■
НОРМАЛЬНАЯ
ФИЗИОЛОГИЯ
УУЧЕБНИК
Поод редакцией
акаадемика РАМН
КХ.В.Судакова
И З Д А Т Е Л Ь С К А Я ГРУППА
«ГЭОТАР-Медиа»
УДК 612(075.8)
ББК 52.52я73
Н83
Авторский коллектив
Судаков К.В., зав. кафедрой нормальной физиологии ГОУ ВПО Первого МГМУ
им. И.М. Сеченова, профессор, академик РАМН;
Андрианов В.В., д-р мед. наук, профессор кафедры нормальной физиологии ГОУ
ВПО Первого МГМУ им. И.М. Сеченова;
Вагин Ю.Е., д-р мед. наук, профессор кафедры нормальной физиологии ГОУ ВПО
Первого МГМУ им. И. М. Сеченова;
Джебраилова Т.Д., д-р биол. наук, профессор кафедры нормальной физиологии
ГОУ ВПО Первого МГМУ им. И.М. Сеченова;
Киселев И.И., канд. мед. наук, доцент кафедры нормальной физиологии ГОУ ВПО
Первого МГМУ им. И.М. Сеченова;
Умрюхин П.Е., д-р мед. наук, доцент кафедры нормальной физиологии ГОУ ВПО
Первого МГМУ им. И.М. Сеченова.
Н83 Нормальная физиология: у ч е б н и к /п о д ред. К. В. С удакова. —
М . ; Г Э О Т А Р -М ед и а, 2012. — 880 с.: ил.
ISBN 978-5-9704-1965-6
У чебник отраж ает соврем енны е представления об организации
нормальных физиологических функций.
В отличие от традиционных учебников по физиологии, в которых
излагаю тся физиологические свойства отдельных органов, в данном
учебнике приведены системные основы построения функций целого
организма, его гомеостатических, поведенческих и психических процессов.
В учебнике отражен многолетний опыт преподавания курса нормальной
физиологии на кафедре нормальной физиологии Первого Московского
медицинского университета им. И.М. Сеченова.
Рекомендовано Первым Московским государственным медицинским
университетом им. И.М. Сеченова в качестве учебника для студентов
учреждений высшего профессионального образования.
УДК 612(075.8)
ББ К 52.52я73
Права на данное издание принадлежат ООО Издательская группа «ГЭОТАР-Медиа».
Воспроизведение и распространение в каком бы то ни было виде части или целого издания не
могут быть осуществлены без письменного разрешения ООО Издательская группа «ГЭОТАРМедиа».
ISBN 978-5-9704-1965-6
О Коллектив авторов, 2012
О ООО Издательская группа«ГЭОТАР-Медиа», 2012
© ООО Издательская группа «ГЭОТАР-Медиа», оформление, 2012
ОГЛАВЛЕНИЕ
Используемые с о к р а щ е н и я ..............................................................................5
Предисловие (Судаков К.В.)................................................................................7
Общая характеристика физиологии как науки {Судаков К .В .)...........10
Глава 1. Основы ж изнедеятельности {Вагин Ю .Е )...................................13
1.1. Общие свойства живых организм ов................................................13
1.2. М ембранные потенциалы клеток возбудимых тканей ......... 17
1.3. Законы раздраж ения возбудимых т к а н е й ...................................23
1.4. Возбудимость; изменение возбудимости при возбуждении . . . 27
1.5. Торможение ...........................................................................................30
1.6. Проведение возбуждения .................................................................30
1.7. С инаптическая передача возбуж дения .......................................33
1.8. М ыш ечное с о к р а щ е н и е ..................................................................... 39
1.9. С е к р е ц и я ................................................................................................. 50
1.10. Р е ц е п ц и я ...............................................................................................51
Глава 2. Общие принципы построения целого организма
(Судаков К .В)..........................................................................................................53
2.1. К о р р ел я ц и я .............................................................................................53
2.2. Р е г у л я ц и я ...............................................................................................53
2.3. Рефлекторные р е а к ц и и ....................................................................5 4
2.4. С а м о о р г а н и за ц и я ................................................................................58
2.5. С ам орегуляци я...................................................................................... 60
2.6. Ф ункциональны е с и с т е м ы ...............................................................60
Глава 3. Нервная система (Андрианов В.В. ) ..................................................80
3.1. Общая физиология центральной нервной си стем ы ................. 80
3.2. Частная физиология центральной нервной системы ............. 108
3.3. В егетативная нервная с и с т е м а ...................................................... 144
Глава 4. В нутренняя среда организма (Джебраилова Т .Д )................. 169
4.1. Ж и дки е среды организма. Г о м ео стази с..................................... 169
4.2. Гормональная регуляция физиологических ф у н к ц и й ...........171
4.3. К ровь........................................................................................................208
Глава 5. Висцеральные ф у н к ц и и .................................................................257
5.1. Ф изиология сердца (Вагин Ю .Е )....................................................257
5.2. Гемодинамика сосудистого русла (Андрианов В.В) ................. 302
5.3. Д ыхание (Андрианов В .В )................................................................... 344
5.4. С истемны е механизмы голода, аппетита и насы щ ения
(Судаков К .В )................................................................................................. 377
4
Оглавление
5.5. П ищ еварение (Умрюхин П .Е .).......................................................... 391
5.6. Выделение (Вагин Ю.Е.)..................................................................... 442
5.7. Регуляция водно-солевого обмена (Вагин Ю .Е .)......................468
5.8. Обмен веществ и энергии (Киселев И.И., Умрюхин П .Е ) .........485
5.9. Терморегуляция (Вагин Ю .Е.)........................................................ 513
Глава 6. Анализаторы (Андрианов В .В .)......................................................536
6.1. С труктурно-ф ункциональная организация
а н ал и зат о р о в ...............................................................................................536
6.2. Зрительный ан ал и зато р ...................................................................546
6.3. Слуховой а н а л и з а т о р ....................................................................... 560
6.4. Вестибулярный ан ал и затор ............................................................ 568
6.5. Кож ный а н а л и з а т о р ......................................................................... 573
6.6. О бонятельны й а н а л и з а т о р ............................................................ 577
6.7. Вкусовой ан ал и зато р ......................................................................... 580
Глава 7. Двигательные ф ункции (Андрианов В.В.).................................. 583
7.1. О рганизация д в и ж е н и й ...................................................................583
7.2. Вегетативное и эндокринное обеспечение поведенческих
ак то в .................................................................................................................. 618
Глава 8. Поведение и психическая д е я т е л ь н о с т ь ................................ 621
8.1. Общие принципы организации поведения (Судаков К.В.) . . . . 621
8.2. Врожденное и приобретенное поведение (Судаков К.В.) . . . . 649
8.3. Торможение условно-рефлекторной деятельности
(Судаков К.В.)................................................................................................. 662
8.4. С истемная архитектоника поведенческих актов
(Судаков К.В.)................................................................................................. 667
8.5. М отивации (Судаков К.В.).................................................................682
8.6. П амять (Судаков К.В.)......................................................................... 710
8.7. Эмоции (Судаков К .В .)....................................................................... 730
^ 8.8. Боль (Андрианов В .В .)......................................................................... 748
8.9. Системное построение психической деятельности
человека (Судаков К. В ) ................................................................................765
8.10. Трудовая деятельность человека (Судаков К .В .)......................787
- 8.11. Сон (Судаков К .В .)..............................................................................800
8.12. Половые ф ункции человека (Джебраилова Т .Д .)....................819
Контрольные вопросы к главам у ч е б н и к а ............................................. 838
Рекомендуемая дополнительная л и т ер ату р а......................................... 849
Предметный у к а з а т е л ь .................................................................................. 852
ИСПОЛЬЗУЕМЫЕ СОКРАЩЕНИЯ
GLUT
р02
pcb2
АД
АДГ
АДФ
АКТГ
АТФ
ВИП
— трансм ембранны й переносчик глюкозы
— парциальное давление (напряж ение) кислорода
— парциальное давление (напряж ение) углекислого газа
— артериальное давление
— антидиуретический гормон
— аденозиндифосфат, аденозиндиф осф орная кислота
— адренокортикотропны й гормон
— аденозинтрифосфат, аденозинтриф осф орная кислота
— вазоактивны й интестинальны й пептид, вазоинтестиналь­
ный пептид
— высшая нервная деятельность
вид
— вегетативная нервная система
вне
ГАМК — гам м а-ам ином асляная кислота
— гуанозинтрифосфат
ГТФ
ДНК
— дезоксирибонуклеиновая кислота
ж кт
— ж елудочно-киш ечны й тракт
— кортиколиберин, кортикотропин-рилизинг-горм он
КРГ
— лю теинизирую щ ий гормон
лг
л п в п — липопротеины высокой плотности
л п н п — липопротеины низкой плотности
л п о н п — липопротеины очень низкой плотности
МРТ
— м агнитно-резонансная томография
— объем циркулирую щ ей крови
ОЦК
ПНУП — предсердный натрийуретический пептид
ПНУФ — предсердный натрийуретический фактор
— паратиреоидны й гормон, паратгормон
птг
— п озитронно-эм иссионная томография
ПЭТ
РААС — ренин-ангиотензин-альдостероновая система
— быстрое движение глаз (от англ. Rapid Eye Movement)
РЕМ
— рибонуклеиновая кислота
РНК
соэ — скорость оседания эритроцитов
— сом атотропны й гормон, соматотропин
стг
— трийодтиронин
тз
Т4
— тетрайодтиронин, тироксин
— тиреолиберин, тиреотропин-рилизинг-горм он
ТРГ
— тиреотропны й гормон, тиреотропин
ттг
ФКГ
— фонокардиограмма, ф онокардиограф ия
— ф олликулостимулирую щ ий гормон
ФСГ
6
Используемые сокращения
цАМФ — ц и кли ч ески й аденозинм онофосфат
цГМ Ф — цикли ческий гуанозинмонофосфат
ЦНС — центральная нервная система
ЧСС
— частота сердечных сокращ ений
ЭКГ
— электрокардиограм ма, электрокардиограф ия
ЭЭГ
— электроэнцеф алограм м а, электроэнцеф алограф ия
ПРЕДИСЛОВИЕ
Ф изиологии принадлеж ит ведущ ая роль в поним ании законом ер­
ностей жизнедеятельности ж ивотны х и человека. Н ормальная ф изио­
логия как раздел общей физиологии изучает ж изнедеятельность здо­
рового человека и его адаптацию к экстремальны м воздействиям.
В последние годы ф изиология все более углубляется в изучение
молекулярных и генетических основ ф ун кц ий организма. Это важ­
ная тенденция ее развития. В то же время в учебниках по ф изиоло­
гии меньше вни м ани я уделяется ф ун кц иям целого организма в его
тесных взаимодействиях с окружающей средой.
Н астоящ ий учебник нацелен не только на изучение основ ж и з­
недеятельности и работы отдельных органов, но главным образом —
на п оним ание системны х механизмов ф ун кц и он и рован и я целого
организма в его неразры вны х связях с окружаю щей средой.
В основе системной организации ф ун кц ий живы х организмов, и в
частности человека, лежит теория ф ункциональны х систем, предло­
женная П.К. Анохиным. С позиций теории ф ункциональны х систем
целый организм определяется совокупной деятельностью множества
слаж енно взаимосодействую щ их ф ункциональны х систем метаболи­
ческого, гомеостатического, поведенческого и психического уровней
организации.
Авторы учебника — ведущие профессора кафедры нормальной
физиологии Первого М осковского государственного медицинского
университета им. И.М. Сеченова.
В учебнике отраж ен многолетний опы т преподавания курса нор­
мальной ф изиологии на основе теории ф ун кц ион альн ы х систем,
использован значительны й опы т системной униф и каци и курса н ор­
мальной физиологии.
По структуре н астоящ и й учебник значительно отличается от
сущ ествующих отечественны х и зарубежных учебников по ф и зи о­
логии.
В учебнике, наряду с описанием общ их физиологических свойств
отдельных органов, рассматриваю тся системные механизмы ведущих
физиологических ф ункций. В нескольких главах приведены сведе­
ния о системных механизмах поведения, психической и трудовой
деятельности.
В основу данного и здания положен учебник «Нормальная ф и зи о­
логия» (М., 2004), в котором недостаточно полно были изложены
новейшие сведения по физиологии отдельных ф ункций. В настоящ ее
8
Предисловие
издание включен современны й материал, характеризую щ ий деятель­
ность сердца, кровообращ ения, физиологию крови, пищ еварения и
выделения. В соответствии с соврем енны ми взглядами на механизмы
гуморальной регуляции физиологических ф ун кц ий в значительной
мере пересмотрено изложение материала данного раздела учебника.
Более подробно рассмотрено взаимодействие биологически ак ти в ­
ны х вещ еств с разн ы м и ти пам и рецепторов клеток-«миш еней».
Изменено построение раздела, касаю щ егося частной физиологии
эндокринны х ф ункций.
Глава «Основы жизнедеятельности» дополнена описанием впи­
санности живого в пространственно-временной континуум и опере­
жающего отраж ения действительности как универсального свойства
живой материи.
В параграфах «Общая физиология центральной нервной системы»
и «Вегетативная нервная система» рассмотрены современные методы
исследования головного и спинного мозга как в экспериментах на
животных, так и в клинической практике. Приведены схемы основных
видов вегетативных рефлексов.
В параграфе «Ф изиология сердца» показана взаимосвязь потен­
циалов действия кардиомиоцитов и зубцов электрокардиограммы.
Д ана подробная характеристика сердечного цикла. Работа кл ап ан ­
ного аппарата сердца связана с фонокардиограф ическим и тонами
сердца. О писана корреляц ия между зубцами электрокардиограммы
и тонами сердца. Объяснены закономерности нервно-гуморальной
регуляции сердечной деятельности. П роанализирована зависимость
между объемом и давлением крови в сердце. О писаны отличия в
работе правой и левой половины сердца. П оказаны особенности
метаболизма миокарда.
В параграф «Гемодинамика сосудистого русла» введены понятия
«напряж ение сдвига» и «растяжимость» сосудов, используемые в
настоящ ее время для оп исан ия сосудистой гемодинамики. Ф акторы,
последовательно обеспечиваю щ ие движение крови в артериальном
и венозном отделах сосудистого русла, представлены в виде общей
схемы.
В параграфе «Пищеварение» добавлены современные данные о
роли молекулярных механизмов в секреции соляной кислоты и вса­
сывания.
В параграфе «Выделение» подробно разобраны нервно-гуморальная регуляция образования мочи и системные механизмы мочевыделе­
Предисловие
9
ния. Дана характеристика потоотделения, процессов выделения через
легкие и пищ еварительный тракт.
В параграфе «Обмен веществ и энергии» приведены данные об учас­
тии основных биологически активных веществ в регуляции обменных
процессов.
В параграфе «Терморегуляция» добавлены сведения об оздорови­
тельной роли тепло-холодовых процедур. Описаны субъективные и
парадоксальные характеристики термоощущений.
В главе «Анализаторы» и параграфе «Боль» приведены ори ги н ал ь­
ные схемы клеточной организации сетчатки и уровней обработки
зрительной инф орм ации в головном мозге, созданны е на основе
современных данны х, а такж е схемы ноцицептивной и ан тиноцицептивной систем и нейрохимического обеспечения их деятельности.
В разделе описан механизм, лежащ ий в основе феномена облегчения
восприятия болевого ощ ущ ения.
В главе «Д вигательны е ф ун кц ии » приведен ан ал и з о сн ов­
ных характеристик аф ф ерентны х волокон, идущ их от проприоцептивны х рецепторов, их участие в динам ической и статической состав­
ляю щ ей движ ения. О писаны быстро утомляю щ иеся и устойчивые к
утомлению моторные единицы . Представлены также данны е о корково-ядерны х зонах мозжечка в ф орм ировании движ ения, ори гин аль­
ные схемы клеточной структуры мозж ечка и его связей. И зложены
новые данны е об особенностях л окали зац и и мотонейронов в меди­
альны х и латеральны х столбах сп и нн ого мозга, иннервирую щ их
мы ш цы проксим альны х и дистальны х отделов конечностей.
В учебник вклю чены и ллю страц ии из изданного авторам и
атласа «Физиология в динамических схемах» (изд-во «ГЭОТАР-Медиа»,
2009 г.).
Существенно сокращены и отредактированы параграфы учебника,
в которых изложены системные механизмы поведения, психическая и
трудовая деятельность человека.
В некоторых других разделах изменен план изложения материа­
ла, добавлен целый ряд новых иллю страций, сделаны сущ ественные
дополнения и проведены необходимые сокращ ения.
В целом настоящ ий учебник содержит материал, даю щ ий учащ им­
ся современные научные знания в области нормальной физиологии.
Авторы учебника с благодарностью прим ут все деловые зам е­
чания.
ОБЩАЯ ХАРАКТЕРИСТИКА ФИЗИОЛОГИИ
КАК НАУКИ
Ф изиология — часть естественно-научного зн ан ия, н аправленно­
го на поним ание механизмов жизнедеятельности на молекулярном,
органном и системном уровнях.
На основе частны х процессов и механизмов физиология строит
системную д ин ам ику работы целого организма в его неразры вны х
связях с окружаю щ ей средой.
В курсе медицинского образования физиология подразделяется на
нормальную физиологию и патофизиологию.
Н орм альн ая ф и зи ологи я изучает ф у н к ц и и орган и зм а п р ак ти ­
чески здорового человека, особ ен н о — м еханизм ы его устой ч и ­
вости к действию р азл и ч н ы х о тк л о н яю щ и х ф акторов. Т аким и
ф ак то р ам и яв л яю тся чрезм ерны е ф и зи чески е и эм оци он альны е
н агрузки , сол н еч н ая р ад и ац и я , электр о м агн и тн ы е поля, техн оген ­
ное загр язн ен и е окруж аю щ ей человека среды и т.д.
Нормальная физиология — теоретическая основа
здравоохранения
В нимание врачей веками приковано, в первую очередь, к организ­
му больного человека. Однако не менее значим ая задача здравоохра­
нения состоит в том, чтобы не дать здоровому человеку, несмотря на
ты сячи разнообразны х причин, заболеть, т.е. в самы х разнообразны х
социальны х, географических и бытовых условиях использовать естес­
твенны е механизмы здоровья. В этом заклю чается проф илактическая
направленность медицины , и нормальной физиологии здесь п рина­
длеж ит первостепенная роль. Понять механизмы здоровья человека
можно только на основе научного зн ан и я — прежде всего нормальной
физиологии.
Во многих странах мира и во Всемирной О рганизации Здра­
воохранения (ВОЗ) осознали, что акцент медицины должен быть
перенесен на практически здорового человека, укрепление его здоро­
вья, что более оправдано в медицинском и экономическом плане по
сравнению с вложением средств и усилий в лечение уже сложившихся
заболеваний.
Отсюда очевидно, что чем лучш е будут познаны механизмы здо­
ровья, тем эф ф ективнее будет здравоохранение. И менно поэтому
Общая характеристика физиологии как науки
11
нормальная ф изиология является научной теоретической основой
здравоохранения.
Патофизиология, в отличие от нормальной физиологии, является
теоретической основой кли н и ч ески х дисци п лин , изучаю щ их меха­
низмы заболевания человека и выздоровления.
Особенности нормальной физиологии как науки
Социальная направленность
О течественная физиология направлена не только на поним ание
ф ун кц ий организма человека, но и на научное реш ение острых соц и ­
альны х проблем на основе физиологического знания.
Н ачи ная с С.Г. Зы белин а — осн овополож н и ка отечественной
физиологии, выдаю щ иеся русские физиологи (С.Я. Мудров, А.М. Ф иломаф итский, И.М . Сеченов, А.А. Ухтомский, И.П. Павлов, В.Я. Д а­
нилевский, М.Н. Ш атерников, JI.A. О рбели, К.М . Быков, И.П. Разенков, П.К. Анохин и др.) стремились прим енить ф изиологичес­
кие зн ан ия д ля п он и м ан и я ф ун кц и й целого организма, зак он о­
мерностей его связей со средой обитания и с общ ественны м раз­
витием.
И.М. Сеченов, И.П. Павлов, А.А. Ухтомский, П.К. Анохин и дру­
гие крупные отечественны е физиологи в объяснении физиологичес­
ких закономерностей поднимались до ф илософ ских обобщ ений.
Новый уровень физиологической науки связан с развитием систем­
ного подхода к поним анию явлений природы и общества. С истемны й
подход заставил изучать отдельные физиологические процессы в их
тесной связи с другим и сторонами жизнедеятельности и с целым
организмом. С истемны й подход в области физиологии наш ел отра­
жение, в частности, в теории ф ункциональны х систем, предложенной
П.К. Анохины м (см. ниже).
Логическое мышление
В отличие от ряда морфологических дисци п лин , в физиологии
ш ироко прим еняется логическое мы ш ление. Ф изиология требует
умения ф ормулировать задачи исследования, логически и экспери­
ментально их разреш ать и сопоставлять различны е научные факты.
Логическое мы ш ление требует такж е точны х ф ормулировок ф и зи о­
логических закономерностей, умения выразить мысль в виде граф ика
или рисунка.
12
Общая характеристика ф изиологии как науки
Ф изиология — наука экспериментальная
В физиологии для познания закономерностей жизнедеятельности
прим еняю тся как острые, так и хронические гуманно обставленные
опыты на животны х. Научное знание приобретается такж е при п ро­
ведении дозволенны х исследований на людях.
Объект физиологического исследования
О бъектом ф и зи ологи чески х исследован и й яв л яется ж ивой орга­
н изм р азл и чн ого уровн я о р ган и зац и и — от п ростей ш и х до ч ело­
века.
Глава 1
ОСНОВЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
1.1. ОБЩИЕ СВОЙСТВА ЖИВЫХ ОРГАНИЗМОВ
В отличие от неорганического мира, живые организмы в процессе
эволю ционного развития воплотили в себе ряд качественно новых
свойств.
Вписанность живого в пространственно-временной континуум окру­
жающего мира. Земля, как планета, сформ ировалась около 4,5 млрд лет
назад. Ж ивые организмы в их самой прим итивной форме возникли
около 0,5—1 млрд лет назад. Следовательно, ж ивые организмы вп и ­
сались в окруж аю щ ие их явл ен и я неорганического мира Земли:
притяж ение Земли, газовую среду атмосферы, температуру воздуха,
морей и океанов, электром агнитное поле Земли. С войства окруж аю ­
щей живые организмы среды отразились в биологических свойствах
живых сущ еств (П .К . Анохин).
Среда, в которую вписались живые организмы, представляет собой
пространственно-временной континуум событий (А. Эйнштейн). Это
значит, что все явления на Земле тесно связаны во времени и п ро­
странстве. Это не хаос событий, а их материальная и информ ационная
организация. О рганизация явлений физического мира определяется,
прежде всего, соотношением планет солнечной системы, и в первую
очередь соотношением Земли и Солнца. В пространственно-времен­
ном континууме окружающего мира есть явления эпизодические,
например, атмосферные осадки, землетрясения и т.д. Наряду с этим,
имеются явления, периодически повторяющиеся с регулярным рит­
мом. Это смена времен года, приливы и отливы океанов, смена д н я и
ночи. Ж ивые существа отразили в своей организации как эпизодичес­
кие, так и повторяющиеся явления окружающего их мира. Особенно
важны ми для жизнедеятельности оказались воздействия, периодичес­
ки повторяю щиеся на протяж ении ж изни одного поколения.
14
Глава 1
Изоляция живых существ. Ж ивые сущ ества не только вписались в
пространственно-временной континуум внешнего мира, но и изоли­
ровали себя от неживой природы.
И золяц и я осущ ествляется универсальны м способом с помощью
ф осф оли п идн ы х мембран. Характерно, что мембраны различны х
клеток — оболочки яй ц еклетки морского ежа и мембраны нервных
клеток — почти одинаковы е по строению . М ембраны позволили
ж ивы м организмам противопоставить себя водной среде, в которой
они первично возникли, активно воздействовать на окружаю щ ую их
среду и соверш енствоваться в своей организации. И золяц и я создала
предпосы лки ф орм ирования ф ункциональны х свойств живых субъ­
ектов.
Раздражимость. На мембраны воздействуют различны е ф ак то­
ры внеш ней среды, которые п олучили название раздраж ителей.
Раздражители подразделяю тся на:
— ф и зические (м еханические, тем пературны е, электрические
и т.п.);
—хи м ические (кислоты , щ елочи, соли, гормоны , медиаторы
и т.п.);
— ф изико-хим ические (изменение осмотического давления, реак­
ц ии среды, ионного состава и др.);
— биологические (бактерии, вирусы и пр.);
— инф орм ационны е, несущ ие, наряду с ф изическим и и хим ичес­
ки м и свойствами, определенную информ ацию , обусловлен­
ную взаимодействием ж ивых сущ еств и происходящ их в них
физиологических процессов. Это разнообразны е эм оциональ­
ные состояния, сигналы призывов и опасности у животны х,
речь человека и т.д.
Раздраж имость — способность живы х существ отвечать н а дейс­
твие раздраж ителей изменением своих протоплазматических свойств,
в первую очередь, изменением структуры составляю щ их их клеточ­
ных мембран. Различаю т два вида раздраж имости: неспециф ическая
и избирательная.
Неспецифическая (триггерная) раздраж им ость не зависит от качес­
тва раздраж ителя. Она обусловлена внутренним и процессами, п ро­
исходящ ими под влиянием внеш них раздраж ителей в самом живом
объекте, в первую очередь, в его клеточны х мембранах. Этот процесс
напом инает процесс спуска курка ружья («триггер» в переводе с
английского — «курок»). Раздраж итель постепенно доводит молеку­
Основы жизнедеятельности
15
лярны е изменения в клеточны х мембранах до критического уровня
и порождает специф ический ответ ж ивого объекта.
Специфическая (избирательная) раздраж имость проявляется пре­
имущественно по отношению к действию биологически активных, в
частности, лекарственных веществ. Показано, что в отдельных учас­
тках мембраны клеток находятся рецепторные зоны — специальные
белковые молекулы, наиболее чувствительные к действию определен­
ных химических веществ, с которыми они вступают во взаимодействие
типа «ключ—замок». Эти структуры получили название молекулярных
рецепторов. Химические вещества — лиганды, специфически взаим о­
действуя с рецептором, вызывая биохимические реакции в мембране
и протоплазме клеток. Избирательная раздраж имость связана также
с диаметром белковых ионных каналов мембран, через которые могут
проникать молекулы только определенного размера.
Р аздраж им ость — первичное проявлен ие отн ош ен и я ж ивы х
существ к действию факторов внеш ней среды. В ходе эволю ции
живых сущ еств раздраж им ость трансф орм ировалась в возбудимость
нервной и мыш ечной ткани.
Память. У ниверсальны м свойством ж ивы х объектов явл яется
память, т.е. способность ф иксировать молекулярны е и зм енения,
вызванные тем или ины м раздраж ителем, хранить следы этих раз­
драж ений и впоследствии воспроизводить эту информацию .
Свойство пам яти наиболее отчетливо проявляется по отнош ению
к повторно действую щ им раздраж ителям. Однако запом инаю тся и
единичны е сильны е воздействия, особенно вызывающ ие у живы х
существ эм оциональны е реакции. Характерно, что механизмы п ам я­
ти прин ц ип и ально одинаковы у ж ивы х объектов различного уровня
организации. Они связаны с ф ункционированием универсального
генетического аппарата.
Опережающее отражение действительности. Общ им свойством
живых объектов, описанны м П.К. А нохины м, является способность
к опережающему отраж ению окруж аю щ их событий. Это свойство
живых сущ еств тесно связано с сигнальны м значением условных
раздражителей, откры ты х И.П. Павловым.
О пережаю щ ее отраж ение действительности имеет отнош ение,
прежде всего, к периодически повторяю щ им ся воздействиям на
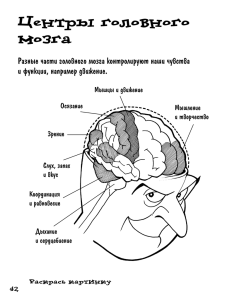
живые организмы (рис. 1.1).
Каждое последующее воздействие внешней среды оставляет в живом
организме соответствующие молекулярные изменения. При многократ-
16
Глава 1
I ----------------------II --------------------------- ► III----------------------►- IV
I
-----------------------------------------------------► Ш ----------------------► IV
Рис. 1.1. Опережающее отражение действительности — одно из свойств
живой материи: А, Б, В — этапы возникновения в организме опережа­
ющего отражения действительности (I, И, III, IV — последовательность
событий во внешней среде; 1, 2, 3, 4 — последовательность внутренних
процессов в организме)
ных повторяющихся воздействиях молекулярные изменения в организ­
ме связываются в химический континуум. Благодаря этому при очеред­
ном действии первого внешнего раздражителя быстро осуществляется
вся последовательная цепь молекулярных изменений. Молекулярные
изменения организма опережают последующие внешние раздражения.
Основы жизнедеятельности
17
1.2. МЕМБРАННЫЕ ПОТЕНЦИАЛЫ КЛЕТОК
ВОЗБУДИМЫХ ТКАНЕЙ
Поляризация клеточных мембран
По отнош ению к внеш нему раздраж ению ткани организма делятся
на невозбудимые (эпителиальная, соединительная, костная) ткан и и
возбудимые (нервная и мы ш ечная) ткани . При раздраж ении нервной
или мыш ечной ткан и в ней возникает возбуждение, которое распро­
страняется по этой ткани от места раздраж ения.
В условиях ф ункционального покоя при отсутствии раздраж ений
поверхностные мембраны клеток возбудимой ткан и п оляри зова­
ны. В нутренняя поверхность мембран заряж ена отрицательно, а
наружная — положительно. Это связано с работой натрий, к а л и е­
вой аденозинтриф осф атазы ( № +,К +-АТФ-азы) — фермента, который
встроен в поверхностную мембрану возбудимой клетки и за счёт
энергии, выделяю щ ейся при расщ еплении аденозинтриф осф атазы
(АТФ), закачивает внутрь клетки ионы К+ и переносит наружу ионы
Na+ (рис. 1.2). Во внутриклеточной ж идкости ионов К+ в 20—30 раз
больше, чем во внеклеточной ж идкости. Внутри клетки они связаны
с отрицательно заряж енны м и белкам и цитоплазм ы . С наружи клетки
Na+ в 10—15 раз больше, чем внутри. В неклеточная ж идкость такж е
содержит в 20—30 раз больше С а2+ и Mg2+, что обусловлено работой
мембранных ферментов. Ионы С1_ уравновеш иваю т полож ительны й
заряд катионов во внеклеточной ж идкости, и поэтому их в 15—25 раз
больше снаружи клетки, чем во внутриклеточной ж идкости.
Основными ионами, формирую щ ими п отенциал покоя и потен­
циал действия, являю тся ионы Na+ и К +. Ионы N a+ стрем ятся по
градиенту концентрации войти внутрь клетки, но в покое мембрана
для них непроницаем а. П оэтому снаруж и мембраны сохраняется
высокая кон ц ен трац ия ионов N a+. Д ля ионов К+ мембрана частично
проницаема, и они пассивно по концентрационном у градиенту выхо­
дят через калиевы е поры из клетки. Эти поры образованы мембран­
ными белкам и и назы ваю тся каналам и утечки ионов К+.^Вышедшие
ионы скапливаю тся около наружной поверхности мембраны, так как
на внутренней поверхности мембраны остаю тся отрицательно заря­
женные крупны е анионы белковых молекул цитоплазм ы , которые
электростатически притягиваю т ионы К+. П оэтому из клетки выхо­
дит менее 1% ионов К+. У станавливается динам ическое равновесие
между концентрационны м градиентом ионов К+, определяю щ им их
Глава 1
18
Na
Клеточная
мембрана
АТФ-аза
10Na+ , К+
*
30К+ , Na+
Цитоплазма
lONa
15С1
,+ К
К
к+
К
к
+
А-
Л
А-
+
+
.
А'
К+
А'
А -
^
А~
' 3 0 К
' С '
А
'
А -
Рис. 1.2. Последовательность ионных процессов спонтанной поляриза­
ции клетки возбудимой ткани: А — активный транспорт ионов Na+ и К+;
Б — неравномерное распределение ионов Na+ и К+ снаружи и внутри
клетки; В — пассивный выход ионов К+ по градиенту концентрации;
Г — возникновение калиевого равновесного потенциала. Ионы К+ движут­
ся наружу по градиенту концентрации и внутрь по электростатическому
градиенту; Д — ионы С1- распределяются асимметрично распределению
ионов К+. Незначительное пассивное вхождение ионов Na+ и С1—изменяет
величину калиевого равновесного потенциала
Основы жизнедеятельности
19
выход из клетки, и электростатическим градиентом, обусловлива­
ющим прекращ ение выхода ионов К+ и скопление их на наружной
поверхности мембраны. На мембране возникает калиевый равновес­
ный потенциал, которы й является одной из наиболее важ ны х причин
поляризации мембран клеток в покое.
Регистрация уровня п оляри зац ии мембраны осущ ествляется с
помощью стекл ян н ы х микроэлектродов, которые вводятся внутрь
клетки, не вы зы вая разруш ения поверхностной мембраны. Разность
потенциалов между макроэлектродом на наруж ной поверхности мемб­
раны, имеющей полож ительны й заряд, и кончиком м икроэлектрода на
внутренней поверхности мембраны, заряж енной отрицательно, вызы­
вает ток между электродами, который регистрируется электронны м и
приборами. В нутриклеточны й способ регистрации уровня заряда кл е­
точной мембраны обусловливает то, что анализ изменений величины
поляризации мембраны ведётся на внутренней поверхности мембра­
ны. В еличина потенциала покоя имеет значения от —50 до —100 мВ,
что вызвано разной активностью А ТФ -азы и разны м количеством пор
для ионов К+ в разли чн ы х возбудимых клетках.
Одиночное возбуждение
Действующие на возбудимую ткань раздраж ения делятся по силе
на подпороговые, пороговые и надпороговые. При подпороговых раз­
драж ениях потенциал покоя мембраны уменьш ается на небольш ую
величину в месте раздраж аю щ их электродов. Это изменение назы ва­
ется локальны м ответом. В еличина локального ответа увеличивается
при увеличении силы подпорогового раздраж ения. При пороговом
раздраж ении локальны й ответ достигает уровня критической депо­
ляризации мембраны и переходит в потенциал действия, который
характеризует изменение заряда на мембране при распростран яю ­
щемся от раздраж аю щ их электродов возбуж дении. В потенциале
действия выделяю т несколько частей: предспайк, спайк, отрицатель­
ный и полож ительны й следовые потенциалы (рис. 1.3).
В нейронах и скелетных миоцитах спайк длится более короткое
время, чем следовые п отенциалы , длительность и наличие которых
непостоянны. Ф орма и длительность потенциалов действия сущ ес­
твенно различаю тся в разли чн ы х возбудимы х тканях: нейронах,
рецепторах, м иоцитах скелетных и гладких мы ш ц и кардиомиоцитах.
Амплитуда п отенциала такж е различна в разны х ткан ях и в то же
время п остоянна в каж дой клетке. А мплитуда п отенциала действия
в нервном волокне равняется примерно 100 мВ.
20
Глава 1
h-ворота открываются
Основы жизнедеятельности
21
Постоянство амплитуды потенциала действия отражено в законе
раздражения возбудимых тканей «Все или ничего», который гласит, что
при подпороговом раздражении потенциал действия не возникает, а при
пороговом и надпороговых раздражениях возникает потенциал действия
максимальной амплитуды. Закону «Все или ничего» подчиняются нейро­
ны, нервные волокна, мышечные волокна, гладкие мышцы и сердечная
мышца. Hg,п о д чиняются этому закону нервы и скелетные мышцы. Это
обусловлено различным порогом раздражения нервных волокон, входящих
в состав нерва, и мышечных волокон, образующих скелетную мышцу.
При возникновении потенциала действия начинаются процессы,
вызывающие изменение поляризации мембраны. Во время предспайка происходит медленная частичная деполяризация мембраны за счет
постепенного откры тия в мембране пор для ионов Na+, вхождения их
в клетку и частичного уменьш ения отрицательного заряда на внутрен­
ней поверхности мембраны (см. рис. 1.3. аБ). При достижении уровня
критической деполяризации происходит открытие всех пор для ионов
Na+, и лавина ионов Na+ пассивно по градиенту концентрации вхо­
дит внутрь клетки, вызывая быструю полную деполяризацию, а затем
инверсию знака заряда на мембране (см. рис. 1.3. аВ). Изменение знака
заряда н а ^мембране вызывает открытие электрозависимых пор ионов
К+, и ионы К+ выходят наружу клетки, вызывая повторную поляри­
зацию мембраны, которая называется реполяризацией (см. рис. 1.3. аГ).
Иногда возникает явление повышенной поляризации или гиперполя­
ризации мембраны, ионные механизмы которого связаны с повыш ен­
ным выходом наружу ионов К+ и иногда с входом в клетку ионов С1_
(см. рис. 1.3. аД). После окончания потенциала действия Na+,K+-ATO-
Рис. 1.3. Динамика ионных процессов при возбуждении: А — состояние
покоя в клетке возбудимой ткани; Б — частичная деполяризация при
подпороговом раздражении; В — деполяризация при пороговом или надпороговом раздражении; Г — реполяризация; Д — гиперполяризация;
Е — возвращение исходного распределения ионов; а — распределение ионов
вокруг клеточной мембраны; б — мембранные потенциалы; в — положение
ворот в натриевом и калиевом электрозависимых каналах (V —напряжение
на внутренней поверхности мембраны; t — время; ПП — потенциал покоя;
КД критическая деполяризация; Р - раздражение; ЛО — локальный
ответ; ПС — предспайк; Деп —деполяризация; И — инверсия; С — спайк;
Реп — реполяризация; ОСП — отрицательный следовой потенциал;
ПСП —положительный следовой потенциал; ПД —потенциал действия)
22
Глава 1
аза перекачивает наружу ионы Na+ и внутрь ионы К+ при сохраняю ­
щемся потенциале покоя мембраны (см. рис. 1.3. аЕ).
Ионные каналы клеточной мембраны образованы мембранны ми
белками. К ажды й канал имеет форму трубочки в молекуле белка.
Отверстие канала перекрыто вы ростами этого белка, назы ваемы ми
воротами канала. Н атриевый канал имеет т - и h-ворота. В состоя­
н ии покоя m -ворота закры ты , и ионы N a+ не проходят через канал
(см. рис. 1.3 вА). При этом h -ворота открыты. Ворота электрозависимых калиевы х каналов такж е закрыты . При этом калий выходит из
клетки через другие каналы , назы ваемы ми каналам и утечки калия.
Раздражение возбудимой клетки изменяет форму белка натриевых
каналов, и m -ворота открываю тся сначала частично (см. рис. 1.3. вБ),
а затем полностью (см. рис. 1.3. вВ). Ионы Na+ входят снаружи внутрь
клетки за счет разности их концентрации. Входящие в клетку поло­
ж ительно заряженные ионы N a+ действуют на электрочувствительную
область h-ворот, и они закрываю тся (см. рис. 1.3. вГ). Одновременно
положительный заряд на внутренней поверхности мембраны вызывает
открытие ворот электрозависимых каналов для ионов К+. Ионы К+
выходят из клетки, вызывая реполяризацию мембраны (см. рис. 1.3. вД).
М ембранный потенциал возвращается к уровню потенциала покоя.
Последующее активное перераспределение ионов № +,К+-АТФ-азой
приводит к закрытию m -ворот и открытию h-ворот в натриевых кан а­
лах и закрытию ворот в калиевых каналах (см. рис. 1.3. вЕ).
Особенности местного и распространяю щ егося
возбуждения
Местное возбуждение возникает при действии подпороговых сти­
мулов, проявляется локально, только в зоне действия стимула и
быстро затухает в смежном с раздражением месте. При повторных
воздействиях местное возбуждение способно к сум м ации и может
перейти в распространяю щ ееся возбуждение. Оно характеризуется
местным потенциалом или локальны м ответом, при котором повы­
ш ается возбудимость клеточной мембраны. Л окальн ы й ответ не
подчиняется закону «Все и ли ничего».
Распространяющееся возбуждение возникает при действии пороговых
и надпороговых раздражений или в результате суммации до критического
уровня местных возбуждений. На клеточной мембране возникает потен­
циал действия. Потенциал действия распространяется на значительное
расстояние от места возникновения без уменьшения исходной величины.
Потенциал действия не способен к суммации и подчиняется закону «Все
Основы жизнедеятельности
23
или ничего». При распрострвняющемся возбуждении возбудимость изме­
няется сложным образом —то увеличиваясь, то уменьшаясь.
1.3. ЗАКОНЫ РАЗДРАЖЕНИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Законы раздраж ения регламентирую т параметры раздраж ения,
которое необходимо для появления распространяю щ егося возбуж­
дения в возбудимой ткани.
Закон «Все или ничего» характеризует необходимую для возбуж де­
ния силу раздраж ения при одиночном раздраж ении прямоугольны м
импульсны м током.
При пороговых и надпороговых раздраж ениях в возбудимых тк а­
нях возникаю т потенциалы действия, ам плитуда которых не зависит
от силы раздраж ения. При подпороговых раздраж ениях ам плитуда
местных потенциалов увеличивается при усилении подпорогового
раздражения.
Закон «Силы—времени» определяет зависимость между амплитудой
и длительностью раздражающего импульса, достаточного для возник­
новения возбуждения. Чем короче время раздраж ения, тем с большей
силой надо раздраж ать ткань,
чтобы вызвать возбуждение.
Кривая «силы—времени» (рис. 1.4)
образована точкам и, каж дая из
которых характеризует в ел и чи ­
ну силы порогового раздраж ения
при определенной длительности
раздражения. Точки пространс­
тва слева от кривой характеризу­
ют подпороговые раздраж ения, а
справа от кривой — надпороговые раздраж ения.
В зависим ости от д л и тел ь­
ности раздраж ен ия возбудимая
ткан ь им еет м нож ество зн а­
Рис. 1.4. Зависимость между силой
чений п о р о го вы х р азд р аж е­
(V) и временем (t) порогового раз­
ний. М и н и м альн ы й порог при
дражения: Р-реоб аза; 2Р—удвоен­
неограниченном врем ени р а з­
ная реобаза; МПВ —минимальное
драж ения н азы вается реобазой.
полезное время; Хр —хронаксия
М инимальное время, в течение
24
Глава 1
которого раздраж ение силой в одну реобазу вызывает возбуждение
ткан и , называется минимальным полезным временем.
Установлено, что реобаза не всегда точно характеризует возбуди­
мость исследуемой ткани. Поэтому предложено измерять возбуди­
мость ткани с помощью определения хронаксии. Хронаксия — время,
в течение которого ток, равны й удвоенной реобазе, вызывает воз­
буждение ткани. В кли н и ч ески х условиях хронаксию определяю т с
помощью прибора хронаксиметра.
Кривые «силы—времени» и хронаксия различны у различны х тканей
в разны х видах ж ивотны х и человека. Хронаксия скелетной мыш цы
человека измеряется в миллисекундах. Хронаксия гладкой мы ш цы
улиток и желудка лягуш ек длится до десятых долей секунды.
Закон аккомодации характеризует скорость увеличения силы раз­
драж ения, необходимой для возбуж дения. Чем ниж е эта скорость,
тем через больш ий интервал времени появится возбуждение. При
акком одации происходит привы кание ткани к действию постепенно
увеличиваю щ ейся силы раздраж ения.
Закон сформулирован французским физиологом Э. Дюбуа-Реймоном.
Он обнаружил, что ткань отвечает на раздражение при резком нараста­
нии тока пороговой силы. Ток пороговой силы при медленном нараста­
нии не оказывает раздражающего действия на ткань. Изменение порога
раздражения во времени получило название аккомодации.
А кком одация определяет повыш ение порога раздраж ения в зави­
симости от скорости нарастания силы раздраж ителя. При медленном
н арастан ии постоянного тока он может не вызвать возбуж дения
вследствие сниж ения возбудимости ткани (рис. 1.5). В основе явл е­
н и я акком одации леж ит уменьш ение натриевой и повыш ение кал и ­
евой проводимости мембраны возбудимой ткани.
Рис. 1.5. Изменение амплитуды потенциала действия и уровня критической
деполяризации нервного волокна при постепенном увеличении силы раздра­
жения: А — мембранный потенциал при замедлении увеличения силы раз­
дражения (V —напряжение на внутренней поверхности мембраны; t —время;
ПП —потенциал покоя; КД —критическая деполяризация; 1-3 —потенциалы
действия; 4 —локальный ответ; 5 — мембранный потенциал при медленно
увеличивающемся подпороговом раздражении); Б — форма раздражающих
пороговых импульсов тока (1 — прямоугольный импульс; 2-5 — импульсы
с разной скоростью нарастания переднего фронта)
Основы жизнедеятельности
Б
,Л
25
26
Глава 1
Разные ткани обладают разной аккомодацией. Так, например,
чувствительные нервные волокна обладают меньшей аккомодацией
по сравнению с двигательны м и волокнами. Болевые волокна имеют
низкую аккомодацию.
Полярный закон определяет зависимость возникновения возбуж­
дения от полярности раздраж аю щ их электродов. Э. Пфлюгер обнару­
жил различия в действии анода и катода при вклю чении и вы клю че­
н ии постоянного тока, раздраж аю щ его ткань. При зам ы кан ии цепи
постоянного тока возбуждение возникает под катодом. При разм ы ка­
н и и цепи постоянного тока возбуждение возникает под анодом.
В озникновения возбуж дения под катодом при вклю чении тока
и под анодом при выклю чении тока обусловлено изменением воз­
будимости клеточны х мембран ткани. При начальном прохождении
постоянного тока через возбудимую ткань под катодом возбудимость
повышается (катэлектротон), а под анодом — пониж ается (анэлектротон). При продолжающемся действии постоянного тока под като­
дом развивается пониж ение возбудимости (католическая депрессия),
а под анодом — повыш ение возбудимости (анодная экзальтация).
Функциональная лабильность
Ф у н кц и ональн ая лабильность — способность возбудимых тканей
воспроизводить частоту поступаю щ их на них раздраж ений без тран с­
ф орм ации ритма раздраж ения (Н.Е. Введенский). Мерой лабильнос­
ти является максим альная частота раздраж ения, которую возбуди­
мая ткань воспроизводит без трансф орм ации ритма раздраж ения.
Р азличны е ткани обладают разной ф ункциональной лабильностью.
Наиболее лабильна нервная ткань. Так, диаф рагм альны й нерв может
воспроизводить частоту раздраж ения импульсов без трансф орм ации
ритма до одной ты сячи импульсов в секунду, скелетные мы ш цы —
до 100—200 и м п /с и нервно-мы ш ечны е синапсы — до 50—100 имп/с.
Оптимум и пессимум раздражения. Если высокая частота ритма раздра­
жения не превышает меру лабильности возбудимой ткани, то ткань отве­
чает на каждый стимул. Состояние возбудимой ткани при этом получило
название оптимума. В случае, когда частота раздражения превышает меру
лабильности ткани, в ней развивается состояние пессимума.
М еханизм оп ти м ум а определяется повы ш ением возбудим ос­
ти ткани при ее возбуждении. Если последующие раздраж аю щ ие
импульсы попадаю т в период повы ш енной возбудимости предш ест­
вующих циклов возбуж дения, ответная реакц ия ткани возрастает до
максим альной величины .
Основы жизнедеятельности
27
М еханизм п ессим ум а обусловлен ум еньш ением возбудим ос­
ти ткани при ее возбуждении. Если последующие раздраж аю щ ие
импульсы попадаю т в период отсутствия возбудимости предыдущ его
возбуждения, новое возбуж дение не возникает.
Парабиоз. И зучая свойства возбудимы х тканей при м нож ест­
венны х раздраж ен иях, Н.Е. В веденский разработал представления
о парабиозе (пара (гр.) — около, при; биоз (гр.) — жизнь). П арабиоз —
уменьш ение возбуди м ости тк а н и с последую щ им тормож ением
за счет застойного возбуж дения. П арабиоз может возни кать при
действии н а возбудимую ткан ь разл и чн ы х хи м и чески х вещ еств и
ф и зических раздраж ителей, таких, к а к н агревание, охлаж дение,
электром агнитное излучение. Я влени я парабиоза возни каю т при
вхождении человека в наркоз, при отравлен и ях, при приеме а л к о ­
голя и наркотиков, при ш оковых состоян иях. К аж дое из этих ф ун к­
ц иональны х со сто ян и й организм а возн и кает благодаря разл и чн ы м
ф изиологическим и биохим ическим механизмам. О днако во всех
случаях общ ей чертой п арабиотических процессов явл яется опреде­
ленная стади й ность н аступ лен и я перевозбуж дения с последую щ им
торможением.
Имеется три стадии парабиоза. В первой, уравнительной, стадии
слабые и сильны е раздраж ения вызываю т одинаковую силу ответов
возбудимой ткани . Вторая стадия является парадоксальной. Слабые
раздраж ения вызываю т сильны е ответные реакц ии , а сильны е раз­
драж ения — слабые. Третья стадия — тормозная, когда возбудимая
ткань перестает отвечать на раздраж ения.
1.4. ВОЗБУДИМОСТЬ; ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ
ПРИ ВОЗБУЖДЕНИИ
Возбуждение — процесс, характеризую щ ийся изменением заряда
клеточных мембран при действии определённой силы раздраж ения.
Возбудимость — свойство возбудимой ткан и возбуждаться при дейс­
твии раздраж ения. Возбудимость ткан и характеризую т величиной
порога раздраж ения, продолж ительностью латентного периода раз­
драж ения, хронаксией, ф ункциональной лабильностью и скоростью
аккомодации. Основной мерой возбудимости является порог раздра­
жения. Порог раздражения — м и н им альн ая сила раздраж ения, кото­
рая вызывает распространяю щ ееся возбуждение, характеризую щ ееся
потенциалом действия. В еличина порога раздраж ения д ля каждой
28
Глава 1
клетки и ндивидуальна и зависит от разности между уровнем заряда
мембраны и уровнем критической деполяризации. Возбудим ость и
порог раздраж ения находятся в обратно пропорциональной зависи­
мости. Чем больше порог, тем меньше возбудимость и наоборот. Так,
например, нервная ткань характеризуется высокой возбудимостью
и н изким порогом, а м ы ш ечная ткань — низкой возбудимостью и
вы соким порогом.
Д ля того чтобы исследовать, что происходит с возбудимостью
при возбуж дении, нуж но после первого раздраж ения, вызывающего
возбуждение, многократно повторно раздраж ать возбудимую ткан ь в
различны е фазы потенциала действия. Необходимо для каждой фазы
потенциала действия найти порог раздраж ения, то есть м и н и м ал ь­
ную силу раздраж ения, которая вызовет второе возбуждение. График
изменения возбудимости во время потенциала действия строится
эксперим ентально по точкам , в отличие от граф и ка п отенциала
действия, которы й регистрируется на электронной аппаратуре при
раздраж ении возбудимой ткан и (рис. 1.6).
Во время предспайка разница между уровнем заряда мембраны
и уровнем критической деполяризации уменьш ается. П оявляется
возможность дополнительны м пороговым раздраж ением меньшей
величины быстрее откры ть поры для ионов N a+, достичь уровня
критической деполяризации мембраны и быстрее вызвать потенци­
ал действия. Возбудимость к ак обратная величина порога при этом
увеличивается и становится супернормальной.
При спайке откры ваю тся все поры для ионов N a+, и натрий посту­
пает снаружи внутрь клетки. Поэтому н икакая величина раздраж е­
ния не способна усилить процесс возбуждения. Порог раздраж ения
увеличивается до бесконечности, а возбудимость, как обратная вели­
чина, падает до нуля. Падение происходит мгновенно при достиж е­
н ии уровня критической деполяризации мембраны. Н аступает пери­
од абсолютной рефрактерности или полной невозбудимое™ . Свойство
возбуждаться исчезает, так к ак мембрана уже возбуждена.
Во время реполяризации поры для ионов Na+ постепенно закрываются,
поэтому появляется возможность их заново открыть дополнительным
раздражением. Однако сила раздражения должна быть выше, чем величи­
на исходного порогового раздражения. Наступает период относительной
рефрактерности, когда возбудимость постепенно восстанавливается до
исходного уровня. Фазы абсолютной и относительной рефрактерности
имеют важное физиологическое значение для понимания механизмов
многих функциональных процессов в возбудимых тканях.
29
Основы жизнедеятельности
V, мВ
С
Рис. 1.6. Изменение возбудимости при одиночном возбуждении: А —потен­
циал действия; Б —динамика возбудимости (V —напряжение на внутренней
поверхности мембраны; t —время; ПП —потенциал покоя; КД критическая
деполяризация; Р —раздражение; ПС —предспайк; С —спайк; ОСП —отри­
цательный следовой потенциал; ПСП —положительный следовой потенциал;
В —возбудимость; СуперВ —суперномальная возбудимость; АР —абсолютная
рефрактерность; ОР — относительная рефрактерность; Э — экзальтация;
СубВ —субнормальная возбудимость)
Во время отрицательного следового потенциала разница между
уровнем заряда мембраны и уровнем критической деполяризации
меньше, чем в состоянии покоя. Порог раздраж ения такж е меньше, а
возбудимость больше. Наступает стадия экзальтации, имеющая сам о­
стоятельное значение в ряде ф изиологических процессов,
Во время положительного следового потенциала разница между
уровнем заряда мембраны и уровнем критической деполяризации
увеличивается, порог раздраж ения увеличивается, и возбудимость
уменьшается. Н аступает период субнормальной возбудимости или
вторичной относительной рефрактерности.
30
Глава 1
1.5. ТОРМОЖЕНИЕ
Торможение — активны й биологический процесс, который пре­
пятствует развитию возбуж дения, либо прекращ ает уже развивш ийся
процесс возбуждения.
Тормозные влияния нервной системы на деятельность сердца при раз­
дражении блуждающих нервов открыли в 1847 г. братья А. и Э. Веберы.
Это пример периферического торможения. Торможение возбуждения
происходит также в центральной нервной системе. Такое торможение
называется центральным. Первый эксперимент по исследованию цент­
рального торможения был описан И.М. Сеченовым в 1862 г.
Торможение — временный процесс, регулирующий уровень возбуж­
дения. В центральной нервной системе и внутренних органах процессы
возбуждения и торможения постоянно сменяют друг друга. Торможение
может возникать как самостоятельный функциональный процесс или
может быть результатом взаимодействия двух возбуждений.
По элетрофизиологической природе различают гиперполяризационное, стойкое деполяризационное и стабилизационное торможение.
Гиперполяризация возникает на постсинаптических мембранах при
действии тормозных медиаторов и приводит к уменьш ению возбуди­
мости ткани. Стойкая деполяризация возникает при высокой частоте
возбуждения, что приводит к накоплению возбуждающего медиатора,
которы й препятствует возвращ ению потенциала постсинаптической мембраны в состояние покоя. При этом увеличивается разница
между зарядом мембраны и уровнем критической деполяризации.
Процесс сходен с явлением парабиоза или аккомодации. Стойкая
деполяризация может также возникать при действии тормозных меди­
аторов за счет специфических свойств постсинаптических мембран.
Стабилизация потенциала покоя возникает при действии ряда био­
логически активны х веществ, которые вызываю т блокаду натриевой
проницаем ости клеточных мембран без изменения потенциала покоя.
Это приводит к невозможности возникновения возбуждения.
1.6. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
Вторым общим свойством возбудимых тканей является их способ­
ность проводить возбуждение по клеточным мембранам от места возник­
новения возбуждения. Существует два способа проведения возбуждения:
непрерывный и сальтаторный. При непрерывном способе проведения воз-
31
Основы жизнедеятельности
р
БМ
лт
I
лт
+ + + + + + + - - - - + + + + ++
+ + + +
+ + + +
Т + 4- 4- + + +
-
-
-
Покой
-----------
-
+
+ + + +
Покой
Возбуждение
---------- ►
M
Рис. 1.7. Двустороннее проведение возбуждения в нервных волокнах
(М — миелиновое нервное волокно; БМ — безмиелиновое нервное волок­
но; Р —раздражение; ЛТ —локальный ток)
буждения оно проводится по всем участкам мембраны. Это происходит
за счет местных токов, возникающих между возбуждённым участком и
невозбуждёнными участками мембраны. Местные токи вызывают час­
тичную деполяризацию невозбужденных участков мембраны, уровень
заряда мембраны достигает критического уровня, открываются все поры
для ионов Na+, и возникает потенциал действия. Между вновь возбуж­
денным участком и последующими невозбужденными участками мемб­
раны возникают местные токи, за счёт которых возбуждение продолжает
распространяться далее по мембране (рис. 1.7). Непрерывное проведение
происходит в безмиелиновых нервах и всех видах мышечной ткани.
Сальтаторный или прыж ковый способ проведения возбуж дения
существует в миелиновы х нервах. Возбуждение распространяется по
перехватам Ранвье такж е за счёт местных токов. При этом возбуж­
дение захваты вает до 3—5 последующих невозбуж денны х перехватов.
Скорость распространения возбуж дения при этом выше, чем при
непрерывном проведении возбуж дения.
32
Глава 1
Законы проведения возбуждения в нервах
Возбуждение, распространяю щ ееся по нервным волокнам, входя­
щ им в нервы, подчиняется определенным закономерностям.
Первый закон, «Морфологической и функциональной непрерывности
нервов», г о в о р и т е необходимости сохранности морф оф ун ки и онального состоян ия неовя-для подведен и я возбуж дения. Возбуж дение не
проводится через место перерезки нерва или через участок нерва с
наруш енны ми ф ун кц и ям и мембраны при сохранении его анатом и­
ческой целостности. Если блокируемый участок нервного волокна
меньше расстояния между 3—5 перехватами Ранвье, то возбуждение
может перепры гнуть через этот участок.
Второй закон^ «Двустороннего проведения возбуждения в нервах»,
говорит о том, что при нанесении раздраж ения возбуж дение в нерве
распространяется в л б е стороны .
Третий закон. «Изолированного проведения возбуждения в нервах»,
говорит о том, что нервные волокна, входящие в нерв, изолированы
друг от друга за счет миелиновы х и соединительнотканны х оболочек,
и возбуждение не распространяется между нервны ми волокнами,
располагаю щ имися пялом.
Четвертый закон. «Относительной неутомляемости нервов», отражает способность нервов длительное время проводить возбуждение
п р и и х ритмическом раздраж ен ии . С инапсы и м ы ш цы воспроизводят
ритмическое раздраж ение значительно более короткое время.
Функциональная классиф икация нервных волокон
Скорость проведения возбуж дения в нервных волокнах зависит
от н ал и чи я или отсутствия в них м иелина и от диаметра нервного
волокна. М иели н овые волокна проводят возбуждение быстрее, чем
безмиелиновые, а толстые — быстрее, чем то н к ие.
Суммарны й потенциал действия нерва вблизи места раздраж ения
имеет синусоидальную форму, а вдали от места раздраж ения — слож­
ную форму. Это связано с неодновременны м приходом возбуж дения
в конечную точку нерва по различны м нервны м волокнам, в которых
выделяю т группы А а, А(3, Ay, А5, В и С (рисГТ.8).
Быстрее всего, (со скоростью порядка 100 м/с) возбуждение проводится в волокнах типа А а и медленнее всего, со скоростью 1м/с, в
волокнах С. Волокна Асх — самые толстые, около Ю мкм в диаметре,
а волокна С — самые тон ки е, около 1 мкм в диаметре. Волокна А и
В миелиновые. а волокна С — безмиелиновые. Волокна А а образуют
афф ерентные волокна от мыш ечных проприорецепторов в централь-
Основы жизнедеятельности
33
Нерв
Р
Рис. 1.8. Изменение формы суммарного потенциала действия нерва за счет
разной скорости проведения возбуждения по разным группам нервных
волокон (Р —раздражение; Аа, Ар, Ay, Аб, В, С - группы нервных волокон)
пую нервную систему (ЦНС) и эфферентные соматические нервы,
идущие из ЦНС к скелетным мышцам. Волокна Ар образуют аф ф е­
рентные волокна от рецепторов прикосновения кожи в ЦНС. Волокна
Ау образуют эфф ерентные волокна к мыш ечным веретенам. Волокна
А5 являю тся преимущ ественно афф ерентны ми соматическими волок­
нами, идущ ими от температурных и болевых рецепторов кожи в ЦНС.
При проведении возбуждения по этим волокнам возникает первич­
ный эпикритический компонент боли. Волокна В и С входят в состав
вегетативной нервной системы: В — преганглионарные, С — постганI шонарные. Волокна С также передают афферентные возбуждения от
внутренних органов в ЦНС. По ним передается возбуждение, форми­
рующее вторичный протопатический компонент боли.
1.7. СИНАПТИЧЕСКАЯ ПЕРЕДАЧА ВОЗБУЖДЕНИЯ
В возбудимых тканях возбуждение проводится с одних клеток
на другие через специальны е контакты между клетками: эфапсы ,
34
Глава 1
синапсы и синапсы на расстоянии. Эфапсы, электрические синапсы
или нексусы — щелевые контакты между клеткам и. Ш ирина щели в
электрических синапсах в 10 раз меньше, чем в химических. В них
возбуждение проводится электрическим способом за счет местных
ионны х потоков между возбужденной пресинаптической и невоз­
бужденной постсинаптической мембраной. М еханизм проведения
возбуж дения п рин ц ип и ально не отличается от механизма проведе­
ния возбуждения между перехватами Ранвье миелинового нервного
волокна. Передача возбуж дения посредством щелевых контактов
осущ ествляется в миокарде, гладкой мускулатуре и частично в ЦНС.
Возбуждение в эф апсах может проходить в обе стороны.
В синапсах проведение возбуж дения осущ ествляется между двумя
клеткам и с помощью химических веществ, назы ваемы х медиаторами.
С инаптический способ проведения возбуж дения — основной в ган ­
глиях нервной системы, между нейронами ЦНС, в нейроорганны х
синапсах и нервно-мы ш ечны х синапсах.
В синапсах на расстоянии медиатор одной клетки за счёт дифф узии
возбуждает одновременно постсинаптические мембраны нескольких
клеток. Это происходит в ганглиях симпатической нервной системы,
в которых каждое преганглионарное нервное волокно возбуждает
около 20 ганглионарны х нейронов, а такж е в нейроорганны х си н ап ­
сах вегетативной нервной системы.
Ф ункции медиаторов
К аж дая нервная клетка синтезирует только один медиатор. Это
свойство назы вается принципом Дейла.
В синапсах периферической нервной системы имеется два медиато­
ра. Ацетилхолин выделяется в нервно-мыш ечных синапсах соматичес­
кой нервной системы, ганглионарных синапсах вегетативной нервной
системы, нейроорганных синапсах парасимпатической нервной систе­
мы и нейроорганных синапсах симпатической нервной системы в пото­
вых железах и кровеносных сосудах скелетных мышц. Норадреналин
выделяется в основной части нейроорганных синапсов симпатической
нервной системы. В синапсах соматической нервной системы ацетил­
холин является возбуждающим медиатором. В нейроорганных синап­
сах вегетативной нервной системы ацетилхолин и норадреналин могут
вызывать и возбуждение, и торможение постсинаптических мембран,
что зависит от свойств постсинаптических рецепторов.
Кроме основного медиатора, п^есинаптические окончания выде­
ляю т комедиаторы, которые настраиваю т чувствительность постси-
Основы жизнедеятельности
35
маптических рецепторов к основному медиатору. В адренергических
синапсах комедиаторами являю тся доф ам ин, АТФ, гонадотропин и
пептид Y. В холинергических синапсах комедиатором является вазоинтестинальны й полипептид.
В синапсах центральной нервной системы имеется множество
медиаторов. Главны м возбуж даю щ им медиатором явл яется глутамат, с помощью которого в ЦНС передается 80% инф орм ации.
А цетилхолин, норадреналин, доф ам и н и серотонин настраиваю т
возбудимость нейронов, определяя уровень бодрствования орган и з­
ма и его м отивационно-эм оциональны й фон.
Гам м а-ам ином асляная кислота (ГАМК) и глицин являю тся тор­
мозными медиаторами, избирательно и временно блокирую щ им и
возбуждение в нервных центрах при различны х ф ункциональны х
состояниях организма.
А цетилхолин поддерживает тонус нервных центров как при бодр­
ствовании, так и во время сна. Н орадреналин дополнительно ак ти ­
вирует нервные центры при напряж ении организма. Д оф ам ин спо­
собствует регуляции движ ений и мы ш ечного тонуса. С еротонин
сдерживает возбуждение нервны х центров в необходимых пределах.
Ряд биологически активны х веществ обладает модулирующ им
действием на основные медиаторы. Эти вещества называю тся нейро­
модуляторы. Гистамин повышает чувствительность нервных центров
к основным медиаторам. Вещество Р разнообразно модулирует дейс­
твие основных медиаторов и принимает прямое участие в проведении
болевого и эмоционального возбуждения. Э нкеф алины и эндорфины
тормозят выделение основных медиаторов при болевом возбуждении.
Классиф икация синапсов
В соответствии с морф ологической кл асси ф и кац и ей синапсы
подразделяю т на нейрональны е (центральны е и ганглионарны е) и
нейроорганные. В нейрональны х синапсах выделяют аксо-сом атические, аксо-дендритические, аксо-аксональны е и дендро-дендритические. В нейроорганны х синапсах различаю т нейро-мускулярны е
и нейро-железистые. В соответствии с физиологической класси ф и ­
кацией имеются возбуждаю щие и тормозные синапсы. В соответс­
твии с фармакологической классиф икацией в зависимости от вида
медиатора выделяю т холин-, адрен-, ГАМК-, серотонин-, доф ам ин,
глутаматергические синапсы. Х олинергические синапсы делятся на
Н-холинергические и М-холинергические. В обоих видах синапсов
возбуждение проводится с помощью ацетилхолина. Часть холинер-
36
Глава 1
гических рецепторов на постсинаптической мембране возбуждается
при действии на них табачного алкалоида никотин а в малых кон­
центрациях, а другая часть возбуждается при действии яда некото­
рых видов ракообразны х и грибов — мускарина. Первые буквы слов
«никотин» и «мускарин» дали обозначение холинергическим си н ап ­
сам. Н -холинергические рецепторы находятся в нервно-мы ш ечны х
синапсах соматической нервной системы, в ганглионарны х синапсах
вегетативной нервной системы и в нейроорганны х холинергических синапсах симпатической нервной системы. М -холинергические
рецепторы находятся в нейроорганны х синапсах парасимпатической
нервной системы. А дренергические рецепторы находятся в нейроор­
ганны х синапсах симпатической нервной системы, и в зависимости
от места локализации делятся на а-, (Зх-, Р2-адренергические. В ЦНС
имею тся все виды постсинаптических рецепторов.
М еханизм проведения возбуждения в синапсах
В синапсе различаю т пресинаптическую мембрану, синаптическую
щель и постсинаптическую мембрану. Часть медиатора синтезируется
в теле нервной клетки, и затем с помощью аксонного транспорта со
скоростью около 40см /сут попадает в пресинаптическое окончание.
Другая часть медиатора синтезируется непосредственно в пресинаптическом окончании. Порции медиатора окружаются мембранами и в
виде везикул хранятся в пресинаптическом окончании. При проведе­
нии возбуждения увеличивается проницаемость мембраны пресинаптического окончания для ионов Са2+ (рис. 1.9.А). Ионы Са2+ входят в
Рис. 1.9. Проведение возбуждения через возбуждающий синапс и торможение
проведения возбуждения в тормозном синапсе: А — проведение возбуждения
по аксону и увеличение проницаемости мембраны для ионов Са2+; Б — Са2+
вызывает движение везикул к пресинаптической мембране. Медиатор выхо­
дит в синаптическую щель; В — медиатор присоединяется к рецепторам
постсинаптической мембраны и возникает возбуждающий или тормозной
постсинаптический потенциал (ВПСП или ТПСП); Г — локальные токи спо­
собствуют возникновению потенциала действия (ПД) в соседних с постсина­
птической мембраной участках, и возбуждение проводится в возбуждающем
синапсе. В тормозном синапсе возбуждение не проводится. Медиатор расщеп­
ляется ферментативно, и большая часть продуктов его расщепления активно
всасывается обратно в пресинаптическое окончание. Пресинаптические
рецепторы оценивают количество выделившегося медиатора
Основы жизнедеятельности
37
Возбуждающий синапс
Аксоплазматический
Потеря
части
медиатора
Проведение
Пресинаптические
рецепторы
Проведение
возбуждения
Тормозной синапс
38
Глава 1
пресинаптическое окончание и присоединяю тся к везикулам, обеспе­
чивая их движение к пресинаптической мебране и слияние с ней (см.
рис. 1.9Б). Медиатор выходит в синаптическую щель, где с помощью
диффузии пассивно проходит к постсинаптической мембране.
В возбуждающих синапсах медиатор действует на постсинаптические рецепторы, образованные белково-липидны ми молекулами, что
приводит к откры тию пор для ионов N a+ и деполяризации постсина­
птической мембраны. При этом на мембране возникает возбуждающий
постсинаптический потенциал (ВПСП) (см. рис. 1.9В). Каждая порция
медиатора в одной везикуле называется квантом медиатора, который
изменяет заряд постсинаптической мембраны на 0,1 мВ. Величина и
длительность ВПСП зависят от количества медиатора и составляю т
1-50 мВ и 10-100 мс. За счёт местных токов ВПСП вызывает потенциал
действия в соседних участках поверхностной мембраны клетки, кото­
рый далее проводится по возбудимой клетке (см. рис. 1.9Г).
Подействовавш ий на постсинаптическую мебрану медиатор раз­
рушается ферментами. Продукты его распада всасываю тся в кровь
или закачиваю тся в пресинаптическое окончание, где происходит
синтез новых порций медиатора. А цетилхолин расщ епляется холинэстеразой, а норадреналин расщ епляется последовательно действу­
ю щ ими ферм ентами котехоламиоксиметилтрансферазой (КОМ Т) и
м оноаминоксидазой (МАО). Тормозные медиаторы увеличиваю т п ро­
ницаемость постсинаптической мембраны для ионов К+ и С1~. Ионы
К+ выходят, а ионы С Г входят в клетку по градиенту концентрации,
в результате чего возникает гиперполяризация мебраны , и на ней
формируется тормозной постсинаптический потенциал (ТП СП) (см.
рис. 1.9В). Порог раздраж ения увеличивается, возбудимость падает, и
возбуждение не проводится, что и характеризует состояние торможе­
ния в синапсах (см. рис. 1.9Г). Так действуют ГАМК и глицин в ЦНС.
Также действуют ацетилхолин и норадреналин в тормозных синапсах
вегетативной нервной системы.
Функциональные свойства синапсов
• В синапсах осущ ествляется одностороннее проведение возбужде­
ния от пре- к постсинаптической мембране.
• За счет диф ф узии медиатора в синаптической щели происходит
синаптическая задержка проведения возбуждения.
• Н аличие постсинаптических рецепторов обусловливает высокую
химическую чувствительность синапсов к биологически активны м
веществам.
39
Основы жизнедеятельности
• П остсинаптические потенциалы в синапсе не подчиняю тся зак о ­
ну «Все или ничего» и способны к суммации. А мплитуда п остсина­
птических потенциалов зависит от количества действующего на
постсинаптическую мембрану медиатора.
• С инапсы обладают повышенной утомляемостью по сравнению с
утомляемостью нервов и м ы ш ц при их длительном ритмическом
раздраж ении. Утомляемость связана с истощ ением запасов меди­
атора в синапсах.
• С инапсы имеют низкую лабильность по сравнению с лабильн ос­
тью нервов и мыш ц. Л абильность или ф ункциональная подвиж­
ность характеризуется быстротой появления и исчезновения воз­
буж дения и количественно отраж ает м иним альную длительность
процесса возбуж дения. Мерой лабильности является м аксим аль­
ная частота раздраж ения, которую возбудимая ткань воспроизво­
дит без трансф орм ации ритма. К оличественно мера лабильности
является обратной величиной от длительности возбуж дения или,
точнее, от длительности рефрактерного периода (табл. 1.1).
Таблица 1.1. Величины лабильности нерва, мышцы и синапса
Возбудимое
образование
Нерв миелиновый
Нерв безмиелиновый
Мышца поперечнополосатая
Синапс нервно-мышечный
Время абсолютного реф­ Мера лабильности,
рактерного периода, мс
имп./с
1
2
5
10
1000
500
200
100
1.8. МЫШЕЧНОЕ СОКРАЩЕНИЕ
М орфологически различаю т три вида мыш ечной ткани:
— скелетную, или поперечнополосатую, или исчерченную;
— гладкую, или неисчерченную;
— м иокардиальную поперечнополосатую.
Все мы ш цы обладаю т свойствами возбудимости, проводимости
и сократимости. С ократимость — это свойство мыш ечной ткани
укорачиваться и увеличивать тонус при пороговом и надпороговом
раздражении.
Виды мышечного сокращения. М ы ш ечны е сокращ ени я бываю т
одиночные и тетанические, возникаю щ ие при ритмическом раздра­
40
Глава 1
ж ении. Кроме того, различаю т изометрические, изотонические и см е­
ш анны е (ауксометрические) виды сокращ ений. При изометрическом
сокращ ении увеличивается тонус мы ш цы без изм енения ее длины .
При изотоническом сокращ ении м ы ш ца укорачивается без измене­
ния тонуса. Эти виды сокращ ений могут возникать при определён­
ны х условиях. Чащ е всего происходят см еш анны е сокращ ения с
изменением как длины , так и тонуса мыш цы.
С о к р ащ ен и я скелетн ы х м ы ш ц п одразделяю тся на ф азн ы е и
тонические. Бы страя (фазная) активн ость скелетны х м ы ш ц опреде­
ляется белыми мышечными волокнами, которые отличаю тся значи­
тельны м содерж анием м иоф ибрилл, гликогена и гликолити чески х
ф ерм ентов. Гликоген обесп ечи вает эн ергети чески е потребности
м ы ш цы . С одерж ание м и оглоби н а в белы х м ы ш ечн ы х волокнах
незначительно.
Тонические сок ращ ен и я определяю тся красными мышечными
волокнами. В них содержится большое количество миоглобина и
цитоплазм атических органелл, особенно митохондрий.
Красны е мы ш ечные волокна способны к длительному н ап ряж е­
нию и к более длительной динам ической работе. Белые волокна,
миоф ибриллы которых толщ е и сильнее, быстро сокращ аю тся, но
и легко утомляю тся. Больш инство мыш ц смеш анны е, они в разных
пропорциях содержат белые и красные волокна.
Энергетические процессы мышечного сокращения. Быстрые и мед­
лен н ы е м ы ш ечны е волокн а имею т разн ы й ти п энергетического
обеспечения. Быстрые белые м ы ш ечны е волокна, богатые гл и ко­
геном, работаю т преим ущ ественно на энергетической основе а н а э ­
робного оки слен и я глю козы. О ни быстро утом ляю тся. М едленные
красны е мы ш ечны е волокна, богатые м и тохон дри ям и и миоглобином, использую т при сокращ ени и в основном энергию аэробны х
р еакц и й . О ни хорош о снабж аю тся кровью и могут долго работать
без утом ления.
При интенсивной работе в мы ш цах окисляю тся в основном угле­
воды, а при легкой работе - свободные жирные кислоты.
Функции скелетной мускулатуры. Работа мускулатуры обеспечи­
вает перемещ ения тела и его отдельных частей в пространстве для
удовлетворения ведущ их потребностей организма. Обладая мыш еч­
ны м тонусом, мускулатура обеспечивает равновесие тела в разли ч­
ны х статических полож ениях и при движ ении организма. М ыш цы
увеличиваю т теплопродукцию при сокращ ении и при мышечной
дрожи. С окращ ение мыш ц способствует току лимфы.
Основы жизнедеятельности
41
Свойства скелетных мышц. В скелетной мыш це каждое мы ш еч­
ное волокно подчиняется закону «Все или ничего». Целая м ы ш ца
не подчиняется этому закону, так как каждое мышечное волокно
имеет свой порог раздраж ения и при увеличении силы раздраж ения
возбуждается большее количество мы ш ечны х волокон, а амплитуда
сокращ ения м ы ш цы увеличивается.
В одиночном мышечном сокращении различаю т три периода: латен­
тный, укорочения и расслабления (рис. 1.10). Л атентны й или скры ­
тый период связан с возникновением и проведением возбуж дения по
мышце, которое потом переходит в сокращ ение.
Одиночное мы ш ечное сокращ ение способно к суммации. Если
второе раздраж ение наносят в период расслабления, то расслабление
сменяется вторым сокращ ением, ам плитуда которого больше ам п л и ­
туды первого сокращ ения (см. рис. 1.10А). Если второе раздраж ение
наносят в период укорочения, то укорочение удлиняется и его ам п ­
литуда увеличивается (см. рис. 1.10Б,В). Если второе раздраж ение
наносят в латентны й период, то сокращ ение не возникает, так как
второе раздраж ение попадает в абсолю тны й реф ракторны й период
первого возбуж дения (см. рис. 1.10Г).
