещества имеют следующие средние молекулярные веса: белки — 35000, липиды— 1000, углеводы — 200, вода— 18.
70—80% сырой массы протоплазмы составляет вода, 10—20% белки, 2—3%
липиды, 1—1,5% углеводы и другие органические вещества. На одну белковую молекулу приходится в среднем 18 000 молекул воды, 100 молекул других неорганических веществ, 10 молекул липидов, 20 молекул других органических веществ. Важнейшими органическими веществами считаются белки, нуклеиновые кислоты, липиды, углеводы.
Белки по химическому составу являются соединениями С (около 50%),
О (около 25%), N (16%), Н (до 8%), S (0,3—2,5%). В состав белков в небольшом количестве входят и другие макро- и микроэлементы. Белки — это полимеры, состоящие из мономеров — аминокислот. Аминокислоты в белках
объединены между собой пептидными связями (—СО—NH—) — связями
между карбоксильной группой одной и аминной группой другой молекулы.
Пептидные связи образуют первичную структуру белков, в них аминокислотные остатки соединены ковалентными силами. Для каждого белка характерно определенное количество аминокислот, их состав и последовательность расположения в молекуле. Возможные комбинации из 20 известных
аминокислот составляют астрономическое число— 1018. Длинные цепи белковых молекул под действием водородных связей скручиваются в спиральные структуры — это вторичная структура белка. Третичная структура белка
поддерживается гидрофобными, электростатическими или дисульфидными
связями и придает белку специфическую форму. Объединение нескольких
молекул белка в одну макромолекулу фибриллярной (нитчатой) или глобулярной (шаровидной) формы — это четвертичная структура белка.
Все белки амфотерны, так как содержат как кислые (карбоксил—
СООН), так и основные (аминные — NH2) группы. В связи с этим характер
белка и его свойства могут меняться в зависимости от рН среды. Если белок
состоит только из аминокислот, его называют простым или протеином (молочный, яичный, сывороточный, альбумины, глобулины, фибриноген, миозин и др.), а если белок кроме аминокислотных остатков включает z себя другие небелковые вещества (так называемую простетическую группу) — сложным белком или протеидом. В зависимости от природы небелковой части
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
различают: 1) нуклеопротеиды комплексы белков с нуклеиновыми кислотами, особо важная для клетки группа; 2) гликопротеиды — комплексы белков
с углеводами (муцин, различные мукоиды, цикозамины, гликозаминогликаны); 3) фосфопротеиды—соединения белка с фосфорной кислотой (казеиноген молока, вителлин яйца и др.); 4) липопротеиды — комплексы белков с
липидами (все мембранные структуры клетки); 5) хромопротеиды — соединения простого белка с тем или иным окрашенным соединением небелкового
характера, иногда содержащие металл — Fe или Сu (гемоглобин, миогло-бин,
некоторые ферменты — каталаза, пероксидаза и др.).
Белки выполняют многочисленные функции: они входят в состав всех
мембранных структур клетки (пластическая функция); обладают каталитическими способностями (все ферменты являются белками); в экстренных случаях используются как источник энергии (глюконеогенез); им присущи защитные свойства (иммунные белки); являются акцепторами и переносчиками
кислорода в процессе дыхания (гемоглобин, миоглобин); образуют структуры, осуществляющие движение клетки и ее частей, органа, организма (актин,
миозин, тубулин).
Нуклеиновые кислоты — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК) — полимеры с молекулярным весом 104—107. Это чрезвычайно
важные соединения. Функциями ДНК являются хранение и передача наследственной информации и регуляция синтеза белка, а РНК—синтез белка. Их
мономеры — нуклеотиды. Каждый нуклеотид состоит из сахара (пентозы), к
которому с одного конца присоединяются азотистое основание (пуриновое
или пиримидиновое), а с другого — фосфат — остаток фосфорной кислоты.
В нуклеотидах, входящих в состав ДНК, сахаром является дезоксирибоза,
пуриновые основания — аденин и гуанин, пиримидиновые — цитозин и тимин.
В нуклеотидах, составляющих РНК, сахар — рибоза, а в азотистых основаниях вместо тимина присутствует урацил. Друг с другом нуклеотиды соединены при помощи фосфата — диэфирными фосфатными связями, в результате образуется длинная цепочка. Так выглядит РНК. ДНК находится в
ядре в виде двух спиралей, закрученных вокруг общей оси и соединенных
между собой водородными комплементарными связями, возникающими между азотистыми основаниями. Причем всегда образуются пары только двух
видов: аденин — тимин (А—Т) и цитозин — гуанин (Ц—Г). В период подготовки клетки к делению происходит удвоение ДНК — редупликация. Процесс
этот идет под действием ферментов, разъединяющих спирали ДНК. При этом
водородные связи азотистых оснований оказываются свободными и к ним по
принципу комплементарности присоединяются нуклеотиды. Из одной молекулы ДНК образуются две, имеющие ту же первичную структуру.
В период активного функционирования клетки, когда в ней происходит
синтез белка, на одноцепочечных участках молекул
ДНК происходит матричный синтез информационной РНК, которая затем, выходя в цитоплазму и участвуя в синтезе белка, определяет его первичную структуру. В этот период ДНК имеет вид длинных неравномерно спираwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лизованных нитей и в световой микроскоп видна в ядре в виде хроматина —
глыбок разного размера, окрашенных основными красителями. В период деления ДНК сильно спирализуется и приобретает вид окрашенных телец —
хромосом. РНК также адсорбирует основные красители, но локализуется как
в ядре (в основном в ядрышке), так и в цитоплазме. Известно три вида РНК:
информационная (иРНК), транспортная (тРНК), рибосомальная (рРНК). Все
они синтезируются на молекулах ДНК.
В клетках существуют и свободные нуклеотиды, играющие большую
роль в процессах обмена веществ и энергии. Это аденозинтрифосфорная кислота (АТФ), а также трифосфаты уридина, цитидина и гуанозина (УТФ,
ЦТФ и ГТФ). Их называют макроэргическими соединениями, так как они являются аккумуляторами и переносчиками энергии. Энергия высвобождается
при отщеплении от молекулы нуклеотида фосфорных остатков. При распаде
АТФ образуется 38 кДж/моль энергии. Определенное значение придается
еще одному нуклеотиду — циклическому аденозинмонофосфату (цАМФ),
играющему большую роль в рецеп-торных функциях клетки, в механизме
транспорта вещестЕ в клетку, в структурных перестройках мембран.
Липиды состоят в основном из С, О, Н, широко распространены в протоплазме, очень разнообразны по своему строению и свойствам. Молекулы
многих липидов имеют полярные по растворимости концы — один из них не
вступает в связь с водой и с белками — гидрофобный, другой — взаимодействует с водой и белками — гидрофильный. Липиды входят в состав всех
мембранных структур клетки, а также в состав биологически активных веществ (стероидных гормонов), являются запасным энергетическим материалом, так как при их окислении высвобождается большое количество энергии.
Углеводы, как и липиды, образованы в основном С, О, Н и повсеместно
распространены в живом веществе в виде моносахаридов— простые сахара
(глюкоза, фруктоза и др.), дисахаридов (сахароза, лактоза и др.), полисахаридов — их полимеров (гликоген, крахмал, клетчатка, мукополисахариды и
др.). Моно- и дисахариды водорастворимы, полисахариды в воде нерастворимы.
Углеводы — это источники энергии в клетке, в соединении с белками и
липидами входят в состав мембранных структур клетки, нуклеиновых кислот, являются составной частью межклеточного вещества соединительных
тканей, образуют биологически активные вещества (гепарин).
Неорганические вещества представлены водой и минеральными солями.
Вода — необходимая составная часть протоплазмы, в ней протекают все
жизненные процессы. Она проникает в клетку легче других веществ, вызывая
ее тургор и набухание. Поступает вода в клетки пассивно. Проницаемость
клеток разных тканей для воды различная. Так, проницаемость эритроцитов в
100 раз выше, чем яйцеклетки. Данное свойство сильно меняется в зависимости от физиологического состояния клетки и внешнего воздействия. В
норме количество воды в клетках животных поддерживается на постоянном
уровне благодаря работе специальных систем организма, обеспечивающих
постоянство осмотического давления тканевой жидкости и плазмы крови.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Вода находится в клетках в свободном и в связанном состоянии. Количество
связанной воды (от 5 до 80%) зависит как от самой ткани, так и от физиологического состояния организма. Связанная вода образует сольватные оболочки макромолекул и удерживается водородными связями. Свободная вода
— растворитель. В форме растворов в клетку и из клетки поступают различные вещества. Свободная вода является той средой, в которой протекают реакции в клетке, а ее высокая теплоемкость предохраняет клетку от резких перепадов температуры.
Из минеральных веществ в организме чаще встречаются соли угольной,
соляной, серной и фосфорной кислот. Растворимые соли обусловливают осмотическое давление в клетках, поддерживают кислотно-щелочное равновесие, определяя этим реакцию среды, влияют на коллоидное состояние протоплазмы. Минеральные вещества могут входить в состав сложных органических соединений (фосфолипиды, нуклеопротеиды и др.).
Физико-химические свойства протоплазмы определяются состоянием
веществ, входящих в ее состав. Плотность протоплазмы 1,09—1,06, показатель преломления света 1,4. Она приобретает свойства коллоидных систем
из-за присутствия большого количества макромолекул, способных к полимеризации и агрегации. Агрегация молекул происходит в результате их способности к адсорбции. С явлением адсорбции связаны такие жизненно важные
процессы, как дыхание и питание клетки. Многие ферменты функционируют
только в адсорбированном состоянии. Протоплазма обладает рядом свойств
типичных коллоидных растворов, но в то же время имеет и специфические
свойства, характерные только для живого вещества.
Коллоидные растворы являются двухфазной системой, состоящей из
растворителя — дисперсионной среды и взвешенных в нем частиц — дисперсной фазы. Коллоидные частицы — мицеллы— удерживаются во взвешенном состоянии благодаря одноименному электрическому заряду и сольватной оболочке.
Уменьшение заряда и частичное разрушение сольватной оболочки приводит к агрегации мицелл с образованием своеобразной решетки, в ячеях которой находится дисперсионная среда. Этот процесс называется желатинизацией, а продукт — гелем. Гель может переходить в более жидкое состояние
— золь при обособлении мицелл, а золь — в гель при агрегации мицелл. В
протоплазме сочетаются различные коллоидные фазы, которые находятся в
очень неустойчивом состоянии и могут легко меняться в зависимости от
функционального состояния клетки и внешних воздействий. При этом значительно изменяется вязкость протоплазмы. Например, при формировании веретена деления, образования псевдоподий, воздействии током вязкость повышается, при изменении температуры — понижается.
Потеря заряда и добавление электролитов приводят к коагуляции (coagulatio — свертывание) —слипанию мицелл и выпадению дисперсной фазы в
осадок. При слабом воздействии коагуляция обратима, при сильном — необратима и приводит к гибели клетки. От неживых коллоидных систем протоплазма отличается высокой лабильностью; составляющие ее мицеллы белков
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
и липидов образуют сложную структурную основу клетки; при уплотнении
протоплазма способна образовывать новые белковые мицеллы и восстанавливать нарушение связи между ними, а при нарушении целостности клетки
— уплотняться на наружной поверхности с образованием мембранноповерхностной реакции преципитации. Поэтому клетка быстро ликвидирует
свои повреждения.
В определенных условиях дисперсная фаза коллоида обособляется не в
виде осадка, а в виде раствора. При этом коллоидный раствор расслаивается
на два раствора разной концентрации. Это коацервация (coacervacio — нагромождение, собирание в кучу). Коацерваты имеют вид капель, веретен,
слоев. В неоднородной по химическому составу протоплазме, представляющей собой сочетание различных, часто противоположно заряженных коллоидов, возникают сложные комплексные коацерваты.
МОРФОЛОГИЯ КЛЕТКИ
Клетка состоит из сложноорганизованного живого вещества— протоплазмы, разделенного с помощью мембран на цитоплазму и ядро. Ядро отграничено от цитоплазмы ядерной оболочкой и состоит из кариоплазмы.
Клетка отделена от внешней среды клеточной оболочкой — плазматической
мембраной (плазмо-или цитолеммой), которая в животных клетках очень
тонкая и видна лишь в электронный микроскоп (рис. 1).
Размеры и форма клеток очень разнообразны. Самые маленькие клетки
не превышают нескольких микрометров (малые лимфоциты, клетки-зерна
мозжечка), самые большие достигают нескольких сантиметров (яйцеклетки
птиц). По форме клетки бывают шаровидные, овальные, кубические, призматические, звездчатые, дисковидные, с разнообразными отростками и т. д.
Форма клеток тесно связана с их функцией и с механическим воздействием
окружающей среды. Клетки, обладающие амебовидной подвижностью (лейкоциты), способны менять свою форму. Размеры ядра и цитоплазмы обычно
находятся в определенных соотношениях, характерных для той или иной
ткани и отражающих состояние клетки. У молодых и активно функционирующих клеток ядра бывают обычно крупнее, чем у клеток того же типа, находящихся в покое или стареющих.
Электронная микроскопия показала, что ядро, цитоплазма и клеточная
оболочка имеют сложное строение. Они выполняют разнообразные функции.
Нормальная жизнедеятельность клетки возможна только при их сохранности
и взаимодействии.
Клеточная оболочка — поверхностный аппарат клетки, в состав которого входят: плазматическая мембрана, надмембранный комплекс и субмембранный опорно-сократительный аппарат (рис. 2).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
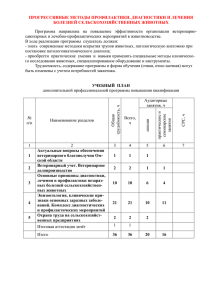
Рис. 1. Схема ультрамикроскопического строения животной клетки:
I — цитолемма: 1 — пиноцитозная вакуоль; 2 — плотный контакт; 3
— десмосома; 4 — контакт по типу замка; 5 —щелевой контакт; 6 — простой контакт; II — цитоплазма; 7 — микроворсинка; 8 —ресничка; 9 — центросома; 10 — микротрубочки; 11 — гранулярная эндоплазматическая
сеть; 12 — агранулярная (гладкая) эндоплазматическая сеть; 13 — аппарат
Гольджи; 14 — митохондрия; 15 — лизосома; 16 — фаголизосома; 17 — полирибосомы; 18 — секреторная вакуоль- 19 — включения гликогена; 20 —
жировые включения; III — ядро; 21 — кариолемма; 22 — пора; 23 — кариоплазма; 24 — хроматин; 25 — ядрышко; а — вндоцитоз; б — экэоцитоз.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 2. Строение поверхностного аппарата клетки:
I — надмембранный комплекс (гликокаликс); II — плазмолемма; III —
субмембранный комплекс; IV — цитоплазма; 1 — гликопротеиды; 2 — гликолипиды; 3 — фосфолипиды; 4 — холестерин; 5 — интегральные. 6 — полуинтегральные и 7 — периферические белки; 8 — микротрубочки; 9 — микрофиламенты; 10 — билипидный слой.
Плазматическая мембрана — плазмолемма или цитолемма имеет толщину около 10 нм и видна лишь в электронный микроскоп. Цитологические,
биологические и физико-химические исследования показали, что она представляет собой белково-липидный комплекс, организованный определенным
образом. Количество белков и липидов в плазмолемме по массе почти одинаковое, но мелких липидных молекул при этом оказывается гораздо больше,
чем крупных белковых. Молекулы липидов полярны. У них незаряженные
гидрофобные хвосты и заряженные гидрофильные головки. В цитолемме они
располагаются в два слоя, гидрофобными хвостами друг к другу, а гидрофильными головками наружу. Такие же структуры образуют молекулы липидов и в неживых объектах, располагаясь на поверхности воды. Молекулы
липидов, входящие в мембраны, разнообразны и очень подвижны. Вдоль
мембраны они могут перемещаться со скоростью более миллиона раз в секунду, образуя как бы вечно подвижное липидное озеро, в котором плавают
молекулы глобулярных белков.
Белки мембран являются ферментными, рецепторными и структурными.
Они также могут перемещаться в мембране в процессе выполнения функции,
в результате чего меняются свойства мембраны и ее функциональная активность. Молекулы белков располагаются в мембране как между молекулами
липидов— интегральные белки, так и под ними — периферические белки. А
так как молекулы белков гораздо крупнее липидных, интегральные белки
выступают наружу, образуя поверхностные слои мембраны. Такая структура
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
плазмолеммы называется мозаичной. В электронном микроскопе она выглядит трехслойной. Два крайних электронно-плотных слоя соответствуют гидрофильным головкам липидных молекул и белковым глобулам, а средний
электронно-прозрачный слой — гидрофобным концам липидных молекул.
Мембраны такого строения принято называть элементарной биологической
мембраной.
Надмембранный комплекс (гликокаликс) — в основном углеводной
природы. Представлен он полисахаридами, образующими с белками мембраны сложные соединения — гликопротеиды, а с ее липидами — гликолипиды.
Длинные ветвящиеся углеводные концы молекул гликопротеидов и гликолипидов соединены в вязкий желеобразный надмембранный чехол толщиной
3—4 нм, который служит своеобразной межклеточной смазкой, склеивающей
клетки между собой. В нем снижается скорость диффузии различных веществ, определенным образом располагаются ферментные комплексы, принимающие участие во внеклеточном расщеплении высокополимерных веществ. Особенно хорошо развит полисахаридный слой в кишечном эпителии,
где очень активны процессы пристеночного (внеклеточного) пищеварения и
всасывания. Считается, что надмембранный комплекс выполняет рецепторную функцию, в том числе и тканевой совместимости, функцию «узнавания»
клетками друг друга, так как благодаря разнообразию химических связей молекул углеводов «рисунок» поверхности клеток тканеспецифичен и даже индивидуален. Он может приводить в действие иммунные механизмы.
Субмембранный комплекс сосредоточен в кортикальном слое цитоплазмы, прилежащем в плазмолемме и тесно с ней связанном. Представляет специализированную периферическую часть опорно-сократительной системы
клетки. Состоит из белков, собранных в нитевидные структуры разной величины: микрофиламенты (диаметром 5—7 нм), микрофибриллы (около 10 нм)
и микротрубочки (диаметром 20—22 нм). Все они вступают друг с другом в
сложные химические и структурные связи, образуя терминальную сеть.
Среди белков терминальной сети основными являются актин, миозин, тубулин, динеин и некоторые другие. Они образуют системы, способные к
скольжению, взаимному перемещению, в результате чего перемещаются
структуры плазмолеммы, связанные с этими системами, то есть осуществляется движение клетки и ее частей. Субмембранный комплекс принимает участие в рецепции, трансмембранном транспорте, стабилизирует белки плазмолеммы и осуществляет их направленное (координированное) перемещение.
Функции плазмолеммы. Плазмолемма вместе с над- и субмембранным
комплексами осуществляет многообразные функции: барьерную, транспортную, рецепторную, двигательную, межклеточных взаимодействий и др.
Барьерная функция, проникновение веществ в клетку. Плазмолемма,
одевая клетку, отграничивает ее от внешней среды, в результате чего вещества внутрь клетки проникают избирательно. С помощью диффузии проникает
в клетку и из клетки лишь вода и некоторые растворенные в ней газы. По
градиенту концентрации (пассивным транспортом) перемещаются некоторые
ионы и мелкие органические молекулы, особенно жирорастворимые. Против
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
градиента концентрации (активным транспортом) поступает большинство
ионов, мелких молекул неорганических и органических веществ (соли, сахара, аминокислоты и др.). Процессы активного транспорта происходят с затратой энергии. В нем принимают участие все элементы поверхностного аппарата клетки. Макромолекулы, их агрегаты и более крупные частицы поступают в клетку благодаря эндоцитозу. При этом частица сначала адсорбируется на мембране клетки, затем окружается ею и отшнуровывается от мембраны с образованием вакуоли. Содержимым вакуоли будет захваченная частица, а ее стенкой — участок плазмолеммы. Если поглощаются более мелкие
частицы и растворы, говорят о пиноцитозе, если более крупные — о фагоцитозе. Процесс выведения из клетки крупных частиц, заключенных в вакуоль, называется экзоцитозом. При этом стенка вакуоли встраивается в плазмолемму и становится ее частью, а содержимое изливается наружу.
Рецепторную функцию клетка выполняет с помощью специальных белков плазмолеммы и элементов гликокаликса (надмембранного комплекса).
Рецепторы клетки разнообразны и многочисленны, что позволяет клеткам
осуществлять взаимные контакты, реагировать на биологически активные
вещества, осуществлять иммунные реакции и т. д.
Движение плазмолеммы осуществляется с помощью субмембранного
комплекса, в котором скапливаются микротрубочки и микрофиламенты. При
передвижении лейкоцитов, в процессе пино-, фагоцитоза и других формах
движения клетки происходит перераспределение опорно-сократимой системы субмембранного комплекса и осуществляется движение клетки и ее частей.
Межклеточные контакты — соединения клеток, особенно распространены в эпителиальных тканях, где клетки плотно прилежат друг к
другу, но имеются и между клетками других тканей. В образовании межклеточных контактов принимают участие все слои клеточной оболочки (поверхностного аппарата клетки). Межклеточные контакты неоднородны по структуре и функции. В зависимости от особенностей строения и функционального назначения различают следующие виды контактов: 1) простой контакт—
клетки прилежат друг к другу, но между ними сохраняется щель 15—20 нм.
Такой контакт широко распространен в различных органах и тканях; 2) зубчатый контакт, или контакт по типу замка, — плазмолеммы клеток образуют
выпячивания, входящие во впячивания соседних клеток. Этот тип соединения способствует креплению клеток в единый пласт и характерен для эпителиев; 3) десмосома, или зона слипания, — сложноорганизованный участок
двух соседних клеток, предназначенный для плотного скрепления клеток.
Десмосомы широко распространены в покровном эпителии, но имеются также в железистом, почечном, кишечном эпителиях, в гладкой и сердечной
мышечных тканях; 4) плотный (замыкающий) контакт — зона плотного прилегания клеток, в которой происходит слияние их мембран. Плотный контакт
обычно окаймляет апикальные полюсы клеток, изолирует внутреннюю среду
организма от внешней и распространен в эпителиях, особенно в кишечном; 5)
щелевой контакт — особый тип соединения клеток, способствующий провеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дению ионов и низкомолекулярных соединений из одной клетки в другую,
минуя межклеточные щели.
Цитоплазма
Цитоплазма — сложная динамическая, многокомпонентная система
клетки, в которой происходят основные метаболические процессы. В цитоплазме различают гиалоплазму, органеллы и включения.
Рис. 3. Участок клетки с фибриллярными структурами цитоплазмы
(схема):
1 — плазмолемма; 2— цистерна эндоплазматической сети; 3 — микротрубочка; 4— микрофиламенты; 5 — полисома; 6 — митохондрия; 7 — ее
наружная и 8 — внутренняя мембраны; 9 — криста; 10 — матрикс; 11 —
мицелла.
Гиалоплазма (hyaline — прозрачный)—жидкая внутренняя среда клетки,
состоящая из воды, низкомолекулярных веществ, растворенных в воде, водорастворимых веществ и высокомолекулярных веществ, рассредоточенных в
виде мицелл и нитей, образующих нежную трехмерную сеть, видимую в
электронный микроскоп лишь при очень большом увеличении (рис. 3). Особенности клетки как коллоидной системы, ее осмотические и буферные свойства в основном определяются составом гиалоплазмы и процессами, происходящими в ней. Гиалоплазма содержит большое количество ферментов (до
25% белков клетки приходится на гиалоплазму), участвующих в процессах
гликолиза, метаболизма сахаров, азотистых оснований, аминокислот, липидов. В гиалоплазме располагаются молекулы АТФ, транспортные РНК и
ферменты активации аминокислот при синтезе белка. Через гиалоплазму
осуществляется большая часть внутриклеточного транспорта ионов, аминокислот, жирных кислот, нуклеотидов, сахаров и других веществ. Здесь откладываются запасные питательные вещества.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Органеллы (маленькие органы) — постоянные составные части цитоплазмы, выполняющие определенные функции. Одни из органелл присутствуют в каждой клетке организма и поэтому называются общими, другие —
лишь в клетках определенного типа и называются специальными. К общим
органеллам относят митохондрии, цитоплазматнческую (эндоплазматическую) сеть, пластинчатый комплекс (аппарат Гольджи), лизосомы, рибосомы,
центросому, микротрубочки, микрофибриллы, а к специальным — миофибриллы, нейрофибриллы, тонофибриллы, реснички, жгутики, микроворсинки.
Большинство общих органелл имеет мембранное строение, то есть представляет собой структуры, стенка которых образована элементарной биологической мембраной. Мембраны органелл устроены так же, как и плазмолемма, и выполняют те же функции: транспортную, разграничительную и
интегрирующую.
Мембранные органеллы — это эндоплазматическая сеть, комплекс
Гольджи, лизосомы и митохондрии.
Эндоплазматическая (цитоплазматическая) сеть — впервые описана в
1945 г. К- Портером с сотрудниками. Эта широко распространенная органелла состоит из вытянутых цистерн, трубочек, мешков, узких каналов, связанных взаимными переходами, в результате чего образуется единая внутриклеточная сеть, которая как бы делит цитоплазму на отсеки, участки. Но при
этом не имеет непосредственных контактов с плазмолеммой. Данная органелла очень подвижная и ранимая, ее составные части легко отшнуровываются, фрагментируются, объединяются, но содержимое при этом не изливается в гиалоплазму, а остается окруженным мембранами цитоплазматической
сети. Различают агранулярную (гладкую) и гранулярную (шероховатую) эндоплазматическую сеть (рис. 4). Цистерны гранулярной эндоплазмати-ческой
сети имеют полости шириной от 20 нм до нескольких микрометров. Со стенками гранулярной эндоплазматической сети связаны рибосомы. Повидимому, первоначально в клетке появляется именно этот вид сети, и лишь
в процессе дифференцировки некоторая часть цистерн теряет рибосомы и
ветвится, превращаясь в гладкую эндоплазматическую сеть. Обратных переходов, образования из агранулярной эндоплазматической сети гранулярной
ее разновидности не наблюдается. Элементы гладкой эндоплазматической
сети уже, их диаметр до 100 нм. Они представляют собой сложную систему
переплетающихся трубочек, канальцев и пузырьков небольшого диаметра.
Цитоплазматическая сеть — своеобразная транспортная система клетки,
по которой перемещаются разнообразные вещества, однако механизм транспорта неясен. Большая поверхность мембран сети говорит о том, что в клетке
может происходить одновременно большое количество разнообразных реакций.
Основная функция гранулярной цитоплазматической сети — синтез
белков. На рибосомах, прикрепленных к ее цистернам, синтезируются белки
секреторных гранул, ферментные белки лизосом и белки, входящие в состав
мембран клетки. Все эти типы белков транспортируются по цистернам цитоплазматической сети, не выходя в гиалоплазму. В гранулярной цитоплазмаwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тической сети синтезируются, кроме того, некоторые углеводные соединения
и липиды, входящие в состав мембран. Синтезированные молекулы подвергаются различным воздействиям (переаминирование, фосфорилирование и
др.), в результате чего меняются их свойства, образуются сложные комплексные соединения. Гранулярная сеть считается местом образования мембран для всей клетки. Войдет ли вновь созданная мембрана в состав стенки
эндоплазматической сети, пластинчатого комплекса, лизосомы или плазмолеммы зависит от того, какие белки и фосфолипиды синтезированы гранулярной цитоплазматической сетью.
Рис. 4. Пластинчатый комплекс Гольджи и эндоплазматическая сеть:
1 — цистерны пластинчатого комплекса; 2— вакуоли; 3 — секреторные гранулы; 4 — цистерны гранулярной эндоплазматической сети; 5 —
гладкая эндоплазматическая сеть; 6 — транспортный пузырек.
Гладкая цитоплазматическая сеть многофункциональна. Она является
местом синтеза ряда углеводов и липидов; поглощает, накапливает и транспортирует ионы (например, ионы кальция в волокнах поперечнополосатой
мышечной ткани); осуществляет детоксикацию вредных продуктов обмена
благодаря определенному набору ферментов, входящих в состав ее мембран
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(в печени). Этот вид цитоплазматической сети очень распространен в клетках
надпочечников, семенников — органах, где образуются стероидные гормоны.
Комплекс Гольджи (аппарат Гольджи), пластинчатый комплекс был
описан К. Гольджи в 1898 г. в виде рыхлой сети, элементы которой имеют
вид палочек, запятых, крючочков. Так, эта органелла выглядит в световой
микроскоп.
Электронная микроскопия показала, что в данный комплекс входят цистерны, вакуоли и пузырьки. Как правило, он представляет собой стопку из
5—10 плоских мешковидных цистерн, узких в центре (7—10 нм) и расширяющихся по краям. По периферии они могут переходить в сеть канальцев и
заканчиваться вакуолями, которые отшнуровываются от пластинчатого комплекса. В клетке насчитывают от одного до нескольких десятков комплексов.
Расположен комплекс обычно вокруг ядра или над ним. Все его структуры
построены из элементарной биологической мембраны, но толщина их и химический состав различны. Наиболее тонкие стенки имеют цистерны, обращенные к ядру и цитоплазматической сети, и по химическому составу подобны последней. Наиболее толстые стенки у цистерн, обращенных к наружным частям клетки. Они по структурно-химической организации подобны
цитолемме.
Пластинчатый комплекс связан со всеми структурами клетки: с поверхностным и ядерным аппаратами, с цитоплазматической сетью, лизосомами.
Выяснено, что в нем накапливаются, конденсируются и созревают продукты,
синтезированные в цитоплазматической сети. При этом происходит их дальнейшая химическая перестройка с образованием секреторных гранул. В цистернах пластинчатого комплекса синтезируются полисахариды и в соединении с белками образуют гликозаминогликаны, мукопротеиды, протеогликаны, иммуноглобулины и другие комплексные соединения. Здесь синтезируются составные части надмембранного комплекса, которые потом без
изменения входят в его состав. С помощью отшнуровывающихся вакуолей
выводятся готовые секреты за пределы клетки. Сами же вакуоли встраиваются в цитолемму и становятся ее частью. Пластинчатый комплекс — место образования лизосом — вакуолей, наполненных гидролитическими ферментами, и пероксисом — пузырьков, содержащих окислительные ферменты.
Лизосомы открыты в 1949 г. де Дювом. Это разнообразные вакуоли
диаметром 0,1—0,4 мкм. Стенка у них образована элементарной биологической мембраной, а содержимое представляет собой скопление гидролитических ферментов. В лизосомах обнаружено около 60 различных гидролаз, действующих в кислой среде. Ферменты лизосом способны разрушать практически1 все природные органические полимеры (белки, углеводы и нуклеиновые
кислоты), но не разрушают клетку и стенки лизосом потому, что находятся в
неактивном состоянии. Лизосомы в клетке очень разнообразны по размерам
и внутреннему содержимому. Связано это с изменением лизосом в процессе
функционирования, а также, возможно, со специализацией лизосом, так как.
они отличаются по набору гидролитических ферментов. Ферменты лизосом в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
течение всей жизни клеток отграничены мембраной. Выполняя гидролитическую функцию, лизосома приближается к пиноцитозной или фагоцитозной
вакуоли и сливается с ней. В результате образуется единая вакуоль, содержащая ферменты и фагоцитированный материал — вторичная лизосома, или
фагосома.
Лизосомы переваривают не только продукты пиноцитозных и фагоцитозных вакуолей, но и разрушенные органеллы клетки (фрагменты митохондрий, цитоплазматической сети и других мембран). Непереваренные остатки
могут накапливаться в лизосомах в виде слоистых телец, пигментных скоплений или удаляются из клетки в процессе экзоцитоза. Лизосомы в клетке
могут передвигаться. В данном процессе какое-то участие принимают микротрубочки, однако механизм перемещения лизосом неясен. Большое количество лизосом бывает в клетках, выполняющих секреторную, экскреторную,
всасывательную и фагоцитарную функции.
Митохондрии (mitos — нить, chondrion — гранула, зерно) впервые описаны в 1850 г. Келликером под названием саркосом, так как он увидел их в
мышечных волокнах. В 1880 г. Альтман вновь привлек к ним внимание, разработав метод специфической окраски, а в 1898 г. Бенда назвал их митохондриями.
Митохондрии видны в световой микроскоп в виде нитчатых, палочковидных, гантелевидных или зернистых образований диаметром 0,2—2 мкм и
длиной 1—10 мкм. В пределах одной клетки они могут иметь разную форму.
Количество их в клетках сильно варьирует и исчисляется от единиц до тысяч.
В соматических клетках млекопитающих обычно насчитывают 500—1000
митохондрий, которые занимают более 20% объема цитоплазмы. В связи с
развитием электронно-микроскопических исследований стали поступать сведения о том, что эти органеллы могут иметь, очень сложную ветвистую
структуру, в результате чего одна митохондрия попадает в срез несколько раз
и создает впечатление большего числа митохондрий, чем на самом деле. Так
или иначе общая поверхность их огромна. В клетках печени, например, она в
4—5 раз превышает площадь поверхности плазматической мембраны. Располагаются митохондрии в клетках в различных местах. Они скапливаются в
тех участках, где расходуется энергия
{под апикальным полюсом всасывающих клеток, у базального полюса
клеток почечных канальцев и т. д.) или где находится энергетический субстрат (около липидных включений).
Независимо от формы и размеров общее строение у митохондрий одинаково в любых клетках. Они ограничены двумя мембранами толщиной по 7
нм. Между мембранами имеется межмембранное пространство шириной
10—20 нм. Полость митохондрии заполнена матриксом тонкозернистого
строения. Мембраны различаются своими структурно-биохимическими и физическими свойствами. Наружная мембрана гладкая, не связана ни с какими
другими мембранами клетки, имеет вид замкнутого мешка. Для нее характерна неспецифическая проницаемость, малое содержание белков и большое
количество липидов. Внутренняя мембрана отличается высокоспецифичной
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
проницаемостью и высоким содержанием белков (до 75% вещества мембраны). У нее многочисленные выросты пластинчатой или трубчатой формы,
направленные внутрь митохондрии — гребни, или кристы. Располагаются
они как вдоль, так и поперек митохондрии, поэтому образуется подобие сети.
Количество и величина крист заметно меняются в зависимости от функционального состояния и возраста митохондрии. У молодых и малоактивных митохондрий крист мало, и они короткие. У зрелых и функционально активных
количество и размеры крист увеличиваются. На поверхности кристы, обращенной к матриксу, обнаружены грибовидные тельца. Матрикс митохондрии
представляет собой коллоидную систему, содержащую большое количество
макромолекул различной природы.
Функция митохондрии — синтез АТФ. Она определяется набором окислительным ферментов, находящихся во внутренней мембране и матриксе. В
последнем сосредоточены ферменты цикла Кребса (трикарбоновых кислот) и
окисления жирных кислот. Во внутренней мембране расположена цепь переноса электронов (дыхательная цепь) и многочисленные транспортные системы, а на грибовидных тельцах локализованы ферменты фосфорилирования
(превращения АДФ в АТФ). Все эти ферментные системы могут синтезировать и накапливать макроэргические (высокоэнергоемкие) соединения (главным образом в виде АТФ), только образуя сложные молекулярные комплексы в составе внутренней мембраны или будучи тесно с ней связанными. В
местах потребления энергии АТФ легко распадается также при участии ферментов митохондрий. При этом образуется свободная энергия для выполнения механической, химической, электрохимической и осмотической работ.
Поэтому митохондрии образно называют «силовыми станциями» клетки.
Помимо этого, они могут участвовать в транспорте воды, накоплении Са, Mg
и Р, в синтезе стероидных веществ.
В матриксе митохондрий выявляются тонкие нити и мелкие гранулы.
Оказалось что нити — это собственная ДНК митохондрий, отличающаяся по
составу и структуре от ДНК ядра, а мелкие гранулы — митохондриальные
рибосомы. Митохондрии, таким образом, имеют свой автономный аппарат
для синтеза собственных митохондриальных белков.
Изучение живых митохондрий с помощью фазово-контрастной микроскопии показало, что они очень динамичны: могут расти в длину, сжиматься,
ветвиться, делиться — и все это за период меньше одной минуты. Жизнь митохондрии продолжается около 20 дней, после чего она утилизируется лизосомами. Гибнущие митохондрии сменяются новыми. Количество митохондрий возобновляется за счет роста и деления предшествующих митохондрий
перетяжкой или почкованием.
Немембранные органеллы — это рибосомы, центросома, микротрубочки, микрофибриллы.
Рибосомы как маленькие плотные тельца были впервые описаны Паладе
в 1955 г., а в 1958 г. Роберте дал им название «рибосома». Их можно увидеть
только в электронный микроскоп.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Это округлые образования диаметром 10—30 нм, состоящие из двух
субъединиц. Обе субъединицы образованы РНК и белками примерно в равных количествах, которые связаны в виде рибонуклеопротеидов (РНП).
Субъединицы неравны по величине. Большая субъединица напоминает полусферу с тремя торчащими выступами, а меньшая — короткую изогнутую палочку с выступом и несколькими полостями.
Рибосома принимает участие в синтезе белка. На ней происходит сборка
полипептидов и белков. При этом к малой субъединице присоединяется молекула иРНК, а к большой — молекулы тРНК Возможно такое присоединение молекул диктуется формой субъединиц (В. Я. Стельмашук). В то время
когда не происходит синтеза белка, субъединицы, как правило, разобщены.
Их объединение происходит под действием ионов магния. Рибосомы синтезируются в ядрышке, но функционируют в основном в цитоплазме. Здесь они
располагаются в гиалоплазме — свободные рибосомы или прикрепляются к
цистернам цитоплазматической сети—связанные рибосомы. Белки, синтезированные свободными и связанными рибосомами, имеют разную судьбу.
Связанные рибосомы синтезируют белки, которые, как правило, попадают в
цистерны цитоплазматической сети, оттуда — в пластинчатый комплекс и в
составе вакуолей покидают клетку (секреторные белки) или становятся белками лизосом. Свободные рибосомы в процессе синтеза белка объединяются
в группы — полисомы (полирибосомы). Они синтезируют белки, входящие в
состав гиалоплазмы, структурные белки немембранных органелл, субмембранного комплекса и другие пластические и ферментативные белки клетки.
Микротрубочки существуют во всех клетках, кроме бактерий и некоторых водорослей, как самостоятельные структуры или в составе немембранных органелл (кроме рибосом). Представляют собой длинные, тонкие, неветвящиеся полые цилиндры с диаметром 22—26 нм и стенкой, построенной из
глобулярного белка тубулина. Микротрубочки очень лабильны. Под влиянием изменения внутриклеточной среды происходит постоянная их самосборка
и саморазборка. В результате одновременного протекания этих процессов в
разных участках цитоплазмы система микротрубочек в целом поддерживается в состоянии динамического равновесия и в интерфазной клетке имеет вид
трехмерной сети. В делящейся клетке сеть микротрубочек цитоплазмы исчезает и вместо нее формируется веретено деления.
В настоящее время микротрубочки относят к скелетным образованиям
клетки, поскольку форма последней в значительной мере зависит от того, как
в ней распределены эти органеллы. -С ними связывают транспортную функцию клеточной оболочки на том основании, что в большом количестве они
обнаруживаются в субмембранном комплексе. Микротрубочки — основные
составные части органелл, осуществляющих движение клетки и ее частей:
центросомы, ресничек, жгутиков. Однако механизм движения их неясен, так
как сами микротрубочки не обладают способностью к сокращению, а движение тем не менее осуществляется. Возможно, своим расположением они создают систему упорядоченного движения внутриклеточных компонентов, являются для них как бы рельсами.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Центросома (клеточный центр) — органелла, присущая животным клеткам. В клетках растений ее нет. Обнаружена и описана практически одновременно Флеммингом в 1875 г. и Бенсденом в 1876 г.
Центросома состоит из центриолей и центросферы (рис. 5). В состав
центросомы обычно входят две центриоли (диплосома), перпендикулярно
расположенные друг к другу. Каждая из них состоит из девяти триплетов
(троек) микротрубочек, уложенных с помощью специальных образований в
виде цилиндра шириной 0,15 мкм и длиной 0,3—0,5 мкм. Микротрубочки
центриолей такого же строения, как и микротрубочки цитоплазмы, и отличаются от них лишь большой стабильностью. Одна из центриолей материнская, другая — дочерняя. На материнской центриоли имеется ряд дополнительных структур: сателлиты, придатки и др., которых нет на дочерней
центриоли. Центросфера — зона цитоплазмы с расходящимися нитчатыми
структурами в виде лучей, окружающая
центриоли. Электронная микроскопия
показала, что лучистое ее сияние — это
многочисленные микротрубочки, отходящие от сателлитов и радиально расходящиеся вокруг центриолей. В реснитчатых клетках центриолей гораздо
больше, так как в основании каждой реснички расположена центриоль в виде базального тельца.
Рис. 5. Центросома:
Центриоли
являются
центрами
1 — материнская центриоль; 2
— дочерняя центриоль; 3 — са- сборки микротрубочек цитоплазмы, микротрубочек веретена деления, выполнятеллит; 4 — микротрубочки,
образующие центросферу; 5 — ют роль базальных телец, от которых оттриплет микротрубочек цен- растают реснички и жгутики. Тем не менее центросома до сих пор — самая затриоли.
гадочная органелла клетки. Высказывается предположение, что центриоли выполняют функцию внутриклеточной
рецепции (Ю. С. Ченцов). Тесная связь ее с ядром с помощью микрофиламентов до сих пор не получила никакого объяснения.
Микрофибриллы и микрофиламенты — нитчатые структуры различной
длины, толщиной от 5 до 10 нм, образованные белками. Располагаются по
всей цитоплазме, но особенно развиты у апикального полюса клетки, где образуют терминальную сеть. Выполняют опорную и двигательную функцию,
в различных тканях образуя специальные органеллы.
Специальные органеллы — постоянные структуры, присущие лишь
клеткам определенных тканей. К ним относятся реснички, жгутики, тонофибриллы, миофибриллы, нейрофибриллы, микроворсинки.
Реснички и жгутики — органеллы движения. Реснички развиты в клетках эпителия дыхательных путей и некоторых отделов половых трактов, а
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
жгутики имеются у спермиев. Структура ресничек и жгутиков сходна, отличаются они лишь размерами. Реснички имеют длину 5—10 мкм, жгутики—
150 мкм. И те и другие представляют собой длинные тонкие выросты цитоплазмы, покрытые цитолеммой, имеющие в основании базальное тельце, а по
всей длине — аксонему. Базальное тельце представляет собой центриоль, у
которой можно наблюдать дополнительные структуры, характерные для материнской центриоли, а иногда можно видеть и дочернюю центриоль, расположенную под прямым углом к материнской. Основная нить ресничек и жгутиков — аксонема организована подобно центриоли, но содержит девять
дублетов (пар) периферических и две центральные микротрубочки. Периферические и центральные микротрубочки связаны между собой в единую подвижную систему. При движении ресничек и жгутиков происходит скольжение микротрубочек друг по другу, в результате чего органеллы изгибаются.
Тонофибриллы, миофибриллы, нейрофибриллы — разновидности микрофибрилл и микрофиламентов, характерные для клеток той или иной ткани.
Тонофибриллы развиты в эпителиальных тканях, где образуют скелет клеток.
Миофибриллы развиты в мышечных тканях, и определяют сократимость мышечных клеток и волокон, а нейрофибриллы — в нервных клетках. Предполагается, что они участвуют в токе аксоплазмы и проведении нервного импульса.
Микроворсинки — выросты цитоплазмы, одетые цитолеммой и содержащие внутри пучок микрофиламентов. Они увеличивают всасывательную
поверхность клетки. Особенно хорошо развиты в кишечном эпителии.
Включения. Это необязательные компоненты клетки, появляющиеся и
исчезающие в зависимости от интенсивности и характера обмена веществ в
клетке и от условий существования организма. Включения имеют вид зерен,
глыбок, капель, вакуолей, гранул различной величины и формы. Их химическая природа очень разнообразна. В зависимости от функционального назначения включения объединяют в группы: трофические включения, секреты
и инкреты, пигменты, экскреты и др. Среди трофических включений (запасных питательных веществ) важную роль играют жиры и углеводы. Белки как
трофические включения используются лишь в редких случаях (в яйцеклетках
в виде желточных зерен). Пигментные включения придают клеткам и тканям
определенную окраску. Секреты и инкреты, накапливаются в железистых
клетках, так как являются специфическими продуктами их функциональной
активности. Экскреты — конечные продукты жизнедеятельности клетки,
подлежащие удалению из нее.
Ядро
Ядро — такая же необходимая составная часть клетки, как и цитоплазма. Цитоплазма без ядра может существовать какое-то время, но вскоре гибнет, а ядро без цитоплазмы погибает еще быстрее. Как оформленная составная часть клетки, ядро возникло не сразу. Существуют клетки, у которых
ядерный материал не отделен от остальной протоплазмы. Это бактерии, неwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
которые водоросли. Клетки без оформленного ядра называют прокариотическими, с оформленным ядром — эукариотическими.
Для большинства эукариотических клеток характерно наличие одного
ядра, но бывают и многоядерные клетки (например, мегакарноциты — специализированные клетки костного мозга, остеокласты в костной ткани и др.).
Многоядерными (обычно двуядерными) могут становиться клетки печени,
надпочечников, обычно одноядерные. При этом резко возрастает функциональная активность клетки, заметно увеличиваются ее размеры.
Форма ядер обычно зависит от формы клетки. У плоских клеток уплощенные, у сильно вытянутых веретеновидные ядра, у равносторонних клеток
они, как правило, округлые, а у цилиндрических— овальные. Встречаются
ядра сегментированные (у зернистых лейкоцитов крови) и даже ветвистые (у
шелкопряда).
В клетках существуют определенные соотношения между размерами
ядра и цитоплазмы, характерные для видовой принадлежности организма,
ткани, органа. Эти соотношения непостоянные, они могут варьировать в широких пределах в зависимости от возраста и функциональной активности организма, органа, клетки. В молодых малодифференцированных клетках ядра
относительно крупнее, чем в зрелых специализированных. У активно функционирующих клеток по сравнению с недеятельными клетками того же типа
ядра более крупные. Изменения размеров ядра и цитоплазмы диктуются особенностями внутриклеточного метаболизма. Высказывается предположение
(Нейфах), что определенное соотношение размеров ядра и цитоплазмы, называемое критическим, является толчком к делению клетки.
Химический состав ядра заметно отличается от состава цитоплазмы. В
нем содержится почти вся ДНК клетки, много основных белков типа гистонов и негистоновых глобулярных белков. РНК и липидов меньше, чем в цитоплазме. Из неорганических веществ большую роль играют вода, ионы Са и
Mg.
Функции ядра определяются наличием в нем ДНК, в которой содержится вся генетическая информация. В ядре происходит редупликация
(удвоение) молекул ДНК, благодаря чему генетическая информация в
полном объеме передается следующим генерациям клеток. В ДНК в виде
последовательности нуклеотидов закодирована информация о первичной
структуре белков, которая реализуется в процессе синтеза. В ядре
синтезируются все три вида РНК: рРНК, иРНК, тРНК и происходит
образование субъединиц рибосом. Следовательно, ядро создает весь аппарат
синтеза белка и регулирует этот синтез как с качественной, так и с
количественной стороны. А это определяет жизнедеятельность клетки, так
как практически все функции в ней обеспечиваются белками. Это хорошо
видно в опытах по пересадке ядер из клетки одного типа в клетку другого
типа. Пересаженное ядро направляет развитие и характер метаболизма в
клетке
Строение
в сторону
ядра.
своего
Ядро
типа.
состоит из ядерной оболочки (кариолеммы), отделяющей его от цитоплазмы, хроматина, ядрышка и кариоплазмы (рис. 6).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Кариолемма (ядерная оболочка, кариотека) состоит из двух элементарных биологических мембран толщиной по 7 нм, разделенных промежутками
10—20 нм. Наружная мембрана по структурно-химической организации подобна мембранам гранулярной цитоплазматической сети. К ней присоединяются рибосомы, она может
образовывать выпячивания, отшнуровываться в цитоплазму. Внутренняя мембрана гладкая, без выпячиваний. К ней прикрепляются нити
хроматина.
Особенность кариолеммы —
наличие пор. Это округлые участки
диаметром 50—100 нм, где сливаются наружная и внутренняя мембраны. Они заполнены, как пробкой,
фибриллярными
и
глобулярными белками, образующими
Рис. 6. Строение ядра:
1 — кариолемма; 2— пора с поровым поровый комплекс. Размеры пор
комплексом; 3— кариоплазма; 4— обычно постоянны, а их количество
конденсированный и 5 — диффузный может уменьшаться в процессе
хроматин; 6 — ядрышко; 7 — рибо- дифференцировки и функциониросомы; 8 — цистерны эндоплазма- вания клетки. В среднем приходится по 10 пар на 1 мкм2 поверхности
тической сети.
ядра. Через поры осуществляется
транспорт веществ между ядром и цитоплазмой, но, несмотря на довольно
большие размеры, далеко не все молекулы могут пройти через них — поровый комплекс обладает избирательной проницаемостью. Он пропускает, например, такие крупные структуры, как белки-гистоны, субъединицы рибосом, но задерживает многие мелкие неорганические молекулы.
Хроматин не виден в живом ядре, но становится заметен в фиксированной интерфазной клетке в виде глыбок, нитей, скоплений, окрашивающихся
основными красителями. Назван так потому, что хорошо воспринимает красители (греч. chroma — цвет).
Химический анализ показал, что хроматин представляет собой комплексное соединение ДНК и белков — дезоксирибонуклеопротеиды (ДНП).
Следовательно, хроматин, это не что иное, как хромосомы в деконденсированном состоянии. Одна молекула ДНК соответствует одной хромосоме.
Длина молекул ДНК очень большая — от 0,5 мкм до 7 см. У человека длина
молекул ДНК всех хромосом одной клетки равна 170 см. Чем более рыхло
уложены хромосомы, тем активнее происходит на них транскрипция РНК и
редупликация ДНК. Деконденсация хромосом неодинаково выражена на
всем протяжении ДНП. Более конденсированные участки окрашены сильнее
и заметны в виде глыбок хроматина (гетерохроматин). Чем заметнее хроматин в ядре, тем он более конденсирован и менее активен. Недеятельный хроwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
матин полностью конденсирован, например, в зрелых эритроцитах птиц, где,
как известно, синтетические процессы отсутствуют. Полностью конденсирован хроматин и в хромосомах во время митоза.
Наиболее активные синтетические процессы происходят в участках
диффузного хроматина (эухроматин)—в полностью деконденсированных
участках хромосом. Отсюда ясно, что по виду и расположению хроматина
можно судить об активности синтетических процессов не только в ядре, но и
в клетке. Чем диффузнее хроматин, тем выше синтез РНК и активнее в клетке идет синтез белка. Такой вид хроматина характерен для молодых, способных к делению клеток. В процессе дифференцировки клетки, по мере ее специализации, увеличивается количество конденсированного хроматина —
происходит выключение участков молекулы ДНК из процесса транскрипции.
Часто такой конденсированный хроматин накапливается в непосредственной
близости к кариолемме. В последние годы появились данные, говорящие в
пользу того, что конденсированный хроматин прочно связан с внутренней
мембраной кариолеммы. В результате этого хромосомы в ядре интерфазной
клетки, по-видимому, не разбросаны как попало, каждая из них занимает определенное пространство.
Ядрышко — наиболее плотный участок ядра округлой формы, диаметром 1—5 мкм. Окрашивается ядрышко основными красителями. Обычно в
ядре 1—2 ядрышка, но может быть и много десятков, как, например, в ядрах
ооцитов рыб. Общий объем ядрышек составляет около 1,5% ядерного объема. Находится ядрышко в непосредственной близости к определенному участку хромосомы, так называемому ядрышковому организатору. У свиньи,
например, он обнаружен на 8-й и 10-й хромосомах, В области ядрышкового
организатора ДНК расположены гены, на которых происходит синтез рибосомной РНК — основного вида РНК ядрышек. В области ядрышка рРНК
синтезируется, накапливается и соединяется с белками — образуются рибонуклеопротеиды (РНП). Здесь же РНП объединяются в субъединицы рибосом. Ядрышко — неоднородная структура. Химически оно состоит на 60—
90% из белков, 2—8% РНК, 5—12% ДНК В нем различают тонкие фибриллы
и гранулы, заключенные в аморфное вещество. Считают, что фибриллы —
это участки молекул ДНК области ядрышкового организатора и РНП, а гранулы — субъединицы рибосом.
Расположение компонентов ядрышка может быть различным, как и его
размеры. Это зависит от активности синтеза, накопления и выведения в цитоплазму рРНК При активном синтезе рРНК ядрышко крупное, его фибриллы лежат рыхло, гранул много. При затухании синтеза рРНК размеры ядрышка уменьшаются, фибриллы лежат плотно, гранул мало или нет совсем.
В период деления клетки ядрышко исчезает, его материал рассеивается в содержимом ядра между хромосомами. Есть данные, что материал ядрышка
входит в состав матрикса хромосом и вместе с ними передается дочерним
клеткам. По окончании митоза, по мере деконденсации хромосом, происходит высвобождение компонентов матрикса и организация новых ядрышек.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Кариоплазма — жидкая фаза ядра, где располагаются все его структуры.
В ней содержится большое количество белков, образующих так называемый
ядерный матрикс. Функции этих белков во многом неясны. Предполагают,
что они играют важную роль в поддержании общей структуры ядра и участвуют в регулировании синтеза нуклеиновых кислот. В состав кариоплазмы
входят и мелкие молекулы органической и неорганической природы.
Вопросы для самоконтроля 1. Дайте характеристику химического состава и физико-химических свойств клетки. 2. Каково строение и функции
клеточной оболочки, органелл, ядра?
Глава 3. ЖИЗНЕДЕЯТЕЛЬНОСТЬ КЛЕТКИ
ПРОЦЕССЫ, ПРОИСХОДЯЩИЕ В КЛЕТКЕ
Все живое неразрывно связано с внешней средой, с которой оно обменивается веществом и энергией, реагирует на внешние воздействия. Не является
исключением и клетка. В основе любых проявлений жизни клеток лежит обмен веществ.
Обмен веществ. Это определенный порядок превращения веществ, направленный на сохранение, самообновление клетки и выполнение ею своих
функций. Интенсивность и направленность обмена веществ клетки в многоклеточном организме в основном определяются надклеточными регуляторными системами: нервной и эндокринной. Но существуют и сложные внутриклеточные системы регуляции процессов жизнедеятельности, что хорошо
видно, например, в таких процессах, как редупликация ДНК, синтез белка. В
процессе обмена веществ поступающие в клетку химические соединения
претерпевают различные превращения (гидролиз, синтез, окисление, восстановление, переаминирование и др.), в результате чего они либо используются
как промежуточные метаболиты, либо входят в состав структур клетки, либо
в состав продуцируемых ею продуктов. Конечные продукты обмена удаляются из клетки.
Внутриклеточная регуляция обмена веществ производится особыми
белками—ферментами, обладающими высокой специфичностью. Они катализируют (ускоряют), как правило, только одну какую-то реакцию. Локализация ферментов в определенном порядке на многочисленных мембранах
клеточных органелл приводит к образованию целых ферментных систем, которые и направляют обмен веществ, ускоряя течение реакций лишь в определенном направлении. Регуляция деятельности ферментных систем осуществляется как извне (гормонами и другими биологически активными веществами), так и изнутри (продуктами, синтезированными в процессе реакции). В
каждой ферментативной системе, как правило, существуют ключевые ферменты, связанные с продуктами их катализа отрицательной обратной связью.
Отрицательная обратная связь — это такая связь, когда фермент своей деятельностью стимулирует образование какого-либо продукта реакции, а продукт реакции, накапливаясь в клетке, тормозит активность фермента. Обмен
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
веществ между клеткой и средой складывается из трех этапов: 1) поступление веществ в клетку; 2) преобразование веществ в процессе внутриклеточного обмена; 3) выведение из клетки продуктов метаболизма.
Поступление веществ в клетку и выведение из клетки метаболитов происходит через цитолемму и при непосредственном ее участии. Наличие внутриклеточных каналов, соединяющих внешнюю среду с цитоплазмой и с
ядром, минуя цитолемму, в настоящее время отрицается. В зависимости от
механизма проникновения вещества различают: пассивный перенос через
мембрану клетки — по градиенту концентрации без затраты энергии, активный перенос — против градиента концентрации с затратой энергии. Например, нормально функционирующая клетка стремится к накоплению ионов
калия и выведению ионов натрия. В результате в клетке калия оказывается
больше, а натрия меньше, чем в межклеточной среде, но калий продолжает
поступать в клетку, а натрий — выводиться из нее благодаря существованию
так называемого натриево-калиевого насоса— определенного механизма переноса ионов с использованием АТФ и цАМФ.
Макромолекулы и более крупные частицы вещества поступают в клетку
с помощью эндоцитоза — процесса
проникновения в клетку веществ в
виде вакуоли, стенки которой образованы отшнуровавшимся участком
плазмолеммы (рис. 7). Разновидностями эндоцитоза являются пиноцитоз (pynein — нить) и фагоцитоз
(phagein — пожирать). Пиноцитоз
— проникновение в клетку более
мелких и жидких частиц, фагоцитоз
— более крупных и плотных частиц. Фагоцитоз у лейкоцитов был
подробно изучен И. И. Мечниковым. Обычно при пиноцитозе возникает углубление цитолеммы, в
которой оказывается захватываемый клеткой материал. Углубление
затем принимает вид мешочка, поРис. 7. Схема эндо- и экзоцитоза:
1 — ядро; 2 — эндоплазматическая том пузырька, наконец, отшнуросеть; 3 — комплекс Гольджи; 4 — вывается от цитолеммы и погрутранспортный пузырек; 5 — секре- жается в цитоплазму клетки. Обраторная гранула; 6 — лизосома; 7 — зуется пиноцитозная вакуоль. При
экзоцитоз; 8 — пиноцитоз; 9 — фа- фагоцитозе клетка для поглощения
гоцитоз; 10 — фаголизосома; 11 — материала может образовывать как
пищеварительная вакуоль (мульти- впячивания, так и выпячивания в
везнкулярное тельце); 12 — остаточ- виде ложноножек или парусов, которые, обволакивая, накрывают соное тельце.
бой частоту вещества, погибшую
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
клетку, бактерию и, таким образом, погружают ее в цитоплазму. Образуется
фагоцитарная вакуоль. Судьба пиноцитозной и фагоцитарной вакуолей
сходна. И та и другая сливаются с лизосомами. Образуется фаголизосома,
или вторичная лизосома, в которой происходит гидролиз (переваривание)
захваченной частицы. Продукты гидролиза — мелкие молекулы простых веществ (сахара, аминокислоты, ионы, нуклеотиды и др.) — выходят через
стенку этой пищеварительной вакуоли в гиалоплазму, а лизосома с оставшимися непереваренными продуктами образуют остаточное тельце, подлежащее удалению из клетки. Процесс выведения веществ из клетки называется экзоцитозом. Он подобен эндоцитозу, но идет в обратном направлении. Иногда при нарушении обмена веществ в стареющем организме можно видеть накопление в клетке остаточных телец в виде миелиновых телец,
липофусциновых гранул, молочных камней и других образований, нарушающих жизнедеятельность клетки. Энергия, необходимая для осуществления процессов поглощения и переваривания веществ, поступает из митохондрий в виде АТФ.
Использование веществ, поступающих в процессе внутриклеточного
обмена можно проследить на примере синтеза белков и образования секретов
(см. «Железистые эпителии»).
Синтез белков (рис. 8) складывается из нескольких этапов: активации,
элонгации и терминации. Первичная структура белка— последовательность
аминокислот в белковой цепи детерминируется (определяется) структурой
ДНК. Участок молекулы ДНК, ответственный за синтез одного вида белка,
называется геном. В период активного белкового синтеза отдельные участки
молекул ДНК деконденсируются, спирали ее расходятся и на них по принципу комплементарности под действием ферментов РНК — полимераз синтезируются молекулы иРНК из запаса нуклеотидов, сахаров, фосфатов, азотистых оснований, имеющихся в ядре. Этот процесс называется матричным
синтезом или транскрипцией (переписывание). Ясно, что нуклеотидная последовательность иРНК точно такая же, как и участка ДНК, на котором она
синтезирована. С одного участка ДНК может сниматься много одинаковых
иРНК. Все они поступают в цитоплазму и вступают в тесную связь с меньшей субъединицей рибосомы. Это стимулирует сборку рибосом и полирибосом. Чем больше рибосом вошло в состав полирибосомы, тем больше синтезируется белковых молекул.
Наряду с этим в цитоплазме происходит активация аминокислот— соединение аминокислот с АТФ и присоединение активированных аминокислот к транспортным РНК. Каждой аминокислоте соответствует своя тРНК и
свой фермент, активирующий эту реакцию. тРНК — небольшая молекула, на
одном конце которой имеется антикодон (триплет)—участок из трех азотистых оснований, комплементарный определенному кодону иРНК. тРНК переносит соединенную с ней аминокислоту на рибосому и прикрепляется к
большой субъединице. При этом антикодон тРНК на короткое время присоединяется к триплету иРНК по принципу комплементарности. иРНК делает
один шаг на рибосоме, аминокислота оказывается присоединенной к строяwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
щейся полипептидной цепочке, а тРНК, освобождаясь от связи с рибосомой,
аминокислотой и иРНК, вновь попадает в гиалоплазму и может присоединять
следующую молекулу аминокислоты. иРНК протягивается через рибосому,
триплет за триплетом, в результате чего идет наращивание белковой молекулы, пока вся иРНК не будет прочитана.
Процесс прочтения информации, закодированной в иРНК, на рибосоме с
помощью тРНК называется трансляцией. Синтезированная белковая молекула попадает либо в гиалоплазму, либо в цистерны цитоплазматической сети и
приобретает специфическую форму.
Считается, что белки, попавшие в гиалоплазму, используются внутри
клетки. Это могут быть структурные белки немембранных органелл, субмембранного комплекса, а также пластические и ферментные. Белки, поступающие в цистерны цитоплазматической сети, являются
белками мембран клетки,
лизосом и секреторных
гранул, которые удаляются
из клетки.
Кругооборот мембран
цитоплазмы. Многие процессы клетки, такие, как
пиноцитоз, фагоцитоз, экзоцитоз, секреция, экскреция связаны с использованием мембранных структур в качестве упаковки
того или иного содержимого. Можно было бы
предположить
большой
расход плазмолеммы у
всасывающих клеток или,
наоборот, приращение ее у
секретирующих. Однако
большинство клеток в
процессе жизнедеятельности мало меняет свои разРис. 8. Схема биосинтеза белка.
меры. Объясняется это
тем, что процессы катаболизма и анаболизма в клетке тесно связаны между
собой и с кругооборотом мембранных структур. Источником всех мембран
клетки считается гранулярная цитоплазматическая сеть. В процессе транспорта веществ ее элементы отрываются и переходят в состав пластинчатого
комплекса. Там они видоизменяются, приобретают сходство с клеточной
мембраной, а их содержимое созревает, превращаясь в секреторную или экскреторную гранулу. Зрелая гранула в составе вакуоли отрывается от пластинчатого комплекса и, подходя к клеточной мембране, изливается за преwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
делы клетки. Оболочка вакуоли встраивается в цитолемму. Параллельно с
этим участки цитолеммы обособляются в качестве оболочки пиноцитозной
или фагоцитарной вакуоли, попадают в цитоплазму и подвергаются гидролизу с помощью лизосом. При этом разбирается и оболочка вакуоли (бывший
участок цитолеммы). Из ее белков синтезируются мембраны цитоплазматической сети, которые затем в районе пластинчатого комплекса вновь видоизменяются. Такой кругооборот мембран существует в течение всего периода
активного функционирования клетки.
Реакции клетки на изменение внешней среды. Клетки обладают раздражимостью — специфической реакцией на воздействия внешней среды.
Характер реакции тканеспецифичен. В ответ на одно и то же, например, механическое воздействие мышечная клетка отреагирует сокращением, нервная
— генерацией нервного импульса, мерцательный эпителий —движением
ресничек. В организме есть клетки, реагирующие на определенные химические вещества активным движением, — лейкоциты.
В то же время на разнообразные воздействия физических и химических
факторов (изменения химического состава поступающих в клетку веществ,
изменение рН среды, колебания температуры, воздействие лучистой энергии,
механические и др.) клетки разных тканей и органов реагируют однотипно.
Реакция клеток зависит не от их тканевой принадлежности, а от силы и длительности действия фактора. Различают нормальную реакцию клетки, паранекроз (или некробиоз) и некроз.
Нормальная реакция клетки может не проявиться в ее морфологических
изменениях, но может выразиться в некоторых изменениях размеров, структуры ядра и органелл, степени желатинизации цитоплазмы. При этом клетка
переходит в более или менее активное состояние. При активации клетки (под
действием гормонов, после кормления животного, при кратковременном
стрессе, действии малых доз УФ-лучей и др.) в ней диспергируется хроматин, увеличивается ядрышко, растет число митохондрий и крист в них, лизосом, цистерн пластинчатого комплекса и цитоплазматической сети, усложняется структура митохондрий, уплотняется их матрикс. Снижение функциональной активности сопровождается конденсацией хроматина, уменьшением
ядрышка и числа органелл. При этом митохондрии набухают, кристы исчезают, матрикс их становится светлее, в мембранах цистерн эндоплазматической сети появляются везикулы.
При более сильном и продолжительном воздействии раздражителя в
клетке развивается состояние паранекроза, при котором значительно нарушаются функции, структура и химизм клетки. При паранекротическом состоянии уменьшается дисперсность коллоидов и возрастает вязкость ядра и
цитоплазмы. Цитоплазма при этом мутнеет, а в ядре появляются нитчатые и
глыбчатые структуры. Реакция цитоплазмы сдвигается в кислую сторону.
Клетка теряет способность связывать прижизненные красители, и они диффузно окрашивают цитоплазму. Такие изменения ставят клетку на грань между жизнью и смертью. Но с прекращением действия раздражителя, если не
повреждены структуры ядра, клетка возращается к нормальному морфофункwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
циональному состоянию. Если же раздражитель действует слишком долго и
сильно, в клетке развиваются необратимые изменения — некроз, ведущие к
смерти.
ВОСПРОИЗВЕДЕНИЕ КЛЕТОК
Способность клеток к воспроизведению — важнейшее свойство живой
материи. Благодаря этой способности обеспечивается непрерывная преемственность клеточных поколений, сохранение клеточной организации в эволюции живого, совершаются рост и регенерация. Способом воспроизведения
клеток считается деление. Известно три типа деления: митоз, мейоз и амитоз.
В результате митоза каждая дочерняя клетка получает то же количество
материала наследственности (ДНК), которое имела материнская клетка. Следовательно, митоз невозможен без предварительного увеличения вдвое (редупликации) количества ДНК в клетке, готовящейся к делению. Удвоение
ДНК и митоз разъединены во времени. Редупликация ДНК происходит в период подготовки к делению — в интерфазе.
Интерфаза и митоз составляют клеточный, или митотический цикл, который для клетки, способной к делению, равен ее жизни, так как в процессе
деления материнская клетка прекращает свое существование, дав начало
двум дочерним клеткам. Их жизнь также заканчивается следующим делением, если они не вступят на путь дифференцировки и выполнения специфических функций. Продолжительность клеточного цикла у разных типов клеток зависит от особенностей ткани, которой принадлежит клетка. Например,
клеточный цикл эпителия кишечника мыши около 19 ч, а эпителия кожи того
же животного почти 586 ч.
Молодые клетки, только что образовавшиеся в результате деления,
имеют в 2 раза меньше ДНК, РНК, белков и других пластических и энергетических материалов по сравнению с материнской клеткой и им необходим определенный период времени для образования и накопления всех этих веществ. Этот период принято называть интермитотической фазой (промежутком между делениями) или интерфазой, составляющей большую часть клеточного цикла. В эпителии кишечника мыши на ее долю приходится 18 ч —
95% времени, в эпителиях кожи — 582 ч — 99%. Интерфазу разделяют на
три периода: пресинтетнческий, синтетический, постсинтетический.
Пресинтетический (поетмитотический) период, или G1-период, наступает сразу после завершения деления клетки и продолжается до начала удвоения ДНК. Это самый длительный период интерфазы. Он составляет от 50 до
90% ее времени. В этот период клетка интенсивно растет, особенно ее цитоплазма, в которой происходит дифференциация органелл, увеличивается количество цистерн цитоплазматической сети, пластинчатого комплекса, митохондрий, удваиваются центриоли центросомы. В клетке совершаются активные процессы синтеза РНК, ферментных и пластических белков, макроэргических соединений и других веществ.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Синтетический период, или S-период, считается ключевым, так как в
это время происходит удвоение ДНК. В природе неизвестно ни одного случая митоза без предварительного прохождения клеткой S-периода. Особым
случаем является мейоз, когда между первым и вторым делениями созревания половых клеток этого периода нет.
Длительность S-периода зависит от скорости редупликации ДНК, а это,
в свою очередь, зависит от размеров молекул ДНК, степени их конденсации и
общего количества ДНК в клетке. Авторадиографическим методом установлено, что у млекопитающих молекулы ДНК имеют линейную форму и редуплицируются одновременно во многих участках, называемых репликонами.
Все хромосомы одной клетки (геном) содержат десятки тысяч репликонов,
что во много раз ускоряет синтез ДНК. Так. редупликация первой хромосомы
человека, длина ДНК которой около 7 см, продолжается 7—12 ч, а если бы
этот процесс шел с одного конца молекулы до другого последовательно, то
занял бы около трех месяцев. Синтез ДНК происходит асинхронно. Раньше
редуплицируются наиболее деконденсированные (деспирализованные) хромосомы или их участки, позже — наиболее конденсированные хромосомы,
например одна из Х-хромосом самок.
В результате редупликации молекул ДНК ее количество в клетке увеличивается вдвое и клетка в конце S-периода содержит тетраплоидное (4n) количество ДНК при диплоидном количестве хромосом (2n), так как редуплицированные молекулы ДНК сохраняют связь друг с другом и теперь каждая
хромосома содержит две молекулы ДНК, называемые хроматидами.
Соответственно увеличению количества ДНК возрастает синтез РНК,
белков-гистонов и других макромолекул. Гистоны переходят в ядро и там
связываются с ДНК, свертывая определенным образом эту гигантскую молекулу. Благодаря описанным выше процессам в S-периоде заметно увеличиваются размеры ядра.
Постсинтетический (премитотический) период, или G2-период, непродолжительный (1—10% времени клеточного цикла) и характеризуется равномерным ростом ядра и цитоплазмы. В этот период происходит накопление
энергии, синтез специальных белков— тубулинов, необходимых для организации митотического аппарата и других макромолекул, используемых при
митозе. По завершении этого периода наступает митотическое деление клетки.
Непрямое деление (митоз или кариокинез).
Митоз — самый распространенный вид деления. Непрямым его называют потому, что окончательному разделению клетки на две дочерние — цитотомии предшествуют сложные изменения и перемещения материала клетки,
особенно ядра. Отсюда второе название деления — кариокинез (по В.
Шлейхеру), что означает движение ядра. Подвижными оказываются структуры, имеющие форму нитей. Отсюда третье название деления — митоз (mytos
— нить), данное В. Флеммингом в 1882 г. Смысл митоза состоит в равном
разделении между дочерними клетками материала наследственности— ДНК,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
заключенного в хромосомах. Термин «хромосома» был предложен В. Вальдейером в 1888 г.
Рис. 9. Различные уровни конденсации хромосомы:
1 — нуклеосомная; 2— нуклеомерная; 3 — хромомерная; 4 — хромонемная.
Строение хромосомы. Хромосома — молекула ДНК, соединенная с белками. Хромосомы существуют в ядре в течение всей жизни клетки, но имеют
разную форму и величину. В период интерфазы это деконденсированные
фибриллярные дезоксирибонуклеопротеиды (ДНП) длиной до нескольких
сантиметров и толщиной 20—30 нм. Двуспиральная молекула ДНК в интерфазной хромосоме уложена с помощью белков — гистонов в виде бусинок на
нитке (рис. 9). Такая укладка ДНК называется нуклеосомной. Это первый
уровень компактизации ДНК, который удается видеть в электронный микроскоп. Обычно интерфазная хромосома уложена более компактно: 8—10 бусинок — нуклеосом компонуются гистонами в «сверхбусину» — нуклеомер.
В таком виде ДНК успешно выполняет свои функции в интерфазной клетке.
В период митоза хромосомы — это компактные тельца характерной
формы, длиной до нескольких микрометров и толщиной 0,5—1 мкм. Для того чтобы хромосома могла принять такие размеры, она должна укоротиться в
10 ООО раз, то есть конденсироваться. В это время гигантская молекула ДНК
изгибается, образуя мелкие, плотно уложенные петли, скрепленные негистоновыми белками. В начальных стадиях конденсации ДНП иногда удается видеть (при мейозе чаще, чем при митозе) утолщения— хромомеры, расположенные на хромосоме через определенные промежутки. В состав одного хромомера входит 50—70 тыс. пар нуклеотидов, что соответствует размеру одного гена. Дальнейшая конденсация приводит к образованию хромонемы —
нити, образующейся за счет сближения хромомеров, видимой уже в световой
микроскоп. Хромонема, в свою, очередь, конденсируется. Возможно, это
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
спирализация или образование петлистых структур. Полностью конденсированную хромонему называют хроматидой. Хромосома содержит одну или
две хроматиды в зависимости от фазы деления (рис. 10).
Полностью конденсированная митоти-ческая
хромосома имеет два плеча, между которыми имеется
первичная перетяжка. В области первичной перетяжки содержится центромера, или кинетохор,—
участок, который присоединяется к микротрубочкам
веретена деления и обеспечивает движение хромосомы. Некоторые хромосомы имеют вторичную перетяжку. Она обычно расположена на одном из концов
и отделяет маленький участок хромосомы — спутник. В области вторичной перетяжки расположена
ДНК, ответственная за синтез рибосомной РНК, поэтому ее также называют ядрышковым организатором.
Митотическая хромосома окружена рибонуклеопротеидами, белками и другими веществами. Это
матрикс хромосомы. С мат-риксом хромосомы переРис. 10. Двухро- дают дочерним клеткам материал, необходимый для
матидная
мета- формирования ядрышка и начального этапа белкового
фазная хромосо- синтеза в молодых клетках.
Совокупность числа, величины и морфологии
ма:
хромосом называется кариотипом. Он видоспецифи1 — плечо хромочен, даже у близких видов отличается числом или
сомы (теломер);
формой хромосом. В кариотип входят соматические и
2 — первичная пеполовые хромосомы. Все соматические хромосомы
ретяжка;
(аутосомы) парные. Пару одинаковых хромосом на3 — центромера
зывают гомологичными. Половые хромосомы непар(кинетохор);
ные. У млекопитающих мужских особей это Х- и Y4 — матрикс хрохромосомы. У женских особей две Х-хромосомы, одмосомы:
на из них находится в неактивном состоянии.
5 — хромонема;
У птиц наоборот: самцы имеют две Z6 — хроматида.
хромосомы, самки W- и Z-хромосомы.
Собственно митоз. В митозе различают четыре фазы (стадии): профазу,
метафазу, анафазу, телофазу. Митоз обычно занимает меньше 10% времени
клеточного цикла, продолжаясь 1—3 ч (рис. 11).
Профаза — первая фаза митоза, во время которой происходит конденсация хромосом — постепенный и довольно продолжительный процесс. Поэтому в профазе различают две стадии: плотного клубка (тонкие нити ДНП
сильно перепутаны в ядре) и рыхлого клубка (более спирализованные хромосомы пространственно отделены друг от друга, но не имеют еще окончательной формы). По мере прохождения профазы удвоенные в интерфазный период молекулы ДНК конденсируются, укорачиваются. При этом в каждой хромосоме и прежняя, и вновь синтезированная молекулы ДНК скручиваются и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
спирализуются одна около другой, образуя единую структуру. Следовательно, в профазе митоза каждая хромосома состоит из двух хроматид, и если количество хромосом в это время равно 2n, то количество ДНК равно 4 n.
Рис. 11. Схема митоза:
А — профаза ранняя (плотного клубка); Б — профаза поздняя (рыхлого клубка); В — метафаза; Г—анафаза ранняя; Д — анафаза поздняя; Е — телофаза; 1 — ядерная оболочка; 2— ядрышко; 3 — центросома; 4 — центросфера:
5 —хромосомы; 6 — веретено деления; 7— экваториальная пластинка (материнская звезда); 8 — дочерняя звезда; 9 —дочерняя клетка.
Одновременно с конденсацией хромосом идет дезинтеграция ядрышка и
включение ядрышкового материала в состав матрикса хромосом. Значительные преобразования в период профазы отмечают и в других структурах клетки. Гранулярная цитоплазматическая сеть и пластинчатый комплекс распадаются на мелкие цистерны и вакуоли, разрушаются полисомы. Все это ведет
к резкому падению синтеза белков. К концу профазы ядерная оболочка
фрагментируется на мелкие мембранные пузырьки. Кариоплазма сливается с
цитоплазмой — образуется миксоплазма. Дезинтегрированные органеллы и
материал, из которого они состояли, равномерно распределен по клетке.
В профазе происходит образование митотического аппарата, без которого было бы невозможно расхождение хромосом. Удвоенные в интерфазе центриоли центросомы начинают расходиться к полюсам клетки и достигают
полюсов к моменту исчезновения ядерной оболочки. К каждому полюсу отходит по две центриоли. От них отрастают микротрубочки, выстраиваясь в
виде веретенообразной структуры. По мере развития веретена все органеллы
клетки и их фрагменты оттесняются на периферию клетки, а ее центр оказыwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
вается занят огромным количеством довольно тесно расположенных микротрубочек, в сумме с центросомой формирующих митотический ахроматиновый аппарат.
Метафаза довольно продолжительная, иногда составляет даже Уз митоза. В этой фазе завершается образование веретена деления. В начале метафазы хромосомы лежат неупорядоченно, но не выходят за пределы ядерной области, где они совершают дрейфующие движения, которые, по-видимому,
являются результатом взаимодействия хромосом с микротрубочками. Постепенно формируется метафазная пластинка — хромосомы выстраиваются по
экватору клетки таким образом, что их центромеры обращены к центру веретена, а концы — к его периферии. Такую фигуру называют материнской
звездой. Каждая хромосома соединена с двумя нитями веретена, так как состоит из двух хроматид, и у каждой хроматиды имеется свой кинетохор. Сестринские хроматиды каждой хромосомы на протяжении метафазы начинают
отходить друг от друга, но в области кинетохора сохраняют связь до конца
метафазы. Поэтому метафазные хромосомы часто имеют Х-образную форму.
Анафаза начинается резко — все хромосомы теряют центромерные
связки, и их хроматиды начинают быстро двигаться к полюсам со скоростью
до 0,5 мкм/мин. Анафаза самая короткая фаза митоза. По-видимому, ахроматиновые нити веретена деления тянут хромосомы к полюсам клетки, отчего
они приобретают V-образную форму. Центромеры хромосом направлены к
полюсам, а противоположные концы плеч — к экватору. Образуются две фигуры, напоминающие материнскую звезду. Их называют дочерними звездами.
Телофаза начинается с остановки хромосом, достигших полюсов клетки.
В ранней телофазе начинается деконденсация хромосом. Они набухают, увеличиваются в объеме, слабее окрашиваются. Их ориентация не меняется в
сравнении с анафазой. Вокруг хромосом обособляется ядерная оболочка,
формируясь из мембранных пузырьков. После образования кариолеммы в
области ядрышкового организатора некоторых хромосом формируются ядрышки.
Одновременно с этим идет разрушение митотического аппарата. Дольше
всех он сохраняется на экваторе клетки, в области бывшей метафазной пластинки. Высказывается предположение, что микротрубочки митотического
аппарата в области экватора вместе с субмембранным комплексом клетки
участвуют в образовании клеточной перетяжки. При разделении клетки —
цитотомии органеллы пассивно распределяются между двумя дочерними
клетками.
По разным причинам (нарушение веретена деления, нерасхождение
хроматид и др.) во многих органах и тканях встречаются клетки с крупными
ядрами или многоядерные клетки. Это — результат соматической полиплоидии. Такое явление называют эндорепродукцией. При этом клетка может содержать во много раз больше ДНК, но оно будет кратно 2n. Чаще полиплоидия встречается у беспозвоночных животных. У некоторых из них распроwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
странено и явление политении — построение хромосомы из многих молекул
ДНК.
У млекопитающих полиплоидные клетки встречаются в печени, эпителии мочевого пузыря, в пигментном эпителии сетчатки глаза, в концевых отделах слюнных и поджелудочной желез, среди мегакариоцитов и других специализированных клеток. Полиплоидии не бывает в период эмбриогенеза
(кроме провизорных органов) и у стволовых клеток. Полиплоидные и политенные клетки, образовавшиеся в результате эндомитоза (нарушения веретена деления), не вступают в митоз и могут делиться только амитозом.
Смысл данного явления в том, что как полиплоидия — увеличение количества хромосом, кратное 2n, так и политения — увеличение количества молекул
ДНК в хромосоме приводят к значительному усилению функциональной активности клетки. А это (эндорепродукция) позволяет без перерыва в функционировании нарастить клеточную массу и увеличить объем работы, выполняемой клеткой, а отсюда и органом.
Амитоз — прямое деление клетки. При данном делении не происходит
конденсации хромосом, не образуется веретено деления. Ядро находится в
интерфазном состоянии. Деление начинается с размножения или разделения
ядрышка перетяжкой. Затем перетяжкой делится ядро. При этом могут возникнуть ядра, неравные по величине, может образоваться сразу несколько
(больше двух) ядер. Дальше следует цитотомия. Однако разделение клеток
при амитозе необязательно. Деление может завершиться образованием многоядерных клеток. Клетки, разделившиеся амитозом, не в состоянии делиться
митотически. Амитоз встречается почти всегда либо в клетках отживающих,
находящихся в конце своего жизненного пути, либо в клетках временно существующих образований (в клетках трофобласта, плодных оболочек, фолликулярных клетках яичника и др.), либо при различных патологических
процессах (воспаление, регенерация, злокачественный рост).
Мейоз — процесс, состоящий из двух делений, быстро следующих друг
за другом, в результате чего образуются половые клетки — гаметы с вдвое
уменьшенным гаплоидным числом хромосом (1n). Встречается он у всех живых организмов (растений и животных), размножающихся половым путем.
Необходимость мейоза в развитии половых клеток диктуется необходимостью сохранения из поколения в поколение кариотипа, характерного для вида. Если бы этого не было, слияние при оплодотворении двух диплоидных
клеток — мужской и женской — привело бы к появлению тетраплоидного
потомства (4n), затем октаплоидного (8n) и т. д., в результате чего каждое последующее поколение представляло бы новые формы, самовоспроизведение
вида было бы невозможно.
Мейоз включает в себя два деления: редукционное — уменьшительное и
эквационное — уравнительное, быстро следующих друг за другом. Деления
разделены короткой интерфазой, в которой отсутствует S-период, то есть перед вторым делением не происходит редупликации ДНК. Каждое деление состоит из четырех фаз: профазы, метафазы, анафазы и телофазы (рис. 12).
Мейоз несравнимо длиннее митоза. Объясняется это сложностью и длительwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ностью преобразований в профазе первого деления мейоза (профазе I), которая может длиться от нескольких дн°й до нескольких лет (в оогенезе).
Профаза I характеризуется такими процессами, как рекомбинация генетического материала, обмен участками между гомологичными хромосомами,
синтез рибосомной и информационной РНК, активация ядрышек. На протяжении профазы I сохраняется кариолемма. Профазу I принято разделять на
пять стадий: лептонема, зигонема, пахинема, диплонема, диакинез.
Рис. 12. Схема мейоза:
1 - профаза I (а – лептонема, б – зигонема, в – пахинема, г – диплонема, д –
диакинез); 2 – метафаза I; 3 – анафаза I;4 – телофаза I; 5 – профаза II; 6 –
метафаза II; 7 – анафаза II; 8 – телофаза II
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Лептонема (leptos — тонкий, пета — нить), или лептотенная (taenia —
лента) стадия, стадия тонких нитей, характеризуется началом спирализации
ДНК. Хромосомы в ней заметны в виде длинных тонких нитей, на которых
видны утолщения,— хромомеры. Каждая хромосома состоит из двух хроматид, но лежат они так плотно друг к другу, что образуют единую нить. Часто
хромосомы связаны с оболочкой ядра и ориентированы таким образом, что
образуют характерную фигуру, называемую «букетом».
Зигонема (zygoo— соединять), или зиготенная стадия, — стадия соединенных нитей, характеризуется продолжающейся конденсацией хромосом,
сближением гомологичных хромосом и их конъюгацией — соединением.
Конъюгация начинается либо с конца хромосом, либо с центромеры. Видимо,
расположение хромосом в виде букета способствует поразительной точности
и специфичности совмещения гомологичных хромосом в пару — бивалент.
Точность эта достигается и благодаря образованию между гомологичными
хромосомами специализированной структуры — синаптонемального комплекса, который и связывает их в биваленты. Поэтому эту стадию еще называют синаптенной.
Пахинема (pachys — толстый), или пахитенная стадия, — стадия толстых нитей, характеризуется завершением конъюгации по всей длине гомологичных хромосом. Они при этом продолжают спирализоваться, принимая
вид коротких, толстых нитей. Становится заметно, что каждая хромосома состоит из двух хроматид, а так как в биваленте объединены две хромосомы, то
фигура эта содержит четыре хроматиды и называется тетрада. Общее количество тетрад в клетке равно In (гаплоидно). Хроматиды одной хромосомы
называются сестринскими. Одновременно с продольным расщеплением хромосом на сестринские хроматиды происходит кроссинговер — обмен участками между несестринскими хроматидами в пределах тетрады. В результате
кроссинговера происходит перекомбинация (рекомбинация) генетического
материала — осуществляется наследственная изменчивость.
Диплонема (diploos — двойной), или диплотенная стадия,— стадия
двойных нитей. Здесь завершается кроссинговер, начавшийся в пахинеме.
Синаптонемальный комплекс разрушается и гомологичные хромосомы начинают отходить друг от друга. Однако в точках перекреста, которые называются хиазмы (мостики), связь сохраняется. Сестринские хроматиды остаются
связанными по всей длине.
В отличие от митоза в профазе мейоза сохраняется оболочка ядра, продолжается синтез всех видов РНК. Наиболее активно синтетические процессы идут на протяжении пахинемы и днплонемы. На этих стадиях ядрышки не
только сохраняются, но и увеличиваются их число и размеры. Увеличиваются при этом и размеры клетки. Особенно ярко эти процессы выражены при
развитии женских половых клеток.
Диакинез («движение вдаль») — последняя стадия профазы I мейоза. В
этой стадии хромосомы резко сокращаются в размеpax, ДНК полностью конденсируется, количество хиазм уменьшается, они сохраняются в основном
только на концах хромосом, исчезают ядрышки, прекращается синтез РНК.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Переход к метафазе I характеризуется разрушением ядерной оболочки и
образованием веретена деления. В метафазе хромосомы выстраиваются в виде экваториальной пластинки. При этом гомологичные хромосомы в биваленте соединены между собой хиазмами. Количество бивалентов в клетке
равно гаплоидному набору (In), а количество хроматид равно 4n. Бивалент
имеет две центромеры. В результате этого при скольжении нитей веретена в
анафазе I к полюсам клетки отходит по целой хромосоме из гомологичной
пары. Число хромосом у полюсов к концу анафазы I оказывается гаплоидным, но каждая хромосома состоит из двух сестринских хроматид, так что
число хроматид диплоидно. Расхождение хромосом из гомологичной пары
случайно, что является еще одним источником рекомбинации отцовского и
материнского геномов.
Телофаза I начинается с того момента, когда анафазные группы хромосом достигнут полюсов клетки. В телофазе происходит формирование ядра,
цитотомия (разделение клетки) и начинается деконденсация хромосом.
Наступает короткая интерфаза — подготовка ко второму делению мейоза. Основное отличие этой интерфазы от интерфазы митотического цикла то,
что в ней не происходит синтез ДНК и белков—гистонов, однако накапливается энергия и синтезируются белки — тубулины, необходимые для формирования веретена деления.
В профазе II мейоза происходят процессы, характерные для профазы
митоза: формируется ахроматиновый аппарат, разрушается ядерная оболочка, конденсируются хромосомы.
В метафазе II нити веретена прикрепляются к хромосомам, число которых гаплоидно (In). Каждая хроматида хромосомы имеет свой кинетохор,
связанный с веретеном.
В анафазе II хроматиды каждой хромосомы расходятся к полюсам, в результате чего у полюсов оказывается гаплоидное число хроматид, называемых теперь хромосомами. В телофазе II образуется оболочка ядра и происходит цитотомия.
В результате из каждой половой клетки, вступающей в мейоз, образуется четыре клетки, содержащие по гаплоидному набору хромосом. Этот набор
содержит по одной из каждой пары гомологичных хромосом одного из родителей, но с измененным благодаря кроссинговеру набором генов, так что каждая из четырех клеток отличается по своей генетической конституции.
ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ
Рост клеток. Независимо от тканевой принадлежности в молодой клетке,
образовавшейся в результате деления, активно происходят процессы синтеза
пластических и энергетических веществ. У такой клетки, как правило, крупное ядро с мелкодисперсным хроматином, показывающим, что ее ДНК деконденсирована и, следовательно, активно синтезирует иРНК и тРНК Ядрышко крупное или их несколько, значит, идет активный синтез рРНК В цитоплазме увеличивается количество мембранных органелл. Активно происwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ходят процессы эндоцитоза (пино- и фагоцитоза). Это приводит к увеличению метаболической активности, к росту цитоплазмы. Рост клетки продолжается до определенных пределов, характерных для ткани, к которой принадлежит клетка. Размеры клеток никак не связаны с размерами организма,
но имеют некоторую связь с классом животного. У земноводных самые
крупные клетки, у птиц и рыб—самые мелкие, у млекопитающих— средние
по размерам клетки. Причины этого неясны. Конечные размеры клеток определяются ядерно-цитоплазматическим соотношением. Рост клетки прекращается, если соотношения этих частей клетки становятся оптимальными для
того, чтобы ядро могло регулировать происходящие в клетке процессы, а цитоплазма могла выполнять свои специфические для данной клетки функции.
Если клетка находится в клеточном цикле, ее рост идет на протяжении
Gi-периода и завершается началом редупликации ДНК — клетка переходит в
S-период и далее проходит все этапы клеточного цикла.
Дифференциация клеток. Если клетка вышла из клеточного цикла
(обычно это отмечают в G1-периоде), то наряду с ростом в ней наблюдается
становление того типа обмена веществ (метаболизма), который характерен
для клеток данного вида, то есть происходит клеточная дифференциация.
Она приводит к появлению специфических структурных, функциональных и
биохимических признаков, характерных именно для данного вида клеток.
Это поразительное единство формы, функции и метаболизма дифференцированных клеток подтверждается в любой ткани, любом органе. Мышечные волокна, например, в соответствии с функцией сокращения имеют вытянутую
форму и содержат специальные органеллы — миофибриллы. Дифференцированные клетки, как правило, имеют структурные признаки своей специализации. Это могут быть реснички, микроворсинки или другие выросты
клетки, определенная форма или сильное развитие какой-либо из органелл
(например, лизосом у зернистых лейкоцитов), характер соединения клеток
(развитие десмосом и плотных контактов в покровном эпителии), накопление
специфических гранул в цитоплазме (зимоген в клетках поджелудочной железы), форма ядер (сегментоядерные лейкоциты), особенности расположений
и структурированности хроматина (хроматин в виде спиц колеса в плазмоцитах) и т. д.
Причины и механизмы дифференциации клеток изучены слабо, тем не
менее ясно, что она определяется взаимодействием генотипа клетки и внешними по отношению к клетке влияниями. По-видимому, в процессе дифференцировки происходит блокировка части генома. Включенными остаются
гены, поддерживающие жизнедеятельность клетки и отправление ею специфических функций. Эта точка зрения подтверждается экспериментально. Если в цитоплазму энуклеированной (с удаленным ядром) яйцеклетки лягушки
пересадить ядро эпителия кишки головастика, из такой клетки вырастает
нормальный лягушонок. Следовательно, геном эпителиальной клетки сохранен полностью, но в дифференцированной клетке работает только его часть.
Если такое ядро попадет в другие условия (в цитоплазму яйцеклетки), геном
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
может быть разблокирован и функционировать в полной мере (дать начало
целому организму).
Дифференциация клеток определяется не только внутриклеточными
факторами, но и межклеточным взаимодействием тканей. В процессе эмбрионального развития отмечают явление индукции, когда эмбриональные
зачатки способствуют развитию друг друга. Так, развивающийся глазной бокал способствует дифференцировке прилежащей эктодермы в хрусталик, а
хрусталик способствует развитию стекловидного тела. Имеются данные о
влиянии на дифференциацию нервной и эндокринной систем, а также питания.
Способность клеток к делению. Жизнь клеток на ранних этапах эмбрионального развития организма, когда бурно увеличивается количество клеток
зародыша, как правило, равна митотическому циклу. По мере роста и развития организма, в период гистогенеза и органогенеза все большее количество
клеток выходит из клеточного цикла, дифференцируется и начинает осуществлять свои специфические функции. Обычно это происходит в Giпериоде. Клетки, вышедшие из митотического цикла, принято называть клетками Go-периода, или покоящимися клетками. Однако в тканях, где физиологичным является короткий срок жизни дифференцированных клеток (например, клеток крови, кишечного эпителия и др.), на протяжении всего онтогенеза сохраняются клетки, функцией которых является пополнение клеточного состава ткани взамен естественно отмирающих клеточных элементов.
Такие клетки называются камбиальными, или стволовыми. Они не выполняют функций, специфических для органа, в котором находятся, остаются малодифференцированными и способны к многократному делению, то есть к
многократному повторению клеточного цикла. Дифференцированные клетки
в таких тканях не способны вступать в митоз.
В других органах (печени, многих эндокринных железах) не обнаружено
стволовых клеток, но их дифференцированные клетки являются долгоживущими и к тому же обладают способностью к делению. При подготовке к делению такие клетки округляются, теряют специфические черты, приобретают
сходство с недифференцированными клетками и вступают в клеточный цикл.
В организме существуют и такие высокоспециализированные тканевые
системы, дифференцированные структурные элементы которых не способны
к делению. Это нервные клетки и мышечные волокна. Неспособность нейронов к делению, видимо, объясняется тем, что деление клетки связано с ее дедифференциров-кой, потерей ею специфичности. Для нервной ткани это означает выпадение каких-либо регуляторных функций, которые могут сказаться на жизнеспособности всего организма. Мышечные волокна— это симпласты, имеющие сложный путь развития, в связи с чем дедифференцировка
до миобластов для них оказывается невозможной. Пути увеличения скелетной мышечной ткани иные (см. в разделе о тканях).
Старение и смерть клетки. Организм и его клетки постоянно подвергаются воздействию самых разнообразных факторов. Эти факторы, несмотря
на саморегуляцию, заложенную в каждой клетке, могут вызвать структурные
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
или метаболические нарушения в клетке. В процессе отправления специфических функций в клетке также могут накапливаться продукты метаболизма,
которые способны изменить ее гомеостаз (постоянство внутренней среды). В
результате этого клетки изнашиваются, стареют и гибнут. Продолжительность жизни клеток в разных тканях может быть очень различной и исчисляться часами и днями (моноциты, эпителий кишки), месяцами (эритроциты,
эпителий кожи), годами и десятилетиями (гепатоциты, остеоциты), а может
быть сравнима и с жизнью особи (невроциты).
В процессе старения клетки изменяются свойства ее протоплазмы—
снижается способность ее коллоидов связывать воду, увеличивается структурированность из-за появления долгоживущих макромолекул взамен метаболически активных короткоживущих макромолекул. В клетках накапливаются
шлаки в виде гранул липофусцина, уменьшается количество белка и гликогена и увеличивается содержание липидов, холестерина и др., конденсируется
хроматин.
К явлениям естественного старения неизбежно примешиваются изменения, вызванные воздействием неблагоприятных факторов. Для поврежденных клеток характерно снижение количества АТФ, активация гликолиза и
протеолиза, сдвиг рН клетки в кислую сторону, нарушение проницаемости
клеточных мембран, вакуолизация и фрагментация органелл, накопление ряда включений (пигментов, слоистых телец и др.)- В результате этого развиваются различные дистрофии (жировая, белковая и др.). При обратимых
изменениях структура и функция клетки возвращаются к норме после снятия
воздействия. При необратимых изменениях клетка гибнет. Явный признак
гибели клетки — активация внутриклеточных гидролитических ферментов,
вышедших из разрушившихся лизосом. Под их влиянием происходит автолиз— самопереваривание клетки.
Вопросы для самоконтроля. 1. Какие вы знаете основные процессы
жизнедеятельности клетки; как участвуют составные части и органеллы клетки в процессах обмена, транспорта (поступления и выведения) веществ? 2.
Каковы реакции клетки на изменения внешней среды? 3. Опишите основные
этапы жизненного цикла клетки: рост, способность к делению, дифференциация, старение и смерть. 4. В чем состоит подготовка клетки к делению; особенности строения хромосомы в разные периоды митотического
(клеточного) цикла? 5. Что такое митоз, мейоз, амитоз; что общего и в чем
различия митоза и мейоза?
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Раздел второй. ОСНОВЫ ЭМБРИОЛОГИИ
Эмбриология (embryon — зародыш, logos — учение) — наука о развитии
организма от оплодотворения до рождения (для живородящих); до вылупления (для яйцекладущих), до окончания метаморфоза (для животных с личиночной стадией). Ее делят на общую и частную. Общая эмбриология изучает
ранние этапы развития зародыша, образование осевых органов и плодных
оболочек (для млекопитающих и птиц это соответствует зародышевому периоду развития). В ее задачу входит также изучение развития половых клеток
и процесса оплодотворения (предзародышевый период или прогенез). Частная эмбриология изучает развитие отдельных органов и их систем, а также
период новорожденности— у амниот. У млекопитающих это последние этапы развития плода, акт рождения и первые недели постнатальной жизни.
Глава 4. РАЗВИТИЕ, СТРОЕНИЕ ПОЛОВЫХ КЛЕТОК И
ОПЛОДОТВОРЕНИЕ (ПРОГЕНЕЗ)
Способность к воспроизводству себе подобных является характерной
чертой, присущей любой живой системе: клетке, одноклеточному и многоклеточному организмам.
Возникновение половых клеток в филогенезе. У организмов известно
два основных типа размножения: бесполое и половое. Бесполое размножение, или апогамия, осуществляется без участия половых клеток путем деления, почкования, фрагментации, спорообразования. Этот способ размножения является наиболее древним и происходит путем прямого обособления от
родительской особи какой-либо ее части (из одной или многих клеток), из
которой развивается целый организм. Половое размножение организмов, или
сингамия, осуществляется путем слияния половых клеток. Данное размножение в мире животных, по-видимому, возникло на ранних этапах эволюции,
так как его подобие встречается у некоторых одноклеточных, например у
инфузорий, когда наблюдается слияние двух особей с последующим их разделением на четыре инфузории.
С появлением многоклеточности и дифференциацией организма на различные ткани возникают специальные клетки для размножения — гаметы,
или половые клетки. Видимо, вначале каждая половая клетка могла развиваться в целостный организм, что мы сейчас отмечаем в случае партеногенеза (partenos — девственница), когда из неоплодотворенной яйцеклетки образуется организм. Партеногенез как естественное явление в той или иной мере
свойствен многим беспозвоночным животным, например пчелам, и даже
встречается у некоторых позвоночных (у отдельных рыб, пресмыкающихся,
птиц). Однако в подавляющем большинстве случаев половое размножение
осуществляется в результате слияния мужской и женской половых клеток.
Если гаметы в мужском и женском организме не отличаются между собой, такие организмы называются изогамными (многие низшие растения).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Организмы, в которых гаметы различаются по своим морфофизиологическим
признакам (большинство растений и животных), называются анизогамными.
Более древней формой полового размножения дифференцированными
гаметами считается гермафродитизм — в одном организме развиваются и
женские, и мужские гаметы, сливаясь, они дают жизнь новой особи. Данное
явление присуще многим растениям, беспозвоночным и некоторым позвоночным (некоторые рыбы) животным.
Наиболее совершенная форма полового размножения — гонохоризм — в
одном организме развиваются либо мужские, либо женские гаметы. Для зарождения нового организма необходимо определенное взаимодействие родительских особей, результатом которого является оплодотворение — слияние
мужской и женской гамет.
РАЗВИТИЕ ПОЛОВЫХ КЛЕТОК В ОНТОГЕНЕЗЕ
МЛЕКОПИТАЮЩИХ
Развитие половых клеток — гамет называется гаметогенезом. Половые
клетки у млекопитающих дифференцируются очень рано — когда организм
состоит примерно из 1000 клеток. В это время среди клеток энтодермы в задней части желточного мешка обнаруживаются первичные половые клетки,
которые вычленяются из нее и мигрируют в дорсальном направлении. От
других клеток первичные половые клетки отличаются округлой формой
(диаметр 20 мкм), очень крупным ядром и высоким содержанием гликогена и
кислой фосфатазы, благодаря чему при специальной гистохимической окраске приобретают черно-коричневый цвет. Ко времени возникновения половых валиков (закладки гонад) первичные половые клетки мигрируют в эндотерму задней части кишки, оттуда по ее брыжейке в дорсальном направлении
— в закладку гонад. Дойдя до половых валиков, первичные половые клетки
вселяются в них, теряют подвижность, округляются, размножаются и дифференцируются в мужские или в женские половые клетки в зависимости от типа половых хромосом, содержавшихся в клетках зародыша. Сначала закладки
гонад индифферентны. После вселения и размножения в них первичных половых клеток, когда в гонадах начинается характерная для данного пола
дифференциация тканей, половые клетки в мужской железе вместе с зачатками семенных канальцев перемещаются во внутреннюю зону, в женской остаются на периферии — в корковом веществе.
Развитие мужских половых клеток — сперматогенез
Мужская половая клетка — спермий, или сперматозоид, была открыта
Л. Левенгуком в 1675 г. Мужские половые клетки развиваются в семенниках.
В эмбриональный период онтогенеза первичные половые клетки вселяются в
закладку семенных извитых канальцев семенника, неоднократно делятся и
превращаются в первичные сперматогонии — стволовые клетки с пониженным обменом веществ, входящие в состав стенки семенных извитых канальцев. Дальнейшее развитие мужских половых клеток приостанавливается
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
до пубертатного периода — до начала полового созревания особи. Начиная с
этого времени и до старости у самцов идет активный сперматогенез.
Рис. 13. Схема сперматогенеза (I) и оогенеза (II).
Отцовские хромосомы темные, материнские — светлые.
А — аутосомы; X, Y — половые хромосомы.
В сперматогенезе принято различать четыре стадии: размножения, роста, созревания и формирования (рис. 13). Полный цикл развития половая
клетка проходит у быка за 63 дня, у барана— за 48, у хряка — за 40 дней.
Процесс сперматогенеза протекает волнообразно. На протяжении одного канальца можно последовательно проследить все его этапы (рис. 14). В пределах же среза в стенке канальца невозможно увидеть одновременно все виды
клеток.
В стадии размножения происходит многократное митотическое деление
сперматогоний. При этом часть половых клеток сохраняет все черты эмбриональности, остается стволовыми клетками и в дальнейшем является источником пополнения сперматогенного эпителия. Другая часть вступает на путь
дифференцировки. И те и другие клетки называются сперматогониями, хотя
и различаются по морфологическим признакам. Все сперматогоний прилежат
к оболочке извитого канальца, располагаясь на базальной мембране. Это уплощенные или овальные клетки со светлой цитоплазмой и округлыми или
овальными ядрами. Хроматин, вначале мелкодисперсный, с каждым делением становится все заметнее. У животных каждого вида наблюдается опредеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ленное количество сперматогониальных делений (от 1 до 14), чаще 4—6 (у
быка — 6, у барана — 5).
Рис. 14. Последовательные этапы развития клеток сперматогенного эпителия (I—
VI) на протяжении семенного извитого канальца:
1—оболочка;
2
—
стенка и 3 — просвет
канальца; 4— сперматогония; 5 — сперматоциты I порядка на
разных стадиях профазы I мейоза; 6 — сперматоцит II порядка в
интерфазе; 7 — ранние
и 8 — поздние сперматиды; 9— спермии; 10
— поддерживающая и
11 — делящиеся клетки.
Отличительная особенность сперматогоний, вступивших на путь дифференцировки, — неполная цитотомия, в результате чего сестринские и дочерние сперматогоний оказываются связанными между собой цитоплазматическими мостиками, то есть являются синцитием (рис. 15). Мостики (фузомы) достаточно длинные, так что клетки могут отходить друг от друга на
расстояние до 25 мкм. Все связанные между собой клетки образуют клон.
Клетки одного клона синхронно проходят стадии сперматогенеза. После завершения сперматогониальных делений все клетки клона вступают в стадию
роста.
В стадии роста половая клетка называется сперматоцит I порядка. В это
время в клетке идут активные процессы ассимиляции, дифференцировки органелл, синтез ДНК, размножение митохондрий делением, увеличивается ядрышко. Клетка округляется, оттесняется от базальной мембраны по направwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лению к просвету канальца, увеличивается в размерах. По количеству хромосом сперматоцит I порядка считается диплоидным, а по содержанию ДНК —
тетраплоидным, так как каждая хромосома состоит из двух хроматид. В
крупном округлом ядре его последовательно происходят преобразования
хромосом, характерные для профазы I мейоза. Потеряв связь с базальной
мембраной семенного извитого канальца, сперматоциты I порядка вступают
в тесный контакт с поддерживающими клетками, которые называют также
клетками Сертоли, фолликулярными, питающими, выстилающими, защитными и т. д. Это однослойный эпителий из очень крупных клеток, высотой во
всю толщину стенки канальца с бледной цитоплазмой и ядрами неправильной формы, лежащими у базального полюса, с большим количеством ветвистых отростков. Поддерживающие клетки выполняют разнообразные функции (опорную, трофическую, защитную, гормональную, разграничительную,
фагоцитарную, регуляторную, секреторную и др.) и строение их подвержено
циклическим изменениям в зависимости от стадии развития половых клеток.
Рис. 15. Взаимоотношения между поддерживающими
и половыми клетками:
1 — сперматогоний; 2 — сперматоцит I порядка; 3— сперматида поздняя; 4
— мостик; 5 — поддерживающая клетка (сертоли); 6 — ее ядро; 7 —
плотный контакт между двумя клетками сертоли.
В стадии созревания происходит редукция числа хромосом в половых
клетках в процессе двух делений созревания, быстро следующих друг за другом, то есть в процессе мейоза. В результате первого деления созревания —
редукционного деления мейоза из каждого сперматоцита I порядка образуетwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ся два сперматоцита II порядка. Это короткоживущие клетки, более мелкие,
чем сперматоциты I порядка. Они содержат гаплоидное число хромосом (но
каждая хромосома состоит из двух хроматид) и тут же вступают во второе
деление созревания — эквационное деление мейоза. Между делениями созревания интерфаза очень короткая. В ней отсутствует S-период, нет деконденсации ДНК. А так как хромосомы сохраняют бивалентную структуру,
профаза II проходит очень быстро. В результате второго деления из каждого
сперматоцита II порядка образуются две сперматиды — истинно гаплоидные
клетки. У них в сравнении с соматическими клетками уменьшено вдвое не
только количество хромосом, которое гаплоидно (1n), но и количество ДНК
(1с). Кроме того, сперматиды различаются по половым хромосомам: половина их содержит Х-хромосому, а половина Y-хромосому. Остальные хромосомы их отличаются и от родительских хромосом и друг от друга в разных
клетках, так как произошел случайный обмен генами во время кроссинговера.
Сперматоциты II порядка, и особенно сперматиды, находятся в тесном
контакте с клетками Сертоли. В процессе созревания они перемещаются все
ближе к просвету канальца и все глубже проникают в цитоплазму поддерживающей клетки.
1. Характеристика спермиев млекопитающих
Вид животного
Скорость
Возраст по- Возраст наиДлина
Длина
Количество в
движеявления зревысшей
спермия, головки,
эякуляте,
ния, мкм,
лых сперми- половой акмкм
мкм
млрд
с
ев, мес
тивности
Жеребец
Бык
Баран
Хряк
35—62 5—10
51—63 5,6—8,5
55—60
4—5
37—62 7—10
90
120
50
—
4—20
5—12
2—10
25—80
_
6
4—5
4
_
2—8 лет
С 1,5 года
1—2 года
В стадии формирования сперматиды превращаются в спермий (сперматозоид). Данную стадию проходят одновременно все сперматиды одного
клона. На нее приходится около 1/3 времени всего сперматогенеза. Процессы, происходящие в половых клетках в стадии формирования, подробно изучены у некоторых видов млекопитающих (в основном у лабораторных животных). При этом выделено 19 этапов, которые проходит сперматида, прежде чем превратится в спермий. Сперматиды, только что появившиеся в результате второго деления созревания, — ранние сперматиды—это мелкие
округлые клетки с небольшим округлым ядром, содержащим мелкогранулярный хроматин. В их цитоплазме имеются все органеллы общего значения.
Сперматиды в период формирования располагаются в карманах — углублениях на апикальном полюсе клетки Сертоли, которые, вероятно, вырабатывают гормон, регулирующий их дифференцировку.
В процессе дифференцировки на переднем полюсе сперматиды в области комплекса Гольджи развивается акросома — специфическая органелла,
обеспечивающая растворение оболочек яйцеклетки благодаря содержанию
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
большого количества гиалуронидазы и других гидролитических ферментов.
Акросома примыкает к ядру и по мере развития прикрывает его. Ядро вытягивается, уплощается и резко уплотняется. Объем его уменьшается в 20—50
раз. Происходит конденсация и специальная упаковка хромосом с помощью
белков — протаминов. Упаковка эта настолько плотная, что дезоксирибонуклеопротеиды приобретают даже некоторые свойства кристаллов. От центриоли отрастает жгутик, направленный в сторону, противоположную акросоме — к просвету канальца. Клетка принимает грушевидную форму. Часть
протоплазмы сползает со сперматиды и фагоцитируется клеткой Сертоли.
Сперматида все более удлиняется. Такая клетка называется поздней сперматидой. Когда процесс дифференцировки сперматиды завершается, она теряет
связь с клеткой Сертоли и другими клетками своего клона и становится
спермием.
Строение спермия. Спермий — свободная,
подвижная клетка, длиной у млекопитающих
40—60 мкм (табл. 1). Состоит из головки, шейки и хвоста. Все его части покрыты цитолеммой
(рис. 16).
Головка имеет видоспецифическую форму.
У сельскохозяйственных животных она лепешкообразная. Акросома в виде шапочки покрывает головку на 2/3. Основу головки составляет
ядро, в котором содержатся ДНК в виде плотно
упакованных хромосом. Цитоплазма очень тонким слоем покрывает ядро.
Шейка — самый короткий отдел, видимый
только в электронный микроскоп. В ней располагается центросома из двух центриолей.
Проксимальная центриоль — та, что ближе к
Рис. 16. Строение
ядру, лежит поперек шейки, а дистальная — боспермия:
I — головка; II — шей- лее удаленная от ядра — вдоль шейки. От нее
ка; III — средний, IV — берет начало осевая нить (аксонема) жгутика.
Хвост составляет основную длину клетки.
главный и V — концевой
отделы хвоста; 1 — ак- Он состоит из среднего (промежуточного),
росомная шапочка; 2 — главного и концевого отделов. На протяжении
ядерный материал; 3 — всего хвоста проходит осевая нить. Она попроксимальная центри- добно ресничкам состоит из двух центральных
оль; 4 — дистальная и девяти пар периферических микротрубочек.
центриоль; 5 — осевая Микротрубочки построены из белков тубулина
нить (аксонема); 6 — и динеина, благодаря которым осуществляется
плотные фибриллы; 7 — волнообразное и бичевидное движение жгутиспиральная нить; 8 — ка. В среднем и главном отделах хвоста микрозамыкающее
кольцо; трубочки сопровождаются расположенными
9—ундулирующая мем- снаружи девятью дополнительными плотными
фибриллами. В среднем отделе хвоста поверх
брана.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
фибрилл лежат митохондрии, упакованные в виде спирали. Это так называемая спиральная нить спермия.
Средний отдел от главного отделяется замыкающим кольцом. Цитоплазма тонким слоем покрывает весь хвост, за исключением концевого отдела (последних 5—7 мкм). Иногда цитоплазма образует 1—2 гребневидных
выроста — ундулирующую мембрану, дополнительное приспособление для
передвижения.
Созревание спермия. Сформированные спермин млекопитающих, находящиеся в семенниках, еще не могут выполнять специфическую функцию —
они не обладают способностью к активному движению и оплодотворению.
Из семенного извитого канальца спермин выносятся с током жидкости, заполняющей его просвет. Процесс созревания спермия продолжается во время
его перемещения в половых путях самца и самки, вплоть до момента оплодотворения. При этом меняется строение оболочки спермия, его физикохимическое состояние. Так, пройдя придаток семенника спермий приобретают способность к активному движению, их оболочка покрывается углеводнобелковым слоем. В половых путях самки в спермиях происходят изменения,
способствующие контакту спермия с яйцеклеткой, акросомной реакции и оплодотворению. При этом жизнеспособность спермиев в половых путях самки
первоначально даже повышается (у спермиев быка в течение двух суток)—
увеличивается потребление кислорода, вырабатывается больше энергии. Однако в дальнейшем жизнеспособность, оплодотворяющая способность спермиев резко падает — происходит их старение. У различных животных оптимальный срок пребывания спермиев в половых путях самок разный. У млекопитающих — это от одного до нескольких дней, у птиц — от одной до нескольких недель, у матки медоносной пчелы — до 3—5 лет.
При оплодотворении «старыми» спермиями увеличивается процент эмбриональной смертности, уродств, снижается оплодотворяемость. Считают,
что это вызвано излишней прочностью связи ДНК с белками в хромосомах у
«старых» спермиев. Подобное следует учитывать при осеменении животных,
синхронизируя время охоты и покрытия самок.
Влияние различных факторов на сперматогенез. Продолжительность
сперматогенеза постоянная для данного вида животных и не зависит от действия эндогенных и экзогенных факторов. У быка она равна 62—63 дня, у
барана — 47—48 дней, у хряка — 39—40 дней. Что же касается активности
сперматогенеза, качества и количества развивающихся половых клеток, то
здесь прослеживается заметное влияние различных факторов.
В процессе сперматогенеза из одной сперматогоний, вступающей на
путь дифференцировки, теоретически может образоваться до 36 768Х215
спермиев. Однако этого не происходит, так как в сперматогенезе отмечено
несколько волн гибели клеток, которые приходятся на стадии размножения
(во время профазы митоза в сперматогониях), созревания (во время деления
созревания) и формирования (во время конденсации хроматина в сперматидах). В результате спермиев образуется 50—70% от теоретически рассчитываемого количества. Причиной дегенерации клеток в период сперматогенеза
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
считаются спонтанные нарушения хромосом в процессе деления. Высказывается также мнение, что на гибель клеток влияет нарушение соотношения между половыми и поддерживающими клетками.
Различные внешние воздействия: жесткое ультрафиолетовое, рентгеновское облучение и другие виды проникающей радиации, низкая или высокая
температуры, обездвиживание, удаление гипофиза, гипоксия, введение гормонов (эстрогенов) и других биологически активных веществ, общие тяжелые заболевания, травма и пересадка семенника, крипторхизм приводят к однотипной реакции. Все эти и многие другие факторы вызывают или тотальную гибель сперматогенного эпителия, или клеток на определенной стадии
развития и тем угнетают сперматогенез; увеличивают число атипичных клеток, неспособных к оплодотворению (с измененным ядром, центросомой, с
измененным содержанием ДНК, с нарушениями веретена деления, митоза и
мейоза). Наиболее чувствительны клетки в период конъюгации хромосом
(профаза I мейоза) и во время второго деления созревания. С. С. Райцина на
основании многочисленных исследований приходит к выводу, что однотипная реакция сперматогенного эпителия на повреждающие внешние воздействия вызвана нарушением гематотестикулярного (haema — кровь, testis — семенник) барьера, регулирующего поступление из крови в семенные канальцы
сывороточных белков, биологически активных веществ, метаболитов, токсинов, лекарственных препаратов. Барьерную функцию в семеннике выполняют: белочная оболочка, собственная оболочка семенных извитых канальцев и клетки Сертоли. Нарушение гематотестикулярного барьера приводит к
гибели или атипичному развитию сперматогенного эпителия. Гематотестикулярный барьер у новорожденных животных несовершенен и достигает
полного развития к началу пубертатного периода, что следует, учитывать при
выращивании самцов-производителей. В консервирующих смесях спермин
могут долго сохранять жизнеспособность. Данное качество используется при
искусственном осеменении животных. Есть опыт хранения спермы быка до
25 лет, барана — до 11, хряка — до 3 лет. Это имеет большое значение для
развития животноводства, а также для сохранения генофонда редких и исчезающих видов.
Развитие женских половых клеток — оогенез
Женская половая клетка — яйцеклетка (ovum — яйцо) — была открыта
К. Бэром в 1825 г. Развитие женских половых клеток— оогенез, происходит в
яичнике и завершается в яйцеводе. В оогенезе различают три стадии: размножения, роста и созревания (см. рис. 13).
Стадию размножения женских половых клеток у млекопитающих отмечают в утробный период жизни особи. Первичные половые клетки, вселившись в закладку яичника, превращаются в оогонии и много раз подряд делятся митозом — идет накопление половых клеток. Оогонии — круглые или
овальные клетки с оксифильной цитоплазмой и крупным ядром, в котором
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
видны 1—2 ядрышка и мелкодисперсный хроматин. Среди соматических
клеток яичника они выделяются своими крупными размерами.
Рис. 17. Рост и созревание женских половых клеток в яичнике (А)
и яйцеводе (Б):
I — корковое вещество яичника; II — мозговое вещество; III — воронка
и IV — просвет яйцевода; 1 — покровный эпителий; 2 — белочная оболочка;
3— интерстициальная ткань; 4 — первичный (примордиальный) фолликул; 5
—растущий (вторичный) фолликул; 6 — пузырчатый фолликул; 7 — тека
фолликула; 8 — фолликулярный эпителий; 9 —зернистый слой; 10— полость
фолликула; 11 — лучистый венец; 12 — блестящая оболочка; 13 — ооцит I
порядка; 14 — овулированный фолликул; 15 — желтое тело; 16 — беловатое
тело; 17 — атретический фолликул; 18 — сосуды; 19 — ооцит II порядка;
20 — зрелая яйцеклетка; 21 —полярное тельце; 22 — зигота.
К концу плодного периода у млекопитающих большинства видов митотическая активность оогониев затухает. У кроликов и свиней, по данным некоторых исследователей, размножение оогониев продолжается некоторое
время после рождения (у свиней — в течение первого месяца). Прекратив делиться, половые клетки вступают в стадию роста (рис. 17).
Стадия роста оогенеза проходит в яичнике, отличается длительностью и
сложностью преобразований. Половая клетка на этой стадии называется
ооцитом I порядка. В его ядре происходят преобразования, характерные для
профазы I мейоза. У большей части ооцитов I порядка отмечают синхронные
последовательные стадии профазы I, вплоть до диплонемы. При этом половая клетка окружается одним слоем клеток — фолликулярным эпителием.
Образуется многоклеточная структура — фолликул. Фолликул, стенки которого образованы одним слоем плоских фолликулярных клеток, плотно прилегающих к ооциту I порядка, называется примордиальным, а затем, когда
фолликулярные клетки увеличиваются в размерах, — первичным. В первые
дни после рождения ооциты проходят диплонему и вступают в проwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
должительную интерфазную стадию диктиотены, предшествующую диакинезу. Она длится до полового созревания и заканчивается незадолго перед
овуляцией. В это время хромосомы становятся невидимыми, их ДНК деконденсируется, активно идет синтез всех видов РНК, различных белков, увеличивается количество органелл, большинство которых располагается вокруг
ядра. В результате всех этих процессов происходит рост ооцита I порядка.
Рост совершается благодаря ассимиляционной деятельности половой клетки.
Идет он медленно, продолжается долго— месяцы и годы и называется периодом малого, или цито-плазматического роста. Первичные фолликулы с
ооцитами I порядка в стадии малого роста располагаются по периферии корковой зоны яичника. У взрослой коровы они составляют почти 55% от всех
фолликулов яичника. Постепенно фолликулярные клетки увеличиваются в
размерах и начинают активно функционировать. Такой фолликул называется
растущим (вторичным). У коровы вторичные фолликулы составляют 43%.
При совместной деятельности фолликулярных клеток и ооцита I порядка вокруг половой клетки формируется блестящая оболочка — вторичная оболочка половой клетки. Через ее поры устанавливается тесная связь половой и
фолликулярных клеток. Последние, размножаясь, покрывают ооцит I порядка
несколькими слоями. Особенно активен фолликулярный эпителий в период
накопления желтка — в период вителлогенеза (vitellus — желток, genesis —
происхождение), который начинается при половом созревании. С помощью
фолликулярного эпителия идет быстрое накопление ооцитом I порядка белковых и других веществ (вителлогенина, фосвитина, липовителлина, вителлина), синтезированных за пределами яичника, в основном в печени и принесенных кровью.
Период вителлогенеза называют еще периодом большого роста, или
трофоплазматическим. У свиньи он протекает в фолликулах с диаметром от
4 до 8 мм. В это время происходит пропорциональное увеличение ооцита I
порядка и фолликула. Фолликулярные клетки осуществляют отбор и концентрацию вокруг ооцита молекул, подлежащих эндоцитозу. Они же, повидимому, выделяют вещества, стимулирующие микропиноцитоз в ооците и
гормоны, стимулирующие синтез вителлогенина печенью. Живые неокрашенные ооциты I порядка в период большого роста имеют коричневый цвет.
Органеллы в них располагаются в основном по периферии цитоплазмы, где в
первую очередь идет отложение желточных гранул и других включений. Постепенно желток заполняет большую часть цитоплазмы ооцита, вителлогенез
затухает. Рост ооцита прекращается, а фолликул продолжает расти.
Между фолликулярными клетками появляется полость, в которой накапливается жидкость, содержащая половые гормоны (эстрогены) и другие
продукты деятельности фолликулярного эпителия. Фолликул становится пузырчатым (Граафов пузырек или третичный фолликул). У коровы полость
начинает появляться в фолликулах, достигших диаметра 1 мм. Место, где в
стенке фолликула расположен ооцит I порядка, называется яйценосным бугорком. Ооцит покрыт слоем фолликулярных клеток или лучистым венцом,
который у сельскохозяйственных животных образован 8—12 рядами клеток.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Остальные фолликулярные клетки формируют зернистый слой фолликула—
гранулезу. Поверх него фолликул одет соединительнотканной оболочкой —
текой. Тека выполняет опорную и трофическую функции, в ней много сосудов и нервов, залегают клетки, вырабатывающие половые гормоны. Созревший пузырчатый фолликул имеет макроскопические размеры (табл. 2).
2. Размеры женских половых клеток и фолликулов
на разных стадиях развития (по данным разных авторов)
Вид животСтруктура
Диаметр
ного
Яйцеклетка
113—135 мкм
Кобыла
Яйцеклетка с оболочками
160 мкм
Созревший фолликул
От 8 до 70 мм
Яйцеклетка
122—140 мкм
Корова
Яйцеклетка с оболочками
157—183 мкм
Созревший фолликул
18 мм
Яйцеклетка
120 мкм
Овца
Созревший фолликул
6—7 мм
Оогония
12 мкм
Ооцит в профазе мейоза
18—25 мкм
Свинья
Яйцеклетка
120—140 мкм
Яйцеклетка с оболочками
146 мкм
Созревший фолликул
8—11 мм
Незадолго до конца стадии роста хромосомы в ядре ооцита I порядка
конденсируются и принимают вид, характерный для диакинеза профазы I,
после чего мейоз идет без остановки.