При увеличении частоты раздражения возникает слитное или тетаническое сокращение мышцы. Последовательно возникаю т зубчатый
тетанус, гладкий тетанус, оптимум и пессимум (рис. 1.11). Зубчатый тета­
нус возникает, когда каждое последующее раздражение попадает в
период расслабления предыдущего одиночного сокращения. Гладкий
тетанус возникает, когда каждое последующее раздражение попадает в
период укорочения предыдущего одиночного сокращения. Оптимум —
максимальная амплитуда тетанического сокращения мышцы при её
ритмическом раздражении, когда каждое последующее раздражение
попадает в период укорочения предыдущего одиночного сокращ ения и
одновременно в фазу экзальтации предыдущего одиночного возбужде­
ния. Пессимум — расслабление мышцы при ее ритмическом раздраже­
нии с частотой больше меры лабильности, когда каждое последующее
раздражение попадает в латентный период предыдущего одиночного
сокращения и одновременно в рефракторный период предыдущего
одиночного возбуждения.
Т етани ческое сокращ ени е — лабораторн ы й ф еном ен, в о зн и ­
каю щ ий при и скусственном ритм ическом раздраж ен ии м ы ш цы .
В организм е сли тн ое сокращ ени е скелетны х м ы ш ц д ости гается
последовательны м , а не одноврем ен ны м раздраж ением м ы ш еч­
ных волокон. Ч астота возбуж дений, приходящ их по сом атическим
Глава 1
42
Потенциал
Г
Абсолютная
рефрактерность
Одиночное
Основы жизнедеятельности
43
Рис. 1.10. Д инам ика суммации двух одиночных сокращ ений скелет­
ной мышцы: А — второе раздражение попадает в период расслабления;
Б — второе раздражение попадает в период укорочения; В — второе раздра­
жение попадает в начало периода укорочения и экзальтацию; Г — второе
раздражение попадает в латентный период и абсолютную рефрактерность
(ЛП —латентный период; ПУ —период укорочения; ПР —период расслаб­
ления; Рр Р2 —первое и второе раздражение)
нервам к м ы ш цам , обы чно в 10 раз меньш е вели чи ны лабильности
мышцы.
Нейромоторные единицы. Установлено, что каждое отдельное нервное
волокно иннервирует несколько мышечных волокон. Нервное волокно
и иннервируемые им мышечные волокна называются нейромоторной
единицей. В различных мышцах количество мышечных волокон в ней­
ромоторной единице различно. В наружных мышцах глазного яблока их
3 -6 , в пальцах руки — 10—25, а в икроножной мышце ноги — до 7000. Чем
меньшее количество мышечных волокон иннервируется одним нервным
волокном, тем точнее движение, которое обеспечивает мышца. Внутри
пейромоторных единиц сокращение отдельных мышечных волокон про­
исходит последовательно в связи с неодновременностью прихода к ним
возбуждения, что обеспечивает плавность сокращения мышцы.
Рис. 1.11. Слитное сокращение скелетной мышцы при увеличении часто­
ты раздражения (А —амплитуда сокращения; t —время; ОС —одиночные
сокращения; ЗТ — зубчатый тетанус; ГТ — гладкий тетанус; Опт — опти­
мум; Пес —пессимум)
44
Глава 1
Каждое мыш ечное волокно иннервируется нескольким и нервны ­
ми волокнам и от разны х мотонейронов. Таким образом, регуляция
сокращ ения мыш ечных волокон осущ ествляется несколькими ней­
ронами центральной нервной системы.
Тонические сокращения скелетных мышц. П роприорецепторы
м ы ш ц, нейроны сп и нн ого мозга и нервные центры экстрапи рам идной системы головного мозга обеспечиваю т тонические сокра­
щ ения скелетных мы ш ц, направленны е на сохранение равновесия
тела в пространстве за счет поддерж ания определенных поз тела.
М оторные единицы экстрапирам идной системы вклю чаю т до 2000—
3000 мыш ечных волокон.
Мышечное утомление. При длительном действии на мы ш цу частых
раздраж ений наблюдается постепенное удлинение всех фаз одиноч­
ных мыш ечных сокращ ений. С ила сокращ ения мы ш цы уменьш ает­
ся. Развивается утомление с отсутствием сокращ ения м ы ш цы в ответ
на раздражение. При этом, несмотря на отсутствие сокращ ений, в
мы ш це все еще регистрирую тся лиопотенциалы , которые не п риво­
дят к сокращ ению . Утомление связано с накоплением в мыш це угле­
кислого газа и молочной кислоты, уменьш ением количества ионов
Са2+ в эндоплазматическом ретикулуме и Т-системах, уменьш ением
запасов гликогена, глюкозы и АТФ.
В целом организме утомляю тся сначала нервные центры, затем
нервно-мы ш ечны е синапсы , и только после этого происходит утом­
ление мы ш ечны х волокон.
Теплообразование в мышце. В м ы ш це энергия хим и чески х реак­
ц и й превращ ается в м еханическую энергию м ы ш ечного со к р а­
щ ения. При этом мы ш цы выделяю т в окруж аю щ ую среду часть
энергии в виде тепла.
В покоящ ейся мы ш це происходит обусловленное метаболически­
ми процессами теплообразование покоя. При сокращ ении мыш цы
теплообразование усиливается. Оно назы вается начальны м теп ло­
образованием сокращ ения. При расслаблении в мы ш це происходит
конечное теплообразование сокращ ения. В еличины начального и
конечного теплообразования сокращ ения равны между собой.
М еханизм сокращ ения в скелетных мышцах и миокарде
М ышечное волокно состоит из миофибрилл, каж дая из которых
разделена Z -дисками на саркомеры. К мембранам прикреплены нити
белка актина. На поверхности актин а расположены тонкие нити
тропомиозина, заканчиваю щ иеся тропонином. Между актином нахо­
45
Основы жизнедеятельности
дятся нити белка миозина, головки выростов которых контактирую т
с тропонином на поверхности актин а (рис. 1.12, рис. 1.13А).
При проведении возбуждения по мышечному волокну изменяет­
ся проницаемость поверхностной мембраны и саркоплазматического
ретикулума. Внутрь саркомеров устремляются ионы Са2+. В скелетных
мышцах ионы Са2+ выходят из саркоплазматического ретикулума, а в
миокарде — из ретикулума и Т-систем. Эти ионы электростатически
взаимодействуют с тропонином, что обеспечивает сдвиг его в сторо­
ну. Головки миозина, в которых находится АТФ, вступают в контакт
с актином, и фермент АТФ -аза расщ епляет АТФ. АТФ превращается
в АДФ с выделением энергии, и АДФ выходит из головки миозина в
цитоплазму (см. рис. 1.13Б). Энергия АТФ используется на изменение
четвертичной структуры головок миозина. Это напоминает гребковые
Z-диск
Саркомер
Z -диск
Рис. 1.12. Изменение длины саркомера при мышечном сокращении:
А — при расслаблении; Б — при сокращении
46
Глава 1
Z-диск
Актин
АТФ-аза
неактивная
=
X
Тропонин
_
АТФ
г
Миозин
,
Са2 _
АТФ-аза
активная
Возбуждение
-Щ ПГ1
/ T v Фосфат
' 0"’
-/
Са^
Укорочение саркомера
......... j Ql
гп
Митохондрия
4
Окислительное
фосфорилирование
IS jr * -
Основы жизнедеятельности
47
движения весел на лодке. При этом миозин остаётся на месте, а нити
актина сдвигаются к центру саркомера, и дли н а саркомера уменьш а­
ется на незначительную величину (см. рис. 1.13В). С помощью окиспительного ф осф орилирования АДФ превращается в митохондриях в
ЛТФ и снова встраивается в головки м иозина (см. рис. 1.13Г). При этом
головки миозина теряю т связь с актином и выпрямляю тся без затраты
жергии. Головки миозина вступают в контакт со следующими моле­
кулами тропонина на поверхности актина (см. рис. 1.13Д). С помощью
ионов С а2+ происходит очередной ц икл уменьш ения длины саркомера.
5а счёт уменьш ения длины всех саркомеров миофибрилл происходит
сокращение мышечного волокна и всей мыш цы.
Места присоединения головок выростов м иозина к актину назы ­
вают поперечны ми мостиками. Количество поперечных мостиков
между актином и м иозином и количество циклов уменьш ения дли н ы
саркомера определяет силу мыш ечного сокращ ения (рис. 1.13).
Образующееся после смерти трупное окоченение связано с исполь­
зованием запасенной при ж изни АТФ на сокращ ение скелетной муску­
латуры. Отсутствие в мышечной ткани окислительного ф осф орилиро­
вания и образования новых молекул АТФ приводит к невозможности
головок миозина выйти из контакта с актином, что сопровождается
отсутствием расслабления мышц. Трупное окоченение исчезает только
после начала некротических процессов в мышечной ткани.
Закон средних нагрузок
С ила сокращ ения зависит от степени растяж ения мы ш цы перед ее
сокращ ением. Н аибольш ая ам плитуда сокращ ения достигается при
средней нагрузке (рис. 1.14). При этом между нитям и актин а и м иози­
на возможно возникновение максимального количества поперечны х
мостиков (см. рис. 1.14Ба). При сильной нагрузке дли н а саркомеров
Рис. 1.13. Изменение положения поперечного мостика между актином и
миозином в динамике мышечного сокращения: А — исходное положение
во время покоя мышцы; Б — выход ионов Са2+ при возбуждении мышцы
и сдвиг тропонина в сторону; В — за счет энергии АТФ головка выроста
миозина поворачивается под новым углом; Г — Са2+ активно закачивается
в Т-трубочки и саркоплазматичеекий ретикулум. Тропонин возвращается
на исходное место. Новые молекулы АТФ заполняют головку миозина;
Д — головка выроста миозина выпрямляется и контактирует со следующей
молекулой тропонина на актине
48
Глава 1
увеличивается, и количество поперечных мостиков между нитями
акти н а и м иозина уменьш ается, что приводит к меньшей силе сокра­
щ ения (см. рис. 1.14В). При отсутствии нагрузки количество попереч­
ных мостиков между актином и миозином такж е уменьш ается из-за
того, что нити актин а наползаю т друг на друга при уменьш ении
дли н ы саркомера (см. рис. 1.14АБ).
Особенности сокращ ения гладкой мышечной ткани
Гладкая мы ш ечная ткань обладает низкой возбудимостью, д ли ­
тельным возбуждением, малой скоростью проведения возбуждения и
самой низкой лабильностью. Одиночное сокращ ение возникает мед­
ленно, длительно продолжается и достигает значительной силы. Эти
свойства используются в организме для длительного поддержания
тонуса стенок и сфинктеров полых органов. Больш ая сила сокращ е­
н ия необходима для выполнения запирательной ф ун кц ии сф и н кте­
ров таких органов, как желудок, ж елчный пузырь, мочевой пузырь.
Гладкие мы ш цы обладают автоматизмом, то есть способностью
самопроизвольно возбуждаться без внеш них раздраж ений. В связи с
этим тонус и сокращ ение гладких мы ш ц сохраняю тся даже в случае
разруш ения у ж ивотны х головного и спинного мозга.
Гладкие мы ш цы не подчиняю тся закону изолированного проведе­
ния возбуж дения. За счёт щелевых контактов (нексусов) возбуждение
распространяется с одного мыш ечного волокна на соседние волокна.
Поэтому гладкая м ы ш ца подчиняется закону «Все или ничего», в
отличие от скелетной мы ш цы , которая ему не подчиняется.
Гладкие мы ш цы обладают пластическим тонусом или пластич­
ностью, то есть способностью сохранять приданную медленным
растяж ением длину. Это свойство необходимо д л я неизм енности
тонуса гладких мы ш ц полых органов, таких, как желудок, желчный
пузырь, мочевой пузырь, при их медленном наполнении. В отличие
от гладкой, скелетная мускулатура обладает упругостью, то есть спо­
собностью возвращ аться к исходной длине после растяж ения.
Одним из адекватны х раздраж ений является быстрое растяжение
мы ш цы . Это приводит к тому, что при быстром наполнении полых
органов гладкие мы ш цы , выстилаю щ ие их стенки, начинаю т сокра­
щаться, вызывая опорожнение этих органов.
Гладкие мы ш цы , в отличие от скелетных мы ш ц, обладаю т повы­
ш енной химической чувствительностью. С келетны е мы ш цы сокра­
щ аю тся под влияни ем только нервны х возбуж дений, а гладкие
м ы ш цы сокращ аю тся за счёт собственного автоматизма, действия
Основы жизнедеятельности
49
Рис. 1.14. Одиночные сокращения скелетной мышцы при предварительном
растяжении ее разной силой: А — при сокращении мышцы без нагрузки сила
мышечного сокращения ниже максимальной, так как нити актина мешают
друг другу контактировать с миозином; Б — при средней нагрузке максималь­
ная сила сокращения обусловлена максимальным количеством поперечных
мостиков между актином и миозином; В — при максимальной нагрузке сила
мышечного сокращения уменьшается за счет снижения количества попе­
речных мостиков между актином и миозином; а — количество контактов
между актином и миозином при изменении длины саркомера; б — амплитуда
сокращения мышцы при изменении нагрузки на мышцу; (Р — раздражение;
Лс —амплитуда сокращения; t —время; Ак — актин; М — миозин)
Глава 1
50
вегетативной нервной системы и под влиянием биологически ак ти в ­
ных веществ, в частности, гормонов.
1.9. СЕКРЕЦИЯ
С екреция — процесс образования в клетках хим ических продук­
тов с последующим выделением этого продукта, назы ваемого сек­
ретом, из клетки. С екреторны е клетки выделяю т самые различны е
по структуре и свойствам вещества: белки, гликопротеиды , липиды ,
ферменты, растворы кислот и оснований, соли и воду.
Различаю т три вида веществ, выделяемых клеткам и. Секреты —
продукты клеточного анаболизма. Их синтез и выделение являю тся
основной ф ункцией секреторной клетки. Экскреты — конечные п ро­
дукты клеточного катаболизма и, как правило, подлежат выделению
из организма. Рекреты — вещества, которые сначала поглощаются
клеткой, а затем выделяю тся ею в неизменном виде.
Процессы секреции начинаю тся с поступления в секреторную
клетку из кровеносного русла предш ественников секретируемого
вещества. В клетке происходит синтез секрета, транспорт его по
цитоплазме, окончательное созревание секрета, накопление и хра­
нение секрета. Процессы секреции заверш аю тся выделением секрета
(экзоцитозом) из секреторной клетки.
Выделение секрета происходит разли чн ы м и способами (рис. 1.15).
При мерокриновой секреции молекулы секрета окруж аю тся мембра­
ной, и в цитоплазм е секреторной клетки образую тся пузырьки или
везикулы , наполненны е секретом. Везикулы перемещ аются к клеточ­
ной мембране, сливаю тся с ней, и порции секрета выходят наружу
клетки. Такой способ выделения секрета назы вается экзоцитозом
(рис. 1.15А). М ерокриновая секрец и я происходит в эн докри н ны х
железах и железах ж елудочно-киш ечного тракта.
Второй способ выделения секрета назы вается апокриновым. От
секреторной клетки отделяется ап и кальная часть, в которой нахо­
дится созревш ий секрет (см. рис. 1.15Б). Это происходит в клетках
потовых и молочных желез.
Третий вид секреции назы вается голокриновым. Такая секреция
происходит в сальны х железах, в которых каж дая клетка полностью
разруш ается (см. рис. 1.15В).
Различаю т внутренню ю и внешнюю секреции. При внутренней
секреции секрет выделяется во внутренню ю среду организм а —
Основы жизнедеятельности
51
Рис. 1.15. Виды секреции железами: А —мерокриновая секреция; Б —апок­
риновая секреция; В —голокриновая секреция
кровь, тканевую ж идкость или лимфу. Этот вид секреции назы вается
эндокринной. При внешней секреции секрет выделяется на поверх­
ность слизистой оболочки, кожи или в просвет пищ еварительного
тракта. Этот вид секреции назы вается экзокринной.
Биоэлектрические процессы в секреторных клетках. Возбуждение
секреторных клеток сопровождается секреторны м и потенциалам и.
Возбуждение одних секреторны х клеток связано с их деполяризаци­
ей, а других — с гиперполяризацией.
Базальная и ап и кал ьн ая мембраны секреторны х клеток характери­
зуются разной степенью поляризации. За счет разности потенциалов
этих мембран создается электрическое поле напряж ением 20—30 мВ,
которое удваивается при возбуж дении клетки. Электрическое поле
способствует перемещ ению зрелых гранул секрета к апикальном у
полюсу секреторной клетки, взаимодействию их с мембраной и вы хо­
ду их из клетки путем дифф узии и экзоцитоза.
1.10. РЕЦЕПЦИЯ
Рецепция — процесс восприятия и преобразования механической,
термической, электромагнитной и химической энергии в сложную после­
довательность мембранных и цитоплазматических процессов, порождаю­
щих нервные сигналы. Существует клеточная и сенсорная рецепция.
52
Глава 1
Клеточная рецепция. Роль клеточны х рецепторов вы полняю т п ро­
низы ваю щ ие мембраны клетки белковые молекулы. Белковые рецеп­
торы мембран по п ри н ц и п у «ключ-замок» связы ваю т соответствую ­
щие их молекулярной структуре вещества, называемые лигандами.
Взаимодействия рецепторов с лигандам и через активны е центры
связы вания составляю т процессы клеточного узн аван ия веществ.
Рецепторы локализую тся не только на поверхности, но и внутри
клетки — в цитоплазм е и ядре. Белки рецепторов постоянно си н ­
тезирую тся в ядры ш ках и после взаимодействий с транспортной и
инф орм ационной рибонуклеиновой кислотой (РН К ) поступаю т в
цитоплазму. Д альнейш ее созревание, ф орм ирование и транспорт
клеточны х рецепторов происходит в эндоплазматической сети с пог­
лощ ением энергии АТФ.
После связы вания лиганда с рецептором откры ваю тся или закры ­
ваются ионны е каналы клеточной мембраны, активирую тся мемб­
ранны е или цитоплазм атические ферменты.
Рецепторы для веществ белковой и пептидной природы распо­
ложены на наружной поверхности клеточны х мембран. Рецепторы
стероидны х и тиреоидны х гормонов расположены в цитоплазм е и
ядре клеток.
Сенсорная рецепция происходит в рец еп торн ы х клетках.
Раздражение сенсорны х рецепторов в ж ивы х организмах осущ ест­
вляется так и м и естественны м и раздраж ителям и , к ак свет, звук,
давление, температура, кислотность среды, осмотическое давление,
электром агнитны е волны . С енсорная рецепция осущ ествляется с
помощью специальны х находящ ихся в мембранах рецепторных к л е­
ток белков, взаимодействие которых с соответствую щ ими раздраж и­
телям и приводит к изменению ионной проводимости рецепторной
мембраны и возникновению локального рецепторного потенциала.
Рецепторные потенциалы вызывают увеличение возбудимости до
критического уровня, что приводит к возникновению потенциалов
действия, распространяю щ ихся по аф ф ерентны м нервам в ЦНС.
Глава 2
ОБЩИЕ ПРИНЦИПЫ ПОСТРОЕНИЯ
ЦЕЛОГО ОРГАНИЗМА
Общие принципы построения организма — корреляция, регуляция,
рефлекторные реакции, самоорганизация и саморегуляция — отрабо­
таны ж ивы м и организмами в течение миллионов лет эволю ционного
развития.
2.1. КОРРЕЛЯЦИЯ
Корреляция — взаимодействие элементов в целом организме, при
котором они приобретают новое качество, и в то же время выполняют
присущие только им функции. Коррелятивные связи подразделяют на
механические и химические. Примером механических коррелятивных
связей является взаимодействие работающих сердца и легких, кишечиика и прилегающих органов — печени, желудка и др. Химическая кор­
реляция осуществляется с помощью веществ-«посредников», к которым
относятся медиаторы и другие биологически активные вещества.
Х имическая корреляция, в свою очередь, может быть кон такт­
ной и дистантной. При контактном взаимодействии прилегаю щ ие
участки мембран соседних клеток образуют зоны сли ян и я — щ еле­
вые контакты , допускаю щ ие обмен протоплазмой и биологически
активны м и вещ ествами. Д истантное взаимодействие осущ ествляется
с помощью биогенных аминов, а такж е веществ белковой природы —
гормонов и олигопептидов — через ж и д ки е среды организм а.
Химическая корреляц ия обеспечивает такж е рост нервных волокон
до иннервируемого органа в эмбриогенезе и регенерацию нервов при
повреждении или оперативны х вмеш ательствах на нервных стволах.
2 .2 . РЕГУЛЯЦИЯ
Регуляция представляет собой такую форму взаимодействия в
организме, когда одна структура или процесс направленно подчиняет
себе другую структуру или процесс в интересах целого организма.
Глава 2
54
Регуляция осущ ествляется нервны м, гуморальны м и нейрогуморальн ы м способами.
Н ервная регуляция осущ ествляется с помощью нервной системы.
К ак правило, она происходит быстро.
Гуморальная регуляция реализуется за счет веществ, циркулирующих
в жидкостях организма, — крови, лимфе, цереброспинальной, тканевой и
других жидкостях. К факторам гуморальной регуляции относятся олиго­
пептиды, цитокины, гормоны, иммунные факторы, медиаторы и другие
биологически активные вещества. Как правило, гуморальные регулиру­
ющие влияния осуществляются относительно медленно.
В целом организме гуморальная и нервная регуляция существуют
обычно в форме нейрогуморальной регуляции. Последняя осущ ест­
вляется путем первичного действия гуморальных факторов на нервные
центры, которые вторично по нервным путям распространяю т вл и я­
ние на периферические органы. С другой стороны, на один и тот же
орган могут распространяться одновременно нервные и гуморальные
регулирующие влияни я, из которых первые реализуются быстро и
действуют кратковременно, в то время как гуморальные влияни я осу­
ществляются позднее, но действуют более продолжительно.
2.3. РЕФЛЕКТОРНЫЕ РЕАКЦИИ
Рефлекс — ответная реакц и я организма на различны е воздейс­
твия, осущ ествляемая с помощью нервной системы.
Ф актором, инициирую щ им лю бой реф лекторны й ответ, является
стимул, который может действовать на организм как извне, так и из
внутренней среды.
Представление об отраженной деятельности и ее обусловленности
внеш ними стимулами ввел ф ранцузский естествоиспытатель, ф изик,
математик и философ Рене Декарт (1595-1650).
Термин «рефлекс» предлож ил в 1743 г. ф ран ц узски й врач и ф и л о­
соф А. М онпелье.
«Рефлекторная дуга»
Структурно-функциональную основу рефлекса любой сложности
составляет «рефлекторная дуга», включающая следующие компонен­
ты: рецептор (чувствительные нервные окончания), афферентный путь
(чувствительный нерв), нервный центр, эфферентный путь (двигательный
нерв) и эффектор. В качестве эффекторов выступают мышечная, секре­
Общие принципы построения целого организма
55
торная и нервная ткани со своими специфическими ответами — мышеч­
ным сокращением, секрецией и электрическим ответом (рис. 2.1, А).
Рефлекс проявляется только при целостности всех составляю щ их
компонентов рефлекторной дуги.
Рефлекторная теория
С ущ ность реф лекторной теории составляет распространение идеи
рефлекса на различны е формы ж изнедеятельности.
О сновны м и проводн икам и реф лекторной теории в Западной
Европе были И. П рохазка и М. Галь, а в России — И.М . Сеченов и
И.П. Павлов. Реф лекторная теория твердо поставила физиологические
явления на материальную структурную основу рефлекторной дуги и пред­
ставила неоспоримые экспериментальны е доказательства тончайш ей
организации реф лекторны х процессов.
С трогий детерм инизм рефлекторны х реакц ий такж е характери­
зует их материальную природу. Рефлекторны е реакц ии последова­
тельно разверты ваю тся в причинно-следственны х отнош ениях от
действующих на рецепторы стимулов до ответной реакции организма.
И.М. С еченов распростран ил п ри н ц и п реф лекса на ф ун кц и и
высших отделов мозга, т.е. на психические ф ункции. Он первый
указал на чрезвы чайную изменчивость рефлексов головного мозга.
(' именем И.М. Сеченова связано такж е откры тие центрального тор­
можения рефлекторной деятельности (см. гл. 8).
И.П. Павлов разделил рефлексы головного мозга на две группы, одни
рефлексы он назвал безусловными, вторые — условными. Безусловные
рефлексы, по Павлову, — врожденные; они генетически детерминирова­
ны, возникают при действии адекватных раздражителей на соответству­
ющие рецепторы и проявляются относительно постоянно. К сложным
ирожденным безусловным рефлексам И.П. Павлов отнес пищевые, обо­
ронительные, половые и ориентировочно-исследовательские рефлексы.
Другая форма рефлекторной деятельности — условные рефлексы,
которые приобретаю тся в индивидуальной ж изни ж ивотны х, связа­
ны с обучением и чрезвы чайно изменчивы (см. гл. 8).
В теорию условного рефлекса И.П. Павлов ввел понятие подкреп­
ления как непременного условия образования условного рефлекса.
11одкрепляющими ф акторам и И.П . П авлов считал все формы сложн их безусловных рефлексов, удовлетворяю щ их ж изненно важны е
потребности ж ивотны х.
И.П. Павлов рассматривал процесс образования условного р еф ­
лекса как взаимодействие двух дуг возбуждений: условного и безу-
56
Глава 2
Эфферентное звено
Афферентное
звено
-Ш Ш г
Рецептор
Б
Общие принципы построения целого организма
57
словного рефлексов. Между этим и «дугами», как считал И.П. Павлов,
при повторных сочетаниях образуется «временная связь». Важная
роль подкрепления состоит в том, что при его отсутствии или отмене
временная связь нарушается и условный рефлекс исчезает.
Другое дополнение, сделанное в развитие рефлекторной теории, —
принцип сигнальности. И.П. Павлов показал, что условный раздра­
житель при его действии на организм несет в себе свойства будущего
подкрепляю щ его безусловного воздействия. Условный раздраж итель,
как на это неоднократно указы вал И.П. Павлов, как бы си гн ал и ­
зирует о последующем безусловном рефлексе. Действие условного
раздраж ителя — пример деятельности организма, направленной на
будущее ж изненно важное событие.
Постулаты рефлекторной теории
Развитие идей рефлекса заверш илось следую щ ими, теперь кл ас­
сическими, постулатами рефлекторной теории.
• В ф орм ировании любой формы рефлекторной деятельности веду­
щ ая роль принадлеж ит внеш ним стимулам.
• Возбуждение по рефлекторной дуге распространяется поступа­
тельно от пункта к пункту, от рецепторов к эффекторам.
• Любой рефлекс заканчивается действием, и только действием —
мыш ечным сокращ ением, секреторны м ответом, биоэлектричес­
кой реакцией.
• С пинальны е рефлексы характеризую тся постоянством: они п р о ­
являю тся всегда при действии адекватного раздраж ителя на соот­
ветствующие рецепторы. Рефлексы головного мозга изменчивы.
Реф лекторное кольцо
Представление о рефлекторной дуге претерпело значительную эводюцию. Работы Ч. Белла, А.М. Ф иломафитского, И.М. Сеченова у ка­
зывали на кольцевую организацию рефлекса за счет си гн ализац ии от
Рис. 2.1. Рефлекторная дуга: А — при действии внешнего стимула через
трехчленную рефлекторную дугу формируется отраженное сокращение
мышцы; Б — от проприоцепторов мышцы при ее сокращении поступа­
ет обратная афферентация в центр рефлекторной дуги (четвертое звено
рефлекса). Это свидетельствует о том, что в основе рефлекторного ответа
лежит своеобразное «рефлекторное кольцо»
Глава 2
58
мыш ц. Эти представления были развиты в трудах Н.А. Бернш тейна и
Ф.В. Бассина, которые дополнили классическую рефлекторную дугу
четвертым звеном — аф ф ерентацией от эф ф екторов, в частности от
мыш ечного действия. Вследствие этого рефлекторная дуга тран сф ор­
м ировалась в «рефлекторное кольцо» (см. рис. 2.1, Б).
В настоящ ее время структурная основа рефлекса часто рассмат­
ривается как кольцевая орган и зац ия, в которой эф ф екторы пос­
тоян н о сигнализирую т в соответствую щ ие центры об изменении
своего состоян ия под действием тех или ины х раздраж ителей. Такие
представления, однако, не изм енили сути п они м ани я рефлекса, его
п роявлен ия «от стимула — к действию» и его поступательного, пос­
ледовательного характера разверты вания.
2.4. САМООРГАНИЗАЦИЯ
Отдельные элементы , представляю щ ие исходно хаотическое
состояние, организуются периодически возникаю щ ими потребностя­
ми живых существ в определенные констелляции, обеспечивающие
деятельность, направленную на удовлетворение этих потребностей.
Эти констелляции фиксирую тся при удовлетворении потребности, т.е.
достижении полезных приспособительных результатов. В итоге ф ор­
мируются функциональны е системы, в которых потребный результат
начинает опережающе предвидеться, что оптим альны м путем направ­
ляет живые существа к наиболее успешному удовлетворению их ж и з­
ненно необходимых потребностей (рис. 2.2).
Рис. 2.2. Роль мотивации и подкрепления в динамике формирования
функциональной системы: А — неупорядоченное хаотическое состояние
элементов; Б — под влиянием потребности и формирующейся на ее осно­
ве мотивации возникает ориентировочно-исследовательская реакция,
направленная на поиск факторов, удовлетворяющих исходную потреб­
ность; В — достижение адаптивного результата формирует на структуре
доминирующей мотивации аппарат предвидения свойств его параметров —
акцептор результатов действия (А.Р.Д). В результате складывается дина­
мическая саморегулирующаяся архитектоника функциональной системы;
Г — при очередном формировании аналогичной потребности в ф ункцио­
нальной системе акцептор результатов действия опережающе программи­
рует свойства потребного результата и постоянно оценивает с помощью
непрерывной обратной афферентации параметры его достижения
Общие принципы построения целого организма
О риентировочно­
исследовательская
реакция
Б
П отребность
В
Адаптивный
результат
Ц еленаправленная
деятельность
П отребность
Г
Потребность
60
Глава 2
2.5. САМОРЕГУЛЯЦИЯ
Для нормального существования организма важно, чтобы его мета­
болизм поддерживался на относительно стабильном уровне. Вместе с
тем, обмен веществ и энергии в организме динамичен, связан с потреб­
лением и выделением определенных веществ и тепла. Для стабилиза­
ции обменных процессов в организме существует специальная форма
взаимодействия отдельных элементов — саморегуляция.
Саморегуляция — такая форма взаимодействия элементов в
организме, при которой отклонение той или иной ф ункции от
уровня, обеспечивающего нормальную жизнедеятельность, и пре­
жде всего оптимальный клеточный метаболизм, является причиной
возвращения этой ф ункции к исходному уровню (рис. 2.3). Основы
принципа саморегуляции физиологических функций заложены в
трудах французского физиолога К. Бернара и американского физио­
лога В. Кэннона. К. Бернар сформулировал представления о посто­
янстве внутренней среды как условии «свободной жизни организма»;
В. Кэннон ввел понятие «гомеостазис».
2.6. ФУНКЦИОНАЛЬНЫЕ СИСТЕМЫ
М орфофункциональными аппаратами саморегуляции в организ­
ме являются функциональные системы. Функциональные системы
открыты в живых организмах П.К. Анохиным.
Рис. 2.3. Схема саморегулирующейся функциональной системы, опре­
деляю щ ей полезный для организма приспособительны й результат:
А — в нормальных условиях результат определяет оптимальный уровень
метаболизма; Б — при отклонении результата от уровня, обеспечиваю щ е­
го нормальную жизнедеятельность, активируются рецепторы результата,
усиливается обратная афферентация, поступающая к центру ф ункцио­
нальной системы; В — в результате возбуждения центра функциональ­
ной системы усиливаются эффекторные вегетативные, гормональные и
поведенческие механизмы, что приводит к возвращению полезного при­
способительного результата к оптимальному для метаболизма организма
уровню (1 — полезный приспособительный результат функциональной
системы; 2 —рецепторы результата; 3 —обратная афферентация; 4 —центр
функциональной системы; 5 — метаболизм; 6— внешнее воздействие;
7 — поведение; 8 — вегетатика; 9 — гормоны)
■
62
Глава 2
Под функциональными системами понимают самоорганизующи­
еся и саморегулирующиеся динамические организации, деятельность
всех составных компонентов которых взаимодействует и взаимосодействует достижению полезных для организма в целом приспосо­
бительных результатов.
Приспособительный результат как ведущ ий фактор
образования функциональных систем
Результат действия любой функциональной системы представляет
собой жизненно значимый адаптивный для организма показатель,
обеспечивающий его нормальные отправления в биологическом и
социальном плане. Результат действия выступает в качестве ведущего
фактора организации функциональных систем различного уровня, в
чем состоит его системообразующая роль.
Полезными приспособительными для организма результатами,
формирующими функциональные системы различного уровня орга­
низации, являются следующие.
• М етаболические результаты представлены обширной группой
результатов деятельности многочисленных, главным образом
метаболических, молекулярных процессов в организме, вследс­
твие которых образуются продукты, необходимые для процессов
тканевого метаболизма, и продукты, нарушающие нормаль­
ную жизнедеятельность и подлежащие удалению из организма.
Конечные продукты останавливают или, наоборот, ускоряют
течение метаболических реакций и выступают таким образом в
форме полезных для организма результатов.
• Гомеостатические результаты представлены показателями крови,
тканевой лимфы, межклеточной и внутриклеточной, внутрикишечной, цереброспинальной и других жидкостей, а именно
уровнем осмотического давления, pH, давления крови, темпера­
туры, содержанием питательных веществ, газов, физиологически
активных веществ: гормонов, олигопептидов, иммунных факто­
ров, нейромедиаторов и др. Все эти показатели в совокупности
обеспечивают разные стороны нормального метаболизма тканей
и составляют динамическую внутреннюю среду организма.
• Результаты поведенческой деятельности животных и человека
удовлетворяют их ведущие метаболические и биологические
потребности. Благодаря активным метаболическим процес­
сам во внутренней среде организма постоянно происходит
расходование, потребление или накопление определенных
Общие принципы построения целого организма
63
веществ. Это формирует пищевые, половые, выделительные,
защитные и другие биологические (метаболические) потреб­
ности живых существ, удовлетворение которых часто требует
активного воздействия живых существ на окружающую их
среду. Многочисленные факторы окружающей среды — пища,
вода, комфортная температура, особи противоположного пола,
защитные приспособления, — удовлетворяющие внутренние
потребности живых существ, составляют находящиеся за пре­
делами организма результаты поведенческих функциональных
систем. Результатами поведенческой деятельности являются и
факторы, препятствующие удовлетворению метаболических и
других потребностей организма, которые живые существа в своей
деятельности пытаются избегать или преодолевать.
• Результаты стадной (зоосоциальной) деятельности животных. При
объединении живых существ в сообщества их индивидуальные
потребности подчиняются интересам сообщества. Результаты зоо­
социальной деятельности животных представлены определенными
иерархическими соотношениями в популяциях, общими убежища­
ми, обобщенным удовлетворением пищевых, половых и других пот­
ребностей. Деятельность животных, направленная на достижение
зоосоциальных результатов, нередко приобретает альтруистический
характер в интересах сообщества и, прежде всего, его выживания. В
стадной деятельности у животных нередко наблюдается изменение
характера их индивидуальных биологических потребностей.
• Результаты социальной деятельности человека представлены,
прежде всего, плодами учебной и производственной деятельнос­
ти, бытовой активности, мероприятиями по защите общества,
общением с предметами культуры и искусства и др.
Роль результата деятельности особенно значима в социальноэкономических отношениях, в частности при взаимоотношениях
человека с орудиями производства. В процессе производственной
деятельности человек с помощью различных функциональных сис­
тем метаболического, гомеостатического и поведенческого уровней
достигает социально значимых результатов.
Социальная деятельность человека в значительной степени стро­
ится специальными функциональными системами, определяющими
его психическую, мыслительную деятельность. Результаты функ­
циональных систем человека представлены отражением в сознании
человека жизненно важных понятий, абстрактных представлений о
ппешних предметах и об их отношениях, инструкций, знаний и др.
Глава 2
64
Совокупная деятельность людей направлена на создание обще­
ственного продукта, охрану окружающей среды, мероприятия по
общественной защите и др.
Архитектоника функциональных систем
Ф ункциональные системы любого уровня организации имеют
принципиально однотипную архитектонику (см. рис. 2.3) и включа­
ют следующие общие, универсальные периферические и централь­
ные узловые механизмы:
• полезный приспособительный результат как ведущий показатель
деятельности любой функциональной системы;
• рецепторы результата;
• обратную афферентацию, поступающую от рецепторов результа­
та в центральные образования функциональной системы;
• узловые механизмы центральной архитектоники;
• исполнительные соматические, вегетативные, эндокринные и
иммунные компоненты, включающие организованное целенап­
равленное поведение.
Обратная афферентация
Любое изменение результата контролируется в функциональных
системах соответствующими рецепторами. Афферентная импульсация, возникающая в рецепторах функциональных систем, по аффе­
рентным путям поступает в соответствующие центры. Она называ­
ется «обратная афферентация», так как постоянно сигнализирует о
полезном приспособительном результате каждой функциональной
системы. Под влиянием обратной афферентации в функциональную
систему избирательно вовлекаются центральные и периферичес­
кие исполнительные механизмы, направленные на восстановление
потребного для метаболизма или социальной деятельности чело­
века результата. Обратная афферентация является, таким образом,
той стержневой основой, за счет которой оцениваются все этапы
деятельности функциональной системы по достижению полезного
результата и которая определяет процессы саморегуляции каждой
функциональной системы. С помощью обратной афферентации ЦНС
может регулировать приспособительные реакции целого организма
в соответствии с потребностями организма и условиями окружаю­
щей среды. Наличие звена обратной афферентации делает каждую
функциональную систему циклической саморегулирующейся орга­
низацией.
Общие принципы построения целого организма
65
Изоморфизм функциональных систем
В функциональных системах различного уровня просматривает­
ся однотипный принцип организации, получивший название и зо­
морфизма. В каждой функциональной системе имеются результат
деятельности, обратная афферентация о результате и исполнитель­
ные механизмы. Вместе с тем, в функциональных системах различ­
ного уровня организации имеются некоторые различия.
В функциональных системах метаболического уровня отсутствует
центральная архитектоника.
Большинство функциональных систем гомеостатического уров­
ня представлены только внутренними генетически детерминиро­
ванными механизмами саморегуляции. Полезные для организма
приспособительные результаты деятельности этих функциональных
систем обеспечиваются в основном вегетативными, не контроли­
руемыми произвольно, механизмами. Такие функциональные сис­
темы определяют, например, оптимальный для метаболизма орга­
низма уровень массы крови, форменных элементов, pH, давление
крови.
Отдельные функциональные системы гомеостатического уров­
ня имеют внешнее звено саморегуляции, за счет которого орга­
низм взаимодействует с внешней средой. Ф ункциональная система
дыхания, например, наряду с внутренним вегетативным звеном
саморегуляции имеет относительно пассивное при наличии доста­
точного содержания газов в окружающей среде внешнее звено само­
регуляции, которое обеспечивает поступление воздуха в альвеолы
легких, поглощение кислорода и выделение двуокиси углерода (С 02).
Внешнее звено саморегуляции имеет также функциональная система
выделения.
Разновидность функциональных систем гомеостатического уровня
представляют функциональные системы с активным внешним звеном
саморегуляции, включающим целенаправленное поведение живых
существ во внешней среде, связанное нередко с ее преобразованием.
Как правило, эти функциональные системы строятся на основе внут­
ренних метаболических потребностей организма и обеспечивают их
удовлетворение. Целенаправленная поведенческая деятельность в них
определяется деятельностью головного мозга, включая его высшие
отделы. Примерами являются функциональные системы, обуслов­
ливающие своей деятельностью оптимальный уровень питательных
веществ в организме, осмотическое давление, содержание продуктов
метаболизма, температуру тела и др.
66
Глава 2
В функциональных системах популяционного уровня функцию
центральной архитектоники выполняют специальные, как правило,
лидирующие особи.
В функциональных системах группового и социального уровней
и в функциональных системах психической деятельности челове­
ка результат часто непосредственно не связан с метаболическими
потребностями, а отражает их и может их косвенно обеспечивать.
Такие функциональные системы могут определяться объединенной
деятельностью людей, функциями их головного мозга и производной
от этих функций поведенческой деятельностью, обеспечивающей
достижение тех или иных полезных для людей приспособительных
результатов. Примером такой функциональной системы является
производственная деятельность человека, направленная на получе­
ние социально значимого для него и общества результата, например
сборка определенных деталей, конструирование специальных уст­
ройств, написание книги и др. Иногда результат деятельности фун­
кциональных систем психического уровня не выносится за пределы
мозга, как, например, в случае творческой, мыслительной деятель­
ности человека.
Изоморфна центральная архитектоника различных функциональ­
ных систем (рис. 2.4) см. главу 8.4.
Несмотря на качественные особенности функциональных систем
разного уровня, они, тем не менее, всегда сохраняют изоморфизм
их саморегулирующихся организаций, обеспечивающих различные,
полезные для функциональной системы и для организма в целом,
приспособительные результаты.
Функциональные системы и адаптация организма
к окружающей среде
Функциональные системы поведенческого уровня направлены на
достижение субъектом или сообществом биологических или соци-
Рис. 2.4. Общая схема центральной архитектоники функциональной сис­
темы, формирующей поведенческий акт: АС — афферентный синтез;
М — мотивация; П — память; ОА — обстановочная афферентация;
ПА — пусковая афферентация; А.Р.Д. — акцептор результатов действия;
А — параметры результата соответствуют; Б — динамика рефлекторной
дуги на структурной архитектонике функциональной системы
Общие принципы построения целого организма
Обратная афферентация
АС
Б
Обратная афферентация
67
Глава 2
68
ально значимых результатов во внешней среде. Они также опреде­
ляются исходными потребностями индивидов или сообщества, а
достигнутые результаты деятельности на основе обратной афферентации по принципу саморегуляции постоянно оцениваются с точки
зрения удовлетворения этой потребности.
Окружающая среда при этом органически включается в деятель­
ность отдельных функциональных систем поведенческого уровня
саморегуляции. Только взаимодействие организма с окружающей
средой позволяет этим функциональным системам, и особенно соци­
ального уровня, обеспечивать полезные для организма результаты и
его адаптацию к условиям существования.
Функциональные системы и гомеостазис
Функциональные системы, обусловливающие своими саморегуляторными механизмами относительную устойчивость различных
показателей внутренней среды, представляют конкретные физиоло­
гические аппараты гомеостазиса.
Различные показатели внутренней среды организма, обеспечи­
вающие различные стороны метаболизма, постоянно или периоди­
чески изменяются под воздействием внутренних факторов и ф акто­
ров внешней среды. И только благодаря механизмам саморегуляции
функциональные системы удерживают эти показатели близкими
к определенному, оптимальному для жизнедеятельности, уровню.
Из этого следует, что «константы» в организме условны. Можно
говорить о «жестких константах», которые активно удерживаются
соответствующими функциональными системами у строго опре­
деленного значения и отклонение которых от этого значения при­
водит к необратимым нарушениям метаболизма и смерти. Наряду
с этим имеются «пластичные константы», отклонение которых от
определенного уровня возможно в относительно широком размер­
ном и временном диапазоне без ущерба для здоровья. Примеры
«жестких констант» — уровень осмотического давления, активная
реакция крови, примером «пластичных» служит уровень кровяного
давления, питательных веществ в крови.
Гомеостазис целого организма определяется содружественной
и согласованной саморегулирующейся деятельностью различных
функциональных систем. Поскольку в организме отсутствует абсо­
лютное постоянство внутренней среды, а все его константы дина­
мичны и взаимосвязаны, принято говорить не о гомеостазисе,
а о гомеокинезисе (В.А. Ш идловский).
Общие принципы построения целого организма
69
Динамика деятельности функциональной системы
Отклонение полезного приспособительного результата от уров­
ня, обеспечивающего нормальную жизнедеятельность, немедленно
воспринимается рецепторными аппаратами и посредством нервной
и гуморальной обратной афферентации избирательно мобилизует
центральные элементы функциональных систем. Последние через
исполнительные механизмы, включая вегетативные реакции и пове­
дение, возвращают полезный приспособительный результат к необ­
ходимому для нормального метаболизма уровню. Все эти процессы
протекают непрерывно с постоянным информированием центра фун­
кциональной системы о достижении или недостижении полезного
приспособительного результата (см. главу 8). Каждая функциональная
система в динамике строится по принципу системного квантования —
от потребности к ее удовлетворению.
Интенсивность процессов саморегуляции определяет эндогенные
ритмы жизнедеятельности. Чем более важен для жизнедеятельности
тот или иной показатель внутренней среды (например, осмотическое
давление, pH среды), тем активнее и быстрее работает функциональ­
ная система, обеспечивающая своей деятельностью оптимальный
для метаболизма уровень этого показателя. Деятельность других
функциональных систем менее напряжена.
В нормальных условиях в деятельности каждой функциональной
системы проявляется следующая закономерность: общая сумма
механизмов, возвращающих отклоненны й результат к исходно­
му уровню, всегда превышает сумму отклоняю щ их механизмов.
Иными словами, в каждой функциональной системе в здоровом
организме имеется «запас прочности», позволяющий ей справиться
с любыми возможными отклоняю щими воздействиями. Так, напри­
мер, в функциональной системе, определяющей оптимальный для
организма уровень кровяного давления, общая сумма депрессорных
механизмов в нормальных условиях с избытком превышает сумму
прессорных механизмов; в функциональной системе питания меха­
низмы насыщения всегда достаточны, чтобы затормозить механиз­
мы голода, и т.д.
Голографический принцип организации
функциональных систем
В функциональных системах каждый входящий в них элемент в
своих свойствах отражает деятельность всей функциональной систе­
мы в целом и особенно — состояние ее полезного приспособительно­
Глава 2
70
го результата. Этот принцип организации функциональных систем
отражает их голографические свойства.
Имеются многочисленные примеры, свидетельствующие о том,
что в ритме деятельности любого органа, например сердца, дыха­
тельного аппарата, пищевода и желудка, а также отдельных нейронов
головного мозга отражается их включение в деятельность различных
функциональных систем организма.
По аналогии с физической голографией сигнализация о потребнос­
ти в функциональных системах может быть представлена в качестве
опорной волны, а сигнализация об удовлетворении потребностей —
предметной волны. Интерференция этих волн осуществляется на
голографических экранах — мембранах клеток, молекулах дезокси­
рибонуклеиновой кислоты и рибонуклеиновой кислоты и др.
Избирательное вовлечение элементов
в функциональные системы
Важным свойством функциональных систем является избира­
тельная мобилизация результатов деятельности отдельных органов и
тканей в целостную организацию функциональной системы. Каждая
функциональная система для обеспечения своего, полезного для
организма в целом, результата избирательно объединяет тканевые
элементы различного уровня, принадлежащие к различным анато­
мическим образованиям. Функциональные системы для достижения
разнообразных приспособительных результатов могут использовать
разные или одни и те же внутренние органы. Так, работа сердца
может быть использована как для поддержания постоянного уровня
кровяного давления, так и для обеспечения газообмена, сохранения
оптимальной температуры тела, питательного гомеостазиса и др.
Работа почек, кроме выделительной функции, может быть исполь­
зована для обеспечения оптимальных для метаболизма pH крови,
осмотического давления, температуры тела и др.
Внутри каждой функциональной системы имеется возможность
чрезвычайной взаимозаменяемости, взаимокомпенсации ее эффекторных механизмов. При выходе из строя одного или нескольких
исполнительных компонентов функциональной системы обеспече­
ние ее конечного приспособительного результата может быть осу­
ществлено другими входящими в нее компонентами. Среди испол­
нительных механизмов есть быстродействующие (оперативные) и
медленнодействующие (стратегические). Например, при повыше­
нии кровяного давления почти сразу изменяются просвет сосудов,
Общие принципы построения целого организма
71
сердечный ритм, скорость кровотока. Если эти регулирующие влия­
ния не обеспечивают нормализацию полезного приспособительного
результата, меняются ОЦК и свойства крови. Значительно позднее
в процессы саморегуляции могут включаться почечные факторы,
кроветворение и кроверазрушение.
Взаимодействие элементов в функциональных системах
Включение отдельных органов и тканей в функциональные сис­
темы всегда происходит по принципу взаимосодействия, т.е. каждый
элемент не просто пассивно включается в функциональную систему,
но, взаимодействуя с другими элементами системы, активно способс­
твует достижению ею полезного приспособительного результата.
Наиболее отчетливо избирательное взаимосодействие осущест­
вляется между объединенными в функциональные системы нервны­
ми элементами различных уровней ЦНС.
Кибернетические свойства функциональных систем
В функциональных системах проявляются общекибернетические
свойства: регулирование по конечному эффекту и обмен инф ор­
мацией. Регулирование по конечному эффекту в кибернетике, как
известно, называется обратной связью. Обратные связи открыты
Н. Винером в технических устройствах и общественных явлениях.
П.К. Анохин обнаружил их в живых организмах в виде обратной
афферентации, что составило признанный приоритет отечественной
науки в области физиологической кибернетики.
Многообразие функциональных систем в организме
Многообразие полезных для организма приспособительных
результатов указывает на то, что число функциональных систем,
организующих различные стороны жизнедеятельности целого орга­
низма, чрезвычайно велико. Одни из них определяют различные
показатели внутренней среды, другие обусловливают поведение
живых существ, их взаимодействие с окружающей средой и себе
подобными. Одни функциональные системы, особенно метаболи­
ческого и гомеостатического уровня, генетически детерминированы;
другие складываются в индивидуальной жизни животных и человека
по мере формирования и удовлетворения ими метаболических и, в
первую очередь, поведенческих потребностей организма.
В нормальном организме деятельность всех функциональных сис­
тем гармонически взаимосвязана.
72
Глава 2
Рис. 2.5. Генетическая детерминация системогенеза. Под влиянием био­
логически активных веществ (А, Б, В) складывается определенная группа
взаимосвязанных клеток и тканей (2, 3), формирующих специальную
функцию. Функция приводит к адаптивному результату (4), который на
основе обратной афферентации (6) формирует функциональную систему
(1 — геном; 5 — рецепторы результата)
Функциональные системы организуются самим ходом процессов
жизнедеятельности: процессами обмена веществ, генетически детер­
минированными и приобретенными механизмами памяти, а также
под воздействием факторов окружающей среды (рис. 2.5).
Взаимодействие функциональных систем
в целом организме
В целом организме взаимодействие различных функциональных
систем строится на основе принципов иерархии, мультипараметрического взаимодействия, последовательного взаимодействия, систе­
могенеза и системного квантования процессов жизнедеятельности.
Иерархия функциональных систем
Иерархия функциональных систем состоит в том, что в каждый
данный момент времени деятельность организма определяет домини­
рующая в плане выживаемости или адаптации к внешней среде функ­
циональная система. Принцип доминанты открыт А.А. Ухтомским.
Доминирование функциональных систем в целом организме опре­
деляется их биологической, а для человека — в первую очередь соци­
альной значимостью.
Общие принципы построения целого организма
73
По отношению к каждой доминирующей функциональной систе­
ме все другие функциональные системы в соответствии с их биоло­
гической и социальной значимостью выстраиваются в определенном
иерархическом порядке, начиная от молекулярного вплоть до оргаиизменного и социального уровней. После удовлетворения ведущей
потребности деятельностью организма завладевает следующая, веду­
щая по социальной или биологической значимости, потребность,
которая строит свою доминирующую функциональную систему. По
отношению к этой функциональной системе другие также выстраи­
ваются в иерархическом порядке. Смена доминирующей функцио­
нальной системы на другую динамически осуществляется в течение
жизни индивидов, отражая сущность непрерывно происходящего
обмена веществ и энергии и постоянного взаимодействия их с окру­
жающей средой (рис. 2.6).
Мультипараметрическое взаимодействие
Взаимодействие различных функциональных систем определя­
ет их обобщенную деятельность. Как правило, изменение одного
показателя, результата деятельности одной функциональной сис­
темы, немедленно сказывается на результатах деятельности других
функциональных систем, или один параметр сложного результата
деятельности функциональной системы приводит к изменению его
лругих параметров.
Принцип мультипараметрического взаимодействия распространяет­
ся практически на все показатели гомеостазиса и объединяет деятель­
ность различных функциональных систем гомеостатического уровня в
единую обобщенную функциональную систему гомеостазиса.
М ультипараметрическое взаимодействие объединяет также
результаты деятельности функциональных систем поведенческого и
гомеостатического уровней (рис. 2.7).
В ф ункциональных системах многосвязного взаимодействия
отклонение от оптимального уровня того или иного параметра
результата является стимулом к направленному перераспределению
(к определенных соотношениях) значений всех других параметров
результатов обобщенной функциональной системы.
Последовательное взаимодействие функциональных
систем
В целом организме проявляется континуум действия различных
функциональных систем, когда деятельность одной функциональ-
74
Глава 2
Рис. 2.6. Динамика доминирования функциональных систем: А — дом и­
нирует функциональная система с адаптивным результатом Рг Другие
функциональные системы либо способствуют деятельности первой, либо
вытормаживаются; Б — после удовлетворения потребности доминиру­
ющей функциональной системы доминирует функциональная система
с адаптивным результатом Р2; В — доминирует функциональная система
с адаптивным результатом Р3
Общие принципы построения целого организма
75
А
Рис. 2.7. Динамика мультипараметрического взаимодействия ф ункцио­
нальных систем, при которой изменение параметров одного из взаимо­
связанных результатов их деятельности изменяет состояние всех связан­
ных с ним результатов: А — доминирует функциональная система № 1;
Б — доминирует функциональная система № 2. Изменяются параметры
функциональных систем, связанных с доминирующей системой, и их
взаимосвязи
76
Глава 2
ной системы во времени сменяется деятельностью другой (рис. 2.8).
Последовательная деятельность функциональных систем гомеоста­
тического уровня жестко генетически программируется специаль­
ными центрами нервной системы, а программирование их последо­
вательного включения осуществляется по опережающему принципу.
Каждый предшествующий результат деятельности соответствую­
щей функциональной системы на основе обратной нервной и гумо­
ральной сигнализации оценивается соответствующими механиз­
мами, после чего происходит смена этой функциональной системы
на другую (рис 2.8).
Процессы последовательной смены ф ункциональных систем
наблюдаются в динамике процессов дыхания, выделения, кровооб­
ращения, поведения.
Систем огенез
Системогенез — избирательное и последовательное созревание
функциональных систем и их отдельных частей в процессе пре- и
постнатального онтогенеза.
Процессы системогенеза определяют также последовательное раз­
витие, становление и взаимодействие функциональных систем и
их деструкцию на довольно длительных отрезках индивидуальной
жизни живых существ.
Системное квантование процессов жизнедеятельности
Периодически возникающие метаболические потребности живых
существ переводят континуум их жизнедеятельности в дискретную
форму. Все многообразие жизнедеятельности в ее динамике разби­
вается на последовательный ряд результативных отрезков (рис 2.9).
Каждый результативный отрезок жизнедеятельности от потребности
к ее удовлетворению, определяемый специальной функциональной
системой, рассматривается как системоквант.
Системокванты обнаруживаются на разных уровнях жизнеде­
ятельности, например, системокванты генома, определяющие разви-
Рис. 2.8. Динамика последовательного взаимодействия функциональных
систем. Результат деятельности каждой предшествующей функциональ­
ной системы (PI, Р2, РЗ) выступает в качестве побуждающего стимула
формирования последующих функциональных систем
Общие принципы построения целого организма
77
78
Глава 2
Рис.2.9. Динамика формирования системокванта функциональной сис­
темы от потребности к ее удовлетворению через каскад промежуточных
результатов деятельности Pt—Р6 — промежуточные Рк — конечные резуль­
таты деятельности каждый результат оценивается с помощью обратной
афферентации
Общие принципы построения целого организма
79
тие эмбриона, плода и растущего организма. На уровне метаболизма
проявляются системокванты различных результативных химических
реакций. На уровне гомеостатических процессов системное квантонание отчетливо проявляется в процессах пищеварения, дыхания,
выделения, кровообращения, в половых функциях и др.
В процессе дыхания, например, можно выделить следующие сис­
темокванты со своими конечными результатами: вдох и поступление
определенного количества воздуха в альвеолы; диффузию газов из
альвеол в легочные капилляры; транспорт кислорода к тканям; диф ­
фузию газов из капилляров в ткани и из тканей в кровь; транспорт
газов к легким; альвеолярный газообмен; выдох.
Принцип системного квантования распространяется и на поведе­
ние живых существ.
Рефлекс и функциональные системы
как единицы жизнедеятельности
Рефлекс и функциональная система выступают в качестве единиц
жизнедеятельности: рефлекс — как единица отраженной деятельнос­
ти, а функциональная система — как единица процессов самооргани­
зации и саморегуляции жизнедеятельности. В отличие от рефлекса,
который строится на основе элементарных процессов возбуждения
и торможения, в организации функциональных систем участвуют
интегративные механизмы.
Глава 3
НЕРВНАЯ СИСТЕМА
3.1. ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ
НЕРВНОЙ СИСТЕМЫ
Согласно современным представлениям, в основе деятельнос­
ти ЦНС лежат процессы, связанные, в первую очередь, с работой
нервных клеток. Нейрональная теория успешно объясняет целый
ряд явлений, протекающих в головном и спинном мозге. Нейрон
является структурной и функциональной единицей нервной систе­
мы. Число нейронов в головном и спинном мозге — около 10 млрд.
Нейроны состоят из тела, отходящих от него дендритов с боль­
шим количеством синапсов и одного аксона, по которому импульсы
идут от тела к периферии. От аксона может отходить несколько
коллатералей, также заканчивающихся синапсами. Нейроны быва­
ют афферентными, вставочными и эфферентными. Афферентные
нейроны обеспечивают проведение информации от органов чувств
к ЦНС. Вставочные нейроны целиком находятся в ЦНС и обеспе­
чивают связь между афферентными и эфферентными нейронами.
Эфферентные нейроны проводят импульсы из ЦНС к мышцам или
железам. Сформулированы следующие принципы нейронной орга­
низации (Р. Кахаль).
• Нейрон с отростками составляет единое морфологическое целое.
• Нейроны генетически едины, происходят из одних нейробластов.
• Н ей рон ед и н ф ун к ц и он ал ь н о.
• Возбуждение по нейрону распространяется в одном направле­
нии — от дендритов к аксону.
• Нейрону присуще трофическое единство. При удалении тел
нейронов наблюдается гибель аксонов. Перевязка аксона также
вызывает гибель клетки.
• Нейрон участвует как единое целое в патологических реакциях.
Деструктивные изменения охватывают весь нейрон.
81
Нервная система
Нейронная теория
Указанные принципы легли в основу нейронной теории построе­
ния ЦНС. Согласно этой теории строение и функции ЦНС определя­
ются множеством взаимосвязанных нейронов с дискретными свойс­
твами. Наряду с этим не утрачивают своего значения представления
о том, что ЦНС имеет сплошное строение типа синцития, присущее,
например, нейропилю, имеющемуся в нервной системе низших бес­
позвоночных (кишечнополостные) и в мозге земноводных (аксолотли).
Гетерогенные нейроны в их взаимодействиях позволяют ЦНС человека
участвовать в миллионах разнообразных реакций. На телах отдельных
нейронов располагается от сотен до тысяч синапсов, причем синапсы
функционируют с помощью различных медиаторов и связаны со спе­
цифическими постсинаптическими химическими реакциями.
Процессы возбуждения нейронов объясняют электрическая и
химическая теории.
Электрическая теория возбуждения нейронов
Исходит из представлений о морфофункциональной неоднород­
ности нейронов. Большинство синапсов расположено на дендритах
нейронов, где они занимают 75% площади мембраны, и только 2%
синапсов — в коре мозга — на телах нейронов. Синапсы на дендритах
получили название интеграторных, а на теле клетки ближе к выходу
аксона — детонаторных. Считалось, что мембрана дендритов элект­
рически невозбудима. В настоящее время доказано, что отдельные ее
участки обладают потенциалоуправляемой проводимостью, обуслов­
ленной наличием кальциевых каналов, активация которых приво­
дит к кальциевым потенциалам действия. У места отхождения аксо­
нов располагается так называемый «светлый аксонный холмик», или
«светлое пятнышко», где синапсы отсутствуют. Этот участок нейрона
положительно заряжен по отношению к другим участкам мембраны
нейронов. При возбуждении синапсов, располагающихся на дендри­
тах и теле нейронов, увеличивается разность потенциалов между
«аксонным холмиком» и остальной частью нейрона. При достижении
определенной величины это порождает спайковую активность аксона.
Чем стремительнее нарастает разность потенциалов тела нейрона и
•■аксонного холмика», тем чаще выходная импульсация аксона.
Химическая теория возбуждения нейронов
Химическая теория связывает процессы возбуждения нейронов
го спецификой выделяемых синапсами нейронов медиаторов, олиго­
Глава 3
82
пептидов и других биологически активных веществ. Возбуждающие
медиаторы (норадреналин, ацетилхолин, глутамат и др.) деполяри­
зуют постсинаптическую мембрану и формируют возбуждающий
постсинаптический потенциал. Тормозные медиаторы (например,
ГАМК) формируют на постсинаптической мембране тормозные постсинаптические потенциалы. Процесс возбуждения нейронов зависит
от специфики постсинаптических химических реакций, распростра­
няющихся к генетическому аппарату ядер нейронов.
Медиаторные функции олигопептидов и аминокислот
Олигопептиды-опиоиды, вещество П, ангиотензин II, вазопрессин, окситоцин, а также цитокины модулируют действие медиа­
торов в синапсах ЦНС. Самостоятельную медиаторную функцию
выполняют аминокислоты — глутаминовая кислота, аспарагино­
вая кислота и ГАМК. У ряда нейронов ЦНС выявлены специ­
фические рецепторы к глутамату (NMDA-рецепторы, аффинные
к N -метил-О-аспартату). ГАМК — самый распространенный тор­
мозной медиатор в ЦНС. Другой тормозной медиатор, точкой при­
ложения которого являются нейроны продолговатого и спинного
мозга, — глицин.
Функции нейронов
• Рецепторная. Способность к восприятию информации обеспе­
чивается рецепторами постсинаптических мембран, входящих
в состав нейро-нейрональных синапсов, количество которых
может достигать нескольких тысяч.
• Память нейрона позволяет хранить информацию как генетичес­
кого, так и приобретенного характера.
• Интегративная. Основная деятельность нейрона заключается во
внутриклеточной обработке информации, которая может быть
использована в данный момент времени.
• Синтезирующая. В теле нейрона идут процессы синтеза биологи­
чески активных веществ — нейромедиаторов и нейромодулято­
ров, первые из которых обеспечивают передачу сигнала на клетку,
а вторые могут влиять на эффективность передачи, во многом
определяя ее характер. Сейчас известно, что процессы синтеза
также имеют место в корковой зоне аксона нейрона.
• Транспортная. Синтезируемые веществатранспортируютсявнутри
аксона нервной клетки к синаптическим окончаниям. Существует
два вида аксонального транспорта: быстрый, со скоростью до
Нервная система
83
20 см/сут и медленный — порядка 10 мм/сут. Транспорт осущест­
вляется при помощи специальных фибриллярных белков.
• Генераторная. Внешняя реакция нейрона заключается в гене­
рации потенциалов действия, которая происходит в начальном
сегменте нейрона. В настоящее время также показано, что генера­
ция электрических потенциалов возможна в местах отхождения
коллатералей от основного аксона.
• Кодирующая. Нейрон обладает свойством кодирования, т.е. пере­
дачи информации в условной форме кода. Рисунок последователь­
ности потенциалов действия — паттерн — есть способ участия
нейрона в конкретной экспериментальной или поведенческой
ситуации.
• Проводниковая. Нейрон обладает свойством проводника, причем
потенциалы действия из области начального сегмента распро­
страняются по аксональной мембране к рабочим синапсам.
• Секреторная. Проявляется в выделении медиаторов и некоторых
пептидов в окончании аксона.
• Трофическая. Следует отметить, что на рост аксонов и функци­
онирование синаптических связей влияют так называемые тро­
фические факторы и, в частности, фактор роста нервов (NGF),
который синтезируется клетками-мишенями, захватывается ней­
ронами и ретроградно транспортируется в сому, где влияет на
ферментативные процессы, синтез медиаторов и рост аксона.
Рассмотренные свойства нейронов позволяют им участвовать в
сложной интегративной деятельности.
Интегративная деятельность нейронов
По мнению академика П.К. Анохина, «интегративная деятельность
нейрона» включает в себя, во-первых, конвергенцию разнообразных
возбуждений на его синапсах, во-вторых, создание интегративного
состояния на основе внутриклеточной обработки информации, втретьих, формирование специфической импульсной активности на
аксоне.
П.К. Анохин сформулировал гипотезу, согласно которой процесс
возбуждения не ограничивается мембраной нейронов, а распростра­
няется в глубь цитоплазмы постсинаптических нейронов. Благодаря
множественной конвергенции возбуждений на отдельных нейронах,
нейроны, включенные в доминирующее возбуждение, избирательно
реагируют только на это возбуждение. При этом каждое из этих возбуж­
дений формирует специфическую постсинаптическую реакцию вплоть
Глава 3
84
до экспрессии геномом нейрона определенного вида белка, формиру­
ющего, в свою очередь, специфическую реакцию мембраны нейрона,
определяющую его реагирование только на те возбуждения, которые
способствуют удовлетворению исходной потребности организма.
Нервные центры
Совокупность нейронов, обеспечивающих какую-либо физиоло­
гическую функцию, называется нервным центром. Нервные центры
могут иметь разную локализацию: спинальные, стволовые, мозжеч­
ковые, подкорковые, корковые. По своим функциям нервные центры
бывают моторные или сенсорные. Они различаются также по степени
сложности: простые (спинальный центр коленного рефлекса) и слож­
ные (корковый центр речи). В зависимости от типа регулируемых
функций нервные центры бывают соматические, вегетативные и пси­
хические. Нервные центры обладают рядом свойств, которые опреде­
ляются свойствами нервных клеток и синапсов их составляющих.
Свойства нервных центров
• Одностороннее проведение возбуждений от рецептора к эффекто­
ру, что связано с локализацией основных мембранных рецепторов
на постсинаптической мембране нейро-нейрональных синапсов.
• Центральная задержка. В основе центральной задержки прове­
дения возбуждений лежит время проведения возбуждения через
один или несколько синапсов.
• Суммация возбуждений в нервном центре. Выделяют пространс­
твенную и временную суммацию, первая представляет собой
суммацию постсинаптических потенциалов от последовательно
приходящих импульсов через одни и те же синапсы, а вторая —
от одновременно приходящих сигналов через разные синапсы.
• Низкая функциональная лабильность нервных центров. Это
свойство определяется значительным временем распространения
возбуждения по нейронам нервного центра.
• Высокая утомляемость нервных центров имеет в своей основе
синаптическую депрессию, связанную с истощением запасов
медиатора в пресинаптическом окончании нейронов центра.
• Высокая химическая чувствительность нервных центров к раз­
личным эндогенным и экзогенным веществам. Химические аген­
ты могут действовать на пресинаптическую и постсинаптическую
области синапсов нейронов мозга, блокируя образование медиа­
торов, их взаимодействие, обратный захват медиатора пресинап-
Нервная система
85
тической областью или постсинаптические химические реакции,
вплоть до экспрессии генов.
• Последействие. Явление последействия в нервном центре объ­
ясняет протекание рефлекторной реакции уже после окончания
действия раздражителя. В основе последействия лежат циркуля­
ция импульсов по замкнутым нейронным цепям, наличие выра­
женной следовой деполяризации и высокоамплитудных, полисинаптических возбуждающих постсинаптических потенциалов в
нейронах ЦНС.
• Тонус нервных центров. Имеет рефлекторную природу и прояв­
ляется наличием фоновой активности в эффекторных нервах,
несущих импульсы от центра.
• При высоком уровне тонического возбуждения в нервном центре
наблюдается состояние доминанты. Доминирующий центр обла­
дает высокой и устойчивой возбудимостью, что позволяет ему
подавлять деятельность других центров. Существование доми­
нанты хорошо иллюстрирует классический опыт А.А. Ухтомского,
который заключается в электрическом раздражении моторной
коры у кошки. Раздражение всегда вызывало сокращение контра­
латеральной лапы. Однако если в прямую кишку кошки был вве­
ден резиновый баллон и его слегка раздували, то же раздражение
моторной коры вместо сокращения лапы вызывало у животного
акт дефекации. Аналогично, если в желудок предварительно вво­
дили слабый раствор отвергаемого вещества, то при раздражении
моторной коры у животных возникала рвота.
• Субординация и реципрокность в работе нервных центров.
Первое из этих свойств означает подчинение нижерасположен­
ных центров вышележащим, а второе — возникновение возбуж­
дения одного центра при одновременном торможении другого
центра.
• Пластичность нервных центров. Представляет собой перестройку
их деятельности для повышения эффективности ф ункциониро­
вания.
Особенности распространения возбуждения
в центральной нервной системе
Возбуждения, поступившие или возникшие в ЦНС, распростра­
няются в ней, используя различные нейрональные механизмы.
• Иррадиация (дивергенция) возбуждений, которая бывает гене­
рализованной, охватывающей большие мозговые территории, и
Глава 3
86
®
>
Рис. 3.1. Механизм иррадиации. Показано распространение импульсов,
поступающих по афферентным волокнам от одного нейрона через коллатерали его аксона к большему числу нервных клеток
направленной, затрагивающей лишь отдельные области мозга
(рис. 3.1).
• Конвергенция возбуждений различной сенсорной модальности и
различного биологического значения имеет место на отдельных
нейронах ретикулярной формации, неспецифических ядрах тала­
муса и коры головного мозга. Нейроны, к которым конверги­
руют возбуждения двух, трех и более сенсорных модальностей,
называются би-, три-, полисенсорными нейронами. Такой вид
конвергенции получил название мультисенсорной. Другой вид
конвергенции — мультибиологический — заключается в схожде­
нии к одному нейрону возбуждений от центров, формирующих те
или иные биологические состояния (голод, жажда, боль). Третий
вид конвергенции — сенсорно-биологический — представляет
собой комбинацию двух первых, например, схождение к нейрону
импульсов от рецепторов сетчатки глаза и гипоталамического
центра голода (рис. 3.2).
• М ультипликация возбуждений, возникающая, когда импульсы
от первого нейрона направляются по параллельным цепям с раз-
Нервная система
87
Рис. 3.2. Механизм конвергенции. Схождение импульсов, идущих по м н о­
жеству афферентных волокон, к одному нейрону: А — мультисенсорная
конвергенция происходит на нейронах, к которым конвергируют возбуж­
дения двух, трех и более сенсорных модальностей; Б — другой вид кон­
вергенции — мультибиологический — заключается в схождении к одному
нейрону возбуждений от центров, формирующих те или иные биологичес­
кие состояния (голод, жажда, боль); В — третий вид конвергенции — сен­
сорно-биологический — представляет собой комбинацию двух первых,
например, схождение к нейрону импульсов от рецепторов глазного яблока
и гипоталамического центра голода; Г — основа аксонально-сенсорнобиологической конвергенции заключается в том, что нейрон может полу­
чать возбуждения по коллатералям пирамидных нейронов моторной коры,
возбуждения от различных рецепторных зон (сенсорные) и различных
биологических центров организма (биологические). Нейрон отвечает спе­
цифическим паттерном разрядов на аксоне в каждом отдельном случае
личным числом вставочных нейронов и последующим выходом
на общий эфферентный нейрон (рис. 3.3).
• Пролонгирование возбуждений обеспечивается циркуляцией
(реверберацией) импульсов в замкнутых нейрональных цепях
типа «нейронной ловушки». Прекращение подобной ревербера­
ции возникает либо вследствие истощения запасов медиатора в
нейронах цепи, либо под действием внешнего тормозного воз­
действия. С пролонгированием возбуждения связано явление
рефлекторного последействия в реакциях организма после пре­
кращения действия раздражителя (рис. 3.4).
• Окклюзия (закупорка) заключается в том, что суммарный резуль­
тат реакции может оказаться меньшим, чем сумма взаимодейс-
Глава 3
88
2
А
3
4
S ------ N.
о------- <о------- <о-------<а*£)
Г
Рис. 3.3. Механизм мультипликации. Благодаря синаптическим задерж­
кам нейрон 4 получает последовательно три возбуждения: А — через
3 синапса; Б — через 4 синапса; В — через 5 синапсов
О---- <(^Л—Л-Л2)
Рис. 3.4. Механизм пролонгирования. Импульс проходит по основной
цепочке, а его копия по коллатерали через промежуточный нейрон пов­
торно возбуждает исходную нервную клетку. Количество импульсов на
конечном нейроне определяется временем циркулирования возбуждений
в системе вставочных нейронов
твующих реакций. Это явление объясняется частичным пере­
крытием рецепторных синаптических полей двух рефлекторных
реакций (рис. 3.5).
• Принцип общего конечного пути — мотонейроны спинного
мозга представляют собой конечный путь самых разнообразных
и многочисленных рефлексов. Ч. Ш еррингтон сравнивал конеч­
ные исполнительные нейроны с «воронкой» и рассматривал их
как «чек на предъявителя». Именно на путях к мотонейронам
Нервная система
89
Рис. 3.5. Механизм окклюзии. Раздельное возбуждение каждого из входов
приводит к активации двух нейронов. При одновременном возбуждении
обоих входов число активирующихся нейронов не достигает простой
арифметической суммы, а равно трем, поскольку существует нейрон,
активирующийся двумя входами
спинного мозга развивается конкуренция различных возбужде­
ний в ЦНС (рис. 3.6).
Торможение в центральной нервной системе
И.М. Сеченов впервые показал, что в ЦНС наряду с процесса­
ми возбуждения протекают процессы торможения. Торможением
называется активный биологический процесс, внешним проявле­
нием которого является прекращение или ослабление наличного
процесса возбуждения. Суть опыта И.М. Сеченова состоит в следу­
ющем (рис. 3.7). Обнажали мозг лягуш ки и поперечным разрезом
в области зрительных бугров отделяли большие полушария. После
подсушивания места разреза определяли время кислотного защ ит­
ного оборонительного рефлекса при погружении задней лапки
лягуш ки в раствор 0,25% соляной кислоты. Затем на место среза
помещали кристаллик поваренной соли и повторно определяли
время кислотного оборонительного рефлекса, которое при этом
увеличивалось. На этом основании был сделан вывод о формиро­
вании в ЦНС процесса торможения, который и влиял на параметры
Рис. 3.6. Механизм общего конечного пути. Возбуждения из различных
источников поступают на мотонейрон спинного мозга, ответ которого
является результатом обработки всей наличной информации (1 — пира­
мидальный тракт; 2 - руброспинальный тракт; 3 — вестибулоспинальный тракт; 4 — ретикулоспинальный тракт; 5 — вставочные нейроны
спинного мозга; 6 - чувствительные рецепторы мышечного веретена;
7 — гамма-афферентные волокна спинного мозга; 8 — альфа-мотонейрон
спинного мозга)
91
Нервная система
HCl
Рис. 3.7. Схема опыта Сеченова.
После выполнения разреза головного
мозга лягушки на уровне зрительных бугров (1) измеряют время рефлекса
отдергивания задней лапы (2) при погружении ее в раствор соляной кисло­
ты. Далее накладывают кристаллик поваренной соли на срез зрительных
бугров и повторяют тот же эксперимент. При этом время рефлекса отдер­
гивания увеличивается. Кристаллы соли, раздражая зрительные бугры,
вызывают возбуждение, которое распространяется к спинальным центрам
и тормозит их при помощи тормозных вставочных нейронов (Т Н )
двигательной реакции животного. Вначале И.М. Сеченов полагал,
что он открыл задерживающие (тормозные) центры в зрительном
бугре. Затем его представления о центральном торможении изме­
нились. И.М. Сеченов пришел к выводу, что торможение является
следствием взаимодействия двух и более возбуждений на спинальных
нейронах: возбуждения, обусловленного раздражением рецепторов соляной
92
Глава 3
кислотой, и возбуждения, обусловленного раздражением зрительных буг­
ров кристалликом соли. В этом случае одно возбуждение неизбежно
становится тормозящим, а другое — тормозимым.
Классификация торможения
• По отделам ЦНС, в которых формируется процесс торможения,
оно может быть:
—спинальное;
—ретикулярное;
—таламическое;
—мозжечковое;
—корковое.