В определенный момент полового цикла под влиянием нейроэндокринной регуляции расширяются кровеносные сосуды теки, резко возрастает давление жидкости внутри фолликула. Стенка его разрывается, и половая клетка вместе с лучистым венцом выпадает в яйцевод. Этот процесс
называется овуляцией.
Стадия созревания у большинства млекопитающих начинается в яичнике и завершается (с метафазы II) в яйцеводе. Исключение составляют лошадь,
собака, у которых половая клетка овулирует стадии диакинеза профазы I. В
стадии созревания половая клетка делится мейозом. В результате первого деления созревания— редукционного деления мейоза из одного ооцита I порядка— образуются две неравнозначные клетки. Одна — крупная половая
клетка, содержит всю цитоплазму и половину хромосом. Это ооцит II порядка. Другая — мелкая, содержит половину хромосом и почти не имеет цитоплазмы, первое редукционное, или полярное, тельце. Сразу после завершения
первого деления созревания наступает второе деление созревания — эквационное деление мейоза, результатом которого вновь являются две неравноценные клетки: зрелая яйцеклетка и второе редукционное тельце. Иногда
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
первое редукционное тельце также делится эквационно. Импульсом для деления созревания у многих животных является совокупление или проникновение спермия в оболочки яйцеклетки. Если этого не произойдет, половая
клетка гибнет на стадии метафазы II. Все редукционные тельца в дальнейшем
отмирают. Таким образом, в оогенезе из одного ооцита I порядка образуется
лишь одна, но очень крупная половая клетка — яйцеклетка, содержащая гаплоидный набор хромосом и полный набор факторов, необходимых для начальных этапов развития зародыша.
Строение зрелой яйцеклетки. Яйцеклетка, как и любая клетка, состоит
из ядра, цитоплазмы и цитолеммы. В то же время по своим свойствам и
структуре она коренным образом отличается от всех других клеток организма.
Ядро яйцеклетки небольших размеров, неактивное, с гаплоидным набором хромосом. Ядерно-цитоплазматическое отношение сильно сдвинуто в
сторону преобладания цитоплазмы. В цитоплазме яйцеклетки происходит
развитие морфогенетической организации, формирование специализированной кортикальной зоны. В ней накоплены разнообразные вещества, необходимые для ранних стадий развития зародыша: все виды РНК, многообразные
ферменты, структурные белки, энергетические резервы, множество факторов,
выполняющих регуляторные функции, благодаря деятельности которых как
бы развертывается план строения раннего зародыша. Особенно в больших
количествах накапливается желток, в состав которого входят белки (вителлин), углеводы и липиды, образуя комплексные соединения (фосвитин, липовителлин и др.). 80% белков цитоплазмы яйцеклетки приходится на долю
желтка, 16% составляют структурные белки рибосом.
Размеры яйцеклетки определяются количеством желтка. Количество
желтка зависит от условий и длительности развития зародыша. У животных с
личиночным (беспозвоночные), коротким (ланцетник) и внутриутробным
(млекопитающие) развитием яйцеклетки содержат мало желтка и называются
олиголецитальные. У животных с прямым наружным развитием (рептилии,
птицы) яйцеклетки полилецитальные, в них много желтка. Если желток расположен в яйцеклетке равномерно, она относится к изолецитальному типу
(ланцетник), и если скапливается у одного полюса — к телолецитальному
типу (птицы). Олиголецитальные яйцеклетки гораздо меньше полилецитальных. Масса яйцеклеток у млекопитающих в среднем 3×10-6 г, диаметр 60—
180 мкм, у ланцетника — 100 мкм, у домашних птиц — 30—50 мм, у страуса— 105 мм, а у одного вида акул — до 220 мм.
Вещества и органеллы цитоплазмы яйцеклетки не разбросаны в беспорядке, а организованы столь определенным образом, что эта структурная организация во многом определяет последующие этапы эмбрионального развития. Вот один пример. Полюс, у которого скапливается желток, называется
вегетативным. Ядро яйцеклетки при этом располагается у противоположного полюса— анимального. Данными экспериментальной эмбриологии установлено, что оси будущего зародыша определяются этой полярностью. Анимальный полюс становится головным концом зародыша, вегетативный —
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
хвостовым. По особенностям структурной организации разных участков цитоплазмы яйцеклетки можно определить брюшную и спинную стороны будущего зародыша, плоскость билатеральной симметрии и другие показатели.
В периферическом слое цитоплазмы ооцита, или в кортикальном, располагается большое количество микрофибрилл, делающих его вязким. Здесь
же скапливаются так называемые кортикальные гранулы, или кортикальные
вакуоли, которые содержат гликозаминогликаны. Кортикальный слой цитоплазмы, формирующийся еще в период роста ооцита, играет важную роль
при оплодотворении.
Яйцеклетка покрыта оболочками. Первичная оболочка синтезируется
самим ооцитом в период роста и называется желточной, или вителлиновой.
Она тонким слоем покрывает ооцит, располагаясь между его микроворсинками. К ней тесно прилегает вторичная оболочка, образованная деятельностью фолликулярных клеток. У млекопитающих первичная и вторичная оболочки столь тесно прилежат и пронизывают друг друга, что известны под общим названием блестящей, или прозрачной, оболочки. Строение ее у сельскохозяйственных животных сходно, толщина 10—: 20 мкм. Она является
мощным иммунным барьером, обладает избирательной проницаемостью для
спермиев. Третичная оболочка продуцируется железами яйцевода и покрывает яйцеклетку, когда она движется по яйцеводу. Третичная оболочка может
иметь сложное строение. У птиц, например, она состоит из белка, нодскорлуповой, скорлуповой и надскорлуповой оболочек, образующихся в разных
отделах яйцевода.
Влияние различных факторов на оогенез. В процессе оогенеза на всех
стадиях развития происходит гибель какой-то части половых клеток под
влиянием внутренних, во многом не выясненных, факторов. Отмечено три
волны массовой дегенерации: на стадии размножения оогонии, ооцитов I порядка в стадии пахитены и в стадии диплотены. В результате число половых
клеток к рождению уменьшается в 7—10 раз. Гибель клеток (так называемая
атрезия) продолжается и в дальнейшем. Считается нормальным, если из 20
растущих фолликулов до овуляции дорастает один. У коров критическим
моментом в стадии роста является также период достижения фолликулом
размеров от 6 до 9 мм. В это время происходит атрезия 55% пузырчатых
фолликулов. По-видимому, так же как в сперматогенезе, основная причина
гибели половых клеток в оогенезе — нарушения в хромосомах и веретене деления. В результате оогония или ооцит теряют способность к дальнейшему
развитию и погибают. К атрезии фолликулов ведет и нарушение гормональных взаимоотношений (между эстрадиолом, тестостероном и прогестероном)
в процессе оогенеза.
Большинство повреждающих внешних факторов (высокая или низкая
температура, действие химических веществ, электрошок) нарушает течение
митоза или мейоза в результате разрушения веретена деления. Ионизирующая радиация и некоторые химические вещества воздействуют непосредственно на хромосомы. Погибают половые клетки или образуются их атипичные формы. Следствием этого может быть бесплодие, снижение плоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
довитости, повышенная эмбриональная смертность и появление уродств у
потомства. Половые клетки плодов и новорожденных животных более чувствительны к повреждающим факторам, чем у взрослых животных.
В современном животноводстве все шире применяются различные методы стимуляции многоплодия и повышения оплодотворяемости. От одной
коровы, стимулируя суперовуляцию, культивируя овулировавшие ооциты и
пересаживая их другим коровам, можно в год получить до 60 телят (Ельчанинов, 1978). Однако на пути реализации этой заманчивой перспективы стоит много нерешенных вопросов. Дело в том, что при искусственно вызываемой полиовуляции (суперовуляции) нарушается синхронность между процессами роста, созревания и овуляции ооцитов. В яйцевод попадают половые
клетки разной степени зрелости. Это приводит к большой гибели эмбрионов
на ранних стадиях развития. У крупного рогатого скота при многоплодии
часто появляются «интерсексы», особенно у молочных пород. Еще меньше
изучены вопросы культивирования и пересадки яйцеклеток.
ОПЛОДОТВОРЕНИЕ И ЕГО БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
Оплодотворение — сложный комплекс физиологических процессов и
реакций, приводящий к слиянию мужской и женской половых клеток в одну
клетку — зиготу (zygotes — объединенный). В нем можно выделить осеменение и собственно оплодотворение.
Осеменение — это биологические процессы, обеспечивающие сближение гамет.
Собственно оплодотворение протекает в четыре стадии: 1) сближение
половых клеток; 2) проникновение спермиев в блестящую оболочку; 3) проникновение в цитоплазму яйцеклетки; 4) слияние ядер половых клеток (рис.
18). В процессе оплодотворения две высокоспециализированные клетки —
спер-мий и яйцеклетка — взаимодействуют друг с другом специфическим
образом. Функция спермия — обеспечение встречи с яйцеклеткой, побуждение ее к развитию и внесение в нее отцовского наследственного материала.
Функция яйцеклетки — обеспечение активации ядра спермия, привнесение
материнского наследственного материала, накопление трофического, энергетического и структурного материала, а также регуляторных факторов (пока
еще не выясненной природы), необходимых для начальных этапов развития
зародыша.
У крупного и мелкого рогатого скота осеменение влагалищное, у лошадей и свиней — маточное. При этом во влагалище (у коровы, овцы и козы)
или шейку матки (у кобылы и свиньи) попадают миллиарды спермиев (см.
табл. 1). В половых путях самок спермин движутся благодаря реотаксису
(способности двигаться против тока жидкости), а также присасывающего
действия, производимого сокращением мускулатуры матки и яйцеводов. В
результате через несколько часов (6—7 — у овец, 5—9—1 у коров) спермин
оказываются в верхней трети яйцевода. Но доходит туда меньше 1 % спермиев. Основная их масса гибнет в пути: внедряются в слизистую оболочку поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ловых путей, фагоцитируются лейкоцитами, склеиваются специальными веществами (фертилизинами), выделяемыми яйцеклеткой. Таким образом происходит как бы отбор спермиев, в процессе которого погибают атипичные и
маложизненные клетки.
Рис. 18. Схема процесса оплодотворения:
1 —спермий; 2— блестящая оболочка; 3 — кортикальный слой цитоплазмы яйцеклетки; 4 — акросомная реакция; 5 — проникновение спермия
через блестящую оболочку; 6 — слияние плазмолемм спермия и яйцеклетки;
7 — проникновение спермия в цитоплазму яйцеклетки; 8 — превращение головки спермия в мужской пронуклеус; 9 — сближение женского и мужского
пронуклеусов; 10— слияние пронуклеусов; 11 — зигота (а и б —редукционные
тельца).
Спермий с яйцеклеткой должны встретиться в верхней трети яйцевода.
Если этого не произойдет и яйцеклетка до встречи со спермиями пройдет
больший путь, она стареет и к концу яйцевода теряет способность к оплодотворению. Чтобы произошло оплодотворение, верхней трети яйцевода должно достигнуть по крайней мере несколько тысяч или десятков тысяч спермиев, так как эти клетки вступают в контакт с яйцеклеткой в результате случайного столкновения. Это следует учитывать при разбавлении спермы во время
искусственного осеменения. Яйцеклетки выделяют вещества, ускоряющие
движение и продлевающие период подвижности спермиев, но не выделяют
специальных привлекающих веществ (хемотаксис отсутствует). Вероятность
встречи увеличивается и тем, что половые клетки имеют разный электропотенциал: положительный у спермиев и отрицательный у яйцеклетки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У большинства сельскохозяйственных животных лучистый венец яйцеклетки, попавшей в яйцевод, начинает разрушаться еще до встречи со спермиями. Однако под влиянием спермиев этот процесс совершается быстрее.
На первой стадии оплодотворения — при контакте яйцеклетки со спермиями в последних происходит акросомная реакция: частично разрушается
цитолемма и наружная оболочка акросомы. Гиалуронидаза, заключенная в
акросомах спермиев, изливается и разрушает остатки лучистого венца. Он
рассыпается, блестящая оболочка яйцеклетки разрыхляется. Эта реакция неспецифическая, в ней могут участвовать и спермий других видов животных
(что используется порой в животноводстве для экономии семени особо ценных производителей при искусственном осеменении).
Вторая стадия оплодотворения видоспецифична. В ней участвует несколько десятков спермиев своего вида. Они проходят сквозь прозрачную
оболочку благодаря ферментам, связанным с внутренней мембраной акросомы. Приблизившись к поверхности яйцеклетки, один из спермиев сливается с
ее цитолеммой боковой поверхностью головки — наступает третья стадия
оплодотворения.
В месте контакта спермия и яйцеклетки частично расплавляются мембраны и цитоплазмы клеток сливаются. Мембрана спермия входит составной
частью в мембрану яйцеклетки. Внутрь яйцеклетки спермий может войти целиком, а может только головкой, шейкой и средним отделом хвоста (у свиньи).
Слияние мембран спермия и яйцеклетки служит своего рода толчком
для пробуждения яйца. С этого момента снимается блокада мейоза и он продолжается с той стадии, на которой остановился во время овуляции (завершается первое и начинается второе деление созревания). Резко активизируется метаболизм яйцеклетки. Развивается кортикальная реакция. Она состоит в
том, что кортикальные гранулы подходят к цитолемме и их содержимое изливается в сторону прозрачной оболочки. Образуется перивителлиновое пространство, в результате чего яйцеклетка отслаивается от прозрачной (желточной) оболочки. Часть гранул входит в состав желточной оболочки, которая при этом утолщается и преобразуется в оболочку оплодотворения. Материал кортикальных гранул видоизменяет и цитолемму яйца. Она становится
плотной и не проницаемой для других спермиев, так как покрывается гиалиновым слоем. Тем самым кортикальная реакция является механизмом, предохраняющим от проникновения сверхчисленных спермиев, и образует оболочку, под которой развивается зародыш до тех пор, пока он не покидает яйцовые оболочки.
Кортикальная реакция начинается от места слияния мембран спермия и
яйцеклетки и распространяется отсюда по всей поверхности яйца. У олиголецитальных яиц это происходит быстро, в результате чего обеспечивается
моноспермия — только один спермий попадает в яйцеклетку. У животных с
полилецитальными яйцами (птицы, рептилии) наблюдается физиологическая
полиспермия (кортикальная реакция у них развивается медленно и несколько
спермиев могут за это время проникнуть внутрь яйца). Однако и в том и друwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
гом случае четвертая стадия оплодотворения (слияния ядер половых клеток)
протекает аналогично. С ядром яйцеклетки сливается ядро лишь одного
спермия.
Дальнейшая судьба частей спермия в цитоплазме яйцеклетки различна.
Одни включаются в развитие, другие дегенерируют. Ядро спермия преобразуется в мужской пронуклеус (предъядрие) и проникает все глубже в цитоплазму, в нем за 20—30 мин реорганизуется и реактивируется хроматин, а
его объем увеличивается в 50 раз. К моменту образования мужского пронуклеуса в ядре яйцеклетки завершается второе деление созревания, выделяется
второе редукционное тельце и ядро преобразуется в женский пронуклеус.
Пронуклеусы сближаются друг с другом. В это время в них происходит редупликация ДНК, а после ее завершения— профаза митоза. На основе центриолей центросомы формируется веретено деления. Сблизившиеся пронуклеусы сливаются. Конечная стадия их слияния — образование единой метафазной экваториальной пластинки уже не отдельных половых клеток, а зиготы. Оплодотворение завершилось — началось развитие нового организма.
Значение оплодотворения состоит в том, что оно приводит к слиянию
отцовского и материнского геномов и таким образом является средством передачи наследственных свойств новому организму. В результате в стаде, породе, популяции возникает множество новых комбинаций наследственных
факторов, создается многообразный генофонд, который служит материалом
для отбора в ходе хозяйственной деятельности человека.
Клетка, появившаяся в результате оплодотворения—зигота, отличается
от яйцеклетки своими биологическими, морфологическими и физикохимическими свойствами. Она обладает большой жизненностью, более широкой наследственной основой. Однако эти качества не абсолютны и во многом зависят как от возраста и состояния здоровья родителей, так и от сроков
осеменения и оплодотворения. Близкородственное спаривание (инбридинг),
как правило, приводит к появлению малочисленного слабого потомства. С
возрастом матери увеличивается количество хромосомных нарушений. Несвоевременное осеменение приводит к «старению» половых клеток и, как
следствие этого, к снижению их жизнеспособности и способности к оплодотворению.
Вопросы для самопроверки. 1. Опишите процессы, происходящие при
сперматогенезе и оогенезе, и влияние на них внешних и внутренних факторов. 2. Какие черты сходства и различия сперматогенеза и оогенеза вы знаете? 3. В чем особенность стадии роста оогенеза? 4. Каково строение зрелого
спермия и зрелой яйцеклетки? 5. Значение оплодотворения и его морфология.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Глава 5. РАННИЕ ЭТАПЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ.
ДРОБЛЕНИЕ
Сразу после образования зигота приступает к делению митозом. Деления быстро следуют одно за другим, митотический цикл клеток короткий,
они не дорастают до размеров материнской, поэтому дочерние клетки с каждым делением становятся все мельче. Такой процесс называется дроблением,
а клетки—бластомерами. Ядра бластомеров не уменьшаются, так как каждому делению предшествует редупликация ДНК. Ясно, что при каждом делении соотношение ядра и цитоплазмы в клетках изменяется. Дробление
прекращается тогда, когда в бластомерах устанавливается ядерно-плазменное
соотношение, типичное для соматических клеток данного вида животного. В
клетках начинается активный белковый синтез, скорость деления снижается,
каждая дочерняя клетка дорастает до размеров материнской, а размеры зародыша резко возрастают.
Скорость и характер дробления неодинаковы у разных животных и зависят от количества желтка в яйцеклетке. Олиголецитальные яйцеклетки (млекопитающие) привносят в зиготу мало желтка, он не мешает зиготе делиться
полностью — от анимального до вегетативного полюса. Если при этом яйцеклетка была изолецитальная (ланцетник) —бластомеры получаются одинакового размера. Такое дробление называется полным равномерным. В телолецитальных яйцах бластомеры разных размеров, так как желток, скопившийся
у вегетативного полюса, мешает делению и здесь образуются более крупные
клетки, как, например, у амфибий. Это полное неравномерное дробление. Если яйца полилецитальные (рыбы, рептилии, птицы), крупные массы желтка
настолько мешают дроблению, что делятся лишь клетки анимального полюса. Вся остальная масса остается в виде нераздробившегося желтка. Такой
тип дробления называется частичным или дискоидальным (бластомеры в
процессе дробления располагаются в форме диска).
В период дробления клетки делятся в определенном порядке. Первая и
вторая борозды дробления идут от анимального к вегетативному полюсу и
называются меридианальными. Третья борозда — широтная. Дальше идут
чередования меридианальных, широтных и тангенциальных (параллельно
поверхности) борозд, в результате чего правильность расположения бластомеров нарушается. Дольше всего правильность их расположения сохраняется
при полном равномерном дроблении. Быстрее всего она нарушается при частичном дроблении (цв. табл. I) уже после 2—3-го деления.
Бластомеры во время дробления тесно связаны друг с другом, образуется как бы плотный комок из клеток — морила, а затем пузырек с полостью —
бластула. Стенка бластулы называется бластодерма, а полость — бластоцель. Клетки бластодермы, расположенные на месте бывшего анимального
полюса, образуют крышу, а расположенные на месте вегетативного полюса
— дно бластулы. Между ними находится краевая зона. При полном равномерном дроблении изоолиголецитальных яиц образуется бластула с бластодермой из одного слоя почти равных клеток и большим бластоцелем. Это цеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лобластула (ланцетник). При полном неравномерном дроблении олиголецитальных яиц формируется бластула с очень маленьким бластоцелем — стерробластула (млекопитающие). Из мезолецитальных яиц амфибий образуется
бластула с толстым дном и небольшим бластоцелем, сдвинутым к крыше —
амфибластула, а при частичном дискоидальном дроблении полителолецитальных яиц — дискобластула, в которой бластоцель имеет вид щели (птицы).
В формировании бластулы имеет значение не только количество желтка
и интенсивность дробления, но и типы сцепления
между бластомерами, размеры пространств между ними, явления взаимного скольжения, образование выростов типа псевдоподий и другие недостаточно изученные явления. На любой стадии развития зародыш представляет
собой интегрированное целое. Например, на стадии двух бластомеров при их
разъединении из каждого бластомера образуется полноценный организм (однояйцовые близнецы). В том случае, если убить один из бластомеров (проколом), но не отъединять его от другого, из оставшегося бластомера разовьется
только половина зародыша. Интеграция возрастает по мере увеличения количества бластомеров и у 8-кле-точного зародыша каждая клетка теряет способность развиваться в целый организм. У подавляющего большинства животных бластула без остановки переходит в следующую стадию развития—
гаструлу. Исключение составляют птицы, а из млекопитающих звери семейства куньих. У птиц зародыш замирает на стадии бластулы в промежутке от
снесения яйца до насиживания (или инкубации). Это время не должно превышать 2—3 недель, так как жизнеспособность зародыша падает. У куньих
задержка в развитии может длиться 2—3 мес и связана с регуляцией сроков
рождения щенят (весной) независимо от сроков оплодотворения.
ГАСТРУЛЯЦИЯ
Гаструляция — совокупность сложных процессов, приводящих к образованию гаструлы —зародыша, состоящего из нескольких пластов клеток.
Она связана с активным перемещением клеточного материала, в результате
чего зародыш из однослойного (бластулы) превращается в двухслойный.
Процессы гаструляции вот уже не один десяток лет находятся под пристальным вниманием эмбриологов, пытающихся разгадать причины активного передвижения и дифференциации клеток и целых клеточных пластов. Существует много теорий, объясняющих эти морфогенетические движения различиями в активности митозов в разных участках зародыша, в способности клеток менять форму, склеиваться друг с другом, осуществлять амебоидные,
скользящие, поступательные движения, в выделении клетками биологически
активных веществ, направленно действующих на их микроокружение, в изменении метаболизма клеток и др. Однако существующие ныне теории говорят лишь об отдельных моментах и не дают общую причинно-следственную
характеристику морфогенетических процессов периода гаструляции, причины которых во многом еще остаются неясными.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Различают четыре типа гаструляции (рис. 19): 1. Инвагинация — впячивание. Таким способом образуется гаструла из целобластулы. Клетки дна
впячиваются внутрь бластоцеля и достигают изнутри клеток крыши бластулы. Зародыш из округлого становится чаше- или мешкообразным, из однослойного—двухслойным. Слои называются зародышевыми листками. Наружный зародышевый листок получил название эктодерма, внутренний —
энтодерма. Мешковидная полость гаструлы является первичной кишкой —
гастроцель, а отверстие, при помощи которого она сообщается с внешней
средой, — первичный рот — бластопор. Этот тип гаструляции характерен
для ланцетника.
Рис. 19. Типы гаструляции:
А — инвагинация; Б — эпиболия; В — миграция; Г — деламинация.
У червей, моллюсков и членистоногих дефинитивный рот образуется на
базе бластопора, поэтому они называются первичноротые. У вторичноротых
(щетинкочелюстные, плеченогие, иглокожие, кишечнодышащие и хордовые)
рот возникает на брюшной стороне головного конца, а бластопор превращается в анальное отверстие или нервно-кишечный канал.
2. Иммиграция — вселение. В этом случае из бластодермы, особенно из
области дна бластулы, клетки выселяются в ее бластоцель, располагаясь под
наружным пластом. Наружный пласт клеток превращается в эктодерму, а
лежащий под ним — в энтодерму. Этот тип гаструляции свойствен кишечнополостным.
3. Деламинация — расслоение. Возникает в том случае, если происходит
синхронное деление бластомеров параллельно поверхности бластулы. При
этом один слой клеток окажется лежащим снаружи, а другой — внутри. Этот
тип гаструляции встречается у кишечнополостных (медузы).
4. Эпиболия — обрастание. Происходит в том случае, когда перегруженный желтком вегетативный полюс зиготы либо не участвует в дроблении
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(частичное дробление), либо при дроблении здесь образуются очень крупные
бластомеры, заполненные желтком и поэтому делящиеся медленно. Более
мелкие, быстро делящиеся клетки крыши бластулы их обрастают. Такой тип
гаструляции встречается у малощетинковых червей.
Тот или иной тип гаструляции редко встречается в животном мире в
чистом виде. Гораздо чаще наблюдается смешанная гаструляция, включающая в себя элементы двух, а то и трех перечисленных типов. В любом случае
гаструляция приводит к образованию одних и тех же зародышевых листков:
эктодермы, энтодермы и мезодермы (у губок и кишечнополостных ее нет).
Дифференцировка зародышевых листков. Вслед за образованием двух
зародышевых листков или параллельно с ними между эктодермой и энтодермой образуется третий зародышевый листок— мезодерма. При нормальном
развитии зародыша, к какому бы классу и типу он ни относился, листки,
взаимодействуя между собой, дифференцируются в строго определенном направлении: из каждого зародышевого листка образуются зачатки определенных гомологичных тканей — гистогенез и органов — органогенез. У
хордовых прежде других образуются так называемые осевые органы: нервная
трубка, хорда и кишечная трубка (см. цв. табл. I). При дальнейшем органогенезе из эктодермы развиваются эпителий кожи и его производные (роговые
образования, кожные железы), нервная система, передний и задний концы
кишечной трубки. Производные энтодермы — эпителиальная выстилка пищеварительного канала и его железы, дыхательная система, а мезодермы —
мышечная система, скелет, органы мочеполовой системы, стенки серозных
полостей тела и др. Мезодерма дает и начало мезенхиме — эмбриональной
соединительной ткани, которая входит в состав большинства органов.
В процессе органогенеза взаимосвязь и взаимопроникновение зародышевых листков настолько тесные, что в образовании практически каждого
органа принимают участие клеточные элементы двух, а то и всех трех зародышевых листков. Кроме того, в период активной дифференцировки зародышевого материала широко распространено явление индукции, когда одна
какая-либо закладка влияет на характер развития другой. Так, материал дорсальной губы бластопора индуцирует развитие нервной трубки, в результате
нервная трубка может возникнуть в любом другом месте, куда был перенесен
материал дорсальной губы.
В процессе эмбриогенеза признаки, характерные для животного, появляются постепенно. Так, в эмбрионе любого позвоночного животного сначала можно определить черты, характерные для типа хордовых, например, характер возникновения и расположения осевых органов. Затем становятся
видны черты, присущие классу, например характер кожного покрова, позже
— отряду, семейству, роду, виду, породе и, наконец, индивидууму.
Поэтому, прежде чем приступить к изучению развития столь сложно организованных животных, как млекопитающие, целесообразно ознакомиться с
более примитивно организованными хордовыми. Самым удобным и хорошо
изученным объектом является ланцетник.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Вопросы для самоконтроля. 1. Чем отличается дробление от обычного
деления клеток и как оно зависит от особенностей строения яйцеклетки? 2.
Что такое бластула и гаструла, какие типы бластул и гаструляции вы знаете?
3. Как протекает дифференцировка зародышевых листков?
Глава 6. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ЖИВОТНЫХ РАЗНЫХ
КЛАССОВ ТИПА ХОРДОВЫХ
РАЗВИТИЕ ЛАНЦЕТНИКА
Ланцетник — примитивное хордовое животное длиной 2— 5 см, обитает
в прибрежных водах. Его развитие подробно изучено А. О. Ковалевским. Яйцеклетка ланцетника диаметром 100—120 мкм, изоолиголецитальная, оплодотворение и развитие внешнее (цв. табл. I). Дробление полное, равномерное.
Первые десять делений происходят синхронно. Первая борозда, разделяющая
зиготу на два бластомера, меридианальная. Она проходит через три точки
(апикальный полюс, место проникновения спермия и вегетативный полюс) и
делит зародыш на правую и левую половины. Вторая борозда тоже меридианальная, но идет перпендикулярно к первой и делит зародыш на спинную
(дорсальную) и брюшную (вентральную) половины. Третья борозда широтная, проходит почти по экватору и делит зародыш на переднюю и заднюю
части, после чего он состоит из восьми бластомеров. Дробление — процесс
быстрый. В течение двух часов после оплодотворения образуется целобластула, которая состоит более чем из 1000 клеток.
Начинается гаструляция путем инвагинации. Дно бластулы уплощается,
а потом вдавливается внутрь. Через 11 ч от оплодотворения образуется двухслойная мешковидная гаструла с широким бластопором. Эктодерма ее состоит из уплощенных клеток, а энтодерма — из более высоких и крупных.
Затем зародыш несколько вытягивается в длину. При этом его дорсальная
сторона уплощается, а бластопор сужается. В нем различают дорсальную губу
— край бластопора, примыкающий к спинной стороне зародыша, противоположную ей — вентральную губу и боковые губы, расположенные между ними. В области дорсальной губы бластопора происходят особенно активные
процессы размножения и перемещения клеток. При этом бластопор становится все меньше, а нарастающий клеточный материал перемещается в головном направлении. Тяж эктодермальных клеток, лежащий на спинной стороне вдоль всего зародыша, превращается в нервную пластинку. Участки эктодермы, расположенные по бокам от нервной пластинки, приподнимаются,
образуя два продольных валика, а нервная пластинка углубляется в виде желоба. Продольные эктодермальные валики все более сближаются между собой, а желоб нервной пластинки углубляется. К тому времени, как края нервного желоба смыкаются и он превращается в нервную трубку, эктодермальные валики также срастаются друг с другом и нервная трубка оказывается
под эктодермой, которая теперь становится кожной эктодермой. Эктодерwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мальные валики преобразуются в ганглиозную пластинку, тесно связанную с
нервной трубкой. Из нее образуются спинномозговые ганглии.
Параллельно с этим из материала дорсальной энтодермы вычленяется,
начиная с головного конца, хордомезодермальный зачаток, который разделяется на хорду и мезодерму. Хорда (греч. chorde — струна) сворачивается в
тяж, идущий по всей длине под нервной трубкой ланцетника. Мезодерма лежит с двух сторон от хорды в виде карманов, обращенных выпуклой стенкой
к эктодерме, а полостью — к энтодерме. Некоторое время она сохраняет
связь с энтодермой, а затем полностью отшнуровывается. Края энтодермы
смыкаются и образуется кишечная трубка, стенка которой называется кишечной энтодермой.
Мезодерма распространяется, разрастаясь, между экто- и энтодермой в
виде двух мешковидных образований. При этом, начиная с головного конца,
она дифференцируется на сомиты, сегментные ножки и спланхнотом.
Сомиты—метамерные сегменты, расположенные по дорсальной стороне зародыша с двух сторон от нервной трубки и хорды. Наружные участки
сомитов, прилежащие к кожной эктодерме, разрыхляются, дают начало соединительнотканной основе кожи— дерме и поэтому называются дерматомы. Средние участки сомитов — миотомы самые развитые. Они дают начало
соматической мускулатуре. Внутренние участки сомитов — склеротомы
принимают вид мезенхимы, из которой впоследствии развивается скелет.
Полость сомитов становится щелевидной, а в дальнейшем зарастает.
Сегментные ножки, или нефротомы, — узкие трубочки из клеток, связывающие сомиты со спланхнотомом. Из них развиваются части мочеполовой системы.
Спланхнотом — вентральная часть мезодермы, сначала имеет вид метамерно сегментированных парных мешков. Затем сегментация исчезает, образуются два мешка, которые с дальнейшим ростом зародыша дорастают друг
до друга и сливаются на вентральной стороне зародыша. Полость спланхнотома становится единой. Это вторичная (серозная) полость тела, или целом.
Стенки целома — листки превращаются в серозные оболочки. Наружный
листок, прилежащий к стенке тела, называется париетальным (пристеночным), внутренний — висцеральным (внутренностным). Далее на переднем и
заднем концах кишки прорываются отверстия, зародыш ланцетника превращается в свободную личинку. Дальнейший органогенез ланцетника видоспецифичен и не представляет интереса для понимания развития млекопитающих.
РАЗВИТИЕ РЫБ И АМФИБИЙ
Развитие бесчерепных через ряд промежуточных стадий дало начало
классу рыб, у большинства которых, как и у ланцетника, свободный личиночный тип развития. Однако отдельные отряды и семейства отличаются по
содержанию желтка в яйцеклетке настолько, что это отражается на всем
дальнейшем эмбриогенезе— приводит к неличиночному типу развития.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У многих костистых рыб, некоторых костных и хрящевых ганоидов и
двоякодышащих кистеперых яйца мезолецитальные. Такие же яйца, а следовательно, и характер эмбриогенеза, у амфибий, произошедших от кистеперых. Поскольку эмбриогенез амфибий изучен несравнимо лучше, чем у рыб,
на его примере рассмотрим развитие животных, имеющих мезолецитальные
яйца.
Яйцеклетки и зигота этих животных мезо- и телолецитальные. Оплодотворение и развитие внешние. Дробление полное, неравномерное. В результате дробления развивается амфибластула. Гаструла образуется путем инвагинации и эпиболии. В районе впячивания — будущем хвостовом конце особи — возникает углубление— гомолог бластопора ланцетника. Активнее всего размножается и прежде всего подворачивается клеточный материал, соответствующий дорсальной губе бластопора, по бокам от него материал боковых губ. Материал дорсальной губы бластопора, распространяющийся снаружи в сторону головного конца зародыша, дифференцируется в нервную
пластинку, которая затем свертывается в трубку. На головном конце ее формируются мозговые пузыри. Материал, подворачивающийся в области боковых губ, превращается в мезодерму, а клетки нижней губы бластопора обрастают крупные инертные бластомеры дна бластулы, формируя вместе с ними
кишечную энтодерму. Дифференциация мезодермы аналогична тому, что
описано у ланцетника.
РАЗВИТИЕ ПТИЦ
У птиц, как и некоторых рыб, яйцеклетки полилецитальные, с неравномерным распределением желтка. Их яйца, кроме первичной и вторичной, покрыты еще и третичными оболочками (белковая, подскорлуповые пленки и
скорлупа). Это создает дополнительные преимущества при питании и защите
зародыша от внешних воздействий. Большое же количество желтка обеспечивает питанием зародыш в течение всего эмбриогенеза. Отпадает необходимость в личиночной стадии. Такой тип развития называется неличиночным, или прямым. Особенно это заметно у выводковых птиц, к которым относятся все бегающие, пингвины, а также все сельскохозяйственные птицы
(куриные, пусиные) и др. У них птенцы зрелорождаемы: к моменту вылупления хорошо развита мускулатура (способны бегать), оперение (покрыты пухом), самостоятельно находят пищу.
Оплодотворение у птиц внутреннее, развитие наружное. Дробление частичное, дискоидальное. В результате дробления образуется дисков ластула
(бластодиск). Дробление и начало гаструляции происходят в яйцеводе. Гаструла возникает путем деламинации, иммиграции и эпиболии.
Клетки эктодермы цилиндрические, плотно прилегают друг к другу в
несколько слоев. Клетки энтодермы плоские, лежат более рыхло. Энтодерма
образуется как путем подворачивания, так и выселения из клеток эктодермы.
В первые 12 ч инкубации очень активно идут процессы эпиболии—
бластодиск разрастается по желтку. Центральная его часть утолщается —
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
развивается зародышевый щиток (рис. 20). В клетках зародышевого щитка
идут активные процессы гаструляции. При этом желток, лежащий под зародышевым щитком, частично резорбируется, используется зародышем. Отделившиеся от желтка клетки кажутся более светлыми. Овальной формы центр
зародышевого щитка называется светлым полем, а его край, где клетки
вплотную прилегают к желтку, — темным полем. Зародыш развивается из
материала светлого поля, темное — является источником внезародышевых
образований.
Рис. 20. Щиток куриного зародыша
(стрелки указывают направление движения клеточного материала в период
гаструляции):
1 — первичная полоска; 2— бороздка; 3— первичный (гензеновский) узелок;
4— первичная ямка; 5 — внеза-родышевая
эктодерма; 6 — закладка нервной пластинки; 7— закладка хорды; 8 — закладка мезодермы.
В результате сложных перемещений клеточного материала зародышевого щитка по средней линии светлого поля на протяжении 3/5 его длины образуется утолщение — первичная полоска. Передний конец ее еще более утолщен и расширен. Это первичный (гензеновский) узелок. В области первичной
полоски и узелка начинается подворачивание клеточного материала внутрь.
При этом в первичном узелке развивается первичная ямка, а в первичной полоске — первичная бороздка. Область ямки соответствует дорсальной губе,
правая и левая сторона бороздки — боковым губам бластопора. В области
первичной ямки подворачивается материал хордального зачатка и головной
энтодермы. Подвернувшись, он движется в сторону будущего головного конца зародыша и называется головным отростком. Параллельно с продвижением головного отростка вперед над ним из дорсальной эктодермы формируется нервная пластинка. В области первичной бороздки подворачивается и
распространяется в стороны и вперед материал мезодермы. По мере перемещения клеточного материала головной отросток удлиняется, а первичная полоска укорачивается. Гензеновский узелок смещается в каудальном направлении, пока не окажется на месте будущего ануса. Мезодерма, сначала
имеющая вид сплошного тяжа, к концу первых суток насиживания делится
на сомиты, сегментные ножки и спланхнотом. Дифференцировка зародышевых листков начинается с головного конца — в области головного отростка и постепенно распространяется к хвостовому — к области гензеновского
узелка. В процессе роста зародыша происходит отделение зародышевых частей от внезародышевых с помощью туловищной складки, окружающей зародыш (см. цв. табл. III).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Параллельно с дифференцировкой зародышевых листков наблюдается
обрастание желтка внезародышевыми частями эктодермы, энтодермы и мезодермы — развивается желточный мешок— внезародышевый орган питания, дыхания и кроветворения эмбриона. Стенками его являются три зародышевых листка, а содержимым — желток. В процессе отделения зародышевых частей от внезародышевых туловищная складка стягивается под зародышем, в результате чего кишечная энтодерма превращается в кишечную
трубку, связанную с желточным мешком — желточным протоком, который
проходит в середине пупочного стебелька. Кроме желточного мешка у наземных животных (рептилий, птиц, млекопитающих), так называемых амниот, образуются и другие внезародышевые органы, прикрывающие зародыш и
поэтому называемые зародышевыми, или плодными, оболочками. У птиц —
это амнион, аллантоис и серозная оболочка. На второй день развития эмбрион слегка погружается в разжижившийся желток в области латебры. При
этом навстречу туловищной складке начинает вырастать амниотическая
складка, состоящая из эктодермы и париетального листка спланхнотома мезодермы. Она становится высокой, обрастает зародыш и на 3—4-е сутки инкубации смыкается над ним таким образом, что наружные склоны складки
срастаются между собой и образуют серозную оболочку, а внутренние склоны — амнион. Однако амниотическая складка срастается не по всей длине. В
заднем ее участке сохраняется отверстие, соединяющее амнион с серозной
оболочкой. Это — серозно-амниотический канал.
Серозная оболочка, разрастаясь под скорлупой, обрастает белок, который по серозно-амниотическому каналу поступает в амнион и там заглатывается зародышем. Оболочки обращены друг к другу внезародышевой мезодермой, между ее листками сохраняется внезародышевый целом. В него от
заднего участка кишки (от клоаки) начинает отрастать аллантоис (allantois —
колбаса), который, быстро распространяясь, к 10-м суткам занимает все пространство между амнионом и серозой. Так как аллантоис — это вырост кишки, его стенки состоят из энтодермы и висцерального листка спланхнотома
мезодермы. Мезодерма аллантоиса срастается с мезодермой серозной оболочкой, образуя серозоаллантоис.
Мезодерма провизорных органов дает начало сосудам, которые образуют внезародышевую сосудистую систему. Она связана с эмбрионом пупочными сосудами, проходящими в пупочном стебельке. По сосудам желточного
мешка поступают питательные вещества из желтка. Здесь же обнаруживаются первые очаги кроветворения и первичные половые клетки. В сосуды серозоаллантоиса поступает кислород воздуха, поскольку эта оболочка тесно
прилегает к скорлупе. В них же поступает резорбируемый из скорлупы кальций. Амнион - водная оболочка является средой, в которой развивается зародыш, предохраняет его от внешних воздействий. Его содержимое заглатывается зародышем, особенно во второй половине плодного периода (с 16—17
суток). Аллантоис— резервуар для некоторых продуктов обмена, в нем накапливается мочевая кислота. Срастаясь с амнионом и серозной оболочкой, алwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лантоис принимает участие в выполнении трофической и дыхательной функций.
В развитии птичьего зародыша имеется несколько критических периодов, которые приходятся на этапы смены характера питания и дыхания. Так,
на вторые сутки инкубации куриного зародыша происходит становление
внекишечного питания через сосуды желточного круга кровообращения; на
13-е сутки начинается внутрикишечное питание заглатываемым белком, на
18-е сутки плод переходит на внутрикишечное питание желтком, а с 4-го дня
после вылупления переходит на активное питание кормом. Меняется и тип
дыхания: в возрасте 7—18 суток эмбрион получает кислород через сосудистые сети серозоаллантоиса, с 18—19 суток — переход к легочному дыханию
кислородом воздушной камеры, в период вылупления с 20-го дня — наклев и
дыхание кислородом воздуха.
РАЗВИТИЕ МЛЕКОПИТАЮЩИХ
У плацентарных млекопитающих яйцеклетка, а следовательно, и зигота
олиголецитальная, как и у ланцетника. Однако на раннее эмбриональное развитие большой отпечаток наложило их филогенетическое развитие — происхождение от ранних рептилий, имеющих телолецитальные яйца. До сих пор
имеются этому достоверные подтверждения: в настоящее время на земле существуют три вида яйцекладущих млекопитающих. Так что утеря желтка яйцеклеткой млекопитающих — это вторичное явление, вызванное их внутриутробным развитием и питанием за счет организма матери. Сумчатые, считаясь как бы переходной ступенью от яйцекладущих к плацентарным, наглядно
демонстрируют начальные этапы становления внутриутробного развития. Беременность у них очень короткая, плацента не образуется, зародыш появляется на стадии предплода, и дальнейшее его развитие протекает в кожной
сумке.
У плацентарных внутриутробное развитие длительное. У крупного рогатого скота оно около 285—290 дней, у лошадей — 335— 338, у овец— 150, у
свиней — 114, у кролика — 30 дней. Плацента сложно устроена и функционирует с конца зародышевого, в течение предплодного и плодного периодов.
Детеныши рождаются гораздо более зрелыми, чем у сумчатых. Но у различных отрядов и семейств плацентарных млекопитающих степень зрелорождаемости различна. Это определяется особенностями экологии того или иного семейства и зависит от длительности внутриутробного вынашивания и
строения плаценты.
Дробление у плацентарных млекопитающих полное, неравномерное.
Бластомеры неодинаковой величины, делятся несинхронно. В результате зародыш может состоять из 3, 5, 9, 12 и т. д. бластомеров. При дроблении появляются мелкие светлые бластомеры и крупные темные. Светлые бластомеры располагаются сверху, темные — под ними. Образуется морула, которая,
однако, существует недолго, так как между светлыми и темными бластомерами возникает полость, заполненная белковой жидкостью—продуктом деяwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тельности светлых бластомеров. Развивается бластодермический пузырек —
стерробластула, или бластоциста. Стенку ее образуют мелкие бластомеры,
выполняющие трофическую функцию — трофобласт. Внутри в виде комочка клеток, прилежащего к трофобласту, находятся более крупные и темные
клетки — это зародышевый узелок, или эмбриобласт. Эмбриобласт является
источником, из которого развивается зародыш и часть внезародышевых органов. Трофобласт — внезародышевая часть бластоцисты, выполняющая
трофическую и защитную функции. Раннее разделение зародышевых и внезародышевых частей — одна из особенностей эмбриогенеза млекопитающих.
Дробление начинается в яйцеводе под прикрытием блестящей оболочки
и оболочки оплодотворения и продолжается в матке. У свиньи дробление
идет 3 дня, у овцы — 4, у коровы дробящаяся зигота движется по яйцеводу 4
дня и на стадии морулы, состоящей из 15—16 бластомеров, поступает в матку, где дробление продолжается еще 4 дня. За это время зародыш практически не увеличивается по сравнению с зиготой. К концу дробления — началу
гаструляции растворяются яйцевые оболочки (у свиньи на 8-й, у овцы на 10й, у коровы на 13-й день), трофобласт активно всасывает жидкость из полости матки и зародыш начинает бурно расти. Особенно значительно увеличивается трофобласт. Он у жвачных приобретает веретеновидную форму, у
свиньи и лошади— овальный. В то время как трофобласт достигает 20 см
длины, эмбриобласт имеет длину только 1 мм (Н. М. Решетникова и др.). Эмбриобласт распластывается под трофобластом в средней его части и становится двухслойным.
Гаструла образуется в основном деламинацией, а также иммиграцией
(как у птиц). В результате образуется зародышевый диск, эктодерма которого
примыкает к трофобласту, а энтодерма— к полости бластоцисты. Над зародышевым диском трофобласт рассасывается и эктодерма с ним смыкается по
периферии диска. По средней линии зародышевого диска в результате морфогенетических движений клеточного материала образуются первичная полоска с бороздкой и первичный узелок с ямкой. Подворачивание клеток и
дифференцировка мезодермы и осевых органов протекают так же, как и у
птиц. В области первичной ямки подворачивается и движется вперед материал прехордальной пластинки и хорды.
Мезодермальный материал подворачивается в области первичной бороздки и движется вперед и в сторону между экто- и энтодермой у свиньи с
9-го дня, у овцы — с 13-го, у коровы — с 15-го дня. Наряду с этим из зародышевых листков выселяются клетки, дающие начало мезенхиме — эмбриональной соединительной ткани. Дифференцировка зародышевых листков начинается с головного конца. На 14-й день у свиньи, на 15-й у овцы, на 19-й у
коровы наблюдается дифференцировка нервной пластинки из клеток дорсальной эктодермы в области головного отростка. Через 3—4 суток нервная
пластинка превращается в нервную трубку, а в головном конце закладываются мозговые пузыри, органы обоняния, зрения и слуха. Мезодерма дифференцируется на сомиты, сегментные ножки и спланхнотом. Энтодерма
дифференцируется на кишечную и желточную. В это время зародыш нахоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дится в одном из рогов матки. На его трофобласте образуются первичные
ворсинки, которыми он внедряется в слизистую оболочку матки и всасывает
секрет маточных желез — маточное молоко. Такое питание характерно для
млекопитающих до формирования плаценты. Вместе с дифференцировкой
зародышевых листков происходит развитие и провизорных (временных) зародышевых органов: желточного мешка, амниона, аллантоиса, хориона и плаценты.
При этом формируются туловищная и амниотическая складки. Энтодерма и висцеральный листок спланхнотома мезодермы обрастают полость бластоцисты и образуют желточный мешок, заполненный белковой жидкостью.
Эктодерма и париетальный листок спланхнотома мезодермы прикрывают зародыш сверху и превращаются в амнион и хорион. От заднего отдела кишки
отрастает аллантоис, стенка которого состоит из энтодермы и висцерального
листка спланхнотома мезодермы. Он врастает во-внезародышевый целом
между амнионом и хорионом.
Желточный мешок не играет сколько-нибудь значительной роли в питании зародышей жвачных и свиней. У лошади на короткое время образуется
желточная плацента, когда желточный мешок дорастает до хориона. Как ни
коротко существование желточного мешка (13—30 суток у овцы, 16—35 у
коровы, 11—23 суток у свиньи, до конца третьего месяца у лошади), он выполняет в это время чрезвычайно важные функции: в нем формируются сосудистые сети, он осуществляет кроветворение, питание, дыхание, является
местом образования первичных половых клеток, вырабатывает гормональные вещества. В конце зародышевого периода желточный мешок совершенно
редуцируется. Перечисленные функции переходят к другим органам.
Амнион — водная оболочка, окружающая эмбрион на протяжении почти всего внутриутробного развития, так как она образуется в начале зародышевого периода на стадии зародышевого диска (у овцы на 15-й, у коровы на
19-й, у свиньи на 13-й день), а прекращает существование в процессе родов.
Амнион выполняет защитную функцию, вырабатывая околоплодные воды,
являющиеся средой, в которой развивается зародыш и плод.
Аллантоис появляется у овцы на 16-е, у коров — на 20-е, у свиней — на
15-е сутки развития. У жвачных он имеет более узкую центральную часть и
два сильно развитых рога, у свиньи и лошади напоминает сплюснутый шар
вокруг задней части тела. В аллантоисе скапливаются жидкие продукты обмена, некоторые биологически активные вещества и другие метаболиты. Он
быстро растет, занимая пространство между амнионом и хорионом. Их мезодермальные слои при этом сливаются, образуются аллантоамнион и аллантохорион. В них развиваются сосудистые сети. Особенно мощные сосудистые поля образуются в аллантоисе со стороны хориона. Они врастают в первичные ворсинки трофобласта, отчего те становятся крупными, ветвистыми.
При этом усложняется строение стенки аллантохориона — образуется зародышевая (дочерняя, плодная) часть плаценты.
Хорион (кожа)—наружная оболочка, существующая практически на
всем протяжении внутриутробного развития плацентарных млекопитающих.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Выполняет защитную и трофическую функции. Еще в бластоцисте эту роль
выполнял трофобласт. После разрушения яйцовых оболочек на трофобласте
формируются первичные ворсинки, с помощью которых устанавливается
связь с железами слизистой оболочки матки — происходит внедрение зародыша и осуществляется его внесосудистое питание. Ко времени смыкания
амниотической складки внезародышевая мезодерма обрастает изнутри трофобласт и внедряется в его ворсинки— образуется хорион. С этого времени
питание осуществляется с помощью сосудистой сети хориона. В дальнейшем
хорион срастается с аллантоисом (аллантохорион), а у рогатого скота, кроме
того, на некотором расстоянии и с амнионом (амниохорион) и входит в состав плаценты в качестве ее зародышевой части. При этом ворсинки еще более разрастаются, стенка аллантохориона усложняется и начинает выполнять
функции, типичные для плаценты.
Плацента возникает на базе вышеописанных провизорных органов в
конце зародышевого периода и существует до конца беременности, обеспечивая ее нормальное течение. Это специфический внезародышевый орган,
присущий только плацентарным млекопитающим и имеющий для них огромное значение. Она выполняет разнообразные функции: трофическую,
экскреторную (для эмбриона), механический и иммунобиологической защиты, является эндокринным органом (вырабатывает хориальный гонадотропин, прогестерон, заменяет функцию желтого тела, соматомаммотропин стимулирует рост молочных желез и Др.).
Плацента состоит из двух частей: зародышевой (плодной) и материнской (маточной). Зародышевая часть образована аллантохорионом, а у
жвачных и амниохорионом. Материнская часть образована слизистой оболочкой стенки матки, где происходят специфические преобразования.
У разных семейств животных плаценты различаются по степени развития и характеру расположения ворсинок на хорионе, а также по глубине их
проникновения в ткани слизистой оболочки матки. По характеру расположения ворсинок различают плаценты: диффузную, котиледонную, кольцевидную, дискоидальную. У диффузной, или рассеянной, плаценты ворсинки
равномерно размещены по всему хориону в виде коротких небольших кустиков. Такая плацента характерна для кобылы, ослицы, верблюдицы, свиньи. У
котиледонной, или множественной, плаценты ворсинки крупные, сильноветвистые, собраны в крупные впячивания, названные котиледонами. Они
вступают в тесную связь с выпячиваниями стенки матки — карункулами. Такого типа плацента у жвачных. У кольцевидной, или зонарной, плаценты ворсинки располагаются в виде пояса, идущего вокруг аллантохориона. Такая
плацента у хищных. У человека, приматов и грызунов плацента дискоидальная. Ворсинки на ней занимают площадь в форме диска.
По характеру связи зародышевой и маточной частей различают эпителиохориальную, десмохориальную, эндотелиохори-альную и гемохориальную плаценты (рис. 21). В эпителиохори-альной плаценте эпителий хориона
контактирует с эпителием слизистой оболочки матки, не разрушая его. Ворсинки хориона проникают в полости маточных желез и всасывают секретиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
руемое железами маточное молоко. При родах ворсинки хориона отъединяются от эпителия матки без ее разрушения. Такая плацента у лошади, ослицы, свиньи. В десмохориальной плаценте ворсинки хориона внедряются в
собственную пластинку слизистой оболочки матки, разрушая ее эпителий.
Такая плацента характерна для жвачных. С развитием плаценты происходит
разрастание капилляров в участках слизистой, примыкающих к котиледонам:
усиливается гемотрофное питание плода. При родах слизистая матки в области карункулов оказывается лишенной эпителия, который быстро регенерирует (восстанавливается).
Рис. 21. Типы гистологического строения плацент:
А — эпителиохориальная; Б — десмохориальная; В —
эндотелиохориальная; Г — гемохориальная; I — зародышевая часть; II —
материнская часть; 1 — эпителий; 2 — соединительная ткань и 3 — эндотелий кровеносного сосуда ворсинки хориона; 4 — эпителий; 5 — соединительная ткань и 6 — кровеносные сосуды и лакуны слизистой оболочки
матки.
В эндотелиохориальной плаценте, присущей хищным, происходит деструкция не только эпителия, но и соединительной ткани слизистой оболочки
матки. В результате эпителий ворсинок контактирует с эндотелием капилляров матки. Этот способ плацентации обеспечивает более совершенное питание и снабжение кислородом эмбриона по сравнению с двумя вышеописанными. Гемохориальная плацента образуется у некоторых насекомоядных, рукокрылых, грызунов, обезьян и человека. В этом случае при плацентации
происходит разрешение всех слоев слизистой оболочки и ворсинки хориона
окунаются в кровь капилляров матки. Обеспечивается наиболее тесная связь
организма матери с плодом. При этом во время родов отмечают обильное
кровотечение, однако оно быстро прекращается в результате процессов регенерации.
При всех типах плацент сохраняется барьер между кровью плода и матери. Их сосудистые русла не сливаются, но всегда остаются разделенными
гематоплацентарным барьером, что имеет большое значение для защиты
плода от неблагоприятных условий, в которые попадает мать. Микробы, токсины, антитела, находящиеся в крови матери, до определенных пределов не
могут проникнуть через этот барьер.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Этапы внутриутробного развития млекопитающих. Зародышевый
период начинается дроблением и завершается плацентацией, гистогенезом и
началом органогенеза. У свиньи он длится 23, у овцы — 25—30, у коровы —
35 дней, у лошади — до конца третьего месяца развития. В предплодный период, который продолжается у свиньи с 23-го до 38-го дня, у овцы с 30-го до
45-го, у коровы с 35-го до 60-го дня, у кобылы до начала четвертого месяца
внутриутробного развития происходит активный органогенез, полное развитие плаценты (у жвачных количество котиледонов увеличивается до 100),
масса эмбриона возрастает в 20 раз.
В плодный период, который длится с 39-го до 114-го дня у свиньи, с 46го до 150-го — у овцы, с 61-го до 285-го у коровы, с 98-го до 295-го дня у кобылы, наблюдается совершенствование структуры и становление функции
органов, активный рост плода (к концу предплодного периода теленок весит
13 г, к концу плодного— 16—60 кг). Материал о развитии органов (органогенез) дан при описании соответствующих систем.
Влияние различных факторов на эмбриогенез. В течение эмбрионального развития животных существуют критические периоды, когда эмбрион наиболее чувствителен к внешним и внутренним воздействиям. Это
ранний зародышевый период (формирование бластоцисты и внедрение ее в
стенку матки), поздний зародышевый период — начало плацентации, вторая
половина плодного периода. Большое значение для развития плода имеет величина плаценты. При быстром ее развитии и больших размерах получаются
более крупные новорожденные, а при ее меньших размерах у копытных наблюдаются признаки недоношенности. Способность к вынашиванию плода
во многом зависит от гормонального фона в организме матери. При нарушении функции яичников, гипофиза, щитовидной и поджелудочной желез, а
также гормональной функции плаценты происходят самопроизвольные аборты, внутриутробная гибель плода или его отставание в развитии. Причиной
подобных явлений может быть и иммунологическая несовместимость матери
и плода, когда в организме овцы или коровы возникает повышенная чувствительность к белкам сыворотки крови плода.
На развитие плода влияет уровень кормления и характер рациона матери. Благотворно сказывается усиленное питание матерей, добавки с учетом
растущего плода, микроэлементов и витаминов.
Изучение закономерностей гаметогенеза, оплодотворения, эмбрионального развития особи крайне необходимо для практики сельскохозяйственного производства. Найдены способы длительного сохранения жизнеспособности спермиев, что дало возможность широко применять искусственное осеменение и тем самым в какой-то мере регулировать состояние генофонда.
Созданы банки генов ценных пород и редких видов животных. Отрабатываются и внедряются в практику методы пересадки яйцеклеток с их оплодотворением в пробирке, пересадки эмбрионов, что дает возможность увеличить во много раз потомство от ценных в хозяйственном отношении коров.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Вопросы для самоконтроля. 1. Каковы особенности развития хордовых
с олиго- и полилецитальными яйцами? 2. Что представляют собой внезародышевые части (провизорные органы) эмбриона млекопитающих? 3. Дифференцировка зародышевых листков и источники развития различных систем
органов. 4. Что такое плацента, какие типы плацент вы знаете?
Табл. VI. Сосудистая система крупного рогатого скота:
(принятые сокращения: а —артерия, в —вена; л. у. — лимфоузел); 1 — внутренняя челюстная а.; 2— околоушный л. у.; 3 — заглоточный л. у.; 4— наружная сонная а.; 5 —
парная яремная в.; 6 — позвоночная а.; 7— глубокая шейная а.; 8 — дорсальная лопаточная (поперечная шейная) а.; 9— реберношейный ствол; 10 — передняя межреберная;
11— краниальная полая в.; 12 — плечеголовная а.; 13 — плечеголовной ствол; 14 — левая
непарная в.; 15 — дуга аорты; 16 — пищеводно-бронхиальный ствол; 17 — легочные вв.;
18 — легочная а.; 19 — грудной лимфатический проток; 20 — каудальная полая в.; 21 —
грудная аорта; 22 — печеночные вв.; 23 — межреберные а. (дорсальные ветви); 24 —
брюшная аорта; 25 — чревная а.; 26 — краниальная брыжеечная а.; 27 — поясничная
цистерна; 23 — поясничные аа. (дорсальные ветви); 29 — почечная а.; 30 — яичниковая
а.; 31 — каудальная брыжеечная а.; 32 — наружная подвздошная; 33 — средняя крестцовая; 34 — внутренняя подвздошная аа.; 35 — глубокая бедренная а.; 36 — хвостовая а.;
37 — поверхностный паховый л. у.; 38 — надчревно-срамной ствол; 39 — бедренная а.:
40— выменная а.; 41 —каудальная надчревная а.; 42 — подколенный л. у.; 43 — краниальная а. вымени; 44 — а. цистерны; 45 — подкожная брюшная в.; 46 — воротная в.; 47 —
молочный колодец; 48 — внутренняя грудная в.; 49 — внутренняя грудная а.; 50 — левая
подключичная а.; 51 — подмышечный л. у.; 52 — подкожная в.; 53 — подмышечная а. и в.;
54 — правый лимфатический ствол; 55 — поверхностный шейный л. у.; 56 — правая общая сонная а.; 57 — наружная челюстная а.; 58 — подчелюстной л. у.; 59 — лицевая а.
Табл. VII. Нервная система крупного рогатого скота:
I— обонятельный и II—зрительный нн.; III — глазодвигательный н. вместе с парасимпатическими волокнами; IV — блоковый. V — тройничный и VI — отводящий нн.; VII — лицевой и IX — языкоглоточный нн. вместе с парасимпатическими волокнами; X — блуждающий, XI — добавочный, XII — подъязычный, нн.; 1 — нижнечелюстной и 2 — верхнечелюстной нн.; 3 — ядра среднего и продолговатого мозга; 4 — второй (аборальный)
слюноотделительный путь; 5 — спинномозговой ганглий; б —дорсальная и 7 — вентральная ветви спинномозгового н.; 8 — спинной мозг; 9 — шейное утолщение спинного мозга;
10 — плечевое сплетение; 11 — центры симпатической нервной системы; 12 — сердечное сплетение; 13 — пищеводные стволы блуждающего н.; 14 — симпатический позвоночный ганглий; 15 — серая соединительная ветвь; 16 — межреберный н.; 17 — пограничный симпатический ствол; 18 — большой внутренностный нерв; 19 — полулунный
ганглий; 20 — малые внутренностные ни.; 21 — подвздошно-подчревный. 22 — подвздошно-паховый и 23 — наружный семенной нн.; 24 — поясничное утолщение спинного мозга;
25 — пояснично-кожный н.; 26 — каудальный брыжеечный ганглий; 27 — крестцовый
центр парасимпатической системы; 28 — бедренный н.; 29 — подчревное сплетение; 30
— краниальный и 31 — каудальный ягодичные нн.: 32 — седалищный; 33 — каудальный
кожный бедра; 34 — срамной; 35 — большеберцовый и 36 — малоберцовый нн.; 37 — солнечное сплетение; 38 — локтевой; 39 — срединный; 40 — лучевой; 41 — подмышечный и
42 — краниальные грудные нн.; 43— звездчатый ганглий; 44 — возвратный н.: 45 —
средний шейный ганглий; 46 — н. вагосимпатикус; 47 — краниальный шейный ганглий; 48
— оральный слюноотделительный путь.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛ. I. РАЗВИТИЕ ЛАНЦЕТНИКА: А — ЗИГОТА; Б, В, Г. Д. - ПОСЛЕДОВАТЕЛЬНЫЕ СТАДИИ
ДРОВЛЕННЯ; Е - ЦЕЛОБЛАСТУЛА. Ж - РАННЯЯ ГАСТРУЛА; 3. И, К-ПОЗДНЯЯ ГАСТРУЛА; Л МОРУЛА. М - ОБРАЗОВАНИЕ ОСЕВЫХ ОРГАНОВ; 1 - АНИМАЛЬНЫЙ И 2- ВЕГЕТАТИВНЫЙ
ПЛЮСА; 3 СКОПЛЕНИЕ ЖЕЛТКА; ;4 - БЛАСТОМЕР; 5 - КРЫША И 6-ДНО БЛАСТУЛЫ; 7 БЛАСТОДЕРМА; 8 - БЛАСТОЦЕЛЬ, 9 - ЭКТОДЕРМА; 10 - ЭНТОДЕРМА; 11 - ГАСТРОЦЕЛЬ; 12 БЛАСТОПОР; 13 - ДОРСАЛЬНАЯ И 14 - ВЕНТРАЛЬНАЯ ГУБЫ БЛАСТОЛОРА; 15 - НЕРВНАЯ
ПЛАСТИНКА; 16 - ХОРДОМЕЗОДЕРМАЛЬНЫЙ ЗАЧАТОК; 17 — ЗАЧАТОК ХОРДЫ; 18 – ЗАЧАТОК
МЕЗОДЕРМЫ; 19 - КИШЕЧНАЯ ЭНТОДЕРМА; 20 - ХОРДА; 21 - МЕЗОДЕРМА; 22 - ЦЕЛОМ; 23 КИШКА; 24 - ВТОРИЧНАЯ КИШЕЧНАЯ ПОЛОСТЬ; 25-НЕРВНАЯ ТРУБКА; 26 МЕЗЕНХИМА; 27СОМИТЫ; 28 - СКЛЕРОТОМ; 29 -МИОТОМ; 30-ДЕРМАТОМ; 31 - СЕГМЕНТНЫЕ НОЖКИ; 32 СПЛАНХНОТОМ; 33 - ВИСЦЕРАЛЬНЫЙ И 34 — ПАРИЕТАЛЬНЫЙ ЛИСТКИ СПЛАНХНОТОМА
МЕЗОДЕРМЫ.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛ. III. РАЗВИТИЕ МЛЕКОПИТАЮЩИХ: А, Б, В - ПОСЛЕДОВАТЕЛЬНЫЕ СТАДИИ ДРОБЛЕНИЯ; Г
- МОРУЛА; Д - СТЕРРОБЛАСТУЛА (БЛАСТОЦИСТА), Е, Ж - ГАСТРУЛЯЦИЯ; (ЗАРОДЫШ СВИНЬИ); З
- ЗАРОДЫШЕВЫЙ ДИСК (ЛОШАДЬ); И - ОБРАЗОВАНИЕ ОСЕВЫХ ОРГАНОВ И
ДИФФЕРЕНЦИРОВКА МЕЗОДЕРМЫ; К - ФОРМИРОВАНИЕ ПЛОДНЫХ ОБОЛОЧЕК; Л. М. ПЛОДНЫЕ ОБОЛОЧКИ ТЕЛЕНКА, ЖЕРЕБЕНКА; 1 - БЛЕСТЯЩАЯ ОБОЛОЧКА; 2-БЛАСТОМЕРЫ (А –
СВЕТЛЫЕ, МЕЛКИЕ, Б - ТЕМНЫЕ, КРУПНЫЕ; 3-РЕДУКЦИОННОЕ ТЕЛЬЦЕ; 4 - ЭМБРИОБЛАСТ; 5 ТРОФОБЛАСТ; 6 - БЛАСТОЦЕЛЬ. 7-ЗАРОДЫШЕВЫЙ ДИСК; 8 - ЭКТОДЕРМА: 9 - ЭНТОДЕРМА; 10 ЗАРОДЫШЕВЫЙ ЩИТОК; 11-СВЕТЛОЕ ПОЛЕ;
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
12-ТЕМНОЕ ПОЛЕ; 13-ПЕРВИЧНЫЙ УЗЕЛОК; 14-ПЕРВИЧНАЯ ЯМКА; 15 - ПЕРВИЧНАЯ ПОЛОСКА;
16 - ПЕРВИЧНАЯ БОРОЗДКА; 17-НЕРВНАЯ ТРУБКА; 18 - ХОРДА: 19 - КИШКА; 20 -СОМИТ; 21 СЕГМЕНТНАЯ НОЖКА; 22 - ПАРИЕТАЛЬНЫЙ И 23 - ВИСЦЕРАЛЬНЫЙ ЛИСТКИ СПЛАНХНОТОМА;
24 - ЦЕЛОМ; 25 - ЗАРОДЫШ; 24 - АМНИОН; 77 - ХОРИОН; 28 - АМНИОТИЧЕСКАЯ СКЛАДКА; 29 ТУЛОВИЩНАЯ СКЛАДКА; 30 - АЛЛАНТОИС; 31 - ЖЕЛТОЧНЫЙ МЕШОК; 32 - ЖЕЛТОЧНЫЙ
ПРОТОК; 33 - ПУПОЧНЫЙ КАНАТИК: 34 - АЛЛАНТОАМНИОН: 35 - АЛЛАНТОХОРИОН; 36 АМНИОХОРИОН; 37- КОТИЛЕДОН
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛИЦА III. РАЗВИТИЕ ПТИЦ
А, Б - ДРОБЛЕНИЕ; 3 - ДИСКОБЛАСТУЛА; Г. Д – ГАСТРУЛА; Е — ОБРАЗОВАНИЕ ТУЛОВИЩНОЙ
СКЛАДКИ; Ж - ОБРАЗОВАНИЕ ПЛОДНЫХ ОБОЛОЧЕК; З – ВЗАИМООТНОШЕНИЕ ПЛОДА С
ПРОВИЗОНЫМИ ОРГАНАМИ; 1 ЗАРОДЫШЕВЫЙ ДИСК; 2 – ЖЕЛТОК; 3- БЛАСТОДЕРМА; 4 ПЕРИБЛАСТ; 5 -БЛАСТОЦЕЛЬ; 6 -ЭКТОДЕРМА: 7 - ЭНТОДЕРМА; 8
- МЕЗОДЕРМА; 9 ГЕНЗЕНОВСКИЙ УЗЕЛОК; 10-ПЕРВИЧНАЯ ПОЛОСКА: 11-ПЕРВИЧНАЯ БОРОЗДКА 12 - ПАРИЕТАЛЬНЫЙ И 13 - ВИСЦЕРАЛЬНЫЙ ЛИСТКИ СПЛАНХНОТОМА; 14 - ЦЕЛОМ; 15 - НЕРВНАЯ
ТРУБКА; 16 - ХОРДА; 17 - ТУЛОВИЩНАЯ И 18 – ЛМНИОТИЧЕСКАЯ СКЛАДКИ; I9 - КИШЕЧНАЯ
ЭНТОДЕРМА; 20-ЖЕЛТОЧНАЯ ЭНТОДЕРМА; 21-КИШКА; 22 —СЕРОЗНАЯ ОБОЛОЧКА; 23 АМНИОН; 24 - ПУПОЧНЫЙ КАНАТИК; 25 - АЛЛАНТОИС: 26 -СЕРОЗОАМНИОТИЧЕЕСКИЙ КАНАЛ;
27 - СЕРОЗОАЛЛАНТОИС; 28 - ТЕЛО ЗАРОДЫША; 29-ЖЕЛТОЧНЫЙ МЕШОК; :30 - БЕЛОК
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛИЦА IV. СРАВНИТЕЛЬНАЯ ТАБЛИЦА КЛЕТОК КРОВИ У НЕКОТОРЫХ ДОМАШНИХ
ЖИВОТНЫХ
ТАБЛ. VI. СОСУДИСТАЯ СИСТЕМА КРУПНОГО РОГАТОГО СКОТА.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛ. V. КРАСНЫЙ КОСТНЫЙ МОЗГ:
1 - ЛИМФОЦИТОПОДОБНАЯ СТВОЛОВАЯ КЛЕТКА; 2 - ГЕМОЦИТОБЛАСТ; 3 - БАЗОФИЛЬНЫЙ. 4 ПОЛИХРОМАТОФИЛЬНЫЙ И 5 – ЭОЗИНОФИЛЬНЫЙ ЭРИТРОБЛАСТЫ, 6 - НОРМОБЛАСТ; 7 ЭРИТРОЦИТ; 8-МЕТАМИЕЛОЦИТ; 9-МИЕЛОЦИТ; 10 - СЕГМЕНТОЯДЕРНЫЕ НЕЙТРФИЛЫ; 11 ЭОЗИНОФИЛ И 12-БАЗОФИЛ; 13 - МЕГАКАРИОЦИТ; 14 - ЖИРОВАЯ КЛЕТКА; 15 - РЕТИКУЛЯРНАЯ
КЛЕТКА; 16-СИНУСОИДНЫЙ КАПИЛЛЯР.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ТАБЛ. VII. НЕРВНАЯ СИСТЕМА КРУПНОГО РОГАТОГО СКОТА
Раздел третий. ОБЩАЯ ГИСТОЛОГИЯ - УЧЕНИЕ О ТКАНЯХ
Эволюционное развитие организмов шло путем дивергенции— расхождения признаков, в результате чего в настоящее время существует огромное
разнообразие видов. Параллельно этому шло и развитие тканей. Теорию дивергенции в развитии тканей обосновал и развивал Н. Г. Хлопин. Однако в
эволюции наблюдается поразительное сходство тканей, выполняющих сходные функции у неродственных животных форм, далеко отстоящих друг от
друга. Это дало основание А. А. Заварзину обосновать теорию параллельных
рядов в развитии тканей. Изучая развитие зародыша, можно видеть, как постепенно происходит усложнение его строения, как сравнительно однородный клеточный материал в результате клеточных перемещений, репрессии и
дерепрессии генов, изменений во внутриклеточном обмене веществ, детерминации и дифференцировки превращается в разнородные зачатки, из которых формируются ткани и органы. Процесс образования тканей называется
гистогенезом (histos—1 ткань, genesis — происхождение). В процессе его
клетки размножаются, растут, приобретают специализацию. Ткани развиваются не изолированно, а во взаимозависимости друг от друга. Сформировавшиеся ткани не являются стабильными — они подвергаются постоянным изменениям на протяжении жизни особи в связи с меняющимися условиями
внутренней и внешней среды.
Ткань — это исторически (филогенетически) сложившаяся система гистологических элементов (клеток и межклеточного вещества), объединенных
на основе сходства морфологических признаков, выполняемых функций и
источников развития.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Ткани обладают множеством признаков, по которым их можно отличить
одну от другой. Это могут быть особенности структуры, функции, происхождения, характера обновления, дифференцировки, пластичности и т. д. Существуют различные классификации тканей, но наиболее распространенной
считается классификация, в основу которой положены морфофункциональные признаки, как дающие наиболее общую и существенную характеристику
тканей. Подавляющему большинству организмов присущи такие функции,
как барьерная, обеспечение гомеостаза (постоянства внутренней среды), двигательная, интегративная (восприятие, передача и анализ раздражений). В
соответствии с этим различают четыре типа тканей: покровные (эпителиальные), внутренней среды (опорно-трофические или соединительные), мышечные и нервная.
Глава 7. ЭПИТЕЛИАЛЬНЫЕ ТКАНИ (ЭПИТЕЛИИ). СЕКРЕЦИЯ.
СТРОЕНИЕ ЖЕЛЕЗ
Эпителии — сборная группа тканей, широко распространенная в организме. Они имеют разное происхождение (развиваются из эктодермы, энтодермы, мезодермы) и выполняют разнообразные функции (защитную, трофическую, секреторную, выделительную и др.). Филогенетически эпителии
— один из наиболее древних типов тканей, первичной функцией которого
является пограничная — отграничение организма от окружающей среды.
В эволюции в связи с усложнением строения организмов дивергентное
развитие эпителиев привело к их многообразию, но, несмотря на это, все
эпителиальные ткани сохранили ряд общих морфофункциональных свойств,
по которым их и объединяют в единый тканевый тип.
1. Все виды эпителиальных тканей состоят только из клеток— эпителиоцитов. Между клетками имеются очень тонкие (шириной 10—50 нм)
межмембранные щели, в которых, однако, нет межклеточного вещества. В
них располагается гликокаликс (надмембранный комплекс), сюда поступают
вещества, проникающие в клетки и выделяемые ими.
2. Клетки всех эпителиев расположены плотно друг к другу, образуя
пласты. Эпителиальные ткани могут функционировать только в виде пластов.
Клетки в пласте соединены между собой различного вида контактами, которые упрочивают связь между ними (контакты по типу замка, десмосомы),
облегчают межклеточный обмен (щелевые контакты), отграничивают внутреннюю среду организма от внешней (плотные контакты и зоны слипания).
3. Эпителии подстилаются базальной мембраной (пластинкой), отделяющей их от подлежащей соединительной ткани. Ба-зальная мембрана (пластинка)—это слой межклеточного вещества толщиной 100 нм— 1 мкм, образованный деятельностью как эпителия, так и соединительной ткани. Электронная микроскопия показала ее сложную структуру, а гистохимия выявила
белково-углеводную природу. Базальная мембрана препятствует врастанию
эпителия, так как кровеносные сосуды не заходят внутрь эпителиального
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
пласта и питательные вещества проникают диффузно через подлежащую соединительную ткань и базальную мембрану.
4. Эпителии обладают морфофункциональной полярностью, направление которой совпадает с током веществ в клетке. Она выражается в изменении структуры и свойств эпителиоцитов, лежащих в разных слоях пласта, а
также в том, что апикальная и базальная части клеток отличаются друг от
друга в структурном и функциональном отношении. У нормально функционирующего эпителиоцита ядро обычно сдвинуто к базальному полюсу, под
ним располагается цитоплазматическая сеть, над ним—пластинчатый комплекс. На апикальном полюсе у многих видов эпителиев имеются микроворсинки, реснички, на базальном полюсе — вдавления плазмолеммы. Морфологической полярности соответствует функциональная полярность: в разных
участках клетки совершаются различные процессы, например, всасывание,
движение ресничек, экструзия связаны с апикальным полюсом, синтез веществ— у базального полюса. У разных видов эпителиальных тканей полярность выражена в неодинаковой степени.
5. У эпителиев хорошо выражена способность к регенерации, в их составе имеются стволовые, камбиальные и дифференцированные клетки.
Классификация эпителиев. Существует несколько классификаций эпителиальных тканей, в основу которых положен тот или иной их признак. По
фило-онтогенетической классификации (предложена Н. Г. Хлопиным и развита В. П. Михайловым и Е. А. Шубниковой) эпителии делятся в зависимости от их происхождения из определенной эмбриональной закладки. Функциональная классификация делит эпителии на ткани в зависимости от выполняемой функции. Эта классификация дает достаточно полную характеристику эпителиев, но получается слишком дробной. В ней выделяются эпителии покровный, всасывающий, выделительный, секреторный, сенсорный,
реснитчатый и др., каждый из которых, в свою очередь, можно разделить на
ряд эпителиев.
Морфологическая классификация делит эпителии на отдельные ткани в
зависимости от формы эпителиоцитов и количества их слоев в пласте. Различают однослойные и многослойные эпителии (рис. 22). Однослойные эпителии образуют пласт толщиной в одну клетку. Если все клетки в пласте эпителия одинаковой высоты, говорят об однослойном однорядном эпителии. В зависимости от высоты эпителиоцитов однорядный эпителий бывает плоский,
кубический и цилиндрический (призматический, столбчатый). Если клетки в
пласте однослойного эпителия разной высоты, говорят о многорядном эпителии. Все без исключения эпителиоциты любого однослойного эпителия расположены на базальной мембране.
Многослойные эпителии образованы несколькими слоями клеток, лежащими друг на друге, так что с базальной мембраной контактирует лишь
самый глубокий, базальный слой эпителиоцитов. В нем, как правило, залегают стволовые и камбиальные клетки. В процессе дифференцировки клетки
перемещаются в наружную сторону. В зависимости от формы клеток наружwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ного (поверхностного) слоя различают многослойный плоский, цилиндрический и переходный эпителии.
Наиболее полную характеристику дает морфофункциональная классификация эпителиев, так как она учитывает их структурные и функциональные особенности. В рамках этой классификации эпителии делят на покровный и выстилающий, а также эпителий паренхиматозных органов, в том числе железистый.
Рис. 22. Виды эпителиев:
I — однослойные эпителии (А— плоский, Б — кубический, В — призматический, Г — многорядный мерцательный); II — многослойные эпителии (Д
— плоский неороговевающий, Е — плоский ороговевающий, Ж —переходный
в свободном органе); 1 — базальная мембрана; 2 — соединительная ткань; 3
— бокаловидная клетка; 4 — реснитчатая клетка; 5 — вставочные клетки;
6 — базальный слой; 7 — шиповатый слой; 8 — слой плоских клеток; 9 —
зернистый слой; 10 — блестящий слой; 11 —роговой слой; 12 — промежуточный слой; 13 — покровный слой.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Покровный и выстилающий эпителии
Покровный эпителий входит в состав покровов тела в виде эпидермиса и
его производных (чешуи, перья, волосы, рога, копыта и др.), а выстилающий
— в состав слизистых и серозных оболочек, выстилая изнутри трубкообразные органы и серозные полости. Основная функция этих эпителиев пограничная. Большинство их располагается на границе между внутренней и
внешней средой, что во многом определяет характер их строения и функционирования. Данные сравнительной гистологии указывают на филогенетическое родство этих эпителиев, поэтому их обычно объединяют в группу покровных эпителиев.
Однослойный плоский эпителий (рис. 22-А). Выстилает респираторные
отделы легких, мелкие протоки желез, сеть семенника, полость среднего уха,
серозные оболочки. В последнем случае он известен как мезотелий, что указывает на его происхождение из мезодермы (из обоих листков спланхнотома). Однослойный плоский эпителий состоит из клеток, высота которых
меньше их ширины, ядра уплощенные. В том месте, где расположено ядро,
клетка несколько выше, чем в остальных участках. Морфо-физиологическая
полярность у этого вида эпителия выражена в меньшей степени, чем у других
видов, особенно у мезотелия, который, погрузившись во внутреннюю среду
организма, ее потерял. При нарушении пласта в мезотелий образуются гигантские многоядерные клетки. Функции мезотелия: разграничительная, так
как, покрывая серозные оболочки, он препятствует сращиванию органов друг
с другом и со стенками тела; способствует образованию серозной жидкости;
благодаря активной пиноцитозной деятельности его клеток происходит быстрый перенос веществ из серозной жидкости в лимфатическое русло.
Однослойный кубический эпителий (рис. 22-Б). Имеет различное происхождение. Выстилает протоки желез, канальцы почек. Все клетки лежат на
базальной мембране. Высота их приблизительно равна ширине. Функции
этого эпителия тесно связаны с функциями органа, в котором он находится.
Это может быть разграничительная (в протоках), осморегулирующая (в почках и солевых железах) и другие функции.
Однослойный призматический (цилиндрический) эпителий (рис. 22-Б).
Имеет различное происхождение в зависимости от местонахождения в организме. Выстилает желудочно-кишечный тракт, протоки желез, собирательные трубочки почек. Все его клетки лежат на базальной мембране и обладают морфофизиологической полярностью. Высота их значительно больше
ширины. Кроме разграничительной, этот эпителий выполняет специфические
функции, присущие тому или иному органу. Так, например, призматический
эпителий слизистой оболочки желудка является железистым — вырабатывает слизь, и поэтому называется призматическим слизистым эпителием.
Призматический эпителий слизистой оболочки кишечника называется каемчатым, так как несет на апикальном полюсе микроворсинки — щеточную каемку, благодаря чему осуществляется пристеночное пищеварение и всасывание питательных веществ.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Однослойный многорядный мерцательный эпителий (рис. 22-Г).
Имеет сложное происхождение. Выстилает воздухоносные пути и некоторые
участки половой системы (семявыносящие пути, яйцеводы). Состоит из трех
видов клеток: коротких реснитчатых, длинных вставочных (базальных) и
бокаловидных. Все клетки эпителиального пласта лежат на базальной мембране, но вставочные клетки не доходят до верхнего края пласта. Это стволовые и камбиальные элементы эпителия, которые в процессе роста дифференцируются и становятся реснитчатыми и бокаловидными. Реснитчатые клетки
несут на апикальном полюсе большое количество (до 270) ресничек — органелл движения. Бокаловидные клетки вырабатывают слизь (см. железистый
эпителий). Слизь покрывает пласт мерцательного эпителия, не только защищая его от внешнего воздействия, но и способствуя передвижению приставших частиц в воздухоносных путях или половых продуктов в половых путях.
Следовательно, мерцательный эпителий, кроме разграничительной, выполняет еще транспортную и защитную функции.
Многослойный плоский неороговевающий (слабоороговевающий)
эпителий (рис. 22-Д). Происходит из эктодермы и покрывает роговицу глаза,
а у некоторых животных, кроме того, ротовую полость, пищевод, преджелудок. В нем различают три слоя: базальный, шиповатый и плоский. Базальный
слой лежит на базальной мембране, образован призматическими клетками с
крупными овальными ядрами, несколько сдвинутыми к апикальному полюсу.
Клетки базального слоя делятся и, выдвигаясь вверх, теряют связь с базальной мембраной, дифференцируются и входят в состав шиповатого слоя. Шиповатый слой образован несколькими слоями клеток неправильной многоугольной формы с овальными или округлыми ядрами и с небольшими отростками в форме пластинок и шипов, которые проникают между клетками,
удерживая их наряду с десмосомами друг возле друга. Из шиповатого слоя
клетки перемещаются в поверхностный — плоский слой толщиной в 2—3
клетки. При этом они становятся плоскими, как бы распластываясь по подлежащим клеткам, в результате чего каждая плоская клетка занимает площадь, равную апикальной поверхности нескольких базальных клеток. Ядра
плоских эпителиоцитов также уплощаются, становятся гиперхромными. Связи между клетками ослабевают. Заканчивая свой жизненный цикл, они отмирают и отпадают с поверхности эпителия. У сельскохозяйственных животных, особенно у жвачных, поверхностные клетки этого эпителия ороговевают (кроме роговицы).
Многослойный плоский ороговевающий (сквамозный) эпителий
(рис. 22-Е). Происходит из эктодермы и образует эпидермис кожи, покрывает
ротовую полость и конечный участок прямой кишки. В нем различают пять
слоев: базальный, шиповатый, зернистый, блестящий и роговой. Базальный
слой, как и у неороговевающего эпителия, состоит из одного ряда призматических клеток, связанных десмосомами как между собой, так и с базальной
мембраной. В цитоплазме клеток много свободных рибосом, видны тонофиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ламенты. Шиповатый слой имеет толщину в 4—8 шиповатых клеток. В этих
клетках увеличивается количество тонофиламентов, которые объединяются в
пучки — тонофибриллы, видимые в световой микроскоп. Шиповатые клетки
еще сохраняют способность к размножению, из-за чего иногда базальный и
шиповатый слои объединяют под одним названием — ростковый слой. Зернистый слой толщиной в 2—3 клетки. Эпителиоциты этого слоя уплощенной
формы с плотными ядрами и резко базофильными зернами кератогиалина,
которые сливаются с тонофибриллами. Зернистый слой переходит в блестящий, состоящий из 1—2 рядов погибающих клеток. При этом кератогиалиновые зерна сливаются, органеллы деградируют, ядра распадаются, кератогиалин превращается в элеидин, окрашивающийся оксифильно и сильно преломляющий свет, что дало название слою. Самый поверхностный роговой
слой образован многими рядами (до 100) плоских мертвых клеток — роговыми чешуйками, заполненными роговым веществом — кератином. Десмосомы, связывающие клетки, видоизменяются, между клетками скапливаются
нейтральные жиры. На коже, покрытой волосами, роговой слой тонкий — из
нескольких рядов роговых чешуек.
Функция этого эпителия — пограничная, защита глубжележащих тканей
от внешних воздействий: химических, термических, механических, патогенных и др., что определяет характер дифференцировки эпителиоцитов. Специализация клетки выражается в ее ороговении и превращении в роговую
чешуйку, которая по площади равна апикальной поверхности 9—10 базальных клеток, содержит большое количество химически устойчивых белков и
липидов и обладает плохой теплопроводностью.
Переходный эпителий (рис. 22-Ж). Происходит из мезодермы. Выстилает лоханки почек, мочеточники, мочевой пузырь — органы, подверженные
значительному растяжению при наполнении мочой. Состоит из трех слоев:
базального, промежуточного и покровного. Клетки базального слоя мелкие,
разной формы, являются камбиальными, лежат на базальной мембране. Промежуточный слой состоит из светлых крупных клеток, количество рядов которых сильно колеблется в зависимости от степени наполнения органа. В
свободном от мочи органе они имеют булавовидную форму и располагаются
друг над другом, в наполненном — растягиваются и проникают в межклеточные промежутки. Клетки покровного слоя очень крупные, многоядерные
или полиплоидные, часто выделяют слизь, предохраняющую поверхность
эпителиального пласта от действия мочи.
Эпителий паренхиматозных органов
Эпителий, входящий в состав таких органов, как легкие, почки, печень,
поджелудочная и слюнные железы, семенники, яичники, гипофиз, щитовидная железа, надпочечники, тимус, очень разнообразен и будет рассмотрен
при изучении соответствующих органов. Здесь мы остановимся на общих
принципах строения и функционирования железистого эпителия — широко
распространенного вида эпителиальной ткани.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Железистый эпителий. Это специализированный эпителий, клетки которого вырабатывают и выделяют вещества различной природы, названные
секретами. Железистый эпителий обладает всеми свойствами эпителиальных
тканей, несмотря на то, что часто не контактирует с внешней средой. Это
объясняется особенностями выполняемой им секреторной функции. По своим размерам, форме, структуре железистые клетки очень разнообразны, как и
вырабатываемые ими секреты. Все же для многих железистых клеток характерны крупные размеры, большая поверхность ядер, крупные ядрышки, высокое содержание РНК и белка в цитоплазме, сильное развитие структур,
принимающих участие в процессе секретообразования и наличие на определенном этапе функционального цикла, гранул, зерен, вакуолей секрета или
его предшественников. Продукты, вырабатываемые железистыми клетками,
также весьма разнообразны по химической природе, физическим свойствам,
по количеству и расположению в клетке.
Рис. 23. Схема типов секреции:
1 — мерокринный; 2 — апокринный; 3 — голокринный.
Процесс секретообразования происходит в несколько фаз и называется
секреторным циклом.
Первая фаза — накопление клеткой исходных продуктов. Через базальный полюс в клетку поступают различные вещества органической и неорганической природы, которые используются в процессе синтеза секрета. Вторая фаза — синтез секрета из поступивших продуктов в цитоплазматической
сети. Синтез белковых секретов происходит в гранулярной, небелковых — в
аграну-лярной ее разновидности. Третья фаза — оформление секрета в гранулы и накопление их в цитоплазме клетки. По цистернам цитоплазматической сети синтезированный продукт поступает в место расположения аппарата Гольджи (пластинчатого комплекса), где происходит его конденсация и
упаковка в виде гранул, зерен, вакуолей. После этого вакуоль с порцией секрета отшнуровывается от пластинчатого комплекса и перемещается к апикальному полюсу клетки. Четвертая фаза — выведение секрета— экструзия
протекает по-разному, в связи с чем различают мерокринный, апокринный и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
голокринный тип секреции (рис. 23). При мерокринном типе секрет выводится без нарушения целостности цитолеммы. Секреторная вакуоль подходит к апикальному полюсу клетки, сливается с ней своей мембраной, образуется пора, через которую содержимое вакуоли изливается за пределы клетки.
При апокринном типе происходит частичное разрушение железистой клетки.
Отмечают макроапокринную секрецию, когда вместе с секреторной гранулой
отторгается апикальная часть цитоплазмы клетки, и микроапокринную секрецию, когда отторгаются верхушки микроворсинок. При голокринном типе
секреции наблюдается полное разрушение железистой клетки и превращение
ее в секрет. Пятая фаза — восстановление исходного состояния железистой
клетки.
Фазы секреторного цикла могут проходить последовательно одна за
другой, а могут совершаться одновременно в разных участках клетки. Это
зависит как от особенностей функционирования клеток, так и от силы стимуляции их деятельности.
Железистые клетки входят в состав некоторых видов покровных и выстилающих эпителиев, а также образуют специализированные органы — железы.
Железы. Это органы, основной функцией которых является секреторная.
В зависимости от того, куда выводится секрет, различают экзокринные и эндокринные железы. Экзокринные железы имеют протоки, по которым секрет
изливается или на поверхность тела, или в полость каких-либо трубкообразных органов. У эндокринных желез протоков нет, и продукты их деятельности— инкреты — выделяются во внутреннюю среду организма — в кровь
или лимфу.
По количеству клеток, образующих железы, последние бывают одноклеточные и многоклеточные. Типичная одноклеточная железа в организме
позвоночного животного — бокаловидная клетка. Данный тип клеток имеется в эпителии кишечника, воздухоносных и половых путей. Они выделяют
слизистый секрет. По форме клетка напоминает бокал. У нее узкая базальная
часть, в которой расположено ядро, цистерны цитоплазматической сети, митохондрии и другие органеллы. Наиболее развитой органеллой считается
комплекс Гольджи, расположенный над ядром. В нем синтезируются и накапливаются мукополисахариды, составляющие основную часть секрета. Его
цистерны постепенно переходят в секреторные вакуоли. По мере накопления
вакуоли занимают всю среднюю и апикальную части клетки. Секреторный
цикл в бокаловидной клетке совершается за 20—30 мин. Секрет выделяется
по мерокринному типу.
По расположению различают эндоэпителиальные и экзоэпителиальные
железы. Одноклеточные железы позвоночных всегда эндоэпителиальные —
залегают в пласте эпителия. Многоклеточные железы обычно экзоэпителиальные — лежат за пределами эпителиального пласта.
Многоклеточные экзокринные железы состоят из концевых отделов, образованных железистыми клетками, и выводных протоков, по которым выводится синтезированный секрет. По форме концевых отделов железы бывают
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
трубчатые, альвеолярные (пузырчатые) и трубчато-альвеолярные (рис. 24).
Если выводной проток оканчивается одним каким-либо концевым отделом,
говорят о простой неразветвленной альвеолярной или трубчатой железе. Если в проток открывается несколько концевых отделов, такую железу называют простой разветвленной. Когда разветвляется выводной проток, образуется
сложная железа. Различают сложные альвеолярные, трубчатые и трубчатоальвеолярные железы. По характеру выделяемого секрета железы делят на
серозные — вырабатывающие белковый секрет, слизистые и смешанные —
белково-слизистые.
Рис. 24. Схема строения желез:
I — простые железы; II — простые железы с разветвленным концевым
отделом; III — сложные железы; а — трубчатая железа; б — альвеолярная
железа; в — альвеолярно-трубчатая железа; IV—одноклеточная железа—
бокаловидная клетка; 1 — микроворсинки; 2 — гранулы секрета; 3— комплекс Гольджи; 4— митохондрии; 5 — ядро; 6 — эндоплазматическая сеть.
Влияние факторов. Паренхима желез однотипно реагирует на воздействие различных факторов. При действии экстремальных перегрузок, токсических или инфекционных поражений, механических повреждений и денерваwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ции в железистых клетках замедляется процесс образования и созревания
секрета, нарушается ритм секреторной деятельности. Наблюдается набухание
митохондрий (укорочение или исчезновение крист и просветление матрикса),
вплоть до разрушения, фрагментация и дегрануляция цитоплазматической
сети, разрушение цистерн комплекса Гольджи и перинуклеарного пространства, появление незрелых секреторных гранул и миелиноподобных структур.
После прекращения действия фактора, если он не был губительным, через
какое-то время происходит восстановление структур желез.
Вопросы для самоконтроля. 1. Что такое ткань? 2. Каковы основные
признаки эпителиальных тканей? 3. Дайте характеристику различных видов
покровного, выстилающего и железистого эпителиев? 4. Как происходит
процесс секретообразования? 5. Дайте классификацию и характеристику желез.
Глава 8. ТКАНИ ВНУТРЕННЕЙ СРЕДЫ, ИЛИ ОПОРНОТРОФИЧЕСКИЕ (СОЕДИНИТЕЛЬНЫЕ) ТКАНИ
К опорно-трофическим относятся самые разнообразные ткани по структуре и функции: кровь, лимфа, собственно соединительные ткани, хрящ,
кость. Все они происходят из эмбриональной соединительной ткани — мезенхимы и находятся во внутренней среде организма.
МЕЗЕНХИМА — ИСТОЧНИК ОПОРНО-ТРОФИЧЕСКИХ ТКАНЕЙ
Мезенхима появляется на ранней стадии развития зародыша. В период
дифференцировки от каждого сомита отделяется по два зачатка мезенхимы: в
направлении стенки тела — дерматом, внутрь — склеротом. Из листков
спланхнотома и из других зародышевых листков также выселяются клетки.
Мезенхима всех источников объединяется и образует внутреннюю среду развивающегося организма. Клетки мезенхимы имеют отростки, которыми устанавливают друг с другом контакты «по типу замка», в результате чего
ткань приобретает вид трехмерной сети, называемой синцитием. Пространства между клетками заполнены аморфным межклеточным веществом — основным веществом, которое и выполняет роль внутренней среды организма.
Через него осуществляется обмен веществ на ранних стадиях развития зародыша.
Клетки мезенхимы могут округляться, терять связь с другими клетками,
становиться подвижными. При этом к одним из них переходит функция переноса веществ, защиты путем фагоцитоза, они преобразуются в клетки крови. Другие клетки мезенхимы формируют стенку вокруг подвижных клеток
— образуются сосуды. Возникает кровеносная система, во много раз ускоряющая продвижение веществ, а следовательно, способствующая быстрому
развитию организма. Бывают клетки, которые специализируются на синтезе
межклеточного вещества как аморфного, так и волокнистого — образуется
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
система соединительных тканей. В процессе дифференцировки мезенхима
как ткань перестает существовать: она превращается в опорно-трофические
ткани.
ОБЩАЯ ХАРАКТЕРИСТИКА ОПОРНО-ТРОФИЧЕСКИХ ТКАНЕЙ
Все ткани данной группы состоят из клеток и межклеточного вещества,
обильно снабжаются кровью. Клетки их не имеют полярности. Межклеточное вещество по массе преобладает над клетками, хотя и является продуктом
деятельности клеток, с их помощью в нем происходит обмен веществ. Вместе с тем физико-химические свойства межклеточного вещества определяют
основные свойства тканей этого типа. У тканей с жидким межклеточным веществом (кровь, лимфа) основными функциями являются трофическая и защитная (фагоцитарной и иммунной защиты). Чем плотнее межклеточное вещество, тем меньше ткань выполняет трофическую функцию, тем больше на
первый план выходит функция опоры и механической защиты. В наибольшей
степени эти функции выражены у костной ткани, имеющей твердое минерализованное межклеточное вещество. В соответствии со степенью уплотнения межклеточного вещества ограничивается подвижность клеток, вплоть до
их полной неподвижности. Функция тканей тесно связана и с количественным соотношением клеток и межклеточного вещества: в тканях с выраженными функциями трофики, фагоцитарной и иммунной защиты клетки более
разнообразны и составляют больший процент, чем в тканях с функцией опоры и механической защиты. В последних значительно более развито межклеточное вещество, особенно его волокна.
Функции опорно-трофических тканей: 1) трофическая — участие в обмене веществ; 2) транспортная — перенос веществ; 3) защитная — способность клеток к фагоцитозу и участие в иммунных реакциях; 4) регуляторная
— участие в регуляции обменных процессов с помощью биологически активных веществ, циркулирующих в крови, а также вырабатываемых некоторыми клетками (базофилами, лаброцитами); 5) опорная и механической защиты.
В связи с особенностями строения и функции тип опорно-трофических
тканей можно разделить на отдельные ткани следующим образом (см. схему
на стр. 110).
Все разновидности опорно-трофических тканей способны быстро регенерировать (восстанавливаться) и приспосабливаться к меняющимся условиям существования. Будучи окружены со всех сторон довольно однородной
средой самого организма, клетки этих тканей (кроме эндотелия) не обнаруживают морфофункциональной полярности, характерной для эпителиев.
Эндотелий (рис. 25)—внутренняя выстилка кровеносных и лимфатических сосудов. Эндотелиальные клетки — эндотелиоциты вытянутые плоские, длиной 5—175 мкм и высотой 1—5 мкм — лежат плотно друг к другу в
виде однослойного пласта. Соединяются между собой черепицеобразными
наложениями, «по типу замка» и десмосомами. Электронная микроскопия
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
показала, что эндотелиоциты (кроме лимфатических капилляров) лежат на
базальной мембране и им присуща полярность в расположении органелл. На
поверхности, обращенной к сосуду, часто можно видеть короткие микроворсинки, особенно в эндотелии вен. В цитоплазме клеток имеются многочисленные пиноцитозные вакуоли, свидетельствующие о переносе продуктов из
крови в межклеточное вещество соединительной ткани. Пластинчатый комплекс лежит над ядром ближе к поверхности клетки, обращенной к просвету
сосуда. Эндотелий считается особым видом соединительнотканных клеток,
адаптированных к условиям функционирования.
КРОВЬ, ЛИМФА
Кровь — ткань внутренней среды жидкой консистенции, в которой
клетки (форменные элементы крови) составляют 40—45%, а на долю межклеточного вещества (плазмы) приходится 55— 60% объема ткани. На начальных этапах онтогенеза кровь тесно связана со своим источником — мезенхимой, а в процессе развития— кроветворения она сосредоточивается в
определенных местах— органах кроветворения. Таким образом, кровь оказывается заключенной в замкнутую систему сосудов. Это так называемая периферическая кровь. Большая ее часть находится в значительном удалении от
органов кроветворения и содержит зрелые форменные элементы, неспособные к делению в сосудистом русле. В организме кровь составляет 7—10% от
массы тела и выполняет разнообразные функции. У новорожденных и молодых животных относительный объем ее (на единицу массы тела) больше, чем
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
у взрослых. У новорожденных жеребят он составляет 23% при общем количестве крови 10 л, у взрослых лошадей 7% при количестве крови 50 л (И. С.
Ковальчук).
Рис. 25. Рыхлая соединительная ткань:
1 — кровеносный сосуд; 2 — эндотелий; 3 — базальная мембрана; 4 — эритроцит; 5 — перицит; 6 — фиброцит; 7 —
фибробласт; 8 — гистиоцит;
9— моноцит; 10 — нейтрофил
и 11 — ретикулоцит; 12 — макрофаг; 13 — плазмоцит; 14 —
малый лимфоцит; 15 — жировая клетка; 16 — пучок коллагеновых волокон; 17 — эластическое волокно; 18 — основное
аморфное вещество; 19 — тучная клетка.
Основные функции крови — транспортная, трофическая, дыхательная,
защитная, регуляторная, экскреторная. Разнося по организму различные вещества, она доставляет их в места утилизации: питательные вещества и кислород — ко всем органам и тканям, СО2 — в легкие для обмена на кислород,
жидкие конечные продукты обмена — в почки для экскреции (удаления),
ядовитые метаболиты — в печень для обезвреживания. Многие клетки крови
обладают фагоцитарной активностью, защищая организм от микробов и их
ядовитых продуктов. С кровью связана система иммунной защиты организма. Через кровь осуществляется гуморальная регуляция процессов жизнедеятельности, так как в ней циркулируют гормоны и другие биологически активные вещества.
Форменные элементы крови. У млекопитающих к ним относятся клетки крови (эритроциты и лейкоциты) и кровяные пластинки (цв. табл. IV).
Форменные элементы в крови находятся в определенных количественных
соотношениях, которые называют формулой крови, или гемограммой, а соотношения лейкоцитов, выраженные в процентах, — лейкоцитарной формулой
(табл. 3).
Эритроциты — самые многочисленные клетки крови. У млекопитающих они мелкие безъядерные, у остальных позвоночных — с недеятельными палочковидными ядрами.
У млекопитающих эритроциты представляют собой двояковогнутый
диск диаметром 3—8 мкм и толщиной от 0,8 мкм в центре до 3,5 мкм по пеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
риферии. Наблюдаются и отклонения в размерах эритроцитов. Примерно
80% имеют типичную для животного величину и называются нормоциты.
Остальные могут быть меньше — микроциты или больше — макроциты.
Обычно между размерами и количеством эритроцитов наблюдается обратная
зависимость: чем крупнее эритроциты, тем количество их меньше. Из позвоночных самые крупные эритроциты — 58 мкм и самое меньшее количество
их в 1 мм3 — 36 тыс. у протея (земноводное). Как правило, более мелкие
клетки свойственны животным с высоким уровнем тканевого метаболизма.
Такая же зависимость наблюдается и в онтогенезе. У телят эритроциты мельче (в среднем 3,5 мкм), но количество их больше (9—10 млн/мм3), чем у
взрослого крупного рогатого скота (4—5 мкм и 6 млн/мм3) (Л. В. Степовик и
др.). Цвет эритроцитов желтовато-зеленоватый, при больших скоплениях
красный.
Оболочка зрелого эритроцита отличается большой эластичностью.
Клетка может менять форму при прохождении узких капилляров, сильно вытягиваясь в длину. В цитоплазме отсутствуют органеллы, вся она заполнена
гемоглобином — комплексным соединением, в состав которого входит липопротеид глобин и железосодержащий пигмент гем. Сухое вещество эритроцита составляет 40%. 90—95% сухого вещества приходится на гемоглобин
(33% от общей массы эритроцита). Общее количество гемоглобина у лошади
массой 500 кг равно 6,8 кг.
Главнейшая функция эритроцитов — перенос кислорода от легких к
тканям и органам с помощью гемоглобина. Гемоглобин способен образовывать нестойкое соединение с кислородом — оксигемоглобин. В органах кислород отщепляется от оксигемоглобина и переходит в ткани тела, а взамен
эритроцит захватывает СО2 и выносит в легкие, где происходит обмен на кислород. Газообмен совершается в результате разности парциальных давлений
О2 и СО2 в легких и тканях организма.
Оболочка эритроцита устроена таким образом, что может адсорбировать
(удерживать) и переносить по сосудистому руслу аминокислоты, антитела,
токсины, лекарственные вещества.
С надмембранный комплексом оболочки эритроцита связана групповая
принадлежность крови и заряд эритроцита. В норме эритроциты заряжены
отрицательно. При снятии заряда происходит склеивание эритроцитов друг с
другом в виде монетных столбиков. При восстановлении заряда эритроциты
разъединяются, не разрушаясь.
3. Морфологический состав и размеры клеток крови
сельскохозяйственных животных (по В. Н. Никитину и др.)
www.timacad.ru
Свинья
Лошадь
0,8—
1,5
4,95,8
3—6
0,5
Коза
13—
17
Олень
7,7
Верблюд 11,0
Кролик
5—6
Курица
3-4
Индейка
2,7
2,9
3,24,5
4—8 5—10
30—
0,1—
6—10
36
0,5
4,3
8—10
30—
32
3-7
12
35
10,7
8—4 10,1
8
13,7 23-35
34
38
35
Количество кровяных
пластинок, 1 мм1,тыс.
моноциты
лимфоциты
базофилы
3—8
7—9,5
36—
72
50—
9—11
60
эозинофилы
Лейкоцитарная формула и размеры лейкоцитов
нейтрофилы
Количество лейкоцитов, 1 мм3/тыс.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
6—6,5 6-8 15—20
Крупный
рогатый 5—7
скот
Овца
8-13
Гусь
Утка
Диаметр эритроцита,
мкм
Животное
Количество эритроцитов 1 мм3/млн
Вракин В.Ф, Сидорова М.В.
2,3240—350
5,3
2,5—
18-35
300—350
3
24—
57
51— 4,5—
59
5
400
0,2— 57—
2-3 66—370
0,5
78
0,03— 55— 2,2—
5—6
500-600
0,5
57
4,5
6,8
50
6,6
4
0,3
42
1,7
1,5
1
55
2,5
240
5
3
60
2
23—130
3—9
37
53
40
30
23—
1—4 2,5
46
35
0
2,5
32
8,3
3,6
44
53
51
6,4
10
1,55,5
48
49
49
Своеобразная форма эритроцита позволяет ему активно участвовать в
процессах всасывания. Набухание эритроцитов приводит к резкому снижению активности всасывания. Эритроцит может участвовать в транспорте воды, так как легко набухает в гипотонической среде и отдает воду в гипертонической среде.
В эмбриональный период эритроциты образуются в желточном мешке,
печени, селезенке. Затем эритропоэз переходит в красный костный мозг, где
и совершается в течение всей жизни. Молодые формы клеток эритроидного
ряда — эритробласты — имеют ядра, общие органеллы, способны к делению и синтезу белков. В процессе дифференциации в них накапливается гемоглобин и происходит редукция органелл. На последующей стадии развития разрушается ядро, клетка превращается в эритроцит и выходит в кровяное русло. В молодых эритроцитах еще некоторое время при специальной
обработке заметна нежная сеть — остатки цитоплазматической сети и митоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
хондрий. Такие клетки называются ретикулоциты. Они могут составлять до
12% всех эритроцитов.
В сосудах в процессе функционирования эритроцит продолжает меняться. Будучи безъядерной клеткой, он не может поддерживать свою жизнедеятельность. В нем снижается содержание липидов и АТФ, увеличивается чувствительность к механическим и осмотическим воздействиям, происходит
нарушение газообменной функции, он стареет, погибает и утилизируется
макрофагами селезенки н красного костного мозга. Продолжительность жизни эритроцита у крупных животных дольше, чем у мелких: у быка — в среднем 120—140 дней, у овцы— 127, у свиньи — 72, у кролика — 30, у курицы
— 28 дней. По мере старения эритроциты уменьшаются в размерах: ретикулоциты имеют диаметр 7 мкм, молодые эритроциты — 5,5 мкм, старые —
3—4 мкм. На количество эритроцитов, продолжительность их жизни и функциональную активность влияют различные внешние и внутренние факторы
(см. ниже).
Лейкоциты, или белые кровяные клетки, — разнородная группа бесцветных ядерных клеток, в цитоплазме которых имеются органеллы и включения. Во взвешенном состоянии они округлой формы, в распластанном —
неправильной, так как образуют псевдоподии, обладая способностью к активному движению. В крови их гораздо меньше, чем эритроцитов. Один лейкоцит приходится на 500—1000 эритроцитов, а общее количество их составляет у млекопитающих 5—20 тыс. (см. табл. 3), у птиц — до 40 тыс. в 1 мм3
крови. Основная функция лейкоцитов—защитная, которую они выполняют
за пределами кровеносного русла. Данные клетки способны проходить через
эндотелий капилляров, через базальную мембрану, между эпителиальными
клетками и в соединительной ткани, резко меняя форму тела и ядра. Направление движения определяется, по-видимому, химическими веществами, выделяемыми при распаде тканей. Лейкоциты обладают способностью к фагоцитозу, вырабатывают биологически активные вещества, в том числе интерферон.
В зависимости от наличия или отсутствия в цитоплазме специфической
зернистости лейкоциты делят на гранулоциты и агранулоциты.
Зернистые лейкоциты — гранулоциты, в свою очередь, делят в зависимости от сродства их зернистости к определенным красителям на нейтрофильные, эозинофильные и базофильные. Зрелые гранулоциты имеют сегментированные ядра и не способны к делению. Развиваются гранулоциты в
красном костном мозге.
Нейтрофильные гранулоциты (нейтрофилы, гетерофилы) составляют у
животных разных видов 30—60% от общего количества лейкоцитов. Размеры
их на мазке равны 9—12 мкм, а в капле крови — в среднем 8,7 мкм. Нейтрофилы, находящиеся в кровеносном русле, имеют округлую форму и сегментированное ядро. С возрастом клетки количество ядерных сегментов увеличивается. У зрелых нейтрофилов их 3—5, а у некоторых животных до 10 сегментов, соединенных между собой тонкими перемычками. В цитоплазме содержится большое количество мелких гранул, воспринимающих как кислые,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
так и основные красители, поэтому на мазках цвет их розово-фиолетовый,
слабо заметный из-за малых размеров гранул (0,2—0,5 мкм). Зернистость
включает ряд гидролитических и окислительных ферментов, антибактериальные вещества, гликоген, а также кейлоны — вещества, регулирующие
процессы дифференцировки лейкоцитов. В периферической крови есть и небольшой процент молодых нейтрофилов: палочкоядерных (с подковообразными ядрами) и юных (с удлиненными несегментированными ядрами).
Продолжительность жизни нейтрофилов 4—20 суток, причем в кровяном русле они проводят от одного до нескольких дней. В очаге воспаления
их жизнь сокращается до нескольких десятков минут. Основная функция
нейтрофилов — фагоцитарная. Из кровеносного русла они активно выселяются и быстро перемещаются к очагу воспаления. Там буквально «набиваются» микробами и переваривают их с помощью лизосом и специальных гранул. Погибшие нейтрофилы выделяют вещества, привлекающие лимфоциты
и макрофаги, чем способствуют дальнейшему очищению очага воспаления.
Количество нейтрофилов увеличивается при беременности, длительной мышечной работе. При инфекционных заболеваниях и гнойных процессах в
крови обнаруживают большее количество юных и палочкоядерных нейтрофилов.
Эозинофильные гранулоциты (эозинофилы) насчитывают у разных животных от 1 до 10% от общего количества лейкоцитов. Они крупнее нейтрофилов: в капле крови их диаметр 9— 10 мкм, на мазке 12—14, а у лошади до
22 мкм. У зрелых эозинофилов обычно 2—3-сегментные ядра. У овцы, верблюда, кролика количество ядерных сегментов доходит до 5—6. Цитоплазма
слегка базофильна, в ней находятся общие органеллы и много специфических крупных (1—1,5 мкм, а у лошади до 3 мкм) гранул. Они содержат основные белки и поэтому окрашиваются кислыми красителями (эозин) в яркооранжевый цвет. В них обнаружены окислительные и гидролитические ферменты. По-видимому, они являются лизосомами. Эозинофилы способны к
фагоцитозу, но в значительно меньшей степени, чем нейтрофилы. Они принимают участие в защитных реакциях организма, обезвреживая токсины.
Участвуя в аллергических реакциях, они могут накапливать гистамин и инактизировать его, тормозить выведение гистамина из базофилов и тучных клеток. Наряду с сегментоядерными в периферической крови могут встречаться
палочкоядерные и юные эозинофилы. Они менее дифференцированы и хуже
выполняют специфические функции. Число эозинофилов увеличивается при
гельминтозных заболеваниях, аллергии, введении чужеродных белков. Даже
белки корма вызывают кратковременное (увеличение количества эозинофилов — алиментарную эозинофилию. При многих инфекционных заболеваниях, а также при действии некоторых гормонов гипофиза и надпочечников наблюдается снижение содержания эозинофилов — эозинопения. В период выздоровления число их увеличивается.
Базофильные гранулоциты самые малочисленные из лейкоцитов: от 0,3
до 3%. Это округлые клетки средних размеров. В капле крови их диаметр 9
мкм, на мазке—10—12, у лошади — до 21 мкм. Ядра базофилов слабее окwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рашиваются, чем у других гранулоцитов, и не обладают такой выраженной
дольчатостью. Зрелые базофилы слабосегментированные, у них лопастное
или бобовидное ядро, которое часто маскируется базофильной красноватосиреневой зернистостью, заполняющей цитоплазму крови. У лошади и крупного рогатого скота зерна средних размеров (0,5—0,8 мкм), у свиньи — более крупные (до 1,5 мкм). В зернах содержится гепарин, гистамин, серотонин
и другие биологически активные вещества. По-видимому, базофилы стимулируют воспалительную реакцию, повышают проницаемость стенок сосудов
и влияют на сокращение их мышечной оболочки. Возможно, они обладают и
противосвертывающим действием.
Незернистые лейкоциты — агранулоциты не содержат в цитоплазме
специальной зернистости, но на мазках крови в них можно видеть азурофильные гранулы, которые являются лизосомами. Агранулоциты — ядерные
клетки, в цитоплазме которых имеются общие органеллы. Но структура и
функция у них разные, потому их подразделяют на лимфоциты и моноциты.
Лимфоциты являются либо преобладающей, либо второй группой по
численности лейкоцитов (см. табл. 3). Это округлые или слегка овальные
клетки с базофильной цитоплазмой и округлым ядром, занимающим большую часть клетки. Лимфоциты делят на большие, средние и малые.
Большие лимфоциты размером около 15 мкм со светлым крупным
ядром, находятся в органах кроветворения — тимусе, селезенке, лимфатических узлах. Средние (7—10 мкм) и малые (4—7 мкм) лимфоциты циркулируют в периферической крови. Основную массу составляют малые лимфоциты, на долю средних приходится 5—10%.
Малые лимфоциты — с округлым плотным ядром и ободком дитоплазмы, в которой имеется небольшое количество органелл. Это разнородная популяция клеток, различающихся по степени дифференцировки и выполняемым функциям. С помощью современных методов исследований (радиоавтографический, иммуно-морфологический и др.) удалось установить, что среди
лимфоцитов есть как короткоживущие (срок жизни 3—6 дней), так и долгоживущие (срок жизни сопоставим с продолжительностью жизни особи).
Встречаются клетки как малодифференцированные, способные к делению и
дальнейшей специализации, так и высокодифференцированные. Попадаются
среди них и вышедшие из костного мозга стволовые клетки: 1 на 10 тыс. в
красном костном мозге и 1 на 100 тыс. в периферической крови. Считается,
что они, поступая в органы кроветворения, могут давать начало любым клеткам крови, а попадая в соединительную ткань, дифференцируются в фибробласты, макрофаги, тучные и другие клетки.
Среди дифференцированных лимфоцитов различают Т- и В-лимфоциты.
В крови преимущественно находятся многократно циркулирующие и долгоживущие Т-лимфоциты (до 70%). Реже встречаются короткоживущие Влимфоциты (до 30%).
При некоторых заболеваниях их соотношение меняется. Они отличаются друг от друга тем, что Т-лимфоциты меньше и у них более темная цитоплазма и более гладкая поверхность, чем у В-лимфоцитов, у которых заметно
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
светлое перинуклеарное пространство, а в электронный микроскоп видны
многочисленные отростки. В онтогенезе лимфоциты дифференцируются
позднее других клеток крови: у крупного рогатого скота на 4—5-м месяце
внутриутробного развития. Развиваются лимфоциты из стволовых клеток,
которые выходят из костного мозга и заселяют тимус, миндалины, лимфатические узлы, пейеровы бляшки и другие лимфатические образования.
Т-лимфоциты размножаются и дифференцируются в тимусе. Они ответственны за клеточный иммунитет: уничтожают чужеродные и патологически
измененные клетки, патогенные грибы и вирусы. Есть сведения о том, что Тлимфоциты определяют направленность кроветворения, играют роль в противоопухолевом иммунитете, препятствуют образованию аутоантител, являются клетками иммунной памяти. В-лимфоциты ответственны за гуморальный иммунитет: вырабатывают антитела против бактерий и вирусов, дифференцируются в плазматические клетки. Лимфоциты осуществляют свои
функции как в кровяном русле, так и за его пределами, выходя в окружающие ткани.
Моноциты составляют 2—5% от всех лейкоцитов. Это самые крупные
клетки крови. Их размер в капле крови 9—12 мкм, на мазке 15—20 мкм. Образуются они в красном костном мозге из моноцитобластов. Циркулирующие
в крови представляют собой малодифференцированные клетки. У них крупные бобовидные, подковообразные или лопастные ядра с мелкодисперсным
хроматином и мелкими ядрышками. Цитоплазма слабобазофильна, содержит
в небольшом количестве общие органеллы, кроме комплекса Гольджи, который хорошо развит. В крови моноцит пребывает от одного до нескольких суток. Моноциты способны к амебоидному движению и фагоцитозу. Выселяясь
из кровеносного русла, они дифференцируются, превращаясь в специальные
макрофаги различных тканей и органов. В настоящее время считается, что
видоизмененными моноцитами являются макрофаги соединительной ткани
(гистиоциты), органов кроветворения (отростчатые макрофаги), нервной системы (микроглия), костной и хрящевой ткачей (остеокласты) и др. В процессе дифференцировки в моноцитах увеличивается содержание гранулярной
цитоплазматической сети, вакуолей, лизосом. Моноциты крови и специальные макрофаги, произошедшие из них, способны фагоцитировать бактерии,
остатки клеток, антигены, чужеродные белки, токсины. Они секретируют антибактериальный белок ли-зоцим и другие биологически активные вещества.
Погибая, моноциты обогащают окружающую среду веществами, стимулирующими размножение клеток.
Кровяные пластинки крови млекопитающих — это безъядерные «осколки» гигантских клеток костного мозга — мегакариоцитов. Участки цитоплазмы, отрываясь от мегакариоцитов, выходят в кровяное русло и принимают участие в свертывании крови. Кровяные пластинки неправильной формы, их размеры в пределах 2—4 мкм, средняя продолжительность жизни 5—
8 суток. В них содержится вещество тромбопластин, без которого свертывание крови невозможно. На поверхности кровяных пластинок имеются рецепторы, обеспечивающие слипание пластинок между собой и прилипание к
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
стенкам сосуда. Ими выделяются вещества, стимулирующие сужение сосуда.
Уменьшение количества пластинок или нарушение их структуры ведет к кровоточивости. У животных остальных классов типа хордовых вместо кровяных пластинок имеются тромбоциты — мелкие овальные ядерные клетки,
выполняющие те же функции, что и кровяные пластинки.
Плазма крови — жидкое межклеточное вещество, соломенно-желтого
цвета, слегка щелочной реакции, на 90—93% состоящее из воды. Из 7—10%
сухого вещества 6—8% приходится на белки, 0,9%—соли и около 1%—на
глюкозу. В плазме крови находятся различные органические соединения, в
том числе питательные вещества, конечные продукты обмена (мочевина,
креатинин, билирубин и др.), биологически активные вещества (гормоны,
ферменты), ионы и др. Среди белков плазмы большое значение имеют альбумины, так как они способствуют переносу плохо растворимых в воде веществ, глобулины, в состав которых входят иммунные белки, и фибриноген,
принимающий участие в свертывании крови.
Лимфа. Как и кровь, она является жидкой тканью и состоит из плазмы и
форменных элементов. Плазма лимфы (лимфоплазма) образуется из тканевой
жидкости и плазмы крови, которая выпотевает из капилляров. По химическому составу она близка плазме крови, но в ней меньше белков.
Основные клетки лимфы — лимфоциты, которыми она обогащается,
проходя через лимфатические узлы. В лимфе встречаются и другие форменные элементы, но в гораздо меньших количествах, чем в крови. Лимфоциты,
как и в крови, неоднородны по составу. Основная масса их имеет диаметр
4—6 мкм и очень высокое ядерно- цитоплазменное отношение. Наряду с
этим встречаются более крупные и светлые лимфоциты размерами 7—18
мкм.
Лимфа течет по системе лимфатических сосудов, которые соединяются с
кровеносной системой (лимфа сливается с кровью). В разных участках тела
лимфа существенно отличается как по количеству и спектру клеток, так и по
химическому составу лимфоплазмы. Так, в лимфе, оттекающей от кишечника, содержится много жиров (до 4%).
Кровь и лимфа как составные части внутренней среды организма. Кровь,
лимфа и рыхлая соединительная ткань являются той внутренней средой организма, в которой происходят процессы его жизнедеятельности. Естественно, что особенности обмена веществ, связанные с особенностями организации и образом жизни животного, изменения в процессах жизнедеятельности
с возрастом, при различных физиологических состояниях организма и под
влиянием изменений окружающей среды сказываются в первую очередь на
химическом составе плазмы, морфологическом составе и функциональной
активности клеток крови.
Видовые и породные особенности состава крови. Из сельскохозяйственных животных больше всего эритроцитов в 1 мм3 крови содержится у козы,
затем в порядке убывания идут верблюд, лошадь, овца, крупный рогатый
скот, свинья, кролик, курица. Однако по размерам эритроцитов этот ряд надо
читать с конца: наибольший размер их у курицы— 100 мкм3, у свиньи — 60,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
у коровы — 55, у лошади — 50 мкм3. В то же время концентрация гемоглобина у лошади наивысшая—14— 16 г/%, у крупного рогатого скота—12—13,
у овцы, свиньи — 10—12, у курицы —6—9 г/%.
Содержание лейкоцитов и их соотношение также заметно различаются у
разных животных. У птиц число лейкоцитов в 1 мм3 в среднем составляет
30—35 тыс., в связи с чем их кровь называется лейкоцитарной, а у домашних
млекопитающих данный показатель обычно не превышает 10 тыс., за исключением свиньи, у которой в 1 мм3 крови содержится 15—20 тыс. лейкоцитов.
При этом у лошади — нейтрофильный профиль лейкоцитарной формулы то
есть у нее наибольший процент среди лейкоцитов составляют нейтрофильные гранулоциты (до 72%), а у рогатого скота кровь имеет лимфоцитарный
профиль: у них лимфоциты составляют до 60—80%- У свиньи, оленя, верблюда в 1 мм3 крови примерно равное количество лимфоцитов и нейтрофилов.
Порода также накладывает отпечаток на состав и количество крови. У
верховых лошадей в 1 мм3 крови содержится 9— 9,5 млн эритроцитов и 10—
11 тыс. лейкоцитов, у рысистых соответственно 8,5—9 млн и 8—9 тыс., а у
тяжеловозов 7—8 млн. У чистокровной верховой породы на 100 кг массы
приходится около 10 л крови, у тяжеловозов — 6,5 л. В 1,5 раза меняется
число лейкоцитов у разных пород свиней.
Возрастные изменения. У новорожденных гемограмма резко отличается
от взрослых животных. У них больше эритроцитов и мало лейкоцитов, особенно эозинофилов и В-лимфоцитов. В первые 2—4 недели жизни резко
снижается количество эритроцитов, уменьшаются их размеры, увеличивается
число лейкоцитов в основном за счет короткоживущих В-лимфоцитов. После
отъема от матери отмечается резкий подъем числа лейкоцитов, затем их снижение и к началу полового созревания лейкоцитарная формула приобретает
вид, характерный для взрослых животных данного вида, типа, породы. При
этом замечено, что быстрорастущие животные имеют большее количество
эритроцитов, лейкоцитов и гемоглобина в крови, чем их медленнорастущие
сверстники. У старых животных происходит уменьшение содержания всех
форменных элементов крови в среднем в 1,5 раза, за исключением Влимфоцитов, число которых не меняется. Эритроциты приобретают более
сферическую форму.