• По конфигурации нейрональных сетей различают три вида тор­
можения:
—возвратное торможение — процесс торможения эфферентных
нейронов вызывается импульсами, возвращающимися к ним
через промежуточные тормозные нейроны (рис. 3.8); такими
нейронами могут быть клетки Реншоу в спинном мозге или
клетки Пуркинье в мозжечке;
< о ^ <
Рис. 3.8. Возвратное торможение — тормозящее действие вставочных ней­
ронов на те нервные клетки, которые их активируют. Первые два импульса
серии проходят по основной цепочке, а их копия по коллатерали аксона
подходит к тормозному нейрону (ТН); после прекращения действия тор­
мозного нейрона прохождение серии импульсов по основной цепочке
продолжается
Нервная система
93
Рис. 3.9. При латеральном торможении вставочные тормозные нейроны
(ТН) соединены таким образом, что при активации афферентных путей
(1) они блокируют проведение возбуждений по путям, параллельным цен­
тральному активирующему (2)
—латеральное торможение — торможение группы нейронов,
вызванное возбуждением соседней группы нейронов (рис. 3.9);
этот вид торможения широко представлен в анализаторах,
где оно обеспечивает их дискриминационную чувствитель­
ность или способность выделять несколько точек в рецептив­
ном поле;
—реципрокное торможение — поочередное торможение эф ф е­
рентных нейронов с участием тормозных клеток имеет место
в сегментах спинного мозга, где возбуждение мотонейронов
мышц-сгибателей сопровождается торможением мотонейронов
мышц-разгибателей (рис. 3.10).
94
Глава 3
Рис. ЗЛО. Реципрокное торможение наблюдается, когда посредством вста­
вочных тормозных нейронов происходит последовательное торможение
мотонейронов с различными функциями. Так, проведение возбуждения
по одной цепочке (1) и сокращение мышцы (2) сопровождаются активаци­
ей посредством коллатерали тормозного нейрона (TH) и соответственно
торможением проведения возбуждения по другой цепочке
• По локализации нателе нейрона:
—постсинаптическое торможение — возникает при обязательном
участии тормозного вставочного нейрона, медиатор которого
вызывает повышение порога возбуждения основного нейрона;
—пресинаптическое торможение — развивается при участии
аксо-аксонального синапса вставочного нейрона на пресинап-
Нервная система
95
Рис. 3.11. Механизмы пресинаптического и постсинаптического торможе­
ния: А — приход возбуждения по аксону 1 активирует тормозной нейрон
(ТН), который блокирует проведение возбуждения по аксону 2 (пресина­
птическое торможение); Б — приход возбуждения по аксону 1 активирует
тормозной нейрон, который непосредственно тормозит пирамидную клет­
ку 2 (постсинаптическое торможение)
тическом окончании основного нейрона, что приводит к выде­
лению ГАМК, выходу ионов С1", деполяризации мембраны,
блокированию проведения импульсов по пресинаптическим
путям к постсинаптической нервной клетке и выделению недо­
статочного количества медиатора (рис. 3.11).
Глава 3
96
• По мембранным механизмам торможения выделяют:
—гиперполяризацию — увеличение знака заряда постсинаптической мембраны под действием тормозного медиатора;
участвующие в процессах торможения тормозные нейроны
бывают трех типов: глицинергические, вызывающие постсинаптическое торможение, ГАМК-эргические, вызывающие
пост- и пресинаптическое торможение, и тормозные нейроны
смешанного типа, выделяющие глицин и ГАМК (в последние
годы обнаружены тормозные нейроны, расположенные глав­
ным образом в стволе мозга, у которых из аксона выделяются и
ГАМК, и глицин);
—устойчивую деполяризацию — как следствие устойчивой акти­
вации натриевых и калиевых каналов с удлинением периода
абсолютной рефрактерности; это явление наступает в условиях
избытка медиатора (пессимальное торможение) или отсутствия
ингибитора медиатора;
—устойчивую поляризацию, которая имеет место на фоне вещес­
тва, взаимодействующего с постсинаптическими рецепторами,
но не изменяющего проницаемость мембраны и не активирую­
щего натриевые и калиевые каналы. Такой механизм торможе­
ния называется конкурентным.
Методы исследования центральной нервной
СИСТемы (С о в м е с т н о е В .Ф . В олковы м )
Изучение топографии структур головного мозга
Стереотаксическая техника
Для введения регистрирующих и раздражающих электродов в глу­
боко расположенные структуры головного мозга при электрофизиологических исследованиях применяется стереотаксический метод.
Принцип метода заключается в том, что положение электрода отно­
сительно структур мозга — ядер и проводящих путей — определяется
по трем координатам, которые отсчитываются при эксперименталь­
ных исследованиях от ориентиров на черепе животного или, при
нейрохирургических операциях, на голове человека. Для успешного
попадания в искомую структуру используются стереотаксические
приборы, состоящие из двух частей: головодержателя для крепления
головы в строго определенном положении и системы, позволяющей
Нервная система
97
перемещать держатели электродов на заданные расстояния в прямо­
угольной или полярной системе координат. Для расчета координат
введения электродов используются атласы мозга экспериментальных
животных и человека, в которых на послойных фронтальных сре­
зах указано расположение ядер и проводящих путей относительно
базальной и сагиттальной плоскостей. Стереотаксическая техника
используется как в остром опыте на наркотизированных животных,
так и при операциях вживления электродов для хронических экспе­
риментов. В последнем случае процедура проводится в стерильных
условиях. Электроды могут применяться для регистрации элект­
рической активности структур мозга, их раздражения или локаль­
ного разрушения. Стереотаксическая техника применяется также
для прицельного введения канюль, через которые в мозг вводятся
вещества или забираются пробы, а также устройств для локального
охлаждения ткани и т.п.
Эхоэнцефалография
В основе метода лежит способность ультразвуковых колебаний
проникать через кости черепа и ткани головного мозга и отражаться
от границ анатомических образований, имеющих разную плотность,
благодаря чему можно определять наличие или отсутствие смещения
срединных структур мозга. На эхоэнцефалограмме при располо­
жении излучателей-приемников ультразвуковых волн в височных
областях головы можно видеть начальный, конечный и срединный
комплекс. Начальный комплекс возникает при отражении импульсов
от кожных покровов головы и костей черепа со стороны излучателя,
срединный комплекс есть результат отражения волны от срединных
структур мозга: III желудочка, шишковидной железы, прозрачной
перегородки. Конечный комплекс представляет собой отражение
ультразвукового сигнала от костно-кожных покровов противополож­
ной стороны головы. Если рисунки эхосигналов, полученных при
помощи излучателей, расположенных справа и слева, симметричны,
то это свидетельствует об отсутствии смещения срединных струк­
тур мозга. Если расстояние до М-эха справа меньше, чем слева, то
срединные структуры мозга смещены в результате кровоизлияния.
Величину смещения можно определить по разности М ,—М2. Данная
методика позволяет увидеть отклонения срединных структур мозга
на самые незначительные величины: 1—2 мм. Достоинства эхоэнцефалографии — простота выполнения обследования и возможность
его экстренного проведения даже по месту жительства пациента.
98
Глава 3
Обнаружение достоверных признаков смещения срединных структур
головного мозга служит основанием для проведения дальнейшего
углубленного обследования.
Нейрорентгенологические методы
Нейрорентгенологические методы основаны на том, что различ­
ные ткани по-разному поглощают рентгеновские лучи. Они широко
используются в клинике, обеспечивая визуализацию нормальных
и патологически измененных структур мозга (нейровизуализацию).
Однако применение рентгенологических методов осложняется тем,
что у структур мозга способность пропускать рентгеновские лучи
различается крайне незначительно, поэтому необходимо применять
дополнительные методы, улучшающие контрастирование исследуе­
мых областей, или специальные приемы анализа получаемых изоб­
ражений. В различных вариантах используют введение рентгенокон­
трастных веществ в сосудистое русло или спинномозговой канал.
Рентгеновская компьютерная томография
Методика рентгеновской компьютерной томографии основана
на том, что источник рентгеновского излучения подвижен, и очень
узкий рентгеновский луч проходит через голову под различными
углами. Расположение структур, задерживающих рентгеновские лучи
в соответствии с их плотностью, моделируется с применением мате­
матических методов послойно. При сканировании нескольких слоев
методы компьютерной пространственной реконструкции расположе­
ния структур позволяют увидеть детальную трехмерную прижизнен­
ную анатомическую картину мозга пациента.
Изучение электрической активности мозга
Электроэнцефалография
ЭЭГ — метод исследования головного мозга, основанный на
регистрации его суммарных электрических потенциалов. С помо­
щью этого метода можно определять локализацию органических
поражений мозга и — что более существенно — оценивать его фун­
кциональную активность.
Распространение возбуждения в мозге, изменение потенциалов на
мембранах нейронов создают неоднородное в пространстве и изменя­
ющееся во времени электрическое поле. В связи с этим между двумя
точками мозга, а также между мозгом и удаленными от него тканями
Нервная система
99
организма возникает разность потенциалов, которая и регистрирует­
ся с помощью метода ЭЭГ.
В стволе мозга и в передних отделах лимбической системы
имеются ядра, активация которых приводит к изменению уровня
функциональной активности практически всего мозга. Эти обра­
зования подразделяются на восходящ ие активирующие системы,
расположенные на уровне ретикулярной формации среднего мозга
и в преоптических ядрах переднего мозга, и подавляющие, тор­
мозящие, сомногенные системы, расположенные главным образом
в продолговатом мозге, нижних отделах моста и неспецифичес­
ких таламических ядрах. Активирующие ретикулокортикальные
и лимбикокортикальны е системы вызывают повышение уровня
функциональной активности мозга, при активации сомногенных
ретикулокортикальных и таламокортикальны х систем уровень
бодрствования снижается вплоть до засыпания. Возбуждение акти­
вирующих ретикулокортикальных систем приводит к возникнове­
нию на ЭЭГ десинхронизации, выражающейся появлением высо­
кочастотной, нерегулярной, низкоамплитудной активности. При
снижении уровня деятельности мозга сокращается афферентный
приток. Отдельные нейроны ф ункционально объединяются при
этом в огромные популяции с синхронизированной активностью,
которая отражается на ЭЭГ медленными, регулярными, высокоамплитудными колебаниями.
При экспериментах на животных регистрируется ЭЭГ коры и
подкорковых структур. В клинических исследованиях ЭЭГ отво­
дится с помощью электродов, расположенных на интактных пок­
ровах головы человека и в некоторых экстракраниальных точках.
Отведение ЭЭГ может быть монополярным, при котором активный
электрод располагается над определенным участком мозга, а пас­
сивный (референтный) — на мочке уха, подбородке или на носу.
При биполярном отведении регистрируется изменение разности
потенциалов между электродами, расположенными над мозгом.
Основными характеристиками ЭЭГ являю тся частота и амплитуда.
Частота определяется количеством колебаний в секунду и выража­
ется в герцах (Гц). Амплитуда измеряется от пика предшествующей
полны до пика последующей волны в противоположной фазе и
ныражается в микровольтах (мкВ). Определеный тип электрической
иктивности, соответствующий тому или иному состоянию мозга,
соответствует понятию «ритм». Для взрослого бодрствующего челоиека наиболее характерны альф а-(а)- и бета-((3)- ритмы (рис. 3.12).
Глава 3
100
a -ритм
\/\/\длллАллллЛЛл/уу\/^у^'Л/\Д/\ЛЛА/^лА/\|уУУу\/\/\^\лУ^\л^чЛ/'^\
p-ритм
25 мкВ
Рис. 3.12. Ритм электроэнцефалограммы.
Альфа-(а)-ритм имеет частоту 8—13 Гц, амплитуду до 100 мкВ.
Ритм лучше всего выражен в затылочных областях, наибольшую
амплитуду имеет в состоянии спокойного расслабленного бодрство­
вания, особенно при закрытых глазах.
Бета-((3)-ритм. Частота 14—40 Гц, амплитуда до 15 мкВ. Ритм появ­
ляется при повышении функциональной активности мозга, напри­
мер, при открывании глаз (реакция десинхронизации).
Тета-(0)-ритм с частотой 4—6 Гц и амплитудой от 40 до 300 мкВ и
дельта-(8)-ритм, имеющий частоту 0,5—3 Гц и амплитуду, как у тета-ритма,
могут в небольшом количестве встречаться на ЭЭГ взрослого здоро­
вого человека; у детей и подростков они наблюдаются чаще. Медлен­
новолновые ритмы характерны для состояния сна. Увеличение доли 6ритма и a -ритма при возрастании их амплитуды у взрослого бодрствую­
щего человека свидетельствует о развитии патологических процессов.
Компрессированный спектральный анализ
Сущность этого метода заключается в компьютерном преобра­
зовании первичной ЭЭГ в спектр мощностей ритмов с дальнейшим
построением трехмерных изображений структуры ЭЭГ с выделением
Нервная система
101
доминирующих частот. Особенностью компрессированного спект­
рального анализа является возможность длительного клинического
наблюдения за динамикой изменений электрической активности
коры полушарий большого мозга в течение часов и суток.
Топоселективное картирование электрической активности мозга
В основе метода лежит перевод значений электрической активности
мозга в числовую форму с последующим выведением показателей на экран
или печатающее устройство в виде карт распределения амплитуд, частот
или мощностей различных ритмов ЭЭГ на поверхности головного мозга.
Картирование распределения потенциалов позволяет определить локали­
зацию очагов максимальной активности в коре больших полушарий.
Регистрация вызванных потенциалов
Данный метод используется для определения состояния аффе­
рентных систем мозга и заключается в регистрации электрических
потенциалов, возникающих в ответ на самые различные раздраже­
ния как периферических рецепторов, так и отдельных структур ЦНС.
В отличие от изменения фоновой ритмики, вызванные потенциалы
представляют собой стереотипные комплексы колебаний, возника­
ющие с определенным латентным периодом в ответ на приход аффе­
рентных импульсов после подачи кратковременного раздражения.
Форма вызванных потенциалов непосредственно связана с характе­
ром активности нервных структур, что отражает функциональное
состояние ЦНС. Метод вызванных потенциалов используется для
определения локализации проекционных зон различных анали­
заторных систем в коре мозга. При этом в условиях регистрации
электрической активности посредством электродов, перемещаемых
но поверхности мозга, осуществляют повторные раздражения раз­
личных уровней сенсорных систем — рецепторов, периферических
чувствительных нервов, релейных ядер. Отмечая территорию макси­
мальной выраженности ответов, выявляют контуры корковой зоны,
и которой заканчиваются афферентные пути данного анализатора.
Магнитоэнцефалография
Магнитоэнцефалография (МЭГ), как и ЭЭГ, относится к функци­
ональным методам исследования ЦНС и применяется для выявления
областей мозга, связанных с решением определенных задач, анализа
пктивности данных областей и для анализа изменения их функцио­
нальной организации при нарушениях. Преимущество МЭГ по срав­
102
Глава 3
нению с ЭЭГ заключается в большей точности пространственного
разрешения — для коры головного мозга диаметр источника элект­
ромагнитного поля составляет 5 мм. По сравнению с функциональ­
ными магниторезонансными методами и ПЭТ (позитронно-эмиссионной томографией), МЭГ обладает гораздо большей скоростью
получения информативного сигнала — в течение 1 мс.
Микроэлектродная техника
Регистрация импульсной активности отдельных нейронов может
осуществляться у нервных клеток в культуре ткани, нейронов ган­
глиев беспозвоночных животных (например, виноградной улитки).
Возможна регистрация импульсной активности нейронов ЦНС у
позвоночных наркотизированных животных.
Наиболее информативной и в то же время наиболее сложной
является регистрация импульсации нервных клеток в условиях сво­
бодного поведения объекта. Наилучшее качество регистрации раз­
рядной деятельности нервных клеток отмечается при использовании
стеклянных микроэлектродов (рис. 3.13).
Оптическая регистрация импульсной активности нейронов
В некоторых экспериментальных ситуациях распространение воз­
буждения по нервным клеткам и волокнам можно зарегистриро­
вать без применения микроэлектродов. Для оптической регистрации
применяются специально разработанные красители, которые при
введении, например, в культуру нервной ткани связываются с кле­
точной мембраной нейронов и изменяют поглощение клеткой про­
ходящего света или вызывают ее флуоресценцию при изменениях
ее мембранного потенциала. Модулированный световой поток с
помощью волоконной оптики передается на оптический усилитель,
преобразуется в электрический сигнал и регистрируется.
Изучение молекулярных механизмов работы мозга
Микроионофорез
Микроионофорез — одна из наиболее тонких и эффективных ней­
рофизиологических методик, предназначенных для изучения характе­
ристик клеток ЦНС. Этот метод позволяет при регистрации импульсной
активности отдельного нейрона дозированно подводить из отдельных
стволов многоканального стеклянного электрода различные биологи­
чески активные вещества в зону расположения или внутрь исследуемой
Нервная система
103
III IV
Рис. 3.13. А — нейронограмма; Б — суммарные гистограммы импульсной
активности нейрона зрительной коры кошки на последовательных этапах
пищедобывательного условного рефлекса. Условный раздражитель - свет;
В — суммарные гистограммы импульсной активности того же нейрона в
случаях применения звукового условного сигнала (I —включение условно­
го сигнала, II —нажатие животным педали, III —момент появления молока
в кормушке, четвертая — начало лакания кошкой молока. По вертикали —
число импульсов в канале. Ш ирина канала 50 мс) (В. Андрианов, 1990)
нервной клетки. Благодаря этому методу можно анализировать чувстви­
тельность нейрона к биологически активным веществам — классичес­
ким и пептидным медиаторам, блокаторам белкового синтеза и др.
Радионуклидные методы
Радионуклидые методы основаны на регистрации излучений, возни­
кающих при радиоактивных превращениях нестабильных изотопов.
104
Глава 3
При динамических исследованиях оцениваются кровоснабже­
ние мозга и его нарушения на основе анализа времени поступ­
ления вещества, содержащего радиоактивны й изотоп, в мозго­
вой кровоток, разности времен поступления вещества в правое и
левое полушарие, времени прохождения через мозг и ряда других
показателей. Излучение изотопа регистрируется томографическими
гамма-камерами, что позволяет создать трехмерную компьютерную
реконструкцию распределения радиофармакологического препара­
та в мозге.
Позитронно-эмиссионная томография
Метод ПЭТ основан на введении короткоживущих радиоактивных
изотопов в кровеносное русло пациента или экспериментального живот­
ного с последующей регистрацией их распределения в мозге по испуска­
емым позитронам примерно в течение получаса после инъекции.
Метод обладает довольно большой пространственной разреша­
ющей способностью, позволяя различать структуры объемом около
0,5 см3, однако для получения изображения требуется от десятков
секунд до нескольких минут. Использование метода ПЭТ позволяет
увидеть места поражения мозга, а также участки, берущие на себя
функции пораженной области.
Магнитно-резонансная томография
В основе метода МРТ лежит явление ядерного магнитного резо­
нанса ядер водорода, которые взаимодействуют с внешними ста­
тическим и переменным магнитными полями. Сущность ядерного
магнитного резонанса заключается в избирательном испускании
веществом электромагнитного излучения при переориентации маг­
нитных моментов атомных ядер водорода, находящихся в постоян­
ном магнитном поле. Метод отличается достаточно высоким про­
странственным разрешением, изображение может быть получено в
течение нескольких секунд. Благодаря различным модификациям
метода МРТ можно получать изображения структуры органов и
тканей, оценивать их состояние, проводить ангиографию крупных
сосудов без введения контрастных веществ.
Функциональная магнитно-резонансная томография
Интенсивность и скорость затухания электромагнитных волн от
резонирующих ядер водорода зависят от однородности или неодно­
родности магнитного поля исследуемой области. Неоднородность
Нервная система
105
поля создается в присутствии диамагнетиков. В крови оксигемоглобин и дезоксигемоглобин имеют разные магнитные свойства:
первый является слабым диамагнетиком, второй — парамагнети­
ком. Изменение соотношения оксигемоглобин/дезоксигемоглобин в
каком-либо омываемом кровью участке ткани приводит к изменению
ее магнитно-резонансного сигнала, на чем и основан метод функ­
циональной МРТ. Благодаря методу функциональной МРТ можно
определять расположение зон, участвующих в восприятии сигна­
лов, выполнении движений, формировании речи, то есть проводить
неинвазивное картирование коры головного мозга.
Термоэнцефалоскопия
Физиологические процессы, протекающие в структурах головно­
го мозга, требуют значительного расхода энергии, в конечном счете
выделяющейся в виде тепла. Головной мозг — достаточно сильный
тепловой генератор, а для нормальной работы нейронов требуются
стабильные температурные условия. Метод термоэнцефалоскопии
основан на регистрации теплового излучения коры головного мозга
с применением техники тепловидения, цифровой обработки распре­
деления температур и построении изображения.
Микродиализ
М икродиализ позволяет исследовать химический состав межкле­
точного пространства в живых тканях и его изменения в процессе
жизнедеятельности организма. Особенности микродиализа состоят
в том, что при помощи этого метода возможно наблюдение за уров­
нем метаболитов в различных тканях в течение длительного времени
в условиях свободного поведения экспериментальных животных.
Принцип микродиализа заключается в диффузии химических соеди­
нений, находящихся в межклеточном пространстве, через полупро­
ницаемую мембрану в просвет микродиализного зонда, имплантиру­
емого в исследуемую ткань.
Картирование функционального состояния мозга по экспресии
ранних генов
Активация различных отделов головного мозга, обеспечивающих
ге или иные физиологические функции, например, формирование
долговременной памяти, сопровождается усилением синтеза спе­
цифических белков. Очевидно, что синтезу белков предшествует
активация определенных генов, кодирующих данные белки. В свою
Глава 3
106
очередь, экспрессии специфических генов предшествует активация
так называемых ранних генов c-fos b c-jun. Следовательно, выявление
в мозге областей экспрессии ранних генов может дать представление
о локализации в мозге областей повышенной активности, сопро­
вождающейся белковым синтезом. Метод основан на образовании
гибридов между матричной РН К искомого гена и комплементарным
олигонуклеотидом, меченным радиоактивным изотопом фосфора-33.
Области связывания меченого олигонуклеотида на сечениях мозга
фиксируются авторадиографической методикой и с помощью скане­
ра вводятся в компьютер для получения трехмерного изображения.
Таким образом осуществляется визуализация процессов, лежащих в
основе создаваемых мозгом различных биологических состояний, а
также тонких механизмов обучения и памяти.
Реоэнцефалография
Кровообращение мозга отражает интенсивность метаболичес­
ких процессов, происходящих в нем. Исследование достаточности
кровообращения головного мозга и его изменения при решении
определенных задач может проводиться с применением реоэнцефалографии. В основе метода лежит измерение электрического
сопротивления ткани мозга при пропускании через него слабого
переменного тока — 0,5—1,0 мА при напряжении 0,75—1,5 В с часто­
той 90—120 кГц. Изменение сопротивления связано и с изменением
объема крови, находящейся в сосудистом русле между располо­
женными на поверхности кожных покровов головы электродами.
Применение реоэнцефалографии в клинической практике и науч­
ных исследованиях позволяет получить информацию об особен­
ностях кровенаполнения бассейнов сосудов головного мозга, их
тонусе, о реакциях на воздействие различных факторов, венозном
оттоке из полости черепа. Проведение функциональных проб дает
дополнительные диагностические возможности, например отсутс­
твие реакции расширения мозговых сосудов при введении сосу­
дорасширяющих препаратов свидетельствует о преимущественно
атеросклеротическом их поражении.
Методы активного воздействия на мозг
Хирургические методы
Экспериментальные хирургические методы включают опера­
ции «подрезки» и «обрезки», суть которых заключается в изоляции
Нервная система
107
одних структур мозга от других. Операция «подрезки» коры позво­
ляет исследовать работу нейронов небольшого участка коры мозга
при нарушении их связей с нижерасположенными структурами.
Операция «обрезки» позволяет исследовать работу нейронов участка
коры, изолированного от соседних участков. Хирургические методы
позволяют прервать как афферентные потоки нервных импульсов,
так и эфферентную импульсацию от головного мозга к спинному и
далее к органам. С помощью данных методов можно делать выводы
о роли структурных связей внутри ЦНС.
Электрическое раздраж ение
Электрическая стимуляция мозга проводится специальными
электродами. Размер электродов должен быть достаточно большим,
чтобы электрический ток не вызывал термического поражения при­
легающих структур и воздействовал бы на достаточно большой объем
ткани мозга для получения ожидаемого эффекта. Чаще всего такие
электроды изготавливают из нихрома, вольфрама, золота. Ствол
электрода изолируется лаком, неизолированнаый активный кон­
чик имеет площадь около 0,1 мм. Раздражающие импульсы прямо­
угольной формы имеют длительность около 1 мс. С целью контроля
попадания электродов в ту или иную структуру с тех же электродов
можно регистрировать различные типы электрических сигналов,
соответствующих различным физиологическим процессам.
Магнитная стимуляция
М агнитная стимуляция головного мозга была разработана как
неинвазивный метод оценки функционального состояния централь­
ных двигательных проводящих путей. Теоретическая основа метода
заключается в том, что мощное магнитное поле, проникающее через
кости черепа, может возбуждать интернейроны головного мозга и
пирамидные клетки. С помощью метода магнитной стимуляции
можно оценить состояние пирамидного пути при ишемическом
инсульте, дегенеративных заболеваниях нервной системы, травме
спинного мозга.
Химическое раздраж ение
Для химического раздражения мозга вещества вводятся внутрь
желудочков мозга или непосредственно в мозговую ткань через спе­
циальные вживленные канюли, прикрепленные к костям черепа.
Исходя из целей исследования в ткани и желудочки мозга могут
Глава 3
108
вводиться самые разнообразные субстанции: глюкоза, медиаторы,
пептиды, блокаторы синтеза белка и лекарственные препараты.
Электрокоагуляционное воздействие
Электрокоагуляция — сочетание хирургического метода воздейс­
твия на мозг и электрического раздражения мозга, но при таких
параметрах тока, когда в тканях происходят необратимые структур­
ные изменения. Она применяется в сочетании со стереотаксической
техникой для деструкции глубокорасположенных мозговых структур
без значительных повреждений вышележащих.
3 .2 . ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ
НЕРВНОЙ СИСТЕМЫ
Ф ункции спинного мозга можно разделить на собственные, осу­
ществляемые при участии сегментарного аппарата, и проводниковые,
связанные с деятельностью проводящих путей. Спинной мозг —
основной канал проведения кожной, температурной, проприоцептивной и болевой сигнализации от периферических рецепторов к
высшим отделам ЦНС, а также ведущий канал проведения возбужде­
ний из ЦНС к мотонейронам и вставочным нейронам боковых рогов,
формирующих двигательные и вегетативные реакции.
Проводниковые функции спинного мозга
Через спинной мозг пролегают основные афферентные пути от
периферических кожных, проприоцептивных и висцеральных рецеп­
торов в высшие отделы ЦНС и эфферентные пути распространения
возбуждений из ЦНС к мышцам и висцеральным нервам.
Ведущие афферентные пути: путь Голля и Бурдаха; спиннотала­
мический тракт; спинно-мозжечковый тракт. Эфферентные пути:
пирамидный тракт; экстрапирамидные пути: ретикулоспинальный
тракт; руброспинальный тракт; тектоспинальный тракт; вестибулоспинальный тракт. Спинальные двигательные центры находятся,
таким образом, под контролирующим влиянием вышележащих отде­
лов ЦНС. Ретикулоспинальный тракт моносинаптически возбуждает
альфа- и гамма-мотонейроны проксимальных мышц конечностей и
туловища. Вестибулоспинальный путь обеспечивает рефлексы под­
держания равновесия и формирования позы. Мотонейроны дисталь-
Нервная система
109
ных мышц конечностей (особенно мышц пальцев кисти и стопы)
находятся под нисходящим влиянием кортикоспинального и руброспинального трактов. Тектоспинальный путь участвует в реализа­
ции зрительных и слуховых реакций четверохолмия.
Спинальный шок. Наблюдается после перерезки спинного мозга.
При этом все ф ункции спинного мозга исчезают. У лягушек реф ­
лекторные реакции спинного мозга быстро (через 10—15 мин) вос­
станавливаются. У человека восстановления ф ункций разорванного
спинного мозга практически не происходит. Ф. Гольтц рассмат­
ривал спинальный шок как результат раздражения. Спинальный
шок также наблюдается при Холодовой блокаде спинного мозга
(Ч. Ш еррингтон, Г. Тренделенбург). Повторная перерезка спинного
мозга ниже первой перерезки не вызывает спинального шока. Все
это указывает на то, что спинальный шок возникает вследствие
отсутствия функциональных связей спинного мозга с вышележа­
щими отделами головного мозга.
Собственные функции спинного мозга
Они включают в себя спинальные соматические рефлексы раз­
личной степени сложности, имеющие центральное звено в тех или
иных сегментах спинного мозга, и так называемые вегетативные
рефлексы. Каждый сегмент спинного мозга получает афферентную
импульсацию через задние чувствительные корешки и посылает воз­
буждение к эффекторам через передние двигательные корешки (закон
Белла—Мажанди). Если на спинальной лягушке в области поясничпо-крестцовых сегментов перерезать справа все задние корешки, а
слева все передние, то правая конечность теряет чувствительность, но
способна к рефлекторному движению, а левая, наоборот, сохраняет
чувствительность, но не совершает движения. Таким образом, уже
и пределах каждого сегмента спинного мозга возможна интеграция
простейших рефлекторных реакций.
Спинальные соматические рефлексы спинного мозга
Одним из принципов работы ЦНС является рефлекторный. Рефлекс
представляет собой ответную реакцию организма на раздражитель, осу­
ществляющуюся при участии ЦНС. Материальная основа рефлекса —
рефлекторная дуга, которая состоит из рецептора, афферентного волокпа чувствительного нейрона, центральных нейронов, расположенных в
структурах мозга, эфферентного волокна моторного нейрона и эффекгора — мышцы или железы. Выделяют простую (рис. 3.14), моно- или
Глава 3
110
Рис. 3.14. П роведение возбуж ­
дения по простой рефлекторной
дуге: возбуждение раздражителем
рецептора; проведение возбужде­
ния по афферентному нейрону;
активация вставочного нейрона;
возбуждение эфферентного ней­
рона; проведение возбуждения по
двигательному волокну; возбуж­
дение и сокращение мышцы
полисинаптическую (рис. 3.15) и
полисегментарную дугу (рис. 3.16).
Время от момента раздражения
рецептора до начала ответной
реакции эффектора называет­
ся временем рефлекса. Рефлексы
можно классифицировать по
ряду признаков: по локализации
их центров — спинальные, буль­
барные, стволовые, корковые; по
отношению к регулируемым орга­
нам — соматические и вегетатив­
ные; по эффектору — мышечные,
секреторные; по структуре цент­
рального звена — моносинаптические и полисинаптические; по
функциональному значению —
сгибательные и разгибательные,
статические и статокинетические,
одиночные и цепные, сухожильные
и миотатические; по биологичес­
кому значению — защитные и обо­
ронительные. Наконец, рефлексы
могут быть аллиированными, или
союзными, если они содействуют
друг другу, и антагонистическими,
когда они оказывают друг на друга
тормозящие влияния.
Вегетативные рефлексы спинного мозга
На уровне спинного мозга осуществляется регуляция ряда веге­
тативных функций:
• дефекация;
• мочеиспускание;
• регуляция сосудистого тонуса;
• эрекция и эякуляция;
• потоотделение.
В боковых рогах грудных и верхних поясничных отделов спинного
мозга располагаются первые нейроны симпатического, а в крестцо­
вых отделах — парасимпатического отдела ВНС.
111
Нервная система
Животные с перерезкой между спинным и продолговатым мозгом
получили название спинальных животных. У спинальных животных:
отсутствует самостоятельное дыхание; наблюдается низкое кровяное
давление; снижен сосудистый тонус; утрачена способность поддер­
живать постоянство температуры тела; исчезают все формы сложной
целенаправленной деятельности (пищевая, половая, оборонитель-
Б
I/
MB
Рис. 3.15. Проведение возбуждений по моносинаптической и полисинаптической рефлекторным дугам: А — возбуждение рецепторов мышечного
веретена и проведение возбуждения по афферентному и эфферентному
нейронам в моносинаптической дуге; Б — возбуждение внешним раздра­
жителем рецептора; проведение возбуждения по афферентному нейрону,
нескольким вставочным и эфферентному нейрону в полисинаптической
дуге. Отмечается большее время рефлекса
Глава 3
112
Рис. 3.16. Проведение возбуждения в полисегментарной рефлекторной
дуге. Возбуждение от рецептора посредством коллатерали распространя­
ется на два сегмента спинного мозга с последующим выходом на эф ф ерен­
тные нейроны
ная). При перерезках ниже люмбальных отделов наблюдается пара­
лич анальных и мочеиспускательных сфинктеров.
В табл. 3.1 представлены методы исследования и рефлекторные
дуги важнейших рефлексов ЦНС.
Таблица 3.1. М етодика исследования основных рефлекторных реакций
человека
Рефлексы
М етодика исследования
Кожные рефлексы Рукояткой неврологического моло­
точка, затупленной иглой или спич­
Брюшные:
■ верхний
кой наносить штриховое раздражение
на три-четыре пальца выше пупка
параллельно реберной дуге, наблю­
дать сокращение брюшных мышц на
соответствующей стороне
Рефлекторная
дуга
т,-т 8
Нервная система
113
Окончание табл. 3.1
Рефлексы
• средний
• нижний
Надкостничные
рефлексы:
• надбровный
• нижнечелюс­
тной
М етодика исследования
Рефлекторная
дуга
Т*9 — Т1 10
Нанести штриховое раздражение
кожи на уровне пупка, наблюдать
сокращение брюшных мышц
Нанести штриховое раздражение на
т111 - т'12
кожу на три-четыре пальца ниже пупка,
наблюдать сокращение брюшных мышц
Неврологическим молоточком нанес­ I ветвь V-VII
ти легкий удар по краю надбровной
черепных нервов
дуги, наблюдать смыкание век
0
1
0
ON
1
0
"J
1
ооо
Неврологическим молоточком слегка Чувствительные
ударить по подбородку при приоткрытом и двигательные
рте, наблюдать сокращение жевательных волокна III ветви
мышц и поднятие нижней челюсти
Vчерепного нерва
• пястно-лучевой Неврологическим молоточком нанес­
(стилорадиаль- ти удар по шиловидному отростку,
ный)
наблюдать сгибание в локтевом суста­
ве и пронацию предплечья
• лопаточноНеврологическим молоточком нанес­
с 3- с 5
плечевой
ти удар по внутреннему краю лопат­
ки, наблюдать приведение, ротацию
плеча кнаружи
• костно-абдо­
минальный
Неврологическим молоточком слег­
ка ударить по краю реберной дуги,
кнутри от сосковой линии, наблюдать
сокращение мышц живота на соот­
ветствующей стороне
Рефлексы со сли­
зистых оболочек:
Кусочком свернутой ватки прикоснуть­
ся поочередно к роговице правого и
Чувствительные
левого глаза, наблюдать смыкание век
и двигательные
Кусочком свернутой ватки прикос­
волокна V—VII
нуться поочередно к конъюнктиве
черепных нервов
правого и левого глаза, наблюдать
смыкание век
Шпателем или чайной ложкой прикос­
нуться к задней стенке глотки, наблю­ Чувствительные
дать кашель или рвотное движение
и двигательные
ядра IX—X череп­
Шпателем или чайной ложкой кос­
нуться небной занавески, наблюдать ных нервов
поднятие небной занавески
• корнеальный
• конъюнкти­
вальный
• глоточный
• небный
т 6- т 7
Глава 3
114
Физиология продолговатого мозга
и варолиева моста
Животные с перерезками головного мозга по переднему краю
варолиева моста называются бульбарными. Бульбарные животные
самостоятельно дышат, поддерживают постоянный уровень АД.
Однако они не могут поддерживать постоянство температуры тела
и передвигаться.
Проводниковые функции продолговатого м озга и моста
Через продолговатый мозг проходят восходящие и нисходящие
пути спинного мозга: пирамидный, руброспинальный, спинотала­
мический, а также заканчиваются пути Голля и Бурдаха. Кроме того,
в продолговатом мозге начинаются вестибулоспинальный, ретикуло­
спинальный и оливоспинальный пути. По этой причине при элек­
трическом раздражении вентромедиальных отделов ретикулярной
формации ствола мозга наблюдается торможение: коленного реф­
лекса; кортикально вызванных движений; передачи возбуждения по
у-афферентным волокнам. При раздражении дорсолатеральных учас­
тков ретикулярной формации ствола мозга облегчаются коленный
рефлекс и кортикально вызванные движения; увеличивается мышеч­
ный тонус мышц сгибателей и разгибателей. Серотонинергические
нейроны локализуются в дорсальном и медиальном ядрах шва про­
долговатого мозга, а также в среднем мозге и варолиевом мосту.
Их аксоны иннервируют гипоталамус, миндалины, бледный шар,
гиппокамп и кору больших полушарий. Высокая концентрация серо­
тонина обнаружена в эпифизе.
Собственные функции продолговатого м озга и моста
В продолговатом мозге расположены ядра VII, IX, X, XI и XII
черепных нервов. В мосту находятся ядра V и VI пар черепных нервов.
При участии структур продолговатого мозга и моста осуществляются
соматические и вегетативные рефлексы.
Соматические рефлексы: совместно с центрами среднего мозга
продолговатый мозг и мост участвуют в реализации тонических реф­
лексов, обеспечивающих формирование позы. Перерезка на уровне
переднего края варолиева моста вызывает у животных усиление тону­
са мышц-разгибателей. Животные не сохраняют равновесия, пада­
ют. Это явление получило название «децеребрационная ригидность».
Полагают, что децеребрационная ригидность связана с устранением
Нервная система
115
нисходящих влияний мозжечка и красных ядер среднего мозга на
бульбарные центры.
Нейроны продолговатого мозга проводят первичный анализ вес­
тибулярного, кожного, слухового и вкусового раздражителя.
Пищевые рефлексы: глотание, сосание, жевание, рефлекторная
деятельность пищеварительного тракта (слюноотделение, секреция
желудка и других пищеварительных органов).
Защитные рефлексы: чихание, кашель, рвота, мигание (моргание),
слезоотделение.
Указанные реакции сложно координированы. За счет объеди­
нения нескольких ядер нервов они охватывают комплекс испол­
нительных мышечных и вегетативных реакций. Каждая из этих
реакций имеет специфическое рецепторное (рефлексогенное) поле
и вызывается специфическим раздражителем. Например, глотание
формируется при пищевом тактильном раздражении задней стенки
ротовой полости, мигание — при раздражении конъюнктивы глаза;
механическое раздражение глотки вызывает рвоту, раздражение той
же области пищей — глотание.
Вегетативные рефлексы: регуляция дыхания осуществляется нахо­
дящимся в продолговатом мозге парным дыхательным центром,
состоящим из отделов вдоха и выдоха; регуляция сосудистого тонуса —
сосудодвигательным центром; регуляция сердечной деятельности —
ядром блуждающего нерва и нейронами, связанными со спинальны­
ми центрами, что обеспечивает тормозные и возбуждающие эффекты
соответственно. Некоторые нейроны продолговатого мозга обладают
автоматизмом. Такие нейроны, в частности, формируют смену вдоха
и выдоха. Причиной автоматизма дыхательных бульбарных центров
являются изменение в них внутренних метаболических процессов в
ответ на поступающую импульсацию блуждающих нервов и гумо­
ральное влияние двуокиси углерода. В мосте располагаются нейроны
пневмотаксического центра, регулирующие смену в работе центров
вдоха и выдоха.
Об участии ствола мозга в обменных процессах свидетельствует
известный опыт К. Бернара. При локальном уколе в дно IVжелудочка
обнаруживается резкое увеличение содержания глюкозы в крови
(«сахарный укол»).
Физиология среднего мозга
Животные с перерезкой мозга на уровне передних бугорков чет­
верохолмия получили название мезенцефальных. М езенцефальные
116
Глава 3
животные отличаются от спинальных поддержанием мышечного
тонуса и положения тела в пространстве. У мезенцефальных живот­
ных отсутствует явление децеребрационной ригидности. Черная
субстанция участвует в регуляции актов жевания, глотания и их
последовательности. В черной субстанции сосредоточены дофа­
миновые нейроны, участвующие в организации эмоционального
поведения и в регуляции тонких движений (например, пальцев рук).
Нейроны серого вещества вокруг сильвиева водопровода обладают
рецепторной функцией по отношению к спинномозговой жид­
кости. Они участвуют в поддержании внутренней среды на пос­
тоянном уровне. Красные ядра, локализованные в среднем мозге,
оказывают тормозное влияние на вестибулярные ядра Дейтерса,
к которым распространяется афферентация от ушных лабиринтов
и мышечных проприоцепторов по вестибулоспинальному тракту.
Разрушение красных ядер вызывает у животных выраженную децеребрационную ригидность. Децеребрационная ригидность опре­
деляется также афферентацией, поступающей к ядрам Дейтерса
от проприоцепторов мышц и вестибулярного аппарата. Перерезка
задних корешков спинного мозга и вестибулярных нервов также
устраняет децеребрационную ригидность. При отсутствии тормозя­
щих влияний красных ядер на уровне заднего мозга избирательно
повышается активность мышц-разгибателей. Норадренергические
нейроны располагаются в голубоватом ядре и прилежащих учас­
тках продолговатого и среднего мозга (вентролатеральные ядра
ретикулярной формации). Аксоны этих нейронов широко распро­
страняются в стволе мозга, мозжечке, гипоталамусе, гиппокампе,
миндалине, поясной и энторинальной коре.
Проводниковая функция среднего мозга
Эта функция осуществляется через ножки мозга, по которым в
восходящем направлении несут импульсы волокна спиноталамичес­
кого тракта и медиальной петли, а в нисходящем — пирамидного и
кортикомостового трактов.
Собственные функции среднего мозга
Тонические рефлексы обеспечивают регуляцию позы. Тонические
рефлексы подразделяются на статические и статокинетические
(Р. Магнус и А. Де-Клейн). Первые выступают как рефлексы положе­
ния (позы) и рефлексы выпрямления.
Нервная система
117
Статические рефлексы положения определяют поддержание удоб­
ной позы в пространстве при лежании, сидении и стоянии животных
за счет изменения мышечного тонуса. Статические рефлексы связаны
с раздражением лабиринтов вестибулярного аппарата, с положением
головы по отношению к туловищу, с сигнализацией от рецепторов
кожи, мышц и суставов всего тела, со зрительной афферентацией о
предметной структуре среды. Так, при поднятой голове усиливается
тонус разгибателей передних и сгибателей задних конечностей, а при
опущенной голове — тонус мышц-разгибателей задних конечностей
и сгибателей передних конечностей.
Статические рефлексы выпрямления определяют возвращение
животного из неустойчивого положения в устойчивое. Эти реак­
ции также определяются раздражением лабиринтов вестибулярного
аппарата, афферентацией от шейных мышц и от кожных рецепторов
поверхности тела. Основной фактор при смене позы — изменение
положения головы. Лежащее животное прежде всего поднимает
голову и устанавливает ее в горизонтальном положении. Вслед за
этим немедленно за счет шейно-тонических рефлексов и афферентации от рецепторов мышц и кожи перераспределяется мышечный
тонус, определяющий вставание животного. Если на мезенцефальное животное слегка надавить сверху доской, то голова его тут же
падает.
Статокинетические рефлексы связаны с изменением тонуса мышц
при прямолинейном или круговом движении, что сопровождает­
ся перераспределением тонуса соматической мускулатуры. При вра­
щательном движении наблюдается нистагм головы и глаз, который
выражается в медленном движении головы и глаз в сторону, про­
тивоположную вращению, и быстром возвращении в исходное сос­
тояние.
Ориентировочные рефлексы обеспечиваются нейронами ядер вер­
хнего и нижнего четверохолмия, которые являются первичными
зрительными и слуховыми центрами и определяют такие компонен­
ты ориентировочных реакций, как прислушивание, перемещение
глазных яблок, регуляцию просвета зрачка, поворот ушных раковин
у животных.
Сторожевые реакции при внезапных световых или звуковых раз­
дражениях обеспечивают у человека старт-реакцию — немедленную
мобилизацию мышечного аппарата к действию. Корковые влияния
могут усиливать или ослаблять эти реакции.
Глава 3
118
Физиология мозжечка
Ф ункции мозжечка связаны, главным образом, с организацией
двигательных актов и регуляцией вегетативных функций.
При осуществлении двигательного акта перемещающиеся части
тела испытывают влияние инерционных сил, что нарушает плав­
ность и точность выполняемого движения. Структуры мозжечка
осуществляют коррекцию движения. Мозжечок определяет пос­
троение быстрых баллистических целенаправленных движений
(например, бросание мяча в цель, прыжок через препятствие, игра
на фортепиано). В таких случаях коррекция по ходу выполнения
движения невозможна из-за малых временных параметров, и бал­
листическое движение будет выполнено только по заранее заготов­
ленной программе.
Афферентные связи мозжечка, посредством которых импульсы пос­
тупают в различные его отделы, включают в себя пути от вестибуляр­
ных ядер, соматосенсорные пути из спинного мозга и нисходящие пути
из коры головного мозга. В структурно-функциональном отношении
мозжечок состоит из трех отделов: архицеребеллюма (древний мозже­
чок), палеоцеребеллюма (старый мозжечок), неоцеребеллюма (новые
структуры мозжечка). Архицеребеллюм включает в себя флокуллонодулярную долю, к которой подходят афферентные вестибулярные
волокна и волокна вестибулярных ядер. Палеоцеребеллюм состоит из
передней доли, простой дольки и задней части тела мозжечка, куда
поступают волокна из сенсомоторной коры мозга и спинного мозга.
При этом спинномозговые и корковомозжечковые пути имеют в про­
межуточной зоне передней доли четкую соматотопическую организа­
цию. Неоцеребеллюм включает среднюю часть тела и большую часть
полушарий мозжечка. Афферентные пути в этот отдел мозжечка идут
от коры мозга, а также от рецепторов зрительного и слухового ана­
лизаторов. В кору мозжечка импульсы поступают по лиановидным
и моховидным волокнам, окончания которых имеют четко выражен­
ную соматотопическую организацию. Лиановидные волокна несут
информацию от кожных и мышечных рецепторов, которая поступа­
ет в мозжечок через нижнее оливарное ядро продолговатого мозга.
Моховидные волокна несут информацию от коры, проприоцепторов
и вестибулоцепторов, которая поступает к мозжечку через ядра ство­
ла мозга. Третьей афферентной системой в коре мозжечка является
моноаминергическая, которая состоит из норадренергических, серотонинергических и дофаминергических волокон.
Нервная система
119
Кора мозжечка имеет выраженное клеточное строение (рис. 3.17).
Основными клеточными элементами являются клетки-зерна, звез­
дчатые клетки, корзинчатые клетки, клетки Гольджи и клетки
Пуркинье. Одно лиановидное волокно (медиатор аспартат) образует
Р ис. 3.17. Схема
основны х м еж нейрональны х связей мозжечка:
КК — корзинчатые клетки; ЗК — звездчатые клетки; КЗ — клеткизерна; КГ — клетки Гольджи; КП — клетки Пуркинье; КЯ — клетки
ядер мозжечка; MB — моховидные волокна; Л В — лиановидные волокна;
«+» — возбуждающ ие и «—» — тормозные влияния
120
Глава 3
контакт с одной клеткой Пуркинье, что создает условия для сильно­
го моносинаптического активационного входа посредством данного
афферентного пути. Моховидные волокна оказывают на клетки
Пуркинье как возбуждающее действие — посредством вовлечения
клеток-зерен (медиатор глутамат), так и тормозное — посредством
звездчатых и корзинчатых клеток. Клетки Пуркинье имеют тор­
мозные входы от корзинчатых клеток и звездчатых клеток, а также
активационный от отростков клеток-зерен. Клетки Пуркинье пред­
ставляют собой высокодифференцированные нейроны, аксоны кото­
рых обеспечивают единственный эфферентный выход тормозного
характера на ядра мозжечка.
Кора мозжечка может изменять активность ядер мозжечка при
помощи, прежде всего, процессов торможения. Основным медиа­
тором клеток Пуркинье является ГАМ К. Разряды клеток Пуркинье
коррелируют с фазой стояния или флексией шагательного цикла.
Функции мозжечка
Соматические функции. Мозжечок, во-первых, строит общую схему
движения, во-вторых, получает информацию от проприоцепторов
о ходе выполнения движения, в-третьих, осуществляет сравнение
параметров выполняемого движения с моделью общей схемы дви­
жения и, в-четвертых, проводит необходимую коррекцию самого
двигательного акта.
Эфферентная система мозжечка определяется существованием
трех корково-ядерных зон: медиальной, промежуточной и лате­
ральной, расположенных в медиолатеральном направлении в соот­
ветствии с проекцией коры мозжечка на его ядра. М едиальная
(червячная) зона проецируется на ядро шатра, вестибулярное ядро
моста и ретикулярную формацию. Импульсы, идущие по вестибулоспинальному пути, вызывают активацию альфа- и гамма-мото­
нейронов разгибателей и торможение мотонейронов мышц-сгибате­
лей. Ретикулярная формация оказывает либо активирующее, либо
тормозящее влияние на мотонейроны спинного мозга. Эти нисхо­
дящ ие влияния обеспечивают регуляцию тонуса и позы как основы
движения. Промежуточная (околочервячная) зона проецируется на
пробковидные и шаровидные ядра, а через них на красное ядро,
которое посылает импульсы по руброспинальному тракту к ней­
ронам спинного мозга. Кроме того, промежуточная зона получает
копию сигналов от пирамидного тракта о готовящемся движении,
Нервная система
121
что п о зв о л я ет м о зж еч к у и зм ен я т ь п о л о ж ен и е тела д л я в ы п о л н ен и я
п о сл ед у ю щ его д в и ж е н и я , а так ж е о б ес п еч и в а ет в за и м н у ю к о р р ек ­
ц и ю п о зн ы х и л ок ал ь н ы х д в и ж е н и й в п о в ед е н ч е ск о м акте. Б лагодаря
н а л и ч и ю зн а ч и т ел ь н о го с е н с о р н о г о п р и т о к а о т р ец еп т о р о в м ы ш ц и
сустав ов м о зж еч о к р а ботает как ап парат ср а в н ен и я , о с у щ е с т в л я ю ­
щ и й о б щ у ю р егу л я ц и ю м отор н ой ф у н к ц и и по п р и н ц и п у о б р а т н о й
св я зи. В сл едстви е э т о г о при у ч а ст и и п р о м е ж у т о ч н о й зо н ы м о зж еч о к
б ы стр о и сп р ав л я ет о т к л о н ен и я в т р а ек т о р и и д в и ж е н и я , ч то о б е с ­
п еч и в ает п о д го т о в к у к д в и ж е н и ю с его п о с л е д у ю щ е й к о р р ек ц и ей .
Л атерал ьн ая зо н а , п ол уч аю щ ая и н ф о р м а ц и ю от всех отд ел о в коры
м озга, п осы л ает и м п ул ь сы к зу бч ат ом у я др у и д а л е е к к р а сн о м у я д р у
ср ед н е г о м озга. В ы р аж ен н ы е т о р м о зн ы е п р о ц ессы в эт о м о т д ел е
м о зж еч к а дел аю т н ев о зм о ж н ы м х р а н е н и е и н ф о р м а ц и и у ж е ч ер ез
30 м с п о сл е ее п о с т у п л е н и я , что с о з д а е т усл о в и я д л я о б е с п е ч е н и я
бы стры х д в и гател ь н ы х актов, н а п р и м ер в сп о р т и в н ы х и гр а х и л и
п ри и гр е н а м узы к ал ь н ы х и н ст р у м ен т а х . Т ак и м о б р а зо м , л а т ер а л ь ­
ная зо н а п р и н и м а е т у ч аст и е в ф о р м и р о в а н и и п р огр ам м ы бы стры х
ф а зи ч е с к и х д в и ж е н и й .
Ф у н к ц и и м озж еч к а п о д р о б н о изуч ен ы на безм озж еч к ов ы х со б а к а х
(Л. Л ю чи ан и ). П ри уд а л ен и и м озж еч к а у со б а к н абл ю дается м ы ш еч ­
ный ги п ер т он ус за счет о сл абл ен и я в л и я н и й к расн ы х ядер ср ед н его
мозга. В дал ьн ей ш ем р азвиваю тся сл ед у ю щ и е си м п том ы .
• А то н и я (ги п о т о н и я ) — с н и ж е н и е м ы ш еч н о го т о н у са . Голова
ж и в о т н ы х п ов и сает н а грудь.
• А бази я — потеря сп о с о б н о с т и п рав и л ьн о п р оец и р ов ать ц ен тр
веса тела на п лощ адь опоры . При эт о м характерн а х одь ба на
ш и р ок о р асстав л ен ны х ногах (как при качке п о п алубе).
• А ст ази я — н ар уш ен и е ст о я н и я . У си л ен н ое качание из ст о р о н ы в
стор ону. У л ю дей астази я п р оя в л я ется при н а р у ш ен и и ф у н к ц и й
м озж еч к а, п ри закры ты х глазах и вы тянуты х в п ер ед руках (п р оба
Р ом берга).
• А таксия — н ар уш ен и е д в и ж ен и я . П оходка стан ов и тся р а сх л я б а н ­
н ой , как у пьяного.
• А ст ен и я — п ов ы ш ен н ая утом л я ем ость . П ри это м зубчаты й т ета ­
н ус вы является при тех частотах р а зд р а ж ен и й м ы ш ц , при к о т о ­
рых о бы ч н о п рояв л яется гл адк и й т етан ус — повы ш ается ф у н к ц и ­
он альн ая лаби л ь н ость м ы ш ц.
• Н абл ю дается д р о ж а н и е — ст ати ч еск и й и д и н а м и ч еск и й трем ор.
• В озрастает т ер м о п р о д у к ц и я .
122
Глава 3
У человека устранение мозжечковых влияний приводит к следу­
ющим явлениям:
• размашистости движений (вместо круга больные рисуют изло­
манную линию);
• отсутствию точных движений;
• больные с поражением мозжечка не могут при закрытых глазах
дотронуться до кончика носа (пальценосовая проба);
• изменению речи (скандирование, прерывание речи на необычных
слогах);
• при откидывании головы назад больной падает.
Вегетативные функции. Мозжечок играет значительную роль в
регуляции функций организма. Так, удаление мозжечка приводит к
нарушению деятельности сердечно-сосудистой системы, повышению
тонуса гладкой мускулатуры кишечника, бронхов и т.д. В основе этих
последствий — связи вегетативных центров гипоталамуса с различ­
ными отделами мозжечка, включая нейроны его коры.
Нейроны зубчатой фасции мозжечка оценивают состояние внут­
ренних органов и приспосабливают их деятельность, так же как и
деятельность мышц, к интересам целого организма.
При повреждении мозжечка нарушаются углеводный, минеральный
и белковый обмен, а также процессы энергообразования, терморегуля­
ции и кроветворения. Мозжечок участвует в репродуктивных функци­
ях. Благодаря обширным связям мозжечка с ретикулярной формацией
среднего мозга, красными и вестибулярными ядрами, гипоталамусом,
лимбическими структурами и корой больших полушарий он высту­
пает в роли своеобразного компаратора, динамически оценивающего
движения и деятельность внутренних органов организма.
Физиология ретикулярной формации
ствола мозга
Ретикулярная (сетчатая) формация охватывает значительное про­
странство ствола мозга. Она включает: латеральные ретикулярные
ядра; ретикулярные вентральные ядра; ядра покрышки мозга (ядра
Бехтерева); парамедиальные ретикулярные ядра; ретикулярные гига­
нтоклеточные ядра; оральные ретикулярные ядра моста; каудаль­
ные ретикулярные ядра моста; ретикулярные мелкоклеточные ядра.
Некоторые исследователи включают в состав ретикулярной формации
ствола мозга задние отделы гипоталамуса и вентральные ядра таламу­
са. На одной клетке ретикулярной формации имеется до 40 ООО синап­
сов, что указывает на ее широкие связи с другими клетками. Кроме
Нервная система
123
того, работу клеток ретикулярной формации ствола мозга определяют
гуморальные факторы. Нейроны ретикулярной формации обладают
рядом особенностей: они полисенсорны, имеют спонтанную актив­
ность, высокую возбудимость, повышенную чувствительность к ряду
фармакологических препаратов. Показано, что инъекции адрена­
лина непосредственно в ростральные отделы ретикулярной форма­
ции вызывают активацию ЭЭГ. Такое же действие оказывает аппли­
кация С 0 2 к структурам ретикулярной формации среднего мозга
(E.JI. Голубева). В структурах ретикулярной формации обнаружены
адренергические, дофаминергические, холинергические нейроны, что
свидетельствует о химической гетерогенности ее структур.
Проводниковые функции ретикулярной формации
Афферентные пути представлены спиноретикулярным, несу­
щим импульсы от температурных и болевых рецепторов, а также
церебеллоретикулярным корково-подкорково-ретикулярным путем
(от коры, базальных ядер и гипоталамуса). Эфферентные пути: нисхо­
дящий ретикулоспинальный, ретикуло-корково-подкорковый, ретикуло-мозжечковый, а также пути к другим отделам ствола мозга.
Собственные функции ретикулярной формации
Соматические функции
Восходящие активирующие влияния ретикулярной формации. В ходе
специальных экспериментов было показано, что раздражение ретику­
лярной формации через вживленные электроды пробуждает спящее
животное, а у животных, находящихся под наркозом, наблюдается
реакция активации на ЭЭГ. Наряду с этим при двустороннем разру­
шении структур ретикулярной формации ствола головного мозга у
животных наблюдается беспробудный сон (Д. Линдсли).
Таким образом, ретикулярная формация ствола мозга оказывает
па кору больших полушарий восходящие генерализованные акти­
вирующие влияния. Они адресуются практически ко всем отделам
головного мозга. При этом генерализованные влияния ретикулярной
формации проявляются в коре через 20—30 мс после того, как воз­
буждения по лемнисковым путям активируют нейроны проекцион­
ных областей коры больших полушарий. Это связано с тем, что от
пемнисковых путей к ретикулярной формации ствола мозга отходит
шачительное количество коллатералей. Именно по этой причине в
ответ на раздражение за вызванными потенциалами в соматической
124
Глава 3
коре кожной поверхности спустя несколько десятков миллисекунд
следуют потенциалы по всей коре мозга (рис. 3.18.).
Тонические активирующие влияния ретикулярной формации
образуются за счет постоянного поступления к ее структурам аффе­
рентных нервных импульсов от различных органов чувств и перифе­
рических рецепторов, а также благодаря наличию в ней многочислен­
ных аппаратов мультипликации импульсов, «нейронных ловушек»
и циклических (реверберирующих) взаимодействий между различ­
ными ее отделами и другими структурами мозга. Установлено, что
любые сенсорные и гуморальные факторы активируют ретикуляр­
ную формацию ствола головного мозга. Это указывает на неспецифи-
Рис. 3.18. Проведение восходящих возбуждений по специфическому сома­
тическому афферентному пути и через ретикулярную формацию. Первое
возбуждение направляется от рецепторов в кору по проекционным сен­
сорным путям и достигает специфической для данного анализатора кор­
ковой зоны, где возникает вызванный потенциал. Второе возбуждение,
следующее от ретикулярной формации, генерализованно направляется в
кору мозга. Активное состояние ретикулярной формации поддерживается
непрерывным потоком афферентных импульсов, поступающих к ней по
коллатералям от проекционных сенсорных путей
Нервная система
125
ческие восходящие активирующие влияния ретикулярной формации
на кору головного мозга и другие его образования.
Вместе с тем подтверждено, что ретикулярная формация изби­
рательно чувствительна к различным наркотическим веществам.
Так, под уретановым наркозом при раздражении седалищного нерва
отчетливо проявляются восходящие активирующие влияния рети­
кулярной формации на кору головного мозга. В то же время они
не проявляются у животных, находящихся под нембуталовым нар­
козом. Все это указывает на химическую неоднородность структур
ретикулярной формации ствола мозга. Об этом свидетельствуют и
результаты экспериментов, показавшие, что неблокируемые уретаном восходящие активирующие влияния ретикулярной формации на
кору головного мозга при ноцицептивных раздражениях животных
блокировались дополнительным введением им а-адреноблокатора —
аминазина. Все это также указывает на наличие в составе ретику­
лярной формации структур, определяющих состояние бодрство­
вания, и химически отличных структур, определяющих состояние
боли. Показано, что аминазин избирательно устраняет активацию
условно-рефлекторного оборонительного состояния у животных и не
влияет на их пищевые условные рефлексы (А.И. Шумилина).
Все вышеизложенное позволило сформулировать представления о
специфических восходящих активирующих влияниях ретикулярной
формации на кору головного мозга при реакциях различного биоло­
гического качества (П.К. Анохин).
Эксперименты продемонстрировали, что специфика восходящих
активирующих влияний ретикулярной формации на кору головного
мозга при реакциях различного биологического качества определя­
ется дополнительными влияниями структур гипоталамуса и лимби­
ческих образований головного мозга.
Нисходящие влияния ретикулярной формации. Структуры ретику­
лярной формации ствола мозга оказывают облегчающее и тормозное
влияния на рефлекторные реакции спинного мозга. Примером таких
нисходящих влияний является описанное выше сеченовское тормо­
жение, которое состоит в угнетении пассивной реакции отдергива­
ния лапки лягуш ки в ответ на действие на нее соляной кислоты при
наложении кристаллика соли на ретикулярные структуры таламуса.
Тормозные влияния на сгибательные рефлексы оказывает преиму­
щественно ретикулярная формация продолговатого мозга, а облегча­
ющие — ретикулярная формация моста (рис. 3.19). Поддержание пос­
турального тонуса мышц и контроль выполнения фазных движений
126
Глава 3
Рис. 3.19. Активирующие и тормозные нисходящие влияния ретикулярной
формации ствола мозга на сегментарный аппарат спинного мозга: А — рит­
мическое раздражение участка моторной коры электрическими импульсами
приводит посредством возбуждения пирамидных нейронов к ритмическим
сокращениям мышцы конечности собаки (1). Одновременное постоянное
раздражение электрическим током мезэнцефалических отделов ретику­
лярной формации (2) сопровождается увеличением амплитуды вызванных
мышечных сокращений; Б — раздражение бульбарных отделов ретикуляр­
ной формации (3) приводит к активации вставочных тормозных нейронов,
что сопровождается снижением амплитуды вызванных сокращений
формация осуществляет через тракты экстрапирамидной системы —
ретикулоспинальный, вестибулоспинальный, руброспинальный.
Активация красноядерно-спинномозговых влияний приводит к
возбуждению а - и у-мотонейронов сгибателей и торможению соот­
127
Нервная система
ветствующих нейронов разгибателей. Центробежные команды от
нейронов вестибулярных ядер вызывают обратные эффекты — акти­
вацию а - и у-мотонейронов разгибателей и торможение соответс­
твующих мотонейронов мышц-сгибателей. Нейроны ретикулярной
формации продолговатого мозга возбуждают а - и у-мотонейроны
сгибателей и тормозят мотонейроны мышц-разгибателей.
Как известно, поза формируется совокупностью рефлексов пере­
распределения мышечного тонуса при раздражении вестибулярных
рецепторов и проприоцепторов мышц шеи и туловища. Таким обра­
зом, возбуждения проприоцепторов скелетной мускулатуры и вес­
тибулярных ядер являются ведущими в механизмах формирования
позы.
Вегетативные функции
Раздражение электрическим током ретикулярных структур у экс­
периментальных животных приводит к изменению ряда вегетатив­
ных функций. В ретикулярной формации продолговатого мозга
расположены нейроны, составляющие отдел вдоха и отдел выдоха
дыхательного центра. Их деятельность контролируется нейрона­
ми пневмотаксического центра ретикулярной формации варолиева
моста. В ретикулярной формации продолговатого мозга находит­
ся сосудодвигательный центр, электрическое раздражение которого
сопровождается сосудистыми реакциями.
Физиология гипоталамуса
Гипоталамус, являясь сложноорганизованной структурой, осущест­
вляет целый ряд вегетативных функций. В нем выделяют до 50 пар
ядер, образующих несколько групп: преоптическую группу (паравенгрикулярные, преоптические, медиальные и латеральные преоптические ядра), переднюю группу (супраоптические, паравентрикулярные
и супрахиазматические ядра), среднюю группу (вентромедиальные и
дорсомедиальные ядра), наружную группу (латеральные ядра и ядра
серого бугра), заднюю группу (задние и перифорникальные ядра,
медиальные и латеральные ядра мамиллярных тел). В гипоталамусе
нет гематоэнцефалического барьера, а его нейроны имеют высокую
чувствительность к изменениям парциального давления кислорода
(р02) и углекислого газа (рС 02), кислотности среды, содержанию кате­
холаминов, ионов натрия и калия. В вентробазальных и латеральных
ядрах также обнаружены глюкорецепторы. Располагающиеся в гипо­
таламусе дофаминергические нейроны посылают короткие аксоны в
128
Глава 3
гипофиз. Другие дофаминергические нейроны располагаются в черной
субстанции. Аксоны этих нейронов проецируются в полосатые тела.
Тела третьих дофаминергических нейронов локализуются в среднем
мозге рядом с черной субстанцией. Их аксоны проецируются в лим­
бические структуры (миндалину, обонятельный бугорок, септальную
область, поясную извилину и энторинальную кору) и кору головного
мозга, особенно ее фронтальные отделы.
Проводниковые функции и связи гипоталамуса
Афферентные связи: гипоталамус имеет связи со структурами
лимбической системы, базальными ганглиями, таламусом, височной,
орбитальной и теменной корой.
Эфферентные связи обеспечивают передачу информации к тала­
мусу, гипофизу, гиппокампу, ретикулярной формации, вегетативным
ядрам ствола и спинного мозга.
Собственные функции гипоталамуса
В гипоталамусе сосредоточены ведущие центры биологических
мотиваций голода, жажды, полового влечения, температурные цен­
тры, центр сна и бодрствования и др. М отивациогенные центры
гипоталамуса при мотивациях различного биологического качества
оказывают на кору больших полушарий специфические восходящие
активирующие влияния, которые при вовлечении ретикулярной ф ор­
мации могут приобретать генерализованный характер. Как высший
отдел вегетативной регуляции гипоталамус корректирует работу ее
парасимпатического и симпатического отделов. Так, раздражение
ядер переднего отдела гипоталамуса вызывает трофотропные эффек­
ты парасимпатической нервной системы: снижение АД, усиление
моторной и секреторной деятельности пищеварительного тракта,
уменьшение силы и ЧСС. Раздражение задней группы ядер гипота­
ламуса оказывает эрготропное влияния через симпатическую нервную
систему, которые имеют противоположный характер в виде повы­
шения кровяного давления, снижения моторной и секреторной
деятельности пищеварительного тракта, а также увеличения силы
сокращений сердца и ЧСС. Здесь же расположены многочислен­
ные группы нейронов, продуцирующих факторы, освобождающие
гормоны — либерины и статины, которые посредством порталь­
ной системы оказывают стимулирующее или тормозное влияние
на выделение тропных гормонов передней долей гипофиза. Задние
отделы гипофиза накапливают гормоны, регулирующие деятель­
Нервная система
129
ность матки и молочных желез, а также водно-солевое равновесие
в организме.
Физиология лимбической системы
Лимбическая система состоит из древней коры (палеокортекса), старой коры (архикортекса) и срединной коры (мезокортекса).
Палеокортекс объединяет препириформную, периамигдалоидную,
диагональную кору, обонятельные луковицы, обонятельный бугорок,
прозрачную перегородку. Архикортекс включает гиппокамп, зубча­
тую фасцию, поясную извилину. Мезокортекс объединяет инсулярную кору и парагипнокампальную извилину. В состав лимбической
системы мозга входят миндалины мозга, ядра перегородки, переднее
таламическое ядро, мамиллярные тела и гипоталамус. Лимбическая
система тесно связана с моноаминергической системой мозга, вклю­
чающей в себя норадренергические, серотонинергические и дофаминергические механизмы. Нейроны данной системы берут начало
в стволе мозга и широко распространяют свои влияния по всему
мозгу. Тела норадренергических нейронов расположены в продолго­
ватом мозге и мосту; серотонинэргических — в ядрах продолговатого
мозга и нижних отделах среднего мозга, а дофаминергических —
в вентральных отделах среднего мозга.
Проводниковые функции и связи лимбической системы
Лимбические структуры получают афферентные возбуждения со
стороны практически всех органов чувств: обонятельного тракта
(перегородка, миндалины); зрительных путей через таламус (гип­
покамп, инсула); слуховых путей (гиппокамп, инсула); тактиль­
ных путей (инсула); от висцеральных органов (инсула). Эфферентные
возбуждения направляются из лимбической системы к таламусу,
подкорковым ядрам, большинству структур ствола и определенным
областям коры головного мозга.
Лимбические образования связаны друг с другом кольцевыми
взаимосвязями. Различают несколько лимбических кругов.
Большой гиппокампальный круг Пейпса включает мамиллярные
тела, таламус, гиппокамп, энторинальную кору, ядра перегородки,
мамиллярные тела гипоталамуса (рис. 3.20).
Септально-амигдалоидный круг: перегородка — диагональный
пучок — миндалина — концевая полоска — перегородка.
Септально-гиппокампальный круг: перегородка — супракалозальные полоски — гиппокамп — свод — перегородка.
Рис. 3.20. Основные лимбические структуры головного мозга и боль­
шой гиппокампальный круг Пейпса: 1 — парагиппокамповая извилина;
2 — гиппокамп; 3 — свод; 4 — перегородка; 5 — мамиллярные тела;
6 — передний таламус
Собственные функции лимбических структур
Циркуляция возбуждений по лимбическим кругам определяет
суточные ритмы многих физиологических функций: голод, насыще­
ние, мочевыделение, секреция гормонов корой надпочечников и т.д.
Лимбическая система — центральное звено в формировании эмоци­
ональных реакций и восприятии обонятельных раздражителей. Она
участвует в формировании эпилептической активности. При этом
блокируются болевые ощущения. Поясная извилина выполняет роль
главного интегратора тех образований мозга, которые участвуют в
формировании эмоций. К важнейшим структурам лимбической сис­
темы относятся гиппокамп и миндалина.
Гиппокамп формируют повторяющиеся модули, ритмическая
деятельность которых приводит к высокоамплитудной ритмической
активности. Нейроны гиппокампа имеют высокий уровень фоновой
Нервная система
131
частоты разрядов. Электрическая активность гиппокампа и новой
коры имеет реципрокный характер: в период усиления медленной
активности в новой коре в гиппокампе возникает десинхронизация.
Если же в неокортексе усиливается десинхронизация, то в гиппо­
кампе появляется 0-ритм. 0-Ритм отражает участие гиппокампа в
ориентировочном рефлексе, настораживании, внимании и процес­
се обучения. Повышенный 0-ритм является показателем страха,
агрессии, сильного голода и жажды. Гиппокамп имеет хорошие
связи с задними зонами лобной коры, что обеспечивает процессы
перехода кратковременной памяти в долговременную. При наруше­
ниях структур гиппокампа утрачиваются старые навыки и с трудом
образуются новые, проявляется дефицит кратковременной памяти.
Пациенты не узнают лечащих врачей, не помнят дату своего рож­
дения, не могут выучить название больницы и даже не запомина­
ют дорогу к своей кровати. Они не помнят счет спортивной игры,
теряют нить разговора.
М индалина находится в глубине височной доли мозга.
Полисенсорные нейроны миндалины имеют рецепторные поля с
широкой конвергенцией на раздражители различной сенсорной
модальности и обладают широкой химической чувствительностью
к биологически активным веществам. Нейрональная активность
может коррелировать с дыханием и ритмом сердечных сокращений.
Миндалина тесно связана с регуляцией таких вегетативных фун­
кций, как пищеварение, работа сердца, деятельность мочеполовой
и дыхательной системы. М индалина тесно связана с вентральной
лобной корой, поражение которой у человека сопровождается нару­
шением эмоций.
Физиология таламуса
Таламус имеет около 120 ядер, которые можно разделить на
три группы: переднюю с проекцией в поясничную кору, медиаль­
ную — в лобную и латеральную — в теменную, височную и заты­
лочную.
Проводниковые функции таламуса
Основная функция таламуса — проводниковая. Таламус — кол­
лектор практически всех афферентных потоков (за исключением
обонятельных), идущих к коре больших полушарий. В таламусе име­
ется четкая локализация представительства различных афферентных
мутей, идущих от экстеро-, проприо-, интерорецепторов.
132
Глава 3
Специфические (лемнисковые) проводящие пути к коре больших
полушарий распространяются через латеральные ядра таламуса. Эти
афферентные пути заканчиваются в определенных специфических
ядрах таламуса (передние вентральные, медиальные, вентролатеральные, постмедиальное, постлатеральное, а также медиальные и
латеральные коленчатые тела), откуда возбуждения направляются по
двум путям: к базальным ядрам конечного мозга и к специфическим
проекционным зонам коры мозга (зрительной, слуховой, соматосен­
сорной и др.). К специфическим ядрам таламуса поступает афферен­
тация от тактильных, болевых, температурных, мышечных рецеп­
торов, а также от блуждающих и чревных нервов. Особенно тесно
таламус связан с соматосенсорной областью коры большого мозга.
Их взаимодействия осуществляются благодаря наличию корково­
таламических волокон, образующих замкнутые циклические связи.
Регуляция передачи возбуждения через релейные ядра осуществля­
ется с помощью как возбуждающих, так и тормозных интернейронов.
Тормозные интернейроны возбуждаются от таламокортикальных
клеток, осуществляя возвратное торможение, и от корковых нейро­
нов, проецирующихся в данное ядро. Этот механизм и обеспечивает
корковый контроль проведения возбуждения. Медиальные и лате­
ральные коленчатые тела выступают как подкорковые центры слуха
и зрения соответственно.
Неспецифические ядра таламуса (срединный центр, парацентральное ядро, центральное, медиальное и латеральное, субмедиальное, переднее вентральное, парафасцикулярное, ретикулярное,
перивентрикулярное ядро и центральная серая масса) являются
местом переключения восходящих активирующих влияний, которые
направляются от ретикулярной формации ствола в кору большого
мозга. Благодаря своим связям неспецифические ядра выступают
в качестве посредника между стволом мозга и мозжечком, с одной
стороны, и лимбической корой, новой корой и базальными ганг­
лиями — с другой. Высокочастотное раздражение неспецифических
медиальных ядер таламуса вызывает локальную кратковременную
активацию коры мозга.
Специфические и неспецифические ядра таламуса находятся в
тесной функциональной взаимосвязи, что обеспечивает первичный
анализ и интеграцию всех афферентных возбуждений, поступающих
в головной мозг. Основная функция неспецифических ядер — облег­
чение или торможение ответов в специфической коре.
Нервная система
133
Ассоциативные ядра (медиодорсальное, латеральное дорсальное
ядро и подушка) обрабатывают информацию различной модальнос­
ти, которая поступает в 1-й и 2-й слои ассоциативной зоны коры
и — частично — в проекционные зоны коры (4-й и 5-й слои).
Собственные функции таламуса
В ядрах таламуса выявлена пространственная топография раз­
личных частей тела, в частности сетчатки, языка и кортиева органа.
Ядра таламуса:
• определяют двигательные и вегетативные реакции, связанные с
сосанием, жеванием, глотанием и смехом;
• участвуют в механизмах медленноволновой фазы сна;
• участвуют в механизмах боли; определяют возникновение как
мучительной каузалгии, так и отсутствие болевых ощущений.
Двигательные реакции организма интегрируются в таламусе с
вегетативными процессами, обеспечивающими выполнение дви­
жения.
Физиология базальных ганглиев
Базальные ганглии головного мозга состоят из полосатого тела, в
которое входят хвостатое ядро и скорлупа, бледного шара и ограды.
Базальные ганглии связывают ассоциативные области коры больших
полушарий с моторной корой головного мозга и моторными ядра­
ми ствола мозга (рис. 3.21). Афферентные связи имеются со всеми
отделами коры больших полушарий, неспецифическими ядрами
таламуса и черным веществом. Эфферентные связи идут от бледного
шара к структурам ствола мозга: ретикулярной формации, красному
ядру и через них к спинному мозгу, а также от бледного шара через
нижнюю оливу к мозжечку. Среди структурных образований экстрапирамидной системы полосатое тело (стриатум) считается высшим
подкорковым регуляторно-координационным центром организации
движений. Полосатое тело и бледный шар (паллидум), влияя на
мотонейроны через структуры среднего и продолговатого мозга,
координируют тонус и фазовую двигательную активность мышц,
в том числе и мышц, обеспечивающих повороты глазных яблок.
11олосатое тело инициирует организацию движения. Удаленность
хвостатого ядра от сенсорных входов определяет его нейронные реак­
ции: они мультимодальны и коррелируют не с параметрами сенсор­
ного раздражителя, а с реакцией внимания и состоянием ожидания.
134
Глава 3
Рис. 3.21. Схема связей между отдельными образованиями базальных ган­
глиев головного мозга: АК — ассоциативная кора; СК — сенсорная кора;
МК — моторная кора; Р.Ф. — ретикулярная формация; К.Я. — красное ядро;
М.О.З. — мозжечок; «+» — возбуждающие и «—» — тормозные влияния
Нервная система
135
Скорлупа формирует специальную форму мотивационной деятель­
ности — пищевое поведение. Бледный шар участвует в обеспечении
ориентировочной деятельности и оказывает тормозящее воздействие
на ядра среднего мозга.