Физиологическое состояние. Беременность заметно влияет на количественный состав крови: в первой половине беременности увеличивается число
гранулоцитов, во второй — количество эритроцитов и всех лейкоцитов. Роды
приводят к резкому снижению содержания эритроцитов и лейкоцитов, особенно лимфоцитов и базофилов. По характеру крови матери можно судить о
крови ее плода, так как обнаружена высокая положительная корреляция по
лейкоцитам и гемоглобину и отрицательная по количеству эритроцитов.
Во время активной мышечной работы (при беге, тренинге, скачках) на
30—40% увеличивается объем циркулирующей крови, содержание в ней
эритроцитов (у верблюда — на 20%, у лошади— на 20—60%), гемоглобина
на 25%, лейкоцитов на 20— 30%- Для возвращения количества эритроцитов
к исходному уровню необходим отдых в течение 3—4 ч, лимфоцитов — в теwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
чение 6 ч. Регулярный тренинг способствует стойкому увеличению количества эритроцитов, их размеров и концентрации в них гемоглобина.
Влияние внешних факторов. Состав крови изменяется в зависимости от
сезона года. Замечено, что у лошадей в зимне-весенние месяцы содержание
эритроцитов большее, чем в летне-осенние. У телят, родившихся летом, количество эритроцитов равно 11 млн/мм3, а зимой — 9,8 млн/мм3, у ягнят же
максимальное количество эритроцитов приходится на осень. Большее число
лейкоцитов у телят обычно бывает осенью. При этом весной и летом лейкограмма имеет нейтрофильный характер, а зимой — лимфоидный.
Даже в течение суток количество форменных элементов в периферической крови не остается постоянным. Утром эритроцитов несколько
больше, чем днем; максимальное число лейкоцитов приходится на 11 —15 и
22 ч, а минимальное — на 5—7 и 19 ч.
Акклиматизация к высокогорным условиям, воздействие постоянного
магнитного поля, ультрафиолетовое облучение, ионизация воздуха способствуют увеличению количества эритроцитов и гемоглобина и уменьшению
лейкоцитов в крови. Воздействие магнитного поля, кроме того, сдвигает лейкограмму в сторону увеличения В-лимфоцитов. От долгого пребывания в
темноте снижается количество эритроцитов и концентрация гемоглобина в
них, а от сильного шума уменьшается число лейкоцитов. Транспортировка и
вызванный ею механический стресс приводят к сдвигу лейкоцитарной формулы: увеличивается содержание нейтрофилов и уменьшается — лимфоцитов. Нормализация крови происходит только через месяц.
Внесение в рацион различных добавок, заменителей, стимуляторов роста способно изменить состав крови. Есть данные о том, что недостаток протеина в рационе приводит к снижению количества эритроцитов и увеличению лейкоцитов у овец, супоросных и подсосных свиноматок, в то время как
у растущих поросят тот же рацион вызывает некоторое увеличение эритроцитов. При использовании таких заменителей белка, как мочевина, карбамид,
диаммонийфосфат, увеличивается количество эритроцитов и лейкоцитов в
крови. Добавки минеральных премиксов, сапропеля, таких микроэлементов,
как Со, Си, J, Fe, приводят к увеличению количества эритроцитов, а по некоторым данным, и лейкоцитов в крови. Добавки витаминов в комплексе с органическими кислотами приводят к сдвигу лейкограммы в сторону нейтрофилов (С. Дурдыев).
Все перечисленные факты говорят о том, что кровь — чрезвычайно лабильная ткань, чутко реагирующая на различные изменения как эндо-, так и
экзогенного характера. Это следует учитывать при исследовании крови и
брать ее в строго одинаковых условиях.
РЫХЛАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Данная ткань практически есть в каждом органе. Она сопровождает сосуды и нервы, образует внутренний каркас — строму большинства компактных органов, формирует основу слизистой оболочки трубкообразных органов и подкожной клетчатки. Ей присущи все основные функции тканей внутренней среды. В разных местах организма преобладает та или иная функция,
что отражается на структуре рыхлой соединительной ткани. Там, где ведущая
— опорная функция, в ней относительно больше волокнистых структур, а где
основная функция защитная — больше клеточных форм.
Рыхлая соединительная ткань состоит из клеток и межклеточного вещества (см. рис. 25). Среди клеток рыхлой соединительной ткани различают как
оседлые, так и подвижные клетки, попадающие из крови. Постоянными клетками являются фибробласты, фиброциты, гистиоциты.
Фибробласт — основная клеточная форма. Функция ее — синтез и экструзия составных частей межклеточного вещества. В период эмбриогенеза
фибробласты дифференцируются из клеток мезенхимы. В постнатальный период онтогенеза их источником могут быть молодые фибробласты, еще способные к делению, ретикулярные и адвентициальные клетки соединительной
ткани и подвижные лимфоцитоподобные стволовые клетки, мигрирующие из
костного мозга в кровоток, а оттуда — в соединительную ткань. В процессе
дифференцировки фибробласты приобретают характерные черты.
Дифференцированный фибробласт, активно синтезирующий элементы
межклеточного вещества, — крупная (30—40 мкм) веретеновидная или отростчатая клетка с овальным ядром, расположенным в средней части. В ядре
видны 1—3 крупных ядрышка и диффузный, слабобазофильный хроматин. В
цитоплазме фибробласта различают две зоны: эктоплазму и эндоплазму. Эндоплазма — часть цитоплазмы, прилежащая к ядру, базофильна, в ней много
таких органелл, как зернистая цитоплазматическая сеть и комплекс Гольджи,
так как основной функцией клетки служит синтез белков на экспорт. Митохондрии и лизосомы развиты умеренно. Эктоплазма — периферический
слой цитоплазмы — слабо окрашивается гистологическими красителями, содержит мало органелл и много секреторных гранул, микротрубочек и микрофибрилл, образующих цитоскелет и определяющих подвижность клетки. В
световой микроскоп границы фибробластов неразличимы.
По мере старения клетки и накопления вокруг нее межклеточного вещества, зона эндоплазмы сужается, количество органелл уменьшается, хроматин конденсируется, ядро уплощается, клетка вытягивается и называется
фиброцитом. Фиброциты не способны к делению, их синтетическая деятельность гораздо ниже, чем у фибробластов.
Гистиоцит, или тканевый макрофаг, — вторая по распространению клеточная форма рыхлой соединительной ткани. В состоянии покоя гистиоцит
веретеновидной или овальной формы с овальным или бобовидным ядром,
несколько сдвинутым в сторону. Хроматин в виде крупных глыбок. Цитоплазма имеет комковатый вид. Субмикроскопически в ней обнаруживается в
умеренных количествах цитоплазматическая сеть, комплекс Гольджи и
большое количество лизосом. При попадании в соединительную ткань разwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дражающего агента (инфекции, чужеродного тела, погибших тканевых элементов) гистиоцит активизируется и приобретает черты, характерные для
любого макрофага. Клетка принимает неправильные очертания. У нее образуются псевдоподии, способность к амебоидному движению и фагоцитарной
активности.
В цитоплазме появляются фагосомы — вакуоли с фагоцитированными
частицами. По современным представлениям, гистиоциты происходят из моноцитов, выселившихся в соединительную ткань.
В рыхлой соединительной ткани всегда имеются малодифференцированные клетки, благодаря чему эта ткань оказывается очень лабильной — быстро регенерирует и легко отзывается на влияние различных
факторов перестройкой своей структуры. К таким клеткам относятся: перициты, адвентициальные, ретикулярные и лимфоцитоподобные, выселяющиеся из сосудистого русла.
Перицит, или периваскулярная клетка, — удлиненная отростчатая клетка с овальным ядром. Тесно прилежит к эндотелию капилляров. Есть данные,
позволяющие думать, что перициты принимают участие в изменении просвета сосудов, вызывая набухание клеток эндотелия при соответствующей нервной регуляции. Они способны к делению и дифференцировке в другие клетки
соединительной ткани и в гладкие миоциты.
Адвентициальная клетка по внешнему виду очень похожа на перицит:
удлиненная, отростчатая, с удлиненным ядром, бедная органеллами. Прилежит к сосудам, способна к делению. Такие клетки дают начало многим оседлым клеткам рыхлой соединительной ткани (фибробластам, тучным, жировым и др.). Одни исследователи считают адвентициальные клетки самостоятельной формой, другие — ранней стадией дифференцировки фибробласта,
третьи отождествляют их с перицитами.
Ретикулярная клетка, или ретикулоцит, — удлиненная отростчатая, с
неправильным овальным ядром. В нем диффузный хроматин и слабо контурированное ядрышко. Способна к делению и дифференцировке в другие
клетки соединительной ткани. Вырабатывает ретикулярные волокна. Ретикулоциты, соединяясь друг с другом своими отростками и тесно переплетаясь с
ретикулярными волокнами, образуют сеть (reticulum). Если преобладающими
клетками в соединительной ткани являются ретикулоциты, а волокнами —
ретикулярные, говорят о ретикулярной ткани (в органах кроветворения).
Лимфоцитоподобная клетка по морфологическим признакам похожа на
малый лимфоцит. В настоящее время ей приписывают роль стволовой клетки
как крови, так и соединительной ткани.
В рыхлой соединительной ткани много различных специализированных
клеток, чем объясняется разнообразие функций, выполняемых этой тканью.
Специализированные клетки являются либо дифференцированными формами оседлых клеток соединительной ткани, развивающимися в ней с самого
начала (плазмоциты, жировые, тучные клетки), либо пришлыми — вышедшими из сосудистого русла для выполнения своей функции (все формы
лейкоцитов).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Плазматическая клетка — небольшая клетка округло-овальной или слегка угловатой формы с округлым, эксцентрично расположенным ядром.
Крупные глыбки хроматина лежат по периферии в виде характерной картины, напоминая спицы колеса. Крупное ядрышко находится в центре ядра. В
цитоплазме большое количество зернистой цитоплазматической сети„ что
определяет ее сильную базофилию. Комплекс Гольджи хорошо развит и лежит вместе с центросомой в светлой околоядерной зоне цитоплазмы. Плазмоциты дифференцируются из В-лимфоцитов, выселившихся в соединительную ткань. Плазматические клетки располагаются обычно недалеко от сосудов, так как вырабатывают иммуноглобулины, которые проникают через
тканевую жидкость и лимфу в кровь.
Тучная клетка (лаброцит, тканевый базофил)—крупная клетка неправильно-овальной формы с небольшим центральным ядром. В цитоплазме
есть митохондрии и хорошо развит комплекс Гольджи. Отличительная особенность лаброцитов — большое количество крупных базофильных гранул,
заполняющих всю цитоплазму и маскирующих структуру клетки. В гранулах
содержится гепарин, гистамин, а у некоторых животных и серотонин. Тучные клетки лежат вблизи капилляров и венул в соединительной ткани печени, легких, кишечного тракта и других органов. Выделяя в окружающую
среду гепарин и гистамин, они регулируют местный обмен веществ, изменяют проницаемость стенок сосудов, препятствуют свертыванию крови, способствуют оводнению (разжижению) основного вещества соединительной
ткани, участвуют в защите организма от инфекции. Происхождение тучных
клеток не выяснено. По своей структуре и функции они близки, но не идентичны базофильным гранулоцитам крови. Продолжительность их жизни колеблется от 9 дней до 12 месяцев.
Жировая клетка, или липоцит — крупная округлая клетка, вид и размеры которой во многом зависят от количества жира в ней. Незрелые жировые
клетки, а также клетки бурой жировой ткани содержат несколько мелких жировых капель. У таких клеток округлое центральное ядро, в цитоплазме хорошо развиты общие органеллы, особенно гладкая цитоплазматическая сеть. В
процессе созревания происходит активный синтез и накопление липидов.
Жировые капли увеличиваются в размерах, сливаются друг с другом и формируют огромную жировую каплю, занимающую большую часть цитоплазмы. При этом клетка увеличивается до 150 мкм, ядро оттесняется к плазмолемме, органелл становится меньше. Скорость синтеза и обмена липидов в
жировой клетке падает. Образуются жировые клетки из адвенгициальных и
фибробластов и располагаются обычно группами или поодиночке вблизи капилляров.
Пигментные клетки в соединительной ткани млекопитающих встречаются редко. Более распространены у рыб, амфибий и рептилий. Это отростчатые клетки с центрально-расположенным округлым ядром. В цитоплазме
содержится большое количество зерен пигмента. У крупного рогатого скота
и свиньи преобладают крупные звездчатые клетки. У лошади они мельче и
встречаются реже, еще мельче, веретеновидной формы — у овец. Под возwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
действием гормонов или нервной системы клетки могут менять форму: втягивать и вытягивать отростки,
что ведет к изменению окраски. Среди пигментных
клеток различают хроматофоры (меланофоры) —
клетки, неспособные к синтезу пигментов, но способные к их накоплению путем фагоцитоза, в меланоциты — способные и к синтезу, и к накоплению пигментов. Происхождение пигментных клеток до конца
неясно. Скопления, их в радужной оболочке глаза и в
некоторых участках кожи (мошонки, вымени и др.)
позволяют говорить о пигментной ткани.
Межклеточное вещество рыхлой волокнистой
соединительной ткани существует в виде аморфного
основного вещества и нескольких видов волокон —
коллагеновых, эластических, ретикулярных. Все виды
межклеточного вещества синтезируются и секретируются фибробластами, а некоторые — ретикулоцитами (основное вещество и ретикулярные волокна).
Коллагеновые волокна (рис.. 26) образованы молекулами белка тропоколлагена. Он относится к фибриллярным белкам и состоит из трех полипептидных
цепочек, скрученных в виде тройной спирали. В состав полипептидов входят различные аминокислоты,
поэтому химически различают несколько десятков
разновидностей коллагена (32 у позвоночных). Основными аминокислотами, определяющими свойства
коллагена, являются пролин, оксипролин (вместе —
30%), лизин и глицин (26% от общего содержания
аминокислот). Молекулы тропоколлагена, располагаясь одна за другой по принципу «хвост к голове»,
формируют протофибриллу коллагена. ПротофибрилРис. 26. Схема строе- лы, склеенные гликозаминогликанами и другими угния коллагенового леводами, представляют собой коллагеновые фибрилволокна:
лы. Фибриллы склеиваются в волокна.
1 — коллагеновое воВ электронный микроскоп видно, что коллагенолокно; 2 — фибрилла
вые волокна обладают поперечной и продольной исколлагена; 3 — протофибрилла; 4— мо- черченностью, которая выявляет упорядоченность
лекула
коллагена расположения молекул тропоколлагена и протофиб(тропочоллаген); 5 — рилл в волокне. Коллагеновые волокна могут быть
одна из трех поли- различной толщины (от 0,1 до 15 мкм и больше) и
пептидных
цепочек длины. Они объединяются в пучки, но никогда не размолекулы коллагена.
ветвляются и не анастомозируют друг с другом. В
коллагеновом волокне спирально закручены не только молекулы тропоколлагена, но и протофибриллы и даже фибриллы. Это дает возможность всему волокну несколько вытягиваться при очень большом
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
натяжении. Прочность коллагеновых волокон велика — 6 кг/мм2. Особенностью коллагеновых волокон, определяющих их название (colla — клей),
является способность при длительном проваривании в воде давать животный
клей — желатин. Коллаген сильно набухает в слабых кислотах и щелочах,
переваривается пепсином и хорошо усваивается. Однако белки коллагена неполноценные, так как содержат мало незаменимых аминокислот.
Фибробласты синтезируют тропоколлаген на зернистой цитоплазматической сети и секретируют их с помощью комплекса Гольджи. Сборка молекул тропоколлагена в фибриллы и волокна происходит за пределами клетки,
хотя и в непосредственной близости от нее. Коллаген один из самых распространенных белков. По массе он составляет около 1/3 белков всего организма
и имеется, кроме рыхлой соединительной ткани, в дерме кожи, сухожилиях,
хрящах, костях, стенках кровеносных сосудов, в органах нервной системы, в
стекловидном теле глаза и в других местах. Основная функция коллагена
опорная.
Эластические волокна (см. рис. 25) по свойствам и внешнему виду сильно отличаются от коллагеновых. Они построены из двух компонентов:
аморфного — в центре волокна и фибриллярного — по его периферии. В них
нет ни продольной, ни поперечной исчерченности, не организуются в пучки,
зато разветвляются и анастомозируют друг с другом, образуя широкопетлистую сеть. Толщина их в рыхлой соединительной ткани равна 1—3 мкм. На
обычных препаратах они слабо окрашены, выявляются специальными красителями (орсеин).
Прочность эластического волокна небольшая — 0,06 кг/мм2. Эти волокна не развариваются, не разрушаются слабыми кислотами и щелочами, не
перевариваются пищеварительными ферментами, за исключением эластазы
поджелудочной железы. Обладают эластичностью — способностью к восстановлению формы и размера после растяжения или сдавливания.
Ретикулярные (ретикулиновые, или аргирофильные) волокна по химическому составу и структуре похожи на коллагеновые, что позволяет многим
авторам считать их преколлагеновыми, или молодыми коллагеновыми волокнами. Однако у них есть и отличительные свойства, из-за чего их выделяют в самостоятельный вид волокон. Они анастомозируют и ветвятся, импрегнируются серебром, не образуют пучки, не перевариваются пепсином, по
растяжимости занимают промежуточное место между коллагеновыми и эластическими волокнами. Распространены ретикулярные волокна в строме органов кроветворения, в базальных мембранах, вокруг кровеносных сосудов,
мышечных и нервных волокон.
Основное аморфное вещество — микроскопически бесструктурная, прозрачная основа, в которую заключены клетки и волокна. Оно имеет вид студенистой массы, но может принимать вид пленок и пластинок. По химическому составу это сложный комплекс веществ различных по молекулярной
массе, природе, свойствам. Основу его составляют гликозаминогликаны,
протеогликаны, гликопротеиды — вещества белково-углеводной природы,
которые придают ему свойства геля. В его состав входят также белки плазмы
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
крови, гормоны, свободные электролиты, пептиды, аминокислоты, сахара,
вода и другие предшественники и продукты распада более сложных соединений. Жидкая часть основного вещества — тканевая жидкость постоянно
циркулирует, поступая из артериальных капилляров и всасываясь венозными
и лимфатическими капиллярами. Тем самым основное вещество соединительной ткани является областью, где осуществляется активный обмен веществ. Нарушение оттока жидкости, уплотнение межклеточного вещества
уменьшают диффузионные процессы, приводят к снижению уровня обмена
веществ.
Возрастные изменения рыхлой соединительной ткани и ее реакция
на различные воздействия. С возрастом в соединительной ткани происходит уменьшение активности фибробластов,
появляется больше малодеятельных фиброцитов, тучных клеток. Количество гистиоцитов увеличивается в ранний период постнатального онтогенеза (в период подсоса), потом снижается. Уменьшается и число малодифференцированных клеток. По данным А. Е. Борисенко, у новорожденных поросят фибробласты составляют около 30% от всех клеток подкожной рыхлой
соединительной ткани, гистиоциты—13 %, малодифференцированные клетки
— более 50%, а у взрослых свиней — соответственно 55, 15 и 30%- В межклеточном веществе увеличивается содержание волокон, уплотняется основное вещество, сшивки коллагеновых фибрилл и волокон уплотняются. Образуются долгоживущие, малоактивные коллаген и эластин. Волокна пропитываются минеральными солями и делаются более ломкими.
У скороспелых животных (свиней) процессы роста клеток и синтеза
межклеточного вещества активно протекают в более раннем возрасте, чем у
позднеспелых, у которых они растянуты во времени. Замечены и некоторые
сезонные колебания клеточного состава. У сельскохозяйственных животных
летом увеличивается количество плазматических клеток.
Физиологическое состояние организма заметно влияет на развитие специализированных клеток. Отмечено увеличение числа плазматических клеток на высоте лактации, при беременности, облучении УФ, повышенной температуре окружающей среды. От низкой температуры (5°С) и голода снижается число плазмоцитов. Малоподвижность беременных животных приводит
к рождению детенышей с эмбриональными чертами в строении соединительной ткани: в ней оказываются недоразвитыми волокнистые структуры, а из
клеток присутствуют лишь фибробласты и макрофаги.
Полноценное сбалансированное кормление способствует своевременному развитию соединительной ткани. Недокорм приводит к снижению активности фибробластов. Сами клетки при этом оказываются мельче, с
мелкими плотными ядрами, узкой зоной эндоплазмы. Уменьшается при недокорме и число тучных клеток. При нормализации кормления не происходит полной компенсации структур. Замена недостающего протеина в рационе
белково-витаминным концентратом ведет к увеличению количества гистиоцитов у растущих цыплят, поросят, бычков с 10 до 30% (В. А. Латышев, А. В.
Букреев). Тип кормления в меньшей степени влияет на характер соединиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тельной ткани, но все же концентратный тип кормления приводит к большему развитию фибробластов, гистиоцитов и жировых клеток, чем рацион с
умеренным содержанием концентратов (А. Е. Борисенко).
МАКРОФАГИЧЕСКАЯ (РЕТИКУЛО-ЭНДОТЕЛИАЛЬНАЯ, ИЛИ
РЕТИКУЛО-ГИСТИОЦИТАРНАЯ) СИСТЕМА
Клетки крови и рыхлой соединительной ткани, принимающие участие в
осуществлении фагоцитарной функции, представляют собой единую систему
неспецифической защиты, рассредоточенную по всему организму. В различных органах клетки ретикуло-гистиоцитарной системы (РГС) имеют специфические черты. Это гистиоциты, эндотелиальные клетки венозных синусов
печени, надпочечника, гипофиза, макрофаги органов кроветворения, легкого,
нервной ткани, серозных оболочек, остеокласты и другие клетки, способные
захватывать из тканевой жидкости частицы коллоидов, инородные частицы,
погибающие клетки, неклеточные структуры, бактерии. Несмотря на свою
разбросанность, система эта работает слаженно, однотипно реагируя на появление раздражающего агента. Она участвует не только в фагоцитозе и переваривании бактерий и продуктов распада, но и в создании иммунитета,
взаимодействуя с лимфоцитами и плазматическими клетками. Регулируется
РГС нервной и эндокринной системами и местными механизмами.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ
Если в соединительной ткани преобладает один какой-либо вид клеток,
которые выполняют специфическую функцию и придают ткани особые свойства, говорят о соединительной ткани со специальными свойствами. Если в
ткани имеется много пигментных клеток, такую ткань называют пигментной.
Ткань, в которой преобладают ретикулярные клетки и вырабатываемые или
ретикулярные волокна, называют ретикулярной, а ткань, образованную жировыми клетками, — жировой.
Жировая ткань является депо жира и воды, источником энергии, принимает участие в терморегуляции, выполняет амортизационную и защитную
функции. Она состоит из скоплений жировых клеток, разделенных на дольки
рыхлой соединительной тканью. В прослойках рыхлой соединительной ткани
проходят сосуды и нервы. Жировые клетки в пределах дольки лежат плотно,
так что долька по форме напоминает виноградную гроздь.
По расположению в организме различают подкожную (шпик у свиньи,
полив, курдюк у овцы, горбы у верблюда), межмышечную и внутреннюю
(околопочечную, околосердечную, большого и малого сальников, внутримошоночную) жировую ткань. Они отличаются друг от друга по морфологическим признакам и химическому составу.
Раньше других в начале плодного периода появляется жировая ткань
около почек, а затем на сальниках и вокруг внутренних органов. Позже всех
при внутриутробном развитии образуется подкожная жировая ткань. У новоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рожденных поросят общее количество жировой ткани в тушке не превышает
10— 15%, а размеры клеток — 20—40 мкм. Основная доля жировой ткани
приходится на межмышечную (60—70%). После рождения жировая ткань
растет столь интенсивно, что к 4-месячному возрасту у поросят ее количество увеличивается в 500 раз, при увеличении массы тела — в 30 раз (А. Ф.
Шурмухин). У скороспелых свиней и свиней сального типа жировая ткань
закладывается раньше и развивается быстрее, чем у позднеспелых и беконных. Кастрация приводит к усиленному отложению жира под кожей и в
мошонке. У 15-месячных бычков-кастратов масса межмышечной жировой
ткани достигает 23 кг, сальниковой — 13, околосердечной — 0,3, околопочечной — 5, подкожной — 6, внутримошоночной — 3,5 кг. Общее количество жировой ткани в организме крупного рогатого скота по массе составляет
10— 15 %, у свиней — до 30 %.
У молодых животных в жировой ткани идет активный липид-ный обмен.
У взрослых животных он снижается, особенно во внутренних жировых депо.
В жировой ткани становится меньше воды и белка, больше жира и каротиноидов — пигментов, придающих ей желтый цвет.
Количество, размеры клеток и химический состав жира изменяются при
различных физиологических состояниях. При родах и при длительном стрессе резко уменьшается количество жировой ткани в организме. У лактирующих коров жировые клетки почти в 2 раза мельче, чем у телок. Неполноценное питание ведет к уменьшению количества жировой ткани прежде всего
под кожей, между мышцами, внутри мошонки и сальников. При этом меняется и химический состав липоцитов. При голодании дольше всего сохраняется жир в жировых подушках глаза, около почек, сердца. Отмечено, что
умеренное кормление в период интенсивного роста жировой ткани (для телят
это возраст от 7 до 12 мес) благоприятно сказывается на их росте и мясной
продуктивности, так как подавление роста жировой ткани не отражается на
росте мускулатуры. Замена молока в подсосный период обратом, сочными и
зелеными кормами приводит к перераспределению жировой ткани в организме. Она в большей степени развивается на внутренних органах в сальниках и в меньшей — под кожей и между мышцами.
Кроме обычной, так называемой белой жировой ткани, у новорожденных, грызунов и зимнеспящих животных имеется бурая жировая
ткань. Она расположена обычно на шее, вдоль позвоночника, за грудиной, в
области лопаток и между мышцами. Состоит из небольших округлых клеток,
в цитоплазме которых встречается несколько (5—15) мелких жировых вакуолей, ядро расположено центрально. В цитоплазме клеток между жировыми вакуолями много митохондрий, которые содержат много ферментов для
окисления липидов и мало — для синтеза АТФ, поэтому образующаяся при
окислении жиров энергия превращается в тепло и служит для поддержания
температуры тела в первые дни жизни и после пробуждения от спячки.
ПЛОТНАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Основную массу плотной соединительной ткани составляют волокна
межклеточного вещества, плотно прилегающие друг к другу. Клеток и основного аморфного вещества мало. Расположение волокон зависит от направления действия сил натяжения. Если силы действуют в одном направлении, то волокна располагаются параллельно друг другу в направлении действия сил, образуя плотную оформленную соединительную ткань. Если действующие силы разнонаправлены, волокна образуют сложные системы и сети.
Такая ткань называется плотной неоформленной соединительной тканью.
Плотная соединительная ткань выполняет опорную (механическую) функцию. Рост и восстановление плотной соединительной ткани происходят за
счет элементов рыхлой соединительной ткани, которая всегда есть вокруг сосудов и нервов.
Плотная неоформленная волокнистая соединительная ткань состоит из
большого количества коллагеновых волокон, объединенных в пучки. Последние идут в различных направлениях, образуя сложную трехмерную систему. Между ними лежат фиброциты, встречаются эластические волокна и
прослойки рыхлой соединительной ткани с проходящими в них сосудами и
нервами. Между рыхлой и плотной неоформленной соединительными тканями нельзя провести четкую границу, так как соотношение клеток и межклеточного вещества меняется постепенно, оно различное в разных участках
тела. Эта ткань образует сетчатый слой дермы, встречается в надхрящнице,
надкостнице и других местах.
Плотная оформленная соединительная ткань встречается в двух вариантах: фиброзной (коллагеновой) и эластической.
Плотная оформленная фиброзная (коллагеновая) соединительная ткань
(рис. 27) образована параллельно лежащими пучками коллагеновых волокон.
Друг от друга пучки отделены рядами фиброцитов. Сильно сдавленные волокнами фиброциты имеют длинные плоские отростки, проникающие между
пучками. Пучок коллагеновых волокон, отграниченный фиброцитами, называется пучком I порядка. Прослойки рыхлой соединительной ткани пронизывают плотную оформленную фиброзную ткань и, окутывая группы пучков
I порядка, формируют пучки II порядка.
1
2
3
4
Рис. 27. Плотная оформленная фиброзная
соединительная ткань (продольный срез
через сухожилие):
1 — пучки коллагеновых волокон; 2—
фиброциты между пучками I порядка; 3 —
прослойка рыхлой соединительной ткани (эндотеноний); 4 — пучок II порядка.
Более широкие прослойки рыхлой соединительной ткани окутывают по
нескольку пучков II порядка, формируя пучки III порядка. В крупных сухожилиях могут быть и пучки IV порядка. В прослойках рыхлой соединительной ткани проходят сосуды и нервы. Из плотной оформленной фиброзной
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
соединительной ткани построены сухожилия, связки, она встречается в фасциях, капсулах органов и других образованиях.
Плотная оформленная эластическая соединительная ткань образована
плотно лежащими толстыми эластическими волокнами с примесью коллагеновых волокон, разделенными фиброцитами и тонкими прослойками рыхлой
соединительной ткани. Пучкового строения эта ткань не имеет, так как эластические волокна идут не прямолинейно, а анастомозируют друг с другом.
Этой тканью образованы выйная и истинные голосовые связки, она встречается в стенках крупных артерий.
ХРЯЩЕВАЯ ТКАНЬ
Хрящевая ткань специализированная, выполняет опорную функцию и
принимает участие в углеводном обмене. Она сочетает в себе прочность, упругость и пластичность, рассчитана на сопротивление давлению, сжатию. В
ней содержится до 60—80°/о воды, 10—15% органических веществ и 4—7%
минеральных солей. Межклеточное вещество по массе преобладает над клетками. Клетки — хондробласты и хондроциты — продуцируют межклеточное вещество. В состав межклеточного вещества входят волокна и основное аморфное вещество — хондромукоид, образованный комплексами
хондроитинсульфатов с белками (сульфатированными гликозаминогликанами, протеогликанами и гликопротеидами). Он во многом определяет свойства хряща и, несмотря на большое содержание воды, делает его вязким и
плотным, связывая воду, придает тургор.
Рис. 28. Гиалиновый хрящ:
1 — надхрящница; 2— кровеносный сосуд; 3 — хондробласты; 4—хондроциты;
5 — изогенная группа; 6 — межклеточное вещество; 7 — оксифильная и 8 —
базофильная зоны.
Хрящ покрыт надхрящницей, состоящей из плотной соединительной
ткани с большим количеством сосудов и нервов. Недифференцированные
клеточные элементы надхрящницы дифференцируются в молодые клетки
хряща — хондробласты. Рост хряща происходит как с периферии за счет
дифференцировки и размножения клеток надхрящницы (аппозиционный
рост), так и в участках молодого хряща за счет размножения клеток и увеличения количества межклеточного вещества (интерстициальный рост). В
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
толще хряща отсутствуют сосуды, его питание осуществляется диффузно со
стороны надхрящницы либо от сосудов подлежащей кости. Развивается хрящевая ткань из мезенхимы в области склеротомов сомитов мезедормы. В связи с особенностями строения и функции различают три вида хрящевой ткани:
гиалиновую, эластическую и волокнистую.
Гиалиновая (стекловидная) хрящевая ткань (рис. 28) молочно-белого
или голубоватого цвета, полупрозрачная, наиболее твердая, упругая и самая
распространенная. Из нее состоит большая часть скелета плода, концы ребер,
носовая перегородка. Она покрывает суставные поверхности костей, имеется
в стенках трахеи, бронхов, гортани. По периферии гиалиновый хрящ покрыт
надхрящницей, клетки которой превращаются в хондробласты, находящиеся
под надхрящницей поодиночке параллельно поверхности хряща. Это удлиненные клетки с хорошо развитыми органелла-ми. Они продуцируют межклеточное вещество. В процессе функционирования все больше расходятся
друг от друга и перемещаются в глубь хряща, оттесняемые вновь нарастающими клетками. Размеры их увеличиваются, они приобретают овальную форму, цитоплазма светлеет. В ней обнаруживаются сильно развитые цистерны
шероховатой цитоплазматической сети, комплекс Гольджи и большое количество митохондрий, гликогена, воды и жировые включения. Молодые хондроциты промежуточной зоны хряща (между поверхностной и глубокой зонами) наиболее активно вырабатывают межклеточное вещество и сохраняют
способность к делению. В глубоких участках хряща у хондроцитов низкая
синтетическая и митотичеокая активность, но еще сохраняется способность к
делению амитозом. Разделившись, такие клетки не расходятся далеко друг от
друга, а образуют изогенную группу (одинаковую по происхождению) из 2—5
клеток. У зрелых хондроцитов уменьшается количество органелл, размеры
ядра, накапливаются липиды. Они приобретают угловатую форму. Изменяется и химический состав образуемого ими межклеточного вещества, в результате чего каждая изогенная группа оказывается как бы отделена от окружающих структур оксифильной и базофильной зонами.
Межклеточное вещество гиалинового хряща имеет однородный вид,
однако состоит не только из аморфного основного вещества, но и из коллагеновых волокон. Последние не образуют пучков, располагаются беспорядочно, имеют коэффициент преломления света, близкий к аморфному веществу,
и поэтому не выделяются из общей массы межклеточного вещества.
Эластическая хрящевая ткань непрозрачная, желтого цвета. Содержится
в некоторых хрящах гортани (надгортаннике, черпаловидных), в ушной раковине и наружном слуховом проходе. По структуре она подобна гиалиновому хрящу, лишь в межклеточном веществе наряду с коллагеновыми имеется много толстых эластических волокон, хорошо заметных в световой микроскоп в виде грубой сети, оплетающей клетки и направленных от поверхностных слоев хряща вглубь. Изогенные группы более мелкие (обычно из двух
клеток), расположены чаще, чем в гиалиновом хряще.
Волокнистая хрящевая ткань имеется в межпозвоночных дисках, симфизе лонных костей, круглой связке бедра и в местах прикрепления мышц к
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
костям. Для волокнистого хряща характерно сильное развитие коллагеновых
волокон, которые объединяются в пучки и располагаются, как и в плотной
оформленной фиброзной соединительной ткани, по направлению действия
сил. Пучки коллагеновых волокон хорошо видны в микроскоп, за что хрящ и
получил свое название. Между ними рядами лежат хондробласты.
КОСТНАЯ ТКАНЬ
Костная ткань возникла в филогенезе позже других тканей. Она имеется
только у позвоночных животных (кроме хрящевых рыб). Наилучшим образом приспособлена к выполнению опорной функции, сочетая в себе прочность и легкость. Образуя скелет у взрослого животного, она защищает органы, расположенные в голове, грудной и тазовой полостях и костный мозг, заполняющий полости костей, принимает активное участие в минеральном обмене.
В костной ткани содержится 10—30% воды, 20—50% органических веществ, 40—70% минеральных компонентов. 95% органических веществ составляет белок коллаген, остальное — гликопротеины, липиды и гликозаминогликаны (хондроитинсульфат, кератансульфат и другие глюкозамины и галактозамины). Костная ткань — основное депо минеральных солей. В ней
концентрируется 98% всех неорганических веществ, содержащихся в организме, в том числе 99% Са, 87% Р, 58% Mg, 46% Na. Обнаружено свыше 30
микроэлементов (Mg, Сu, Sr, Zn, Ва, Al, Si, F и др.), которые имеют значение
для жизнедеятельности клеток костной ткани, для процессов ее оссификации
и декальцинации. В костной ткани много различных солей кальция: фосфаты,
карбонаты, хлориды, соединения Са с фтором и органическими кислотами.
Наиболее распространенные фосфаты кальция. Они могут быть как в виде
аморфного фосфорнокислого кальция (Са3(РО4)2), так и в виде кристаллических солей — костных апатитов, в основном гидроксиапатитов
Ca10(PO4)6×(ОН)2.
Минеральные вещества кости образуют упорядоченные конструкции с
ее органическими веществами и представляют собой систему с огромной поверхностью (у крупного рогатого скота 15000 м2), участвующую в поддержании ионного равновесия в организме.
Как все опорно-трофические ткани, костная ткань происходит из мезенхимы и состоит из клеток и межклеточного вещества.
Клетки костной ткани — остеобласты, остеоциты и остеокласты (рис.
29).
Остеобласты — молодые клетки костной ткани цилиндрической, призматической или угловатой формы диаметром 15— 20 мкм. Ядро крупное с
рыхлым хроматином, 1—2 ядрышками, лежит эксцентрично. Цитоплазма базофильная, в ней хорошо развит комплекс Гольджи, гранулярная цитоплазматическая сеть, много митохондрий, то есть органеллы, обеспечивающие
клетке высокую жизнедеятельность и активный синтез белка. Видны секреторные гранулы с электронно-плотным материалом. От тела клетки отходят
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
отростки, которыми она контактирует с другими клетками. В ней много РНК,
высокая активность ферментов, особенно щелочной фосфатазы, играющей
важную роль в минеральном обмене кости. Остеобласты продуцируют все
составные части межклеточного вещества и, видимо, способствуют его минерализации. К делению не способны. По мере созревания превращаются в
остеоциты. Обнаруживаются в местах образования кости в процессе ее развития или перестройки.
Рис. 29. Клетки костной ткани:
Схема строения остеобласта (I), остеоцита (II) и остеокласта (III) — А, Б,
В — то же, в световом микроскопе; 1 — ядро; 2 — комплекс Гольджи; 3—
гранулярная эндоплазматическая сеть; 4— митохондрии; 5 — отростки; 6
— лакуна; 7 — костный каналец; 8 — лизосома; 9— гофрированная каемка;
10— межклеточное вещество.
Остеоциты — зрелые клетки, конечный этап дифференцировки остеобластов, преобладающая клеточная форма. Они неправильно-овальной или
многоугольной формы с многочисленными отростками. Ядро крупное, темноокрашенное. Цитоплазма слабобазофильная, органелл мало, но много
электронно-плотных пузырьков. В процессе минерализации межклеточного
вещества клетки костной ткани оказываются в нем замурованными. Места,
где расположены их тела, называются лакунами, а их отростки — костными
канальцами. Остеоциты не способны продуцировать межклеточное вещество,
но активно поддерживают жизнедеятельность кости, регулируют солевой состав межклеточного вещества. С их помощью в костной ткани совершается
обмен веществ. По костным канальцам, которые связывают клетки друг с
другом и достигают сосудов, к клеткам доставляются питательные вещества.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Остеоциты живут от нескольких месяцев до нескольких лет, затем погибают.
Участок костной ткани с погибшими остеоцитами, по-видимому, воспринимается организмом как чужеродный и подвергается уничтожению с помощью остеокластов.
Остеокласты — крупные клетки диаметром 90—100 мкм, содержащие
от трех до нескольких десятков мелких ядер. В цитоплазме развит комплекс
Гольджи, много митохондрий и лизосом с гидролитическими ферментами.
На той стороне клетки, которой она примыкает к разрушаемой костной ткани, у нее имеются многочисленные цитоплазматические выросты, формирующие гофрированную каемку, а в цитоплазме много вакуолей, из которых
в межклеточные пространства выделяются вещества, разрушающие кость.
Ядра скапливаются на противоположном конце клетки. Остеокласты обнаруживаются только в местах разрушающегося хряща или кости при активной
ее перестройке или развитии. Считается, что они происходят из моноцитов
крови путем их слияния и являются фагоцитами костной ткани.
Межклеточное вещество костной ткани состоит из коллагеновых волокон и основного аморфного вещества, пропитанных минеральными солями.
По количеству оно значительно преобладает над клетками и придает ткани
прочность и упругость. Коллагеновые волокна пропитаны кристаллами костного апатита и поэтому называются оссеиновыми. Между молекулами основного вещества располагаются молекулы аморфного фосфата кальция и все
вместе называется оссеомукоид. Закономерное расположение оссеиновых волокон и кристаллов оссеомукоида обусловливает жесткость кости и сопротивляемость силам сжатия, растяжения, кручения, изгиба. В зависимости от
характера расположения волокон различают грубоволокнистую, пластинчатую и дентиноидную костную ткань.
Дентиноидная костная ткань образует дентин зуба, и мы ее рассмотрим
при изучении строения зубов.
Грубоволокнистая костная ткань характеризуется неупорядоченным
расположением грубых оссеиновых волокон, формирующих толстые пучки,
видимые в световой микроскоп между разбросанными в беспорядке клетками. Этой тканью образован скелет плода и новорожденного животного. В течение всей жизни она сохраняется в швах между костями черепа, в местах
прикрепления сухожилий и связок к костям, в цементе зуба.
Пластинчатая костная ткань (рис. 30) характеризуется упорядоченным
параллельным расположением тонких оссеиновых волокон (толщина 10—
150 нм), сцементированных аморфным основным веществом и плотно упакованных в виде костных пластинок. Между пластинками упорядоченными рядами залегают остеоциты. Эта ткань в раннем постнатальном периоде замещает грубоволокнистую ткань в скелете животного. Костные пластинки образуют упорядоченные структуры разной формы. Наиболее распространенной являются остеоны.
Остеон — цилиндрическая структура, образованная концентрическими
костными пластинками, вложенными друг в друга. Между ними в лакунах
лежат остеоциты. Пластинки образованы межклеточным веществом с больwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
шим количеством тонких коллагеновых волокон. Направление волокон в соседних костных пластинках взаимно перпендикулярное, что увеличивает
прочность ткани. Пластинки пронизаны костными канальцами, по которым
осуществляется связь между остеоцитами. В середине— в канале остеона—
проходит сосуд, окруженный небольшим количеством недифференцированных клеточных элементов, таких, как перициты и др. Обычно диаметр остеона не пре вышает 200 мкм, так как именно на такую глубину могут диффузно распространяться вещества по костным канальцам. По данным разных
авторов, у взрослой коровы диаметр остеонов 100—140 мкм, у свиньи—
150—170, у овцы— 120—140 мкм. Каждый остеон состоит из 3—8 костных
пластинок и имеет канал диаметром 20—30 мкм.
Другой структурой являются вставочные пластинки. Это остатки старых разрушенных остеонов. Они более минерализованы и на препарате выделяются, так же как и старые остеоны, их окраска светлее молодых остеонов. По структуре от остеонов они отличаются тем, что не имеют центрального канала с проходящим в нем кровеносным сосудом.
У копытных встречаются циркулярно-параллельные структуры —
сложные костные образования, включающие несколько сосудистых каналов,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
окруженных костными пластинками. И, наконец, генеральные пластинки —
это костные пластинки, разделенные остеоцитами, идущие вокруг всей кости.
Развитие костной ткани. Костная ткань развивается из мезенхимы в
области склеротомов сомитов мезодермы. Клетки области склеротомов активно делятся, образуя уплотненный скелетогенный мезенхимный зачаток.
Из него могут развиваться и хрящ, и кость.
Если костная ткань развивается непосредственно из мезенхимы, говорят
о перепончатом остеогенезе, в процессе которого образуются покровные
кости. Это большинство костей черепа и часть ключицы. При перепончатом
остеогенезе в скелетогенный зачаток прорастают сосуды, мезенхимные клетки активно делятся и дифференцируются в остеобласты, которые продуцируют межклеточное вещество, раздвигаются, замуровываются им и превращаются в остеоциты. Образуется грубоволокнистая костная ткань, которая
вскоре замещается пластинчатой.
Если костная ткань развивается из мезенхимы, но на месте ранее образовавшегося хряща, говорят о хрящевом остеогенезе. Так развивается основная
масса костей скелета. В этом случае в мезенхимном зачатке закладывается и
начинает развиваться гиалиновый хрящ (см. выше), который принимает форму будущей кости. Сосудов в нем нет, питание хряща диффузное. С развитием кровеносной системы зародыша вокруг и внутри надхрящницы разрастаются сосуды, она становится надкостницей, и ее клетки начинают дифференцироваться не в хондробласты, а в остеобласты. Остеобласты продуцируют межклеточное вещество и откладывают его поверх хрящевого зачатка
кости. Этот процесс называется перихондральным окостенением. В результате образуется костная манжетка, которая нарушает питание хряща и приводит к его разрушению. В очаг разрушающегося хряща проникают сосуды
вместе с мезенхимными и другими недифференцированными клетками.
Здесь они превращаются в костные клетки: остеокласты и остеобласты.
Остеокласты разрушают обызвествленный хрящ, в результате чего возникают лакуны, туннели и полости, а остеобласты по их стенкам слоями в
виде костных пластинок откладывают межклеточное вещество, замуровывая
в нем более ранние генерации остеобластов. Этот процесс называется эндохондральным окостенением. В результате его образуется пластинчатая костная ткань, замещающая собой разрушенный гиалиновый хрящ.
Возрастные изменения костной ткани и ее реакция на различные
воздействия. В течение внутриутробного развития идет замещение хрящевых зачатков костной тканью, формирование, нарастание и перестройка остеонов и других структур кости. В раннем постнатальном периоде продолжает увеличиваться диаметр остеонов, нарастает число костных пластинок в остеоне от 2—4 (у новорожденных) до 4—10, уменьшается диаметр канала остеона с 50 до 20 мкм. В дальнейшем размеры остеонов остаются постоянными, в то время как диаметр канала остеона уменьшается на протяжении всей
жизни. Это приводит к увеличению количества остеоновна единицу площади
кости: с 16 шт. на 1 ммв при рождении до 34 шт. — у взрослых овец. В течение жизни повышается (в 5 раз) степень минерализации костной ткани.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У взрослых животных остеоны на одном и том же участке кости многократно перестраиваются, что сопровождается обменом минеральных веществ, входящих в состав ее структур.
Развитие и рост костной ткани регулируются эндокринной системой.
Гормон паращитовидной железы (паратирин) способствует активации деятельности остеобластов. Гормон роста гипофиза (соматотропин) усиливает
синтез белков в костных клетках, а следовательно, и их жизнедеятельность.
Структура костной ткани связана с типом конституции животного и направленностью его продуктивности. Так, дикий баран архар отличается от
мериносов более мелкими, тесно расположенными остеонами с узкими каналами. Остеоны у шерстных овец крупнее, с широкими каналами и расположены реже, чем у овец других типов. Плотнее и мельче всех остеоны у мясных овец. У коров молочного и молочно-мясного типов остеоны более крупные, у них более широкие каналы, но расположены плотнее, чем у коров
мясного и мясо-молочного типов. У помесных животных, как правило, процессы развития костной ткани протекают активнее, чем у их чистопородных
сверстников, то же — у одинцов по сравнению с двойнями.
Содержание и кормление отражается на процессах роста, перестройке и
структуре костной ткани. При стойловом содержании у животных костные
пластинки неравномерной толщины, остеоны расположены более рыхло по
сравнению с животными выгульного и пастбищного содержания. Улучшенное кормление беременных животных и молодняка приводит к более интенсивному росту костной ткани у последних, увеличению размеров остеонов
при их более плотном (на 25—30%) расположении и лучшем кровоснабжении. Недокорм в раннем возрасте задерживает рост костной ткани. Недостаток витаминов D тормозит всасывание Са, перестройку кости и ее рост, задерживает резорбцию хряща. Избыток витамина D также тормозит перестройку кости, но резко ускоряет резорбцию хряща и нарастание костной
ткани. Недостаток витамина С снижает синтез коллагеновых волокон, а отсюда и рост костной ткани. Избыток витамина А усиливает деятельность остеокластов, что приводит к деструкции костной ткани. При выращивании в
подсосный период на рационе с уменьшенным количеством молока, но с
преобладанием грубых и сочных кормов над концентратами у телят и поросят образуется больше рядов остеонов и костных пластинок.
Добавление к рациону молодняка макро- и микроэлементов способствует минерализации костной ткани, активной ее перестройке. Несбалансированный по минеральному составу рацион вызывает нарушения костной ткани, особенно у молодых и беременных животных. Недостаток Са и Р
может вызвать у плодов и молодняка полнее прекращение минерализации
костной ткани.
Вопросы для самоконтроля. 1. Каковы основные признаки, функции и
классификация опорно-трофических тканей? 2. Как характер межклеточного
вещества влияет на структуру и функции опорно-трофических тканей? 3. Каково происхождение, строение и значение мезенхимы? 4. В чем особенности
структуры и функции эндотелия? 5. Классификация, строение и функции
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
клеток крови. 6. Чем лимфа отличается от крови? 7. Каково происхождение,
строение, распространение в организме и функции рыхлой соединительной
ткани? 8. Какие виды клеток и межклеточного вещества встречаются в соединительной ткани? Их строение и функции. 9. Какие виды жировой ткани
вы знаете и чем отличаются друг от друга? 10. Виды хрящевой ткани, их
строение, распространение и различия? 11. Развитие, строение и перестройка
пластинчатой костной ткани. Где она находится?
Глава 9. МЫШЕЧНЫЕ ТКАНИ
К этому типу относятся ткани, основным свойством которых является
способность к сокращению. Они обеспечивают движение животных в пространстве и двигательные функции органов (сердца, кишечника, мочеточников и др.). Мышечные ткани различают по происхождению, строению, особенностям функционирования. Общее у них — сходная форма эмбриональных клеток— миобластои и наличие большого количества специальных органелл — миофибрилл. В их состав входят распространенные в животных
клетках сократительные белки (актин и миозин). Последние участвуют в
движении клетки (лейкоциты) и ее частей (хвост спермия, отростки миоэпителиальных клеток, реснички эпителия), однако только в мышечных тканях
эти белки образуют специфический актомиозиновый комплекс, наилучшим
образом осуществляющий функцию движения. Мышечные ткани как более
специализированные в филогенезе возникли позже эпителиальных и соединительных. Они способны к возбуждению, вызывающему их сокращение.
Кроме собственно мышечных структур, в состав мышечных тканей всегда входят нервные и соединительнотканные элементы. На основании особенностей строения и функции мышечные ткани позвоночных делят на гладкую и поперечнополосатые— скелетную и сердечную.
ГЛАДКАЯ (НЕИСЧЕРЧЕННАЯ) МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань образует мышечную оболочку трубкообразных органов пищеварения (кроме глотки и части пищевода), дыхания, выделения, размножения, находится в стенках кровеносных сосудов,
протоков желез, в селезенке, коже, ресничном теле глаза и в других органах.
Основная структурная единица гладкой мышечной ткани — клетка —
гладкий миоцит. Это сильно вытянутая веретеновидная, иногда отростчатая
(в мочевом пузыре, в эндокарде) клетка длиной 20—500 мкм (в матке беременных животных) и шириной 6—20 мкм. Вытянутое ядро находится в средней, утолщенной ее части. Здесь же, вокруг ядра, располагаются общие органеллы: митохондрии, комплекс Гольджи, центросома, рибосомы, эндоплазматическая сеть и включения гликогена. По всей клетке разбросаны протофибриллы — нитчатые структуры, образованные актином или миозином. Актиновые протофибриллы тоньше миозиновых и их в несколько раз (в 3—24
раза) больше. В расслабленной клетке протофибриллы ориентированы в осwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
новном продольно, но не образуют миофибриллярных комплексов. При сокращении образуется миофибриллярный актомиозиновый комплекс, приводящий к укорочению клетки.
Оболочка гладких миоцитов двухслойная. Она состоит из плазмолеммы
обычного строения и базальной мембраны (пластинки) углеводно-белковой
природы, выполняющей опорную функцию. Между ними имеется пространство шириной 15— 20 нм.
Функциональная единица гладкой мышечной ткани — пучок из 10—15
миоцитов, связанных с нервным волокном. В пучке мышечные клетки тесно
связаны между собой с помощью десмосом, плотных и щелевых контактов.
Это позволяет всем клеткам пучка одновременно реагировать на нервное
раздражение, несмотря на то, что нервное окончание имеется лишь на одной
из его клеток. Между мышечными клетками даже внутри пучка залегают
тонкие коллагеновые и эластические волокна. Коллагеновые волокна оплетают миоциты, вплетаются в базальную пластинку, тем самым удерживая
клетки от чрезмерного растяжения или сжатия. Пучки клеток отделены друг
от друга прослойками соединительной ткани, в которой проходят сосуды и
нервы.
Гладкая мышечная ткань иннервируется вегетативной нервной системой. Регуляция ее деятельности контролируется корой полушарий, но без
участия сознания, сокращения осуществляются непроизвольно. Сокращается
гладкая мышечная ткань медленно, ритмично. Период одного сокращения
длится от 3 с до 5 мин. Может длительное время находиться в состоянии сокращения без заметного утомления. Такой характер сокращения называется
тоническим.
Происходит гладкая мышечная ткань из мезенхимы, клетки которой,
дифференцируясь в гладкие миоциты, удлиняются и прилегают плотно друг
к другу. Гладкие миоциты сохраняют способность к делению. Кроме того, на
протяжении всего онтогенеза гладкие миоциты могут образовываться из недифференцированных клеток соединительной ткани, сопровождающих кровеносные сосуды.
ПОПЕРЕЧНОПОЛОСАТАЯ (ИСЧЕРЧЕННАЯ) СКЕЛЕТНАЯ
МЫШЕЧНАЯ ТКАНЬ
Из данной ткани состоит скелетная мускулатура, мышцы языка, гортани,
глотки, передней части пищевода, диафрагмы, глазного яблока, уха и некоторых других органов. Основная функция этой ткани — движение животного в
пространстве и движение отдельных частей тела. Кроме того, она является
основным источником тепла в организме, так как только 30—40% энергии
мышечного сокращения используется в виде механической работы (движение), остальное превращается в тепловую энергию.
Структурная и функциональная единица поперечнополосатой скелетной
мышечной ткани — мышечное волокно (рис. 31).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 31. Строение поперечнополосатой мышечной ткани:
А — гистосрез; Б — схема; В — электроннограмма участка мышечного волокна; I — мышца; II — пучок мышечных волокон; III — мышечное волокно;
IV— миофибрилла; V — саркомер; VI — актомиозиновый комплекс:1 — эндомизий В; J — пернмизий; S — сарколемма; 4 — саркоплазма; 5 — ядро; 6 —
И-диск; 7 — А-диск; 8 — Z-полоска (телофрагма); 9 — М-полоска (мезофрагма); 10 — саркомер; 11 — протофибрилла из молекул миозина; 12 —
протофибрилла из молекул актина.
Это крупное симпластическое образование цилиндрической формы шириной 15—150 мкм (у крупного рогатого скота 16—95 мкм, у свиньи 20—90,
у овцы 20—50, у лошади 50—80 мкм) и длиной от 1—2 мм до 10—15 см, с
закругленными концами. Толщина волокон, возможно, ограничивается условиями их питания. Несмотря на то, что кровеносные капилляры прилегают к
волокнам очень плотно, центральные участки слишком толстых волокон могут страдать от недостатка трофических и энергетических веществ. Оболочка
волокна называется сарколеммой (sarcos — мясо), а содержимое — саркоплазмой. В волокне имеется от нескольких десятков до нескольких сотен
ядер, которые у млекопитающих располагаются под сарколеммой. Ядра могут быть округлой, овальной и удлиненной формы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Форма ядер зависит от возраста животного (у молодых животных ядра
более округлые), степени сокращения волокна (в сокращенных волокнах ядра
короче), от характера работы мышц. Если мышца с ярко выраженной динамической функцией, ее ядра более округлой конфигурации и лежат скоплениями. У мышцы со статической функцией ядра удлиненные, лежат цепочкой. В световом микроскопе каждое мышечное волокно выглядит исчерченным поперечными чередующимися светлыми и темными полосами, за что
ткань получила свое название.
Мышечные волокна объединяются в пучки I порядка с помощью тонких
прослоек рыхлой соединительной ткани (эндомизий). В эндомизии эластические волокна преобладают над коллагеновыми. В нем проходят кровеносные
и лимфатические капилляры и нервные волокна. На одно мышечное волокно,
как правило, приходится один капилляр, идущий вдоль волокна.
Пучки I порядка объединяются соединительной тканью в пучки II порядка, а те — в пучки III порядка. Эти прослойки соединительной ткани называют перимизием, а соединительную ткань, одевающую мышцу сверху, — эпимизием. В перимизии и эпимизии проходят крупные эластические и коллагеновые волокна, встречаются скопления жировых клеток. Мышечные волокна
очень прочно связаны с одевающим их соединительнотканным каркасом. В
местах соединения в мышечных волокнах образуются впячивания и складки,
куда вклиниваются коллагеновые волокна и вплетаются там в сарколемму
мышечных волокон.
Субмикроскопическое строение мышечного волокна. Сарколемма
мышечного волокна состоит из плазмолеммы толщиной 7—10 нм и базальной мембраны (пластинки) толщиной 15— 50 нм. Плазмолемма через равные
промежутки образует глубокие впячивания в виде трубок диаметром 4 нм (у
свиньи)„ идущих поперек волокна и названных Т-каналами. Они играют
большую роль в обмене веществ и проведения нервного импульса. Базальная
мембрана (пластинка) выполняет опорную функцию и принимает участие в
поддержании гомеостаза волокна. С ней связаны коллагеновые и эластические волокна, оплетающие вдоль и поперек мышечное волокно, образующие
его внешний каркас.
В ядрах хроматин в виде глыбок. Во время сокращения ядра могут принимать штопорообразную форму, в них видны глубокие впячивания кариолеммы. В саркоплазме под сарколеммой вокруг ядер локализуются элементы
гранулярной эндоплазматической сети, пластинчатого комплекса. В ней же
находится пигмент миоглобин, который (как и гемоглобин в крови) связывает и депонирует кислород, поступающий в волокно, и отдает его при работе
мышцы. Чем больше миоглобина в саркоплазме, тем больший резерв кислорода в мышце. Значительная часть саркоплазмы занята миофибриллами —
специальными органеллами, ответственными за сокращение волокна. Вокруг
них лежат митохондрии (саркосомы), гладкая эндоплазматическая (саркоплазматическая) сеть, включения гликогена.
Миофибрилла представляет собой нитчатую структуру с диаметром
0,5—2 мкм и длиной, соответствующей длине волокна, построенную из упоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рядочение расположенных белков актина, миозина, тропомиозина, тропонина, актинина. По всей длине миофибриллы чередуются участки из белков актина и миозина. Миозин обладает двойным лучепреломлением (анизотропией), поэтому участки миофибриллы, построенные из этого белка, названы
анизотропными дисками (А-диски). В световом микроскопе при проходящем
свете они выглядят темными. Участки, построенные из актина, не обладают
двойным лучепреломлением— это изотропные диски (И-диски). Они под
микроскопом светлые. А-диски соседних миофибрилл волокна (так же, как и
И-диски) расположены на одном уровне, в результате чего они воспринимаются как одна полоса, светлая или темная. От этого все волокно кажется поперечно-исчерченным чередующимися темными и светлыми полосами.
Электронная микроскопия показала, что диски миофибрилл состоят из
параллельно лежащих нитей, более толстых в А-диске и более тонких в Идиске. Нити эти названы протофибриллами. Тонкие протофибриллы построены из глобулярных молекул белка актина. Молекулы актина образуют
цепочку, подобную двойной нитке бус, скрученной в виде веревочки, длиной
1 мкм и шириной 5—7 нм. Толстые протофибриллы построены ив молекул
фибриллярного белка миозина, уложенных в виде пучка длиной 1,5—2 мкм и
толщиной 10—25 нм.
Середину каждого И-диска пересекает темная Z-полоска, или телофрагма, а в средней части А-диска имеется светлая Н-зона, в центре которой проходит М-полоска, или мезофрагма. Z- и М-полоски имеют сложное, не до
конца выясненное строение и выполняют опорную функцию, способствуя закреплению протофибрилл в таком положении, при котором каждая толстая
про-тофибрилла оказывается окруженной шестью тонкими, а вокруг каждой
тонкой лежат три толстые протофибриллы.
Тонкие протофибриллы одним своим концом прикреплены в области Zполоски, другим — заходят в А-диск между толстыми протофибриллами,
вплоть до Н-зоны. Следовательно, краевые участки А-диска образованы как
миозиновыми, так и актиновыми протофибриллами, где они перекрывают
друг друга, образуя зону перекрытия. Лишь в Н-зоне нет актиновых протофибрилл.
При мацерапии волокна или воздействием на него трипсином его миофибриллы распадаются по Z-полоскам на отдельные кусочки, названные саркомерами. Следовательно, саркомер — элементарная структурная единица
миофибриллы. Его границами являются две соседние Z-полоски. Состоит он
из половины И-диска, целого А-диска и половины другого И-диска, длина
его 2,5—4 мкм. При сокращении происходит взаимное скольжение протофибрилл. Тонкие протофибриллы еще глубже втягиваются между толстыми,
И-диск и Н-зона становятся уже. Длина саркомера уменьшается на 20—40%.
При сильном сокращении И-диск и Н-зона исчезает совсем. Вся миофибрилла становится темной. Такую картину можно наблюдать в узлах сокращения
— местах наибольшего сокращения волокна.
Во время скольжения происходит перестройка (конформационные реакции) всех белков, входящих в состав миофибриллы, и между протофибрилwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лами возникают временные связи в виде мостиков, которые позволяют удерживать миофибриллы в их новом положении — образуется актомиозиновый
комплекс. Процессы эти протекают со значительной затратой энергии и при
обязательном присутствии ионов Са++ и Mg—Са — АТФазы.
По наиболее обоснованной теории сокращения, предложенной Р. Дэвисом, которая учитывает процессы превращения энергии, данные электронной
микроскопии и рентгеноструктурного анализа, мышечные волокна сокращаются следующим образом. Нервное возбуждение через двигательное нервное
окончание достигает сарколеммы. При этом происходит деполяризация мембраны волокна, которая с помощью Т-каналов быстро распространяется по
всему волокну и приводит к изменению направления ионных потоков — возникает ток действия. Т-каналы тесно связаны с цистернами саркоплазматической сети. На уровне границы А- и И-дисков (у насекомых, рептилий и млекопитающих) или Z-полосок (у амфибий) они образуют характерные комплексы, названные триадами. Под влиянием тока действия из цистерн саркоплазматической сети в гиалоплазму выходят ионы Са++, и когда их концентрация поднимается выше пороговой величины, Са включает механизм
перемещения протофибрилл друг относительно друга. Сокращение идет с затратой энергии АТФ до тех пор, пока длится потенциал действия и в гиалоплазме достаточно много ионов Са++. Чем их больше, тем интенсивнее сокращаются миофибриллы. При исчезновении потенциала действия Са++ устремляются из гиалоплазмы в сарко-плазматическую сеть и мышечное волокно расслабляется.
Энергия для работы мышечного волокна образуется в процессе гликолиза и окислительного фосфорилирования. В зависимости от того, какой источник используется, в волокне будут преобладать включения гликогена или
миоглобина. Волокна, содержащие большое количество миоглобина, имеют
более красную окраску (в нефиксированной ткани) и названы красными. Волокна, в которых миоглобина меньше, имеют более светлый цвет и названы
белыми. Между этими крайними типами есть промежуточные формы. Волокна отличаются друг от друга не только количеством миоглобина, но и
структурно-функциональными особенностями. Как правило, каждая мышца
содержит и белые, и красные, и промежуточные волокна. Ее свойства, хотя и
зависят от того, какой тип волокон в ней преобладает, будут складываться не
просто из суммы свойств составляющих ее волокон, но и зависеть от их
взаимодействия в процессе работы.
Развитие поперечнополосатой скелетной мышечной ткани.
Данная ткань развивается из миотомов сомитов мезодермы. Миотомы
состоят из плотно расположенных удлиненных клеток — миобластов. В период эмбриогенеза миобласты активно делятся, перемещаются в места закладки мускулатуры, выстраиваются в цепочки и сливаются друг с другом,
образуя мышечные трубочки. В процессе дальнейшей дифференцировки
происходит наращивание и совершенствование структур органелл и мышечные трубочки превращаются в мышечные волокна — дефинитивные симплаwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
стические структуры поперечнополосатой скелетной мышечной ткани. Часть
миобластов не проходит указанный путь, а сохраняется в виде удлиненных
одноядерных малодифференцированных клеток, плотно прилежащих к волокну и одетых общей с ним базальной пластинкой. Такие клетки называют
миосателлитами. Считают, что они могут быть источником образования новых волокон в течение всей жизни животного. Есть данные о том, что новые
волокна образуются путем продольного расщепления толстых волокон и увеличения количества миофибрилл в тонких волокнах.
Возрастные и другие изменения скелетной мышечной ткани
выражаются прежде всего в увеличении размеров волокон. Диаметр волокна увеличивается за счет прибавления протофибрилл по бокам миофибрилл. В длину волокно растет в результате присоединения протофибрилл по
концам миофибрилл. Увеличивается и количество миофибрилл в основном за
счет продольного расщепления. Процессы эти активно протекают как в эмбриональный, так и в постнатальный период. За это время мышечные волокна утолщаются в 2—4 раза. У крупного рогатого скота особенно активно растут мышцы от рождения до 6 и от 12 до 18 месяцев. С возрастом у всех животных увеличивается относительное содержание в скелетных мышцах белых волокон по сравнению с красными.
Пучки мышечных волокон у новорожденных животных плохо сформированы, так как прослойки рыхлой соединительной ткани имеют вид нежной
сети и не содержат жира. С ростом животного увеличивается количество
пучков благодаря их разделению в результате врастания соединительной
ткани. Мышечные пучки хорошо очерчены, лежат плотно. Прослойки перимизия тонкие, но более плотные, чем у новорожденных, в них больше становится коллагеновых волокон. Общее содержание соединительной ткани в
мышцах колеблется в широких пределах (от 5 до 20%) в зависимости от типа
мышцы. Старческие изменения в мышечной ткани выражаются в появлении
большого количества ядер в центре волокон, разрастании плотной соединительной ткани, истончении и нарастании дегенеративных изменений в мышечных волокнах, снижении васкуляризации.
Мышечная ткань у животных разных пород и направления продуктивности может отличаться толщиной волокон, степенью развития и плотностью
перимизия и эпимизия, хотя различия эти в основном незначительны. Все же
замечено, что у животных скороспелых пород и мясного типа мышечные волокна более толстые: в их межпучковой соединительной ткани чаще встречаются жировые клетки. От кастрации замедляется рост волокон, сохраняется рыхлость перимизия. Стойловое содержание приводит к увеличению диаметра мышечных волокон и разрастанию жира в перимизии.
Значительно меняется структура мышечной ткани под влиянием кормления. Сбалансированное оптимальное кормление способствует нормальному развитию мышечной ткани. При обильном кормлении и откорме утолщаются мышечные волокна, между ними появляется много жировых клеток.
Несбалансированное и неполноценное кормление несколько сдерживает рост
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мышечной ткани. Пагубно сказывается на ней недостаточное питание, особенно в период наиболее интенсивного роста: диаметр мышечных волокон в
этом случае меньше на 20—35%.
СЕРДЕЧНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ (ИСЧЕРЧЕННАЯ)
МЫШЕЧНАЯ ТКАНЬ
Сердечная ткань дифференцируется из висцерального листка спланхнотома мезодермы и образует основной слой стенки сердца— миокард. По
структуре и функции различают две ее разновидности: рабочую и проводящую.
Рабочая мышечная ткань по структуре и физиологическим свойствам
занимает как бы промежуточное положение между скелетной и гладкой. Сокращается она ритмично, медленнее скелетной мышечной ткани и мало
утомляется. Состоит из клеток — сердечных миоцитов, объединенных в сердечные волокна. Последние анастомозируют друг с другом так, что образуют
единую систему. Между волокнами имеются прослойки рыхлой соединительной ткани — эндомизий, в котором проходят сосуды и нервы.
Одеты сердечные волокна сарколеммой, подобной сарколемме скелетного мышечного волокна. Однако в отличие от него внутренний слой сарколеммы — плазмолемма — одевает каждую клетку отдельно. В местах соединения соседних клеток друг с другом их плазмолеммы образуют пальцевидные выпячивания и контакты типа десмосом, плотные и щелевые, а также
специализированные контакты на уровне Z-полосок, которые способствуют
функционированию сердечной мышцы как единого целого. Границы соседних миоцитов в волокнах представлены в виде вставочных (темных) полосок, идущих поперек волокна. С возрастом они утолщаются, особенно у
крупного рогатого скота и лошадей.
Сердечные 1миоциты — клетки цилиндрической формы длиной 50—120
мкм и шириной 15—20 мкм. Одно- два овальных ядра лежат в центре клетки.
Миофибриллы занимают ее периферию. Строение их такое же, как в скелетной мышечной ткани, а количество меньшее. Поэтому поперечная исчерченность сердечных миоцитов видна хуже, чем продольная.
Сердечная мышечная ткань темно-красного цвета, так как цитоплазма
сердечных миоцитов богата миоглобином. В ней много митохондрий и
включений гликогена. Следовательно, она получает энергию как в процессе
окислительного фосфорилирования, так и гликолиза. Благодаря мощной
энергетической оснащенности сердечная мышца работает непрерывно всю
жизнь. Замечено, что у животных, обитающих в высокогорных условиях,
увеличивается количество гликогена, митохондрии крупнее, с волнистыми
или сетчатыми кристами. Т-каналы в сердечных миоцитах расположены на
уровне Z-полосок, а не на границе дисков, как в скелетных мышечных волокнах. Саркоплазматическая сеть развита хуже, поэтому сердечная мышца
нуждается в постоянном притоке Са извне.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Строение сердечной ткани несколько различное у животных разных видов. Из домашних животных у лошади мышечные волокна уложены наиболее компактно, имеют лентовидную форму, боковые перемычки редки, эндомизий развит слабо, кровоснабжение обильное, миоциты узкие (10—21 мкм)
и длинные (110—130 мкм), с большим количеством миофибрилл, которые
часто лежат в центре клеток, оттесняя длинные узкие ядра на периферию.
Поперечная исчерченность хорошо видна. У рогатого скота волокна полигональные, миоциты короче и шире, боковые перемычки встречаются чаще, а
количество миофибрилл меньше, чем у лошади. Располагаются они по периферии миоцитов. У свиньи сетчатость сердечной мышечной ткани наиболее
выражена, волокна округлой формы, эндомизий хорошо развит, но капилляры встречаются реже, чем у лошади, миофибрилл меньше, поперечная исчерченность слабо выражена.
Своеобразие сердечной мышечной ткани состоит в том, что она, представляя собой по существу симпласт и сокращаясь как единое целое, в то же
время мало страдает при повреждении отдельных миоцитов. Сердечная мышечная ткань не имеет камбиальных элементов и на тренинг или травму отвечает физиологической гипертрофией миоцитов. Поврежденные миоциты
погибают и замещаются соединительной тканью.
Интенсивность и частота сокращений сердечной мышцы регулируются
нервными импульсами. Однако сердечная мышца обладает и собственной
системой регуляции движений. Правда, без регуляции извне частота сердечных сокращений уменьшается вдвое. Обеспечивается автоматизм сокращений проводящей мускулатурой, построенной из атипичных мышечных волокон (Пуркинье). Состоят они из крупных клеток с малым количеством миофибрилл и образуют проводящую систему сердца, которая делает согласованной сокращения предсердий и желудочков сердца, обеспечивает ритмичную смену рабочего акта (систолы и диастолы) восстановительным периодом
(расслабление сердечной мышцы).
Вопросы для самоконтроля. 1. Каково происхождение, строение, распространение, особенности функционирования гладкой мышечной ткани? 2.
Происхождение и строение поперечнополосатой скелетной мышечной ткани? 3. Строение мышечного волокна. 4. Что такое саркомер, каково его
строение и функция? 5. В чем особенности строения и функций сердечной
поперечнополосатой мышечной ткани?
Глава 10. НЕРВНАЯ ТКАНЬ
Нервная ткань высокоспециализированная, из нее построена вся нервная
система. В центральной нервной системе она образует серое и белое вещестwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
во головного и спинного мозга, в периферической— ганглии, нервы, нервные
окончания. Нервная ткань способна воспринимать раздражения из внешней и
внутренней среды, возбуждаться под их влиянием, вырабатывать, проводить
и передавать импульсы, организовывать ответные реакции. Сумма этих
свойств нервной ткани проявляется в основной функции нервной системы:
регуляции и координации деятельности различных тканей, органов и систем
организма.
Развивается нервная ткань из нейроэктодермы. Из нее образуется сначала нервная пластинка, а затем нервная трубка, вдоль которой с двух сторон лежат нервные гребни (валики). В нервной трубке и гребнях формируются все клетки нервной ткани. Строение нервной ткани в различных участках
нервной системы сильно различается. Тем не менее она везде состоит из нейронов и нейроглии. Между ними имеются межклеточные пространства, заполненные тканевой жидкостью. Межклеточные пространства мозга составляют 15—20% его объема. В тканевой жидкости происходит диффузия веществ между капиллярами и клетками нервной ткани. Нейроны — нервные
клетки, способные к выработке и проведению нервного импульса. Нейроглия
состоит из клеток, выполняющих вспомогательные функции.
Строение и виды нейронов. Нейрон (нейроцит) — основная структурная
и функциональная единица нервной ткани (рис. 32). В нем различают тело
— перикарион и отростки. Нейроны разных отделов нервной системы отличаются друг от друга по функции, форме, размерам, количеству и характеру
ветвления отростков, по выделяемому медиатору. По функции нейроны бывают чувствительные (рецепторные, или афферентные), двигательные (эффекторные, или эфферентные) и вставочные (ассоциативные). Размеры нейроцитов колеблются от 4 мкм у клеток-зерен мозжечка до 130 мкм (у гигантопирамидных клеток коры).
Нейроны в основном одноядерные клетки. Ядро крупное, округлое, расположено обычно в центре клетки. Кариоплазма светлая, так как хроматин не
образует крупных глыбок. Содержит 1—2 крупных ядрышка. Комплекс
Гольджи расположен вокруг ядра. Много митохондрий, микротрубочек, есть
центросома, лизосомы. Хорошо представлен аппарат синтеза белка: рибосомы и гранулярная цитоплазматическая сеть. Адсорбция основных красителей
на скоплениях этих органелл образует характерную картину в виде крупных
глыбок, напоминающую шкуру тигра (при изучении в световой микроскоп),
за что и названа тигроидным (базофильным) веществом или субстанцией
Ниссля (по имени описавшего ее гистолога). Есть и специальные орга-неллы
— нейрофиламенты. Пучки нейрофиламентов и микротрубочек (нейротубул)
благодаря адсорбции на них красителей видны в световой микроскоп в виде
нейрофибрилл. Эти органеллы участвуют в формировании цитоокелета, в передвижении веществ по клетке и ее отросткам.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 32. Схема строения нейрона:
А — на светооптическом и Б —
на
ультрамикроскопическом
уровне:
1 — перикарион; 2 — ядро; 3 —
ядрышко; 4 — дендриты; 5 —
аксон; 6 — конечные разветвления аксона; 7— комплекс Гольджи; 8 — гранулярная эндоплазматическая сеть; 9 — митохондрии; 10 — нейрофибриллы.
Форма перикариона во многом определяется количеством отростков.
Различают униполярные — с одним отростком, ложно-униполярные, биполярные — с двумя отростками и мультиполярные нейроны — с несколькими
(3—20) отростками. Тела униполярных и ложноуниполярных клеток округлые, биполярных — веретеновидные, мультиполярных — разнообразные.
Отростки — обязательная принадлежность нейронов. Без них нейроциты не
могут выполнять свои функции, так как отростки обеспечивают проведение
нервного импульса от одной части тела в другую. Их длина бывает от нескольких микрометров до 1—2 м. По морфологическим и функциональным
свойствам отростки неравнозначны. В нейроне различают дендриты и аксон
(нейрит). Аксон в клетке всегда один, дендритов может быть разное количество. По аксону возбуждение распространяется от тела, по дендриту— к телу
нервной клетки. Дендриты, как правило, сильно ветвятся и в них присутствуют все органеллы, которые есть и в теле клетки. Аксон не ветвится, но может отдавать коллатерали — ответвления, идущие параллельно. В нем нет
базофильного вещества. Нейрофиламенты и нейротрубочки располагаются
упорядочение — вдоль аксона. Униполярными считаются недифференцированные нервные клетки на ранней стадии развития, когда дендриты еще не
образовались. Среди дифференцированных клеток униполярные нейроны
встречаются редко.
От тела ложноуниполярного нейрона отходит один отросток, который Тобразно разветвляется на дендрит и нейрит. Такие клетки распространены в
спинномозговых узлах (ганглиях). Это чувствительные нейроны, дендриты
которых идут на периферию, где заканчиваются в органах чувствительными
нервными окончаниями (рецепторами), а нейриты несут возбуждение от тела
клетки в центральную нервную систему. Как видим, эти клетки по своим
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
структурно-функциональным свойствам приближаются к биполярным нейронам, которые встречаются в органе зрения, обоняния и среди ассоциативных
нейронов. Самыми распространенными являются мультиполярные нейроны.
Это все двигательные (моторные) и большинство ассоциативных нейронов.
Среди их отростков только один аксон, а остальные дендриты. У ассоциативных нейронов аксон не покидает центральной нервной системы, у двигательных — идет на периферию —к органам (мышцам, железам), где и оканчивается двигательным нервным окончанием.
Нервные клетки рано дифференцируются в онтогенезе, утрачивают способность к делению, в норме продолжительность их жизни равна продолжительности жизни особи. Для поддержания жизнедеятельности и способности
к выполнению функций на протяжении столь длительного времени в нейронах развита система внутриклеточной регенерации. При этом макромолекулы
и их ансамбли постоянно разрушаются и создаются вновь. Белковые синтезы
идут в основном в теле клетки. Высокий уровень жизнедеятельности отростков поддерживается постоянным таком цитоплазмы в отростки и обратно.
Плазмолемма нейрона выполняет все функции, присущие ей в любых
клетках. Кроме того, она способна к возбуждению при деполяризации (снижении величины заряда) в результате перемещения ионов Na+ в клетку. Деполяризация возникает локально (в одном месте) и волнообразно перемещается от дендрита к телу и аксону. С какой скоростью движется волна деполяризации, с такой же скоростью передается и нервный импульс. Торможение
наступает при противоположном явлении: увеличении заряда мембраны под
влиянием ионных потоков (О-— в клетку и К+ — из клетки). В нервной ткани
нейроны образуют ансамбли, характерные для определенных участков нервной системы. Характер их расположения носит название цитоархитектоника.
Рис. 33. Синапс:
1 — пресинаптический полюс; 2 —
синаптические пузырьки; 3— митохондрия; 4 — пресинаптическая
мембрана; 5 — синаптипеская щель;
6 — постсинаптический полюс; 7—
постсинаптическая мембрана.
Передача нервного импульса от одного нейрона к другому осуществляется в месте их контакта—синапсе (sinapsis — соединение) (рис. 33). В зависимости от того, какие участки нейронов вступают в контакт, различают аксодендритические (аксон одного нейрона контактирует с дендритом другого
нейрона), аксосоматические (аксон контактирует с телом другого нейрона) и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
аксоаксо-нальные (контактируют аксоны двух нейронов) синапсы. Описаны
также дендросоматические и дендродендритные синапсы. Примерно половина поверхности тела нейрона и почти вся поверхность его дендритов бывает занята синапсами.
В результате каждый нейрон имеет обширные контакты. Так, на одной
грушевидной клетке мозжечка насчитывают до 200 000 синапсов. Синапсы
бывают как возбудительные, так и тормозные.
У всех синапсов общие принципы строения: концевые веточки аксона,
передающего импульс нейрона в месте синапса, образуют колбовидные
утолщения — это пресинаптический полюс. В нем содержится много митохондрий и синаптических пузырьков, которые различаются по виду и размерам в зависимости от содержащегося в них медиатора — вещества, возбуждающего второй нейрон. Это могут быть серотонин, ацетилхоллин, адреналин и другие вещества. Участок второго нейрона, воспринимающий импульс, называется постсинаптическим полюсом. В нем нет синаптических
пузырьков и митохондрий. Между двумя полюсами находится узкая синаптическая щель (около 20 им), ограниченная контактирующими мембранами
двух полюсов: пресинаптической и постсинаптической. Эти мембраны
имеют утолщения и другие специальные структурные приспособления, обеспечивающие успешную передачу нервного импульса только в одном направлении. Нервный импульс, пришедший в пресинаптический полюс, приводит
к выбросу медиатора в синаптическую щель. Вызванный им нервный- импульс переходит на второй нейрон.
Нейроглия заполняет в нервной ткани все пространства между нейронами, их отростками, кровеносными капиллярами. Тесно прилегает к перечисленным структурам, образуя их оболочки. Она выполняет разнообразные
функции: опорную, изолирующую, разграничительную, трофическую, защитную, обменную, гомеостатическую. Нейроглиальные клетки — глиоциты
— называют вспомогательными клетками нервной ткани, так как они не проводят нервный импульс. Тем не менее их функции жизненно необходимы,
поскольку отсутствие или повреждение нейроглии делает невозможной работу нейронов. Существуют две разновидности нейроглии: макроглия и микроглия.
Макроглия (глиоциты), как и нейроны, развивается из клеток нервной
трубки. Среди глиоцитов различают: эпендимоциты, астроциты, олигодендроциты.
Эпендимоциты — глиальные клетки кубической или цилиндрической
формы, на их апикальном полюсе имеются реснички, от базального полюса
отходит длинный отросток, который пронизывает всю толщу мозга. Они
плотно прилегают друг к другу, выстилая сплошным слоем стенки желудочков мозга и спинномозгового канала. Движениями ресничек создается ток
цереброспинальной жидкости. В некоторых эпендимоцитах обнаруживают
секреторные гранулы. Предполагают, что эпендимоциты выделяют секрет в
цереброспинальную жидкость и регулируют ее состав.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Астроциты — основная разновидность глиоцитов центральной нервной
системы. Это клетки с диаметром тела 10—25 мкм, с округлыми или овальными ядрами, с многочисленными, расходящимися в разные стороны отростками. Различают плазматические и волокнистые астроциты. Плазматические
астроциты залегают в сером веществе мозга (то есть там, где находятся тела
нейронов). У них светлая цитоплазма, короткие и толстые отростки, которые,
прилегая к телам нейронов и сосудам, частично распластываются и принимают вид пластинок. Волокнистые астроциты залегают в белом веществе
мозга, то есть там, где находятся нервные волокна. У этих клеток цитоплазма
темнее, более длинные, тонкие и слабоветвящиеся отростки по сравнению с
плазматическими астроцитами. Они также образуют расширения в виде пластинок на стенках сосудов и нервных волокон, отграничивая их друг от друга
и в то же время удерживая в определенном положении. Оба вида астроцитов
выполняют опорную и разграничительную функции. Есть данные, что они
участвуют в водном обмене и транспорте веществ из капилляров к нейронам.
Олигодендроциты — многочисленная и довольно разнообразная группа
глиоцитов. Это мелкие клетки угловатой или овальной формы с небольшим
количеством коротких тонких отростков. Они окружают тела и отростки
нейронов, сопровождая их вплоть до нервных окончаний. Функции их разнообразны. Они участвуют в образовании оболочек вокруг дендритов и аксонов, в питании нейронов. При сильном возбуждении передают часть своей
РНК в тело нейрона. Способны накапливать в себе большое количество жидкости и других веществ, поддерживая гомеостаз нервной ткани. Следовательно, олигодендроциты выполняют разграничительную, трофическую и
гомеостатическую функции.
Микроглия (глиальные макрофаги)—мелкие клетки, происходящие из
мезенхимы, а затем из клеток крови, по-видимому, путем трансформации
моноцитов. Количество их невелико— около 5% глиальных клеток. В спокойном состоянии у них удлиненное тело и небольшое число ветвящихся отростков. При возбуждении отростки втягиваются, клетки округляются, увеличиваются в объеме, приобретают подвижность и способность к фагоцитозу.
Нервные волокна — отростки нервных клеток (аксоны и дендриты), покрытые оболочками из глиоцитов. В головном и спинном мозге оболочку волокон образуют олигодендроциты, в остальных частях— их разновидность,
называемая леммоцитами (шванновскими клетками).
В зависимости от особенностей строения различают миелиновые и безмиелиновые нервные волокна. Безмиелиновые волокна распространены в вегетативной нервной системе и в сером веществе мозга, миелиновые — в периферической (соматической) нервной системе и в белом веществе. При образовании волокна клетки олигодендроглии располагаются вдоль отростка
нейрона, плотно прилегая как к отростку, так и друг к другу. Отросток нервной клетки, входящий в состав волокна, называется осевым цилиндром.
Безмиелиновые нервные волокна. В случае образования безмиелинового
нервного волокна отросток нейрона продавливает в месте прилегания к лемwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
моциту его оболочку в виде желобка. По мере опускания отростка желобок
становится глубже, плазмолемма леммоцита одевает его со всех сторон в виде муфты. В конце концов осевой цилиндр, погруженный в леммоцит, как бы
повисает в складке (мезаксоне) его плазмолеммы. Мезаксон и плазмолемму
леммоцита, окружающую осевой цилиндр, видно только в электронный микроскоп. В безмие-линовых волокнах, как правило, проходит несколько осевых цилиндров (3—20). Они могут быть погружены в леммоцит на разную
глубину и иметь разной длины мезаксон. Такие волокна называются волокнами кабельного типа. Толщина их 1—5 мкм. Ядра леммоцитов располагаются как сбоку, так и в центре волокна. Изоляция осевых цилиндров внутри
волокон кабельного типа невелика, нервный импульс может распространяться диффузно — на все осевые цилиндры волокна. Осевые цилиндры переходят из одного безмиелинового волокна в другое, что также способствует распространению нервного импульса по волокнам. Скорость прохождения нервного импульса сравнительно невелика — 0,2— 2 м/с.
Рис. 34. Схема строения миелинового нервного волокна:
1 — осевой цилиндр; 2 — неврилемма:
3 — ядра и 4— отростки леммоцита;
5 — миелиновая оболочка; 6 — узловой
перехват; 7— межузловой сегмент.
Миелиновые нервные волокна устроены сложнее (рис. 34). В центре каждого миелинового волокна проходит осевой цилиндр, одетый миелиновой
оболочкой. Верхний слой волокна называется неврилеммой. Миелиновая
оболочка и неврилемма— это составные части леммоцитов, окружающих
осевой цилиндр. При образовании миелинового волокна леммоциты, прилегающие к отростку нейрона, уплощаются и накручиваются вокруг осевого
цилиндра, обертывая его несколько раз. При этом из намотавшегося участка
леммоцита цитоплазма выдавливается в свободные участки, а плазмолемма
спадается, слипается и образует слой миелиновой оболочки. В процессе накручивания на осевой цилиндр леммоцит растет, все больше вытягивается,
количество слоев миелина увеличивается. Оставшаяся ненамотанной часть
клетки с ядром и цитоплазмой оказывается сверху. Это и будет неврилемма
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(невролемма). Леммоциты несравнимо меньше осевого цилиндра. Располагаются они В волокне поочередно, соединяясь друг с другом пальцеобразными выростами. В месте контакта соседних леммоцитов волокно резко истончается, так как миелиновая оболочка здесь отсутствует и волокно покрыто
только неврилеммой — узловые перехваты. Участки волокна, покрытые
миелиновой оболочкой, называются межузловыми сегментами.
Миелиновые волокна толще безмиелиновых. Их диаметр 7— 20 мкм.
Нервный импульс по ним проходит гораздо быстрее (5—120 м/с). Чем толще
волокно, тем быстрее идет по нему импульс. В ускорении прохождения
нервного импульса большую роль играет миелиновая оболочка. В узловых
перехватах плазмолемма (аксолемма) осевого цилиндра возбуждается, как и в
безмиелиновых нервных волокнах, в результате деполяризации под влиянием
ионных потоков. В области же межузловых сегментов миелиновая оболочка,
действуя как изолятор, способствует молниеносному прохождению нервного
импульса, подобно тому, как это происходит в электрическом проводнике. В
результате нервный импульс как бы перескакивает от одного узлового перехвата до другого и таким образом движется с большой скоростью.
Безмиелиновые и миелиновые нервные волокна за пределами центральной нервной системы одеты базальной мембраной, подобной базальной мембране эпителия. В нервной ткани нервные волокна образуют ансамбли, характерные для того или иного участка нервной системы. Характер расположения нервных волокон называется миелоархитектоникой. В центральной
нервной системе волокна образуют проводящие пути, на периферии — нервные стволы или нервы.
Нерв. Нервные волокна, объединенные соединительной тканью, образуют нерв, а тончайшие прослойки соединительной ткани, расположенные
между нервными волокнами, — эндоневрий. Он тесно связан с базальными
мембранами волокон, в нем залегают капилляры. Эндоневрий связывает
нервные волокна в пучок. Пучки нервных волокон одеты периневрием — более широкими прослойками соединительной ткани с упорядоченным расположением волокон и с проходящими в ней сосудами. Снаружи нерв покрыт
эпиневрием — волокнистой соединительной тканью, богатой фибробластами,
макрофагами, жировыми клетками. В нем разветвляются кровеносные и
лимфатические сосуды и нервы нервов.
В состав нервов входят как миелиновые, так и безмиелиновые волокна.
Бывают нервы чувствительные, образованные дендритами чувствительных
нейронов (чувствительные черепномозговые нервы), двигательные — образованные аксонами моторных нейронов (двигательные черепномозговые
нервы) и смешанные — в состав которых входят отростки различных по
функции и структуре нейронов (спинномозговые нервы). Размеры нервов и
их состав зависят во многом от размеров и функциональной активности органов, иннервируемых ими. Замечено, что нервы мышц динамического типа
с активной двигательной функцией состоят из толстых миелиновых волокон
с небольшим количеством безмиелиновых. Так же устроены вентральные
ветви спинномозговых нервов. В дорсальных ветвях спинномозговых нервов
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
и в нервах, иннервирующих динамостатические мышцы, более тонкие миелиновые и больше безмиелиновых волокон.
Рис. 35. Типы нервных
окончаний:
I — чувствительные нервные
окончания — неинкапсулированные: А — в эпителии роговицы;
Б— в эпителии пятачка свиньи;
В — в перикарде лошади; инкапсулированные:
Гфатерпачиниево тельце; Д — тельце
Мейснера; Е — тельце из соска
овцы; 11 — двигательные нервные окончания; Ж —в поперечнополосатом волокне; 3 —в
гладкой мышечной клетке; 1 —
эпителий; 2 — соединительная
ткань; 3 — нервные окончания:
4 — меркелевская клетка; 5 —
дискоидальное концевое расширение нервного окончания; 6 —
нервное волокно; 7 — разветвление осевого цилиндра; 8 — капсула; 9 — ядро леммоцита; 10 —
мышечное волокно.
Нервные окончания (рис. 35). Нервное окончание — место контакта отростка нервной клетки с различными структурами не нервной природы. Это
могут быть мышечные волокна, клетки железистого или покровного эпителия и др. В зависимости от функциональной направленности различают чувствительные (рецепторные, афферентные) и двигательные (эффекторные,
эфферентные) нервные окончания.
Чувствительные нервные окончания — рецепторы образованы конечными разветвлениями дендритов чувствительных нейронов и воспринимают
раздражения, идущие к ним от разных участков организма или извне. Они
рассеяны по всему организму. В зависимости от того, откуда рецепторы получают раздражение, их делят на экстерорецепторы, воспринимающие раздражения из внешней среды, проприорецепторы, несущие возбуждения от
органов движения, и интерорецепторы, воспринимающие раздражения от
внутренних органов.
Рецепторы чувствительны лишь к определенному виду раздражителей.
В связи с этим различают механо-, термо-, фото-, баро-, хемо- и другие рецепторы. Наиболее распространенные механорецепторы. Они присутствуют
в коже, мышцах, внутренних органах. Болевые ощущения воспринимаются
как болевыми рецепторами, так, по-видимому, и любыми другими рецептоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рами при их чрезмерном раздражении. По структуре рецепторы делят на свободные и несвободные. Несвободные рецепторы, в свою очередь, бывают инкапсулированные и неинкапсулированные.
Свободные нервные окончания образованы только конечными разветвлениями дендритов, которые ничем не покрыты, и в виде кустиков, клубочков, петелек, колечек располагаются между клетками иннервируемой ткани.
Чаще всего свободные нервные окончания встречаются в эпителии и соединительной ткани. Их много в эпидермисе носового зеркала у овец и лошадей,
носогубного зеркала у коровы, вокруг волосяных фолликулов. Они обладают
разнообразной чувствительностью.
Несвободные нервные окончания представляют собой конечные разветвления дендрита, окруженные специальными рецепторными клетками. Неинкапсулированные нервные окончания — это такая разновидность несвободных рецепторов, в которой разветвления осевого цилиндра (дендрита) окружены эпителиальными или глиальными клетками. Подобные нервные окончания хорошо развиты в пятачке свиньи. Это осязательные мениски (диски
Меркеля), в которых конечные разветвления дендрита оплетают особые
клетки в многослойном эпителии, чувствительные к прикосновению и давлению.
Инкапсулированные нервные окончания устроены наиболее сложно. В
них осевой цилиндр окружен не только клетками глии, но и соединительнотканной капсулой. Существует много разновидностей инкапсулированных
нервных окончаний: осязательные тельца (Мейснера)—тактильные рецепторы, пластинчатые тельца (Фатера — Пачини)—барорецепторы, луковицеобразные тельца (Гольджи — Маццони), генитальные тельца (Догеля), концевые колбы (Краузе)—терморецепторы, нервно-мышечные веретена и др.
Лучше других изучено пластинчатое тельце (Фатера - Пачини) и нервномышечное веретено.
В пластинчатом тельце конечные разветвления дендрита (телодендрии)
окружены глиальными клетками, которые, распластываясь и плотно наслаиваясь друг на друга, образуют внутреннюю колбу (луковицу). Внутренняя
колба покрыта слоями распластанных фибробластоподобных клеток, в совокупности формирующих наружную капсулу тельца. Между внутренней колбой и наружной капсулой и около нервного окончания имеется пространство,
в котором обнаруживаются чувствительные отростчатые (ресничные)
клетки. Пластинчатые тельца реагируют на любые изменения давления в
тканях (давление жидкостей, при опоре, нажатии, ударе и т. д.), кодируя при
этом направление, частоту раздражающего стимула и вид его энергии. Они
очень распространены в организме — залегают в соединительной ткани органов опорно-двигательного аппарата, внутренних органов, кровеносных сосудов, нервных стволов, встречаются в лимфатических узлах, вегетативных
ганглиях, эндокринных железах. Количество и размеры их колеблются в зависимости от возраста, места расположения и частоты возбуждения (0,1—6
мм).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Другие инкапсулированные рецепторы построены по такому же принципу, различаясь характером ветвления осевого цилиндра, количеством и
расположением пластинок во внутренней колбе и капсуле. Особенности
строения определяют характер чувствительности того или иного нервного
окончания. В поперечнополосатой мышечной ткани разветвления осевого
цилиндра оплетают сверху группу видоизмененных мышечных волокон, образуя подобие веретена. Сверху нервно-мышечное веретено покрыто соединительнотканной капсулой.
Двигательные нервные окончания — эффекторы в гладкой мышечной
ткани и железах обычно построены по типу свободных нервных окончаний.
В поперечнополосатой мышечной ткани они имеют сложное строение и называются нервно-мышечными синапсами, или моторными бляшками. Подойдя к мышечному волокну, нервное волокно видоизменяется. Его осевой цилиндр, являющийся аксоном моторного нейрона, разветвляется на терминали, которые вдавливаются в мышечное волокно и образуют с его плазмолеммой контакт, подобный синапсу. Плазмолемма аксона в месте контакта —
это пресинаптическая мембрана нервно-мышечного синапса, плазмолемма
мышечного волокна — постсинаптическая. Между ними находится синоптическая щель шириной около 50 нм. Базальные мембраны нервного и мышечного волокна соединяются, переходя одна в другую и покрывают сверху
моторную бляшку. Плазмолемма мышечного волокна в месте контакта образует многочисленные складки. Предполагают, что с их развитием связана
скорость сокращения мышцы. Один двигательный нейрон (и его аксон) вместе с иннервнруемыми им мышечными волокнами создает двигательную
единицу — мион. Сила сокращения мышцы зависит от того, сколько двигательных единиц участвует в сокращении.
Рис. 36. Рефлекторная дуга:
1—спинной мозг; 2 — дорсальный и 3 — вентральный рог серого вещества; 4— спинномозговой ганглий; 5 — чувствительный и 6 —
двигательный корешки спинномозгового нерва; 7 — смешанный спинномозговой нерв; 8 —
кожа; 9 — мышца; 10 — чувствительное
нервное окончание: 11—дендрит; 12 — тело и
13— аксон чувствительного нейрона; 14 —
вставочный нейрон и его (15) аксон; 16 — двигательный нейрон и его (17) аксон; 18 — двигательное нервное окончание.
В нее входят от 3 до 2000 мышечных волокон. Мышечные волокна, относящиеся к одной двигательной единице, распределены по всей мышце. В
результате при возбуждении небольшого числа нейронов сокращается вся
мышца, а не какая-то ее часть.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рефлекторная дуга (рис. 36). Возбуждение в нервной ткани и в нервной
системе распространяется не хаотично, а по определенным путям — рефлекторным дугам. Рефлекторная дуга образована чувствительным, одним или
несколькими ассоциативными и двигательными нейронами. Возбуждение в
рефлекторной дуге идет всегда в строго определенном направлении: от рецептора (чувствительного нервного окончания) по центростремительному
отростку чувствительного нейрона (обычно дендриту) к его телу, расположенному в ганглии (нервном узле), откуда по его центробежному отростку
(аксону) — к дендриту ассоциативного нейрона. Между аксоном чувствительного нейрона и дендритом ассоциативного нейрона образуется синапс,
пропускающий нервный импульс только в одном направлении: от пресинаптического полюса к постсинаптическому. Нервный импульс последовательно переходит на дендрит, тело и аксон ассоциативного нейрона, а оттуда
— через синаптическую связь на дендрит, тело и аксон моторного нейрона.
Ассоциативные нейроны с отростками, дендриты и тела моторных нейронов
располагаются в центральной нервной системе. Аксоны же моторных нейронов покидают ее и направляются к иннервируемым тканям и органам, где
их конечные разветвления формируют двигательные нервные окончания —
эффекторы. Раздражение рецептора (например, надавливание на кожу возбуждает пластинчатые
тельца) приводит к волне возбуждения, которая проходит путь по рефлекторной дуге и, дойдя до эффектора, организует ответное действие, названное рефлексом. (В нашем примере—сокращение мышц в ответ на надавливание и как следствие этого — движение.)
Возрастные и реактивные изменения в нервной ткани. У новорожденных животных структурные элементы нервной ткани дифференцированы в такой степени, что могут выполнять все функции, присущие нервной системе (особенно у зрелорождающихся копытных): рецепцию,
интеграцию рецепторного сигнала, передачу нервного импульса, секрецию
медиатора в синаптическую щель и организацию эффекторной реакции. Тем
не менее и в постнатальный период онтогенеза идет увеличение размеров и
усложнение структуры нейронов, что, видимо, связано со спецификой их
функционирования. Пропорционально увеличиваются размеры тел и ядер
нейронов, накапливается базофильное вещество. У овец прослежено увеличение его количества до 3-летнего возраста, а с 5—6 лет — возрастное снижение. Утолщаются миелиновая оболочка и размеры леммоцитов, образующих эту оболочку.
Вопросы для самоконтроля. 1. Каково происхождение и принципы
строения нервной ткани? 2. Что такое нейрон, какие бывают нейроны по
структуре и функции? 3. Что такое синапс, его виды и строение? 4. Какие
клетки нейроглии вы знаете, чем они отличаются друг от друга? 5. Что такое
нервное волокно, как оно устроено, чем отличаются и где встречаются миелиновые и безмиелиновые волокна? 6. Что такое нервное окончание? 7.
Классификация и строение нервных окончаний. 8. Состав рефлекторной дуги.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Раздел четвертый. МОРФОЛОГИЯ ОРГАНОВ И ИХ СИСТЕМ
Глава 11. ОБЩИЕ ПРИНЦИПЫ ПОСТРОЕНИЯ И РАЗВИТИЯ
ОРГАНИЗМА
Современная морфология, стоящая на позициях диалектического материализма, рассматривает организм — объект своего изучения — как единое
целое, все части которого взаимосвязаны, взаимообусловлены и взаимозависимы. Кроме того, организм рассматривается не в статике, а в процессе его
роста и развития (в онтогенезе), в свете эволюционных преобразований (в
филогенезе), в зависимости от условий жизни и влияния функций.
Такой всеобъемлющий подход к (Изучению организма с помощью целого ряда наук и направлений, входящих как составные части в морфологию,
дал основание В. А. Домбровскому (1946) определить ее как интегральную
морфологию. Следовательно, организм — это живая, целостная, существующая самостоятельно, исторически сложившаяся система, имеющая свое особое строение и развитие, обусловленное наследственными свойствами, взаимодействием его частей и влиянием среды. Состоит организм из органов,
объединенных в системы и аппараты, которые обеспечивают все проявления
его жизни: реактивность, обмен веществ, размножение, рост и развитие.
Орган (organon — орудие) — часть организма, построенная из закономерно взаимосвязанных тканей; имеет определенную форму, занимает определенное положение в организме и выполняет специфическую функцию. Органы единого происхождения, сходного строения, как правило, морфологически тесно связаны и взаимозависимы, выполняют общую функцию, составляют систему органов, например нервная, сосудистая, костная, мышечная и другие системы. Органы, обеспечивающие определенный жизненный процесс, но имеющие разное строение и происхождение, объединяются
в аппарат. Например, аппарат движения, пищеварения, дыхания, крово-,
лимфообразования и др. Системы органов могут входить в аппараты как их
составная часть.
Системы органов и аппараты в зависимости от их морфофункциональных особенностей делят на три группы: соматическую, висцеральную и интегрирующую. В соматическую группу входят скелет, мускулатура (объединяемые в аппарат движения) и органы кожного покрова. Они
образуют сому — стенки тела. В висцеральную (спланхническую) группу входят пищеварительный, дыхательный и мочеполовой аппараты. В совокупности они составляют внутренности (греч. splanchna, лат. viscera), расположенные большей частью в естественных полостях тела. В группу интегрирующих систем входят эндокринная, сердечно-сосудистая и нервная системы
с органами чувств. Сердечно-сосудистая система пронизывает все органы и
ткани организма (за редким исключением), выполняет транспортную функцию и объединяет все системы. Через нее осуществляется гуморальная регуляция. Нервная система регулирует и координирует деятельность всех систем, в том числе и сосудистой, обеспечивая гармоничную целостность оргаwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
низма и адекватную связь его с окружающей средой с помощью органов
чувств.
Для всех хордовых характерны единые принципы построения тела: а)
биполярность (одноосность)—тело имеет два полюса — головной (краниальный) и хвостовой (каудальный); б) билатеральность — двусторонняя симметрия — правая и левая стороны тела являются зеркальным отражением
друг друга; в) сегментность (метамерия) — близлежащие сегменты близки
построению; г) четырехногость (тетраподия); д) расположение большинства
непарных органов вдоль основной оси тела.
Направление сил тяжести четвероногого животного совпадает с анатомическими границами, разделяющими скелет на его естественные отделы:
череп, шейный, грудной, поясничный, крестцовый и хвостовой, а на конечностях проходит через отделы конечностей (пояс, стилоподий, зейгоподий,
автоподий). Общий центр тяжести проходит через печень, что у рогатого
скота соответствует уровню 11-го грудного позвонка.
Взаимосвязь организма со средой. Организм не может существовать в
отрыве от окружающей среды, так как постоянно обменивается с ней веществом и энергией. На изменения внешней среды организм отвечает приспособительными реакциями. Однако они небезграничны и всегда находятся в
пределах нормы данной реакции.
Норма реакции — это пределы способности организма реагировать изменением своих морфологических и физиологических свойств на изменения
среды обитания без нарушения основных морфофункциональных систем.
Нормой строения считается наиболее часто встречающийся вариант
строения организма. Для животных разных видов, пола, возраста, конституции характерны свои нормы строения, отличающие их от других групп (возрастных, половых и т. д.). Так, для молодняка копытных характерна относительная высоконогость, а для взрослых — растянутость тела. Норма строения
закреплена в генотипе более жестко, чем норма физиологическая. Следовательно, структурные изменения под влиянием среды будут менее выражены,
чем функциональные. Но и структуры в разной степени генетически обусловлены. Так, размеры головы, длина трубчатых костей, форма мышц в
большей степени определяется генотипом, чем, например, размеры поясницы, толщина трубчатых костей, масса мышц. Индивидуальная изменчивость
организма и органов, если она не нарушает их жизнедеятельности, находится
в пределах нормы реакции и строения. Если изменения среды превышают
адаптивные возможности организма, развивается патология, которая выражается в болезнях, уродствах, преждевременной смерти и т. д.
Онтогенез как диалектический процесс. Организм в течение всего периода своего существования закономерно изменяется. Процесс индивидуального развития — онтогенез — начинается с появления зиготы и кончается со
смертью организма. Развитие организма является диалектическим процессом,
то есть таким процессом, в котором взаимосвязаны и взаимозависимы противоречивые явления. Противоречивость внутренне присуща онтогенезу— это
его движущая сила. Так, противоречие наследственности и изменчивости опwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ределяет весь ход индивидуального развития; ассимиляция (анаболизм) и
диссимиляция (катаболизм) неразрывно связаны в ходе обмена веществ
(энергия, освобождающаяся при катаболизме, тратится в процессе анаболического синтеза); взаимодействие между отмирающими и нарождающимися структурами, между прогрессивными и регрессивными процессами наблюдается на всех стадиях развития (появление дочерних клеток
на базе материнской, образование кости на месте резорбирующегося хряща и
т. д.).
Развитие особи — это отражение развития вида, что было сформулировано Э. Геккелем в виде биогенетического закона, который гласит, что онтогенез повторяет филогенез. Ч. Дарвин в 1842 г. писал, что зародыш является
как бы свидетелем прошлых столетий, через которые прошел вид. А. Н. Северцов дополнил и расширил это положение, показав, что онтогенез не только результат, но и основа филогенеза, так как филогенез — это ряд онтогенезов и изменения, происходящие в генотипе особи, передаваясь потомству,
влияют на направление филогенеза. Изменения же в процессе развития особи, происходящие под влиянием окружающей среды, считаются приспособительными. Выраженность изменений зависит от индивидуальных реакций организма, и тогда начинает действовать естественный отбор, сохраняя особей,
изменения которых оказались наиболее адаптивными, повысили их жизнеспособность, способствовали активному размножению и т. д. Чем шире норма реакции и структуры, чем шире гено- и фенотипическая изменчивость,
тем больше возможности морфофункционального приспособления, а следовательно, процветания и эволюции вида. Наглядно влияние внешней среды
на морфофункциональную организацию животного видно в процессе одомашнивания: у рыжих лис уже через несколько поколений появляются полиэстричность, пестрота окраски шерсти, меняется поведение, появляется способность махать хвостом; у овец меняются длина и структура желудочнокишечного тракта, качество шерсти.
Онтогенез осуществляется по определенному плану: у свиньи рождаются поросята, из которых вырастают свиньи, а не, скажем, короЕЫ. Закономерность и направленность онтогенеза определяются генетической программой, взаимовлиянием частей организма в процессе развития и функционирования, а полнота ее реализации зависит от влияния внешней среды и проявляется в фенотипе.
Взаимовлияние частей организма совершается путем регуляции и происходит на всех уровнях — от молекулярного до системоорганного. Регуляторные процессы всегда основываются на принципе отрицательной обратной
связи и являются саморегулирующимися: регулируемый орган накоплением
продуктов своей деятельности подавляет активность регулятора. На уровне
клетки регуляция осуществляется через клеточный метаболизм и межклеточные взаимодействия. На органном и организменном уровне — с помощью
эндокринной и нервной систем.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рост и дифференцировка — две стороны единого процесса развития.
Индивидуальное развитие (онтогенез) включает в себя качественную (дифференцировка) и количественную (рост) стороны.
Дифференцировка, или дифференциация, — возникновение в процессе
развития организма биохимических, морфологических и функциональных
различий между клетками, тканями и органами. В результате происходит
расчленение целого на части, образуются и приобретают специализацию различные клетки, ткани, органы. В онтогенезе многоклеточных животных специализация возникает на стадии нескольких бластомеров. В процессе дифференцировки органеллы, клетки, ткани, органы приобретают специфические
черты строения и присущую им функцию. При этом оказывается, чем более
специализирована часть организма, тем больше она зависит от других частей.
Р о с т — увеличение массы и размеров тела и его частей, органически
связанное с формативными процессами. В биологии до настоящего времени
нет единой теории роста, хотя сам процесс хорошо изучен у животных различных видов, классов, типов. Рост — это количественный показатель изменения структур, поэтому его легче, чем другие характеристики онтогенеза,
выразить в математической форме. Характер роста у животных разных видов
и даже у одного животного на разных стадиях онтогенеза неодинаков.
Подавляющее большинство теплокровных животных (кроме крыс), достигнув определенных размеров, прекращают расти. Такой рост называется
ограниченным. Для большинства хладнокровных хордовых животных (в том
числе рыб) характерен неограниченный рост. Эти животные растут в течение
всей жизни.
Рост и дифференцировка связаны между собой обратной зависимостью:
чем более дифференцирован организм, тем меньше скорость его роста. Наибольшая скорость роста наблюдается у зародыша, меньше у плода, еще
меньше — после рождения, а с достижением физиологической и морфологической зрелости рост прекращается. Следовательно, в онтогенезе происходит
постоянное убывание скорости роста. Однако убывание это неравномерно,
так как периоды активного роста и дифференцировки чередуются, в результате чего рост носит характер угасающих колебаний, а дифференцировка
также ступенчато возрастает до периода морфофункциональной зрелости.
У разных животных скорость и продолжительность роста неодинаковы,
причем между этими показателями роста имеется обратная зависимость: чем
быстрее скорость роста, тем короче его продолжительность, и наоборот.
Особенно высокая скорость роста и короткая его продолжительность у птиц.
У млекопитающих отмечена связь между размерами животного, скоростью и
продолжительностью роста. Виды мелких животных растут обычно интенсивно, но непродолжительно, виды крупных животных менее интенсивно, но
длительно. Масса тела, которой достигает тот или иной вид не случайна, а
определяется взаимоотношением между поверхностью тела, его объемом,
поверхностью слизистой оболочки кишечника и интенсивностью обменных
процессов. Эти взаимоотношения накладывают строгие пределы на размеры
животного. Если во время роста тело не меняет форму, такой рост называется
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
пропорциональным. Он характерен лишь для самых ранних стадий дробления зародыша и в период, близкий к завершению роста (от периода полового
созревания до морфофизиологической зрелости). Для всех остальных периодов характерен непропорциональный рост, когда одни органы растут быстрее, чем другие. При этом меняются пропорции тела, соотношения тканей
в органах.
Неравномерность роста определяется сроком закладки органов (одни органы закладываются рано, например глаза, мозг, другие гораздо позже —
кишечник, мышцы), величиной закладки (у глаз, мозга — большая, у легких,
мышц — сравнительно меньшая), сроками и длительностью гистологической
дифференцировки. Если у органа большая величина закладки и эта закладка
формируется рано, для такого органа характерен медленный темп эмбрионального и постнатального роста (глаз, мозг). К тому же такие органы генетически высокодетерминированы, то есть их морфологическое развитие определяется в основном генотипом. Если закладка органа формируется поздно, но ее гистологическая дифференцировка идет быстро, для такого органа
характерен быстрый эмбриональный и короткий постнатальный рост (например, печень). Если закладка органа формируется поздно, а гистологическая
дифференцировка у нее медленная, для такого органа характерен быстрый
эмбриональный и долгий постнатальный рост (аппарат движения, размножения). При этом чем позже закладывается орган, тем развитие его больше
зависит от условий среды. А так как кости, мышцы и кожа являются одним
из наиболее поздно дифференцирующихся органов, это дает в руки зооинженера ключ к управлению процессами формирования мясной и шерстной продуктивности.
Периодизация развития. В процессе индивидуального развития выделяют определенные этапы, когда в результате нарастающих количественных
изменений организм переходит в новое качественное состояние. Периоды
внутриутробного развития прослежены нами в разделе «Эмбриология». В постнатальном онтогенезе выделяют периоды новорожденность, молочный, роста и развития молодняка, полового созревания, морфофизиологической зрелости, расцвета функциональной деятельности (взрослого состояния), старения. В процессе индивидуального развития есть критические периоды, когда
те или иные органы наиболее чувствительны к внешнему воздействию. Оказалось, что критическим периодом для органа является время наиболее интенсивного его роста, а также моменты периодических (ритмичных) повышений скорости роста на протяжении онтогенеза. В ритмичности проявляется неравномерный характер роста. Давно известны годичные ритмы, связанные со сменой времен года. У диких животных, а из домашних у северного
оленя, верблюда рост приостанавливается зимой и возобновляется в теплое
время года. Обнаружены околомесячные, околосуточные и околочасовые
ритмы у всех млекопитающих, а также 12-дневные циклы активности и затухания роста у телят и цыплят. Применение дифференцированного кормления
в соответствии с обнаруженными ритмами позволило увеличить продуктивность на 20—30% при 20% экономии кормов. Знание закономерностей роста
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
и использование их в практике животноводства позволяют управлять не
только процессами роста животного, но и его развития в целом, что является
одним из резервов интенсификации животноводства.
ПЛОСКОСТИ ТЕЛА И ТЕРМИНЫ ДЛЯ ОБОЗНАЧЕНИЯ
РАСПОЛОЖЕНИЯ ОРГАНА
Для более точного определения расположения органов и частей тело
животного расчленяют тремя воображаемыми взаимоперпендикулярными
плоскостями — сагиттальной, сегментальной и фронтальной (рис. 37). Срединная сагиттальная (медианная) плоскость проводится вертикально вдоль
середины тела животного от рта до кончика хвоста и рассекающая его на две
симметричные половины. Направление в теле животного к срединной плоскости называется медиальным, а от нее — латеральным (lateralis — боковой).
Рис. 37. Плоскости и направления в теле животного.
Плоскости: I — сегментальная; II — сагиттальная; III — фронтальная. Направления: 1 — краниальное; 2 — каудальное; 3 — дорсальное; 4 — вентральное; 5 — медиальное; 6 — латеральное; 7 — ростральное (оральное); 8
— аборальное; 9 — проксимальное; 10 — дистальное; 11 — дорсальное
(спинковое, тыльное); 12 — пальмарное; 13 — плантарное.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Сегментальную плоскость проводят вертикально поперек тела животного. Направление от нее в сторону головы называется краниальным (cranium
— череп), в сторону хвоста — каудальным (cauda — хвост). На голове, где
все краниальное, раз дичают направление к носу — назальное или хоботку—
ростральное и противоположное ему — каудальное. Фронтальную плоскость (Irons — лоб) проводят горизонтально вдоль тела животного (при горизонтально вытянутой голове), то есть параллельно лбу. Направление в
данной плоскости в сторону спины называется дорсальным (dorsum — спина), к животу — вентральным (venter — живот).
Для определения положения участков конечностей существуют термины
проксимальный (proximus — ближайший) — более близкое положение к осевой части тела и дистальный (distalus — удаленный) — более отдаленное положение от осевой части тела. Для обозначения передней поверхности конечностей приняты термины краниальный или дорсальный (для лапы), а для
задней поверхности — каудальный, а также пальмарный или волярный
(palma, vola — ладонь) —для кисти и плантарный (planta — стопа) — для
стопы.
ОТДЕЛЫ И ОБЛАСТИ ТЕЛА ЖИВОТНОГО И ИХ КОСТНАЯ
ОСНОВА
Тело позвоночных животных делят на осевую часть и конечности. У рыб
осевая часть состоит из головы, туловища и хвоста. Начиная с амфибий, у
животных осевую часть тела делят на голову, шею, туловище и хвост. Шея,
туловище и хвост в совокупности составляют ствол тела. Каждая из частей
тела, в свою очередь, разделена на отделы и области (рис. 38). Основой их в
большинстве случаев являются кости скелета, имеющие те же названия, что и
области.
Голова (лат. caput, греч. cephale) делится на череп (мозговой отдел) и
лицо (лицевой отдел).
Череп (cranium) представлен областями: затылочной (затылок), теменной (темя), лобной (лоб) с областью рога у рогатого скота, височной (висок)
и околоушной (ухо) с областью ушной раковины.
На лице (fades) различают области: глазничную, (глаза) с областями
верхнего и нижнего век, подглазничную, скуловую с областью большой жевательной мышцы (у лошади — ганаши), межчелюстную, подбородочную,
носовую (нос) с областью ноздрей, ротовую (рот), в состав которой входят
области верхней и нижней губ и щеки. Над верхней губой (в области ноздрей) находится носовое зеркальце, у крупных жвачных оно распространяется
на область верхней губы и становится носогубным.
Шея (cervix, collum) простирается от затылочной области до лопатки и
делится на области: верхнюю шейную, лежащую над телами шейных позвонков; боковую шейную (область плечеголовной мышцы), идущую вдоль тел
позвонков; нижнюю шейную, вдоль которой тянется яремный желоб, а также
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
гортанную и трахейную (на вентральной ее стороне). У копытных шея сравнительно длинная в связи с необходимостью питания подножным кормом.
Поэтому же у них существует прямая корреляция между длиной конечностей
и длиной шеи. Самая длинная шея у высоконогих быстроаллюрных лошадей.
Самая короткая — у свиньи. Форма шеи у травоядных овальная, вытянутая в
дорсовентральном направлении, у свиньи (всеядное) более округлая.
Рис. 38. Области тела крупного рогатого скота:
1 — лобная; 2— затылочная; 3— теменная; 4 — височная; 5 — околоушная; 6— ушной раковины; 7 — носовая; 8 — области верхней и нижней
губ; 9 — подбородочная; 10 — щечная; 11 — межчелюстная; 12 — подглазничная; 13 — скуловая; 14 — область глаза; 15 — большой жевательной
мышцы; 16 — верхняя шейная; 17 — боковая шейная; 18 — нижняя шейная;
19 — холки; 20 — спины; 21 — реберная; 22 — предгрудинная; 23 — грудинная; 24 — поясничная; 25 — подреберья; 26 — мечевидного хряща; 27 — околопоясничтая (голодная) ямка; 28 — боковая область; 29 — паховая; 30 —
пупочная; 31 — лонная; 32 — маклок; 33— крестцовая; 34 — ягодичная; 35
— корень хвоста; 36 — седалищная область; 37 — лопатка; 38 — плечо; 39
— предплечье; 40 — кисть; 41 — запястье; 42 — пясть; 43 — пальцы; 44 —
бедро; 45 — голень; 46 — стопа; 47 — заплюсна; 48 — плюсна.
Туловище (truncus) состоит из грудного, брюшного и тазового отделов.
Грудной отдел включает области холки, спины, боковые реберные,
предгрудинную и грудинную. Он прочный и одновременно подвижный. В
каудальном направлении прочность уменьшается, а подвижность нарастает
благодаря разной степени развития костей скелета и особенностям их соединения. Костной основой холки и спины являются грудные позвонки. В обласwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ти холки у них наиболее высокие остистые отростки. Чем выше и длиннее
холка, тем больше площадь прикрепления мышц позвоночника и пояса грудной конечности, тем размашистее и более упруги движения. Между длиной
холки и спины существует обратная зависимость. Самая длинная холка и короткая спина у лошади, у свиньи — наоборот.
Брюшной отдел включает поясницу (lumbus) и живот (abdomen), или
брюхо (venter), поэтому еще называется пояснично-брюшным отделом. Поясница — продолжение спины до крестцовой области. Ее основой являются
поясничные позвонки. Живот имеет мягкие стенки и делится на ряд областей: правого и левого подреберий, мечевидного хряща, верхняя граница которой проходит по реберной дуге; парную боковую (подвздошную) с голодной ямкой, примыкающую снизу к пояснице, спереди— к последнему ребру,
а сзади — переходит в паховую область; пупочную, лежащую снизу живота
позади области мечевидного хряща к впереди от лонной области. На вентральной поверхности областей мечевидного хряща, пупочной и лонной у
самок расположены молочные железы. У лошади наиболее короткая поясница и менее обширный брюшной отдел. У свиньи и рогатого скота поясница
более длинная. Наиболее объемистый брюшной отдел у жвачных.
Тазовый отдел (pelvis) делится на области: крестцовую, ягодичную,
включая маклок, седалищную и промежностную с примыкающей мошоночной областью. В хвосте (cauda) различают корень, тело и кончик. Области
крестцовая, две ягодичных и корень хвоста у лошади образуют круп.
Конечности (membra) подразделяются на грудные (передние) и тазовые
(задние). Состоят они из поясов, которыми соединяются со стволовой частью
тела, и свободных конечностей. Свободные конечности делятся на основной
поддерживающий столб и лапу. Грудная конечность состоит из плечевого
пояса, плеча, предплечья и кисти.
Области плечевого пояса и плеча примыкают к боковой грудной области. Костной основой плечевого пояса у копытных является лопатка, поэтому
его часто называют областью лопатки.
Плечо (brachiuni) расположено ниже плечевого пояса, имеет форму треугольника. Костной основой является плечевая кость.
Предплечье (antebrachium) находится вне кожного туловищного мешка.
Его костная основа — лучевая и локтевая кости.
Кисть (marius) состоит из запястья (carpus), пясти (metacarpus) и пальцев
(digiti). Последние называются порядковыми номерами, счет их ведут с внутренней стороны. У животных разных видов их бывает от 1 до 5. Каждый палец (кроме первого) состоит из трех фаланг: проксимальной, средней и дистальной (которые у копытных называются соответственно путовой, у лошади
— бабка), венечной и копытцевой (у лошади — копытной).
Тазовая конечность состоит из тазового пояса, бедра, голени и стопы.
Область тазового пояса (таз) входит в состав осевой части тела в качестве ягодичной области. Костная основа — тазовая или безымянная кости.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Область бедра (femur) расположена под тазом и так же, как плечо, в
своей значительной части покрыто вместе с туловищем общим пластом кожи. Костная основа — бедренная кость.
Область голени (crus) находится вне кожного туловищного мешка. Костная основа — большая и малая берцовые кости.
Стопа (pes) состоит из заплюсны (tarsus), плюсны (metatarsus) и пальцев
(digiti). Количество их, строение и названия у копытных такие же, как на кисти.
Вопросы для самоконтроля. 1. Что такое орган, система органов, организм? 2. Каковы принципы построения тела хордовых? 3. В чем выражается
взаимосвязь организма со средой? 4. Почему рост и дифференциация — две
взаимосвязанные стороны онтогенеза? 5. Какие плоскости тела и термины
для обозначения расположения органов и частей тела вы знаете? 6. Перечислите области головы, шеи, туловища, конечностей.
Часть I. СОМАТИЧЕСКИЕ СИСТЕМЫ
Кожный покров, скелетная мускулатура и скелет, формируя собственно
теле — сому животного, — объединяются в группу соматических систем организма.
Аппарат движения (система органов произвольного движения) образован двумя системами: костной и мышечной. Кости, объединенные в скелет,
представляют собой пассивную часть аппарата движения, являясь рычагами,
на которые действуют прикрепленные к ним мышцы. Мышцы действуют
только на кости, подвижно соединенные с помощью связок. Мышечная система— активная часть аппарата движения. Аппарат движения обеспечивает
движение организма, его перемещение в пространстве, поиск, захват и пережевывание пищи, нападение и защиту, дыхательные движения, движения
глаз, ушей и др. Аппарат движения формирует внешний облик, поскольку на
его долю приходится от 40 до 60% массы организма. Он определяет форму
тела животного (экстерьер), пропорции, обусловливая типовые особенности
конституции, что имеет огромное практическое значение в зоотехнии, так как
с особенностями экстерьера, типом конституции связаны выносливость, приспособляемость, способность к откорму, скороспелость, половая активность,
жизнестойкость, и другие качества животных.
Глава 12. СКЕЛЕТ, СОЕДИНЕНИЕ КОСТЕЙ СКЕЛЕТА (УЧЕНИЕ О
КОСТЯХ — ОСТЕОЛОГИЯ)
Общая характеристика и значение скелета. Скелет (греч. skeleton— иссохший, мумия) образован костями и хрящами, закономерно расположенными в теле и соединенными между собой соединительной, хрящевой или костной тканями. Скелет млекопитающих называется внутренним, так как он
расположен под кожей и, как правило, покрыт слоем мышц. Выполняет разнообразные функции. Прежде всего он является твердой основой тела и слуwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
жит футляром для головного, спинного и костного мозга, для сердца, легких
и других органов. Упругие деформации (сжатие и растяжение), возникающие
при движении животного, играют роль присасывающего механизма и этим
помогают сердцу перемещать кровь. Упругость и рессорные свойства скелета
обеспечивают плавность движений, предохраняют мягкие органы от толчков
и сотрясений. Скелет принимает активное участие в минеральном обмене. В
нем содержатся большие запасы неорганических солей кальция, фосфора и
других веществ, которые постоянно обмениваются в ходе перестройки костной ткани. Скелет — наиболее точный показатель степени развития и возраста животного. Многие прощупываемые кости являются по' стоянными
ориентирами при проведении зоотехнических измерений животного.
ДЕЛЕНИЕ СКЕЛЕТА
Скелет в соответствии с отделами тела животного делится на осевой и
скелет конечностей (периферический) (рис. 39).
Осевой скелет включает скелет головы, шеи, туловища и хвоста. Скелет
туловища состоит из скелета грудной клетки, поясницы и крестца. Периферический скелет образован костями поясов и свободных конечностей. Количество костей у животных разных видов, пород и даже особей неодинаково
(табл. 4).
4. Количество костей у разных видов животных
Крупный
Отделы скелета
Лошадь
рогатый
Овца
Свинья
скот
Скелет головы
31
31
31
32
Позвоночный столб
53—56
49—51
35—55
51—58
Ребра
18—19 пар 13—14 пар 12—14 пар 14—15 пар
Грудные конечности
42—40
48
48
82
Тазовые конечности
40—42
46
46
82
Масса скелета составляет у взрослого животного от 6 % (свиньи) до
12—15% (лошадь, бык). У новорожденного теленка достигает 20 %, у поросенка 30 % от массы тела. У новорожденных животных более развит периферический скелет. На его долю приходится 60—65 % массы всего скелета, а на
долю осевого 35—40 %- После рождения активнее растет, особенно в молочный период, осевой скелет и у 8—10-месячного теленка отношения этих отделов скелета выравниваются, а затем осевой начинает преобладать: К 18 месяцев у крупного рогатого скота он составляет 53—55%. У свиньи масса осевого и периферического скелета примерно одинакова.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 39. Скелет коровы (А), свиньи (Б), лошади (В).
Осевой скелет: 1 — кости мозгового отдела (черепа): 2— кости лицевого отдела (лица); 3 — шейные позвонки; 4 — грудные позвонки; 5 — ребра;
6— грудина (4—6 — грудная клетка); 7— поясничные позвонки; 8 — крестцовая кость; 9 — хвостовые позвонки (3, 4, 7, 8, 9 — позвоночник). Скелет
конечностей: 10— лопатка; 11 — плечевая кость; 12 — кости предплечья
(лучевая и локтевая); 13 — кости запястья; 14 — кости пясти; 15—кости
пальцев (13—15 — кости кисти); 16 — тазовая кость; 17 — бедренная
кость; 18 — коленная чашка; 19 — кости голени (большая и малая берцовые); 20 — кости заплюсны; 21 — кости плюсны; 22— кости пальцев (20—
22 — кости стопы).
КОСТЬ КАК ОРГАН. ФОРМА И СТРОЕНИЕ КОСТЕЙ
Кость (лат. os) считается органом костной системы. Как всякий орган,
она имеет определенную форму и состоит из нескольких видов тканей. Форма костей определяется особенностями ее функционирования и положением
в скелете. Различают длинные, короткие, плоские и смешанные кости.
Длинные кости бывают трубчатыми (многие кости конечностей) и дугообразными (ребра). Длина тех и других гораздо больше ширины и толщины.
Длинные трубчатые кости напоминают по форме цилиндр с утолщенными
концами (рис. 40). Средняя, более узкая часть кости называется телом —
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
диафиз (греч. diaphysis), расширенные концы — эпифизы (epiphysis). Эти
кости играют основную роль в статике и динамике животного, в кроветворной функции (содержат красный костный мозг).
Короткие кости обычно небольшой величины, их высота, ширина и
толщина близки по размеру. Они часто выполняют рессорную функцию или
являются сесамовидными костями.
Плоские кости имеют большую поверхность (ширину и длину) при малой толщине (высоте). Обычно они служат стенками полостей, защищая помещенные в них органы (черепномозговая коробка) или это обширное поле
для прикрепления мышц (лопатка).
Смешанные кости имеют сложную форму, совмещая в себе элементы
перечисленных форм. Эти кости, как правило, непарные и размещаются по
оси тела. При распиле по срединной сагиттальной плоскости они разделяются на две симметричные половины — правую и левую. Таковы затылочная,
клиновидная кости, позвонки. Парные смешанные кости несимметричны, например височная кость.
Строение кости. Основная ткань, образующая кость, — пластинчатая
костная. В состав кости входят также ретикулярная, рыхлая и плотная соединительные ткани, гиалиновый хрящ, кровь и эндотелий сосудов, нервные
элементы.
Снаружи кость одета надкостницей, или периостом, за исключением
места расположения суставного хряща. Наружный слой надкостницы фиброзный, образован соединительной тканью с большим количеством коллагеновых волокон; определяет ее прочность. Внутренний слой содержит недифференцированные клетки, которые могут преобразовываться в остеобласты и
являются источником роста кости. Через надкостницу в кость проникают сосуды и нервы. Надкостница во многом определяет жизнеспособность кости.
Кость, очищенная от надкостницы, погибает; надкостница же с вылущенной
костью дает начало костным структурам.
Под надкостницей залегает слой кости, образованный плотно уложенными костными пластинками. Это компактное вещество кости. В трубчатых костях в нем различают несколько зон. К надкостнице примыкает зона
наружных генеральных пластинок толщиной 100—200 мкм. Она придает
кости большую твердость. Затем следует наиболее широкая и важная в
структурном отношении зона остеонов. Чем толще слой остеонов, тем лучше
рессорные свойства кости. В этом слое между остеонами залегают вставочные пластинки — остатки старых разрушенных остеонов. У копытных в нем
часто встречаются циркулярно-параллельные структуры, которые рассматривают как структуры высшего порядка, чрезвычайно устойчивые к сопротивлению на изгиб. Не случайно они широко распространены в длинных
трубчатых костях копытных, испытывающих большое давление со стороны
грузного тела, значительные гравитационные, нагрузки при размашистых маятникообразных движениях. Толщина внутреннего слоя компактного вещества 200—300 мкм, образован он внутренними генеральными пластинками
или же переходит в губчатое вещество кости.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 40. Строение трубчатой кости.
А— продольный распил; Б — участок стенки кости; 1 — эпифиз; 2 — диафиз; 3 — метаэпифизарный хрящ; 4 — суставной хрящ; 5 — надкостница; 6
— компактное и 7 — губчатое вещество кости; 8 — костномозговая полость; 9— сосуды и 10 — нервы, входящие в кость; 11 — наружные генеральные пластинки; 12 — остеоны; 13 — сосуд остеона; 14 — внутренние
генеральные пластинки; 15 — костные перекладины.
Губчатое вещество представлено костными пластинками, которые не
плотно примыкают друг к другу, а формируют сеть из костных перекладин
(трабекул), в ячеях которой располагается красный костный мозг. Губчатое
вещество особенно развито в эпифизах. Его перекладины располагаются не
беспорядочно, а строго следуют линиям действующих сил (сжатия и растяжения). При изменении направления постоянно действующей силы постепенно изменяется и расположение трабекул.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
В середине диафиза трубчатой кости имеется костная полость, распространяющаяся у взрослых животных до эпифизов. Она образовалась в результате резорбции костной ткани остеокластами во время развития кости и
заполнена желтым (жировым) костным мозгом. Стенки костной полости
выстланы эндостом, выполняющим, как и периост, камбиальную функцию.
Им же покрыты перекладины губчатого вещества.
Кость богата сосудами, которые образуют сеть в ее надкостнице, пронизывают всю толщину компактного вещества, находясь в центре каждого остеона, и разветвляются в костном мозге. В кости кроме сосудов остеонов
имеются так называемые питательные сосуды (фолькмановы), прободающие
кость перпендикулярно ее длине. Вокруг них не образуется концентрических
костных пластинок. Особенно много таких сосудов около эпифизов. Нервы в
кость проникают от надкостницы через те же отверстия, что и сосуды. Поверхность кости, обращенная в полость сустава, покрыта гиалиновым хрящом без надхрящницы. Толщина его 0,2—6 мм и прямо пропорциональна нагрузке на сустав.
Строение коротких, сложных и плоских костей такое же, как и трубчатых, с той лишь разницей, что у них обычно нет костных полостей. Исключение составляют некоторые плоские кости головы, в которых между пластинками компактного вещества имеются обширные пространства, заполненные воздухом,— синусы или пазухи.
ФИЛОГЕНЕЗ СКЕЛЕТА
Развитие опорной системы в филогенезе многоклеточных животных шло
двумя путями: образования наружного и внутреннего скелета. Наружный
скелет закладывается в покровах тела. Наивысшего развития достигает у
членистоногих. Внутренний скелет развивается вне связи с покровами тела,
под кожей и обычно прикрыт мышцами. Его элементы встречаются и у беспозвоночных, но о развитии внутреннего скелета можно говорить со времени
появления хордовых. У примитивных хордовых (оболочники, ланцетник)—
спинная струна — хорда представляет собой опорную систему. У бесчерепных (ланцетник) внутренний скелет дополняется соединительнотканным футляром хорды, от которого отходят пластинки — миосепты, разделяющие мышечные пласты на отдельные сегменты (у ланцетника их около
60).
С усложнением организации животных соединительнотканный скелет
замещается хрящевым, а затем костным. Наряду со сменой тканевой основы
скелета происходило и его структурное усложнение.
Филогенез стволового скелета. В филогенезе позвоночных раньше других элементов скелета появляются позвонки. У наиболее примитивных позвоночных— круглоротых (миноги) зачатки позвонков развиваются в виде
хрящевых дуг, метамерно расположенных над хордой, выполняющей роль
основного опорного стержня тела (рис. 41—А—Д). Хрящевые дуги по своему
строению однотипны, так как движения круглоротых однообразны и мускуwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
латура недифференцирована на отдельные мышцы, в результате чего осевой
скелет нечетко разделяется на головной, туловищный и хвостовой отделы.
Рис. 41. Схема последовательных стадий образования позвонка.
Схема позвоночника: А — миноги; Б — зародыша селахии; В — взрослой
селахии; Г — рыбы. Схема позвонка: Д — миноги; Е, Ж — селахии; З —
рыбы; И —рептилии; К—млекопитающего; 1 — передняя и 2— задняя дорсальные дуги; 3 — миосепты; 4 — дорсальный и 5 — вентральный нервные
корешки; 6 —хорда; 7— оболочка хорды; 8 — вентральные дуги; 9 — ребро;
10— сосудистые отростки; 11— поперечный отросток позвонка; 12 — тело
позвонка; 13 — дужка позвонка; 14 — отверстие позвонка; 15 — остистый
отросток; 16 — суставной отросток.
С усложнением организации животных, увеличением активности и разнообразия движений вокруг хорды развиваются не только дуги, но и тела позвонков, в результате она оказывается сдавленной, а у многих взрослых
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
форм и в различной степени редуцированной (Б, В, Г). Постепенное замещение хорды позвонками можно проследить, сравнивая строение осевого
скелета у хрящевых и костистых рыб. У хрящевых рыб весь скелет образован
хрящом, иногда обызвествленным. Кроме верхних дуг, также метамерно под
хордой у них развиваются нижние дуги. Концы верхних дуг каждого сегмента, сливаясь, образуют остистый отросток. Появляются тела позвонков в виде
двояковогнутых дисков (амфицельные) с отверстием в центре (Е, Ж). В канале, образованном отверстиями тел позвонков, проходит хорда, которая теперь теряет значение опорного стержня. Она принимает четкообразную форму с сужениями в телах позвонков и расширениями между позвонками. Появляются зачаточные ребра.
У некоторых видов хрящевых и костных (кистеперых, двоякодышащих,
осетровых) рыб тела позвонков не развиваются, ребра отсутствуют. У всех
костистых рыб происходит замещение хрящевого скелета костным, позвонки
амфицельного типа с хорошо развитыми телами, дорсальными дугами и
длинными остистыми отростками. Развиваются поперечные отростки, к которым причленяются хорошо развитые длинные ребра. Появляются суставные отростки, которыми позвонки сочленяются друг с другом, чем обеспечивается прочность осевого скелета при сохранении его подвижности. Осевой
скелет четко разделяется на головной, туловищный с ребрами, охватывающими полость тела с органами, и сильно развитый хвостовой отдел — локомоторный.
Предки современных наземных позвоночных, по-видимому, произошли
от древних кистеперых рыб, о чем говорит строение их ископаемых остатков.
Переход к наземному образу жизни приводит к прогрессивному развитию
одних частей скелета и редукции других. Скелет туловища дифференцируется на шейный, грудной (спинной), поясничный и крестцовый отделы, частично редуцируется скелет хвоста, так как основная нагрузка при движении
по земле падает на конечности (кроме безногих амфибий и рептилий). В
грудном отделе в тесной связи с ребрами развивается грудина, формируется
грудная клетка.
Постепенность усложнения стволового скелета видна при сравнении
классов наземных позвоночных. У земноводных шейный и крестцовый отделы позвоночника имеют только по одному позвонку, поясничный отдел отсутствует. Ребра очень короткие, у многих срастаются с поперечными отростками позвонков. Общее число позвонков сильно различается у амфибий
разных отрядов: от 21 у бесхвостых до 300 у безногих. Тела их спереди вогнутые, сзади выпуклые (процельные). У грудины нет связи с ребрами, грудная клетка не образуется.
У рептилий шейный отдел удлиняется до восьми позвонков и приобретает большую подвижность. Два первых позвонка атлант и осевой (эпистрофей) сильно отличаются от остальных, к трем последним причленяются шейные ребра. Позвонки процельные (И). В грудном отделе 1—5 пар ребер соединены с грудиной — образуется грудная клетка. Поясничный отдел длинный, имеет ребра, величина которых уменьшается в каудальном направwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лении. Крестцовый отдел образован двумя позвонками, хвостовой отдел
длинный, хорошо развит.
Предками млекопитающих, по-видимому, были мелкие зверозубые пресмыкающиеся (величиной с крысу или кролика) пермского периода, которые
наряду с крайне примитивными признаками имели ряд черт сходства с млекопитающими. У млекопитающих, независимо от образа жизни, постоянное
число шейных позвонков, равное 7. Исключение составляют ламантин и двупалый ленивец, имеющие 6 шейных позвонков, а трехпалый ленивец — 8—
10. Относительно постоянно число позвонков и в остальных отделах: 12—19
грудных, 5—7 поясничных, 3—9 крестцовых. Хвостовых позвонков насчитывают от 3 до 46. Позвонки, за исключением первых двух, соединены при
помощи хрящевых дисков (менисков), связок и суставных отростков.
Поверхности тел шейных позвонков часто имеют выпукло-вогнутую
форму — опистоцельные (К). В остальных отделах позвонки обычно плоские
— платицельные.
Ребра сохраняются только в грудном отделе. В пояснице они редуцируются и срастаются с поперечными отростками позвонков. В крестцовом
отделе срастаются и позвонки, образуя крестцовую кость. Хвостовой отдел
облегчен, его позвонки сильно редуцируются.
Филогенез скелета головы (рис. 42). Скелет головного конца тела развивается вокруг нервной трубки — осевой (мозговой) скелет головы и вокруг
головной кишки — висцеральный. У примитивных хордовых (круглоротые)
они не связаны друг с другом. Осевой скелет головы представлен хрящевыми
пластинками, окружающими нервную трубку снизу и с боков, крыша черепа
перепончатая. Висцеральный скелет головы состоит из хрящевых жаберных
дуг, связанных с аппаратом дыхания и пищеварения; челюстей нет.
Филогенетическое развитие скелета головы шло путем объединения
мозгового и висцерального скелетов и усложнения их строения в связи с прогрессивным развитием головного мозга, органов чувств, расположенных на
головном конце тела (обоняния, зрения, слуха), преобразованиями органов
захвата и удержания пищи и дыхательного аппарата. Мозговой череп хрящевых рыб представляет собой сплошную хрящевую коробку, окружающую
головной мозг. Висцеральный скелет образован хрящевыми жаберными дугами, которые подобно ребрам в туловище опоясывают собой головную
часть кишечного канала. Передние висцеральные дуги превратились в губные хрящи, челюстную и подъязычную дуги.
Черепная коробка у костистых рыб сложного строения. Наряду с первичными костями, появившимися в большом количестве, на месте хрящевого
черепа развиваются покровные кости. Первичные кости формируют затылочную область, часть основания черепа, обонятельную и слуховую капсулы и
стенку глазницы. Покровные кости покрывают первичную черепную коробку
сверху, снизу и с боков. Висцеральный скелет костистых рыб построен из
большого количества первичных и вторичных костей и представляет собой
очень сложную систему рычагов, участвующих в хватательных, глотательных и дыхательных движениях. С черепной коробкой висцеральный скелет
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
сочленяется при помощи подвеска (hyomandibulare), в результате чего образуется единый скелет головы. С помощью костей плечевого пояса он неподвижно соединяется со стволовым скелетом.
Рис. 42. Филогенетические преобразования скелета головы (покровные
кости белые, первичные кости и хрящи, из которых они развились, в
точку):
А—селахии; Б — костистой рыбы; В — млекопитающего: а — обонятельная, б — глазничная, в — слуховая и г — затылочная области черепа;1
— губные хрящи; 2 — нёбно-квадратный хрящ; 2' — квадратная кость; 2" —
наковальня; 3— челюстной хрящ; 3' — сочленовная кость; 3" — молоточек;
4 — подъязычночелюстной хрящ; 4' — подвесок; 4" — стремечко; 5 — подъязычный хрящ; 5" — подъязычная; 6 — носовая; 7 — лобная; 8 — теменная;
9 — межтеменная; 10—затылочная; 11 — височная; 12 — клиновидная и 13
— слезная кости; 14 — сошник; 15 — крыловидная. 16—нижнечелюстная, 17
— нёбная; 18 — решетчатая; 19 — верхнечелюстная и 20 — резцовая кости;
I—V — жаберные дуги; Г — тело подъязычной кости; 1Г—IIГ — щитовидный хрящ; IV—V — остальные хрящи гортани.
С выходом на сушу, с резкой сменой среды обитания, а следовательно, и
образа жизни животных происходят значительные изменения в скелете головы. Череп теряет связь с плечевым поясом и подвижно причленяется к шейwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ному отделу. Уменьшается количество костей черепа за счет их сращения,
увеличивается его прочность. Смена типа дыхания (с жаберного на легочное)
приводит к редукции жаберного аппарата и преобразованию его элементов, а
также некоторых покровных костей висцерального скелета, в подъязычную и
слуховые кости.
Преобразования в строении черепа приводят к срастанию челюстного
аппарата с основанием черепа, к появлению слуховых косточек в обособившейся полости среднего уха, к объединению обонятельной капсулы с носовой полостью, к разделению носовой и ротовой полостей с помощью твердого нёба и изменению характера причленения нижней челюсти к осевому черепу.
В ряду наземных хордовых животных прослеживается постепенность
этих усложнений. В черепе взрослых земноводных много хрящей, слуховая
кость одна — стремечко (столбик). По числу костей черепа рептилии скорее
приближаются к рыбам, чем к амфибиям, однако строение черепа типично
для наземных животных. Для черепа млекопитающих характерно прогрессивное развитие, выраженное в уменьшении количества костей за счет их
сращения (например, затылочная кость образуется путем сращения 4, а каменистая— 5 костей), в стирании граней между первичными и покровными
(вторичными) костями, в мощном развитии обонятельной области и сложном
звукопроводящем аппарате, в крупных размерах черепной коробки, резко
выраженной разнозубости, самостоятельном причленении нижней челюсти к
слуховой области черепа (без каких-либо промежуточных костей).
Филогенез скелета конечностей (рис. 43). Гипотеза о происхождении
конечностей наземных животных на базе парных плавников рыб оказалась
плодотворной и сейчас имеет широкое признание. Парные плавники в типе
хордовых впервые появились у рыб (Л). У круглоротых они отсутствуют,
есть, как и у ланцетника, только непарные плавники. Костной основой парных плавников рыб служит целая система хрящевых и костных элементов,
которые могут быть подразделены на несколько отделов. Самый проксимальный отдел, называемый поясом грудных плавников, у хрящевых рыб
имеет вид изогнутой пластинки, прилежащей к осевому скелету и соединяющей правый и левый плавники в единую систему. У костистых рыб он более
сложной формы и прочнее связан с осевым скелетом. Среди его костей есть
две небольшие кости, гомологичные костям плечевого пояса наземных животных: лопатка — лежащая на спинной стороне и коракоид — на брюшной.
Тазовый пояс у рыб развит в меньшей степени. Имеет вид хрящевой или
костной пластинки, лежащей в мускулатуре брюшной стенки. К его боковой
поверхности причленяется скелет брюшного плавника. У современных лучеперых рыб плавники претерпели редукцию и видоизменение в сравнении с
костными гоноидами и двудышащими. По строению к конечностям наземных позвоночных ближе всего стоят плавники и их пояса у ископаемых кистеперых рыб (Б).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 43. Филогенетическое преобразование скелета конечностей:
А —парный грудной плавник селахии; Б — грудной плавник кистеперой
рыбы; В — скелет грудной конечности примитивного наземного позвоночного; Г — скелет грудной конечности стопоходящего млекопитающего (медведя); Д — изменения кисти у предков лошади: а — эогиппус; б — мезогиппус; в — гиппогиппус; г — гиппарион; д — современная лошадь; 1—плечевой
пояс: 2 — базалии; 3— радиалии; 4 — лопатка; 5 — ключица; 6 — коракоидная кость; 7 — плечевая кость; 8 — кости предплечья; 9 — кости кисти;
10— кости запястья; 11 — кости пясти; 12 — кости пальцев.
С выходом на сушу полностью исчезает скелет непарных плавников. На
базе парных плавников развивается скелет конечностей, расчлененный на отделы, типичные для пятипалой конечности (В). Пояса конечностей состоят из
трех пар костей и укрепляются связью с осевым скелетом: плечевой пояс — с
грудиной, тазовый с крестцом. Плечевой пояс состоит из коракоида, лопатки
и ключицы, тазовый — из подвздошной, лонной и седалищной костей. Скелет свободных конечностей расчленен на три отдела: у передней конечности
— это кости плеча, предплечья и кисти, у задней — бедра, голени и стопы.
Число костей в гомодинамных звеньях конечностей одинаково и увеличивается в дистальном направлении.
Дальнейшие филогенетические преобразования связаны с характером
передвижения, его скоростью и маневренностью. У земноводных пояс грудных конечностей, причленяясь к грудине, не имеет жесткой связи с осевым
скелетом. В поясе тазовых конечностей развита в основном его вентральная
часть. Причленяется он к единственному крестцовому позвонку. Свободные
конечности присоединяются к поясам в сегментальной плоскости, так что
плечо и бедро отходит от тела под прямым углом, а лапы направлены латероwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
каудально. У рептилий в скелете поясов в равной степени развиты дорсальная и вентральная части. Свободные конечности отходят от тела, как и у амфибий, под прямым углом, но их дистальные звенья развернуты, особенно на
грудной конечности, в латеро-краниальном направлении.
Плечевой пояс млекопитающих (Г) сильно редуцирован, особенно его
вентральное звено. Он состоит из двух или даже одной кости. У животных с
развитыми отводящими движениями грудной конечности (например, кротов,
летучих мышей, обезьян) развиты лопатка и ключица, а у животных с однообразными движениями (например, у копытных)—только лопатка. Тазовый
пояс млекопитающих упрочен тем, что лонные и седалищные кости соединяются вентрально с соименными костями. Скелет свободных конечностей
млекопитающих развернут в сагиттальной плоскости, лапы направлены краниально. Построен он по типу пятипалой конечности, с длинными проксимальными звеньями, в результате чего тело животного высоко поднято над
землей. Приспособление же к различным видам передвижения (бег, лазанье,
прыжки, полет, плавание) привело к сильной специализации конечностей у
разных групп млекопитающих, что выражается в основном в изменении длины и угла наклона отдельных звеньев конечностей, формы суставных поверхностей, срастании костей и редукции пальцев.
Изменение в строении конечностей в филогенезе в связи с повышением
специализации — приспособленностью к определенному виду движения подробнее всего изучено в ряду лошадей (В. О. Ковалевский). Предполагаемый
предок лошади — эупротогония, совмещая в себе черты копытных и хищных, была размером с лисицу и имела пятипалые конечности с когтями, по
форме приближающимися к копытцам. На протяжении нескольких периодов
(от нижнего эоцена до нижнего плиоцена) происходила постепенная смена
условий существования и параллельно этому шло приспособительное изменение структуры конечностей, связанное с изменением характера движения
(Д). От разнообразных мягких движений по рыхлой земле с высокой растительностью (эоценовый лес) до широких размашистых быстрых движений по
сухим открытым пространствам (миоценовая степь). При этом происходило
удлинение основного поддерживающего столба конечностей из-за раскрытия
(увеличения) углов между ее звеньями. Лапа приподнималась, животное переходило от стопо- к пальцехождению (эогиппус — четырехпалый, мезогиппус—трехпалый). При этом наблюдалась постепенная редукция нефункционирующих пальцев: первого у эогиппуса, первого и пятого у мезогиппуса.
При переходе от пальце- к фаланго-(копыто-) хождению в состав основного
поддерживающего столба входит уже вся лапа, а редукция пальцев достигает
максимума. У лошади полноразвитым сохраняется на конечности лишь III
палец. У рогатого скота развитыми оказываются два пальца — III и IV.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ОНТОГЕНЕЗ СКЕЛЕТА
В процессе индивидуального развития особи — в онтогенезе — скелет в
общих чертах проходит те же три стадии развития и в той же последовательности, что и в филогенезе: соединительнотканного, хрящевого и костного скелета.
Хорда как один из первых осевых органов закладывается в зародышевый
период внутриутробного развития в результате дифференцировки энтодермы
и мезодермы в период гаструляции. Вскоре вокруг нее образуется сегментированная мезодерма — сомиты, внутренняя часть которых — склеротомы,
прилежащие к хорде, являются скелетогенными зачатками.
Соединительнотканная стадия. В области склеротомов идет активное
размножение клеток, которые принимают вид мезенхимных, разрастаются
вокруг хорды и превращаются в ее соединительнотканный футляр и в миосепты — соединительнотканные тяжи, метамерно отходящие от футляра
хорды. Соединительнотканный скелет у млекопитающих существует очень
короткое время, так как параллельно процессу обрастания хорды в перепончатом скелете происходит размножение мезенхимных клеток, особенно вокруг миосепт, и дифференцировка их в хрящевые.
Хрящевая стадия. Дифференцировка мезенхимных клеток в хрящевые
начинается с шейной области. Первыми закладываются хрящевые дуги позвонков, которые образуются между хордой и спинным мозгом, обрастают
спинной мозг сбоку и сверху, формируя его футляр. Смыкаясь между собой
попарно над спинным мозгом, дуги формируют остистый отросток. В это же
время из сгущений мезенхимных клеток, размножающихся в футляре хорды,
развиваются хрящевые тела позвонков, а в миосептах — хрящевые зачатки
ребер и грудины. Процесс замещения соединительной ткани хрящом начинается у свиньи и овцы на 5-й, у лошади и крупного рогатого скота — на 6-й
неделе эмбрионального развития. Затем в такой же последовательности, в какой шло образование хрящевого скелета, идет его окостенение.
Развитие кости на месте хряща (рис. 44). В хрящевой закладке (модели)
кости сосудов нет. С развитием кровеносной системы зародыша идет образование сосудов вокруг и внутри надхрящницы, в результате чего ее клетки
начинают дифференцироваться не в хондробласты, а в остеобласты, то есть
она становится надкостницей—периостом. Остеобласты продуцируют межклеточное вещество и откладывают его поверх хрящевого зачатка кости.
Формируется костная манжетка. У овец это происходит на 45-м дне внутриутробного развития. Костная манжетка построена из грубоволокнистой костной ткани. Процесс образования и роста манжетки вокруг хрящевого зачатка называется перихондральным окостенением. Костная манжетка затрудняет питание хряща и начинает разрушаться. Первые очаги обызвествления и деструкции хряща обнаруживают в центре (диафизе) хрящевого зачатка, где клетки вакуолизируются, набухают, ядра их сморщиваются. В очаг
разрушающегося хряща от надкостницы проникают сосуды вместе с недифференцированными клетками. Здесь они размножаются и превращаются в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
костные клетки — возникает первый очаг (центр) окостенения. У каждой
кости обычно несколько очагов окостенения (в позвонках копытных их насчитывают 5—6, в ребрах — 1—3).
В очаге окостенения остеокласты разрушают обызвествленный хрящ,
образуя лакуны и туннели, шириной 50—800 мкм. Остеобласты вырабатывают межклеточное вещество, которое откладывается по стенкам лакун и
туннелей. Мезенхима, проникающая вместе с капиллярами, дает начало следующей генерации остеобластов, которые, откладывая межклеточное вещество в сторону стенок туннелей, замуровывают предыдущие генерации остеобластов — развиваются костные пластинки. Поскольку лакуны и туннели
образуют сеть, костная ткань, выстилающая их,
повторяет их форму и в целом напоминает губку,
состоящую из переплетающихся костных тяжей,
перекладин или трабекул. Из них формируется
губчатое вещество кости. Образование кости
внутри хрящевого зачатка на месте разрушенного хряща называется эндохондральным (энхондральным) окостенением. У плодов овец оно начинается с 49-го дня.
Часть недифференцированных клеток, проникающих вместе с капиллярами в туннели и лакуны, превращается в клетки костного мозга (у
овец с 52-го дня), который по мере развития заполняет пространства между костными трабекулами губчатого вещества.
Процесс энхондрального окостенения, начавшись в области диафиза, распространяется к
концам зачатка — эпифизам. Параллельно с
этим утолщается и нарастает костная манжетка.
В таких условиях хрящевая ткань может расти
Рис. 44. Развитие кости на только в продольном направлении. При этом
месте хряща:
хондробласты, размножаясь, выстраиваются
1 — надкостница; 2 —
друг над другом в виде клеточных колонок (моперихондральная манжетка; 3 — лакуны, запол- нетных столбиков).
Закладка хрящевых моделей и их окостенененные мезенхимой; 4 — энхондральная кость; 5 — зо- ние происходят быстро в тех участках тела, где
на обызвествления и разру- потребность в опоре появляется очень рано. У
шения хряща; 6 — клеточ- разных животных эти сроки неодинаковы (обуные колонии; 7 — неразрусловлены их экологией) и прямо связаны со стешенный гиалиновый хрящ.
пенью зрелорождаемости. По срокам закладки и
темпам дифференцировки костного скелета млекопитающих можно разделить на несколько групп. Копытные относятся к
группе, у которой закладка и формирование очагов окостенения почти заканwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
чиваются к моменту рождения, 90% кости образовано костной тканью. После
рождения продолжается лишь рост этих очагов. Новорожденные таких животных активны, сразу могут самостоятельно передвигаться, следовать за матерью и добывать себе корм.
Первичные очаги окостенения в предплодный период отмечают в скелете туловища у свиньи на 5—6-й, у крупного рогатого скота и лошади — на
7— 8-й неделе эмбрионального развития. У рогатого скота вначале окостеневают ребра. Окостенение позвонков начинается с дуги атланта и распространяется в каудальном направлении. Тела окостеневают прежде всего у средних грудных позвонков. У 4,5—5-месячных плодов телят тела костей представляют собой лакунарные поля, выстланные тонкими костными пластинками первичных остеонов. Во второй половине эмбрионального развития активно образуются остеоны и у новорожденных телят средняя часть кости
представляет собой остеонный слой с остатками лакун. Выделяются слои наружных и внутренних генеральных пластинок. Периостальная поверхность
ребристая. В постнатальном онтогенезе идет нарастание новых слоев костной
ткани до завершения роста животного, а также перестройка уже существующих остеонов.
Зона клеточных колонок постоянно нарастает со стороны эпифизов благодаря дифференцировке хрящевых клеток от надхрящницы. Со стороны же
диафиза отмечается постоянное разрушение хряща из-за нарушения его питания и изменения химизма ткани. До тех пор, пока эти процессы уравновешивают друг друга, кость растет в длину. После закладки вторичных очагов
окостенения (в эпифизах и апофизах) хрящ оказывается расположен между
двумя фронтами окостенения. Называется он эпифизарным (метаэпифизарным), считается источником роста трубчатых костей в длину. Когда же скорость энхондрального окостенения становится больше, чем скорость роста
метаэпифизарного хряща, он (у 8—9-месячных баранов) истончается и, наконец, полностью исчезает. С этого времени прекращается линейный рост
животного. В осевом скелете дольше всего сохраняются хрящи между эпифизами (головкой и ямкой) и телом позвонков, особенно в крестце.
Рост кости в ширину происходит благодаря перестройке энхондральной
и перихондральной кости. В энхондральной кости этот процесс начинается
из середины диафиза и выражается в росте, разрушении старых и образовании новых остеонов, в формировании костной полости. В перихондральной кости перестройка заключается в том, что грубоволокнистая костная
ткань манжетки замещается нарастающей со стороны периоста пластинчатой
костной тканью в виде остеонов, циркулярно-параллельных структур и генеральных пластинок, составляющих в совокупности компактное вещество
кости. В процессе перестройки образуются вставочные пластинки. Это остатки старых, частично разрушенных остеонов, циркулярно-параллельных
структур и генеральных пластинок, расположенных между молодыми остеонами. У крупного рогатого скота и свиньи осевой скелет начинает окостеневать в 3—4 года, а полностью процесс завершается в 5—7 лет и даже в 10 лет
(лицевые кости), у лошади — в 4—5 лет, у овцы — в 3—4 года.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Развитие черепа. Начало осевому черепу дают 7—9 сомитов, которые
закладываются в области головы. Вокруг конечного участка хорды склеротомы этих сомитов образуют сплошную перепончатую пластинку без следов
сегментации (рис. 45). Эта пластинка распространяется вперед дальше хорды
(прехордально) и охватывает снизу и с боков мозговые пузыри, слуховые и
обонятельные капсулы и глазные бокалы. Замещение соединительнотканного
осевого черепа хрящевым начинается около переднего конца хорды под основанием головного мозга. Здесь закладывается пара околохордовых (парохордалии) хрящей. Дальше в оральном направлении закладываются две хрящевые балки или трабекулы. Так как они лежат впереди хорды, этот отдел
осевого черепа называется прехордальным. Вскоре трабекулы и парахордалии, разрастаясь, сливаются вместе, образую основную хрящевую пластинку. К трабекулам прирастают охрящевевшие обонятельные капсулы, а к
парахордалиям — слуховые капсулы. В оральной части вдоль основной хрящевой пластинки закладывается хрящевая носовая перегородка, по обе стороны от которой развиваются носовые раковины. Затем хрящи замещаются
первичными, или примордиальными, костями. Начинается это с закладки отдельных очагов окостенения на 5-й неделе у свиньи, 6-й у овцы, 7—8-й неделе у эмбриона крупного рогатого скота и лошади. Первичными костями осевого черепа являются затылочная, клиновидная, каменистая и решетчатая,
образующие дно, переднюю и заднюю стенки черепномозговой полости, а
также носовая перегородка и раковины. Остальные кости вторичные, кожные, или покровные, так как возникают из мезенхимы, минуя хрящевую стадию. Это теменные, межтеменные, лобные, височные (чешуя), формирующие
крышу и боковые стенки черепномозговой полости (см. рис. 42).
Параллельно развитию осевого черепа идет преобразование висцерального скелета головы. Из парных сегментированных спланхнотомов головной
мезодермы закладываются соединительнотканные зачатки висцеральных (жаберных) дуг. Большая часть зачатков висцеральных дуг подвергается полной
редукции, а часть их материала идет на образование слуховых косточек,
подъязычной кости и хрящей гортани. Основная же масса костей висцерального скелета — вторичные, покровные. Осевой и висцеральный скелет головы млекопитающих в процессе развития настолько тесно связывается друг с
другом, что кости одного входят в состав другого. Поэтому череп млекопитающих независимо от происхождения костей делят на мозговой отдел (собственно череп), являющийся вместилищем мозга, и лицевой отдел (лицо),
образующий стенки носовой и ротовой полостей. В плодный период определяется форма черепа, характерная для вида и породы. Роднички — неокостеневшие участки — закрываются плотной соединительной тканью или хрящом.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 45. Развитие черепа.
Схема строения черепа на хрящевой стадии развития:
А — вид сбоку; Б — вид сверху; В — череп зародыша свиньи (вид сверху
со снятой крышкой): 1 — ноздря; 2 — носовая ямка; 3 — носовая капсула; 4
— трабекула; 5 — передний мозг; 6 — глаз;. 7 — глазничный хрящ; 8 — зрительный нерв; 9 — отверстие зрительного нерва; 10— средний мозг; 11 —
гипофиз; 12— отверстие гипофиза; 13 — ямка гипофиза; 14 — задний мозг;
15 — слуховая капсула; 16 — ямка мозгового моста (средний мозг); 17— ямка продолговатого мозга (задний мозг); 18 — парахордальный хрящ; 19 —
спинной мозг; 20 — большое затылочное отверстие; 21 — хорда; 22 — склеротомы; 23— хрящевые закладки позвонков; 24 — закладки хрящей висцерального черепа; 25 — кишечная трубка.
Развитие конечностей. Конечности у млекопитающих закладываются в
виде каплеобразных выростов шейно-грудных и пояснично-крестцовых сомитов. У крупного рогатого скота это происходит на 3—4-й неделе. Сегментация их не выражена. Закладки имеют вид скоплений мезенхимы, которые
быстро увеличиваются в длину, превращаясь в лопастевидные выросты. Сначала эти выросты подразделяются на два звена: закладку поясов и свободных
конечностей, не расчлененных на отделы и кости. Затем из сгущений мезенхимы дифференцируются соединительнотканные и хрящевые закладки костей, что у зародыша теленка видно на 6-й неделе. В процессе дифференцировки скелет конечностей проходит те же три стадии, что и стволовой скелет, но с некоторым отставанием. Окостенение конечностей у плода теленка
начинается на 8—9-й неделе и протекает аналогично со стволовым скелетом.
Каждая кость развивается не менее чем из трех очагов окостенения. Многие
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
выросты костей — апофизы имеют собственные очаги окостенения. В процессе окостенения в трубчатых костях образуется губчатое и компактное вещество. Перестройка из центра кости распространяется на ее периферию.
При этом в области диафиза благодаря деятельности остеокластов почти
полностью исчезает губчатое вещество, сохраняясь лишь в эпифизах. Увеличивается костная полость. Красный костный мозг в ней становится желтым.
Слои компактного вещества становятся заметны в течение первых месяцев жизни. Степень его развития зависит от вида животного. У копытных в
нем хорошо развиты генеральные пластинки и циркулярно-параллельные
структуры, у хищных преобладают остеоны. Это связано с различиями в
функциональных нагрузках костей, особенно конечностей. У копытных они
приспособлены к прямолинейному движению и удержанию массивного тела,
у хищных — к более легкому телу и разнообразным движениям.
В конечностях очаги окостенения появляются прежде всего в костях
поясов, распространяясь затем в дистальном направлении. Окончательное же
окостенение (синостоз) прежде всего происходит в дистальных звеньях. Так,
у крупного рогатого скота окостенение дистальных звеньев конечности
(плюсны и пясти) завершается к 2—2,5 годам, к 3—3,5 годам окостеневают
все кости свободной конечности, а кости тазового пояса — лишь к 7 годам.
По срокам и темпам окостенения скелета самки несколько опережаю.т самцов, за исключением симфиза тазовых костей.
Возрастные изменения скелета. В связи с различиями в сроках закладки,
скорости роста и окостенения костей скелета на протяжении онтогенеза происходит закономерное изменение пропорций тела животного. В период эмбрионального развития кости растут с разной интенсивностью. У копытных в
первую половину интенсивнее растет осевой скелет, во вторую — скелет конечностей. Так, у 2-месячных плодов телят осевой скелет составляет 77%,
скелет конечностей — 23%, а к рождению — соответственно 39 и 61%. По
данным Н. Н. Третьякова, у советского мериноса от времени хрящевой закладки (1-месячный эмбрион) до рождения скелет тазовой конечности с поясом увеличивается в 200 раз, грудной конечности — в 181 раз, таз — в 74
раза, позвоночник — в 30 раз, череп — в 24 раза. В результате молодняк копытных отличается высоконогостью, коротким туловищем и относительно
широкой головой с коротким лицевым отделом. После рождения на смену
усиленному росту периферического скелета приходит линейный рост осевого
скелета. Интенсивно растет лицевой отдел черепа, в ряде костей появляются
синусы. Выступы, гребни, шероховатости и отростки костей увеличиваются,
что приводит к более выраженному рельефу тела, свойственному виду и породе.
В постнатальном онтогенезе скелет растет с меньшей скоростью, чем
мышцы и многие внутренние органы, поэтому его относительная масса снижается с 23—25% у новорожденного теленка, с 30% у жеребенка и поросенка, с 18% у ягненка, с 15% у кролика до 10—15%, у взрослой коровы, до 13%
у лошади, до 7% у овцы, свиньи, до 8% у кролика.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
В процессе роста и дифференцировки костей увеличивается их прочность, что связано с изменением внутренней структуры (увеличивается число
остеонов на единицу площади), химизма (количество кристаллической фазы
повышается в онтогенезе в 9 раз), толщины компакты и соотношения компакты с костной полостью. От рождения до взрослого состояния толщина
компактного вещества увеличивается в 3—4 раза (у овец с 2 до 6 мм, у коров
до 9 мм), содержание в нем минеральных солей — в 5 раз, предельная нагрузка — в 3—4 раза, достигая у овец 280, у коров 1000 кг на 1 см2. Толщина
компакты тем больше, чем большие нагрузки приходятся на данный участок.
Например, в плечевой кости овец это задняя сторона, в лучевой и пястной —
медиальная сторона костей. Окончательной прочности кости крупного рогатого скота достигают уже к 12-месячному возрасту.
Отмечена интересная закономерность: чем крупнее животное, тем меньше у него запас прочности костей, который исчисляется как отношение предельной прочности кости к массе животного. У самцов кости, как правило,
толще, чем у самок, но недокорм сказывается на них сильнее.
Улучшенные породы овец и свиней имеют более короткие и широкие
кости конечностей, особенно пясть и плюсну, укороченные остистые отростки позвонков, более широкую с укороченным лицевым отделом голову. У
скороспелых животных кости толще, чем у одновозрастных позднеспелых.
Кости коров молочного типа лучше снабжаются кровью. В то же время у коров мясного и мясо-молочного типов больше площадь компактного вещества
кости и толщина ее стенки, что обусловливает большую прочность кости при
статических нагрузках. Прочность кости на изгиб обусловливает структура
остеонов. У свиней породы ландрас, например, прочность кости на изгиб
выше, чем у крупной белой и сибирской северной пород, в связи с тем что у
ландрас более плотное расположение остеонов, большее количество костных
пластинок, уже каналы остеонов.
Из всех внешних условий на развитие скелета наибольшее влияние оказывает кормление и моцион. Улучшение кормления в период интенсивного
роста костей ускоряет, недокорм угнетает их темпы роста, особенно в ширину, но не нарушает общих закономерностей роста скелета. У животных пастбищного содержания компактное вещество кости плотнее, в нем преобладают пластинчатые структуры, трабекулы губчатого вещества толще, равномернее по ширине и направлены строго по действию сил сжатия — растяжения. Метаэпифизарный хрящ лучше выражен, перестройка и нарастание
слоев костной ткани идут активнее, а резорбция — медленнее, чем у животных стойлового содержания. При стойловом и клеточном содержании животных замедляется рост и внутренняя перестройка костей, снижается их плотность и прочность по сравнению с выгульным, напольным содержанием и с
животными, подвергающимися дозированному принудительному движению.
Добавление к рациону молодняка макро- и микроэлементов способствует образованию костей с более толстым компактным веществом и трабекулами и меньшей костной полостью. При недостатке минеральных веществ
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
происходит деминерализация скелета, размягчение и резорбция позвонков,
начиная с хвостовых.
СТРОЕНИЕ ОСЕВОГО СКЕЛЕТА
В состав осевого скелета входит скелет головы и ствола тела (см. рис.
39). Стволовой скелет образован костями шеи, туловища (грудной клетки,
поясницы, крестца) и хвоста. Состоит он из костных сегментов, которые полностью развиты лишь в передней части грудного отдела туловища. Такой
сегмент состоит из позвонка, пары ребер и сегмента грудины. В краниальном
и каудальном направлениях костный сегмент подвергается редукции: отсутствуют сегменты грудины, редуцируются ребра и их остатки прирастают к
позвонкам. В хвостовом и шейном отделах редуцируются и позвонки, так что
у первого шейного позвонка отсутствует тело, а у большей части хвостовых
— дуга. Все позвонки стволового скелета образуют позвоночный столб или
позвоночник, внутри которого, в позвоночном канале, залегает спинной мозг.
У позвоночника млекопитающих три характерных изгиба: шейный, грудной
и поясничный. Шейный изгиб образуется между головой и двумя первыми
шейными позвонками. Грудной изгиб — между последним шейным и первым грудным позвонками, поясничный изгиб находится в области поясницы.
Изгибы увеличивают рессорные свойства позвоночника. Шейный и грудной
изгибы, кроме того, являются приспособительными рычагами для удержания
на весу головы и обеспечения более широкого обзора. Здесь нужна подвижность, которая обеспечивается особенностями в строении позвонков (даны
при описании скелета шеи).
Подвижность позвоночника снижается в каудальном направлении, так
как для туловища важно прежде всего удержание тяжелых внутренностей и
передача энергии движения с задних конечностей на передние. Туловищный
отдел позвоночника по своим механическим свойствам напоминает арку
моста, противодействующую переразгибанию. Особенно большое значение
для прочности туловищного отдела имеет холка. Остистые отростки позвонков области холки, налегая один на другой, удерживают позвоночник от переразгибания. Чем они длиннее и шире, тем больше и крепче оказываются
рычаги, к которым прикрепляются мышцы спины и плечевого пояса. Короткая поясница улучшает механические свойства скелета туловища, длинная,
наоборот, ослабляет сопротивление переразгибанию и требует дополнительного упрочения, что мы увидим при изучении особенностей строения поясничных позвонков рогатого скота и свиньи. Для выполнения активной мышечной работы крестец в противоположность грудному и поясничному отделам желателен более длинный, так как на данный участок позвоночника при движении передается сила, развиваемая тазовыми конечностями.
Он же является рычагом, обеспечивающим перенесение силы тяжести для
облегчения передней половины тела при вынесении грудных конечностей
вперед. При этом скелет туловища впереди крестца считается плечом скорости, начиная с крестца — плечом силы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Скелет грудного отдела туловища (грудной клетки)
Скелет грудной клетки крупного рогатого скота состоит из 13—14 грудных позвонков, такого же количества пар ребер и грудины. У свиньи 13—15
(до 17), у лошади 17—19 (чаще 18 грудных позвонков) и такое же количество
пар ребер. В целом грудная клетка представляет собой конус с усеченной
вершиной и широким косым основанием. На вершине, обращенной вперед,
имеется краниальное грудное отверстие — вход в грудную полость, ограниченное со всех сторон первым костным сегментом грудного отдела: сверху
— позвонком, сбоку — ребрами, снизу — рукояткой грудины. Через краниальное отверстие в грудную полость проходят пищевод, трахея, крупные
кровеносные сосуды и нервы, а у молодого животного тимус. Выход из грудной клетки в брюшную полость — каудальное грудное отверстие гораздо
шире краниального. Оно ограничено последним грудным позвонком, последними ребрами и их хрящами, образующими реберную дугу, и затянуто
диафрагмой, в которой имеются отверстия для прохода пищевода, сосудов и
нервов.
Грудной позвонок — vertebra thoracica — по форме является смешанной симметричной костью и состоит из тела, дуги и отростков.
У крупного рогатого скота (рис. 46) длина тела грудного позвонка больше его ширины. На теле различают краниально направленную уплощенную
головку, каудально — неглубокую ямку. Головки позвонков становятся более
выпуклыми, а ямки — более глубокими у первых грудных позвонков. Справа
и слева на головке и ямке видны неглубокие вдавления— краниальные и каудальные реберные ямки. Краниальная и каудальная реберные ямки двух соседних позвонков формируют глубокую реберную ямку, в которую входит головка ребра. На дорсальной и боковых поверхностях тела позвонка видны отверстия для прохождения артерий внутрь кости и вен из нее. На вентральной
поверхности тела грудного позвонка виден вентральный гребень, лучше развитый на первых и последних позвонках в связи с наличием в этих участках
вентральной мускулатуры позвоночного столба.
Дорсально от тела позвонка поднимается дуга (дужка) позвонка, соединенная с телом ножками (корнями) дуги. Между телом и дугой образуется
пространство — отверстие позвонка. Отверстия всех позвонков позвоночника (исключая последние хвостовые) формируют позвоночный канал.
Спереди на ножках дуги имеется краниальная позвоночная вырезка, сзади — латеральное позвоночное отверстие. Между позвонками в этом месте
образуется межпозвоночное отверстие для выхода нервов и сосудов. По бокам от дуги отходят небольшие поперечные отростки с плоскими поперечнореберными ямками для сочленения с бугорком ребра. Вверх направлен остистый отросток, у основания которого на дуге спереди и сзади видно по
паре суставных краниальных и каудальных отростков в виде небольших
плоских фасеток, которыми соседние позвонки причленяются друг к другу.
Лишь у первого, второго и последних грудных позвонков суставные отростки
хорошо выражены. Остистые отростки длинные, в виде широких пластин с
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
острыми каудальными краями, сильно наклонены каудально. Особенно развиты остистые отростки на 2—6 позвонках, образующих остов холки. У 13го позвонка, который называется диафрагмальным, остистый отросток направлен прямо вверх. У расположенных за ним позвонков остистые отростки
наклонены краниально.
У свиньи тела грудных позвонков короткие и широкие, несколько
сплюснутые в дорсовентральном направлении, вентральный гребень не развит, головки и ямки плоские. Остистые отростки плоские и широкие. Самые
длинные, три первых, направлены несколько вперед, остальные — назад.
Диафрагмальный позвонок 11-й или 12-й.
На поперечных отростках имеется поперечное отверстие, идущее дорсовентрально.
Рис. 46. Грудной позвонок крупного рогатого скота:
1 — остистый отросток; 2 — позвоночная
дужка; 3 — поперечный отросток; 4 — поперечнореберная ямка; 5 — краниальный суставной отросток; 6 — краниальная позвоночная вырезка; 7 — краниальная реберная
ямка; 8 — головка позвонка; 9 — каудальный
суставной отросток; 10— латеральное позвоночное отверстие; 11 — каудальная реберная ямка; 12 — ямка позвонка; 13 — тело
позвонка; 14 — вентральный гребень.
Рис. 47. Ребро крупного рогатого скота:
I— реберная кость; II— реберный хрящ; 1 — головка; 2
— шейка; 3 — бугорок; 4 —
мышечный желоб; 5 — угол
ребра; 6 — тело ребра.
У лошади тела грудных позвонков по форме напоминают трехгранную
призму из-за хорошо развитых вентральных гребней, особенно на пяти первых и на трех последних позвонках. Вместо латерального позвоночного отверстия — каудальная позвоночная вырезка, которая вместе с краниальной
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
позвоночной вырезкой формирует межпозвоночное отверстие. Остистые отростки высокие, с расширенным задним краем и булавовидным расширением
на верхнем конце. Высота их увеличивается до 4—5-го позвонка, затем постепенно снижается. Диафрагмальный позвонок обычно 15-й, но может быть
14-й и 16-й.
Ребро — costa — по форме является длинной изогнутой костью. Состоит из реберной кости и реберного хряща (рис. 47). Реберная кость имеет тело и два конца — дорсальный (вертебральный), обращенный к позвонкам, и
вентральный (стернальный)—обращенный к грудине. Ребра, концы которых
присоединяются к грудине, называются стернальными (истинными), которые до нее не доходят, а прикрепляются к хрящам впереди лежащих ребер —
астернальными (ложными).
У крупного рогатого скота на дорсальном конце ребра различают головку с суставными поверхностями для сочленения с глубокой реберной ямкой
позвонков и бугорок для сочленения с поперечнореберной ямкой позвонка.
Между головкой и бугорком имеется суженный участок — шейка ребра. Она
короткая на первом ребре, удлиняется на последующих и исчезает на последних ребрах, на которых головка и бугорок сливаются. Тело ребра плоское, широкое, особенно в его вентральной части. На нем различают выпуклую наружную и вогнутую внутреннюю поверхности, выпуклый каудальный
и вогнутый краниальный края. На каудальном крае ниже бугорка заметно небольшое возвышение — угол ребра. По телу ребра проходит два пологих желоба: с латеральной стороны около краниального края — мышечный желоб,
с медиальной стороны около каудального края — сосудистый желоб. Длина
реберной кости увеличивается до 7—9, затем резко уменьшается, а длина реберных хрящей, наоборот, становится больше. Стернальных ребер 8 пар. Их
хрящи сочленяются с реберными вырезками грудины. Со 2-й по 10-ю реберные кости сочленяются с реберными хрящами тугими суставами. Астернальные ребра имеют длинные тонкие хрящи, которыми они прикрепляются к
хрящам впереди лежащих ребер.
У свиньи ребра узкие, с хорошо выраженным углом, а потому имеют
форму запятой. Стернальных ребер 6—8 пар. Со 2-й по 7-ю реберные кости
причленяются к хрящам с помощью суставов.
У лошади ребра узкие, равномерной ширины, почти округлые в поперечном сечении, кроме первых; шейка ребра короткая, реберный угол выражен слабо, а сосудистый желоб — хорошо. Стернальных ребер 8 пар.
Грудина — sternum — образует вентральную стенку грудной клетки
(рис. 48). Состоит из тела, рукоятки и мечевидного хряща.
У крупного рогатого скота (А) тело грудины уплощено в дорсовентральном направлении и расширяется к каудальному концу. Образовано тело 7
грудинными сегментами, соединенными у молодых животных хрящевой тканью, у взрослых— костной. Впереди тела расположена рукоятка грудины
трехгранной формы. Краниальный конец ее приподнят вверх, каудальный
причленяется к телу с помощью сустава. На рукоятке имеется парное углубление — реберные вырезки для сочленения с хрящами первой пары ребер. По
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
бокам тела, между грудинными сегментами есть еще по 6 реберных вырезок
для сочленения с хрящами стернальных ребер. Шестая реберная вырезка
двойная — в нее входят хрящи 7—8-го ребер. Каудальнее тела имеется удлиненный мечевидный отросток, переходящий в широкий мечевидный хрящ
Рис. 48. Грудина крупного рогатого скота (А) и лошади (Б):
1 — рукоятка; 2 — тело (его дорсальная поверхность); 3— реберные
вырезки; 4 — мечевидный отросток; 5 — мечевидный хрящ; 6 — соколок; 7
— гребень грудины.
У свиньи грудина состоит из 6 сегментов, рукоятка длинная, направлена
вперед. На ней общая реберная вырезка для первой пары ребер. Мечевидный
хрящ короткий и неширокий.
У лошади (Б) тело грудины сдавлено с боков, состоит из 6 сегментов и
имеет 7 пар реберных вырезок. Рукоятка срастается с телом и несет на себе
хрящевой добавок — соколок, который в виде гребня продолжается по вентральной поверхности тела грудины. Мечевидный отросток очень короткий,
а его хрящ — широкий.
Скелет поясничного отдела туловища
Скелет поясницы образован у рогатого скота 6, а у лошади 5—6, у свиньи 5—7 поясничными позвонками. В поясничном отделе ребра редуцированы, а их дорсальные концы срослись с поперечными отростками — образовались поперечнореберные отростки. Реберные ямки в силу этого отсутствуют.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Поясничный позвонок — vertebra lumbalis — смешанная симметричная
кость, состоит из тела, дуги и отростков.
У крупного рогатого скота (рис. 49)
длина тела поясничного позвонка больше его
ширины,
головка
и
ямка
плоские,
вентральный гребень выражен хорошо на
первых
позвонках,
поперечнореберные
пластинчатые,
длинные,
Рис. 49. Поясничный по- отростки
звонок крупного рогатого расположены во фронтальной плоскости, а их
концы повернуты краниолатерально. Вместо
скота:
1 — тело; 2— ямка; 3 — латерального позвоночного отверстия может
вентральный гребень; 4 — быть глубокая каудальная позвоночная
позвоночная вырезка; 5 — вырезка. Краниальная позвоночная вырезка
поперечнореберный
от- пологая. Краниальные суставные отростки
росток; 6— краниальный имеют форму дуги, каудальные — цилиндсуставной отросток; 7 — рические. Такая форма суставных отростков
остистый отросток; 8 — способствует очень прочному соединению
отверстие позвонка; 9 — поясничных позвонков друг другом и с
каудальный суставной от- крестцовой костью. Остистые отростки
широкие пластинчатые, направлены вверх
росток.
или слегка отклонены вперед.
У свиньи тело позвонка более округло в поперечном сечении из-за слабого развития вентрального гребня, ширина его больше длины. У основания
поперечнореберных отростков имеется поперечное отверстие или вырезка, а
концы отростков направлены вентрально.
У лошади тело позвонка короче, чем у крупного рогатого скота, в поперечнике клиновидной формы благодаря хорошо развитому вентральному
гребню. Поперечнореберные отростки развернуты во фронтальной плоскости, прямые, на 2—3 последних отростках есть суставные площадки для более прочного соединения позвонков. Краниальные и каудальные позвоночные вырезки хорошо выражены. Краниальные суставные отростки имеют
слегка вогнутые суставные поверхности, каудальные — конической формы.
Остистые отростки высокие, вершины их направлены краниально.
Скелет крестцового отдела туловища
Скелет крестца составляет крестцовая кость (рис. 50) — os sacrum,
сросшаяся у крупного рогатого скота и лошади из 5, у овцы и свиньи из 3—4
крестцовых позвонков. Места сращения видны в виде поперечных линий на
вентральной стороне тела крестцовой кости, образованного из тел позвонков.
Остатки ребер сохранились только у первых двух сегментов крестцовой кости.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 50. Крестцовая кость:
А—крупного рогатого скота; Б — свиньи; В — лошади;
1 — головка; 2 — крыло; 3 — ушковидная поверхность; 4 — дорсальные и 5
—вентральные крестцовые отверстия; 6 — боковые части; 7 — тело; 8 —
краниальный суставной отросток; 9 — межкостное пространство; 10 —
гребень; междуговые пространства; 12— краниальные суставные поверхности.
У крупного рогатого скота (А) тело крестцовой кости выгнуто вверх в
виде дуги. На его краниальном конце имеется головка, на каудальном — ямка. Дуги крестцовых позвонков и их остистые отростки срослись, образовав
гребень крестца с утолщенным свободным краем. Между телом и гребнем
проходит крестцовый канал, сформированный позвоночными отверстиями.
Поперечнореберные отростки двух первых крестцовых сегментов срослись и
образовали крылья крестцовой кости, развернутые в дорсолатеральном направлении. На них есть шероховатая ушковидная поверхность для сочленения с тазовыми костями. Поперечные отростки остальных сегментов срослись с образованием латеральных (боковых) гребней. Межпозвоночные отверстия превратились в ряд дорсальных и вентральных (тазовых) крестцовых
отверстий. На краниальном конце у основания гребня крестцовой кости
имеются краниальные суставные отростки дуговой формы, на каудальном
— маленькие каудальные суставные отростки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У свиньи (Б) тело крестцовой кости выгнуто слабо, срастается поздно.
Остистые отростки отсутствуют. Имеются большие междуговые отверстия,
крылья развернуты в сагиттальной плоскости так, что их ушковидные поверхности направлены латерально.
У лошади (В) тело крестцовой кости лежит горизонтально. Остистые отростки срастаются редко, расширены, иногда раздвоены. Крылья развернуты
во фронтальной плоскости, ушковидная поверхность обращена дорсально.
На краниальном крае крыльев есть суставные поверхности для сочленения с
суставными поверхностями ноперечнореберных отростков последнего поясничного позвонка. Суставные отростки по форме такие же, как на поясничных позвонках.
Скелет хвоста
Костной основой хвоста являются хвостовые позвонки — vertebrae caudales, количество которых колеблется в широких пределах. Хвостовые сегменты сильно редуцированы. Лишь на нескольких первых хвостовых позвонках есть тело, дуга и отростки. Начиная с 3—4-го позвонка, идет редукция дуги и отростков, затем редукция захватывает и тело. У крупного рогатого скота от 2-го до 5-го хвостовые позвонки имеют гемальные дуги с вентральной стороны тела для прохождения сосудов и нервов. Последний позвонок приобретает вид маленького конуса.
Скелет шеи
Скелет шеи образован у всех млекопитающих 7 позвонками за редким
исключением (у ленивца — 6—9, у ламантина — 6), с которыми рано срастаются остатки редуцированных ребер. 1-й и 2-й шейные позвонки резко отличаются от остальных, 3, 4 и 5-й сходны по строению и называются типичными. У 6-го позвонка иная форма поперечнореберного отростка, а 7-й сочетает в себе черты как шейного, так и грудного позвонка. В связи с длиной и
подвижностью шеи позвонки этого отдела отличаются сильно выпуклой головкой, вогнутой ямкой, значительной толщиной межпозвоночных дисков,
сильным развитием всех отростков, кроме остистых, крупными межпозвоночными отверстиями. Все это, в том числе слабое развитие остистых отростков, способствует подвижности шеи.
У типичного шейного позвонка — vertebra cervicalis — крупного рогатого скота сравнительно короткое тело с выпуклой головкой, глубокой ямкой,
выраженным вентральным гребнем (кроме 6-го позвонка). На дуге глубокие
краниальные и каудальные позвоночные вырезки. Хорошо развиты краниальные и каудальные суставные отростки. Остистый отросток сравнительно высокий, направлен краниально. Поперечнореберный отросток раздвоен в дорсовентральном направлении. Между ним и телом позвонка есть межпоперечное отверстие. Позвоночное отверстие широкое. У 7-го позвонка длинный остистый отросток, есть каудальные реберные ямки. У свиньи тело поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
звонка очень короткое, вентральный гребень не выражен. У основания поперечнореберных отростков есть поперечное отверстие.
У лошади тело позвонка в 2 раза длиннее, чем у крупного рогатого скота, вентральный гребень хорошо развит. Остистый отросток очень мал, шероховат. Поперечнореберные отростки раздвоены в краниокаудальном направлении.
2-й шейный позвонок — осевой (эпистрофей)—axis — массивный, с
длинным телом, на краниальном конце которого вместо головки имеется зуб,
или зубовидный отросток (рис. 51).
Рис. 51. 2-й шейный позвонок (сбоку):
А — крупного рогатого скота; Б — свиньи; В — лошади; 1 — зубовидный
отросток; 2 — суставные поверхности; 3 — тело; 4 — вентральный гребень; 5 — межпоперечное отверстие; 6 — поперечнореберный отросток; 7
— ямка; 8 — каудальная позвоночная вырезка; 9 — каудальные суставные
отростки; 10 — гребень; 11 — межпозвоночное отверстие.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У крупного рогатого скота (А) зуб позвонка полулунной формы. По обе
стороны от него — вентральная суставная поверхность для сочленения с
атлантом. На каудальном конце тела — глубокая ямка, на вентральной поверхности тела — вентральный гребень. На ножках дуги спереди — латеральное отверстие, сзади глубокая каудальная позвоночная вырезка. По бокам — поперечные отростки. Между ними и телом—межпоперечное отверстие. Остистый отросток имеет вид мощной прямоугольной пластины и называется гребнем. Сзади у его основания видны парные дорсальные суставные поверхности, соответствующие каудальный суставным отросткам других позвонков.
У свиньи (Б) тело позвонка сравнительно короткое, зуб имеет форму конуса. Гребень очень высокий и приподнят к заднему краю. Поперечные отростки короткие, вентральная суставная поверхность разъединена на две. Вентральный гребень развит слабо.
У лошади (В) тело осевого позвонка длинное, зуб в форме полуцилиндра
с острым краем, вентральный гребень хорошо развит. Гребень позвонка
мощный, раздвоенный, несет на себе дорсальные суставные поверхности.
1-й шейный позвонок (атлант)—atlas — не имеет тела. Его тело в процессе филогенеза переместилось и стало зубом осевого позвонка. В результате у атланта образовалась форма кольца, состоит он из двух дуг: дорсальной
и вентральной.
У крупного рогатого скота (рис.52) дорсальная и вентральная дуги атланта, срастаясь, формируют овальной формы отверстие позвонка. На дорсальной дуге есть дорсальный бугорок, на вентральной — вентральный бугорок. У рогатого скота они не выступают за края дуг. На краниальном крае атланта обширная краниальная суставная ямка для сочленения с затылочной
костью черепа. На каудальном крае — парная плоская каудальная суставная
ямка для сочленения с осевым позвонком. Поперечные отростки атланта широкие, называемые крыльями. На них имеются два отверстия, лежащие в общем углублении; медиально расположенное межпозвоночное и более латерально — крыловое.
Рис. 52. 1-й шейный позвонок
крупного рогатого скота (сверху):
1— дорсальная дуга; 2— дорсальный бугорок; 3 — межпозвоночное отверстие; 4 — крыловое
отверстие; 5 — крыло; 6 — каудальная суставная поверхность;
7 —вентральная дуга; 8 — вентральный бугорок.
У свиньи атлант короткий, с узкими крыльями. Дорсальный бугорок
широкий, вентральный—длинный, направлен каудально и выступает за край
дуги. На крыле кроме межпозвоночного и крылового (часто в виде вырезки)
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
есть поперечное отверстие (иногда неполное), расположенное у каудального
края позвонка.
У лошади дорсальная дуга сильно выпуклая, отверстие позвонка округлое, крылья широкие, тонкие, края их опущены вниз. У каудального края
крыла имеется крупное поперечное отверстие. Дорсальный бугорок сглажен.
Скелет головы
Скелет головы, или череп, — cranium — является вместилищем головного мозга, органов слуха, зрения и обоняния, образует стенки ротовой и носовой полостей. Кости черепа преимущественно пластинчатые, многие из
них пневматизированы— имеют пазухи (синусы) — полости, заполненные
воздухом. Это облегчает массу черепа, сохраняя обширные площади прикрепления мышц. Скелет головы подразделяют на мозговой и лицевой отделы,
граница между ними проходит на уровне глазниц. Граница эта условная, так
как происходит взаимное проникновение костей одного отдела в другой.
Мозговой отдел черепа (кости черепа). К костям мозгового отдела относятся 4 непарные кости: затылочная, клиновидная, решетчатая и межтеменная и 3 парные: височная, теменная и лобная. Все они участвуют в образовании стенок черепной полости. Крыша черепной полости у рогатого скота
образована лобными костями. У свиньи и у лошади еще и теменными и межтеменной. Боковые стенки образованы теменными и височными костями, передняя стенка — решетчатой костью. В состав задней стенки у рогатого скота входят затылочная, межтеменная и теменные кости, а у свиньи и лошади
только затылочная кость. Со стороны черепной полости на костях есть следы
от головного мозга: передняя, средняя и задняя ямки от вдавливания частей
мозга, пальцевые вдавления в местах прилегания извилин, мозговые возвышения там, где проходят борозды. В местах прохождения сосудов и нервов на
костях видны борозды и отверстия.
Затылочная кость — os occipitale (рис. 53) —состоит из основной части
(тела), двух боковых частей и чешуи. Между телом и боковыми частями находится большое затылочное отверстие, через которое головной мозг соединяется со спинным. Тело лежит в задней части основания черепа в форме
уплощенного цилиндра с несколько расширенным концом (особенно у свиньи). На его дорсальной (внутренней) поверхности имеются вдавления продолговатого мозга и мозгового моста, входящие в состав задней черепной
ямки. На наружной (вентральной) поверхности тела в месте его срастания с
телом клиновидной кости заметны мышечные бугры для прикрепления длинной мышцы головы. Они наиболее развиты у рогатого скота и наименее — у
лошади. По бокам тело заостренное и частично формирует рваное отверстие.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 53. Череп крупного рогатого скота:
А — вид сзади; Б — сверху со вскрытыми слева лобной и носовой костями; В
— сагиттальный распил с медиальной стороны;
I — затылочная кость; 1 — ее тело; 2 — боковые части; 3— чешуя: 4—
мыщелки: 5 — большое затылочное отверстие; 6— яремные отростки; 7—
подъязычное отверстие; 8— ямка продолговатого мозга; 9— ямка мозгового моста; II — теменная кость; III — межтеменная кость; IV — лобная
кость; 10— лобный гребень; 11— надглазничное отверстие; 12 - скуловой
отросток; 13 — лобные пазухи; V — височная кость; 14 — скуловой отросток; 15 — скалистая часть; VI — скуловая кость; VII — слезная кость; VIII
— носовая кость; IX — резцовая кость; 16 — носовой отросток; 17 — тело;
18 — нёбный отросток; X — сошник: XI—дорсальная носовая раковина; XII
— вентральная носовая раковина; XIII — верхнечелюстная кость; 19 — беззубый край; 20 — коренной край; 21 — нёбный отросток; 22 — нёбный синус: XIV — нёбная кость (ее перпендикулярная пластинка); XV — крыловидная кость; XVI — решетчатая кость; 23 — лабиринт; 24 — продырявленная пластинка; XVII — клиновидная кость; 25 — овальное отверстие; 26 —
ямка гипофиза.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
На боковых частях различают мыщелки, расположенные по обеим сторонам от большого затылочного отверстия. Ими череп сочленяется с атлантом. Латеральнее мыщелков с каждой стороны имеется по яремному отростку, свободным концом направленному вниз. Они особенно сильно развиты
у свиньи. В углублении между мыщелком и яремным отростком есть подъязычное отверстие (у рогатого скота парное) для прохождения одноименного
нерва.
Чешуя затылочной кости образует заднюю стенку черепной полости. Ее
верхний край является выйным (затылочным) гребнем. У лошади и свиньи
он идет по каудодорсальному краю черепа. У рогатого скота он спускается
ниже в связи с развитием роговых отростков, которые оттесняют межтеменную и частично теменные кости на заднюю стенку черепа. На чешуе закрепляются канатиковая часть выйной связки и некоторые мышцы, разгибающие
голову.
Клиновидная кость — os sphenoidale (см. рис. 53 и 55) —по форме является симметричной смешанной костью и состоит из тела, двух пар крыльев и
крыловидных отростков. Тело клиновидной кости цилиндрическое, образовано двумя частями, между которыми долго сохраняется хрящевая прослойка. Передняя часть тела приподнята по отношению к задней и снаружи прикрыта сошником. Вместе с крыльями тело принимает участие в формировании средней и передней мозговых ямок. В области средней мозговой ямки на
теле имеется углубление, переходящее в возвышение, — турецкое седло. Углубление турецкого седла— это ямка гипофиза, а его возвышение — спинка
турецкого седла. Она высокая у рогатого скота и свиньи и низкая у лошади.
Впереди турецкого седла в месте перехода передней части тела в заднюю
возвышается желоб перекреста зрительных нервов, переходящий на наружной поверхности кости в зрительные отверстия. Через желоб, перекрещиваясь, проходят зрительные нервы, направляясь к головному мозгу. В теле клиновидной кости имеется пазуха, заходящая и в нёбную кость.
По бокам от тела каудодорсально направлены височные крылья, ростродорсально глазничные крылья, в которых имеется ряд мозговых вдавлений,
сосудистых и нервных желобов и отверстий. Глазничные крылья граничат с
лобной костью и вместе с нею образуют медиальную стенку глазницы. Височные крылья прилежат к височной кости. На их внутренней поверхности у
лошади видны два желоба, ведущие в глазничное (медиальное) и круглое (латеральное) отверстия, разъединенные перегородкой, иногда неполной. У рогатого скота и свиньи они объединены в одно круглоглазничное отверстие.
Через эти отверстия прохо дят с III до VI черепномозговые нервы. Несколько
выше находится зрительное отверстие для зрительного нерва. Каудальный
край височного крыла вместе с телом затылочной и каменистой костями
формирует рваное отверстие. Оно сильно развито у лошади, узко и прикрыто барабанной частью каменистой кости у свиньи и особенно у рогатого скота, у которого из его передней части сформировалось овальное отверстие.
Вперед и вниз от тела отходят парные крыловидные отростки. Латерально к
ним прилегает нёбная кость, а медиально — крыловидная.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 54. Череп с латеральной стороны:
А— череп крупного рогатого скота и Б — его височной области; В —
лошади; Г —свиньи; I — резцовая кость; 1 — ее тело; 2 — носовой отросток; 3 — резцы; II—верхнечелюстная кость; 4— клык; 5 — беззубый край;
6— лицевая поверхность; 7 — подглазничное отверстие; 8 — тело; 9 — альвеолярный край; 10 — лицевой бугор (у лошади — гребень); 11 — челюстной
бугор; III — носовая кость; IV — слезная кость; 12 — лицевая пластинка; 13
— глазничная пластинка; V — лобная кость; 14 — скуловой отросток; VI —
теменная кость; VII — затылочная кость; 15 — мыщелок; 16 — яремный
отросток; VIII — височная кость; 17 — чешуя височной кости; 18 — наружный слуховой проход; 19 — костный барабан; 20 — мышечный отросток; 21 — скуловой отросток; IX — клиновидная кость; 22 — крыловидный
отросток; 23 — височное крыло; 24 — круглоглазничное отверстие; 25 —
глазничное крыло; X — нёбная кость; XI — скуловая кость; 26 — височный
отросток; 27 — лобный отросток; XII — хоботковая кость.
Височная кость — os temporale (рис. 54, 55) —парная, по форме несимметричная смешанная кость, состоит из чешуйчатой, каменистой (скалистой)
и барабанной частей, которые у крупного рогатого скота рано сливаются в
одну кость, а у овцы, свиньи и лошади некоторые части сохраняют самостоятельность. Кроме того, эта кость входит в состав боковой стенки черепной
полости и скуловой дуги. Она является вместилищем органа слуха и образует
сустав с нижней челюстью, а потому имеет очень сложное строение и большое количество щелей и каналов для прохождения нервов, сосудов.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 55. Череп крупного рогатого скота с базальной поверхности:
I — затылочная кость; 1 — ее тело; 2— мышечные отростки; 3—
яремный отросток; 4 — мыщелок; 5 — большое затылочное отверстие; II—
височная кость; 6— костный барабан; 7 — мышечный отросток; 8 — подъязычный отросток; 9 — челюстная ямка; 10 — суставной бугорок; 11 —
скуловой отросток; III — клиновидная кость; 12 — ее тело; 13 — височные
крылья; 14 — овальное отверстие; 15 — глазничные крылья; 16 — крыловидные отростки; IV — сошник; V — крыловидная кость; VI — лобная кость;
17 — слезный мешок; VII — скуловая кость; 18 — височный отросток; VIII
— нёбная кость; 19 — вертикальная пластинка; 20 — горизонтальная пластинка; 21 — клинонёбная ямка; IX — верхнечелюстная кость; 22 — челюстной бугор; 23— тело; 24 — альвеолярный край; 25 — лицевой бугор; 26 —
нёбный отросток; 27 — беззубый край; X — резцовая кость; 28 — носовой
отросток; 29 — нёбный отросток; 30 — тело.
Чешуйчатая часть образует большую часть боковой стенки черепной полости. Вбок от височной поверхности чешуи отходит скуловой отросток, коwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
торый затем поворачивает вперед и вместе с височным отростком скуловой
кости образует скуловую дугу. Между скуловой дугой и височной поверхностью чешуи видна обширная, особенно у хищных, височная ямка.
На вентральной поверхности скулового отростка (до его поворота вперед) находится пологий суставный бугорок с суставной поверхностью для
сочленения с мыщелковым отростком нижней челюсти, позади которого лежит неглубокая нижнечелюстная ямка.
Барабанная часть — место прикрепления ушной раковины и вместилище
среднего уха. Наружный слуховой проход расположен позади и немного ниже скулового отростка чешуи. Его отверстие у свиньи направлено вверх, у
рогатого скота — вбок и дополняется хрящом слухового прохода, который
переходит в хрящ ушной раковины. Сзади и ниже наружного слухового прохода в углублении расположен короткий, напоминающий пенек, подъязычный отросток для сочленения с подъязычной костью.
Наружный слуховой проход соединен с медиально расположенным барабанным пузырем (костным барабаном). У лошади он округлой формы, у
рогатого скота и свиньи — овальной. Внутри барабанного пузыря имеется
барабанная полость с заключенными в ней слуховыми косточками: молоточком, наковальней, стремечком. От наружного слухового прохода барабанная
полость отделена барабанной перепонкой. На наружной поверхности барабанного пузыря имеется тонкий мышечный отросток, своей заостренной
вершиной направленный ростровентрально. У его основания заметно полулунное отверстие мышечно-трубного канала (глоточнобарабанной, или евстахиевой трубы), который соединяет глотку с барабанной полостью для
выравнивания давления в последней.
Каменистая (скалистая) часть углублена в черепную полость и своим
сосцевидным отростком прикреплена к боковой части затылочной кости
около яремного отростка (у рогатого скота и свиньи сосцевидный отросток
не развит). На медиальной поверхности каменистой части есть внутренний
слуховой проход, своим отверстием направленный в черепную полость. В его
глубине видны два отверстия. То, что лежит каудальнее, открывается в канал
слухового (преддверно-улиткового) нерва и ведет во внутреннее ухо. То, что
лежит ростральнее, ведет в лицевой канал, в котором внутри каменистой части проходит лицевой нерв. На другом конце лицевой канал открывается отверстием, расположенным между подъязычным и сосцевидным отростками.
Теменная кость — os parietale (см. рис. 54)—парная плоская кость, входит в состав крыши, боковых стенок черепной полости и вместе с височной
костью образует височную ямку. У рогатого скота она в виде узкой полосы,
сдвинутой вниз и назад, где срастается с межтеменной и затылочной костями. У свиньи и лошади теменные кости срастаются на некотором
расстоянии между собой медиальными краями, формируя сагиттальный гребень. С внутренней стороны на кости видны пальцевые вдавления и мозговые возвышения.
Межтеменная кость — os interparietale (см. рис. 53)—маленькая непарная кость треугольной или квадратной формы, рано срастается с теменными
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
и затылочной костями. У свиньи сращение происходит еще в плодный период развития.
Лобная кость — os frontale (см. рис. 53, 54)—парная, плоская кость. На
ней различают чешую, височную и носовую части. Лобная чешуя образует
свод черепной коробки. У крупного рогатого скота она плоская, у лошади и
овцы — выпуклая, у свиньи — вогнутая. У крупного рогатого скота каудально доходит до затылочной области, формирует лобный бугор и несет роговые
отростки. У мелкого рогатого скота роговые отростки отходят от лобной
чешуи позади глазниц, не доходя до затылочной области. У лошади и свиньи
лобная чешуя не доходит до затылочной области, так как формирует свод
вместе с теменными костями.
В области глазницы от чешуи отходит скуловой отросток, который формирует задний край орбиты глаза. 'На его внутренней поверхности имеется
мелкая ямка слезной железы. У крупного рогатого скота и лошади скуловой
отросток достигает скуловой кости, поэтому говорят, что у этих животных
орбита замкнутая. У свиньи и овцы орбита незамкнутая — у них скуловой
отросток не достигает скуловой кости. Над глазницей имеется надглазничное
отверстие для прохождения лобных артерий и нерва.
Глазничная часть участвует в образовании медиальной стенки глазницы,
а носовая часть входит в состав крыши носовой полости.
Решетчатая кость — os ethmoidale (см. рис. 53)—непарная смешанная
кость, состоит из ряда пластинок. Ее продырявленная пластинка образует переднюю стенку черепной полости и имеет два углубления с многочисленными отверстиями для прохождения нитей обонятельного нерва. От ее середины в глубь носовой полости отходит перпендикулярная пластинка, передний
конец которой переходит в хрящевую носовую перегородку. От перпендикулярной и продырявленной пластинок справа и слева отходят пластинки, к которым прикрепляются очень тонкие пористые костные листочки, завитки и
трубочки, в сумме формирующие решетчатый лабиринт. Кости решетчатого лабиринта покрыты обонятельным эпителием, образующим рецепторное
поле органа обоняния.
Лицевой отдел черепа (кости лица). Кости лица образуют стенки носовой и ротовой полостей. К ним относятся 3 непарные кости (сошник, хрящевая носовая перегородка и подъязычная кость) и 10 парных (носовая, верхнечелюстная, резцовая, слезная, скуловая, дорсальная и вентральная носовые
раковины, нёбная, крыловидная, нижнечелюстная кости), а у свиньи еще хоботная кость.
Крыша носовой полости образована лобной (частично) и носовой костями. Боковые стенки — верхнечелюстной, слезной, скуловой. Вход в носовую
полость ограничен резцовой и носовой костями, выход (хоаны)—сошником,
крыловидной, нёбной, лобной и клиновидной костями. В носовой полости
имеются внутренние перегородки, образованные решетчатой костью, хрящевой носовой перегородкой, сошником, носовыми раковинами. Дно носовой и крышу ротовой полости образуют резцовая, верхнечелюстная, нёбная
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
кости. Нижнечелюстная и подъязычная кости входят в состав дна ротовой
полости.
Носовая кость — os nasale (см. рис. 53, 54) —парная, плоская, имеет
вид длинной пластинки с заостренным передним концом. Вдоль ее внутренней поверхности идет решетчатый гребень для прикрепления дорсальной
носовой раковины.
Слезная кость — os lacrimale (см. рис. 53, 54) —парная плоская, состоит из двух пластинок, расположенных под углом друг к другу: лицевой и
глазничной. Лицевая пластинка входит в состав стенки носовой полости, а
глазничная пластинка — в состав глазницы. На глазничной пластинке у ее
переднего края имеется воронкообразное углубление — ямка слезного мешка, на дне которой открывается слезное отверстие, ведущее в слезно-носовой
канал.
Скуловая кость — os zygomaticum (см. рис. 53, 54) —парная, плоская,
образует боковую часть лицевого отдела черепа и входит в состав стенки орбиты. Имеет два отростка. Ее височный отросток вместе со скуловым отростком височной кости формирует скуловую дугу, а лобный отросток вместе
со скуловым отростком лобной кости — задний край орбиты.
Он хорошо развит у рогатого скота, слабо у свиньи и отсутствует у лошади.
Верхнечелюстная кость — os maxillare (см. рис. 53, 54) — парная, плоская, имеет тело, в котором расположены зубы, несколько поверхностей, образующих боковые стенки и дно носовой и крышу ротовой полостей. На теле
различают альвеолярный край с альвеолами для коренных зубов и клыки (у
жеребцов и свиней обоего пола). Часть тела между клыком и коренными зубами не содержат альвеол и называется межальвеолярным (беззубым) краем.
Он длинный у лошади, рогатого скота и короткий у свиней. Позади последней альвеолы на теле виден широкий верхнечелюстной бугор. Его медиальная поверхность участвует в формировании крылонёбной ямки, глубокой у
рогатого скота и свиньи, широкой у лошади. В ней открывается несколько
отверстий, ведущих в каналы (нёбный, подглазничный и др.) для сосудов и
нервов.
Вверх от тела отходит широкая лицевая поверхность (пластинка), на которой различают лицевой бугор (у лошади и свиньи — лицевой гребень) для
прикрепления большой жевательной мышцы, а также подглазничное отверстие, ведущее в подглазничный канал. По нему сосуды и нервы подходят к
зубам верхней челюсти. С медиальной стороны вдоль лицевой поверхности
проходит гребень вентральной раковины.
Медиально от тела находится носовая поверхность (нёбный отросток),
соединяющаяся с соименной поверхностью другой верхнечелюстной кости и
входящая в состав твердого нёба.
Резцовая кость — os incisivum (см. рис. 53, 54, 55) —парная, состоит из
тела, носового и нёбного отростков. Тело резцовой кости рогатого скота
плоское, в виде пластинки, из-за отсутствия резцов, у лошади и свиньи массивное, несет три альвеолы для резцов. У свиньи к телу прилежит небольшая
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
хоботковая кость треугольной формы. Каудодорсально от тела отходит носовой отросток, образующий боковую границу входа в носовую полость.
Каудально от тела направлен нёбный отросток, входящий в состав костной
основы твердого нёба.
Нёбная кость — os palatinum (см. рис. 55)—парная, плоская, состоит из
горизонтальной и перпендикулярной пластинок. Горизонтальная пластинка
широкая у рогатого скота и свиньи, узкая у лошади. Соединяясь рострально с
нёбной поверхностью верхнечелюстной кости, входит в состав твердого нёба, образует вентральный край хоаны и является местом прикрепления сошника. Перпендикулярная пластинка отходит вверх от горизонтальной пластинки в сагиттальной плоскости. Хорошо развита у рогатого скота и лошади, слабо у свиньи. Она является боковой стенкой хоаны и вместе с крыловидным отростком клиновидной кости и верхнечелюстным бугром образует
крылонёбную ямку.
Крыловидная кость — os pterygoideum (см. рис. 55)—парная, плоская
кость, в виде тонкой пластинки, широкой и длинной у крупного рогатого
скота, широкой и короткой у свиньи, узкой и длинной у лошади. Прилегает с
медиальной стороны к крыловидному отростку клиновидной кости и перпендикулярной пластинке нёбной кости. Входит в состав боковой стенки хоаны.
Сошник —vomer (см. рис. 55)—непарная плоская кость, располагается в
средней сагиттальной плоскости на дне носовой полости в виде желоба, в котором проходит хрящевая носовая перегородка. Каудально сошник делит выход из носовой полости на две хоаны и прикрывает снизу тело клиновидной
кости.
Носовая перегородка является продолжением в ростральном направлении перпендикулярной пластинки решетчатой кости. Из костной она переходит в хрящевую, разделяет носовую полость на две половины. Дорсально соединяется с носовыми костями, вентрально входит в желоб сошника.
Носовые раковины (см. рис. 53)—парные. В каждой половине носовой
полости есть вентральная и дорсальная носовые раковины. Они являются
продолжением лабиринта решетчатой кости в ростральном направлении. Образованы очень тонкими пористыми костными (бумажными) пластинками,
закрученными в виде спиральных трубок. Дорсальная раковина прикрепляется к решетчатому гребню носовой кости, вентральная — к гребню вентральной раковины лицевой поверхности верхнечелюстной кости. Раковины являются остовом, на котором располагается слизистая оболочка носа.
Нижнечелюстная кость — os mandibulare (рис. 56)—парная плоская. У
свиньи и лошади обе кости рано срастаются. У рогатого скота между ними
сохраняется хрящ. Кости нижней челюсти каудально расходятся одна от другой. Между ними образуется треугольной формы межчелюстное пространство.
Каждая нижнечелюстная кость состоит из тела и ветви. Тело подразделяют на резцовую и щечную (коренную) части, на дорсальном — альвеолярном крае которых есть альвеолы для зубов: по 3 альвеолы для резцов у лошади и свиньи, по 4 у рогатого скота; по 6 альвеол для коренных зубов у рогаwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
того скота и лошади, по 7 у свиньи. Участок дорсального края между ними не
содержит альвеол и называется межальвеолярным краем. На нем ближе к
резцовой части у свиней обоего пола и у жеребцов имеется альвеола для
клыка. На наружной и внутренней поверхности тела около резцовой части
имеются подбородочные отверстия. Вентральный край тела у свиньи и лошади ровный, у рогатого скота — несколько выпуклый. На нем около перехода в ветвь видна пологая лицевая сосудистая вырезка, где из межчелюстного пространства на латеральную сторону проходят проток околоушной
слюнной железы, лицевые артерии и вена.
Рис. 56. Нижнечелюстная кость крупного
рогатого скота:
1 — тело; 2 — резцовая часть: 3 — коренная
часть; 4 — альвеолярный край; 5 — межальвеолярный край; 6 — подбородочное отверстие; 7 — вентральный край; 8 — сосудистая вырезка; 9 — ветвь нижней челюсти;
10— челюстной угол; 11 — ямка большой
жевательной мышцы; 12 — нижнечелюстное отверстие; 13 — мышечный отросток;
14 — суставной отросток.
От каудального конца тела вверх под тупым углом отходит ветвь нижней челюсти. В месте перехода у вентрального края образуется челюстной
угол. На латеральной поверхности видна обширная пологая ямка большой
жевательной мышцы, на медиальной поверхности подобная ей ямка крыловой мышцы для прикрепления соответствующих мышц. На медиальной поверхности ветви видно нижнечелюстное отверстие, ведущее в нижнечелюстной канал. По нему проходят сосуды и нервы и выходят из канала через подбородочные отверстия. На дорсальном конце ветви видны два отростка: венечный (мышечный) отросток — служит местом прикрепления височной мышцы и мыщелковый (суставной) отросток для сочленения с суставным бугорком височной кости с образованием височно-нижнечелюстного
сустава. У рогатого скота венечный отросток длинный, загнут в каудальную
сторону; у лошади длинный, направлен вверх, у свиньи короткий, по высоте
равный мыщелковому отростку.
Подъязычная кость — os hyoideum (рис. 57)—непарная, смешанная,
располагается в межчелюстном пространстве, соединена с барабанной частью височной кости. На ней прикрепляются мышцы гортани, глотки, языка
и некоторые мышцы шеи, осуществляющие согласованные движения во время глотания, что позволяет называть всю эту структуру подъязычным аппаратом. Состоит из тела, рогов и ветвей. Тело имеет вид пластинки и лежит в
области корня языка. От него в толщу языка у рогатого скота и лошади отходит язычный отросток. Каудально от тела направлены большие рога, которые соединяются со щитовидным хрящом гортани. Дорсокраниально от тела
идут малые рога, к которым прикрепляются ветви, состоящие из трех члениwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ков. Дистальный членик небольшой. У рогатого скота и у лошади костный, у
свиньи — в виде связки, соединяющей малые рога со средним члеником.
Средний членик длинный, плоский у рогатого скота и лошади, у свиньи столбикообразный, частично образован хрящом. Проксимальный членик короткий, хрящевой, им кость присоединяется к подъязычному отростку височной
кости.
Рис. 57. Подъязычная кость.
А — крупного рогатого скота; Б—лошади; В — свиньи; 1 — тело; 2—
язычный отросток; 3 — большие рога; 4 — малые рога; 5 — дистальный
членик ветви; 6 — средний членик ветви; 7 — проксимальный членик ветви.
Костные пазухи. Многие кости черепа пневматизированы. У всех пневматизированных костей головы между наружной и внутренней пластинками
имеются пространства, выстланные слизистой оболочкой и заполненные воздухом — пазухи или синусы (см. рис. 53). Они поделены перегородками на
отдельные камеры. Многие пазухи соединены между собой и с носовой полостью. Их функцией является облегчение костей черепа, увлажнение и согревание воздуха. Лучше всего развиты пазухи у крупного рогатого скота.
Наиболее крупные пазухи: верхнечелюстная— расположена в верхнечелюстной, слезной, скуловой и нёбной костях; лобная — находится в лобной и теменной костях (у лошади — только в лобной); клинонёбная — в клиновидной и нёбной костях.
СКЕЛЕТ КОНЕЧНОСТЕЙ (ПЕРИФЕРИЧЕСКИЙ СКЕЛЕТ)
Скелет конечностей состоит из скелета поясов и свободных конечностей. Пояса плечевой и тазовый служат для связи конечностей с туловищем.
Строение плечевого и тазового поясов резко отличается, что связано с особенностями функций конечностей. Грудная конечность является основной
при опоре и рычагом, размах которого определяет ширину шага при движении. Тазовая конечность — основной толкатель туловища вперед при движении. При распрямлении углов тазовой конечности во время движения сила
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
отталкивания передается через пояс конечности на ствол тела, который приподнимается и толкается вперед.
Скелет свободных грудной и тазовой конечностей состоит из основного
поддерживающего столба и лапы. Поддерживающий столб включает плечо и
предплечье на грудной, бедро и голень на тазовой конечности. У копытных в
его состав входит и большая часть лапы. Конечности представлены одинаковым числом тождественных звеньев, соединенных суставами. Отличия заключаются в том, что углы аналогичных суставов на грудной и тазовой конечностях обращены в противоположные стороны. К тому же кости тазовой
конечности несколько длиннее и массивнее костей грудной конечности и соединены друг с другом под более острыми углами. Количество костей в гомодинамных звеньях свободных конечностей одинаково: самое верхнее звено состоит из одной кости (плечевой на грудной и бедренной— на тазовой
конечности); второе — из двух костей (лучевой и локтевой на грудной, большой и малой берцовых — на тазовой конечности). Третье звено (лапа) состоит из запястья (заплюсны), пясти (плюсны) и пальцев. Здесь число костей в
каждом звене, кроме запястья и заплюсны, соответствует количеству развитых пальцев и может быть от 1 до 5. Каждый палец состоит из трех фаланг. В
основе каждой фаланги лежит по одной кости. Большинство сельскохозяйственных животных относятся к копытным. Это фалангоходячие животные с
частичной редукцией костей лапы и основного поддерживающего столба.
Скелет грудной конечности
Скелет пояса грудной конечности у примитивных (однопроходных)
млекопитающих состоит из трех костей: лопатки, кора-коидной кости и ключицы. У домашних млекопитающих полноразвитой является только лопатка.
Коракоидная кость редуцировалась до небольшого бугорка, сросшегося с лопаткой. От ключицы осталась у кролика и хищных небольшая косточка в виде узкой пластинки, вправленной в плечеголовную мышцу. Плечевой пояс не
сочленяется с костями осевого скелета, а присоединяется к нему с помощью
мышц. Такой вид соединения называется синсаркозом.
Лопатка — scapula — парная, плоская кость в виде треугольника, вершиной направлена краниовентрально, а основанием каудодорсально. На ней
различают латеральную и медиальную (реберную) поверхности, дорсальный,
краниальный и каудальный края, краниальный, каудальный и вентральный
(дистальный) углы.
У крупного рогатого скота (рис. 58) по латеральной поверхности лопатки тянется длинный узкий гребень, высота которого постепенно повышается,
— ость лопатки. Около вентрального угла ость лопатки круто обрывается и
заканчивается выступом — акромионом. На середине ости есть шероховатое
утолщение — бугор ости. Ость делит латеральную поверхность лопатки на
две ямки: предостную — более узкую и заостную — более широкую. В них
лежат одноименные мышцы. На медиальной (реберной) поверхности лопатки
имеется широкая подлопаточная ямка, в которой лежит подлопаточная мышwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ца. Над ямкой видна зубчатая поверхность, к которой прикрепляется зубчатая вентральная мышца. На дорсальном крае лопатки закреплен надлопаточный хрящ.
Около вентрального угла лопатка сильно сужается, образуется шейка
лопатки. Затем на вентральном углу лопатка вновь несколько расширена и
имеет суставную впадину для сочленения с плечевой костью. Краниально от
суставной впадины имеется надсуставной бугорок (бугор лопатки), на медиальной поверхности которого виден небольшой коракоидный отросток (рудимент коракоидной кости).
У свиньи лопатка треугольная короткая и широкая. Бугор ости сильно
развит и наклонен к заостной ямке. Акромиона нет.
Рис. 58. Лопатка крупного рогатого скота с латеральной (А)
и с медиальной (Б) поверхности:
1 — краниальный угол; 2 — ость лопатки; 3 — предостная ямка; 4 —
бугор ости; 5 — акромион; 6—шейка лопатки; 7 — бугор лопатки; 8 — коракондный отросток; 9— суставная впадина: 10 — заостная ямка; 11 —
каудальный угол; 12 — надлопаточный хрящ; 13 — подлопаточная ямка;
14— зубчатая поверхность.
У лошади надлопаточный хрящ очень широкий. Бугор ости хорошо развит и слегка наклонен к заостной ямке. Акромион отсутствует. Коракоидный
отросток хорошо выражен.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Плечевая кость — humerus (os brachii)—длинная трубчатая кость, состоит из диафиза и двух эпифизов (рис. 59). На ней различают краниальную,
каудальную, медиальную и латеральную поверхности. На проксимальном
эпифизе у крупного рогатого скота находится крупная округлая головка,
сдвинутая к медиальной стороне. Под ней — шейка, а около головки два
мышечных бугорка, небольшой медиальный — малый бугорок и крупный
латеральный — большой бугорок, нависающий над малым. Между ними
межбугорковый желоб, по которому проходит сухожилие двуглавой мышцы.
От латерального бугорка на тело спускается гребень, который переходит в
дельтовидную шероховатость, выступающую на латеральной поверхности
тела, и продолжается после нее до дистального эпифиза. На медиальной поверхности тела заметна округлой формы шероховатость большой круглой
мышцы (круглая шероховатость).
Рис. 59. Плечевая кость с краниальной поверхности:
А — крупного рогатого скота; В — лошади; В — свиньи: 1 — головка;
2— шейка; 3 — малый бугорок; 4 — большой бугорок; 5 — межбугорковый
желоб; 6 — гребень; 7— дельтовидная шероховатость; S — круглая шероховатость; 9 — мыщелок; 10 — медиальный надмыщелок; 11—связочные бугорки и ямки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
На дистальном эпифизе есть поперечно поставленный мыщелок плечевой кости с блоком, образующим сустав с костями предплечья. По бокам от
мыщелка располагаются латеральный и медиальный надмыщелки с глубокой
локтевой ямкой между ними, а также связочные бугорки и ямки.
У свиньи большой бугорок сильно нависает над малым, так что межбугорковый желоб почти принимает форму кольца. Гребень, дельтовидная и
круглая шероховатости выражены слабо.
У лошади мышечные бугорки невысокие, двойные. Межбугорковый желоб тоже двойной, его разделяет средний бугорок.
Кости предплечья — ossa antebrachii — длинные трубчатые (рис. 60).
Это лучевая и локтевая кости, которые у копытных животных развиты в разной степени. Более развита лучевая кость. Локтевая кость редуцирована в
разной степени, особенно ее дистальный конец.
Рис. 60. Кости предплечья с дорсолатеральной стороны:
А — крупного рогатого скота; Б — лошади; В — свиньи; I — лучевая
кость; 1 — головка; 2 — ямка головки с суставной поверхностью; 3 — шейка; 4 — лучевая шероховатость; 5 — тело; 6 — суставной блок; II — локтевая кость; 7 — локтевой отросток; 8 — локтевой бугор; 9 — блоковая вырезка; 10 — крючковидный отросток; 11 — тело; 12 — грифелевидный отрос-ток; 13 — межкостное пространство.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У крупного рогатого скота лучевая и локтевая кости срослись, и лишь в
двух участках между ними имеется межкостная щель.
Лучевая кость — radius — состоит из тела, проксимального и дистального эпифизов. На проксимальном эпифизе различают головку, на которой
имеется ямка головки с суставной окружностью. Под головкой — слабо выраженная шейка. На ней спереди шероховатость лучевой кости, по бокам —
связочные бугорки. Тело в поперечнике имеет форму овала с более выпуклой
краниальной поверхностью и более плоской — каудальной. На дистальном
эпифизе расположен суставной блок для сочленения с костями запястья.
Локтевая кость — ulna — имеет тело и два эпифиза, хотя и в редуцированном виде. Проксимальный эпифиз развит лучше других частей. На нем
есть хорошо выраженный локтевой отросток с блоковой вырезкой (для сочленения плечевой кости) и локтевым бугром. Над блоковой вырезкой нависает крючковидный отросток, входящий при разгибании в локтевую ямку
плечевой кости. Тело редуцировано, срослось с лучевой костью и в виде
трехгранной палочки лежит на ее латерокаудальном крае. В дистальном направлении кость все более истончается и от дистального эпифиза остается
лишь грифелевидный (шиловидный) отросток с суставной поверхностью для
костей запястья.
У свиньи кости предплечья короткие, массивные. Локтевая кость прилежит к лучевой широкой шероховатой поверхностью и у взрослых животных прирастает к ней.
У лошади лучевая кость длинная, мощная, локтевая сильно редуцирована. От нее остался лишь проксимальный эпифиз и часть тела, сросшаяся с
лучевой костью. В одном месте между ними есть межкостное пространство.
Скелет кости состоит из костей запястья, пясти и пальцев (рис. 61).
Кости запястья — ossa carpi — короткие, лежат в два ряда. В проксимальном ряду 4 кости: с медиальной стороны — лучевая кость запястья — os
carpi radiale, посередине — промежуточная кость запястья — os carpi intermedium, с латеральной стороны — локтевая кость запястья — os carpi ulnare,
сзади — добавочная кость запястья — os carpi accessorium. В дистальном ряду у животных с пятипалой конечностью 5 костей, счет которых ведется с
медиальной стороны: I запястная кость — os carpale primum, II запястная
кость — os carpale secundum, III запястная кость — os carpale tertium, IV запястная кость — os carpale quartum, V запястная кость — os carpale quintum. У
домашних копытных в проксимальном ряду сохранились все кости. В дистальном ряду у крупного рогатого скота остались 2 кости, образовавшиеся в
результате слияния II с III и IV с V, I кость отсутствует. У свиньи 4 кости: I,
II, III, а IV срослась с V. У лошади те же 4 кости, что и у свиньи, но резко
различаются по размерам — I очень маленькая, часто отсутствует, III — самая крупная, плоская.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 61. Кости кисти с дорсальной поверхности:
А—свиньи; Б— крупного рогатого скота; В — лошади; а—кости запястья; б
— кости пясти; в — кости пальцев; 1— лучевая кость; 2— локтевая кость;
3— лучевая запястная; 4 — промежуточная; 5— локтевая запястная; 6' —
добавочная; 7 — I запястная; 8 — II запястная; 9 — III запястная; 10— IV
запястная; 11— V запястная; 12— I пястная; 13 — II пястная; 14 — III пястная; 15 — IV пястная; 16 — V пястная; 17 — кость проксимальной фаланги пальца (путовая); 18 — кость средней фаланги пальца (венечная); 19
— кость дистальной фаланги пальца (копытцевая, у лошади — копытная); I
— первый, II — второй, III — третий, IV — четвертый и V — пятый пальцы.
Кости пясти — ossa metacarpi — длинные, трубчатые, на проксимальном
эпифизе имеют суставную поверхность для костей запястья и связочные бугорки на боковых поверхностях. На дистальном эпифизе — суставной блок и
по бокам связочные ямки. Количество костей пясти может быть 1—5.
У крупного рогатого скота III и IV пястные кости срослись в одну. В
месте сращения по дорсальной и пальмарной поверхностям проходят дорсальный и пальмарный продольные желоба. Проксимальный эпифиз несет
плоскую суставную поверхность, на дорсальной его поверхности — пястная
шероховатость, а по бокам — связочные бугорки. На дистальном эпифизе —
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
двойной суставной блок с межблоковой вырезкой и связочными ямками по
бокам.
У свиньи 4 пястные кости. Они короткие, массивные. III и IV развиты
сильнее, трехгранные, II и V — меньше, изогнутые, I отсутствует.
У лошади 3 пястные кости. Полно развита III. Это длинная мощная
кость, овальная в поперечнике. На ее проксимальном эпифизе — пястная шероховатость, плоская суставная поверхность, связочные бугорки и фасетки
для прикрепления II и IV пястных костей. На дистальном эпифизе суставной
блок. II и IV пястные кости — грифелевидные, сильно редуцированы, имеют
вид тонких палочек, достигающих 1/2—2/3 III пястной кости, часто с ней срастаются.
Кости пальцев — ossa digitorum-—состоят из трех фаланг в каждом
пальце: проксимальной, средней и дистальной.
У крупного рогатого скота и овцы развиты III и IV пальцы, II и V висячие, часто не имеют костной основы, а являются лишь производными кожи.
У III и IV пальца по три фаланги. Проксимальная фаланга — путовая кость,
длинная, трубчатая. На проксимальном ее конце желобоватая суставная поверхность, по бокам—связочные бугорки, на дистальном — суставной блок,
по бокам — связочные ямки. Средняя фаланга — венечная кость, короткая,
имеет те же черты строения, что и у путовой. Дистальная — копытцевая
кость, в форме трехгранной пирамидки. На ней различают боковую, межпальцевую, подошвенную и суставную поверхности. Впереди над суставной
поверхностью — разгибательный отросток, сзади — сгибательный бугорок.
У свиньи развиты II, III, IV и V пальцы. III и IV — основные, II и V —
висячие. В каждом из них по три фаланги, сходные по строению с соответствующими фалангами пальцев крупного рогатого скота.
У лошади развит лишь III палец. Путовая и венечные кости его аналогичны этим фалангам рогатого скота. Дистальная фаланга— копытная кость
— симметричная. На ней различают стенную, суставную, подошвенную поверхности, венечный (верхний) и подошвенный края. Впереди и выше суставной поверхности— разгибательный отросток, сзади и ниже — сгибательная поверхность, а позади нее — суставная поверхность с сесамовидной (челночной) костью. Стенная поверхность сзади сужается и переходит в пальмарные отростки.
Сесамовидные кости пальцев бывают проксимальные и дистальные.
Проксимальные — на каждом пальце парные, лежат на пальмарной поверхности пястно-путового сустава; дистальная (у лошади — челночная)—на каждом пальце по одной,, лежит на пальмарной поверхности копытцевого (копытного) сустава.
Скелет тазовой конечности
Скелет пояса тазовой конечности, или таз, образован двумя тазовыми,
или безымянными, костями — ossa coxae, — которые соединяются друг с
другом тазовым сращением — симфизом (рис. 62). Каждая из них, в свою
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
очередь, образована тремя костями: подвздошной, лонной и седалищной.
Они рано срастаются между собой таким образом, что подвздошная кость
образует краниальный участок, седалищная — каудальный, а лонная— медиальную часть безымянной кости. В месте их сращения образуется вертлужная ямка (суставная впадина), в глубине которой видна ямка впадины. Над
ней в месте слияния подвздошной и седалищной костей возвышается крупный гребень полулунной формы — седалищная ость. Между лонной и седалищной костями имеется обширное запертое отверстие. Кости таза, крестец
и первые хвостовые позвонки формируют костную стенку тазовой полости.
Широкий вход в тазовую полость ограничен сверху крестцовой костью, с боков — подвздошными, снизу — лонными костями. Более узкий выход обрамлен сверху четвертым хвостовым позвонком, с боков и снизу — седалищными костями.
Рис. 62. Тазовые кости с дорсальной стороны: А — крупного рогатого
скота; Б — лошади;
I — подвздошная кость; 1 — крыло; 2 — медиальный бугор; 3 — подвздошный гребень; 4 — маклок; 5— ягодичная поверхность крыла; 6— тело подвздошной кости; 7 — большая седалищная вырезка; 8 — поясничный бугорок; 9— седалищная ость; 10 — суставная впадина; 11 — малая седалищная
вырезка; 12 — запертое отверстие; II — седалищная кость; 13 — впадинная
ветвь; 14 — тело седалищной кости; 15 — шовная ветвь; 16 — седалищный
бугор; 17 — седалищная дуга; III — лонная кость; 18 — ее тело; 19 — впадинная ветвь; 20 — каудальная ветвь; 21 — подвздошно-лонное возвышение;
22 — лонный гребень.
Подвздошная кость — os ilium (I)—состоит из крыла и тела.
У крупного рогатого скота крыло подвздошной кости имеет вид широкого косо поставленного треугольника, медиальным углом нависающего над
крестцом. Наружная поверхность крыла вогнутая и называется ягодичной поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
верхностью, внутренняя выпуклая— крестцово-тазовая поверхность. На
ней видна шероховатая ушковидная поверхность для сочленения с соименной поверхностью крестцовой кости. Острый передний край крыла называется подвздошным гребнем. Медиально он переходит в крестцовый бугор, латерально — в латеральный подвздошный бугор, или маклок, — место прикрепления ряда мышц туловища и конечности. Он хорошо прощупывается у
животных и используется как опознавательная точка в зоотехнических промерах. Тело столбчатой формы каудальным концом участвует в образовании
суставной впадины. На нем с медиальной стороны заметен небольшой бугорок малой поясничной мышцы (поясничный бугорок). На каудодорсальном
крае подвздошной кости большая седалищная вырезка, которая продолжается
до седалищной ости.
Седалищная кость — os ischii (II)—состоит из тела и ветви. У крупного рогатого скота тело пластинчатой формы. Краниально оно участвует в образовании суставной впадины. Каудальный конец тела несет трехгранный седалищный бугор По краю тела от седалищного бугра до седалищной ости
проходит малая седалищная вырезка. Ветви двух седалищных костей направлены друг к другу и соединяются своими шовными поверхностями, участвуя в образовании симфиза. У быков симфиз срастается рано, у коров — к
7—10 годам. Между седалищными буграми двух костей образуется седалищная дуга.
Лонная кость — os pubis (III)—состоит из тела и двух ветвей. У крупного рогатого скота тела двух лонных костей прилежат друг к другу по средней линии дна таза. У коров срастаются позже, чем у быков. На вентральной
поверхности есть вентральный лонный бугорок, хорошо выраженный у самцов. От тела краниолатерально отходит краниальная ветвь, которая участвует в формировании суставной впадины. На ее переднем крае расположен
лонный гребень с подвздошнолонным возвышением, к которому прикрепляется прямая брюшная мышца, Каудально от тела отходит каудальная ветвь.
Каудальные ветви двух лонных костей срастаются своими шовными поверхностями.
У свиньи тазовая полость цилиндрической формы. Крыло подвздошной
кости округлое и узкое, развернуто в сагиттальной плоскости и прилежит к
крестцу сбоку. Подвздошный гребень округлый, выпуклый. Маклок и крестцовый бугор выражены нечетко. Маклок обращен вентрально. Седалищная
ость, высокая. Подвздошнолонное возвышение сильно развито. Лонный бугорок направлен краниально. Большая и малая седалищные вырезки и седалищная дуга глубокие.
У лошади тазовая полость конусовидная. Крылья крестцовой кости развернуты во фронтальной плоскости и налегают на крестец сверху. Крестцовые бугры сближены. Маклок и седалищный бугор пластинчатые, с двумя
углами.
Бедренная кость — os femoris — длинная трубчатая, состоит из тела
(диафиза) и двух эпифизов (рис. 63). У крупного рогатого скота на проксимальном эпифизе различают округлой формы головку с углублением почти в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
центре — ямкой головки. Головка сидит на шейке. Латерально и выше головки расположен большой вертел, каудомедиально и ниже головки — малый
вертел. Между вертелами — вертлужная ямка. На каудолатеральной поверхности тела у его дистального конца видна плоская надмыщелковая ямка
овальной формы.
Рис. 63. Бедренная кость:
А—крупного рогатого скота с каудальной и Б — с латеральной стороны; В — лошади и Г —свиньи с каудальной стороны; 1 — головка; 2—ямка
головки; 3 — шейка; 4 — малый вертел; 5 — большой вертел; 6 — вертлужная ямка; 7 — третий вертел; 8 — надмыщелковая ямка; 9— латеральный
надмыщелок; 10— латеральный мыщелок; 11 — межмыщелковая ямка; 12 —
медиальный мыщелок; 13 — медиальный надмыщелок; 14 — связочные бугорки и ямки; 15 — ямка подколенной мышцы; 16—разгибательная ямка;
17—латеральный гребень; 18 — желоб коленной чашки; 19 — медиальный
гребень; 20 — средний вертел.
На дистальном эпифизе различают латеральный и медиальный мыщелки, обращенные к каудальной поверхности бедренной кости, и блок бедренной кости (блок коленной чашки), обращенный к дорсальной поверхности
кости. Мыщелки разделены межмыщелковой ямкой с ямками для крестовидной связки в глубине ее. Блок бедренной кости состоит из большого — медиального гребня, меньшего — латерального гребня и желоба между ними, по
которому движется коленная чашка. Между мыщелками и гребнем с каждой
стороны образуется надмыщелок. На латеральном и медиальном надмыщелках имеются бугорки и ямки для прикрепления мышц и связок.
Коленная чашка — patella—короткая сесамовидная кость треугольной
формы, вправленная в четырехглавую мышцу бедра.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У свиньи шейка бедренной кости хорошо выражена, большой вертел не
выступает выше головки. Ямка головки маленькая. Надмыщелковая ямка
имеет вид шероховатости. Гребни блока бедренной кости одинаковой высоты.
У лошади ямка головки глубокая, треугольная, расположена сбоку. От
большого вертела обособляется средний вертел, ниже имеется третий вертел. Надмыщелковая ямка глубокая. Медиальный гребень блока сильно развит и на проксимальном конце имеет площадку для фиксации коленной чашки. Коленная чашка четырехугольной формы.
Кости голени — ossa cruris — представлены большой и малой берцовыми костями. Малая берцовая кость у животных разных видов в различной
степени редуцирована (рис. 64).
Рис. 64. Кости голени с дорсолатеральной стороны:
А — крупного рогатого скота; Б— лошади; В —свиньи; I — большая
берцовая кость; 1— медиальный мыщелок; 2— межмыщелковое возвышение;
3— латеральный мыщелок; 4— гребень; 5 —медиальная лодыжка; 6 — суставной блок; II — малая берцовая кость; 7 — головка; 8 — тело; 9 — латеральная лодыжка (лодыжковая кость); 10 — межкостное пространство.
У крупного рогатого скота большая берцовая кость — tibia — длинная
трубчатая, состоит из тела (диафиза) и двух эпифизов (I). На проксимальном
эпифизе различают латеральный и медиальный мыщелки, разделенные межмыщел-ковым возвышением, в котором имеются ямки для крестовидной
связки. Краниальный край дорсальной поверхности тела сильно выступает
вперед в виде гребня, идущего от медиального мыщелка. По каудальной поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
верхности проходит линия подколенной мышцы. На дистальном эпифизе имеется суставной блок, отграниченный справа и слева лодыжками. На блоке,
латеральной и медиальной лодыжках есть желоба и гребни, идущие по суставной поверхности спереди назад в сагиттальной плоскости.
Малая берцовая кость — fibula — редуцировалась почти полностью
(II). Остаток ее головки прирастает с латеральной стороны к латеральному
мыщелку большой берцовой кости. Остаток дистального эпифиза превращается в латеральную лодыжковую кость, туго сочленяющуюся с дистальный
эпифизом большой берцовой кости.
У свиньи большая берцовая кость имеет сбоку у латерального мыщелка
и у дистального суставного блока суставные поверхности для сочленения с
малой берцовой костью. Малая берцовая кость самостоятельная, развита на
всем протяжении, имеет вид тонкой уплощенной палочки с утолщениями на
концах. Дистальный конец малой берцовой кости образует латеральную лодыжку.
У лошади большая берцовая кость имеет сбоку у латерального мыщелка
суставную поверхность для сочленения с головкой малой берцовой кости.
Малая берцовая кость редуцирована, обычно самостоятельна, но может и
прирастать к латеральному мыщелку большой берцовой кости. Тело ее в виде
шиловидного отростка, истончается и к середине большой берцовой кости
сходит на нет. Между костями находится межкостное пространство. Дистальный конец малой берцовой кости срастается с эпифизом большой берцовой кости, образуя латеральную лодыжку.
Скелет стопы состоит из костей заплюсны, плюсны и пальцев.
Кости заплюсны — ossa tarsi — короткие, лежат в 3 ряда (рис. 65). Проксимальный ряд образован двумя костями. С медиальной стороны и несколько впереди лежит таранная кость — talus. Она имеет большое количество
суставных площадок для сочленения с другими костями заплюсны, а впереди
— суставной блок для сочленения с костями голени. С латеральной стороны
и несколько сзади лежит пяточная кость — calcaneus — с большим, выступающим назад пяточным бугром, держателем таранной кости, направленным
медиально, и рядом суставных поверхностей.
В среднем ряду заплюсны у млекопитающих лишь одна плоская широкая кость — центральная — os tarsi centrale. В дистальном ряду заплюсны
пятипалой конечности имеется 5 костей, называемых порядковыми номерами: заплюсневые I, II, III, IV и V — ossa tarsi primum, secundum, tertium, quartum et quin-tum. Счет ведется с медиальной стороны.
У крупного рогатого скота в заплюсне б свободных костей. Суставной
блок таранной кости расположен в сагиттальной плоскости. Заплюсневая I
кость очень маленькая, заплюсневые II и III срослись. IV и V срослись между собой и с центральной костью.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 65. Кости заплюсны:
А — крупного рогатого скота; Б— лошади; В — свиньи; 1 — таранная
кость; 2 — блок таранной кости; 3 — пяточная кость; 4 — пяточный бугор; 5 — держатель таранной кости: Ь — центральная кость; 7— I заплюсневая кость; 8— II заплюсневая кость; 9 — III зaплюсневая кость; 10—
IV и V заплюсневые кости; 11— II плюсневая кость; 12 — III плюсневая
кость; 13 — IV плюсневая кость; 14 — V плюсневая кость.
У свиньи 7 свободных костей: в проксимальном ряду — таранная и пяточная, в среднем — центральная, в дистальном — I, II, III и IV+V, сросшиеся в одну.
У лошади 6 свободных костей. Суставной блок таранной кости поставлен косо. В дистальном ряду I срастается со II, а IV с V.
Кости плюсны — ossa metatarsi — длинные трубчатые, напоминают пястные. На проксимальном эпифизе имеют суставную поверхность для костей
заплюсны и связочные бугорки по бокам. На дистальном эпифизе — суставной блок и связочные ямки по бокам. Количество костей плюсны у животных
разных видов может быть от 1 до 5.
У крупного рогатого скота II плюсневая кость — os metatarsal secundum
— имеет вид горошины и лежит с медиальной стороны, III и IV плюсневые
кости — ossa metatarsale tertium et quartum — срослись в одну. В месте сращения по дорсальной поверхности кости проходит продольный желоб, а дистальнын суставной блок разделен межблоковой вырезкой. На проксимальном
конце плоская суставная поверхность и шероховатость III плюсневой кости.
Плюсневая кость рогатого скота длиннее и мощнее пястной, в поперечнике
имеет вид четырехугольника.
У свиньи 4 плюсневые кости: II, III, IV и V. Все они самостоятельные. III
и IV развиты лучше, так как они являются основными.
У лошади развита полностью только III плюсневая кость-. На ее проксимальном эпифизе видна шероховатость. Она длиннее и массивнее пястной
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
кости и в поперечнике почти округлая. По бокам расположены II и IV плюсневые кости в виде палочек, утолщенных на проксимальном конце. С возрастом срастаются с III плюсневой костью.
Кости пальцев — ossa digitorum — имеют такое же строение и то же количество, как и на грудной конечности. Сесамовидные кости пальцев такие
же, как на грудной конечности.
СОЕДИНЕНИЕ КОСТЕЙ СКЕЛЕТА (АРТРОЛОГИЯ)
Виды соединений. Кости в скелете соединены друг с другом с разной
степенью подвижности в зависимости от выполняемой функции: являются ли
они защитой для нежных органов, площадью для прикрепления мышц или
рычагами при движении и опоре.
Соединения костей бывают непрерывные и прерывные.
Непрерывные соединения, или синартрозы, возникают при сращении
костей или их частей друг с другом. В зависимости от того, какой тканью соединены кости, различают следующие виды синартрозов:
1. Мышечное соединение (синсаркоз)—присоединение лопатки к осевому скелету с помощью мышц, практически не ограничивающее движений.
2. Фиброзное соединение — сращение с помощью соединительной ткани. Если между соединенными костями имеются сравнительно широкие соединительнотканные прослойки, образуются синдесмозы. Таковы соединения
костей предплечья у всеядных, атланта с затылочной костью, дуг позвонков.
Синдесмозы в наименьшей степени ограничивают подвижность.
3. Хрящевое соединение (синхондроз) —сращение с помощью хрящевой
ткани. Подвижность синхондрозов во многом зависит от толщины и вида
хряща, соединяющего кости. Волокнистый хрящ, соединяющий тела позвонков, позволяет большую подвижность, чем гиалиновый хрящ. Разновидность
хрящевого соединения — симфиз — соединение тазовых костей с образованием щелевидной полости внутри хрящевой основы.
4. Костное соединение (синостоз)—полное неподвижное сращение костей с помощью костной ткани. У взрослого животного синостозами замещены многие фиброзные и хрящевые соединения: швы между костями черепа,
эпифизарные хрящи в трубчатых костях, соединение костей таза в безымянную кость.
Прерывные (синовиальные) соединения, или суставы (диартрозы)—
это такие сочленения, когда кости, сохраняя самостоятельность и подвижность, оказываются объединенными в единую структуру — сустав, обеспечивающую движения в том или ином участке скелета. В каждом суставе различают: суставную капсулу, герметически отграничивающую сустав от окружающих структур; суставную полость, расположенную между сочленяющимися костями и заполненную синовиальной жидкостью; суставной гиалиновый хрящ, покрывающий суставные; поверхности костей. Многие суставы
имеют связки — веревчатые тяжи из плотной соединительной ткани, соединяющие кости и расположенные снаружи от капсулы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Суставная капсула двухслойная. Наружный слой — фиброзный, внутренний — синовиальный. Фиброзный слой образован плотной соединительной тканью, которая является продолжением надкостницы и переходит с одной кости на другую. Он выполняет функцию механической защиты. Синовиальный слой представляет собой пластинку рыхлой соединительной ткани,
выстланную 1—3 слоями клеток, вырабатывающих синовиальную жидкость.
В рыхлой соединительной ткани синовиального слоя проходит большое количество нервов, кровеносных и лимфатических сосудов. Синовиальная
жидкость вязкая, тягучая, соломенно-желтого цвета, служит в качестве смазки суставных поверхностей костей, уменьшая трение и выполняя буферную
функцию.
Виды суставов по строению. Построению суставы делят на простые и
сложные. Простые суставы образованы двумя костями, между которыми нет
никаких прослоек или прокладок. В сложных суставах между длинными сочленяющимися костями находятся прокладки в виде хрящей или коротких
костей, или внутрисуставные связки.
Виды суставов по движению. Основные движения в суставах: разгибание, сгибание, отведение, приведение, вращение. Разгибание — экстензия
(extensio)—движение, при котором угол, образуемый костями сустава, увеличивается, противоположные концы костей расходятся. Сгибание — флексия (flexio)—движение, при котором суставной угол уменьшается, противоположные концы костей сближаются. Отведение — абдукция (abductio)—
движение в латеральную сторону (от срединной сагиттальной плоскости).
Приведение — аддукция (adductio)—движение в медиальную сторону (к срединной сагиттальной плоскости). Вращение — ротация (rotatio) —может
быть либо передней поверхностью сустава наружу — супинация (supinatio),
либо внутрь — пронация (pronatio).
Характер движения в суставе в основном определяется формой трущихся поверхностей сочленяющихся костей. Различают одно-, двух- и многоосные суставы. Для одноосных суставов характерными являются на одной стороне — цилиндрическая или блоковая поверхность, на другой стороне —
плоское или соответствующее ей по форме углубление. В таких суставах возможно движение по одной оси в одной плоскости: разгибание и сгибание
(например, локтевой сустав). В двухосных суставах на одной стороне эллипсоидной формы выпуклость, а на противоположной стороне — такой же
формы углубление. Движение в них возможно в двух плоскостях по двум
осям: разгибание, сгибание, отведение, приведение (например, атлантозатылочный сустав). В многоосных суставах на одной стороне шарообразная
головка, на другой — чашеобразная ямка. В них возможны различные виды
движений по многим осям во многих плоскостях, в том числе разгибание,
сгибание, отведение, приведение, супинация, пронация (например, тазобедренный сустав).
Соединение костей черепа. Большинство костей черепа соединено непрерывно с помощью швов. У молодых животных швы выполнены соединительной тканью (фиброзное соединение), которая с возрастом замещается
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
хрящевой, а потом костной, возникает синостоз. Подвижными в течение всей
жизни остаются нижнечелюстная и подъязычная кости.