Ограда уменьшает пороги восприятия на раздражители слуховой,
зрительной и соматической модальности. Наличие прямых связей
между бледным шаром и таламусом, а также другими стволовыми
реле специфических афферентных путей свидетельствует о том, что
настройка анализаторов управляется не просто наличием объекта,
но манипуляциями с ним. Подобная моторно-сенсорная регуляция
составляет специфику выхода стриопаллидума на анализаторы.
Полосатое тело и бледный шар, влияя на нейроны спинного мозга
через структуры среднего и продолговатого мозга, координируют
тонус и фазовую двигательную активность мышц. При участии стриарных образований мозга осуществляется синергизм всех элементов
таких сложных двигательных актов, как ходьба, бег, лазанье, переша­
гивание через препятствие, вдевание нитки в иголку, а также дости­
гаются плавность движений и установка исходной позы. Базальные
ядра не имеют прямых выходов к мотонейронам спинного мозга,
а опосредуют свои влияния на них через ретикулоспинальный тракт.
Эти влияния во многом определяют возбудимость у-мотонейронов спинного мозга, которые регулируют поток проприоцептивных
афферентных импульсов, поступающих в спинной мозг от мышечных
веретен. Кроме того, ретикулярные эфферентные импульсы влияют
на возбудимость а-мотонейронов, активность которых определяет
рабочее состояние скелетных мышц.
Базальные ганглии имеют обширные связи с областями коры и
таламическими ядрами в виде трех замкнутых петель. Первая из
них — скелетно-моторная петля — служит для регуляции таких пара­
метров движения, как амплитуда, сила, направление. Деятельность
стриопаллидарных структур связана с выполнением медленных или
червеобразных движений, таких как медленная ходьба. Полосатое
тело дает также начало многим двигательным путям экстрапирамидной системы, среди которых выделяют эффекторный путь к
паллидуму, далее — красное ядро — руброспинальный тракт —
спинной мозг. В свою очередь, бледный шар оказывает тормозящее
воздействие на ядра среднего мозга. Неостриатум включает хвос­
татое ядро и скорлупу. Эти образования и бледный шар вызывают
торможение моторного компонента условных и безусловных реакций
организма, а нейроны головки хвостатого ядра принимают участие
Глава 3
136
в организации предпусковых процессов, которые включают пере­
стройку позы, предшествующей произвольному двигательному акту.
Возбуждающим медиатором, осуществляющим корково-стриатные
связи, является глутамат, а ГАМК между базальными ганглиями и
таламусом осуществляет тормозное действие. Считается, что базаль­
ные ганглии выполняют также роль общего «моторного тормоза» в
двигательной сфере.
Вторая петля — глазодвигательная — участвует в регуляции саккад,
дрейфа и вестибулярных движений глазных яблок.
Наконец, сложные петли участвуют в обеспечении контроля
мотивации и прогнозирования, а в целом в координации мотиваци­
онных и познавательных процессов, необходимых для организации
движения. Хвостатое ядро, скорлупа и бледный шар участвуют не
только в регуляции моторной деятельности, но и в анализе афф е­
рентных потоков, в регуляции ряда вегетативных функций, в осу­
ществлении сложных форм врожденного поведения, в механизмах
кратковременной памяти, а также в регуляции цикла «бодрство­
вание-сон». Ш ирокое участие стриопаллидарных образований в
организации сложного поведения базируется на обширной мультисенсорной и гетерогенной конвергенции возбуждений к их отде­
льным нервным клеткам. На нейронах базальных ядер происходит
взаимодействие афферентных потоков, поступающих практически
от всех сенсорных структур, от многих областей коры головного
мозга, от таламических, ретикулярных, нигральных, лимбичес­
ких и других структур мозга. Все это позволяет сделать вывод
об участии базальных ганглиев в эмоциональных (аффективных)
и познавательных (когнитивных) процессах. Симптомы, связан­
ные с нарушением деятельности базальных ганглиев, проявляю тся
в гипофункции — акинезии (отсутствии движений) и гиперфунк­
ции — хорее (быстрых подергиваниях).
Физиология коры больших полушарий
В коре большого мозга (неокортекс) человека различают пять
долей: лобную, теменную, затылочную, височную и островковую,
каждая из которых имеет проекционные и ассоциативные области
(рис. 3.22).
Лобная доля
Лобная доля отвечает, прежде всего, за формирование двига­
тельной активности. В ней располагаются двигательная, премотор-
Нервная система
137
ная, дополнительная двигательная и глазодвигательная области.
Расположенная в прецентральной извилине коры большого мозга
двигательная область (рис. 3. 23) дает начало пирамидному тракту.
Пирамидные клетки моторной коры являются пунктом широкой
А
Б
Рис. 3.22. Карта коры больших полушарий мозга человека: А — латераль­
ная поверхность; Б — медиальная поверхность. Цифры обозначают изви­
лины коры мозга
138
Глава 3
Рис. 3.23. Сенсорные (А) и моторные (Б) представительства в коре боль
ших полушарий (X. Пенфилд, Т. Расмуссен, 1950)
L
Нервная система
139
конвергенции возбуждений различных сенсорных и биологических
модальностей. В пирамидной системе, являющейся общим конечным
путем выхода на периферию многих сенсорных сигналов, мультисенсорная конвергенция создает благоприятные условия для интег­
рации возбуждений в системных процессах формирования целенап­
равленного поведения. Двигательные области коры имеют сложную
организацию. Так, активация отдельных участков первичной двига­
тельной коры приводит к дискретным движениям групп мышц на
противоположной стороне тела. Данная область задействована при
выполнении самого движения. Стимуляция дополнительной двигатель­
ной области приводит к возникновениям сложных позных движений.
Эта область коры формирует двигательные команды. Стимуляция
лобных глазодвигательных полей сопровождается содружественными
движениями обоих глаз, а раздражение премоторной коры сопровож­
дается двигательными реакциями только при действии интенсивных
стимулов. Считается, что эта область коры управляет осевой мус­
кулатурой туловища. Интеграция возбуждений индивидуальна для
каждого нейрона коры и подчинена полезному результату деятель­
ности организма. Наконец, в нижней лобной извилине домини­
рующего полушария расположен двигательный центр речи Брока.
Нейроны, расположенные в префронтальной области, при целе­
направленном поведении отражают динамику поведенческого акта,
связанную главным образом с моментом выбора цели, механизмами
внимания и кратковременной памяти. Передний отдел лобной доли
также принимает участие в формировании личностных качеств и
творческих процессов.
Теменная доля
Ф ункции теменной коры больших полушарий определяются
наличием соматосенсорной области с поступающей к ней информа­
цией от специфических сенсорных ядер таламуса, а также наличием
теменной ассоциативной области (см. рис. 3.23). Соматосенсорная
область коры мозга получает также сигналы от неспецифических
ядер таламуса. Эти неспецифические влияния определяют уровень
активного состояния коры большого мозга. Общим для сенсорных
областей коры большого мозга является наличие большого количест­
ва специфических нейронов, которые дают реакции на раздражители
строго определенной сенсорной модальности. Среди специфических
нейронов выделены проекционные нейроны, имеющие однозначную
связь с проекционным рецептивным периферическим полем на теле,
140
Глава 3
и непроекционные нейроны, возбуждающиеся с различных рецеп­
тивных полей. При поступлении возбуждения в первичные сенсор­
ные области специфические проекционные нейроны дают реакции
с коротким стабильным латентным периодом, которые сопровожда­
ются последующей активацией или торможением. Неспецифические
нейроны как на специфический, так и на неспецифический раз­
дражители отвечают после длительного вариабельного латентно­
го периода, причем реакция, обычно превышает время действия
раздражителя. В специфических проекционных зонах наблюдается
сенсорно-биологическая конвергенция возбуждений, когда к одним
и тем же нейронам поступают сенсорные возбуждения и возбуждения
от мотивациогенных центров гипоталамуса и лимбических структур
головного мозга. В настоящее время не говорят ни об узкой локали­
зации корковых центров, ни об их эквипотенциальности (равнознач­
ности). Благодаря тесному взаимодействию возбуждений в коре мозга
по вертикальному и горизонтальному типу можно сделать вывод о
динамической локализации функций, что лежит в основе процессов
компенсации нарушенных функций коры больших полушарий при
ее повреждении.
Затылочная и височная доли
В затылочной коре проводится переработка зрительной, а в височ­
ной — слуховой информации. Данные области коры мозга получают
также сигналы от неспецифических ядер таламуса.
Исследование структурно-функциональной организации зритель­
ной области коры больших полушарий головного мозга было начато в
фундаментальных работах Д. Хюбела и Т. Визела (1977), показавших
наличие в этой структуре «колонок» нервных клеток с одинаковыми
по своим характеристикам рецептивными полями. В основе форми­
рования рецептивного поля лежит принцип конвергенции к нейрону
возбуждений из различных отделов ЦНС. Согласно классическим
представлениям ориентационная избирательность «простых» корко­
вых нейронов создается в связи с приходом к одному и тому же нейрону
импульсов от нервных элементов латерального коленчатого тела,
обладающих круговыми рецептивными полями. В свою очередь,
формирование «сложных» и «сверхсложных» клеток, реагирующих
на все более сложные зрительные сигналы, определяется конверген­
цией возбуждений от нейронов с «простыми» рецептивными полями.
Так, при световой стимуляции обоих глаз активируется до 80% ней­
Нервная система
141
ронов зрительной коры. Кроме того, в 18-м поле зрительной области
коры мозга кошки, в отличие от 17-го и 19-го полей, обнаружены
нейроны, реагирующие на степень освещенности.
Зрительные поля 17 и 18 кошки получают входы в основном из
слоев дорсального ядра наружного коленчатого тела. В обоих полях
ретинотопически представлено все поле зрения. Основываясь на
функциональных отличиях нейронов, предполагают, что в поле 17
происходит анализ статических объектов в зрительном пространстве,
а в поле 18 — движущихся. Предполагается, что поле 17 анализирует
и передает в поле 18 информацию о горизонтальной составляющей
изображения, а поле 18 анализирует и передает в поле 17 информа­
цию о вертикальной составляющей этого же изображения. Согласно
современным данным, рецептивные поля клеток ЦНС и, в частности,
зрительной области коры головного мозга не являются стабильными,
но в определенных условиях могут существенно изменяться.
Слуховая кора имеет тонотопическую организацию, что поз­
воляет ей проводить всесторонний анализ звукового раздражителя.
В коре расположены «колонки» нейронов, реагирующих на звук одной
частоты. Имеются суммационные «колонки» с преимущественной
активацией на бинауральный раздражитель и ослабляющие «колонки»,
преимущественно реагирующие на моноуральный раздражитель.
Ассоциативные функции коры принадлежат тем енно-височно­
затылочным и лобным областям. Именно здесь осуществляется
высший анализ соматосенсорных, вкусовых и зрительных сигналов.
Ассоциативная кора занимает до 70% неокортекса. Ассоциативные
функции лобной области коры большого мозга играют главную
роль в выработке стратегии поведения. Выбор стратегии поведе­
ния особенно нагляден при смене одних поведенческих актов на
другие. Основным источником возбуждений, поступающих к ассо­
циативным областям коры мозга, являются другие проекционные
и ассоциативные корковые зоны. На нейроны ассоциативной коры
головного мозга конвергируют возбуждения различной сенсор­
ной модальности (мультисенсорная конвергенция). Благодаря гене­
рализованным восходящим активирующим влияниям на многие
нейроны различных отделов ассоциативной коры головного мозга
конвергируют возбуждения различной биологической модальности
(мультибиологическая конвергенция). Обширные зоны ассоциа­
тивной височной доли коры мозга включаются в механизмы долго­
временной памяти. Оценка достигнутых результатов в ассоциатив­
142
Глава 3
ных зонах коры осуществляется специальным видом конвергенции
возбуждений на нейронах (аксонально-сенсорно-биологическая
конвергенция). При этом к одним и тем же нейронам, особенно
лобных долей коры мозга, передаются возбуждения сенсорной и
биологической модальностей, а также возбуждения от коллатералей
пирамидного тракта. В коре заднего отдела верхней височной изви­
лины левого полушария расположен слуховой центр речи (центр
Вернике), поражение которого приводит к нарушению понимания
устной речи.
Среди мозговых структур именно коре головного мозга посвяще­
но наибольшее число работ по изучению тонких нейрональных и
нейрохимических механизмов, лежащих в основе формирования и
осуществления поведения. Так, было показано, что нервные клетки
коры мозга могут принимать участие в регуляции достаточно слож­
ного поведенческого акта либо на всех, либо на отдельных этапах.
При этом вовлечение корковых нейронов в центральные механизмы
целенаправленной деятельности происходит на нейрохимических
путях, что подтверждается особенностями чувствительности отдель­
ного нейрона к различным веществам на разных этапах поведения
(рис. 3.24). Интегративная деятельность нейрона — способность ней­
рона к получению и обработке многочисленной информации различ­
ного качества с последующей генерацией собственного возбуждения,
представляющего собой вклад данного нейрона в общую систему
возбуждений целого мозга, обеспечивающую формирование целос­
тного приспособительного поведенческого акта. Корковые нейроны
отражают в своей импульсной активности не только пространственно-временные и вероятностные параметры окружающей среды, но
и отношение живого организма к этой среде, вплоть до активной
трансформации самой предметной среды.
У необученного животного имеется лиш ь основа поведения —
мотивационное возбуждение, поступающее к нервным элементам
коры головного мозга, а также обстановочная афферентация от
органов чувств. По мере обучения к ним присоединяется обратная
афферентация, т.е. информация о различных параметрах движе­
ния. Достижение конечного полезного для организма результата —
подкрепления — сопровождается эмоциональной оценкой, а также
приводит к колебаниям уровня мотивационного возбуждения.
Функциональная асимметрия коры мозга. Исторически сформирова­
лись две концепции локализации функций в коре головного мозга.
Первая заключалась в узком локализационизме, а вторая — в функ-
143
Нервная система
I —нажатие н а педаль
II — м омент появления м олока в кормуш ке
III - начало л акания молока
Рис. 3 .2 4 . Импульсная активность нейрона зрительной коры мозга кошки
на различны х этапах осущ ествления животны м целенаправленного
пищедобывательного поведенческого акта: А — разрядная деятельность
в поведенческом акте; Б, В, Г — суммарные гистограммы нейронной
активности по 10 выполнениям поведенческих актов: Б — без подведения
веществ; В — на фоне микроионофоретического подведения к нейрону
ацетилхолина; Г — норадреналина. У нейрона, импульсная активность
которого повышалась с момента нажатия животным педали, м икроионофоретическое подведение к нейрону ацетилхолина устраняло его разряд
при нажатии животным педали и снижало импульсную активность в
процессе подкрепления. На фоне подведения норадреналина наблюда­
лось общ ее сниж ение фоновой активности нейрона на всех этапах пищ е­
добывательного поведения (В. Андрианов, 1990)
Глава 3
144
циональной эквипотенциальности корковых структур. Современная
концепция основана на многофункциональности корковых полей.
Считается что правое и левое полушария не равноценны по своему
значению в физиологической и психической деятельности головного
мозга. Выделяют сенсорную, моторную и психическую межполушарную функциональную асимметрию. У большинства людей домини­
рующим является левое полушарие, которое обеспечивает функцию
речи, контроль действий правой руки, вербальное, логическое мыш­
ление. Такой человек тяготеет к теории, имеет большой запас слов,
ему присущи целеустремленность, повышенная двигательная актив­
ность, способность предвидеть события. Правое полушарие головно­
го мозга специализировано для восприятия формы и пространства и
участвует в интуитивном мышлении. Доминирование правого полу­
шария проявляется у человека в конкретных видах деятельности, в
способности тонко чувствовать и переживать.
В системной организации функций оба полушария взаимосодействуют для достижения полезных для организма приспособительных
результатов. Вместе с тем известно, что в функциональном отно­
шении левое и правое полушария отличаются. При этом одно из
полушарий в большинстве случаев доминирует, что проявляется в
психологических свойствах личности.
3.3. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Ф ункции ВНС имеют свои отличительные особенности. Эта сис­
тема:
• регулирует вегетативные (растительные) ф ункции организма
(обмен веществ, трофику, рост тканей и органов);
• влияет на работу внутренних органов, гладкую мускулатуру,
железистый аппарат, кровеносные и лимфатические сосуды;
• обладает длительными и стойкими эффектами;
• отчасти оказывает влияние на деятельность скелетной мускула­
туры и наружных половых органов;
• является автономной, так как ее деятельность лиш ь в слабой сте­
пени регулируется произвольно.
В целостных системных реакциях организма вегетативные функ­
ции всегда тесно взаимодействуют с соматическими. Все это застав­
ляет говорить о вегетативных компонентах целостных приспособи­
тельных реакций.
Нервная система
145
Морфофункциональная организация
вегетативной нервной системы
На основании структурно-функциональных особенностей разли­
чают три отдела ВНС: симпатический, парасимпатический и метасимпатический. Афферентные волокна ВНС несут от периферичес­
ких рецепторов информацию, которая, как правило, не достигает
сознания. Быстродействующие медиаторы, выделяемые из афферен­
тных окончаний, представлены возбуждающей аминокислотой глутаматом и такими пептидами, как ангиотензин II, аргинин-вазопрессин, бомбезин, холецистокинин, вещество П и пр.
Симпатический и парасимпатический отделы ВНС состоят из
центральных и периферических частей (рис. 3.25). Структурно-функциональная характеристика отделов вегетативной, а также сомати­
ческой нервной системы представлена в табл. 3.2.
Эффекторные волокна ВНС иннервируют практически все ткани
и внутренние органы. Часть органов имеет двойную и тройную
иннервацию: симпатическую, парасимпатическую и метасимпатическую. Так, по медиаторному обеспечению мышцы Ж КТ имеют
тройную иннервацию — адренергическую, холинергическую и
серотонинергическую . В последние годы специально выделяют
моноаминергическую систему, которая представляет собой сово­
купность нейронов ствола мозга, посылающих свои аксоны в гипо­
таламус, таламус, лимбические образования и новую кору. Система
эта состоит из серотонинергических, дофаминергических и норадренергических нейронов и, по мнению ряда авторов, обеспечивает
различные эмоциональные состояния за счет тесных контактов с
лимбическими структурами. Причем серотонин участвует в ф ор­
мировании отрицательных эмоциональных состояний, а норадре­
налин — положительных.
В отличие от соматической нервной системы, которая построена
по сегментарному принципу, ВНС не имеет сегментарной органи­
зации. Нейроны ВНС располагаются в специальных отделах ЦНС,
образуя так называемые вегетативные центры.
Центры вегетативной нервной системы
Центры ВНС располагаются на спинальном, бульбарном, мезэн­
цефалическом уровнях, в гипоталамусе, лимбической системе, моз­
жечке и коре большого мозга. В основе их взаимодействия лежит
принцип иерархии. Гипоталамус управляет всеми вегетативными
146
Глава 3
Рис. 3.25. Строение симпатического (красный) и парасимпатического
(зеленый) отделов вегетативной нервной системы (ВНС)
Нервная система
147
Таблица 3 .2 . Сравнительная характеристика структурно-функциональных
особенностей вегетативной и соматической нервной системы
Приз­
нак
55
S
Я
rS
D-
5 8
О
Ч
cd
н
m
1 s
& У
м В*
Л
а
03
°п
X
S
S
!=Г
ей
CQ
Он
0)
Д
I
в н е
парасимпати­
ческая
метасимпатическая
Задний гипота­
ламус, нейроны
боковых рогов
грудных и пояс­
ничных сегмен­
тов С П И Н Н О Г О
мозга
Передний гипо­
таламус, сред­
ний мозг, мост,
продолговатый
мозг, боковые
рога I—V крест­
цовых сегментов
спинного мозга
Стенки полых
внутренних ор­
ганов
Сероевещество
спинного мозга
(вставочные и
д в и га тел ь н ы е
нейроны)
Прерываются в
ганглиях; преганглионарные
короткие, тип
В, постганглионарные длин­
ные, тип С
Прерываются в
ганглиях; преганглионарные
длинны е, тип
В, постганглионарные корот­
кие, тип С
Прерываются в
пределах орга­
на; пре- и постганглионарные
короткие, тип
В и С, обиль­
но ветвятся в
толще органов
Аксоны дли н ­
ные, на протя­
жении не пре­
рываются, тип
А
Диффузная (ин­
нервирует все
внутренние
органы)
Ограничена
(нет в надпочеч­
никах и стенках
большинства
сосудов)
Строго огра­
ничена только
теми органами,
которые имеют
собственную
моторную ак­
тивность (серд­
це, кишечник,
мочеточник,
матка)
Сегментарная
Паравертебральные (погра­
ничный столб),
превертебральные (чревное,
солнечноеспле­
тение, брыжееч­
ные узлы)
В иннервиру­
емых органах
(интрамурально) или рядом
с ними
Только в орга­
нах
Нет эффекторных ганглиев
Ацетилхолин и
норадреналин
Ацетилхолин
Ацетилхолин,
норадреналин,
ВИП, АТФ,
АДФ, аденозин
Только ацетил­
холин
S
аЗ
Д
О
го
о?
S
д
СЗ
со
S
1=5
аЗ
О
Он
о
н
СЗ
S
п
<D
Соматическая
нервная система
симпатическая
148
Глава 3
функциями организма. Он получает информацию как по афферен­
тным путям спинного мозга от различных периферических рецеп­
торов, так и через кровь. Известно, что при раздражении передней
группы ядер проявляются парасимпатические реакции, а задней —
симпатические. В гипоталамусе находятся центры биологических
мотиваций, голода, страха, имеющие в своих проявлениях сильные
вегетативные реакции. Лимбическая система теснейшим образом свя­
зана с деятельностью внутренних органов, за что и получила назва­
ние висцерального мозга. Через контроль вегетативных реакций
лимбическая система участвует в формировании различных эмоци­
ональных состояний. Вовлечение мозжечка в регуляцию внутренних
органов определяется его способностью оказывать активирующие и
тормозные влияния на их деятельность. Именно мозжечок ответс­
твенен за вегетативное обеспечение поведения. В латеральной части
задневентрального ядра таламуса представлен основной коллектор
импульсов, идущих от брюшных органов, — чревный нерв, а в его
медиальной части — блуждающий нерв. Соматосенсорные области
коры головного мозга S I и S II являются представительством чревно­
го нерва, а зоны S II и передней части крестовидной борозды — тазо­
вого. В височной коре представлены моторные центры регуляции
сердца и органов брюшной полости. Высшим центром автономной
иннервации считается лобная кора. Регулируемые ею вегетативные
функции в полном объеме проявляются в процессах обучения и осу­
ществления сложных двигательных актов.
Условно выделяемые «нижние этажи» этой иерархии, обладая доста­
точной автономностью, осуществляют местную регуляцию физиоло­
гических функций. Каждый более высокий уровень регуляции обес­
печивает более высокую степень интеграции автономных функций.
Тонус центров вегетативной нервной системы
Центры ВНС находятся в постоянном тоническом напряжении,
обусловленном спонтанной активностью их нейронов, афферент­
ными влияниями со стороны соматической или самой ВНС или
прямым действием на нейроны биологически активных гумораль­
ных веществ, в частности гормонов, олигопептидов и витаминов.
Наличие постоянных тонических влияний симпатической нервной
системы демонстрирует так называемый синдром Горнера. После
перерезки постганглионарных симпатических волокон верхнего и
среднего шейного симпатического узла у животных на стороне пере­
резки наблюдается эндофтальм, сужение зрачка и расслабление тре­
Нервная система
149
тьего века. При низкочастотном электрическом раздражении перифе­
рического конца перерезанного симпатического нерва эти явления
исчезают.
Вегетативные показатели
Вегетативный тонус представляет собой относительно устойчивую
характеристику состояния вегетативных показателей. Количественно
его можно определить несколькими способами. Во-первых, по ф ор­
муле Кердо. Индекс Кердо позволяет оценить состояние вегетативного
тонуса по параметрам, характеризующим состояние сердечно-сосу­
дистой системы (величине АД и ЧСС):
ВИК= (1 - АДд/ЧСС) х 100%,
где ВИК — вегетативный индекс Кердо, %; АДд— величина диасто­
лического АД, мм рт. ст.; ЧСС — частота сердечных сокращений.
Оценка результата : ВИК от —10 до + 10% — вегетативное равно­
весие; ВИК более + 10% — симпатикотония; ВИК менее — 10% —
ваготония.
Во-вторых, вегетативный тонус определяют методом кардиоинтервалометрии с последующим расчетом индекса напряжения по
формуле P.M. Баевского (1992). Индекс напряжения отражает состояние
вегетативного гомеостазиса организма:
ИН = AM ,/2M 0 ДХ,
где ИН — индекс напряжения, уел. ед.; М0 — мода — наиболее часто
встречающееся в ЭКГ значение интервала R—R, с; АМ0 — ампли­
туда моды — наиболее часто встречающиеся в ЭКГ интервалы R—R
(в процентах от общего количества в выборке), характеризует тонус
симпатического отдела ВНС; ДХ — вариационный размах — разность
между минимальными и максимальными значениями кардиоинтер­
валов, характеризует тонус парасимпатического отдела ВНС.
Оценка результата: ИН =50—200 уел. ед. — нормотония; ИН менее
50 усл.ед. — ваготония; ИН = 200-500 усл.ед. — симпатикотония; ИН
свыше 500 уел. ед. — гиперсимпатикотония.
Вегетативная реактивность — величина и направленность реакций
ВНС на внешние или внутренние раздражители.
Вегетативное обеспечение — вегетативное сопровождение различ­
ных форм деятельности.
150
Глава 3
Общие свойства вегетативной нервной системы
По сравнению с соматической нервной системой ВНС характери­
зуется некоторыми особенностями.
Волокна ВНС обладают:
• меньшей возбудимостью;
• удлиненной фазой абсолютной рефрактерности нервных волокон;
• меньшей функциональной лабильностью (не более 15 имп./с);
• меньшей скоростью проведения возбуждения: преганглионарные
волокна группы В со скоростью 3—20 м/с и постганглионарные
волокна группы С со скоростью 0,5—3 м/с;
• длительным потенциалом действия за счет следовых фаз
(до 150 мс).
Ганглиям ВНС свойственны:
• временная и пространственная суммация возбуждений;
• процессы конвергенции, окклюзии, мультипликации;
• синаптическая задержка от 1 до 25 мс;
• низкая фоновая импульсная активность ганглионарных нейро­
нов (не более 15 имп./с);
• высокая чувствительность ганглионарных нейронов к адренали­
ну крови, выступающему модулятором синаптической передачи.
Особенности химической организации вегетативной
нервной системы
ВНС осуществляет свои регулирующие влияния главным образом
путем химических медиаторных и рецепторных механизмов (рис. 3.26).
На основании различий в химических свойствах медиаторов и рецепто­
ров ВНС может быть разделена на холинергическую, норадренергическую, серотонинергическую, дофаминергическую и пуринергическую.
М едиаторно-рецепторные соотношения в вегетативной нервной системе
Эффекты ВНС зависят как от характера выделяющегося медиа­
тора, так и от соответствующего рецептора в эффекторном органе.
Кроме постсинаптических рецепторов, выделяют пресинаптические
и внесинаптические рецепторы. Первые из них регулируют выде­
ление медиатора по принципу обратной связи, а вторые — путем
взаимодействия с различными биологически активными вещест­
вами, поступающими в околоклеточное пространство. Воздействия
различных отделов ВНС на функции органов показаны в табл. 3.3.
В разных тканях имеются различные рецепторы к медиаторам ВНС.
Нервная система
151
Рис. 3.26. Схема выделения медиаторов в вегетативной нервной системе:
А — первым и вторым нейронами парасимпатической нервной систе­
мы; Б — первым и вторым нейронами симпатической нервной системы;
АХ — ацетилхолин; НА — норадреналин; Н-ХР — ни котин-холи норе цептор; М-ХР — мускарин-холинорецептор; а - и (3-АР — адренорецепторы
Согласно общепринятой классификации, все холинорецепторы
делятся на мускарино- (М) и никотиночувствительные (Н), а адрено­
рецепторы — на а - (а,- и а 2-) и р- (Р,- и Р2-) адренорецепторы. Данные
о локализации холино- и адренорецепторов в различных структурах
организма представлены в табл. 3.3.
Показано, что перерезка симпатических нервов приводит к повы­
шению чувствительности соответствующих тканей к циркулирующе-
Глава 3
152
Таблица 3.3. Рецепторы и эффекты холинергических и адренергических вли­
яний вегетативной нервной системы
Холинергические
эффекты
Эффекторный
орган
тип
рецептора
характер
ответа
Норадренергические
эффекты
тип
рецептора
характер
ответа
а
Сокращение (мидриаз)
р
Расслабление
для дальнего
видения
Глаз
• радиальная
мышца радуж­
ки
• сфинктер
радужки
М
м
Сокращение
(миоз)
• цилиарная
мышца
м
Сокращение для
ближнего виде­
ния
• синоатри­
альный узел
м
р.
Учащение сердце­
биения
• предсердия
м
Вагусный эффект.
Урежение сердце­
биений
Снижение сокра­
тимости и прово­
димости
р,
Снижение
проводимости,
атриовентрику­
лярная блокада
р,
Возрастание про­
водимости и сокра­
тимости
Возрастание
проводимости
Сердце
• атриовент­
рикулярный
узел и прово­
дящая система
• миокард
м
р2
Возрастание сокра­
тимости и прово­
димости
а
Сужение.
Расширение
Сосуды
Артериолы
• коронарные,
поперечнопо­
лосатых мышц,
легочные,
абдоминаль­
ные, внутрен­
них органов,
почек
м
Расширение
Р2
Нервная система
153
Продолж ание табл. 3.3
• кожи и сли­
зистых обо­
лочек, мозга,
слюнных
желез
Вены
а
Сокращение
а
Р2
Сужение.
Расширение
Легкие
•бронхиаль­
ная мускула­
тура
М
Сокращение
Р2
Расслабление
•бронхиаль­
ные железы
м
Стимуляция
Р2
Торможение
«1. Р2
Ослабление
Ж елудок
•сокращение
и тонус
•сфинктеры
Усиление
м
•секреция
Расслабление
а.
Сокращение
Усиление
Р2
Торможение
Усиление
а,, Р2
Ослабление
Кишечник
•сокращение
и тонус
• сфинктеры
м
•секреция
Желчный
пузырь и про­
токи
Расслабление
а1
Сокращение
Усиление
м
Сокращение
Р2
Р2
Торможение
м
Сокращение
Р
Расслабление
Расслабление
а
Сокращение
Усиление
а
Усиление
Расслабление
Мочевой пузырь
• сжимающая
мышца
•треугольник
и сфинктер
Мочеточник
подвижность и
тонус
м
Зависит от поло­
вого цикла
М атка
Половой член
м
Эрекция
Р2>ai
а
Зависит от поло­
вого цикла
Эякуляция
Глава 3
154
Окончание табл. 3.3
Кожа
•пиломотор­
ные мышцы
• потовые
железы
М
Генерализованная
секреция
С елезенка
М озговой слой
надпочечников
м
Сокращение
а
Слабая, локализо­
ванная секреция
Р2
Сокращение.
Расслабление
а |? р?
Гликогенолиз
Секреция адрена­
лина и норадре­
налина
Печень
П одж елудоч­
ная железа
а
м
•ацинусы
•островки
Увеличение сек­
реции
Увеличение сек­
реции инсулина
и глюкагона
а
а
Слюнные
железы
м
Профузная сек­
реция
Слезные
железы
м
Секреция
Н осоглоточ­
ные железы
м
Секреция
а
Снижение секре­
ции
Уменьшение сек­
реции инсулина
и глюкагона.
Увеличение секре­
ции инсулина
и глюкагона
Слабая вязкая сек­
реция. Секреция
амилазы
Жировая
ткань
Р,
Липолиз
Ю кстагломерулярные
клетки
Р,
Увеличение секре­
ции ренина
Ш ишковид­
ная железа
Р
Ганглии ВНС
н
Синаптическая
передача
Увеличение син­
теза и секреции
мелатонина
Активация синап­
Нрецепто- тической передачи
в нейронах ганг­
ры
лиев вне
Нервная система
155
му в крови иорадреналину, а парасимпатических структур — к ацетилхолину (У. Кэннон и А. Розенблют). Это происходит главным образом
за счет уменьшения содержания в тканях деградирующих фермен­
тов — аминооксидазы и ацетилхолинэстеразы соответственно.
Формы вегетативного влияния.
• Изменение функциональной активности органа.
• Изменение кровотока в регулируемом органе посредством изме­
нения просвета сосудов.
• Изменение трофических процессов в иннервируемом органе.
Парасимпатический отдел
Морфология парасимпатического отдела
Тела преганглионарных нейронов парасимпатического отдела ВНС
расположены в среднем и продолговатом мозге среди моторных нейро­
нов окуломоторного, лицевого, языкоглоточного и блуждающего нервов,
а также в сером веществе крестцового отдела спинного мозга.
Вместе с черепными нервами парасимпатические преганглионарные волокна распространяются к ганглионарным нейронам, распо­
ложенным в цилиарном, крылонебном, подчелюстном и ушном ган­
глиях. Отходящие от них постганглионарные волокна иннервируют
глазное яблоко, аккомодационную мышцу глаза и сфинктер зрачка,
подчелюстную, подъязычную и околоушную слюнные железы.
Преганглионарные волокна блуждающего нерва распространяют­
ся к нейронам, расположенным в ганглиях органов грудной и верхней
части брюшной полости.
Аксоны нейронов крестцового отдела спинного мозга через тазо­
вые нервы адресуются к нейронам, располагающимся в ганглиях
органов нижней части брюшной полости, таза и в наружных половых
органах. В отличие от симпатических постганглионарных волокон,
волокна парасимпатической нервной системы не иннервируют глад­
кие мышцы кровеносных сосудов, за исключением сосудов половых
органов, артерий мозга, слюнных желез и брыжейки.
В результате особенностей строения симпатические нервы пред­
ставлены в основном постганглионарными, а парасимпатические —
преганглионарными нервными волокнами. Преганглионарные волок­
на симпатической и парасимпатической нервной системы образованы
слабомиелинизированными волокнами типа В. Постганглионарные
волокна — «серые», представлены безмиелиновыми волокнами
типа С.
156
Глава 3
Физиологические особенности парасимпатического
отдела
Процессы иррадиации и мультипликации парасимпатических
возбуждений наблюдаются в ганглиях, расположенных в стенке
или около иннервируемого органа, поэтому их влияние местное.
Вследствие этого парасимпатическая нервная система оказывает
ограниченное воздействие в пределах иннервируемых органов.
Парасимпатическая нервная система оказывает в основном
успокаивающее, расслабляющее действие на функции организма.
Парасимпатические влияния проявляются преимущественно в ослабле­
нии работы внутренних органов, снижении возбудимости ЦНС, умень­
шении интенсивности метаболизма, снижении возбудимости миокарда,
уменьшении силы сердечных сокращений и ЧСС, снижении кровяного
давления, уменьшении объема легочной вентиляции и температуры
тела, увеличении секреции инсулина и, как следствие, снижении кон­
центрации глюкозы в крови и увеличении внутриклеточного депони­
рования ее в виде гликогена. При этом одновременно усиливаются
моторная, секреторная и всасывательная функции ЖКТ.
Парасимпатическая нервная система представлена на периферии
в основном преганглионарными волокнами типа В. Вследствие этого
возбуждения по парасимпатической нервной системе распространя­
ются быстрее, чем по симпатической нервной системе.
Медиатором в окончаниях преганглионарного и постганглионарного нейронов служит ацетилхолин. Холинергические влияния ВНС
обычно дискретны и кратковременны, потому что ацетилхолин быс­
тро разрушается в постсинаптических окончаниях имеющейся там в
избытке ацетилхолинэстеразой. Однако при определенных условиях
ацетилхолин, выделяемый нейронами ВНС, может появляться в
крови. Так, если заблокировать эзерином холинэстеразу в подчелюс­
тной слюнной железе, то при раздражении нерва барабанной стру­
ны можно наблюдать не только секрецию подчелюстной слюнной
железы, но и брадикардию, падение кровяного давления, урежение
дыхания, торможение моторной деятельности Ж КТ (В.П. Бабкин).
В синапсах, образуемых постганглионарными холинергическими
нейронами на органах, функционирует эффективный механизм кон­
троля выделения медиатора (рис. 3.27).
Эффекты ВНС пролонгируются вторичными влияниями гормонов
желез внутренней секреции, которые иннервируются той же ВНС.
Полагают, что парасимпатическая нервная система усиливает
преимущ ественно анаболические реакции. Парасимпатические
Нервная система
157
Р и с. 3 .2 7 . М еханизм
п ресин аптич еск ой
регуляции
вы свобож де­
ния медиаторов нейронам и парасимпатической нервной системы:
АХ — ацетилхолин; НА — норадреналин; а-А Р — адренорецептор;
Н-ХР — никотин-холинорецептор; М-ХР — мускарин-холинорецептор;
ГЦ — гуанилатциклаза; ГТФ — гуанозинтрифосфат; цГМФ — цикличес­
кий гуанозинмонофосфат
влияния определяют состояние сна и любого удовлетворения.
В парасимпатической системе медиатором в преганглионарных и
постганглионарных волокнах служит ацетилхолин. Эффекты пара­
симпатической нервной системы по сравнению с симпатической
менее продолжительны, поскольку действие ацетилхолинэстеразы
более сильное, чем действие моноаминоксидазы и катехолортометилтрансферазы — ферментов, разрушающих медиаторы симпатической
нервной системы в иннервируемых органах.
Симпатический отдел
Морфология симпатического отдела
Тела преганглионарных нейронов симпатического отдела BHC
располагаются в сером веществе боковых рогов спинного мозга,
158
Глава 3
начиная с I грудного сегмента и до II—III поясничных сегментов.
Миелинизированные аксоны этих нейронов представлены медлен­
но проводящими возбуждения В-волокнами. Аксоны преганглио­
нарных нейронов симпатической нервной системы через синапсы
взаимодействуют с нейронами, расположенными в периферических
ганглиях.
Околопозвоночные (вертебральные) ганглии расположены по обе
стороны позвоночника в виде цепочек, составляющих правый и левый
симпатический ствол. Продольно ганглии соединяются межганглиевыми соединительными ветвями, состоящими из миелиновых и
безмиелиновых нервных волокон. Имеются и поперечные связи между
ганглиями правой и левой сторон. Кроме того, ганглии симпатическо­
го ствола соединяются со спинномозговыми нервами, благодаря чему
они получают возможность иннервировать скелетную мускулатуру.
Часть постганглионарных волокон направляется к внутренним орга­
нам. Определенное количество преганглионарных волокон, выходя­
щих из центров спинного мозга, проходит через вертебральные ган­
глии, не прерываясь, и заканчивается в превертебральных ганглиях.
Последние расположены на большем, чем паравертебральные, рассто­
янии от позвоночника и, вместе с тем, на значительном отдалении от
иннервируемых органов, например, сердца, желудка и др.
Вегетативные ганглии, подобно ЦНС, защищены от воздействия
биологически активных веществ, циркулирующих в крови, барьером,
сходным с гематоэнцефалическим.
Каждый преганглионарный аксон разветвляется и контактирует с
10—30 ганглионарными нейронами.
Аксоны ганглионарных нейронов симпатической нервной систе­
мы представлены немиелинизированными С-волокнами.
Существует три варианта передачи симпатических влияний на
органы: прямой контакт медиатора с клеткой органа; действие меди­
атора через кровь на клетки органа; влияние медиатора на нейроны
метасимпатической нервной системы.
Физиологические основы симпатического отдела
Из симпатических ганглиев возбуждения широко распростра­
няются сразу на несколько органов. В результате разветвленных
влияний на иннервируемые органы симпатическая нервная система
оказывает на них и на организм в целом генерализованное воздейс­
твие. Симпатическая нервная система представлена на периферии в
основном постганглионарными волокнами типа С, вследствие этого
Нервная система
159
возбуждение по симпатической нервной системе распространяется
медленнее, чем по парасимпатической нервной системе.
Эффекты симпатической нервной системы по сравнению с пара­
симпатической более продолжительны. Электрические потенциалы
в симпатических ганглиях характеризуются продолжительными сле­
довыми явлениями.
При стимуляции преганглионарных волокон в симпатических
ганглиях сначала регистрируется быстрый возбуждающий постсинаптический потенциал, который часто сменяется медленным тормоз­
ным постсинаптическим потенциалом. Вслед за этим снова регист­
рируется медленный возбуждающий постсинаптический потенциал.
Медленный возбуждающий постсинаптический потенциал продол­
жается минуты.
В симпатической нервной системе передача сигнала с пре- на
ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — посредством норадреналина. При этом на
норадреналин приходится 90%, 7% — на адреналин и 3% — на
дофамин. Исключение составляют постганглионарные симпати­
ческие волокна, идущие к потовым железам и отдельным сосудам,
в окончаниях которых выделяется ацетилхолин. В последние годы
получены данные о существовании серотонинергических нервных
волокон, идущих в составе симпатических стволов и усиливающих
сокращения гладких мышц ЖКТ. Наряду с норадреналином из
симпатических окончаний выделяются АТФ, а также нейропептиды
соматостатин и нейропептидУ.
Установлено, что начальная деполяризация обусловлена действи­
ем ацетилхолина на никотиновые рецепторы. Медленный тормоз­
ной постсинаптический потенциал определяется действием дофа­
мина, который секретируется малыми интернейронами ганглия.
Интернейроны, в свою очередь, активируются через мускариновые рецепторы. Поскольку медленный тормозной постсинаптичес­
кий потенциал не зависит от действия циклического АТФ, можно
думать, что он определяется (3-,-рецепторами. 80% выделившегося
в синаптическую щель норадреналина захватывается обратно пресинаптической мембраной и поступает в синаптические пузырьки.
Остальной медиатор разрушается ферментами — моноаминооксидазой и катехолортометилтрансферазой. Считается, что а,- и р^рецепторы постсинаптической мембраны активируются норадреналином.
а 2- и р2-рецепторы находятся на пресинаптических окончаниях постганглионарного волокна и выполняют функцию контроля выделения
160
Глава 3
синапсом медиатора (рис. 3.28). Кроме того, эти рецепторы преиму­
щественно активируются циркулирующим в крови адреналином.
Симпатическая нервная система оказывает преимущественно
активирующие влияния на функции иннервируемых органов: уси­
ливаются сердечные сокращения и увеличивается ЧСС; повышает­
ся АД; улучшается оксигенация тканей; усиливается метаболизм;
нарастает содержание глюкозы в крови; увеличивается скорость про­
ведения возбуждения в скелетных мышцах и повышается их тонус;
расширяются бронхи; увеличивается объем легочной вентиляции;
расширяются зрачки; увеличивается секреция катехоламинов надпо­
чечниками. При этом одновременно снижается тонус пищеваритель­
ного тракта, ослабляются процессы всасывания и ферментативного
расщепления в кишечнике. В целом симпатическая нервная система
усиливает катаболические реакции организма.
Симпатическая нервная система участвует в формировании эмоций
и таких целостных состояний, как агрессия, стресс, болевые реакции.
Рис. 3.28. Механизм пресинаптической регуляции высвобождения меди­
аторов нейронами симпатической нервной системы: АХ — ацетилхолин;
НА — норадреналин; М-ХР — мускарин-холинорецептор; а - и р-АР — адренорецепторы; АЦ — аденилатциклаза; ИФ — инизитолтрифосфат;
АТФ —аденозинтрифосфат
Нервная система
161
Влияние симпатической нервной системы мобилизует организм на борьбу
и бегство, активное взаимодействие с окружающим миром (В. Кэннон).
Адаптационно-трофическая роль симпатической нервной
системы
n ' t 0 ^ * 4 с c-ot-e-e^e>
.
JI.A. Орбели и А.Г. Гинецинский обнаружили, что раздражения
постганглионарных волокон симпатического нерва восстанавливают
сокращение утомленной раздражением двигательного нерва попе­
речнополосатой мышцы. Этот эффект обусловлен прямым действи­
ем симпатической нервной системы на обмен веществ мышечной
ткани и не обусловлен сосудистыми влияниями. Заполнение сосудов
мышцы вазелином не снимает этот эффект (рис. 3.29).
В синаптических окончаниях симпатических нервов предполага­
ют наличие специальных трофогенов. К ним относятся: нуклеотиды,
некоторые аминокислоты, простагландины, катехоламины, серо­
тонин, ацетилхолин, сложные липиды, ганглиозиды, вещество П и
пептид, родственный кальцитонину.
Считают, что адаптационно-трофическое действие оказывают многие
олигопептиды: либерины, статины, энкефалины, эндорфины, брадикинин, нейротензин, холецистокинин, фрагменты АКТГ, окситоцин и др.
Трофическое действие на ткань присуще всем нервам, однако
наиболее заметно проявляется действие симпатической нервной сис­
темы. Трофическая функция ВНС в значительной степени связана с
выделяющимися при раздражении постганглионарных волокон сим­
патического нерва трофогенами: факторами роста нервов, эпителия,
инсулиноподобным фактором, фактором некроза опухолей и т.п.
Мозговой слой надпочечников
Мозговой слой надпочечников представляет собой видоизменен­
ный симпатический ганглий, состоящий из хромаффинной ткани.
Преганглионарные нейроны симпатической нервной системы
образуют на этих клетках возбуждающие холинергические синапсы.
При возбуждении преганглионарных нейронов в кровь выбрасывает­
ся смесь, состоящая на 90% из адреналина и на 10% — из норадре­
налина. Сам адреналин вызывает бинарный эффект — посредством
а 2-адренорецепторов он ингибирует, а посредством р-адренорецепторов — активирует аденилатциклазу, что приводит к накоплению
циклического аденозинмонофосфата (цАМФ). Катехоламины моз­
гового вещества надпочечников можно рассматривать как мета­
болические гормоны, поскольку они усиливают гликогенолиз,
162
Глава 3
А 'J------------------- L
Рис. 3.29. Влияния симпатической нервной системы на силу сокраще­
ния поперечнополосатой мускулатуры (адаптационно-трофическая роль
симпатической нервной системы): А — снижение амплитуды сокращения
скелетной мышцы вследствие утомления, вызванного ее длительным рит­
мическим раздражением (1); Б — восстановление амплитуды сокращения
той же мышцы на фоне присоединившегося раздражения электрическим
током (2) подходящих к ней симпатических волокон
увеличивают высвобождение жирных кислот подкожной жировой
клетчатки и ускоряют расщепление углеводов. Симпатические волок­
на и иннервируемые ими клетки коры надпочечников представляют
собой симпат-адреналовую систему, активирующую и мобилизующую
деятельность целого организма.
Функциональный антагонизм симпатического
и парасимпатического отделов вегетативной
нервной системы
Большинство внутренних органов имеет двойную — симпатичес­
кую и парасимпатическую — иннервацию. В эффекторных влияниях
Нервная система
163
на внутренние органы симпатическая нервная система оказывает
мобилизующее действие. Парасимпатическая нервная система про­
тивостоит влиянию симпатической нервной системы и способствует
восстановлению измененной функции. Влияние симпатического и
парасимпатического отделов ВНС на внутренние органы всегда про­
исходит содружественно. Характер воздействия зависит от состояния
иннервируемой ткани и возбудимости ее рецепторов.
Некоторые органы (например, мочевой пузырь) иннервируются в
основном парасимпатической нервной системой, а потовые железы,
волосковые мышцы кожи, селезенка и надпочечники — симпатичес­
кой. Считают, что мышцы ЖКТ имеют тройную иннервацию — сим­
патическую (адренергическую), парасимпатическую (холинергическую) и серотонинергическую.
Выделяющиеся при возбуждении ВНС медиаторы не только воз­
действуют на постсинаптические мембраны, где они быстро разруша­
ются соответствующими ферментами, но и могут, особенно при бло­
каде этих ферментов, поступать в кровь, оказывая генерализованное
влияние на многие органы, имеющие соответствующие рецепторы.
Н орадреналин имеет более продолжительное действие по
сравнению с ацетилхолином. Ацетилхолин в крови отсутствует.
Норадреналин, адреналин и дофамин содержатся в плазме крови.
Значительное количество норадреналина поступает в кровоток бла­
годаря диффузии из норадренергических нервных окончаний.
Двойственность влияния ВНС на внутренние органы поддержи­
вается вторичными влияниями гормонов-антагонистов желез внут­
ренней секреции. Например, у кошки при виде лающей собаки уси­
ливается секреция как адреналина мозговым слоем надпочечников,
так и инсулина поджелудочной железой.
Взаимодействие между отделами вегетативной нервной
системы
Поддержание оптимального соотношения тонуса симпатической
и парасимпатической систем в процессе регуляции деятельности
внутренних органов обеспечивается взаимодействием данных отде­
лов ВНС на четырех уровнях. Первый уровень включает в себя пос­
тупление возбуждений к синапсам эффекторной клетки. Второй —
реципрокное торможение на уровне нервных терминалей. Третий —
на уровне нейронов вегетативных ганглиев. Четвертый — взаимо­
действие между соответствующими структурами, расположенными
в различных отделах ЦНС. При этом возможна мобилизация сим­
Глава 3
164
патических или парасимпатических влияний в масштабе целого
организма, первые из которых формируют эрготропную, а вторые —
трофотропную регуляцию физиологических функций.
Метасимпатический отдел
Морфология метасимпатического отдела
М етасимпатическая (внутриорганная) часть ВНС имеет ряд
признаков, отличающих ее от других отделов нервной системы
(А.Д. Ноздрачев). К метасимпатической нервной системе относит­
ся комплекс микроганглионарных образований, расположенных в
стенках внутренних органов, обладающих двигательной активностью
(сердца, бронхов, мочевого пузыря, пищеварительного тракта, матки,
желчного пузыря и желчных путей).
Участки метасимпатической нервной системы называются по
месту их расположения: кардиометасимпатический — в сердце, энтерометасимпатический — в кишечнике и т.д. По своей структурной
организации микроганглии внутренних органов подобны ядерным
образованиям ЦНС. М икроганглии изолированы от окружающих
тканей специальными барьерами, их внеклеточное пространство
незначительно. В микроганглиях имеются многочисленные гли­
альные элементы. Структурной основой процессов, протекающих в
метасимпатической нервной системе, являются афферентные, вста­
вочные и эфферентные нейроны, а также генератор спонтанной
импульсной активности. Тела нейронов метасимпатической нервной
системы имеют множество синапсов, а отростки нервных клеток
содержат большое количество пузырьков с медиаторами.
Физиологические особенности метасимпатического отдела
• Метасимпатическая нервная система имеет входы от обоих отде­
лов ВНС. Ганглионарные нейроны метасимпатической нервной
системы имеют рецепторы к ацетилхолину и норадреналину.
• Ганглионарные нейроны осуществляют влияние на иннервиру­
емые ткани с помощью секретируемых ими АТФ и аденозина.
Кроме того, в синапсах постганглионарных волокон выделяются
разнообразные вещества — ацетилхолин, норадреналин, серото­
нин, дофамин, адреналин, гистамин и т.д.
• Физиологическая роль пуринергических нейронов (рецепторы кото­
рых воспринимают АТФ и аденозин) заключается в осуществлении
тормозных влияний, противоположных активирующим влияниям
Нервная система
165
холинергических нейронов. Так, активация пуринергических ней­
ронов вызывает уменьшение тонуса желудка и кишечника.
• Метасимпатическая нервная система иннервирует только те орга­
ны, которые обладают самостоятельной моторной активностью.
М икроганглионарные образования определяют свойство автома­
тизма — способности органов ритмически сокращаться с опреде­
ленной частотой и амплитудой без воздействия извне.
• Деятельность метасимпатической нервной системы еще в мень­
шей степени зависит от соматической нервной системы, чем
деятельность симпатической и парасимпатической систем.
• Метасимпатическая нервная система не находится в антагонис­
тических отношениях с другими отделами нервной системы.
Вегетативные рефлексы
Нейроны ВНС участвуют в рефлекторных реакциях, называемых
вегетативными рефлексами. Вегетативный рефлекс — ответная реак­
ция организма на раздражитель, осуществляющаяся при участии
ЦНС, внутренних органов или кровеносных сосудов. Некоторые
из них используются в качестве функциональных проб: глазо-сердечный рефлекс, дыхательно-сердечный рефлекс, ортостатическая
реакция, кожный дермографизм и т.д. Таким образом, вегетативный
рефлекс — способ, при помощи которого ВНС вовлекается в целос­
тные функции организма.
Вегетативная рефлекторная дуга
ВНС функционирует по рефлекторному принципу. Как и сома­
тическая, вегетативная рефлекторная дуга состоит из трех звеньев,
имеющих более сложное строение, чем рефлекторная дуга сомати­
ческой нервной системы. Она состоит из рецептора, афферентного
волокна чувствительного нейрона, вставочного нейрона, нейрона,
расположенного в структурах мозга и посылающего преганглионарное волокно к вегетативному ганглию, нейрона, тело которого распо­
ложено в ганглии и постганглионарного волокна, идущего к рабочему
эффектору — мышце или железе.
Первое звено вегетативных рефлексов включает в себя термо-,
механо- и хеморецепторы висцеральных органов, воспринимающие,
соответственно, температурные, механические и химические раздра­
жители, а также ноцицептивные сигналы. Характер ответной реак­
ции рецептора определяется силой, конфигурацией и длительностью
раздражителя. Периферические вегетативные афферентные нервные
166
Глава 3
пути составляют 90% волокон блуждающего нерва, 50% волокон
тазового нерва и столько же в симпатических нервных стволах.
Импульсы от рецепторов по афферентным волокнам направляются к
клеткам спинномозговых или периферических ганглиев. В симпати­
ческой нервной системе афферентные волокна начинаются в органах,
а тела, расположенные в симпатических ганглиях, направляют аксо­
ны либо в спинной мозг, либо к вставочным ганглиозным нейронам,
а через них к эфферентным нейронам. В парасимпатической нервной
системе основным коллектором чувствительных волокон является
блуждающий нерв и его сенсорное ядро, а также волокна языкогло­
точного нерва. В некоторых случаях афферентное звено может быть
общим для соматического и вегетативного рефлекса.
Второе — ассоциативное — звено представлено вставочными и
преганглионарными нейронами, расположенными в боковых рогах
спинного мозга, в продолговатом или среднем мозге. Отростки вста­
вочных преганглионарных нейронов выходят из спинного мозга в
составе вентральных корешков, вступают в соматические нервные
стволы и отсюда достигают узлов пограничного симпатического
ствола или парасимпатических узлов в органах, где переключаются
на эффекторные клетки. Третье звено образовано эффекторными
клетками периферического ганглия и эффектором. Следует отме­
тить, что местом переключения импульсов в вегетативном рефлексе
могут быть самые различные уровни ЦНС.
Виды вегетативных рефлексов
Вегетативные рефлексы бывают: сомато-висцеральные, висцеровисцеральные, висцеро-соматические, висцеро-кожные (рис. 3.30).
Рис. 3.30. Вегетативные спинальные рефлексы: А — висцеро-висцераль­
ный, при котором возбуждение одного внутреннего органа или его участка
приводит к изменению работы другого органа или его участка (например:
кишечно-кишечный рефлекс); Б — висцеро-кожный, при котором, напри­
мер, может наблюдаться покраснение кожи или изменение ее тактильной
чувствительности в случаях заболевания внутренних органов; В — висцеро-соматический, при котором возникает защитное напряжение мышц
брюшной стенки в условиях возбуждения спинальных мотонейронов
афферентами от внутренних органов брюшной полости; Г — кожно-висцеральный, который проявляется в изменении деятельности внутренних
органов при тактильных раздражениях кожной поверхности
I
Нервная система
167
168
Глава 3
• Кожно-висцеральный рефлекс заключается в изменении деятель­
ности внутренних органов при активации соматических нервов.
Примером такого рефлекса может служить реакция внутренних
органов при поверхностном кожном массаже.
• Висцеро-висцеральный рефлекс заключается в изменении
деятельности одного внутреннего органа при активации другого
внутреннего органа. Примером такого рефлекса является реф­
лекс Гольца, при котором механическое раздражение брыжейки
кишечника приводит к урежению сердечных сокращений.
• Висцеро-соматический рефлекс проявляется как увеличение
тонуса мышц передней стенки живота под влиянием возбужде­
ний (возможно, патологической природы), исходящих из органа,
расположенного в брюшной полости. Висцеро-соматические реф­
лексы лежат в основе отраженных болей и связанных с ними зон
Захарьина—Геда, при раздражении которых эти боли возникают.
При этом кожные и висцеральные возбуждения конвергируют на
общих нейронах, входящих в состав спиноталамического тракта.
• Висцеро-кожный рефлекс состоит, в частности, в изменении так­
тильной чувствительности участка кожи, который иннервируется
сегментом спинного мозга, куда поступают импульсы от раздра­
жаемого внутреннего органа.
Глава 4
ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА
4.1. ЖИДКИЕ СРЕДЫ ОРГАНИЗМА.
ГОМЕОСТАЗИС
Вода в организме человека составляет около 60% массы тела и
распределена по двум основным водным пространствам: внутрикле­
точному, на которое приходится около 40%, и внеклеточному (20%).
Внутриклеточная жидкость представлена жидкой фазой цитоплазмы
и ядра. Основными компонентами внеклеточной жидкости являю т­
ся плазма, интерстициальная (тканевая, межклеточная) жидкость и
лимфа.
Плазма, лимфа и интерстициальная жидкость образуют внут­
реннюю среду организма, отличающуюся относительным постоянс­
твом своего состава и физико-химических характеристик и созда­
ющую оптимальные условия сущ ествования клеток организма.
Положение о постоянстве внутренней среды организма как условии
его независимого существования было сформулировано физиоло­
гом К. Бернаром. Впоследствии У. Кэннон для обозначения посто­
янства внутренней среды предложил термин «гомеостазис». Говоря о
постоянстве состава, биологических и физико-химических свойств
внутренней среды, следует понимать, что оно является относи­
тельным и динамичным. В настоящее время под гомеостазисом
понимают динамическое постоянство внутренней среды организма в
совокупности с механизмами, обеспечивающими поддержание этого
постоянства; принят также термин «гомеокенезис».
В крови, лимфе и межтканевой жидкости сосредоточены «предконечные» по отношению к метаболическим процессам в тканях
результаты деятельности многочисленных функциональных систем
гомеостатического уровня, такие как ОЦК и количество форменных
элементов, кровяное и осмотическое давление, температура, реакция
среды (pH), уровень питательных веществ, газов, продуктов выделе­
170
Глава 4
ния и т.д. Каждый из этих показателей поддерживается на оптималь­
ном для жизнедеятельности уровне специфической функциональной
системой, благодаря чему надежно обеспечивается обмен веществ
непосредственно в тканях организма.
В результате содружественной деятельности указанных функцио­
нальных систем обеспечивается гомеостазис в целом (рис. 4.1).
Рис. 4.1. Содружественная саморегуляторная деятельность различных
функциональных систем, обеспечивающих гомеостазис
171
Внутренняя среда организма
4.2. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ
ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
Гуморальная регуляция процессов жизнедеятельности, осущест­
вляемая с помощью биологически активных веществ через жидкие
среды организма, является филогенетически наиболее ранней час­
тью единой системы нейрогуморальной регуляции физиологических
функций. К факторам гуморальной регуляции относятся различные
биологически активные вещества: ионы, метаболиты, эйкозаноиды,
гормоны и др.
Гормоны
Гормоны — группа биологически активных веществ, синтезиру­
емых специализированными эндокринными клетками, секретируемых во внутреннюю среду организма (преимущественно в кровь)
и специфически действующих на клетки-«мишени», содержащие
рецепторы к соответствующему гормону.
Следует сказать, что классическое определение понятия «гормон»
становится все более расплывчатым по мере накопления новых
сведений о механизмах гуморальной регуляции. Иногда трудно про­
вести четкую грань между понятиями «гормон» и «медиатор». Так,
например, многие гипоталамические гормоны (рилизинг-гормоны,
АДГ, окситоцин и др.) не только действуют как гормоны, но и
играют роль медиаторов или модуляторов других медиаторных сис­
тем. Катехоламины норадреналин и адреналин рассматриваются как
гормоны, если они вырабатываются в клетках мозгового вещества
надпочечников, или как медиаторы, если они выделяются в симпа­
тических нервных окончаниях. Одни и те же гормоны могут секретироваться в кровь, а могут воздействовать на рядом расположенные
клетки путем диффузии (например, кортизол).
Гормоны регулируют все процессы жизнедеятельности, включая
метаболизм, репродукцию, рост и развитие, гомеостазис, поведение.
Гормональная регуляция — неотъемлемый компонент любой функ­
циональной системы. Выделяют следующие основные типы физио­
логического действия гормонов:
• кинетическое (вызывают определенную деятельность эффекторных органов);
• метаболическое (вызывают изменение обмена веществ, как пра­
вило, путем регуляции активности ферментов);
172
Глава 4
• морфогенетическое (оказывают воздействие на процессы диф­
ференциации тканей и органов, на рост тканей, формообразова­
ние);
• корригирующее (приводят к изменению интенсивности функций
органов и тканей);
• поведенческое (участвуют в формировании поведения, например,
пищевого, питьевого, полового и др.).
Функционально гормоны делят на три группы:
• эффекторные — гормоны, которые оказывают влияние непос­
редственно на клетки, ткани и органы-«мишени»;
• тропные (гландотропные) — гормоны, основной функцией кото­
рых является регуляция синтеза и секреции эффекторных гормо­
нов периферическими железами внутренней секреции (тропные
гормоны аденогипофиза);
• рилизинг- и ингибирующие гормоны — либерины и статины
нейросекреторных клеток гипоталамуса, регулирующие синтез и
секрецию тропных гормонов аденогипофиза.
Особенности гормональной регуляции:
• гормоны обладают высокой биологической активностью, оказы­
вая физиологическое действие при очень малых концентрациях в
крови (10—6—10“12 моль/л);
• гормональные эффекты в регулируемых клетках развиваются
с большим латентным периодом и более продолжительны, чем
нервные регулирующие влияния;
• гормоны, как правило, имеют множественную локализацию мест
синтеза в организме (например, одни и те же гормоны синтезиру­
ются в ЦНС и ЖКТ);
• гормоны оказывают избирательное действие на клетки и органы«мишени», обладающие соответствующими рецепторами;
• гормоны обладают вариабельностью действия в различных тка­
нях и органах, обусловленной полиморфизмом рецепторов;
• гормоны оказывают генерализованное действие, обеспечивая
согласованность физиологических процессов в организме.
Стероидные гормоны и производные аминокислот не имеют
видовой специфичности. Белково-пептидные, как правило, обладают
видовой специфичностью, введение гормонов животного происхож­
дения может вызвать у человека образование специфических антител,
а также аллергические реакции.
■
Внутренняя среда организма
173
Источники гормонов
Гормоны вырабатываются секреторными клетками различного
происхождения и локализации:
• специализированными железистыми клетками — гландулоцитами;
• клетками, сочетающими морфологические признаки нейронов и
гландулоцитов, — нейросекреторными клетками;
• хромаффинными клетками;
• неспециализированными клетками, секретирующими, напри­
мер, тканевые гормоны.
Важнейшим источником гормонов являются нейросекреторные
клетки гипоталамуса, продуцирующие рилизинг- и ингибирующие
гормоны (либерины и статины), эффекторные гормоны (АДГ, окситоцин) и другие биологически активные вещества.
Клетки, продуцирующие гормоны, могут составлять специали­
зированные тканевые образования — железы внутренней секреции,
основной функцией которых является выработка гормонов, а могут
быть распределены в различных тканях и органах, образуя так назы­
ваемую диффузную эндокринную систему.
Железы внутренней секреции подразделяют на центральные,
к которым относят гипофиз и эпифиз, расположенные в пределах
головного мозга, и периферические. Последние, в свою очередь, делят
на гипофиззависимые (кора надпочечников, щитовидная и инкре­
торные клетки половых желез) и гипофизнезависимые (мозговое
вещество надпочечников, паращитовидные, эндокринная часть под­
желудочной железы). Выработка эффекторных гормонов гипофиззависимыми железами внутренней секреции находится под контролем
тропных гормонов аденогипофиза.
Основные морфофункциональные особенности желез внутренней
секреции:
• отсутствие выводных протоков для секретируемых веществ;
• интенсивный метаболизм;
• густая капиллярная сеть.
Поджелудочная и половые железы характеризуются смешанным
типом секреции (экзо- и эндокринные).
Частью диффузной системы являю тся эндокринные клетки
ЖКТ (так называемая энтериновая система), продуцирующие гас­
троинтестинальные гормоны (гастрин, холецистокинин, секре­
тин, ВИП, соматостатин, энкефалин и другие пептиды), многие из
174
Глава 4
которых имеют и другие места синтеза (например, нейроны ЦНС).
Гастроинтестинальные гормоны участвуют в регуляции секреции,
моторики и всасывания в органах пищеварения, обмена веществ,
высвобождения регуляторных пептидов, пищевого поведения и мно­
гих других процессов жизнедеятельности.
К диффузной эндокринной системе относят и секреторные клет­
ки, расположенные в других органах. Кардиомиоциты преимущест­
венно правого предсердия продуцируют ПНУФ (атриопептид), обла­
дающий широким спектром почечных и сосудистых эффектов. В
почках образуются эритропоэтин, стимулирующий кроветворение,
кальцитриол, участвующий в регуляции обмена кальция, ренин —
фермент, являющийся инициативным звеном ренин-ангиотензинальдостероновой системы (РААС). В печени синтезируются соматомедины, предшественник ангиотензина I — белок ангиотензиноген.
Источником ряда биологически активных веществ являются эпите­
лиальные клетки бронхов, легких, жировые клетки и другие ткани.
Классификация гормонов
П о химической структуре принято выделять три класса гормонов:
белково-пептидные, стероидные и производные аминокислот.
Белково-пептидные гормоны : пептиды (кортикотропин, вазопрес­
син); белки (инсулин СТГ); гликопротеиды ТТГ (фоллитропин).
Стероидные гормоны (производные холестерина): глюкокортикоиды, минералкортикоиды, андрогены, эстрогены, прогестерон.
Производные аминокислот : тирозина (дофамин, норадреналин,
адреналин; иодсодержащие гормоны щитовидной железы); трипто­
фана (серотонин, мелатонин); гистидина (гистамин).
Особо следует выделить класс биологически активных веществ,
производных арахидоновой кислоты (эйкозаноиды). К ним относятся про­
стагландины, тромбоксаны, лейкотриены и другие вещества, обла­
дающие широким спектром действия и высокой физиологической
активностью.
Химическая структура определяет особенности синтеза, транс­
порта, механизм взаимодействия с клетками-миш енями, период
полужизни и способ инактивации гормона.
Синтез всех гормонов осуществляется в результате ряда последо­
вательных этапов. Белково-пептидные гормоны являются прямыми
продуктами экспрессии их генов, синтез происходит через прогормо­
ны. Синтез стероидных гормонов и производных аминокислот осу­
Внутренняя среда организма
175
ществляется в результате модификации молекул-предшественников
под действием энзимов.
Секреция гормонов
Пептидные гормоны накапливаются в секреторных гранулах и
выделяются путем экзоцитоза. Выделяют две формы секреции белково-пептидных гормонов: постоянную, так называемую «базальную»,
и вызванную какими-либо дополнительными гуморальными или
нервными влияниями «стимулируемую» секрецию. После адекватно­
го раздражения секретируется путем экзоцитоза небольшое, необхо­
димое для конкретной ситуации, количество гормона.
Катехоламины мозгового вещества надпочечников по мере син­
теза накапливаются в везикулах и выделяются также путем экзо­
цитоза. Тиреоидные гормоны хранятся в клетках в виде капельных
включений.
Стероидные гормоны не накапливаются в секреторных грану­
лах, а секретируются по мере синтеза. Специфическая стимуляция
железы, продуцирующей стероиды, индуцирует синтез активных
форм гормона, которые диффундируют через мембрану клеток во
внутреннюю среду организма.
Ритмы секреции гормонов
Для большинства гормонов не существует стабильного базального
уровня концентрации. Динамика секреции большинства гормонов в
той или иной степени подчиняется определенным ритмам.
Выделяют так называемую эпизодическую, или импульсную,
секрецию, при которой интервалы между пиками секреции состав­
ляют от нескольких минут до часов (например, импульсная секреция
гонадолиберина имеет период около 90 мин). Секреция ряда гормо­
нов имеет выраженную суточную ритмику (мелатонин, кортизол,
СТГ). Например, кортизол максимального уровня в плазме достигает
утром, а за 2—3 ч до этого увеличивается концентрация АКТГ —
тропного гормона гипофиза, стимулирующего синтез и секрецию
кортизола. В реализации циркадианных (близких к суточным) рит­
мов гипофизарных и других гормонов участвует гипоталамус, осо­
бенно супрахиазматические ядра. Циклические изменения содер­
жания в крови у женщин гонадотропинов, ингибинов, эстрогена и
прогестерона имеют периодичность около 28 сут (овариально-менструальный цикл).
176
Глава 4
Циркуляция гормонов в крови
Гормоны циркулируют в крови либо свободно (в основном белково-пептидные гормоны), либо в комплексе со связывающими их
транспортными белками (гормоны щитовидной железы, стероидные
гормоны), либо адсорбированными на мембране клеточных элемен­
тов крови.
Связанный с белками гормон является запасной циркулирующей
формой, так как биологически активен только свободно циркулиру­
ющий гормон. Между этими двумя формами циркуляции гормонов
существует равновесие.
Время полужизни гормона в плазме — время, в течение которого
50% биологически активного вещества удаляется из плазмы. Этот
процесс является следствием метаболизма, выделения и интернали­
зации гормона клеткой-«мишенью».
Гормональный метаболизм осуществляется специфическими фер­
ментными системами. Белково-пептидные гормоны быстро разру­
шаются пептидазами и в целом имеют короткое время полужизни в
крови (от нескольких минут до часов). Благодаря наличию связи с
транспортными белками, предотвращающей быстрое расщепление
гормонов, время полужизни стероидных гормонов составляет часы,
а гормонов щитовидной железы — дни.
Экскретируются гормоны в основном (до 90%) уже в инактивиро­
ванной форме. Большая часть гормонов (до 80%) выделяется почка­
ми, некоторое количество выводится печенью с желчью и далее через
ЖКТ, а также в составе пота и слюны.
Внутриклеточное взаимодействие гормонов
Синергизм — однонаправленное действие гормонов. Например,
адреналин и глюкагон являются синергистами в регуляции уровня
глюкозы, стимулируя распад гликогена и приводя к повышению
содержания глюкозы в крови.
Антагонизм — противоположно направленное действие гормонов.
Например, глюкагон и инсулин по отношению к содержанию глю­
козы в крови.
Пермиссивный эффект. Гормон, сам не вызывая физиологического
эффекта, создает условия для действия других гормонов на клет­
ки-миш ени». Например, глюкокортикоиды значительно повышают
чувствительность адренорецепторов к катехоламинам.
Внутренняя среда организма
177
Рис. 4.2. Основные формы передачи сигнала: А — эндокринная; Б — паракринная; В — аутокринная
Основные формы передачи сигнала
Эндокринная — сигнальное вещество (гормон) секретируется в кровь
и через кровь достигает отдаленных клеток-«мишеней». Эндокринная
форма передачи сигнала характерна для большинства гормонов эндок­
ринных желез, диффузно расположенных эндокринных клеток, ней­
росекреторных и хромаффинных клеток (рис. 4.2, А).
Паракринная — секретируемое клеткой в межклеточное про­
странство биологически активное вещество действует путем диф ­
фузии на соседние клетки. Паракринная форма передачи сигнала
характерна, например, для некоторых гастроинтестинальных гор­
монов, регулирующих функции системы пищеварения, тканевых
гормонов (гистамин) и других биологически активных веществ
(см. рис. 4.2, Б).
Аутокринная — синтезируемое и секретируемое клеткой биологичес­
ки активное вещество действует на саму клетку. Аутокринная передача
сигналов показана, например, для некоторых пептидов ЖКТ, которые
могут влиять на свою собственную секрецию (см. рис. 4.2, В).
Взаимодействие гормонов
с клетками-«мишенями»
Биологически активные вещества оказывают влияние на свои
клетки-«мишени» в результате взаимодействия со специфическими
рецепторами, связывающими только определенные лиганды (лиган­
178
Глава 4
дами называют все специфически связывающиеся с рецепторами
вещества). Взаимодействие лиганда со своим рецептором и образо­
вание лиганд-рецепторного комплекса является ключевым иници­
ативным моментом, индуцирующим проявление физиологического
эффекта. В зависимости от расположения в клетке-«миш ени» рецеп­
торы подразделяются на мембранные и внутриклеточные (цитозоль­
ные и ядерные).
Мембранная рецепция
С мембранными рецепторами взаимодействуют белково-пептидные гормоны, катехоламины и другие нейротрансмиттеры.
Среди мембранных рецепторов можно выделить ионотропные,
представляющие собой трансмембранные белки, сочетающие свойс­
тва рецептора и ионного канала. Взаимодействие нейротрансмиттера
с этими рецепторами непосредственно приводит к активации ион­
ного канала в составе рецептора и возникновению лигандиндуцированных ионных токов. Следует отметить, что данный тип рецеп­
торов наиболее задействован в механизмах синаптической передачи,
например, Н-холинорецепторы нервно-мышечных синапсов или
вегетативных ганглиев.
Взаимодействие лигандов с другим типом рецепторов (метаботропные рецепторы) индуцирует активацию характерного для этих
рецепторов каскада посредников (вторичных мессенджеров).
К основным группам метаботропных рецепторов могут быть отне­
сены каталитические рецепторы и рецепторы, связанные с G -белка­
ми (ГТФ-связывающ ие белки).
Каталитические рецепторы — трансмембранные белки, наружная
часть которых связывает лиганд, а цитоплазматическая часть либо
сама является активным центром фермента, ли бо связана с молеку­
лой фермента. Например, рецепторы П Н У Ф (рецепторные гуанилатциклазы), рецепторы инсулина (рецепторные тирозинкиназы).
Мембранные рецепторы, связанные с G -белками, составляют м но­
гочи слен ное семейство, в котором более 500 представителей.
К ним относятся а - и p-адренергические, мускариновые холинергические, серотонинергические, аденозиновые, рецепторы
больш инства пептидных гормонов и др. Д ля м ногих лигандов
(например, ацетилхолина, адреналина, норадреналина, серотони­
на) показано наличие разных подтипов связанных с G -белками
рецепторов.
Внутренняя среда организма
179
Эти рецепторы представляют собой полипептидные цепочки,
пронизывающие мембрану 7 раз, образуя по 3 вне- и внутриклеточ­
ных петли. Внеклеточный фрагмент содержит участок, связываю­
щий лиганд, а одна из расположенных в цитозоле петель связывает
G -белок. G -белки служат посредниками между рецепторами плаз­
матической мембраны и внутриклеточными процессами, которые
контролируют биологически активные вещества, взаимодействую­
щие с рецепторами.
Связывание регуляторного вещества с его рецептором активи­
рует G -белок, а тот, в свою очередь, взаимодействует с какой-либо
из систем вторых посредников (ц А М Ф , цГМ Ф , инозитолтрифосфат,
диацилглицерин, Са2+ и др.), активируя или ингибируя фермент или
ионный канал.
Одним из основных путей передачи сигнала, опосредованного
G -белкам и, является регуляция активности аденилатциклазы,
фермента, отвечающего за синтез второго посредника — цА М Ф .
Активация или ингибирование аденилатциклазы приводит соответс­
твенно к возрастанию или снижению внутриклеточного содержания
цА М Ф , что, в свою очередь, влияет на активность цАМ Ф -зависимы х
протеинкиназ. Вследствие этого изменяется уровень фосфорилирования различных белков (преимущественно ферментов), обусловли­
вающих конечный физиологический ответ клетки (рис. 4.3, А).
Некоторые G -белки регулируют работу ионных каналов как опос­
редованно (через зависимые от вторых посредников протеинкиназы),
так и путем прямого взаимодействия с каналами. Например, связыва­
ние ацетилхолина с мускариновыми (М 2) рецепторами пейсмекерных
клеток миокарда ведет к активации G -белка, его взаимодействию с
К+-каналами и повышению проницаемости мембраны атипичных
кардиомиоцитов для К +.
В функциональном отношении G -белки разделяют на активаторы
аденилатциклазы, ингибиторы аденилатциклазы, активаторы фосфолипазы, активаторы ионных каналов (К +; Са2+ ) и др.
Общим свойством систем вторых посредников является м ного­
кратное усиление сигнала, благодаря чему гормоны оказываются
эффективными, находясь в крови в очень низких концентрациях.
Внутриклеточная рецепция
Принципиальным отличием взаимодействия с клетками-«миш енями» стероидных и иодсодержащих гормонов щитовидной железы
Глава 4
180
>еяковьн
гормон
Рецептор
G -белок
Клеточная
мембрана
Аденилат-циклаза
Белковый синтез
i
Изменение функции клетки
Цитоплазма
Б
Цитоплазма
Изменение функции клетки
Рис. 4.3. Мембранная (А ) и внутриклеточная (Б ) рецепция гормонов
Внутренняя среда организма
181
является их проникновение внутрь клетки. Полагают, что стероид­
ные гормоны, будучи неполярными веществами, свободно проходят
через мембрану. Наряду с этим, имеются данные о существовании
мембранных рецепторов, взаимодействующих со стероидами, обес­
печивающих узнавание гормона и его транспорт в клетку.