Височно-нижнечелюстной сустав образован суставным бугорком чешуи височной кости и мыщелковым (суставным) отростком нижнечелюстной
кости. По строению он сложный, так как между сочленяющимися поверхностями костей имеется хрящевая прокладка — суставной диск. По движению
сустав двухосный, поскольку форма трущихся поверхностей эллипсоидная.
Кроме капсулы, сустав имеет латеральную и каудальную связки.
В подъязычной кости суставными капсулами соединены ее членики. С
шиловидным (подъязычным) отростком височной кости проксимальный членик подъязычной кости образуют височно-подъязычный сустав.
Соединение костей стволового скелета. Соединения позвонков разнообразны. Между затылочной костью и двумя первыми шейными позвонками
образуются суставы. Остальные позвонки в области тел связаны друг с другом синхондрозом, в области суставных отростков — суставами. Остальные
отростки связаны синдесмозами.
Атланто-затылочный сустав образован мыщелками затылочной кости
и краниальной суставной ямкой атланта. По строению он простой, по движению двухосный. Кроме капсулы, имеет две слабо натянутые боковые связки
и две мембраны — вентральную и дорсальную.
Атланто-осевой сустав образован каудальной суставной ямкой атланта
и вентральной суставной поверхностью осевого позвонка. Этот сустав одноосный, позволяющий вращательные движения вокруг зуба осевого позвонка,
и сложный, так как под капсулой внутри сустава проходят связки, соединяющие между собой позвонки и затылочную кость.
Тела остальных позвонков связаны между собой синхондрозом в виде
межпозвоночных дисков (рис. 66). Периферическая часть межпозвоночного
диска образована волокнистым хрящом и называется волокнистым кольцом.
В центре его находится пульпозное ядро — остаток хорды. Чем толще волокнистые кольца, тем подвижнее связь между позвонками и лучше их рессорные свойства. Наиболее толстые межпозвоночные диски между шейными позвонками. По дорсальной поверхности тел позвонков от 2-го шейного позвонка до крестца проходит дорсальная продольная связка, по вентральной
поверхности тел в области поясницы — вентральная продольная связка.
Суставные отростки соседних позвонков связаны между собой капсулами. Остальные отростки позвонков связаны синдесмозами: между остистыми
отростками натянуты межостистые связки, между поперечными и поперечнореберными — межпоперечные. В области шеи развита (не у свиньи) мощная выйная связка. Она образована эластической тканью и состоит из двух
частей: канатика и пластинки. Канатик начинается от чешуи затылочной
кости капюшонообразно, двойным тяжом тянется по шее, формируя ее дорсальную границу и прикрепляется к самым длинным остистым отросткам
грудных позвонков области холки. Далее переходит в надостистую связку,
которая соединяет вершины остистых отростков грудных, поясничных и крестцовых позвонков и заканчивается на крестцовых буграх подвздошных косwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тей. Пластинка в виде отдельных эластических тяжей отходит от остистых
отростков 2—7 шейных позвонков и вплетается в канатик выйной связки.
Рис. 66. Соединения костей стволового скелета:
А — соединения позвонков и ребер; Б— выйная связка; 1 — волокнистое
кольцо; 2— пульпозное ядро; 3— вентральная продольная связка; 4 — соединительная связка головок ребра; 5 — дорсальная продольная связка; 6 — соединение шейки ребра с телом позвонка; 7— соединение бугорка ребра с поперечным отростком позвонка; 8 — канатик выйной связки; 9 — пластинка
выйной связки; 10 — межостистые связки; 11— надостистая связка.
Соединение частей полного костного сегмента. Головка ребра с глубокой реберной ямкой позвонка образуют сустав и соединены капсулой и
связками; между головками двух ребер, входящих в сегмент, проходит соединительная связка; бугорок ребра с поперечным отростком позвонка образуют сустав и соединены капсулой и связкой; шейка ребра соединена связкой
с телом позвонка. Костные ребра с реберными хрящами соединены синхондрозами, кроме 2—5 у свиньи и 2—10 у крупного рогатого скота, где между
ними находятся тугие суставы с суставными капсулами. Реберные хрящи образуют простые суставы с реберными вырезками грудины. Ребра между собой соединены межреберными мышцами и внутригрудной фасцией. Сегменты грудины соединяются синхондрозом, а ее рукоятка причленяется к телу суставом у свиньи и жвачных.
Соединение костей грудной конечности. Лопатка причленяется к осевому скелету синсаркозом — с помощью мышц. Между звеньями конечности
образуются суставы (рис. 67).
Плечевой сустав (А) образован суставной впадиной лопатки и головкой
плечевой кости, простой, многоосный. У копытных все виды движения, кроме разгибания и сгибания, в нем ограничены утолщениями капсулы, подлоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
паточной и заостной мышцами, которые приобрели роль боковых связок изза большого количества пронизывающих их сухожильных прослоек.
Локтевой сустав (Б) образован мыщелками плечевой кости с одной
стороны, проксимальными эпифизами лучевой и локтевой костей с другой
стороны. У копытных с редуцированной и приросшей локтевой костью —
это простой одноосный сустав. Имеет капсулу, тугие межкостную и боковые
связки — медиальную и латеральную.
Запястный сустав (В) образован блоком лучевой кости, костями запястья и суставной поверхностью проксимальных эпифизов костей пясти. Одноосный, сложный сустав, так как в нем роль прокладок играют короткие
кости запястья. Фиброзный слой капсулы сустава перекидывается от лучевой
кости к пясти, не закрепляясь на костях запястья. Синовиальный слой капсулы крепится в каждом ряду костей, формируя отдельные полости. Капсула
утолщена с пальмарной стороны, что служит предохранением от переразгибания сустава. По бокам расположены латеральные и медиальные боковые
связки: длинные соединяют кости предплечья с пястью, а короткие — кости
разных рядов запястья. Связки также имеются между костями в пределах одного ряда — межкостные и между костями разных рядов — межрядовые.
Кроме того, добавочная запястная кость соединяется специальными связками
с костями предплечья, запястья и пясти.
Рис. 67. Соединение костей грудной конечности:
А — плечевой сустав лошади с дорсальной поверхности; Б — локтевой
сустав с медиальной поверхности; В — запястный сустав с дорсальной поверхности; Г — суставы пальцев с латеральной поверхности; 1 — капсула
сустава; 2— утолщение капсулы; 3— боковая связка; 4— длинная боковая
латеральная связка; 5—короткая боковая медиальная связка; 6 — длинная
боковая медиальная связка; 7 — межкостная связка; 8 — межрядовая связwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ка; 9 — связки сесамовидной кости; 10 — дорсальная связка; 11 — пальмарная связка; 12 — третья межкостная мышца; 13 — сухожилия мышц разгибателей пальцев; 14 — сесамовидная кость.
Суставы пальцев (Г) простые, одноосные. Путовый (пястнопальцевый)
сустав образован блоком пястных костей и проксимальными эпифизами путовых костей. Имеет капсулу, тугие боковые связки и большое количество
связок сесамовидных костей, лежащих на пальмарной поверхности сустава:
прямая, косые, крестовидные связки сесамовидных костей, которые у рогатого скота соединяют III и IV пальцы. К числу связок сесамовидных костей
следует отнести и третью межкостную мышцу, превратившуюся в мощную
связку на пальмарной поверхности, предохраняющую сустав от переразгибания.
Рис. 68. Соединение костей тазовой конечности (крупного рогатого скота):
А—соединение костей таза (с латеральной стороны); Б— коленный сустав с
дорсолатеральной поверхности; В — срединный сагиттальный распил коленного сустава; Г — заплюсневый сустав с латеральной поверхности; 1 —
крестцово-подвздошная дорсальная короткая связка; 2 — крестцовоподвздошная дорсальная длинная связка; 3 — крестцово-бугорковая связка; 4
— боковые связки; 5 — прямые связки коленной чашки; 6 — крестовидная
связка; 7 — бедроменисковая связка; 8 — берцовоменисковая связка; 9— боковая длинная связка; 10 — боковая короткая связка; 11 — межкостная
связка; 12 — межрядовая связка; 13 — заплюсневая плантарная связка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Венечный (межфаланговый проксимальный) сустав образован дистальным концом путовой и проксимальным концом венечной кости. Имеет капсулу, боковые и пальмарные связки на каждом пальце. Копытцевый (копытный, дистальный межфаланговый) сустав образован дистальным концом венечной кости и суставной поверхностью копытцевой (копытной) кости. Имеет капсулу, тугие боковые связки и связки сесамовидной (челночной) кости,
лежащей в соответствующей ямке на пальмарной стороне: боковые путовочелночные, челночнокопыт-ную. У рогатого скота имеются межпальцевые
связки.
Соединение костей тазовой конечности (рис. 68). Две безымянные (тазовые) кости соединяются тазовым симфизом, образованным волокнистым
хрящом. Запертое отверстие затянуто запирательной мембраной, в которой
осталось отверстие (у лошади — два) для прохождения сосудов и нервов. С
крестцом безымянная кость соединяется (А) крестцово-бугорковой (крестцово-седалищной) связкой, протянувшейся между боковой частью крестца, седалищной остью и бугром седалищной кости, а также крестцовоподвздошными связками и некоторыми другими. Крестцово-подвздошный
сустав, образованный ушковидными поверхностями крестцовой и подвздошной костей тугой, неподвижный, укреплен связками.
Тазобедренный сустав образован суставной ямкой (впадиной) безымянной кости и головкой бедренной кости, простой, многоосный. Внутри
сустава для его упрочения имеется круглая связка головки, а у лошади, кроме
того, добавочная связка. У копытных отведение, приведение и ротация в суставе ограничены в результате тесной связи сустава с глубокой ягодичной и
наружной запирательной мышцами и связками.
Коленный сустав (Б, В) образован дистальный эпифизом бедренной
кости, проксимальным эпифизом костей голени и коленной чашкой; он
сложный одноосный. Делят его на два сустава: бедроберцовый и бедрочашечный. Бедроберцовый сустав образован мыщелками бедренной кости и
костями голени, сложный одноосный. Между сочленяющимися костями расположены латеральный и медиальный мениски. Внутри сустава имеются крестовидные, бедро- и берцовоменисковые связки. Снаружи от капсулы проходят боковые связки. Бедрочашечный сустав образован блоком бедренной
кости и коленной чашкой, простой одноосный. Кроме капсулы, имеет боковые связки и три прямые связки коленной чашки.
Заплюсневый (скакательный) сустав (Г) образован дистальный концом костей голени, костями заплюсны и проксимальным концом костей
плюсны. Одноосный сложный сустав, так как роль прокладок в нем играют
короткие кости заплюсны. Как и в запястном суставе, фиброзный слой капсулы перекидывается от костей голени к плюсне, минуя кости заплюсны. Синовиальный слой образует отдельные полости, закрепляясь на каждом ряду
костей заплюсны. Связочный аппарат: латеральные и медиальные длинные и
короткие связки, межкостные, межрядовые, плантарные и дорсальные
связки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Суставы пальцев по строению, движению и связочному аппарату такие
же, как на грудных конечностях.
Глава 13. МУСКУЛАТУРА (УЧЕНИЕ О МЫШЦАХ — МИОЛОГИЯ)
ОБЩАЯ ХАРАКТЕРИСТИКА И ЗНАЧЕНИЕ МУСКУЛАТУРЫ
Мускулатура — активная часть аппарата движения. С ее помощью осуществляется движение животного в окружающей среде — локомоция и разнообразные движения отдельных частей организма и его органов (например,
головы, шеи, дыхательные, жевательные, перистальтические, сердцебиение и
т. д.). В зависимости от характера двигательной функции, особенностей происхождения и иннервации мускулатуру делят на соматическую и висцеральную.
Соматическая мускулатура построена из поперечнополосатой мышечной
ткани; произвольная, иннервируется периферической (соматической) нервной системой. Основная ее масса оснащает скелет (скелетная мускулатура).
Встречается она под кожей — подкожная мускулатура, образует грудобрюшную преграду — диафрагму, содержится в гортани, среднем ухе, наружных
половых органах, приводит в движение глазное яблоко.
Висцеральная мускулатура в основном построена из гладкой мышечной
ткани, непроизвольная, иннервируется вегетативной нервной системой. Образует мышечные пучки, слои, оболочки внутренних органов и составляет
около 8 % массы организма. Особое место занимает она в висцеральном аппарате головы, появившись в филогенезе в стенке передней кишки. Параллельно со сложными преобразованиями висцерального скелета эта мускулатура дифференцировалась в поперечнополосатую и образовала большинство
мышц головы, часть мышц шеи (трапециевидную, плечеголовную, грудиноголовную), мышцы глотки, простираясь на более или менее значительную
часть пищевода (у жвачных до желудка).
Скелетная мускулатура как составная часть входит в локомоторный аппарат, или систему органов произвольного движения. На ее долю в аппарате
движения приходится 68—75 %. что составляет у рогатого скота 30—37 %
массы животного, у лошади 35—38 %, у свиньи 30—35 %. Скелетная мускулатура состоит из органов — многочисленных и разнообразных мышц. У копытных их насчитывается более 500 (250 парных и несколько непарных).
Мышцы обладают раздражимостью, сократимостью и упругостью. Раздражаясь под влиянием нервного импульса, они сокращаются, а в расслабленном состоянии сохраняют упругость. Скелетные мышцы сокращаются быстро, энергично, но кратковременно. Такой тип сокращения называется тетаническим. Следующие друг за другом волны сокращения приводят к утомлению. Но и в состоянии покоя мышцы находятся в некотором напряжении —
тонусе. Благодаря тонусу мышцы немедленно реагируют на раздражение, не
затрачивая энергию и время на предварительную подготовку к сокращению.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Мускулатура в своей деятельности тесно связана со многими системами
организма. Наиболее тесная связь между мышечной и нервной системами.
Она устанавливается с момента появления мышечной ткани как в фило-, так
и в онтогенезе и никогда не нарушается. Разрыв этой связи приводит к прекращению функционирования мышцы. Филогенетические преобразования
той или иной мышцы сопровождаются коррелятивными изменениями связанного с ней нерва: при увеличении мышцы — увеличивается, при уменьшении — уменьшается и нерв. При расчленении на несколько мышц разъединяется и нерв на несколько ветвей. Мышцы, образованные слиянием нескольких мышц, иннервируются несколькими нервами. Столь тесная нерушимая связь позволяет прослеживать пути развития тех или иных мышц.
Значение и функции мышечной системы. Основная функция мускулатуры — динамическая. Сокращаясь, мышца укорачивается на 20—50 % своей
длины и тем самым меняет положение связанных с ней костей. Производится
работа, результатом которой является движение. Двигательная функция
чрезвычайно важна для жизнедеятельности организма. Можно сказать, что
движение является условием существования организма. Без него нарушается
деятельность нервной и сердечно-сосудистой систем, аппаратов пищеварения, дыхания, выделения, размножения, желез внутренней секреции.
Другая функция мышечной системы — статическая. Проявляется она в
фиксации тела в определенном положении, в сохранении формы тела и его
частей. Одно из проявлений этой функции — способность спать стоя, развитая у некоторых животных (лошадь). Не менее важная функция — участие
мускулатуры в обмене веществ. При сокращении мышцы лишь 30 % энергии
превращается в механическую (движение), а 70%- в тепловую. Следовательно, работа мышц — это основной источник тепла в организме. Мышечная
система играет роль жирового и водяного депо. В ней удерживается до 2/3 воды организма, а между мышцами и внутри их при откорме накапливается
большое количество жира. В состоянии покоя мускулатура экономно тратит
ресурсы организма: в то время как на ее массу приходится около 40 % массы
организма, обмен веществ в ней составляет не более 25 % общего метаболизма. Во время же активной мышечной работы он возрастает до 70 %.
Скелетная мускулатура сельскохозяйственных животных имеет большое
значение и как источник полноценной белковой пищи для человека. Среди
органических веществ мышц от 16 до 22 % приходится на долю белков, ценность которых увеличивается еще и тем, что многие из них содержат высокий процент незаменимых аминокислот.
СТРОЕНИЕ МЫШЦЫ КАК ОРГАНА
Скелетная мышца (лат. musculus) состоит из двух различных по функции и строению частей: мышечного брюшка и сухожилия. Мышечное брюшко, сокращаясь, производит работу, а сухожилие служит для закрепления
брюшка на костях, как рычагах движения. Мышечное брюшко выдерживает
нагрузку
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
7 кг/см2, сухожилие — 500—700 кг/см2. Ясно, что основная работа мышечного брюшка — динамическая — сокращение, а сухожилия— статическая, удержание тела. В аппарате движения копытных, особенно лошади,
проводящей практически всю жизнь стоя, имеются специальные приспособления для удержания тела без затраты мышечной энергии (см. ниже).
Мышца как компактный орган состоит из стромы и паренхимы. Паренхима мышечного брюшка — это поперечнополосатая мышечная ткань (см.
рис. 31). Мышечные волокна объединены соединительной тканью в пучки. В
состав пучка входят как красные (быстрые фазные), так и белые (медленные
фазные) мышечные волокна. От их соотношения зависят функциональные
качества мышцы. Так, камбаловидная мышца, состоящая преимущественно
из красных волокон, способна сохраняться в напряженном состоянии 2,5 ч, в
то время как икроножная, в которой красные волокна составляют 15—20 %,
— лишь 20мин.
8 то же время скорость сокращения белых мышечных волокон больше,
поэтому такие мышцы называются быстрыми. Тонкие коллагеновые и эластические волокна, оплетающие каждое мышечное волокно вдоль и поперек
и вступающие в тесную связь с его гликокаликсом, называются эндомизием.
Он объединяет мышечные волокна в пучки I порядка. Последние окутаны
прослойками рыхлой соединительной ткани, которая отделяет их друг от
друга и называется внутренним перимизием. Он, в свою очередь, формирует
пучки II, III порядков и т. д. Снаружи мышца одета футляром из плотной соединительной ткани — наружным перимизием, или эпимизием. В местах его
сильного развития образуется сухожильное зеркало — слой плотной соединительной ткани с перламутровым блеском. Эпи-, пери- и эндомизий образуют единый соединительнотканный каркас мышцы, который защищает
мышцу от чрезмерного утолщения или растяжения. По нему внутрь органа
входят и в нем разветвляются сосуды и нервы.
Сухожилие состоит из плотной соединительной ткани и построено по
тому же принципу, что и мышечное брюшко, с той лишь разницей, что вместо мышечных волокон его пучки содержат плотно упакованные коллагеновые волокна. Сухожилие обладает огромной прочностью на разрыв 600—900
кг на 1 см2. Прослойки соединительной ткани внутри сухожилия носят названия эндо- и перитенония. Соединительная ткань, одевающая сухожилие
снаружи и являющаяся продолжением эпимизия, называется наружный перитеноний, или эпитеноний. Коллагеновые волокна сухожилия прочно соединяются своими концами с мышечными волокнами брюшка, вплетаясь в
выросты их сарколеммы и гликокаликса. Соединительнотканные тяжи от сухожилия могут проникать на разную глубину внутрь мышечного брюшка и
даже пронизывать его насквозь, увеличивая прочность и силу мышцы.
Сосуды и нервы проникают в мышцу в области ворот — участок мышечного брюшка, где механическое воздействие оказывается наименьшим.
Обычно это проксимальный участок мышцы, несколько выше ее геометрического центра.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Спинномозговые (или черепномозговые) нервы, разветвляясь в мышце,
достигают мышечных волокон и образуют на них двигательные (моторные
бляшки) или чувствительные (мышечные веретена) нервные окончания (проприорецепторы). Терминали одного нервного волокна обычно бывают связаны со многими мышечными волокнами (от 10 до 3000). Нервное волокно с
ин-нервируемыми им мышечными волокнами образует нервно-мышечный
комплекс, или мышечную функциональную моторную единицу — мион. Количество мышечных волокон, входящих в один мион, зависит от характера
двигательной активности мышцы и тонкости ее дифференцировки.
Так, в прямой латеральной мышце глаза одно нервное волокно приходится на 19 мышечных, а в трехглавой мышце голени — 227. Существует
мнение, что волокна, входящие в состав одного миона, могут располагаться
на значительных расстояниях друг от друга, входить в состав разных мышечных пучков. Нервы вегетативной симпатической нервной системы, разветвляясь в мышце, иннервируют ее сосуды, регулируют тонус сосудистых стенок, а следовательно, и уровень кровоснабжения мышцы.
В мышцу обычно входит несколько артерий, коллатерально разветвляющихся в мышце, что обеспечивает бесперебойное кровоснабжение и быстрый отток и приток крови в зависимости от функциональной необходимости. В покое в мышце может функционировать лишь 1/10 часть ее капилляров. При активном движении кровоснабжение ее увеличивается в 30 раз.
Цвет мышцы определяется как составом образующих ее волокон, так и количеством крови, протекающей через нее. Из сельскохозяйственных животных
наиболее темные мышцы у лошади.
В соединительнотканном каркасе мышцы встречаются скопления жировых клеток, придающих мясу мраморный цвет. Размеры жировых клеток
(40—60 мкм) сопоставимы с размерами волокон (50—70 мкм). В пучках II и
III порядков жировой ткани больше, чем в эндомизии и перимизии пучков I
порядка. Много жировых скоплений по ходу сухожилий, особенно в мышцах
статодинамического типа.
КЛАССИФИКАЦИЯ СКЕЛЕТНЫХ МЫШЦ
Мышцы делят по происхождению, топографии (расположению на теле),
форме, выполняемой функции, по особенностям внутренней структуры.
По происхождению мышцы бывают соматические и висцеральные.
По топографии различают мышцы отдельных областей тела (шеи, плеча
и т. д.).
Форма мышц разнообразна и зависит от положения на теле, происхождения, выполняемой функции. В осевой части тела располагаются обычно
пластинчатые мышцы, которые могут быть широкими и узкими, длинными и
короткими, треугольными, ромбовидными, веерообразными и др. Они имеют
широкие пластинчатые сухожилия — апоневрозы. Если мышцы кончаются
на нескольких, следующих друг за другом однотипных костях (ребрах, позвонках) отдельными порциями или пучками, они называются многораздельwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ными, зубчатыми. Вокруг естественных отверстий залегают кольцевидные
или полукольцевидные мышцы. На конечностях форма мышц в основном веретенообразная— в виде цилиндрических или конических образований с хорошо развитым брюшком, коротким начальным и длинным веревчатым конечным сухожилием.
По функции мышцы делят в зависимости от вида движения, осуществляемого с их помощью, на разгибатели — экстензоры, сгибатели — флексоры, отводящие — абдукторы, приводящие— аддукторы, вращатели — ротаторы (в том числе вращающие наружу — супинаторы и внутрь — пронаторы), расширители— дилятаторы, сжиматели — констрикторы, запиратели — сфинктеры, подниматели — леваторы, опускатели — депрессоры,
оттягивающие — ретракторы, подтягивающие — протракторы, напрягатели — тензоры.
Мышцы, выполняющие одинаковую функцию, называют синергистами,
противоположную — антагонистами. Мышца может действовать как на
один, так и на несколько суставов. В последнем случае функция ее в разных
суставах оказывается неодинаковой. Так, двуглавая мышца плеча является
экстензором плечелопаточного сустава и флексором — локтевого. Мышца
может совершать основное и побочное действие. Так, для средней ягодичной
мышцы основной функцией будет разгибание тазобедренного сустава. Разгибая сустав, она одновременно несколько отводит конечность в сторону (побочное действие).
По внутренней структуре мышцы делят в зависимости от количества
внутримышечных сухожильных прослоек и различного направления связанных с ними пучков мышечных волокон на одноперистые (нет сухожильных
прослоек), двуперистые (одна прослойка), многоперистые (две и больше).
Названия мышцам дают не произвольно, а по какому-либо характерному признаку, связанному с их классификацией: по функции (сгибатель), по
форме (круглая), по направлению мышечных волокон (поперечная), по величине (большая), по названиям костей и областей, на которые действуют или
около которых лежат (плеча, поясничный), по названиям ямок, площадок, где
они залегают (предостная), по количеству головок (трехглавая), брюшек
(двубрюшная). В названии мышцы для более точного отличия ее от других
мышц обычно отражено несколько признаков, поэтому оно состоит из нескольких слов: лучевой разгибатель запястья, остистая и полуостистая мышца спины и шеи.
ТИПЫ МЫШЦПО ВНУТРЕННЕЙ СТРУКТУРЕ
Структура мышцы тесно связана с ее функцией (рис. 69). Функционирование мышц в определенных условиях (силовой нагрузки, расположения точек прикрепления и т. д.) обусловливает особенности ее внутренней структуры. Точно так же внутренняя структура мышцы во многом определяет особенности ее функционирования: одни мышцы больше приспособлены для
силовой, другие — для скоростной работы. На этом основании П. Ф. Лесгафт
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
предложил делить мышцы на сильные и ловкие и ввел понятие анатомического и физиологического поперечников, с помощью которых можно было
бы определять силу мышцы и производимую ею работу. Анатомический поперечник — это площадь поперечного сечения, проведенная через середину
(в веретенообразных мышцах — в самом широком месте) мышцы. Физиологический поперечник— площадь поперечного сечения, проведенная перпендикулярно ко всем пучкам мышечных волокон. В простых одно-перистых
мышцах анатомический и физиологический поперечники или равны, или отличаются несильно. С усложнением внутренней структуры (в двух- и особенно в многоперистых мышцах) физиологический поперечник оказывается
намного больше анатомического. Мышечные волокна в таких мышцах короткие, так как идут не вдоль мышцы (как в одноперистой), а прикрепляются
к сухожильным прослойкам, располагаясь под разными углами. Количество
же мышечных волокон в многоперистой мышце оказывается в несколько раз
больше, чем в одноперистой, имеющей одинаковый с ней анатомический поперечник.
Рис. 69. Типы мышц по внутренней структуре:
А — динамический тип; Б— динамостатический тип, одноперистая
мышца: В — полустатодинамический тип, двуперистая мышца: Г — статодинамический тип, многоперистая мышца; а—а — анатомический поперечник; б—б — физиологический поперечник; 1 — мышечные пучки; 2 — сухожильные прослойки; 3 — сухожильное зеркало
Усложнение внутренней структуры мышцы увеличивает ее силу, но
уменьшает размах сокращения. Сила мышцы рассчитывается как соотношение физиологического и анатомического поперечников. Однако для полной
характеристики мышцы определения только ее силы недостаточно, тем более
что этот показатель в разных мышцах колеблется в широких пределах: от 5
до 13,7 кг на 1 см2.
Большое внимание изучению внутренней структуры мышц уделяли
школы А. Ф. Климова и П. А. Глаголева. Их трудами обосновано, что тип
мышцы должен характеризоваться комплексом показателей, включающих
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
анатомические, гистологические и биохимические данные. При этом мышцы
разделяются на пять морфофункциональных типов: динамический, динамостатический, полустатодинамический, статодинамический, статический.
Для мышц динамического типа характерно: наличие нежного соединительнотканного остова; отсутствие внутри мышечного брюшка развитых сухожильных прослоек; направление мышечных пучков, параллельное длине
мышечного брюшка; меньшее количество мышечных пучков; большая относительная длина мышечных пучков; меньшая площадь их прикрепления;
почти равное отношение физиологического поперечника (ФП) к анатомическому поперечнику (АП); наименьший диаметр мышечных волокон;
количество мышечной ткани во много раз превышает количество соединительной ткани; незаменимые аминокислоты преобладают над заменимыми,
высокий белковый качественный показатель (БКП). Мышцы динамического
и динамостатического типов одноперистые, быстрые, приспособленные к
динамической работе. Сокращаются почти на половину своей длины, но при
этом утомляются быстрее, чем более статичные мышцы. В мышцах динамического типа у свиней площадь одного волокна равна 150 мм2, площадь пучка I порядка— 13 мм2, мышечная ткань составляет 74%, соединительная— 11
%, жировая — около 13%. По мере увеличения статичности мышцы происходит (табл. 5): постепенное уплотнение соединительнотканного остова; все
большая выраженность мышечного зеркала и увеличение количества сухожильных прослоек внутри мышечного брюшка; косое направление мышечных пучков; увеличение угла наклона мышечных пучков к месту их прикрепления; увеличение количества мышечных пучков; уменьшение относительной длины мышечных пучков; увеличение отношения ФП к АП; увеличение
диаметра мышечных волокон; уменьшение отношения мышечной ткани к соединительной; уменьшение доли незаменимых и увеличение доли заменимых
аминокислот, снижение БКП; снижение содержания белка и увеличение содержания жира.
Мышцы более статичных типов двух- и многоперистые, сильные, приспособленные как к динамической, так и к статической работе. Сокращаясь
на 25%, а некоторые лишь на 10% своей длины, они мало утомляются и способны к длительной работе. Мышцы статического типа утрачивают мышечные элементы, как, например, 3-я межкостная мышца взрослых копытных
животных, и превращаются в мышцы-связки, выполняющие только статическую работу. Указанная мышца предохраняет пальцевые суставы от переразгибания. Преимущественно статическую работу выполняют и некоторые
мышцы статодинамического типа: заостная, подлопаточная, 3-я малоберцовая и др., пронизанные насквозь сухожильными прослойками, которые составляют единый тяж с начальным и конечным сухожилиями этих мышц. В
статодинамических мышцах свиней волокна в 2 раза крупнее, чем в динамических (390 мкм2), а пучки в 2,5 раза мельче (6 мм2), мышечная ткань составляет 74%, соединительная 20 %, жировая 5 % (А. Г. Корсан).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
5. Морфохимические показатели мышц разных типов
Типы мышцы
динамо- полуста- статоПоказатели
динамистатичестатиче- тодина- динамический
ский
ский мический ческий
Отношение ФП к АП
8 (и
Мышеч1—1,7: 1 2—4:1
4-8:1 больше):
ные
1
Отношение длины мышеч40 и
Пучки и
20—40 10—20 1 — 10
ного пучка к длине мышечбольше
волокна
ного брюшка, %
Угол наклона мышечного
35 и Отсутст5—20 15—30 25—40
пучка, град
больше
вуют
Количество сухожильных
2 и боНет
0—1
1—2
прослоек, шт.
лее
Диаметр мышечных воло40 и бокон (мкм) у крупного рога- 20—40 25—45 30—35
лее
того скота
Отношение незаменимых
5—12:1 3—8:1
1,5-3:1 0,7—1:1
аминокислот к заменимым
Мышцы пяти морфофункциональных типов располагаются на теле неравномерно. Наиболее массивные и более близкие к динамическому типу
лежат на туловище и верхних звеньях конечностей. Причем на тазовых конечностях мышцы ближе к динамическому типу, чем на грудных. Чем дистальнее расположены мышцы на конечностях копытных и чем на более дистальное звено они действуют, тем они все больше уклоняются в сторону статичности. Так, у лошади мышцы, действующие на лопатку, имеют в среднем
ФП : АП = 1,7:1, действующие на плечо — 3,7:1; на пясть — 8,2:1, на пальцы
— 17,6:1. С возрастом мышцы динамического типа становятся еще динамичнее, статодинамического — статичнее.
В волокнах динамостатических и статодинамических мышц почти в 2
раза увеличивается плотность расположения миофибрилл за счет возрастания
их количества. В динамических мышцах в волокнах преобладает саркоплазма
(70—63 % от площади волокна).
Образ жизни животного и способ его передвижения накладывают отпечаток на типологический характер мускулатуры. У пальцеходящих хищных,
способных к разнообразным ловким движениям и питающихся малообъемистой концентрированной пищей, мышцы преимущественно динамического и
динамостатического типов. У всеядных (свинья) мускулатура более статична,
чем у хищных, но более динамична, чем у других копытных. Из жвачных самая динамичная мускулатура у овцы. У крупного рогатого скота и козы —
более статичная. Наибольшей сложностью внутренней структуры отличаются мышцы у лошади.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ДЕЙСТВИЕ МЫШЦ ПРИ ДВИЖЕНИИ И СТОЯНИИ ЖИВОТНОГО
Движения животного можно разделить на две группы: локомоторные,
приводящие к перемещению животного в пространстве (ходьба, бег, прыжки), и частные, деформирующие — изменяющие расположение частей организма или органов (головы, конечности, губ, щек, дыхательные движения и т.
д.) без перемещения всего животного.
Любое движение в организме всегда (в норме) осуществляется координированно и связано, как правило, с работой нескольких или даже многих
мышц. Все мышцы, участвующие в том или ином движении, объединены в
так называемую кинематическую цепь из-за того, что сокращаются в определенной последовательности. Определенная последовательность включения
той или иной мышцы в двигательный акт зависит от нескольких причин, в
том числе от строения сустава, расположения мышцы, ее морфологической
связи (с помощью сухожилий, апоневрозов, фасций, общей иннервации) с
другими мышцами. На тазовой конечности в единую кинематическую цепь
объединены мышцы, действующие на свободную конечность, начиная от тазобедренного сустава, на грудной — не только мышцы, действующие на ее
суставы, но и присоединяющие конечность к осевой части тела. Сгибание
грудной конечности начинается под действием тяжести тела с поворота лопатки с помощью мышц, подвешивающих ее к туловищу, шее и голове (трапециевидной, ромбовидной, плечеголовной, широчайшей спины) таким образом, что ее дистальный (суставной) угол двигается вперед и вверх, а задний
— назад и вниз. Это автоматически вызывает сгибание лопаткоплечевого
сустава и включение его мышц-сгибателей в кинематическую цепь. Это движение (сгибание) с чуть заметным запаздыванием (на доли секунды) распространяется последовательно на локтевой, запястный, пальцевые суставы.
При этом чем дистальнее сустав, тем больше его сгибание. Так, угол плечевого сустава при сгибании уменьшается на 2° (по сравнению с фазой опоры),
локтевого —на 26°, запястного и пястнопальцевого — на 78° каждого. Движения в суставах настолько зависят друг от друга, что сгибание или разгибание одного из них (локтевого или коленного) автоматически влечет за собой
такое же движение в других суставах (запястном, заплюсневом, пальцевых).
Особенно жестко организованы мышцы в кинематические цепи у копытных,
приспособленных к однообразным поступательным движениям — быстрому
бегу.
Грудная и тазовая конечности при движении животного несут различную нагрузку. Это отражается и на их массе. Масса тазовой конечности
обычно на 5—15 % больше, чем грудной, так как она является основным
толкателем. Мощность ее мышц, особенно разгибателей тазобедренного, коленного и скакательного суставов, очень велика. По массе экстензоры в тазобедренном суставе в 3 раза, в коленном в 18—20 раз, в заплюсневом в 5 раз
больше флексоров. При толчке движение с тазовой конечности через крестцово-тазовое сочленение, ягодичную и заднебедренную группы мышц передается на туловище. Сокращается дорсальная мускулатура позвоночного
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
столба, что приводит к подниманию головы и шеи. При этом центр тяжести
перемещается в каудальной направлении, облегчается передняя половина тела и создаются условия для отрыва от земли, разгибания грудной конечности
и выноса ее вперед. Чем шире размах движений грудной конечности, тем
шире шаг. Это достигается широким прикреплением плечевого пояса и плеча
к осевой части тела: от атланта до поясницы. Грудная конечность выносится
вперед и подтягивает туловище.
Разгибатели в плечевом и локтевом суставах также развиты лучше, чем
сгибатели. В дистальных звеньях (в запястном и пальцевых суставах) лучше
развиты флексоры, их масса в 2 раза больше,
чем экстензоров, так как они предохраняют эти
звенья от излишнего переразгибания.
Мышцы действуют на скелет как на систему рычагов движения, прикрепляясь к подвижно соединенным костям (рис. 70). Работа мышц
осуществляется при помощи рычагов различных систем. Одни и те же части локомоторного
аппарата выступают в роли различных рычагов
в зависимости от фазы движения. Это могут
быть двуплечие — рычаги первого рода, когда
действующая и противодействующая силы находятся по разные стороны от точки опоры, что
наблюдается при разгибании суставов (например, локтевого). Чаще же образуются рычаги
второго рода, когда точка опоры находится по
одну сторону от линии действия обеих сил —
действующей и противодействующей. Среди
рычагов второго рода различают: 1) рычаг силы, когда плечо приложения силы (действующей силы) длиннее плеча сопротивления (противодействующей силы), что наблюдается в фазу опоры конечности. При этом получается выигрыш в силе, но проигрыш в скорости; 2) рыРис. 70. Мышцы как
чаг скорости, когда плечо сопротивления длинрычаги движения.
нее плеча приложения силы, что наблюдается
при сгибании сустава в фазу перенесения конечности. При этом получается
выигрыш в скорости при проигрыше в силе.
Во время стояния животного на грудную конечность падает 55% тяжести тела, на тазовую — 45%, что зависит от положения центра тяжести. В
связи с этим мышцы грудной конечности более приспособлены для статической работы, что отражается в их внутренней структуре: большая часть их
относится к статодинамическому типу. Особенно сильно развиты сухожильные прослойки в мышцах грудных конечностей лошади, у которой формируется единая сухожильная система, связывающая плечевой, локтевой, запястный и пальцевые суставы. Она является статическим аппаратом грудной
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
конечности, позволяет полностью выключить мышцы при разогнутых суставах во время отдыха.
На тазовой конечности также существует механизм фиксации суставов в
возможно более разогнутом положении, удобном для отдыха стоя. Ярче всего он выражен у лошади, хотя имеется и у крупного рогатого скота, осла,
верблюда и заключается в закреплении коленной чашки на так называемой
статической площадке, расположенной на проксимальном конце ее медиального гребня блока. Коленная чашка заводится на статическую площадку сокращением медиальной головки четырехглавой мышцы бедра. Отпирается
этот механизм при совместном сокращении четырехглавой и двухглавой
мышц бедра.
Грудная клетка выполняет три функции: защита для расположенных в
ней органов, фиксация грудных конечностей и вентиляция легких. В связи с
этим она должна одновременно обладать прочностью, устойчивостью и подвижностью. Обеспечивается это особенностями конструкции ее скелета, расположением мышц и направлением их мышечных пучков. В результате передний отдел грудной клетки оказывается более прочным и менее подвижным, особенно у копытных, а задний — более подвижным.
ВСПОМОГАТЕЛЬНЫЕ ОБРАЗОВАНИЯ МЫШЕЧНОЙ СИСТЕМЫ
К вспомогательным образованиям мышечной системы, которые улучшают условия работы мышц, относятся фасции, бурсы, связки, влагалища
мышц и сухожилий, блоки и сесамовидные кости.
Фасции (fascia — обертка)—соединительнотканные пленки, одевающие
и разобщающие мышцы. В целом фасции образуют соединительнотканный
остов сомы в дополнение к костному остову— скелету. Функции фасций
многообразны. Они являются местом прикрепления некоторых мышц; образуют футляры, обеспечивающие оптимальные условия растяжения и утолщения мышц; удерживают мышцы от сдвигания, особенно на выступах и гребнях костей; одевая группы мышц, создают условия для одновременной работы синергистов; разделяя мышцы, препятствуют их срастагшю; натяжение
фасций способствует проталкиванию крови по сосудам. Фасции образованы
как толстыми, так и тонкими пластинками плотной фиброзной соединительной ткани. Волокна в фасциях преимущественно идут поперек длины мышц
или в виде сети.
В зависимости от расположения и размеров различают поверхностную
(или общую), глубокую (или частную) и собственные (или специальные)
фасции. Поверхностная фасция расположена под кожей. Во многих местах
она разделяется на 2 листка, между которыми разрастается жировая ткань и
залегает подкожная мускулатура. Местами она соединяется с глубокой фасцией, прикрепляется к коже и костям, что позволяет при сокращении подкожной мускулатуры подергивать кожей. Из сельскохозяйственных животных лучше всего развита подкожная мускулатура у лошади, поэтому она обwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ладает самой подвижной кожей. В зависимости от расположения различают
поверхностную фасцию головы, шеи, туловища и конечностей.
Глубокая фасция отделяет группы мышц друг от друга и от серозной
оболочки, выстилающей полости тела. Ее также делят в зависимости от положения на глубокую фасцию головы, шеи, груди, брюха и конечностей. В
области туловища она формирует желтую брюшную фасцию, в состав которой входит большое количество эластических волокон, придающих ей повышенную упругость.
В тех местах, где требуются более дифференцированные движения, от
глубокой грудной фасции отходят прослойки, отделяющие отдельные мышцы друг от друга. В результате образуется целая система соединительнотканных футляров — собственных, или специальных, фасций. Наиболее выраженным из этих фасций даны названия: широкая фасция бедра, фасция предплечья и др.
Связки и влагалища. Поперечные связки
мышц (рис. 71) —утолщенные участки фасций конечностей в виде лент, которые перекидываются
через отдельные мышцы, удерживая их в определенном положении и изменяя направление действующей силы.
Фиброзные влагалища сухожилий — футляры, образованные фасциями вокруг сухожилий
мышц. Особенно развиты они вокруг сухожилий
мышц, действующих на пальцы. Для лучшего
скольжения сухожилий внутри фиброзного влагалища организуется синовиальное влагалище сухожилия — удлиненное мешковидное образование с
полостью, заполненной синовиальной жидкостью.
Синовиальное влагалище в виде кольца охватывает сухожилие, отделяя его, таким образом, от
фиброзного влагалища.
Бурсы (bursa — сумка) (см. рис. 71 и 72)—
замкнутые мешочки с полостью, заполненной
Рис. 71. Вспомога- жидкостью. Располагаются в местах активного
тельные приспособ- движения и значительного трения мышц, связок,
ления мышц в об- кожи о выступы костей. Различают врожденные,
ласти заплюсневого или постоянные, и приобретенные, или непостоянсустава лошади:
ные, бурсы. Постоянные бурсы имеются уже у но1 — поперечные ворожденных животных, непостоянные — обрасвязки; 2 — бурсы; 3 зуются в большинстве случаев в условиях непра— синовиальные вла- вильной эксплуатации животного. Наружная стенгалища сухожилие.
ка бурсы образована фиброзной соединительной
тканью. Полость бурсы выстлана клетками, вырабатывающими слизь или синовию. В зависимости от этого различают слизистые и синовиальные бурсы. Полости синовиальных бурс часто сообщаются
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
с полостями суставов. Одной своей стороной бурса закрепляется на костях,
другой — на коже, мышце, связке, сухожилии. В связи с этим различают
бурсы подкожные, подмышечные, подвязочные, подсухожильные. Последние могут преобразовываться в синовиальные влагалища сухожилий.
Рис. 72. Подкожная мускулатура, поверхностные фасции и расположение
подкожных и подсвязочных бурс на теле лошади:
1 — подкожная м. лица; 2— подкожная лопатко-плечевая м.; 3— подкожная м. туловища; 4 — поверхностная шейная фасция; 5 — грудобрюшная фасция; 6 — поверхностная фасция бедра. Овалами показаны места расположения и размеры бурс.
Блоки — определенной формы участки эпифизов трубчатых костей, выстланные гиалиновым хрящом, через которые перекидываются мышцы. Благодаря блокам мышцы и их сухожилия не смещаются в сторону. Между сухожилием и блоком обычно располагается слизистая бурса. Блоки и расширенные эпифизы костей, меняя направление проходящих мышц, особенно если мышцы закрепляются возле основания эпифиза, улучшают условия их работы. Известно, что наилучшие условия создаются при отхождении мышцы
от кости под углом 90°. Такую же функцию выполняют и сесамовидные косwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ти, располагающиеся на дистальном конце противоспинковой поверхности
костей пясти, плюсны и дистальных фаланг пальцев (см. скелет). К сесамовидным костям относятся коленная чашка и добавочная кость запястья.
КРАТКИЕ СВЕДЕНИЯ ПО ФИЛО И ОНТОГЕНЕЗУ МУСКУЛАТУРЫ
Филогенетические преобразования. Мышечные элементы в ряду развития живых существ появляются рано у кишечнополостных. Они еще не
обособлены в самостоятельные морфологические единицы, а являются лишь
сократимыми мышечными элементами эпителиальных клеток. В дальнейшем
они обособляются от эпителия, образуя несколько слоев гладкомышечных
клеток, тесно соединенных с кожей, в результате чего образуется так называемый кожно-мышечный мешок (плоские черви). Источником образования
мышечных клеток становится мезодерма.
С появлением вторичной полости тела происходит разделение мускулатуры на соматическую, входящую в состав кожно-мышечного мешка, и висцеральную, окружающую кишечник и сосуды. Несмотря на такое разделение,
она может быть либо вся гладкая (кольчатые черви), либо вся поперечнополосатая (насекомые). Это свидетельствует о том, что в филогенезе поперечнополосатая мускулатура почти не отличаются от гладкой ни по происхождению, ни по функции. С дальнейшим усложнением организации соматическая и висцеральная мускулатура развиваются дивергентно, все более
расходясь друг от друга структурно и функционально.
У примитивных хордовых (ланцетник, круглоротые) вся соматическая
мускулатура развивается из сомитов мезодермы и является поперечнополосатой. Она представляет собой парную правую и левую продольные мышцы,
идущие вдоль всего тела, поделенные соединительнотканными перегородками— миосептами на ряд миомеров — коротких сегментов из прямых мышечных пучков. Такое (сегментарное) деление единого мышечного пласта
называется метамерией (рис. 73).
С обособлением головы и развитием конечностей (в виде плавников)
дифференцируется и мускулатура. Продольная мышца у рыб делится горизонтальной септой на дорсальную и вентральную мышцы. Они иннервируются соответственно дорсальными и вентральными ветвями спинномозговых
нервов. Иннервация эта сохраняется при всех дальнейших преобразованиях
мышц. Дорсальные и вентральные продольные мышцы в связи с однообразием движений первичноводных животных имеют миомерное строение. Каждому миомеру соответствует обычно свой позвонок и парный спинномозговой нерв. У высших рыб (сельдевые и др.) можно видеть их продольное расщепление на отдельные слои. Обособляется и мускулатура плавников, однако по сравнению с мышцами ствола тела она развита слабо, поскольку основная нагрузка при движении у водных животных ложится на хвост и туловище.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 73. Мышцы тела хордовых животных:
А — ланцетник; 5 —рыба; В — хвостатая амфибия; Г —рептилия; 1 — миомеры (миотомы); 2— миосепты; 3— дорсальная м.. туловища; 4— продольная боковая перегородка; 5— дорсальная м. хвоста; 6 — поверхностный
сжиматель; 7— трапециевидная м.; 8 — вентральная м. туловища; 9 — вентральная м. хвоста; 10— мм. грудной конечности; 11 — широчайшая м, спины; 12, 13, 14 — вентральные мм. (12 — косая наружная, 13—косая внутренняя, 14 — прямая); 15 — мм. тазовой конечности.
С выходом на сушу и увеличением разнообразия движений растет расчленение мышечных пластов на отдельные мышцы как вдоль, так и поперек.
При этом метамерность постепенно исчезает. Она хорошо прослеживается в
мускулатуре рыб, заметна еще и у амфибий, слабо — у рептилий. У млекопитающих сохранена лишь в глубоких слоях, где короткие мышцы соединяют между собой элементы двух соседних костных сегментов (межостистые,
межпоперечные, межреберные мышцы).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Прежде всего метамерия начинает исчезать в брюшном отделе туловища, где уже у амфибий отдельные миомеры сливаются с образованием широких пластинчатой формы брюшных мышц. Вместе с этим идет продольное
расщепление мышечной брюшной стенки с формированием четырехслойного
брюшного пресса. В дорсальной мускулатуре туловища амфибий можно вычленить два тяжа: латеральный и медиальный, метамерия которых затушевывается лишь в шейном отделе, где обособляются самостоятельные
мышцы.
У рептилий мышечные пучки латерального и медиального мышечных
тяжей приобретают разную направленность. Миомерия сохраняется лишь в
глубоких слоях. Чем ближе к голове, тем яснее раздробление дорсальных тяжей на отдельные мышцы.
У млекопитающих соматическая мускулатура дифференцирована в наибольшей степени. В дорсальной мускулатуре формируется 4 слоя за счет расслоения латерального и медиального мышечных тяжей. При этом наблюдается четкая закономерность: чем глубже находится мышца, тем лучше
выражена ее метамерия; чем ближе к наружной поверхности тела лежит
мышца, тем больше она теряет метамерию, распространяясь по телу широким пластом. Расчлененность дорсальной мускулатуры возрастает и в краниальном направлении, что связано со степенью подвижности позвоночника.
Если в области крестца — самого неподвижного отдела стволового скелета
— дорсальные мышцы абсолютно не расчленены, то в области холки, и особенно шеи, мышечные комплексы состоят из большого количества самостоятельных мышц.
Вентральная мускулатура стволовой части тела также имеет 4 слоя,,
правда, не везде выраженные в полной мере. В грудной клетке это внутренние и наружные межреберные, прямая и поперечная грудные мышцы, в пояснично-брюшном отделе — мышцы брюшного пресса.
Локомоторная функция мускулатуры хвоста с выходом па сушу становится все меньше и у млекопитающих полностью утрачивается. Это приводит к значительному уменьшению массы мышц при сохранении высокой
степени их дифференцированности в связи с подвижностью хвоста.
Конечности наземных позвоночных произошли из кистеперого плавника, очень подвижного, с хорошо развитым скелетом и сильной мускулатурой
(латимерия). Метамерность мускулатуры конечностей, хорошо заметная у
лучеперых рыб, в филогенезе теряется очень рано, особенно с выходом на
сушу. С превращением конечности в сложный рычаг, поддерживающий и
передвигающий тело животного по суше, происходит обособление большого
числа мышц.
Для примитивных четвероногих характерно отхождение плечевой и бедренной костей в сторону и вверх от пояса. При таком расположении отделов
конечностей требуются большие затраты мышечной энергии для поддержания тела на вису. На грудной конечности наибольшая нагрузка падает на коракоидную кость, к которой в результате крепится основная масса мускулатуры плечевого и локтевого суставов.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Приспособления к быстрому бегу, манипуляции грудной конечностью и
способность отдыхать стоя, развившиеся у млекопитающих, сопровождались
поворотом конечности из сегментальной в сагиттальную плоскость раскрытием суставов и все более высоким приподниманием тела над землей. При
этом изменялись условия действия силы тяжести и работы мышц при стоянии и движении животного. У копытных приспособление конечностей к быстрому поступательному движению и к экономному расходованию мышечной энергии при стоянии привело к утере разнообразия движения. Выразилось это в еще большей редукции плечевого пояса (исчезновение ключицы) и
выпрямлении свободной конечности. Плечевой пояс утерял костную связь с
осевой частью тела и приобрел обширную площадь опоры с помощью мышц,
связавших его с головой, шеей, холкой, спиной и грудной клеткой. Так что
мускулатура конечностей по массе начала преобладать над мускулатурой туловища. Мышцы поясов и проксимальных отделов конечностей в значительной мере покрывают сверху туловищную мускулатуру и частично ее вытесняют. Развитие мышц дистальных звеньев во многом определяется особенностями механики движения и экологии животного (хождение, ползание, прыгание, рытье и т. д.). У копытных в связи с редукцией пальцев, выпрямлением суставов произошло уменьшение количества к усложнение строения
мышц дистальных звеньев конечностей.
И наконец, самым поверхностным и наименее расчлененным мышечным
пластом является подкожная мускулатура — впервые возникшая у рептилий
часть соматической мускулатуры. У млекопитающих она сильно развита,
особенно у животных, которые могут свертываться (еж, броненосец). Из домашних животных она хорошо развита у лошади и имеет вид широких пластов, лежащих под кожей в области шеи, холки, лопатки, груди и брюха (см.
рис. 72). На голове подкожная мускулатура вступает в тесную связь с висцеральной и входит составной частью в мышцы лица, век, носа, ушной раковины.
Сложные преобразования в мускулатуре головы происходят параллельно со сложными филогенетическими преобразованиями черепа. В результате
соматическая мускулатура в области головы в значительной степени вытесняется висцеральной мускулатурой, окружающей головную кишку. Соматическая мускулатура головы уже у рыб, представлена лишь мышцами глаза и
некоторыми над- и поджаберными мышцами с продольным направлением
мышечных волокон (участвуют в дыхательных движениях жаберного аппарата).
Висцеральная мускулатура, окружающая головной конец кишечной
трубки, подверглась значительной дифференцировке, приобрела свойства
поперечнополосатой мышечной ткани, но сохранила свое кольцевое направление волокон. Ею образованы кольцевые мышечные пласты челюстной,
подъязычной и жаберных дуг, на базе которых развивается основная масса
мышц головы: челюстные, подъязычные, жаберные, некоторые мышцы плечевого пояса с хватательной, жевательной и другими функциями.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У млекопитающих соматическая мускулатура головы представлена
мышцами глаза, среднего уха, языка и некоторыми мышцами подъязычной
кости. Висцеральная мускулатура образует мимические (лицевые) и жевательные (челюстные) мышцы.
И наконец, только у млекопитающих имеется мышечная грудобрюшная
преграда — диафрагма.
Онтогенетическое развитие. Соматическая мускулатура в основном происходит из миотомов сомитов мезодермы (рис. 74). В области головы из трех
предушных миотомов образуются мышцы глазного яблока. Передние заушные миотомы исчезают, а из задних (затылочных) развивается подъязычная
мускулатура. Висцеральная мускулатура головы мезенхимного происхождения. Шейные, грудные, поясничные, крестцовые и хвостовые миотомы закладываются в соответствии с количеством метамерных сегментов тела. Они
разрастаются в дорсальном и вентральном направлениях и дают начало всей
соматической мускулатуре шеи, туловища и хвоста. Мышцы конечностей
образуются путем выростов вентральных участков миотомов, к которым
присоединяется клеточный материал, выселяющийся из париетального листка спланхнотома мезодермы. Закладка мускулатуры несколько отстает от закладки скелета и в определенной мере зависит от него.
Рис. 74. Метамерная закладка
мускулатуры у зародыша млекопитающего миотомы:
1— затылочные. 2 — шейные, 3 —
грудные. 4 — поясничные, 5 — крестцовые, 6 — хвостовые.
В зародышевый период, с 20—22-го дня развития, у крупного рогатого
скота в миотомах происходит размножение миобластов. В предплодный период начинается анатомическая дифференцировка: обособляются мышцы и
группы мышц. Параллельно с этим, но значительно дольше идет гистогенез
мышечной ткани. Миобласты сливаются в мышечные трубочки, в них появляются миофибриллы. Анатомическая дифференцировка в основном заканчивается в предплодном периоде — к 50—55-му дню. Закладка и дифференцировка мышц идет в определенной последовательности. Раньше других закладывается осевая мускулатура. В ней дифференцировка идет от головного
конца к хвостовому. При этом глубокие мышцы дифференцируются раньше
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
поверхностных. В процессе дифференцировки мышц в них врастают соответствующие черепно- или спинномозговые нервы. Эта связь устанавливается очень рано и сохраняется в течение всей жизни. Закладки конечностей появляются в виде валикообразных утолщений около вентральных участков от
5-го шейного до 1-го грудного миотома — зачаток грудной конечности и от
1-го поясничного до 3-го крестцового миотома — зачаток тазовой конечности. Вскоре валики стягиваются и принимают вид сплющенных конических
выростов — почек. Образование мышц на грудной конечности у зародыша
теленка начинается с 32-го дня, а на задней — с 34-го дня эмбрионального
развития. Раньше других закладываются мышцы поясов, затем свободной
конечности, где процесс распространяется от проксимальных звеньев к дистальным. Как и в осевой части тела, дифференцировка глубоких мышц идет
раньше, поверхностных — позже. На латеральной стороне конечности закладываются экстензоры, абдукторы и супинаторы, на медиальной — флексоры,
аддукторы и пронаторы. Брюшки мышц закладываются раньше сухожилий.
К концу предплодного периода мышцы конечностей оказываются анатомически сформированы, гистологически же незрелы — состоят из мышечных
трубочек, лежащих пучками. На протяжении плодного периода продолжается
гистологическая дифференцировка мышц: увеличиваются число и размеры
мышечных трубочек, происходит превращение трубочек в мышечные волокна, нарастает в них количество миофибрилл; оформляется эндомизий и перимизий мышц, развиваются капиллярные сети, оформляются пучки I, II и III
порядков.
В результате анатомо-гистологической дифференцировки из дорсальных
участков миотомов образуется дорсальная мускулатура позвоночного столба,
лежащая над телами позвонков. Она иннервируется дорсальными ветвями
спинномозговых нервов. Из вентральных участков миотомов образуется вентральная мускулатура позвоночного столба, лежащая под телами позвонков,
мускулатура грудной клетки, брюшной стенки и диафрагма. Из мышечных
почек развивается вся мускулатура конечностей.
В процессе органогенеза происходит обособление мышц по длине, толщине, дробление или слияние, образование комплексных и многораздельных
мышц, формирование их перистой структуры. В ранний плодный период быстрее растут мышцы туловища, а в поздний — мышцы конечностей, особенно их самых дистальных звеньев — лапы.
У копытных животных к рождению полностью сформирован аппарат
движения, который тотчас же начинает функционировать: через несколько
часов новорожденный теленок, ягненок, жеребенок, поросенок может следовать за матерью. Однако это не значит, что в локомоторном аппарате завершены процессы роста и дифференцировки. Они продолжаются до возраста
морфофизиологической зрелости, а адаптивная перестройка аппарата движения происходит в течение всей жизни.
Постнатальный рост мускулатуры. После рождения продолжается интенсивный рост мускулатуры, которая по скорости роста обгоняет скелет.
Особенно усиленно этот процесс протекает в первые два месяца после рожwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дения. Следующие пики роста у крупного рогатого скота приходятся на 6-й и
12-й месяцы жизни, у овец — на 3-й и 9-й месяцы. Осевая мускулатура растет быстрее, чем мышцы конечностей, особенно с наступлением половой
зрелости. У новорожденных телят масса осевой мускулатуры составляет
46%. а у 14-месячных — 53%. На конечностях большая скорость роста мышц
в проксимальных звеньях (по сравнению с дистальными). На грудной конечности они растут несколько интенсивнее, но быстрее завершают рост, чем
мышцы тазовой конечности. Разгибатели растут быстрее сгибателей, причем
периоды подъема скорости их роста не совпадают.
С возрастом в мышцах уменьшается количество мышечных волокон на
единицу площади и в первичных мышечных пучках, так как наряду с утолщением мышечных волокон (примерно в 15—20 раз) происходит прорастание мышц соединительной тканью, она становится плотнее, мышечные пучки. I порядка включают меньше волокон. Тем не менее относительное количество соединительной ткани в мышцах с возрастом снижается, а мышечной
— увеличивается. Так, за 18 месяцев у бычков количество соединительной
ткани увеличивается в 8 раз, а мышечной — в 17 раз. Меняется и химический
состав: увеличивается количество протеина и жира, меньше становится воды.
Для каждого типа мышц характерна своя динамика химических показателей.
Не только группы мышц, но и каждая мышца обладает собственным характером роста, что связано как с особенностями ее внутренней структуры
так и функционирования. Самые высокие темпы роста у мышц динамического типа. Неравномерность роста мышц во многом определяет изменение пропорций и статей тела.
Влияние внутренних и внешних факторов на рост мышц. Образ
жизни животного, способ добычи и характер корма накладывают отпечаток
на рост и дифференцировку мускулатуры. Так, у свиней больше развивается
дорсальная мускулатура, особенно шеи. У лошади лучше, чем рогатого скота, развиты жевательные мышцы. Брюшные мышцы, наоборот, более развиты у рогатого скота.
На характер роста мышц накладывает отпечаток и пол животного. При
одинаковой упитанности у бычков мышцы развиты лучше и составляют в
туше больший процент, чем у телок и бычков-кастратов. К тому же рост мускулатуры у бычков продолжается дольше, следовательно, от них в конечном
итоге можно получить больше мяса. У бычков более развитой оказывается
мускулатура шеи, холки и плечевого пояса (что важно для силы животного
при установлении иерархии в стаде). У телок более развита мускулатура
брюшного пресса и задней половины тела. Кастраты по характеру роста мускулатуры приближаются к телкам, а по росту длиннейших и полуостистых
мышц отстают от животных обоих полов. У бычков в мышцах меньше жировых включений, а у телок и кастратов тоньше мышечные волокна, хорошо
выражена мраморность мяса.
Наблюдаются и некоторые различия в темпах роста и развития мускулатуры между породами с различным направлением продуктивности. Для
скороспелых пород характерна высокая энергия роста, зато у позднеспелых
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дольше сохраняются высокие приросты. У помесных животных при правильном подборе родительских пар темпы роста мускулатуры выше, а волокна—
тоньше. Поэтому промышленное скрещивание считается одним из резервов
увеличения производства мяса.
Кормление оказывает, пожалуй, наибольшее влияние на рост и развитие
мускулатуры. Обильное кормление в молодом возрасте способствует не
только интенсивному росту мышц, но и большему отложению жира. Мышцы
реагируют и на недокорм. Замечено, что при голодании питательные вещества распределяются не в равной мере по всей мускулатуре, а с таким расчетом,
чтобы обеспечить развитие мышц функционально наиболее необходимых на
данном этапе онтогенеза. Так, при недокорме телят-молочников от рождения
до 5 месяцев в наибольшей степени страдала большая жевательная мышца,
масса которой оказалась на 44% ниже, чем у нормально кормившихся телят,
в то время как другие крупные мышцы мало отстали в своем развитии. При
недокорме же в возрасте от 11 до 15 месяцев, когда телят перевели на кормление более грубым кормом, эта мышца практически не снизила скорость
роста, а вот мышцы плечевого пояса отстали по массе на 23%.
Перевод на обильное питание стимулирует рост мускулатуры. Особенно
интенсивно увеличивают свою массу наиболее отставшие в росте мышцы.
Однако полной компенсации их массы и размеров мышечных волокон не
происходит. Оптимальному росту мышц в постнатальном онтогенезе способствует и двигательная активность. Есть данные, что в условиях промышленного животноводства можно избежать отрицательного влияния гиподинамии
на рост мускулатуры с помощью принудительного движения животных.
ПОДКОЖНЫЕ МЫШЦЫ
Самый поверхностный пласт соматической мускулатуры образован подкожными мышцами (см. рис. 72). Они не имеют никакой связи со скелетом,
прикрепляются к коже поверхностной фасцией, а порой залегают и между
листками фасции. Их сокращение вызывает подергивание кожи, позволяет ей
собраться в мелкие складки. У некоторых млекопитающих (еж, броненосец,
дикообраз) эти мышцы представляют собой единый мышечный пласт, лежащий под кожей и тесно с ней соединенный, что позволяет этим животным
свертываться клубком. У млекопитающих большинства видов подкожные
мышцы в той или иной степени редуцировались. Из сельскохозяйственных
животных в наибольшей степени они развиты у лошади и называются в соответствии с местом расположения: подкожная мышца туловища, шеи, лица,
лопатко-плечевая, молочной железы, бедра, препуциальная, мошонки.
МЫШЦЫ ОСЕВОЙ ЧАСТИ ТЕЛА (ГОЛОВЫ, ШЕИ, ТУЛОВИЩА И ХВОСТА)
Мышцы осевой части тела делят на несколько групп, исходя из их расположения и выполняемой функции. Это дорсальные и вентральные мышцы
позвоночного столба, мышцы грудной и брюшной стенок. Сюда же относятwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ся мышцы плечевого пояса и плеча, связывающие грудную конечность с осевой частью тела.
Мышцы головы
Мышцы головы (рис. 75, 76) делят на несколько групп: лицевые (мимические) и жевательные, короткие мышцы головы, действующие с шеи, мышцы подъязычной кости, обеспечивающие движения корня языка, глотки и
гортани.
Лицевая мускулатура образует комплексы мышц, расположенные вокруг естественных отверстий: рта, ноздрей, глаз, ушей. Вокруг каждого отверстия, сформированного кожными складками головы, мышцы составляют
двухслойный пласт. Более глубокий слой образует кольцевые мышцы, выполняющие роль сфинктеров отверстий. Поверхностный слой разделяется на
многочисленные, радиально расположенные пластинчатые мышцы, которые
одним концом вплетаются в кольцевые мышцы, а другим закрепляются на
костях черепа. Эти мышцы, сокращаясь, способствуют открыванию отверстий, то есть выполняют роль дилататоров — расширителей. Разная степень
дифференцировкн радиально расположенных мышц обусловливает неодинаковую подвижность губ, ноздрей, ушных раковин. Наиболее подвижны губы,
ноздри и уши у лошади, менее подвижны— у рогатого скота (особенно у
крупного) и у свиньи.
Круговая мышца рта — m. orbicularis oris — составляет основу губ.
Снаружи покрыта кожей, изнутри — слизистой оболочкой, с которыми плотно срастается. Мышечные пучки идут вдоль губ. Хорошо развита у лошади и
мелкого рогатого скота. У крупного рогатого скота и свиньи развита слабо.
Иннервируется лицевым нервом. Васкуляризируется лицевой артерией. В нее
вплетаются мышцы — дилататоры, другой конец которых заканчивается на
костях черепа: носо-губной подниматель, опускатель нижней губы, подниматель верхней губы, клыковая, опускающая угол рта и др. Названия этих
мышц говорят об их функции. Из сельскохозяйственных животных лучше
развиты у лошади, козы, овцы. Функция — сжимает ротовое отверстие.
Щечная мышца — m. buccinator — пластинчатая, составляет основу
щек. Снаружи прикрыта подкожными и большой жевательной мышцами. Изнутри выстлана слизистой оболочкой. Состоит из двухперистых пластов. Косо идущие мышечные пучки закрепляются на верхнечелюстной и нижнечелюстной костях на уровне коренных зубов. Рострально сплетаются в круговую мышцу рта. У травоядных развита лучше, чем у всеядных. Иннервируется лицевым нервом. Васкуляризируется лицевой артерией. Функция — способствует перемещению корма при жевании; образует мягкую
стенку ротовой полости.
Жевательная мускулатура немногочисленна, но мощно развита. Ее составляют всего 4 мышцы, основной функцией которых является смыкание
(большая жевательная, крыловидная, височная) и размыкание (двубрюшная)
челюсти, а также некоторое отведение, приведение и выдвигание вперед
нижней челюсти, особенно развитое у травоядных. Все жевательные мышцы
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(кроме двубрюшной) имеют сложноперистое строение, обладают большой
силой и относятся по внутренней структуре к статодинамическому типу.
Рис. 75. Мышцы головы:
А — с латеральной стороны у
коровы. Б — с медиальной
стороны у лошади; 1— носогубный подниматель; 2 —
круговая м. глаза; 3— лобная
м.; 4 — ушные мм.; 5 — круговая м. рта; 6 — клыковая
м.; 7 — щечная м.; 8 — скуловая м.; 9 — большая жевательная м.; 10 — крыловидная
м.; 11—двубрюшная м.; 12 —
височная м.
Большая жевательная мышца — m. masseter — мощная пластинчатая
мышца, лежит на латеральной поверхности ветви нижней челюсти, прикрыта
фасцией и подкожной мышцей. Состоит из двух не полностью разделенных
частей.
Поверхностная часть начинается от лицевого бугра (лицевого гребня —
у лошади), пучки ее идут каудовентрально. Глубокая часть берет начало на
скуловой дуге. Ее пучки идут дорсовентрально. Обе части, срастаясь, оканчиваются в ямке большой жевательной мышцы. Иннервация — ветвями
тройничного нерва. Васкуляризация — ветвями наружной челюстной артерии. Ее двухслойное строение, особенно выраженное у травоядных, при попеременном сокращении поверхностной и глубокой частей позволяет совершать боковые, жерновообразные движения нижней челюстью при перетираwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
нии корма. Наиболее сильно развита данная мышца у лошади (перетирание
необработанного корма). Масса ее составляет 52% от всех жевательных
мышц, у крупного рогатого скота — 50%, у свиньи — 47%.
Функция — поднимание нижней челюсти (смыкание челюстей).
Рис. 76. Поверхностная
мускулатура:
А — крупного рогатого скота; Б — лошади; В — свиньи; 1— круговая м. рта;
2— щечная м.; 3— большая
жевательная м.; 4— грудино-головная м.; 5— плечеголовная м.; 6— плечеатлантная м.; 7— пластыревидная; 8 — ромбовидная; 9
— шейная и 10— грудная
части трапециевидной м.; 11
— широчайшая м. спины;
12 — шейная и 13 — грудная части зубчатой вентральной м.; 14 — плечевая
и 15 — предплечная части
поверхностной грудной м.;
16 — предлопаточная и 17
— плечевая части глубокой
грудной м.; 18 — межреберные наружные мм.; 19 —
косая наружная м. живота;
20 — предостная м.; 21 —
дельтовидная м.; 22 — трехглавая м. плеча; 23 — напрягатель фасции предплечья; 24 — лучевой разгибатель запястья; 25 — разгибатели пальцев; 26 — напрягатель широкой фасции
бедра; 27 — средняя ягодичная м.; 28—поверхностная ягодичная м.; 29 —
двуглавая м. бедра; 30 —
полусухожильная м.; 31 —
полуперепончатая м.; 32 —
стройная м.; 33 — икроножная м.; 34 — мм. сгибатели
пальцев.
Крыловидная мышца — m. pterygoideus — пластинчатая, лежит на медиальной ветви нижней челюсти. Начинается от крыловидной, нёбной костей
и крыловидного отростка клиновидной кости. Состоит из двух слоев, мыwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
шечные пучки которых идут под углом друг к другу. Оканчивается, веерообразно расширяясь по всей ямке крыловидной мышцы нижнечелюстной кости.
Иннервируется ветвями тройничного нерва. Васкуляризируется челюстными
артериями. Функция — синергист большой жевательной мышцы. Боковые
перетирающие движения усиливаются при одновременной работе левого
massetera и правой крыловидной и наоборот.
Височная мышца — m. temporalis — мощная, лежит в височной ямке,
заполняя ее. Начинается от височного гребня. Мышечные пучки, направляясь
с разных сторон, оканчиваются на венечном (мышечном) отростке нижнечелюстной кости. Особенно развита у хищных для которых характерно дробящее движение нижней челюсти (снизу вверх). Из сельскохозяйственных копытных лучше развита у свиньи.
Двубрюшная мышца — m. digastricus — тонкая веретеновидная мышца,
лежит между затылочной и нижнечелюстной костями, на медиальной поверхности крыловидной мышцы. Начинается от яремного отростка, идет ростровентрально и закрепляется на вентральном крае тела нижнечелюстной
кости. У крупного рогатого скота и лошади в середине разделяется сухожильным тяжом, образуя 2 брюшка, следующих друг за другом. У свиньи
одно брюшко, так как сухожильного тяжа нет. У лошади мышца имеет дополнительное брюшко, оканчивающееся на углу нижней челюсти. Иннервируется ветвями лицевого и тройничного нерва. Васкуляризируется ветвями
челюстных артерий. Функция — опускание нижней челюсти.
Дорсальные мышцы позвоночного столба
Дорсальные мышцы позвоночного столба (рис. 77) лежат над телами позвонков и расчленяются на два тяжа: латеральный и медиальный, каждый из
которых состоит из двух слоев: поверхностного и глубокого. Глубокий слой
медиального мышечного тяжа образует комплекс остистых мышц с продольным расположением мышечных пучков, прикрепляющихся к остистым отросткам позвонков. На нем сверху лежит более поверхностный слой медиального мышечного тяжа, образованный комплексом полуостистых мышц с краниодорсальным направлением мышечных пучков (от поперечнореберных
или поперечных отростков позади лежащих позвонков к остистым отросткам
впереди лежащих позвонков). К нему сверху прилежит глубокий слой латерального мышечного тяжа, образованного комплексом длиннейших мышц с
краниовентральным направлением мышечных пучков (от остистых отростков
позади лежащих позвонков к поперечным отросткам впереди лежащих позвонков или вертебральным концам ребер). И самый поверхностный слой латерального тяжа образован комплексом подвздошнореберных мышц с продольным расположением мышечных пучков (вдоль поперечнореберных отростков позвонков и углов ребер). Каждый слой, в свою очередь, формирует
ряд длинных и коротких мышц.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 77. Дорсальная мускулатура позвоночного столба:
1 — длиннейшая м. спины; 2 —длиннейшая м. шеи; 3 —длиннейшая м. головы
и атланта; 4 — остистая и полуостистая м. спины и шеи; 5 — полуостистая м. головы; 6 — подвздошнореберная м.; 7 — длинная м. головы; 8 — лестничная м.; 9 — поверхностные межреберные им.
За начало дорсальных мышц принята наименее подвижная область —
крестец. Отсюда прослеживается расположение мышц в краниальную сторону — к голове и в каудальную — к хвосту. Все дорсальные мышцы по форме
комплексные, что во много раз увеличивает их силу благодаря множественному прикреплению к костям.
Дорсальные мышцы позвоночного столба разгибают позвоночник или
его отдельные участки (шею, поясницу, хвост), осуществляют, насколько это
возможно, вращательные движения позвоночника. При одностороннем действии изгибают позвоночник или его участки вбок. При совместном действии
с вентральными мышцами укрепляют позвоночник. При движении, действуя
совместно с мышцами тазовой конечности, способствуют перенесению центра тяжести и попеременному облегчению движения то грудной, то тазовой
конечности.
Длиннейшая мышца — m. longissimus — длинная толстая мышца протяженностью от крестца до головы. Подразделяется на длиннейшие мышцы
поясницы и груди (спины), шеи, головы и атланта.
Длиннейшая мышца поясницы и груди (длиннейшая мышца спины)
—m. longissimus lumborum et thoracis — наиболее мощная мышца спины. Начинается от гребня подвздошной кости и от остистых отростков крестцовых,
поясничных и последних грудных позвонков. Основной массой лежит в поясничном отделе, где выполняет треугольное пространство между поперечнореберными и остистыми отростками позвонков. По направлению к шее
становится тоньше и все более вытесняется остистой и полуостистой мышцами. По ходу закрепляется отдельными пучками на суставных, поперечнореберных и поперечных отростках позвонков. Оканчивается у рогатого скота
и лошади на 7-м, у свиньи на 5-м шейном позвонке. Направление мышечных
пучков краниовентральное.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Длиннейшая мышца шеи — m. longissimus cervicis — лежит на полуостистой мышце. Прикрыта длиннейшей мышцей груди (спины) и шейной
частью зубчатой вентральной мышцы. Начинается от первых 5—7 грудных
позвонков, по ходу закрепляется на поперечных отростках впереди лежащих
позвонков. Оканчивается отдельными зубцами на поперечнореберных отростках от 4-го до 7-го шейных позвонков.
Длиннейшая мышца головы и атланта — m. longissimus capitis et atlantis — лежит под длиннейшей мышцей шеи и пластыревидной мышцей.
Начинается от первых грудных и последних шейных позвонков, по ходу закрепляется на поперечнореберных отростках впереди лежащих позвонков.
Оканчивается на каменистой части височной кости и крыле атланта, сливаясь
с сухожилием пластыревидной мышцы.
Комплекс длиннейших мышц в основной своей массе относится к динамостатическому типу. Его краниальные порции — длиннейшая головы и атланта у свиньи и крупного рогатого скота динамического типа. Иннервируется дорсальными ветвями поясничных, грудных и шейных спинномозговых
нервов. Васкуляризируется глубокой шейной, позвоночной, межреберными,
поясничными, краниальной ягодичной артериями. Функция — основной разгибатель позвоночного столба, выполняет все функции, присущие дорсальной мускулатуре позвоночного столба.
Пластыревидная мышца — m. splenius (см. рис. 76—7) — пластинчатая, в виде широкого треугольника лежит на шее на полуостистой
мышце головы под трапециевидной и ромбовидной мышцами. По направлению мышечных волокон ее следует отнести к комплексу длиннейших мышц.
Начинается на холке от канатиковой части выйной связки, остистых отростков 3—5-го и поперечных отростков 5—7-го грудных позвонков. Оканчивается у рогатого скота на затылочной кости и крыле атланта, у свиньи, кроме
того, на каменистой части височной кости; у лошади— на поперечнореберных отростках 3—5-го шейных позвонков и крыле атланта. У свиньи и крупного рогатого скота относится к динамическому, у овцы и лошади — к динамостатическому типу. Иннервация шейными и грудными спинномозговыми
нервами. Васкуляризация глубокой шейной и позвоночной артериями. Функция — разгибание, а также поворачивание в сторону шеи и головы.
Остистая и полуостистая мышцы груди (спины) и шеи — mm. spinalis et semispinalis thoracis (dorsi) et cervicis — длинный пласт не полностью
разделенных мышц (см. рис. 77—4). Лежит на остистых отростках под и выше длиннейшей мышцы. Обособляется от длиннейшей мышцы поясницы и
груди у крупного рогатого скота на уровне первого поясничного — последних грудных позвонков, у овцы — на уровне 8—9-го грудного позвонков.
Мышечные пучки идут продольно и краниодорсально. У лошади и свиньи на
уровне 12—13-го грудного позвонка дифференцируется только остистая
мышца груди (спины) и шеи с продольным направлением мышечных пучков.
По ходу мышечные пучки закрепляются на впереди лежащих позвонках через несколько сегментов. Оканчивается на 4—5 последних шейных позвонках. У крупного рогатого скота и свиньи эта мышца относится к диwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
намостатическому, а у овцы и лошади — к полустатодикамическому типу.
Иннервация — шейными и грудными спинномозговыми нервами. Васкуляризация — позвоночной, поперечной шейной и межреберными артериями.
Функция — как у остальных дорсальных мышц позвоночного столба.
Полуостистая мышца головы — m. semispinalis capitis (5) — пластинчатая, лежит в виде широкого пласта по дорсальной части шеи, прикрыта
пластыревидной мышцей. Мышечные пучки идут краниодорсально, начинаясь у рогатого скота от поперечных отростков первых десяти грудных и последних шейных позвонков. Закрепляются по ходу на остистых отростках
позвонков через несколько сегментов. У лошади эта мышца короче: начинается от первых 6—8 грудных позвонков, у свиньи состоит из двух частей и
имеет несколько сухожильных прослоек. Оканчивается на чешуе затылочной
кости. У свиньи и крупного рогатого скота относится к динамостатическому
типу, у овцы и лошади— к полустатодинамическому. Иннервация — шейными спинномозговыми нервами. Васкуляризация — глубокой шейной артерией. Функция — разгибание, а также отведение в сторону шеи и головы.
Подвздошнореберная мышца —гл. iliocostalis (6)—длинная узкая комплексная мышца, лежит латеральнее длиннейшей мышцы, частично прикрыта зубчатыми дорсальными и широчайшей мышцами. В ней различают поясничную, грудную и шейную части, следующие друг за другом. Начинается от
маклока подвздошной кости и отдельными зубцами от поперечнореберных
отростков поясничных позвонков. Вся мышца состоит из большого числа ясно различимых длинных пучков, идущих продольно или с небольшим краниовентральным уклоном. По ходу пучки закрепляются на углах ребер.
Оканчивается на поперечнореберных отростках последних шейных позвонков. Относится к статодинамическому типу. Иннервируется шейными, грудными, поясничными спинномозговыми нервами. Васкуляризация — межреберными и поясничными артериями. Функция — как у других дорсальных
мышц позвоночного столба.
Короткие дорсальные мышцы позвоночного столба. Самое медиальное положение среди дорсальных мышц занимают короткие сегментированные мышцы. Они соединяют между собой одноименные и разноименные отростки близлежащих позвонков на протяжении всего позвоночного столба:
межостистые, межпоперечные, поперечноостистые и др. Количество их возрастает в краниальном направлении. Особенно дифференцированы короткие
мышцы, соединяющие голову с атлантом и осевым позвонком, а также вентральные, дорсальные, боковые, прямые и косые мышцы головы. Эти мышцы, как правило, относятся к динамическому типу. Иннервируются дорсальными ветвями шейных, грудных, поясничных и хвостовых нервов. Васкуляризиру-ются позвоночной, ветвями межреберных и поясничных артерий.
Принимают участие в выполнении функций, присущих дорсальной мускулатуре.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Вентральные мышцы позвоночного столба
Данная мускулатура лежит под телами позвонков (рис. 78). Сгибает позвоночник или его отдельные части. При совместном действии с дорсальными мышцами способствуют повороту вбок или вращению головы, шеи, хвоста, а также фиксации позвоночника (при двустороннем совместном действии). Вентральная мускулатура развита в меньшей степени, чем дорсальная,
и расположена в основном в области шеи и поясницы.
Длинная мышца шеи — m. longus colli — многораздельная мышца,
лежит по вентральной поверхности тел всех шейных и первых грудных позвонков. По срединной линии соседствует с соименной мышцей другой стороны. В шейной и грудной частях мышечные пучки идут в разных направлениях. Начинается от тел грудных позвонков — с 5—6-го и направляется краниально. Мышечные пучки лежат косо и и прикрепляются через
2—3 сегмента к поперечнореберным отросткам впереди лежащих позвонков.
Оканчивается мышца на вентральном бугре атланта. Мышца имеет сложную
структуру и относится к полустатодинамическому (свинья, крупный рогатый
скот) или статодинамическому (овца) типу. Иннервируется шейными и грудными спинномозговыми нервами. Васкуляризируется позвоночной и межреберной артериями. Функция — сгибает шею.
Рис. 78. Вентральная мускулатура позвоночного столба в области поясницы:
1— внутренние межреберные мм.; 2 —
малая и 3 — большая поясничные мм.; 4
— квадратная поясничная м.; 5 — подвздошная и.
Длинная мышца головы — m. 1оngus capitis — комплексная мышца,
лежит на вентральной поверхности тел средних шейных позвонков, сбоку от
длинной мышцы шеи. Начинается от поперечнореберных отростков средних
шейных позвонков. Оканчивается на мышечных бугорках затылочной кости.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Иннервируется шейными спинномозговыми нервами. Васкуляризируется позвоночной и общей сонной артериями. Функция — сгибает голову и шею.
Квадратная мышца поясницы — m. quadratus lumborum (см. рис. 78—4) —
лежит отдельными мышечными зубцами на вентральной поверхности поперечнореберных отростков поясничных позвонков. Прикрыта большой поясничной мышцей. Начинается от вертебральных концов двух последних ребер
и от поперечнореберных отростков поясничных позвонков. Пучки направляются назад, по ходу закрепляясь на последующих позвонках. Оканчивается
мышца на вентральной поверхности крыльев крестцовой кости. Она обильно
пронизана сухожильными прослойками и относится к статодинамическому
типу. Иннервируется поясничными спинномозговыми нервами. Васкуляризируется поясничными артериями. Функция — сгибание поясницы.
Малая поясничная мышца — m. psoas minor (2) —небольшая мышца,
лежит с вентральной стороны поясницы, медиально от большой поясничной
мышцы. Начинается от тел последних грудных и первых поясничных позвонков. Оканчивается на поясничном бугорке подвздошной кости. Относится к полустатодинамическому типу. Иннервируется поясничными спинномозговыми нервами. Васкуляризируется поясничными артериями. Функция— подтягивает таз и сгибает поясницу.
Большая поясничная мышца (3), см. мм. тазовой конечности.
Короткие вентральные мышцы позвоночного столба — расположены в области соединения головы с шеей и на хвосте. Участвуют в опускании и вращении головы и хвоста.
Мышцы грудной клетки
В мускулатуре грудной клетки можно различить четыре мышечных слоя
с различным направлением мышечных пучков, хотя и развитых неповсеместно (см. рис. 76, 77 и 79). Мышцы с каудовентральным и продольным направлением пучков являются вдыхателями — инспираторами, так как при их сокращении ребра поворачиваются таким образом, что грудная клетка расширяется. Диафрагма считается инспиратором, увеличивающим грудную клетку
в длину. Мышцы с краниовентральным и поперечным направлением пучков
— выдыхатели — экспираторы. При их сокращении ребра поворачиваются
таким образом, что грудная клетка сужается. При поверхностном дыхании
действуют короткие мышцы: межреберные, подниматели ребер. При глубоком дыхании включаются и длинные мышцы грудной стенки: дорсальные
зубчатые, поясничнореберная, лестничная, прямая грудная. Если в гкте дыхания активное участие принимают мышцы брюшного пресса, дыхание называется брюшным.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 79. Глубокая мускулатура туловища:
1 — пластырсвидная м.; 2 —зубчатый дорсальный вдыхатель; 3 — подвздошнореберная м.; 4 — зубчатый дорсальный выдыхатель; 5 — поясничнореберная м.; 6 — подвздошная м.: 7 —лестничная м.; 8 — прямая м. груди;
9 — межреберные наружные мм.; 10 — межреберные внутренние мм.; 11 —
прямая м. живота; 12 — внутренняя косая м. живота.
Мышцы-вдыхатели — инспираторы (см. рис. 79). Краниальная дорсальная зубчатая мышца (зубчатый дорсальный вдыхатель)—m. serratus
dorsalis cranialis (2)—комплексная, пластинчатая. Лежит на дорсальной мускулатуре и вертебральных концах ребер, прикрыта широчайшей мышцей
спины. Начинается широким пластинчатым сухожилием от остистых отростков грудных позвонков области холки. Переходит в плоское мышечное
брюшко, составленное из 4—6 зубцов у крупного рогатого скота, 3 зубцов у
овцы, 4—5 зубцов у свиньи, 7—8 зубцов у лошади. Направление мышечных
пучков каудовентральное. Оканчивается на краниальных краях с 4—5-го по
8—9-е ребро у крупного рогатого скота, с 4-го по 6-е ребро у овцы, с 4—5-го
по 8-е ребро у свиньи, с 5—6-го по 11—12-е ребро у лошади. Относится к
динамическому типу. Иннервируется межреберными нервами Васкуляризируется межреберными артериями.
Подниматели ребер — mm. levatores costarum — короткие мышцы треугольной формы, прикрыты длиннейшей мышцей спины и подвздошнореберной мышцей. Начинаются от поперечных отростков позвонков. Оканчиваются на вертебральных концах позади лежащих ребер. Направление мышечных пучков каудовентральное. Относятся к динамическому типу. Иннервируются межреберными нервами. Васкуляризируются межреберными артериями.
Межреберные наружные мышцы — mm. intercostales externi (9) — являются прямым продолжением поднимателей ребер на боковые стенки грудной клетки, образуя ее мягкие стенки. Лежат на межреберных внутренних
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мышцах, прикрыты зубчатой вентральной и широчайшей мышцей спины.
Расположены между соседними ребрами так, что мышечные пучки идут каудовентрально от каудального края предыдущего до краниального края последующего ребра. Относятся к динамическому типу. Иннервируются межреберными нервами. Васкуляризируются межреберными артериями.
Лестничная мышца — m. scalenus (7) —пластинчатая, в виде 2—3
сравнительно узких лент, лежащих ступенчато. Является продолжением
межреберных наружных мышц на области шеи. Начинается от поперечнореберных отростков 4—5 последних шейных позвонков. Мышечные пучки
идут каудовентрально и закрепляются у рогатого скота и свиньи двумя порциями: на первом и 2—4 ребрах. У лошади есть только часть мышцы, которая заканчивается на первом ребре. Относится к динамическому типу.
Иннервируется шейными и межреберными нервами. Васкуляризируется
общей сонной артерией. Функция — кроме инспирации при двустороннем
сокращении опускает шею, при одностороннем — сгибает ее вбок.
Прямая мышца груди — m. rectus thoracis (8)—пластинчатая, в виде
небольшой ленты. Лежит на межреберных мышцах ниже зубчатой вентральной, прикрыта глубокой грудной мышцей. Начинается на стернальном конце
первого ребра, мышечные пучки направлены каудально. По ходу отдельными
порциями оканчивается на 2—4 реберных хрящах и переходит в сухожилие
прямой брюшной мышцы. Относится к динамическому типу. Иннервируется
межреберными нервами. Васкуляризируется наружной грудной артерией.
Диафрагма (грудобрюшная преграда)—diaphragma — пластинчатая
мышца куполообразной формы. Лежит поперек полости тела, закрывая собой
широкий выход из грудной клетки (каудальное грудное отверстие), и разделяет грудную и брюшную полости. Вершиной выпуклого сухожильного центра направлена в грудную полость. Периферическая часть диафрагмы состоит из мышечных пучков, направленных к центру, и в зависимости от места прикрепления делится на поясничную, реберную и грудинную части. Поясничная часть начинается под телами поясничных и последних грудных позвонков, образуя ножки диафрагмы. Правая ножка длиннее левой. Реберная
часть начинается от медиальной поверхности ребер, а грудинная часть — от
мечевидного отростка грудины. Все три части сходятся в сухожильном центре. В диафрагме имеется три отверстия для сосудов и пищевода: между
ножками диафрагмы аортальное отверстие, ниже его, на границе с сухожильным центром — пищеводное отверстие, в сухожильном центре — отверстие каудальной полой вены. По внутренней структуре диафрагма относится к статодинамическому типу. Иннервируется диафрагмальным нервом.
Васкуляризируется диафрагмальными и межреберными артериями. Функция
— является инспиратором, одновременно оказывая прессорное действие на
аорту и полые вены. Действуя вместе с брюшными мышцами, способствует
дефекации, мочеиспусканию, родам. При сокращении диафрагмы грудная
полость увеличивается в длину.
Мышцы - выдыхатели — экспираторы. Каудальная дорсальная зубчатая мышца (зубчатый дорсальный выдыхатель) — m. serratus dorsalis cauwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
dalis (4)—комплексная, пластинчатая. Лежит на дорсальной мускулатуре позади краниальной дорсальной зубчатой мышцы, прикрыта широчайшей
мышцей спины. Начинается пластинчатым сухожилием от остистых отростков последних грудных и поясничных позвонков. Переходит в плоское мышечное брюшко, составленное из 3—4 зубцов у крупного рогатого скота, из 5
— у овцы, 5—6 — у свиньи, 7—8 — у лошади. Направление мышечных пучков краниовентральное. Оканчивается на каудальных краях ребер с 10—11-го
по 13-е у крупного рогатого скота, с 9—13-го ребра у овцы, с 9—10-го по
14—15-е у свиньи, с 11—12-го по 18-е ребро у лошади. Относится к динамическому типу. Иннервируется межреберными нервами. Васкуляризируется
межреберными артериями.
Мышца, оттягивающая ребро (поясничнореберная мышца),— m. retractor costae—пластинчатая, в виде небольшого треугольника лежит на поперечной брюшной мышце под косой наружной брюшной и каудальной дорсальной зубчатой мышцами. Начинается от поперечнореберных отростков
1—3 поясничных позвонков. Оканчивается на каудальном крае последнего
ребра. Направление мышечных пучков краниовентральное. Относится к динамическому типу. Иннервируется межреберными нервами. Васкуляризируется межреберными артериями.
Межреберные внутренние мышцы — mm. intercostales interni (10)—
лежат между ребрами под межреберными наружными мышцами. Расположены между соседними ребрами так, что мышечные пучки идут краниовентрально от краниального края позади лежащего ребра к каудальному краю
впереди лежащего ребра. У овцы направление пучков горизонтальное. Относится к динамическому типу. Иннервируется межреберными нервами. Васкуляризируется межреберными артериями.
Поперечная мышца груди — m. transversus thoracis — пластинчатая,
треугольной формы, лежит на дне грудной полости по грудине. Начинается
от внутренней поверхности грудины. Оканчивается на хрящевых концах истинных ребер. Направление мышечных пучков поперечное. Относится к динамическому типу. Иннервируется межреберными нервами. Васкуляризируется внутренней грудной артерией.
Мышцы брюшной стенки
Брюшная стенка в отличие от грудной в основном мягкая, образована
четырьмя пластами мышц с различным направлением мышечных волокон,
так что объем ее может значительно меняться. Прочность мышечной стенки
усилена фасциями: поперечной брюшной и поверхностной, называемой
желтой брюшной фасцией из-за содержания большого количества эластических волокон. Это дает возможность значительного растяжения брюшных
стенок без снижения их прочности и быстрого возврата в исходное состояние
благодаря тяге эластических волокон. Пластинчатые сухожилия мышц, срастаясь и накладываясь друг на друга, формируют по средней линии белую линию живота. Основная функция брюшных мышц — удержание внутренноwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
стей и оказание на них прессорного действия, отчего мускулатура живота называется брюшным прессом. Сокращение мышц брюшной стенки при расслабленной диафрагме способствует выдоху, при сокращенной диафрагме —
опорожнению внутренностей (отрыжке при жвачке, дефекации, мочеиспусканию, родам).
Наружная косая мышца живота — m. obliquus externus abdominis (см.
рис. 76—19) —пластинчатая, мощная, широкая лежит поверхностно на боковой стенке живота. Начинается от стернальных концов 4—5-го и до последних ребер, вклиниваясь между зубцами вентральной зубчатой мышцы. Мышечные пучки идут каудовентрально, в нижней части брюшной стенки переходят в пластинчатое сухожилие, которое закрепляется на маклоке, лонном
бугорке, вплетается с медиальной стороны в фасцию бедра, а по средней линии живота срастается с сухожилием одноименной мышцы другой стороны и
сухожилием внутренней косой мышцы живота, входя в состав белой линии
живота.
По внутренней структуре относится к динамическому типу. Иннервируется межреберными и поясничными нервами. Васкуляризируется межреберными, краниальной и каудальной надчревными артериями.
По краю мышцы, между ее закреплением на маклоке и лонном бугорке,
образуется щель — поверхностное паховое кольцо, отграниченное паховой
связкой. Эта щель ведет в паховый канал— длинное щелевидное пространство, косо идущее между косыми мышцами живота, пронизывающее брюшную
стенку и открывающееся в брюшную полость внутренним паховым кольцом.
Через этот канал у самцов в конце плодного периода опускаются в мошонку
семенники, а в течение всей жизни в нем залегает семенной канатик. При неправильной эксплуатации животного возможно выпадение в него внутренностей — паховая грыжа. У самок паховый канал недоразвит и в норме рано зарастает.
Внутренняя косая мышца живота — m. obliquus internus abdominis
(см. рис. 79—12) — пластинчатая, в виде широкого веера лежит под наружной косой мышцей живота. Начинается у рогатого скота и свиньи от фасции
на уровне поперечнореберных отростков поясничных позвонков и от маклока. У лошади — только от маклока. Мышечные пучки направлены краниовен-трально. Переходит в пластинчатое сухожилие, которое заканчивается на
реберной дуге с медиальной стороны и по средней линии живота. Относится
к динамическому типу. Иннервируется межреберными и поясничными нервами. Васкуляризируется поясничными, краниальной и каудальной надчревными артериями.
Прямая мышца живота — m. rectus abdominis (11) —пластинчатая, в
виде широкой ленты лежит вдоль нижней поверхности живота. Прикрыта сухожилиями обеих косых мышц живота. Начинается сухожильно на хрящах
4—9-го ребра и на вентральной поверхности грудины. Оканчивается на лонном бугорке и лонном гребне. Мышечные пучки направлены каудально
вдоль тела. С соименной мышцей соединяется по средней линии живота.
Мышца пронизана поперечными сухожильными перемычками. В результате
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
этого она приобретает большую силу и относится к статодинамическому типу. Около второй перемычки имеется щелевидное отверстие — «молочный
колодец», через которое проходит подкожная брюшная (молочная) вена. Иннервируется межреберными и поясничными нервами. Васкуляризируется
межреберными, поясничными и надчревными артериями.
Поперечная мышца живота — m. transversus abdominis — пластинчатая, самая глубокая мышца брюшного пресса, лежит на поперечной
брюшной фасции. Начинается от поперечнореберных отростков поясничных
позвонков и по краю реберной дуги с медиальной стороны. Мышечные пучки направлены вентрально, поперек брюшной стенки. Переходит в пластинчатое сухожилие, которое по средней линии живота срастается с сухожилием
соименной мышцы и входит в состав белой линии живота, образованной сухожилиями всех мышц брюшного пресса. Относится к динамическому типу.
Иннервируется межреберными и поясничными нервами. Васкуляризируется
межреберными, поясничными и надчревными артериями.
МЫШЦЫ, СОЕДИНЯЮЩИЕ ГРУДНУЮ КОНЕЧНОСТЬ с ОСЕВОЙ ЧАСТЬЮ ТЕЛА
У копытных животных плечевой пояс утратил костную связь с осевой
частью тела и присоединяется к ней синсаркозом, то есть с помощью мышц.
В связи с редукцией коракоидной кости и ключицы мышцы, ранее закреплявшиеся на них, стали присоединяться к плечевой кости или грудине.
По расположению мышцы плечевого пояса можно разделить на дорсальные и вентральные. Дорсальные мышцы: трапециевидная, ромбовидная,
грудино-плечеголовная, плечеатлантная, широчайшая, спускаются с головы,
шеи и туловища к лопатке и плечевой кости. Вентральные мышцы: поверхностная и глубокая грудные, зубчатая вентральная, поднимаются от грудины
и боковой стенки тела к плечевой кости и лопатке. Мышцы плечевого пояса
не только привязывают грудную конечность, но и участвуют в ее работе. Передвигая лопатку и плечо, способст-ствуют маятникзобразному движению
грудной конечности. Удерживают туловише между лопатками.
Мышцы, соединяющие плечевой пояс с осевой частью тела. Трапециевидная мышца — m. trapezius — пластинчатая по форме, в виде вытянутого треугольника (см. рис. 76—9, 10). Лежит поверхностно в дорсальной
части шеи и холки на ромбовидной мышце и мышцах лопатки. Состоит из
шейной и грудной частей. Шейная часть у рогатого скота и лошади начинается тонким пластинчатым сухожилием, отходящим от канатчковой части
выйной связки и растянутым от 1—2-го шейного до 3-го грудного позвонка.
У свиньи начало простирается от затылочной кости до 3-го грудного позвонка. Заканчивается вдоль краниального края всей ости лопатки. Грудная часть
начинается широко от надостистой связки на протяжении от 3-го до 10-го (у
лошади — до 11-го) грудного позвонка. Оканчивается на каудальном крае
верхней трети ости лопатки. По внутренней структуре относится к динамическому типу. Иннервируется добавочным нервом. Васкуляризируется дорсальной артерией лопатки (поперечной шейной). Функция — прикрепляет
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лопатку к туловищу, при двустороннем сокращении во время стояния несколько опускает туловище между лопатками (у лошади этого не происходит). Во время движения при сокращении шейной части лопатка выносится
вперед и вверх, а при сокращении грудной части —назад и вверх; при сокращении всей мышцы лопатка поднимается вверх, грудная конечность отрывается от земли.
Плечеатлантная мышца — m. omotransversarius (6)—пластинчатая по
форме, в виде широкой ленты лежит на боковой поверхности шеи под плечеголовной мышцей, вентральнее шейной части трапециевидной мышцы. У рогатого скота и свиньи начинается от крыла атланта, оканчивается на фасции
лопатки в области акромиона. У овцы срастается с плечеголовной мышцей. У
лошади обычно отсутствует. По внутренней структуре относится к динамическому типу. Иннервируется добавочным нервом. Васкуляризируется поперечной шейной и общей сонной артериями. Функция — синергист шейной
части трапециевидной мышцы. При стоянии поворачивает голову вбок. При
движении выносит вперед и вверх вентральный угол лопатки, способствуя
отрыву конечности от земли и вынесению вперед.
Ромбовидная мышца — m. rhomboideus (8) — пластинчатая мышца, в
виде длинного толстого ремня лежит на пластыревидной и краниальной зубчатой дорсальной мышцах, под трапециевидной мышцей. Состоит из шейной
и грудной частей с различным направлением мышечных пучков. Шейная
часть (ромбовидная мышца шеи) начинается от канатиковой части выйной
связки на протяжении от 2-го шейного до 2—3-го грудного позвонка. Оканчивается на медиальной поверхности надлопаточного хряща. У свиньи вычленяется еще головная часть, начало которой простирается до затылочной
кости. Оканчивается она вместе с шейной частью. Грудная часть (ромбовидная мышца груди) начинается от надостистой связки на протяжении от 2—3го до 7—8-го грудного позвонка у рогатого скота, от 2— 3-го до 8—9-го позвонка у лошади, от 1—2-го до 5—6-го позвонка
у свиньи. Оканчивается на медиальной поверхности надлопаточного
хряща, опускаясь несколько на дорсальный край лопатки. По внутренней
структуре относится к динамическому типу. Иннервируется дорсальными
ветвями шейных и грудных нервов. Васкуляризируется глубокой шейной и
дорсальной артерией лопатки (поперечной шейной). Функция — полный синергист трапециевидной мышцы.
Вентральная зубчатая мышца — m. serratus ventralis (12, 13) —
пластинчатой формы, в виде широкого веера с выступающими на концах
зубцами, лежит на боковой поверхности шеи и грудной клетки. Прикрыта
плечеголовной, плечеатлантной, трапециевидной и широчайшей мышцами.
Состоит из шейной и грудной частей. Шейная часть (вентральная зубчатая
мышца шеи) начинается у рогатого скота и лошади от поперечнореберных
отростков с 4-го по 7-й шейный позвонок, у свиньи — от всех шейных позвонков отдельными зубцами. Грудная часть (вентральная зубчатая мышца
груди) начинается у рогатого скота от первых 6—7, у свиньи — 8, у лошади
— 8—9 ребер зубцами. Усилена мощной эластичнофиброзной связкой, расwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тянутой между зубчатой линией лопатки и ребрами. Особенно хорошо она
выражена у лошади. Мышечные зубцы шейной и грудной частей направлены
в сторону лопатки и закрепляются на ее зубчатой поверхности. По внутренней структуре у рогатого скота и свиньи мышца динамостатического, а у лошади — статодинамического типа.
Иннервируется каудальными шейными и краниальными грудными нервами. Васкуляризируется поперечной шейной и межреберными артериями.
Функция — основной держатель туловища между лопатками. Эта функция
осуществляется с помощью эластичнофиброзной связки и не требует затраты
мышечной энергии. Сокращаясь в момент вынесения конечности вперед, она
тянет лопатку вентрокраниально, чем значительно увеличивает шаг. Служит
амортизатором при ударе конечности о землю.
Мышцы, соединяющие плечо с осевой частью тела (см. рис. 76). Грудино-плечеголовная мышца — состоит из двух не полностью разделенных
мышц: грудино-головной и плечеголовной, соединенных друг с другом краниальными участками.
Плечеголовная мышца — m. brachiocephalicus (5)— пластинчатой формы. В виде широкой ленты лежит поверхностно на боковой сюроне шеи,
прикрыта лишь поверхностной фасцией и подкожной мышцей. В ней различают несколько частей, которые начинаются у крупного рогатого скота от затылочной, височной, нижнечелюстной костей и выйной связки, у свиньи —
от затылочной и височной костей, у лошади — от затылочной, височной костей и от поперечнореберных отростков 2—5-го шейных позвонков. Оканчивается у всех на гребне плечевой кости под дельтовидной шероховатостью,
закрывая плечевой сустав с дорсальной стороны.
Грудино-головная мышца — m. sternocephalicus (4) — пластинчатой
формы. В виде узкой ленты идет по вентролатеральной стороне шеи. Вентрально обе грудино-головные мышцы соприкасаются друг с другом. Выше
грудино-головной мышцы, между нею и плечеголовной мышцей проходит
яремный желоб, в котором залегают сонная артерия и яремная вена.
Начинается мышца у крупного рогатого скота от угла нижнечелюстной
и височной костей, где переплетается с плечеголовной мышцей; у овцы и
свиньи — от височной кости; у лошади — от угла нижней челюсти, не связана с плечеголовной мышцей. Оканчивается у всех на рукоятке грудины.
Обе мышцы относятся к динамическому типу, иннервируются добавочным и подмышечным нервами, васкуляризируются позвоночной и общей
сонной артериями. Функция — при стоянии опускают голову вниз и в сторону (при одностороннем сокращении), а действуя с конечностями, способствуют опусканию нижней челюсти (открыванию рта), при движении разгибают плечевой сустав и выносят конечность вперед.
Широчайшая мышца спины — m. latissimus dorsi (11)—пластинчатой
формы. Широким треугольником лежит на дорсолатеральной поверхности
грудной стенки, прикрыта лишь подкожной мышцей. Начинается широким
пластинчатым сухожилием от надостистой связки и остистых отростков от
3—5-го грудного до последнего поясничного позвонка, а у рогатого скота и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
свиньи, кроме того, отдельными порциями от трех последних ребер. Оканчивается на круглой шероховатости плечевой кости, а у лошади — и на медиальном мышечном бугре плечевой кости. Относится к динамическому типу.
Иннервируется каудальными грудными нервами. Васкуляризируется межреберными и поясничными артериями. Функция — антагонист плечеголовной
мышцы; в момент опоры продвигает тело вперед, разгибая плечевой сустав, а
в момент висения конечности сгибает плечевой сустав и оттягивает конечность назад, слегка ее пронируя.
Поверхностная грудная мышца — m. pectoralis superficialis (14, 15) —
пластинчатой формы. Лежит с краниовентральной стороны грудной клетки
между грудными конечностями. Состоит из двух сросшихся частей: плечевой
и предплечной. Плечевая часть (нисходящая грудная мышца) начинается на
рукоятке грудины, оканчивается на гребне плечевой кости рядом с плечеголовной мышцей. Между ними с каждой стороны образуется боковая
грудная борозда, хорошо заметная у лошади и крупного рогатого скота. Медиальными краями левая и правая плечевые части соприкасаются и между
ними образуется срединная грудная борозда. Предплечная часть (поперечная
грудная мышца) начинается от передней половины грудины, оканчивается на
фасции предплечья с медиальной стороны. По внутренней «структуре относится к динамическому типу. Иннервируется краниальными грудными нервами. Васкуляризуется глубокой грудной артерией. Функция — аддуктор конечности; разгибая плечевой сустав, вместе с другими мышцами выносит
поднятую конечность вперед; при опоре оттягивает туловище назад.
Глубокая (восходящая) грудная мышца — m. pectoralis profundus (17)
— пластинчатой формы. Лежит в виде мощного треугольника на латеровентральной стороне грудной клетки каудальнее поверхностной грудной мышцы
и частично ею прикрыта. Начинается на вентральной поверхности брюшной
стенки в области мечевидного хряща, от боковой поверхности грудины и на
реберных хрящах (с 3—5-го по 8—9-й). Направляется краниально, доходит
до плечевой кости, огибает ее медиодорсально так, что прикрывает плечевой
сустав с дорсальной поверхности и оканчивается на латеральном и медиальном буграх плечевой кости. По внутренней структуре относится к динамическому типу. Иннервируется краниальными грудными нервами. Васкуляризируется глубокой грудной артерией. Функция — аддуктор конечности; в
фазу опоры разгибает плечевой сустав и продвигает туловище вперед; в фазу
висения конечности сгибает плечевой сустав.
МЫШЦЫ КОНЕЧНОСТЕЙ
Основная масса мускулатуры конечностей расположена в проксимальных звеньях. Мышцы дистальных звеньев в связи с редукцией пальцев и
включением лапы в основной поддерживающий столб сильно редуцированы,
особенно короткие. Сохранившиеся мышцы, действующие на дистальные
звенья, приобрели длинные сухожилия. В результате мускулатура конечностей напоминает треугольник с основанием на уровне поясов и вершиной,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
опущенной до земли. Такая форма конечностей зависит не только от неравномерного распределения массы мышц, но и от различий в их расположении.
В проксимальных звеньях (в плечевом, локтевом, тазобедренном, коленном
суставах) кости расположены таким образом, что мышцы действуют на них
почти под прямым углом — в условиях, наилучших для функционирования.
В дистальных звеньях мышцы лежат параллельно костям, и лишь около конечной точки прикрепления угол изменяется благодаря перебрасыванию через сесамовидные кости.
Форма суставных поверхностей и особенности связочного аппарата суставов конечностей таковы, что облегчают разгибание— сгибание и затрудняют все остальные движения. Ясно, что среди мышц наибольшее развитие
получили экстензоры и флексоры. Экстензоры располагаются снаружи угла
сустава, флексоры— внутри; абдукторы — с латеральной, аддукторы — с медиальной стороны конечности. Супинаторы и пронаторы лежат косо по отношению к оси сустава, на который действуют.
Основная сила мышц конечностей сосредоточена в проксимальных
звеньях. Здесь же преобладают мышцы пластинчатой формы, динамического
и динамостатического типов. В дистальных звеньях статичность мышц возрастает, преобладают мышцы веретенообразной формы.
Мышцы грудной конечности
Мышцы, связывающие плечевой пояс с осевой частью тела, расправлены широкими веерами и покрывают собою мускулатуру шеи, грудной стенки, частично позвоночного столба, поэтому они описаны выше.
Мышцы, лежащие в области лопатки и действующие на плечевой
сустав. Плечевой сустав по форме трущихся поверхностей многоосный, следовательно, среди мышц, действующих на этот сустав, должны быть экстензоры, флексоры, абдукторы, аддукторы, супинаторы и пронаторы (рис. 80).
ЭСТЕНЗОРЫ. Предостная мышца — m. supraspinatus (1)— толстая
пластинчатая мышца, заполняет предостную ямку, прикрыта трапециевидной
и плечеатлантной мышцами. Начинается от предостной ямки, оканчивается
на большом и малом бугорках плечевой кости, огибая плечевой сустав с краниальной стороны. У овцы относится к динамостатическому типу, у свиньи
— к полустатодинамическому, у крупного рогатого скота и лошади — к статодинамическому типу. Иннервируется подлопаточным нервом. Васкуляризация ветвями подмышечной и подлопаточной артерий.
Коракоидноплечевая мышца — m. coracobrachialis (4) — лентовидная
мышца, лежит на медиальной поверхности плеча под глубокой грудной
мышцей. Начинается от коракоидного отростка лопатки, кончается на медиальной поверхности плечевой кости около большой круглой мышцы. У
большинства сельскохозяйственных животных относится к полустатодинамическому типу, у лошади — к статодинамическому. Иннервируется мышечно-кожным нервом. Васкуляризируется ветвями плечевой артерии.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Кроме этих мышц, в разгибании плечевого сустава принимают участие
плечеголовная, грудные мышцы и двуглавая мышца плеча.
Рис. 80. Мышцы грудной конечности крупного рогатого скота (А — с
латеральной и Б — с медиальной сторон):
1 — предостная м.; 2 — заостная м.; 3 — дельтовидная м.; 4— коракоидноплечевая м.: 5 — подлопаточная м.; 6— широчайшая м. спины; 7 —большая и
8 — малая круглые ми.; 9 — напрягатель фасции предплечья; 10, 11, 12 —
трехглавая м. плеча (10 — ее длинная головка,11 — латеральная и 12 — медиальная головка); 13 — локтевая м.; 14 — двуглавая м. плеча; 15 — плечевая
м.; 16 — лучевой разгибатель запястья; 17 — специальный разгибатель III
пальца; 18 — общий пальцевый разгибатель; 19 — боковой пальцевый разгибатель; 20 — локтевой разгибатель запястья; 21 — лучевой сгибатель запястья; 22 — локтевой сгибатель запястья; 23 — длинный абдуктор I пальца; 24 — поверхностный пальцевый сгибатель; 25 — глубокий пальцевый
сгибатель; 26 — межкостная средняя м.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ФЛЕКСОРЫ. Дельтовидная мышца — т. deltoideus (3) — пластинчатая, треугольной формы, лежит поверхностно на заостной мышце. У рогатого
скота и свиньи имеет две части: лопаточную и акромиальную. Лопаточная
часть начинается пластинчатым сухожилием от ости лопатки и от заостной
мышцы, акромиальная часть — от акромиона. Обе части оканчиваются на
дельтовидной шероховатости плечевой кости. У лошади есть только лопаточная часть. Мышца динамостатического типа, с возрастом становится статичнее. У лошади — статодинамическая. Иннервируется подмышечным нервом. Васкуляризируется подлопаточной артерией.
Малая круглая мышца — m. teres minor (8) — небольшая, веретеновидная, лежит каудальнее заостной мышцы под дельтовидной мышцей. Начинается от нижней трети каудального края лопатки, оканчивается на
круглой шероховатости плечевой кости. По внутренней структуре сильно
различается: от динамического типа у овцы до статодинамического — у лошади. Иннервируется подмышечным нервом. Васкуляризируется ветвями
подлопаточной артерии.
Большая круглая мышца — m. teres major (7)—пластинчатая, в форме
ленты, лежит позади лопатки на медиальной поверхности трехглавой мышцы
плеча. Начинается от каудального угла лопатки, оканчивается на круглой
шероховатости плечевой кости. Относится к динамическому типу. Иннервируется подлопаточным нервом. Васкуляризируется подлопаточной артерией.
Перечисленные мышцы являются флексорами только при их совместном
(групповом) действии. Кроме того, флексорами плечевого сустава являются
широчайшая мышца спины и длинная головка трехглавой мышцы плеча.
АБДУКТОР. Заостная мышца — m. infraspinatus (2)—толстая, лежит в
заостной ямке под дельтовидной мышцей, с которой срастается. Начинается
от заостной ямки, оканчивается на большом (латеральном) бугорке плечевой
кости. Относится к полустатодинамическому типу, а у лошади — к статодинамическому и даже статическому типу, превратившись в мышцу-связку.
Иннервируется подлопаточным нервом. Васкуляризируется подлопаточной
артерией.
АДДУКТОРЫ. Подлопаточная мышца — m. subscapularis (5) — многоперистая, лежит в подлопаточной ямке, на которой и закрепляется. Оканчивается на малом бугорке плечевой кости. У свиней и овец она полустатодинамического, у крупного рогатого скота статодинамического, а у лошади
— статического типа, то есть является мышцей-связкой, ограничивающей
боковые движения. Иннервируется подлопаточным нервом. Васкуляризируется подлопаточной артерией.
В качестве побочного действия функцию приведения выполняет коракоидноплечевая мышца.
СУПИНАТОРЫ И ПРОНАТОРЫ в качестве самостоятельных мышц у
копытных отсутствуют. Функцию супинации наряду со своей основной
функцией выполняют дельтовидная и круглая малая мышцы. Функцию пронации наряду со своей основной функцией выполняют большая круглая и
широчайшая мышцы спины.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Мышцы, лежащие в области плеча и действующие на локтевой сустав. По форме трущихся поверхностей локтевой сустав одноосный. В нем
возможно лишь разгибание-сгибание.
ЭКСТЕНЗОРЫ. Трехглавая мышца плеча — m. triceps brachii (10—
12)—мощная пластинчатая, самая крупная мышца свободной грудной конечности. Заполняет треугольное пространство между лопаткой и плечевой костью. Начинается тремя головками: длинной (массивной) — от каудального
края лопатки, латеральной — под шейкой плечевой кости с латеральной стороны и медиальной — от середины медиальной поверхности плечевой кости.
Срастаясь, все головки заканчиваются на локтевом бугре локтевой кости.
Длинная головка относится к динамостатическому типу, а латеральная и медиальная— к динамическому. Иннервируется мышца лучевым нервом. Васкуляризируется подлопаточной артерией.
Локтевая мышца — m. anconeus (13) —маленькая, лежит под длинной
головкой трехглавой мышцы, с которой срастается. Начинается по краям
локтевой ямки, оканчивается на локтевом бугре локтевой кости. Относится к
динамическому типу. Иннервируется лучевым нервом. Васкуляризируется
плечевой артерией.
Напрягатель фасции предплечья — m. tensor fasciae antebrachii (9) —
тонкая лентовидная мышца, лежит вдоль каудального края длинной головки
трехглавой мышцы плеча. Начинается от каудального угла лопатки, оканчивается сухожилием на локтевом бугре локтевой кости и далее вплетается в
фасцию предплечья. Относится к динамическому типу. Иннервируется лучевым нервом. Васкуляризируется плечевой и локтевой артериями.
ФЛЕКСОРЫ. Двуглавая мышца плеча — m. biceps brachii (14) —
толстая, веретенообразная. Лежит на передней поверхности плечевой кости.
Начинается мощным сухожилием от бугра (надсуставного бугорка) лопатки,
проходит по межбугорковому желобу и оканчивается на шероховатости лучевой кости. Имеет перистое строение, пронизана сухожильными тяжами. У
большинства сельскохозяйственных животных она статодинамическо-го, а у
свиньи — полустатодинамического типа. У лошади от нижней трети мышцы
отходит сухожильный тяж, который направляется дистально, срастается с
конечным сухожилием лучевого разгибателя запястья. В результате образуется единый сухожильный тяж, идущий от надсуставного бугорка лопатки
через плечевой и локтевой суставы к проксимальному концу пястной кости.
Этот механизм, во-первых, связывает в единую кинематическую цепь плечевой, локтевой и запястный суставы (которые в результате могут разгибаться
и сгибаться как единое целое) и, во-вторых, позволяет животному находиться
в стоячем положении без затраты мышечной энергии. Иннервируется мышца
мышечно-кожным нервом. Васкуляризируется подмышечный и плечевой артериями.
Плечевая мышца — m. brachialis (15)—веретенообразная, лежит на
плечевой кости винтообразно. Начинается на задней поверхности плечевой
кости под шейкой. С задней поверхности переходит на латеральную, затем на
краниальную и оканчивается на шероховатости лучевой кости. Относится к
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
динамическому типу. Иннервируется мышечно-кожным нервом. Васкуляризируется плечевой и лучевой артериями.
Мышцы, лежащие в области предплечья и действующие на запястный сустав. Запястный сустав по форме трущихся поверхностей одноосный.
В нем возможно сгибание-разгибание.
ЭКСТЕНЗОРЫ. Лучевой разгибатель запястья — m. extensor carpiradialis (16)—веретенообразная мышца, лежит на предплечье дорсолатерально.
Начинается на латеральном надмыщелке плечевой кости, переходит на дорсальную поверхность предплечья и сухожильно заканчивается на шероховатости 3-й пястной кости. У лошади в его сухожилие вплетается сухожильный
тяж от двуглавой мышцы плеча. Относится к динамостатическому типу. Иннервируется лучевым нервом. Васкуляризируется лучевой и межкостной артериями.
Длинный абдуктор I пальца — m. abductor digitiprimilongus (22) —
тонкая пластинчатая мышца, лежит косо в виде узкой ленточки на дорсальной стороне запястья. Начинается на латеральной поверхности лучевой кости, переходит на дорсальную поверхность и заканчивается на проксимальном
конце 2-й пястной кости. Мышца динамостатического типа у свиньи, у остальных статодинамического.
Иннервируется лучевым нервом. Васкуляризируется лучевой и межкостной артериями.
ФЛЕКСОРЫ. Лучевой сгибатель запястья — m. flexor carpi radialis
(21) —узкая лентовидная мышца, лежит на медиальной поверхности предплечья впереди локтевого сгибателя. Начинается на медиальном надмыщелке
плечевой кости. Оканчивается у рогатого скота и свиньи на 3-й, у лошади —
на 2-й пястной кости. У свиньи динамического типа, у остальных — динамостатического. Иннервируется срединным нервом. Васкуляризируется срединной артерией.
Локтевой сгибатель запястья — m. flexor carpi ulnaris (22) — веретенообразная, лежит на медиопальмарной стороне предплечья. Начинается двумя
головками: одной от медиального над-мыщелка плечевой кости, другой — от
локтевого бугра локтевой кости. Обе головки вскоре соединяются. Оканчивается мышца на добавочной кости запястья. Относится к статодинамическому
типу. Иннервируется локтевым нервом. Васкуляризируется срединной, локтевой и межкостной артериями.
Локтевой разгибатель запястья — m. extensor carpi ulnaris (20)—
веретенообразная, лежит по заднему краю латеральной поверхности предплечья. Начинается на латеральном надмыщелке плечевой кости, оканчивается на добавочной кости запястья. Мышца статодинамического типа. Иннервируется лучевым нервом. Васкуляризируется локтевой, межкостной и
срединной артериями. У копытных мышца выполняет функцию флексора в
связи со смещением дистальной точки прикрепления на добавочную кость
запястья.
Мышцы, лежащие в области предплечья и действующие на пальцы.
Суставы пальцев одноосные, позволяющие сгибание-разгибание. Все экстенwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
зоры иннервируются лучевым нервом, кровоснабжение — от лучевой и межкостной артерий. Флексоры соответственно срединным и локтевым нервами
и срединной артерией.
ЭКСТЕНЗОРЫ. Общий пальцевый разгибатель — m. extensor digitalis
communis (18)—веретенообразная мышца, лежит на латеральной поверхности предплечья. Начинается от латерального надмыщелка плечевой кости. У
рогатого скота затем разделяется на два сухожилия. У свиньи начинается
тремя брюшками, которые затем формируют четыре сухожилия. У всех кончается на разгибательных отростках копытцевых (копытных) костей каждого
пальца. Относится к динамостатическому типу.
Боковой пальцевый разгибатель — m. extensor digitalis lateralis (19)—
веретенообразная мышца, лежит между общим пальцевым разгибателем и
локтевым разгибателем запястья. Начинается от проксимального конца лучевой и локтевой костей. Оканчивается у рогатого скота на венечной и копытцевой костях IV пальца, у свиньи — на IV и V пальцах, у лошади — на путовой кости.
Относится к полустатодинамическому типу.
Специальный разгибатель III пальца — m. extensor digiti ter-tii proprius (17) —имеется у рогатого скота в виде тонкой веретенообразной мышцы, лежащей сбоку от общего пальцевого разгибателя, частью которой ее
считают. Начинается на латеральном надмыщелке плечевой кости, оканчивается на копытцевой кости III пальца. У свиньи является одним из брюшек
общего пальцевого разгибателя. У лошади отсутствует.
ФЛЕКСОРЫ. Поверхностный пальцевый сгибатель — m. flexor digitalis superficialis (24)—веретенообразная мышца, проходит по пальмарной
стороне предплечья, прикрыта локтевым сгибателем запястья. Начинается от
медиального надмыщелка плечевой кости. У всех сельскохозяйственных животных, кроме лошади, имеет по нескольку брюшек, которые срастаются между собой, а также с глубоким пальцевым сгибателем и межкостными мышцами. В области путового сустава сухожилие разделяется по числу пальцев и
оканчивается на венечной кости каждого пальца, образуя 2 сухожильные
ножки. Между ними проходит сухожилие глубокого пальцевого сгибателя.
Относится к статодинамическому типу.
У лошади в сухожилие поверхностного пальцевого сгибателя вплетается
сухожильный тяж, идущий от дистального конца лучевой кости, в результате
чего конструкция из сухожильных тяжей, объединяющая все суставы грудной конечности (см. двуглавую мышцу и лучевой разгибатель запястья), оказывается завершенной. В результате этой конструкции и сухожильно-фиброзной связки зубчатой вентральной мышцы лошадь не затрачивает мышечной энергии при опоре на грудные конечности. Конечность удерживается на
разогнутых суставах благодаря натяжению общей сухожильной конструкции.
Глубокий пальцевый сгибатель — m. flexor digitalis profundus (25) —
лежит на пальмарной поверхности предплечья. Имеет несколько головок, которые начинаются на медиальном надмыщелке плечевой, от локтевой и лучевой костей. Все они сливаются в единое мощное сухожилие, которое у дисwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тального конца пястнопальцевого (путового) сустава делится по количеству
пальцев, проходит через сухожильное влагалище поверхностного пальцевого
сгибателя и оканчивается на сгибательном бугорке копытцевой (копытной)
кости. Разные головки имеют разную внутреннюю структуру — от динамостатического до статодинамического типов.
Короткие пальцевые мышцы сильно редуцированы. Более выражены из
них межкостные мышцы.
Межкостные мышцы — mm. interossei (26)—лежат на пальмарной поверхности пястных костей. Начинаются на пальмарной поверхности запястного сустава, по ходу отдают сухожильные связки к сухожилиям общего
пальцевого разгибателя, поверхностного и глубокого пальцевых сгибателей.
Оканчиваются на путовой кости несколькими ветвями и вплетаются в концы
сухожилий пальцевых сгибателей. Содержат мало мышечных пучков, особенно у лошади.
Относятся к статическому типу.
Мышцы тазовой конечности
На тазовую конечность приходится основная работа при движении животного. В связи с этим мышцы, действующие на тазобедренный сустав, достигли особенно сильного развития. Основание мышечного треугольника тазовой конечности на теле не такое широкое, как на грудной, но гораздо более
мощное.