В цитоплазме гормоны связываются с цитозольным рецептором , то
необходимо для поступления гормона в ядро, где он образует гормонрецепторный комплекс с ядерным рецептором. Этот активированный
комплекс, в свою очередь, приводит к активации транскрипции
матричной Р Н К и синтеза транспортной Р Н К и, в конечном итоге,
влияет на синтез белков и ферментов, обеспечивающих клеточный
ответ (см. рис. 4.3, Б).
Транспорт гормонов щ итовидной железы [трииодтиронина
(ТЗ) и тироксина (Т4)] кровью осущ ествляется с помощью спе­
циальных белков (глобулинов и альбуминов). П осле связывания
с рецептором на поверхности мембраны происходит диссоциация
транспортного гормон-белкового комплекса. Гормоны щ итовид­
ной железы транспортируются через клеточную мембрану с помо­
щью активного переносчика. Внутри клетки Т4 деиодируется до
ТЗ, после чего связывается с белковыми ядерными рецепторами.
Активируемый с помощью ТЗ рецептор является важным регулято­
ром транскрипции, модулируя таким образом синтез больш ого числа
белков.
Регуляция функции рецепторов
Активность всех типов рецепторов не является неизменной, регу­
ляция рецепторной функции осуществляется путем изменения п лот­
ности и аффинности специфических рецепторов.
Количество рецепторов в клетках-«миш енях» не остается посто­
янным. Рецепторы инактивируются или разрушаются в процессе
функционирования, реактивируются и постоянно синтезируются.
Ч исло рецепторов и их чувствительность к лигандам изменяются
в зависимости от концентрации действующего вещества, продолжи­
тельности воздействия и других регуляторных влияний (например,
других гормонов). Недостаток биологически активного вещества при­
водит к увеличению числа активных рецепторов. При избыточном
количестве гормона или нейромедиатора число действующих рецеп­
торов или их чувствительность к лигандам уменьшается. У некото­
рых рецепторов белково-пептидных гормонов и катехоламинов после
присоединения лиганда происходит интернализация (инактивация
Глава 4
182
лиганд-рецепторного комплекса путем транспортировки в клетку) —
рецепторы инсулина, Р-адренорецепторы.
Регуляция содержания гормонов в крови
Д ля эффективной регуляции физиологических функций уровень
эффекторных гормонов в крови должен очень точно соответствовать
потребностям организма. Особая роль в регуляции эндокринных
функций принадлежит гипоталамусу как ведущему подкорковому
вегетативному центру. Регулирующие влияния гипоталамуса реали­
зуются как гуморальным путем — через гипоталамо-гипофизарную
систему, так и нервным — через ВНС.
Основной механизм регуляции содержания эффекторных гор­
монов в крови — обратная связь, преимущественно отрицательная,
когда гормон прямым или опосредованным путем влияет на свою
секрецию. Классическим примером отрицательной обратной связи
является регуляция уровня глюкагона. Понижение концентрации
глюкозы в крови стимулирует а-клетки островков Лангерганса под­
желудочной железы, приводя к увеличению секреции глюкагона.
Глюкагон инициирует процессы гликогенолиза, приводящие к повы­
шению содержания глюкозы в крови. При увеличении концентрации
глюкозы в крови секреция глюкагона уменьшается.
Этот же принцип отрицательной обратной связи лежит в основе
регуляции секреции гормонов в системе гипоталамус—гипофиз—
периферические железы внутренней секреции. Повышение концен­
трации периферического эффекторного гормона в крови тормозит
на уровне гипоталамуса секрецию соответствующего либерина, на
уровне аденогипофиза — соответствующего тропного гормона и на
уровне периферической эндокринной железы — самого эффекторно­
го гормона (так называемые «длинны е», «короткие» и «ультракорот­
кие» петли обратной связи).
В лияния ВНС в значительной степени могут реализовывать­
ся через изменения тонуса сосудов, перфузирующих эндокринные
органы и ткани. Гипоталамические влияния через симпатическую
нервную систему непосредственно регулируют функцию мозгового
вещества надпочечников.
Секреция некоторых гормонов инициируется импульсацией от
периферических или центральных рецепторов. Например, секреция
АДГ, П Н У Ф и альдостерона во многом определяется афферентацией
от баро-, осмо- и волюморецепторов.
Внутренняя среда организма
183
Гипоталамо-гипофизарная система
Гипоталамус, часть промежуточного мозга, имеет многочисленные
двусторонние связи с другими структурами ЦНС. Нейросекреторные
клетки гипоталамуса, синтезирующие многие биологически актив­
ные вещества, не только находятся под контролем механизма обрат­
ной связи как одного из общ их принципов регуляции уровня би оло­
гически активных веществ в крови, но и регулируются различными
влияниями со стороны ЦНС. Именно гипоталамусу принадлежит
ведущая роль в интеграции нервных и гуморальных механизмов
регуляции физиологических функций.
Гипоталамус и отходящий от него гипофиз морфологически и
функционально объединены в гипоталамо-гипофизарную эндокрин­
ную систему. Гипофиз, расположенный в турецком седле, состоит
(у взрослого человека) из двух долей, различных по эмбриональному
происхождению: передней (аденогипофиз) и задней (нейрогипофиз).
Расположенная между ними промежуточная доля у человека имеется
только в эмбриональном периоде.
Гормоны гипоталамуса
Нейросекреторные клетки различных областей гипоталамуса син­
тезируют эффекторные гормоны, либерины и статины, управляющие
деятельностью аденогипофиза, а также другие регуляторные пептиды
и нейротрансмиттеры (рис. 4.4).
Нейросекреторные клетки крупноклеточных ядер (паравентрикулярные и супраоптические) переднего гипоталамуса синтезируют
эффекторные гормоны : вазопрессин (А Д Г ) и окситоцин, которые по
аксонам транспортируются в нейрогипофиз, откуда секретируются
в кровь. Небольшая часть аксонов оканчивается в срединном воз­
вышении и секретирует в воротную вену гипофиза некоторое коли­
чество АДГ. Помимо гипоталамуса А Д Г и окситоцин синтезируются
и на периферии (в мозговом веществе надпочечников и в половых
железах).
АДГ, взаимодействуя с V-2 рецепторами собирательных трубочек,
повышает реабсорбцию воды в почках, через V-l рецепторы оказывает
вазоконстрикторный эффект, а попадая в воротную систему гипофиза,
стимулирует секрецию АК ТГ. Показано участие А Д Г в формировании
питьевого поведения и в механизмах памяти. Наряду с этим А Д Г дейс­
твует как нейротрансмиттер и нейромодулятор в ЦНС.
184
Глава 4
Рис. 4.4. Схема гипоталамо-гипоф изарной системы
Окситоцин у женщин в процессе родов усиливает сокращения
матки, а в период грудного вскармливания, наряду с пролактином, стимулирует секрецию молока. У мужчин окситоцин вызывает
сокращение семявыводящих протоков при семяизвержении.
М едиальная область гипоталамуса содержит мелкоклеточные
ядра, нейроны которых синтезируют пептидные гормоны, управля­
ющие деятельностью аденогипофиза (рилизинг- и ингибирующие
гормоны, или либерины и статины). Аксоны этих нейронов проеци­
руются в срединное возвышение и выделяют нейросекрет в воротную
систему гипофиза (рис. 4.5).
Либерины гипоталамуса [КРГ, тиреолиберин (Т Р Г ), гонадолиберин, пролактолиберин] стимулируют синтез и секрецию гормонов
аденогипофиза, статины (соматостатин, пролактостатин-дофамин)
тормозят синтез и секрецию гландотропных гормонов.
Гипофизотропные гормоны гипоталамуса могут принимать учас­
тие в регуляции секреции различных гормонов аденогипофиза. Так,
Т Р Г стимулирует тиреотропные и соматотропные клетки гипофиза,
соматостатин тормозит не только секрецию СТГ, но и А К Т Г , Т Т Г и
185
Внутренняя среда организма
Гипоталамус.
Рилизинг- и
Кортико-
l
Аденогипофиз.
^ ^Тропны е
гормоны:
Кортикотропин
(А К ТГ)
^ ингибирирующие либерин
. гормоны:
Тиреолиберин
Соматолиберин
Сомато­
статин
ПролакгоПролактостатин
(дофамин) либерин
Гонадолиберин
-А 5 Г Ж З
Соматотропин
(С ТГ)
Тиреотропин
(ТТ Г)
Лютенизирующий
гормон (ЛГ)
Фолликулости­
Пролакмулирующий
тин
гормон (Ф С Г)
/
I
I
I
Перефирические
^железы:
/
Кора
надпочечников
+
У Эффекторные
у гормоны:
Глюкокорти коиды
Андрогены
Альдостерон*
Щитовидная
железа
Половые
железы
1
Трииодтиронин
Тетраиодтирон и н
Андрогены
Эстрогены
Прогестерон
Клетки, ткани и органы — «мишени»
Рис. 4.5. Р егуля тор н ы е взаимодействия между ги п оталам усом , аде­
ноги поф изом и периф ерическими ж елезам и внутренн ей секреции:
---- ► — стим улирую щ ие влияния; — -► — ингибирую щ ие влияния;
---- ► — вли я н и я эф фекторных гормонов на к летк и -«м и ш ен и »
пролактина. Дофамин, также поступающий в клетки аденогипофиза
через кровь, ингибирует секрецию Ф С Г, ЛГ, Т Т Г и пролактина.
На синтез и секрецию каждого конкретного гормона аденогипо­
физа могут оказывать влияние многие гипоталамические факторы:
либерины , статины, опиоиды, другие нейропептиды, дофамин.
В целом можно считать, что синтез и секреция каждого гормона
аденогипофиза находятся под контролем определенной комбинации
гипоталамических факторов.
Особого обсуждения требует достаточно обширная группа био­
логически активных веществ, синтезируемых нейросекреторными
клетками гипоталамуса, часто объединяемых термином «регулятор­
ные гипоталамические пептиды », функции которых полностью еще не
186
Глава 4
изучены. К этой группе относят энкефалины и эндорфины, ВИП,
нейротензин, вещество П, холецистокинин и др. М ногие из нейро­
пептидов гипоталамуса синтезируются и в других местах, прежде
всего, в эндокринных клетках Ж КТ.
Известно, что некоторые из этих веществ обладают гипофизотропным влиянием (опиоиды), являются компонентами ноци­
цептивной (вещество П ) и эндогенной антиноцицептивной систем
(энкефалины и эндорфины). Совместно с либеринами и эффекторными гормонами гипоталамуса они участвуют в центральной
регуляции А Д (В И П , опиоиды, А Д Г ), в формировании специфи­
ческих форм поведения, таких, например, как пищевое (холецис­
токинин, нейротензин), питьевое (АД Г, ангиотензин II), половое
(гонадолиберин). Некоторые нейропептиды гипоталамуса оказывают
влияние на такие функции, как внимание, обучение, память (АДГ,
окситоцин, А К Т Г ).
Гормоны аденогипофиза
По химической структуре гормоны аденогипофиза можно раз­
делить на пептидных (А К Т Г , СТГ, пролактин) и гликопротеиды
(Т Т Г, ФСГ, Л Г). Ф ункционально гормоны аденогипофиза делятся
на эффекторные (СТГ, пролактин) и гландотропные, которые стиму­
лирую т выработку гормонов периферическими гипофиззависимыми
железами внутренней секреции (А К Т Г , ТТГ, Ф СГ, Л Г ).
Соматотропный гормон
С Т Г секретируется клетками аденогипофиза с четкой суточной
периодикой. Базальный уровень секреции поддерживается сек­
реторными выбросами, которые ночью происходят чаще и имеют
больш ую амплитуду. Повышение секреции С Т Г совпадает с нача­
лом фазы глубокого сна. Основными регуляторами секреции С Т Г
являются гипоталамические нейропептиды — соматолиберин, ока­
зывающий стимулирующее действие, и соматостатин, тормозящий
секрецию. Стимулируют продукцию С Т Г вазопрессин, эндорфин
и ТРГ. Тормозное влияние оказывает дофамин. В регуляции секре­
ции С Т Г принимают участие циркулирующие в крови инсулино­
подобные факторы роста, глюкоза, жирные кислоты и аминокис­
лоты. Так, гипогликемия активирует секрецию соматолиберина и
СТГ, а гипергликемия — тормозит. Избыток аминокислот и сни­
жение содержания свободных жирных кислот в крови активируют
секрецию СТГ.
Внутренняя среда организма
187
Около половины С Т Г циркулирует в крови в связанной с белком
форме, образуя так называемый гормональный резерв. Время полужизни С Т Г составляет около 20 мин.
В отличие от других белково-пептидных гормонов, С Т Г имеет
выраженную видовую специфичность.
С Т Г — анаболический гормон, стимулирующий рост всех тканей
за счет увеличения поступления в клетки аминокислот и усиления
синтеза белка. С Т Г стимулирует продукцию в печени инсулиноподоб­
ного фактора роста (инсулиноподобный фактор роста I, соматомедин),
совместно с которым стимулирует рост костной ткани, особенно в
юношеской фазе развития. В эмбриональном периоде развития дейс­
твие С Т Г опосредует инсулиноподобный фактор роста II. Недостаток
С Т Г в детском и юношеском периодах приводит к ограничению роста,
при избыточной продукции С Т Г развивается гигантизм.
Влияние С Т Г на углеводный и жировой обмен имеет две фазы.
В начальной фазе С Т Г увеличивает поглощение глюкозы мышцами и
жировой тканью, одновременно тормозя в последней липолиз (инсу­
линоподобный эффект). С Т Г может непосредственно стимулировать
секрецию инсулина р-клетками поджелудочной железы.
Через несколько минут развивается вторая фаза, для которой харак­
терны контринсулярные эффекты СТГ. Тормозятся поглощение и ути­
лизация глюкозы, усиливается липолиз. Усиление липолиза приводит
к освобождению жирных кислот. Гормон повышает активность а-клеток островков Лангерганса и продукцию глюкагона. Одновременно
активируется инсулиназа печени и усиливается разрушение инсулина.
Таким образом С Т Г оказывает гипергликемическое действие.
Пролактин
Синтез и секреция пролактина регулируются гипоталамическими
нейропептидами — пролактолиберином (стимулирует продукцию
пролактина) и пролактостатином-дофамином. Секреция пролактина
зависит от уровня в крови эстрогенов, глюкокортикоидов и гормонов
щитовидной железы. В период беременности и лактации синтез про­
лактина увеличивается.
Основным орган ом -«м и ш енью » являю тся м олочны е железы.
Пролактин совместно с эстрогенами и С Т Г стимулирует рост молоч­
ных желез и пролиферацию молочных протоков. Данный эффект
наиболее выражен во время беременности. В период лактации
пролактин стимулирует синтез молока. Секреция пролактина рефлекторно стимулируется актом сосания.
188
Глава 4
Пролактин участвует также в регуляции полового созревания и
овариально-менструального цикла. Пролактин способствует подде­
ржанию секреторной активности желтого тела в яичниках и образо­
ванию прогестерона.
Пролактин принимает участие в регуляции водно-солевого обме­
на, повышая реабсорбцию натрия, хлора и воды в почках. Гормон
усиливает эффекты альдостерона и вазопрессина. Пролактин сти­
мулирует синтез белка, повышает образование жира из углеводов,
способствуя послеродовому ожирению.
Гландотропные гормоны после секреции в кровяное русло достигают
органов-«мишеней», взаимодействуют со специфическими рецепто­
рами эндокринных желез и через различные системы вторых пос­
редников активируют ферменты, необходимые для синтеза эффекторных гормонов. Помимо действия на периферические железы
внутренней секреции, гормоны аденогипофиза выполняют и многие
другие функции.
К гландотропным гормонам аденогипофиза относятся А К Т Г , ТТГ,
гонадотропные гормоны (Ф С Г и Л Г ). Ф ункционально могут быть
выделены три эндокринные организации:
• гипоталамус—аденогипофиз—кора надпочечников;
• гипоталамус—аденогипофиз—щитовидная железа;
• гипоталамус—аденогипофиз—половые железы.
В системе гипоталамус—гипофиз—периферические железы основ­
ным принципом регуляции является механизм отрицательной обрат­
ной связи. Эффекторные гормоны, циркулирующие в крови, посредс­
твом отрицательной обратной связи вызывают уменьшение секреции
соответствующих либеринов, тропинов и самого периферического
гормона.
Система гипоталамус-аденогипофиз-кора
надпочечников
Эндокринная система гипоталамус-аденогипофиз—кора надпо­
чечников продуцирует гормоны, регулирующие многие функции
организма, но особого рассмотрения требует система важнейших гор­
монов: К Р Г —кортикотропин—кортизол. Эти гормоны играют особую
роль в обеспечении жизнедеятельности организма при физических,
эмоциональных, умственных нагрузках, действии стрессорных фак­
торов, во время болезни.
КРГ(кортикотропин-рилизинг-гормон) синтезируется в мелкоклеточ­
ных ядрах гипоталамуса. Помимо механизма отрицательной обрат­
Внутренняя среда организма
189
ной связи особое значение для активации нейросекреторных клеток
гипоталамуса, продуцирующих КРГ, имеют влияния других структур
ЦНС, прежде всего, лимбической системы.
К Р Г оказывает стимулирующее влияние на клетки аденогипофи­
за, продуцирующие проопиомеланокортин, из которого образуют­
ся А К Т Г , меланоцитстимулирующий гормон, (З-эндорфин и другие
пептиды. Как нейротрансмиттер и нейромодулятор К Р Г участвует в
центральной регуляции вегетативных функций, активируя симпати­
ческий отдел ВНС.
Адренокортикотропный гормон
Секреция А К Т Г имеет суточный ритм, увеличиваясь после засы­
пания и достигая максимума перед пробуждением. Стимулирующее
действие на секрецию А К Т Г , помимо КРГ, оказывают АДГ, норадреналин, ангиотензин II. Тормозит секрецию соматостатин. П о меха­
низму отрицательной обратной связи синтез и секрецию А К Т Г тор­
мозит циркулирующий в крови кортизол.
Рецепторы к А К Т Г — мембранные, связанные с G -белком, акти­
вирующим аденилатциклазную систему.
Физиологические эффекты А К Т Г делят на надпочечниковые и
вненадпочечниковые.
А К Т Г , стимулируя клетки пучковой зоны коры надпочечников,
регулирует синтез и секрецию глюкокортикоидов. В существенно
меньшей степени А К Т Г оказывает влияние на клетки клубочковой
(синтез минералкортикоидов) и сетчатой (синтез андрогенов) зон
коры надпочечников.
Важнейшие вненадпочечниковые функции А К Т Г заключаются
в стимуляции липолиза, а также в повышении синтеза меланина и
усилении пигментации кожи.
Гормоны коры надпочечников
Кора надпочечников морфологически и функционально состоит
из трех слоев. В клубочковой зоне синтезируются минералкортикоиды
(главным образом, альдостерон), в пучковой зоне — глюкокортикоиды
(в основном, кортизол), в сетчатой зоне — андрогены.
Предшественником кортикоидов является холестерин, из которо­
го они синтезируются с помощью специфических ферментативных
систем.
Рецепторы всех стероидных гормонов внутриклеточные, ядерные,
активирующие транскрипцию.
190
Глава 4
Глюкокортикоиды
Синтез и поступление в кровь глюкокортикоидов, так же как и
регулирующих эти процессы К Р Г и А К Т Г , имеют суточную ритмику.
При нормальном цикле «сон —бодрствование» после засыпания сек­
реция кортизола в кровь начинает возрастать, достигая максимума в
утренние часы (примерно через 1—2 ч после максимума А К Т Г ).
Выделение кортизола быстро увеличивается при активации сек­
реции К Р Г гипоталамусом и А К Т Г аденогипофизом в условиях
действия на организм стрессорных факторов, физической работы,
во время заболеваний.
Глюкокортикоиды, циркулирующие в крови, по механизму отри­
цательной обратной связи подавляют секрецию К Р Г на уровне гипо­
таламуса и А К Т Г на уровне гипофиза, что приводит к уменьшению
синтеза глюкокортикоидов корой надпочечников.
В крови более 90% глюкокортикоидов связано с транспортны­
ми белками (альбумином или кортизолсвязывающим глобулином —
транскортином). Время полужизни кортизола составляет около 2 ч.
Инактивированные в печени глюкокортикоиды выделяются из орга­
низма в основном через почки и, в небольшом количестве, через ЖКТ.
Ф ункции глюкокортикоидов
Эффекты глюкокортикоидов многообразны, поскольку практи­
чески все клетки организма имеют рецепторы к ним.
Глюкокортикоиды прямо или опосредовано регулируют практи­
чески все составляющие обмена веществ в организме человека.
Углеводный обмен. Глюкокортикоиды повышают уровень глюкозы
в крови. Эти гормоны стимулируют глюконеогенез в печени, акти­
вируя синтез соответствующих ферментов. При этом глюкокортико­
иды усиливают распад белков скелетных мышц, соединительной и
костной ткани, а также липолиз. Образовавшиеся метаболиты (ами­
нокислоты и свободные жирные кислоты) поступают в печень, где
включаются в процессы глюконеогенеза. Глюкокортикоиды также
подавляют утилизацию глюкозы тканями (за исключением ЦНС).
Белковый обмен. На белковый обмен глюкокортикоиды оказывают
в основном катаболическое и антианаболическое действие, что может
привести к отрицательному азотному балансу.
Жировой обмен. Непосредственно глюкокортикоиды стимулируют
липолиз и подавляют синтез жира из углеводов. Однако вызываемая
гормонами гипергликемия и связанное с ней повышение секреции
Внутренняя среда организма
191
инсулина приводят к повышению липогенеза в верхней части туло­
вища, на шее и на лице.
Минеральный обмен. При высоких концентрациях глюкокортикоиды могут взаимодействовать с рецепторами минералкортикоидов и
вызывать аналогичные эффекты (повышение реабсорбции Na+ и Н 20
в почечных канальцах при усилении выведения К+).
Глюкокортикоиды:
• понижают число эозинофилов, базофилов и лимфоцитов в
крови;
• ослабляю т клеточный, а при длительном терапевтическом воз­
действии — и гуморальный иммунитет, в высоких дозах являются
иммунодепрессантами;
• усиливая катаболизм белка в лимфоидной ткани и угнетая им м ун­
ные реакции, оказывают антиаллергические эффекты;
• тормозят воспалительные процессы;
• стимулируют синтез катехоламинов, особенно адреналина, в
мозговом веществе надпочечников;
• повышают чувствительность адренорецепторов к катехоламинам;
• повышают сенсорную чувствительность и возбудимость ЦНС.
Глю кокортикоиды участвуют в формировании стрессорного
ответа, повышая энергетические резервы, способствуя мобилизации
физиологических функций, поддерживая оптимальную регуляцию
сердечно-сосудистых функций, повышая в целом устойчивость орга­
низма к действию стрессорных факторов.
М инералкортикоиды
Основной минералкортикоид, синтезируемый в коре надпочечни­
ков — альдостерон, который является компонентом РААС (см. ниже),
участвующей в регуляции многих процессов в организме, прежде
всего, водно-солевого обмена и гемодинамики. Синтез и высвобож­
дение альдостерона стимулируются главным образом ангиотензинном II. Активация РААС, приводящая в том числе и к увеличению
секреции альдостерона, происходит при уменьшении О Ц К и пони­
жении кровяного давления, гипонатриемии в крови и повышении
концентрации Na+ в моче дистальных канальцев.
Дополнительное стимулирующее действие на секрецию альдосте­
рона оказывает А К Т Г , особенно в стрессовых ситуациях.
Тормозит образование альдостерона предсердный натрий-уретический фактор (П Н У Ф ).
192
Глава 4
По механизму отрицательной обратной связи гиперкалиемия сти­
мулирует, а гипокалиемия тормозит секрецию альдостерона.
В крови альдостерон в основном циркулирует свободно, поэтому
время полужизни гормона составляет не более 15 мин. Инактивируется
в печени и экскретируется преимущественно почками.
Рецепторы, как и для всех стероидов, внутриклеточные, ядерные,
активирующие транскрипцию генов транспортных белков-переносчиков и ферментов.
Эффекты альдостерона
Основная функция альдостерона — поддержание баланса элект­
ролитов в жидкостях организма.
Главными органами-«миш енями» являются почки, где альдо­
стерон влияет на реабсорбцию в дистальных извитых канальцах и
начальных отделах собирательных трубочек. Альдостерон увеличи­
вает реабсорбцию Na+, Cl- и бикарбонатов в канальцах, что при­
водит к усилению реабсорбции Н 20 , увеличению О Ц К и давления.
Одновременно альдостерон усиливает выведение Н+ и К+.
Аналогичное влияние на обмен натрия и калия альдостерон ока­
зывает в слюнных и потовых железах.
Альдостерон обладает провоспалительным действием, связанным
с усилением экссудации жидкости из просвета сосудов в ткани и
появлением отечности тканей.
П оловы е гормоны коры надпочечников
Андрогены надпочечников в клетках-«миш енях» с помощью фер­
ментов частично превращаются в тестостерон и эстрогены.
Существенную роль андрогены надпочечников играют в основном
в детском возрасте, когда еще мало развита инкреторная функция
половых желез. С возрастом продукция андрогенов корой надпочеч­
ников сокращается (примерно на 30%). У женщин андрогены надпо­
чечников составляют основную долю мужских половых гормонов.
Секреция гормонов регулируется по механизму отрицательной
обратной связи, стимулируется А К Т Г и, соответственно, имеет суточ­
ную периодичность. М аксимальный уровень наблюдается утром
(6—8 ч), минимальный — вечером (19—20 ч).
Функции:
• способствуют развитию вторичных половых признаков;
• стимулируют синтез белка (анаболическое действие);
• стимулируют окостенение эпифизарных хрящей.
Внутренняя среда организма
193
Система гипоталамус-гипофиз-щитовидная
железа
Т Р Г секретируется не только нейросекреторными клетками гипо­
таламуса, но и нейронами других структур ЦНС, в частности амигдалы и ствола мозга. На синтез и секрецию Т Р Г в гипоталамусе, наряду
с механизмами отрицательной обратной связи с уровнем циркулиру­
ющих в крови ТЗ и Т4, оказывают влияние и другие факторы, напри­
мер, секрецию Т Р Г стимулирует сильное холодовое воздействие.
В аденогипофизе Т Р Г стимулирует синтез и секрецию не только
ТТГ, но и пролактина. Кроме того, Т Р Г играет роль нейротрансмит­
тера и нейромодулятора в ЦНС.
Т Т Г — гликопротеид, гландотропный гормон аденогипофиза, регу­
лирует процессы секреции и синтеза гормонов щитовидной железы и
ее метаболизм. Синтез Т Т Г стимулирует ТРГ, а тормозит — соматоста­
тин гипоталамуса. П о механизму отрицательной обратной связи про­
дукцию Т Т Г регулируют иодсодержащие гормоны щитовидной желе­
зы (Т4 и ТЗ). Глюкокортикоиды тормозят выработку ТТГ. Показано
снижение секреции Т Т Г при травмах, болях, наркозе. Усиливает
секрецию ТТГ, так же как и ТРГ, охлаждение организма. Секреция
Т Т Г имеет суточную периодику, максимум наблюдается в часы, пред­
шествующие засыпанию.
Т Т Г увеличивает кровоснабжение щитовидной железы, влияет
на рост и развитие ф олликулярного эпителия щитовидной железы,
стимулирует все этапы синтеза и секреции иодсодержащих гормонов,
а также поглощение иодидов. Избыток Т Т Г проявляется гиперфунк­
цией щитовидной железы.
Гормоны щитовидной железы
Щитовидная железа, состоящая из двух соединенных перешейком
долей, расположена по обеим сторонам трахеи ниже щитовидного
хряща. Эндокринные функции осуществляют два типа клеток щ ито­
видной железы: образующие ф олликулы тироциты, синтезирующие
иодсодержащие тиреоидные гормоны ТЗ и Т4, и парафолликулярные
К-клетки, синтезирующие пептидный гормон кальцитонин.
Внутри образованных тироцитами фолликулов находится коллоид­
ная масса тиреоглобулина. В тироцитах, способных захватывать иодиды, осуществляется синтез тиреоглобулина и окисление иодидов в
атомарный иод. Тиреоглобулин содержит на поверхности остатки кис­
лоты тирозина, которые подвергаются иодированию. Иодированный
194
Глава 4
тиреоглобулин — это прогормон иодсодержащих гормонов, форма их
хранения в коллоиде. При необходимости секреции капли коллоида
путем эндоцитоза поступают в тироциты, где подвергаются гидролизу
ферментами лизосом. Два продукта гидролиза — ТЗ и Т4 — секретируются через базальную мембрану тироцитов в кровь.
Основным продуктом, образующимся исклю чительно в щ ито­
видной железе, является Т4. Вне щитовидной железы Т4 путем деио­
дирования превращается в высокоактивный ТЗ или в биологически
неактивный реверсивный ТЗ (гТЗ). Синтез иодсодержащих гормонов
зависит от поступления иода и эффективности его обмена.
Под влиянием Т Т Г осущ ествляется синтез тиреоглобулина и
тиреоидных гормонов. Активируя гидролиз тиреоглобулина, Т Т Г
увеличивает секрецию тиреоидных гормонов.
По принципу отрицательной обратной связи циркулирующие в
крови тиреоидные гормоны подавляют секрецию Т Р Г гипоталамусом
и Т Т Г аденогипофизом.
К периферическим механизмам регуляции относится активация
и инактивация гормонов щитовидной железы путем превращения
малоактивного Т4 в высокоактивный ТЗ или в биологически неак­
тивный гТЗ с помощью специфических ферментных систем.
Нервная регуляция функций щитовидной железы играет сущест­
венно меньшую роль, чем гуморальная. Симпатическая нервная сис­
тема стимулирует, а парасимпатическая — тормозит синтез гормонов
щитовидной железы.
Транспорт циркулирующего в крови Т4 более чем на 99% связан
с белками плазмы (альбумином и глобулином) и имеет достаточно
длительны й период полужизни, составляющий около 7 сут.
ТЗ образуется преимущественно в клетках-«миш енях» путем дейо­
дирования Т4. ТЗ в значительно меньшей степени связывается с
белками плазмы, период полужизни этого гормона составляет около
одних суток.
После связывания тиреоидных гормонов с мембранными рецептора­
ми они транспортируются через мембрану с помощью переносчиков. В
клетке происходит деиодирование Т4 до ТЗ, который связывается с ядерными рецепторами. Активированный гормон-рецепторный комплекс
оказывает влияние на транскрипцию, следствием чего является синтез
специфических белков, опосредующих клеточный эффект гормонов.
Иодсодержащие гормоны инактивируются (конъюгируются) в
печени, экскретируются с желчью в кишечник, откуда всасываются,
деиодируются в почках и выводятся с мочой.
Внутренняя среда организма
195
Ф ункции иодсодержащих гормонов
Иодсодержащие гормоны оказывают очень разнообразные влия­
ния на клетки-«миш ени» в организме человека, принимают участие
в регуляции обмена веществ и многих физиологических функций.
• Активируют клеточный метаболизм, увеличивают потребление
кислорода, повышают основной обмен.
• Вызывают увеличение синтеза белков в результате активации
транскрипции в клетках-«мишенях», а также повышения прони­
цаемости мембраны для аминокислот. При этом очень высокие
концентрации иодтиронинов стимулируют катаболические про­
цессы, что может привести к отрицательному азотному балансу.
• Стимулируют все этапы метаболизма углеводов, вызывая в целом
гипергликемический эффект. Под влиянием иодтиронинов уси­
ливается распад гликогена, повышается всасывание глюкозы в
кишечнике, ускоряется инактивации инсулина в печени. При
этом гормоны увеличивают потребление и окисление глю ко­
зы клетками. Одновременно развивающаяся гипергликемия по
механизму отрицательной обратной связи стимулирует секре­
цию инсулина. Таким образом, тиреоиды, стимулируя секрецию
инсулина и одновременно оказывая контринсулярные эффекты,
могут способствовать развитию сахарного диабета.
• Оказывают липолитический эффект, усиливают окисление жир­
ных кислот.
• Усиливают эффекты катехоламинов, повышая чувствительность
адренорецепторов и подавляя активность монаминоксидазы
(фермента, инактивирующего катехоламины). Эти влияния тиреоидов проявляются в увеличении ЧСС, сужении периферических
сосудов и усилении других симпатических эффектов.
• Вызывают повышение теплопродукции, обусловленное как повы­
шением интенсивности клеточного метаболизма, так и усилением
симпатических эффектов. Иодтиронины участвуют в формиро­
вании ответной реакции на охлаждение.
• Обеспечивают нормальный рост, развитие и дифференцировку
тканей и органов. Тиреоиды являются синергистами С Т Г в регуля­
ции роста тканей, в частности, скелета. Кроме того, ТЗ увеличивает
синтез СТГ, специфически стимулируя транскрипцию. Тиреоиды
обеспечивают нормальное созревание и развитие нервной системы.
• Повышают возбудимость Ц Н С и активацию психических про­
цессов.
• Участвуют в поддержании половых функций человека.
196
Глава 4
Недостаточный синтез тиреоидных гормонов (гипотиреоз) может
наблюдаться, например, при нарушениях продукции Т Т Г гипофизом
или дефиците иода в пище и воде. Гипотиреоз проявляется в сниже­
нии основного обмена, гипогликемии, снижении температуры тела,
торможении нервно-психической активности, снижении активности
симпатической нервной системы и других нарушениях.
Избыточная продукция тиреоидны х гормонов (гипертиреоз)
сопровождается повышением основного обмена, гипергликемией,
тахикардией, повышением температуры тела, повышенной возбуди­
мостью ЦНС, проявлениями повышенного симпатического тонуса.
Калъцитонин — пептидный гормон парафолликулярных К-клеток
щитовидной железы. Помимо этого, образуется в легких и в тимусе.
Является одним из кальцийрегулирующих гормонов. Секреция кальцитонина в основном регулируется уровнем ионизированного каль­
ция в крови по механизму обратных связей. Стимулятором секреции
служит значительное увеличение концентрации Са2+. На секрецию
кальцитонина оказывают влияние некоторые гастроинтестинальные
гормоны, в частности, гастрин.
Рецепторы кальцитонина мембранные, реализующие эффект
через системы вторых посредников ц А М Ф и цГМ Ф .
Кальцитонин понижает концентрацию Са2+ в крови, облегчая
минерализацию и подавляя резорбцию костной ткани. Гормон спо­
собствует выведению кальция из организма, понижая его реабсорб­
цию в почках.
Система гипоталамус-аденогипофиз-половые
железы
Гонадолиберин, синтезируемый нейросекреторны ми клетками
гипоталамуса, поступает в кровь воротной вены аденогипофиза в
пульсирующем режиме, оказывая влияние на клетки аденогипофиза,
синтезирующие Л Г и ФСГ.
У мужчин синтез гонадолиберина по механизму отрицательной
обратной связи подавляют тестостерон и ингибины.
Влияние гонадолиберина на секрецию Ф С Г и Л Г у женщин
зависит от циклических изменений содержания в крови гормонов
яичников — эстрогенов, прогестерона и ингибинов. При низком
содержании эстрогенов гонадолиберин стимулирует синтез ФСГ, а
при высоком — синтез ЛГ.
Гонадотропные гормоны аденогипофиза влияют на функции поло­
вых желез у мужчин и женщин. У мужчин Ф С Г и Л Г регулируют
Внутренняя среда организма
197
сперматогенез и синтез половых гормонов. У женщин Ф С Г и Л Г учас­
твуют в регуляции овариально-менструального цикла, беременности,
секреции половых гормонов. К гонадотропным гормонам в части
своих функций близок пролактин, так как он, помимо прямого вли­
яния на развитие и функции молочных желез, стимулирует синтез и
секрецию прогестерона желтым телом яичников.
Половые железы человека выполняют две основные тесно связан­
ные функции — экзокринную и эндокринную. Экзокринная функция
заключается в образовании сперматозоидов в семенниках у мужчин и
яйцеклеток в яичниках у женщин. Эндокринная функция состоит в
синтезе и секреции мужских и женских половых гормонов.
Мужские половые гормоны — андрогены (тестостерон и дигид­
ротестостерон) — образуются в интерстициальных клетках яичек
(клетках Лейдига). При этом некоторое количество андрогенов в
клетках Сертоли путем ароматизации превращается в эстрогены.
В этих же клетках образуются ингибины.
Женские половые гормоны — эстрогены (эстрадиол, эстриол, эст рон) — образуются преимущественно в ф олликулах яичников, а про­
гестерон — в основном в желтом теле яичников. В половых железах
у женщин также синтезируется некоторое количество андрогенов и
ингибины.
Основная физиологическая роль половых гормонов заключается в
обеспечении половых функций, но при этом они обладают широким
спектром других эффектов.
Подробно функции и механизмы регуляции гормонов системы
гипоталамус—аденогипофиз—половые железы рассмотрены в главе
«Половые функции человека».
Симпатоадреналовая система
Гормоны симпатоадреналовой системы — катехоламины — обра­
зуются из аминокислоты тирозина в последовательности: тироз и н -Д О Ф А -д о ф а м и н -н о р а д р е н а л и н -а д р е н а л и н .
О сн овн ы м и
периферическими областями синтеза катехоламинов являются моз­
говое вещество надпочечников и нейроны симпатических ганглиев.
Эндокринную функцию мозгового вещества надпочечников осу­
ществляют хромаффинные клетки, эмбриогенетически родственные
клеткам симпатической нервной системы и являющиеся, по сути,
видоизмененными ганглионарными нейронами. М елкие скопления и
одиночные хромаффинные клетки находятся также в крупных арте­
риях, в некоторых вегетативных ганглиях и в некоторых органах.
198
Глава 4
В хромаффинных клетках синтезируется и накапливается в сек­
реторных гранулах преимущественно адреналин и в небольшом коли­
честве — норадреналин. Одновременно с катехоламинами хромаффинные клетки секретируют в кровь А Т Ф и энкефалины.
Постганглионарные симпатические окончания, напротив, выде­
ляю т в основном норадреналин, который играет роль медиатора,
однако некоторое его количество попадает в кровоток, что и позво­
ляет рассматривать норадреналин как гормон. В целом в спокойном
состоянии норадреналина содержится в крови больше, чем адрена­
лина. При этом секреция катехоламинов в условиях покоя невелика,
но она резко повышается в ситуациях, вызывающих активацию
симпатической нервной системы.
Нервные центры, регулирую щ ие секреторную ф ункцию хромаффинной ткани надпочечников, расположены в гипоталамусе.
Возбуждение симпатических ядер гипоталамуса через симпати­
ческие преганглионарны е волокна (в составе чревного нерва),
окончания которых выделяют медиатор ацетилхолин, взаимодейс­
твую щ ий с никотиновы ми холинорецепторами ганглионарного
типа, стимулирует выделение катехоламинов. При этом вначале
преобладает секреция адреналина, а при д ли тельн ом возбуждении
секреция адреналина уменьшается, а норадреналина — ув ели ч и ­
вается.
На синтез катехоламинов в мозговом веществе надпочечников
паракринное стимулирующ ее влияние оказывает кортизол, син­
тезируемый корковым веществом под непосредственным влияни­
ем А К Т Г . Кортизол главным образом стимулирует метилирование
норадреналина, приводящее к образованию адреналина.
Инактивация катехоламинов осуществляется под влиянием моноаминооксидазы и катехолортометилтрансферазы. Продукты метабо­
лизма выводятся почками.
Время полуж изни катехоламинов в плазме составляет около
2 мин.
Особенности физиологических эффектов катехоламинов обуслов­
лены их взаимодействием с различными типами адренорецепторов,
относящихся к мембранным рецепторам, связанным с G -белками.
Взаимодействие катехоламинов с а,-адренорецепторами приводит к
образованию инозитолтрифосфата и увеличению внутриклеточного
[Са2+], а с а 2-адренорецепторами — вызывает ингибирование адени­
латциклазы и уменьшение внутриклеточного [ц А М Ф ]. Образование
комплекса с p-адренорецепторами сопровождается активацией аде-
Внутренняя среда организма
199
нилатциклазы и увеличением внутриклеточного [ц А М Ф ], а также
активацией Са2+-каналов и увеличением внутриклеточного [Са2+].
Р-Адренорецепторы, в зависимости от блокирующего действия анта­
гонистов, разделяют на подтипы р, и р2.
Адреналин обладает большим сродством к Р-адренорецепторам,
а норадреналин — к а-адренорецепторам.
Количество адренорецепторов (особенно p-рецепторов) непосто­
янно, оно может уменьшаться вследствие интернализации и возрас­
тать, например, при денервации. В присутствии агонистов они десенситизируются благодаря фосфорилированию. Глюкокортикоиды и
иодсодержащие гормоны щитовидной железы повышают чувстви­
тельность адренорецепторов к катехоламинам.
Эффекты адреналина
• Адреналин усиливает окислительные процессы в тканях, уве­
личивает потребление кислорода и теплопродукцию в тканях,
повышает основной обмен.
• Липолитическое действие адреналина приводит к повышению
содержания свободных жирных кислот в плазме.
• Под влиянием адреналина повышается уровень глюкозы в крови.
К гипергликемии приводят усиление расщепления и подавление
синтеза гликогена в печени и мышцах, снижение потребления
глюкозы тканями, усиление глюконеогенеза в печени.
• Адреналин повышает работоспособность скелетных мышц.
• Взаимодействуя с Р^рецепторами миокарда, адреналин повы­
шает возбудимость кардиомиоцитов, вызывает увеличение силы
сердечных сокращений и ЧСС.
• Влияние адреналина на периферические сосуды может быть
различным в зависимости от его концентрации в крови. В диа­
пазоне физиологической концентрации адреналина, циркули­
рующего в крови, у сосудов скелетной мускулатуры и печени
преобладает опосредуемая p-адренорецепторами реакция рас­
ширения, а у сосудов органов брюшной полости — опосредуемая
а-адренорецепторами реакция сужения. При высокой концен­
трации адреналина преобладает повышение тонуса резистив­
ных сосудов скелетных мышц (а-адренорецепторный эффект).
При этом адреналин не сужает сосуды работающих скелетных
мышц. В сосудах кожи адреналин вызывает только вазоконстрикцию.
• Адреналин стимулирует секрецию ренина в почках.
200
Глава 4
• Под влиянием адреналина происходят снижение тонуса бронхи­
альной мускулатуры и расширение бронхов и бронхиол, наблюда­
ется увеличение глубины дыхания.
• Адреналин оказывает стимулирующее действие на ретикулярную
формацию, которое проявляется реакцией активации на ЭЭГ и
сопровождается усилением внимания, умственным возбуждени­
ем, состоянием тревоги. Под влиянием адреналина повышается
возбудимость рецепторов сетчатки, слухового и вестибулярного
анализаторов. При этом повышаются пороги болевой чувстви­
тельности (адреналовая антиноцицептивная система).
• А дреналин вызывает экстренную перестройку функций, обес­
печивающую улучш ение взаимодействия организма с окружа­
ющей средой, повышение работоспособности в чрезвычайных
ситуация.
Действие норадреналина на функции организма сходно с действи­
ем адреналина, но не полностью идентично. В частности, у человека
норадреналин повышает периферическое сосудистое сопротивление,
оказывая вазоконстрикторный эффект во всех участках сосудистого
русла, повышает систолическое и диастолическое АД . Адреналин
вызывает подъем только систолического давления. Адреналин вызы­
вает больш ий метаболический эффект, чем норадреналин. Слабее
выражен гипергликемический эффект норадреналина. Адреналин в
больш ей мере стимулирует сердечную деятельность, так как лучше
связывается с Р-рецепторами.
В целом активация симпатоадреналовой системы обеспечивает
мобилизацию организма в ситуациях, требующих активной деятель­
ности, борьбы, бегства, эмоционального напряжения, при охлажде­
нии и действии других стрессорных факторов.
Эндокринная функция поджелудочной железы
Эндокринная часть поджелудочной железы представлена спе­
цифическими клетками островков Лангерганса. Основными гор­
монами подж елудочной железы, регулирую щ им и углеводны й
обмен, являются инсулин и глюкагон. И нсулин оказывает сущ ес­
твенное влияние на обмен белков и жиров. Секреция инсулина
и глюкагона, определяемая главным образом концентрацией глю ­
козы в крови, зависит также от влияния ВНС и некоторых дру­
гих гормонов. В частности, модулирующее влияние на секрецию
инсулина и глюкагона оказывает соматостатин, синтезируемый в
клетках островков Лангерганса. Благодаря тому что гормоны ост-
■
Внутренняя среда организма
201
ровковых клеток секретируются в кровь панкреатической вены,
впадающей в воротную вену печени, в печени концентрация этих
гормонов в 2—3 раза выше, чем в остальной системе кровообра­
щения.
Инсулин
Пептидный гормон, синтезируется в (3-клетках островков Лангерганса. Контролирует многие процессы метаболизма, стимулируя
синтез соединений, использующихся в организме для депонирования
питательных веществ (гликоген, триглицериды и др.).
Рецепторы инсулина относятся к семейству мембранных тирозинкиназных рецепторов.
Влияние инсулина на углеводный обмен
Активация инсулинозависимых переносчиков глюкозы стиму­
лирует транспорт глюкозы через мембрану в скелетных мышцах,
сердце, жировой ткани. В мышечных клетках инсулин стимулирует
образование мышечного гликогена, а в жировых — образование
триглицеридов.
В печени инсулин усиливает поглощение глюкозы и ее накопление
в форме гликогена, тормозит распад гликогена и глюконеогенез.
Инсулин стимулирует гликолиз и окисление углеводов в печени,
мышцах и жировой ткани.
Следствием этих процессов является понижение уровня глюкозы
в крови.
Влияние инсулина на обмен жиров
Инсулин способствует поступлению жирных кислот в клетки и
их превращению в триглицериды, тормозит липолиз триглицеридов.
Таким образом, инсулин обладает липогенным действием.
Влияние инсулина на обмен аминокислот и белков
Инсулин оказывает анаболическое и антикатаболическое дейс­
твие. Гормон стимулирует синтез белка в мышцах, увеличивая пог­
лощение аминокислот, а также блокирует гидролитическое расщеп­
ление белков.
Основной фактор, регулирующий секрецию инсулина, — концен­
трация глюкозы в крови, увеличение которой прямо стимулирует
секрецию инсулина. Стимулируют секрецию инсулина некоторые
гастроинтестинальные гормоны (гастрин, секретин и др.). Тормозит
202
Глава 4
секрецию инсулина соматостатин поджелудочной железы (паракринно).
Парасимпатические влияния стимулируют секрецию инсулина,
а симпатические преимущ ественно тормозят.
Глюкагон
Синтезируется в а-клетках островков Лангерганса. В регуляции
ряда процессов, прежде всего углеводного обмена, глюкагон высту­
пает антагонистом инсулина.
Глюкагон, активируя гликогенфосфорилазу, стимулирует рас­
щепление гликогена, высвобождение глюкозы и ее поступление в
кровь. Ингибируя гликогенсинтетазу, тормозит образование гликоге­
на. Стимулирует глюконеогенез.
Глюкагон обладает липолитическим действием, снижает количес­
тво триглицеридов и приводит к появлению кетоновых тел.
Таким образом, глюкагон обладает гипергликемическим, ли п оли ­
тическим и кетогенным действием, обеспечивая снабжение тканей
глюкозой и кетоновыми телами.
Секреция глюкагона усиливается при понижении концентра­
ции глюкозы в крови, при продолж ительной физической нагрузке,
в условиях стресса. С Т Г посредством соматомедина повыша­
ет активность а-клеток и продукцию глюкагона. Симпатические
влияния стимулирую т секрецию глюкагона. Подавляют секрецию
глюкагона инсулин, соматостатин, глюкоза крови.
Соматостатин
Синтезируется в 8-клетках островков Лангерганса поджелудоч­
ной железы. Соматостатин тормозит процессы секреции в большом
количестве разных клеток, активируя тормозной G -белок системы
цА М Ф . В островках Лангерганса соматостатин паракринно тормозит
секрецию инсулина и глюкагона.
Эпифиз
Эпифиз является железой нейроглиального происхождения. В эпи­
физе синтезируются многие пептидные гормоны и биогенные амины:
мелатонин, серотонин, норадреналин, гистамин и др.
Основная функция эпифиза — регуляция циркадных (суточных)
биологических ритмов, обеспечивающих адаптацию организма к
меняющимся условиям освещения. В условиях дневного освеще­
Внутренняя среда организма
203
ния тормозится превращение серотонина в мелатонин, в темноте,
наоборот, синтез мелатонина усиливается. Учитывая, что мелато­
нин принимает участие в регуляции многих функций организма,
цикличность его продукции определяет и циклические изменения
регуляторных влияний эпифиза.
Мелатонин тормозит секрецию гормонов, усиливающих обмен
веществ, — А К Т Г , ТТГ, СТГ. Гормон определяет цикличность гона­
дотропных эффектов, в том числе продолжительность овариально­
менструального цикла у женщин.
Полагают, что повышение уровня мелатонина способствует переходу
от бодрствующего состояния ко сну, вызывает седативный эффект.
Регуляция секреции мелатонина осуществляется рефлекторно с
участием симпатической нервной системы в зависимости от освещен­
ности, изменения которой воспринимаются сетчаткой.
Паращитовидные железы
Две пары околощитовидных желез расположены сзади или внутри
щитовидной железы. П Т Г является одним из кальцийрегулирующ их
гормонов организма человека и поддерживает нормальный уровень
кальция в крови.
Ф ункции П Т Г :
• усиливает функцию остеокластов в костной ткани, что приводит
к ее деминерализации и повышению уровня кальция в крови;
• усиливает реабсорбцию кальция в почках;
• стимулирует синтез кальцитриола в почках;
• уменьшает реабсорбцию фосфатов в почках и усиливает их выве­
дение с мочой.
Продукция П Т Г в основном определяется содержанием кальция
в крови. При увеличении концентрации кальция в плазме секреция
П Т Г снижается и, наоборот, уменьшение содержания кальция в
крови стимулирует образование ПТГ.
Эндокринная функция почек
В почках отсутствуют специализированные эндокринные клетки,
однако определенные клетки тканей почек обладают способностью
к синтезу и секреции биологически активных веществ. Гормонами
принято считать кальцитриол и эритропоэтин. Ренин, начальное
звено РААС, обычно относят к ферментам. Помимо этих веществ в
204
Глава 4
почках синтезируются тромбопоэтин, простагландины, калликреин
и другие биологически активные вещества.
Ренин
В юкстагломерулярном аппарате почек синтезируется фермент
ренин, являющийся инициативным звеном РААС. Секреция ренина
определяется несколькими факторами:
•давление крови в приносящей артериоле (степень растяжения
артериолы); понижение давления активирует, а повышение —
подавляет секрецию ренина;
• концентрация натрия в моче дистального канальца. Роль рецеп­
торов Na+ выполняют клетки так называемого плотного пятна
( macula densa). Секреция ренина усиливается при повышении
концентрации Na+ в моче дистального канальца;
• симпатические влияния через p-адренорецепторы стимулируют
секрецию ренина; стимулирующее действие оказывает и адре­
налин;
• уменьшает секрецию ренина предсердный натрий-уретический
фактор.
Регуляция секреции ренина осуществляется также по механиз­
му отрицательной обратной связи, определяемой содержанием в
крови компонентов РААС (ангиотензина и альдостерона), а также их
эффектами (содержанием в крови Na+, К+).
Помимо почек, продукция ренина осуществляется в стенках сосу­
дов других органов и тканей, головном мозге, слю нных железах.
Под влиянием ренина из ангиотензиногена, синтезируемого в
основном в печени и циркулирующего в плазме, образуется м ало­
активный пептид ангиотензин I. Ангиотензин I, в свою очередь,
под влиянием ангиотензин-превращающего фермента (А П Ф ), обра­
зующегося преимущественно в легких, превращается в ангиотен­
зин II.
Ангиотензин II обладает широким спектром ф изиологических
эффектов. Прежде всего следует отметить стимуляцию клубочко­
вой зоны коры надпочечников, секретирующей минералкортикоиды, главным образом, альдостерон. Помимо этого ангиотензин II
вызывает сужение артериальных сосудов, активирует симпатичес­
кую нервную систему, способствует формированию чувства жажды
и питьевого поведения. Учитывая эффекты альдостерона (увеличе­
ние реабсорбции N a+, C l- и бикарбонатов в почечных канальцах,
приводящее к усилению реабсорбции Н 20 и задержке жидкости в
■
Внутренняя среда организма
205
организме), понятно, что РААС является одним из важнейших меха­
низмов регуляции А Д , О Ц К , водно-солевого обмена и поведения.
Активация РА А С приводит к задержке в организме натрия, хлора,
бикарбонатов и воды, сужению сосудов, повышению А Д , ф ормиро­
ванию питьевого поведения и ряду других эффектов.
Кальцитриол
Кальцитриол, метаболит витамина D, имеет стероидную природу
и образуется в три этапа. Первый этап протекает в коже, где под
влиянием ультрафиолетовых лучей из провитамина образуется вита­
мин D. В печени витамин D превращается в соединение (25-ОН-холекальцитриол), которое как запасающая форма циркулирует в крови
в связи с а-глобулином. Завершающий, третий этап, осуществляется
в почках, где в митохондриях проксимальных канальцев образуется
собственно кальцитриол.
Основным стимулятором образования кальцитриола является
ПТГ, освобождающийся при гипокальциемии.
Основной эффект кальцитриола заключается в активации всасыва­
ния кальция и фосфатов в кишечнике, а также в стимуляции реабсор­
бции кальция и фосфатов канальцевым эпителием в почках. Влияние
кальцитриола на костную ткань связано с прямой стимуляцией осте­
областов и обеспечением костной ткани усиленно всасывающимся в
кишечнике и реабсорбирующимся в почках кальцием, что, в конечном
итоге, приводит к активации роста и минерализации костей.
Инактивация кальцитриола происходит в печени.
Эритропоэтин
У взрослых людей большая часть эритропоэтина (около 90%)
образуется в почках, в клетках эндотелия капилляров и в фибробластах коркового вещества почек. Оставшаяся часть эритропоэтина
продуцируется в печени.
Эритропоэтин — гликопротеин, который стимулирует эритропоэз
в костном мозге, регулируя цикл пролиферации и дифференцировки
клеток-предшественников эритроцитов.
Стимулируют секрецию эритропоэтина гипоксия и анемия.
Эндокринная функция сердца
В кардиомиоцитах преимущественно правого предсердия син­
тезируется пептидный гормон — П Н У Ф , имеющий и другое наиме­
нование — атриопептид. По мере секреции гормон накапливается в
206
Глава 4
специфических гранулах саркоплазмы кардиомиоцитов. Основные
стимуляторы секреции П Н У Ф :
• повышение АД ;
• растяжение предсердий (увеличение объема притекающей
крови);
• повышение [N a +] в крови;
• повышение содержания вазопрессина в крови.
Основные физиологические эффекты атриопептида:
• понижение тонуса сосудов и вазодилатация;
• подавление канальцевой реабсорбции натрия и хлора, и соответс­
твенно усиление их экскреции почками;
• повышение диуреза за счет увеличения клубочковой фильтрации
и уменьшения реабсорбции воды в почках;
• подавление секреции ренина, ингибирование эффектов ангиотензина II и альдостерона.
Таким образом, атриопептид принимает участие в регуляции АД,
О Ц К , водно-солевого обмена, являясь при этом антагонистом РААС.
В целом П Н У Ф способствует выведению из организма натрия, хлора
и воды, расширению сосудов и понижению АД.
Эндокринная функция желудочно-кишечного тракта
Эндокринные клетки Ж К Т рассеяны в слизистой оболочке и
пищеварительных железах, образуя так называемую энтериновую
систему, которая, в свою очередь, является частью диффузной эндок­
ринной системы. К летки энтериновой системы продуцируют гас­
троинтестинальные гормоны (гастрин, холецистокинин, секретин,
желудочный ингибирующий пептид, ВИП, соматостатин, энкефалин
и другие пептиды и амины), многие из которых имеют в организме
человека и другие места синтеза (например, нейроны ЦН С). В насто­
ящее время известно более 30 гормонов Ж КТ.
В пищеварительном тракте гастроинтестинальные гормоны учас­
твуют в регуляции секреции, моторики и всасывания, высвобожде­
ния других регуляторных пептидов, влияют на пролиферативные
процессы, изменяют количество гландулоцитов в слизистой оболочке
желудка и в поджелудочной железе.
Гастроинтестинальные гормоны принимают участие в регуляции
обмена веществ, пищевого поведения, деятельности эндокринной и
сердечно-сосудистой систем и других процессов жизнедеятельности.
Подробно механизмы регуляции секреции и функции гастроин­
тестинальных гормонов описаны в параграфе «Пищеварение».
■
Внутренняя среда организма
207
Роль желез внутренней секреции
в функциональных системах организма
В деятельности ф ункциональных систем гормоны выполняют
информационные и регуляторные функции и обеспечивают интегра­
цию физиологических процессов, направленных на достижение фун­
кциональными системами организма полезных приспособительных
результатов. Каждая функциональная система избирательно объеди­
няет гормоны различных желез внутренней секреции и включает их
в процессы саморегуляции.
С участием гормонов происходят кодирование метаболических
потребностей и их трансформация в мотивационное возбуждение
головного мозга и поведение.
Гормоны направленно изменяют состояние нервных центров и
способны влиять на активность рецепторов в афферентном звене
нейроэндокринных реакций.
Гормоны воздействуют на территориально разобщенные органы
и клетки-«миш ени», синхронизируют ритмы их работы и опреде­
ляю т временную последовательность физиологических процессов,
осуществляют прямые и обратные связи исполнительных органов с
центрами их регуляции.
Различные функциональные системы включают одни и те же гор­
моны в регуляцию разных приспособительных результатов на основе
изменения свойств рецепторов в соответствующих органах и тканях.
В функциональных системах железы внутренней секреции зани­
мают особое место среди вегетативных исполнительных механизмов.
Выделяемые ими гормоны вызывают множественные центральные и
периферические, а также локальные реакции функциональных сис­
тем, осуществляют системное объединение вегетативных органов и
соматических реакций, обеспечивающих активную поведенческую
деятельность для достижения полезных для организма гомеостати­
ческих или поведенческих результатов.
Системное объединение желез внутренней секреции происходит
на основе их избирательной химической чувствительности к эндо­
генным регуляторам. Железы внутренней секреции объединяются
в функциональных системах по принципу избирательного взаимосодействия.
Отклонение того или другого гомеостатического показателя от
оптимального для метаболизма уровня вызывает активацию деятель­
ности специальных желез внутренней секреции, в итоге регулиру­
208
Глава 4
емый показатель гомеостазиса удерживается в оптимальных для
метаболизма границах.
Каждая функциональная система организма имеет свою эндок­
ринную «аранжировку», обеспечивающую полезный приспособи­
тельный результат.
Это значит, что в разных функциональных системах действует
свой, специфический комплекс желез внутренней секреции, обеспе­
чивая как оптимальный уровень различных показателей внутренней
среды, так и достижение поведенческих адаптивных результатов.
Так, в функциональную систему, определяющую оптимальный уро­
вень глюкозы в крови, включаются поджелудочная железа, щитовидная
железа, гипофиз, надпочечники. В функциональной системе, обеспечи­
вающей оптимальный уровень температуры тела, задействованы щито­
видная железа, надпочечники и другие железы внутренней секреции.
Гормональный профиль организма — это обобщенные показатели
содержания гормонов во внутренних средах и жидкостях, обус­
ловленные согласованной деятельностью разнообразных ф ункцио­
нальных систем организма. Гормональный проф иль определяется
спектром различных гормонов, вырабатываемых различными типа­
ми эндокринных клеток и поступающих в кровь с определенным
ритмом, их взаимодействием, уровнем активности тканей и плазмы
крови, где реализуется «жизненный ци кл» гормонов, т.е. их связыва­
ние, расщепление и выведение.
4.3. КРОВЬ
Кровь — важнейшая внутренняя жидкая среда организма, пред­
ставленная циркулирующей в сосудистой системе плазмой и взве­
шенными в ней клеточными элементами.
Система крови включает в себя:
• собственно кровь, которая является жидкой разновидностью
соединительной ткани, состоящей из двух фракций — циркули­
рующей и депонированной;
• органы кроветворения и кроверазрушения;
• нейрогуморальный аппарат регуляции.
Функции крови
Многочисленные функции крови определяются свойствами плаз­
мы и клеточных элементов, а также тем обстоятельством, что кровь
Внутренняя среда организма
209
циркулирует в сосудистой системе, пронизывающей все ткани и
органы, и находится в постоянном обмене с интерстициальной жид­
костью. Кровь выполняет следующие функции:
• ды хательная функция заключается в транспорте кислорода
(в виде оксигемоглобина и в растворенном виде) и углекислого
газа (в составе бикарбонатов, в виде карбгемоглобина и в раство­
ренном виде);
• трофическая функция состоит в транспорте питательных веществ
(аминокислот, липидов, моно- и дисахаридов, витаминов, мик­
роэлементов и т.д.);
• экскреторная функция связана с транспортом продуктов мета­
болизма к органам выделения (почки, потовые железы, легкие,
Ж К Т );
• терморегуляторная функция связана с высокой теплоемкостью и
теплопроводностью крови, обеспечивающей поддержание пос­
тоянной температуры разных частей тела;
• регуляторная функция обусловлена транспортом ферментов,
витаминов, биологически активных веществ (в свободном виде,
в связи с белками, адсорбированными на мембране клеточных
элементов); посредством крови осущ ествляется гуморальная
регуляция физиологических функций организма;
• гомеостатическая функция направлена на сохранение относи­
тельного постоянства важнейших показателей внутренней среды
организма (например, кислотно-основного состояния);
• защитная функция реализуется в двух формах: во-первых, иммун­
ные реакции, включающие клеточный и гуморальный имму­
нитет, а во-вторых — сложный комплекс свертывающих и противосвертывающих механизмов, обеспечивающих остановку
кровотечения при повреждении сосудов и поддержание жидкого
агрегатного состояния крови в нормальных условиях.
Основные физиологические показатели крови
Общий объем крови у взрослого человека составляет примерно 6—8%
массы тела (без учета жира). Так, у взрослого мужчины массой 70 кг
объем крови составляет около 5600 мл. При этом 3,5—4 л обычно
циркулируют в сосудистом русле и полостях сердца (О Ц К ), а 1,5-2 л
депонированы в сосудах органов брюшной полости, легких, подкож­
ной клетчатки и других тканей (депонированная фракция). Объем плазмы
составляет примерно 55% общего объема крови. Клеточны е элементы
составляют около 45% общего объема крови.
210
Глава 4
Гематокрит — часть объема крови, приходящаяся на ее клетки.
Нормальное значение у женщин составляет 0,37—0,47, а у мужчин
0,40—0,54. Гематокрит, который на 99% составляют эритроциты, опре­
деляет реологические (вязкие) свойства крови. Помимо форменных
элементов, вязкость крови связана с наличием белков плазмы. Если
принять вязкость воды за единицу, то вязкость цельной крови составит
примерно 5 уел. ед. Вязкость крови, в свою очередь, влияет на скорость
ее движения по сосудам. Согласно уравнению Пуазейля, сопротивле­
ние току крови по сосудам прямо пропорционально вязкости, поэтому
увеличение гематокрита приводит к возрастанию сопротивления току
крови и, следовательно, к повышению нагрузки на сердце.
Осмотическое давление — сила, с которой молекулы (ионы) рас­
творенного вещества удерживают воду в своей гидратной оболочке
или притягивают ее через полупроницаемую мембрану из-за раз­
ницы концентраций растворенных веществ. Осмотическое давление
традиционно измерялось в атмосферах или миллиметрах ртутного
столба. Однако в настоящее время в биологической и медицинской
литературе, а главное, в клинической практике используются поня­
тия «осмолярность» и «осмоляльность» и соответствующие единицы
измерения.
Осмолярность — концентрация растворенного вещества в единице
объема раствора (мОсм/л раствора).
Осмоляльность — концентрация осмотически активных веществ в
расчете на килограмм воды (мОсм/кг Н 20).
О см оляльн ость плазмы составляет около 300 мОсм/кг Н 20.
Внутриклеточная и внеклеточная жидкости в целом осмотически
равновесны, хотя возможны кратковременные отклонения от поло­
жения равновесия.
Осмотически активны вещества, плохо проникающие через мем­
брану клеток. Основными ионами, определяющими осмоляльность
внеклеточной жидкости, являются катионы натрия, анионы хлора и
гидрокарбоната, причем наибольший вклад (более 90%) в осм оляль­
ность плазмы вносят ионы Na+. Коллоидны е соединения, преиму­
щественно белки, не проникают через мембрану и благодаря спо­
собности удерживать воду в своей гидратной оболочке создают так
называемое онкотическое (коллоидно-осмотическое) давление. Доля
онкотического давления, создаваемая белками плазмы, очень мала
по сравнению с ее общим осмотическим давлением. При нормальном
содержании белка в плазме (70 г/л) онкотическое давление составляет
всего 1,65 мОсм/кг Н 20 (из 300 мОсм/кг Н 20 ), или около 25 мм рт. ст.
Внутренняя среда организма
211
Однако величина онкотического давления имеет очень больш ое
значение, являясь одним из важнейших факторов, определяющих
транспорт воды и растворенных в ней веществ через капиллярную
стенку между кровью и межклеточной жидкостью.
П лазм а крови состоит из воды (90%) и растворенных в ней веществ
(10%, органические — 9%, неорганические — 1%; в твердом остатке на
долю белков приходится примерно 2/3, а 1/3 — низкомолекулярные
вещества и электролиты). Химический состав плазмы сходен с соста­
вом интерстициальной жидкости (преобладающий катион — N a+,
преобладающие анионы — С1~, Н С 0 3~), но концентрация белка в
плазме выше (около 70 г/л).
Содержащиеся в плазме осмотически активные вещества, т.е.
электролиты низкомолекулярных (неорганические соли, ионы) и
высокомолекулярных (коллоидны е соединения, преимущественно
белки) веществ определяют важнейшие характеристики крови —
осмотическое и онкотическое давление.
В плазме содержится более сотни различных белков, образующих­
ся в основном в печени, костном мозге, селезенке, а также поступа­
ющих из циркулирующих в крови клеточных элементов и из других
источников.
Выделяют пять электрофоретических фракций плазменных бел­
ков (альбумины и 4 фракции глобулинов).
• Альбумины (концентрация в плазме 40—45 г/л) образуются пре­
имущ ественно в печени; это белки с относительно низкой
молекулярной массой; в значительной степени они определяют
онкотическое (коллоидно-осм отическое) давление (25 мм рт. ст.,
или 3,3 кПа) крови (в 5 раз более онкотического давления меж­
клеточной жидкости). Разница в величине онкотического дав­
ления противодействует гидростатическому давлению и удер­
живает жидкость в системе кровообращения. И менно поэтому
при массивной потере альбуминов (гипоальбуминемия) через
почки развиваются «почечны е» отеки, а при голодании — «г о л о ­
дны е» отеки.
• Глобулины (25—30 г/л) — крупномолекулярные белки, подразделя­
ются на фракции:
— а,-глобулины (а,-антитрипсин, о^-ЛПВП, протромбин и др.);
— а 2-глобулины (а 2-макроглобулин, а 2-антитромбин III, а 2-гаптоглобулин, плазминоген и др.);
— р-глобулины (Р -Л П Н П , трансферрин, фибриноген и др.);
— у-глобулины (иммуноглобулины IgA, IgD, IgE, IgG, IgM ).
Глава 4
212
Ф ункции плазменных белков:
• определяют онкотическое давление крови;
• являются компонентами свертывающей (плазменные факторы
свертывания, например, протромбин, фибриноген), фибринолитической (плазминоген) и противосвертывающей (антитром­
бин I I I ) систем крови;
• участвуют в иммунных реакциях (иммуноглобулины , белки сис­
темы комплемента, макроглобулины);
• участвуют в транспорте продуктов метаболизма, жиров, гормо­
нов и других биологически активных веществ, витаминов, солей
тяжелых металлов и др. Например: альбумины (жирные кислоты),
липопротеины (холестерин), трансферрин (железо), гемоглобин,
транскортин (кортизол), транскобаламины (витамин В12) и м но­
жество других;
• влияют на вязкость крови;
• являются резервом аминокислот для построения тканевых бел­
ков;
• обладая буферными свойствами, принимают участие в регуляции
кислотно-основного состояния крови.
Л ипопротеины плазмы обеспечивают транспорт холестерина,
фосфоглицеридов, триацилглицерина и других веществ. По п лот­
ности различают Л П О Н П Л П Н П и Л П В П , а также хиломикроны.
Различная плотность липопротеинов связана с процентным содер­
жанием жиров (90% у Л П О Н П и 50% у Л П В П ). С клинической точки
зрения существенное значение имеет содержание в крови Л П Н П ,
концентрация которых в плазме прямо коррелирует с вероятностью
развития атеросклероза (атерогенность).
В плазме находятся гормоны и другие биологически активные
вещества, а также питательные вещества и продукты метаболизма
(органические кислоты, азотсодержащие вещества и др.), которые
переносятся кровью к разным органам.
Функциональная система, поддерживающая
оптимальный для метаболизма
объем циркулирующей крови
Полезный приспособительный результат
О Ц К составляет в обычный условиях больше половины общего
объема — 3,5—4,0 л.
Внутренняя среда организма
213
Постоянство О Ц К включает два фактора: постоянство объема
плазмы и постоянство массы форменных элементов, в основном
эритроцитов (рис. 4.6).
О Ц К , как и объем других жидкостных сред организма (внекле­
точных и внутриклеточных), относительно постоянен, однако может
претерпевать значительные изменения в соответствии с потребнос­
тями организма.
Увеличение О Ц К вызывает физическая деятельность. На величи­
ну О Ц К влияет изменение положения тела: при переходе человека в
вертикальное положение в результате депонирования крови в венах
конечностей и брюшной полости объем кратковременно уменьша­
ется. В состоянии сна, физического покоя О Ц К также уменьшается.
Уменьшение О Ц К может наблюдаться при повышении температуры
окружающей среды в результате интенсивного потоотделения, при
Рис. 4.6. Ф ункци ональн ая система, поддерживающая оптим альны й для
метаболизма объем циркулирующ ей крови
214
Глава 4
кровотечениях, в том числе внутренних, и при потере значительного
количества плазмы при отеках поврежденных конечностей.
Рецепция и регуляция результата
Отклонение величины О Ц К от оптимального для метаболизма
уровня воспринимается различными рецепторными образованиями.
На изменение объема крови реагируют барорецепторы сосудов, в пер­
вую очередь дуги аорты и синокаротидной области. К чувствитель­
ным образованиям, воспринимающим изменения О Ц К , относятся
и рецепторы объема — волюмрецепторы, расположенные в сердце и
сосудистом русле. При уменьшении О Ц К и ухудшении кровоснаб­
жения тканей в рецепцию результата вовлекаются также хеморецеп­
торы сосудистого русла, прежде всего синокаротидной и аортальной
областей. В связи с тем, что О Ц К в значительной степени связан с
водно-электролитным балансом организма, в рецепцию включаются
также периферические и центральные осморецепторы.
Нервные центры
Афферентация от аортальной и синокаротидной рецепторных
сосудистых зон по депрессорному и синокаротидным нервам посту­
пает в продолговатый мозг к сосудодвигательному центру. От волюмо- и осморецепторов информация об изменении О Ц К достигает
ядер гипоталамуса, оказывающих регулирующее влияние на водно­
солевой обмен организма.
Эндокринное звено саморегуляции
Гипоталамо-гипоф изарны е механизмы регуляции секреции
вазопрессина и, в меньшей степени, выделения корой надпочечников
альдостерона приводят к изменениям в деятельности почек, скорости
реабсорбции воды в Ж К Т, деятельности потовых желез.
Важная роль в регуляции объема крови принадлежит П Н У Ф
(атриопептиду) и РААС (см. раздел «Гормоны и биологически актив­
ные вещества»),
П Н У Ф секретируется кардиомиоцитами преимущественно пра­
вого предсердия при их растяжении. П Н У Ф участвует в регуляции
водно-солевого баланса, увеличивая экскрецию Na+ и воды путем
снижения их реабсорбции в проксимальных канальцах и собиратель­
ных трубочках, что приводит к уменьшению ОЦК. П Н У Ф снижает
секрецию ренина и альдостерона, а также образование ангитензинов.
Атриопептид обладает также сосудорасширяющим действием.
Внутренняя среда организма
215
Активация РА А С приводит к повышению реабсобции Na+ и воды
в почках, их задержке в организме и, соответственно, увеличению
ОЦК. Ангиотензин II, являясь дипсогенным веществом, стимули­
рует потребление воды. Кроме того, ангиотензин II оказывает вазоконстрикторный эффект.
Исполнительные механизмы
Центральные, в первую очередь гипоталамические, структуры,
получая сигнализацию об изменении О Ц К от рецепторных аппара­
тов, обеспечивают включение исполнительных механизмов, с помо­
щью которых осуществляется возвращение О Ц К к оптимальному
для жизнедеятельности уровню. Поступление афферентации к сосу­
додвигательному центру способствует включению в процесс само­
регуляции объема крови таких механизмов, как изменение Ч СС и
силы сердечных сокращений, изменение тонуса и просвета сосудов,
скорости кровотока.
Исполнительные механизмы рассматриваемой функциональной
системы следующие.
• Изменение работы сердца и скорости кровотока. При изменении
О Ц К первоначально возникают приспособительные изменения в
деятельности сердечно-сосудистой системы, имеющие компенса­
торный характер.
• Изменение просвета сосудов. В процессе саморегуляции О Ц К
существенная роль принадлежит изменению тонуса венозного
отдела сосудистого русла, в котором содержится более 60% общего
объема крови. При уменьшении О Ц К в первую очередь уменьша­
ется емкость вен, а только потом отмечается сужение артериаль­
ных сосудов как компенсаторный механизм, поддерживающий
соотношение объема крови и просвета сосудистого русла.
• Депонирование крови и перераспределение регионарного кро­
вотока. Общий О Ц К регулируется в значительной степени кро­
вяными депо — накоплением или выделением из них крови.
К основным депо крови относятся селезенка, печень, легкие, кожа.
• Перераспределение жидкости в процессе транскапиллярного
обмена зависит в основном от соотношения гидростатического и
онкотического давления крови капилляров и интерстициального
пространства, электролитного состава, pH крови, степени прони­
цаемости мембран капилляров.
• Изменение процессов выделения воды из организма (работа
почек, потовых желез). Изменение процессов мочеобразования
216
Глава 4
(в меньшей степени порообразования) и выведения воды из
организма приводит к изменению водного баланса организма и,
соответственно, ОЦК.
• Изменение интенсивности процессов кровообразования и кроверазрушения.
Внешнее звено саморегуляции — питьевое поведение
Внешнее звено саморегуляции данной функциональной системы
определяется активацией нервных центров гипоталамической облас­
ти, обусловливающих формирование ощущения жажды, питьевой
мотивации и поведения, направленного на потребление воды из
внешней среды.
М естны е механизмы саморегуляции
К местным механизмам, в первую очередь, относится гемодинамическая регуляция сердечной деятельности (см. раздел
«Кровообращ ение»),
Исполнительные механизмы рассматриваемой функциональной
системы, в первую очередь при отклонении О Ц К от оптимального для
метаболизма уровня, включают: изменение работы сердца, изменение
просвета сосудов, скорости кровотока, депонирование крови и пере­
распределение воды в организме. Более медленно происходит вовлече­
ние внешнего звена саморегуляции, а также процессов кровообразова­
ния и кроверазрушения во внутреннем звене саморегуляции.
О Ц К тесно связан с величинами кровяного и осмотического
давления в организме. Соответственно, ф ункциональные систе­
мы, направленные на поддержание оптимальных для метаболизма
величин О Ц К , А Д и осмотического давления крови, имеют неко­
торые общие центральные и периферические звенья (см. разделы
«Кровообращ ение» и «Выделение»),
Динамика работы функциональной системы при изменении объема цир­
кулирующей крови в сосудистом русле
Уменьшение ОЦК. При уменьшении О Ц К (при кровопотере, теп­
ловом стрессе и др.) первоначально возникают приспособитель­
ные изменения в деятельности сердечно-сосудистой системы.
Уменьшение объема крови приводит к снижению венозного при­
тока к сердцу, что уменьшает сердечный выброс. Компенсаторно
происходят учащение деятельности сердца, рефлекторное сужение
сосудов, выход из депо (селезенки, печени, кожи и др.) в сосудистый
Внутренняя среда организма
217
кровоток дополнительного количества крови. Происходит регио­
нальное перераспределение ОЦК.
При уменьшении О Ц К он восполняется за счет перехода воды в
сосудистое русло из интерстициальных пространств. Этот процесс
связан с повышением онкотического давления за счет усиления про­
дукции печенью белков плазмы, в частности альбуминов, которые по
лимфатическим сосудам попадают в кровь.