Мышцы, лежащие в области крестца (крупа) и действующие на тазобедренный сустав (рис. 81). Тазобедренный сустав многоосный, поэтому
среди мышц, действующих на него, должны быть экстензоры, флексоры, абдукторы, аддукторы, супинаторы и пронаторы. Характер суставных поверхностей, особенности связочного аппарата и прикреплений мышц ограничивают у копытных все движения, кроме разгибания-сгибания, поэтому часть
мышц с иной функцией редуцировалась, а их функцию в качестве побочного
действия стали выполнять экстензоры и флексоры.
ЭКСТЕНЗОРЫ из всех мышц крупа особенно многочисленны и мощны,
так как именно на них падает основная нагрузка при разгибании конечности.
Их разделяют на две группы по месту расположения: ягодичную и заднебедренную. В ягодичную группу входит несколько ягодичных мышц, которые
действуют на тазобедренный сустав. Заднебедренную группу составляют:
двуглавая бедра, полусухожильная, полуперепончатая, квадратная мышца
бедра.
Они действуют на несколько суставов.
Поверхностная ягодичная мышца — m. gluteus superficialis (3) — пластинчатая, треугольная, лежит в ягодичной области между напрягателем широкой фасции бедра и двуглавой мышцей, срастаясь с ними. Самостоятельна
только у лошади. Начинается сухожильно от фасции, маклока и крестцовой
кости. Кончается на III вертеле бедренной кости. Одновременно с разгиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
банием пронирует тазобедренный сустав. Иннервируется ягодичными нервами. Васкуляризируется ветвями наружной подвздошной артерии.
Рис. 81. Мышцы тазовой
конечности крупного рогатого скота
(А, Б— с латеральной поверхности; В — с медиальной
поверхности;
Г—
сзади; Д — спереди): 1—
напрягатель широкой фасции бедра; 2 — средняя
ягодичная и.; 3 — поверхностная ягодичная м.; 4—
двуглавая м. бедра; 5 —
полусухожильная м.; 6 —
полуперепончатая м.; 7 —
малая поясничная м.; 8 —
подвздошная м.; 9— внутренняя запирательная м.;
10 — прямая, 11 —
медиальная и 12 — латеральная головки четырехглавой м. бедра; 13 —
портняжная м.; 14 — гребешковая м.; 15 — стройная м.; 16 — икроножная
м.; 17 — пяточная м.; 18
— большеберцовая передняя м.; 19 — малоберцовая
третья м.; 20 — малоберцовая длинная м.; 21 —
длинный пальцевый разгибатель; 22 — боковой
пальцевый разгибатель; 23
— сгибатель глубокий
пальцевый; 24 — поверхностный пальцевый сгибатель; 25 — межкостная
средняя м.
Средняя ягодичная мышца — m. gluteus medius (2) — толстая, мощная, заполняет ягодичную ямку подвздошной кости, частично прикрыта поверхностной ягодичной мышцей. Начинается от маклока и крестцового бугра
подвздошной кости. У рогатого скота и лошади заходит в область поясницы,
где срастается с длиннейшей мышцей спины. Оканчивается на большом вертеле бедренной кости. Относится к динамостатическому типу. Иннервируется краниальным ягодичным нервом. Васкуляризируется ветвями наружной и
внутренней подвздошных артерий. Одновременно с разгибанием несколько
отводит конечность.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Глубокая ягодичная мышца — m. gluteus profundus — небольшая, веерообразная, лежит в ягодичной ямке подвздошной кости под средней ягодичной мышцей. Начинается от седалищной ости тазовой кости. Оканчивается на большом вертеле бедренной кости. Участвует в разгибании вместе с
другими экстензорами. Основная функция — отведение конечности. Относится к полустатодинамическому типу. Иннервируется краниальным ягодичным нервом. Васкуляризируется ветвями внутренней подвздошной артерии.
Двуглавая мышца бедра — m. biceps femoris (4)—мощная, толстая, пластинчатая мышца, лежит под кожей в области крупа и бедра позади тазобедренного сустава. Имеет 2 головки с разным направлением мышечных пучков,
сросшихся друг с другом. Крестцовая головка начинается от гребня крестцовой кости и крестцово-седалищной связки, а седалищная головка — от седалищного бугра седалищной кости. Массивное брюшко оканчивается пластинчатым сухожилием у рогатого скота и свиньи на гребне большой берцовой кости и на бугре пяточной кости, у лошади, кроме того, на коленной
чашке. При действии с другими экстензорами является мощным разгибателем тазобедренного, коленного и заплюсневого сустава. При этом супинирует бедро и отводит конечность. У всех животных относится к динамостатическому типу. Иннервируется каудальный ягодичным и большеберцовым нервами. Васкуляризируется ягодичной, глубокой и каудальной бедренными артериями.
Полусухожильная мышца — m. semitendinosus (5) — толстая длинная,
лежит под кожей позади двуглавой мышцы бедра. Начинается у всех от седалищного бугра седалищной кости, а у свиньи, кроме того, от фасции в области крупа, у лошади от седалищного бугра и крестцовой кости. Мясистое
брюшко переходит в сухожилие, которое оканчивается на гребне большой
берцовой кости и бугре пяточной кости с медиальной стороны. При совместном действии с другими экстензорами разгибает тазобедренный и заплюсневый суставы. При обособленном действии пронирует и приводит конечность.
Относится к динамическому типу. Иннервируется каудальным ягодичным и
болыиеберцовым нервами. Васкуляризируется каудальной ягодичной, глубокой и каудальной бедренными артериями.
Полуперепончатая мышца — m. semimembranosus (6)—толстая пластинчатая, лежит на каудомедиальной поверхности бедра под приводящей
мышцей. Вместе с полусухожильной мышцей формирует задний контур бедра. Начинается от седалищного бугра седалищной кости, а у лошади, кроме
того, от первых хвостовых позвонков и крестцово-седалищной связки. Оканчивается на медиальных мыщелках бедренной и большой берцовой костей.
При совместном действии с другими экстензорами разгибает тазобедренный
и коленный суставы. При обособленном действии приводит и пропирует суставы. Относится к динамическому типу. Иннервируется большеберцовым
нервом. Васкуляризируется каудальной ягодичной, глубокой и каудальной
бедренной артериями.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Квадратная мышца бедра — m. quadratus femoris — небольшая, лежит
под двуглавой мышцей и между приводящей и наружной запирательной
мышцами. Начинается от вентральной поверхности седалищной кости.
Оканчивается на задней поверхности бедренной кости. Относится к динамическому типу. Иннервируется седалищным нервом. Васкуляризируется запирательной артерией (ветвью внутренней подвздошной артерии).
ФЛЕКСОРЫ. Большая поясничная мышца — m. psoas major (см. рис.
78—3) —лежит в области поясницы. Начинается от тел последних грудных и
поясничных позвонков, оканчивается на малом вертеле бедренной кости. Относится к динамическому типу. Иннервируется грудными и поясничными
нервами. Васкуляризируется глубокой бедренной артерией. Сгибает тазобедренный сустав и поясницу.
Подвздошная мышца —m. iliacus — мясистая, треугольной формы,
лежит на передней поверхности подвздошной кости (рис. 81—8). Начинается
от подвздошной и крыла крестцовой костей, от сухожилия малой поясничной
мышцы. Оканчивается на малом вертеле бедренной кости, срастаясь с большой поясничной мышцей. Относится к динамическому типу. Иннервируется
бедренным нервом. Васкуляризируется ветвями внутренней подвздошной артерии. Сгибает и одновременно супинирует тазобедренный сустав.
Напрягатель широкой фасции бедра — m. tensor fasciae latae (11) —
треугольной формы, лежит поверхностно вдоль переднего края бедра. Начинается от маклока, веерообразно расширяется и сливается с широкой фасцией бедра. У лошади тянется до коленной чашки и гребня большой берцовой
кости. Относится к динамическому типу. Иннервируется краниальным ягодичным нервом. Васкуляризируется ветвями внутренней подвздошной артерии. Сгибает тазобедренный и разгибает коленный суставы.
Портняжная мышца — m. sartorius (13)— пластинчатая, в виде ленты
лежит на медиальной поверхности бедра вдоль переднего края стройной
мышцы. Начинается от сухожилия малой поясничной мышцы и тела подвздошной кости до фасции около коленной чашки. Относится к динамическому типу. Иннервируется бедренным нервом. Васкуляризируется краниальной бедренной и ясной артериями. Является флексором и аддуктором тазобедренного и экстензором коленного сустава.
Гребешковая мышца — m. pectineus (14)—небольшая, треугольной
формы, лежит позади портняжной мышцы. Начинается на подвздошнолонном возвышении, оканчивается на медиальной поверхности бедренной кости.
Относится к динамостатическому типу. Иннервируется запирательным и
бедренным нервами. Васкуляризируется глубокой бедренной артерией. В зависимости от фазы движения мышца выполняет функции флексора, аддуктора и супинатора.
АДДУКТОРЫ. Стройная мышца — m. gracilis (15) —толстая пластинчатая, лежит на медиальной поверхности бедра. Начинается на вентральной
поверхности лонной кости, оканчивается на медиальной поверхности гребня
большой берцовой кости. Относится к динамическому типу. Иннервируется
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
запирательным и бедренным нервами. Васкуляризируется глубокой бедренной и ясной артериями.
Приводящая мышца — m. adductor — лежит под стройной мышцей
позади гребешковой. Начинается от вентральной поверхности лонной кости,
оканчивается на медиальной поверхности нижней половины бедренной кости. Относится к динамическому типу. Иннервируется запирательным и бедренным нервами. Васкуляризируется глубокой бедренной и бедренной артериями.
Кроме этих мышц, в приведении конечности участвуют портняжная и
гребешковая мышцы.
АБДУКТОРЫ. Функцию абдукторов выполняют средняя и глубокая
ягодичные и двуглавая мышца бедра.
СУПИНАТОРЫ. Наружная запирательная мышца — m. obturatorius
externus — небольшая мышца, веерообразно развернутая на вентральной поверхности тазовой кости. Начинается медиально от запертого отверстия,
оканчивается сухожильно в вертлужной впадине. Относится к полустатодинамическому (рогатый скот) или динамостатическому (свинья) типу. Иннервируется запирательным нервом. Васкуляризируется ветвями глубокой бедренной артерии.
Внутренняя запирательная мышца — m. obturatorius inter nus (9) —
плоская, треугольная, лежит на дорсальной поверхности тазовой кости. Начинается у рогатого скота от седалищной, у лошади и свиньи и от подвздошной костей. Проходит через запертое отверстие, сливаясь с сухожилием наружной запирательной мышцы. Относится к полустатодинамическому типу.
Иннервируется запирательным нервом. Васкуляризируется ветвями глубокой
бедренной артерии.
В супинации тазобедренного сустава участвуют, кроме того, подвздошная, большая поясничная, гребешковая, квадратная, двуглавая мышцы.
ПРОНАТОРЫ. Функцию пронаторов наряду с основной функцией разгибания тазобедренного сустава выполняют поверхностная ягодичная, полусухожильная, полуперепончатая мышцы.
Мышцы области бедра, действующие на коленный сустав. Коленный
сустав одноосный, в нем действуют разгибатели и сгибатели.
ЭКСТЕНЗОРЫ. Четырехглавая мышца бедра — m. quadriceps femoris
(10—12) —самая крупная мышца свободной конечности. Лежит на краниальной, латеральной и медиальной поверхностях бедренной кости, образуя передний контур бедра. Состоит из четырех головок веретенообразной формы,
объединенных у дистального конца. Прямая головка начинается на теле подвздошной кости, над суставной ямкой, лежит поверхностно на краниальной
поверхности бедренной кости; латеральная головка — от латеральной поверхности бедренной кости; медиальная головка — от медиальной поверхности бедренной кости; промежуточная головка — от краниальной поверхности
бедренной кости. Все головки, сливаясь, закрепляются на коленной чашке, а
их сухожилия продолжаются в качестве трех прямых связок коленной чашки
и оканчиваются на большой берцовой кости. Относится к динамостатическоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
му типу. Иннервируется бедренным нервом. Васкуляризируется бедренной
артерией. Прямая головка участвует в сгибании тазобедренного сустава.
В разгибании коленного сустава, кроме того, участвуют двуглавая мышца бедра (у лошади), напрягатель широкой фасции, портняжная, а при опирающейся конечности — полуперепончатая и полусухожильная мышцы.
ФЛЕКСОРЫ. Подколенная мышца — m. popliteus — небольшая треугольная, лежит на каудальной поверхности большой берцовой кости. Начинается на латеральном надмыщелке бедренной кости, оканчивается на линии
подколенной мышцы большеберцовой кости. Иннервируется большеберцовым нервом. Васкуляризируется передней большеберцовой артерией.
В сгибании коленного сустава в различные фазы движения принимают
участие двуглавая мышца бедра, полусухожильная, полуперепончатая, трехглавая мышца голени.
Мышцы области голени, действующие на заплюсневый (скакательный)
сустав. Заплюсневый сустав одноосный, на него действуют разгибатели и
сгибатели.
ЭКСТЕНЗОРЫ. Трехглавая мышца голени — m. triceps surae — образует задний контур голени и состоит из двух слившихся мышц: икроножной
и подошвенной. Икроножная мышца — m. gastrocnemius (16) — начинается
двумя головками по краям надмыщелковой (плантарной) ямки бедренной кости. Подошвенная (пяточная) мышца — m. soleus (17) — слабая, начинается у
рогатого скота и лошади на малоберцовой кости, у свиньи — на бедренной
кости. Мышцы вскоре сливаются и образуют сухожилие, к которому добавляются сухожилия поверхностного пальцевого сгибателя, двуглавой, полусухожильной и полуперепончатой мышц. В результате формируется мощное
общее пяточное, или ахиллово, сухожилие, которое закрепляется на бугре пяточной кости.
Трехглавая мышца голени участвует в сгибании коленного сустава. Относится к динамостатическому типу. Иннервируется большеберцовым нервом. Васкуляризируется бедренной и подколенной артериями.
В разгибании заплюсневого сустава принимают участие двуглавая мышца бедра и полусухожильная мышца.
ФЛЕКСОРЫ. Краниальная (передняя) большеберцовая мышца— m.
tibialis cranialis (anterior) (18)—слабая тонкая, начинается у рогатого скота и
свиньи на проксимальном конце и на гребне большеберцовой кости, у лошади — еще и от малоберцовой кости. Оканчивается на I заплюсневой, а также
у рогатого скота на III—IV плюсневых костях, у свиньи — на II плюсневой, у
лошади — на II и III плюсневых костях. У крупного рогатого скота и свиньи
относится к динамическому, у овцы и лошади — к статодинамическому типу.
Иннервируется малоберцовым нервом. Васкуляризируется передней большеберцовой артерией.
Малоберцовая третья мышца — m. peroneus (fibularis) tertius (19)—
веретенообразная, лежит на передней поверхности голени, прикрывая разгибатели пальцев. Начинается на латеральном надмыщелке бедренной кости,
оканчивается у рогатого скота на II—III заплюсневой и III—IV плюсневой
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
костях; у свиньи— на I и II заплюсневых и II плюсневой костях; у лошади —
на III и IV заплюсневых и на II и III плюсневых костях. Относится у свиньи и
полустатодинамическому, у рогатого скота — к статодинамическому, у лошади — к статическому типу.
Малоберцовая длинная мышца — m. peroneus (fibularis) longus (20)—
длинная тонкая мышца. Лежит на латеральной поверхности голени. Начинается от малоберцовой кости и латерального мыщелка большеберцовой кости.
Оканчивается на I и II заплюсневых костях. У лошади отсутствует. Относится к динамостатическому типу. Иннервируется малоберцовым нервом. Васкуляризируется передней большеберцовой артерией.
Мышцы области голени, действующие на суставы пальцев. Суставы
пальцев одноосные, на них действуют экстензоры и флексоры.
ЭКСТЕНЗОРЫ. Длинный пальцевый разгибатель — m. extensor digitalis longus (21) —веретенообразная мышца, ее брюшки (1—3 у разных видов
животных) прикрыты малоберцовой третьей мышцей. Начинается на латеральном надмыщелке бедренной кости. Сухожилие принимает и отдает сухожильные связки к другим пальцевым мышцам, перед окончанием разветвляется и оканчивается на разгибательном отростке копытцевой (копытной)
кости каждого пальца. Относится к динамостатическому типу. Иннервируется малоберцовым нервом. Васкуляризируется передней большеберцовой артерией.
Боковой пальцевый разгибатель — m. extensor digitalis lateralis (22) —
слабая веретенообразная мышца, лежит на боковой поверхности голени рядом с длинным пальцевым разгибателем. Начинается на латеральном мыщелке большеберцовой кости (рогатый скот) и на малоберцовой кости (свинья, лошадь) 1— 2 брюшками. Сухожильно заканчивается на венечной кости
IV пальца у рогатого скота, IV и V у свиньи, III у лошади, срастаясь с сухожилием длинного пальцевого разгибателя. Относится к полустатодинамическому типу. Иннервируется малоберцовым нервом. Васкуляризируется передней большеберцовой артерией.
ФЛЕКСОРЫ. Поверхностный пальцевый сгибатель — m. flexor digitalis superficialis (24)—лежит на каудальной поверхности голени, прикрыта
икроножной мышцей, с которой частично срастается. Начинается от надмыщелковой (плантарной) ямки бедренной кости. Сухожилие сплетается с сухожилием икроножной мышцы, перекидывается через пяточный бугор. На
уровне плюснопальцевой (путовой) фаланги делится по числу пальцев и закрепляется на венечных костях, формируя по 2 ножки, через которые проходит сухожилие глубокого пальцевого сгибателя. Относится к статодинамическому типу. Иннервируется малоберцовым нервом. Васкуляризируется бедренной, подколенной и передней большеберцовой артериями.
Глубокий пальцевый сгибатель — m. flexor digitalis profundus (23) —
лежит на каудальной поверхности голени, прикрыт трехглавой мышцей голени и поверхностным пальцевым сгибателем. Имеет несколько веретенообразных брюшек. Начинается на латеральном мыщелке и плантарной поверхности большеберцовой кости у рогатого скота, а также на малоберцовой косwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ти у свиньи и лошади. Их общее сухожилие проходит по плантарной поверхности заплюсны, минуя пяточный бугор. В области плюсны разветвляется по
количеству пальцев, проходит между ножками сухожилия поверхностного
пальцевого сгибателя и оканчивается на сгибательном отростке копытцевых
(копытной) костей каждого пальца. Разные брюшки относятся к разным типам по внутренней структуре: от динамостатического до статодинамического. Иннервируется малоберцовым нервом. Васкуляризируется
задней большеберцовой артерией.
Межкостные мышцы — mm. interossei — сухожильно-мышечные тяжи,
идущие по плантарной поверхности плюсневой кости. Начинаются на плантарной поверхности заплюсневого сустава и плюсны. По ходу соединяются
сухожильными связками с другими пальцевыми мышцами. Оканчиваются на
путовой кости, а также, переходя на переднюю поверхность пальцев, сливаются с сухожилиями пальцевых разгибателей. Относятся к статодинамическому и статическому типам. Иннервируется малоберцовым нервом.
Глава 14. СИСТЕМА ОРГАНОВ КОЖНОГО ПОКРОВА
Общий кожный покров — integumentum commune — одевает все тело
животного, отграничивая его от внешней среды. Его состав и строение резко
отличаются у животных разных классов в связи с их экологическими и морфофизиологическими особенностями. У млекопитающих в систему органов
кожного покрова входят кожа — cutis — и ее производные: волосы, потовые,
сальные, пахучие и молочные (вымя) железы, рога, копытца, копыта, когти,
мякиши, каштаны, шпоры (у лошади), кожные ушные складки, кожные перепонки на конечностях (летучая мышь, бобр), иглы (еж, дикобраз).
Находясь на границе между внутренней и внешней средой и подвергаясь
прежде других органов воздействию внешней среды, кожный покров приобрел ряд свойств, позволяющих противостоять этим воздействиям, способствуя сохранению постоянства внутренней среды организма. Кожный покров
обладает плотностью, прочностью, упругостью, непроницаемостью для
большинства веществ, имеет кислую реакцию (рН 3,2—5,2).
Основная функция кожного покрова — защитная во всех ее проявлениях: предохраняет тело от избытка влаги (не набухает в воде) или ее недостатка (не сморщивается при сухости), от проникновения микроорганизмов,
ультрафиолетовых лучей, а также от различных механических и химических
воздействий. Осуществляет терморегуляционную функцию благодаря развитым сосудистым сетям. Принимает участие в выделительной функции и водно-солевом обмене (с помощью желез), в функции дыхания, в витаминном
обмене (синтез витамина D). Является депо крови (в сосудах кожи может содержаться до 20%. крови организма), жира (в подкожной жировой клетчатке,
горбах у верблюда, в курдюке у курдючной овцы), воды (в соединительной
ткани кожи может скапливаться до 1/6 всей воды организма; источник воды
— это и жир в процессе его расщепления). Считается огромным рецепторным полем, образуя орган осязания, который осуществляет связь организма
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
со средой обитания. Благодаря рецепторной функции кожного покрова организм получает информацию о тактильном (контактном) внешнем воздействии (боль, тепло, холод, зуд и др.).
Благодаря столь разнообразным функциям кожный покров оказывается
связан со многими системами организма: нервной (рецепторная деятельность
кожи), выделительной (удаление веществ через потовые и сальные железы),
дыхательной (проникновение кислорода через неповрежденную кожу), половой. Связь с половой системой проявляется в развитии вторичных половых
признаков — изменении участков кожи или ее производных, свойственных
лишь определенному полу.
Кожный покров имеет и прикладное значение. Кожа с производными,
снятая с животного, называется шкурой. Из нее выделывают меха, кожи. Волосяной покров овец (руно) и коз идет на изготовление шерстяных изделий.
Молоко является полноценным продуктом питания.
КРАТКИЕ СВЕДЕНИЯ О РАЗВИТИИ СИСТЕМЫ ОРГАНОВ КОЖНОГО ПОКРОВА
Филогенез. У беспозвоночных наружный покров происходит из эктодермы. У хордовых кожный покров имеет сложное происхождение. У низших хордовых (ланцетник) поверхностный слой — эпидермис образован однослойным эпителием, а лежащая под ним основа кожи состоит преимущественно из студенистой ткани. С усложнением организации хордовых
происходит увеличение слоев и дифференцировка клеток эпидермиса. У рыб
эпидермис представлен многослойным эпителием, в нем развивается большое количество одноклеточных слизистых желез. Еще большее их количество в коже амфибий, они стали многоклеточными и опустились глубже — в
дерму кожи. В эпидермисе проходят протоки желез, открывающиеся на поверхности кожи. В дерме все большее распространение получают рыхлая и
плотная соединительные ткани. Появляются такие производные кожного покрова, как чешуя. Наиболее древняя ее форма — плакоидная чешуя, в образовании которой принимают участие эпидермис и дерма. Чешуя современных
рыб и зубы всех хордовых произошли из плакоидной чешуи.
С выходом на сушу происходит значительное уплотнение кожного покрова с целью увеличения его прочности, уменьшения испарения и сохранения влаги в организме. Эпидермис сильно ороговевает или покрывается
сложными ороговевшими выростами: чешуей, перьями, волосами. В определенных местах появляются роговые образования: щитки, когти, копытца, рога. Дерма подразделяется на два слоя: сосочковый и сетчатый. Сосочковый
слой, образованный рыхлой соединительной тканью, содержит большое количество сосудов и нервных окончаний. Он выполняет трофическую функцию, так как в эпидермис сосуды не заходят, и осуществляет связь эпидермиса с дермой. Сетчатый слой образован плотной соединительной тканью о
разным направлением пучков коллагеновых волокон. Он обеспечивает прочность кожного покрова. У рептилий и птиц кожа сухая, содержит только спеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
циализированные железы: копчиковую у птиц, бедренные у ящериц, мускусную у крокодилов и некоторые другие.
У млекопитающих эпидермис образован многослойным плоским ороговевающим эпителием. Дерма значительно толще эпидермиса, состоит из сосочкового слоя, представленного рыхлой соединительной тканью, и сетчатого слоя из плотной неоформленной соединительной ткани. Производные кожи разнообразны по структуре и функции. На конечности — это мякиши,
когти, копытца, копыта, на голове — рога. Разнообразны и кожные железы.
Это сальные, нескольких видов потовые и, наконец, молочные железы, характерные только для млекопитающих. По-видимому, они развиваются на
базе видоизмененных апокринных потовых желез, что хорошо видно у низших млекопитающих ехидны и утконоса, у которых молочная железа имеет
вид железистого поля. У сумчатых и еще в большей степени у плацентарных
молочная железа дифференцируется в крупной паренхиматозный орган.
Онтогенез. Развитие кожи. Кожный покров развивается из эктодермы
и мезодермы. После отшнуровывания нервной трубки оставшаяся эктодерма
становится кожной эктодермой. Из нее образуется эпидермис кожи. У эмбриона в зародышевом периоде эпидермис имеет вид однослойного эпителия. На базе дерматомов сомитов мезодермы развивается основа кожи—
дерма. В зародышевый период мезодерма сомитов и частично париетального
листка спланхнотомов разрыхляется, превращается в мезенхиму, которая в
предплодный период начинает дифференцироваться в соединительную
ткань.
В течение зародышевого и предплодного периодов кожный покров формируется довольно медленно, К концу предплодного — началу плодного периода эпидермис состоит из двух слоев клеток, дерма представлена в виде
мезенхимного синцития, в котором мезенхимные клетки превращаются в
фибробласты. Сосуды образуют в дерме нежную сеть. В плодный период
развитие кожного покрова ускоряется: рост кожи происходит почти в 3 раза
интенсивнее роста плода в целом. Ее масса увеличивается в 240 раз, относительная масса — с 4 до 12%, а площадь в 816 раз. В течение плодного периода происходит нарастание слоев эпидермиса и превращение мезенхимы в соединительную ткань. У 2-месячных плодов свиней и овец, у 3-месячных плодов телят число слоев эпидермиса достигает 5—7. Он дифференцирован на
базальный, шиповатый и роговой слои. В дерме видны фибробласты и тонкие
рыхло лежащие пучки коллагеновых волокон. Происходит закладка волосяных фолликулов, потовых желез, несколько позже — сальных желез в виде
тяжей эпителия, опускающихся в глубь дермы. Еще через месяц эпидермис,
луковицы волос, потовые и сальные железы сформированы, в дерме начинают различаться по структуре сосочковый и сетчатый слои. Все слои кожи активно растут в толщину, но у разных видов животных с разной скоростью. У
плодов всех животных эпидермис составляет 3— 4%, сетчатый же слой у
плодов телят равен 65% от толщины кожи, у овцы—35%, у свиньи —27%.
В период выхода зачатков волос на поверхность кожи, что наблюдается
у плодов свиней в 3,5 месяца, овец в 4, телят в 7 месяцев, происходит слущиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
вание поверхностных слоев эпидермиса, он резко истончается. К концу утробного развития все структуры кожи полностью сформированы. Эпидермис
безволосых участков кожи состоит из пяти слоев, кожи с волосами из 3—4
слоев. Потовые и сальные железы уже функционируют, стержни волос отрастают за пределы корня.
Развитие волос. Закладка волос происходит в начале плодного периода,
когда эпидермис трехслойный. В месте будущего волоса разрастаются клетки
базального слоя эпителия. По мере размножения клеток зачаток волоса —
волосяной фолликул в виде колбовидного утолщения врастает в глубь дермы
несколько под углом к поверхности кожи. Из нижней, расширенной части зачатка волоса формируется луковица — участок корня волоса, состоящий из
молодых делящихся клеток, Участок фолликула выше луковицы преобразуется в два влагалища — наружное и внутреннее. В про« цессе дифференцировки фолликула часть клеток отмирает и стержень волоса оказывается отъединенным от окружающих его эпителиальных клеток. Соединительная ткань
врастает снизу в луковицу, формируя волосяной сосочек, а вокруг образует
сумку волоса. Из окружающей мезенхимы дифференцируются сосуды волосяного сосочка и мышца, поднимающая волос.
Раньше других закладываются вибриссы — синуозные осязательные волосы, расположенные в разных местах морды. Из кроющих волос сначала закладываются так называемые первичные фолликулы, из которых развиваются грубые остевые волосы в наиболее толстых участках кожи — на беках,
спине, крупе. Более тонкие пуховые волосы развиваются из вторичных фолликулов, которые закладываются позже вокруг первичного фолликула и
имеют меньшие размеры. У свиней фолликулы расположены поодиночке.
Лучше всего группы фолликулов развиты у овец. У грубошерстных вокрур
одного первичного фолликула развивается 3—5, у тонкорунных—10—15
вторичных фолликулов.
Потовые железы закладываются и развиваются одновременно с волосами, сальные железы — несколько позже.
Развитие молочной железы. Закладка молочных желез обнаруживается в
предплодный период в виде парных утолщений эпидермиса на брюшной стороне зародыша — млечных валиков. Мезенхима в области млечных валиков
также интенсивно нарастает, в результате чего образуется выпячивание в виде линзы, а затем колбочки. В зачатке молочной железы по мере ее роста
идет дифференцировка эпителия, который, проникая внутрь, выстилает формирующийся канал соска и сосковую цистерну. Мезенхима, отрастая кнаружи, формирует соединительнотканную оболочку соска. Под зачатком соска
разрастаются сосудистые сплетения и жировые островки, У плодов коров
молочных пород зачаток молочной железы дифференцируется быстрее, чем у
мясных.
Несмотря на раннюю закладку, молочные железы в утробный период отстают от роста тела и к рождению имеют вид небольших сосочков, в которых
залегают зачатки будущих цистерн. Альвеолы отсутствуют. В постнатальный период до полового созревания молочные железы развиваются
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
очень медленно. Разрастаются в основном жировые островки, в меньшей степени — соединительнотканная строма и еще меньше — система протоков
железы. С наступлением половой зрелости начинают усиленно развиваться
соединительная ткань, железистый эпителий и сосудистая сеть вымени. Соединительнотканная строма обгоняет в развитии железистую паренхиму, и
только во время беременности эпителиальная ткань растет быстрее соединительной, она дифференцируется во все элементы железистой паренхимы и к концу беременности начинает продуцировать молозиво. Наивысшей же функциональной активности молочная железа достигает во время
лактации.
СТРОЕНИЕ КОЖНОГО ПОКРОВА
Кожа —cutis —прочная, упругая оболочка, покрывающая тело животного, в области естественных отверстий переходящая в слизистую оболочку.
Масса кожи у большинства взрослых животных находится в пределах 5—7%
(без руна у овец) от массы тела, что составляет у крупного рогатого скота
20—40 кг, у овцы 1,5—2,5, у свиньи 7—10, у лошади 8—20 кг. У новорожденных животных кожа относительно тяжелее: у телят 11—12%, У поросят 12—18%. Толщина кожи различается у животных разных видов и на разных участках тела: у крупного рогатого скота кожа плотная, средней толщины (3—6 мм), у овец — тонкая (0,7—2 мм), у свиней грубая и толстая (с подкожной клетчаткой 5—7 см), у лошадей 1—7 мм. Обычно более толстая кожа
находится на дорсальной поверхности шеи (загривке), спине, крупе; средней
толщины — по бокам; наиболее тонкая — на брюхе и медиальных поверхностях конечностей, особенно в области паховой складки. Имеются отличия в
толщине кожи, связанные с полом и особенностями поведения: у хряков, кабанов и козлов наиболее толстая кожа на вентральной части шеи и предгрудинной области, так называемый щит.
Состоит кожа из эпидермиса, дермы и подкожной клетчатки (рис. 82).
Эпидермис — epidermis (1)—поверхностный слой кожи, которым она
контактирует с окружающей средой. Толщина его составляет у крупного рогатого скота 30—60 мкм, у овцы 10—30 мкм, у лошади 40—50 мкм. Образован он многослойным плоским ороговевающим эпителием. В участках тела,
не покрытых волосами, состоит из 5 слоев: базального, шиповатого, зернистого, блестящего и рогового (см. раздел третий). Базальный слой выполняет
камбиальную функцию за счет размножения стволовых клеток, входящих в
его состав. Между клетками эпителия — эпидермоцитами залегают отростчатые пигментные клетки — меланоциты, вырабатывающие и накапливающие
пигменты. Они обусловливают цвет кожи и выполняют функцию защиты от
избытка ультрафиолетовых лучей.
Защитные свойства эпидермиса определяются толщиной и состоянием
его рогового слоя. Повышение температуры, воздействие щелочами, кислотами, органическими растворителями увеличивает проницаемость эпидермиса. Он пополняется за счет клеток, поступающих из нижележащих слоев, коwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
торые по мере выдвижения на поверхность кожи подвергаются ороговению.
Стимулом к ороговению служат затруднение питания и изменения веществ в
клетках по мере их удаления от базальной мембраны (питание эпителия осуществляется диффузно). При этом в клетках накапливаются кератиноидные
белки, гранулы которых заметны в верхних рядах эпидермоцитов шиповатого слоя. В клетках зернистого слоя гранулы кератогиалина заполняют всю
цитоплазму, клетки уплощаются и как бы распластываются. В процессе гибели клетки зерна кератогиалина сливаются и подвергаются химическим
превращениям — образуется элеидин, заполняющий плоские широкие клетки блестящего слоя.
Рис. 82. Строение кожи (А— без волос, Б — с волосами):
I — эпидермис; II дерма; III— подкожная клетчатка; 1—роговой слой;
2—блестящий слой; 3 — зернистый слой; 4 — шиповатый слой; 5 — базальный слой - дерма; 6 — сосочковый слой; 7 — сетчатый слой; 8—выводной
проток и 9 — концевой отдел потовой железы; 10 — кровеносный сосуд; 11
— лимфатический сосуд; 12 — 14 — скопления жировых клеток; 15 — стержень волоса; 16 — корень волоса; 17 — волосяная луковица; 18 — нерв; 19 —
нервные окончания; 20 — сменяющийся волос. пучки коллагеновых волокон.
Кератинизация сопровождается потерей воды. Зрелый элеидин становится кератином — белком, богатым серой, очень устойчивым к различным
воздействиям. Клетки к этому времени отмирают и имеют вид роговых чешуек — плоских распластанных по поверхности так, что площадь одной чешуйки равна площади, занимаемой 5—15 базальными клетками. Чешуйки
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
содержат кератин, пузырьки воздуха, между ними обнаруживаются нейтральные жиры. При нарушении связи между клетками (деградация десмосом) чешуйки слущиваются. Роговой слой состоит из многих рядов чешуек.
В участках, подвергающихся постоянному трению, давлению и другим механическим воздействиям, роговой слой значительно утолщается, что можно
видеть в строении мякишей. В норме изнашивание и слущивание эпителия с
поверхности рогового слоя восполняются нарастающими и дифференцирующимися клетками.
В участках кожи, покрытых волосами, эпидермис тоньше и состоит из
базального, шиповатого, зернистого и рогового слоев.
Дерма — derma (II), или собственно кожа, подстилает эпидермис, отграничена от него базальной мембраной, образована соединительной тканью и
состоит из сосочкового и сетчатого слоев.
В участках кожи, лишенных волос, граница между эпидермисом и дермой неровная. Врастание эпидермиса в глубь дермы называется гребешками.
Дерма вклинивается в эпидермис в виде сосочков, отчего прилежащий к эпидермису слой называют сосочковым. В коже с волосами гребешки и сосочки
выражены слабо. Этот слой сильно различается по толщине у животных разных видов: от 100 мкм у свиньи до 2 мм у рогатого скота и составляет у коровы 18—20%, у свиньи 3%, У лошади 30—35% от толщины дермы. Для сосочкового слоя характерно то, что он состоит из рыхлой соединительной ткани с большим количеством разнообразных клеточных элементов (фибробласты, фиброциты, гистиоциты, лаброциты, пигментные клетки и др.), кровеносных и лимфатических капилляров, нервов и нервных окончаний.
Между слоями дермы нет четких границ. Деление основано на том, что
сетчатый слой в отличие от сосочкового образован плотной неоформленной
соединительной тканью. Эта ткань содержит большое количество коллагеновых и гораздо меньше эластических волокон. Среди соединительнотканных
белков этого слоя 98% приходится на коллаген. В зависимости от величины,
плотности, направления и характера переплетения пучков коллагеновых волокон различают несколько типов вязи. От характера вязи зависит прочность
сетчатого слоя дермы, его механические свойства. У большинства животных
этот слой является основным по толщине.
Подкожная основа (клетчатка) — tela subcutanea (III)—образована
рыхлой соединительной тканью. Связывает кожу с поверхностной фасцией и
подкожной мускулатурой. В подкожной основе часто наблюдается скопление
жировой ткани, особенно при откорме. Чаще всего жир откладывается в области шеи, плеча, холки, узкой полосой вдоль позвоночника, на крестце, по
бокам, у заднего края грудной клетки, в паху, вокруг ануса. У свиньи в подкожной клетчатке по всему телу нарастает большое количество жира —
шпик. У верблюда жир откладывается в горбах, у курдючных овец — у корня
хвоста и на ягодицах, образуя курдюк. Подкожная основа обеспечивает коже
подвижность, является жировым депо. Благодаря скоплению жира кожа принимает участие в терморегуляции.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Васкуляризация и иннервация кожи. Кожа обильно снабжена сосудами, которые образуют в ней несколько сплетений: под эпидермисом, между
слоями дермы, на границе с подкожной основой и в самой основе. Как между
сплетениями, так и внутри имеются многочисленные анастомозы, участвующие в перераспределении крови и терморегуляции.
Иннервируется кожа как нервами соматической, так и вегетативной
нервной системы. Они образуют сплетения в подкожной основе, в дерме и
под эпидермисом. Чувствительные нервы соматической нервной системы образуют в коже многочисленные и разнообразные чувствительные нервные
окончания как свободные, так и несвободные. Свободные нервные окончания
преимущественно разветвляются в эпидермисе. В дерме присутствуют свободные прямолинейные, клубочковые, древовидные и несвободные (Фатера
— Пачини, Краузе, Мейснера) нервные окончания. Особенно богата нервными окончаниями кожа век, губ, хоботка, носогубного зеркальца, ушей,
брюшной стенки, пальцевых мякишей, вымени, половых органов. Вегетативная нервная система иннервирует сосуды, железы, мышцы- подниматели волос.
Волосяной покров — особенность кожного покрова млекопитающих.
Он образован производными эпидермиса — волосами и защищает кожу от
влаги, механических, электрических и других воздействий. В различных участках тела и у животных разных родов волосяной покров сильно различается
по составу, густоте, длине волос. У овец тонкая кожа покрыта длинным, густым волосяным покровом. У свиней кожа толстая с редко расположенными
грубыми волосами, которые на определенных участках становятся щетиной.
У крупного рогатого скота сравнительно однообразный, средний по густоте
волосяной покров.
По особенностям строения и функции различают осязательные, длинные
и кроющие волосы. Осязательные (синуозные, вибриссы) волосы — толстые
чувствительные, корень их окружен сосудами и нервными окончаниями.
Располагаются на морде около рта и глаз. Выполняют рецепторную функцию
Длинные волосы растут в определенных участках тела: на хвосте, на челке и
щетках, гриве у лошадей, по бокам тела у яков. Выполняют они защитную
функцию. Основная масса волос, покрывающих тело, называется кроющими,
или покровными. Они располагаются на коже в определенном порядке, образуя круговые и линейные потоки. Густота волос у животных разных родов и
семейств очень различна. Есть животные со слаборазвитым волосяным покровом. Из домашних животных это свинья. Количество волос у нее составляет 60—100 на см2 (из них 3—30 щетин), у крупного рогатого скота 800—
1500, у овец 4000—8000. Среди покровных волос различают щетину, ость,
полупух и пух. У крупного рогатого скота, свиньи и лошади волосы растут
более или менее равномерно, у овцы — группами. В группе один остевой и
несколько пуховых (у тонкорунных — до 15 шт.) волос.
Волос — pilus (рис. 83)—твердая ороговевшая эластичная нить конической, цилиндрической, веретеновидной, ланцетовидной формы и различной
степени извитости и окраски, образованная эпителиальными клетками. В нем
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
различают стержень и корень. Стержень волоса выступает над кожей. Корень находится в коже и окружен волосяным фолликулом. Фолликулы остевых волос закладываются первыми и называются поэтому первичными. Фолликулы пуховых волос закладываются позже и называются вторичными.
Корни волос у большинства животных залегают в сетчатом слое дермы. У
свиньи корни щетин проникают в подкожную клетчатку.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 83. Развитие (А) и
строение (Б) волоса (о, б,
в, г, д — последовательные стадии развития
волоса):
1 — эпидермис; 2— дерма; 3— волосяной фолликул; 4 — волосяная луковица; 5 — волосяной сосочек; 6—корень волоса; 7
— сумка волоса; 8 — наружное корневое влагалище; 9 — внутреннее
корневое влагалище; 10 —
подниматель волоса; 11
— сальная железа; 12 —
роговое вещество; 13 —
потовая железа; 14 —
кутикула; 15 — норковое
вещество; 16—жировые
клетки; 17 — воронка
фолликула.
Фолликул волоса имеет сложное строение. Полость фолликула, в которой расположен корень волоса, называется каналом фолликула, а его расширенная верхняя часть—воронкой. Наружная часть его стенки образована соединительной тканью и называется сумкой волоса. Дно сумки волоса утолщено, состоит из рыхлой соединительной ткани, пронизанной сосудами, и вдается в глубь фолликула в виде сосочка волоса. Сосу ды сосочка питают волос. Внутренняя часть фолликула эпителиальная. Она состоит из двух влагалищ — наружного и внутреннего — и переходит в колбообразное утолщение
— луковицу волоса. Наружное корневое влагалище является продолжением
росткового слоя эпидермиса и, постепенно истончаясь, покрывает луковицу
1—2 слоями клеток. Внутреннее корневое влагалище отрастает от луковицы
и доходит только до воронки фолликула. У крупного рогатого скота оно имеется по всей длине фолликула и переходит в роговой слой эпидермиса. На
уровне воронки в канал фолликула волоса открываются протоки сальной и
апокринной потовой желез. В сумку волоса (ости и щетины) вплетается
мышца — подниматель волоса из гладкой мышечной ткани.
Клетки волосяной луковицы, размножаясь, продвигаются вверх по каналу
фолликула, формируя корень, а затем стержень волоса. Существует мнение:
чем крупнее луковица, тем толще растущий волос. Клетки вскоре ороговевают и стержень волоса состоит из ороговевших клеток, лежащих в несколько
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
концентрических слоев. Самый наружный слой клеток волоса — кутикула—
представлена однослойным плоским эпителием, имеющим вид чешуек, налегающих друг на друга, как черепицы. Она химически нейтральна, выполняет
защитную функцию. Клетки кутикулы у животного каждого вида имеют характерную форму, так что можно определить, кому принадлежит волос.
Под кутикулой расположено корковое вещество (кора) волоса из нескольких рядов концентрически наслоенных эпителиальных клеток. В процессе дифференцировки клетки захватывают пигментные зерна из отростков
меланоцитов, залегающих в луковице, и быстро ороговевают. Ороговевшие
клетки коры, как и кутикулы, содержат твердый кератин и остатки ядер. Корковое вещество волоса определяет его цвет, прочность и эластичность. В
центре волоса находится мозговое вещество, образованное крупными, медленно ороговевающими клетками. Ороговение завершается к переходу корня
волоса в стержень. Клетки содержат мягкий кератин, пузырьки воздуха.
Пуховые и полупуховые волосы состоят только из кутикулы и коркового
вещества. В ости имеются все три слоя.
Смена волос происходит периодически, так как срок жизни волоса от 50
дней до 5 лет. Массовая смена волос называется линькой. Она может быть
ювенальной — до полового созревания и сезонной — обычно весной и осенью. Бывает и постоянная смена волос (перманентная), не зависящая от годового цикла. Это относится в первую очередь к длинным волосам. Смена
начинается с нарушения питания волоса, затем происходит перестройка луковицы, после чего старый волос теряет с ней связь, а от нее отрастает новый
волос.
Сальные железы — glandulae sebaceae (см. рис.82— 13) —пристенные
простые альвеолярные с разветвленными концевыми отделами, секретируют
по голокринному типу. Развиваются из эпителия наружного волосяного влагалища первичного фолликула и открываются в канал фолликула волоса на
уровне его воронки. С фолликулами пуховых волос сальные железы не связаны. Концевые отделы железы — альвеолы — не имеют полости. Стенка
альвеолы выстлана одним слоем мелких малодифференцированных камбиальных клеток. При активном делении часть клеток вытесняется из пласта в
глубь альвеолы и вступает на путь жирового перерождения. Такие клетки укрупняются, в их цитоплазме накапливаются капли жира. По мере выталкивания клеток все ближе к протоку железы происходит все более выраженное
жировое перерождение клеток. Наконец, клетки, лежащие ближе других к
просвету, гибнут, распадаются, превращаясь в жировой секрет.
Проток железы выстлан многослойным плоским эпителием, открывается
в канал фолликула волоса. Жировой секрет предохраняет кожу и волосы от
высыхания, мацерации, химических воздействий. У овец жировой секрет
смешивается с секретом потовых желез, образуя жиропот, количество которого может достигать 40% от массы руна. Жиропот участвует в теплоотдаче,
увеличивает испаряющую поверхность кожи и шерсти, предохраняет кожу от
избыточного влияния солнечных лучей и влаги, придает эластичность, споwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
собствует формированию штапельного строения руна. Сальных желез нет на
мякишах, сосках (у коров), в коже носового зеркальца.
Потовые железы — glandulae sudoriferae (8, 9)— пристенные, простые,
трубчатые, иногда с ветвящимися концевыми отделами (особенно у свиньи и
лошади). У крупного рогатого скота концевые отделы желез слабо изогнуты,
у лошади, овцы и свиньи свернуты в виде клубочка. Встречаются по всей поверхности тела. По типу секреции различают мерокринные и апокринные потовые железы. Мерокринные расположены в участках кожи без волос, апокринные — в коже с волосами. Концевые отделы желез залегают в глубоких
слоях дермы на уровне волосяных луковиц, проникая в подкожную клетчатку. У апо-кринных желез они в 4—5 раз крупнее, чем у мерокринных. Выводные протоки имеют вид прямых или спирально извитых трубочек, проходящих через все слои кожи. У мерокринных желез они открываются порами
на поверхности кожи. У большинства апокринных желез протоки открываются в воронку волосяного фолликула, иногда — самостоятельно вблизи волоса.
Трубчатые концевые отделы образованы однослойным кубическим или
цилиндрическим эпителием, вырабатывающим секрет, 98% которого составляет вода, а 2% минеральные соли и некоторые органические вещества (белки, мочевина и др.). Апокринные железы содержат белки, которые обусловливают запах животного. Выводится секрет из концевых отделов при сокращении миоэпителиальных клеток, покрывающих снаружи железистый эпителий. Количество потовых желез у крупного рогатого скота в среднем около
1000 шт. на 1 см2 кожи. У коров молочных пород потовых желез на 5—25%
больше, чем у мясных. У свиней на 1 см2
приходится 8—25 желез, у овцы — 360.
Развитие апокринных потовых желез у телок используют как признак, позволяющий судить об их будущей молочной продуктивности, так как есть работы, свидетельствующие о положительной корреляции между развитием потовых и молочных желез.
В кожном покрове имеются и другие железы: серозные в но-согубном зеркальце крупного рогатого скота, железы
пятачка у свиньи, подглазничные, паховые и межкопытцевые железы у овцы.
Рис. 84. Вымя коровы:
СТРОЕНИЕ МОЛОЧНОЙ ЖЕЛЕЗЫ
а — чашеобразное, б — ваннообразное брюшное в — бедМолочная железа сельскохозяйстренное, г — «козье»; 1 — основание вымени; 2 — тело; 3 — дно; венных животных — вымя — uber (рис.
4 — сосок; 5 — молочное зерка- 84) —крупная, сложная, застенная, альвеолярно-трубчатая. У свиньи вымя
ло.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
множественное. Оно состоит из 4—8 пар долей, лежащих на вентральной поверхности грудной и брюшной стенки в виде отдельных холмов вдоль белой
линии живота. У рогатого скота и лошади вымя простое — его доли слились
в единый орган, расположенный в паховой области между бедер. У крупного
рогатого скота и лошади 4 доли вымени, у овцы и козы — 2. У рогатого скота
каждая доля имеет свой сосок. У лошади и свиньи один сосок связан с двумя
долями.
Вымя коровы (рис. 85). Поверхность вымени, прилежащая к животу, называется основанием вымени. Каудальная складчатая поверхность вымени —
молочное зеркало. Основная масса железы — тело вымени, вентральная
часть — дно вымени. Вымя покрыто кожей с потовыми и сальными железами
и редкими нежными волосами. Под кожей лежит поверхностная фасция вымени, а под ней глубокая — продолжение желтой брюшной фасции с большим количеством эластических волокон. Глубокая фасция, спускаясь от белой линии живота в виде подвешивающей связки, разделяет вымя на правую
и левую половины. Каждая половина состоит из двух долей: передней и задней, нечетко отделенных одна от другой. Каждая доля имеет свою систему
протоков и свой сосок, в котором различают основание, тело и верхушку.
Рис. 85. Схема строения (А) и
развития (Б) вымени коровы:
а — основание вымени; б — тело;
в — дно; г — сосок; 1 — кожа; 2
— поверхностная и 3—глубокая
фасции; 4— подвешивающая
связка; 5— строма вымени; 6—
дольки вымени; 7 — кровеносные
сосуды; 8 — лимфатические сосуды; 9 — нервы; 10 — альвеолотрубки; 11 — молочный канал
(проток); 12 — молочный ход; 13
— железистая цистерна; 14 —
сосковая цистерна; 15 — сосковый канал; 16 — сфинктер соска;
17 — миоэпителиальная клетка;
18 — млечный валик; 19 — зачаток соска; 20 — пробка из ороговевших клеток.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Под глубокой фасцией расположена соединительнотканная капсула, от
которой внутрь железы отходят соединительнотканные прослойки — трабекулы. Капсула и трабекулы составляют остов — строну вымени. В ней постоянно обнаруживаются скопления жировых клеток.
Железистая часть — паренхима — образована эпителием, из которого
построены концевые отделы и система выводных протоков железы. Трабекулы, проникая между железистыми элементами паренхимы, делят вымя на
дольки. В соединительной ткани трабекул к долькам подходят нервы, кровеносные и лимфатические сосуды. Основной объем дольки составляют концевые отделы — альвеолотрубки, или альвеолы, эпителий которых вырабатывает составные части молока. Тончайшие внутридольковые прослойки соединительной ткани с идущими в них нервами и сосудами оплетают каждую
альвеолотрубку.
Альвеолотрубки лактирующей молочной железы (рис. 86) имеют широкий просвет, заполненный альвеолярным молоком. Стенка альвеолы образована однослойным кубическим или цилиндрическим эпителием. Клетки
имеют крупные округлые или овальные ядра, хорошо развитую цитоплазматическую сеть и аппарат Гольджи, большое количество митохондрий и рибосом. На апикальном полюсе, обращенном в полость альвеолы, есть микроворсинки. В период накопления секрета высота клетки увеличивается, после выведения — уменьшается, так как часть ее секрета— жир и окружающая его цитоплазма с прилежащей цитолеммой — отрывается от клетки по
апокринному способу. Другие составные части молока — углеводы, белки —
выводятся из клетки микроапокринным (с отрывом участков микроворсинок)
или мерокринным способом. Не все клетки альвеолы выводят секрет одновременно. Секреция осуществляется асинхронно. Каждая клетка секретирует
все составные части молока.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Вымя пронизано большим количеством сосудов — разветвлением наружной срамной артерии. Каждая альвеола оплетена сетью капилляров.
Альвеолярное молоко удаляется из альвеолотрубки благодаря сокращению
миоэпителиальных отростчатых клеток, которые оплетают концевой отдел
снаружи наподобие корзинки. Из альвеол и альвеолотрубок молоко поступает в выводные протоки — узкие трубочки, образованные однослойным кубическим эпителием. Их диаметр меньше диаметра альвеолотрубок.
Внутридольковые выводные протоки объединяются в междольковые
выводные протоки — молочные каналы, а эти, в свою очередь, сливаются в
короткие широкие молочные ходы. Молочные ходы выстланы двухслойным
эпителием, видны невооруженным глазом и открываются в молочную цистерну. Молочная цистерна имеется в каждой доле вымени. Она представляет
собой полость, размеры и форма которой зависят от характера ветвления выводных протоков. Цистерна доли вымени переходит в сосковую цистерну,
расположенную в соске. В стенках молочных каналов, ходов, цистерн имеются гладкомышечные клетки, количество увеличивается по мере укрупнения протока. Сосок сверху покрыт кожей, но в отличие от кожи вымени в ней
нет (у коровы и свиньи) потовых и сальных желез и волос, много нервных
окончаний. Внутри соска расположена сосковая цистерна, переходящая в
сравнительно короткий (0,5— 2 см) узкий сосковый проток (синус). Кожа
соска в области отверстия соскового протока переходит в слизистую оболочку. Слизистая оболочка соскового протока выстлана многослойным плоским
эпителием, сосковой цистерны — двухслойным эпителием. В собственной
пластинке слизистой оболочки кровеносные сосуды образуют сплетения. В
стенке соска гладкая мускулатура представлена четырьмя слоями. В области
соскового протока сильноразвитый кольцевой слой формирует сфинктер соска.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 86. Гистоструктура
вымени коровы:
А — в период лактации; Б
— сухостоя; В — схема
строения альвеолотрубки;
Г — схема субмикроскопического строения альвеолярного эпителия у сухостойной (б) и лактирующей
(а) коровы:
1— долька; 2— междольковые и 3 —внутридольковые
прослойки соединительной
ткани; 4 — альвеолотрубки; 5 — молочные каналы; 6
— ядро альвеолоцита; 7 —
гранулярная эндо-плазматическая сеть; 8 — аппарат Гольджи; 9 — жировая
вакуоль; 10 — белковые гранулы; 11 — миоэпителиальная клетка; 12 — кровеносный сосуд; 13 — жировые
клетки; 14 — молочные
камни.
У лошади сосок связан с двумя долями вымени, так что в каждом соске
имеется по 2 цистерны и 2 протока. У свиньи в каждом соске по 1—3 цистерны и столько же протоков.
Форма вымени коровы в определенной степени связана с продуктивностью животного. У высокопродуктивных коров и кобыл оно в основном ванно- и чашеобразной формы. Чаще других встречается округлое и
чашеобразное вымя. Коровы и кобылы «с козьим» выменем обычно малопродуктивны.
Изменение внутренней структуры вымени при различных физиологических состояниях животного. Наивысшей функциональной активности
вымя достигает в период лактации. В это время паренхима составляет 70—
80% массы железа. Альвеолотрубки имеют широкий просвет, высокий эпителий. Тесно прилежат друг к другу, разделяясь нежными прослойками рыхлой соединительной ткани. К концу лактации уменьшаются размеры альвеол,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
утолщаются внутри- и междольковые прослойки рыхлой соединительной
ткани. В ней образуются скопления жировых клеток.
В период сухостоя альвеолы спавшиеся, эпителий их низкий, соединительнотканные прослойки широкие с большими жировыми скоплениями. У
старых и малопродуктивных коров лучше развита строма по сравнению с паренхимой, построенная из более плотной соединительной ткани, чем у молодых и продуктивных животных. У хорошо упитанных животных строма увеличивается за счет отложения жира — жировое вымя.
МЯКИШИ
Мякиши — tori — плотные, подушкообразные утолщения кожи на задней поверхности лапы. Из сельскохозяйственных животных только у лошади
и осла есть запястные и заплюсневые мякиши, или каштаны, пястные и
плюсневые мякиши, или шпоры. Каштан расположен на медиальной поверхности дистального конца предплечья и заплюсны, шпора — на задней поверхности I фаланги пальца, прикрыта щеткой. Каштаны и шпоры — безволосые участки кожного покрова. Состоят из эпидермиса, дермы и подкожной клетчатки. Эпидермис толстый, сильно ороговевающий (мягкий кератин).
Пальцевые мякиши (рис. 87) имеются на дистальной фаланге каждого
пальца. Состоят из эпидермиса, дермы, подкожной клетчатки. Эпидермис
сильно ороговевающий, его толстый роговой слой образован трубчатым рогом. В дерме много эластических волокон, сосудов и потовых желез. Большое количество нервных окончаний делает пальцевый мякиш органом осязания. В подкожной основе из жировых скоплений образуется подушка мякиша, выполняющая роль амортизатора. Она особенно развита в задней его
части. У лошади кроме подушки в мякише различают стрелку и хрящи.
Стрелка мякиша состоит из вершины, направленной вперед, и ножек, расходящихся назад и в стороны. Между ними — межножковая борозда. Мякишные хрящи в виде неправильных прямоугольников лежат по бокам копытной
кости и соединены связками с путовой, венечной, копытной и челночной
костями.
РОГОВЫЕ ОБРАЗОВАНИЯ КОЖИ
Роговые образования — это безволосые участки кожи, в которых эпидермис образует чрезвычайно толстый роговой слой. Роговые производные
кожи развиваются на дистальных фалангах пальцев — копытца, копыта —
как защитные приспособления при фалангохождении и на голове — рога —
как орган защиты.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 87. Палец:
А — сагиттальный распил пальца коровы; Б — со снятым роговым башмаком; В — роговой башмак коровы; Г — подошва пальца лошади; Д — гистологическое строение стенки копыта; 1— пястная кость; 2— путовая
кость; 3— венечная кость; 4— копытцевая (копытная) кость; 5 — сесамовидные кости; 6 — копытцевая кайма; 7— копытцевый венчик; в — копытцевая стенка; 9 — подошва; 10 — мякиш; 11 —подушка мякиша; 12 —
стрелка мякиша; 13 — верхушка стрелки; 14 — ножка стрелки; 15 — межножковая борозда; 16 — ветви копытной подошвы; П — пяточные углы; 18
— заворотные части; 19 — заворотные углы; 20 — глазурь; 21— Дерма; 22
— сосудистый слой дермы; 23 — листочковый слой дермы; 24 — эпидермис;
25 — ростковый слой эпидермиса; 26 — листочковый рог; 27 —трубчатый
рог; 28 — роговая трубочка; 29 — промежуточный рог.
Копытце — ungulicula — у рогатого скота и свиньи состоит из каймы,
венчика, стенки и подошвы. Копытцевая кайма у крупного рогатого скота тянется в виде полосы шириной 6—7 мм, у свиньи 3—5 мм по верхнему краю
копытца. При переходе в венчик кайма образует складку, нависающую в виде
козырька. Переход венчика в стенку копытца происходит постепенно, в реwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
зультате он распространяется вниз до половины стенки. Стенка составляет
переднюю, боковые и частично подошвенную поверхности копытца.
Гистологическое строение копытца (Д) подобно строению кожи, но имеет свои особенности. Кайма состоит из эпидермиса, дермы и подкожной основы. Эпидермис каймы имеет все слои, присущие многослойному плоскому
эпителию: ростковый, зернистый, блестящий и роговой. Благодаря наклонному положению роговой слой, отрастая, сползает вниз по копытцу, образуя
его глазурь. Дерма состоит из сосочкового и сетчатого слоев. Сосочки высокие, опущены вниз. Это определяет направление роста рога и его вид. В дерме много сосудов. Подкожная основа развита незначительно, переходит в
надкостницу пальца.
Венчик состоит из эпидермиса, дермы и подкожной основы. В эпидермисе ростковый слой сразу переходит в роговой, очень толстый слой. Сосочки дермы длинные, расположены довольно редко и сильно изогнуты вниз. В
результате эпидермис образует трубчатый рог и растет вниз, покрывая копытцевую стенку. Рог этот очень крепкий. Состоит из роговых трубочек, в
которых клетки плотно прилежат друг к другу. Трубочки спаиваются промежуточным рогом. Трубчатый рог пигментирован. В дерме много сосудов.
Подкожная клетчатка переходит в надкостницу пальца.
Стенка состоит из эпидермиса и дермы. В эпидермисе развиты ростковый и роговой слои. В дерме сосочки слились и образовали листочки в виде
длинных полос, направленных по стенке сверху вниз. Форма листочков дермы определяет рельеф эпидермиса. В результате роговой слой тоже имеет
форму листочков— образуется листочковый рог. Сетчатый слой дермы копытцевой стенки, который называют часто сосудистым из-за обильного развития сосудов, переходит в надкостницу копытце-вой кости.
Подошва состоит из эпидермиса и дермы. Эпидермис образует трубчатый рог. Дерма переходит в надкостницу копытцевой кости.
Роговая капсула (башмак) копытца таким образом имеет сложное
строение. Стенки ее трехслойны: самый поверхностный слой тонкий, быстро
разрушается — глазурь; средний слой самый толстый и прочный — трубчатый рог; внутренний слой не содержит пигмента — листочковый рог. Слои
стенки видны и на подошве при расчистке копытца. Роговая капсула в норме
нарастает с такой же скоростью, как и стирается. Нарушение обмена веществ
ведет к избыточному ороговению. Содержание на твердых полах (железный,
цементный) может привести к слишком быстрому стиранию рогового башмака и воспалению копытца. Нарушение в питании может вызвать снижение
эластичности рогового башмака и замедление роста. При этом на копытцах
появляются борозды, поверхность их становится матовой, шероховатой из-за
ослабления роста и истончения глазури.
Копыто лошади состоит из тех же частей, что и копытце: каймы, венчика, стенки и подошвы. Копытная кайма имеет ширину 5—6 мм, продуцирует
глазурь. Копытный венчик шириной 15 мм имеет развитую дерму, которая
нависает в виде венечного валика над стенкой копыта. Поэтому на роговом
башмаке в области венчика образуется венечный желоб. Сосочки дермы венwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
чика хорошо развиты, направлены вниз. Эпидермис венчика формирует
очень толстый, прочный пигментированный трубчатый рог, который сползает по стенке до подошвы копыта, образуя основную массу рога копыта. В
стенке копыта различают зацепную, латеральную и медиальную боковые и
заворотные части, заворотные углы и подошвенный край. На подошве копыта различают тело и подошвенные ветви (ножки), между которыми вклинивается роговой пальцевый мякиш со стрелкой.
Гистологическое строение копыта подобно гистологическому строению
копытца. Прочность удержания рогового башмака усиливается тем, что дермальные листочки стенки копыта несут на себе вторичные листочки, в результате чего площадь соприкосновения эпидермиса с дермой на стенке одного копыта достигает 1 м2.
Скорость нарастания рогового башмака составляет в среднем 8—9 мм в
месяц. Зимой несколько меньше, чем летом. Роговой башмак трехслойный.
На подошвенном крае стенки на расчищенном копыте хорошо видны слои
башмака. Белая зона (линия), образованная листочковым .рогом, является
линией, указывающей границу, за которую не следует заходить при ковке
лошади, чтобы не повредить живые ткани пальца.
Рог—cornu — сильно ороговевший кожный чехол рогового отростка
лобной кости. В роге различают корень, тело и верхушку. Гистологически он
построен из эпидермиса и дермы. Дерма состоит из сосочкового и сетчатого
слоев. Последний переходит в надкостницу рогового отростка. Высота сосочков дермы увеличивается по направлению к верхушке рогового отростка.
Эпидермис продуцирует прочный трубчатый рог, толщина которого пропорциональна величине дермальных сосочков. Рог растет интенсивно в молодом
возрасте. При недокорме, холоде, беременности рост рога замедляется, что
приводит к образованию колец, заметных у крупного рогатого скота около
корня, а у овец и коз по всему рогу. По роговым кольцам можно определить
примерный возраст коровы, прибавив к числу колец цифру 2.
ВОЗРАСТНЫЕ, ПОЛОВЫЕ, ПОРОДНЫЕ И ДРУГИЕ ИЗМЕНЕНИЯ КОЖИ
У новорожденного животного относительная масса кожи составляет
11—12%, у поросят — до 18%. С трех месяцев у телят интенсивно нарастает
толщина дермы, усложняется ее вязь, увеличивается количество желез на
единицу площади. Однако в течение первого года площадь кожи растет интенсивнее, чем нарастание ее производных. В результате у годовалых телят
количество желез и волосяных фолликулов на единицу поверхности кожи
уменьшается, у половозрелых животных — стабилизируется.
Зимой утолщается сетчатый слой дермы, волосы становятся гуще и
длиннее. У северного аборигенного скота волосы зимой в 2,5 раза длиннее
(25 мм), пуха в 3 раза больше (83%), чем летом. Особенно длинные волосы с
широкой мозговой зоной отрастают на брюхе. У ягнят, родившихся зимой, в
3-месячном возрасте шерсть в 2 раза длиннее (8—10 см), чем у родившихся
летом (4,5 см). Большое значение имеет степень адаптации животного к усwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ловиям существования. У аборигенного калмыцкого скота зимой густота и
длина волос на 25% больше, чем у казахского белоголового. Для скота, приспособленного к жаркому климату, характерно утолщение эпидермиса, очень
сильное развитие потовых и сальных желез, изреженность и глубокое залегание волосяных луковиц, слабое развитие подкожной клетчатки, хорошо
развитый мозговой слой волос, мало пуха по сравнению с животными умеренного климата.
Вместе с тем есть и породные различия в степени развития тех или иных
структур. Так, развитие сосудов, толщина кожи у различных мясных пород
крупного рогатого скота различается на 25—30%- Породные различия закладываются в середине плодного периода и выражаются в разной скорости и
продолжительности развития первичных и вторичных фолликулов. У грубошерстных овец фолликулы залегают глубже, чем у тонкорунных. Значительны и половые различия в строении кожи; у хряков кожа составляет 13%
(31 кг), у свиноматок — 5% (10 кг). Толщина кожи, коллагеновых пучков у
самцов в 2 раза, а эпидермиса на 20% больше, чем у самок.
Полноценное кормление способствует увеличению густоты шерсти овец
за счет увеличения количества вторичных фолликулов, утолщению пучков
коллагеновых волокон, усложнению вязи дермы. Недокорм, особенно в молодом возрасте (до полового созревания), сдерживает рост сетчатого слоя
дермы, приводит к формированию слабого типа вязи, снижает скорость закладки и роста фолликулов. Методы современного животноводства, стимулирующие многоплодие, не ухудшают качество шкурки каракульских ягнят,
однако у многоплодных ягнят затягивается формирование фолликулов. Доместикация (одомашнивание) приводит к утолщению кожи и более рыхлой
вязи коллагеновых волокон, к выравненности шерсти. У диких представителей (архар) кожа более тонкая, плотная, эластичная, с лучше развитым эпидермисом и сетчатым слоем дермы, с более короткими волосами и тонкими
коллагеновыми волокнами.
Часть II. ВИСЦЕРАЛЬНЫЕ СИСТЕМЫ (СПЛАНХНОЛОГИЯ)
Основными свойствами жизни являются обмен веществ между организмом и внешней средой и размножение. Эти функции выполняются системами
органов, имеющими связь с внешней средой и расположенными основной
массой внутри стволовой части в естественных полостях тела. Это — системы органов пищеварения, дыхания, мочевыделения и размножения. По своему местоположению внутри ствола тела они получили название внутренних
органов или внутренностей — лат. viscera, греч. splanchna. В стволовой части
тела млекопитающих имеется три полости: грудная, брюшная и тазовая, где
расположены внутренности, сердце с крупными сосудами, экстрамуральные
нервные ганглии, некоторые железы внутренней секреции и серозные полости.
Грудная полость — cavum thoraci — находится в грудной клетке. Сверху она отграничена грудными позвонками, с боков— ребрами и межреберwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ными мышцами, снизу — грудиной, сзади — диафрагмой. Купол диафрагмы,
обращенный в грудную полость, доходит до уровня 6—7-го ребра. Стенки
грудной полости изнутри выстланы внутригрудной фасцией, снаружи —
мышцами и кожей. В грудной полости находятся: сердце, легкие, трахея, пищевод, тимус, сосуды, лимфатические и нервные узлы, нервы и их сплетения,
серозные оболочки и полости перикардиальная и 2 плевральные.
Брюшная полость — cavum abdominis — находится в поясничнобрюшном и частично в спинно-грудном отделах тела. Сверху отграничена поясничными позвонками с мышцами, спереди— диафрагмой и последними ребрами, с боков и снизу — брюшными мышцами. Сзади переходит в тазовую
полость. Границей между ними считается вход в таз. Стенки брюшной полости изнутри выстланы фасциями, снаружи покрыты кожей. В брюшной полости находятся: пищевод, желудок, кишечник, печень, поджелудочная железа, селезенка, надпочечники, почки, мочеточники, часть органов половой
системы, сосуды, лимфатические и нервные узлы, нервы и их сплетения, серозная оболочка с перитонеальной полостью.
Деление брюшной полости на области. Для лучшей ориентации в расположении органов в брюшной полости ее делят на три отдела: передний,
средний и задний. Каждый отдел, в свою очередь, подразделяется на несколько областей (рис. 88).
Передний брюшной отдел отделяется от среднего воображаемой сегментальной плоскостью, опущенной по касательной и наибольшей выпуклости
последней пары ребер. Он делится на три области. Фронтальная плоскость,
проведенная по реберной дуге, отделяет снизу область мечевидного хряща.
Остальная часть переднего брюшного отдела делится срединной сагиттальной плоскостью на правое и левое подреберья.
Средний брюшной отдел (В) отделяется от заднего воображаемой сегментальной плоскостью, опущенной по касательной к маклокам. Он делится
на четыре области. Две боковые сагиттальные, плоскости, проведенные по
касательной и поперечнореберным отросткам поясничных позвонков, отсекают правую и левую подвздошные области. Оставшаяся средняя часть делится фронтальной плоскостью, проведенной на уровне середины 1-го ребра
на поясничную и пупочную области.
Задний брюшной отдел (Г) граничит с тазовой полостью. Граница проходит по подвздошным и лонным костям, ограничивающим вход в таз. Этот
отдел делится на три области. Боковые сагиттальные плоскости, проведенные
по касательной к поперечнореберным отросткам поясничных позвонков,
продолженные в каудальном направлении, отсекают правую и левую паховые
области. Часть, оставшаяся между ними, является лонной областью.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 88. Деление брюшной полости на области:
А —с боковой стороны; Б — сагиттальный разрез на уровне переднего
брюшного, В — среднего брюшного и Г — заднего брюшного отделов; I —
грудная полость; 1 — контур диафрагмы; II — брюшная полость; III— тазовая полость; 2— левое подреберье; 3— правое подреберье; 4 — реберная
дуга; 5 — область мечевидного отростка; 6 — поясничная, 7 —пупочная, 8
—правая и 9 — левая подвздошные; 10 — маклок; 11 — правая паховая область; 12 — лонная; 13 — левая паховая.
Тазовая полость — cavum pelvis — отграничена сверху крестцом и
первыми хвостовыми позвонками, с боков и снизу — тазовыми костями и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
связками. Каудальная граница тазовой полости проходит у травоядных на
уровне 3-го, у свиньи — 2-го хвостового позвонка. Изнутри выстлана фасциями. Снаружи одета мышцами и кожей. В ней расположены части прямой
кишки, мочевого пузыря, половых органов, сосуды и нервы. Сюда частично
заходит перитонеальная серозная полость.
Серозные полости и их оболочки. Серозная полость — это замкнутое
со всех сторон пространство, вдавленное внутренностями само в себя. Серозные полости появляются на ранних этапах эмбрионального развития, в
период дифференцировки мезодермы из спланхнотома. Полость спланхнотома— целом и есть серозная полость. Париетальный листок спланхнотома выстилает изнутри стенки полостей тела, а висцеральный прилежит к внутренностям,
образуя их серозные оболочки. Сначала
закладывается парная серозная полость —
туловищно-головной целом, париетальные листки которого выстилают полость
тела, а висцеральные примыкают с двух
сторон к кишечной энтодерме. Соединяясь над и под кишечной трубкой, висцеральные листки правой и левой серозных
полостей образуют дорсальную и вентральную брыжейки. В процессе развития
зародыша единый туловищно-головной
целом разделяется на головной, грудной и
туловищный. Головной целом формирует
вокруг сердца перикардиальную полость,
а его стенки образуют листки перикарда.
Рис. 89. Схема расположения Вместе с сердцем он перемещается в
внутренних органов и сероз- грудную полость.
Грудной целом сдавливается растуных телостей в грудной пощими легкими и имеет вид узкой плевлости:
1 — аорта; 2 — пищевод; 3 — ральной полости (рис. 89). Его висцеральтрахея; 4 — легкие; 5— сосу- ный листок, срастаясь с поверхностью
ды; 6— сердце; 7 — средо- легкого, превращается в легочную плевру,
стение; 8 — плевральная се- а париетальный — в реберную и диафрагрозная полость; 9 — внутри- мальную плевру. Часть органов грудной
грудная фасция; 10 — реберная полости — трахея, тимус, аорта — не соплевра; 11 — легочная плевра; прикасаются с серозной оболочкой и ле12—перикардиальная серозная жат в так называемом средостении.
В туловищном целоме вентральная
полость; 13 — сердечная сумка; 14 — париетальный и 15 — брыжейка рассасывается, правая и левая
висцеральный листки перикар- полости объединяются. Образуется брюшинная, или перитонеальная, полость. Ее
да; 16 — сердечная связка.
париетальный листок — брюшина— выwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
стилает стенки брюшной полости и в виде бухтообразных выпячиваний вдается в тазовую полость. Растущие внутренности продавливают серозный мешок на разную глубину и оказываются подвешенными в складках его висцерального листка. Складки висцерального листка, подвешивающие кишки, называются брыжейками, переходящие с одного органа на другой — связками.
Складка между печенью и желудком называется малым сальником, а свешивающаяся с желудка — большим сальником.
Внутренние органы никогда не заходят в полости серозных мешков. Они
лишь сдавливают их, разрастаясь, и прирастают к их висцеральному листку,
который при этом входит в состав серозной оболочки органов.
Строение систем внутренних органов.
Системы органов, входящие в состав внутренностей, несмотря на специфические особенности в строении и функции каждой из них, имеют много
общего: 1) все они представляют собой трубки, сообщающиеся с внешней
средой; 2) стенки трубкообразных органов всех систем внутренних органов
имеют принципиально
сходное строение; 3) в толще стенок трубкообразных органов есть железы; 4) каждая система имеет хотя бы один (парный) паренхиматозный орган;
5) в стенках органов часто встречаются скопления ретикулярной ткани, образующие лимфатические одиночные узелки (солитарные) или в виде скоплений; 6) во внутренних органах проходят кровеносные и лимфатические сосуды; 7) иннервация внутренних органов осуществляется вегетативной нервной
системой. Системы внутренних органов морфологически связаны между собой: пищеварительная и дыхательная системы имеют общий орган — глотку,
а мочевыделительная и половая — общий участок — мочеполовой синус (канал).
Все внутренние органы по характеру строения принято подразделять на
два типа: трубкообразные и паренхиматозные.
Трубкообразные (полые) органы — это пути для прохождения определенного содержимого (воздуха, пищи, мочи и др.). Стенки их состоят из нескольких оболочек: слизистой, подслизистой основы, мышечной и серозной
или адвентиции.
Слизистая оболочка — внутренняя, состоит из эпителия, собственной и
мышечной пластинок. Эпителий выстилает просвет органов, может быть одно- и многослойным. От подлежащего слоя он отделяется базальной мембраной. Собственная пластинка слизистой оболочки образована рыхлой соединительной и ретикулярной тканями. В ней находится большое количество сосудов и нервов, которые образуют подэпителиальные и внутрислизистые сети и сплетения, имеются лимфоидные скопления, залегают железы. Мышечная пластинка слизистой оболочки образована гладкой мышечной тканью,
которая в одних органах располагается отдельными пучками, в других —
имеет вид одно- и даже двухслойного пласта.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Слизистая оболочка выстилает полости трубкообразных органов, соприкасается с их содержимым, и это определяет особенности ее строения. Поверхность ее всегда увлажнена или покрыта слизью (муцином) — продуктом
выделения бокаловидных клеток эпителия или желез, расположенных в собственной пластинке. Слизь обеспечивает скольжение содержимого и предохраняет поверхность органа от повреждения. Слизистая оболочка светлорозового цвета из-за просвечивающих сквозь эпителий кровеносных капилляров.
Она собрана в мелкие складки в результате тонуса мышечной пластинки.
Подслизистая основа образована рыхлой соединительной тканью, в которой сосуды и нервы образуют подслизистые сети и сплетения. В ней залегают концевые отделы пристенных желез. В органах, где подслизистая основа хорошо развита, слизистая оболочка может собираться в крупные складки.
Мышечная оболочка в большинстве органов состоит из гладкой мышечной ткани, формирующей 2 слоя: внутренний (кольцевой) и наружный (продольный). Попеременное сокращение слоев — перистальтика, приводит к
перемешиванию и продвижению содержимого.
Серозная оболочка состоит из собственной пластинки и мезотелия. Собственная пластинка — слой рыхлой соединительной ткани с сосудами и
нервами, образующими подсерозные сплетения. Мезотелий — однослойный
плоский эпителий — производное висцерального листка стенки серозной полости. Серозная оболочка предохраняет органы от срастания. Она увлажнена
серозной жидкостью, продуцируемой ею, и потому скользкая. Это имеет
большое значение для движения органов в процессе их функционирования.
Если орган или его часть не соприкасается с серозными полостями или
находится за пределами полостей тела, то наружной оболочкой будет адвентиция — соединительная ткань, связывающая его с соседними органами
(шейная часть пищевода, аорта и др.).
Паренхиматозные (компактные) органы обычно крупные, округлоовальной или уплощенной формы. Состоят из стромы и паренхимы.
Строма — это соединительнотканный остов органа. В ней различают: 1)
капсулу, покрывающую орган снаружи; 2) междольковые соединительнотканные прослойки (трабекулы), разделяющие орган на дольки; 3) внутридольковые прослойки из рыхлой соединительной ткани. Строма, кроме роли
мягкого каркаса, является местом вхождения и разветвления в органе сосудов
и нервов, местом скопления лимфоидной ткани. В ней же проходят выводные
протоки, если этот орган железа внешней секреции.
Паренхима — определенным образом организованная специфическая
ткань, обычно эпителий, составляющая главную массу долек и выполняющая
функции, свойственные органу (газообмен, выработка секрета и др.). В состав паренхимы входит и система пронизывающих орган ходов в виде каналов: система выводных протоков желез, воздухоносные пути легких, мочевые канальцы и собирательные трубочки почек, семенные канальцы семенwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ника и др. Паренхиматозные органы, как правило, хотя бы частично покрыты
серозной оболочкой, которая тесно срастается с их капсулой.
Глава 15. СИСТЕМА ОРГАНОВ ПИЩЕВАРЕНИЯ (АППАРАТ
ПИЩЕВАРЕНИЯ)
В основе жизненных процессов лежит обмен веществ, который совершается только при постоянном поступлении в организм питательных веществ с
помощью аппарата пищеварения. Он осуществляет передвижение (моторная
функция), механическую и химическую (секреторная функция) переработку,
всасывание расщепленных пищевых веществ и выталкивание непереваренных твердых остатков.
Рис. 90. Топография внутренних органов коровы:
А — слева; Б — справа; 1—трахея; 2 —пищевод; 3 — сердце; 4 — легкое; 5
—диафрагма; 5 — селезенка; 7 — сетка; 8 —рубец; 9 — тощая кишка; 10 —
мочевой пузырь; 11 — матка; 12 — прямая кишка; 13 — влагалище;14 — мочеполовое преддверие; 15 — слепая кишка; 16 — правая почка; 17 — двенадцатиперстная кишка;18 — сычуг; 19 — книжка; 20 — яичник; 21 — ободочная кишка; 22 — печень.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 91. Топография внутренних органов лошади (А—слева, Б —справа)
1 — легкие- 2 — сердце: 3 —аорта; 4 — пищевод; 5 — диафрагма при выдохе; 6 — печень; 7 —желудок; 8 — селезенка; 9 — почка; 10 — петли малой
ободочной кишки; 11 —яичник; 12 -рог матки- 13 - влагалище; 14 - прямая
кишка; 15 — мочеполовое преддверие; 16 — реберная дуга- 17 — петли тощей кишки; 18 — двенадцатиперстная кишка; 19 дорсальное и 20 - вентральное левые положения большой ободочной кишки; 21 — мочевой пузырь;
22 -слепая кишка; 23 — дорсальное и 24 - вентральное правые положения
большое ободочной кишки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 92. Топография внутренних органов свиньи (слева):
1 — пищевод; 2— трахея; 3— сердце; 4— легкие; 5 — диафрагма; 6— печень;
7 — желудок; 8 — селезенка; 9 — почка; 10 — ободочная кишка; 11—слепая
кишка; 12 — рог матки; 13 — мочевой пузырь; 14 — прямая кишка; 15 — мочеполовое преддверие; 16 — тощая кишка; 17 — реберная дуга.
Пищеварительную систему млекопитающих делят на четыре отдела: головную, переднюю, среднюю и заднюю кишки (рис. 90, 91, 92). Головная
кишка, или ротоглотка, располагается в лицевом отделе головы. В нее входят
рот с ротовой полостью и глотка. С ее помощью осуществляется захват пищи
и воды из внешней среды, предварительная механическая и начальная химическая обработка, формирование пищевого кома и эвакуация его в переднюю кишку. Передняя кишка, или пищеводо-желудочный отдел, начинается
в шее, тянется через всю грудную полость и заканчивается в брюшной полости. Состоит из пищевода и желудка. В нем происходят начальные этапы
переваривания белков и всасывание воды и некоторых растворимых солей.
Средняя кишка включает в себя тонкий отдел кишечника (двенадцатиперстную, тощую и подвздошную кишки) и застенные железы (печень и
поджелудочную железу, связанные протоками с двенадцатиперстной кишкой). Все они лежат в брюшной полости. В средней кишке идут наиболее интенсивные процессы переваривания и всасывания всех питательных веществ.
Задняя кишка, или толстый отдел кишечника, состоит из слепой, ободочной и
прямой кишок. Лежат они в брюшной полости, а прямая кишка заходит в тазовую полость и открывается наружу заднепроходным отверстием — анусом.
В задней кишке продолжаются процессы всасывания, а у некоторых животных (лошадь) и расщепления веществ, формирование и выбрасывание каловых масс.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
КРАТКИЕ СВЕДЕНИЯ О РАЗВИТИИ СИСТЕМЫ ОРГАНОВ ПИЩЕВАРЕНИЯ
Филогенез. Пищеварительная система — одна из самых древних систем
организма, так как осуществляет одно из основных свойств живого — обмен
веществ с окружающей средой. Одноклеточные организмы захватывают пищевые частицы либо всей поверхностью, после чего вокруг частиц образуется пищеварительная вакуоль, подобная фаголизосоме (амеба), либо пища
попадает в ротовую вакуоль, где также происходит переваривание (инфузории). В обоих случаях пищеварение внутриклеточное. У простейших многоклеточных животных — кишечнополостных (гидра) пищеварение принципиально не отличается от пищеварения одноклеточных. Мешкообразная полость их тела, образованная клетками энтодермы, является в то же время кишечной полостью. Пищевые вещества с током жидкости при движении щупалец попадают через первичный рот в кишечную полость, а при сокращении
тела через него же выбрасываются наружу. Пищеварение внутриклеточное.
У некоторых представителей типа кишечнополостных (ктенофоры) и у
ресничных червей имеется приспособление для принудительного продвижения воды с взвешенной в ней пищей. На переднем конце их тела к энтодермальной кишке добавляется глотка эктодермального происхождения в виде
трубки с мышечными стенками, способная впячиваться внутрь кишки и выворачиваться наружу, создавая этим двусторонний ток жидкости. Дальнейшие усложнения пищеварительной системы происходили, видимо, в определенной последовательности. На заднем конце тела образовалось выходное
отверстие (что наблюдается в типе круглых червей) и кишка приобретает вид
трубки, идущей вдоль всего тела. Большая ее часть образована энтодермой, а
краниальный и каудальный концы — эктодермой. С появлением вторичной
полости тела — целома (у кольчатых червей) кишечная трубка обособляется
от кожно-мышечного мешка. Вокруг нее из элементов мезенхимы формируется собственная мышечная оболочка. Кишка приобретает возможность самостоятельно занимать различное положение в полости тела, образовывать
изгибы, петли независимо от формы тела животного. В единой пищеварительной трубке при этом различают переднюю (эктодермальную), среднюю
(энтодермальную) и заднюю (эктодермальную) кишки.
Следующее чрезвычайно важное усложнение в строении пищеварительной системы мы находим у кишечнодышащих: переход от внутриклеточного
к внеклеточному пищеварению. Связано это со специализацией клеток эпителия, выстилающего кишку. Пищеварительную систему кишечнодышащих
можно разделить на переднюю, среднюю и заднюю кишки. В состав передней кишки входят рот, глотка с жаберными щелями и продольный желоб с
эпителием, клетки которого дифференцируются на мерцательные и железистые, вырабатывающие слизь. Слизь с приставшими частицами пищи продвигается в среднюю кишку, в стенке которой имеются участки в виде кармашков, называемых печеночными выростами. Их клетки вырабатывают
пищеварительные ферменты. Большая часть заглоченной воды не попадает в
кишку, а удаляется через жаберные щели. Пища же с током слизи попадает в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
среднюю кишку, где и подвергается внеклеточному перевариванию. Переваренные продукты всасываются стенкой кишки. Непереваренные остатки удаляются через анальное отверстие, которым заканчивается задняя кишка на
каудальном конце тела.
У примитивных хордовых строение пищеварительной системы очень
похоже на ее строение у высших червей. У ланцетника (рис. 93—А) кишечная трубка начинается ротовым отверстием, отграниченным от глотки складкой слизистой оболочки. Вода с пищей загоняется в рот движением мерцательных усиков и далее в глотку. Глотка — это широкая трубка с многочисленными жаберными щелями, расположенными по бокам. На дне глотки
имеется желобок, выстланный мерцательным и железистым эпителием (выделяет слизь). Вода со взвешенными частичками пищи прогоняется через
глотку и увлекается наружу движением ресничек, покрывающих стенки жаберных щелей.
Частицы же пищи склеиваются слизью, выделяемой эпителием и движением ресничек мерцательного эпителия желобка, и продвигаются в среднюю
кишку.
Средняя кишка ланцетника короткая, прямая, имеет мешкообразный печеночный вырост, соединенный с кишкой протоком. Клетки печеночного выроста вырабатывают пищеварительный сок, который по протоку поступает в
кишку. Здесь происходит внеклеточное пищеварение. Эпителий кишки осуществляет всасывание переваренных продуктов, а непереваренные остатки
продвигаются к анальному отверстию, снабженному кольцевой мышцей—
сфинктером. Расположено оно не на конце тела, а около хвостового отдела.
Хвост таким образом освобождается от внутренностей и становится основным органом движения.
Дальнейшее усложнение пищеварительной системы шло за счет прогрессивного развития всех ее участков. Это привело к увеличению длины кишечной трубки (она стала в 5—50 раз длиннее тела); формированию различных видоспецифических изгибов, петель и положений; к утолщениям к
расширениям определенных участков (желудка, ободочной кишки у некоторых животных); к формированию пристенных и застенных пищеварительных желез; к совершенствованию всасывающего аппарата — формированию
ворсинок.
Особенно большие преобразования происходят в передней кишке, которая с формированием головы у рыб делится на головную и собственно переднюю (Б, В).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 93. Схема расположения внутренних органов у животных разных
классов типа хордовых:
А — ланцетник; Б —рыба; В— млекопитающее; 1—ротоглотка; 2 — жаберные щели; 3 — эндостиль (желобок); 4 — печеночный вырост; 5 —кишка;
6 — анальное отверстие; 7— половые железы; 8 — пищевод; 9— плавательный пузырь; 10— печень; 11 — желудок; 12 — поджелудочная железа; 13 —
средняя кишка; 14 — задняя кишка; 15 — почка; 16 — мочеточник; 17 —
щитовидная железа; 18 — селезенка; 19 — яичник; 20 — яйцевод; 21 — клоака; 22 — носовая полость; 23 — глотка; 24 — гортань; 25 — трахея; 26 —
тимус; 27 — легкие; 28 — тонкий кишечник; 29 — слепая кишка; 30 — ободочная кишка; 31 — надпочечник; 32 — прямая кишка; 33 — мочевой пузырь;
34 — мочеполовой синус; 35 —матка; 36 — влагалище.
Головная кишка, или ротоглотка, состоит из ротовой полости и глотки и
первоначально тесно связана с дыхательной системой. Стенки ротовой полости образованы висцеральным скелетом и связанными с ним мышцами,
сверху покрыты кожей. Вход в ротовую полость у рыб, амфибий и рептилий
— рот обрамлен губами — кожными складками, плотно сидящими на челюстях. Только у млекопитающих губы — это кожно-мышечные складки, в основе которых залегает круговая мышца рта. Развиваются они как приспособление для сосания молока. Такую же роль выполняют щечные мышцы. Язык
у рыб не имеет собственной мускулатуры, являясь складкой слизистой оболочки дна ротовой полости. У земноводных, рептилий и птиц подвижность
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
языка обеспечивается в основном мышцами подъязычного аппарата. Лишь у
млекопитающих язык с мощной собственной мускулатурой, как орган, приспособленный для сосания молока.
Рис. 94. Схема усложнения зубов в филогенезе:
1— простой конический зуб; 2, 3, 4 — трехзубчатые зубы; 5 — трехбугорчатый; 6, 7 — четырехбугорчатый; 8 — складчатый; 9 — лунчатый.
Зубы в типе хордовых появляются у рыб. Это производное плакоидной
чешуи, которая состоит из дентинной пластинки с шипом, покрытым эмалевым чехликом. Такие зубы имеют форму конусов, вершиной обращены в
сторону глотки, располагаются в различных местах ротовой полости, в течение жизни сменяются много раз. Простые однотипные конические мелкие
многосменные зубы типичны и для амфибий, и для рептилий (рис. 94). Они
расположены на челюстях, служат для удержания добычи. И только млекопитающие имеют разнотипные крупные несменяемые или двухсменные зубы. Связано это с тем, что только млекопитающие размельчают и пережевывают пищу. Каждый зуб расположен в своей альвеоле — углублении челюстных или резцовых костей. Для удержания добычи служат клыки, конические по форме и обычно крупные по размерам, особенно у хищных. Для
разрезания, отрывания, скобления пищи используются резцы, наиболее развитые у грызунов. Коренные зубы служат для измельчения, перетирания пищи. Они имеют наиболее сложное строение.
Процесс усложнения простого конического зуба, как считают, мог идти
двумя путями: либо срастанием нескольких конических зубов, либо постепенным отрастанием дополнительных вершин у конического зуба под влиянием изменившейся функции. В обоих случаях могли появиться трех-, четырех-, пятивершинные зубы. При этом увеличивается и количество корней.
В зависимости от остроты вершин такие зубы могут быть остро- или тупобугорчатыми, что характерно для всеядных. Если вершины зуба соединены
между собой острым краем, он называется режущим (у хищных). При формировании коренных зубов травоядных бугорки сильно вытягиваются и, соединяясь между собой, образуют рельеф зуба, характерный для каждого рода
животных. У лошади, например, складчатые зубы, для рогатого скота — лунчатые.
Железы полости рта у хордовых появляются с выходом последних на
сушу. У амфибий это мелкие пристенные железы, выделяющие слизь. У ряда
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рептилий наряду с железами, вырабатывающими слизистый секрет, имеются
специальные ядовитые железы. У птиц слюнные железы многочисленные, но
мелкие. Секрет их слизистой не содержит ферментов. У мле-. копитающих
наряду с пристенными образуются и крупные застенные слюнные железы, в
том числе и такие, которые вырабатывают серозный секрет, содержащий
ферменты (околоушная слюнная железа).
Нёбо, разделяющее ротовую и носовую полости, появляется у животных
только с выходом на сушу. У первичноводных животных носовой полости
нет, так как аппарат дыхания (жабры) у них тесно связан с ротоглоткой. У
земноводных носовая полость почти не отделена от ротовой полости.
У рептилий появляется твердое нёбо, в значительной степени разобщающее
дыхательные и пищеварительные пути. В наибольшей степени они разобщены у млекопитающих, у которых наряду с твердым развивается и мягкое нёбо.
Глотка с выходом животных на сушу значительно видоизменяется. Она
перестает быть местом расположения органов дыхания — жабр. С развитием
нёбно-глоточной дуги и нёбной занавески (у крокодилов) глотка четко отделяется от ротовой полости. Однако у млекопитающих она остается местом,
где анатомически связаны и перекрещиваются дыхательный и пищеварительный пути.
Передняя и средняя кишки в процессе филогенеза подвергались меньшим изменениям по сравнению с головной кишкой. Изменения эти выражаются в основном в удлинении пищевода и кишечника; в увеличении диаметра желудка, изменении его формы и положения в теле; в развитии и дифференцировке железистого и всасывающего аппарата. У рыб и амфибий части
кишечной трубки нечетко отделены друг от друга. Веретенообразной формы
желудок лежит вдоль тела, содержит однотипные железы. Печень является
самостоятельным компактным органом, связанным с начальным участком
кишечника. Поджелудочная железа в виде органа вычленяется у некоторых
рыб и у всех амфибий. У рептилий органы пищеварительной системы четко
отграничены друг от друга, имеют большое количество пристенных желез и
аппарат всасывания — ворсинки. У млекопитающих желудок мешкообразной формы, лежит поперек тела, часто в нем образуются выпячивания, а у
некоторых и дополнительные камеры (многокамерный желудок жвачных).
Железы желудка дифференцируются на кардиальные, донные (фундальные,
собственные) и пилорические. Кишечник делится на тонкий и толстый. Тонкий кишечник включает двенадцатиперстную, тощую и подвздошную кишки,
без резких границ, переходящие друг в друга. В стенке двенадцатиперстной
кишки развиваются собственные железы. В нее открываются протоки печени
и поджелудочной железы.
Задняя кишка в филогенезе выделилась позднее других участков кишечной трубки. У рыб, амфибий и рептилий она не дифференцирована на отдельные кишки. Представляет собой по существу место формирования каловых масс — прямую кишку, которая открывается в клоаку. Лишь у рептилий
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
отдельных видов имеется слабый дорсальный выступ — прототип слепой
кишки. У птиц слепые кишки обычно парные, развиты в разной степени. И
только у млекопитающих задняя кишка преобразуется в толстый кишечник, в
состав которого входят: слепая, ободочная и прямая кишки Клоака есть у
всех хордовых, включая однопроходных млекопитающих. Это конечный, ампулообразно расширенный участок кишечной трубки, куда открываются
трубки мочевыделительной и половой систем. Только у сумчатых и плацентарных млекопитающих происходит разделение пищеварительной и мочеполовой систем.
Онтогенез (рис. 95). Пищеварительная трубка в онтогенезе закладывается очень рано — в период дифференцировки зародышевых листков. Энтодерма зародыша формирует первичную кишку, сообщающуюся в своей
средней части с желточным мешком. У зародыша теленка это происходит на
17-й день, у овцы — на 14-й, у свиньи — на 12—13-й день развития. Краниальный и каудальный концы первичной кишки заканчиваются слепо —
бухтами. Связь среднего участка кишки с желточным мешком сначала широкая, к концу зародышевого периода сужается до узкого желточного протока.
От задней бухты отрастает аллантоис. Между ним и кишкой сохраняется
связь с помощью протока аллантоиса.
Стенка кишки приобретает сложное строение. Изнутри она выстлана энтодермой, снаружи покрыта висцеральным листком спланхнотома мезодермы. Между ними врастает мезенхима.
Рис. 95. Положение кишечной трубки у зародыша (А) и дальнейшая стадия дифференцировки клоаки (Б):
1 — ротовая ямка: 2 —язык; 3 — глотка; 4 — пищевод; 5 —зачаток
трахеи; 6 — зачаток легких; 7 —селезенка: 8 — желудок; 9 — зачаток печени; 10 — зачаток поджелудочной железы; 11 — нисходящее колено первичной кишки; 12 — отрезок аорты: 13 — краниальная брыжеечная артерия;
14 — зачаток слепой кишки; 15 — восходящее колено первичной кишки; 16 —
клоака; 17 — хвостовая кишка; 18 — анальная ямка; 19 — проток аллантоиса; 20 — желточный проток; 21 — яичник; 22 — яйцевод; 23 — почка;
24 — мочеточник; 25 — прямая кишка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Зародыш заметно растет в длину, на его переднем и заднем концах кожная эктодерма впячивается внутрь, навстречу энтодерме образуются ротовая
и анальная ямки. Они углубляются, так что стенка между кишкой и ямками
принимает вид перепонки. Ротовая ямка является зачатком ротовой полости.
Она обрамлена валиками, из которых образуются челюсти, губы и язык. На
головном конце глоточная перепонка отделяет эктодермальный зачаток ротовой полости от краниальной кишечной бухты — будущей глотки. Вскоре
перепонка прорывается, на ее месте формируется нёбно-глоточная дуга и
мягкое нёбо. В заднюю кишечную бухту кроме аллантоиса открывается проток промежуточной почки (вольфов проток) — образуется клоака. В нее прорывается анальная ямка.
На 18—19-й день зародышевого развития у овцы, на 22—23-й у крупного рогатого скота в вентральной стенке кишки закладывается печень. Соответственно на 22-й и 28-й день — желудок, на 24-й и 30-й день — поджелудочная железа. Желудок жвачных сначала имеет вид веретеновидного
расширения, но уже к концу зародышевого периода в нем намечаются все 4
камеры. В это же время в кишечнике образуется первый изгиб. От желудка
он направляется каудовентрально — нисходящее колено, а после желточного
протока — поднимается каудодорсально — восходящее колено. На восходящем колене образуется выступ слепой кишки. В конце зародышевого — начале предплодного периода в кишке можно различить зачатки ротоглотки,
пищевода, желудка, кишечника, печени и поджелудочной железы.
На протяжении предплодного и плодного периодов продолжается анатомо-гистологическое формирование, рост и изменение расположения органов пищеварения. Они закладываются в разные сроки и растут неравномерно. В желудке жвачных в период закладки и до начала плодного периода
наиболее крупной камерой является рубец. Затем темпы роста сычуга нарастают, и к рождению он оказывается самой крупной камерой. После рождения вновь увеличивается скорость роста преджелудков. С переходом животных с молочной пищи на растительную рубец вновь становится самой
крупной камерой.
Кишечник в предплодный период уже можно разделить на тонкий и толстый. Границей является закладка слепой кишки. Тонкий кишечник образует
целую гирлянду петель у предплодов свиньи к 30-му дню, овцы — к 40-му,
.коровы — к 50—60-му дню. Толстый кишечник дифференцируется позднее:
лабиринт ободочной кишки формируется у овцы к 40—50-му дню, у коров —
к 60-му, у свиней — к 45-му дню. Клоака делится продольной перегородкой
на 2 отдела: верхний — прямую кишку и нижний — мочеполовой синус. Развивающаяся промежность отделяет друг от друга анальное и мочеполовое
отверстие.
Печень — в зародышевый и предплодный периоды растет настолько интенсивно, что является самым крупным органом и занимает почти всю
брюшную полость. С переходом функции кроветворения к селезенке и костному мозгу в плодном периоде печень начинает отставать от общего роста
плода, уступая место интенсивнее растущим в это время желудку и киwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
шечнику. Поджелудочная железа растет равномерно на протяжении всего
эмбриогенеза.
В процессе развития желудок делает два поворота. У жвачных первый
поворот происходит в горизонтальной плоскости и заканчивается к началу
плодного периода. Второй поворот — вентрально завершается в поздний
плодный период. При поворотах желудка камеры меняют свое положение в
разной степени и после второго поворота занимают окончательное положение. Однокамерный желудок свиньи и лошади сначала обращен вверх большой кривизной. В предплодный период происходит продольный поворот желудка влево на 180о так, что дорсально расположенной оказывается малая, а
вентрально — большая кривизна. В плодный период желудок поворачивается
вокруг вертикальной оси влево так, что вход в желудок оказывается слева, а
выход — справа.
Одновременно с желудком меняют свое положение и расположенные
близко от него, связанные с ним серозной оболочкой печень, поджелудочная
железа и селезенка. Печень, расположенная сначала в вентральной брыжейке,
перемещается вправо, вперед и вверх, вследствие того что брыжейка оттягивается в этом направлении развивающейся диафрагмой. Дорсальная
брыжейка, разрастаясь, образует слева слепой мешок — большой сальник.
Сюда же перемещаются селезенка и поджелудочная железа. При втором повороте желудка печень располагается впереди желудка и связана с ним
складкой серозной оболочки — малым сальником. Большой сальник оказывается сзади желудка и, сильно вытягиваясь, подстилает кишки.
Одновременно со вторым поворотом желудка делает поворот и кишечник на 180° вправо вокруг краниальной брыжеечной артерии, которая проходит в брыжейке к кишечнику. При этом двенадцатиперстная кишка ложится в правой части брюшной полости, а поджелудочная железа располагается в ее петле. В связи с удлинением кишечника и формированием петель
продолжается его перемещение в том же направлении еще на 180°. В результате слепая кишка ложится сзади справа, верхушкой назад. Петли кишечника принимают определенное положение, характерное для каждого вида
животного.
Гистогенез органов пищеварения. Эктодермальная выстилка кишечной
трубки преобразуется в многослойный эпителий ротовой полости и конечного участка прямой кишки, а энтодермальная выстилка — в однослойный эпителий слизистой оболочки желудочно-кишечного тракта и желез. Мезенхима
дает начало соединительной и гладкой мышечной тканям органов пищеварения и их сосудам. Висцеральный листок спланхнотома мезодермы становится серозной оболочкой органов. Позднее всех формируется мышечная пластинка слизистой оболочки и слизистая отделяется от подслизистой основы.
В плодный период закладываются и развиваются железы желудка. Дифференцировка слизистой оболочки кишечника начинается в конце предплодного — начале плодного периода. Совершается постепенно, начинаясь в двенадцатиперстной кишке и распространяясь отсюда в краниальном и каудальной направлениях. Закладываются пристенные железы и ворсинки перwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
воначально по всему кишечнику, затем в толстом кишечнике ворсинки редуцируются.
Рис. 96. Последовательные стадии (А, Б, В, Г)
развития зуба:
1 — эпителий десны; 2—
зубная пластинка; 3 —
зубной орган; 4— зубной
сосочек; 5 — зубной мешочек; 6 — кутикула эмали; 7 — многоотростчатые клетки; 8 — энамелобласты; 9 — одонтобласты; 10 — эмаль; 11—
дентин; 12 — дентинные
трубочки; 13 — предентин; 14 — пульпа зуба; 15
— кровеносный сосуд.
В поджелудочной железе первыми к концу предплодного периода дифференцируются островки и островковые клетки. В начале плодного периода
появляются первые ацинусы. Дольчатость железы хорошо выражена в поздний плодный период. В печени в предплодный период формируются печеночные пластинки, а в начале плодного периода — дольки печени.
Зубы закладываются в предплодный период (рис. 96). От многослойного
плоского эпителия десны в подлежащую мезенхиму впячивается эпителиальный тяж — зубная пластинка. На ее передней поверхности (обращенной в сторону губ и щек) образуются выпячивания в виде колпачков — зубные (эмалевые) органы. Внутрь колпачка врастает мезенхима, образуя зубной
сосочек. Зубных органов закладывается столько, сколько будет зубов. Первыми у крупного рогатого скота закладываются премоляры, вслед за ними —
резцы, потом — моляры. Вокруг зачатка каждого зуба мезенхима формирует
зубной мешочек. Зубной (эмалевый) орган состоит из эпителиальных клеток,
которые на наружной и внутренней поверхностях его лежат однослойным
пластом, а в середине — образуют многоотростчатый синцитий. Внутренние
клетки дифференцируются в клетки-эмалеобразователи — энамелобласты
(амелобласты). Многоотростчатые клетки редуцируются, а клетки наружного
слоя превращаются в кутикулу эмали.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Мезенхима зубного сосочка дает начало дентину и пульпе зуба. В участках зубного сосочка, прилежащих к зубному органу, мезенхимные клетки
превращаются в клетки-дентинообразователи — одонтобласты. Мезенхима
более глубоких участков зубного сосочка дифференцируется в соединительную ткань, богатую кровеносными сосудами. В нее врастают нервы. Образуется пульпа зуба.
Энамелобласты откладывают эмаль в сторону дентина. В процессе развития столбик эмали над каждым энамелобластом растет, а сама клетка укорачивается и несколько отодвигается. Продуцированная эмаль минерализуется и принимает вид длинных, слегка S-образно изогнутых эмалевых
призмочек, спаянных промежуточным веществом. К моменту прорезывания
зубов энамелобласты редуцируются и эмаль оказывается покрытой кутикулой эмали. Во время прорезывания кутикула разрушается и эмаль становится
поверхностным слоем зуба в области коронки.
Одонтобласты откладывают в сторону эмали дентин — разновидность
костной ткани. Сами при этом отступают в сторону пульпы, оставляя в слое
дентина свои отростки. В процессе обызвествления дентина вокруг отростков
образуются дентинные трубочки. По системе этих трубочек осуществляется
питание твердых тканей зуба. Дентин — основная ткань зуба, распространенная и в корне, и в коронке. Мезенхима зубного сосочка дифференцируется в разновидность грубоволокнистой костной ткани — цемент,
покрывающий снаружи корень, и периодонт — плотную соединительную
ткань, связывающую зуб со стенкой альвеолы.
Смена зубов. Зачатки постоянных зубов, кроме моляров, закладываются
позади каждого молочного зуба и лежат сначала в общей альвеоле. Затем
между ними образуется перегородка. В период смены зубов остеокласты разрушают перегородку и корень молочного зуба. Постоянный зуб при этом начинает усиленно расти и выталкивает молочный зуб. Постоянные моляры
развиваются без молочных предшественников. Закладка постоянных зубов
животных разных видов происходит в разное время. У крупного рогатого
скота все постоянные зубы закладываются после рождения, резцы — начиная
с 6-, а коренные — с 9-месячного возраста. Прорезываются постоянные зубы
в той же последовательности: сначала резцы (в 25 месяцев — зацепы, в 44
месяца — окрайки), потом — премоляры и последними — моляры. Стираются зубы приблизительно на 1,5—2 мм в год. Резцы у рогатого скота не
выдвигаются. Стирание приводит к их укорочению и разъединению, так как
в области шейки они уже, поэтому при стирании создается впечатление расхождения зубов. Коренные зубы рогатого скота выдвигаются по мере стирания. При этом ослабевает их укрепление в челюсти, зубы расшатываются и у
старых животных могут выпадать.
Структура органов пищеварения у плодов свидетельствует о возможности в них процессов секреции, экскреции, всасывания. О том же говорит наличие содержимого в желудочно-кишечном тракте плодов и новорожденных
животных. Однако и у зрелорожденных животных, какими являются копытные, органы пищеварения к рождению не достигают морфофизиологической
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
зрелости. В первые месяцы и даже годы происходит усложнение структуры
органов, совершенствование их функции.
Возрастные изменения органов пищеварения. У крупного рогатого скота
до 3—5 лет происходит рост и усложнение структуры органов. В ротоглотке
утолщается соединительнотканный и эпителиальный слои слизистой оболочки, формируются крипты миндалин, увеличиваются размеры и густота расположения сосочков языка и пристенных желез. Происходит смена зубов.
Интенсивно увеличивается емкость желудка и устанавливается окончательное соотношение его камер. Рубец составляет 59% от массы желудка, книжка
— 22, сычуг—11, сетка — 8%. Кишечник в постнатальный период растет гораздо медленнее, чем до рождения. В плодный период его длина увеличивается в 30 раз, а с рождения до 18 месяцев — в 3 раза. У новорожденных кишечник в 16—17 раз длинее тела животного, у взрослого—в 25—26 раз. Толстый кишечник после рождения растет интенсивнее тонкого: у новорожденного соотношение длины тонкого и толстого кишечника составляет 8 : 1, в
4,5 года — 4 : 1.
У старых животных в возрасте 10—12—14 лет частично атрофируются
железы рта и фолликулы миндалин, замещаясь соединительной тканью, Стираются сосочки языка, а также зубы, что приводит к укорочению резцов и
расшатыванию коренных. Изменяется соотношение камер желудка: увеличивается объем сычуга и книжки, рубца и сетки уменьшается. Снижается ферментативная активность поджелудочной железы и печени.
РОТОГЛОТКА
Ротовая полость — cavum oris (рис. 97)—является начальным отделом
пищеварительной трубки. Стенки ее образованы костями лицевого (висцерального) черепа и мышцами, связанными с ним. К органам ротовой полости
относятся: губы, щеки, зубы, десны, язык, твердое и мягкое нёбо, слюнные
железы, миндалины. Функции ротовой полости: захват пищи, ее предварительная механическая обработка (пережевывание), формирование пищевого кома, его увлажнение и ослизнение, начальные этапы расщепления углеводов, определение вкуса пищи. Кроме того, ротовая полость участвует в
осуществлении защитной функции, в издавании звуков.
Костный остов ротовой полости образован верхнечелюстной, резцовой и
нижнечелюстной костями. Вход в ротовую полость — ротовая щель— находится между губами. Выход в глотку — зев — между мягким нёбом и корнем
языка. Часть ротовой полости, отграниченная снаружи губами и щеками, а
изнутри зубами и деснами, называется преддверием ротовой полости.
Губы — labia — кожно-мышечные складки, ограничивающие ротовую
щель. Место соединения верхней и нижней губ называется спайкой. Спайки
обрамляют углы рта. Снаружи губы покрыты кожей с волосами, изнутри —
слизистой оболочкой с многослойным плоским эпителием. В основе губ залегает круговая мышца рта, в которую вплетаются лицевые мышцы. Это позволяет не только сжимать ротовое отверстие, но и двигать губами. В основе
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
слизистой оболочки губ, особенно в углах рта, залегают пристенные губные
железы, открывающиеся своими протоками в преддверие ротовой полости.
Губы богаты чувствительными нервными окончаниями.
У крупного рогатого скота губы безволосые, малоподвижные. Верхняя
губа сливается с областью ноздрей, образуя носогубное зеркальце. Нижняя
губа короткая толстая прикрыта верхней. Подбородок раздвоен. У овцы губы
очень подвижны, покрыты волосами. Верхняя губа несколько раздвоена. У
свиньи губы малоподвижные, узкие, с небольшим количеством волос. Верхняя губа переходит в хоботок, нижняя заострена. У лошади губы тонкие,
длинные, очень подвижные и чувствительные, поросшие вибриссами и нежными короткими волосами.
Рис. 97. Сагиттальный разрез головы (полусхема):
1 — губы; 2— круговая мышца рта; 3— преддверие ротовой полости; 4 —
резцы; 5— нижнечелюстная кость; 6— голодная бородавка; 7 — проток
подчелюстной слюнной железы; 8— подъязычная слюнная железа; 9— уздечка языка; 10 — язык;11 — сосочки языка; 12 — твердое нёбо; 13 — подушка языка; 14 — мягкое нёбо; 15 — язычная миндалина; 16 — зев; 17 —
надгортанник; 18 — глотка; 19 — гортань; 20 — глоточная миндалина; 21
— пищевод; 22 — трахея; 23 — преддверие носовой полости; 24 — носовая
полость; 25 — дорсальный носовой ход; 26 — дорсальная носовая раковина;
27 — средний носовой ход; 28 — нейтральная носовая раковина; 29 — вентральный носовой ход; 30 — хоана; 31 — лабиринт решетчатой кости.
Щеки — bассае — кожно-мышечные пластинки, образующие боковые
стенки ротовой полости. Снаружи они покрыты кожей с волосами, среди которых встречаются чувствительные синуозные волосы. Под кожей располоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
жена щечная мышца. Изнутри щеки выстланы слизистой оболочкой с многослойным плоским эпителием. У жвачных слизистая оболочка образует многочисленные выросты — ороговевшие щечные сосочки длиной до 1 см, предохраняющие слизистую от травм и способствующие удержанию корма. В
собственной пластинке слизистой и между мышечными пучками залегают
пакеты пристенных щечных желез. Их протоки открываются в преддверие
ротовой полости. Слизистая оболочка щеки переходит в слизистую десны. У
свиньи и лошади слизистая оболочка щек гладкая.
Десны — gingivae — слизистая оболочка, покрывающая челюсти вокруг
основания видимой части коронки (у короткокоронковых — в области шейки) зубов. Десны бедны чувствительными нервными окончаниями, но богаты
сосудами, имеют розовый цвет. У жвачных на верхней десне в области тела
резцовых костей вместо отсутствующих резцов образуется толстая пластинка
ороговевшего эпителия — зубная пластинка, что позволяет животным захватывать большие пучки травы.
Зубы — dentes — расположены на верхней и нижней частях в виде зубных аркад или дуг. Каждый зуб находится в отдельной луночке — альвеоле
кости. Количество, размеры и форма зубов у млекопитающих разных семейств резко различаются. Связано это с образом жизни животного и характером корма. Так, например, у одного из видов дельфинов насчитывается
свыше 230 зубов, у рогатого скота — 32, у свиньи — 44, у лошади — 36—40
зубов. Зубы млекопитающих двухсменны, разнотипны. Среди них различают
молочные и постоянные резцы, клыки и коренные.
Резцы — dentes incisivi (I) имеют уплощенную спереди назад форму и
располагаются в альвеолах резцовой и нижнечелюстной костей обычно в количестве 6 (по 3 на каждой половине челюсти). У жвачных 4 пары резцов и
только на нижней челюсти. Два самых передних резца называются зацепы,
лежащие рядом с ними — средние, а крайние — окрайки.
Клыки — dentes canini (С)—конусовидной формы, сильно развиты.
Кроме разрывания или выкапывания пищи являются орудием защиты и нападения. На каждой челюсти может быть 2 клыка, которые лежат по одному
сбоку от окрайков на некотором расстоянии от них.
Коренные зубы делятся на премоляры, или малые коренные, и моляры,
или большие коренные. Широкие, многобугорчатые. Приспособлены для
размалывания, перетирания пищи. Премоляры— dentes praemolares (Р) расположены на боковых сторонах зубных аркад. Между ними и резцами обычно имеется беззубое пространство. На каждой челюсти по 6 премоляров (по 3
с каждой стороны). Иногда впереди первого прехмоляра имеется дополнительный — волчий зуб. Моляры — dentes molares (М) — самые крупные зубы, не имеют молочных предшественников. Расположены в аборальной части челюстей, по 3 с каждой стороны. Иногда позади последнего моляра вырастает зуб мудрости.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 98. Зубы:
I—резец 10-днсвного теленка; II — резец 7-летней
коровы (а — на разрезе; б
—внешний вид); III — 1-й
моляр коровы; IV—моляр
лошади; V —моляр свиньи; VI — клык свиньи; 1
— коронка; 2 — шейка;
3— корень; 4 — эмаль; 5
— дентин; 6 — цемент; 7
— зубная полость.
Строение зуба (рис. 98). Каждый зуб образован эмалью, дентином и цементом и имеет коронку, шейку, корень и внутри-зубную полость, заполненную пульпой.
Эмаль — поверхностный слой коронки. Ее образуют эпителиальные
клетки зубного органа — энамелобласты в период формирования зуба. В это
же время эмаль пропитывается минеральными солями и приобретает свои
уникальные свойства. Это самая I крепкая, наиболее минерализованная ткань
организма. Она состоит на 96—97% из неорганических веществ, в основном
фосфатов и карбонатов кальция, довольно много (до 4%) фторида кальция.
Органические вещества составляют 3—4% и образуют матрикс эмали. Пропитанный минеральными солями, он приобретает форму эмалевых призм —
S-образно изогнутых палочек толщиной 3—5 мкм, спаянных склеивающим
веществом.
Дентин образует основную массу зуба. Он имеется в коронке, шейке и
корне. Это разновидность костной ткани, 78% которой составляют минеральные вещества (преимущественно фосфаты кальция и магния с примесью
фторида кальция) и 28% — органические, в основном коллаген. Клетки этой
ткани — одонтобласты расположены на границе с пульпой зуба. Продуцированное ими межклеточное вещество и есть дентин. Масса дентина пронизана
многочисленными дентинными трубочками, в которых проходят отростки
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
одонтобластов. По трубочкам осуществляется питание зуба. Между дентином и одонтобластами имеется полоска предентина — необызвествленного
дентина. Рост дентина продолжается и у взрослого животного. Одонтобласты
отступают к пульпе, предентин, обызвествляясь, превращается в дентин, а
между дентином и одонтобластами вновь накапливается предентин, продуцируемый ими.
Цемент образуется в области корня. По химическому составу и строению напоминает грубоволокнистую костную ткань. Около 70% его массы составляют неорганические вещества. Отличие от костной ткани состоит в том,
что в цементе нет сосудов. Питание его диффузно.
Пульпа — рыхлая соединительная ткань, которая заполняет полость зуба
и каналы корней. В пульпе проходят сосуды и нервы. Здесь же имеются малодифференцированные клетки, способные превращаться в одонтобласты.
Различают зубы короткокоронковые и длиннокоронковые.
Короткокоронковые зубы не меняют своего положения в альвеоле. Их
коронка целиком выдвинута из челюсти и покрыта эмалью толщиной 0,5—4
мм. В течение жизни по мере стирания коронка становится короче. Под эмалью в коронке расположен дентин, а в центре — полость зуба, заполненная
пульпой. В области шейки зуб окружен десной. Эмаль здесь истончается и
сходит на нет. На нее сверху нарастает цемент. Корень зуба находится в альвеоле. Он образован дентином, который сверху покрыт цементом. В корне
проходит канал, заполненный пульпой. У многобугорчатых зубов по нескольку корней.
Длиннокоронковые зубы выдвигаются из альвеол по мере стирания, так
что часть их тела, расположенная над десной, остается неизмененной по высоте. В длиннокоронковом зубе различают тело и корень, в нем нет выраженной шейки. Тело зуба состоит из эмали и дентина на всем протяжении и
часто поверх эмали покрыто цементом, что придает зубам желтый цвет. Корни относительно короче, чем у короткокоронковых зубов.
У рогатого скота резцы на резцовой кости отсутствуют, на нижней челюсти их 4 пары: зацепы, средние медиальные, средние латеральные, окрайки. Самые крупные из них — зацепы, самые мелкие — окрайки. Предполагают, что это видоизмененные клыки. Молочные и постоянные резцы относятся к короткокоронковым зубам. У старых животных коронки стираются
до уровня десны. По размерам, форме и степени стертости резцов определяют возраст животного. Коренные молочные зубы короткоко-ронковые, коренные постоянные длиннокоронковые, лунчатого типа. Тело сравнительно
короткое, а корни длинные, хорошо выраженные. Премоляры в 2 раза тоньше
моляров. Зубная формула у рогатого скота, то есть количество зубов на одной половине верхней (числитель) и нижней (знаменатель) челюстей, следующая:
У свиньи все зубы короткокоронковые. Среди молочных зубов отсутствуют: первый премоляр и все моляры. Постоянные коренные зубы относятся
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
к многобугорчатым. Клыки развиты у свиней обоего пола. Особенно мощны
у самцов. Они глубоко сидят в альвеолах и длительно растут. Зубная формула
У лошади все молочные зубы короткокоронковые, все постоянные—
длиннокоронковые. Тела зубов длинные, массивные, глубоко погружены в
альвеолы, корни короткие. При выдвигании зубы прочно удерживаются в
альвеолах. На жевательной поверхности резцов имеется углубление — зубная чашка. Нестертая зубная чашка на верхних резцах имеет глубину 14 мм,
на нижних— 7 мм. По мере стирания глубина и форма чашки меняются, чем
основано определение возраста животных. Клыки имеются у самцов, у самок
часто отсутствуют. Они отделены беззубым краем от резцов и от коренных.
Имеют конусовидную форму. Молочные клыки развиваются, но обычно не
прорезываются, коренные зубы складчатого типа. Зубная формула
Твердое нёбо — palatum durum — крыша ротовой полости, отделяет ее
от носовой. Костной основой его являются нёбные
отростки резцовых, верхнечелюстных и горизонтальные пластинки нёбных костей. Слизистая оболочка покрыта многослойных: плоским эпителием
и образует поперечные валики. Вдоль нёба проходит нёбный шов. У жвачных поперечные валики в задней части нёба на уровне моляров отсутствуют.
Это не мешает отры гивать пищу во время жвачки.
Мягкое нёбо, или нёбная занавеска, — palatum molle — складка слизистой оболочки, являющаяся продолжением слизистой оболочки твердого нёба. Задний свободный конец мягкого нёба называется нёбной дугой (дужкой).
Она формирует выход из ротовой полости в глотку—зев. На мягком нёбе
различают ротовую поверхность, выстланную многослойным плоским эпителием, и глоточную поверхность, выстланную мерцательным эпителием. В
толще мягкого нёба имеются мышцы, железы и скопления лимфоидной ткани — миндалины.
У рогатого скота и свиньи мягкое нёбо короткое, зев широкий-, у лошади оно длинное, довольно плотно закрывает вход в глотку, соприкасаясь с
надгортанным хрящом. Поэтому лошади практически не могут дышать ртом.
Миндалины — tonsillae — крупные скопления лимфоидной ткани в
складках слизистой оболочки в виде множества лимфатических фолликулов
на границе ротовой полости и глотки. Различают непарную нёбную миндалину — в слизистой оболочке мягкого нёба, парные нёбные миндалины на
границе нёбной занавески и корня языка справа и слева, непарную глоточную — между входами в глоточнобарабанные (евстахиевы) трубы и непарную язычную — на корне языка. В совокупности они образуют лимфоэпителиальное глоточное кольцо, выполняющее функцию защитного барьера.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Дно ротовой полости выстлано слизистой оболочкой с многослойным
плоским эпителием. Оно почти целиком занято языком. Между языком и
деснами имеются боковые щелевидные пространства. От переднего участка
дна ротовой полости к языку тянется складка слизистой оболочки — уздечка
языка. По бокам от нее у рогатого скота и лошади видны два возвышения —
подъязычные или голодные бородавки. В них открываются протоки подчелюстной слюнной железы, а у рогатого скота и свиньи также протоки однопротоковой подъязычной слюнной железы. По бокам языка в дне ротовой полости открываются выводные протоки многопротоковой подъязычной слюнной
железы.
Язык—linqua (рис. 99)—массивный подвижный мышечный орган, служащий для захвата, перемешивания, продвижения и определения вкуса корма. У языка различают верхушку, тело и корень, а также спинку, боковые и
вентральную поверхности.
У рогатого скота несколько заостренная верхушка языка занимает переднюю часть ротовой полости, упираясь в резцовые зубы. На ней различают
все 4 поверхности. К вентральной поверхности прикрепляется двойная уздечка языка. Тело языка расположено между коренными зубами. На нем различают спинку и две боковые поверхности. На спинке заметно выступает эллипсоидное утолщение — подушка языка. Корень простирается от конца коренных зубов до надгортанника. Он прикреплен к к подъязычной кости, от
которой в язык заходит язычный хрящ; глубоко лежит в ротовой полости, так
что свободной оказывается лишь спинка. Вентральная поверхность языка
гладкая, спинка — шероховатая, так как усеяна многочисленными сосочками. Сосочки делят на механические и вкусовые. Механические сосочки двух
видов: нитевидные и конические. Они выполняют функцию осязания и способствуют удержанию и слизыванию корма. Нитевидные сосочки длинные
сильно ороговевшие, наиболее густо расположены на верхушке. На подушке
они уплощены. Среди них встречаются и мелкие конические сосочки. Спинка корня гладкая.
Вкусовые сосочки грибовидные и валиковидные. Грибовидные сосочки
разбросаны на верхушке и теле между нитевидными сосочками. Мелкие округлые с диаметром около 1 мм, слегка напоминают грибок на толстой ножке, содержат вкусовые луковицы. Желобоватые (валиковидные) сосочки
имеют округло-овальную форму, не выступают над поверхностью, окружены
валиком, отделенным от сосочка рвом. Содержат большое количество вкусовых луковиц. В дне рва открываются протоки вкусовых желез, секрет которых, растворяя частицы пищи, увеличивает возможности ее вкусового анализа. Лежат валиковидные сосочки позади подушки — у коров по 8—17, у овец
по 18— 28 сосочков с каждой стороны. Каждый сосочек имеет диаметр 2—5
мм.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 99. Язык крупного рогатого
скота:
I — внешний вид; II — гистологическое строение; А — нитевидный сосочек; Б — листовидный сосочек; В —
вкусовая луковица; 1 — верхушка; 2 —
тело; 3 — корень; 4— подушка; 5 —
нитевидные сосочки; 6 — грибовидные
сосочки; 7— валиковидные сосочки; 8
— эпителий сосочка; 9— собственная
пластинка слизистой; 10— вкусовая
луковица; 11—вкусовая пора; 12 —
вкусовая ямка; 13 — вкусовая клетка;
14 — поддерживающая клетка.
У свиньи язык относительно узкий с длинной верхушкой. На теле подушки нет. Язычный хрящ хорошо выражен. Нитевидные сосочки мягкие и
тонкие. На корне языка многочисленные крупные конические сосочки, направленные вершинами назад. Грибовидные сосочки есть не только на спинке, но и по бокам тела, где они особенно многочисленны. Два валиковидных
сосочка диаметром до 5 мм расположены около корня языка. По бокам от
них, на боковых поверхностях тела, имеется по одному вкусовому листовидному сосочку. Листовидный сосочек овальной формы, до 1 см длиной. Представляет собой комплекс складок слизистой оболочки, расположенных друг
около друга наподобие листков книги. Содержит наибольшее количество
вкусовых луковиц (до 7 тыс.).
У лошади язык подвижный, с закругленной верхушкой и длинным телом
без подушки. Уздечка языка одинарная. Язычкового хряща нет. Нитевидные
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
сосочки длинные, мягкие и тонкие. Конических сосочков нет. Валиковидных
сосочков два — около корня языка. Два крупных листовидных сосочка (1,5—
2 см) лежат по сторонам корня языка.
Гистологическое строение языка (II). Основная масса языка образована
поперечнополостной мышечной тканью, пучки волокон которой расположены вертикально и горизонтально, вдоль и поперек языка, что позволяет ему
осуществлять сложные движения. Кроме того, к языку подходят мышцы от
подъязычной кости и подбородка, обеспечивающие движения языка вперед,
назад и вбок.
Сверху язык покрыт слизистой оболочкой с многослойным плоским
эпителием. Собственная пластинка слизистой оболочки образована рыхлой
соединительной тканью с большим количеством сосудов и нервов. В ней залегают сложные альвеолярные и альвеолярно-трубчатые слюнные железы,
вырабатывающие серозный, слизистый и смешанный секрет. Концевые отделы желез заходят и в мышцу языка, располагаясь между мышечными пучками. В определенных местах (у корня) слизистой оболочки имеются крупные скопления лимфоидной ткани, формирующие миндалины. На спинке и
боковых поверхностях слизистая оболочка образует выросты — механические и вкусовые сосочки.
Механические сосочки — нитевидные и конические — покрыты многослойным плоским ороговевшим эпителием. Особенно толстый роговой слой
на вершине сосочка, что придает сосочку жесткость, языку шероховатость и
предохраняет слизистую от повреждения. Соединительнотканная основа сосочка является продолжением собственной пластинки слизистой внутрь сосочка.
Вкусовые сосочки — грибовидные, валиковидные (желобоватые), листовидные имеют сходное гистологическое строение, различаясь формой, размерами и количеством вкусовых почек (луковиц). Каждый вкусовой сосочек
представляет собой определенной формы складку слизистой оболочки. У
грибовидных сосочков складка слизистой расширена сверху и сужена внизу.
У валиковидных сосочков такая же складка, но в глубине слизистой. Она окружена кольцевидными бороздой (рвом) и валиком. Листовидные сосочки
состоят из ряда продольных складок, разделенных бороздами. Эпителий сосочков многослойный плоский неороговевающий. На боковых сторонах сосочков в слое эпителия залегают вкусовые почки (луковицы)—
хеморецепторы, реагирующие на вкус корма. На дне борозд (рвов) желобоватых и листовидных сосочков открываются протоки вкусовых желез. Вкусовые железы разветвленные трубчатые, выделяют жидкий серозный секрет,
омывающий эпителий сосочков и вкусовых луковиц и удаляющий раздражающие вещества.
Вкусовая луковица (почка) — яйцевидное (у жвачных), веретеновидное
(у свиньи), овальное (у лошади) тельце, состоящее из плотно уложенных вытянутых клеток, расположенное поперек эпителиального пласта. От подлежащей соединительной ткани она отделена базальной мембраной. На поверхности эпителия вкусовая почка открывается отверстием — вкусовой поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рой, которая ведет в небольшое углубление — вкусовую ямку. С помощью
электронной микроскопии в почке различают несколько видов клеток, функции которых окончательно не разграничены. Основную массу (до 70%) составляют более темные клетки с удлиненными ядрами и микроворсинками на
апикальном конце. По-видимому, они выполняют опорную, защитную и фагоцитарную функции.
Рецепторные вкусовые клетки составляют 10—15% от общего количества дифференцированных клеток вкусовой почки. Их апикальный конец электронно-плотный, снабжен микроворсинками, ядра овальные, смещены базально, у базального полюса можно видеть типичные синаптические связи с
нервными окончаниями. Жизненный цикл клеток вкусовой почки равен в
среднем 10 дням. Затем они погибают и фагоцитируются. Их место занимают
молодые клетки, дифференцирующиеся из так называемых базальных клеток
— коротких, не достигающих вкусовой ямки, лежащих на дне вкусовой почки. В каждую почку входит 50— 60 нервных окончаний — это разветвления
чувствительных нервов, идущих к головному мозгу.
Рис. 100. Слюнные железы крупного рогатого скота:
1—околоушная железа и 2 — ее
проток; 3 — нижнечелюстная железа; 4 — щечные железы.
Слюнные железы — glandulae salivales (рис. 100) —вырабатывают секрет — слюну, основное назначение которой — увлажнение корма. У крупного рогатого скота за сутки выделяется до 60 л слюны, у лошади— 40 л, у
свиньи — 15 л. По составу секрет может быть серозным, слизистым и смешанным. Слюнные железы располагаются как в стенках органов ротовой полости — пристенные, так и за ее пределами —застенные железы. К пристенным желе зам относят многочисленные мелкие губные, щечные, язычные,
нёбные железы, залегающие в слизистой оболочке этих органов. Застенные
сложные железы — околоушные, нижнечелюстные, подъязычные — парные
компактные органы.
Околоушная железа — gl. parotis — у рогатого скота имеет вид вытянутого утолщенного треугольника буровато-красного цвета, расположенного
под наружным слуховым проходом между ветвью нижней челюсти и атлантом, основанием направленного дорсально, а вершиной вентрально, простираясь до угла нижней челюсти. Прикрывает собой частично подчелюстную
железу и большую жевательную мышцу. Выводный проток околоушной жеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лезы проходит в межчелюстном пространстве, затем через сосудистую вырезку проходит на латеральную поверхность и открывается в преддверии ротовой полости на уровне 4-го коренного зуба. У свиньи это самая крупная
слюнная железа. Имеет форму треугольника, широким основанием направленного вниз, а вершиной — вверх. Впереди доходит до большой жевательной мышцы.
У лошади околоушная железа имеет форму четырехугольника. Проток
открывается против третьего коренного зуба.
Нижнечелюстная (подчелюстная) железа — gl. mandibularis — у рогатого скота самая крупная из слюнных желез, удлиненная, желтоватого цвета.
Лежит в межчелюстном пространстве от атланта до сосудистой вырезки.
Прикрыта околоушной железой, двубрюшной и крыловой мышцами. Проток
нижнечелюстной железы проходит в мускулатуре межчелюстного пространства по медиальной поверхности подъязычной железы и открывается в подъязычной (голодной) бородавке на дне ротовой полости. У свиньи железа небольшая, округлой формы, лежит на уровне угла нижней челюсти, целиком
закрыта околоушной слюнной железой. Проток открывается на дне ротовой
полости возле уздечки языка. У лошади она удлиненная, слегка серповидно
изогнутая. Лежит на протяжении от атланта до тела подъязычной кости в
межчелюстном пространстве.
Подъязычная железа — gl. sublingualis — у рогатого скота лежит в слизистой дна ротовой полости сбоку от корня языка до подбородочного угла.
Делится на две части: однопротоковую (длиннопротоковую) и многопротоковую (короткопротоковую). Однопротоковая железа длиннее и лежит вентральнее и впереди от многопротоковой. Ее большой подъязычный проток
открывается в голодной бородавке. Малые подъязычные протоки многопротоковой железы короткие, открываются сбоку от языка в дне ротовой полости.
У свиньи железа красновато-желтоватого цвета. Однопротоковая железа
лежит позади многопротоковой. Большой подъязычный проток иногда сливается с подчелюстным протоком. Открывается в дне ротовой полости около
уздечки языка.
У лошади подъязычная железа состоит только из короткопротоковой ее
части. Лежит она на протяжении от третьего коренного зуба до подбородочного угла.
Гистологическое строение застенных слюнных желез (рис. 101). Застенные слюнные железы — компактные органы, состоят из стромы и паренхимы. Строма образована соединительной, паренхима — эпителиальной тканью. В состав стромы входят: капсула, одевающая железу снаружи; отходящие от капсулы широкие междольковые прослойки; разветвляющиеся в
дольках нежные тонкие внутридольковые прослойки. В соединительнотканной строме проходят выводные протоки железы, сосуды и нервы вегетативной нервной системы (симпатической и парасимпатической), регулирующие приток крови к железе и ее секреторную деятельность.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 101. Схема строения паренхимы околоушной (А), нижнечелюстной (Б), подъязычной (В) слюнных желез:
1 — междольковый выводной проток; 2 — исчерченный проток; 3 —
вставочный проток; 4 — альвеола; 5
— трубка; 6 — альвеолотрубка; 7 —
миоэпителиальная клетка; а — слизистые и б — серозные клетки.
Паренхима состоит из концевых отделов и выводных протоков. Концевые отделы — это замкнутые конечные структуры в виде трубочек и мешочков (альвеол), образованных однослойным железистым эпителием, который
секретирует слюну. Выведение секрета из клеток осуществляется по мерокринному типу. Из просвета концевых отделов секрет удаляется при сокращении миоэпителиальных (корзинчатых) клеток, покрывающих снаружи железистый эпителий. Миоэпителиальные клетки — это специализированные отростчатые клетки, функцией которых является сокращение. Соединяясь друг
с другом отростками, они образуют нечто вроде сетчатой корзинки вокруг
железистого эпителия. Концевые отделы составляют основную массу долек.
Друг от друга они отделены тончайшими прослойками рыхлой соединительной ткани. Из концевых отделов секрет поступает в систему выводных
протоков. Несколько концевых отделов, объединенных общим выводным
протоком, называется ацинусом.
Самые мелкие — вставочные протоки представляют собой узкие трубочки из уплощенного или кубического эпителия, покрытого снаружи миоэпителиальными клетками. Диаметр вставочных протоков меньше диаметра
концевых отделов. Вставочные протоки переходят в исчерченные протоки.
Исчерченный проток имеет больший диаметр, стенка его построена из цилиндрического эпителия, клетки которого имеют ацидофильную цитоплазму
и содержат секреторные гранулы. Исчерченные протоки называются так потому, что плазмолемма базальных частей их эпителия образует глубокие впячивания внутрь клеток, а между складками плазмолеммы располагаются митохондрии. Такая структура говорит об их секреторной деятельности. Исчерченные протоки, как и вставочные, являются внутридольковыми выводными
протоками. Они объединяются в междольковые выводные протоки, а те — в
главный выводной или выводные протоки железы.
Различия между тремя застенными железами заключаются в основном в
строении концевых отделов и начальных участков системы выводных протоков.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Околоушная железа — альвеолярная, вырабатывает серозный (белковый) секрет, который у свиньи и лошади содержит ферменты диастазу и амилазу, расщепляющие крахмал. Одета плотной соединительнотканной капсулой. Соединительнотканная строма хорошо развита, отчего паренхима железы четко разделена на дольки. Дольки образованы концевыми отделами альвеолярной формы и системой внутридольковых протоков. Эпителий альвеол
однослойный призматический. Ядра в клетках эпителия округлые, содержат
глыбчатый хроматин, лежат в середине клетки или несколько сдвинуты к базальному полюсу. Цитоплазма умеренно базофильная в базальной части, где
расположена хорошо развитая цитоплазматическая сеть и оксифильная в
апикальной части при накоплении секреторных гранул. Просвет альвеол небольшой. Вставочные протоки хорошо развиты. Главный околоушной выводной проток выходит за пределы железы, одет плотной соединительной
тканью.
Нижнечелюстная железа — альвеолярно-трубчатая, вырабатывает серозно-слизистый секрет. Одета капсулой. Строма менее развита, чем в околоушной железе, дольчатость выражена. Концевые отделы нескольких типов:
альвеолярные — серозные, трубчатые — слизистые, альвеолярно-трубчатые
— смешанные. Строение альвеолярных концевых отделов такое же, как в
околоушной слюнной железе. Трубчатые концевые отделы имеют вид одиночных или разветвленных коротких и широких трубок, стенка которых образована цилиндрическим или высокопризматическим эпителием. Просвет
трубок сравнительно широкий. Слизистые клетки со светлой пенистой цитоплазмой, уплощенным ядром, сдвинутым к базальному полюсу. Альвеолярно-трубчатые концевые отделы содержат как серозные, так и слизистые
клетки. В большинстве случаев серозные клетки лежат в глубине, на самом
дне альвеолотрубки. Вставочные протоки короче и встречаются реже. Повидимому, происходит их ослизнение и превращение в трубчатые концевые
отделы. Остальные участки протоковой системы развиты хорошо.
Подъязычная железа не имеет общей капсулы, разделена на дольки.
Подъязычная однопротоковая слюнная железа по строению и характеру секрета подобна нижнечелюстной железе. Подъязычная многопротоковая
слюнная железа чисто слизистая, состоит из трубчатых концевых отделов.
Вставочные протоки отсутствуют.
Глотка — pharynx — трубкообразный орган конической формы, расположенный на перекрестии пищеварительного и дыхательного путей. Спереди
глотка граничит с ротовой и носовой полостями, сзади — с гортанью и пищеводом. Пограничное расположение глотки наложило отпечаток на ее
строение: глотку делят на пищеварительную (ротовую) и дыхательную (гортанную) части, различающиеся характером эпителия слизистой оболочки. В
пищеварительной части он многослойный плоский, в дыхательной— многорядный мерцательный. Основу глотки образуют мышцы, расширяющие и
сужающие ее просвет. Снаружи глотка покрыта фасциями и рыхлой соединительной тканью, связывающей ее с соседними органами. В глотку ведут 3
отверстия: спереди и сверху —из носовой полости — хоаны (парные), спереwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ди и снизу — из ротовой полости — зев. Из глотки — 4 отверстия: назад и
вниз — в гортань, назад и вверх — в пищевод, по бокам и вверх — к среднему уху по глоточно-барабанным трубам.
Во время глотания нёбная занавеска (мягкое нёбо) напрягается и закрывает хоаны. Нёбно-глоточная дужка, напрягаясь, отделяет ротовую часть
глотки от гортанной. Мышцы подъязычного аппарата, гортани и языка, сокращаясь, двигают язык и гортань навстречу друг другу. Корень языка при
этом надавливает на надгортанник, вход в гортань закрывается, и пищевой
ком эвакуируется в пищевод.
ПЕРЕДНЯЯ КИШКА, ИЛИ ПИЩЕВОДНО-ЖЕЛУДОЧНЫЙ ОТДЕЛ
Пищевод — oesophagus — трубкообразный орган, который обеспечивает перемещение пищевого кома из ротоглотки в желудок. Максимальная ширина пищевода в растянутом состоянии у крупного рогатого скота—17 см, у
овцы — 9, у свиньи — 7, у лошади — 6.5 см. В нем различают шейную,
грудную и брюшную части. Шейная часть пищевода лежит сначала сверху от
трахеи, затем на уровне 5-го шейного позвонка он опускается на левую сторону трахеи, образуя петлю. Петля пищевода обеспечивает его расправление
при движении шеи. Грудная часть пищевода проходит в средостении. В
брюшную полость пищевод проходит через пищеводное отверстие диафрагмы. Брюшная часть пищевода проходит по тупому краю печени, образуя на
нем пищеводное вдавление и входит с левой стороны в желудок, у рогатого
скота — в преддверие рубца. Перед входом в желудок образуется расширение — приспособление для облегчения отрыгивания корма при жвачке.
У свиньи и лошади пищевод входит в кардиальную часть желудка. У
свиньи перед входом в желудок несколько расширяется. У лошади, наоборот,
просвет пищевода сужается, а стенка утолщается, формируя сфинктер, препятствующий рвотным движениям.
Иннервируется пищевод блуждающим нервом и постганглионарными
волокнами каудального шейного и звездчатого ганглиев, васкуляризируется
общими сонными и пищеводной артериями.
Гистологическое строение пищевода (рис. 102). Как всякий трубкообразный орган, пищевод состоит из слизистой оболочки подслизистой основы,
мышечной оболочки и адвентиции.
Слизистая оболочка у млекопитающих после рождения покрыта многослойным плоским эпителием. У копытных степень ороговения эпителия прямо пропорциональна качеству корма: чем он грубее, тем больше ороговение.
Наибольшее ороговение наблюдается у лошади и крупного рогатого скота,
наименьшее— у свиней. Ороговение эпителия уменьшается, а его толщина
увеличивается в каудальном направлении. Собственная пластинка слизистой образована рыхлой соединительной тканью.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 102. Схема строения пищеварительной трубки:
А— пищевод; Б — кардиальная и В — фундальная зоны желудка; Г —
двенадцатиперстная, Д — тощая. Б — подвздошная кишки тонкого кишечника; Ж— толстый кишечник; З — конец прямой кишки; 1— слизистая оболочка; 1—покровный эпителий; 2 — собственная пластинка слизистой оболочки; 3 — мышечная пластинка слизистой оболочки; 4 — железы желудка;
5 — желудочные ямки; 6 — ворсинки; 7 — крипты; 8 — лимфатические фолликулы; II — подслизистая основа; 9 — пищеводные железы; 10 — железы
двенадцатиперстной кишки; 11 — кровеносные сосуды; 12 — подслизистое
(мейснерово) нервное сплетение; 13 — лимфатические фолликулы; III —
мышечная оболочка; 14 — кольцевой слой; 15 — продольный слой; 16 — межмышечное (ауэрбахово) сплетение; IV — серозная оболочка; 17 — собственная пластинка серозной оболочки; 18 — мезотелий; V — адвентиция.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Она вдается сосочками в слой эпителия. Сосочки гуще расположены в
начальной части пищевода. Содержит ретикулярную ткань со скоплениями
лимфоцитов. Мышечная пластинка слизистой образована продольно расположенными пучками гладкой мышечной ткани. У рогатого скота и лошади
она развита на всем протяжении пищевода, у свиньи — в дистальных 2/3 органа. Утолщается в каудальном направлении, так что в брюшной части пищевода толщина ее составляет 1 мм. Наличие мышечной пластинки позволяет слизистой оболочке собираться в собственные складки.
Подслизистая основа образована рыхлой соединительной тканью с
большим количеством сосудов и нервов, которые образуют здесь сплетения.
В ней залегают сложные трубчато-альвеолярные железы, содержащие преимущественно слизистые клетки и вырабатывающие слизистый секрет у рогатого скота, смешанный — у свиньи. Располагаются железы у рогатого скота и лошади лишь в начальном участке пищевода — около глотки, у свиньи
— в передней половине органа.
Мышечная оболочка у животных, способных к частому отрыгиванию,—
жвачных и хищных — на всем протяжении пищевода образована поперечнополосатой мышечной тканью, которая обеспечивает быстрое расширение
пищевода. У лошади и свиньи в передней половине поперечнополосатая, а
затем гладкая мышечная ткань. Переход происходит постепенно. В мышечной оболочке различают два слоя: внутренний — кольцевой и наружный—
продольный. В обоих слоях мышечные пучки располагаются спирально, но
во внутреннем в виде крутой спирали, а в наружном — пологой. Попеременное сокращение мышечных слоев приводит к перистальтическим движениям
органа, что и продвигает пищу.
Наружной оболочкой шейной и частично грудной частей пищевода является адвентиция — рыхлая соединительная ткань. Задний участок грудной
и брюшная части пищевода снаружи покрыты серозной оболочкой с толстой
собственной пластинкой из рыхлой соединительной ткани, покрытой снаружи однослойным плоским эпителием — мезотелием.
Нервы вегетативной нервной системы и сосуды образуют несколько
сплетений: в слизистой, в подслизистой основе, мышечной и наружной оболочках.
Желудок — ventriculus (лат.), gaster (греч.) (рис. 103) — мешковидной
формы полостной орган, в котором корм задерживается и частично переваривается. По количеству камер различают желудки однокамерные — у большинства млекопитающих (из сельскохозяйственных животных — у лошади,
свиньи) и многокамерные— у жвачных. В зависимости от характера слизистой оболочки желудки делят на безжелезистые, или пищеводного типа, железистые, или кишечного типа, и смешанные, или пищеводно-кишечного типа.
Однокамерные желудки. Желудок свиньи крупный, пищеводнокишечного типа. Он имеет две поверхности: париетальную, или диафрагмальную, — обращенную к диафрагме и печени, и висцеральную — обращенную к кишечнику. Влево, назад и вниз желудок направлен выпуклой большой
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
кривизной, вправо, вперед и вверх — вогнутой малой кривизной. В желудке
различают: вход из пищевода в желудок — кардиальное отверстие — с левой стороны желудка, выход из желудка в двенадцатиперстную кишку— пилорическое отверстие. Воронкообразное расширение вокруг кардиального
отверстия является кардиальной частью желудка. Область желудка вокруг
пилорического отверстия — пилорическая часть. Область между ними —
тело желудка, вентральная часть которого называется дном. В кардиальной
области имеется слепое выпячивание — дивертикул желудка. В пилорической части при переходе в двенадцатиперстную кишку имеется сфинктер пилоруса. Слизистая пищеводного типа занимает небольшой участок от кардиального отверстия до дивертикула желудка. Она более светлая и жесткая на
ощупь по сравнению со слизистой кишечного типа, выстлана многослойным
плоским эпителием и не содержит желез.
Рис. 103. Распределение железистых зон в желудке свиньи (Л), лошади
(Б) и крупного рогатого скота (В):
1— конец пищевода; 2— кардиальная часть; 3— тело (фундальная
часть); 4 — пилорическая часть; 5 — начало двенадцатиперстной кишки; 6
— малая кривизна; 7 — большая кривизна; 8 — дивертикул; 9 — слепой мешок; 10 — преддверие рубца; 11 — дорсальный мешок; 12 — дорсальный слепой мешок; 13 — вентральный слепой мешок; 14 — вентральный мешок; 15
— сетка; 16 — книжка; 17 — сычуг.
Иннервируется желудок блуждающим нервом и нервными волокнами
солнечного сплетения. Васкуляризируется желудочной артерией, отходящей
от чревной артерии.
Расположен желудок кардиальной частью в левом подреберье, пилорической — немного заходит в правое подреберье, а дно лежит в области мечевидного отростка, достигая с левой стороны брюшной стенки.
Желудок лошади относительно небольшой, емкостью-6—15 л, пищеводно-кишечного типа. В нем различают те же части, что и в желудке свиньи.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Правда, отсутствует воронкообразное расширение в кардиальной части. Вместо него в стенке желудка формируется мощный мышечный кардиальный
сфинктер, охватывающий вход пищевода в желудок. Чем больше наполняется желудок, тем сильнее мышечная петля сфинктера сжимает пищеводное отверстие, препятствуя рвотным движениям. В кардиальной части желудка имеется большое выпячивание — слепой мешок, который занимает
весь левый конец желудка и имеет слизистую пищеводного типа. Она резко
отделяется от слизистой кишечного типа складкой и более светлым цветом.
Расположен желудок почти целиком в левом подреберье, с небольшим
заходом в правое.
Гистологическое строение однокамерного желудка (рис.104). Стенка
желудка состоит из слизистой оболочки, подслизистой основы, мышечной и
серозной оболочек.
Слизистая оболочка в смешанных желудках двух типов: пищеводного и
кишечного. Она имеет неровный рельеф из-за складок и ямок.
Слизистая оболочка пищеводного типа покрыта многослойным плоским
эпителием. Собственная пластинка слизистой состоит из рыхлой соединительной ткани. Желез в ней нет. Там, где мышечная пластинка развита, слизистая оболочка образует мелкие складки. У свиньи данная оболочка занимает небольшой участок в кардиальной части желудка, идущий от кардиального отверстия до дивертикула. У лошади она выстилает всю кардиальную
часть желудка, включая слепой мешок.
Слизистая оболочка кишечного типа покрыта однослойным цилиндрическим эпителием, клетки которого вырабатывают слизь. Слизь находится на
всей поверхности слизистой оболочки, предохраняя ее от повреждений.
Рельеф слизистой оболочки неровный, образует складки и ямки. Собственная
пластинка слизистой оболочки толстая, сплошь занята железами, разделенными тончайшими прослойками рыхлой соединительной ткани. Они вырабатывают желудочный сок. В зависимости от расположения, особенностей
строения и вырабатываемых секретов железы делят на кардиальные, фундальные и пилорические. Мышечная пластинка слизистой оболочки из гладкой мышечной ткани. Хорошо развита. Ее сокращение способствует выведению секрета из желез.
Фундальные (собственные) железы наиболее распространенные, расположены в теле и дне желудка. Это простые разветвленные или неразветвленные трубчатые железы длиной 1,5—2,5 мм. В них различают перешеек (шейку), тело и дно. Тело и дно железы являются ее секреторными отделами, а
перешеек — выводным протоком, открывающимся в дно желудочной ямки.
У свиньи и лошади в одну желудочную ямку открывается по нескольку желез. Просвет железы очень узкий, а стенки образованы однослойным эпителием, в состав которого входит несколько разновидностей клеток: главные,
париетальные (обкладочные), слизистые (добавочные), эндокринные.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 104. Строение фундальной (собственной) железы желудка:
I— желудочная ямка; II — шейка; III—тело и IV — дно железы; 1 — покровный эпи телий; 2 — добавочная клетка; 3— париетальная клетка; 4 —
главная клетка; 5 — ядро; 6 — митохондрии; 7— цитоплазматическая сеть;
8 — комплекс Гольджи; 9 — внутриклеточные канальцы; 10 — секреторные
гранулы.
Главные клетки кубической или несколько удлиненной (в период накопления секрета) формы, с округлым ядром, лежащим ближе к базальному полюсу. У этого же полюса расположена хорошо развитая гранулярная цитоплазматическая сеть, определяющая его базофилию. Апикальный полюс занят крупными (до -3 мкм) гранулами секрета, содержащими пепсиноген —
фермент в неактивной форме, который под влиянием соляной кислоты превращается в пепсин — ключевой фермент, участвующий в расщеплении белков. Экструзия секрета происходит мерокринным способом через апикальный полюс клетки. Главные клетки расположены в теле и дне железы, составляя здесь 60—70% всех: клеток. Париетальные (обкладочные) клетки
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
крупнее остальных,, овальной, яйцевидной или пирамидной формы с резко
оксифильной цитоплазмой и округлым ядром. Благодаря крупным размерам
они выпучиваются из единого пласта эпителия. Цитоплазма их богата крупными митохондриями и пронизана внутриклеточными секреторными канальцами с многочисленными микроворсинками. Канальцы открываются на апикальном полюсе. Париетальные клетки вырабатывают хлориды, которые в
просвете желудка превращаются в соляную кислоту и активизируют пепсиноген, превращая его в пепсин. Располагаются париетальные клетки преимущественно в теле, но встречаются и в других участках железы.
Слизистые (добавочные) клетки кубической или цилиндрической
формы с вытянутым уплотненным ядром, лежащим у основания клетки.
Апикальный полюс имеет пенистый вид в результате накопления секреторных гранул, содержащих слизистые (мукоидные) вещества. Среди этих клеток часто наблюдаются митозы. Считается, что наряду с выработкой слизи,
предохраняющей поверхность железы от воздействий соляной кислотой и
пепсином, эти клетки выполняют камбиальную функцию — являются источником клеток покровного и железистого эпителия желудка в процессе его
физиологического изнашивания. Средняя продолжительность жизненного
цикла клеток эпителия желудка равна 10 дням. Располагаются слизистые
клетки преимущественно в шейке, но могут встречаться и в других частях
железы. Эндокринные клетки очень разнообразны по образуемым ими биологически активным веществам. Могут быть дифференцированы с помощью
специальных методов исследования. Располагаются: в теле и дне во всех видах желез желудка. Продуцируемые ими гормоноподобные вещества участвуют в регуляции не только деятельности желудка, но и всей пищеварительной системы.
Кардиальные железы у свиньи занимают всю кардиальную область и
часть тела желудка. У лошади лежат узкой полосой на границе с безжелезистой зоной желудка. Это простые разветвленные трубчатые железы. По
структуре они подобны фундальным железам, но короче их, с более широким
просветом и расположены реже. Состоят в основном из слизистых клеток,
подобных добавочным клеткам фундальных желез. Содержат эндокринные
клетки. Изредка в них встречаются главные и париетальные клетки, а у свиньи также клетки, вырабатывающие ферменты, расщепляющие крахмал, так
что наряду со слизью кардиальные железы вырабатывают биологически активные вещества и в небольшом количестве пепсиноген и хлориды.
Пилорические железы располагаются в пилорической части желудка.
Резкой границы между областью фундальных и пилорическпх желез нет. Это
тоже простые разветвленные трубчатые железы. У них длинная шейка (от 1/3
до 2/3 длины железы) и короткий сильно ветвящийся секреторный отдел. Образованы эти железы слизистыми клетками нескольких типов. Среди эпителия пилорических желез встречаются и эндокринные клетки.
Подслизистая основа хорошо развита, что обеспечивает подвижность
слизистой оболочки. Состоит она из рыхлой соединительной ткани с большим количеством кровеносных и лимфатических сосудов и нервов, которые
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
образуют в ней сплетения. В рыхлой соединительной ткани слизистой оболочки и подслизистой основы можно видеть скопления лимфоцитов.
Мышечная оболочка образована гладкой мышечной тканью. Состоит из
2—3 слоев: внутреннего — косого, среднего — кольцевого и наружного —
продольного. Слои неодинаково развиты в разных участках желудка.
В межпучковых прослойках рыхлой соединительной ткани проходят сосуды и нервы межмышечного сплетения.
Серозная оболочка состоит из рыхлой соединительной ткани и однослойного плоского эпителия (мезотелия). Перекидываясь с малой кривизны
желудка на печень, серозная оболочка формирует малый сальник, а спускаясь
с большой кривизны желудка— образует большой сальник.
Многокамерный желудок жвачных пищеводно-кишечного типа (см. рис.
103). Большого объема — у крупного рогатого скота емкость 100—300 л. Состоит из четырех камер: рубца, сетки, книжки и сычуга. Первые 3 камеры
имеют слизистую оболочку пищеводного типа и называются преджелудком.
Собственно желудок— сычуг имеет слизистую оболочку кишечного типа.
Пред-желудок является резервуаром для временного хранения и предварительной обработки грубого корма. Поступившая в рубец и сетку слабо
пережеванная и смоченная слюной пища мацерируется и частично расщепляется под действием ферментов корма и микрофлоры, обитающей в преджелудке (инфузории, целлюлозолитические и другие микроорганизмы). Корм
после отрыгивания и вторичного пережевывания поступает в последующие
камеры — книжку и сычуг.
Рубец — rumen — начальная, у взрослых животных самая крупная камера. На нем различают: париетальную поверхность— прилежащую к левой
стенке брюшной полости, висцеральную поверхность — обращенную к внутренностям, дорсальную и вентральную кривизны, краниальный и каудальный
концы. Рубец сплющен с боков и состоит из двух сообщающихся между собой мешков — вентрального и дорсального, отграниченных друг от друга
правым и левым продольными желобами. В краниальный край дорсального
мешка входит пищевод. Это место называется преддверием рубца, а отверстие, через которое пищевод сообщается с рубцом, — кардиальным. От дорсального и вентрального мешков ближе к каудальному краю отделяются желобами каудодорсальный и каудовентральный слепые мешки.
Слизистая оболочка рубца образует многочисленные листовидные выросты — сосочки. В разных участках рубца они имеют разную толщину и
высоту — от 3 до 12 мм. Рядом с преддверием в дорсальном мешке рубца
имеется широкое отверстие, ведущее в сетку.
От кардиального отверстия вдоль правой стенки преддверия рубца по
его внутренней поверхности к сетке проходит желоб желудка (пищеводный)
или желоб сетки, книжки и сычуга, отграниченный губами — двумя близко
лежащими, спирально изогнутыми складками слизистой оболочки. Он продолжается по дну сетки и книжки до сычуга.
По нему большая часть жидкости, особенно у молодняка, стекает в сычуг, минуя преджелудок.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Сетка — reticulum — самая маленькая округлая камера у крупного рогатого скота, третья по величине и овальная по форме— у овцы. Лежит впереди рубца и открывается в него широким отверстием в области преддверия.
С книжкой связана узким отверстием. По правой стенке сетки проходит желоб желудка. Слизистая оболочка сетки образует выступы разной высоты, создающие характерный рельеф в виде ячеек разного размера.
Функцией сетки является перемешивание и сортировка корма, поступающего в нее из рубца. Более крупные частицы, видимо, вновь возвращаются в рубец, измельченные поступают в книжку.
Предполагают, что сортировка корма осуществляется с помощью ячеек
разного размера.
Книжка — omasum — вторая или третья по величине камера округлой
формы у крупного рогатого скота и самая маленькая овальная у овец. Имеет
2 отверстия — одно из сетки, другое — в сычуг. Между ними по дну книжки
идет желоб желудка. При входе в сычуг мышечная оболочка желоба утолщается, образуется сфинктер. Слизистая оболочка книжки образует большое количество складок — листочков четырех размеров, закономерно чередующихся и подвижных, что позволяет дополнительно перемешивать и перетирать пищу перед поступлением ее в сычуг. Поверхность листочков покрыта
мелкими грубыми сосочками, которые при движении листочков работают
как терка.
Сычуг — abomasum — вторая или третья по величине камера грушевидной формы у взрослого крупного рогатого скота, вторая— у овцы. В молочный период — самая крупная камера. На нем различают вогнутую — малую кривизну и выпуклую — большую кривизну. Краниальным расширенным
концом направлен к книжке, каудальный суженным концом — пилорусом к
двенадцатиперстной кишке. При переходе его в двенадцатиперстную кишку
образуется сфинктер пилоруса. Слизистая оболочка кишечного типа нежнорозового цвета, образует продольные нерасправляющиеся складки.
Иннервируется и снабжается кровью желудок жвачных из тех же источников, что и однокамерный желудок: ветвями блуждающего нерва, нервами
солнечного сплетения, ветвями чревной артерии.
Расположение желудка у крупного рогатого скота. Рубец занимает всю
левую половину брюшной полости и заходит в правую половину, располагаясь в области мечевидного хряща, левых подреберья, подвздоха, паховой, поясничной и пупочной областях. Краниальным концом он доходит до диафрагмы, дорсальной кривизной касается диафрагмы и поясничных мышц, париетальной поверхностью прилежит к левой стенке тела, отделяясь от нее
только селезенкой; висцеральной поверхностью граничит с кишечником.
Каудальным концом доходит до входа в таз, вентральным мешком заходит в
правую половину брюшной полости. Сетка лежит в области мечевидного отростка впереди рубца, касаясь диафрагмы, а книжка — в правом подреберье
над сеткой и сычугом, справа от рубца, слева от печени и ниже ее. Сычуг
расположен в правом подреберье, частично заходит в область мечевидного
отростка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Гистологическое строение многокамерного желудка. Стенка желудка
состоит из слизистой оболочки, подслизистой основы, мышечной и серозной
оболочек. В отдельных камерах имеются свои особенности в строении оболочек, (рис. 105).
Рубец имеет слизистую оболочку пищеводного типа. Она покрыта многослойным плоским ороговевающим эпителием. Роговой слой его невелик,
так как постоянно слущивается при трении о пищу. Собственная пластинка
слизистой образована рыхлой волокнистой соединительной тканью с большим количеством клеток, среди которых встречаются и лимфоциты. Слизистая оболочка образует выросты в виде сосочков. Мышечная пластинка слизистой отсутствует, поэтому четкой границы между слизистой оболочкой и
подслизистой основой провести нельзя. Желез в стенке рубца нет. Подслизистая основа построена из рыхлой соединительной ткани, в которой заметно
больше волокон и меньше клеток, крупнее сосуды по сравнению с собственной пластинкой слизистой. Мышечная оболочка состоит из двух слоев гладкой мышечной ткани. Пучки внутреннего слоя по преимуществу располагаются поперек рубца, наружного — вдоль. Серозная оболочка представлена
тонкой собственной пластинкой и мезотелием.
Рис. 105. Внешний вид стенки сетки
(А), книжки (Б) и гистологическое
строение стенки рубца (В):
I — слизистая оболочка; 1 — эпителий; 2 — собственная пластинка; 3 —
мышечная пластинка; II — подслизистая основа; III—мышечная оболочка;
4— кольцевой слой; 5 — продольный
слой; IV — серозная оболочка.
Сетка по гистологическому строению очень похожа на рубец. Отличается лишь тем, что в стенках наиболее крупных ячеек слизистой оболочки
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
имеются пучки гладкомышечных клеток. Сетка имеет наиболее толстую
мышечную оболочку.
В книжке в отличие от других камер преджелудка хорошо развита мышечная пластинка слизистой оболочки, которая в виде сплошного слоя отделяет собственную пластинку от подслизистой основы и заходит в листочки.
В крупные листочки заходит подслизистая основа и даже внутренний слой
мышечной оболочки.
В результате листочки могут укорачиваться и удлиняться.
Сычуг — собственно желудок со слизистой оболочкой кишечного типа.
Строение его стенки подобно строению стенки однокамерного желудка кишечного типа. В сычуге также различают зоны кардиальных, фундальных и
пилорических желез. Зона кардиальных желез имеет вид узкой полосы у краниального края сычуга. Пилорические железы лежат в пилорической части
сычуга.
Наиболее распространенные железы фундальные. Отличия от желудка
свиньи и лошади в том, что желудочные ямки в сычуге узкие. В каждую ямку
открывается только по одной железе. У фундальных желез длинная шейка и
сравнительно короткое тело. Пилорические железы длиннее, чем у других
сельскохозяйственных животных.
СРЕДНЯЯ КИШКА, ИЛИ ТОНКИЙ КИШЕЧНИК
Средняя кишка (тонкий кишечник) имеет большую длину (свыше 40 м у
крупного и около 30 м у мелкого рогатого скота и лошади, свыше 20 м у свиньи) и состоит из двенадцатиперстной, тощей, подвздошной кишок (рис. 106)
и застенных желез, связанных с двенадцатиперстной кишкой: печени и поджелудочной железы. Диаметр тонкого кишечника относительно небольшой.
В тонком кишечнике происходят наиболее активные процессы переваривания и всасывания пищи.
Двенадцатиперстная кишка — intestinum duodenum (рис.107)—у
крупного рогатого скота длиной 90—120 см, у мелкого — около 50 см.
Шириной 5—7 см у крупного и 2—3 см у мелкого рогатого скота. Подвешена
на короткой брыжейке, а местами лежит экстраперитониально. Вследствие
этого она не меняет своего положения в брюшной полости.
Располагается в основном в правом подреберье и лишь немного заходит
в поясничную область. Начинаясь от сычуга, она направляется вперед до печени. Около ворот печени в правом подреберье делает S-образный изгиб,
поднимается каудодорсально, доходит до правой почки, отсюда направляется
назад до подвздошной кости, после чего поворачивает налево и вперед и без
резких границ переходит в тощую кишку. Примерно на середине в двенадцатиперстную кишку впадает желчный проток, а несколько дальше его — проток поджелудочной железы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 106. Кишечник крупного рога, того скота (А), свиньи (Б) и лошади
(В):
1 — пилорус желудка; 2 — двенадцатиперстная кишка; 3 — тощая кишка; 4
— подвздошная кишка; 5 —слепая кишка и ее 6 —верхушка; 7 —тело; 8 —
головка (основание); 9 —тении; 10— кармашки; 11 — ободочная кишка и ее
12 — начальная, 13 — центрипетальная, 14 — центрифугальная и 15 — конечная петли; 16 — левые вентральное и дорсальное положения; 17 — диафрагмалъные положения; 18 — правые вентральное и дорсальное положения; 19 — малая ободочная кишка; 20 — прямая кишка.
Иннервируется кишка ветвями блуждающего нерва и чревного сплетения. Кровь получает от ветвей чревной артерии.
У свиньи кишка длиной 40—80 см, лежит в правом подреберье и поясничной области. Направляясь назад, не доходит до подвздошной кости, делает поворот около правой почки и возвращается к печени, где переходит в
тощую кишку. Желчный проток открывается в начале двенадцатиперстной
кишки, а поджелудочный — ближе к середине.
У лошади кишка имеет длину около 1 м, лежит в правом подреберье и
поясничной области. Начальный участок ее несколько расширен. Позади
правой почки кишка поворачивает налево, где и переходит в тощую кишку.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Печеночный и поджелудочный протоки впадают рядом на расстоянии 10—12
см от пилоруса.
Рис 107. Участок слизистой
оболочки двенадцатиперстной
кишки:
А— ворсинка; Б— крипта; I —
эпителий; 1 — каемчатая; 2 —
бокаловидная; 3 — эндокринная
и 4 — бескаемчатая клетки; 5
— клетка Панета; II — собственная пластинка слизистой
оболочки; 6 — кровеносный капилляр: 7— лимфатический капилляр; 8 — гладкомышечные
клетки; III — мышечная пластинка слизистой оболочки.
Тощая кишка — intestinum jejunum — самая длинная и узкая кишка. У
крупного рогатого скота ее длина равна 37—39 м, у овцы — около 25 м. Висит на брыжейке, образуя множество петель и завитков. Располагается в виде
гирлянды вокруг лабиринта ободочной кишки преимущественно в правой
половине брюшной полости: в подреберье подвздошной и паховой областях.
Без резких границ переходит в подвздошную кишку. Иннервируется ветвями
брыжеечного сплетения, васкуляризируется краниальной брыжеечной артерией.
У свиньи кишка длиной 15—20 м висит на длинной брыжейке, легко
смещаема, занимает все свободные пространства в брюшной полости между
печенью и ободочной кишкой. Обследовать ее лучше в подреберьях. Кроме
того, ее петли заходят в область мечевидного отростка, пупочную, подвздошные и паховые области.
У лошади кишка длиной 20—30 м, шириной 6—7 см. Висит на длинной
брыжейке (до 50 см), располагаясь в чашеобразном углублении, образованном большой ободочной и слепой кишками. Обследовать лучше в подреберьях и левой подвздошной области.
Подвздошная кишка—intestinum ilium — короткая, лежит в правой
подвздошной области. У сельскохозяйственных животных ее длина составwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ляет около 50 см. Подвешена на короткой брыжейке. Начинается от последнего витка тощей кишки и заканчивается при впадении в толстый кишечник
на границе слепой и ободочной кишок. У лошади впадает в головку слепой
кишки. Иннервируется ветвями брыжеечного сплетения, васкуляризируется
ветвями краниальной брыжеечной артерии.
Гистологическое строение тонкого кишечника. Кишки тонкого отдела — трубкообразные органы. Их стенка образована слизистой оболочкой,
подслизистой основой, мышечной и серозной оболочками.
Слизистая оболочка состоит из трех слоев: эпителия, собственной пластинки и мышечной пластинки. Эпителий, выстилающий просвет кишок, однослойный цилиндрический. В состав его входит несколько разновидностей
клеток. Наиболее распространенными являются каемчатые — всасывающие
и бокаловидные, вырабатывающие слизь.
Собственная пластинка слизистой оболочки образована рыхлой волокнистой соединительной и ретикулярной тканями с большим количеством
разнообразных клеток. В ней распространены как клетки фибробластического ряда (фибробласты, гистиоциты), так и клетки крови (лимфоциты и другие
лейкоциты). В ней разветвляется большое количество кровеносных и лимфатических сосудов, образующих сети, и нервных волокон, а также расположены общекишечные железы (крипты). В собственной пластинке много
лимфоцитов и лимфатических фолликулов. Одиночные или солидарные
лимфатические фолликулы микроскопических размеров. Скопления лимфатических фолликулов — лимфатические (пейеровы) бляшки — выпуклые
макроскопические образования, имеющие длину от нескольких сантиметров
до нескольких метров.
Мышечная пластинка слизистой оболочки состоит из гладкой мышечной ткани, пучки которой образуют 2 слоя: кольцевой и продольный.
Рельеф слизистой оболочки негладкий. Она собирается в складки в результате сокращения мышечной пластинки и мышечной оболочки, которые
расправляются при прохождении пищи. Образуются нерасправляющиеся
складки и многочисленные выпячивания— ворсинки, во много раз увеличивающие поверхность слизистой, что очень важно для интенсификации процессов переваривания и всасывания.
Ворсинка — выпячивание слизистой оболочки, образованное всеми ее
слоями. У крупного рогатого скота листовидной, у остальных сельскохозяйственных животных — пальцевидной формы. На площади в 1 мм2 располагается 25—40 ворсинок высотой 0,4—0,5 мм.
Каждая ворсинка покрыта эпителием, в состав которого входят каемчатые, бокаловидные и эндокринные клетки.
Основной вид клеток — каемчатые (энтероциты с исчерченной каемкой) — цилиндрической формы, плотно присоединены друг к другу замыкающими (плотными) контактами, чем изолируется внутренняя среда организма от полости кишки. Удлиненное ядро сдвинуто к базальному полюсу.
На апикальном полюсе имеется несколько сотен микроворсинок, которые в
совокупности с надмембранным комплексом клетки (гликокаликсом) формиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
руют исчерченную (щеточную) каемку. Она почти в 30 раз увеличивает всасывающую поверхность клетки. Кроме того, в ней обнаружено большое количество ферментов, участвующих в расщеплении и транспорте всасывающихся веществ (фосфатазы, аминопептидазы, глюкозидазы и др.), благодаря
чему эти процессы наиболее активно идут на микроворсинках исчерченной
каемки. Такое пищеварение называется пристеночным в отличие от полостного, протекающего в просвете кишки. В каемчатых энтероцитах хорошо выражен субмембранный комплекс из микрофиламентов и микрофибрилл. Он, видимо, способствует транспортировке всосавшихся веществ в
глубь клетки. В более глубоких частях цитоплазмы хорошо развита гранулярная цитоплазматическая сеть, много лизосом. Митохондрии локализуются
в основном в базальной части клетки.
Бокаловидные клетки имеют типичное строение (см. железистый эпителий). Они вырабатывают слизь, которая увлажняет внутреннюю поверхность
кишечника и тем способствует продвижению пищевых масс. Количество бокаловидных клеток увеличивается от двенадцатиперстной к подвздошной
кишке.
Эндокринные (аргентаффинные) клетки распределены неравномерно.
Встречаются по всему эпителиальному пласту слизистой оболочки. Они бывают нескольких типов: секретирующие серотонин, энтерогастрин, холецистокинин и другие биологически активные вещества, регулирующие деятельность пищеварительной системы.
Эпителий ворсинки отделен от подлежащей стромы базальной мембраной. Строма ворсинки образована рыхлой соединительной и ретикулярной
тканью. Под эпителием находится сосудистая сеть, в капилляры которой поступают вещества из каемчатых энтероцитов. В строме вдоль ворсинки проходят пучки гладкомышечных клеток, а в ее середине — лимфатический капилляр— синус (у овцы — их несколько). При сокращении мышечных клеток
ворсинка меняет свою форму, в результате чего перемешивается пищевая
масса и новые ее порции вступают в контакт с исчерченной каемкой, а также
проталкивается в капиллярах кровь и лимфа, вынося поступившие в них питательные вещества за пределы ворсинки.
Кишечные крипты (общекишечные железы)—углубления эпителия слизистой оболочки в виде простых трубчатых, иногда разветвленных желез.
Вокруг одной ворсинки может располагаться несколько крипт. Общее число
их в тонком кишечнике превышает 100 млн. Эпителиальный слой слизистой
оболочки единый для ворсинок и крипт. В криптах встречаются те же виды
клеток, что и в ворсинках. В верхних участках крипт преобладают каемчатые
и встречаются бокаловидные клетки. Глубже расположены бескаемчатые
недифференцированные клетки, среди которых часты митозы. Эти клетки
являются камбиальными для клеток ворсинок и крипт, так как жизненный
цикл кишечного эпителия не превышает 2—3 дней. На дне крипт залегают
эндокринные (аргентаффинные), апикальнозернистые, клетки с ацидофильной зернистостью (Панета) и др.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Подслизистая основа образована рыхлой волокнистой соединительной
тканью, в которой наряду с коллагеновыми встречаются и ретикулярные волокна, образующие трехмерную сеть. В подслизистой основе много сосудов,
развито подслизистое нервное сплетение. В подслизистой основе начального
участка тонкого кишечника залегают железы двенадцатиперстной кишки—
дуоденальные (подслизистые) железы.
Дуоденальные (подслизистые) железы у крупного рогатого скота и лошади развиты на протяжении 4—5 м, у овцы — 60— 70 см, у свиньи — 3—5
м, то есть находятся в стенках двенадцатиперстной кишки и в начале тощей
кишки. Это сложные трубчатые (у жвачных — альвеолярно-трубчатые) железы, вырабатывающие слизистый (у жвачных — смешанный) секрет. У
жвачных железы расположены отдельными пакетами, у свиньи образуют
сплошной слой из плотно лежащих концевых отделов. Концевые отделы могут быть светлыми и темными, из высоких и низких клеток в зависимости от
фазы секреции. Протоки желез открываются на поверхность слизистой либо
самостоятельно, либо в дно крипт.
Мышечная оболочка из гладкой мышечной ткани образует кольцевой и
продольный слои, разделенные соединительнотканной прослойкой с межмышечным нервным сплетением.
Серозная оболочка состоит из рыхлой соединительной ткани и мезотелия, переходит в брыжейки и связки кишок.
Печень — hepar (рис. 108)—самая крупная застенная железа организма,
особенно в эмбриональный период, когда она является органом кроветворения и занимает большую часть брюшной полости. Функции печени разнообразны.
Рис. 108. Схема долей печени с висцеральной
поверхности:
А — крупного рогатого скота; Б — лошади; В —свиньи; 1 — левая доля; 2 — левая латеральная и 3 — левая медиальная доли; 4 — пищевод; 5 — каудальная полая вена; 6 — хвостатая
доля с хвостатым отростком; 7— воротная вена; 4 — печеночная артерия; 9 — печеночный
проток; 10 — желчный проток; 11 — желчный
пузырь; 12 — квадратная доля; 13 — круглая
связка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Как пищеварительная железа, она вырабатывает желчь, которая эмульгирует жиры, омыляет жирные кислоты, усиливает действие ферментов поджелудочной железы. Печень выполняет барьерную функцию, обезвреживая
экзогенные и эндогенные токсины, попадающие в кровь из желудочнокишечного тракта, в том числе ядовитые продукты белкового метаболизма,
превращая их в мочевину. В печени депонируются углеводы, витамины А, Е,
D, К, кровь (до 20%), синтезируются важнейшие белки плазмы крови (фибриноген, глобулины, альбумины, протромбин), фосфопротеины, витамин B12.
В общей сложности печень в организме выполняет свыше 500 функций.
Печень крупного рогатого скота массивна, буро-красного цвета, слабо
поделена на доли (правую и левую). На ней различают выпуклую диафрагмальную поверхность, прилежащую к диафрагме, и вогнутую — висцеральную поверхность, обращенную к внутренностям. Желудок, кишечник и правая почка образуют на ней вдавления. Почти в центре висцеральной поверхности имеется углубление — ворота печени. В области ворот в печень
входят печеночная артерия, воротная вена, нервы, выходят печеночный проток и лимфатические сосуды. Здесь же располагаются лимфатические узлы.
Вентральнее ворот лежит желчный пузырь. От него отходит пузырный проток, при слиянии которого с печеночным протоком образуется желчный
проток, впадающий в двенадцатиперстную кишку.
Медиодорсальный край печени тупой, остальные — острые. На тупом
крае имеются вдавления пищевода и каудальной полой вены, которая врастает
в ткань печени. В нее открываются печеночные вены. На остром вентральном
крае есть неглубокая вырезка, которая отделяет левую долю. У многих животных в этом месте видна запустевшая пупочная вена в виде круглой связки
печени, которая идет к пупку. Между левой и правой долями различают хвостатую долю, расположенную выше ворот, с большим хвостатым отростком, свешивающимся вниз у правого конца печени, и квадратную долю —
участок печени ниже ворот, между круглой связкой и желчным пузырем.
Иннервируется печень ветвями блуждающего нерва и солнечного сплетения. Васкуляризируется печеночной артерией, отходящей от чревной артерии. Расположена в правом подреберье, доходя справа до 2—3-го поясничного позвонка, а слева до грудины. К диафрагме, почке, двенадцатиперстной
кишке печень присоединена связками. С желудком связана малым сальником.
У свиньи печень относительно крупнее (до 2,5%), чем у рогатого скота,
желтоватого цвета. Правая и левая доли отделены друг от друга глубокой
вырезкой. Каждая из них, в свою очередь, делится вырезками на латеральную и медиальную доли. Каудальная полая вена врастает в ткань печени. Ворота печени сдвинуты к дорсальному краю. Квадратная доля треугольной
формы. Хвостатая доля небольшая, хвостатый отросток выражен нечетко.
Почка не касается печени. Расположена печень в правом и левом подреберьях почти поровну. Вентральным краем заходит в область мечевидного отростка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
У лошади левая доля разделена вырезкой на левую латеральную и левую
медиальную доли. Хвостатый отросток сравнительно небольшой. Квадратная
доля слева отграничена круглой связкой, справа — вырезкой. У лошади, как
и у других непарнокопытных (а также верблюда, оленя, многих грызунов и
др.), отсутствует желчный пузырь. Желчь поступает в двенадцатиперстную
кишку по печеночному протоку.
Гистологическое строение печени (рис. 109). Печень — компактный орган. Состоит из соединительнотканной стромы и паренхимы, образованной
эпителием энтодермального происхождения. В печени чрезвычайно развито
сосудистое русло, так как практически все функции печени выполняются в
тесном контакте с кровью. Тонкая соединительнотканная капсула печени
сверху одета серозной оболочкой. В области ворот соединительная ткань
капсулы проникает внутрь органа и разветвляется, деля его на дольки. Из
сельскохозяйственных животных дольчатость печени хорошо видна у свиньи, у остальных животных ее прослеживают благодаря своеобразному закономерному расположению междольковых сосудов: междольковые артерия,
вена и желчный проток лежат рядом, образуя триаду. Располагаются триады
на границе между дольками. Внутридольковая соединительная ткань отсутствует. В дольках при специальной обработке заметна лишь тончайшая сеть
ретикулярных волокон.
Дольки печени полигональной формы, на срезе имеют вид 5—6угольников диаметром 0,7—2 мм. Образованы они печеночными пластинками, состоящими из клеток печени — гепатоцитов. Печеночные пластинки
разделены широкими кровеносными капиллярами — синусоидами, образованными в результате слияния артериальных и венозных капилляров. В центре дольки находится центральная вена дольки, куда стекает кровь из синусоид-ных капилляров. Печеночные пластинки состоят из двух рядов клеток и
имеют радиальное направление. На своем протяжении от периферии дольки
к ее центру печеночные пластинки анастомозируют друг с другом, образуя
сеть, но при этом каждый гепатоцит обязательно контактирует с синусоидным капилляром хотя бы одной своей стороной.
Внутри печеночных пластинок проходят желчные капилляры. У них нет
собственных стенок, ими являются желобки, образованные плазмолеммой
соседних гепатоцитов. Соединяясь друг с другом, желобки образуют тончайшие трубочки, видные только в электронный микроскоп. Участки плазмолеммы гепатоцитов, образующие стенки желчного капилляра, отграничены зоной слипания и плотной зоной, что исключает контакт желчи с кровью.
Желчные капилляры слепо начинаются в центре дольки. Желчь по ним течет
на периферию дольки, где попадает в междольковый желчный выводной
проток, выстланный кубическим эпителием. Междольковые выводные протоки объединяются и образуют печеночный проток.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 109. Схема гистологического строения печени:
А — долька печени; Б —
расположение
желчных
путей и кровеносных сосудов в дольке; В — гепатоцит; 1— печеночная пластинка; 2 — синусоид; 3 —
центральная вена; 4 —
междольковая
соединительная ткань; 5 —
артерия; 6 — вена и 7 —
желчный проток междольковый; 8 — желчный
капилляр; 9 — микроворсинки; 10 — вокругсинусоидное пространство (Диссе); 11—митохондрии; 12
— гранулярная цитоплазматическая сеть; 13 —
комплекс
Гольджи;
14_лизосомы; 15 — агранулярная цитоплазматическая сеть; 16 — эндотелий капилляра; 17 — звездчатая клетка.
Тесный контакт, существующий между паренхимой печени и ее сосудистым руслом, выражается в особенностях их структуры. Между гепатоцитами и эндотелием капилляров отсутствует базальная мембрана и имеется воwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
кругсинусоидное пространство (Диссе), заполненное тканевой жидкостью.
Здесь происходит обмен веществ между кровью и гепатоцитами. В него направлены многочисленные микроворсинки гепатоцитов, достигающие эндотелия.
В состав эндотелия синусоидных капилляров кроме обычных эндотелиальных клеток входят отростчатые звездчатые ретикулоэндотелиоциты
(купферовские клетки) — видоизмененные макрофаги крови, вошедшие в состав стенки капилляра. Они являются макрофагами печени. С ними прежде
всего контактируют микробы, токсины, чужеродные вещества, попавшие в
кровь. Фагоцитируя их, они затем передают гепатоцитам (через пространство
Диссе) продукты их переработки для дальнейшей утилизации или использования в процессах обмена. Активизируясь, звездчатые клетки увеличиваются
в размерах, вычленяются из эндотелиального пласта, их отростки глубоко
проникают в просвет капилляра и пространство Диссе.
Гепатоциты — полигональные клетки с крупным круглым ядром, лежащим в центре. Встречаются и двухъядерные клетки. У рогатого скота и лошади их количество невелико (8%), у свиньи достигает 40%. С возрастом количество двухъядерных и полиплоидных гепатоцитов увеличивается до 80%.
В цитоплазме развиты все органеллы общего значения. Особенно многочисленны митохондрии, количество которых доходит до 2500, свободные и связанные рибосомы, цистерны цитоплазматической сети. В гепатоцитах накапливаются различные включения: гликоген, жир, витамины и др. Существует
определенный цикл функциональной активности и направленности: днем
преобладает выделение желчи, ночью — синтез гликогена; гликоген откладывается сначала в центре дольки, а затем на периферии, а жир — наоборот.
В гепатоцитах различают 2 функциональных полюса: желчный, обращенный
к желчному капилляру, и сосудистый, контактирующий с синусоидным кровеносным капилляром. На обоих полюсах имеются микроворсинки.
Кровообращение в печени. В печень входят и в ней параллельно разветвляются воротная вена, несущая кровь от органов пищеварения и селезенки, и печеночная артерия. Между дольками их ветви образуют междольковые вены и артерии. Вместе с междольковыми желчными протоками они образуют триады. От междольковых сосудов на разном уровне отходят вокругдольковые артерии и вены, разветвляющиеся на капилляры. Капилляры
входят внутрь долек и сливаются друг с другом, образуя широкие синусоидные капилляры, несущие смешанную кровь. Кровь медленно движется от периферии дольки к ее центру. При этом состав ее меняется, а печень выполняет свои многочисленные функции. В центре кровь сливается в центральную
вену дольки. Таким образом, в печени капилляры оказываются между двумя
венами. Такое явление называется чудесной сетью печени. Из центральной
вены дольки кровь стекает в поддольковую вену. Последние объединяются и
через ряд сосудов впадают в печеночные вены, открывающиеся в каудальную
полую вену. В сосудах печени имеются сфинктеры, которые регулируют ток
крови в ней. Ток крови от вен кишечника, желудка, поджелудочной железы и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
селезенки через печень до каудальной полой вены называется воротным кровообращением.
Поджелудочная железа — pancreas (рис. 110)—относится к железам с
двойной секрецией — внешней и внутренней.
Как железа внешней секреции, она вырабатывает поджелудочный (панкреатический) сок, содержащий трипсин, хемотрипсин, карбоксипептидазу,
рибонуклеазу, липазу и другие ферменты, расщепляющие белки, жиры и углеводы корма. Как железа внутренней секреции, она вырабатывает гормоны
(инсулин, глюкагон, липокаин), регулирующие углеводный обмен, участвующие в регуляции белкового и жирового обмена. Внешнесекреторная
часть железы составляет 97% ее массы и поэтому определяет ее размеры и
форму.
У крупного рогатого скота железа желто-бурого цвета, имеет бугристую
поверхность и состоит из тела (средней части), левой и правой долей. Расположена она в правом подреберье и в правой части поясничной области. Тело
и правая доля лежат вдоль двенадцатиперстной кишки. Левая доля отходит
влево и прилегает к рубцу. Выводной проток открывается в конце двенадцатиперстной кишки. Между долями проходит каудальная полая вена. Иннервируется железа блуждающим и симпатическими нервами. Кровь получает
от ветвей чревной артерии.
У свиньи правая доля тянется по двенадцатиперстной кишке до правой
почки, левая — прилегает к селезенке и левой почке, так что железа лежит в
обоих подреберьях и заходит в поясничную область. Выводной проток открывается в двенадцатиперстную кишку на расстоянии 15—25 см от пилоруса.
У лошади железа серовато-розового цвета. Лежит в обоих подреберьях.
Тело расположено в S-образном изгибе двенадцатиперстной кишки. Правая
доля слабо отделена от тела, левая лежит на малой кривизне желудка. Главный выводной проток открывается в двенадцатиперстную кишку вместе с
печеночным протоком. Иногда встречается добавочный выводной проток,
который открывается отдельно.
Гистологическое строение. Поджелудочная железа — компактный орган, сложная застенная альвеолярная железа. Сверху покрыта серозной оболочкой. Соединительнотканная строма образует тонкую капсулу, от которой
отходят междольковые прослойки соединительной ткани. Внутридольковые
прослойки из рыхлой соединительной ткани развиты слабо. В строме проходят сосуды и нервы, имеются мелкие вегетативные ганглии (скопления нервных клеток) и нервные окончания.
Экзокринная паренхима образована эпителием энто-дермального происхождения и состоит из концевых отделов и выводных протоков.
Концевые отделы имеют форму альвеол, образованы однослойным
призматическим эпителием. Обычно в самый мелкий выводной проток, который называется вставочным протоком, открывается несколько альвеол. Такая структура называется ацинусом. Экзокринная часть железы, состоящая из
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ацинусов, носит название ацинозной паренхимы, а клетки, ее составляющие,— ацинозные.
Рис. 110. Поджелудочная железа крупного рогатого скота:
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
А — внешний вид; Б — гистологическое строение участка дольки железы; 1
— двенадцатиперстная кишка; 2 — правая доля поджелудочной железы; 3
—тело; 4 — левая доля; 5 — воротная вена; 6 — краниальная брыжеечная
артерия; 7— альвеола; 8 —ацинозная клетка; 9 — зимогенная и 10 — базофильная зоны; 11—вставочный проток; 12 — центроацинозная клетка; 13
— внутридольковая и 14 — междольковая соединительная ткань; 15 —
внутридольковый и 16 — междольковый выводные протоки;17 - островок;
18 – В – клетка; 19 — А-клетка; 20 — капилляр.
Ацинозная клетка имеет четко выраженную апикобазальную дифференцировку. Ее базальная, более расширенная часть заполнена массой плоских
мешочков гранулярной цитоплазматической сети, воспринимает основные
красители, за что называется базофильной, или гомогенной, зоной. Круглое
ядро сдвинуто к базальной части клетки. Над ним расположен комплекс
Гольджи. В апикальном полюсе клетки накапливаются крупные округлые
гранулы секрета — зимоген, в который входят разнообразные ферменты в
неактивной форме. Гранулы окрашиваются кислыми красителями, за что эта
часть клетки названа оксифильной, или зимогенной, зоной. Секрет выделяется через апикальный полюс мерокринным способом и попадает в систему
выводных протоков.
Выводной проток наименьшего диаметра назван вставочным, так как
вдавливается на разную глубину в альвеолы. В результате клетки его стенки
оказываются как бы вставленными в центр ацинуса и потому называются
центроацинозными. На срезах их удлиненные светлые ядра можно видеть
лежащими внутри альвеол. Диаметр вставочных протоков меньше альвеол.
Их стенка образована уплощенными клетками со светлой цитоплазмой, бедной органеллами. Вставочные протоки объединяются сначала в мелкие внутридольковые выводные протоки, затем в более крупные внутридольковые
протоки и, наконец, в междольковые выводные протоки. Последние образуют общий выводной проток поджелудочной железы, идущий к двенадцатиперстной кишке. По мере увеличения диаметра протока высота его эпителия
увеличивается, а окружающие их прослойки соединительной ткани утолщаются и уплотняются.
Эндокринная паренхима составляет около 3% массы поджелудочной
железы, представлена панкреатическими островками (Лангерганса). Они
имеют неправильно-округлую, эллипсоидную или отростчатую форму, хорошо снабжаются кровью. В островке обычно можно видеть несколько капилляров. Состоят островки из клеток нескольких типов, вырабатывающих
разные гормоны. Друг от друга отличаются только при специальной обработке. Островковые клетки мельче ацинозных и светлее. Все они содержат
мелкие митохондрии, хорошо развитый комплекс Гольджи и секреторные
гранулы, характерные для каждого вида клеток.
Самые многочисленные — до 70—75%—В- или β-клетки. Специальными красителями эти клетки окрашиваются базофильно. Их зернистость растворима в спирте и нерастворима в воде. Вырабатываемый ими инсулин споwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
собствует утилизации глюкозы тканями и превращению ее в гликоген, чем
снижает уровень глюкозы в крови. А- или α-клетки составляют 20—25%. Их
гранулы при специальной обработке окрашиваются оксифильно, растворимы
в воде и нерастворимы в спирте. Эти клетки вырабатывают глюкагон — гормон, противоположный по действию инсулину. Он способствует расщеплению гликогена до глюкозы и выведению ее в кровь. Среди остальных клеток
различают еще несколько типов, вырабатывающих гормоны, по-видимому,
регулирующие белковый и липидный обмены. Предполагают, что среди этих
клеток находятся и камбиальные элементы. Вместе с тем на границе островков найдены переходные формы клеток — островково-ацинозные. В них есть
как зимогенные гранулы, так и гранулы, характерные для эндокринных клеток. Возможно, эти клетки также являются источником пополнения клеток
островков. У островков нет оболочек, отделяющих их от экзокринной паренхимы.
ЗАДНЯЯ КИШКА, ИЛИ ТОЛСТЫЙ КИШЕЧНИК
Толстый кишечник у сельскохозяйственных животных в среднем в 4
раза короче тонкого. У крупного рогатого скота его длина достигает 11 м, у
овцы — 7, у лошади — 9, у свиньи — 4 м. На всем протяжении он имеет неодинаковый диаметр. В его состав входят слепая, ободочная и прямая кишки
(см. рис. 106). В толстом кишечнике происходит всасывание в основном воды и растворенных в ней солей, а также формирование каловых масс. У некоторых травоядных с объемистым кишечником (лошадь) идут и процессы
расщепления корма с участием обитающей здесь микрофлоры.
Слепая кишка — intestinum caecum — у крупного рогатого скота имеет
длину 30—70 см и наибольший по сравнению с другими кишками диаметр.
Она гладкая, цилиндрической формы. Ее слепой конец называется верхушкой, остальная часть — телом. Концом слепой кишки считается место впадения подвздошной кишки в толстый кишечник. Расположена кишка в правой
половине брюшной полости над ободочной кишкой. Ее вершина направлена
каудально и заходит в паховую область.
Тело лежит в правой подвздошной и поясничной областях. Иннервируется ветвями блуждающего нерва и краниального брыжеечного сплетения. Васкуляризация краниальной брыжеечной артерией.
У свиньи слепая кишка короткая и широкая. В ее стенке проходят 3 продольных мышечных тяжа — тении, которые собирают стенку в поперечные
складки и карманы. Располагается кишка в поясничной области, касаясь телом левой почки. Верхушкой направлена каудально и вправо.
У лошади кишка в форме запятой огромных размеров. В ней различают
краниовентрально расположенную верхушку, тело и наиболее широкое, каудодорсально направленное основание (головку), на нем вогнутую малую кривизну и выпуклую большую кривизну. Верхушка и основание оканчиваются
слепо. У верхушки конец конусовидный, у основания округлый. На дорсальной поверхности малой кривизны открываются 2 отверстия: меньшее веwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
дет из подвздошной кишки в слепую, большее — из слепой в ободочную. В
стенке слепой кишки имеются 4 тении и ряды карманов.
Расположена она верхушкой в области мечевидного отростка, отделяясь
от него ободочной кишкой. Тело и основание — в пупочной, поясничной,
правой подвздошной и правой паховой областях. Телом слепая кишка соприкасается с вентральной брюшной стенкой, а ее основание впереди доходит до
печени, сзади до входа в таз, сверху прилегает к поясничной мускулатуре,
срастаясь с ней рыхлой соединительной тканью (адвентицией).
Ободочная кишка — intestinum colon — у крупного рогатого скота длиной 6—9 м, гладкая, небольшого диаметра, имеет форму диска. В ней различают начальную петлю, спиральный лабиринт и конечную петлю. Началом
ее считается место впадения подвздошной кишки в толстый кишечник. Отсюда начальная или проксимальная петля идет сначала вперед, затем назад, а
потом снова вперед и переходит в спиральный лабиринт (спиральную петлю). Последний закручивается на 1,5—2 оборота к центру — центрипетальная петля, затем на столько же оборотов от центра — центрифугальная петля и после этого переходит в конечную (дистальную) петлю. Она направляется назад, потом вперед и снова назад — без видимой границы переходит в
прямую кишку. У мелкого рогатого скота лабиринт делает по 3 оборота в ту
и другую сторону, и его петли расположены ступенчато, так что ободочная
кишка принимает форму дискоко-нуса. Расположена кишка в правой половине брюшной полости, заходя в правое подреберье, поясничную, пупочную и
правую подвздошную области. Диск ее лежит вертикально, левой стороной
соприкасаясь с правой поверхностью рубца. Иннервируется брыжеечными
сплетениями, содержащими ветви симпатических и парасимпатических нервов. Васкуляризируется ветвями краниальной брыжеечной артерии, а конечный участок — каудальной брыжеечной артерией.
У свиньи — ободочная кишка в форме конуса, основанием направленного каудодорсально, а вершиной — краниовентрально. Конус состоит из двух
петель. Наружная — центрипетальная петля начинается от места впадения
подвздошной кишки в толстый кишечник и идет от основания конуса к его
вершине. Она имеет широкий диаметр, 2 тении и 2 ряда карманов. У вершин
конуса диаметр кишки уменьшается, тении исчезают. Кишка переходит в
центрифугальную петлю, гладкую и узкую, идущую внутри конуса от вершины к основанию. Выйдя из конуса, ободочная кишка делает конечную
петлю, которая идет вперед до желудка, потом поворачивает назад и переходит в прямую кишку. Расположена кишка в левой половине брюшной полости, основанием прикрепляется к поясничной мускулатуре, а вершиной доходит до брюшной стенки в области мечевидного отростка — у молодняка или
в пупочной области — у взрослых животных.
У лошади ободочная кишка самая крупная по своим размерам, в форме
двойной подковы. Диаметр ее в разных местах неодинаков, в связи с чем ее
делят на большую и малую ободочные кишки. Большая ободочная кишка
очень широкого диаметра, имеет тении и карманы. Начинается кишка в правой подвздошной области правым вентральным положением. Направляясь
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
вперед в области мечевидного отростка, переходит в диафрагмальное вентральное положение, прилежащее к диафрагме, а затем в левую подвздошную область в виде левого вентрального положения, проходит через левую
паховую область до входа в таз, здесь заворачивается вверх, сильно сужается.
После этого расширяется и ложится на вентральные положения, повторяя ход
в обратном направлении: левое дорсальное положение, диафрагмальное дорсальное положение и правое дорсальное положение. Вентральные положения
шире, несут 2 тении и 4 ряда карманов. На дорсальных положениях 3 тении и
3 ряда карманов. В поясничной области, значительно сужаясь, кишка переходит в малую (нисходящую) ободочную. Ее диаметр в несколько раз меньше,
чем у большой ободочной. Она имеет 2 тении и 2 ряда карманов, висит на
длинной брыжейке и занимает свободное пространство в поясничной, левой
подвздошной, лонной и левой паховой областях.
Прямая кишка — intestinum rectum — короткий прямой участок толстого кишечника, расположенный в тазовой полости. Краниальная часть
кишки подвешена на короткой брыжейке, каудальная — одета адвентицией,
соединяющей ее с окружающими органами. В конце прямой кишки у свиньи
и лошади есть ампулообразное расширение. Заканчивается прямая кишка
анусом — заднепроходным отверстием. Прямая кишка и анус соединены
мышцами с костями таза. Они помогают при дефекации изгнанию кала и возвращению кишки в исходное положение. В стенке ануса имеются мышцы,
образующие его сфинктер. Иннервируется тазовым сплетением, васкуляризируется артериями прямой кишки, отходящими от брюшной аорты и внутренней подвздошной артерии.
Гистологическое строение толстого кишечника. Как у всяких трубкообразных органов, у кишок толстого кишечника стенка состоит из слизистой
оболочки, подслизистой основы, мышечной и серозной оболочек. Основная
особенность слизистой оболочки — отсутствие ворсинок. Эпителий у нее
однослойный цилиндрический каемчатый, за исключением конечного участка прямой кишки, где он становится многослойным плоским и происходит из
эктодермы. Кишечные крипты глубже и лежат чаще, чем в тонком кишечнике. Они построены из таких же видов клеток, но преобладают бокаловидные.
Вырабатываемая ими слизь способствует эвакуации более плотных масс, заполняющих толстый кишечник. В собственной пластинке слизистой оболочки имеются большие скопления лимфоидной ткани в виде солитарных и агрегатных (пейеровых бляшек) лимфоузлов. Мышечная пластинка представлена кольцевым и продольным слоями, она толще, чем в тонком кишечнике.
Подслизистая основа также отличается сильной инфильтрацией лимфоидными элементами. Мышечная оболочка состоит из кольцевого и продольного
слоев, образованных пучками гладкомышечных клеток. Там, где стенка кишок толстого кишечника гладкая, слои мышечной оболочки расположены
равномерно. В кишках, имеющих тении, кольцевой слой развит равномерно,
а продольный формирует тении — лентовидные утолщения, идущие вдоль
кишки. В промежутках между тениями продольный слой развит слабо. В конечном участке прямой кишки мышечная оболочка образует ряд мышц и
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
сфинктер ануса. Серозная оболочка без особенностей. В прямой кишке за
пределами тазовой полости и на некоторых участках (экстраперитонеальных)
ободочной кишки наружной оболочкой является адвентиция.
Вопросы для самоконтроля. 1. Этапы развития пищеварительной системы в фило- и в онтогенезе. 2. Что входит в состав ротоглотки и каково макро- и микроскопическое строение ее органов? 3. Строение пищевода — типичного трубкообразного органа. 4. Каковы особенности макро- и микроскопического строения желудка всеядных, травоядных и жвачных? 5. Состав
и расположение кишечника у разных видов сельскохозяйственных животных. 6. Изменения в строении стенки кишечной трубки на протяжении от
двенадцатиперстной кишки до ануса. 7. Макро- и микроскопическое строение печени, особенности ее кровоснабжения. 8. Макро- и микроскопическое
строение и топография поджелудочной железы у разных видов животных.
Глава 16. СИСТЕМА ОРГАНОВ ДЫХАНИЯ (АППАРАТ ДЫХАНИЯ)
Система органов дыхания состоит из воздухоносных путей (нос с носовой полостью, гортань, трахея и бронхи легкого) и респираторных отделов
(ацинусы легкого и альвеолы). Она осуществляет газообмен между организмом и окружающей средой, так называемое внешнее дыхание. В процессе
этого дыхания происходит поступление кислорода из окружающей среды в
кровь, а СО2 в обратном направлении. У млекопитающих и птиц внешнее
дыхание осуществляется легкими, у рыб — жабрами. Внутреннее, или тканевое, дыхание происходит в клетках в виде окислительных реакций, в которых
участвует кислород, приносимый к тканям артериальной кровью.
Кроме газообмена, аппарат дыхания выполняет и другие функции: очищает вдыхаемый воздух от пыли, микроорганизмов, согревает и увлажняет
его; участвует в терморегуляции, водно-солевом обмене, иммунологической
защите. В гортани расположен голосовой аппарат, в носовой полости — орган обоняния и орган регуляции осмотического давления — латеральная носовая солевая железа, особенно развитая у морских животных.
КРАТКИЕ СВЕДЕНИЯ О РАЗВИТИИ СИСТЕМЫ ОРГАНОВ
ДЫХАНИЯ
Филогенез. Дыхание в процессе эволюции усложнялось и совершенствовалось, как и органы газообмена. Более простое — диффузное дыхание. Осуществляется она всей поверхностью тела без участия кровеносной системы.
Свойственно одноклеточным и наиболее примитивным многоклеточным
(губки, кишечнополостные). С усложнением организма и развитием кровеносной системы появились 2 разновидности диффузного дыхания: кожное и
кишечное.
Кожное дыхание происходит всей поверхностью тела с участием кровеносной системы. Обязательное условие для осуществления кожного дыхания
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
— нежные и влажные покровы. Такой тип дыхания свойствен многим беспозвоночным с тонким эктодермальным покровом: планариям, нематодам, личинкам многих насекомых. Кожное дыхание сохраняет свое значение и у
многих позвоночных: у разных видов земноводных на его долю приходится
от 15 до 55% потребляемого кислорода. У большинства млекопитающих оно
не превышает 1%, но у лошади при энергичной работе возрастает до 8%.
Кишечное дыхание осуществляется энтодермальной выстилкой кишечной трубки с участием сильно развитой сосудистой сети кишки. Этот тип
дыхания основной для таких водных животных, как кишечнодышащие. Сохраняется в различной степени у некоторых рыб: сом, вьюн. У земноводных
на его долю приходится 10—15% (через слизистую оболочку ротоглотки) .
Возможности всех разновидностей диффузного дыхания ограничиваются двумя факторами: размерами организма и интенсивностью его жизнедеятельности. С увеличением размера поверхность тела увеличивается медленнее» чем его объем, в результате все больше снижается удельная поверхность
тела (часть площади, приходящаяся на единицу объема) и газообмен оказывается недостаточным для поддержания жизнедеятельности крупного организма. Увеличение интенсивности обменных процессов тоже требует более
интенсивного дыхания, чем могут обеспечить разные формы диффузного дыхания. При интенсивных проявлениях жизни и крупных размерах тела необходимо увеличение площади соприкосновения со средой, несущей кислород
(с водой или воздухом), и приспособления для принудительного продвижения этой среды. Этим условиям отвечает специализированное местное дыхание, которое встречается в двух формах: жаберное и легочное.
Жаберное дыхание свойственно первичноводным хордовым. Наибольшего совершенства достигает у костистых рыб. Оно встречается и у земноводных на личиночной стадии развития. Жабры являются выростами головной кишки — ротоглотки. Наиболее примитивные жабры у ланцетника:
большое количество (до 150) узких открытых щелей, в стенках которых
имеются кровеносные сосуды. Вода, проходя через щели, обменивается газами (02 и COj) с кровью жабр. Совершенствование жаберного дыхания шло по
пути уменьшения количества (до 4—5 пар у костных рыб) и усложнения
структуры жабр. Каждая жабра у костистых рыб состоит из двух полужабр с
большим количеством лепестков. Они ярко-розового или красного цвета изза большого количества поверхностно расположенных сосудов.
Легочное дыхание развивается с выходом на сушу. Зачатки этого типа
дыхания встречаем у кистеперых и двоякодышащих рыб, у которых парный
плавательный пузырь открывается в вентральной стенке пищевода и служит
наряду с жабрами органом дыхания, особенно в период спячки во время пересыхания водоемов (6—9 месяцев в году). В стенках такого плавательного
пузыря разветвляются сосуды. Рыба заглатывает воздух, который из ротоглотки по пищеводу попадает в видоизмененный плавательный пузырь, где и
происходит газообмен. У земноводных через легкие поступает от 35 до 75%
кислорода. Их легкие имеют вид простых тонкостенных мешков, разделенных септами на участки. Поверхность таких легких меньше поверхности теwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ла. На базе органа обоняния образуется примитивная носовая полость, а наружные отверстия обонятельных ямок становятся ноздрями. Обонятельные
ямки, которые у большинства рыб не соединены с ротовой полостью, у земноводных открываются в ее начальный участок отверстиями — первичными
хоанами. Через них воздух попадает в ротоглотку, В ротоглотке имеется
щель, ведущая в гортанно-трахейную камеру, стенки которой образованы
двумя черпаловидными, одним кольцевидным и несколькими трахейными
хрящами. Затем воздух поступает в легкие, являющиеся выростами задней
части глотки.
Рис. 111. Онтогенез дыхательной системы у млекопитающих:
А — носовая полость зародыша; Б, В — две фазы развития легких свиньи; 1
— лобный валик; 2 — зачаток глаза; 3— зачаток ноздри; 4 — носовые отростки; 5 — челюстной отросток; 6 — первичная ротовая полость; 7 — первая жаберная дуга; 8 — первая жаберная щель; 9— вторая жаберная дуга;
10— четвертый жаберный карман; 11 — зачаток пищевода; 12 — зачаток
трахеи; 13 — закладка легких; 14 — бронхиальное дерево; 15 — легочная артерия; 16 — плевра.
Дальнейшее разделение дыхательного и пищеварительного путей происходит с развитием твердого и мягкого нёба. При этом хоаны все больше
отодвигаются назад и называются вторичные, или задние, хоаны, что можно
наблюдать у некоторых рептилий (крокодилы) и у всех млекопитающих. Наличие твердого нёба позволяет дышать во время пережевывания пищи. Носовая полость увеличилась, в ней появились носовые раковины. Усложнилось и
строение гортани. У млекопитающих она образована пятью хрящами. С развитием шеи удлиняется трахея. Легкие в ряду позвоночных прогрессивно усложняются: увеличивается количество перегородок, ячей, появляются альвеолы (у млекопитающих). В стенках этих образований внутрь легких врастают
сосуды, увеличивается поверхность соприкосновения с воздухом, особенно у
млекопитающих, у которых она в 50—100 раз превышает поверхность тела.
Параллельно с изменением строения аппарата дыхания изменяется и дыхательный механизм. Сначала он осуществляется мышцами плавательного
пузыря, затем мышцами ротоглотки (у земноводных), а в дальнейшем —
мышцами грудной и брюшной стенок (у рептилий и млекопитающих).
Онтогенез. Органы дыхания закладываются в период дифференцировкн
кишечной трубки (рис. 111). Ротовая ямка, обрамленная пятью валиками
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(лобными и парными челюстными), в процессе дифференцировкн превращается в носовую и ротовую полость (А). Передние концы верхнечелюстных
отростков сближаются, формируя твердое нёбо и отделяя эти полости друг от
друга. Параллельно с этим в зародышевый и предплодный период закладываются и остальные органы дыхания из вентральной стенки головной кишки
в области глотки. Позади последней (IV) жаберной дуги на 23-й день развития теленка отрастает непарный бугорок. Он удлиняется, и нз его передней
части развиваются гортань и трахея. Его задняя часть раздваивается на 2
вздутия, из которых развиваются легкие. Из энтодермального выроста кишки
формируется эпителиальная выстилка гортани, трахеи, легких. Из мезенхимы
дифференцируется межуточная (интерстициальная) ткань, эластические волокна, мышечные элементы и сосуды. Оба источника — энтодермальный и
мезенхимный — образуют единую бронхолегочную закладку. Сначала развивается бронхиальное дерево, затем альвеолярное. Увеличиваясь и разветвляясь, закладка легких проникает в грудную полость и вступает в контакт с
висцеральным листком плевры (В). Затем идет образование долей, сегментация легких, присущая определенному виду животных.
В течение плодного периода масса легких увеличивается в 150—200 раз.
Особенно интенсивно растут легкие в последний месяц внутриутробного
развития. Формирование и перестройка бронхиального дерева и респираторных отделов происходят и после рождения. В соответствии с меняющимися
функциональными потребностями увеличивается или уменьшается числе
альвеол на респираторных бронхиолах. В постнатальном онтогенезе масса
легких возрастает в 4—6 раз, у взрослого крупного рогатого скота равна 3—
4,5 кг и не зависит от породы, конституции, пола. Это говорит о том, что легкие — рано закладывающийся и рано созревающий орган.
НОС И НОСОВАЯ ПОЛОСТЬ
Нос — nasus (см. рис.97) —начальный участок дыхательных путей. В
нем воздух согревается, увлажняется, анализируется на запах. Нос имеет
спинку, боковые стенки, кончик (верхушку) и корень, в основе которых лежат
кости черепа. В спинке — носовая и частично лобная, в боковых стенках —
верхнечелюстная и резцовая. Границей между носом и черепно-мозговой по
лостью служит продырявленная пластинка решетчатой кости
От ротовой полости носовая полость отделена нёбными отростками
верхнечелюстных, резцовых и нёбными костями.
Снаружи нос покрыт кожей с волосами, лишь на кончике носа волос нет
(не у лошадей). Кожа здесь толстая, пигментированная, влажная и прохладная от секрета серозно-слизистых желез, расположенных в дерме. Этот участок называется носовым зеркальцем у мелкого рогатого скота, носогубным
зеркальцем у крупного рогатого скота, хоботковым зеркальцем — у свиньи.
У свиньи в кончике носа — пятачке — заключена хоботковая кость.
Носовая полость — cavum nasi — имеет 2 входных отверстия— ноздри
и два выходных — хоаны. В стенках ноздрей лежат хрящи, благодаря котоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
рым ноздри сохраняют зияние. Подвижность ноздрей обеспечивается специальными мышцами, которые хорошо развиты у лошадей. У последних латеральнее каждой ноздри расположен кожистый карман глубиной до 7 см —
носовой дивертикул.
Внутри полость носа разделена продольной хрящевой носовой перегородкой, которая является продолжением в назальном направлении перпендикулярной пластинки решетчатой кости. В каждой половине носовой полости
имеется по 2 носовые раковины: — более узкая — дорсальная и более широкая — вентральная. В задневерхнем участке носовой полости расположен
лабиринт решетчатой кости.
Пространство носовой полости раковинами и перегородками разделяется на 4 хода. Дорсальный (обонятельный) носовой ход расположен между
крышей носовой полости и дорсальной носовой раковиной. По нему воздух
идет к лабиринту решетчатой кости, где анализируется на запах (см. Органы
чувств). Средний носовой ход проходит между носовыми раковинами. Он ведет в си-лусы (пазухи) черепных костей (см. Скелет головы) и называется синусным. В синусах воздух увлажняется и согревается. Вентральный носовой
ход проходит между вентральной носовой раковиной и дном носовой полости. Он ведет в хоаны и является собственно дыхательным. Общий носовой
ход имеет вид узкой цели между носовой перегородкой и раковинами. По
нему воздух идет во всех направлениях, так как он сообщается с остальными
ходами.
Стенки носовой полости, носовые раковины, лабиринт решетчатой кости и носовая перегородка покрыты слизистой оболочкой. У входа в носовую
полость — в преддверии носовой полоти— слизистая оболочка выстлана
многослойным плоским эпителием, в области лабиринта решетчатой кости
— обонятельным эпителием желтоватого цвета, в остальных участках —
многорядным мерцательным эпителием розового цвета. В собственной пластинке слизистой оболочки залегают многочисленные слизистые и серозные
железы, секрет которых, смешиваясь с секретом бокаловидных клеток, способствует прилипанию пыли и других частиц. Сюда же по носо-слезному
протоку стекают слезы, омывающие глаз. Реснички мерцательного эпителия,
двигаясь, создают ток этой жидкости в направлении носоглотки. Общее количество ее может достигать 500 мл в сутки. В собственной пластинке слизистой оболочки разветвляется большое количество сосудов, что способствует
лучшему согреванию воздуха. В слизистой оболочке носа встречаются лимфатические фолликулы, количества которых увеличивается в носоглотке. У
овцы и свиньи в слизистой оболочке среднего носового хода залегает латеральная носовая солевая железа — сложная, трубчатая железа, участвующая
в регуляции водно-солевого обмена. Глубокие слои слизистой оболочки, богатые коллагеновыми и эластическими волокнами, переходят в надкостницу
или надхрящницу.
Пройдя носовую полость, воздух попадает в глотку, а оттуда— в гортань.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ГОРТАНЬ
Гортань — larynx (рис. 112) —лежит под глоткой близ углов нижней
челюсти. Гортань проводит воздух в трахею, не допускает попадания пищи в
дыхательные пути и содержит голосовой аппарат. Состоит гортань из 5 хрящей: надгортанника, щитовидного, двух черпаловидных и кольцевидного.
Хрящи между собой и с подъязычной костью соединены суставами и связками (Б). Движениями гортани, ее расширением и сужением управляют мышцы
гортани и тесно с ней связанной подъязычной кости. Надгортанник, закрывающий вход в гортань при прохождении пищевого кома, и частично черпаловидные хрящи состоят из эластического хряща, остальные — из гиалинового хряща, обеспечивая зияние гортани. В полости гортани между черпаловидными и щитовидным хрящами расположены 2 голосовые складки с голосовой щелью между ними. В основе голосовой складки лежит эластическая
голосовая связка и голосовая мышца. Голосовой аппарат покрыт слизистой
оболочкой, как и вся полость гортани.
ТРАХЕЯ
Рис. 112. Гортань крупного рогатого
скота снаружи (А) и изнутри (Б):
1 — надгортанник; 2 — щитовидный
хрящ; 3— черпаловидный хрящ; 4—
кольцевидный хрящ; 5 — хрящи трахеи; 6 — голосовая связка; 7 —другие
связки гортани.
Трахея — trachea — трубка,
в стенке которой заключены незамкнутые хрящевые кольца,
обеспечивающие ее зияние. Хрящи соединены друг с другом
кольцевидными связками. Их незамкнутые края обращены дорсально и соединены трахейными
мышцами, которые могут уменьшить просвет трахеи. В области
шеи? трахея лежит под позвонками и пищеводом или рядом с ним.
В грудной полости на уровне
4—6-го ребра трахея делится на 2
главных бронха. Место деления
называется бифуркацией. У рогатого скота трахея содержит
46—50 хрящей, сжата с боков так,
что в поперечнике имеет яйцевидную форму. До бифуркации от
нее отходит трахейный бронх. У
свиньи в трахее 32—36 колец округлой формы, их незамкнутые
концы заходят друг за друга.
Имеется трахейный бронх. У лошадей в трахее 48—60 колец поwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
перечноовальной формы, которые заходят друг за друга. Трахейного бронха
нет.
Иннервируется трахея блуждающим нервом, васкуляризируется ветвями
общей сонной артерии.
Гистологическое строение. Стенка трахеи состоит из слизистой оболочки, подслизистой основы, фиброзно-хрящевой оболочки и адвентиции.
Слизистая оболочка. Эпителий слизистой оболочки многорядный мерцательный. В его состав входят реснитчатые, бокаловидные, эндокринные
(энтерохромаффинные) и базальные клетки (см. главу 7). Реснитчатые клетки
цилиндрической формы, высокие, на апикальном полюсе несут 250—300
ресничек, продвигающих слизь в сторону глотки. Бокаловидные клетки типичные, вырабатывают слизь, увлажняющую поверхность эпителия. Эндокринные клетки подобны таким же клеткам желудочно-кишечного тракта.
Они невысокие, достигают поверхности эпителиального пласта, треугольной
формы, с округлым ядром и гранулами в цитоплазме. Вырабатывают пептидные гормоны и биогенные амины: норадреналин, дофамин, серотонин. Регулируют тонус мышечных элементов трахеи. Базальные клетки низкие, лежат
на базальной мембране. Их ядра образуют самый нижний ряд. Считается, что
они выполняют камбиальную функцию. От собственной пластинки слизистой оболочки эпителий отделен хорошо выраженной базальной мембраной.
Собственная пластинка слизистой оболочки образована тонким слоем
рыхлой волокнистой соединительной ткани с большим количеством эластических волокон. В ней встречаются скопления лимфоцитов и лимфатические
фолликулы. Сколько-нибудь значительного количества мышечных элементов
слизистая не содержит и не может собираться в складки. На границе с подслизистой эластические волокна лежат слоем наподобие эластической мембраны.
Подслизистая основа образована рыхлой волокнистой соединительной
тканью. В ней залегают сложные серозно-слизистые железы, заходящие и в
промежутки между хрящевыми кольцами. Их секрет увлажняет поверхность
слизистой. В подслизистой основе расположены сосудистые и нервное сплетения. Без резких границ она переходит в фиброзно-хрящевую оболочку.
Фиброзно-хрящевая оболочка образована соединительной тканью с правильно расположенными в ней хрящевыми кольцами. В области колец соединительной ткани мало, в основном это надхрящница. Между кольцами
соединительная ткань образует мощный слой, куда заходят концевые отделы
желез. Хрящевые кольца образованы гиалиновым хрящом, покрытым со всех
сторон надхрящницей. Между незамкнутыми концами хрящей лежит поперечная мышца трахеи из гладкой мышечной ткани.
Адвентиция покрывает трахею на всем протяжении, так как в грудной
полости она проходит в средостении.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ЛЕГКИЕ
Легкие — pulmones (рис. 113)—крупный парный орган дыхания, в котором осуществляется газообмен между вдыхаемым воздухом и кровью. Легкие имеют форму конуса — вершина об ращена вперед, а основание лежит на
диафрагме. Они окружают сердце и заполняют собой практически всю грудную клетку, повторяя ее форму, за исключением средостения, в котором находятся сердце, пищевод, трахея и крупные сосуды.
Рис. 113. Доли и бронхи легкого:
А — крупного рогатого скота; Б— свиньи; В — лошади; 1— трахея; 2—
трахейный бронх; 3— бифуркация; 4— главный бронх; 5 — крупный бронх; б
—средний бронх; 7— верху, шечная доля; 8 — сердечная доля; 9— добавочная
доля; 10— диафрагмальная доля; 11 — острый край.
У рогатого скота относительная масса легких составляет 0,6—0,8%. Они
умеренно длинные, суженные в краниальной части. На них различают дорсальный — тупой край и вентрокаудальный — острый край. Выпуклая поверхность легких, прилежащая к ребрам, называется реберной, вогнутая, обращенная к диафрагме, — диафрагмальной, обращенная друг к другу — медиальной (средостенной). На медиальной поверхности каждого легкого имеется сердечное вдавление и углубление — ворота легкого, где внутрь легкого
проникают главный бронх, легочная артерия и нервы, а из легкого выходят
легочные вены. Главный бронх, нервы и сосуды образуют корень легкого.
Правое легкое больше левого. На каждом легком различают краниальную
(верхушечную), среднюю (сердечную) и каудальную (диафрагмальную) доли.
Друг от друга доли отделяются довольно глубокими вырезками. Краниальная
доля правого легкого состоит из двух частей. К ней подходит трахейный
бронх, который отходит от трахеи до бифуркации. Краниальная доля левого
легкого одинарная, небольшая. Средние (сердечные) доли узкие, длинные,
сильно выступают в латеральные стороны. Каудальные (диафрагмальные)
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
доли самые массивные. На правом легком с медиальной стороны имеется
еще одна небольшая доля — добавочная.
У свиньи легкие более округлые. Их относительная масса 0,85%. Краниальная доля правого легкого одинарная, воздух к ней подходит через трахейный бронх. Средние (сердечные) доли не выступают за край дальше каудальных.
У лошади легкие очень крупные, длинные, слабо разделенные на доли.
Их относительная масса 1,3—1,5%. Трахейного бронха нет. На каждом легком краниальная доля одинарная, средняя и каудальная доли соединены в
единую сердечно-диафрагмальную долю.
Сосуды легких. Все функции легких теснейшим образом связаны с кровью. Этим объясняется чрезвычайно обильное кровоснабжение органа. На
долю крови приходится до 1/3 его массы. Функциональные сосуды легких образуют легочный (малый) или дыхательный круг кровообращения. Начинается он легочной артерией, которая несет венозную кровь в легкие. Здесь она
разветвляется до капилляров, где и происходит газообмен. Артериальная
кровь собирается в легочные вены, несущие кровь к сердцу. Сосудом, питающим ткань легкого, является бронхиальная артерия, отходящая от грудной аорты. У более подвижных видов животных артерии имеют более широкий просвет, более тонкую стенку, но с большим содержанием эластических
мембран. У менее подвижных видов в стенках артерий лучше развита мышечная ткань.
Плевра — парная серозная оболочка грудной полости. Ее париетальный
листок прирастает к внутригрудной фасции и в зависимости от расположения
называется реберной и диафрагмальной плеврой. Ее висцеральный листок
прирастает к легким, образуя легочную плевру, с легких переходит на средостение — средостенная плевра и на сердце — сердечная плевра. Между париетальным и висцеральным листками сохраняется щелевидная, герметично
замкнутая парная плевральная полость, имеющая отрицательное давление и
небольшое количество плевральной жидкости, достаточное для смазывания
ее стенок. При расширении грудной клетки под действием мышцинспираторов легкие растягиваются благодаря присасывающему действию
отрицательного давления в плевральных полостях. В легкие засасывается
воздух— происходит вдох. Выдох осуществляется пассивно при расслаблении мышц - вдыхателей и под воздействием эластической тяги легкого.
Гистологическое строение легких (рис. 114). Легкое — компактный орган, покрытый снаружи серозной оболочкой — легочной плеврой. Соединительнотканная строма легкого развита слабо. Она образует тонкую, но плотную капсулу, междольковые прослойки, в которых проходят сосуды, нервы и
бронхи и нежный остов долек. Дольчатость легких лучше других выражена у
рогатого скота и свиньи. Дольки неправильной конической формы своими
более широкими основаниями направлены наружу, что придают поверхности
легкого ячеистый вид. Паренхима легкого образована воздухоносными путями — бронхиальным деревом и респираторными (дыхательными) отделами
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
— альвео лярным деревом. В состав бронхиального дерева входят круп« ные,
средние, мелкие бронхи и бронхиолы. Ветвление бронхиального дерева происходит закономерно. У копытных насчитывается до 16 порядков бронхов.
Диаметр их при ветвлении (при переходе от одного к другому) может
уменьшаться постепенно (у свиньи) или скачкообразно (у лошади).
Рис. 114. Схема строения бронхиального и альвеолярного дерева легких:
Т — трахея; а —хрящевое кольцо; ГБ — главный бронх; КБ — крупный
бронх; СБ —средний бронх; МБ — мелкий бронх и бронхиола; ТБ — терминальная бронхиола; АЦ— ацинус; АБ — альвеолярная (респираторная) бронхиола; Ах — альвеолярный ход; Ам — альвеолярный мешок; А — альвеола. В
прямоугольниках показано гистологическое строение воздухоносных и респираторных отделов легкого: Э — эпителий; С — собственная и М—мышечная пластинки слизистой оболочки; Л— подслизистая основа с железами; X— фибрознохрящевая оболочка; З — надхрящница и наружная соединительнотканная оболочка.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Крупные бронхи по строению своей стенки похожи на трахею. Эпителий
слизистой оболочки многорядный мерцательный с теми же видами клеток,
что и в трахее. В собственной пластинке слизистой оболочки имеются пучки
гладкомышечных клеток. Эластические волокна хорошо развиты. В подслизистой основе имеются сложные трубчато-альвеолярные слизисто-серозные
железы. Гиалиновый хрящ фиброзно-хрящевой оболочки имеет вид сплошных пластин. Адвентиция переходит в междольковую соединительную ткань
легкого.
С уменьшением диаметра бронхов видоизменяется и упрощается их
структура. В средних бронхах эпителий многорядный мерцательный, но высота его меньше, чем в крупных. Уменьшается толщина слизистой оболочки
и подслизистой основы. Увеличивается количество мышечных элементов и
желез. Сохраняется сеть эластических волокон. Редуцируется хрящ. Он принимает вид отдельных островков. Стенка малых бронхов гораздо тоньше.
Эпителий однорядный. В его состав входят наряду с другими безреснитчатые
и специальные секреторные клетки. Мышечная пластинка слизистой хорошо
развита и может регулировать просвет бронха. Подслизистая основа и фиброзно-хрящевая оболочка со всеми их структурами отсутствуют. Сеть эластических волокон хорошо развита. В бронхиолах эпителий кубический. В
его состав входят реснитчатые, безреснитчатые и секреторные клетки. Собственная пластинка слизистой тонкая, в ней имеются гладкомышечные клетки, фибробласты, эластические, ретикулярные и коллагеновые волокна.
Большая часть бронхиального дерева разветвляется за пределами долек легкого. В дольку легкого вступает бронхиола или дольковый бронх и разветвляется на несколько терминальных бронхиол.
Терминальная бронхиола является конечным звеном воздухоносного пути. Альвеолярное дерево составляет дольки легкого. Каждая терминальная
бронхиола разделяется на респираторные бронхиолы. От каждой респираторной бронхиолы идут 2 респираторных (альвеолярных) хода, оканчивающиеся альвеолярными мешками. В стенках респираторных бронхиол, ходов и
мешков есть тонкостенные пузырьки с широким отверстием — альвеолы.
Участок респираторного отдела, связанный с одной терминальной бронхиолой, называется ацинус. В состав дольки обычно входит несколько (10—20)
ацинусов. Количество альвеол в ацинусе увеличивается от его начальных
участков к конечным, так что стенки воздухоносных мешков сплошь состоят
из альвеол.
Альвеола — тонкостенный пузырек, стенка которого образована двумя
видами клеток альвеолярного эпителия, расположенного на базальной мембране, а широкое отверстие открыто в полость альвеолярного мешка. Большая поверхность альвеолы покрыта респираторными альвеолоцитами —
очень тонкими (0,2— 0,3 мкм в высоту) и обширными по площади клетками,
несколько утолщенными (5 мкм) лишь около ядра. Клетки эти богаты органеллами и покрыты сурфактантом — веществом, препятствующим слипанию альвеол. Другой тип клеток — большие альвеолоциты — крупные клетки с большим ядром, секреторными гранулами в цитоплазме и микроворсинwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ками. Они вырабатывают сурфактант. К базальной мембране эпителия альвеол тесно прилежат эластические и коллагеновые волокна, макрофаги и многочисленные кровеносные капилляры. Все эти структуры имеют очень малую толщину, так что воздушно-гематический барьер не превышает 0,3—0,5
мкм и газообмен совершается путем диффузии благодаря разности парциальных давлений О2 и СО2 в крови и воздухе альвеол.
С возрастом в легких увеличивается количество эластина, общее количество желез, хотя плотность их на единицу поверхности бронхов становится
меньше. Существенный отпечаток на строение легких накладывают условия
существования. У домашних видов копытных легкие короче и шире, асимметрия их меньше выражена, чем у диких родственных видов. У ягнят зимнего окота легкие крупнее, имеют лучше развитые железы, более крупный
диаметр бронхов и сосудов, чем у ягнят весеннего окота. При содержании
ягнят в двухъярусных клеточных батареях без выгула и естественного солнечного освещения легкие приобретают красно-коричневый цвет из-за сильного расширения сосудов с утолщенными стенками. Увеличивается содержание лимфоцитов в тканях легкого, происходит редукция желез и ресничек
эпителия, утолщение плевры по сравнению с ягнятами, содержащимися на
пастбище.
Вопросы для самоконтроля. 1. В чем выражаются изменения дыхательной системы в фило- и онтогенезе? 2. Строение носовой полости, гортани, трахеи. 3. Анатомо-гистологическое строение легких. 4. Общие закономерности макро- и микроскопического строения системы органов дыхания.
Глава 17. СИСТЕМА ОРГАНОВ МОЧЕВЫДЕЛЕНИЯ
В состав системы органов мочевыделения входят почки, мочеточники,
мочевой пузырь, мочеиспускательный канал, мочеполовой синус (у самок)
или мочеполовой канал (у самцов). Органы мочевыделения осуществляют
выработку, временное хранение и выделение из организма жидких конечных
продуктов обмена—мочи. Выполняют экскреторную функцию, извлекая из
крови и удаляя из организма вредные продукты азотистого обмена (мочевину, мочевую кислоту, аммиак, креатин, креатинин), инородные вещества
(краски, лекарства и др.), некоторые гормоны (пролан, андростерон и др.).
Удаляя избыток воды, минеральные вещества и кислые продукты, почки регулируют водно-солевой обмен и поддерживают относительное постоянство
осмотического давления и активной реакции крови. В почках синтезируются
гормоны (ренин, ангиотензин), участвующие в регуляции кровяного давления и диуреза (мочеотделения).
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
КРАТКИЕ СВЕДЕНИЯ О РАЗВИТИИ СИСТЕМЫ ОРГАНОВ
МОЧЕВЫДЕЛЕНИЯ
Филогенез. У наиболее примитивно организованных многоклеточных
животных (гидра) выделительная функция осуществляется диффузно всей
поверхностью тела без каких-либо структурных приспособлений. Однако у
большинства бесполостных (плоские черви) и первичнополостных беспозвоночных в паренхиме тела имеется система первичных выделительных трубочек— протонефридий. Это система очень тонких канальцев, проходящих
внутри длинных клеток. Один конец канальца открывается порой на поверхности тела, другой — замыкается особыми отростчатыми клетками. Из
окружающих тканей клетки всасывают жидкие продукты обмена и продвигают их по канальцам с помощью жгутиков, опущенных в каналец. Собственно
выделительная функция здесь присуща клеткам. Канальцы являются лишь
выводящими путями.
С появлением целома — вторичной полости тела (у личинок кольчатых
червей) протонефридиальная система морфологически оказывается связанной с ним. Стенки канальцев несколько вдаются в целом, омываются тканевой жидкостью. К ним переходит функция избирательного всасывания и
выведения продуктов обмена. Отростчатые клетки редуцируются. В них сохраняются мерцательные жгутики, продвигающие жидкость по канальцу. В
дальнейшем замкнутый конец канальца прорывается отверстием во вторичную полость тела. Образуется мерцательная воронка. Сами канальцы
утолщаются, удлиняются, изгибаются, продолжаясь из одного сегмента целома в другой (целом сегментирован). Такие видоизмененные канальцы называются нефридиями. Последние расположены по двум сторонам тела метамерно и соединяются друг с другом своими конечными участками. Это
приводит к формированию с каждой стороны тела продольного протока —
примитивного мочеточника, в который отрываются по пути его хода все сегментальные нефридии. Примитивный мочеточник открывается наружу либо
самостоятельным отверстием, либо в клоаку. В полости тела по соседству с
нефридиями кровеносные сосуды образуют густую сеть капилляров в виде
клубочков. Подобное строение имеет выделительная система у примитивных
хордовых — ланцетника, круглоротых, личинок рыб. Располагается она в передней части тела животного и называется предпочти — pronephros, или головной почкой.
Дальнейший ход изменений выделительной системы характеризуется
постепенным сдвиганием ее элементов в каудальном направлении с одновременным усложнением структур и оформлением в компактный орган. При
этом различают туловищную, или промежуточную почку (mesonephros), и
тазовую, или дефинитивную, почку (metanephros). Усложнение в строении
почечного канальца выражается в его удлинении, большей извитости, а главное, более тесной связи с кровеносном системой. В стенке канальца появляется слепое выпячивание, в которое вдается сосудистый капиллярный клубочек — появляется почечное (мальпигиево) тельце. К нему постепенно переwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ходит фильтрационная функция. Воронка запустевает и редуцируется. Почечные канальцы все больше удлиняются. Их эпителий в разных участках
приобретает специфические черты. В целом каналец осуществляет резорбционную функцию. Для туловищной почки характерна редукция воронки, переход фильтрационной функции к почечному тельцу, а резорбционной функции — к эпителию почечного канальца. Орган теряет метамерность, приобретает черты компактного органа — нефрогенного тяжа с обилием кровеносных сосудов. Проток промежуточной почки (вольфов) часто является отщеплением протока предпочки (мюллерова) и также впадает в клоаку. Промежуточная почка функционирует в течение всей жизни у рыб и амфибий, а в эмбриональный период развития у рептилий, птиц и млекопитающих.
Дефинитивная почка или метанефрос развивается только у амниот: рептилий, птиц и млекопитающих. Развивается она из двух зачатков: мочеотделительного и мочеотводящего. Мочеотделительная часть образована
нефронами — сложными извитыми мочеотделительными трубочками, несущими на конце капсулу, в которую вдается сосудистый клубочек. От канальцев туловищной почки нефроны отличаются большей длиной, извитостью,
большим количеством капилляров в сосудистом клубочке.
Нефроны и оплетающие их кровеносные сосуды объединены соединительной тканью в компактный орган. Мочеотводящая часть развивается из
заднего конца протока промежуточной почки и называется дефинитивным
мочеточником. Дорастая до компактной массы нефрогенной ткани, мочеточник формирует почечную лоханку, стебельки и чашечки и вступает в контакт
с мочеотделительными канальцами почки. Другим концом дефинитивный
мочеточник объединяется с половым каналом в мочеполовой канал и у рептилий, птиц и однопроходных млекопитающих открывается в клоаку. У плацентарных млекопитающих открывается самостоятельным отверстием мочеполового канала (синуса). Промежуточный участок отводящих путей между
мочеточником и мочеполовым каналом образует мешкообразное расширение
— мочевой пузырь. Он образуется у плацентарных млекопитающих из участков стенок аллантоиса и клоаки в месте их соприкосновения.
Онтогенез (рис. 115). У млекопитающих нефрогенная ткань дифференцируется в области сегментных ножек мезодермы всех сомитов последовательно, начиная с головных и кончая тазовыми. При этом на протяжении
внутриутробного развития особи закладывается сначала головная почка, затем туловищная и, наконец, тазовая с характерными для них структурами.
Предпочка закладывается на ранней стадии развития зародыша в области
первых 2—10 сомитов из материала сегментных ножек, существует несколько десятков часов и не функционирует как орган мочевыделения. В процессе
дифференцировкн материал сегментных ножек отшнуровывается от сомитов,
вытягивается по направлению к эктодерме в виде трубочек, сохраняющих
связь с целомом. Это и есть каналец предпочки с воронкой, обращенной в
целом. Противоположные концы канальцев сливаются и образуют трубчатые
протоки, идущие каудально. Вскоре предпочка редуцируется. На базе ее протоков образуются яйцеводы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
После закладки предпочки начинает дифференцироваться нефрогенная
ткань следующих 10—29 сегментов с образованием промежуточной (туловищной) почки. Мезонефридиальные канальцы в отличие от канальцев предпочки имеют почечные тельца с развитыми капсулами и сосудистыми клубочками. Канальцы растут по направлению к протоку предпочки, прорываются в него и с этого момента проток предпочки называется протоком промежуточной почки (вольфов). Промежуточная почка функционирует как орган выделения. Продукты выделения (мочевина, мочевая кислота и др.) по
протоку промежуточной почки стекают в клоаку, а оттуда — в аллантоис, где
и накапливаются.
Рис. 115. Развитие почек:
А — схема строения трех генераций почек; Б, В, Г, Д — схема последовательных стадий эмбрионального развития нефрона; I — пронефрос; II — мезонефрос; III — метанефрос; 1 — Мюллеров канал; 2— вольфов канал; 3—
дефинитивный мочеточник; 4— клоака; 5 — мочеотделительная трубочка
предпочки с воронкой; 6 — целом; 7 —аорта; 8 — почечные артерии; 9 —
наружный сосудистый клубочек; 10 — воронка; — внутренний сосудистый
клубочек; 12 — почечное тельце; 13 — каналец нефрона; 14 — эктодерма; 15
— нервная трубка; 16 — энтодерма; 17 — мезодерма; 18 — сомит; 19 —
сегментная ножка; 20 — спланхнотом.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
К концу зародышевого периода происходит бурный рост и дифференцировка нефрогенной ткани задних сегментов — тазовой почки. Функция мезонефроса при этом затухает. У 2-месячного плода теленка почки по своему
положению соответствуют взрослым животным, но лишь у 4-месячных плодов их форма похожа на форму у взрослых животных. Нефроны начинают
формироваться с 3-го месяца, и новообразование их продолжается не только
в течение утробного развития, но и после рождения (у лошади до 8 лет, у
свиньи до 1,5 года). Дифференцировка нефрона начинается с закладки почечного тельца. Затем развивается каналец нефрона и, наконец, собирательная трубочка. В плодный период масса почек увеличивается в 94 раза, с рождения до взрослого состояния — в 10 раз. Относительная масса почек при
этом снижается с 0,4 до 0,2%.
Одновременно с закладкой дефинитивной почки от протока промежуточной почки отрастает дивертикул — зачаток мочеточника. Врастая в нефро-генный зачаток, он формирует лоханку и почечные чашечки. Одновременного с дифференцировкой нефрогенного эпителия происходит развитие
сосудистого русла почки и тесное соединение его со структурами нефрона.
Основная масса нефронов развивается в периферических участках почки
— в корковом веществе. Корковое вещество в начале плодного периода растет очень интенсивно. Затем по скорости роста его обгоняет мозговое вещество — центральные участки органа, где сосредоточены структуры, отводящие мочу. У новорожденных животных по сравнению со взрослыми
корковый слой развит слабо. Его рост и дифференцировка нефронов происходят активно в первый год жизни и продолжаются, хотя и с меньшей интенсивностью, до полового созревания. У старых животных в почке нарушаются процессы клеточного обновления, снижаются способности почечного эпителия к реабсорбции веществ.
Типы почек. В процессе филогенеза животных разных семейств и родов
образовалось несколько типов дефинитивной почки в зависимости от степени срастания ее участков: множественная, бороздчатая многососочковая,
гладкая много- и однососочковая. Множественная почка наиболее раздробленная. Она состоит как бы из отдельных почечек (до 100 и более), объединенных прослойками соединительной ткани и капсулой в единый компактный орган. Каждая почечка состоит из коркового и мозгового вещества и
соединена с собственной чашечкой. От каждой чашечки отходит стебелек.
Стебельки объединяются в мочеточник, отводящий мочу от почки. Множественная почка присуща медведю, выдре, китообразным. В бороздчатой многососочковой почке отдельные почечки — дольки почки соединены друг с
другом средними участками. Корковое вещество долек отграничено бороздами друг от друга, а мозговое образует большое количество сосочков, каждый из которых опущен в свою чашечку. Такие почки у крупного рогатого
скота. В гладких многососочковых почках корковое вещество почечных долек
слилось, а мозговое образует отдельные сосочки. Такие почки у свиньи, человека. В гладких однососочковых почках слилось не только корковое, но и
мозговое вещество с образованием одного крупного валико-образного сосочwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ка. Такие почки у большинства млекопитающих, а из домашних животных у
лошади, мелкого рогатого скота, собаки.
ПОЧКИ
Почка — ren (рис. 116)—в большинстве случаев бобовидной формы,
буро-красного цвета. На почке различают дорсальную и вентральную поверхности, латеральный и медиальный края, краниальный и каудальный концы.
На медиальном крае есть углубление— ворота почки, ведущие в почечную
ямку — синус. В ворота почки входят артерии, выходят вены и мочеточник.
В синусе расположена лоханка и другие разветвления мочеточника. Сверху
почка одета фиброзной капсулой, которая плотно прирастает лишь в области
ворот. Поверх капсулы и в синусе почки скапливается большое количество
жировой ткани, формирующей жировую капсулу почки. Вентральная поверхность почки покрыта серозной оболочкой. На продольном разрезе в почке видны 3 зоны: корковая, мозговая и промежуточная. Корковая зона лежит
на периферии, буро-красного цвета и является мочеотделительной, так как в
основном состоит из нефронов. Мозговая зона лежит в центральных участках
органа, буровато-желтоватого цвета и является мочеотводящей. Пограничная
зона расположена между корковой и мозговой зонами, темно-красного цвета,
содержит большое количество крупных сосудов.
Рис. 116. Почка:
А—крупного рогатого скота; Б—лошади; В — свиньи; 1 — корковая, 2 —
промежуточная и 3— мозговая зоны; 4 — почечная пирамида; 5 — почечный
сосочек; 6 — почечная чашечка; 7— стебелек; 8 — проток; 9 — лоханка;
10— мочеточник; 11 — почечная артерия; 12—почечная вена; 13—ворота
почки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Почки крупного рогатого скота (А) овальные, относятся к типу бороздчатых многососочковых. Фиброзная капсула почки заходит в глубь борозд.
Краниальный конец почки уже каудального. Ворота почки широкие. Левая
почка перекручена по продольной оси, висит на брыжейке, которая позволяет
ей смещаться за правую почку при наполнении рубца. Масса каждой почки
500—700 г, а относительная масса 0,2—0,3%.
Корковая мочеотделительная зона почки разделена на доли. Пограничная зона хорошо выражена. Мозговая зона в каждой доле имеет форму пирамиды, основанием направленной к корковой зоне, а вершиной, называемой
сосочком, — в чашечку. В почке крупного рогатого скота насчитывается 16—
35 почечных пирамид. Вершины почечных сосочков испещрены сосочковыми отверстиями, через которые моча стекает в почечные чашечки — конечные разветвления мочеточника. Из чашечек моча стекает по стебелькам в 2
протока, которые в области ворот объединяются в один мочеточник.
Правая почка соприкасается с печенью, лежит на уровне от 12-го ребра
до 2—3-го поясничного позвонка (левая — от 2-го до 5-го поясничного позвонка). Иннервируется блуждающим и симпатическими нервами. Васкуляризуется почечной артерией.
У свиньи (Б) почки гладкие многососочковые, бобовидные, уплощенные
дорсовентрально. Пирамид 10—12, столько же сосочков. Некоторые сосочки
могут слиться. К сосочкам подходят чашечки, открывающиеся непосредственно в почечную лоханку, расположенную в синусе почки. Обе почки лежат
в поясничной области, на уровне 1—4 поясничных позвонков.
У лошади (В) почки гладкие, однососочковые. Правая почка сердцевидной формы, левая — бобовидная. Пограничная зона широкая, хорошо выражена. Количество почечных пирамид достигает 40—64. Сосочки слиты в
один, направленный в почечную лоханку. Правая почка почти целиком лежит в подреберье, на уровне от 16-го (14—15-го) ребра до 1-го поясничного
позвонка. Левая почка лежит на уровне 1—3 поясничных позвонков и редко
заходит в подреберье.
Гистологическое строение. Почка — компактный орган (рисунок
117).Строма образует капсулу и тончайшие прослойки внутри органа, которые идут в основном по ходу сосудов. Паренхима образована эпителием,
структуры которого могут функционировать лишь в тесном контакте с кровеносной системой. Почки всех типов делятся на доли. Доля— это почечная
пирамида с участком покрывающего ее коркового вещества. Доли отделены
друг от друга почечными колонками — участками коркового вещества, проникающего между пирамидами. Доли состоят из долек, не имеющих четких
границ. Долькой считается группа нефронов, впадающих в одну собирательную трубочку, которая проходит по центру дольки и называется мозговым
лучом, так как опускается в мозговое вещество (А). Кроме ветвящейся собирательной трубочки, мозговой луч содержит прямые канальцы (петли) нефрона.
Нефрон — основная структурно-функциональная единица почки. В почках крупного рогатого скота до 8 млн нефронов. 80% из них находятся в корwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ковом веществе — это корковые нефроны. 20% располагаются в мозговом
веществе и называются юкстамедуллярными. Длина одного нефрона от 2 до
5 см. Нефрон образован однослойным эпителием и состоит из капсулы нефрона, проксимального отдела, петли нефрона (Генле) и дистального отдела.
Капсула нефрона имеет вид двустенной чаши, ее внутренняя стенка (внутренний листок) тесно связана с кровеносными капиллярами. Наружный листок капсулы построен однослойным плоским эпителием. Между листками
капсулы имеется щелевидная полость капсулы.
Рис. 117. Схема строения почечной дольки (А) и нефрона (Б):
I — корковая, II — промежуточная и III—мозговая зоны; а — корковый нефрон; б — юкстамедуллярный нефрон; в — собирательная трубочка (мозговой луч); 1 — почечное тельце; 2— капсула нефрона; 3— проксимальный извитой каналец; 4 — петля нефрона; 5 — дистальный извитой каналец; 6 —
междольковые артерия и вена; 7— дуговые артерия и вена; 8— наружный и
9 — внутренний листки капсулы нефрона; 10— капилляр сосудистого клубочка; 11 — приносящая и 12—выносящая артериолы; 13 — эндотелий капилляра; 14 — подоцит внутреннего листка капсулы нефрона; 15 — цитотрабекула; 16 — цитоподия; 17 — базальная мембрана; 18 — клетка проксимального отдела; 19 — щеточная каемка; 20 — митохондрии; 21 — базальная исчерченность; 22 — эндоплазматическая сеть; 23 — аппарат Гольджи.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Капилляры анастомозируют друг с другом, образуя сосудистый клубочек из 50—100 петель. Кровь к сосудистому клубочку поступает по приносящей артриоле. Капилляры клубочка объединяются в выносящую артериолу.
Расположение капилляров между двумя артериолами называется чудесной
артериальной системой почки.
Капсула нефрона вместе с сосудистым клубочком называется почечным
тельцем. Все почечные тельца расположены в корковом веществе почки. В
почечном тельце происходит образование первичной мочи — клубочкового
фильтрата, путем фильтрации составных частей плазмы крови. Это становится возможным благодаря структурным особенностям почечного тельца.
Приносящая артериола имеет просвет большего диаметра, чем выносящая
артериола. Это создает повышенное давление в капиллярах сосудистого клубочка. В эндотелии капилляров имеются щели и многочисленные фенестры
— подобия очень мелких пор, что способствует просачиванию плазмы. Эпителий внутреннего листка капсулы тесно прилежит к эндотелию капилляров,
повторяя все их изгибы, отделяясь лишь базальной мембраной. Он образован
своеобразными плоскими отростчатыми клетками диаметром 20—30 мкм —
подоцитами. Каждый подоцит имеет несколько крупных отростков — цитотрабекул, от которых отходят многочисленные мелкие отростки —
цитоподии, прикрепляющиеся к базальной мембране. Между цитоподиями
имеются щели. В результате образуется биологический почечный фильтр,
обладающий избирательной способностью. В норме через него не проходят
клетки крови и крупные молекулы белков. Остальные части плазмы могут
войти в состав первичной мочи, которая поэтому мало отличается от плазмы
крови. Количество первичной мочи — клу-бочкового фильтрата у крупных
животных составляет несколько сотен литров в сутки. Клубочковый фильтрат попадает в просвет капсулы почечного тельца, а оттуда — в каналец нефрона. В нем происходит обратное избирательное всасывание в кровоток—
реабсорбция составных частей клубочкового фильтрата, так что вторичная
моча, удаляющаяся из организма, по объему составляет лишь 1—2% от первичной мочи и совершенно не соответствует ей по химическому составу. Во
вторичной моче в 90 раз меньше воды, натрия, в 50 раз меньше хлоридов, в
70 раз больше концентрация мочевины, в 30 раз — фосфатов, в 25 — мочевой кислоты. Сахар и белок в норме отсутствуют. Реабсорбция начинается и
активнее всего протекает в проксимальном отделе нефрона.