Снижение величины А Д , которое возникает при уменьшении
О Ц К , вызывает усиленный синтез в почках ренина. Активация РААС
способствует задержанию натрия и воды в организме, усиливая их
реабсорбцию в почках, и приводит к сужению сосудов. Помимо того,
ангиотензин II способствует формированию чувства жажды.
Снижение О Ц К приводит к увеличению секреции нейронами
гипоталамуса вазопрессина, что повышает реабсорбцию воды в дис­
тальных канальцах и собирательных трубочках, способствуя задерж­
ке жидкости в организме. Вазопрессин обладает также сосудосужи­
вающим эффектом.
Если О Ц К уменьшается, то активация нервных центров гипоталамической области формирует ощущение жажды и поведение орга­
низма, направленное на потребление воды из внешней среды.
Длительная и значительная кровопотеря включает в приспо­
собительные процессы регуляцию эритроцитарной массы крови.
Возникающая при уменьшении объема крови гипоксия является
стимулирующим фактором образования эритропоэтинов, которые
стимулируют эритропоэз. В свою очередь, уменьшение количества
эритроцитов по принципу обратной связи вызывает усиленную про­
дукцию эритропоэтина, и наоборот.
Однако существует предел потери крови, когда уже никакие при­
способительные реакции не способны восстановить О Ц К до нор­
мального уровня. Потеря 1/3 крови может привести к гибели орга­
низма. В этом случае необходимо срочное переливание крови или
кровозамещающих жидкостей.
Увеличение ОЦК. При увеличении О Ц К происходят обратные про­
цессы, способствующие снижению О Ц К . Увеличивается депони­
рование крови, происходит ее перераспределение. При увеличении
О Ц К не только вены, но и артерии способны вмещать некоторое
добавочное количество крови. Увеличение степени растяжения кар­
диомиоцитов стимулирует секрецию предсердного натрий-уретического фактора, что приводит к снижению реабсорбции натрия и воды
в почках, и, как следствие, увеличению диуреза. П Н УФ , антагонист
Глава 4
218
РААС, уменьшает выработку ренина, ангиотензинов и секрецию
альдостерона, что также способствует выведению жидкости из орга­
низма. Помимо того, П Н У Ф вызывает понижение тонуса сосудов.
Происходит снижение секреции вазопрессина, также приводящее к
повышению диуреза.
Функциональная система,
поддерживающая оптимальный уровень
клеточных элементов в крови
Функциональная система, определяющая оптимальное для мета­
болизма количество форменных элементов крови (рис. 4.7), имеет
только внутреннее звено саморегуляции. Особое место в данной фун­
кциональной системе занимают местные механизмы саморегуляции.
Распад форменных элементов, происходящий не непосредственно
в крови, а в органах кроверазрушения (селезенке, лимфатических
узлах, костном мозге), стимулирует соответственно эритропоэз, лейкопоэз и тромбоцитопоэз.
Гипоталамолимбикоретикулярные
структуры
Рецепторы
костного мозга,
селезенки,
лимфатических
узлов
Депонирование крови
Скорость кровотока
Кроворазрушение
Кровеобразован ие
Рис. 4.7. Ф ункци ональн ая система, поддерживающая оптим альны й ур о ­
вень клеточны х элементов в крови
Внутренняя среда организма
219
Клетки крови у взрослого человека образуются из гемопоэтических
стволовых клеток в красном костном мозге. Их потомки — полипотентные клетки—предшественницы миелопоэза и лимфоцитопоэза —
в процессе дифференцировки через разное количество клеточных
поколений превращаются в зрелые клеточные формы (рис. 4.8).
Сигнальными молекулами, регулирующим количество эритроци­
тов в крови, являются эритропоэтины. Рецепторы к эритропоэтинам
обнаружены в костном мозге, селезенке, лимфатических узлах, поч­
ках, а также в гипоталамусе. Перфузия этих изолированных органов
кровью с пониженным содержанием эритроцитов за счет образова­
ния эритропоэтинов вызывает в организме, в частности в костном
мозге, усиленное образование эритроцитов.
Рис. 4.8. Схема гемопоэза: C F U — колони еобразую щ ие единицы ;
B F U — взры вообразую щ ие единицы . П оли п отен тн ы е предш ест­
венники: C F U -G E M M — м иелопоэза; C F U -L y — лим ф оцитопоэза;
C F U -G M — гранулоцитов и моноцитов; C F U -G — нейтрофилов и базофилов. Унипотентны е предшественники: B F U -Е и C F U -E — эритроцитов;
C F U -M eg — м егакариоцитов;С Ри-Е о — эозиноф илов; C F U -M — м оно­
цитов
220
Глава 4
Гормоны вилочковой железы ~~ сигнальные молекулы, регулиру­
ющие содержание лейкоцитов в крови.
Синтезируемый в печени тромбопоэтин стимулирует образование
мегакариоцитов, от которых отделяются кровяные пластинки — тром­
боциты.
Интерлейкин 3, вырабатываемый Т-лимфоцитами и клетками
стромы костного мозга, поддерживает размножение практически
всех классов ранних клеток-предшественниц. Интерлейкин 3 дейс­
твует на стволовые кроветворные клетки и полипотентные клет­
ки-предш ественницы миелопоэза, а также на больш инство к ле­
ток—предшественниц миелоидного ряда, стимулируя формирование
эритроцитов, гранулоцитов, моноцитов, тромбоцитов.
В механизмах саморегуляции количества форменных элементов
крови принимают участие нервные и гуморальные регуляторные меха­
низмы — симпатическая и парасимпатическая нервная система, гормо­
ны гипофиза (АКТГ, гормон роста), вилочковой и щитовидной желез.
Эритроциты
Из красного костного мозга в кровь поступают преимущественно
незрелые эритроциты — ретикулоциты, окончательная дифференцировка которых происходит в течение 24—48 ч после выхода в крово­
ток. Зрелые эритроциты (нормоциты) — безъядерные клетки диамет­
ром 7—8 мкм, имеющие форму двояковогнутого диска. Количество
эритроцитов у женщин — 3,9-4,9х1012/л, у мужчин — 4,0-5,2х10|2/л.
Более высокое содержание эритроцитов у мужчин обусловлено тем,
что андрогены стимулируют эритропоэз. Продолжительность жизни
(время циркуляции в крови) — 100—120 сут.
Ф ункции эритроцитов
• Основная функция эритроцитов — дыхательная, а именно, транс­
порт кислорода от альвеол легких к тканям и углекислого газа от
тканей к легким. Форма двояковогнутого диска создает наиболь­
шую площадь поверхности по отношению к объему, что обеспе­
чивает максимальный газообмен между плазмой крови и эритро­
цитом. Кислород транспортируется эритроцитами в соединении
с гемоглобином. Около 20% углекислого газа также переносится
с участием гемоглобина. Примерно 2/3 С 0 2 транспортируется в
виде бикарбоната (Н С 0 3_), который является продуктом диссо­
циации Н 2С 0 3, образующейся в эритроцитах из поступающего в
них С 0 2и Н20 под влиянием фермента карбоангидразы.
Внутренняя среда организма
221
• Гемоглобин эритроцитов является важнейшей буферной систе­
мой крови, обеспечивающей поддержание кислотно-основного
равновесия.
• Участие эритроцитов в процессе свертывания крови обусловлено
содержащимися в них факторами свертывающей и противосвертывающей систем, а также их включением в состав фибринового тромба.
• Защитная функция обусловлена способностью эритроцитов
адсорбировать на своей поверхности токсические вещества.
• Эритроциты — носители разнообразных ферментов (карбоангидраза, фосфатаза и др.) и витаминов (группы В, аскорбиновая
кислота).
• Трофическая функция эритроцитов связана с переносом на их
поверхности аминокислот к клеткам тканей.
• Эритроциты адсорбируют на поверхности и осуществляют транс­
порт ряда биологически активных веществ.
• Эритроциты несут групповые признаки крови (антигены).
Практически весь объем эритроцита заполняет гемоглобин (Н Ь) —
сложное химическое соединение, состоящее из белка глобина и
четырех молекул гема, содержащего двухвалентное железо (F e2+).
Гемоглобин синтезируется эритро- и нормобластами в костном мозге,
для его синтеза необходимо достаточное поступление железа с пищей.
При разрушении эритроцитов гемоглобин превращается в билирубин
и экскретируется преимущественно в составе желчи или почками с
мочой в виде уробилина.
У мужчин в крови содержится в среднем 130-160 г/л гемоглобина,
у женщин — 120—150 г/л. В клинике принято вычислять цветовой
показатель — относительную величину, характеризующую степень
насыщения гемоглобином каждого эритроцита. Нормальными счи­
тают значения цветового показателя от 0,8 до 1,1, в этом случае эрит­
роциты считаются нормохромными. Повышение цветового показателя
называют гиперхромазией, снижение — гипохромазией.
Главная функция гемоглобина — транспорт О, к тканям и, в мень­
шей степени, перенос СО,.
В капиллярах легких при высоком р 0 2 гемоглобин связывает 0 2,
образуя оксигемоглобин (Н Ь 0 2), и в этой форме переносит 0 2 к тканям,
где происходят диссоциация оксигемоглобина и освобождение кис­
лорода, а гемоглобин становится дезоксигенированным (Н ЬН ). В виде
оксигемоглобина переносится большая часть (до 90%) кислорода.
Соединение гемоглобина с СО, с образованием карбгемоглобина
происходит в капиллярах больш ого круга кровообращения в тканях
222
Глава 4
организма. В виде карбгемоглобина транспортируется около 20%
углекислого газа.
В особых условиях происходит реакция гемоглобина с угарным
газом (СО) с образованием карбоксигемоглобина, очень прочного соеди­
нения. Гемоглобин, прочно связанный с СО, теряет способность пере­
носить 0 2. Слабое отравление угарным газом обратимо — при вды­
хании чистого воздуха происходит постепенное отсоединение СО, и
способность гемоглобина присоединять кислород восстанавливается.
Метгемоглобин (M etH b) образуется при включении в гем трехва­
лентного железа (F e3+), прочно связывает 0 2, так что диссоциация
последнего затруднена. Это приводит к нарушениям газообмена.
Образование метгемоглобина может быть наследственным или при­
обретенным, например в результате воздействия на эритроциты
сильных окислителей (нитраты и неорганические нитриты, сульфа­
ниламиды и местные анестетики, например, лидокаин).
Помимо транспорта кислорода и углекислого газа гемоглобин
участвует в поддержании кислотно-основного состояния, являясь
основной буферной системой крови.
Скорость оседания эритроцитов
Эритроциты обладают свойством осаждаться при сохранении
несвертывающейся крови (с добавлением антикоагулянта) в верти­
кальном капилляре в виде так называемых «монетных столбиков»,
над которыми образуется слой прозрачной плазмы.
Эритроциты оседают потому, что их относительная плотность боль­
ше относительной плотности плазмы. СОЭ зависит от белкового соста­
ва плазмы, а именно, от соотношения глобулинов и альбуминов. При
увеличении количества глобулинов СОЭ увеличивается. Эритроциты
в крови отталкиваются друг от друга, так как имеют на поверхнос­
ти отрицательный заряд. Уменьшение поверхностного отрицательного
заряда эритроцитов приводит к их агрегации, такие агрегаты менее
устойчивы в гравитационном поле, так как увеличена их эффективная
плотность. Белки в разной степени снижают отрицательный заряд мем­
браны эритроцитов, способствующий их взаимному отталкиванию и
поддержанию во взвешенном состоянии. Причем наибольшее влияние
оказывают фибриноген и иммуноглобулины. Поэтому в тех ситуациях,
когда увеличивается содержание этих белков (при стрессе, беремен­
ности, воспалении различного генеза, туберкулезе, онкологических
заболеваниях и др.), увеличивается и СОЭ. Повышение СОЭ вызывают
стероидные гормоны (глюкокортикоиды, эстрогены). СОЭ у здоровых
■
Внутренняя среда организма
223
мужчин составляет 2—10 мм/ч, а у женщин — 2—15 мм/ч. Изменения
СОЭ имеют большое диагностическое и прогностическое значение.
Эритропоэз
Решающий стимул для образования эритроцитов — гипоксия,
запускающая синтез эритропоэтина в почках, а у плода также в
печени. Эритропоэтин выходит в кровь и поступает в костный мозг,
где стимулирует размножение и дифференцировку унипотентного
предшественника эритроцитов и дифференцировку последующих
клеток эритроидного ряда. Дальнейш ая дифференцировка при­
водит к уменьшению размеров клеток и количества органелл, но
к увеличению содержания гемоглобина и потере ядра. Длительность
эритропоэза (от стволовой клетки до эритроцита) — 2 нед. В резуль­
тате количество эритроцитов в крови увеличивается. Соответственно
возрастает количество кислорода, поступающего в почку, что тормо­
зит образование эритропоэтина.
Удаление эритроцитов из кровотока происходит путем фагоцито­
за, в результате гемолиза и при тромбообразовании.
Ф агоцитоз
Закончившие жизненный цикл и поврежденные эритроциты
фагоцитируются макрофагами селезенки, печени и костного мозга.
Поскольку в эритроцитах нет синтезирующего белок аппарата и синтез
белка de novo невозможен, со временем в них происходит деградация
белков, снижается обмен веществ, нарушается их форма, а на поверх­
ности клетки появляются новые антигены. Такие стареющие, а также
поврежденные клетки распознаются макрофагами и фагоцитируются.
Нормально за сутки из кровотока удаляется 0,5—1,5% общей массы
эритроцитов (40 000—50 000 клеток/мкл, или около 4,2хЮ10/л).
Гемолиз эритроцитов
Гемолизом называют процесс разрушения мембраны эритроцитов
и выход гемоглобина в плазму. Виды гемолиза:
• осмотический гемолиз возникает в гипотонической среде вследс­
твие поступления в эритроцит воды по осмотическому градиенту;
• химический гемолиз вызывают вещества, разрушающие белково­
липидную мембрану эритроцитов (эфир, хлороформ и др.);
• механический гемолиз может возникать при сильных механичес­
ких воздействиях на кровь, например, при ее транспортировке или
при использовании в системах искусственного кровообращения;
224
Глава 4
• биологический гемолиз происходит при действии ядов биологи­
ческого происхождения (змей, насекомых и т.д.);
• температурный гемолиз возникает при замораживании и размо­
раживании крови в результате образования кристалликов льда.
Лейкоциты
Лейкоциты — ядерные клетки шаровидной формы. В 1 л крови
взрослого здорового человека содержится 3,8—9,8х109лейкоцитов.
В зависимости от типа гранул, находящихся в цитоплазме, лейкоци­
ты подразделяют на гранулоциты (нейтрофилы, эозинофилы, базофилы) и агранулоциты (лимфоциты и моноциты).
Увеличение числа лейкоцитов в единице О Ц К называется лейко­
цитозом, а уменьшение — лейкопенией.
Лейкоцитоз может быть обусловлен выполнением организмом опре­
деленной функции, например, лейкоцитоз во время беременности,
увеличение числа лейкоцитов в крови после приема пищи или после
длительной физической работы (так называемый функциональный
лейкоцитоз). Лейкоцитоз развивается при воспалительных процес­
сах, повреждении клеток и тканей (например, после инфарктов
или инсультов, травмы мягких тканей). Патологический лейкоцитоз
наблюдается при лейкозах и характеризуется нарушением ф ункцио­
нальной активности лейкоцитов.
Лейкопения может быть первичной (врожденной) и вторичной
(развившейся вследствие радиационного поражения, отравлений,
применения лекарственных средств).
Выраженная лейкопения характеризуется снижением им м уни­
тета. Это обусловлено тем, что лейкоциты участвуют в реализации
гуморального и клеточного звеньев иммунитета, а также фагоци­
тарной реакции.
Лейкоцитарная формула — процентное содержание в периферической
крови отдельных форм лейкоцитов. Сдвиг лейкоцитарной формулы
влево свидетельствует об увеличении количества молодых и незрелых
форм нейтрофилов, что указывает на воспалительный процесс.
Ф ункции лейкоцитов
Основная функция лейкоцитов — защитная. Лейкоциты осущ ест­
вляют фагоцитоз микроорганизмов, инородных частиц и продук­
тов распада тканей, реализуют реакции гуморального и клеточного
иммунитета, продуцируют и инактивируют различные биологически
активные вещества.
Внутренняя среда организма
225
Отличительной чертой лейкоцитов является их подвижность.
Лейкоциты имеют сократительные белки (актин, миозин) и способны
к активному перемещению, что позволяет им выходить из кровеносных
сосудов, проникая между эндотелиальными клетками (диапедез) и
разрушая секретируемыми ими ферментами базальную мембрану эндо­
телия. Направленную миграцию лейкоцитов (хемокинез, хемотаксис)
контролируют различные вещества (в том числе хемоаттрактанты).
Н ей троф и лы — наиболее м ногочисленн ы й тип лей к оц и ­
тов. Они составляю т 40—75% общ его количества лейкоцитов.
Продолжительность жизни — около 8 сут. Нейтрофилы образуются
в костном мозге и выходят в кровоток, где находятся всего несколько
часов, а свойственные им функции выполняют за пределами сосу­
дистого русла, выход из которого происходит в результате хемотакси­
са. Главная функция нейтрофилов — фагоцитоз тканевых обломков
и уничтожение опсонизированных микроорганизмов. Вместе с мак­
рофагами нейтрофилы осуществляют первичный неспецифический
иммунный ответ.
Эозинофилы — зернистые лейкоциты; составляют 1—5% лей ко­
цитов, циркулирующих в крови. Эозинофилы в течение нескольких
дней после образования остаются в костном мозге, затем несколько
часов циркулируют в крови, после чего больш инство из них мигри­
рует в ткани, контактирующие с внешней средой (слизистые о б о ­
лочки дыхательных и мочеполовых путей, кишечника). Эозинофилы
участвуют в аллергических, воспалительных и антипаразитарных
реакциях, способны к фагоцитозу. Эозинофилы вырабатывают гистаминазы, инактивирующие гистамин, и ингибитор, блокирующ ий
дегрануляцию тучных клеток.
Базофилы составляют 0—1% общего числа лейкоцитов циркулирую­
щей крови. В крови находятся 1—2 сут. Базофилы содержат в гранулах
гистамин и гепарин. Активированные базофилы покидают кровоток и
в тканях участвуют в аллергических реакциях. Выделение гистамина и
других вазоактивных факторов при дегрануляции базофилов вызывает
развитие аллергической реакции немедленного типа.
Моноциты — самые крупные лейкоциты, их количество составляет
2—9% всех лейкоцитов циркулирующей крови. Образуются в костном
мозге, выходят в кровоток и циркулируют около 2—4 сут, после чего
попадают в ткани. В тканях моноциты превращаются в макрофа­
ги. Главная функция моноцитов и образующихся из них макрофа­
гов — фагоцитоз. Макрофаги найдены во всех тканях и органах, это
мобильная популяция клеток. Продолжительность жизни макрофа­
226
Глава 4
гов — месяцы. Макрофаги фагоцитируют из крови денатурированные
белки, состарившиеся эритроциты (фиксированные макрофаги пече­
ни, селезенки, костного мозга). Макрофаги фагоцитируют обломки
клеток и тканевого матрикса. Альвеолярны е макрофаги захваты­
вают пылевые частицы различной природы, попадающие в легкие.
Специфический фагоцитоз происходит при взаимодействии макрофа­
гов с опсонизированной бактерией. Активированные макрофаги секретируют большое количество разнообразных факторов (ферменты,
транспортные белки, интерлейкины, факторы роста, тромбоксаны
и многие другие). Макрофаги проявляют антибактериальную актив­
ность, выделяя лизоцим, секретируют эндогенные пирогены.
Лимф оциты составляют 20—45% общ его числа лейкоцитов крови.
К летк и —предшественники лимфоцитов, образующиеся в костном
мозге, проходят дальнейш ую дифференцировку в первичных л и м ­
фоидных органах — тимусе (Т-лим ф оциты ) или костном мозге
(В-лимфоциты). Лимфоциты поступают в систему кровообращения
постоянно с лимфой из лимфатических узлов. Через несколько часов
они перемещаются в ткани посредством диапедеза, а затем опять
возвращаются с лимфой в кровь. Таким образом осуществляется пос­
тоянная циркуляция лимфоцитов через ткань. Продолжительность
жизни лимфоцитов составляет месяцы и даже годы.
Лимфоциты — иммунокомпетентные клетки, обеспечивающие
специфический иммунитет. С функциональной точки зрения разли­
чают В-лимфоциты, Т-лимфоциты и N K -клетки.
В-лимфоциты составляют менее 10% лимфоцитов крови. После
встречи с антигеном В-лимфоциты мигрируют во вторичные ли м ­
фоидные органы, где трансформируются в плазматические клетки.
Каждый клон синтезирует и секретирует антитела только против
одного антигена. При этом образуется клон лимфоцитов, обладаю­
щих иммунной памятью о встрече с соответствующим антигеном. Влимфоциты, трансформирующиеся в плазматические клетки, и син­
тезируемые ими антитела обеспечивают гуморальный иммунитет.
Т-лимфоциты (80% и более всех лимфоцитов). Зрелые Т-лимф о­
циты покидают тимус, находятся в крови и лимфоидных органах.
Основные функции Т-лимфоцитов — участие в клеточном и гумо­
ральном иммунитете. Т-киллеры осуществляют реакции клеточно­
го иммунного ответа. Т-хелперы поддерживают пролиферацию и
дифференцировку В-лимфоцитов, стимулируют образование цитотоксических Т-лимфоцитов, а также способствуют пролиферации и
дифференцировке супрессорных Т-лимфоцитов.
Внутренняя среда организма
227
N K -клетки — лимфоциты, лишенные характерных для Т- и В-клеток поверхностноклеточных детерминант. Эти клетки составляют
около 5—10% всех циркулирующих лимфоцитов, уничтожают транс­
формированные (опухолевые) и инфицированные вирусами, а также
чужеродные клетки.
Тромбоциты
Количество тромбоцитов в циркулирующей крови — 190—405х109/л.
Две трети кровяных пластинок находятся в крови, остальные депони­
рованы в селезенке. Продолжительность жизни тромбоцитов — 8 сут.
Старые тромбоциты фагоцитируются в селезенке, печени и костном
мозге. Циркулирующие в крови тромбоциты при повреждении сосудов
активируются и участвуют в свертывании крови и восстановлении
целостности стенки сосуда. Одно из важнейших свойств активиро­
ванных кровяных пластинок— их способность к взаимной адгезии
и агрегации, а также адгезии к стенке кровеносных сосудов. Гранулы
тромбоцитов, наряду с тромбоцитарными факторами свертывания
крови, содержат и другие вещества, в частности факторы роста, сти­
мулирующие процесс заживления ран.
Тромбоцитопоэз
Из мегакариобластов развиваются самые крупные (30—100 мкм)
клетки костного мозга — мегакариоциты. При дифференцировке мегакариоцит увеличивается в размерах, его ядро становится дольчатым.
Образуется развитая система демаркационных мембран, по которым
происходит отделение («отш нуровка») тромбоцитов. Пролиферацию
предшественников мегакариоцитов — мегакариобластов — стимули­
рует синтезируемый в печени тромбопоэтин.
Функциональная система, поддерживающая
оптимальный для метаболизма
уровень pH крови
К ислотн о-основн ое равновесие внутренней среды организма
определяется соотношением концентрации водородных Н+ и гид­
роксильных ОН~~ ионов (рис. 4.9). Кислотно-основное равновесие
оценивают по величине pH — водородному показателю, являю щ е­
муся отрицательным десятичным логарифмом концентрации ионов
водорода Н+.
pH = - lg[H +]
Глава 4
228
Д ля чистой воды pH = 7,0; для кислого раствора pH <7,0 ([Н +] в 1 л
больше 10-7); для основного pH >7,0 ([Н +] в литре меньше 10~7).
Поддержание кислотно-основного равновесия в жидких средах
жизненно важно для организма, а pH — одна из самых жестких
гомеостатических констант. Даже незначительные сдвиги pH сопро­
вождаются существенными изменениями активности ферментов,
интенсивности окислительно-восстановительных реакций, чувстви­
тельности клеточных рецепторов к биологически активным вещест­
вам, проницаемости мембран. Только при определенных значениях
pH возможен оптимальный метаболизм.
Полезный приспособительный результат
У здоровых людей pH крови может меняться в пределах 7,35—7,40.
Крайними пределами изменений pH, совместимых с жизнью, явля­
Поведение
Выделительная
деятельность почек
Потоотделение
Легочная вентиляция
Деятельность Ж КТ
Метаболическая
нейтрализация печенью
Гидрокарбонатный буфер
Фосфатный буфер
Гемоглобиновый буфер
Белковый буфер
Рис. 4.9. Ф ункци ональн ая система, поддерживающая оптим альны й для
метаболизма уровень pH крови
229
Внутренняя среда организма
ются величины от 7,0 до 7,8. Однако несовместимым с жизнью может
быть длительное изменение pH даже на 0,1—0,2.
Оптимальные (нормальные) значения pH:
• артериальной крови 7,4;
• венозной крови 7,35;
• внутриклеточной жидкости 6,8—7,2;
• интерстициальной жидкости 7,26—7,38;
• цереброспинальной жидкости 7,4—7,5.
Образующийся в процессе метаболизма С 0 2 повышает концентра­
цию ионов Н + вследствие образования и последующей диссоциации
угольной кислоты с образованием ионов водорода:
со 2+ н2о
н2со 3^ нсо3- + н+
В ходе метаболизма образуются и другие продукты, изменяющие
концентрацию Н +. Например, интенсивная мышечной работа сопро­
вождается накоплением молочной и пировиноградной кислот. Ионы
Н+ и ОН - образуются при расщеплении или перестройке (например,
при глюконеогенезе или синтезе триглицеридов в печени) некоторых
аминокислот. При гидролизе некоторых веществ, поступивших с
пищей (например, солей органических кислот), также образуются
ионы Н+ и О Н - .
Ацидоз — уменьшение pH ниже нормальных значений (увеличение
[Н +]).
Алкалоз — увеличение
pH выше норм альны х значений
(уменьш ение[Н+]).
Нарушения кислотно-основного равновесия могут быть метабо­
лическими (нереспираторными), когда первично изменяется концен­
трация водородных Н + или гидроксильных О Н ” ионов, ли бо респи­
раторными, связанными с первичными изменениями р С 0 7.
Рецепция результата
Периферические рецепторы чувствительные к изменению кон­
центрации ионов водорода Н + и парциального давления углекислого
газа р С 0 2, находятся в кровеносных сосудах и тканях. Основные
рецептивные зоны — область бифуркации сонных артерий, где рас­
положены каротидные тельца, и дуга аорты.
Центральные хеморецепторы расположены в продолговатом мозге.
Гематоэнцефалический барьер, разделяющий кровь и ткань мозга,
относительно непроницаем для ионов Н+ и Н С 0 3” , поэтому цент­
Глава 4
230
ральные хеморецепторы непосредственно не реагируют на изменение
концентрации этих ионов в крови. Однако С 0 2 легко диффундирует
через гематоэнцефалический барьер, поэтому изменение р С 0 2 в
крови достаточно быстро приводит к соответствующему изменению
р С 0 2 в межклеточной жидкости мозга и сопряженному изменению в
ней [Н+]. Увеличение или уменьшение [Н +] во внеклеточной мозговой
жидкости изменяет активность центральных хеморецепторов.
Сигнализация от рецепторов в нервные центры
Активация периферических рецепторов, расположенных в облас­
ти бифуркации сонной артерии и в дуге аорты, приводит к увели­
чению импульсации, поступающей в продолговатый мозг по синус­
ным нервам и аортальной ветви блуждающего нерва. Центральные
хеморецепторы связаны с нейронами дыхательного центра прямыми
гуморальными связями.
Нервные центры
Афферентная импульсация от периферических и прямые гум о­
ральные влияния от центральных хеморецепторов в зависимости
от параметров кислотно-основного состояния изменяют активность
нейронов дыхательного центра продолговатого мозга, что, в свою
очередь, вызывает изменение легочной вентиляции.
Исполнительные механизмы
Буферные системы крови
Буферные системы — это системы соединений, обладающих спо­
собностью связывать или отдавать ионы Н+ и О Н ", сохраняя таким
образом кислотно-основное состояние раствора при добавлении
небольших количеств кислоты или основания. Буферные системы
образованы слабой кислотой и сопряженным с ней основанием в
определенном соотношении.
Гидрокарбонатная буферная система
Эта система образована слабой угольной кислотой и гидрокарбо­
нат-ионом в соотношении 1:20.
н2со 3+ нсо3При увеличении концентрации ионов Н+ они связываются с осно­
ванием Н С 0 3_ с образованием Н 2С 0 3, которая в условиях закисления
Внутренняя среда организма
231
под влиянием карбоангидразы распадается на Н 20 и С 0 2, избыток
которого удаляется легкими (так называемая «открытая» буферная
система). При увеличении концентрации ионов О Н - они взаимодейс­
твуют с ионами водорода угольной кислоты с образованием Н 20.
Емкость гидрокарбонатной буферной системы составляет 7—9%
общей буферной емкости крови и 97—98% буферной емкости внекле­
точной жидкости (плазма, лимфа, цереброспинальная жидкость).
Фосфатная буферная система
Компонентами фосфатной буферной системы (в соотношении 1:4)
являются
н 2р о 4-
+ н р о 42-
В крови фосфатный буфер в основном обеспечивает поддержание
оптимального соотношения компонентов гидрокарбонатного буфера:
н 2со 3+ н р о 42- <-» н 2р о 4- + нсо3Белковая буферная система
Буферные свойства белков определяются их амфотерностью.
В основном растворе они диссоциируют с образованием ионов Н+
(ведут себя как кислоты), а в кислом растворе они диссоциируют
с образованием ионов О Н - . При увеличении концентрации ионов
Н + образуются малодиссоциирующие кислоты, а при увеличении
концентрации ионов ОН~ происходит образование Н 20. В обоих
случаях это будет способствовать поддержанию постоянного уровня
pH. Буферная емкость белков плазмы невелика — около 2% буферной
емкости внеклеточной жидкости.
Гемоглобиновая буферная система
Гемоглобиновая буферная система обеспечивает 3/4 (75%) всей
буферной емкости крови. Роль гемоглобина как буферного вещества
определяется не только амфотерностью, присущей белкам, но и вли­
янием оксигенации на способность гемоглобина поглощать или осво­
бождать ионы водорода. Восстановленный гемоглобин легче присо­
единяет ионы водорода, чем оксигемоглобин, являясь более сильным
основанием. В свою очередь, оксигемоглобин является более сильной
кислотой, чем восстановленный гемоглобин. Проходя через ткани
и поглощая С 0 2, кровь одновременно теряет 0 2, при этом оксиге­
моглобин переходит в восстановленный гемоглобин. Поступившая
углекислота почти полностью нейтрализуется восстановлением окси-
232
Глава 4
гемоглобина в более сильное основание, которое поглощает ионы
водорода, образующиеся при диссоциации угольной кислоты.
Изменение легочной вентиляции
Регуляция уровня pH с помощью изменения легочной вентиляции
осуществляется благодаря тому, что концентрация ионов водорода, так
же как и р С 0 2, в крови оказывает существенное влияние на дыхательный
центр. Увеличение [Н+] и р С 0 2 в крови стимулирует инспираторные ней­
роны дыхательного центра, вызывая увеличение легочной вентиляции.
Чувствительность дыхательного центра такова, что объем вентиляции
удваивается, если pH падает на 0,1 единицы, и снижается вдвое, если pH
возрастает на 0,1 единицы при прочих равных условиях.
Повышение [Н +] в крови и внеклеточной мозговой жидкости,
воздействуя на периферические и центральные хеморецепторы, при­
водит к активации нейронов инспираторного отдела дыхательно­
го центра и увеличению частоты и глубины дыхания. Повышение
легочной вентиляции приводит к усилению выведения из организма
С 0 2, образующегося при диссоциации угольной кислоты, что сопро­
вождается снижением [Н +] в крови и других жидкостях организма.
При уменьшении [Н+] возбуждение инспираторных нейронов сни­
жается, при этом альвеолярная вентиляция уменьшается, что способс­
твует задержке в организме С О э. В связи с задержкой СО., в жидких
средах организма возрастает содержание Н 2С 0 3, диссоциирующей с
образованием ионов водорода, и их концентрация [Н +] повышается.
Изменение легочной вентиляции способно достаточно быстро
(в течение нескольких минут) нормализовать содержание Н+ в крови,
предотвратив таким образом развитие ацидоза или алколоза.
Вы делительная функция почек
В почках регулируется процесс реабсорбции Н С 0 3_ и других
продуктов метаболизма из первичной мочи. В канальцах происходит
также секреция ионов водорода в обмен на реабсорбируемые ионы
натрия. Интенсивность реабсорбции и секреции кислых и основных
продуктов метаболизма меняется в зависимости от ряда факторов, в
том числе и от сдвигов кислотно-основного равновесия.
Нейтрализация кислот в печени
Печень играет основную роль в метаболической нейтрализации
кислот (окисление органических кислот, дезаминирование кислот,
синтез мочевины из аммиака и хлорида аммония).
Внутренняя среда организма
233
П отоотделение
В процессе потоотделения из организма выводится определенное
количество кислых и основных продуктов, что до некоторой степени
влияет на кислотно-основное состояние. Однако потоотделение в
механизмах регуляции уровня pH играет вспомогательную роль.
Деятельность желудочно-киш ечного тракта
Этот механизм, так же как и потоотделение, играет вспомогатель­
ную роль в регуляции кислотно-основного равновесия. Поджелудочная
железа в составе панкреатического сока секретирует значительное
количество НСОэ~. Через пищеварительный тракт из организма выво­
дится некоторое количество кислых и основных продуктов.
Внешнее звено саморегуляции
Внешнее звено саморегуляции представлено механизмами пищевого,
питьевого и солевого поведения. Кислотно-основное состояние может
изменяться за счет принимаемых питательных веществ, воды и солей.
Динамика работы функциональной системы
О тклонение pH крови от оптим ального для метаболизма уровня
прежде всего приводит к включению местных механизмов саморе­
гуляции — буферных систем крови, главным образом гидрокарбо­
натной и гемоглобиновой. Если емкости буферных систем оказы­
вается недостаточно для возвращения pH к нормальному уровню,
приходят в действие механизмы внутреннего звена саморегуляции.
Изменение [Н +] воспринимается периферическими и центральными
хеморецепторами, афферентация от которых поступает в нервные
центры. Происходят изменения легочной вентиляции, деятельности
почек, печени, пищеварительного тракта, процессов потообразования, направленные на возвращение pH к оптимальному уровню.
Функциональная система, поддерживающая pH, за счет механиз­
мов саморегуляции обеспечивает постоянство кислотно-основного
состояния крови для того, чтобы не допустить сдвига этого показате­
ля во внутриклеточной жидкости и сохранить оптимальный уровень
метаболизма тканей.
Система гемостаза
Поддержание ж идкого состоян и я крови и восстановление
целостности кровеносной системы при повреждении сосудов о б е с ­
печиваются слож ным комплексом механизмов, составляю щ их вза­
Глава 4
234
имосвязанные свертывающую, ф ибринолитическую и противосвертывающую системы, объединяемые в систему гем остаза.
Факторы свертывания крови
Свертывание крови осуществляется при взаимодействии поврежден­
ных тканей, в первую очередь, эндотелия сосудов, клеточных элементов
и плазмы крови. Соответственно, можно выделить три группы факто­
ров свертывания крови: тканевые, форменных элементов и плазменные.
При отсутствии повреждений сосудистой системы факторы свертыва­
ния находятся, как правило, в неактивном состоянии. Согласно между­
народной номенклатуре, тромбоцитарные факторы свертывания крови
нумеруются арабскими цифрами, а плазменные — римскими. К номеру
активированной формы фактора добавляют букву «а». Некоторые фак­
торы имеют собственные названия, данные по фамилиям больных, у
которых впервые был обнаружен их дефицит.
Плазменные факторы свертывания крови
• Ф актор I — фибриноген. Белок, синтезируемый в печени. Поддейс-
твием тромбина переходит в нерастворимый фибрин. Участвует в
процессе агрегации тромбоцитов.
• Ф актор I I — протромбин. Образуется в печени с участием витами­
на К. Под действием протромбиназы превращается в тромбин.
• Ф а к тор I I I — тканевой тром бопластин. Ф осф олипопротеид.
Входит в состав клеточных мембран, освобождается из клеток
тканей при их разрушении. Катализирует свертывание крови по
внешнему механизму (образование кровяной протромбиназы).
• Ф актор IV — ионы кальция. Участвует во всех фазах свертывания
крови.
• Ф актор V — Ас-глобулин плазмы, проакцелерин. Белок, синтезирует­
ся в печени. Предшественник акцелерина (Va), белка, участвующего
в образовании ферментативного комплекса — протромбиназы.
• Ф актор V I — исключен из номенклатуры.
• Ф актор V I I — проконвертин. Гликопротеид, образуется в печени
с участием витамина К. Активирует фактор X; необходим для
образования тканевой протромбиназы.
• Ф актор V I I I — антигемофильный глобулин А. Гликопротеид, необ­
ходимый для образования кровяной протромбиназы. Генетически
обусловленный дефицит этого фактора — причина гемофилии А.
• Ф актор IX — фактор Кристмаса, антигемофильный глобулин В.
Гликопротеид, синтезируется в печени в присутствии витамина К.
Внутренняя среда организма
235
Участвует в активации факторах. Генетический дефицит — при­
чина гемофилии В.
• Фактор X— фактор Стюарта-Прауэра,тромботропин. Гликопротеид,
образуется в печени с участием витамина К. Участвует в формиро­
вании тканевой и кровяной протромбиназы и входит в их состав.
• Ф актор X I — фактор Розенталя (плазменный предшественник кро­
вяного тромбопластина). Гликопротеид. После взаимодействия с
фактором X I I становится активатором фактора IX. Дефицит —
причина гемофилии С.
• Ф актор X I I — фактор Хагемана. Гликопротеид, активируется при
контакте с поврежденными тканями. Инициирует образование
кровяной протромбиназы. Активирует фактор XI.
• Ф актор X I I I — фибринстабилизирующий фактор, фибриназа.
Стабилизирует фибрин.
• Фактор Флетчера — плазменный прекалликреин. Белок, синтезируе­
мый в печени. Участвует в активации фактора X II и плазминогена.
• Фактор Фитцджеральда — высокомолекулярный кининоген. Белок,
синтезируемый в тканях. Участвует в активации факторов XI, X II и
плазминогена.
Среди плазменных выделяют группу факторов, синтез которых
зависим от витамина К (II, V II, IX, X), и факторы, образование кото­
рых не связано с наличием витамина К. Недостаточность витамина К
тормозит свертывание крови и сопровождается кровоточивостью,
подкожными и внутренними кровоизлияниями.
Ф акторы свертывания форменных элементов крови
В тромбоцитах обнаружен ряд веществ, участвующих в процессе
свертывания крови и заживления ран (факторы роста тромбоцитов).
В настоящее время известно 12 тромбоцитарных факторов свертыва­
ния, остановимся на некоторых из них.
• Тромбоцитарный тромбопластин (фактор 3), фосфолипид, компо­
нент мембраны тромбоцитов, освобождающийся при их разру­
шении. Один из наиболее важных факторов, участвует в иници­
ации образования кровяной протромбиназы.
• Антигепариновый фактор (фактор 4). Связывает гепарин.
• Фибриноген тромбоцитов (фактор 5) участвует в агрегации тромбо­
цитов. Под влиянием тромбина из фибриногена образуется фиб­
рин, нити которого входят в состав тромбоцитарного тромба.
• Тромбостенин (фактор 8) — комплекс сократительных белков,
обеспечивающих ретракцию тромба.
236
Глава 4
• Сосудосуживающий фактор (фактор 9), серотонин.
• Ф актор агрегации тромбоцитов (фактор 12), АДФ . Способствует
агрегации тромбоцитов.
Из тромбоцитов освобождается тромбоксан А 2, стимулятор агре­
гации тромбоцитов, оказывающий и вазоконстрикторный эффект.
Помимо тромбоцитов факторы свертывания крови содержатся в
эритроцитах и лейкоцитах. Так, эритроциты являются источником
эритроцитарного тромбопластина и ряда других факторов.
Тканевые факторы свертывания крови
Один из наиболее активных тканевых факторов — тромбопластин, инициирующий образование протромбиназы. Эндотелиальный
фактор Виллебранда необходим для адгезии тромбоцитов. Ткани
содержат антигепариновый фактор, соединения, подобные некото­
рым плазменным факторам (V, VII, X), вещества, стимулирующие
агрегацию тромбоцитов, активаторы фибринолиза и другие.
Сосудисто-тромбоцитарный гемостаз
Образующийся в результате сосудисто-тромбоцитарного гемос­
таза тромбоцитарный тромб способен остановить кровотечение из
сосудов микроциркуляторного русла с низким давлением крови. На
этом этапе гемостаза ведущая роль принадлежит кровяным пластин­
кам — тромбоцитам. Сосудисто-тромбоцитарный гемостаз включает
несколько процессов.
• Спазм поврежденного сосуда, способствую щ ий уменьшению
кровотечения. Первичный спазм поврежденного сосуда проис­
ходит рефлекторно за счет усиления симпатических влияний.
В дальнейшем сужение сосуда (вторичный спазм) поддерживает­
ся адреналином и вазоконстрикторами — эндотелиального про­
исхождения и освобождающимися из тромбоцитов как при их
разрушении, так и путем секреции (серотонин, тромбоксан А 2).
• Адгезия тромбоцитов. В результате повреждения сосуда обнажа­
ются волокна коллагена, к которым прилипают тромбоциты.
Прикрепление тромбоцитов происходит с помощью образуемого
клетками эндотелия белка фактора Виллебранда, взаимодейс­
твующего со специфическими рецепторами на мембране тром­
боцитов. Адгезии способствует изменение отрицательного заряда
мембраны поврежденного эндотелия сосудов на положительный.
• Обратимая агрегация тромбоцитов. Непосредственно после адгезии
происходит активация тромбоцитов, которые из дискообразных
Внутренняя среда организма
237
становятся округлыми и образуют псевдоподии, с помощью кото­
рых соединяются друг с другом. Образование псевдоподий связано
с переходом глобулярного актина в фибриллярный актин, ини­
циируемым ионами Са2+ и АДФ. В результате образуется рыхлый
тромбоцитарный тромб, который пропускает плазму.
• Необратимая агрегация тромбоцитов происходит с участием тром­
бина, образовавшегося из протромбина под влиянием тканевой
протромбиназы, которая появляется через 5—10 с после повреж­
дения сосуда. Тромбоциты теряют свою структуру и образуют
гомогенную массу. Агрегация сопровождается высвобождением
содержимого гранул тромбоцитов как в результате разрушения
пластинок, так и путем секреции (реакция высвобождения).
Высвобождаются все тромбоцитарные факторы свертывания
крови, в том числе и инициирующие коагуляционный гемостаз
(тромбопластин), и обладающие сосудосуживающим эффектом
(серотонин, тромбоксан А,), а также факторы роста.
• Ретракция тромбоцитарного тромба (уплотнение) происходит за счет
комплекса сократительных белков тромбоцитов — тромбостенина.
Сосудисто-тромбоцитарный гемостаз за счет спазма сосуда и
образования тромбоцитарного тромба приводит к остановке кро­
вотечения из сосудов микроциркуляторного русла. В норме время
остановки кровотечения из этих сосудов составляет 2—4 мин.
Образовавшийся тромбоцитарный агрегат нестабилен и не может
обеспечить остановки кровотечения из крупных сосудов с более высо­
ким давлением, так как вымывается током крови. В крупных сосудах
остановка кровотечения достигается путем образования в ходе коагу­
ляционного гемостаза более прочного фибринового тромба.
Коагуляционный гемостаз
В коагуляционном гемостазе (рис. 4.10) принято выделять три
фазы. Первая, наиболее сложная, заключается в образовании тка­
невой и кровяной протромбиназы — ферментативного комплекса,
включающего ряд факторов (Ха + Va + Са2+ + фосфолипиды).
Эта фаза представляет собой каскад реакций активации фермен­
тов, центральное место в котором занимает фактор X — тромботропин. Активация ф ак торах может происходить посредством внешней
и внутренней систем свертывания.
Внешняя система свертывания запускается тканевым тромбопластином из поврежденных стенок сосудов и окружающих тканей. Тканевой
тромбопластин активирует фактор VII, который как фактор Vila образу-
238
Глава 4
1
Плазменные
активаторы
Фибринопептид
Рис. 4.10. Схема коагуляционного гемостаза и фибринолиза
Внутренняя среда организма
239
ет комплекс с Са2+ и фосфолипидами и активирует фактор X. Фактор Ха
образует совместно с фактором Va, Са2+ и фосфолипидами фермен­
тативный комплекс — протромбиназу, в данном случае, тканевую.
Образование тканевой протромбиназы происходит за 5—10 с.
Внутренняя система свертывания, представляющая собой кас­
кад ферментативных реакций, запускается активацией фактора XII,
которая осуществляется при его контакте с волокнами коллагена,
обнажающимися при повреждении сосуда. С участием активирован­
ного им калликреина и кинина фактор ХПа активирует фактор XI и
образует с ним комплекс (ХПа + Х1а). Дальнейшие реакции внутрен­
ней системы свертывании протекают на матрице фосфолипидов раз­
рушенных тромбоцитов и эритроцитов. Под влиянием фактора Х1а
активируется фактор IX, который образует комплекс с фактором VIII
и Са2+ (IXa + V illa + Са2+). Этот комплекс адсорбируется на фос­
фолипидах и активирует фактор X. Активированный фактор Ха на
матрице фосфолипидов образует ферментативный комплекс — кро­
вяную протромбиназу (Ха + Va + Са2+ + фосфолипиды). Образование
кровяной протромбиназы происходит за 5—7 минут.
Вторая фаза коагуляционного гемостаза заключается в образова­
нии тромбина из протромбина, она протекает очень быстро, за 2—5 с.
Образовавшийся в результате первой фазы ферментативный комп­
лекс — протромбиназа (Ха + Va + Са2+ + фосфолипиды) адсорбирует
протромбин и превращает его в ферментативно активный тромбин.
Третья фаза — образование нерастворимого фибрина из фибри­
ногена.
Тромбин отщепляет от фибриногена низкомолекулярные пепти­
ды, что приводит к появлению мономеров фибрина, которые затем
под влиянием Са2+ образуют фибрин-полимер. В результате воздейс­
твия ф актораX III, активируемого тромбином, а также фибриназы
тканей и клеточных элементов образуется нерастворимый фибрин.
Нити фибрина образуют сеть, связанную с тромбоцитарным агрегатом
и окружающими тканями. Фибриновая сеть включает клеточные эле­
менты крови, главным образом, эритроциты, образуя красный тромб.
Формированием фибринового сгустка завершается этап коагуляцион­
ного гемостаза, после чего начинается дополнительный этап свертывания
крови, включающий два процесса — ретракцию тромба и фибринолиз.
Ретракция фибринового тромба и фибринолиз
Ретракция приводит к уменьшению объема сгустка крови и окон­
чательному закреплению тромба.
240
Глава 4
Образующийся в процессе коагуляционного гемостаза тром­
бин активирует актомиозиновый комплекс сократительных белков
(тромбостенин) тромбоцитов. Сокращение тромбоцитов приводит
к стягиванию сети фибриновых нитей, тромб уплотняется и умень­
шается в объеме, происходит его дальнейшее укрепление в месте
повреждения сосуда. Ретракция заканчивается через 2—3 ч после
образования сгустка.
Начинается практически одновременно с ретракцией, но идет
существенно медленнее, фибринолиз — ферментативное расщепление
фибрина. Благодаря фибринолизу происходит восстановление про­
света закупоренного фибрином сосуда. Фибринолитическое дейс­
твие оказывает фермент плазмин, находящийся в плазме в форме
неактивного плазминогена. Активный плазмин образуется из плазминогена под влиянием тканевых активаторов (внешняя система) и
активаторов крови (внутренняя система). Фибринолиз проходит в
три фазы.
Первая фаза фибринолиза заключается в образовании, освобожде­
нии и активировании активаторов плазминогена. Естественным сти­
мулятором фибринолиза является сам процесс свертывания крови.
В ответ на гемокоагуляцию происходит освобождение или активация
активаторов плазминогена.
Внешняя система активации плазминогена представлена ткане­
выми активаторами, синтезируемыми в основном клетками эндоте­
лия сосудов. Эпителий отводящих мочевых путей образует активатор
плазминогена урокиназу.
Важнейший активатор внутренней системы — активированный
фактор Хагемана (ХНа).
Активаторы плазминогена продуцируются также клеточными
элементами (лейкоцитами, эритроцитами и тромбоцитами) крови.
Во второй фазе фибринолиза профермент плазмы крови плазминоген под влиянием активаторов преобразуется в активный плазмин.
В частности, фактор ХПа высвобождает из прекалликреина калликреин, который совместно с кининогеном переводит плазминоген в
плазмин. При этом плазминоген адсорбируется фибрином, и обра­
зование плазмина происходит в фибриновом сгустке.
Третья фаза : под влиянием плазмина происходят разрыв пептид­
ных связей фибрина и его расщепление до пептидов и аминокислот.
Помимо фибрина плазмин расщепляет фибриноген, а также факто­
ры V и VIII.
Внутренняя среда организма
241
Противосвертывающие системы крови
Помимо комплекса факторов, необходимых для свертывания, в
крови содержатся вещества, препятствующие свертыванию крови, —
естественные антикоагулянты, которые делят на две группы.
Первичные, постоянно присутствующие в крови, образуют первич­
ную противосвертывающую систему, обеспечивающую поддержание
жидкого состояния крови. Важнейшие первичные антикоагулянты:
• антитромбины, ингибирующие протеазы системы свертывания,
в частности антитромбин-Ш , важнейший ингибитор тромбина;
• гепарин, продуцируемый базофилами и тучными клетками.
Гепарин тормозит тромбоцитарный гемостаз и практически все
фазы коагуляционного гемостаза, подавляя активность боль­
шинства плазменных факторов, резко усиливает ингибирующее
действие антитромбина-Ш ;
• антитромбопластины тормозят образование и действие протромбиназы.
Вторичные антикоагулянты образуются в процессе свертывания
и фибринолиза, составляя вторичную противосвертывающую сис­
тему, ограничивающую свертывание местом повреждения сосуда и
препятствующую распространению тромбообразования. Важнейшие
компоненты вторичной системы:
• образовавшийся в результате гемостаза фибрин (антитромбин-1)
адсорбирует и инактивирует до 90% тромбина;
• пептиды, отщепляемые от фибриногена тромбином при образо­
вании фибрина обладают антигоагуляционным действием;
• продукты деградации фибрина и фибриногена образующиеся при
фибринолизе являются сильными антикоагулянтами, ингибируя
тромбин и тормозя образование фибрина, а также препятствуя
агрегации тромбоцитов.
Группы крови
Антигены, определяющие группы крови, локализованы на наружной
поверхности мембраны эритроцитов (а также других клеток, например,
тромбоцитов или лейкоцитов). В плазме крови имеются (или могут выра­
батываться) антитела к соответствующим антигенам. Взаимодействие
антител с антигенами приводит к агглютинации эритроцитов. Обычно
этого не происходит, так как в крови каждого конкретного человека не
встречаются одновременно антигены и антитела к ним. Антигены групп
крови генетически детерминированы и индивидуальны.
Глава 4
242
В настоящее время известно более 300 систем «антиген—антитело»,
однако существенное практическое значение имеют две из них —
система АВО и резус-система.
Система АВО
На мембране эритроцитов присутствуют гликолипиды, молекулы
которых могут иметь антигены (агглютиногены) — специфические
углеводные компоненты (или А, или В, или А и В), определяющие
соответствующую группу крови. Молекула гликолипида может не
содержать ни компонента А, ни компонента В.
В плазме крови содержатся агглютинины (антитела, иммуноглобу­
лины класса М) к отсутствующему на мембране эритроцитов антигену.
Агглютининов в плазме крови новорожденных детей нет, они образу­
ются в течение первого года жизни.
В соответствии с типом экспрессии на поверхности эритроци­
тов агглютиногенов А и В в системе АВО выделено 4 группы крови.
Эритроциты группы крови 1 (0) не содержат ни агглютиногена А, ни
агглютиногена В. Эритроциты группы крови II (А) содержат агглютиногенА, группы крови III (В) имеют агглютиноген В, а на мембране
эритроцитов группы крови IV (АВ) представлены оба агглютиногена.
Плазма крови группы I (0) содержит а - и р-агглютинины; груп­
пы II (А) — р-агглютинины, III (В) — а-агглю тинины, плазма крови
группы IV (АВ) агглютининов не содержит (табл. 4.1).
При взаимодействии одноименных агглютиногенов и агглютини­
нов (А и а , или В и Р) происходит реакция агглютинации (склеивание
эритроцитов).
Переливание крови, не совместимой по группе, приводит к агглю­
тинации и гемолизу эритроцитов, появлению свободного гемоглоби­
на в крови и моче, может вызвать сосудистый шок и нарушение фун­
кций почек, а в тяжелых случаях — привести к гибели реципиента.
Таблица 4.1. Содержание в крови разных групп (система АВО) агглютиноге­
нов и агглютининов
Группа
Агглютиногены
Агглютинины
1(0)
Нет
а , (3
11(A)
А
III (В)
В
Р
а
IV (АВ)
А, В
нет
Внутренняя среда организма
243
Резус-система
У человека резус-систему групп крови образуют три разных анти­
гена — С, D и Е. Наиболее сильным действием обладает антиген D.
Если на мембране эритроцитов у человека присутствует антиген D,
кровь считается резус-положительной (Rh+), если нет — резус-отрицательной (Rh—). Среди европейцев примерно 85% человек имеют
резус-положительную кровь и около 15% — резус-отрицательную.
Врожденных антител к антигену D в плазме не обнаруживается.
Образование антител возможно в двух ситуациях.
Во-первых, при переливании крови резус-положительного донора
резус-отрицательному реципиенту. После этого повторное перелива­
ние резус-положительной крови будет сопровождаться агглютинаци­
ей и гемолизом эритроцитов донора.
Во-вторых, образование анти-Р-антител возможно при беремен­
ности у резус-отрицательной матери в том случае, если плод имеет
резус-положительную кровь. При этом на течение первой беремен­
ности (если не нарушена целостность плаценты) такое сочетание
крови матери и плода влияния не оказывает, поскольку исходно в
крови матери антитела отсутствуют. В процессе родов эритроциты
резус-положительного ребенка попадают в систему кровообраще­
ния его резус-отрицательной матери. В этом случае в крови матери
образуются анти-Э-антитела, которые, являясь иммуноглобулинами
класса G, имеют небольшой молекулярный вес и способны про­
никать через плацентарный барьер. При следующей беременности
резус-положительным плодом антитела из крови матери, проникая
через плаценту в систему кровообращения плода, могут вызвать
агглютинацию и гемолиз его эритроцитов. Гемолитическая болезнь
может привести к гибели плода.
Правила переливания крови определяются соответствую щи­
ми нормативными документами М инистерства здравоохранения
Минздравсоцразвития РФ, выдержки из которых приводятся ниже.
Переливание (трансфузия) компонентов крови (эритроцитсодер­
жащие переносчики газов крови, тромбоцитсодержащие и плазмен­
ные корректоры гемостаза и фибринолиза, лейкоцитсодержащие и
плазменные средства коррекции иммунитета) — лечебный метод,
заключающийся во введении в кровеносное русло реципиента ком­
понентов, заготовленных от донора или самого реципиента (аутодо­
норство), а также крови (и ее компонентов), излившейся в полости
тела при травмах и операциях (реинфузия).
244
Глава 4
Переливают компоненты, а не цельную кровь, так как показаний
к переливанию цельной консервированной донорской крови нет, за
исключением случаев острых массивных кровопотерь, когда отсутс­
твуют кровезаменители или свежезамороженная плазма, эритроцитная масса или взвесь. Цельная консервированная донорская кровь
используется при проведении обменного переливания в терапии
гемолитической болезни новорожденных.
Компоненты крови должны переливаться только той группы
системы АВО и той резус-принадлежности, которая имеется у реци­
пиента.
По жизненным показаниям и при отсутствии одногруппных по
системе АВО компонентов крови (за исключением детей) допус­
кается переливание резус-отрицательных переносчиков газов
крови группы I (0) реципиенту с любой другой группой крови в
количестве до 500 мл. Резус-отрицательная эритроцитная масса
или взвесь от доноров группы II (А) или III (В) по жизненным
показаниям могут быть перелиты реципиенту с IV (АВ)-группой,
независимо от его резус-принадлежности. При отсутствии одногруппной плазмы реципиенту может быть перелита плазма груп­
пы IV (АВ).
Во всех без исключения случаях переливания эритроцитсодержа­
щих компонентов крови абсолютно обязательным является проведе­
ние до начала переливания проб на индивидуальную совместимость
и в начале трансфузии — биологической пробы.
Запрещается переливание компонентов крови, предварительно не
исследованных на ВИЧ, гепатиты В и С, сифилис.
Врач, производящий трансфузию компонентов крови, обязан,
независимо от произведенных ранее исследований и имеющихся
записей, лично провести следующие контрольные исследования
непосредственно у постели реципиента.
• Перепроверить группу крови реципиента по системе АВО и
Rh, сверить полученный результат с данными в истории бо­
лезни.
• Перепроверить группу крови по системе АВО и Rh из донорского
контейнера и сопоставить результат с данными на этикетке кон­
тейнера.
• Провести пробы на индивидуальную совместимость по системам
АВО и резус эритроцитов донора и сыворотки реципиента.
• Провести биологическую пробу.
Внутренняя среда организма
245
Функциональная система, поддерживающая
оптимальное для метаболизма количество
глюкозы в крови
Углеводы играют роль важнейшего источника энергии, обеспе­
чивающего энергетические потребности и, соответственно, нор­
мальное функционирование организма. Углеводы способны быстро
извлекаться из депо, распадаться и окисляться, что особенно важно
для обеспечения быстро нарастающих энергетических затрат в усло­
виях интенсивной физической деятельности, при эмоциональном
возбуждении, при понижении температуры тела. Особенно большое
значение содержание глюкозы в крови имеет для клеток нервной
системы и мышц.
Углеводы выполняют в организме и пластические функции. Так,
например, образующиеся при окислении глюкозы пентозы входят в
состав нуклеотидов и нуклеиновых кислот.
Уровень глюкозы в крови является одним из интегральных пока­
зателей внутренней среды, отражающим обмен углеводов, белков и
жиров в организме человека. Глюкоза служит энергетическим суб­
стратом, необходимым для процессов синтеза жиров и белков, источ­
ником синтеза жиров. С другой стороны, в печени из аминокислот и
жирных кислот происходит новообразование углеводов.
Ф ункциональная система, поддерживающая оптимальный для
метаболизма уровень глюкозы в крови, представлена на рис. 4.11.
Полезный приспособительный результат функциональной
системы
Оптимальное содержание глюкозы в крови, обеспечивающее
нормальный уровень метаболических процессов в тканях, — 3,3—
5,5 ммоль/л.
В венозной крови содержание глюкозы обычно несколько ниже,
чем в артериальной, так как часть ее утилизируется тканям и.
Задержка глюкозы из крови разными органами неодинакова. Больше
всего глюкозы задерживает мозг (до 12%), некоторые органы (селе­
зенка и легкие) практически не задерживают глюкозу, а используют
гликоген, поступающий к ним с кровью. Артериовенозная разница
содержания глюкозы прямо связана с интенсивностью метаболизма
в тканях, при ее увеличении в период активной работы поглощение
глюкозы возрастает, и артериовенозная разница увеличивается.
246
Глава 4
Глю-
О козорецепторы
Л сосу­
дов
Рис. 4.11. Функциональная система, поддерживающая оптимальный для
метаболизма уровень глюкозы в крови
Снижение содержания глюкозы в крови называется гипогликеми­
ей, повышение — гипергликемией.
Запасов глюкозы в циркулирующей крови достаточно для обес­
печения жизнедеятельности на 3—5 мин, поэтому необходимо пос­
тоянное восполнение потребляемой глюкозы за счет мобилизации
углеводов из депо и новообразования глюкозы. Гипогликемия может
возникнуть у здорового человека при истощении запасов гликогена
во время длительного голодания, при уменьшении секреции гормо­
нов контринсулярного действия, а у больных сахарным диабетом —
при передозировке инсулина. Наиболее чувствительны к гипогли­
кемии нейроны ЦНС, для которых глюкоза служит практически
единственным источником энергии, а депо гликогена в ЦНС отсутс­
твует. Незначительная гипогликемия проявляется общей слабостью
Внутренняя среда организма
247
и быстрой утомляемостью. Характерные симптомы гипогликемии —
мышечная слабость, тремор, потоотделение, учащение сердцебиения,
головокружение, сильный голод. При снижении содержания глюкозы
в крови ниже 3,0 ммоль/л появляются судороги, угнетение функций
мозга вплоть до потери сознания (гипогликемическая кома). Резкая
гипогликемия может привести к смерти. Проявления гипогликемии
ослабляются или исчезают при приеме сахара или введении в кровь
глюкозы.
Кратковременное повышение уровня глюкозы в крови не является
непосредственной угрозой для жизни, но может приводить к повы­
шению осмотического давления крови.
Периодические изменения уровня глюкозы в крови у человека и
животных обусловлены суточными колебаниями, приемом пищи,
эмоциональным состоянием и возрастными особенностями.
Суточные колебания. В полночь содержание глюкозы в крови у
человека примерно на 0,6 ммоль/л выше, чем днем.
После приема пищи, особенно богатой углеводами, содержание глю­
козы в крови быстро увеличивается (алиментарная гипергликемия).
Гипергликемия, обусловленная как всасыванием углеводов в кишечнике,
так и выделением глюкозы из печени в ответ на поступление с кровью из
воротной системы, инициирует активацию механизмов саморегуляции,
обеспечивающих снижение концентрации глюкозы в крови (см. ниже).
Для оценки динамики содержания глюкозы в крови у человека
после приема углеводов проводят тест на толерантность к глюкозе.
Определив уровень глюкозы натощак, пациенту предлагают выпить
раствор, содержащий 50-100 г сахарозы или глюкозы. Концентрацию
глюкозы в крови определяют через каждые 30 мин на протяжении
3—4 ч. Полученные результаты позволяют определить ход так назы­
ваемой «сахарной кривой».
У здорового человека через 30 мин после приема глюкозы содер­
жание ее в крови возрастает до 9,6—10,8 ммоль/л, после чего концен­
трация глюкозы начинается уменьшаться и через 2—2,5 ч наступает
кратковременное снижение содержания глюкозы по отношению к
исходному уровню. Гипогликемическая фаза «сахарной кривой» крови
обусловлена избыточной секрецией инсулина в ответ на гипергли­
кемию. К концу третьего часа содержание глюкозы возвращается к
исходному уровню. У больных сахарным диабетом при недостаточной
секреции инсулина исходный и максимальный уровни глюкозы выше,
гипогликемическая фаза отсутствует, к окончанию третьего часа после
приема углеводов концентрация глюкозы превышает исходную.
248
Глава 4
Возрастные особенности
Возрастные различия содержания глюкозы в крови связаны с
особенностями продукции глюкагона и инсулина поджелудочной
железой в разные возрастные периоды, а также со зрелостью меха­
низмов саморегуляции. У новорожденных содержание глюкозы в
крови натощак колеблется в пределах 1,8—3,0 ммоль/л, у грудных
детей — 4—5 ммоль/л, у более старших —4,8—6,0 ммоль/л. Относительно
высокое содержание глюкозы (5,4—7,2 ммоль/л) наблюдается у детей в
возрасте 12—14 лет, соответствующем пубертатному периоду.
В детском и юношеском возрасте в поджелудочной железе преоб­
ладают островки, содержащие р-клетки, продуцирующие инсулин.
В старческом возрасте, напротив, преобладают островки, содержа­
щие а-клетки, вырабатывающие глюкагон.
Уровень глюкозы в крови новорожденных и грудных детей изза несовершенства нейроэндокринных механизмов саморегуляции
очень неустойчив.
После 50 лет выносливость (толерантность) к углеводам снижа­
ется, изменяется ход «сахарной кривой» после приема пищи, харак­
теризующийся увеличением максимальных значений алиментарной
гипергликемии.
Повышение уровня глюкозы в крови, наблюдаемое при эмоцио­
нальном возбуждении, в ситуации мобилизации к деятельности, а также
при болевых раздражениях, связано, в первую очередь, с активацией
симпатоадреналовой системы.
Изменение уровня глюкозы в крови отмечается при многих инфек­
ционных заболеваниях: гриппе, скарлатине, дифтерии, туберкулезе.
В целом уровень глюкозы в крови является достаточно пластич­
ным гомеостатическим показателем, колебания которого могут быть
обусловлены различными внешними и внутренними факторами.
Изменение содержания глюкозы в крови обеспечивает адаптацию
организма к условиям окружающей среды, к характеру и интенсив­
ности деятельности.
Рецепция уровня глюкозы в крови
Хеморецепторы, чувствительные к изменению концентрации глю­
козы в крови (глюкорецепторы), расположены в печени, сосудах, ЖКТ
и ЦНС. Подтверждением этому служат опыты с изолированными
органами, сохранившими нервную связь с организмом. При введении
в сосуды этих органов растворов глюкозы наблюдается снижение ее
содержания в общем кровотоке. Рефлекторно изменяется уровень
Внутренняя среда организма
249
глюкозы в крови при воздействии растворов глюкозы различной кон­
центрации на слизистые оболочки ротовой полости и желудка.
Центральные глюкорецепторы расположены в вентромедиальном
отделе гипоталамуса.
Нервные центры
Начало изучения нервных механизмов регуляции содержания глю­
козы в крови было положено знаменитым экспериментом выдающего­
ся французского физиолога К. Бернара в 1849 г., вошедшим в историю
физиологии под названием «сахарного укола». Сущность опыта состо­
ит в том, что при уколе в участок продолговатого мозга в области дна
1Ужелудочка, ограниченного местом выхода слуховых и блуждающих
нервов, происходит увеличение содержания глюкозы в крови.
Центральная роль в механизме регуляции содержания глюкозы в
крови принадлежит гипоталамусу, регулирующие влияния которого
на углеводный обмен реализуются через BHC и гипоталамо-гипофизарную систему. Посредством рилизинг- и ингибирующих гормонов
гипоталамус оказывает влияние на продукцию клетками аденогипо­
физа АКТГ, СТГ и ТТГ, которые, в свою очередь, влияют на деятель­
ность периферических желез внутренней секреции, в частности щито­
видной железы, надпочечников и поджелудочной железы.
О
важной роли мозжечка в регуляции концентрации глюкозы в
крови свидетельствуют результаты экспериментальных исследований
и клинико-морфологических наблюдений, выявившие изменения
содержания глюкозы при поражениях данной структуры мозга.
Кора больших полушарий головного мозга координирует уро­
вень содержания глюкозы в крови с поведением организма, которое
обеспечивает его адекватное приспособление к окружающей среде.
Повышение концентрации глюкозы в крови отмечено у спортсменов
в предстартовом состоянии и во время соревнований, а также у сту­
дентов во время экзаменов.
В целом регуляция уровня глюкозы в крови происходит при
участии ряда структур, расположенных на разных уровнях ЦНС, в
частности, продолговатого мозга, промежуточного мозга, мозжечка,
коры больших полушарий.
Исполнительные механизмы
Печень регулирует направленность углеводного обмена, поддержи­
вая концентрацию глюкозы в крови на оптимальном для метаболиз­
ма уровне.
250
Глава 4
Роль печени в углеводном обмене была исследована К. Бернаром
(1853), установившим, что в печени содержится гликоген, из которого
образуется глюкоза крови. Было показано, что животные, у которых
удалена печень, умирают от недостатка в крови глюкозы и жизнь
животного в первые часы после такой операции можно поддержать
только повторным внутривенным введением глюкозы. Впоследствии
было установлено, что при перфузии изолированной печени рас­
твором глюкозы количество гликогена в ней увеличивается. Эти и
другие исследования позволили выявить важнейшую роль печени в
углеводном обмене.
В печени происходит гликогенез — синтез гликогена из глюкозы и
продуктов промежуточного распада углеводов, которые ресинтезируются в глюкозу. Гликоген печени — форма депонирования углеводов,
его количество у взрослого человека может достигать 150—200 г.
Для восполнения постоянно убывающей (вследствие потребления
тканями) глюкозы крови в печени происходит гликогенолиз — рас­
щепление гликогена до глюкозы, поступающей в кровяное русло.
Также в печени осуществляется глюконеогенез — образование угле­
водов из продуктов метаболизма белков и жиров.
Происходящие в печени гликогенез, гликогенолиз и глюконеоге­
нез — взаимосвязанные процессы, интенсивность которых определя­
ется концентрацией глюкозы в притекающей крови. При понижении
уровня глюкозы в крови усиливаются распад гликогена и глюко­
неогенез, повышение концентрации глюкозы стимулирует синтез
гликогена.
Благодаря наличию таких механизмов печень способна самостоя­
тельно регулировать содержание глюкозы в крови. Однако в естествен­
ных условиях в целостном организме этот местный регуляторный про­
цесс находится под контролем нервной системы и эндокринных желез.
Трансмембранный транспорт глюкозы
Транспорт глюкозы через клеточные мембраны осуществляется с
помощью специальных переносчиков. К настоящему времени иден­
тифицировано 8 трансмембранных переносчиков глюкозы (GLUT), в
том числе GLUT1 в эритроцитах, GLUT2 — в островках Лангерганса
и печени, GLUT5 — в канальцах почек. Особо следует указать пере­
носчик, чувствительный к инсулину — GLUT4. Инсулин активирует
поступление глюкозы в клетки печени, скелетных мышц, жировой
ткани, способствуя встраиванию в плазматическую мембрану клеток«мишеней» дополнительного трансмембранного переносчика GLUT4.
Внутренняя среда организма
251
Всасывание моносахаридов в кишечнике
Углеводы в тонком кишечнике всасываются в форме моноса­
харидов. Глюкоза и некоторые другие моносахариды всасываются
активно (с затратой энергии) с помощью встроенных в апикальную
плазматическую мембрану энтероцитов К а+-зависимых транспор­
теров сочетанного переноса ионов натрия и глюкозы (вторично­
активный натрий-зависимый транспорт). Выход глюкозы из энтеро­
цитов через базальную мембрану во внутреннюю среду организма
происходит с помощью облегченной диффузии. Транспорт глюкозы,
являясь активным процессом, зависит от интенсивности обмена
веществ в тонком киш ечнике и тормозится ингибиторами тканевого
дыхания.
Всасывание фруктозы и некоторых других моносахаридов не зави­
сит от натрия и осуществляется пассивно.
Выделение глюкозы почками
Фильтрация глюкозы из кровеносных капилляров почечных телец
в полость капсулы осуществляется пропорционально ее концентра­
ции в плазме крови.
Реабсорбция глюкозы происходит преимущественно в началь­
ных отделах проксимальны х извитых канальцев со скоростью
300—375 мг/мин. Обычно у здоровых людей глюкоза реабсорбиру­
ется полностью. В эпителий канальцев глюкоза поступает пос­
редством активного сочетанного транспорта с Na+ с помощью
специальных котранспортеров, а выходит из клеток путем облег­
ченной диффузии через натрий-независимые транспортеры GLUT.
Глюкоза относится к так называемым «пороговым» веществам,
реабсорбция которых зависит от их концентрации в крови. Порог
выведения — такая концентрация вещества в крови, при превы­
шении которой оно не может быть полностью реабсорбировано в
канальцах и попадает в конечную мочу. Порог выведения глюкозы
составляет около 10 ммоль/л. Если концентрация глюкозы в крови
превышает пороговую величину, она частично выводится с мочой
(глюкозурия). Секреции глюкозы в просвет канальцев нефрона у
здоровых людей не происходит.
Распад углеводов в организме человека и животных происходит
как путем бескислородного расщепления (анаэробный гликолиз),
так и окисления до углекислого газа и воды (аэробный гликолиз).
Скорость окисления глюкозы может увеличиваться в несколько раз,
например под влиянием физической нагрузки.
252
Глава 4
Эндокринное звено саморегуляции
Основные эндокринные железы, принимающие участие в регу­
ляции углеводного обмена, — гипофиз, поджелудочная и щитовид­
ная железы, а также корковое и мозговое вещество надпочечников.
Секреторная функция этих желез находится под контролем гипо­
таламуса, реализуемого через гипоталамо-гипофизарную систему и
ВНС. Гуморальная регуляции уровня глюкозы в крови осуществля­
ется комплексом гормонов, только один из которых — инсулин —
обладает выраженным гипогликемическим действием. Действие
остальных гормонов, принимающих участие в регуляции углеводно­
го обмена, направлено на повышение содержания сахара в крови.
Гипофиз
СТГ. Влияние СТГ на углеводный и жировой обмен имеет две
фазы. В начальной фазе СТГ оказывает инсулиноподобный эффект,
увеличивая поглощение глюкозы мышцами и жировой тканью, одно­
временно тормозя в последней липолиз.
Для второй фазы, развивающейся через некоторое время, харак­
терны контринсулярные эффекты СТГ. Тормозятся поглощение и
утилизация глюкозы тканями. Усиливается липолиз, приводящий
к освобождению жирных кислот, стимулируется глюконеогенез.
Гормон повышает активность а-клеток островков Лангерганса и про­
дукцию глюкагона. Одновременно активируется инсулиназа печени
и усиливается разрушение инсулина. Таким образом, СТГ оказывает
гипергликемическое действие.
Уменьшение концентрации глюкозы в плазме вызывает подъем
уровня СТГ, напротив, повышение концентрации глюкозы в крови
подавляет нормальную выработку СТГ.
АКТГ гипофиза оказывает влияние на углеводный обмен, главным
образом, усиливая продукцию глюкокортикоидов корковым вещес­
твом надпочечников.
ТТГ гипофиза влияет на углеводный обмен, контролируя продук­
цию иодсодержащих гормонов щитовидной железы.
Кора надпочечников
Глюкокортикоиды повышают уровень глюкозы в крови. Эти гормо­
ны стимулируют глюконеогенез в печени, активируя синтез соответс­
твующих ферментов. При этом глюкокортикоиды усиливают распад
белков скелетных мышц, соединительной и костной ткани, а также
Внутренняя среда организма
253
липолиз. Образовавшиеся метаболиты (аминокислоты и свободные
жирные кислоты) поступают в печень, где включаются в процессы
глюконеогенеза. Глюкокортикоиды также подавляют утилизацию
глюкозы тканями (за исключением ЦНС).
Мозговое вещество надпочечников
Под влиянием адреналина повышается уровень глюкозы в крови.
К гипергликемии приводят усиление расщепления и подавление
синтеза гликогена в печени и мышцах, снижение потребления глю­
козы тканями, усиление глюконеогенеза в печени. Липолитическое
действие адреналина приводит к повышению содержания свободных
жирных кислот в плазме.
Одновременно адреналин через а-адренорецепторы, преобладаю­
щие в островках Лангерганса, подавляет секрецию инсулина.
Щ итовидная железа
Иодсодержащие гормоны щитовидной железы стимулируют все
этапы метаболизма углеводов, вызывая в целом гипергликемический
эффект. Под влиянием иодтиронинов усиливается распад гликогена,
повышается всасывание глюкозы в кишечнике, ускоряется инакти­
вации инсулина в печени. При этом гормоны увеличивают потреб­
ление и окисление глюкозы клетками в печени, жировой ткани и
мышцах.
Эндокринная функция поджелудочной железы
Основными гормонами поджелудочной железы, регулирующими
углеводный обмен, являются инсулин и глюкагон, секреция которых
определяется, главным образом, концентрацией глюкозы в крови, а
также влияниями со стороны ВНС и со стороны некоторых других
гормонов. В частности, модулирующее влияние на секрецию инсу­
лина и глюкагона оказывает соматостатин, синтезируемый в клетках
островков Лангерганса. Так как гормоны островковых клеток секретируются в кровь панкреатической вены, впадающей в воротную
вену печени, в печени концентрация этих гормонов существенно
выше, чем в остальной системе кровообращения.
Инсулин. Пептидный гормон, синтезируется в (3-клетках ост­
ровков Лангерганса. Контролирует многие процессы метаболизма,
стимулируя синтез соединений, являющихся формой депонирования
питательных веществ в организме (гликоген, триглицериды и др.).
254
Глава 4
Активация инсулинозависимых переносчиков глюкозы (GLUT4)
стимулирует транспорт глюкозы через мембрану в скелетных мыш­
цах, сердце, жировой ткани. В мышечных клетках инсулин стимули­
рует образование мышечного гликогена, а в жировых — образование
триглицеридов.
В печени инсулин усиливает поглощение глюкозы и ее накопление
в форме гликогена, тормозит распад гликогена и глюконеогенез.
Инсулин стимулирует гликолиз и окисление углеводов в печени,
мышцах и жировой ткани.
Следствием этих процессов становится понижение уровня глю­
козы в крови.