В состав проксимального отдела нефрона входит проксимальный извитой каналец и прямой каналец, который в то же время является частью петли
нефрона. Просвет капсулы почечного тельца переходит в просвет проксимального извитого канальца. Его стенки образованы однослойным кубическим эпителием, который является продолжением эпителия наружного листка капсулы нефрона. Проксимальные извитые канальцы имеют диаметр около 60 мкм, лежат в корковом веществе, изгибаясь в непосредственной близости от почечного тельца. Клетки проксимального извитого канальца на апикальном полюсе, обращенном в просвет канальца, несут большое количество
микроворсинок, формирующих щеточную каемку — приспособление для акwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тивного всасывания веществ. Округлое ядро сдвинуто к базальному полюсу.
Плазмолемма базального полюса образует глубокие впячивания в виде складок внутрь клетки. Между этими складками рядами лежат удлиненные митохондрии. На световом уровне эти структуры имеют вид базальной исчерченности. Клетки активно всасывают глюкозу, аминокислоты, воду и соли и
имеют мутную, оксифильную цитоплазму. На протяжении проксимального
отдела реабсорбируется все количество сахара, аминокислот и мелких белковых молекул, попавших в клубочковый фильтрат, 85% воды и натрия.
Проксимальный извитой каналец переходит в петлю нефрона (Генле).
Это прямой каналец, который на разную глубину заходит в мозговое вещество. В петле нефрона различают нисходящую и восходящую части. Нисходящая часть сначала образована кубическим эпителием, таким же по структуре
и функции, как в проксимальном извитом канальце, и потому этот участок
относят и к проксимальному отделу нефрона в качестве его прямого канальца. Нижний участок нисходящей части петли нефрона имеет диаметр 15
мкм, образован плоским эпителием, ядра которого выступают в просвет канальца и называется тонким канальцем. Его клетки имеют светлую цитоплазму, мало органелл, единичные микроворсинки и базальную исчерченность. Тонкий каналец петли нефрона продолжается и в ее восходящую
часть. В нем происходит всасывание солей и выведение их в тканевую жидкость. В верхнем участке эпителий становится кубическим и переходит в
дистальный извитой каналец с диаметром до 50 мкм. Толщина его стенок
меньше, а просвет больше, чем в проксимальном извитом канальце.
Стенки дистального извитого канальца образованы кубическим эпителием со светлой цитоплазмой без щеточной каемки, но с базальной исчерченностью. В нем происходит реабсорбция воды и солей. Дистальный извитой каналец расположен в корковом веществе и одним своим участком соприкасается с почечным тельцем между приносящей и выносящей артериолами. В этом месте, называемом плотным пятном, клетки дистального извитого канальца высокие и узкие. Предполагают, что они улавливают изменение содержания натрия в моче.
Во время нормальной работы почек активно функционирует 30—50%
нефронов. При введении мочегонных средств — 95— 100%.
Юкстамедуллярные нефроны отличаются по структуре и функции от
корковых нефронов. Их почечные тельца крупнее, лежат в глубоких участках
коркового вещества. Приносящая и выносящая артериолы имеют одинаковый диаметр. Петля нефрона, особенно ее тонкий каналец, гораздо длиннее,
достигает глубоких слоев мозгового вещества. В области плотного пятна
имеется юкстагломерулярный (околоклубочковый) аппарат — скопление нескольких видов клеток, в сумме образующих эндокринный комплекс почек,
регулирующий почечный кровоток и мочеобразование. Он участвует в синтезе ренина — гормона, стимулирующего выработку в организме сосудосуживающих веществ (ангиотензинов), а также стимулирует продукцию гормона альдостерона в надпочечниках. Из дистального отдела нефрона моча
поступает в собирательную трубочку.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Собирательные трубочки не являются составными частями нефронов.
Это конечные разветвления мочеточника, проникающие в паренхиму почки и
срастающиеся с концами нефронов. Участки собирательных трубочек, лежащие в корковом веществе, образованы кубическим эпителием с очень светлой цитоплазмой, в мозговом веществе — цилиндрическим, эпителием. В собирательных трубочках продолжается некоторое всасывание воды из-за гипертоничности окружающей тканевой жидкости. В результате моча становится еще более концентрированной. Собирательные трубочки образуют разветвленную систему. Они проходят в центре мозговых лучей коркового вещества и в мозговом веществе и объединяются в сосочковые протоки, открывающиеся отверстиями на вершине сосочков.
Кровоснабжение почки осуществляется крупной парной почечной артерией, которая входит в почку в области ворот и разветвляется на междолевые
артерии. В пограничной зоне почки они переходят в дуговые артерии. От них
отходит в корковое вещество большое количество междольковых артерий.
Эти артерии разветвляются на внутридольковые артерии, от которых отходят
приносящие артериолы, разветвляющиеся на капилляры сосудистого клубочка. Капилляры собираются в выносящую артериолу. Здесь мы видим чудесную артериальную систему почки — капилляры между двумя артериями. В
этих капиллярах происходит фильтрация крови с образованием первичной
мочи. Выносящая артериола вновь ветвится на капилляры, оплетающие канальцы нефрона. В эти капилляры из канальцев нефрона попадают реабсорбированные вещества. Капилляры объединяются в вены, выносящие
кровь из почки.
МОЧЕТОЧНИКИ, МОЧЕВОЙ ПУЗЫРЬ И
МОЧЕИСПУСКАТЕЛЬНЫЙ КАНАЛ
Мочеточники — ureteres (рис. 118)—длинные узкие трубочки, идущие
от ворот почек до мочевого пузыря вдоль боковых стенок брюшной полости.
Входят в дорсальную стенку мочевого пузыря, некоторое время идут косо в
толще его стенки между мышечной и слизистой оболочками и открываются в
его полость в области шейки. Из-за этого при растяжении мочевого пузыря
поступающей мочой мочеточники ущемляются и поступление мочи в мочевой пузырь прекращается. Мочеточники имеют хорошо развитую мышечную
оболочку. Благодаря ее перистальтическим сокращениям (1—4 раза в минуту) моча прогоняется по мочеточнику к мочевому пузырю.
Мочевой пузырь — vesica urinaria — полый орган грушевидной формы.
В нем различают краниально направленную верхушку, основную часть —
тело и суженную, каудально направленную шейку. Ненаполненным он лежит
на дне тазовой полости. При наполнении верхушка мочевого пузыря опускается в лонную область. Шейка мочевого пузыря переходит в мочеиспускательный канал.
Мочеиспускательный канал — uretra — короткая трубка, отходящая
от мочевого пузыря и впадающая в каналы половых путей. У самок открываwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ется щелевидным отверстием в вентральной стенке влагалища, после чего общий участок мочевыделительных и половых путей называется мочеполовым
преддверием или синусом. У самцов недалеко от начала мочеиспускательного
канала в него впадают семяпроводы, после чего он называется мочеполовым
каналом и открывается на головке полового члена.
Гистологическое строение мочеотводящих
органов. Мочеточники, мочевой пузырь и мочеиспускательный канал являются трубкообразными органами. Их слизистая оболочка выстлана многослойным переходным эпителием (см.
главу 7). Собственная пластинка слизистой образована рыхлой соединительной тканью. Мышечная оболочка образована гладкой мышечной тканью, хорошо развита, особенно в мочеточниках, мочевом пузыре, где она образует три
слоя. Снаружи мочеточники и краниальная
часть мочевого пузыря покрыты серозной оболочкой. Каудальная часть мочевого пузыря и
мочеиспускательный канал покрыты адвентицией.
Рис. 118. Органы мочеотделения лошади:
1 — левая почка; 2 —
правая почка; 3 — мочеточник; 4 — мочевой пузырь; 5 — мочеиспускательный канал; 6 — аорта; 7 — почечная артерия; 8 — надпочечник.
Вопросы для самоконтроля. 1. Фило- и
онтогенез выделительной системы. 2. Анатомогистологическое строение почек сельскохозяйственных животных. 3. Какова структура и
функция нефрона и его частей? 4. Строение мочеточника, мочевого пузыря, мочеиспускательного канала.
Глава 18. СИСТЕМА ОРГАНОВ
РАЗМНОЖЕНИЯ (ПОЛОВОЙ АППАРАТ)
Половая система плацентарных млекопитающих включает в себя: половые железы —
семенники у самцов и яичники у самок, в которых развиваются половые клетки и вырабатываются половые гормоны; половые пути — семяпроводы у самцов и яйцеводы у самок, по которым проходят половые продукты; органы совокупления—
половой член у самцов, влагалище у самок — для внутреннего оплодотворения. У самок, кроме того, развивается орган внутриутробного развития плода
— матка.
Половая система обеспечивает передачу жизни от родителей к потомству из поколения в поколение, то есть является системой, обеспечивающей
существование вида. Это ее основная функция и потому половую систему
иногда называют системой видовой жизни. Половые гормоны участвуют в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
регуляции белкового, липидного обменов, являются анаболическими гормонами и определяют развитие вторичных половых признаков.
КРАТКИЕ СВЕДЕНИЯ О РАЗВИТИИ СИСТЕМЫ ОРГАНОВ
РАЗМНОЖЕНИЯ
Филогенез. Воспроизведение себе подобных является одним из фундаментальных признаков живого. Одноклеточные организмы размножаются
путем деления, которое время от времени сменяется прообразом полового
размножения — конъюгацией (временным соединением двух особей) или копуляцией (постоянным слиянием двух особей).
Половое размножение, или сингамия, то есть размножение с помощью
специализированных мужских и женских половых клеток, появляется у многоклеточных организмов, закрепляется и совершенствуется в процессе эволюции. В филогенезе раньше других элементов половой системы появляются
половые клетки — гаметы. У некоторых многоклеточных (низшие растения)
гаметы внешне не отличаются друг от друга. Такие организмы называются
изогамными. Организмы, гаметы которых различаются своими морфофизиологическими признаками называются анизогамными (большинство растений
и животных). У губок, например, уже можно различить яйцеклетки— макрогаметы и спермин — микрогаметы, но у них еще нет специальных половых
органов. Гаметы могут образовываться в различных участках тела и, созревая, выпадают во внешнюю среду. Если в организме развиваются и мужские,
и женские гаметы, такие организмы называются гермафродитными. Более
совершенная форма полового размножения — гонохоризм, когда мужские и
женские гаметы развиваются в разных организмах.
Вторым приобретением в филогенезе половой системы явилось образование гонад — органов, в которых развиваются гаметы. Уже у медуз половые
клетки созревают в определенных местах. Такие гонады имеют вид замкнутых мешочков и не имеют путей для выведения половых продуктов. Они
прорываются в кишку, целомическую полость в момент выхода из них зрелых гамет. Подобное строение имеет половая система и у кольчатых червей,
у которых гонады открываются в нефридиальные канальцы, и у низших хордовых. Связь половой и выделительной системы прослеживается на протяжении всего филогенеза. У ланцетника, который является раздельнополовым, гонады имеют вид мешочков, одинаковых у обоих полов. Расположены они метамерно в стенке тела в области жаберных щелей рядом с
нефридиями. Зрелые половые продукты выпадают в околожаберную полость
через прорыв ее стенки. Как видим, у низших хордовых развиты лишь две
составные части половой системы: гаметы — половые клетки и гонады —
половые железы. Оплодотворение внешнее, развитие наружное.
Дальнейшее развитие половой системы привело к появлению трубкообразных органов, по которым продвигаются зрелые половые продукты. Связь
ыочевыделительной и половой систем сохраняется. В качестве половых путей выступают выводные протоки почки — мюллеровы у самок и вольфовы
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
— у самцов. Такую систему, состоящую из половых клеток, половых желез
(семенников и яичников) и половых путей имеют рыбы и земноводные. Оплодотворение может быть внешнее или внутреннее (в таком случае копуляция осуществляется с помощью клоаки, так как специальных копу-лятивных
органов нет), развитие зародыша наружное.
Дальнейшее усложнение половой системы мы наблюдаем у амниот,
окончательно вышедших на сушу. Семенники у рептилий и птиц по-прежнему находятся в полости тела, но путь спермиев удлинился благодаря появлению придатка семенника. В его канальцах происходит накопление и дозревание спермиев. Впервые в эволюции у самцов некоторых видов рептилий
появляются в области клоаки специальные, но еще очень примитивные органы совокупления, обеспечивающие внутреннее оплодотворение. Развитие зародыша наружное. В силу этого у самок яйцеводы достигают высокой степени дифференцировки, в них секретируются вещества, идущие на образование
третичных оболочек яйцеклетки.
У плацентарных млекопитающих половая система стала еще более
сложной, обеспечивая не только развитие гамет, их транспортировку и внутреннее оплодотворение, но и внутриутробное вынашивание плода. Семенники и их придатки у большинства млекопитающих опускаются из брюшной
полости и находятся в специальном семенниковом мешке. Развиваются добавочные половые железы, половой член, препуций. У самок появляется орган для вынашивания плода — матка.
Онтогенез. Органы размножения у млекопитающих закладываются в
конце зародышевого — начале предплодного периода в виде двух мезодермальных половых валиков на вентромедальных сторонах промежуточных
почек. Несмотря на различия в генетическом поле зародыша (наличие XYполовых хромосом у самца и ХХ-хромосом у самки), начальные этапы развития семенников и яичников одинаковы (рис. 119). Это стадия индифферентных гонад, когда они у зародышей обоих полов имеют вид округлоовальных скоплений мезенхимы, покрытых мезотелием промежуточной почки. У зародыша теленка гонады закладываются на 34—38-й день, у овцы —
на 23-й день. Рядом с протоком промежуточной почки закладывается сначала
в виде продольной складки мезодермального эпителия, а затем — трубки
проток предпочки — мюллеров проток. Оба протока сохраняются после инволюции провизорных (временных) почек и входят в состав половой системы. Различия гонад по полу начинают проявляться у зародыша теленка с
40—45-го дня, у овцы — с 35-го.
Развитие органов размножения самца. Когда индифферентная гонада
превращается в семенник, в ней очень рано (у 45—55-дневного зародыша теленка) путем врастания мезотелия закладываются семенные извитые канальцы и эндокринные клетки, вырабатывающие тестостерон. Под влиянием
этого мужского полового гормона происходит дифференцировка органов половой системы самца. Промежуточная почка редуцируется. Часть ее канальцев вступает в тесную связь с канальцами растущего семенника и обwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
разует прямые канальцы, объединенные в семенниковую сеть. Часть канальцев промежуточной почки не проникает внутрь семенника.
Рис. 119. Развитие половых желез:
А—схема; Б — гистосрез через закладку гонады; I — индифферентная стадия; последующее развитие у самок (II) и у самцов (III); 1 — дефинитивная
почка; 2— дефинитивный мочеточник; 3 — мюллеров канал; 4— закладка
гонады; 5— промежуточная почка (вол-фово тело); 6— проток промежуточной почки (вольфов); 7— мочевой пузырь; 8 — отверстия мочеточников;
9 — отверстия вольфова протока; 10 — мочеполовой синус; 11 — половой
бугорок; 12 — половая щель; 13 — половые валики; 14 — проток аллантоиса: 15 — яичник; 16 — придаток яичника; 17 — яйцепровод; 18— связка
яичника; 19 — редуцированный вольфов проток; 20 — матка; 21 — мочеиспускательный канал; 22 — клитор; 23 — семенник; 24 — придаток семенника; 25 — семяпровод; 26 — связка семенника; 27 — редуцированный мюллеров проток: 28 — добавочные половые железы; 29 — половой член; 30 —
мошонка; 31 — корковое и 32 — мозговое вещество гонады; 33 — пфлюгеровские мешки; 34 — семенные извитые канальцы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Находясь рядом с семенником, они входят в состав его придатка в качестве выносящих канальцев семенника. Зачаток придатка становится заметен
у плода бычка
на 3-м месяце. Проток промежуточной почки — вольфов проток — превращается в проток придатка и семяпровод. Мюллеров проток превращается
в редуцированную мужскую матку. Семенник связан с брюшной стенкой —
направляющей связкой, часть которой затем преобразуется в паховую связку.
Другой конец паховой связки закрепляется на брюшной стенке, в месте формирования мошонки. В результате роста плода и сокращения паховой связки
семенник вместе с придатком перемещается в мошонку начиная с 4-месячного возраста плода бычка, увлекая за собой оба листка серозной оболочки. В
процессе перемещения семенники проходят через паховые каналы, которые
затем сужаются и семенники остаются в мошонке. Опускание семенников в
мошонку происходит к концу плодного периода, перед рождением или даже
после рождения.
Наружные половые органы закладываются в зародышевый период н
сначала имеют вид индифферентного полового возвышения, расположенного
по средней линии впереди отверстия клоаки. Вскоре оно превращается в половой бугорок, с двух сторон которого образуются половые складки, а позади
— половые валики. К этому времени клоака разделяется на прямую кишку и
мочеполовой синус. Отверстие мочеполового синуса видно между половыми
складками. У плода самца половой бугорок сильно удлиняется и превращается в половой член, половые складки срастаются, образуя мочеполовой канал,
половые валики становятся мошонкой.
Развитие органов размножения самки. При дифференцировке индифферентной гонады в сторону яичника врастающие тяжи мезотелия образуют
пфлюгеровские мешки. Мигрирующие первичные половые клетки вселяются
и гонаду. Отсутствие Y-хромосомы и тестостерона стимулирует развитие половых структур по женскому типу. В яичнике размножаются половые .клетки
и формируются фолликулы. От промежуточной почки сохраняется яичниковая сеть, которая затем тоже подвергается редукции. Протоки промежуточной почки редуцируются, зато мюллеровы протоки прогрессивно
развиваются и дифференцируются. Из передних участков мюллеровых протоков образуются яйцеводы, из средних — матка, на задних — влагалище,
открывающееся в мочеполовой синус. Деление матки на отделы становится
заметным у плода овцы на 4-м месяце. В яичнике в это время уже имеются
первичные, вторичные и третичные фолликулы.
Типы маток. В зависимости от степени срастания мюллеровых протоков
различают несколько типов маток (рис. 120). Если мюллеровы протоки не
слиты на всем протяжении, образуется двойная матка с двойным влагалищем
(сумчатые), а если они срослись каудальными частями с образованием одного влагалища, то матка двойная. Она может открываться двумя или одним
(слившимся) отверстиями во влагалище (хоботные, многие грызуны). При
слиянии задних и частично средних участков моллюровых протоков образуются матки: двураздельная — когда шейка матки единая, а тело на большую
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
глубину разделено внутренней перегородкой (жвачные, оленевые, мозоленогие); двурогая — когда единые шейка и тело матки и парные рога (хищные,
свиньи, непарнокопытные). При слиянии задних и средних частей мюллеровых каналов образуется простая матка, в которую впадают парные яйцеводы
(приматы, некоторые рукокрылые).
Рис. 120. Типы маток:
А—двойная с двойным влагалищем; Б— двойная; В — двураздельная; Г —
двурогая: Д — простая; 1 — яичник; 2 — яйцевод; 3 — рог, 4 — тело и 5 —
шейка матки; 6 — влагалище; 7 — клоака; 8 — отверстие мочеиспускательного канала; 9 — мочеполовой синус.
Наружные половые органы закладываются, как и у самцов, в виде полового бугорка, половых складок и половых валиков. Половой бугорок становится клитором, половые складки и валики — половыми губами.
Изменение половой системы в постнатальном онтогенезе. За время внутриутробного развития семенники увеличиваются по массе в 250 раз, по объему— в 350 раз. У новорожденных бычков масса семенников 8—10 г, В первые месяцы после рождения их рост происходит медленно. С 6 месяцев начинается сперматогенез. В период полового созревания масса семенников,
диаметр семенных канальцев и толщина сперматогенного эпителия интенсивно нарастают. У взрослых быков семенники весят 350—500 г при длине
10—11 см. Полного развития семенники достигают у быков к двум, у баранов — к 1,5 года, у хряков — к 1 году. Наблюдается прямая корреляция между массой животного, массой семенников, количеством полевых клеток и активностью сперматогенеза. Гиподинамия приводит к полнокровию сосудов
семенника, избыточному накоплению зрелых спермиев в канальцах семенника и придатка, торможению сперматогенеза. После 9 лет у быков происходит
снижение активности сперматогенеза, а с 15 лет — угасание.
У новорожденных телочек масса яичника составляет 0,3—1 г, поверхность может быть неровной из-за возвышающихся отдельных третичных
фолликулов. Первые 1—3 месяца яичники активно растут, в них совершаются процессы перестройки: часть фолликулов подвергается атрезии (обратному развитию), другие вступают в стадию роста. Следующий скачок в росте
половых органов происходит в период полового созревания: у 10—14-месячwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ных телок и ярок. Овуляция у телок наблюдается с 18—20 месяцев, у ярок —
с 15 месяцев. Расцвет приходится на 3—6-й год у коровы, 2,5— 4-й год — у
овцы. В это время увеличиваются размеры яичников, матки и влагалища.
Утолщается мышечная оболочка органов, увеличивается количество желез. В
яичнике одновременно растет до нескольких десятков пузырчатых фолликулов, в результате чего масса яичника увеличивается до 7—10 г. У овец возрастает многоплодие до 4,5 года.
С 6—7 лет у овцы, с 10—13 у коровы наблюдаются изменения в структуре органов размножения, снижающие их функциональную активность. В
яичнике разрастается соединительная ткань, уменьшаются размеры фолликулярной зоны (коркового вещества). Резко снижается количество первичных фолликулов (в 10—25 раз по сравнению с новорожденными). В яйцеводах увеличиваются и усложняются складки, происходит их сращение,
уменьшается количество реснитчатых клеток. Истончаются стенки яйцевода
и матки, в них уменьшается количество желез и мышечных пучков, разрастается соединительная ткань. Тем не менее способность к плодоношению сохраняется у овец до 8—9 лет, у коров —до 13—15 лет. У самок половые органы наряду с возрастными подвергаются ярко выраженным циклическим
изменениям, связанным с половым циклом, беременностью, родами. Чрезвычайно резко реагируют яичники на стимуляцию многоплодия и суперовуляции: их масса увеличивается у телок в 10 раз (до 80 г), размеры —
до 8×3 см, число пузырчатых фолликулов достигает 54, овуляции —25.
ПОЛОВАЯ СИСТЕМА САМЦА
В нее входят семенники, придатки семенников, семенные канатики, семяпроводы, добавочные половые железы, семенниковый мешок, мочеполовой канал, половой член и препуций. Семенник — лат. testis, греч. didymis —
парный орган эллипсоидной формы, в котором у половозрелых животных
происходит сперматогенез и вырабатываются половые гормоны (рис. 121). С
ним тесно связан придаток семенника. На семеннике различают: свободный и
придатковый края, головчатый конец, с которым связана головка придатка;
хвостатый конец, к которому прилежит хвост придатка; латеральную и медиальную поверхности.
У быка головчатый конец семенника направлен дорсально, хвостатый —
вентрально. Придатковый край обращен каудально. Латеральная поверхность
выпуклая, медиальная— уплощена. У хряка головчатый конец направлен
краниовентрально, хвостатый — каудодорсально, придатковый край — дорсально. У жеребца семенники заметно сдавлены с боков, головчатый конец
направлен краниально, хвостатый — каудально, придатковый край — дорсально. Гистологическое строение семенника. Семенник — компактный орган, снаружи одетый серозной или собственной влагалищной оболочкой (А).
Под ней расположена белочная оболочка, образованная плотной соединительной тканью. В области головчатого конца соединительная ткань от белочной оболочки проникает в центр семенника, формируя средостение сеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
менника. От средостения во все стороны отходят тонкие соединительнотканные перегородки — септы, или трабекулы, которые делят семенник на дольки. Белочная оболочка, средостение и септы — это строма семенника. Паренхима органа состоит из долек.
Рис. 121. Схема строения семенника (А) и его придатка (Б):
а — головчатый и б — хвостатый
конец семенника; в — свободный и г
— придатковый край семенника; д
— головка, е — тело и ж — хвост
придатка; 1 — собственная влагалищная оболочка; 2— белочная оболочка; 3 — междольковая трабекула; 4 — долька; 5 — семенные извитые канальцы: 6 — прямой каналец;
7— средостение с сетью семенника; 8 — выносящие канальцы; 9 —
проток придатка; 10 — семяпровод; 11 — семенной канатик.
Каждая долька образована 2—3 извитыми семенными канальцами и расположенной между ними интерстициальной тканью с небольшим количеством рыхлой соединительной ткани. Каждый извитой семенной каналец имеет
длину до нескольких десятков сантиметров, ширину 0,1—0,2 мм, покрыт
тонкой собственной соединительнотканной оболочкой. Его стенка состоит из
сперматогенного эпителия на разных стадиях развития и поддерживающих
клеток (см. раздел «Основы эмбриологии»). Сперматогенез идет волнообразно, поэтому на протяжении извитого канальца можно увидеть его разные
стадии. В интерстициальной ткани находятся группы эндокринных клеток.
Эти крупные светлые клетки с крупными круглыми ядрами вырабатывают
мужские половые гормоны — андрогены (тестостерон). По выходе из долек в
средостение извитые канальцы переходят в прямые семенные канальцы. Эти
канальцы переплетаются друг с другом, образуя сеть семенника. Васкуляризация семенника осуществляется семенниковой (внутренней семенной) артерией. Иннервация — нервами вегетативной нервной системы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Придаток семенника — epididymis — часть семявыносящих путей (Б).
Состоит из головки, тела и хвоста. Головка придатка образована 12—20 выносящими канальцами, в которые вливаются канальцы сети семенника. Выносящие канальцы впадают в проток придатка, который находится в теле и
хвосте, где сложен в систему сложных завитков. В развернутом состоянии он
имеет длину от 25 до 85 м при ширине около 1 мм. В хвосте придатка проток
расширяется, поворачивает к головчатому концу семенника и переходит в
семяпровод. В придатке семенника происходит накопление и дозревание
спермиев. Продвижение спермиев по придатку осуществляется благодаря сокращению гладкомышечных клеток его стенки. Сами спермин неподвижны.
Семенники с придатками подвешены на семенных канатиках и заключены в
семенниковый мешок.
Семенниковый мешок — saccus testicularis (рис. 122)—выпячивание
брюшной стенки со всеми ее слоями, последовательно одевающими семенники. Внутренние слои, непосредственно прилежащие к семеннику и придатку, образованы серозными оболочками. Наружные слои семенникового мешка образуют мошонку.
Опускаясь из брюшной полости в мошонку, семенники увлекают за собой висцеральный листок серозной (перитонеальной) оболочки. Тесно прилегая к семеннику и придатку, она становится специальной влагалищной оболочкой семенника и придатка семенника. Париетальный листок серозной
оболочки выстилает изнутри семенниковый мешок и вместе с листком глубокой (поперечной брюшной) фасции образует общую влагалищную оболочку
семенника и придатка. Между влагалищными оболочками имеется щелевидная влагалищная полость. Небольшое количество серозной жидкости, находящейся в полости, способствует скольжению семенников и уменьшает возможность их ущемления. Снаружи к общей влагалищной оболочке прикрепляется мышца — подниматель семенника, которая отходит от внутренней косой брюшной мышцы. Общая влагалищная оболочка рыхло соединена с мошонкой и при разрезе мошонки во время кастрации легко от нее отделяется
вместе с заключенными в нее семенником и придатком.
Мошонка—scrotum — состоит из кожи и мышечно-эластической оболочки, сросшихся между собой. Внутренний слой мошонки— мышечноэластическая оболочка образована за счет фасции, подкожного слоя и гладкой мышечной ткани. Она образует перегородку мошонки, разделяющую полость мошонки на 2 камеры. В каждой камере заключено по одному семеннику с придатком. Кожа мошонки розоватая или пигментированная, содержит потовые и сальные железы и покрыта редкими короткими волосами.
Расположена мошонка у быка, барана, жеребца между бедрами, у хряка —
каудальнее бедер. Расстояние между анусом и мошонкой называется промежностью. У хряка промежность короткая, у быка — наиболее длинная.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 122. Половая система:
А — быка; Б — хряка; В — жеребца; Г — семенниковый мешок быка; 1 —
пузырьковидная железа; 2— ампула семяпровода; 3— мочевой пузырь; 4 —
семяпровод; 5 — сосуды и нервы семенного канатика; 6 — пенис; 7 —
препуций; 8 — головка пениса; 9 — мочеполовой канал; 10— кожа мошонки;
11 — анальное отверстие; 12 — предстательная железа; 13 — луковичная
железа; 14 — тазовая часть мочеполового канала; 15 — промежность; 16
— мочеиспускательный канал;17 — кожа пениса; 18 — мышца — ретрактор пениса; 19 — лонная кость; 20 — семенной канатик; 21 — семенник; 22
— придаток семенника; 23 — дивертикул препуция; 24 — мышечноэластическая оболочка мошонки; 25 — фасция мошонки; 26 — наружный
подниматель семенника; 27 — общая влагалищная оболочка; 28 — влагалищная полость; 29 — специальная влагалищная оболочка; 30 — перегородча
мошонки.
Семенной канатик — funiculus spermaticus (20)—это складка брыжейки
семенника в виде сплющенного конуса, куда заключены сосуды, нервы,
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мышца — внутренний подниматель семенника и семяпровод. Семенной канатик отходит от придатка семенника и через паховый канал проникает в
брюшную полость, где его составные части расходятся в разные стороны.
Семяпровод—ductus deferens (4)—отходит от хвоста придатка, идет в
составе семенного канатика. Достигнув брюшной полости, направляется дорсокаудально в тазовую полость, проходит дорсально от мочевого пузыря и
впадает в мочеиспускательный канал, который с этого места называется мочеполовым. Семяпровод — парный узкий трубкообразный орган. Его стенка
состоит из слизистой, мышечной и серозной оболочек. Спермин передвигаются по семяпроводу благодаря сокращению его мышечной оболочки. Конечный участок семяпровода у быка, барана и жеребца утолщен. Это железистая часть или ампула семяпровода. В ее стенке развиваются железы, секрет
которых, добавляясь к семени, питает спермин.
Мочеполовой канал — canalis urogenitalis (рис. 123)—или мужская
уретра служит для выведения мочи и спермы. Начинается с впадения семяпроводов в мочеиспускательный канал и заканчивается на головке полового
члена. В нем различают тазовую и половочленную части. Тазовая часть лежит на дне тазовой полости, каудально доходит до седалищной дуги, перегибается через нее, переходит на вентральную поверхность полового члена—
становится половочленной частью мочеполового канала. В тазовую часть
мочеполового канала открываются добавочные половые железы. Их секрет
вместе со спермиями составляет сперму. Полозочленная часть проходит в
вентральной части полового члена. Конец мочеполового канала у быка и
хряка не выходит за пределы головки полового члена. У барана образует мочеполовой отросток, выходящий за пределы головки. Стенка мочеполового
канала образована слизистой оболочкой, кавернозным телом и мышечной
оболочкой. Слизистая оболочка покрыта многослойным эпителием. Кавернозное тело представляет собой соединительнотканную оболочку с большим
количеством коллагеновых и эластических волокон. В этом слое заложено
густое венозное сплетение с расширениями — лакунами. При их наполнении
кровью расширяется просвет мочеполового канала.
Добавочные половые железы: пузырьковидные, предстательная, луковичные (см. рис. 122).
Пузырьковидная железа — парная, сложная, трубча-то-альвеолярная. У
быка, барана и хряка крупная, бугристая, длиной 8—15 см, у жеребца гладкая, грушевидная. Расположена дорсальнее мочевого пузыря. Главный выводной проток железы открывается вместе с семяпроводом в начало мочеполового канала. Железа вырабатывает густой клейкий секрет. Высказывается
мнение, что он служит для питания спермиев, а также может образовывать
влагалищную пробку в половых путях самки, препятствующую вытеканию
спермы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 123. Строение полового члена и препуция:
А—головка полового члена быка; Б — барана; В —хряка; Г — поперечный
разрез полового члена быка; Д — головка пениса и препуций жеребца; 1 —
отросток мочеполового канала; 2— колпачок головки; 3— шов головки; 4—
шейка головки; 5— шов препуция; 6 — препуциальный мешок; 7 — венчик головки; 8 — листок препуция, переходящий в брюшную стенку; 9 — внутренний препуций; 10 — мочеполовой канал; 11 — наружный препуций; 12 —
кавернозное тело пениса; 13 — кавернозное тело мочеполового канала; 14 —
луковично-кавернозная мышца; 15 — сосуды пениса; 16 — белочная оболочка
пениса.
Предстательная железа состоит из застенной и пристенной частей. Застенная часть — тело железы длиной 2—4 см расположено поперек шейки
мочевого пузыря и мочеполового канала. У барана тело железы не развито. У
жеребца с двух сторон от тела (перешейка) отходят крупные боковые доли.
Пристенная, или рассеянная, часть залегает в стенке тазовой части мочеполового канала. Железа сложная, альвеолярно-трубчатая, протоки ее открываются в мочеполовой канал. Ее секрет поддерживает двигательную активwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ность спермиев и, по некоторым данным, нейтрализует кислую среду влагалища.
Луковичная (бульбоуретральная) железа парная, сложная, альвеолярно-трубчатая, расположена на конце тазовой части мочеполового канала. У
быка и жеребца имеет размеры 3—4 см, у барана —2—3 см. Прикрыта луковично-ка-вернозной мышцей. У хряка длина железы достигает 12 см. Секрет
ее, вероятно, предохраняет спермин от остатков мочи в мочеполовом канале.
Половой член — penis (рис. 122, 123)—орган совокупления. В нем различают корень, тело и головку. На всем протяжении он построен из плотной
соединительной ткани и кавернозных тел и содержит мочеполовой канал.
Плотная соединительная ткань формирует белочную оболочку, от которой отходят перегородки — трабекулы. В трабекулах много коллагеновых и эластических волокон, гладкомышечных клеток. Между трабекулами образуются ходы и расширения, выстланные эндотелием, — каверны. Обильный приток крови к кавернам во время полового возбуждения приводит к
увеличению и напряжению полового члена— эрекции. В дорсальной стороне
пениса проходят сосуды. На вентральной его стороне имеется желобок, в котором залегает мочеполовой канал.
У быка половой член длинный (до 100 см) и сравнительно тонкий (2,5—
3 см). Корень пениса начинается ножками от седалищных бугров, прикрыт
луковично-кавернозными мышцами. Объединяясь, ножки переходят в длинное тело с S-образным изгибом. Кавернозные тела в пенисе быка развиты
слабо, поэтому при эрекции не происходит заметного утолщения пениса. Удлиняется он за счет распрямления S-образного изгиба. Возвращается в исходное положение в результате сокращения мышцы ретрактора пениса, которая закрепляется одним концом на крестцовых и хвостовых позвонках, другим — на S-образном изгибе пениса. Перед головкой пенис сужается — формируется шейка головки. На головке пениса быка, барана, козла различают
колпачок головки. У быка мочеполовой канал не выходит за пределы колпачка головки. Конечный его участок проходит в спиралеобразном желобе с
левой стороны головки. У барана и козла мочеполовой канал выходит на 2—
3 см дальше головки в виде узкого (2—3 мм), у барана изогнутого, у козла
прямого отростка.
У хряка половой член по строению сходен с пенисом быка, но меньших
размеров (45—50 см) и головка спиралевидно закручена.
У жеребца половой член не имеет S-образного изгиба, тело уплощено с
боков, хорошо развиты кавернозные тела в теле и головке. Головка широкая
с утолщением на конце — венчиком головки. Мочеполовой канал не выступает за пределы головки — открывается в ямке головки. При эрекции пенис
сильно увеличивается в объеме и длине, диаметр венчика головки достигает
12— 15 см. Мышца — ретрактор пениса — тянется по вентральной стороне
пениса от первых хвостовых позвонков до головки пениса.
Корень и тело полового члена закрыты снаружи кожей туловища. В области головки кожа образует складку — препуций.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Препуций — praeputium, или крайняя плоть, — складка кожи, скрывающая передний конец пениса. Препуций быка и хряка имеет длину 30—45
см, у барана — 10—18 см. В препуции самцов рогатого скота и свиньи различают 2 листка — наружный и внутренний. Оба они имеют такое же строение, как и кожа. Внутренний листок переходит в слизистую оболочку мочеполового канала, содержит трубчатые препуциальные железы и лимфатические узелки. Между листками препуция имеется щелевидное пространство.
Наружное отверстие препуция расположено немного дальше пупка и окружено длинными волосами. Передняя часть препуция более широкая, задняя
— узкая. Между ними — кольцевые поперечные складки, сокращаясь они
отгораживают препуций от окружающей среды. Препуций хряка в дорсальной стенке имеет мешковидное выпячивание — дивертикул, достигающий
глубины 9 см и ширины 12 см. Препуций жеребца двойной — наружный и
внутренний. Каждый из них состоит из двух листков— наружного и внутреннего. При эрекции внутренний препуций расправляется, допуская значительное удлинение пениса.
ПОЛОВАЯ СИСТЕМА САМКИ
В половую систему самки входят яичники, яйцеводы, матка, влагалище,
мочеполовое преддверие (синус), наружные половые органы.
Яичник — ovarium (рис. 124, 125) — парный орган, в котором у половозрелых животных происходит оогенез н вырабатываются половые гормоны. У коровы эллипсоидной формы, с гладкой поверхностью, 2×4 см в диаметре, массой 5—10 г. Краниальным концом яичник обращен к воронке яйцевода, каудальным — к матке, дорсальным краем прикреплен к брыжейке,
которая формирует связку яичника, вентральный край свободен. По всему
свободному краю происходит овуляция. Преовуляторные пузырчатые фолликулы выпячиваются над поверхностью яичника в виде бугорков диаметром
от 0,5 до 2 см. Подвешены яичники на уровне крестцовых бугров подвздошной кости. У свиньи, животного многоплодного яичники бугристые.
Расположены на уровне 5— 6-го поясничного позвонка, имеют массу 5— 16
г. У кобылы яичник крупный (5— 8 см), массой до 70— 80 г, бобовидной
формы, почти весь покрыт серозной оболочкой. Лишь в одном месте — в области овуляционной ямки — происходит овуляция фолликулов. Расположены
яничики позади почек. Иннервация — яичниковым сплетением вегетативной
нервной системы. Васкуляризация — яичниковой артерией.
Гистологическое строение яичника. Яичник — компактный орган. Сверху покрыт поверхностным эпителием — производным мезотелия полового
валика. Под ним залегает слой плотной соединительной ткани — белочная
оболочка яичника. За исключением белочной оболочки, строма яичника образована рыхлой соединительной и ретикулярной тканями. В яичнике различают корковое вещество, или фолликулярную зону, и мозговое вещество, или
сосудистую зону.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 124. Схема строения яичника:
1—связка яичника: 2— поверхностный эпителий: 3 — белочная
оболочка; 4 — примордиальный, 5
— первичный, 6 — растущий и 7 —
пузырчатый фолликулы; 8 — атретическое тело; 9 —зрелый фолликул; 10 — тека; 11 — гранулеза;
12 — яйценосный бугорок с ооцитом I порядка; 13 — полость фолликула; 14 — овулировавший фолликул; 15 — ооцит II порядка; 16
— желтое тело в стадии формирования и 17 — расцвета; 18 — беловатое тело; 19 — кровеносные
сосуды мозгового вещества.
Корковое вещество у большинства животных лежит под белочной оболочкой по периферии яичника, у кобылы — в центре яичника, лишь в области овуляционной ямки выходит на периферию. Строма коркового вещества
образована соединительной тканью с незначительным количеством волокон
и большим количеством крупных веретеновидных клеток. Основную массу
коркового вещества составляют фолликулы на разных этапах стадии роста.
Ближе к белочной оболочке залегают примордиальные и первичные фолликулы.
Примордиальные фолликулы—мелкие округлые структуры, в которых
ооцит I порядка окружен одним слоем низких фолликулярных клеток. Число
примордиальных фолликулов у половозрелых животных постоянно уменьшается, так как часть их вступает в рост и становится первичными фолликулами. В первичных фолликулах ооцит I порядка окружен одним слоем кубических или цилиндрических фолликулярных клеток. Последние размножаются и одевают ооцит I порядка несколькими слоями. Они питают половую клетку и вырабатывают гормон эстроген. Образуется вторичный (растущий) фолликул. Эти фолликулы и их ооциты I порядка крупнее первичных
и располагаются в корковом веществе глубже.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 125. Половая система коровы (Л), кобылы (Б), свиньи (В):
1— яичник; 2— воронка яйцевода; 3— яйцевод; 4 — связка яичника; 5 — рог.
6— тело и 7 — шейка матки; 8 — влагалищное отверстие шейки; 9 — влагалище; 10 — отверстие мочеиспускательного канала; 11 — мочеполовой синус; 12 — преддверные железы; 13 — губа; 14 — клитор; 15 — вентральная
комиссура; 16 — широкая маточная связка; 17 — мочевой пузырь.
В процессе деятельности фолликулярных клеток между ними накапливается жидкость. Образуется пузырчатый (третичный) фолликул, имеющий
крупные размеры и сложное строение (см. раздел «Основы эмбриологии»).
Стенка его образована зернистым слоем из фолликулярного эпителия — гранулезой, базальной мембраной и соединительной оболочкой — текой. У коровы полость появляется в фолликулах с диаметром 1 мм. Пузырчатый фолликул быстро растет, распространяясь не только на всю глубину коркового
вещества, но и, выпячиваясь наружу, достигает в диаметре 15—18 мм и становится зрелым фолликулом, готовым к овуляции. В момент овуляции созревающая половая клетка выпадает в просвет яйцевода, а полость фолликула
заполняется кровью. В нее прорастают клетки текальной оболочки, видоизменяются фолликулярные клетки — они превращаются в лютеоциты —
клетки, вырабатывающие гормон прогестерон — появляется желтое тело —
временная железа внутренней секреции. Если беременность не наступила,
желтое тело редуцируется, а если наступила — желтое тело развивается и
поддерживает ее до развития плаценты. В период расцвета плаценты (у овец
— на 3—4-м, коров на 4—5-м месяце беременности) желтое тело беременности редуцируется, замещается соединительной тканью — становится беловатым телом, которое может существовать несколько лет, постепенно рассасываясь.
В корковом веществе яичника наряду с ростом фолликулов происходит
и обратный процесс — их гибель, или атрезия. Механизм и причины атрезии
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
до конца неясны, несмотря на то, что атрезии подвергается большое количество фолликулов на всех этапах их роста. Атретический фолликул часто неправильной формы, содержит атипичные фолликулярные клетки, погибшую
половую клетку. В ходе инволюции атретических фолликулов на их месте
остаются скопления интерстициальных клеток. Есть интерстициальные клетки и между растущими фолликулами. Они, как и клетки фолликулярного
эпителия, и атретического тела, вырабатывают гормон эстроген. Сочетанным
действием гормонов эстрогена и прогестерона под регулирующим влиянием
гонадотропных гормонов гипофиза осуществляются процессы роста и созревания половых клеток, рост и атрезия фолликулов, овуляция.
Мозговое вещество лежит у всех сельскохозяйственных животных (кроме кобылы) в центре яичника. У кобылы оно расположено по периферии органа (за исключением овуляционной ямки яичника). Строма мозгового вещества образована рыхлой соединительной тканью. В мозговом веществе разветвляется большое количество сосудов, проникающих в яичник со стороны
брыжейки. Их ветви из мозгового вещества проникают в корковое, принося к
фолликулам питательные вещества.
Яйцепровод (яйцевод)—oveductus (маточная труба) (рис. 125)—парный
трубкообразный орган, в котором женские половые клетки завершают стадию созревания, происходит оплодотворение и начинается дробление зиготы.
Он имеет вид извилистой трубки, расположенной между яичником и маткой,
длиной у коровы 20—30 см, овцы—15—20, свиньи—15—30, кобылы — 10—
30 см с двумя отверстиями: яичниковым и маточным. На яичниковом конце
имеется воронкообразное расширение — воронка яйцевода с изрезанными
краями — бахромкой воронки. В него из яичника в момент овуляции попадает яйцеклетка. Маточное отверстие яйцепровода открывается в конце рога
матки. Яйцепровод лежит в брыжейке яйцепровода, которая является частью
широкой маточной связи.
Стенка яйцепровода образована слизистой, мышечной и серозной оболочками. Слизистая оболочка выстлана однослойным мерцательным эпителием. Он состоит из реснитчатых и безреснитчатых — секреторных клеток,
вырабатывающих слизистый секрет. Собственная пластинка слизистой оболочки образована рыхлой соединительной тканью с многочисленными продольными складками. Мышечная оболочка образована гладкой мышечной
тканью и состоит из двух слоев. Кольцевой слой развит хорошо, продольный
слой лучше развит у маточного конца яйцепровода. Серозная оболочка имеет
типичное строение. Движение яйцеклетки по яйцеводу осуществляется током
слизи, движением ресничек эпителия и сокращением мышечной оболочки.
Матка — лат. uterus, греч. metra (рис. 125) — непарный орган для внутриутробного развития зародыша и плода. У сельскохозяйственных животных
относится к типу двурогих. Она состоит из рогов, тела и шейки. У коров рога
матки имеют длину 20— 30 см, закручены спиралью в виде рогов барана и к
концу заострены. В краниальный заостренный конец каждого рога открывается маточным отверстием яйцепровод. Каудальные концы рогов сливаются, переходят в непарное тело матки. Снаружи тело матки имеет длину окоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ло 10 см, но внутри его проходит перегородка, в результате чего полость его
равна 5—6 см и даже 1— 3 см.
Рога и тело матки содержат полость матки, которая переходит в канал
шейки матки. Шейка толстостенная, длиной 7— 11 см. Канал шейки обрамлен продольными складками, что способствует замыканию канала. В шейке
имеется внутреннее маточное отверстие, открывающееся в тело, и наружное маточное отверстие, окрывающееся во влагалище. Шейка несколько
выступает во влагалище, образуя влагалищную порцию матки.
Матка подвешена на маточной брыжейке или широкой маточной связке,
которая краниально продолжается в брыжейку яичника и яйцепровода. Матка расположена в лонной области. При беременности распространяется в
правую половину брюшной полости. Иннервируется тазовыми нервами подчревного сплетения. Васкуляризуется передней, средней и задней маточными
артериями.
У свиньи рога матки достигают 2—3 м и образуют петли, подобные петлям кишечника. Тело короткое — до 5 см. Шейка длинная—15—18 см, с
волнообразными складками, плотно закрывающими канал шейки. Шейка без
границ переходит с одной стороны в тело матки, с другой — во влагалище.
У кобылы рога направлены краниовентрально, широкие. Переход в яйцепровод обозначен резко, так как краниальный конец рога тупой. Тело матки по длине лишь немного меньше рогов. Шейка матки с продольными
складками и значительной влагалищной порцией.
Гистологическое строение матки (рис. 126). Матка — трубкообразный
орган. Ее стенка образована слизистой, мышечной и серозной оболочками.
Слизистая оболочка — эндометрий — выстлана у большинства млекопитающих однослойным цилиндрическим эпителием, вырабатывающим
слизь. В определенные периоды цикла эпителий может меняться на многорядный или многослойный. Встречаются в нем и реснитчатые клетки, особенно в рогах матки. Собственная пластинка слизистой образована соединительной тканью с большим количеством разнообразных клеток. В ней в области рогов и тела залегают простые разветвленные трубчатые железы, секретирующие по мерокринному типу. Секрет желез используется для питания
зародыша на ранних этапах развития. У жвачных стенка слизистой образует
выпячивания — карункулы, богатые кровеносными сосудами, но не содержащие желез. В период беременности они становятся крупными макроскопическими образованиями и как шапочками покрываются выростами хориона — котиледонами. Слизистая оболочка шейки образует многочисленные
крупные и мелкие складки, продольные у крупного рогатого скота и лошади,
волнообразные у свиньи. В ее эпителии много слизистых клеток.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 126. Поперечный разрез рога матки овцы (А) и соединение карункула с котиледоном у коровы (Б):
I — слизистая оболочка; 1 — эпителий; 2— железы; 3 — карункул; II — мышечная оболочка; 4 — кольцевой, 5 — сосудистый и 6 — продольный слои; III
— серозная оболочка; 7— широкая маточная связка; 8 — котиледон; 9 — хорион.
Мышечная оболочка — миометрий — образована гладкой мышечной
тканью, расположенной в два слоя. Гладкие миоциты миометрия имеют
крупные размеры (до 0,5 мм), могут ветвиться. Внутренний кольцевой слой
развит лучше продольного. В области шейки кольцевой слой особенно мощный, формирует сфинктер, удерживающий канал шейки в закрытом состоянии. Открывается канал во время охоты и при изгнании плода. Между кольцевым и продольным слоями у хищных, а из сельскохозяйственных животных у жвачных, особенно в области рогов, заметен средний сосудистый слой,
в котором разветвляется большое количество сосудов. У свиньи этот слой не
выражен, у кобылы заметен плохо. Мышечная оболочка матки сильно утолщается во время беременности.
Серозная оболочка — периметрии — состоит из собственной пластинки
и мезотелия. Собственная пластинка, построенная из рыхлой соединительной
ткани, утолщается и перестраивается в период беременности.
Влагалище — vagina — непарный трубкообразный орган длиной у коровы 22—28 см, у свиньи — 10—12 см. Лежит оно в тазовой полости каудальнее матки. Краниально сообщается с шейкой матки, каудально переходит в
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
мочеполовое преддверие. Стенка влагалища образована мышечной, слизистой оболочками и адвентицией. Лишь небольшой краниальный участок покрыт серозной оболочкой. Слизистая оболочка влагалища выстлана многослойным плоским эпителием, не имеет желез и образует продольные складки. Мышечная оболочка состоит из кольцевого и продольного слоев, адвентиция — из рыхлой соединительной ткани, удерживает орган в его положении.
Мочеполовое преддверие (синус, преддверие влагалища)— vestibulum
urogeniiale — продолжение влагалища после впадения в него мочеиспускательного канала. Отверстие мочеиспускательного канала открывается в вентральной стенке мочеполового преддверия. Каудально мочеполовое преддверие граничит с наружными половыми органами—-вульвой. Стенка его образована теми же слоями и тканями, что и у влагалища. В слизистой оболочке
преддверия содержатся лимфатические узелки и залегают преддверные железы. У коровы вентральные, или малые, преддверные железы слабо выражены, их отверстия открываются в каудальную часть преддверия по бокам от
клитора. Латеральные, или большие, преддверные железы открываются
крупными отверстиями между складками слизистой оболочки. Мышечная
оболочка в каудальной части преддверия дополняется кольцевым слоем поперечнополосатых волокон, формирующих сжиматель преддверия.
Наружные половые органы самки — vulva—состоят из половых губ,
соединенных комиссурами, половой щели и клитора. Вульва расположена
ниже ануса. Расстояние между ними называется промежностью. Половые губы — складки кожи, в основе которых залегает мышца — сжиматель половой щели. Между половыми губами — вход в мочеполовое преддверие —
половая щель. Места соединения половых губ называют вентральной и дорсальной комиссурой, или спайкой. У коров с вентральной спайки свисает пучок волос. У вентральной комиссуры выступает головка клитора, который
является гомологом пещеристого тела пениса. Тело клитора погружено в
стенку преддверия.
Вопросы для самоконтроля. 1. Какие этапы развития проходит система
органов размножения в филогенезе? 2. Закладка и дифференцировка органов
половой системы зародыша самца и самки млекопитающего. 3. Каково анатомо-гистологическое строение семенника? 4. Опишите строение и расположение семенникового мешка, семенного канатика, семяпровода, добавочных половых желез, пениса, препуция. 5. Перечислите и опишите строение и положение органов размножения самки. 6. Каково анатомогистологическое строение яичника и матки у коровы, кобылы, свиньи? 7.
Особенности строения шейки матки и влагалища у коровы, свиньи и кобылы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Часть III. ИНТЕГРИРУЮЩИЕ СИСТЕМЫ
Регуляция процессов, происходящих в организме на всех его уровнях,
поддержание постоянства внутренней среды, адекватные реакции организма
как целостной системы на меняющиеся условия внешней среды осуществляются интегрирующими, регуляторными системами: эндокринной, сосудистой, нервной. Ведущая среди них — нервная система. Пронизывая все ткани
и органы, осуществляет тройной контроль за деятельностью организма: изменяют интенсивность работы органа, вплоть до полного ее прекращения;
действуя на сосуды, может изменять интенсивность кровоснабжения и тем
самым трофику органа, а также характер и интенсивность обмена веществ.
Эндокринная система осуществляет гормональную регуляцию функций
организма. Происходит это в тесной связи с сосудистой системой, так как
гормоны— биологически активные вещества, вырабатываемые железами
внутренней секреции, поступают в кровь и достигают органов-мишеней. Регуляция с помощью жидких тканей организма — крови и лимфы называется
гуморальной. Гуморальная регуляция функций организма очень сложна и
многообразна. Гормональная (эндокринная) регуляция является лишь ее частью, филогенетически наиболее молодой и более специализированной по
сравнению с более древними ее формами: неспецифической химической и
тканевой гуморальной регуляцией.
В подбугорной области головного мозга — в гипоталамусе — осуществляется теснейшая связь нервной и эндокринной систем. С помощью нейросекреторных нейронов этой области происходит перевод нервного импульса
в гуморальные стимулы, возбуждающие или тормозящие деятельность желез
внутренней секреции. Нервно-проводниковая и гуморальная регуляции
функционируют в тесном взаимодействии, составляя единую нейрогуморальную регуляцию и интеграцию функций организма, осуществляемую
нервной, сосудистой и эндокринной системами.
Глава 19. ЭНДОКРИННАЯ СИСТЕМА (СИСТЕМА ОРГАНОВ
ВНУТРЕННЕЙ СЕКРЕЦИИ)
Эндокринная система образована органами, основной функцией которых является выработка гормонов (греч. hormao — побуждаю, возбуждаю)—
биологически активных веществ. Гормоны стимулируют рост и развитие органов и тканей, регулируют скорость физиологических процессов, способствуют повышению сопротивляемости организма факторам внешней среды,
участвуют в поддержании гомеостаза.
Эндокринные железы не имеют протоков. Они выделяют гормоны в
кровь — во внутреннюю среду организма, и поэтому их называют еще железами внутренней секреции. Железы внутренней секреции расположены в различных местах организма, и не связаны между собой морфологически. Однако тесная функциональная связь, соподчинение и взаимная зависимость их
друг от друга заставляют говорить о железах внутренней секреции как о едиwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ной системе. К железам внутренней секреции относят: эпифиз, гипофиз, надпочечники, щитовидную и паращитовидную железы. В эндокринную систему включают эндокринные части поджелудочной железы (клетки островков
Лангерганса — вырабатывают инсулин и глюкагон) яичников и семенников
(половые гормоны), почек (ренин и др.), тимуса (тимозин), плаценты (прогестерон и др.). Сюда же включают и одиночные эндокринные клетки неэндокринных органов. Так, в желудочно-кишечном тракте выделено 15 типов
эндокринных клеток, синтезирующих до 30 гормонов, регулирующих деятельность пищеварительной системы.
Секреты эндокринных желез и клеток — гормоны имеют ряд общих
свойств. 1. Все гормоны обладают высокой биологической активностью и
действуют в чрезвычайно малых количествах. Например, в норме у взрослого
крупного рогатого скота количество оксикортикостероидов (гормонов коры
надпочечников) в 1 л крови равно 70 мкг. 2. У многих гормонов нет видовой
специфичности, но все они строго специфичны по своему действию на ткани
и органы. Органы и клетки, на которые тот или иной гормон оказывает действие, называются органы-мишени и клетки-мишени. Так, половые гормоны
способствуют развитию вторичных половых признаков, и никакие другие
гормоны не могут их заменить и вызвать те же изменения в организме. 3. Для
гормонов характерно дистантное действие: органы, вырабатывающие гормоны, и органы-мишени расположены на некотором расстоянии друг от друга.
Иногда это расстояние минимально: инкреторный аппарат желудочнокишечного тракта находится в стенке кишечника; клетки, вырабатывающие
половые гормоны, находятся в половых железах. Иногда расстояние значительно: гипофиз и зависящая от него кора надпочечников.
Принципы строения желез внутренней секреции. Это компактные органы, состоящие из соединительнотканной стромы и железистой паренхимы.
Внутриорганные прослойки соединительной ткани, как правило, тонкие и
нежные, сопровождают сосуды. Паренхима образована эпителиальной или
нервной тканью. Клетки паренхимы формируют тяжи, фолликулы или скопления, тесно прилежащие к многочисленным капиллярам. Выводных протоков у эндокринных желез нет, так как секреты — гормоны — выделяются
непосредственно в кровь. В связи с этим кровоснабжение желез внутренней
секреции чрезвычайно обильно. Масса крови, протекающей по сосудам железы, может в несколько раз превосходить массу железистой паренхимы.
Классификация желез внутренней секреции производится по нескольким
признакам. По происхождению из определенной ткани железы делят на: эпителиальные (щитовидная и паращитовидная железы, передняя и средняя доли
гипофиза, кора надпочечников, островки поджелудочной железы, тимус),
нервные (мозговое вещество надпочечников, параганглии) и нейроглиальные
(задняя доля гипофиза, эпифиз). В функциональном отношении в эндокринной системе различают центральные и периферические звенья. Центральной
железой внутренней секреции является гипофиз. Он, в свою очередь, находится под регулирующим влиянием таких структур мозга, как эпифиз и гипоталамус.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Гипоталамус — участок промежуточного мозга. В нем различают несколько десятков ядер (скоплений нервных клеток), нейроны которых вырабатывают рилизинг-гормоны. Поступая в кровь, они достигают гипофиза, где
стимулируют (либерины) или тормозят (статины) деятельность определенных клеток передней доли гипофиза. Нейроны некоторых ядер гипоталамуса
вырабатывают нейросекрет, стекающий по их аксонам в заднюю часть гипофиза. Столь тесная связь гипофиза и гипоталамуса позволяет говорить о нейроэндокринном звене эндокринной системы.
В периферических звеньях различают железы, зависимые от передней
доли гипофиза и независимые от нее. От передней доли гипофиза зависят:
щитовидная железа, кора надпочечников, гонады (семенники и яичники). Не
зависят: мозговое вещество надпочечников, паращитовидные железы, эндокринные клетки неэндокринных органов. В пределах эндокринной системы
центральные и периферические звенья связаны, как правило, отрицательной
обратной связью. Например, тиреотропный гормон гипофиза стимулирует
активность щитовидной железы. Увеличивается синтез и секреция тиреоидных гормонов. Высокая же концентрация тиреоидных гормонов в крови тормозит секрецию гипофизом тиреотропного гормона.
Гипофиз — hypophysis (рис. 127)—непарный орган серовато-красного
цвета, плотной консистенции, яйцевидной формы.
Рис. 127. Схема строения гипофиза крупного рогатого скота (А) и участок передней доли (Б):
1 — воронка; 2 — туберальная, 3— передняя и 4 — промежуточная доли
аденогипофиза; 5 — стебель воронки и 6 — задняя доля нейрогипофиза; 7—
щель между долями; 8 — твердая мозговая оболочка; 9 — хромофобные, 10
— ацидофильные и 11—базофильные клетки; 12 — синусоидный капилляр.
Расположен в ямке гипофиза (турецкого седла) на теле клиновидной
кости под базальной поверхностью промежуточного мозга, прикрепляясь к
его серому бугру туберальной частью с воронкой гипофиза. Гипофиз покрыт
соединительнотканной капсулой, которая в области ямки гипофиза срастается с твердой мозговой оболочкой. Над гипофизом твердая мозговая оболочка
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
образует утолщение — диафрагму, отделяющую гипофиз от мозга. Связь
между гипофизом и мозгом сохраняется через отверстие диафрагмы, диаметр
которого у крупного рогатого скота около 3 мм. Через это отверстие и проходит туберальная часть гипофиза. У крупного рогатого скота гипофиз в длину
равен 2—2,5 см, в ширину и высоту— 1,5—2, его масса — 3—5 г. У мелкого
рогатого скота и у свиней диаметр гипофиза не превышает 1 см, а масса— 0,5
г. У лошади размеры гипофиза в среднем 2,5x0,7 см, масса — около 3 г.
Развивается гипофиз из двух зачатков: эпителиального и нейроглиального. Эпителиальный зачаток — это карманообразный вырост дорсальной стенки первичной ротовой полости — гипофизарный карман (Ратке).
Отшнуровываясь от стенки первичной ротовой полости на ранней стадии
развития зародыша, он растет навстречу дна воронки — выступу вентральной стенки промежуточного мозга. Срастаясь, обе части формируют гипофиз, в котором эпителиальный зачаток развивается в железистую часть, или
аденогипофиз, а нейроглиальный — в нервную часть, или нейрогипофиз.
Аденогипофиз состоит из передней, туберальной и промежуточной долей. Наибольшее развитие получает передняя доля гипофиза. Она составляет
основную массу аденогипофиза и производит наибольшее количество гормонов.
Передняя доля гипофиза (Б) имеет структуру компактного органа с
очень нежным соединительнотканным остовом и паренхимой из тяжей эпителиальных железистых клеток. Тяжи клеток расположены плотно друг к
другу, без определенного порядка, разделяясь лишь многочисленными синусоидными капиллярами.
Клетки, входящие в состав тяжей, неоднородны. По способности к окраске на гистосрезах их разделяют на хромофобные — слабо красящиеся и
хромофильные — хорошо красящиеся. Хромофобные клетки составляют
40—50% всех клеток передней доли и распространены во всех ее участках
группами или поодиночке. Это сборная группа клеток. Среди хромофобных
клеток есть мелкие и более крупные, отростчатые и без отростков, с крупными округлыми ядрами. В состав хромофобных клеток входят молодые недифференцированные клетки, а также зрелые функционирующие клетки в
период дегрануляции — выведения гормона.
Хромофильные клетки делят на ацидофильные — воспринимающие
кислые красители и базофильные — воспринимающие основные красители.
В каждом типе хромофильных клеток по гистофизиологическим признакам, в
свою очередь, различают несколько видов клеток. Клетки аденогипофиза
секретируют гормон общего характера — гормон роста и кринотропные гормоны— стимулирующие деятельность определенных желез внутренней секреции. В науке уже долгое время ведутся исследования по идентификации
клеток гипофиза и вырабатываемых ими гормонов. В настоящее время считается, что каждый тип клеток производит какой-либо один гормон.
Ацидофильные клетки, на долю которых приходится 35—45% клеток
передней доли гипофиза, вырабатывают полипептидные гормоны. Это
овальные или округлые клетки средних размеров с центрально расположенwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ным округлым ядром. В клетках сильно развита гранулярная цитоплазматическая сеть, небольшое количество довольно крупных митохондрий и умеренных размеров комплекс Гольджи. Среди ацидофильных клеток различают
соматотропоциты и лактотропоциты. Соматотропоциты вырабатывают соматотропный или ростовой гормон (СТГ). У молодого животного он способствует росту, стимулируя белковый синтез. В цитоплазме соматотропоцитов
можно видеть большое количество крупных гранул красно-оранжевого цвета.
Лактотропоциты (маммотропные клетки) вырабатывают лактотропный гормон пролактин (ЛТГ). Он стимулирует процессы образования молока, развития желтого тела (у овец), поддерживает инстинкты, связанные с заботой о
потомстве. В цитоплазме лактотропоцитов в стадии накопления видно большое количество очень крупных (в 2 раза крупнее, чем в соматотропоцитах)
ярко-оранжевых гранул. Оба вида ацидофильных клеток распространены в
латеральных участках передней доли группами и поодиночке. У рогатого
скота по морфологическим признакам трудноразличимы.
Базофильные клетки составляют 7—8%, и лишь у свиньи их количество
достигает 27% от общего числа клеток передней доли гипофиза. Они вырабатывают гормоны гликопротеидной природы и делятся на тиреотропоциты и
гонадотропоциты. Тиреотропоциты (тиротропные клетки) продуцируют тиреотропный гормон (ТТГ), стимулирующий функцию щитовидной железы.
Это крупные клетки неправильно-овальной или угловатой формы с округлым
ядром. Цитоплазма густо заполнена пылевидной фуксинофильной зернистостью. При гипофункции щитовидной железы размеры клеток увеличиваются,
цитоплазма приобретает пенистый вид. У рогатого скота распространены в
центральных участках передней доли вблизи капилляров. Гонадотропоциты
вырабатывают гонадотропные гормоны, стимулирующие деятельность половых желез. Это овальные, грушевидные или отростчатые клетки с ядром,
сдвинутым к одной стороне, сильноразвитым комплексом Гольджи, с большим количеством мелких митохондрий и мелкой зернистостью в цитоплазме.
Имеется две разновидности базофильных гонадотропоцитов, различимые с
помощью электронного микроскопа по характеру зернистости. Фолликулостимулирующие гонадотропоциты продуцируют фоллитропин (фолликулостимулирующий гормон, ФСГ), стимулирующий у самок рост и созревание
фолликулов, у самцов — сперматогенез. Лютеинизирующие гонадотропоциты продуцируют лютропин (лютеинизирующий гормон, ЛГ), регулирующий процесс овуляции и развития желтого тела в яичниках у самок и синтез
мужского полового гормона тестостерона у самцов. Гонадотропоциты встречаются во всех участках передней доли гипофиза. При кастрации клетки резко увеличиваются в размерах, в их цитоплазме появляются крупные вакуоли.
Кортикотропоциты продуцируют адренокортикотропный гормон (кортикотропин, АК.ТГ), стимулирующий развитие и функции коры надпочечников. Клетки неправильно-овальной или отростчатой формы, с округлым
ядром и специфической зернистостью, по которой они четко отличаются на
электронограммах от других клеток передней доли, однако до сих пор не
найдено критерия, позволяющего четко идентифицировать эти клетки на свеwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
товом уровне. В результате разные исследователи относят их к хромофобным, базофильным и даже ацидофильным клеткам.
Промежуточная доля гипофиза имеет вид узкой полосы, сросшейся с
нейрогипофизом. Как и передняя, она также образована эпителием. Встречаются здесь и фолликулоподобные фигуры с полостью, заполненной коллоидом. Среди клеток промежуточной доли есть несколько разновидностей. Они
называются меланотропными клетками, так как продуцируют меланоцитостимулирующий гормон (МСГ) или интермедии. Он регулирует
пигментный обмен, функции пигментных клеток и, возможно, участвует в
формировании реакции напряжения организма. Сосудистое русло промежуточной доли не столь обширно, как передней доли, капилляры щелевидны.
Между передней и промежуточной долями у многих животных имеется щель
— остаток гипофизарного кармана, заполненная желеобразной массой. У
лошади щели нет. Все доли гипофиза срастаются между собой.
Нейрогипофиз состоит из серого бугра, стебля воронки и задней доли.
Задняя доля гипофиза образована нейроглией и пучками нервных волокон.
Клетки нейроглии — питуициты (питуитоциты) имеют разную форму и размеры. Выполняют фагоцитарную, опорную и трофическую функции, возможно, участвуют в выведении гормона из задней доли гипофиза. Нервные
волокна, расположенные в задней доле гипофиза, являются аксонами нейросекреторных нейронов крупноклеточных ядер гипоталамуса.
В перикарионах нейросекреторных нейронов вырабатывается нейросекрет, который стекает по аксонам в заднюю долю и там накапливается в конечных разветвлениях нервных волокон — накопительных тельцах (Херринга). Отсюда ясно, что задняя доля является лишь депо, а не местом синтеза
гормонов. По мере необходимости гормоны, входящие в состав нейросекрета, поступают в близлежащие кровеносные капилляры. Нейросекрет содержит гормоны окситоцин и вазопрессин. Окситоцин стимулирует сокращение
мышечной оболочки матки (миометрия) и миоэпителия молочной железы,
увеличивая молокоотдачу. Вазопрессин (антидиуретический гормон, АДГ)
повышает тонус сосудов и тем самым кровяное давление и уменьшает мочеотделение.
Связь гипофиза с гипоталамусом осуществляется как нервнопроводниковым, так и гуморальным путем. Нейроны мелкоклеточных ядер
гипоталамуса продуцируют нейропептиды, регулирующие деятельность гипофиза. В настоящее время известно 7 нейропептидов — либеринов, стимулирующих деятельность всех клеток передней и промежуточной долей гипофиза, и 3 нейропептида — статина, угнетающих деятельность соматотропных, лак-тотропных и меланотропных клеток гипофиза. Нейропептиды поступают в сосуды гипоталамической области, а оттуда попадают в капилляры
аденогипофиза, где и осуществляют свое регулирующее действие. В заднюю
долю гипофиза, как сказано выше, нейрогормоны поступают по аксонам нейросекреторных нейронов. Аналогичный путь обнаружен и для нейронов, регулирующих работу промежуточной доли, по аксонам которых рилизинггормоны достигают меланотропных клеток.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Эпифиз — epiphysis — входит в состав промежуточного мозга, является
выростом крыши третьего мозгового желудочка (рис. 128). Имеет вид удлиненного бугристого тела, за что был назван шишковидной железой. Из сельскохозяйственных животных бугристость заметна только у свиньи, у остальных эпифиз гладкий. Его верхушка заходит между оральными (передними)
буграми четверохолмия. Длина эпифиза варьирует от 6 до 50 мм, масса равна
у быков 120 мг, коров 280, у свиней 100—200, у лошадей 400—1300 мг.
Гистологическое строение. Эпифиз имеет структуру компактного органа. Сверху одет капсулой из соединительной ткани, переходящей на него с
мягкой мозговой оболочки. От капсулы внутрь отходят тонкие прослойки —
септы, создающие впечатление дольчатости органа. Септы заметны у лошади. У других животных внутриорганная соединительная ткань развита слабо.
Паренхима железы образована нейроглией и содержит различные формы
глиальных клеток, выполняющих функции, типичные для нейроглии: опорную, трофическую, разграничительную. Специфическими клеточными элементами эпифиза являются пинеалоциты— отростчатые клетки, содержащие
базофильные или ацидофильные гранулы. Они, по-видимому, происходят из
эпендимоцитов. Ядра пинеалоцитов крупные, округлые, иногда с неровной
поверхностью. В цитоплазме хорошо развита гладкая цитоплазматическая
сеть, много крупных митохондрий. На концах отростков имеются булавовидные расширения, которыми пинеалоциты контактируют с капиллярами и
клетками глии.
Рис. 128. Эпифиз крупного рогатого скота (А) и
участок паренхимы эпифиза (Б):
1 — верхушка эпифиза; 2—
капсула; 3 —третий мозговой желудочек; 4— прослойка
соединительной
ткани; 5 — глиальные
клетки; 6 — пинеалоцит.
В эпифизе образуется большое количество биологически активных веществ, в том числе фактор, тормозящий выработку рилизинг-гормонов гипоталамусом, чем регулируется деятельность гипофиза и периферических эндокринных желез. В нем синтезируются такие гормоны, как серотонин, мелатонин, адреногломерулотропин. Эти гормоны изменяют активность гипоталамо-гипофизарно-надпочечниковой системы, участвуют в регуляции кровяного давления, водно-солевого обмена, температуры тела, проницаемости соwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
судов. Мелатонин, кроме того, предотвращает преждевременное развитие
половой системы.
Щитовидная железа — gl. thyreoidea (рис. 129)—закладывается как непарное выпячивание энтодермального эпителия вентральной стенки глотки в
области между I и II парами жаберных
карманов. Затем эпителиальный тяж раздваивается и его боковые части усиленно
растут, что определяет форму щитовидной железы. У большинства млекопитающих она состоит из двух долей, соединенных перешейком. У лошади и овцы
перешеек не развит. У свиньи средняя
часть щитовидной железы — тело хорошо развито, а боковые доли имеют вид
небольших выростов. Цвет железы красновато-коричневый, консистенция плотная, поверхность бугристая. Расположена
железа на боковых поверхностях первых
2—4 колец трахеи, краниально касаясь
кольцевидного хряща гортани. Масса щитовидной железы и ее гистоструктура
сильно варьируют у животных одного вида и пола в зависимости от внешних усРис. 129. Щитовидная железа
ловий (температуры, состава рациона,
крупного рогатого скота (А)
длины светового дня) и состояния животи ее гистологическое строеного (беременность, лактация, стресс и
ние (Б):
др.).
1 — доля; 2 — перешеек; 3 —
Гистологическое строение. Щитофолликул; 4 — интерфолликувидная железа — компактный орган. Ее
лярный острозок; 5 — капилкапсула образована плотной соединиляр; 6 — тиреоциты; 7 — колтельной тканью, от которой внутрь пролоид; S—резорбционные вакуоходят соединительнотканные прослойки,
ли.
делящие железу на дольки. Из сельскохозяйственных животных дольчатость лучше всего выражена у крупного рогатого скота. Паренхима образована эпителиальной тканью, организованной в
виде фолликулов и интерфолликулярных островков. Внутри долек тонкие
прослойки рыхлой соединительной ткани пронизаны многочисленными сосудами и нервами. Капилляры оплетают каждый фолликул. Через щитовидную железу в 1 мин протекает крови в 5,5 раза больше ее объема.
Фолликул представляет собой пузырек диаметром от 0,02 до 0,7 мм.
Стенка фолликулов в нормально функционирующей железе образована однослойным кубическим эпителием. Полость его заполнена желеобразной
массой — коллоидом. От окружающих тканей эпителий отделен базальной
мембраной. Клетки фолликулярного эпителия — тиреоциты имеют светлое
округлое ядро, слегка сдвинутое к базальному полюсу. На апикальном полюwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
се, обращенном в полость фолликула, имеются многочисленные короткие
микроворсинки. В клетке хорошо развита гранулярная цитоплазматическая
сеть, комплекс Гольджи, много митохондрий. В фолликулах вырабатываются
тиротоксин и трийодтиронин — гормоны, стимулирующие процессы окисления в организме. Они влияют на белковый, углеводный, жировой и водно-солевой обмен, на рост и дифференцировку тканей, в том числе мозга. Основными веществами, из которых образуются гормоны, являются аминокислота
тирозин и йод. Оба эти вещества активно захватывают тиреоциты из притекающей крови. На их основе синтезируется тиреоглобулин, накапливающийся и дозревающий в составе коллоида. По мере надобности тиреоглобулин с
помощью эндоцитоза вновь поступает в тиреоциты, где от него отщепляются
гормоны и выводятся в кровь через базальный полюс клетки. Активность эндоцитоза можно косвенно определять по разжижению коллоида, наличию резорбционных вакуолей — светлых пузырьков, появляющихся по краю, в коллоиде.
Морфология щитовидной железы резко меняется в зависимости от ее
функционального состояния. При гиперфункции (повышенной активности)
фолликулы уменьшаются в размерах, эпителий увеличивается в высоту —
становится цилиндрическим, ядра тиреоцитов укрупняются, коллоид приобретает пенистый вид из-за большого количества резорбционных вакуолей.
При гипофункции (пониженная активность) фолликулы увеличиваются в размерах, растягиваются, коллоид уплотняется.
Эпителиальная ткань, расположенная между фолликулами, называется
интерфолликулярными островками. Они состоят из молодых малодифференцированных клеток, способных при необходимости формировать новые
фолликулы. Между фолликулами, а иногда и в их стенке (не достигая просвета фолликула) находятся одиночные или небольшими группами парафолликулярные или К-клетки. Они вырабатывают кальцитонин — гормон, стимулирующий поступление Са из крови в кость. Это овальные светлые клетки
с округлым, центрально лежащим ядром, не способны захватывать йод. Кклетки — элементы ультимобранхиальных телец, которые у млекопитающих
интегрируют со щитовидной железой на ранних стадиях эмбрионального
развития и не существуют как самостоятельные органы. Их же элементы,
рассеявшись по щитовидной железе, выполняют свою функцию. Васкуляризуется щитовидная железа несколькими щитовидными артериями, отходящими от сонной артерии. Иннервируется волокнами вегетативной нервной
системы.
Околощитовидные (паращитовидные) железы — gll. parathyreoideae
— представляют собой обычно 2 пары мелких
(диаметр от 0,4 см у свиньи до 1,3 см у лошади), удлиненно-овальных
органов красноватого цвета, мягкой консистенции. Краниальная железа лежит впереди щитовидной железы, каудальная погружена в щитовидную железу, одета общей с ней капсулой. У лошади обе пары околощитовидных желез самостоятельны. Иннервация и васкуляризация их такая же, как и у щитовидной железы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Гистологическое строение. Снаружи околощитовидная железа одета соединительнотканной капсулой, внутрь от которой идут тонкие прослойки.
Паренхима железы образована переплетающимися тяжами эпителиальных
клеток — паратироцитов. Между тяжами проходят тонкие соединительнотканные прослойки с многочисленными сосудами. Среди паратироцитов различают главные, промежуточные и ацидофильные, которые являются одним
видом клеток, находящихся в разном функциональном состоянии. Главные
клетки с базофильной цитоплазмой, светлым ядром и небольшим количеством митохондрий. С переходом в ацидофильное состояние цитоплазмы, количество митохондрий сильно увеличивается, ядра уплотняются. Паратироциты секретируют паратгормон (паратирин). Он увеличивает содержание Са
в крови путем активации остеокластов, снижения реабсорбции Са в почках,
усиления всасывания Са в кишечнике. Паратгормон является антагонистом
кальцитонива. Благодаря их сочетанному действию псчдерживается постоянный уровень Са в крови.
Надпочечники — gll. suprarenales (рис. 130)—парные органы, расположенные впереди почек в забрюшинном пространстве, покрыты общей с почками жировой капсулой, красно-бурого цвета, удлиненно-овальной или треугольной формы. У крупного рогатого скота, лошади, свиньи длина надпочечника колеблется в пределах 4—9 см, у овцы— 1,5—2 см. Масса надпочечников у крупного рогатого скота 26—36 г, у лошади — 20—22, у свиньи
— 5—13, у овцы — 3,6—4 г. С одной стороны, надпочечник имеет вдавление
— ворота надпочечника. В этом месте входит артерия и выходит вена. В надпочечнике объединены, по сути, две железы, различающиеся по происхождению, структуре и функции и образующие в органе корковое и мозговое вещество. Корковое вещество, или интерреналовая железа, развивается из мезодермального эпителия, общего с зачатком промежуточной почки и гонады.
Это определяет функциональную направленность коркового вещества надпочечников: в нем синтезируются гормоны, регулирующие водно-солевой обмен и половые гормоны. Мозговое вещество, или супрареналовая железа,
развивается из участка нервной ткани, общего с симпатическими ганглиями.
У рыб интерреналовая и супрареналовая железы существуют самостоятельно. У остальных классов хордовых они сливаются в единую железу — надпочечник.
Гистологическое строение. Надпочечник — компактный орган. Его капсула образована плотной соединительной тканью, от которой внутрь проходят тонкие прослойки рыхлой соединительной ткани, проходящие по ходу
сосудов между тяжами эпителиальных клеток. В 1 мин через надпочечник
проходит количество крови, в 5 раз превышающее его объем. Под капсулой
часто встречаются скопления нервных клеток — микроганглии.
Корковое вещество. Паренхима образована эпителиальными клетками
— адренокортикоцитами и подразделяется на три зоны: клубочковую, пучковую и сетчатую. Корковое вещество составляет 75—90% органа. На долю
клубочковой зоны приходится 10—15%. пучковой — 80, сетчатой — 5—
10%.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 130. Надпочечники крупного рогатого скота (А) и их гистологическое строение (Б):
1 — капсула; 2 — клубочковая, 3— пучковая и 4 — сетчатая зоны коркового
вещества; 5 — мозговое вещество; 6 — капилляры; а —правый и б — левый
надпочечники; в — почка; г — каудальная полая вена; д — аорта.
Клубочковая зона образована сравнительно мелкими адренокортикоцитами цилиндрической формы со светлой цитоплазмой и плотными ядрами.
Они формируют тяжи, свернутые в виде арок или клубочков. В цитоплазме
клеток клубочковой зоны у некоторых животных встречаются липидные
включения. У свиньи и лошади их нет. Клетки этой зоны вырабатывают минералокортикоиды — альдостерон, дозоксикортикостерон — регулирующие
водно-солевой обмен. Возможно, клетки клубочковой зоны пограничные с
пучковой зоной являются камбиальными для коркового вещества.
Пучковая зона самая обширная. Образована тяжами или пучками, состоящими из адренокортикоцитов, цилиндрической или полигональной формы, более крупных, чем в клубочковой зоне. Среди клеток пучковой зоны
встречаются темные и светлые клетки. Светлые клетки имеют более крупные
ядра и большое число липидных включений, из-за чего цитоплазма приобретает пенистый вид. Темные клетки имеют более плотные ядра, базофильную цитоплазму. В них лучше развита гладкая цитоплазматическая сеть,
больше митохондрий и аскорбиновой кислоты, практически отсутствуют липидные капли. Считается, что светлые клетки находятся в покое или стадии
накопления предшественников гормонов, а темные — более активные — в
стадии синтеза и секреции гормонов. Соотношение темных и светлых клеток
меняется в зависимости от функционального состояния органа. Под действием АКТГ все клетки теряют липидные включения, становятся темными. В
пучковой зоне синтезируются глюкокортикоиды—кортизол (гидрокортизон),
кортизон, кортикостерон и др. — гормоны широкого спектра действия. Они
влияют на белковый, углеводный, жировой обмены, стимулируют глюкоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
неогенез, липолиз, обладают противовоспалительным действием, уменьшают
боль, отек, повышают устойчивость организма в период стресса, подавляют
аллергические реакции. В больших дозах приводят к инволюции лимфоидной ткани в тимусе, селезенке, лимфоузлах.
Сетчатая зона характеризуется тем, что тяжи клеток переплетаются, между ними более заметны прослойки соединительной ткани и синусоидные
капилляры. Адренокортикоциты меньше по размерам и разнообразнее по
форме по сравнению с пучковой зоной. Преобладают темные клетки. Встречаются клетки, содержащие пигментные включения бурого цвета (липохромы). В этой зоне образуются половые гормоны, преимущественно андрогены.
Эстрогены здесь синтезируются более слабые и в меньших количествах. Сетчатая зона граничит с мозговым веществом. Граница эта в большинстве случаев неровная: оба вещества взаимно проникают друг в друга. В некоторых
участках видна между ними соединительнотканная прослойка.
Пучковая и сетчатая зоны находятся под регулирующим влиянием кортикотропных клеток гипофиза. АКТГ стимулирует функцию. Гормоны коры
надпочечников, наоборот, снижают активность кортикотропных клеток.
Мозговое вещество. Паренхима образована клетками, произошедшими
из нейробластов. Это крупные округлые, овальные или полигональные клетки с округлым крупным, эксцентрично лежащим ядром и светлой цитоплазмой. При воздействии солями тяжелых металлов в цитоплазме появляется
бурый осадок, за что клетки получили название хромаффинные. Какой-либо
упорядоченности в расположении хромаффинных клеток не обнаруживается,
однако каждая клетка граничит с капилляром. В цитоплазме клеток в электронный микроскоп видна специфическая зернистость. По характеру гранул,
гистохимическим свойствам и вырабатываемым ими гормонам хромаффинные клетки делят на 2 вида: адреналиновые, вырабатывающие адреналин, и
норадреналиновые, вырабатывающие норадреналин. Эти гормоны относятся
к катехоламинам. Они действуют на организм подобно медиаторам симпатической нервной системы, являются гормонами тревоги — стимулируют организм для борьбы и для этого возбуждают ЦНС, расслабляют мышечную оболочку бронхов, повышают работоспособность мышц, сердца, повышают давление крови, мобилизуют энергетические ресурсы, повышают агрессивность.
Регуляция деятельности мозгового вещества осуществляется симпатической
нервной системой.
В разных местах организма могут встречаться небольшие тельца, состоящие из скоплений клеток либо коркового вещества— интерреналовые
тельца, либо мозгового вещества — параганглии. Они дополняют работу
надпочечников, но заменить их не в состоянии.
Изменение эндокринных желез с возрастом и под влиянием различных факторов. С возрастом происходит некоторое увеличение массы эндокринных желез. В гипофизе молодняка крупного рогатого скота в период интенсивного роста зона расположения ацидофильных клеток (периферия аденогипофиза) растет быстрее, чем зона расположения базофильных клеток
(центральные участки аденогипофиза). В щитовидной железе увеличиваются
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
размеры фолликулов с одновременным увеличением высоты эпителия и резорбцией коллоида. Старческие изменения желез внутренней секреции характеризуются разрастанием соединительнотканных прослоек. В гипофизе,
кроме того, значительно уменьшается количество ацидофилов и менее значительно — базофилов аденогипофиза. В щитовидной железе снижается васкуляризация и индекс активности. В надпочечниках резко разрастается пучковая зона и уменьшается клубочковая и сетчатая.
Внешние воздействия. Повышение температуры окружающей среды
приводит к снижению высоты эпителия и увеличению диаметра фолликулов
щитовидной железы — ее активность снижается. Понижение температуры
действует противоположным образом. В надпочечниках низкая температура
приводит к кровоизлияниям и жировому перерождению клеток коры у овец и
поросят. Снижение двигательной активности уменьшает активность щитовидной железы. В надпочечниках при продолжительном обездвиживании
животного можно видеть признаки усиления активности: увеличение размеров ядер и клеток, количества лизосом в них, уменьшение числа липосом.
При неподвижности в течение 24 и более часов — явления деструкции с увеличением в 30 раз числа аутофагосом, уменьшением крист в митохондриях,
кровоизлияниями (В. М. Гордиенко и др.) - Большое влияние оказывает
кормление. Недокорм и белковая недостаточность приводят к гипофункции
эндокринных желез. При обильном кормлении увеличивается высота эпителия фолликулов щитовидной железы, в надпочечниках возрастает содержание липидов в клетках коры, в гипофизе увеличивается число соматотропоцитов. Биологически активные вещества увеличивают массу эндокринных
желез, приводят к гипертрофии их аденоцитов.
Вопросы для самоконтроля. 1. Дайте характеристику эндокринной системы как одной из регуляторных систем организма. 2. Каково строение и
положение гипофиза, эпифиза, щитовидной, околощитовидной желез, надпочечников? 3. Какие еще органы выполняют эндокринные функции?
Глава 20. СЕРДЕЧНО СОСУДИСТАЯ СИСТЕМА И ОРГАНЫ
КРОВЕТВОРЕНИЯ (АППАРАТ КРОВО- И ЛИМФООБРАЩЕНИЯ)
Сердечно-сосудистая система включает в себя кровеносную и лимфатическую системы, связанные между собой генетически, морфологически и
функционально. Источником их является мезенхима. Обе они состоят из системы сосудов, которые объединяются при впадении лимфатических стволов
в краниальную полую вену. Жидкое содержимое сосудов — кровь и лимфа
— главная функциональная часть сердечно-сосудистой системы и в своей
основной функции (участие в обмене веществ) тесно связаны с тканевой
жидкостью. Выходя из капилляров, составные части крови и лимфы входят в
состав тканевой жидкости, и, наоборот, тканевая жидкость, проникая в капилляры, становится составной частью крови и лимфы. Тканевая жидкость,
кровь и лимфа считаются внутренней средой организма, они составляют до
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
50% массы тела. На долю крови приходится 7—8% массы тела. Жидкие ткани чутко реагируют на воздействия внешней среды на организм и на изменение гомеостаза. На этом основано широкое использование крови в клинических и научных исследованиях (см. раздел третий).
Клеточные элементы крови и лимфы недолговечны и постоянно пополняются за счет деятельности органов кроветворения: красного костного мозга, селезенки, лимфатических узлов, а у плодов и молодняка, кроме того, печени и тимуса.
СИСТЕМА ОРГАНОВ КРОВООБРАЩЕНИЯ
Кровеносная система состоит из сердца и кровеносных сосудов. Сосуды,
по которым кровь движется от сердца, называются артериями, к сердцу —
венами. Конечные разветвления артерий — артериолы распадаются на прекапилляры; прекапилляры — на капилляры, которые затем собираются в посткапилляры; последние— в венулы и наконец — в вены.
Кровеносной системе присущи все функции, выполняемые кровью. Основные из них: транспортная, участие в обмене веществ, терморегуляции,
гуморальной регуляции (благодаря транспорту гормонов и других биологически активных веществ), иммунологической защиты.
Краткие сведения о развитии системы органов кровообращения
Филогенез. У простейших многоклеточных животных обмен веществ
вполне обеспечивается диффузионными токами в неоформленной диффузной
внутренней среде. С усложнением организации и повышением жизнедеятельности фомируются специальные каналы с собственной стенкой, пронизывающие паренхиму тела, по которым в определенном направлении движется
жидкое содержимое — гемолимфа. Движение это осуществляется благодаря
движениям всего тела. При недостаточности такого перемещения в сосудах
начинают развиваться пульсирующие участки с мышечной оболочкой —
сердца. Примитивная сосудистая система может быть незамкнутой, если она
прерывается лакунами — полостями в теле, не имеющими собственной оболочки, или замкнутой, когда концы сосудов соединяются друг с другом в
единую систему, как, например, у кольчатых червей.
Простая замкнутая кровеносная система, характерная для аннелид
(кольчатых червей), состоит из двух продольных сосудов — дорсального .и
вентрального, соединенных друг с другом на концах тела и в каждом сегменте. Метамерные сегментарные сосуды разделяются на висцеральные,
идущие вокруг кишечной трубки, и париетальные, проходящие в стенках тела (рис. 131—А).
С развитием жаберного дыхания в головном конце тела происходит усложнение сосудистой системы (Б). В области жабр сегментарные сосуды делятся на приносящие и выносящие, и между ними у низших хордовых (ланцетник) появляются капилляры, обеспечивающие газообмен. В сосудистой
системе появляются участки с артериальной кровью. В пульсирующем вентральном сосуде к голове течет венозная кровь. С развитием жабр его пульwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
сации становится недостаточно и в основании приносящих жаберных сосудов развиваются пульсирующие мышечные участки — жаберные сердца. В
результате в выносящих жаберных артериях течет артериальная кровь, которая собирается в парные сонные артерии, несущие кровь в головной конец
тела и в непарную дорсальную аорту, лежащую под хордой. Из нее кровь попадает в сегментарные париетальные и висцеральные сосуды, а оттуда — в
парные кардинальные каудальные вены. С головного конца кровь собирается
в парные кардинальные краниальные (яремные) вены. Все кардинальные вены объединяются в проток Кювье (общую кардинальную вену), из которого
кровь стекает в венозный синус.
Рис. 131. Схема строения кровеносной системы у червя (А), первичноводного (Б), у наземного животного (В) и изменение жаберных артерий у
рыбы (Г), двоякодышащих (Д), амфибий (Е), рептилий (Ж), млекопитающих (З):
1 — дорсальная аорта; 2— вентральная аорта; 3— сегментарный сосуд; 4— сонная артерия; 5 — приносящие и выносящие жаберные сосуды; 6
— левая кардинальная краниальная вена; 7 — париетальные и висцеральные
сегментарные сосуды; 8 — левая кардинальная каудальная вена; 9 — проток
Кювье; 10— артериальный конус; 11 — желудочек; 12 — предсердие; 13 —
венозный синус; 14 — печеночная вена; 15 ~ капилляры печени; 16 — подкишечная вена; П — дуга аорты; 18 — легочная артерия; 19 — капилляры легких; 20 — легочная вена; 21 — аорта; 22 — каудальная и 23 — краниальная
полые вены; 24 — воротная вена; 25 — правый и 26—левый желудочки; 27 —
правое и 28 — левое предсердие; I, II, III, IV, V, VI — жаберные артерии.
Черным цветом обозначена венозная кровь, серым — смешанная, белым —
артериальная.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
С усложнением организации у первичноводных животных (рыб) уменьшается число жабр и жаберных артерий до 6 пар, увеличивается протяженность и ветвление капилляров, уменьшается количество висцеральных сосудов. Жаберные сердца сменяются одним более крупным сердцем, расположенным в основании головы позади жаберного аппарата. Сердце двухкамерное, состоит из предсердия и желудочка. К предсердию примыкает венозный синус, в который вливается кровь из протока Кювье, а также сильно
развитых печеночных вен. Желудочек сообщается с артериальным конусом,
который переходит в брюшную аорту.
С развитием легких происходит дальнейшая перестройка и усложнение
сосудистой системы, что намечается уже у двоякодышащих рыб. У них VI
пара жаберных артерий отходит к зачаткам легких, развивается новый крупный сосуд — каудальная полая вена, собирающая кровь с задней половины
тела в венозный синус. Кардинальные каудальные вены начинают терять
свое значение. С выходом на сушу и переходом к легочному типу дыхания
редуцируются I, II и V пары жаберных артерий. Из III пары развиваются сонные артерии, из VI — легочные артерии, из IV — дуги аорты, У рептилий
функционируют обе дуги аорты, у птиц сохраняется лишь правая, у млекопитающих — левая дуга аорты.
Перестройка сердца идет путем разделения его на правую и левую половины. У амфибий и рептилий это деление — неполное, сердце трехкамерное: имеет 2 предсердия и желудочек. У рептилий в нем возникает неполная
перегородка. У птиц и млекопитающих перегородка полностью разделяет
желудочек, образуя 4-камерное сердце, в левой половине которого циркулирует артериальная, в правой — венозная кровь. Формируются 2 круга кровообращения: малый, или легочный, и большой, или системный.
У наземных животных в связи с редукцией хвоста, сменой почек, развитием ногообразных конечностей и особенностями эмбрионального развития существенные изменения наблюдаются н в венозном русле. Кардинальные вены редуцируются, замещаются другими сосудами. Кардинальные краниальные вены становятся яремными, а их конец вместе с протоком Кю1ье
преобразуется в краниальную полую вену, куда течет кровь с головы, шеи,,
грудных конечностей и частично со стенок тела. Венозный синус входит в
состав правого предсердия. Кардинальные каудальные вены замещаются
каудальной полой веной — в нее впадают вены, несущие кровь с задней половины тела и от органов брюшной полости, в том числе крупная воротная
вена. Грудные отрезки кардинальных вен частично сохраняются в виде непарной вены, собирающей кровь с грудных стенок.
Кроме деления сосудистой системы на артериальное и венозное русло, у
позвоночных наблюдается деление ее на кровеносную и лимфатическую системы. Лимфатическая система у круглоротых и хрящевых ганоидов появляется как придаток кровеносной системы и у костистых рыб, амфибий и
рептилий имеет вид сосудистых лимфатических сетей. Развивается лимфатическая система в связи с прогрессивным развитием тканей внутренней среды, Наиболее полного развития она достигает у млекопитающих, где наряду
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
с лимфатическими сосудами, синусами, пространствами, полостями, капиллярами существует множество лимфатических узлов и лимфоидных фолликулов в стенках органов.
Параллельно с развитием и усложнением сердечно-сосудистой системы
происходило развитие крови и органов кроветворения. У ланцетника — простейшего хордового кровь бесцветна без клеточных элементов (кроме амебоидных клеток). У рыб появляются форменные элементы крови и орган
кроветворения — селезенка. Однако кроветворение происходит во многих
органах, содержащих производные мезенхимы. В дальнейшем круг органов
кроветворения сужается и у млекопитающих основным таким органом оказывается красный костный мозг. Кроме него, отдельные виды клеток крови
образуются в селезенке и лимфатических узлах.
Онтогенез. Сердечно-сосудистая система развивается из мезенхимы. На
ранних стадиях развития зародыша происходит обособление некоторых клеток мезенхимы в виде кровяных островков. Их клетки дифференцируются в
клетки крови. Другие мезенхимные клетки образуют стенки вокруг кровяных
островков, превращаются в клетки эндотелия — так в массе мезенхимы закладываются сосуды. Раньше других закладываются сосуды желточного
мешка. Вскоре после их появления начинает развиваться к сосудистая система зародыша. Одновременно с закладкой кровеносных сосудов образуется
сердце. Оно закладывается в шейной области (рис. 132) в слое мезенхимы,
лежащем между энтодермой и висцеральным листком спланхнатома мезодермы в виде двух эндотелиальных трубок. С углублением туловищной
складки трубки сближаются, срастаются и впячиваются в целом, который,
окружая единую теперь сердечную закладку, превращается в серозную околосердечную полость. Стенка сердца утолщается и .дифференцируется на
три слоя: внутренний слой — эндокард образован слоем эндотелиальных
клеток мезенхимного происхождения, мощный средний слой — миокард и
наружный слой — эпикард происходят из висцерального листка спланхнотома мезодермы.
Вначале сердце в виде трубки лежит вдоль тела зародыша, является
двухкамерным и по строению аналогично сердцу рыб: впереди желудочка
расположен артериальный конус, позади предсердия — венозный "синус.
Между предсердием и желудочком образуется перетяжка с узким отверстием. Еще в зародышевый период сердце из двухкамерного становится трех, а потом четырехкамерным и опускается в грудной отдел туловища. В процессе перестройки сердце становится S-образным. При этом желудочек и артериальный конус оказываются внизу, предсердие и венозный синус — сверху. Венозный синус входит в состав предсердия, которое при изгибах сердца
охватывает с боков артериальный конус. В артериальном конусе, предсердии
и желудочке появляются перегородки. Артериальный конус при этом разделяется на аорту и легочную артерию (легочный ствол). В предсердии и желудочке перегородки растут навстречу друг другу, разделяя сердце на левую и
правую половины. Между предсердием и желудочком в каждой половине
формируется клапан, позволяющий движению крови лишь в одном направwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
лении. В течение всего внутриутробного развития между предсердиями сохраняется овальное отверстие.
Рис. 132. Развитие сердца:
А и Б —закладка сердца; В, Г,
Д — изгибы сердца, вид снаружи; 1 — нервная трубка; 2 —
хорда; 3 — аорта; 4 — париетальный и 5 — висцеральный
листки спланхнотома мезодермы; 6 — целом; 7 — мезенхима; 8 — закладки сердца; 9 —
головная кишка; 10— дорсальная брыжейка; 11 — полость перикарда; 12 — эндокард; 13 — миокард; 14 — эпикард; 15 — париетальный листок перикарда; 16 — артериальный ствол; 17 — артериальный конус; 18 — желудочек
(18а — правый, 18б — левый);
19 — предсердие (19а — правое, 19б — левое); 20 — венозный синус; 21—проток Кювье;
22 — закладка межпредсердножелудочковой перегородки; 23
— межпредсердное (овальное)
отверстие.
Параллельно закладке сердца и под его влиянием закладываются крупные сосуды, характерные для низших классов хордовых: жаберные артерии,
дорсальная и вентральная аорты, кардинальные вены. Желточные сосуды
вступают в связь с сосудистой системой зародыша, принося с собой питательные вещества из желточного мешка. У плацентарных млекопитающих
желточное кровообращение существует кратковременно в связи с малыми
запасами питательных веществ в желточном мешке и развитием аллантоиса и
плаценты. Вместе с перестройкой сердца идет активная перестройка сосудистой системы. Часть жаберных артерий (I, II и V пары) редуцируется, оставшиеся превращаются в сонные артерии (III пара), дугу аорты (IV левая),
легочные артерии (VI пара). Кардинальные вены редуцируются. Развиваются
краниальная и каудальная полые вены, собирающие кровь со всего организма
и впадающие в венозный синус правого предсердия. Развиваются артерии и
вены, соединяющие сосудистую систему зародыша с сосудами аллантоиса, а когда тот срастается с хорионом — с сосудами
плаценты.
Кровообращение у плода (рис. 133). После развития плаценты сосудистая система плода устанавливает теснейшую связь с ее сосудами. От каудального участка брюшной аорты отходят парные пупочные артерии. Они
образуют в плаценте густую капиллярную сеть, где и происходит обмен веществ между плодом и матерью. Кровь, насыщенная кислородом и питательwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ными веществами, собирается в пупочную вену и идет к телу зародыша, в составе пупочного канатика. В его состав входят также пупочные артерии и
проток аллантоиса. По пупочной вене кровь попадает в воротную систему
печени, а оттуда по печеночным венам — в каудальную полую вену, где
смешивается (первый раз) с венозной кровью плода. Лишь у немногих видов
животных (крупный рогатый скот, собака) часть крови пупочной вены минует печень и по венозному (аранциевому) протоку попадает прямо в каудальную полую вену. Смешанная кровь из каудальной полой вены попадает в
правое предсердие, где смешивается (второй раз) с венозной кровью, поступающей с переднего конца тела по краниальной полой вене. Из правого
предсердия кровь выходит двумя путями. Большая часть крови через широкое овальное отверстие проходит в левое предсердие. Меньшая часть крови
поступает по стволу легочных артерий в легкие. Однако она не вся доходит
до легких. Между стволом легочных артерий и дугой аорты у плода существует артериальный (боталлов) проток, по которому значительная часть крови оттекает в аорту, не доходя до легких.
Рис. 133. Система кровообращения у плода:
1 — плечеголовной ствол; 2— каудальная полая вена; 3— артериальный
проток; 4— левое предсердие; 5— легочная вена: 6 — капилляры в легких; 7—
аорта; а — печеночная вена; 9 — венозный проток; 10 — чревная артерия;
11— пупочная артерия; 12 — легочная артерия; 13 — краниальная полая вена; 14 — правое предсердие; 15 — овальное отверстие; 16 — правый и 17—
левый желудки; 18 — сосуды печени; 19 — воротная вена; 20—пупочная вена; 21 — плацента.
В левое предсердие, кроме крови, попавшей через овальное отверстие,
поступает кровь из малого круга кровообращения, из легочных вен. Затем
она идет в левый желудочек, оттуда — в аорту, где к ней и присоединяется
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
кровь из артериального протока (с большим содержанием венозной крови) —
это третье смешение крови у плода. У плода в полной мере функционирует
большой круг кровообращения, в состав которого включается плацентарное
кровообращение, и в очень небольшой степени — малый круг кровообращения, еще не выполняющий своей функции.
При рождении происходит перекручивание, сдавливание и разрыв пупочных сосудов. Кровь по плацентарным сосудам перестает поступать, и в
правом предсердии резко падает давление крови. Первый вдох расправляет
легкие и по легочному стволу кровь устремляется в малый круг кровообращения, минуя артериальный проток. Возвращаясь сильной струей в левое
предсердие, она приводит к повышению давления в левом предсердии. Возникает резкая разница давлений в предсердиях, что ведет к захлопыванию и
зарастанию клапана овального отверстия и полному разделению правой и левой половин сердца. Таким образом, устанавливаются два полно функционирующих круга кровообращения. Артериальный и венозный протоки запустевают, зарастают и превращаются в связки. Пупочная вена превращается в
круглую связку печени, а пупочные артерии и проток аллантоиса — в связки
мочевого пузыря.
Возрастные изменения сосудов
В период роста организма закономерно увеличиваются размеры сосудов,
утолщается их стенка. Продолжается это до завершения роста или несколько
дольше с увеличением функциональной нагрузки. У старых животных могут
наблюдаться неравномерные расширения и сужения по ходу сосудов. Отмечают утолщение и пропитывание солями и жиром интимы, снижение содержания эластических волокон и мышечных клеток в медии артерий и вен и
замена их волокнистой соединительной тканью, уменьшение количества клапанов в венах.
Строение кровеносных сосудов
Кровеносные сосуды имеют форму трубок разного диаметра « строения.
Это артерии, несущие кровь от сердца, вены, несущие кровь к сердцу, и сосуды микроциркуляторного русла, которые, кроме транспортной, выполняют
функцию обмена веществ и перераспределения крови в организме. Сосудистая система обладает большой пластичностью. Изменение скорости кровотока ведет к перестройке сосудов, образованию новых сосудов, коллатералей, анастомозов либо к запустеванию и облитерации сосудов. Артерии и
вены имеют одинаковый принцип строения. Их стенка образована тремя оболочками: внутренняя — интима, средняя — медия, наружная — адвентиция.
Однако в зависимости от расположения сосудов и особенностей их функционирования строение оболочек значительно отличается.
Артерии имеют более толстые неспадающиеся стенки и меньший просвет по сравнению с венами, что обусловлено необходимостью противостоwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ять большому давлению крови в артериях (до 200 мм рт. ст.), особенно крупных, несущих кровь непосредственно от сердца, и большей скоростью движения крови (0,5—1 м/с). Толщина стенки артерий составляет 1/3—1/4 ее диаметра. Стенки артерий обладают упругостью и прочностью. Это обеспечивается развитием в них эластической и мышечной тканей. В зависимости от
преобладания той или другой артерии делят на три типа: эластические, мышечные и смешанные (мышечно-эластические).
В артериях эластического типа интима состоит из эндотелия, подэндотелиального слоя из рыхлой соединительной ткани, отделенного от эндотелия
базальной мембраной, и слоя переплетающихся эластических волокон. Средняя оболочка состоит из большого количества слоев эластических волокон и
окончатых эластических мембран, соединенных пучками гладкомышечных
клеток. Это самая толстая оболочка артерий эластического типа. Сильно растягиваясь при поступлении порции крови из сердца, эта оболочка своей эластической тягой проталкивает кровь дальше по артериальному руслу. Наружная оболочка состоит из соединительной ткани, удерживает артерию в
определенном положении и ограничивает ее растяжение. В ней расположены
сосуды, питающие стенки артерий, и нервы. К артериям эластического типа
относятся сосуды крупного калибра: аорта, легочные артерии, плечеголовной
ствол, ствол сонных артерий. По мере удаления от сердца и ветвления артерий их диаметр уменьшается, давление в крови падает. В стенках артерий все
больше развивается мышечная ткань и становится меньше эластической ткани.
В артериях мышечного типа (рис. 134-А) границы между оболочками
хорошо видны. Интима состоит из тех же слоев, но гораздо тоньше, чем в артериях эластического типа. Слой эластических волокон внутренней оболочки
формирует внутреннюю эластическую мембрану. Средняя оболочка толстая,
содержит пучки мышечных клеток, лежащих в несколько слоев под разными
углами (30—50°). Это дает возможность при сокращении мышечных пучков
в определенных условиях либо уменьшать просвет, либо повышать тонус,
либо даже увеличивать просвет сосуда. Между мышечными пучками имеется
сеть эластических волокон. На границе с наружной оболочкой проходит наружная эластическая мембрана, хорошо выраженная в крупных артериях
мышечного типа. К артериям мышечного типа относится большинство артерий, несущих кровь к внутренним органам, и артерии конечностей.
Артерии активно участвуют в продвижении крови, недаром эластическая и мышечная ткани названы «периферическим сердцем». Двигательная
деятельность их столь велика, что без их помощи сердце не в состоянии перекачивать кровь — наступает его паралич.
Вены в сравнении с соответствующими артериями имеют больший просвет и тоньше стенку. Кровь в венах течет медленно (около 10 мм/с), под низким давлением (15—20 мм рт. ст.) с помощью присасывающего действия
сердца, сокращений диафрагмы, дыхательных движений, натяжения фасций
и сокращений мышц тела. Стенка вен состоит из тех же оболочек, но границы между ними видны плохо, мышечная и эластическая ткани в стенках
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
вен менее развиты, чем в артериях. Вены отличаются большим разнообразием в строении своих стенок, порой даже на протяжении одной вены
(например, каудальной полой). Все же можно выделить несколько типов вен,
в том числе вены мышечного и волокнистого (безмышечного) типов.
Вены мышечного типа (Б) обычно расположены в конечностях и других
местах тела, где кровь движется вверх (против силы тяжести). Внутренняя
оболочка у них тонкая. У многих вен она образует кармашковые клапаны,
препятствующие обратному току крови. Средняя оболочка образована в основном соединительной тканью с пучками коллагеновых волокон, пучками
гладкомышечных клеток, которые могут образовывать сплошной слой, и сетью эластических волокон. Внутренняя и наружная эластические мембраны
не развиты.
Наружная оболочка из соединительной ткани, широкая, содержит нервы
и сосуды сосудов.
Рис. 134. Схема строения стенки
артерии (А) и вены (Б) мышечного типа:
I — интима; 1 — эндотелий; 2 —
подэндотелиальный слой; 3 —
внутренняя эластическая мембрана; II —медия; 4 — пучки гладкомышечных клеток; 5 — эластические волокна; 6— коллагеновые волокна; III — адвентиция;
7 — наружная эластическая мембрана; 8 — соединительная
ткань; 9— сосуды сосудов.
Вены безмышечного типа имеют еще более тонкую стенку, состоящую
из эндотелия и соединительной ткани. Это вены мозговых оболочек, сетчатки
глаза, костей, селезенки.
Микроциркуляторное русло включает в себя сосуды с диаметром менее
0,1 мм: артериолы, прекапилляры, капилляры, посткапилляры, венулы. Микроциркуляторное русло превышает по емкости артериальную и венозную
систему сосудов. Конечные участки артерий, стенка которых состоит из инwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
тимы и одного слоя гладкомышечных клеток, называют артериолами. Переходя в прекапилляры и капилляры, они разветвляются, теряют мышечные и
соединительнотканные элементы. В органах на площади 1 мм2 насчитывается
от нескольких сот до нескольких тысяч капилляров.
У капилляров диаметр 4—8 мкм, длина около 200 мкм. В печени, селезенке, костном мозге капилляры имеют диаметр до 50 мкм и называются синусоидными. Стенка капилляра состоит из эндотелия, базальной мембраны и
перицитов. В безъядерных участках толщина стенки капилляра не превышает
0,1—0,5 мкм. Основное назначение капилляров — обмен веществ между содержимым капилляров и окружающими тканями. Он совершается благодаря
медленному течению крови в капиллярах, тонкой стенке, а в капиллярах некоторых органов и наличию пор (костный мозг, почки) или прерывистой базальной мембраны (печень), явлениям пиноцитоза, развитию микроворсинок.
Посткапилляры и венулы несколько шире артериол. Они объединяются в вены и выносят кровь из органа. Между артериолами и венулами имеются артериоловенулярные
(артериовенозные)
анастомозы
(шунты)
—
приспособления для перераспределения крови в организме. В покоящемся
органе они открыты и кровь устремляется в них, минуя капилляры. Так, в неработающей мышце функционируют не более 10% капилляров. В работающем органе артериоловенулярные анастомозы закрываются, и кровь растекается по капиллярам.
Закономерности хода и ветвления сосудов
Развитие организма по принципам одноосности, двусторонней симметрии и сегментального расчленения (метамерии) обусловливает ход сосудистых магистралей и их боковых ветвей. Обычно сосуды (артерия, 1—2 вены,
лимфатические сосуды) идут вместе с нервами, образуя сосудисто-нервные
пучки.
Магистральные сосуды всегда идут кратчайшим путем, чем облегчается
работа сердца и осуществляется быстрая доставка крови к органам. Эти сосуды проходят по вогнутой стороне тела или на сгибательных поверхностях
суставов, в желобках костей, углублениях между мышцами или органами с
тем, чтобы подвергаться меньшему давлению окружающих органов и растяжению при движении. Магистрали отдают боковые ветви ко всем органам,
мимо которых проходят. Величина ветвей зависит от функциональной активности органа и может меняться с изменением этой активности (например,
маточные артерии при беременности). К выступающим частям тела (пальцам,
ушам), как правило, идут две артерии, обеспечивая потребность в их повышенном обогреве.
Коллатерали, обходные сети, углы отхождения артерий.
Часть боковых ветвей, отходя от магистрали, идет параллельно с магистралью и анастомозирует с другими ее ветвями. Это коллатериальные сосуwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
ды. Они имеют большое значение для восстановления кровоснабжения при
нарушении или закупорке основного ствола. К коллатералям относят и обходные сети в области суставов. Они всегда лежат на разгибательной поверхности сустава и поддерживают нормальное кровоснабжение его тканей
во время движения, когда часть сосудов оказывается излишне сдавленной
или растянутой. Боковые ветви от магистралей отходят под разными углами.
Под острым углом идут артерии к удаленным органам. По ним обычно кровь
движется с большей скоростью. Под более прямым углом отходят сосуды к
близлежащим органам, а под тупым углом — возвратные артерии, которые
образуют коллатерали и обходные сети.
Типы ветвления сосудов и их анастомозы. Различают несколько типов ветвления сосудов (рис. 135). Магистральный тип ветвления, когда от
магистрального сосуда последовательно отходят боковые ветви, как, например, артерии, отходящие от аорты. Дихотомический тип ветвления, когда
магистральный сосуд делится на 2 равных сосуда, например, деление ствола
легочной артерии. Рассыпной тип ветвления, когда обычно короткий магистральный сосуд резко делится на несколько крупных и мелких ветвей, что
характерно для сосудов внутренних органов.
Рис. 135. Типы ветвления и анастомозов сосудов:
А — магистральный, Б — дихотомический, В — рассыпной тип ветвления;
Г а, б — анастомозы; Д—артериальные сети; 1 — артериальная дуга; 2 —
артериальная сеть; 3— чудесная сеть (в почке); 4 — капиллярная сеть; 5 —
вена; 6 — артерия; 7 — артериовенозный анастомоз; 8 — артериальный
сфинктер.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Сосуды часто соединяются друг с другом соединительными ветвями —
анастомозами, которые выравнивают кровяное давление, регулируют и перераспределяют ток крови, образуют коллатерали. Анастомозы бывают нескольких типов. Широкое соустье— анастомоз большого диаметра, соединяющий 2 крупных сосуда, например артериальный (боталлов) проток между
аортой и легочным стволом. Артериальная дуга объединяет артерии, идущие
к одному и тому же органу, например пальцевые артерии. Артериальная
сеть — сплетение концевых ветвей сосудов, например дорсальная сеть запястья. Если анастомозы объединяют ветви сосудов, идущих в разных плоскостях, образуется сосудистое сплетение, как в паутинной оболочке мозга. Чудесная сеть — разветвление по ходу сосуда с последующим объединением в
одноименный сосуд, например разветвление приносящей артериолы почечного тельца на капилляры клубочка и последующее объединение их в выносящую артериолу. Объединение концевых участков артерий и вен — артериоловенулярные (артерио-венозные) анастомозы приводят к выключению
участков капиллярной сети и быстрому сбросу крови в венозное русло.
Сердце
Сердце — cor (рис. 136)—конусовидный мышечный орган, расположенный в грудной полости в средостении на уровне 3— 6-го ребра, несколько
сдвинуто влево. Широкое основание сердца направлено вверх. С ним связаны
крупные сосуды, подходящие и отходящие от сердца. Удлиненная заостренная верхушка направлена вниз, назад и влево. Доходит у рогатого скота до 5го, у лошади — до 6-го, у свиньи — до 7-го реберного хряща, немного не доходит до грудины и диафрагмы. Передняя стенка сердца более выпуклая, чем
задняя. Масса его у взрослого крупного рогатого скота составляет 2—3 кг
(0,4—0,6% от массы тела). У лошади сердце относительно более короткое и
широкое, верхушка более округлая, масса 3—6 кг.
У свиньи сердце удлиненное, с короткой заостренной верхушкой, масса
сердца 0,2—0,4 кг.
Двумя продольными бороздами снаружи и мышечной перегородкой
внутри сердце делится на правую (ушковую) и левую (предсердную) половины, не сообщающиеся (после зарастания овального отверстия) между собой.
Каждая половина состоит из предсердия atrium и желудочка — ventriculus,
сообщающихся между собой атриовентрикулярными (предсердожелудочковым) отверстием. Поперек сердца проходит венечная борозда, отмечающая
границу между предсердиями, расположенными в основании сердца, и желудочками, составляющими его основную массу. Внешние размеры левого желудочка больше, чем правого, из-за большей толщины его стенки. Ему всегда
принадлежит верхушка сердца. В продольных и венечных бороздах проходят
сосуды и откладывается жировая ткань. Особенно много ее лежит по венечной борозде. Общее количество жировой ткани у крупного рогатого скота
может достигать 10—15% массы сердца.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Рис. 136. Сердце крупного рогатого скота справа (А) и вскрытое (Б):
1 — верхушка сердца; 2 — продольная борозда с сосудами; 3 — правый желудочек; 4 — правое предсердие; 5—непарная вена; 6 — каудальная полая вена; 7— легочные вены; в — легочная артерия; 9 — аорта; 10 — левый желудочек; 11 — веночная борозда; 12 — венечная артерия; 13 — левое ушко; 14
— венозный сииус; 15 — краниальная полая вена; 16 — плечеголовной ствол;
17 — гребешковые мышцы; 18 — правый атриовентрикулярный клапан; 19
— поперечная мышца; 20 — межжелудочковая перегородка; 21 — эндокард;
22 — миокард; 23 — эпикард; 24 — сосочковая мышца; 25 — сухожильные
струны; 26 — створка митрального клапана; 27 — кармашковый клапан.
Стенка сердца состоит из трех слоев: внутренний — эндокард, средний
— миокард, наружный — эпикард. Эндокард состоит из тонкого слоя соединительной ткани, выстланного со стороны полости сердца эндотелием. Миокард — мощный слой сердечной поперечнополосатой мышечной ткани (см.
раздел третий). Толщина миокарда в левом желудочке в 2—5 раз больше, чем
в правом. Эпикард состоит из тонкого слоя соединительной ткани, покрытого
снаружи мезотелием.
Предсердия в сравнении с желудочками являются тонкостенными камерами. Они имеют выпячивания — ушки предсердий, которые изнутри имеют
вид губки из-за гребешковых мышц. Мышцы способствуют полному выжиманию крови при сокращении камеры. В межпредсердной перегородке гребешковые мышцы отсутствуют. В правое предсердие впадают одна против
другой 2 самые крупные вены организма — краниальная и каудальная полые
вены. Между ними в стенке предсердия имеется межвенозный бугорок, препятствующий образованию турбулентных вихрей при слиянии их кровяных
потоков. Невдалеке от полых вен в правое предсердие впадают непарная вена
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
(или в основание краниальной полой вены) и сердечные вены. В левое предсердие впадают 4—7 легочных вен.
Через атриовентрикулярное отверстие кровь поступает в желудочки, а
оттуда — в артерии. Из левого желудочка выходит самая крупная артерия
организма — аорта, из правого — ствол легочной артерии.
Перемещение крови в сердце обеспечивается последовательным сокращением (систола) и расслаблением (диастола) предсердий и желудочков.
Клапанный аппарат сердца обеспечивает движение крови в одном направлении. Клапаны имеются в атриовентрикулярных отверстиях и в основании артерий — в артериальных отверстиях. В правой половине атриовентрикулярный клапан трехстворчатый, в левой— двустворчатый (митральный).
Клапаны могут иметь дополнительные створки, представляющие собой тонкие, но прочные соединительнотканные пластинки, покрытые эндотелием и
прикрепленные сухожильными струнами к сосковым (сосочковым) мышцам,
выступающим из стенки желудочков. При сокращении предсердий створки
текущей кровью прижимаются к стенкам желудочков. При сокращении желудочков давлением крови створки поднимаются как паруса и закрывают атриовентрикулярные отверстия. Кровь устремляется в артерии. В основании
артерий имеются трехстворчатые кармашковые клапаны. Каждый такой клапан состоит из трех полулунных створок в виде кармашков.
При попытке обратного тока крови они наполняются кровью и закрывают отверстие аорты и легочного ствола.
Желудочки сердца имеют мощный миокард, в разных участках которого
мышечные пучки образуют от 2 до 5 разнонаправленных слоев. В полость
желудочков выступают нечетко отграниченные от стенки утолщения — мышечные перекладины. Они выполняют ту же функцию, что и гребешковые
мышцы предсердий. Межжелудочковая перегородка вдается в правый желудочек, поэтому на разрезе полость левого желудочка оказывается округлой, а
правого — полулунной. В полости желудочков можно видеть 1—2 сравнительно тонких мышечных тяжа, перекинутых между межжелудочковой перегородкой и боковой стенкой поперечные мышцы.
Фиброзный скелет сердца расположен между предсердиями и желудочками на уровне венечной борозды. Он представляет собой каркас вокруг атриовентрикулярных и артериальных отверстий. В фиброзном кольце аорты у
крупного рогатого скота имеются 2 кости сердца, у лошади — 2—3 хряща, у
свиньи — 1 хрящ. К фиброзному скелету прикрепляются мышечные пучки
предсердий и желудочков.
Околосердечная сумка и серозная оболочка сердца. Сердце покрыто серозной оболочкой, которая образует вокруг него мешок— серозный перикард. Он состоит из висцерального и париетального листков, между которыми находится щелевидная перикардиальная полость. Висцеральный листок
перикарда вплотную прилежит к миокарду и называется эпикардом. Париетальный листок срастается с фиброзным листком, отходящим от внутригрудной фасции, с наружной стороны фиброзный листок фасции покрыт перикардиальной плеврой. В результате сращения париетального листка периwww.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
карда, фиброзного листка внутригрудной фасции и перикардиальной плевры
образуется околосердечная сумка. Она изолирует сердце от окружающих органов; укрепляет сердце в определенном положении, так как от нее отходят
связки к грудине и диафрагме; создает оптимальные условия для функционирования, поскольку клетки серозного мешка секретируют небольшое
количество серозной жидкости, снижающей трение при движении сердца.
Сосуды и нервы сердца. От основания аорты отходят правая и левая
венечные артерии (аа. coronariae). В них поступает 10% крови, выталкиваемой левым желудочком. От них по продольным бороздам отходят нисходящие ветви, распадающиеся на более мелкие артерии и капилляры, питающие
сердце. Кровь собирается в малые сердечные вены с правого желудочка, в
среднюю сердечную вену и в большую сердечную вену (v. cordis magna) с остальных участков.
Вены открываются в правое предсердие.
Сердце иннёрвируется вегетативной нервной системой. Симпатические
нервы идут от звездчатого узла (сердечные ветви), стимулируют сердечную
деятельность (учащают сокращения). Парасимпатические ветви идут от блуждающего нерва и замедляют деятельность сердца.
В тесном контакте с вегетативными нервами находится нервномышечная проводящая система сердца. Она обеспечивает ритмичность работы сердца и состоит из двух узлов: синоатриального (синусопредсердного)
и атриовентрикулярного (предсердно-желудочкового) и отходящих от них
волокон, вступающих в тесный контакт с атипичными мышечными волокнами. Они в 2— 3 раза крупнее типичных волокон, содержат мало миофибрилл
и много гликогена. Синоатриальный узел расположен под эпикардом правого
предсердия между краниальной полой веной и правым ушком. Он связан с
мускулатурой предсердий. Атриовентрикулярный узел крупнее, лежит в
межпредсердной перегородке и связывает в работе предсердия и желудочки.
От него отходит атриовентрикулярный пучок, который делится на правую и
левую ножки. Ножки идут в межжелудочковой перегородке и отдают ветви к
наружным стенкам сердца, проходя в поперечных мышцах.
Круги кровообращения
В кровеносной системе различают 2 круга кровообращения; большой и
малый, объединенные с помощью сердца. Большой, системный или трофический круг кровообращения начинаете» аортой, в которую из левого желудочка сердца поступает артериальная кровь. От аорты отходят многочисленные артерии, несущие кровь к органам и стенкам тела. В органах артерии
разветвляются до капилляров. Капилляры объединяются в вены, которые несут кровь к сердцу. С передней половины тела кровь поступает в краниальную полую вену, с задней — в каудальную полую вену. Обе вены вливаются
в правое предсердие. Отсюда кровь идет в правый желудочек, из него в ствол
легочных артерий— начинается малый круг кровообращения.
www.timacad.ru
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Малый, легочный или дыхательный круг кровообращения начинается
стволом легочных артерий, который вскоре делится на две легочные артерии, несущие к легким венозную кровь. В легких легочные артерии разветвляются до капилляров. В них кровь насыщается кислородом. Капилляры объединяются в легочные вены, несущие артериальную кровь и впадающие в левое предсердие. Поступая отсюда в левый желудочек и вливаясь в аорту,
кровь переходит в большой круг кровообращения.
Сосуды малого круга кровообращения следующие.
Ствол легочных артерий — truncus pulmonalis — отходит от правого
желудочка к корням легких, около которых делится на правую и левую легочные артерии — аа. pulmonales dextra et sinistra. Каждая артерия сопровождает главный бронх и ветвится параллельно ветвлению бронхиального дерева.
Легочные вены — vv. pulmonales — идут параллельно легочным артериям и несут артериальную кровь в левое предсердие.
Основные артерии большого круга кровообращения
Аорта—aorta — основной магистральный сосуд большого круга кровообращения. Начинается от левого желудочка сердца. От основания аорты отходят венечные артерии — аа. coronaria, после чего аорта поворачивает назад и вверх и называется дугой аорты. Поднявшись к позвоночнику на уровне 5—6-го грудного позвонка, она становится грудной аортой, а пройдя через диафрагму— брюшной аортой. Вся артериальная