Основной фактор, регулирующий секрецию инсулина, — концен­
трация глюкозы в крови. Увеличение концентрации глюкозы прямо
стимулирует секрецию инсулина, а уменьшение — тормозит. При
снижении концентрации глюкозы в плазме ниже 3,6 ммоль/л секре­
ция инсулина прекращается.
Стимулируют секрецию инсулина некоторые гастроинтестиналь­
ные гормоны (гастрин, секретин, холецистокинин). Адреналин и
норадреналин через а 2-адренорецепторы, преобладающие в остров­
ках Лангерганса, угнетают секрецию инсулина. Тормозит секрецию
инсулина соматостатин поджелудочной железы (паракринно).
Парасимпатические влияния стимулируют секрецию инсулина,
а симпатические — преимущественно тормозят.
Глюкагон. Синтезируется в а-клетках островков Лангерганса.
В регуляции углеводного обмена глюкагон является антагонистом
инсулина, обладая гипергликемическим действием и обеспечивая
снабжение тканей глюкозой.
Глюкагон, активируя гликогенфосфорилазу, стимулирует рас­
щепление гликогена, высвобождение глюкозы и ее поступление в
кровь. Ингибируя гликогенсинтетазу, тормозит образование глико­
гена. Стимулирует глюконеогенез.
Наряду с а-клетками островков Лангерганса глюкагон выраба­
тывается эндокринными клетками слизистой оболочки желудка и
двенадцатиперстной киш ки (диффузная эндокринная система).
Секреция глюкагона усиливается при понижении концентра­
ции глюкозы в крови, при продолжительной физической нагрузке,
в условиях стресса. СТГ повышает активность а-клеток и продук­
цию глюкагона. Симпатические влияния стимулируют секрецию
глюкагона. Подавляют секрецию глюкагона инсулин, соматостатин,
глюкоза крови.
Внутренняя среда организма
255
Динамика работы функциональной системы при
гипер- и гипогликемии
Гипергликемия у здорового человека развивается в результате поступ­
ления углеводов с пищей или при внутривенном введении глюкозы.
После приема пищи глюкоза и другие моносахариды, всосавшиеся
из кишечника в кровь, по воротной системе поступают в печень, где
поглощаются гепатоцитами. При этом различные моносахариды пре­
вращаются в глюкозу. Повышение концентрации глюкозы активиру­
ет ферменты углеводного обмена печени, превращающие глюкозу в
гликоген, тормозя одновременно распад гликогена и глюконеогенез
(гомеостатическая функция печени). Глюкоза, не захваченная гепа­
тоцитами, поступает в общую систему циркуляции.
В ответ на повышение уровня глюкозы в крови, протекающей
через поджелудочную железу, возрастает секреция инсулина р-клет­
ками островков Лангерганса. Секрецию инсулина при поступлении
пищи стимулируют такие гастроинтестинальные гормоны, как гас­
трин, холецистокинин, секретин. Повышение содержания глюкозы в
плазме, воспринимаемое периферическими и центральными глюко­
рецепторами, приводит к активации нейронов гипоталамуса, кото­
рые путем усиления парасимпатических влияний на поджелудочную
железу усиливают секрецию инсулина.
Инсулин увеличивает проницаемость клеточных мембран для
глюкозы, повышает степень утилизации глюкозы тканями, активи­
рует в печени и мышцах образование гликогена, в жировой ткани
стимулирует образование жира из глюкозы, тормозит распад глико­
гена в печени и глюконеогенез.
Если концентрация глюкозы в плазме и, соответственно, в пер­
вичной моче превышает пороговое значение (около 10 ммоль/л), она
не реабсорбируется в почечных канальцах полностью и частично
выводится из организма с мочой.
Повышение концентрации глюкозы в крови подавляет выработку
контринсулярных гормонов, в первую очередь, глюкагона и СТГ.
В совокупности все эти процессы приводят к понижению уровня
глюкозы в крови.
Гипогликемия может возникнуть при длительном голодании, при
длительной изнурительной мышечной работе, при истощении резер­
ва гликогена в печени.
Понижение уровня глюкозы в крови активирует ферментативные
системы печени, усиливающие распад гликогена и глюконеогенез, и
тормозит образование гликогена (гомеостатическая функция печени).
256
Глава 4
Уменьшение концентрации глюкозы в крови непосредственно
влияет на островковые клетки поджелудочной железы, тормозя сек­
рецию инсулина и стимулируя секрецию глюкагона. Снижение сек­
реции инсулина тормозит процессы образования гликогена из глю­
козы в печени и мышцах, уменьшает утилизацию глюкозы тканями.
Глюкагон, активируя гликоген-фосфорилазу и ингибируя гликоген-синтетазу в гепатоцитах, стимулирует распад гликогена печени,
одновременно гормон усиливает глюконеогенез и подавляет гли­
когенез в печени, способствуя, таким образом, повышению уровня
глюкозы в крови.
Импульсация от периферических глюкорецепторов и непосредс­
твенное влияние гипогликемической крови на центральные глю­
корецепторы активируют гипоталамические центры, реализующие
свое влияние на углеводный обмен за счет повышения активности
симпатической нервной системы и через гипоталамо-гипофизарную
систему, стимулируя секрецию СТГ и АКТГ.
М обилизация симпатической нервной системы и увеличение сек­
реции адреналина надпочечниками приводят к торможению про­
дукции инсулина и стимуляции секреции глюкагона поджелудоч­
ной железой. Адреналин стимулирует распад гликогена в печени и
мышцах, усиливает глюконеогенез. Таким образом, активация симпатоадреналовой системы приводит к повышению уровня глюкозы
в крови.
Под влиянием СТГ уменьшается проницаемость клеточных мемб­
ран для глюкозы, усиливается глюконеогенез, активируется секреция
глюкагона, в результате чего содержание глюкозы в крови возрастает.
АКТГ стимулирует в коре надпочечников секрецию глюкокортикоидов, которые, в свою очередь, активируя глюконеогенез в печени,
способствуют повышению содержания глюкозы в крови.
Гипогликемия — один из факторов, стимулирующих нейроны
центра голода гипоталамуса, что приводит к формированию пищевой
мотивации и поведения, направленного на прием пищи и удовлетво­
рение потребности в питательных веществах.
Глава 5
ВИСЦЕРАЛЬНЫЕ ФУНКЦИИ
5.1. ФИЗИОЛОГИЯ СЕРДЦА
Общие представления о циклической работе сердца
Работа сердца происходит ритмически. Сердечный цикл — это
работа правой и левой половин сердца, которая складывается из
автоматически возникающего в сердце возбуждения, распростра­
нения возбуждения по сердцу, последовательного сокращения (сис­
толы) предсердий и желудочков и последовательного расслабления
и отдыха (диастолы) предсердий и желудочков. Результатом сер­
дечного цикла является выброс из сердца ударного объема крови.
Понятие «сердечный цикл» отражает периодичность работы сердца.
Правая и левая части сердца работают содружественно, сокращение
и расслабление их происходит одновременно. Это обусловлено про­
водящей системой сердца, которая обеспечивает одновременность
работы правой и левой половин сердца. Однако кровь дважды
проходит через сердце. Поступившая в правую половину сердца
кровь выбрасывается в малый круг кровообращения. Вернувшаяся
в левую половину сердца кровь выбрасывается в большой круг кро­
вообращения.
Сердце представляет собой мышечный орган для перекачки крови
с собственной проводящей системой, в которой самопроизвольно
возникает возбуждение. После наполнения камер сердца кровью про­
исходит сокращение миокарда, и кровь под давлением выбрасывается
в кровеносные сосуды. Таким образом, строение сердца обеспечивает
нагнетание крови в сосуды порциями.
Ритмическое возбуждение сердца
Периодичность работы сердца связана с ритмическим возник­
новением в нем возбуждения и проведением его ко всем кардиомиоцитам, что вызывает их сокращение. Возбуждение возникает и
258
Глава 5
проводится в проводящей системе сердца (рис. 5.1.1А). Проводящая
система сердца состоит из малодифференцированных атипических
мышечных волокон, которые возбуждаются, проводят возбуждение,
но не сокращаются. В проводящую систему сердца входят синоат­
риальный (синусный) или синусно-предсердный узел, проводящие
волокна предсердий, атриовентрикулярный или предсердно-желудоч­
ковый узел, пучок Гиса и волокна Пуркинье.
Синусно-предсердный узел состоит из сердечных проводящих
кардиомиоцитов, расположенных под эпикардом между ушком пра­
вого предсердия и местом впадения в сердце верхней полой вены.
Синусно-предсердный узел является очагом автоматизма сердца, в
котором первично возникает возбуждение. От синусно-предсердного
узла возбуждение проводится по проводящим волокнам предсердий,
возбуждая в них типичные кардиомиоциты, и достигает предсердножелудочкового узла. От предсердно-желудочкового узла возбуждение
проводится к желудочкам по пучку Гиса, который в перегородке
сердца делится на правую и левую ножки. Далее возбуждение прово­
дится по волокнам Пуркинье и достигает типичных кардиомиоцитов
желудочков сердца.
Автоматизм сердца
Автоматизм сердца — это способность сердца самопроизвольно
возбуждаться без действия внешних раздражителей, а под влиянием
процессов, происходящих в нем самом.
Мера автоматизма — частота возникновения возбуждения в месте
возникновения спонтанных возбуждений. Способность сердца к
автоматизму видна в эксперименте на изолированном от тела сердце,
которое продолжает периодически сокращаться.
Рис. 5.1.1. Изменение ритма сердечных сокращений при наложении на
сердце лягушки лигатур Станниуса: А — нормальная кардиограмма сер­
дца лягушки; Б — остановка сердца после наложения первой лигатуры,
которая изолирует синоатриальный узел; В — возобновление сердечных
сокращений после наложения второй лигатуры, которая раздражает атри­
овентрикулярный узел. При этом предсердия и желудочки сокращаются
одновременно; Г — отсутствие сокращений верхушки сердца после нало­
жения третьей лигатуры, которая изолирует верхушку сердца (СП — сис­
тола предсердий; СЖ — систола желудочков; Д — диастола; САУ — сино­
атриальный узел; АВУ — атриовентрикулярный узел)
Висцеральные функции
259
сж
260
Глава 5
Различные отделы проводящей системы сердца обладают различ­
ным автоматизмом. Градиент автоматизма направлен от основания к
верхушке сердца. Это означает, что наибольшим автоматизмом обла­
дают клетки синусного узла, меньшим — клетки предсердно-желудочкового узла, и еще меньшим — другие части проводящей системы
сердца, в частности, сердечные проводящие кардиомиоциты.
Типичные кардиомиоциты, которые обеспечивают сокращение
сердца, спонтанно не возбуждаются. Однако они обладают потен­
циальной способностью к автоматизму. Если выращивать культуру
миокардиальной ткани на питательной среде, то отдаленные друг от
друга клетки миокарда самопроизвольно возбуждаются и сокращают­
ся. При этом каждая клетка сокращается с отличной от других клеток
частотой. Если эти клетки объединить вместе, то они сокращаются
синхронно с одной самой высокой частотой. Это происходит потому,
что возбуждение клетки, обладающей самым высоким автоматизмом,
распространяется на остальные клетки, в которых возбуждение еще
не успело возникнуть.
Аналогичным образом в сердце синусный узел подавляет авто­
матизм остальных отделов проводящей системы сердца и сократи­
тельных кардиомиоцитов. Поэтому синусный узел является води­
телем ритма сердца или пейсмекером. Возбуждение в нем возникает
в первую очередь и затем распространяется на остальные отделы
сердца.
Автоматизм предсердно-желудочкового узла может проявляться
в условиях патологии при различных заболеваниях, когда возбуж­
дение не возникает в синусном узле или, возникая, не доходит до
предсердно-желудочкового узла. В этих случаях начинает прояв­
ляться способность к автоматизму предсердно-желудочкового узла.
Спонтанные возбуждения в предсердно-желудочковом узле возни­
кают в два раза реже. При этом возбуждение распространяется из
предсердно-желудочкового узла в две стороны: на желудочки и на
предсердия. Вследствие одновременного прихода возбуждения к
предсердиям и желудочкам сокращения предсердий и желудочков
происходят не последовательно, как при нормальной работе сердца,
а одновременно. Это делает работу сердца по перекачиванию крови
менее эффективной.
Автоматизм различных отделов проводящей системы сердца
исследован на обнаженном сердце лягуш ки с помощью наложения
лигатур Станниуса. Миограмму сердца лягушки записывают с помо­
Висцеральные функции
261
щью рычажка Энгельмана на кимографе. При этом на миограмме различают сокращения предсердий и сокращения желудочков
(см. рис. 5.1.1 А). Затем на сердце последовательно накладывают три
лигатуры, то есть перевязывают сердце ниткой в трех местах.
Первую лигатуру накладывают между венозным синусом и пред­
сердиями. Это изолирующая лигатура — она отделяет синусно-пред­
сердный узел от всех остальных отделов сердца, поэтому лигатуру
крепко затягивают. Сердце перестает сокращаться (см. рис. 5.1.1Б).
При этом миокард выше места перевязки продолжает ритмически
сокращаться.
Не снимая первой, накладываю т вторую лигатуру между пред­
сердиями и желудочками. Это раздражающая лигатура. Нитку не
затягиваю т до конца, а с помощью слабого надавливания ниткой
на сердце раздражаю т предсердно-желудочковый узел. В клетках
этого узла начинаю т появляться спонтанные ритмические воз­
буждения. Частота сокращ ений сердца становится примерно в
два раза ниже исходной частоты. На кардиограмме регистриру­
ется только одна волна сокращ ения сердца, так как сокращ ения
предсердий происходят одновременно с сокращ ениями желудоч­
ков (см. рис. 5.1.1В). В некоторых случаях автоматизм предсердножелудочкового узла может проявляться через несколько секунд
или минут после первой лигатуры без наложения второй лигатуры.
Это обусловлено спонтанным проявлением автоматизма предсердно-желудочкового узла.
Третью лигатуру накладывают на нижнюю треть желудочков,
не снимая две предыдущие лигатуры. Третья лигатура изолирую­
щая. Она отделяет верхушку сердца от остальных частей сердца.
После наложения этой лигатуры верхушка сердца не сокращает­
ся (см. рис. 5.1.1 Г). В отдельных случаях наблюдается подергивание
отдельных частей миокарда. За счет автоматизма сердечных про­
водящих кардиомиоцитов частота этих сокращений еще ниже, чем
частота сокращений сердца после наложения второй лигатуры.
Эксперимент с лигатурами Станниуса показывает, что градиент
автоматизма убывает от основания к верхушке сердца.
М еханизм автоматизма связан с отсутствием потенциала покоя
на клеточных мембранах синусного узла. На наружной мембра­
не этих клеток происходит постоянное уменьшение мембранного
потенциала, называемое медленной диастолической деполяризацией
(рис. 5.1.2А).
Глава 5
262
V, мВ
Висцеральные функции
263
Известно, что главными причинами потенциала покоя в возбу­
димых клетках являются высокая концентрация ионов К+ внутри
клеток, наличие некоторой проницаемости мембраны для ионов
К+, выход ионов К+ по концентрационному градиенту из клетки и
возникновение калиевого равновесного потенциала на клеточной
мембране.
Медленная диастолическая деполяризация обусловлена несколь­
кими ионными токами через клеточные мембраны пейсмекерных
клеток (см. рис. 5.1.2Б). Во время расслабления и покоя (дистолы)
предсердий сердца происходит постепенное уменьшение проница­
емости мембраны для ионов К+. В результате этого уменьшается
калиевый равновесный потенциал, и мембранный потенциал сни­
жается. В межспайковый период имеется довольно высокий мед­
ленный постоянны й входящий ток ионов Na+ и в меньшей степени
ионов Са2+. Поэтому в клетках синусного узла возникаю т натрие­
вый и кальциевый равновесные потенциалы, противодействующие
калиевому равновесному потенциалу. В связи с этим максим аль­
ная величина мембранного потенциала клеток синусного узла
(-60 мВ) ниже, чем потенциал покоя клеток миокарда желудочков
(-90 мВ). Уменьшение калиевой проницаемости наклады вается на
постоянны й входящий ток Na+ и С а2+. Кроме того, в самом конце
медленной диастолической деполяризации происходит увеличение
входящего тока Na+ и Са2+, что способствует спонтанной деполя­
ризации.
Клетки синусного узла содержат большое количество ионов С1“,
что отличает их от всех остальных клеток сердца. В межспайковый
период проницаемость мембраны для ионов С1~ медленно увеличива­
ется, и С1“ начинает выходить из клеток по градиенту концентрации.
Это способствует деполяризации мембраны.
В межспайковый период постепенно снижается активность Na+,K+АТФ-азы, что уменьшает градиент концентрации этих ионов снару-
Рис. 5.1.2. Механизмы автоматизма клеток синоатриального узла: А — мем­
бранный потенциал клеток синоатриального узла; Б — ионные процессы в
клетках синоатриального узла; В — изменение возбудимости клеток сино­
атриального узла (V — напряжение на внутренней поверхности мембраны;
t - время; МДД — медленная диастолическая деполяризация; ПД — потен­
циал действия; В —возбудимость; АРП — абсолютный рефрактерный пери­
од; СуперВ —супернормальная возбудимость)
264
Глава 5
жи и внутри клеток синусного узла и постепенно снижает потенциал
покоя.
В результате этих процессов мембранный потенциал приближает­
ся к критическому уровню деполяризации -4 0 мВ, что сопровожда­
ется уменьшением порога раздражения и увеличением возбудимости
(см. рис. 5.1.2В). При достижении величины критической деполя­
ризации в клетках синусного узла возникает потенциал действия
(см. рис. 5.1.2А).
Проведение возбуждения по сердцу
Автоматически возникающее возбуждение клеток синусного узла
распространяется на все другие отделы сердца, и в результате этого
миокард начинает сокращаться.
Проведение возбуждения во всех отделах сердца осуществляется
при помощи местных токов между возбужденными и покоящимися
кардиомиоцитами. Передача возбуждения с одной клетки проводя­
щей системы на другие клетки, а также между типичными кардиоми­
оцитами происходит электрическим способом. Между клеточными
мембранами соседних кардиомиоцитов имеются места тесных кон­
тактов, которые называются нексусами. Нексусы представляют собой
мембрану, пронизанную специальными каналами, что способствует
высокой проницаемости этой мембраны для ионов.
Наличие в сердце нексусов определяет ложно-синцитиальное
строение миокарда. Кардиомиоциты анатомически отделены друг от
друга мембранами, но физиологически они объединены с помощью
нексусов. Благодаря этому возбуждение распространяется из синус­
ного узла на все кардиомиоциты. И именно поэтому сердце подчи­
няется закону «Все или ничего».
Возбуждение может распространяться по атипическим и по типи­
ческим волокнам миокарда. Однако проводящая система сердца
обладает большей скоростью проведения возбуждения, чем типич­
ные кардиомиоциты.
Возбуждение в сердце проводится однонаправлено по проводящей
системе сердца из предсердий в желудочки. Обратному проведению
возбуждения препятствует рефрактерность уже возбужденных частей
миокарда, в особенности атипичных проводящих кардиомиоцитов
в желудочках и участках предсердия выше предсердно-желудочкового узла. Возбуждение в этих двух областях длится значительно
дольше, чем в остальных участках сердца. Длительное возбуждение
Висцеральные функции
265
обусловливает продолжительную невозбудимость этих структур, что
полностью исключает возможность распространения возбуждения в
обратном направлении. Достигается последовательность сокращения
отдельных частей сердца. Сначала сокращаются предсердия, а затем
желудочки.
Возбуждение диффузно распространяется из синусного узла
сначала по правому, а затем по левому предсердиям. Затем воз­
буждение достигает предсердно-желудочкового узла. В желудочках
возбуждение достигает, прежде всего, сосочковых мышц и трабе­
кул, прикрепленных к предсердно-желудочковым клапанам, что
приводит к натяжению трабекул и не дает клапанам вывернуться
в область предсердий при сокращении желудочков. Затем возбуж­
дение охватывает перегородку и верхушку сердца, и в последнюю
очередь — боковые стенки и основание желудочков. В толще мио­
карда возбуждение переходит с проводящих кардиомиоцитов на
типичные кардиомиоциты, а затем распространяется к эндокарду
и эпикарду.
В предсердиях возбуждение исчезает в ходе распространения его
по желудочкам. В желудочках оно исчезает от верхушки к основанию
желудочков. В последнюю очередь возбуждение исчезает в сосоч­
ковых мышцах и трабекулах предсердно-желудочковых клапанов
(рис. 5.1.3).
В предсердиях возбуждение распространяется со скоростью 1 м/с
(см. рис. 5.1.3Б), по атипическим проводящим кардиомиоцитам желу­
дочков — со скоростью 3 м/с (см. рис. 5.1.3Г)и по миокарду желудоч­
ков — со скоростью 1 м/с (см. рис. 5.1.3Д).
Особенностью распространения возбуждения в сердце является
наличие задержки при проведении возбуждения по предсердножелудочковому узлу. Это — предсердно-желудочковая или атрио­
вентрикулярная задержка проведения возбуждения. Возбуждение
проводится со скоростью 0,05 м/с (см. рис. 5.1.3В). Низкая скорость
проведения возбуждения обусловлена отсутствием нексусов между
клетками этого участка миокарда. Поэтому клетки предсердно-желудочкового узла обладают пониженной возбудимостью. Возбуждение
все-таки «перескакивает» с одной клетки на другую, но с меньшей
скоростью, чем в других участках сердца. Кроме того, атипичные кар­
диомиоциты расположены в предсердно-желудочковом узле попереч­
но к направлению проведения возбуждения, что замедляет скорость
проведения.
Глава 5
266
А
Б
Рис. 5.1.3. Динамика проведения возбуждения по проводящей системе
сердца: А — возникновение возбуждения в клетках синоатриального узла
(САУ); Б — проведение возбуждения по атипическим волокнам предсер­
дий; В — проведение возбуждения в атриовентрикулярном узле (АВУ);
Г — проведение возбуждения по ножкам пучка Гиса (ПГ)и по волокнам
Пуркинье (ВП); Д — проведение возбуждения по миокарду желудочков
*
Висцеральные функции
267
Задержка проведения возбуждения обеспечивает начало сокра­
щения желудочков после окончания сокращения предсердий. Это
позволяет крови пройти из предсердий в желудочки при систоле
предсердий до начала сокращения желудочков.
Распространение возбуждения в сердце характеризуется измене­
нием знака заряда мембран кардиомиоцитов, то есть возникнове­
нием потенциалов действия. На графиках мембранных потенциалов
типичных кардиомиоцитов желудочков различают 5 фаз: нулевая
фаза — деполяризация, 1-я фаза — начальная быстрая реполяри­
зация, 2-я фаза — медленная реполяризация (плато), 3-я фаза —
конечная быстрая реполяризация и 4-я фаза — потенциал покоя
(рис. 5.1.4А).
Потенциалы действия клеток синусного узла не имеют плато.
Кроме того, отсутствует потенциал покоя и происходит медленная
диастолическая деполяризация.
Деполяризация связана с быстрым вхождением в клетки ионов
Na+ (см. рис. 5.1.4Б). Реполяризация обусловлена инактивацией вхо­
дящего в клетку тока Na+ и активацией выходящего из клетки
тока К+. Однако реполяризация задерживается в связи с входом в
клетку ионов Са2+, которые участвуют в дальнейшем в процессе
сокращения кардиомиоцитов. Ток ионов Са2+ замедляет реполя­
ризацию и удлиняет процесс возбуждения сердца. Длительность
возбуждения становится равной длительности сокращения пред­
сердий и желудочков сердца. Длительность потенциалов действия
типичных кардиомиоцитов сопровождается длительным периодом
невозбудимости миокарда, предохраняющим сердце от суммации
одиночных сердечных сокращений (см. рис. 5.1.4В). Следовательно,
ионные механизмы возбуждения определяют одно из важных
свойств миокарда — неспособность к слитному тетаническому сок­
ращению.
Длительность потенциалов действия кардиомиоцитов синусного
узла — 230 мс, кардиомиоцитов предсердий — 180 мс, кардиомиоци­
тов желудочков — 330 мс, проводящих кардиомиоцитов и небольшого
участка миокарда предсердий, расположенного чуть выше предсердно-желудочкового узла, — 450 мс. Абсолютные рефрактерные перио­
ды этих двух последних участков сердца — 400 мс, как указывалось
выше, исключают возможность распространения возбуждения в сер­
дце в обратном направлении. Длительный абсолютный рефрактер­
ный период в желудочках — 270 мс — препятствует тетаническому
сокращению сердца.
268
Глава 5
А
Рис. 5.1.4. Возбуждение и возбудимость кардиомиоцитов желудочков:
А — мембранный потенциал кардиомиоцитов желудочков; Б — ионные
процессы в кардиомиоцитах желудочков; В — изменение возбудимости
кардиомиоцитов желудочков (Д — деполяризация; БН Р — быстрая началь­
ная реполяризация; МР — медленная реполяризация; БКР — быстрая
конечная реполяризация; ПП — потенциал покоя; В — возбудимость;
АРП — абсолютный рефрактерный период)
I
Висцеральные функции
269
Электрокардиограмма как показатель возбуждения сердца
Электрокардиограмма (ЭКГ) — это последовательная запись разно­
сти потенциалов между различными участками тела, возникновение
которой обусловлено электромагнитным полем вокруг сердца при
распространении и исчезновении в нем возбуждения.
Появление возбуждения характеризуется деполяризацией мем­
бран кардиомиоцитов. Наружная поверхность их становится заря­
женной отрицательно. Соседние, невозбужденные, мембраны оста­
ются заряженными с наружной поверхности положительно. Между
отрицательными и положительными участками миокарда возникают
местные токи, сила которых сначала увеличивается, а затем умень­
шается. Этот процесс аналогичен процессам проведения возбужде­
ния по мембране любой возбудимой клетки. Изменяющаяся сила
местного тока приводит к возникновению увеличивающегося, а
затем уменьшающегося магнитного поля вокруг направления дви­
жения тока. Согласно законам физики, переменное магнитное поле
вызывает в сердце и в окружающих тканях вихревое электрическое
поле. Вихревое электрическое поле — переменное поле с замкнуты­
ми силовыми линиями; оно тем больше, чем быстрее изменяется во
времени магнитное поле. Переменное магнитное поле и вихревое
электрическое поле всегда связаны между собой, и вокруг сердца
возникает электромагнитное поле при проведении возбуждения.
С помощью электрокардиографии измеряется только электрическая
составляющая этого поля. При возникновении вихревого электри­
ческого поля изменяются заряды в различных участках тела, и между
различными участками тела возникает разность потенциалов, или
электродвижущая сила, которая и регистрируется с помощью элект­
рокардиографии в виде зубцов ЭКГ.
Появление зарядов в различных участках тела связано не только
с проведением возбуждения, но также с последовательным исчезно­
вением этого возбуждения в различных частях сердца. При исчез­
новении возбуждения мембраны кардиомиоцитов реполяризуются
до уровня потенциала покоя. Наружная поверхность этих мембран
становится заряженной опять положительно, в то время как возбуж­
денные мембраны еще имеют снаружи отрицательный заряд. Разница
зарядов вновь вызывает местные токи, вокруг которых возникает
электромагнитное поле, обусловливающее возникновение разности
потенциалов в различных частях тела.
270
Рис. 5.1.5. Изменение амплитуды и
направления электрического век­
тора (ЭВ) сердца: Р, QRS, Т — три
петли векторкардиограммы
Глава 5
Напряженность электричес­
кой составляющей электромаг­
нитного поля сердца характери­
зуется электрическим вектором
сердца, который ориентирован
в пространстве от минуса к
плюсу. В связи со сложностью
проведения возбуждения по
различным структурам сердца
электрический вектор сердца
постоянно меняется по ам п­
литуде и величине (рис. 5.1.5).
Временные изменения проек­
ции мгновенных значений век­
тора на прямые, соединяющие
место расположения электро­
дов при электрокардиографии,
образуют последовательность
зубцов ЭКГ.
При неизменности заряда на
мембранах, независимо от его
величины, на ЭКГ регистриру­
ется изоэлектрическая линия
между зубцами. Это происходит
во время потенциала покоя и в
ходе плато потенциала действия
мембран кардиомиоцитов. При
этом электрическое поле вокруг
сердца отсутствует. Зубцы на
ЭКГ возникают при появлении
и исчезновении возбуждения, а
если возбуждение уже охвати­
ло структуры сердца, то на ЭКГ
регистрируется изоэлектричес­
кая линия между зубцами.
Разность потенциалов в раз­
личных участках тела возникает
только при одновременном воз­
буждении значительной части
мышечных волокон в сердце.
Висцеральные функции
271
Поэтому процессы возбуждения и проведения возбуждения по про­
водящей системе сердца прямо не отражаются на ЭКГ в связи с
недостаточным количеством атипичных кардиомиоцитов проводя­
щей системы для возникновения электромагнитного поля вокруг
сердца.
Отражая процессы возникновения, исчезновения и проведения
возбуждения по сердцу, ЭКГ не связана прямо с процессом сокраще­
ния сердца, поскольку в ряде случаев возбуждение может происхо­
дить без сокращения. Так, например, при длительном пропускании
через изолированное сердце физиологического раствора без ионов
Са2+ сокращение сердца прекращается. Однако при этом регистриру­
ется ЭКГ и потенциалы действия. В умирающем сердце сокращения
могут прекратиться раньше, чем процессы возбуждения, и зубцы ЭКГ
могут регистрироваться после остановки сердца.
Возникновение зубцов и сегментов ЭКГ зависит от места проведения
возбуждения в сердце
На ЭКГ различают зубцы и сегменты между зубцами (изоэлектрическую линию). Возникновение зубцов и сегментов ЭКГ зависит
от наличия или отсутствия возбуждения в различных отделах сердца
(рис. 5.1.6).
Зубец Р возникает при проведении возбуждения из синоатриально­
го узла сначала на правое и затем на левое предсердие (рис. 5.1.6А).
Когда оба предсердия охвачены возбуждением местные токи
отсутствуют, электромагнитное поле вокруг сердца не возникает, и
зубцы на ЭКГ отсутствуют. Возбуждение распространяется с малой
скоростью только в небольшой группе клеток предсердно-желу­
дочкового узла, что не вызывает зубцов на ЭКГ. При этом на ЭКГ
возникает сегмент PQ (рис. 5.1.6Б). Малая скорость проведения воз­
буждения обусловливает атриовентрикулярную задержку. В этот
момент существует разность потенциалов между предсердиями и
желудочками, в связи с чем, возникает электростатическое поле.
Однако зубцы ЭКГ возникают при появлении электромагнитного
поля, которое отсутствует, так как возбуждение с предсердий на
желудочки не проводится.
Зубец Q связан с проведением возбуждения по предсердно-желу­
дочковому пучку и обусловлен возбуждением части межжелудочковой перегородки, сосочковых мышц и трабекул атриовентрику­
лярных клапанов. Зубец Q отрицателен потому, что направление
местных токов по сосочковым мышцам и трабекулам противопо-
/"V
Висцеральные функции
273
ложно основному направлению распространения возбуждения в
сердце от основания к верхушке (рис. 5.1.6В). Небольшая ам пли­
туда зубца Q связана с небольшим количеством возбуждающихся
кардиомиоцитов. Возбуждение и следующее за ним сокращение
сосочковых мышц и трабекул в начале систолы желудочков пре­
дохраняет атриовентрикулярные клапаны от открытия их в сторону
предсердий.
Восходящая часть зубца R обусловлена возбуждением всей пере­
городки и верхушки сердца. Возбуждение распространяется по
перегородке к верхушке сердца и от эндокарда к эпикарду в облас­
ти верхушки сердца. Зубец Л имеет самую большую амплитуду по
сравнению с другими зубцами ЭКГ. Большая амплитуда зубца R
обусловлена большим количеством возбуждающихся кардиомио­
цитов в желудочках сердца (рис. 5.1.6Г). Нисходящая часть зубца R
обусловлена проведением возбуждения через верхушку на боковые
стенки желудочков.
Зубец S связан с распространением возбуждения по боковым
поверхностям желудочков от верхушки сердца к основанию, в связи
с чем, он отрицательный. Вершина зубца S соответствует охва­
ту возбуждением практически всех кардиомиоцитов желудочков.
При этом асинхронное сокращ ение сменяется изометрическим.
Исчезновение распространяющейся волны возбуждения в желудоч­
ках сопровождается исчезновением электрического вектора сердца,
что вызывает возвращение зубца S к изоэлектрической линии на
ЭКГ (рис. 5.1.6Д).
Когда все желудочки охвачены возбуждением на ЭКГ возникает
сегмент ST. Местные токи в сердце не возникают, и электрическое
поле вокруг сердца отсутствует (рис. 5.1.6Е).
Зубец Т возникает при исчезновении возбуждения в желудочках.
Сначала возбуждение исчезает в верхушке сердца, затем в его пере-
Рис. 5.1.6. Зависимость между местом проведения возбуждения в сердце и
возникновением зубцов на электрокардиограмме (ЭКГ): А — проведение
возбуждения в предсердиях; Б — атриовентрикулярная задержка проведе­
ния возбуждения; В — проведение возбуждения по сосочковым мышцам;
Г — проведение возбуждения к верхушке сердца; Д — проведение возбуж­
дения по боковым стенкам желудочков; Е — продолжение возбуждения
желудочков; Ж — исчезновение возбуждения в желудочках; 3 — отсутствие
возбуждения в сердце (Р, Q, R, S, Т — зубцы ЭКГ)
274
Глава 5
городке, потом в боковых стенках желудочков. В последнюю очередь
возбуждение исчезает в сосочковых мышцах и трабекулах. Это спо­
собствует длительному сокращению сосочковых мышц и трабекул, что
предохраняет створки атриовентрикулярных клапанов от открытия в
сторону предсердий в конце систолы желудочков (рис. 5.1.6Ж).
Сегмент ТР возникает при отсутствия возбуждения в сердце
(рис.5.1.63).
Зубцы, сегменты и интервалы ЭКГ коррелируют с фазами мембранных
потенциалов кардиомиоцитов
Кроме зубцов и сегментов на ЭКГ выделяют интервалы. Интервалы
ЭКГ образованы комплексом зубцов и сегментов. Зубцы, сегменты и
интервалы ЭКГ связаны с фазами потенциалов действия типических
кардиомиоцитов предсердий и желудочков (рис. 5.1.7).
Зубец Р длится 40 мс при длительности сердечного цикла 800 мс
и частоте сердечных сокращений 75 уд/мин (рис. 5.1.7А, Б). Он
соответствует деполяризации и быстрой начальной реполяризации
клеточных мембран в предсердиях.
Сегмент PQ длится 60 мс. Он соответствует медленной реполяриза­
ции (плато) в предсердиях (рис. 5.1.7А, Б). Предсердия возбуждены, и
возникает атриовентрикулярная задержка проведения возбуждения
от предсердий к желудочкам. Интервал PQ длится 100 мс и является
суммой зубца Р и сегмента PQ. Длительность интервала PQ соответс­
твует времени систолы предсердий.
Комплекс QRS длится 80 мс. Он соответствует деполяризации и
начальной быстрой реполяризации желудочков, а также конечной
деполяризации предсердий (рис. 5.1.7А, В).
Сегмент S T длится 120 мс. Он связан с фазой плато потенциалов
действия в кардиомиоцитах желудочков (рис. 5.1.7А, В). Желудочки
возбуждены, и возбуждение не исчезает в них.
Зубец Т длится 130 мс. Он соответствует фазе быстрой конечной
реполяризации потенциалов действия в кардиомиоцитах желудочков
(рис. 5.1.7А, В). При этом возбуждение в желудочках исчезает.
Интервал QT длится 330 мс и соответствует времени систолы желу­
дочков.
Сегмент ТР длится 370 мс. Он связан с потенциалом покоя мембран
кадриомиоцитов сердца и соответствует общей диастоле предсердий
и желудочков.
Интервал ТР длится 470 мс и включает в себя сегмент ТР и зубец Р.
Он отражает длительность диастолы желудочков.
I
'
Висцеральные функции
275
Рис. 5.1.7. Корреляция между зубцами ЭКГ и фазами потенциалов дейс­
твия миокардиальных волокон: А — ЭКГ; Б — потенциал действия кардиомиоцита предсердий; В —потенциал действия кардиомиоцита желудочков
(Р, Q, R, S, Т — зубцы ЭКГ. Д — деполяризация; БНР — быстрая начальная
реполяризация; МР — медленная реполяризация; БКР — быстрая конеч­
ная реполяризация; П П — потенциал покоя)
Электрокардиографические отведения
Общепринятая практика электрокардиографии включает в себя
12 отведений: 3 стандартных двухполюсных отведения от конечнос­
тей, 3 однополюсных усиленных отведения от конечностей и 6 одно­
полюсных грудных отведений.
Глава 5
276
Обследуемого человека заземляют, чтобы избавиться от наводок
при регистрации ЭКГ. Электрод, связанный с землей, наклады­
вают на правую голень. Отводящие электроды при отведениях от
конечностей закрепляют на правом и левом предплечьях и левой
голени. I стандартное отведение ЭКГ осуществляется между правой
и левой руками, II отведение — между правой рукой и левой ногой,
и III отведение — между левой рукой и левой ногой.
Прямые, соединяющие места наложения электродов на конечнос­
тях, образуют треугольник Эйнтховена. Электрический вектор сердца
проецируется на стороны этого треугольника. Запись величины
этой проекции во времени образует последовательность зубцов ЭКГ
(рис. 5.1.8А).
А
пр
Б
1
ЛР
ПР
ЛР
Рис. 5.1.8. Проекции электрического вектора (ЭВ) сердца: А - проекции
на стороны треугольника Эйнтховена при регистрации ЭКГ в I, II и III
стандартных отведениях; Б — проекции на перпендикуляры из вершин
к сторонам треугольника Эйнтховена при регистрации ЭКГ в усиленных
клинических отведениях aVR, aVL, aVF (ПР — правая рука; JIP — левая
рука; ПН —правая нога, J1H —левая нога)
Висцеральные функции
277
По ЭКГ в стандартных отведениях определяют электрическую
ось сердца. Электрическая ось сердца — главное направление рас­
пространения возбуждения в сердце во фронтальной плоскости тела
человека. Электрическая ось сердца совпадает с анатомической осью
сердца и может быть вертикальной, наклонной, горизонтальной, что
зависит от положения сердца в грудной полости.
Направление электрической оси сердца определяют по величи­
не зубца R двух любых стандартных отведений. Величину зубца R
откладывают в виде вектора в прямоугольной системе координат
из начала координат в направлении, соответствующем отведению
ЭКГ. Затем в местах окончания этих двух векторов проводят линии,
перпендикулярные направлению векторов. Точка пересечения этих
линий соответствует концу суммарного электрического вектора сер­
дца при возникновении зубца R. Направление этого вектора указы­
вает главное направление распространения возбуждения в сердце, а
значит, показывает направление электрической оси сердца.
Нельзя смешивать понятие «электрический вектор сердца» с
понятием «электрическая ось сердца». Электрический вектор серд­
ца, изменение величины и направления его мгновенного значения
являются причиной ЭКГ. Электрическая ось сердца постоянна по
величине и направлению и определяется по ЭКГ. В течение одно­
го сердечного цикла эти две величины совпадают по величине и
направлению только при возникновении зубца R.
При усиленных отведениях один электрод (активный) находится
на одной из трех конечностей, а другой электрод (индифферентный)
подсоединяют через сопротивления к двум другим конечностям. При
отведении aVR активный электрод находится на правом предплечье,
при отведении аУГ — на левом предплечье, при отведении aVF —
на левой голени (рис. 5.1.8Б).
При грудных отведениях активный электрод прикрепляют пос­
ледовательно в шести местах на грудной клетке. Второй электрод
через сопротивления подсоединяют к трем конечностям челове­
ка. Регистрация ЭКГ в 12 отведениях в клинике дает возможность
выявить характер и локализацию нарушений проведения возбужде­
ния в сердце при обследовании пациентов.
Ритмическое сокращение сердца
Распространяющееся по сердцу возбуждение вызывает сокраще­
ние типичных кардиомиоцитов. Механизм сопряжения возбуждения
с сокращением в сердце несколько отличается от перехода возбуж­
278
Глава 5
дения в сокращение в скелетной мышце. И в скелетной мышце, и в
миокарде сокращение происходит вследствие взаимодействия между
актином и миозином под влиянием ионов Са2+. Однако, если в ске­
летной мышце ионы Са2+ поступают к актину и миозину преиму­
щественно из эндоплазматической сети, то в миокарде Са2+ выходит
в саркоплазму частично из эндоплазматической сети и частично
снаружи кардиомиоцитов из Т-систем. Вхождение в кардиомиоциты
ионов Са2+ называют кальциевым залпом. Он происходит во время
плато потенциала действия. Это отличие обусловливает ряд важных
особенностей сокращения миокарда.
Фазы сердечного цикла
Периодическая работа сердца отличается высокой ритмичностью.
Параметры результата каждого сердечного цикла (ударный объем
крови, давление крови, длительность цикла) почти в точности повто­
ряют параметры результата предыдущего кардиоцикла. При частоте
сердечных сокращений, соответствующей норме и равной 75 ударам в
минуту, длительность одного сердечного цикла равна 60 с/75 = 0,8 с
=800 мс, что принимают за исходную величину при анализе длитель­
ности фаз сердечного цикла. Следует помнить, что при увеличении
частоты сердечных сокращений длительность сердечного цикла и
длительность каждой фазы сердечного цикла уменьшаются.
Работа правой и левой половины сердца происходит содружес­
твенно, систола и диастола их происходят почти одновременно.
Однако длительность этих и более дробных фаз сердечного цикла
у правой и левой половины сердца имеет незначительную разницу,
что связано с некоторой неодновременностью распространения в них
возбуждения.
Работа предсердий складывается из двух фаз: систолы предсердий
(длительностью 100 мс для левого предсердия и 110 мс для правого
предсердия) и диастолы предсердий (700 мс и 690 мс соответствен­
но). Систола правого предсердия начинается на Юме раньше, чем
систола левого предсердия. В работе желудочков различают 9 фаз,
которые заканчиваются промежуточными для сердечного цикла
результатами.
Фаза пресистолы желудочков соответствует систоле предсердий,
с которой начинается сердечный цикл. Каждая фаза сердечного
цикла приводит к определенным изменениям давления и объема
крови в камерах сердца и изменению положения клапанов сердца
(рис. 5.1.9).
279
Висцеральные функции
О
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8 с
Рис. 5.1.9. Динамика давления в камерах сердца и аорте, положение кла­
панов сердца и фонокардиограмма в ходе сердечного цикла: I. Давление
крови в аорте (а — анакрота, к — катакрота, и — инцизура. д — дикрота);
II. Давление крови в левом предсердии; III. Давление крови в левом желу­
дочке; IV. Положение митрального клапана; V. Положение аортального
клапана (3 — клапан закрыт; О — клапан открыт); VI. Фонокардиограмма
(1, 2, 3, 4 — тоны); VII. Фазы работы желудочков: 1 — пресистола 100 мс,
2 —асинхронное сокращение 50 мс, 3 —изометрическое сокращение 30 мс,
4 — быстрое изгнание крови 120 мс, 5 — медленное изгнание крови 130 мс,
6 —протодиастола 40 мс, 7 —изометрическое расслабление 80 мс, 8 —быс­
трое наполнение кровью 40 мс, 9 — медленное наполнение кровью 170 мс
280
Глава 5
2,3
4,5
6,7
I I ТОН
8,9
Висцеральные функции
281
Систола предсердий
Фаза систолы предсердий начинается сокращением кардиомио­
цитов предсердий, продолжается 110 мс в правом и 100 мс в левом
предсердиях и заканчивается переходом сокращения в расслабление
предсердий. Результатом систолы предсердий является выброс крови
из предсердий в желудочки (рис. 5.1.10А).
Систола обоих предсердий начинается с сокращения участков
предсердий в местах впадения в них полых вен, и эти места пере­
жимаются. Это препятствует движению крови из предсердий в вены
во время систолы предсердий. Затем сокращаются ушки сердца,
что способствует более эффективному сокращению предсердий.
Уже через 50 мс после начала систолы предсердий кровь полностью
вытесняется из сокращающихся ушек сердца. Это способствует
дополнительному растяжению стенок предсердий, что, согласно
закону Старлинга, увеличивает силу их сокращения. После этого
сокращаются кардиомиоциты боковых стенок предсердий, и волна
уменьшения объема предсердий движется в сторону желудочков.
Вследствие сокращения предсердий давление крови повышается,
и кровь переходит в желудочки, в результате чего давление крови
в желудочках также повышается до 5 мм рт.ст. в правом и 10 мм рт.
ст. в левом желудочках. Объем желудочков при этом увеличивается
до максимальной величины — конечного диастолического объема,
равного 125 мл.
Если бы в сердце не было предсердий, то низкое венозное давле­
ние крови было бы неспособно растянуть толстостенные желудочки
до нужного объема, в связи с чем, эффективность работы сердца
снизилась, и произошел бы застой крови в венах. Поэтому сердце
имеет многокамерное строение, и повышение давления происходит в
нем ступенчато. Предсердия — тонкостенный отдел сердца, что бла­
гоприятствует заполнению их кровью. Затем, сокращаясь, они нака­
чивают дополнительное количество крови в желудочки, что обеспе-
Рис. 5.1.10. Движение крови, положение клапанов и возникновение тонов
сердца во время сердечного цикла: А: 1 — систола предсердий; Б: 2 — асин­
хронное сокращение желудочков. 3 — изометрическое сокращение желу­
дочков; В: 4 — быстрое изгнание крови из желудочков, 5 — медленное
изгнание крови из желудочков; Г: 6 — протодиастола, 7 — изометрическое
расслабление желудочков; Д: 8 — быстрое наполнение кровью желудочков,
9 — медленное наполнение кровью желудочков
282
Глава 5
чивает последним необходимое растяжение. Ушки сердца выполняют
по отношению к предсердиям такую же насосную функцию, какую
предсердия имеют по отношению к желудочкам.
Напряжение желудочков
Систола правого и левого желудочков начинается одновременно
с их напряжения. Результатом напряжения желудочков является
повышение давления крови в них, под влиянием чего открываются
полулунные клапаны, и кровь начинает выбрасываться из право­
го желудочка в легочный ствол, а из левого желудочка — в аорту.
Напряжение желудочков складывается из двух фаз: асинхронного и
изометрического сокращений желудочков (см. рис. 5.1.10Б).
Асинхронное сокращение желудочков. Фаза асинхронного сокра­
щения желудочков начинается с сокращения некоторых волокон
миокарда желудочков, продолжается 70 мс в правом и 50 мс в левом
желудочке и заканчивается сокращением всех кардиомиоцитов
желудочков.
Асинхронное (неодновременное) сокращение кардиомиоцитов
возникает в связи с неодновременностью прихода возбуждения в
различные участки миокарда желудочков. Прежде всего сокращают­
ся участки миокарда, ближайшие к окончаниям проводящих кар­
диомиоцитов предсердно-желудочкового пучка Гиса. Сокращение
начинается с сосочковых мышц, перегородки и верхушки сердца.
Под влиянием сокращения соседние участки желудочков, в которых
возбуждение еще не возникло, растягиваются. Поэтому общая длина
и объем желудочков не изменяются.
В связи с расслаблением предсердий давление в них начинает
уменьшаться, и кровь из желудочков пытается перейти обратно в
предсердия. При этом створки атриовентрикулярных клапанов пас­
сивно всплывают над кровью в желудочках, так как они легче крови.
Однако окончательное их захлопывание происходит только в начале
следующей фазы.
Изометрическое сокращение желудочков. При проведении возбуж­
дения на все кардиомиоциты желудочков асинхронное сокращение
переходит в синхронное. Фаза изометрического сокращения желу­
дочков продолжается 30 мс. Она начинается с закрытия атриовент­
рикулярных клапанов. Фаза заканчивается открытием полулунных
клапанов и выбросом крови из правого желудочка в легочный ствол,
а из левого желудочка — в аорту.
Висцеральные функции
283
В правой половине сердца атриовентрикулярные клапаны состоят
из трех створок (трехстворчатый клапан), а в левой — из двух створок
(двухстворчатый, или митральный клапан). Клапаны открываются
при движении створок в полость желудочков и закрываются при
движении створок из полости желудочков в сторону атриовентри­
кулярного отверстия. Со стороны полости желудочков к клапа­
нам прикреплены сухожильные нити (трабекулы), которые другим
концом прикреплены к сосочковым мышцам стенок желудочков.
Сокращение сосочковых мышц и напряжение сухожильных нитей
препятствуют движению клапанов в сторону предсердий при увели­
чении давления крови в желудочках в ходе систолы желудочков.
Увеличению долговечности работы клапанов способствует отно­
сительно медленная скорость их закрытия и трабекулы, амортизиру­
ющие удар крови о них. Натягиваясь, трабекулы уменьшают скорость
закрытия этих клапанов.
Несмотря на синхронность сокращения всех кардиомиоцитов
желудочков, общая длина миокарда не уменьшается, потому что
кровь, находящаяся в желудочках, несжимаема, и объем ее не меня­
ется, так как все клапаны сердца закрыты.
Давление крови увеличивается в правом желудочке до 15 мм рт.ст.
и в левом до — до 80 мм рт.ст., вследствие чего полулунные клапаны
открываются, и кровь начинает выбрасываться из желудочков.
Изгнание крови из желудочков
Изгнание крови из желудочков начинается после открытия полу­
лунных клапанов. Клапаны состоят из трех кармашков на стенке
сосудов. Строение клапанов напоминает накладные карманы на
одежде человека. При выходе крови из желудочков кармашки легоч­
ного и аортального клапанов прилегают к стенкам легочного ствола
и аорте. Таким образом, клапаны не мешают выходу крови из желу­
дочков (рис. 5.1.11).
Изгнание крови из желудочков делят на две фазы: быстрое и
медленное изгнание. Большее количество крови выбрасывается из
сердца в фазе быстрого изгнания и меньшее — в фазе медленного
изгнания. В фазе быстрого изгнания крови из желудочков приток
крови из правого желудочка в легочный ствол и из левого желудочка
в аорту превышает отток крови из легочного ствола и аорты в мелкие
артерии и капилляры. В связи с этим давление крови в легочном
стволе и аорте растет. Затем в фазе медленного изгнания крови из
284
Глава 5
Ток крови
Рис. 5.1.11. Положение частей аортального клапана на поперечном сечении
аорты: 3 —клапан закрыт; О —клапан открыт
желудочков отток крови превышает приток крови в легочный ствол
и аорту. Поэтому давление крови в легочном стволе и аорте начинает
уменьшаться. Однако в течение этих обеих фаз в каждый отдельный
момент давление крови в желудочках выше, чем давление крови в
легочном стволе и аорте, что обеспечивает выброс крови из сердца в
сосуды (см. рис. 5.1.10В).
Быстрое изгнание крови из желудочков. Фаза быстрого изгнания
крови из желудочков начинается в момент открытия полулунных
клапанов, продолжается ПОмс для правого и 120 мс — для левого
желудочка и заканчивается при максимальном подъеме давления в
легочном стволе и аорте. Давление крови в легочном стволе увеличи­
вается с 15 до 30 мм рт.ст., а в аорте — с 80 до 120 мм рт.ст.
Медленное изгнание крови из желудочков. Фаза медленного изгнания
крови из желудочков начинается в момент максимального давления
в желудочках, продолжается в правом и левом желудочках 130 мс
и заканчивается переходом миокарда желудочков от сокращения к
расслаблению.
Объем желудочков уменьшается до минимальной величины —
конечного систолического объема, равного 60 мл. Следовательно, из
Висцеральные функции
285
правого желудочка в легочный ствол и из левого желудочка в аорту
выбрасывается 125 - 60 = 65 мл крови. Объем крови, выбрасываемой
из каждого желудочка называют ударным объемом крови. Он равен
разнице между конечным диастолическим объемом и конечным сис­
толическим объемом каждого желудочка.
Известно, что основание желудочков фиксировано по отношению
к окружающим его тканям, а остальная часть желудочков не закреп­
лена. Поэтому при наполнении желудочков кровью длина сердца
увеличивается. При сокращении сердца длина желудочков умень­
шается, что напоминает движение поршня при выбросе жидкости
из шприца. Это позволяет сравнить работу желудочков с работой
поршневого насоса.
При поступлении крови в желудочки, а также при выбросе крови
из желудочков кровь течет не прямолинейно, а совершает в желудочках
вращательные движения. Это происходит благодаря соответствующе­
му строению внутренней поверхности желудочков и ротации (поворо­
ту вокруг своей оси) желудочков при их сокращении. Поэтому сердце
работает не только как поршневой, но и как центробежный насос, рас­
кручивая кровь в полости желудочков перед ее выбросом из сердца.
При изгнании крови из желудочков в них происходят волнооб­
разные мышечные сокращения от верхушки к основанию сердца.
Поэтому считается, что сердце обладает одновременно свойством
перистальтического насоса.
Расслабление желудочков
Диастола желудочков предназначена для отдыха миокарда, вос­
становления его энергетических запасов и подготовки сердца к оче­
редной систоле. Диастола складывается из расслабления желудочков
и наполнения их кровью. В ходе расслабления желудочков напряже­
ние миокарда желудочков падает до минимума, что сопровождается
резким падением давления в их полостях. Расслабление желудочков
состоит из двух фаз: протодиастолы и изометрического расслабления
(см. рис. 5.1.ЮГ).
Протодиастола — первая фаза диастолы. Фаза протодиастолы
начинается с момента расслабления желудочков, длится 50 мс в пра­
вом и 40 мс в левом желудочке и заканчивается закрытием полулун­
ных клапанов.
Давление крови в легочном стволе и аорте продолжает понижать­
ся вследствие оттока крови к органам и тканям организма. Однако
286
Глава 5
уменьшение давления в желудочках идет быстрее, так как они рас­
слабляются. Поэтому давление в желудочках становится меньше, чем
в легочном стволе и аорте, и часть крови из этих сосудов устремля­
ется в сторону желудочков. Устремившаяся в обратном направлении
кровь расправляет кармашки полулунных клапанов и наполняет их
кровью. Кармашки смыкаются, происходит закрытие клапанов, и
кровь в желудочки не попадает.
Долговечности полулунных клапанов способствует находящийся
в стенке аорты специальный комплекс эластичных и коллагеновых
волокон, прикрепленных к основанию клапана. При ударе крови о
клапаны они смещаются в сторону желудочков, а затем комплексом
волокон подтягиваются в прежнее положение. Знания об аморти­
зирующей роли волокон у основания полулунных клапанов учиты­
вается при разработке искусственного аортального клапана сердца.
Чем лучше амортизирующая функция искусственного клапана, тем
больше его долговечность.
Фаза изометрического расслабления начинается закрытием полулун­
ных клапанов, продолжается 60 мс в правом и 80 мс в левом желудоч­
ках и заканчивается открытием атриовентрикулярных клапанов.
Изометрическое расслабление — расслабление мышцы с уменьше­
нием ее напряжения, но без удлинения ее. Во время фазы изометри­
ческого расслабления длина кардиомиоцитов желудочков не меняет­
ся, так как желудочки не могут увеличиваться в объеме. Оставшаяся в
желудочках кровь, как и любая жидкость, не растягивается, и кровь в
желудочки не поступает, так как клапаны сердца закрыты. Давление
в правом желудочке падает до 5 мм рт.ст. и в левом — до 10 мм рт.ст.
Наполнение желудочков кровью
Кровь поступает из вен в сердце не постоянно, а периодически. Во
время систолы кровь в сердце не входит. В предсердия кровь влива­
ется в период диастолы предсердий. В желудочки кровь поступает в
фазе быстрого и медленного наполнения. При этом кровь проходит в
желудочки из вен через предсердия (см. рис. 5.1.10Д). Окончательное
наполнение желудочков кровью происходит во время систолы пред­
сердий.
Фаза быстрого наполнения желудочков кровью начинается откры­
тием атриовентрикулярных клапанов, продолжается 80 мс и закан­
чивается сменой снижения давления в предсердиях и желудочках
на его повышение. Эта фаза начинается синхронно в правой и
Висцеральные функции
287
левой половинах сердца. Желудочки наполняются кровью быстро
в результате повышенного давления крови в предсердиях, полых и
легочных венах, что связано с застоем крови в них во время систолы
желудочков. Быстрому наполнению способствует возможность резко­
го увеличения объемов расслабляющихся желудочков. Кроме того,
расправлению желудочков (в особенности левого) помогает наполне­
ние коронарных сосудов кровью в начале расслабления сердца, что
создает «гидравлический каркас сердца».
Увеличение объемов желудочков приводит к уменьшению дав­
ления в правом желудочке до 0 мм рт.ст. и в левом желудочке —
до 5 мм рт.ст.
Фаза медленного наполнения желудочков кровью начинается подъемом
давления в предсердиях и желудочках, продолжается 160 мс в правом
и 170 мс — в левом желудочках и заканчивается сокращением пред­
сердий.
Наполнение желудочков происходит медленно из-за меньшей
разности давлений между предсердиями и желудочками, чем в фазе
быстрого наполнения, и незначительного увеличения объемов уже
частично наполненных кровью желудочков вследствие упругости
стенок желудочков. Давление крови в них при этом увеличивается:
в правом — до 3 мм рт.ст. и в левом — до 7 мм рт.ст.
Зависимость давления от объема крови в сердце
Физическая величина работы сердца равна произведению измене­
ния давления на изменение объема крови в сердце в ходе сердечного
цикла. Ее можно вычислить как площадь многоугольника на графи­
ке давление—объем крови левого желудочка сердца (рис. 5.1.12А). При
увеличении венозного притока крови работа сердца и ударный объем
крови увеличиваются согласно закону Старлинга без повышения
артериального давления (рис. 5.1.12Б). При увеличении сосудистого
сопротивления выбросу крови из сердца артериальное давление и
работа сердца увеличиваются без изменения ударного объема крови
(рис. 5.1.12В).
Отличия в работе правой и левой половины сердца
М инимальное диастолическое давление в правой половине сер­
дца равно нулю и может становиться ниже атмосферного в связи с
дыханием. В левой половине сердца давление не уменьшается менее
5 мм рт.ст.
288
Глава 5
Висцеральные функции
289
Несмотря на то что обе половины сердца перекачивают равное
количество крови, правая половина сердца совершает значительно
меньше работы, чем левая. Это связано с тем, что правый желудочек
выбрасывает кровь в легочный ствол под давлением 25—30 мм рт.
ст., а левый желудочек выбрасывает кровь в аорту под давлением
110—120 мм рт.ст. В связи с этим мышечная стенка правого желудочка
тоньше, чем левого.
Отличия заключаются также в незначительной несинхронности
работы правой и левой половины сердца. В связи с тем, что воз­
буждение раньше распространяется на правое предсердие, систола
правого предсердия начинается на 10 мс раньше, чем левого пред­
сердия. Систола правого и левого желудочков начинается одно­
временно. Однако в связи с невысоким конечным систолическим
давлением в правой части сердца длительность фаз напряжения и
расслабления в правой половине сердца несколько меньше, чем в
левой. Длительность систолы правого желудочка несколько больше,
а диастолы — несколько меньше, чем левого. Правая половина сердца
работает медленнее и, кроме того, меньше отдыхает, так как тратит
меньше энергии на свою работу.
Равенство сердечного выброса левого и правого желудочков
сердца — необходимое условие нормального кровообращения.
У здорового человека практически вся кровь, протекающая через
большой круг кровообращения, поступает в малый круг. Однако
часть крови (1—2%) минует правую половину сердца. Это связано с
поступлением крови из коронарных вен в левое предсердие. Кроме
того, часть крови попадает в легких из большого круга в малый, а
затем по легочным венам доставляется в левое предсердие. Поэтому
Рис. 5.1.12. Зависимость давления от объема левого желудочка сердца:
А — в норме; Б — при увеличении венозного давления; В — при увеличении
артериального давления (1 — асинхронное сокращение; 2 — изометричес­
кое сокращение; 3 — быстрое изгнание крови; 4 — медленное изгнание
крови; 5 — протодиастола; 6 — изометрическое расслабление; 7 — быст­
рое наполнение кровью; 8 — медленное наполнение кровью; 9 — систола
предсердий (КДО — конечный диастолический объем желудочка 125 мл;
КСО — конечный систолический объем желудочка 60 мл; УО — ударный
объем крови, выбрасываемой из желудочка, 65 мл; СД — систолическое
давление крови; ДД — диастолическое давление крови; ПД — пульсовое
давление крови)
290
Глава 5
правая половина сердца перекачивает чуть меньшее количество
крови, и ее ударный систолический объем составляет 64 мл, в то
время как ударный систолический объем для левой половины сердца
равняется 65 мл.
Фонокардиограмма как показатель сокращения сердца
Фонокардиограмма (ФКГ) — это последовательная запись звуковых
явлений в сердце. Звуковые явления в сердце связаны с колебаниями
клапанов, сосудов и различных отделов сердца в процессе его работы
в диапазоне звуковых частот.
Все звуковые явления в сердце разделяют на тоны и шумы. Тоны
возникают в норме, а шумы — в условиях патологии. Различают
четыре тона. Первый и второй тоны возникают всегда, а третий
и четвертый можно зарегистрировать у меньшей части людей (см.
рис. 5.1.9).
Первый тон происходит во время фазы изометрического сокра­
щения и в самом начале быстрого изгнания крови из желудочков.
Он возникает в момент закрытия атриовентрикулярных клапанов,
и обусловлен, главным образом, колебаниями натянутых створок
атриовентрикулярных клапанов и прикрепленных к ним сухожиль­
ных нитей. Частично первый тон связан с колебаниями полулунных
клапанов аорты и легочного ствола при их открытии.
Второй тон возникает в фазе изометрического расслабления желу­
дочков. Он связан с колебаниями аортальных клапанов и клапанов
легочного ствола при их захлопывании.
Третий тон можно зарегистрировать в фазе быстрого наполнения
кровью желудочков в результате колебания стенок желудочков при
наполнении их кровью из предсердий после открытия атриовентри­
кулярных клапанов.
Четвертый тон возникает во время систолы предсердий и связан
с колебаниями стенок желудочков при добавочном наполнении их
кровью из предсердий.
ЭКГ и ФКГ — основные инструментальные методы исследования
работы сердца у больных и здоровых людей. Поэтому имеет значение
соответствие зубцов ЭКГ и тонов ФКГ (см. рис. 5.1.13).
I
тон сердца соответствует комплексу зубцов QRS и возникает сразу
после вершины зубца R. При этом возбуждение желудочков переходит
в их сокращение. Атриовентрикулярные клапаны закрываются, это
сопровождается вибрацией их створок и трабекул, прикрепленных
Висцеральные функции
291
I ТОН
Рис. 5.1.13. Соответствие между зубцами ЭКГ и тонами сердца ФКГ (Р, Q,
R, S, Т — зубцы ЭКГ; I, II — тоны сердца)
к вершине створок и сосочковым мышцам. II тон сердца возникает
через протодиастолический период после окончания зубца Т. При
этом обратный ток порции крови, направленный в расслабляющиеся
желудочки, закрывает полулунные клапаны аорты и легочного ство­
ла, вызывая колебания стенок этих клапанов.
Функциональные особенности сердца
• В отличие от скелетной мускулатуры, сердце обладает автоматиз­
мом. Оно способно самопроизвольно возбуждаться и сокращаться
без внешних раздражений. Спонтанные возбуждения возникают
за счет медленной диастолической деполяризации атипических
клеток синоатриального узла.
• Особенностью сокращения и расслабления миокарда по сравне­
нию с функционированием скелетных и гладких мышц является
наличие в каждом сердечном цикле фаз изометрического сокра-
292
Глава 5
щения и изометрического расслабления, в ходе которых не изме­
няется длина кардиомиоцитов, а увеличивается или уменьшается
напряжение в них.
• Сила сокращения миокарда, так же как и сила сокращения
скелетной мышцы, зависит от исходной длины мышечных кле­
ток, от величины преднагрузки. Эта закономерность сердечной
ткани имеет значение для работы сердца, так как сила сокраще­
ния сердца зависит от количества поступающей в него крови.
Применительно к работе сердца эта закономерность называется
законом сердца Старлинга. Закон Старлинга заключается в том,
что сила сокращения сердца максимальна при средней степени
растяжения кардиомиоцитов. Изменение исходной длины кар­
диомиоцитов увеличивает силу сокращения, но не влияет на
скорость сокращения.
• Сердечная мышца не способна к тетаническому сокращению, воз­
никающему вследствие суммации отдельных одиночных сокра­
щений. Это свойство необходимо для бесперебойного ритмичес­
кого выброса крови из сердца. Отсутствие способности к тетанусу
связано с длительным абсолютным рефрактерным периодом в
кардиомиоцитах. В отличие от миокарда, в скелетных мышцах
период рефрактерности значительно короче, чем длительность
сокращения, что дает возможность суммироваться отдельным
сокращениям, в результате чего возникает тетаническое сокра­
щение. Потенциал действия кардиомиоцитов предсердий длится
180 мс, что превышает длительность сокращений правого и лево­
го предсердий (110 и 100 мс соответственно). Поэтому очередное
возбуждение предсердий возможно только после их расслабле­
ния. Потенциал действия миоцитов желудочков длится столько
же, сколько систола желудочков — 330 мс. Однако тетаническое
возбуждение желудочков невозможно в связи с большой продол­
жительностью потенциала действия (450 мс), а следовательно, и
длительностью абсолютного рефрактерного периода (400 мс) ати­
пичных кардиомиоцитов проводящей системы сердца. Поэтому
при увеличении частоты возбуждений, приходящих из синусного
узла, сначала частота сердечных сокращений увеличивается.
Затем при дальнейшем увеличении частоты возбуждений вмес­
то тетануса возникает сокращение отдельных кардиомиоцитов.
Это явление называется фибрилляцией кардиомиоцитов сердца.
Следовательно, при нормальной работе сердца следующее сокра­
Висцеральные функции
293
щение миокарда может возникнуть только после окончания пре­
дыдущего.
• За счет проведения возбуждения через нексусы от одних карди­
омиоцитов к другим миокард полностью охватывается распро­
страняющимся возбуждением. Поэтому сердце не подчиняется
закону изолированного проведения возбуждения, которому
подчиняются скелетные мышцы и нервы. Сердце подчиняется
закону «Все или ничего». Это означает, что при подпороговом
раздражении мышца сердца не возбуждается и не сокращается.
При пороговом или надпороговом раздражении сердце отвечает
максимальным сокращением, так как возбуждение передается
сразу на все кардиомиоциты. Это свойство отличает миокард
от скелетной мышцы, которая не подчиняется закону «Все
или ничего», так как возбуждение в ней не переходит с одного
мышечного волокна на другое, и каждое волокно имеет свой
порог раздражения.
• Особенностью сердечной мышцы, делающей ее уникальной по
сравнению с другими типами мышц, является зависимость силы
сокращения от частоты сокращения. Эта закономерность назы­
вается хроноинотропной зависимостью или феноменом лестницы
Боудича. Хроноинотропная зависимость выражается в том, что
с ростом частоты сердечных сокращений растет сила сокраще­
ний миокарда при неизменной исходной длине кардиомиоцитов.
Помимо увеличения силы сокращений сердца увеличивается
скорость этих сокращений, а также скорость расслабления мио­
карда. Механизм хроноинотропной зависимости связан с тем,
что не весь кальций поступает при возбуждении из Т-систем к
актиновым и миозиновым нитям. Частично он поступает в эндоплазматическую сеть и используется в ходе последующего сокра­
щения. Следовательно, ионные процессы каждого предыдущего
сокращения влияют на силу последующих сокращений. Знание
этой закономерности используется в практической кардиологии
в виде парной сочетанной стимуляции сердца для усиления его
работы. При возникновении возбуждения в сердце на него пода­
ется раздражение от кардиосинхронизатора, что вызывает допол­
нительный выход ионов Са2+ из Т-систем и потенцированное
сокращение сердца.
• В сердце могут наблюдаться внеочередные сокращ ения.
Внеочередное сокращение сердца называется экстрасистолой.
294
Глава 5
Экстрасистолы могут возникать у людей при физическом или
эмоциональном напряжении и при различных заболеваниях.
Экспериментально экстрасистолы могут быть получены при
прямом раздражении током сердца лягуш ки. Различают пред­
сердные (рис. 5.1.14А) и желудочковые (рис. 5.1.14Б) экстрасис­
толы.
При возникновении предсердной экстрасистолы источник внеоче­
редного возбуждения находится в предсердиях, чаще в синусном узле.
Внеочередное возбуждение возникает, когда рефрактерный период
окончился, но очередной автоматический потенциал действия еще не
появился. Пауза, следующая за предсердной экстрасистолой, длится
столько же времени, сколько и обычная пауза.
При желудочковой экстрасистоле источник внеочередного воз­
буждения находится в желудочках, чаще в предсердно-желудочковом узле. Внеочередное возбуждение желудочков не отражается на
автоматизме синоатриального узла, и он своевременно генерирует
очередное возбуждение, передающееся к кардиомиоцитам, когда те
находятся еще в рефрактерном состоянии во время экстрасистолы.
Поэтому кардиомиоциты не отвечают сокращением на очередное
возбуждение, поступающее из синоатриального узла. После окон­
чания экстрасистолы возбудимость кардиомиоцитов восстанавли­
вается, и они отвечают сокращением на последующее возбужде­
ние, распространяющееся из синусно-предсердного узла. Поэтому
после желудочковой экстрасистолы возникает компенсаторная пауза,
которая длится дольше, чем обычная пауза (рис. 5.1.14В). Таким
образом, при желудочковой экстрасистоле появляется неравномер­
ность ритма работы сердца, а частота сокращений сердца остается
прежней.
Экстрасистолы характеризуются слабостью сокращения, так как
к моменту начала экстрасистолы сердце только частично напол-
Рис. 5.1.14. Д инамика сердечных сокращений при экстрасистолах:
А — предсердная экстрасистола (ПЭ); Б — желудочковая экстрасистола
(ЖЭ); В — компенсаторная пауза после желудочковой экстрасистолы
(КП); а — направление распространения возбуждения в сердце челове­
ка; б — кардиограмма сердца лягушки (Р — внеочередное раздражение;
САУ — синоатриальный узел; АВУ — атриовентрикулярный узел;
ОВ — очередное возбуждение; ПэП — постэкстрасистолическая потенциация)
295
Висцеральные функции
кп
296
Глава 5
няется кровью, а актиновые и миозиновые нити находятся не в
оптимальном для сокращения положении. Сокращение возникает
при малой нагрузке на миокард, что по закону Старлинга вызывает
слабое сокращение. При желудочковой экстрасистоле вслед за ком­
пенсаторной паузой следует сильное сокращение сердца, превыша­
ющее по амплитуде обычное сокращение. Это явление называется
постэкстрасистолической потенциацией. Постэкстрасистолическая
потенциация связана с повышенным наполнением сердца кровью во
время компенсаторной паузы, что приводит к растяжению миокарда
и, согласно закону Старлинга, вызывает более сильное сокращение
сердца (рис. 5.1.14В).
• Зависимость силы сокращения сердца от частоты и ритмич­
ности возбуждения в сердце указывает на то, что ритм работы
сердца интегрирует работу сердца в единый сложно работающий
механизм. Управление ритмом автоматически приспосабливает
сердце к требуемому режиму работы. Поддержание одинакового
объемного кровотока в большом и малом круге кровообращения
осуществляется сердцем без участия нервной системы. С помо­
щью гемодинамической саморегуляции согласуется мощность
работы правой и левой половин сердца. Это хорошо видно на
опыте, в ходе которого левая и правая половины сердца живот­
ного заменяются двумя сердцами от разных животных. Одно сер­
дце накачивает кровь в малый круг кровообращения, а другое —
в большой круг. Каждое сердце имеет свой автоматизм и сокра­
щается с частотой, отличной от частоты сокращений второго
сердца. Систолы происходят несинхронно. Ударный объем крови,
выбрасываемой сердцами, различается. Однако минутный объем
крови, проходящей через оба сердца, одинаков.
Особенности метаболизма миокарда
В ходе метаболизма сердце использует различные источники энер­
гии: глюкозу, свободные жирные кислоты и молочную кислоту. В
покое миокард окисляет равное количество этих веществ. Важная
особенность метаболизма миокарда — использование молочной кис­
лоты, транспортируемой к сердцу кровью. Если в покое значение
этого явления невелико, то при работе 2/3 энергии сердце получает
при окислении молочной кислоты, которая поступает к сердцу из
работающих скелетных мышц. Остальную энергию сердце получает
при окислении свободных жирных кислот (1/5) и при окислении
глюкозы (1/7).
Висцеральные функции
297
Кровоток по коронарным сосудам сердца происходит только в
диастолу, а во время систолы коронарные артерии пережимают­
ся. Поэтому снабжение сокращающегося миокарда кислородом не
может осуществляться напрямую от гемоглобина крови. В карди­
омиоцитах находится белок миоглобин, который присоединяет к
себе кислород во время диастолы. Во время систолы окисление в
миокарде происходит за счет кислорода, который отщепляется от
миоглобина.
Нервно-гуморальная регуляция
функций сердца
Работа сердца связана с потребностями целого организма, с
деятельностью всех его функциональных систем, поддерживающих
гомеостаз. Поэтому на механизмы саморегуляции работы сердца
накладываются нервные и гуморальные влияния. Изменяя длитель­
ность и силу возбуждения и сокращения сердца, организм обеспечи­
вает необходимое в данный момент времени кровообращение.
Внутрисердечная нервная регуляция дополняет миогенную регуля­
цию сердечной деятельности по закону Старлинга. При умерен­
ном увеличении венозного притока крови к сердцу адренергичес­
кие нервные волокна увеличивают силу сердечных сокращений,
и венозное давление нормализуется (рис. 5.1.15А). При чрезмерном
увеличении венозного притока холинергические волокна притор­
маживают чрезмерное увеличение силы сердечных сокращений,
предохраняя сердце от истощения в нем энергетических запасов
(рис. 5.1.15Б).
Симпатические или парасимпатические рефлексы оказывают положи­
тельные или отрицательные влияния на функциональные процессы
сердца: хронотропный (изменение частоты), инотропный (изменение
силы сокращений), батмотропный (изменение возбудимости), дромотропный (изменение скорости проведения возбуждения), тонотропный
(изменение тонуса) (рис. 5.1.16Г).
Симпатические нервы иннервируют все сердце. Они ускоряют
медленную диастолическую деполяризацию в клетках синусного
узла (см. рис. 5.1.16А) и увеличивают возбудимость типичных и ати­
пичных кардиомиоцитов, вызывая положительные хроно-, батмо- и
дромотропные эффекты. Увеличивая вхождение в кардиомиоци­
ты ионов Са2+(см. рис. 5.1.16В), симпатические нервы увеличивают
длительность потенциалов действия кардиомиоцитов желудочков
Глава 5
2 98
А
Б
Рис. 5.1.15. Этапы внутрисердечной нервной регуляции сердечных сокра­
щений: А —при незначительном увеличении венозного давления; Б —при
значительном увеличении венозного давления (САУ — синоатриаль­
ный узел; АВУ — атриовентрикулярный узел; ВД — венозное давление;
НА — норадреналин; АХ —ацетилхолин)
Висцеральные функции
299
(см. рис. 5.1.16Б), что приводит к положительным ино- и тонотропным эффектам (см. рис. 5.1.16Г).
Блуждающие нервы иннервируют предсердия и уменьшают воз­
будимость клеток синоатриального и атриовентрикулярного узлов
за счет уменьшения проницаемости клеточных мембран кардиоми­
оцитов для ионов Са2+ (см. рис. 5.1.16В). В клетках синусного узла
уменьшается скорость медленной диастолической деполяризации
(см. рис. 5.1.16А), что обусловливает возникновение отрицательных
эффектов в сердце (см. рис. 5.1.16Г).
При продолжающемся ритмическом раздражении блуждающих
нервов возникает эффект ускользания сердца из-под влияния блуждающих
нервов. Влияние на функциональные процессы сердца ослабевает и
затем исчезает (рис. 5.1.17А). Ускользание может происходить за счет
рефлекторного возбуждения симпатической нервной системы, исто­
щения запасов ацетилхолина в блуждающих нервах, уменьшения
чувствительности холинорецепторов миокарда, выработки кардиомиоцитами холинолитических веществ.
Гуморальная регуляция ф ункции сердца происходит с помощью
гормонов, ионов и других биологически активных веществ, действу­
ющих через кровь коронарных сосудов. Положительные эффекты в
сердце вызывают гормоны: адреналин, глюкагон, кортикостероиды,
тироксин, кинины , простагландины. Ионы натрия обеспечивают
процесс возбуждения в сердце. Ионы калия формируют потенци­
алы покоя кардиомиоцитов. При увеличении концентрации калия
в крови и межклеточной жидкости происходит уменьшение уровня
потенциала покоя и возбудимости кардиомиоцитов. Это свойство
ионов калия используется для снижения у людей с повышенной
возбудимостью миокарда с помощью приема ими пищи богатой
калием. Ионы кальция обеспечивают электромеханическое сопря­
жение возбуждения с сокращением в сердце и увеличивают силу
сокращения миокарда (см. рис. 5.1.17Б). Ф ункциональная деятель­
ность сердца существенно угнетается ионами водорода, избыток
которых возникает при интенсивной физической работе, гипоксемии и гиперкапнии.
Рефлекторная регуляция функции сердца происходит при раздра­
жении механорецепторов сердца и сосудов. В регуляции прини­
мает участие сосудодвигательный центр и вегетативная нервная
система. При повышении артериального давления возникает депрессорный рефлекс, при повышении венозного давления — рефлекс
Бейнбриджа.
300
Глава 5
Норма
снс
пене
Са2+
Эффекты
1. Инотропный
(сила)
2. Хронотропный
(частота)
3. Батмотропный
(возбудимость)
4. Дромотропный
(проводимость)
5. Тонотропный
(тонус)
Висцеральные функции
301
Рис. 5.1.16. Влияние симпатической (СНС) и парасимпатической (ПСНС)
нервной системы на функции сердца: А — изменение скорости медленной
диастолической деполяризации (М ДД) в клетках синоатриального узла;
Б — изменение длительности потенциала действия в кардиомиоцитах
желудочков; В — влияние медиаторов на проницаемость клеточной мемб­
раны для ионов Са2+; Г — эффекты влияния СНС и ПСНС (НА — норадреналин; АХ — ацетилхолин)
А
V I/
а
б
/г 1U /1Wя1г/ У / у / у я у л
в
Р азд раж ение п. vagus
Рис. 5.1.17. Примеры нервно-гуморальной регуляции сердца. А — усколь­
зание сердца лягушки от действия блуждающего нерва: а — ритмичес­
кие сокращения сердца в норме; б — остановка сердца при раздражении
блуждающего нерва; в — возобновление и учащение сокращений сердца
при продолжающемся раздражении блуждающего нерва; Б — влияние на
сердце лягушки растворов КС1 и СаС12
Глава 5
3 02
Депрессорный рефлекс — возвращение артериального давления
к исходному уровню при его увеличении за счет раздражения баро­
рецепторов сосудов (рис. 5.1.18А). От барорецепторов возбуждение
проводится по депрессорному нерву в сосудодвигательный центр
продолговатого мозга, что приводит к возбуждению депрессорного и торможению прессорного отдела центра. Далее возбуждается
парасимпатическая и тормозится симпатическая нервная система.
Торможение сердечной деятельности и уменьшение сосудистого
тонуса вызывает снижение артериального давления до исходного
уровня.
Рефлекс Бейнбриджа — возвращение венозного давления к
исходному уровню при его повышении за счет раздражения меха­
норецепторов правого предсердия (см. рис. 5.1.18Б). От рецепторов
возбуждение проводится в сосудодвигательный центр продолгова­
того мозга, что приводит к возбуждению прессорного и торможе­
нию депрессорного отдела центра. Далее возбуждается симпати­
ческая и тормозится парасимпатическая нервная система. Усиление
сердечной деятельности снижает венозное давление до исходного
уровня.
Депрессорный рефлекс и рефлекс Бейнбриджа — часть ф ункцио­
нальной системы, поддерживающей оптимальный для метаболизма
уровень артериального давления.
5 .2 . ГЕМОДИНАМИКА СОСУДИСТОГО РУСЛА
Скорость кровотока
Гемодинамические показатели кровотока формируются биофи­
зическими параметрами всей сердечно-сосудистой системы в целом:
характеристиками сердечной деятельности, морфологией сосудов и
Рис. 5.1.18. Рефлекторная регуляция сердечной деятельности при увели­
чении артериального или венозного давления крови: А — процессы воз­
буждения и торможения вегетативной нервной системы при депрессорном
рефлексе; Б — процессы возбуждения и торможения вегетативной нервной
системы при рефлексе Бейнбриджа (СДЦ — сосудодвигательный центр;
П — прессорный отдел; Д — депрессорный отдел; АД — артериальное дав­
ление; ВД — венозное давление; АХ — ацетилхолин; НА — норадреналин;
ТН — тормозной нейрон)
Висцеральные функции
Работа сердца |
303
304
Глава 5
непосредственно свойствами самой крови. Как известно, движение
жидкости по трубам определяется разностью давлений в начале и в
конце трубы, ее диаметром и сопротивлением, которое испытывает
текущая жидкость вследствие трения между отдельными слоями
жидкости и о стенки трубы. Разность давлений способствует движе­
нию жидкости, а сопротивление препятствует ему. При этом общее
сопротивление труб, последовательно переходящих одна в другую,
равно сумме сопротивлений в каждой из них:
R = R\ + R2 + R3.
При параллельно соединенных трубах складываются их прово­
димости:
С = Cl + С2 + СЗ,
где С — проводимость в трубе; R — сопротивление стенки трубы
протекающей жидкости (законы Кирхгофа).
При этом, общее сопротивление параллельно расположенных
сосудов должно быть меньше, чем сопротивление в каждом отде­
льном сосуде.
С = 1/Л.
Отношение этих величин определяет объемную скорость, т.е.
объем жидкости, протекающей в единицу времени. Объемную ско­
рость (Q) вычисляют по первому уравнению Пуазейля (1854).
Объемная скорость — гидродинамический показатель, характеризу­
ющий количество жидкости, протекающее через поперечное сечение
трубы (сосуда) за 1 мин.
Q = (/>, - P2)/R,
где Q — объемная скорость движения жидкости (крови), мл/мин,
R — гидродинамическое сопротивление, дин*с/см; /*, —Р2 — разность
давлений в начале и конце сосуда, мм рт. ст.
Объемная скорость кровотока различна в разных органах (табл. 5.2);
она зависит от просвета сосуда: самая высокая скорость кровотока в
аорте и полых венах, самая низкая — в каждом отдельном капилля­
ре. Однако объемная скорость кровотока постоянна во всех сосудах
одного калибра, так как количество крови, протекающей через раз­
305
Висцеральные функции
ные участки сосудистого русла, например, через все артерии и вены,
одинаково в единицу времени.
Таблица 5.2. Объемная скорость кровотока в разных органах
Объемная скорость кровотока,
мл/мин
Орган
Щитовидная железа
Почки
Печень
Сердце (через коронарные сосуды)
Кишечник
560
420
150
85
50
65
Мозг
Селезенка
Желудок
Мышцы конечностей (в покое)
70
35
2 -3
Для расчета величины сопротивления току крови на определен­
ном участке сосудистой сети можно использовать приведенную выше
формулу.
Сопротивление току крови тем больше, чем больше ее вязкость,
чем больше длина сосуда, по которому течет кровь, и чем меньше
радиус этого сосуда. Зависимость сопротивления R от этих величин
отражает второе уравнение Пуазейля.
Уравнение Хагена—Пуазейля (1846) отражает зависимость сопротив­
ления тока жидкости от вязкости крови, длины и радиуса трубки
(сосуда) — объемная скорость пропорциональна, а гидродинами­
ческое сопротивление обратно пропорционально радиусу сосудов в
четвертой степени.
R = 8/ri/jtr4,
где R — гидродинамическое сопротивление; / — длина сосуда;
г — радиус сосуда; г| — вязкость крови; л — отношение окружности
сосуда к диаметру.
В соответствии с уравнением максимально большое сопротивление
движению крови оказывают артериолы и несколько меньшее — капил­
ляры в связи с их малой длиной по сравнению с артериолами.
Глава 5
306
Высокое сопротивление артериол и капилляров обусловливает то,
что именно на этом участке сосудистого русла давление крови значи­
тельно падает. 85% энергии, затрачиваемой сердцем на продвижение
крови по организму, расходуется в артериолах и капиллярах, а 10 и
5% — соответственно в артериях и венах. Увеличение вязкости крови
приводит к уменьшению линейной скорости:
V = 1/л,
где V — линейная скорость.
Вязкость (внутреннее трение) — свойство жидкости оказывать
сопротивление перемещению одной ее части относительно другой.
Вязкость крови, как и любой другой жидкости, возрастает при сни­
жении температуры.
Линейная скорость кровотока. Кроме объемной скорости кровото­
ка, важным показателем гемодинамики является линейная скорость
кровотока, т.е. расстояние, которое частица крови проходит за еди­
ницу времени. Линейная скорость кровотока V обратно пропорцио­
нальна площади поперечного сечения сосудов я г2 одного калибра:
V = 0 /я г2.
Уравнение, отражающее закон неразрывности струи. Если жидкость с
постоянной объемной скоростью движется по системе трубок (сосудов)
различного диаметра, то линейная скорость движения жидкости обрат­
но пропорциональна площади поперечного сечения трубы (сосуда):
Vx = 52 V2 или S \/S 2 = V2
где 5 р S2 — площадь сечения трубок (сосудов), V{, V2 — линейная
скорость движения жидкости (крови).
Поскольку объемная скорость кровотока не меняется по ходу
сосудистого русла, линейная скорость зависит только от общей попе­
речной площади сосудов одного калибра. Чем больше площадь,
тем меньше скорость. Во время выброса крови из сердца линейная
скорость крови равняется 50—60 см/с. В артериях максимальная
Висцеральные функции
скорость кровотока равня­
ется 25—40 см/с. В артериолах толчкообразное течение
крови сменяется непрерыв­
ным. Самая низкая скорость
кровотока в капиллярах —
0,5 мм/с. В венах линейная
скорость кровотока возрас­
тает до 20 см/с.
В связи с наличием сил
трения между кровью и
стенкой сосуда линейная
скорость максим альна в
центре сосуда и минимальна
у его стенок. Графики сум­
марного просвета сосудов и
линейной скорости крово­
тока в различных участках
кровеносного русла пред­
ставлены на рис. 5.2.1. Сила,
создаваемая движущ ейся
кровью, текущ ей парал­
лельно поверхности сосуда,
Рис. 5.2 .1 . Динамика изм е­
нений основных гемодинамических показателей при
движении крови через пос­
ледовательные отделы сосу­
дистого русла: А — поступ­
ление крови в артериальный
отдел сосудистого русла;
Б — переход крови в веноз­
ный отдел сосудистого русла;
1 — суммарный просвет
сосудов; 2 — структура сосу­
дистого русла; 3 — линей­
ная скорость кровотока;
4 — кровяное давление
307
см/сек
Глава 5
308
создает напряжение сдвига т (shear stress), которое кровь производит на
сосудистую стенку.
х = 4ri<2/p/-i,
где г| — вязкость крови; Q — объемный кровоток; р — плотность крови;
г — радиус сосуда.
По этой причине текущая кровь производит смещение и деформи­
рование слоев эндотелия, что вызывает высвобождение эндотелийзависимых сосудорасширяющих факторов.
Кровяное давление
В реальном сосудистом русле на чисто механические закономернос­
ти движения жидкости по сосудам накладываются физиологические
факторы: работа сердца, изменения тонуса сосудов, объема циркули­
рующей крови и вязкости крови, которые определяют особенности
кровообращения в различных частях организма. Важнейшие показа­
тели кровообращения — кровяное давление (Р), сила (F), действующая
со стороны крови на сосуды, приходящаяся на единицу площади (S).
Р = F/S.
Давление связано со скоростью движения крови. Согласно закону
Бернулли давление, оказываемое движущейся жидкостью, обратно
пропорционально скорости ее движения:
рК2/2 + Р = const,
где р — плотность жидкости (крови); V — линейная скорость движения
жидкости (крови); м/с, Р — гидростатическое давление, мм рт. ст.
Давление крови в артериях прямо зависит от объема крови Q,
поступающей из сердца, и сопротивления оттоку крови R перифе­
рических сосудов:
Р = QR-
В последние годы введено новое понятие «compliance», которое
можно трактовать как «растяжимость», представляющее собой отно­
шение увеличения объема крови в сосуде к увеличению развивае­
мого в нем давления. Тогда АД прямо пропорционально зависит от
Висцеральные функции
309
величины систолического объема и обратно пропорционально —
от растяжимости артериального русла.
Необходимо отметить, что сосудистая стенка не является пас­
сивным элементом кровообращения. Под влиянием движущейся
по сосуду крови возникает напряжение сдвига в клетках эндотелия
сосудов. Вследствие этого эндотелиальные клетки усиленно синте­
зируют оксид азота (NO), который диффундирует в гладкие мышцы
и вызывает расширение сосуда.
Кровяное давление в аорте и крупных артериях постоянно колеб­
лется. На кривой АД, зарегистрированной прямым методом, разли­
чают волны первого, второго и третьего порядка (рис. 5.2.2).
Волны первого порядка — периодические увеличения и уменьшения
АД, связанные с ритмическим выбрасыванием крови из левого желу­
дочка. Давление крови в аорте повышается с 80 до 120 мм рт. ст. при
выбросе крови из левого желудочка в фазе быстрого изгнания. В этот
период приток крови в аорту из сердца больше, чем отток в артерии.
Затем давление в аорте уменьшается. Весь период уменьшения связан
с оттоком крови из аорты на периферию.
п
hi
1'^MWVWWVWVVMM/4M//n/WMWWW\avv\A/v'a/vVv^'Ww
I— i
Время,2c
Рис. 5.2 .2 . Колебания артериального давления: I — волны первого порядка
(пульсовые); II — волны второго порядка (дыхательные); III — волны тре­
тьего порядка (центральные)
Глава 5
310
Максимальное давление в аорте и в артериях во время систолы
желудочков называется систолическим, а минимальное давление во
время диастолы — диастолическим. Нормальными значениями АД у
человека, измеренного на плечевой артерии, считают систолическое
АД 110—130 мм рт. ст., а диастолическое АД 70—80 мм рт. ст. Разница
между систолическим и диастолическим АД называется пульсовым
давлением. Среднестатистически это давление равно 40—50 мм рт. ст.
Пульсовое давление показывает, насколько систолическое давление
превышает диастолическое, что необходимо для открытия полулун­
ных клапанов аорты во время систолы левого желудочка.
ПД = АДс - АДд,
где ПД — пульсовое давление, мм рт. ст.; АДс — систолическое
АД, мм рт. ст.; АДд — диастолическое АД, мм рт. ст.
Нормативные показатели ПД = 35—55 мм рт. ст. Пульсовое дав­
ление можно рассматривать как приращение артериального объема
(Q). Это увеличение равно объему крови, выбрасываемому в аорту
левым желудочком во время фазы быстрого изгнания, минус объем
крови, который вытекает из артерий и микроциркуляторного русла
во время той же фазы.
В ряде случаев в артериальном русле определяют среднее кровяное дав­
ление, отражающее величину средней энергии, которой обладает кровь во
время систолы и диастолы. Среднее давление не есть средняя арифме­
тическая величина между систолическим и диастолическим давлением.
Для его определения необходимо измерить площадь под кривой АД и
разделить ее на длину этой кривой. Продолжительность систолического
давления меньше, чем уменьшение диастолического, поэтому среднее
давление ближе к величине диастолического давления. Среднее кровяное
давление выражает энергию непрерывного движения крови, представля­
ет собой довольно постоянную величину для данного сосуда и данного
организма и отражает степень эластичности артериальной стенки:
СКД = АДд + (АДс - АДд)/3,
или
СКД = АДд + ПД/3,
где СКД — среднее кровяное давление, мм рт. ст.; ПД — пульсо­
вое давление, мм рт. ст.; АДс — систолическое давление, мм рт. ст.;
АДд — диастолическое давление, мм рт. ст.
г
Висцеральные функции
311
Среднее кровяное давление определяется работой сердца, ОЦК
(объем циркулирующей крови), вязкостью крови и сосудистым сопро­
тивлением.
Волны второго порядка — периферическое изменение систолического
и диастолического давления, связанное с дыхательными движениями
грудной клетки. Волны второго порядка соответствуют дыхательным
волнам и, следовательно, имеют меньшую частоту и большую дли­
тельность, чем волны первого порядка. Происхождение волн второго
порядка связано с изменением внутригрудного давления. Во время
вдоха увеличивается объем грудной клетки, что ведет к уменьшению
давления в плевральной полости. При этом изменяется давление крови
в сосудах большого круга кровообращения в грудной полости.
Сосуды большого круга кровообращения, прилегающие своими стен­
ками к плевральной полости (аорта, артерии), изменяют свою конфи­
гурацию при уменьшении давления в плевральной полости. Изменение
диаметра сосудов приводит к уменьшению в них сопротивления току
крови. При вдохе наблюдается обратное явление. Повышение давления
в плевральной полости приводит к повышению давления в сосудах
большого круга кровообращения. При вдохе давление в плевральной
полости равно — 6 мм рт. ст., а при выдохе — 3 мм рт. ст.
Волны третьего порядка — медленные колебания (1—3 в минуту), на фоне
которых проявляются изменения волн первого и второго порядка, связан­
ные с изменением тонуса дыхательного и сердечно-сосудистого центров.
Давление в различных отделах сосудистого русла
Физическим фактором, обеспечивающим передвижение крови
в системе кровообращения, является разница давления в начале и
конце как всего сосудистого русла, так и на его отдельных участках.
Эта разница может создаваться повышением давления в начальном
отделе сосуда, снижением давления в конце сосуда, либо сочетанием
того и другого. Динамика изменения давления крови в сосудах раз­
личного типа представлена на рис. 5.2.1.
При продвижении крови от сердца к периферии колебания давле­
ния ослабевают в связи с эластичностью аорты и артерий, поэтому
кровь в аорте и артериях продвигается толчками, а в артериолах и
капиллярах — непрерывно.
Наибольшее падение давления происходит в артериолах и затем в
капиллярах. Несмотря на то что капилляры имеют меньший диаметр,
чем артериолы, уменьшение давления на более значительную величину
происходит в артериолах. Это связано с их большей длиной по сравне­
312
Глава 5
нию с капиллярами. В артериальной части капилляра (на «входе») дав­
ление крови равно 35 мм рт. ст., а в венозной (на «выходе») — 15 мм рт. ст.
В полых венах давление приближается к -3 — 5 мм рт. ст.
Классиф икация сосудов
Все составляющие сердечно-сосудистой системы, в том числе
сосуды, классифицируют по морфофункциональным признакам
(по Фолкову).
Сердце — главный «насос», который ритмически выбрасывает
кровь в аорту.
Амортизирующие сосуды — аорта, крупные и средние артерии
эластического типа, обладающие большой растяжимостью, функция
которых заключается в превращении пульсирующего сердечного
выброса в непрерывный равномерный кровоток.
Резистивные сосуды — прекапиллярные сосуды — мелкие арте­
рии и артериолы, имеющие в составе сосудистой стенки гладкие
мышечные волокна и обладающие высоким внутренним (миогенным)
базальным тонусом. Эти сосуды обеспечивают большую часть сопро­
тивления кровотоку, реагируют на симпатические влияния и обла­
дают химической чувствительностью к ряду биологически активных
веществ — метаболитам, гормонам. И.М. Сеченов назвал эти сосуды
«главными кранами» сосудистой системы, которые влияют на снабже­
ние тканей кровью. Прекапиллярные сфинктеры — мелкие, они изме­
няют число капилляров, перфузируемых в данный момент. Базальный
миогенный тонус этих сосудов находится под контролем метаболитов,
особенно сосудорасширяющих. Сокращение гладкомышечных клеток
этих сосудов может привести к перекрытию просвета мелких сосудов
и ограничить поступление крови в капиллярное русло. Таким образом
регулируется количество капилляров, участвующих в кровотоке.
Обменные сосуды — капилляры — выполняют функцию транс­
капиллярного обмена веществ и газов между кровью и тканями на
основе процессов фильтрации, диффузии, осмоса.
Шунтирующие сосуды, или артериоловенулярные анастомозы,
обеспечивают прямое поступление артериальной крови в венозную
часть русла, минуя капилляры.
Емкостные сосуды представлены сосудами венозного ложа. Они
не играют существенной роли в создании общего сопротивления току
крови, но существенно влияют на емкость сосудистого русла, изменяя
конфигурацию и диаметр просвета. Даже самое небольшое изменение
просвета этих сосудов вызывает значительные изменения распределения
Висцеральные функции
313
объема крови, центрального венозного давления, величины венозного
возврата к сердцу и, следовательно, последующего сердечного выброса.
Мелкие, средние и крупные вены называются сосудами возврата,
они обеспечивают возврат крови к сердцу и вмещают до 20% всей
циркулирующей крови.
Пульс
Артериальный пульс. Периодические колебания стенок артерий,
вызванные волной повышения давления, называются артериальным
пульсом. Пульс имеет несколько характеристик: частоту, амплитуду,
быстроту, ритм, напряжение. Частота пульса определяется частотой
сердечных сокращений. Амплитуда пульса — показатель систоли­
ческого объема и эластичности стенок артерии. Быстрота пульса
(быстрый, медленный) отражает скорость увеличения и уменьшения
давления крови в артериях при сокращении левого желудочка. Ритм
пульса (правильный, неправильный) — показатель регулярности
работы сердца. Напряжение пульса (твердый, мягкий) определяется
силой нажатия пальпирующего пальца, которой противодействует
сопротивление стенки артерии. Запись артериального пульса назы­
вается сфигмографией. На сфигмограмме (рис. 5.2.3) различают анакроту, катакроту, инцизуру и дикротический подъем, природа кото­
рых связана с волнами первого порядка, т.е. с изменением давления
крови в аорте при выбросе крови из сердца. Стенка аорты при этом
несколько растягивается, а затем возвращается к исходному размеру
вследствие своей эластичности. Механическое колебание стенки
аорты, называемое пульсовой волной, передается далее на артерии,
артериолы и здесь, не доходя до капилляров, затухает.
Скорость распространения пульсовой волны выше скорости течения
крови и в среднем равна 10 м/с. Пульсовая волна достигает лучевой арте­
рии в области запястья (наиболее часто используемое место регистрации
пульса) примерно за 100 мс при расстоянии от сердца до запястья 1 м.
Следовательно, при синхронной регистрации пульса лучевой артерии и
процессов сокращения сердца пульсовые колебания будут запаздывать
на 100 мс. Скорость пульсовой волны, вызванной выбросом крови из
левого желудочка в период систолы, рассчитывается по формуле:
где V — скорость распространения пульсовой волны, м/с; г — внут­
ренний радиус сосуда, мм; d — толщина стенки сосуда, мм; р — плот-
314
Глава 5
Рис. 5.2 .3 . Запись артериального пульса (сфигмограмма): cd — анакрота;
de — систолическое плато; gh — катакрота; / — инцизура; g —дикротичес-
кий зубец
ность крови; К — коэффициент пропорциональности; е — модуль
упругости стенки сосуда.
Если колебания давления, распространяясь от сердца к периферии,
постепенно затухают, то амплитуда каждой фазы пульса в перифери­
ческих артериях увеличивается. В артериолах пульс затухает оконча­
тельно и отсутствует в капиллярах, венулах, мелких и средних венах.
Венный пульс. В крупных венах, расположенных рядом с сердцем,
на флебограмме регистрируют венный пульс, в котором различают
волны первого и второго порядка (рис. 5.2.4).
К волнам первого порядка относят зубцы а, с, v Первая положитель­
ная волна а обусловлена застоем крови в полых венах во время систолы
правого предсердия. Вторая положительная волна с связана с ударом
крови в сонную артерию и в стенку яремной вены. Первая отрицатель­
ная волна х определяется разряжением в предсердиях в начале систолы
желудочков и притоком крови из вен. Третья положительная волна v
обусловлена переполнением предсердия и застоем крови в полых венах.
Вторая отрицательная волна у отражает фазу быстрого наполнения
правого желудочка и быстрого опорожнения вен.
Висцеральные функции
315
Рис. 5.2 .4. Одновременная запись ЭКГ (А) и пульсации яремной вены (Б)
Плетизмография. Последовательная запись кровенаполнения отде­
льных частей тела называется плетизмографией. Кровенаполнение
зависит от разности между величинами притока крови по артериям
и оттока ее по венам. Так, например, объем кровенаполнения паль­
ца руки постоянно меняется вследствие неодинакового количества
крови, притекающей к пальцу в систолу и диастолу сердца. Однако
эти изменения в пальце будут запаздывать по сравнению с процес­
сами в сердце примерно на 2 с, так как скорость течения крови в
среднем составляет 0,5 м/с, а расстояние от сердца до кисти — при­
мерно I м. Изменения на плетизмограмме аналогичны изменениям
давления в аорте и изменениям пульсовой волны. На плетизмограм­
ме также различают анакроту, катакроту, инцизуру и дикротический
подъем. Однако вследствие различной скорости распространения
пульсовой волны и скорости течения крови аналогичные участки
будут возникать в разное время на сфигмо- и плетизмограммах при
их одновременной регистрации. По плетизмограмме можно оценить
объемную скорость кровотока в исследуемой области.
Факторы, обеспечиваю щ ие движение крови по сосудистому
руслу
Существует два режима течения крови: ламинарный и турбулент­
ный. Ламинарное течение — это упорядоченное течение жидкости, при
котором она перемещается слоями, параллельными направлению тече­
316
Глава 5
ния. С увеличением скорости движения ламинарное течение переходит
в турбулентное, при котором происходит перемешивание всех слоев
жидкости с образованием многочисленных завихрений. Режим течения
можно предсказать путем расчета числа Рейнольдса (NR) по формуле:
NR = pDv/r\,
где р — плотность жидкости; D — диаметр трубки; v — средняя ско­
рость движения жидкости; г| — вязкость.
При NR менее 2000 течение будет ламинарным, а при NR больше
3000 — турбулентным. Кроме того, при увеличении скорости кро­
вотока увеличивается и скорость, при которой один слой жидкос­
ти сдвигается относительно другого. Это явление получило назва­
ние утончение сдвига. Таким образом, турбулентность возникает при
понижении вязкости жидкости, при повышении скорости движения
жидкости, большом диаметре сосуда, высокой плотности жидкости
и при неровностях сосудистых стенок.
Следует отметить, что вихри потока изначально существуют, когда
кровь выталкивается из желудочка в аорту. У мест разветвления
сосудов, а также при возрастании скорости кровотока (при физи­
ческой работе) течение может стать турбулентным и в артериях.
Турбулентное течение связано с дополнительной затратой энергии,
что может привести к дополнительной нагрузке на сердце.
Известно, что кровь движется по большому и малому кругу кро­
вообращения под влиянием различных гемодинамических факторов
(рис. 5.2.5). Время, в течение которого частичка крови проходит большой
и малый круг кровообращения, называется временем кругооборота крови, и
оно составляет в покое 21—23 с. Различают следующие факторы, действу­
ющие в артериальном и венозном коленах системы кровообращения.
В артериальном колене к таким факторам относят:
• работу сердца;
• эластическую силу крупных артерий;
• силу притяжения порции крови, поступившей в сосуды туловища
и верхних конечностей.
В венозном колене возврат крови к сердцу обеспечивают:
• в момент диастолы правого предсердия давление в нем и в полых
венах снижается, и кровь устремляется сюда из нижерасположен­
ных вен;
• присасывающая сила грудной клетки в фазе вдоха, когда давление
в грудной полости снижается, а внутригрудные вены расширяют-
Висцеральные функции
317
Рис. 5.2.5. Факторы, обеспечивающие динамику движения крови по сосу­
дистому руслу: А — в аорте кровь движется вследствие сокращения сердца,
а в начальном участке венозного русла — вследствие работы «мышечного
насоса» нижних конечностей; Б — в артериальном отделе кровь движется
за счет давления, создаваемого «компрессионными» камерами крупных
артерий, в венозном — за счет давления со стороны органов брюшной
полости; В — в артериальном отделе кровь перемещается также под дей с­
твием силы тяжести, в венозном — при участии «дыхательного насоса»;
Г — наконец, в венозном отделе кровь возвращается к сердцу вследствие
ритмической деятельности «предсердного насоса»
Глава 5
318
ся и давление в них падает до —5 мм рт. ст., что приводит к заса­
сыванию крови в правое предсердие;
• капиллярные силы поверхностного натяжения;
• наличие клапанов в венах, которые находятся ниже уровня сердца;
• активность скелетных мышц нижних конечностей, поскольку
при их сокращении тонкостенные вены сжимаются, и кровь из
них выдавливается вверх через клапаны;
• сила тяжести крови, поступающей по яремным венам к предсер­
дию. В венах, расположенных ниже сердца, гидростатическое
давление крови суммируется с давлением крови, создаваемым
сердцем. В венах, расположенных выше сердца, венозное дав­
ление уменьшается на величину гидростатического давления и
может быть ниже атмосферного;
• перистальтика киш ечника, действующая на вены брюшной
полости.
Микроциркуляция
М икроциркуляция представляет собой движение жидкостей по
микрососудам от артериального отдела к венозному. Структурной
основой — модулем — процессов микроциркуляции являются: артериола, прекапиллярная артериола (прекапилляр), капилляр, посткапиллярная венула (посткапилляр), артериолярно-венулярные анас­
томозы и венула.
Из уравнения Лапласа:
Т = Рг,
где Т — напряжение стенки сосуда; Р — трансмуральное давление;
г — радиус сосуда, следует, что из-за узкого просвета тонкостенные
капилляры способны выдержать большое давление. Так, при нор­
мальном кровяном давлении в аорте и капилляре напряжение стен­
ки капилляра в 12 ООО раз меньше, чем напряжение стенки аорты.
В микроциркуляторном русле формируется около 70—80% всего
внутрисосудистого сопротивления. В большинстве тканей перфузируется только часть (20—30%) капилляров. Изменение числа перфузируемых капилляров возникает как адаптивный ответ тканей на
гипоксию. Большую роль при этом играют вазомоции — медленные
колебания, обусловленные изменением просвета прекапиллярны х
артериол.
Выделяют три вида капилляров: соматические капилляры, у которых
стенка образована сплошным слоем эндотелиальных клеток с редки­
Висцеральные функции
319
ми порами. Такие капилляры характерны для соматической и гладкой
мускулатуры, кожи, легких, ЦНС, жировой и соединительной тканей.
Они пропускают воду и растворенные в ней соли. Висцеральные капилля­
ры, обладающие «окнами» размером 25% площади поверхности клеток,
через которые проходит большое количество воды с растворенными в
ней веществами и макромолекулы сложных соединений. Такие капил­
ляры встречаются в клубочках почек, эпителиоцитах кишечника,
эндокринных железах. Наконец, синусоидные капилляры с прерывистой
эндотелиальной оболочкой, которая пропускает макромолекулы и
форменные элементы крови, и большими межклеточными пространс­
твами. Это капилляры селезенки, печени, костного мозга.
Кровь, протекающая по микрососудам, представляет собой гете­
рогенную мультикомпонентную суспензию форменных элементов,
взвешенных в растворе электролитов, белков и липидов. Имеется
прямая связь между вязкостью крови и показателем гематокрита, а
также обратная связь со значением кислотно-щелочного состояния
крови. Гиперкапния способствует усилению вязкости крови, что
определяет большую вязкость венозной крови, чем артериальной.
Вязкость крови — основное свойство, обеспечивающее перфузию
микрососудистого русла. Как было отмечено выше, вязкость крови
зависит от скорости ее движения. В артериях эта зависимость незна­
чительна, а в микрососудах — существенна. В микрососудах вязкость
крови значительно ниже, чем в крупных сосудах. Причина этого —
параболический профиль тока форменных элементов, когда пос­
ледние из-за поперечной разницы давления в сосуде смещаются от
периферии к центру. Вследствие этого скорость движения клеток
крови через капилляр выше, чем у цельной крови. Кроме того, эрит­
роциты имеют на своей поверхности дзета-потенциал около 35 мв,
что обеспечивает их отталкивание от стенки сосуда и от других
эритроцитов. Это препятствует склеиванию клеток крови в сосу­
дистом русле. Отсутствие ядра дает дополнительную возможность
эритроцитам проходить через капилляры, имеющие размеры меньше
поперечника клетки. Эндотелий сосудов занимает 5000 м2 при массе
до 3 кг. Он служит источником вазоактивных веществ. К сосудорас­
ширяющим веществам относят оксид азота (N 0), простациклины,
простагландины, эндотелий-зависимый фактор гиперполяризации.
К сосудосуживающим веществам относят эндотелии, супероксиданион, вазоконстрикторные простаноиды и ангиотензин II.
Между микрососудами и клетками ткани возникает транссосу­
дистый обмен веществ. Роль основных механизмов обмена исполня­
3 20
Глава 5
ют фильтрация, реабсорбция, диффузия и микропиноцитоз. Фильтрация
и реабсорбция основаны на разности гидростатического давления
в капилляре и окружающей ткани, а также на разности онкотического давления, создаваемого белками плазмы крови и тканей
(рис. 5.2.6). При этом в каждом капилляре идут процессы фильтра­
ции и реабсорбции. В настоящее время установлено, что имеются
капилляры, которые только фильтруют, тогда как есть и другие
капилляры, которые только реабсорбируют. Результирующее давле-
Рис. 5.2.6. Обмен жидкостью между кровеносным капилляром и межкле­
точным пространством в процессе прохождения по нему крови: А — век­
торы сил, действующих на стенку капилляра; Б — направление движения
жидкости между капилляром и межклеточным пространством; Ргд— гид­
равлическое давление в начале и конце капилляра; Ронк кр — онкотическое
давление крови; Ротк тк — онкотическое давление в тканях; Рф — фильтра­
ционное давление; Рр — реабсорбционное давление (мм. рт. ст.)
Висцеральные функции
321
ние в артериальном конце капилляра определяет выведение из него
веществ в ткань; результирующее давление в его венозном конце —
поступление веществ в просвет сосуда. Диффузия обеспечивается
разностью концентраций веществ между капилляром и окружа­
ющей тканью. Диффузия определяет переход кислорода и воды.
В основе пиноцитоза лежит активный транспорт при помощи вези­
кул эндотелиальных клеток. Пиноцитоз обеспечивает транспорт
у-глобулинов, гликогена и миоглобина. Степень затрудненности про­
хождения вещества через стенку капилляра выражается коэффициен­
том отражения (а). Коэффициент отражения для воды равен нулю,
а для альбумина (который практически не проходит через стенку) —
единице. Проницаемость сосудистой стенки увеличивается под
влиянием серотонина, гистамина, брадикинина, а уменьшается —
под влиянием ионов кальция, витаминов РР и С, катехоламинов.
Лимф атическая система
В лимфатических сосудах взрослого человека находится до 3 л
лимфы. По своему составу лимфа сходна с плазмой крови, но содер­
жит меньше белка. Сама лимфатическая система представляет собой
совокупность лимфатических сосудов и лимфатических узлов, обес­
печивающих всасывание межклеточной жидкости и возврат веществ
в кровяное русло. Лимфатические сосуды практически отсутствуют в
ЦНС, мозговых оболочках, хряще, роговице, глазном яблоке. Стенки
сосудов обладают большой проницаемостью для молекул белка и
крупных частиц. Лимфатическая система выполняет ряд функций:
• в лимфоидной ткани продолжаются процессы кроветворения,
происходит формирование новых лимфоцитов;
• лимфатическая система обеспечивает дренаж (удаление) из интерстиция продуктов обмена и избытка воды;
• участвуя в транспорте антигенов и формировании первичного и
вторичного иммунного ответа, лимфатическая система выполня­
ет защитную функцию;
• лимфатическая система осуществляет транспорт продуктов гид­
ролиза (липидов) из пищеварительной системы и возврат белков
и электролитов в кровь.
Движение лимфы происходит за счет сократительной активности
лимфатических сосудов, присасывающего действия грудной клетки,
наличия клапанов в лимфатических сосудах. Регуляция движения
лимфы осуществляется за счет сокращения миоцитов лимфангионов
и влияния на них симпатической нервной системы.
Глава 5
3 22
Коронарное кровообращение
Специфика коронарного кровообращения заключается в доставке
кислорода и питательных веществ сердцу, выполняющему постоян­
ные ритмические сокращения. Главный источник энергии для мио­
карда — жирные кислоты. Большое значение имеет наличие в крови
глюкозы и лактата, которые обеспечивают потребности миокарда в
энергии на 30—40%. Коронарный круг кровообращения начинается
от аорты и заканчивается венозным синусом, который впадает в пра­
вое предсердие, или отдельными венами, проникающими в полость
сердца (табезиевы вены). Венечный кровоток потребляет 4—8% всего
систолического объема крови. Для венечного круга характерны сле­
дующие особенности:
• коронарный кровоток зависит от фазы сердечного цикла;
• кровь в венечные сосуды поступает во время диастолы. Это связа­
но с тем, что в фазе систолы устья капилляров закрываются полу­
лунными клапанами аорты, а также с тем, что во время систолы
миокард сокращен, венечные сосуды пережаты, и поступление
крови в них затруднено;
• в период диастолы миоглобин сердечной мышцы насыщается кис­
лородом, который он очень легко отдает сердцу в фазе систолы;
• высокое давление, поскольку венечные сосуды начинаются от аорты;
• венечные сосуды образуют в сердечной мышце густую капилляр­
ную сеть с множеством сосудов конечного типа, что представляет
опасность при их закупорке, особенно в преклонном возрасте;
• наличие артериоловенулярных анастомозов и артериоло-синусоидных шунтов;
• коронарный кровоток ограничивают укорочение диастолы при
тахикардии и падение АД; при брадикардии наблюдается мень­
шее ограничение коронарного притока (больше время пребыва­
ния в диастоле), но также меньше и метаболические потребности
миокарда в кислороде;
• минимальное кровоснабжение миокарда имеет место во время
изометрического сокращения;
• коронарные капилляры имеют высокий базальный тонус;
• сосуды сердца имеют двойную иннервацию — симпатическую и
парасимпатическую.
Регуляция коронарного кровообращения
Кровообращение сердца находится под влиянием многих факто­
ров. Сердце непрерывно сокращается, и просвет венечных сосудов
Висцеральные функции
323
зависит не только от нервных и гуморальных влияний, но и от меха­
нических факторов и локальных метаболических процессов. При
усилении или ослаблении сердечной деятельности взаимодействие
этих факторов и значение каждого из них могут изменяться.
Гемодинамическая регуляция
Как известно, величина венечного кровотока находится в линейной
зависимости от величины перфузионного (АД) давления в крупных
сосудах, и, прежде всего, в аорте, в связи с нагнетательной функцией
сердца. Экстраваскулярное сжатие сосудов миокарда угнетает крово­
ток в них. Венечный кровоток уменьшается во время систолы и быстро
увеличивается с началом диастолы. Наибольшей величины он дости­
гает в фазе изометрического расслабления. Суммарный кровоток в
течение систолы составляет 40,8%, а во время систолы и фазы изомет­
рического расслабления — 68,7% всего венечного кровотока. Учащение
сердечных сокращений приводит к увеличению венечного кровотока.
При этом действуют два противоположных фактора: прирост ЧСС
вызывает уменьшение кровотока, поскольку при этом относительно
увеличивается общее время систолического кровотока по сравнению с
диастолическим; увеличение ЧСС изменяет работу сердца и повышает
обмен в миокарде, что способствует вазодилатации.
Нервная регуляция
В эксперименте раздражение блуждающих нервов ведет к неболь­
шому уменьшению коронарного кровотока. Коронарные эффекты
блуждающих нервов маскируются их влиянием на сердце — замедле­
нием ритма, что приводит к уменьшению АД. Раздражение симпати­
ческих нервов ведет к сужению коронарных сосудов сопротивления.
Полагают, что влияние раздражения симпатических нервов двух­
фазно. Вначале отмечается вазоконстрикция, которая затем сменяется
мощной вазодилатацией, обусловленной изменением обменных про­
цессов в миокарде. Сильное раздражение симпатических нервов ока­
зывает вазоконстрикторный эффект в коронарных сосудах. Влияние
на венечный кровоток спинальных центров симпатического отдела
ВНС и ядер блуждающих нервов находится под контролем гипотала­
муса и коры большого мозга.
Гуморальная регуляция
Гуморальная регуляция является основной в определении просве­
та коронарных сосудов.
Глава 5
3 24
Влияние ацетилхолина и адреналина. Известно, что ацетилхолин
обладает сосудосуживающим действием на коронарные сосуды.
Влияние адреналина, по мнению ряда авторов, имеет двухфазный
характер — первоначальное уменьшение с последующим увеличени­
ем уровня коронарного кровотока. Показано, что в малых дозах адре­
налин расширяет, а в больших — сужает венечные сосуды. Однако
подавляющее большинство исследователей считают, что адреналин
расширяет сосуды и увеличивает кровоснабжение сердца.
Влияние кислорода. Миокард функционирует только в аэробных
условиях. Венечные сосуды высокочувствительны к газовому соста­
ву крови: даже очень небольшое снижение содержания кислорода в
крови вызывает их расширение. Снижение насыщения крови кисло­
родом в эксперименте со 100 до 10% увеличивало кровоток в обеих
венечных артериях на 300%, а после прекращения гипоксии крово­
ток возвращался к исходному уровню. Одновременно повышалось
АД. Некоторые исследователи полагают, что основной причиной
увеличения венечного кровотока является не возрастание притока
крови в результате повышения давления, а активная вазодилатация
сосудов.
Влияние двуокиси углерода. Вдыхание газовой смеси с 30% содер­
жанием двуокиси углерода приводит к тому, что вначале венечное
давление увеличивается, а затем значительно падает. Считается, что
увеличение тонуса венечных сосудов и, следовательно, уменьше­
ние кровотока в первой фазе обусловлены центральным влиянием
двуокиси углерода, а уменьшение во второй фазе — его местным
действием.
Влияние ионов. В здоровом организме поддерживается строго опре­
деленное соотношение различных ионов (К+, Na+, Са2+ и др.). Как
показали исследования, ионы в растворах КС1, М п04, MgCl2 уве­
личивают сопротивление венечных сосудов и снижают венечный
кровоток, а ионы Н+ и Са2+ увеличивают его.
Продукты метаболизма. Величина коронарного кровотока определя­
ется также метаболическими процессами, в результате которых обра­
зуются такие вещества, как аденозин, брадикинин, простагландины,
молочная кислота, N 0, увеличивающие коронарный кровоток.
Легочное кровообращение
Кровообращение в легком решает две задачи: во-первых, создание
эффективного газообмена между альвеолярным воздухом и кровью,
а во-вторых, обеспечение естественной профилактики возможности
Висцеральные функции
3 25
разрыва сосудов. Последнее обстоятельство связано с тем, что порис­
тая ткань легкого не может служить естественной опорой стенки
сосуда при резком повышении в нем кровяного давления. Именно по
этой причине легочное кровообращение имеет свои особенности:
• давление крови в различных отделах малого круга в 5—7 раз ниже,
чем в сосудах соответствующего калибра большого круга, хотя
правый желудочек выбрасывает крови столько же, сколько левый.
В легочном стволе систолическое давление составляет 25—30 мм
рт. ст., а диастолическое — 5—10 мм рт. ст. Давление в левом пред­
сердии и крупных легочных венах колеблется в пределах 4—8 мм
рт. ст. Таким образом, разность давления, обеспечивающая дви­
жение крови в малом круге, составляет примерно 10—12 мм рт. ст.
Давление в легочных капиллярах составляет примерно 7 мм рт.ст;
• интенсивность кровообращения зависит от фазы дыхания; она
уменьшается на выдохе и увеличивается на вдохе;
• общая площадь контактной поверхности капилляров и альвеол
составляет 150 м2;
• из общего количества крови (500 мл), находящейся в единицу
времени в легком, непосредственно альвеолы омывает не более
100 мл;
• легкие обладают двойным кровообращением. Специфическая
газообменная функция легких обеспечивается малым кругом;
питание всей легочной ткани осуществляется группой артерий
большого круга — бронхиальными артериями, отходящими от
грудной аорты. В тканях легких ветви легочных и бронхиальных
артерий связаны между собой системой бронхопульмональных
анастомозов;
• в малом круге широкие (17 мкм) и короткие капилляры; среднее
время газообмена составляет примерно 0,75 с. При физической
работе и увеличении кровотока эта величина может уменьшаться
до 0,33 с;
• малое сопротивление току крови в легочных капиллярах;
• наличие артериоловенулярных анастомозов (шунтов), которые
способствуют сдерживанию повышения давления в легочном
стволе;
• величина кровотока через малый круг равна минутному объему
сердца и составляет в покое 3,5—5,5 л/м ин, а при физической
работе может достигать 30—40 л/м ин. Следовательно, даже в
покое кровоток на единицу массы легочной ткани в 200 раз выше,
чем средний кровоток в других тканях;
326
Глава 5
• при повышении внутрисосудистого давления увеличивается про­
свет сосудов, по которым течет кровь, и вследствие этого общее
сопротивление кровотоку в легком снижается;
• в вертикальном положении человека величина кровотока умень­
шается по направлению снизу вверх;
• при резком повышении давления в легочном стволе рефлекторно
возникают брадикардия, выход крови в большой круг кровообра­
щения и поступление крови в депо (рефлекс Парина);
• кислород, который вдыхается легкими, используется собственно
легочной тканью в объеме не более 10%.
Таким образом, наряду с высокоэффективным газообменом при
резком увеличении кровотока одновременное снижение сопротивле­
ния сосудов малого круга может предотвратить значительный подъем
давления в легочном стволе. Это естественная приспособительная
реакция организма в условиях возросшего при физической нагрузке
легочного кровотока, предотвращающая чрезмерное повышение дав­
ления в сосудах легкого.
Регуляция легочного кровообращения
Кровообращение в малом круге в значительной степени зависит
от механической (вентиляционной), нервной и метаболической регу­
ляции.
М еханическая(вентиляционная)регуляция
В специальных экспериментах показано увеличение легочного
кровотока в интенсивно вентилируемых участках легочной ткани и
его прекращение в участках, где вентиляция снижена или отсутс­
твует. При форсированном вдохе, когда давление в грудной полости
и полых венах снижается, увеличивается приток крови к сердцу.
В результате этого систолическое давление в правом желудочке и
легочном стволе, а также разность давления между легочным ство­
лом и левым предсердием возрастают, а кровоток через малый круг
увеличивается.
Нервная регуляция
Изменения сердечной деятельности и тонуса сосудов большого
круга вызывают рефлекторные изменения гемодинамики малого
круга. Вегетативная иннервация также оказывает влияние непос­
редственно на тонус легочных сосудов. Так, при возбуждении симпа­
тического отдела нервной системы усиливается деятельность сердца
Висцеральные функции
327
и повышается давление в легочном стволе. Кроме того, раздражение
шейных симпатических узлов и симпатических нервов в эксперимен­
тах на животных приводит к сужению легочных артериол и подъему
давления в легочном стволе.
При острой гипертензии в большом круге кровообращения кровь
вытесняется в малый круг, резко увеличиваются объем крови в
легких и давление в легочных артериях. Известно, что блуждающие
нервы, кроме парасимпатических, содержат симпатические волокна,
направляющиеся к легочным сосудам. С помощью капилляроскопии
установлено, что раздражение холинергических волокон блуждающих
нервов приводит к увеличению кровотока в артериолах и капилля­
рах малого круга, а ваготомия дает обратный результат. Повышение
давления в легочном стволе при раздражении его барорецепторов
вызывает брадикардию, гипотензию и расширение сосудов селезенки
(рефлекс Парина). Рефлексы с рецепторов мелких легочных сосудов
осуществляются через нервные узлы, заложенные в легочной ткани,
или по механизму аксон-рефлекса. Умеренная физическая нагрузка,
сопровождающаяся увеличением минутного объема в 2 раза, вначале
приводит к повышению давления в легочном стволе на 4—6 мм рт.
ст., а затем в связи с падением сопротивления давление падает ниже
нормального уровня. При большой физической нагрузке давление в
легочном стволе повышается, несмотря на снижение сопротивления.
Легочное кровообращение изменяется в широких пределах в зави­
симости от функционального состояния организма и под действием
гормонов, других биологически активных веществ и лекарственных
средств.
Гуморальная регуляция
К гуморальным факторам, влияющим на уровень легочного кро­
вообращения, в первую очередь относят состав альвеолярного воз­
духа. При общей альвеолярной гипоксии, вызванной заболеванием
легких или вдыханием газовой смеси с низким содержанием кисло­
рода, возникает падение р 0 2 во всех альвеолах и происходит одно­
временное сужение всех легочных артериол, вследствие чего давление
в легочном стволе резко повышается. Ацетилхолин расширяет легоч­
ные артериолы, снижает давление, увеличивает кровоток в малом
круге. Гистамин сужает легочные артериолы, вызывая подъем давле­
ния в легочном стволе. Адреналин и норадреналин при неизменном
минутном объеме вызывают повышение давления в легочном стволе
за счет сужения легочных артериол и увеличения их сопротивления.
Глава 5
328
Образовавшаяся в эндотелиях сосудов или вдыхаемая NO приводит
к повышению содержания цГМФ в гладких мышцах сосудов легких,
которые при этом расширяются.
Мозговое кровообращение
Задача мозгового кровообращения заключается в создании тако­
го уровня обменных процессов между капиллярами и окружающей
тканью, который создает в данный момент оптимальный уровень
функционирования структур головного мозга, формирующих теку­
щую деятельность.
Мозг человека составляет около 2% массы его тела, однако он
потребляет до 20% всей энергии, расходуемой организмом в покое.
Интенсивность метаболизма мозга настолько велика, что полное пре­
кращение кровообращения по его сосудам приводит спустя 5—15 с к
потере сознания, а через 5—7 мин — к необратимым изменениям в
коре больших полушарий. Именно поэтому исключительное значение
имеет поддержание устойчивого и достаточного кровообращения.
Особенности мозгового кровообращения
• При нормальной ЧСС в мозговую ткань поступает крови в сред­
нем около 750 мл/мин, или 15% общего сердечного выброса.
• В отличие от других органов (легкие, сердце), в мозге отсутству­
ют анастомозы между артериями и венами, а также «дежурные»
капилляры, т.е. все капилляры постоянно функционируют.
• Во всех мозговых артериях отсутствует пульсация, что достига­
ется постоянным объемом черепной коробки и, соответственно,
постоянным уровнем внутричерепного давления. Артерии мозга
имеют резко выраженную извилистость, что позволяет демпфи­
ровать (сглаживать) исходную пульсацию, наблюдающуюся в
сонных артериях.
• Наибольший кровоток имеет место в сосудах тех отделов головно­
го мозга, которые включены в текущую деятельность.
• Существует связь между внутричерепным давлением и систем­
ным давлением. Так, повышение внутричерепного давления при
опухоли мозга приводит к увеличению давления в большом круге
кровообращения.
• Объем крови в мозге постоянен, поскольку черепная коробка
герметична и сохраняет постоянный объем. Изменение кровос­
набжения отдельных областей мозга достигается перераспределе­
нием крови в сосудах и изменением скорости кровотока.
Висцеральные функции
329
• Венозная система не выполняет емкостных функций. В верти­
кальном положении тела в крупных внутричерепных венах имеет
место отрицательное давление крови. Однако вены при этом не
спадаются из-за их фиксации к костям черепа.
Регуляция мозгового кровообращения
Нервная регуляция
Роль ВНС в регуляции мозгового кровообращения незначитель­
на и распространяется только на крупные сосуды. Раздражение
симпатических нервов вызывает незначительное сужение, а раздра­
жение парасимпатических нервов — слабое расширение просвета
этих сосудов.
Гуморальная регуляция
Влияние кислорода. В состоянии физического и психического покоя
потребление кислорода тканями мозга строго постоянно — 3,5 мл
на 100 г мозговой ткани в 1 мин. Снижение содержания кислорода в
притекающей крови вызывает резкое расширение сосудов мозга за счет
напряжения элементов сосудистой стенки и увеличения кровотока.
Влияние двуокиси углерода и ионов водорода и калия. При повыше­
нии содержания двуокиси углерода в крови, притекающей в мозг,
возрастает мозговой кровоток. Это связано с выделением ионов Н+
вследствие диссоциации образовавшейся угольной кислоты, которые
вызывают расширение сосудов и увеличение кровотока в мозговых
артериях. Сами ионы водорода не могут проникать через гематоэнцефалический барьер. Другим важным фактором, влияющим на моз­
говой кровоток, является аденозин, уровень которого заметно повы­
шается в ответ на ишемию, гипокапнию, гипоксимию и гипотензию.
На уровень мозгового кровообращения влияют также ионы К+.
Функциональная система, поддерживающая
оптимальное для метаболизма артериальное
давление
Кровяное давление — пластичный показатель. Эта особенность
возникла и закрепилась в процессе эволюции как результат постоян­
ного приспособления организма к изменяющимся условиям внешней
среды, когда необходимы регулирование и перераспределение жид­
кости, кислорода, питательных веществ в зависимости от характера
330
Глава 5
реакции организма на внешние воздействия. Так, например, при
интенсивной физической работе значительно возрастает потребление
мышечной тканью различных веществ. Это требует в первую очередь
перераспределения их внутри организма, что достигается изменением
основных гемодинамических показателей и, в первую очередь, кро­
вяного давления. Возвращение этого жизненно важного показателя
к норме и удержание его в определенных физиологических границах
осуществляются функциональной системой (рис. 5.2.7).
Полезный приспособительный результат
Конечным приспособительным результатом, формирующим дан­
ную систему, является оптимальный для метаболизма уровень давле­
ния крови. Измерение АД в различных участках кровеносного русла
показывает, что по мере удаления от сердца к периферии величина его
постепенно снижается. В аорте и крупных артериях человека давление
максимальное — 120 и 80 мм рт. ст., в мелких артериях оно снижается до
85 мм рт. ст., в артериолах происходит дальнейшее его снижение с 75 до
40 мм рт. ст., а в капиллярах давление приближается к 30—10 мм рт. ст.
Рецепция результата
Колебания АД воспринимаются специальными образованиями,
расположенными в стенке сосудов, — барорецепторами. По принципу
реагирования эти рецепторы относятся к механорецепторам, пос­
кольку возбуждение их происходит в результате растяжения арте-
Рис. 5.2.7. Динамика работы функциональной системы поддержания кровя­
ного давления в различных ситуациях: А —в нормальных условиях депрессорные влияния в функциональной системе поддержания кровяного давления,
идущие от сосудистых барорецепторов, сильнее прессорных возбуждений,
вызванных эмоциональными раздражителями; Б —в конфликтной ситуации,
сопровождающейся увеличением кровяного давления депрессорные влия­
ния, идущие от сосудистых барорецепторов, при участии парасимпатической
нервной системы вызывают снижение сердечной деятельности (1), поступле­
ние части крови в депо (2) и расширение просвета сосудов (3), что приводит к
снижению повышенного кровяного давления; В — в условиях исходного сни­
женного давления импульсация от барорецепторов уменьшена и повышается
тонус симпатической нервной системы, что приводит к усилению сердечной
деятельности (1), выходу крови из депо (2) и сужению просвета сосудов (3), что
сопровождается повышением кровяного давления
▼
Висцеральные функции
331
Эмоциональные
раздражители
Депрессорная импульсация
Конфликтная
ситуация
332
Глава 5
риальной стенки при повышении давления. Барорецепторы пред­
ставляют собой нервные окончания остроконечного типа, свободно
заканчивающиеся в адвентиции сосудистой стенки.
Барорецепторы рассеяны по всему кровеносному руслу. Однако в
отдельных участках крупных сосудов они образуют своеобразные скоп­
ления — барорецептивные рефлексогенные зоны, которые обнаружены
в обоих сонных синусах, дуге аорты, мезентериальных сосудах брыжей-'
ки. Барорецепторы имеют различный порог срабатывания. Так, бароре­
цепторы каротидного синуса более чувствительны, чем барорецепторы
дуги аорты. В барорецептивных зонах стенки сосудов, как правило,
более тонки и эластичны, содержат меньшее количество мышечных
элементов, что способствует лучшему восприятию изменений АД.
Барорецепторы в малом круге кровообращения по своей струк­
туре сходны с рецепторами синокаротидных зон и дуги аорты и
концентрируются вблизи бифуркации главных легочных артерий.
Электрофизиологическое и морфологическое исследование показало
наличие рецепторов растяжения в камерах сердца. В предсердиях эти
рецепторы расположены субэндокардиально; в правом предсердии — в
области впадения полых вен, а в левом предсердии — легочных вен.
В предсердиях по характеру импульсной активности различают два
вида рецепторов — рецепторы типа А, в которых максимум импульсаций возникает в момент систолы предсердий, и рецепторы типа Б,
разряд которых приходится на время диастолы, т.е. при заполнении
предсердий кровью (рис. 5.2.8). Для рецепторов желудочков сердца
характерны следующие особенности: выраженная асимметрия (в левом
желудочке их значительно больше, чем в правом) и иное расположение
по слоям, чем в предсердиях (больше всего в эпикарде, меньше в эндо­
карде и очень мало в миокарде). Полагают, что А-рецепторы повышают
тонус симпатического отдела центра кровообращения, что ведет к тахи­
кардии. Именно деятельность этих рецепторов обусловливает рефлекс
Бейнбриджа, который заключается в увеличении ЧСС при увеличении
венозного притока к правому предсердию. Б-рецепторы предсердий
вызывают торможение симпатического и возбуждение парасимпатичес­
кого отдела, что ведет к ослаблению деятельности сердца и снижению
тонуса сосудов. Основная локализация барорецепторов — устье аорты
и легочного ствола; область разделения общей сонной артерии на вне­
шнюю и внутреннюю. Для активности барорецепторов желудочков
характерен короткий разряд, состоящий из одного, реже нескольких
импульсов в ритме сокращений желудочков. Следует отметить, что
небольшое значение в регуляции кровяного давления имеют перифе-
333
Висцеральные функции
Б
Рис. 5.2.8. Характер импульсной активности барорецепторов разных типов
(А, Б) в динамике сердечного цикла (объяснение в тексте)
рические хеморецепторы аортального и каротидного тельца. Например,
уменьшение р 0 2 в артериальной крови через хеморецепторы оказывает
влияние на сосудосуживающую область ствола мозга, что приводит к
усилению тонуса резистивных и емкостных сосудов.
Увеличение р С 0 2 стимулирует центральные хеморецепторы про­
долговатого мозга, что также сопровождается сужением сосудов и
увеличением сосудистого сопротивления.
Классификация барорецепторов
В зависимости от специфики реакций на изменение давления
различают три группы барорецепторов:
• ритмические барорецепторы, воспринимающие ритмические
колебания АД, обусловленные систолой и диастолой. Имеются
барорецепторы, воспринимающие, соответственно, колебания
давления крови от 0 до 20, от 20 до 30 и т.д. вплоть до 240 мм рт. ст.
Отдельные барорецепторы в дуге аорты воспринимают колебания
АД в полосе ниже 80 и выше 120ммрт. ст. В обычных условиях эти
рецепторы не работают и включаются только в случаях резкого
снижения или повышения АД;
334
Глава 5
• статические барорецепторы, реагирующие только на постоянное
давление крови;
• вибрационные барорецепторы, воспринимающие колебания давле­
ния, связанные с вихревыми движениями крови.
Физиологические свойства барорецепторов
Все барорецепторы обладают рядом физиологических свойств,
которые позволяют им выполнять основную функцию — слежение
за величиной АД.
• Каждый барорецептор или каждая группа барорецепторов вос­
принимает только свои определенные параметры изменения АД.
• При быстром перепаде давления барорецепторы отвечают более выра­
женными изменениями залповой активности, чем при медленном
изменении давления (рис. 5.2.9). При резком нарастании давления уже
на небольшой его прирост наблюдается тот же прирост импульсации,
как и при плавном на значительно большие величины.
• Барорецепторы обладают свойством наращивать импульсацию в
геометрической прогрессии на одинаковую величину прироста
АД в зависимости от его исходного уровня. Например, подъем
давления на 10 мм рт. ст. (от 130 до 140) вызывает увеличение час­
тоты импульсации в барорецепторах на 5 имп./с. Однако подъем
давления на те же 10 мм рт. ст., но от 180 до 190 мм рт. ст., увеличи­
вает частоту импульсации уже на 25 имп./с (рис. 5.2.10).
• При воздействии на ритмические барорецепторы постоянного дав­
ления, что наблюдается в случае его стойкого повышения, они
перестают реагировать учащением импульсации, т.е. адаптируются
(рис. 5.2.11). Так, по мере увеличения давления от 0 до 140 мм. рт. ст.
частота импульсации нарастает. Однако в дальнейшем при стой­
ком повышении давления в диапазоне 140—200 мм. рт. ст. частота
импульсации не меняется или даже несколько снижается. Это имеет
приспособительное значение, особенно в тех случаях, когда подъем
АД жизненно необходим, например, при значительных физических
нагрузках. При этом продолжают посылать импульсацию только те
барорецепторы, которые попадают в зону колеблющегося давления
на другом функциональном уровне. Адаптированные барорецепто­
ры снова включаются в функционирование, как только вновь попа­
дают в зону колеблющегося давления.
Таким образом, благодаря свойствам барорецепторов кровенос­
ного русла ЦНС получает постоянную информацию обо всех изме­
нениях АД.
Висцеральные функции
335
Рис. 5.2.9. Изменение импульсации барорецептора при различной ско­
рости нарастания артериального давления в артерии. При повышении
давления на одну и ту же величину в случаях резкого повышения давления
число импульсов в разряде будет больше
Глава 5
3 36
J
Барорецептор
&
Сосуд
ЭКГ
Рис. 5.2.10. Динамика импульсной активности барорецептора при измене­
нии давления (1). Изменение импульсации в барорецепторе дуги аорты (2)
при постепенном увеличении артериального давления. При повышении
давления на одинаковую величину число импульсов, идущих от баро­
рецептора, зависит от исходного уровня кровяного давления в сосуде.
3 — электрокардиограмма
Висцеральные функции
Сигнализация о результате
337
АД, мм рт. ст.
По афферентным буферным
нервам импульсы, возникающие
при возбуждении барорецепторов,
200
распространяются в ЦНС. Центро­
стремительные нервные волокна от
рецепторов, расположенных в дуге
аорты, образуют левый аортальный
нерв. Правый аортальный нерв берет
начало от основания безымянной,
или правой, подключичной артерии.
Оба аортальных (депрессорных) нерва
в составе гортанных нервов идут в
центростремительном направлении
Время, мс
к своим клеточным телам в узлова­
тых ганглиях блуждающих нервов.
Второе афферентное волокно этих
Рис. 5.2.11. А даптация баро­
биполярных нейронов направляется
рецепторов синокаротидной
к продолговатому мозгу. У различных
области при высоком (200 мм.
животных каждый депрессорный нерв
рт. ст.) систолическом давле­
включает от 150 до 600 миелиновых
нии в сонной артерии
нервных волокон диаметром от 6—8
(группа А) до 2—4 мкм (группа В), про­
водящих возбуждение со скоростью
до 10 м/с. Кроме того, в состав этих нервов входят безмиелиновые волок­
на группы С со значительно меньшей скоростью проведения. Нервная
сигнализация от рецептивных зон сонного синуса, расположенных в
месте разветвления общей сонной артерии, распространяется по нервам
Геринга, которые входят в мозг в составе языкоглоточных нервов.
Электрофизиологические исследования показали, что в нормаль­
ных условиях в буферных нервах импульсация носит фазный харак­
тер, совпадающий с ритмом работы сердца. Каждый залп импульсов
соответствует повышению давления во время систолы. При повы­
шении давления, например при механическом пережатии сонной
артерии, импульсация приобретает непрерывный характер.
--
Нервные центры
Информация от барорецепторов поступает к нейронам различных
уровней ЦНС, в первую очередь к структурам продолговатого мозга,
имеющим выход через вегетативные ганглии на сосуды и сердце.
Глава 5
Висцеральные функции
339
В случае повышения давления (рис. 5.2.12) наблюдается увеличение
частоты импульсации от периферических барорецепторов, что при­
водит к торможению клеток, оказывающих активирующее влияние
на спинальные симпатические центры, постоянно поддерживающие
тонус сосудов, а также усиливающие и ускоряющие работу сердца.
Параллельно этому по принципу сопряженности и за счет импульса­
ции от барорецепторов возбуждаются эффекторные парасимпатические
нейроны блуждающих нервов, и деятельность сердца затормаживается.
Одновременно с этим возбуждение клеток сосудорасширяющего отдела
продолговатого мозга вызывает торможение нейронов спинальных сим­
патических центров, что также способствует снижению тонуса сосудов
и увеличению их просвета. Резкое замедление сердечной деятельности и
значительное снижение АД можно наблюдать в эксперименте и при раз­
дражении блуждающего нерва у кролика. Сосудосуживающее действие
симпатических нервов не распространяется на сосуды головного мозга,
сердца, легких и работающих мышц. Сосуды этих органов при возбуж­
дении симпатической нервной системы расширяются. Это связано с тем,
что среди симпатических волокон есть холинергические, выделяющие
не норадреналин, а ацетилхолин. Это обеспечивает опережающее снаб­
жение мышц кислородом и питательными веществами при их быстрой
мобилизации во время движения. Следует отметить, что не все пара­
симпатические нервы являются вазодилататорами, например, волокна
парасимпатического блуждающего нерва сужают сосуды сердца.
В случае падения АД в магистральных сосудах и, соответствен­
но, при уменьшении частоты импульсации от барорецепторов по
принципу сопряженности возбуждается симпатический сердечно­
сосудистый центр ствола мозга, что вызывает усиление и ускорение
работы сердца, уменьшение просвета сосудов и, в конечном резуль-
Рис. 5.2.12. Механизмы нервной регуляции сосудистого тонуса и кровя­
ного давления: А — симпатические сосудодвигательные центры продол­
говатого мозга посылают импульсы к первым спинномозговым нейронам
симпатической нервной системы, активация которых поддерживает н еоб­
ходимый тонус кровеносных сосудов; Б — при повышении давления в
артериях возникает усиленная импульсация от расположенных в их стен­
ках барорецепторов; В — усиленная импульсация приводит к пессимальному торможению нейронов центра, вследствие чего последние перестают
посылать импульсы к спинальным нейронам симпатической нервной
системы, и тонус кровеносных сосудов снижается
340
Глава 5
тате, повышение системного АД до нормального уровня. Состояние
вегетативных центров продолговатого мозга координируется высши­
ми отделами вегетативной саморегуляции, к которым относятся
структуры лимбико-гипоталамо-ретикулярного комплекса.
Исполнительные механизмы функциональной системы
Для обеспечения своего жизненно важного для организма резуль­
тата функциональная система включает внутреннее и поведенческое
звенья саморегуляции.
Внутреннее звено саморегуляции функциональной системы под­
держания кровяного давления осуществляется за счет внутренних,
генетически детерминированных, вегетативных механизмов. В про­
цессах возвращения АД к исходному уровню ведущая роль принадле­
жит изменению работы сердца и просвета сосудов.
Изменение работы сердца. Вся приспособительная деятельность
сердца достигается в основном за счет внесердечной (экстракардиальной) регуляции. Совокупность экстракардиальных факторов,
влияющих на деятельность сердца, условно разделяют на 3 типа в
соответствии с обеспечиваемой ими регуляцией: гемодинамической,
нервной и гуморальной (см. выше).
Изменение просвета сосудов — важный фактор, влияющий на уровень
кровяного давления. Следует отметить, что просвет сосудов связан с
наличием у них тонуса. При этом до 60% общего тонуса сосудов при­
ходится на так называемый базальный тонус, связанный с автоматией
их гладких мышц и растяжением эластической стенки. Кроме того,
на величину базального тонуса влияют высокое р 0 2 артериальной
крови и наличие ионов кальция. Особая роль в управлении работой
сердца и просветом сосудов принадлежит гуморальным влияниям —
различным гормонам и биологически активным веществам. Следует
отметить, что эндотелий сосудов представляет собой сложное образо­
вание площадью до 5000 м2 и массой от 2 до 3 кг. Клетки эндотелия
играют исключительную роль в регуляции сосудистого базального
тонуса. Они выделяют как сосудорасширяющие вещества: оксид азота,
простациклин, эндотелий-зависимый фактор гиперполяризации, так
и сосудосуживающие: эндотелии, супероксид-анион, простаноиды,
ангиотензин II. Такие продукты метаболизма, как углекислый газ и
молочная кислота, также расширяют кровеносные сосуды.
Оксид азота диффундирует в гладкомышечные клетки стенки
сосудов, где активирует связанную с протеином G гуанилатциклазу,
что приводит к увеличению содержания внутриклеточного цГМФ с
Висцеральные функции
341
последующим расслаблением стенки сосуда. Во время сокращения
скелетной мышцы высвобождаются ионы калия и фосфат, вызываю­
щие расширение мелких сосудов.
Простациклин синтезируется в стенке сосуда, стимулируя образо­
вание оксида азота. Он также вызывает расслабление гладких мышц
сосудов за счет активации аденилатциклазы и увеличения образо­
вания цАМФ.
Среди веществ с прессорным действием можно назвать гормон
мозгового вещества надпочечников — адреналин и гормон нейро­
гипофиза — вазопрессин. Адреналин оказывает стимулирующее
влияние на деятельность сердца, усиливая или учащая его работу,
суживает артериолы скелетной мускулатуры при высокой концен­
трации (а-адренергический эффект) и расширяет их при низкой
концентрации (Р-адренергический эффект). Он также сужает сосуды
органов брюшной полости, легких, кожи. Вазопрессин преимущес­
твенно сужает артериолы и капилляры. Мощное прессорное дейс­
твие оказывает ренин-ангиотензиновая система. Так, ангиотензин II
обладает выраженным сосудосуживающим действием и повышает
систолическое и диастолическое давление, а также усиливает секре­
цию глюкокортикоидов и альдостерона путем прямого воздействия
на кору надпочечников. Наряду с сосудосуживающим эффектом
ангиотензин II даже при пороговых значениях усиливает влияние
симпатической нервной системы на сердце и сосуды.
К веществам с депрессорным воздействием относятся ацетилхолин,
образующийся в окончаниях всех парасимпатических нервов и симпа­
тических вазодилататоров, а также гистамин. Оба вещества обладают
местным действием на сосуды. Ацетилхолин расширяет преимущест­
венно просвет мелких артерий, гистамин — капилляров. Реакция сосу­
дов на ацетилхолин связана в основном с образованием оксида азота.
Кинины. Брадикинин, лизил-брадикинин принимают участие в
регуляции АД несколькими путями. Основное действие кининов
состоит в расширении мелких артериальных сосудов и прекапиллярных сфинктеров, увеличении венозного оттока и повышении сброса
крови в венозное русло через артериоловенулярные анастомозы.
Кроме того, увеличивая почечный кровоток и изменяя проницае­
мость сосудов, кинины повышают выведение с мочой ионов К+, Na+,
С1—и воды, что также может приводить к снижению АД.
Простагландины. Это группа ненасыщенных жирных кислот, обнару­
женных в легких, мозге, почках, печени и других тканях. В зависимос­
ти от количества ненасыщенных связей их разделяют на три группы:
г
342
Глава 5
А, Е, F. Простагландины группы А и Е вызывают расширение мелких
артерий путем непосредственного воздействия на сосудистую стенку.
Установлено, что простагландины группы Е находятся в конкурен­
тных взаимоотношениях с некоторыми прессорными гормонами —
вазопрессином и адреналином. Влияние простагландинов на АД про­
исходит за счет изменения деятельности сердца. Увеличение ударного
объема сердца достигается усилением коронарного кровотока и изме­
нением метаболизма миокарда. Кроме того, простагландины играют
роль в регуляции почечного кровотока, в его перераспределении, в
выведении натрия и воды. Так, простагландин Е увеличивает диурез
и экскрецию натрия с мочой без изменения уровня фильтрации.
Натрийуреттеский пептид (НУП). В настоящее время выделяют
мозговой, предсердный и печеночный НУП. Причиной секреции НУП,
вырабатывающегося в миоцитах предсердий, является механическое
растяжение ткани миоцитов. Выделяющийся НУП поступает в кровя­
ное русло, усиливает диурез и приводит к существенному снижению
давления крови. Парасимпатическая нервная система и ацетилхолин
тормозят, а симпатическая — усиливает секрецию НУП. Установлено,
что секреция ПНУП в кардиомиоцитах зависит от содержания ионов
Са2+, цАМФ и цГМФ, инозитолтрифосфатов и протеинкиназы С.
НУП вызывает снижение АД, увеличивает экскрецию ионов и воды,
облегчает поступление жидкости из плазмы крови в интерстициаль­
ную соединительную ткань. НУП тормозит выделение альдостерона,
ангиотензина II, катехоламинов, эндотелина, ренина и вазопрессина.
В действии многих веществ, снижающих тонус сосудистой стенки,
участвует оксид азота (N0), выделяющийся эндотелием сосудов.
Изменение массы циркулирующей крови. В обычных нормальных
условиях около 1/5 всей крови выключено из общей циркуляции
и находится в депо. Наиболее крупными депо в организме явля­
ются селезенка, печень, подкожные сосудистые сплетения, легкие.
В селезенке кровь может выключаться полностью из общей цир­
куляции на довольно длительный период (истинное депо крови)
благодаря наличию синусоидов. Эти капилляры имеют сфинкте­
ры, сужение или расширение которых и обеспечивает наполнение
или опорожнение селезенки. В других депо полного выключения
крови из общего кровотока не происходит. Депонирование в этих
резервуарах достигается за счет сокращения венозных сфинктеров,
уменьшения оттока крови и размещения ее в легко расширяющихся
сосудах. Включение этих эффекторных механизмов происходит по
эфферентным путям симпатической и парасимпатической нервной
Висцеральные функции
343
системы параллельно с включением других исполнительных органов
в общий процесс саморегуляции АД. Емкостные сосуды более
чувствительны к возбуждению симпатической нервной системы,
приводящей к их сужению. В то же время у емкостных сосудов
отсутствуют p-адренорецепторы, и по этой причине они не реаги­
руют на сосудорасширяющие метаболиты.
Процессы кроветворения и кроверазрушения. При длительных изме­
нениях кровяного давления в процессы его регуляции вовлекаются
механизмы образования и разрушения форменных элементов крови,
что способствует стабилизации уровня кровяного давления.
Поведенческое звено саморегуляции функциональной системы про­
является, как правило, только в условиях значительных отклонений
кровяного давления. Результаты поведения направлены в этом слу­
чае на обеспечение нормальных для жизнедеятельности организма
обменных процессов между кровью и тканями.
Динамика работы функциональной системы в разных режимах
В целом организме в естественных условиях происходит постоянное
динамическое взаимодействие двух противоположных тенденций —
прессорных и депрессорных. В случае воздействия какого-либо фак­
тора окружающей среды, когда жизненно необходимым условием для
организма становится повышение АД, активируются все нервные и
гуморальные прессорные механизмы. Устранение этого воздействия
немедленно включает весь комплекс депрессорных механизмов, что
приводит АД к исходному, оптимальному для нормального метаболиз­
ма, уровню. В здоровом организме депрессорные влияния преобладают
над прессорными, т.е. максимальное возвращение всегда сильнее мак­
симального отклонения. По скорости оптимизации уровня кровяного
давления выделяют механизмы быстрого, небыстрого и медленного
реагирования. Быстрое реагирование происходит в течение нескольких
секунд и включает в себя рефлекторные реакции со стороны сердца и
сосудов. Небыстрое реагирование осуществляется в течение минут и
десятков минут и осуществляется транскапиллярным переходом жид­
кости, изменением депонирования крови и уровнем выработки ангиотензина. Медленное реагирование заключается в изменении выделения
воды почками на протяжении нескольких часов или даже дней.
При эмоциональных или физических нагрузках нисходящие
возбуждения из лимбико-ретикулярного комплекса адресуются к
симпатическим сосудосуживающим отделам сосудодвигательного
центра. Благодаря этому усиливаются тонические и симпатические,
Глава 5
344
а затем и гормональные влияния на артериолы и сердце, что при­
водит к повышению АД. После устранения причины гипертензии
наблюдается усиление импульсации от барорецепторов со снижением
тонуса симпатического отдела ВНС и повышением тонуса парасим­
патической, что приводит к торможению сердечной деятельности и
расширению просвета сосудов. Параллельно этому осуществляются
регионарное перераспределение крови, выключение из общей цирку­
ляции некоторого объема крови путем его депонирования (селезенка,
кожа, легкие), изменение вязкости крови, а, соответственно, и пери­
ферического сопротивления. Согласованное взаимодействие всех
исполнительных механизмов приводит к нормализации АД.
В случае падения АД, например при кровопотерях, уменьшение
импульсации от барорецепторов вызывает по принципу сопряжен­
ности повышение тонуса симпатического отдела ВНС, что приводит
к сужению просвета сосудов, усилению сердечной деятельности,
выбросу дополнительных порций крови из депо, увеличению вяз­
кости крови и, соответственно, периферического сопротивления.
В конечном итоге все это способствует повышению АД до вели­
чин, необходимых для оптимального метаболизма. В обоих случаях
наряду с нервной регуляцией имеют место и перечисленные выше
гуморально-гормональные механизмы.
5.3. ДЫХАНИЕ
В процессе дыхания осуществляется газообмен между организмом
и внешней газовой средой. Газовая среда представляет собой смесь
газов, свойства которых моделируются условным, так называемым
идеальным газом.
Идеальный газ подчиняется основному уравнению идеального газа:
PV=MRT,
где Р — давление; V — объем; М — количество газа в молях; R — газо­
вая постоянная; Т — температура в градусах Кельвина.
В реальных условиях все физиологически значимые газы ведут
себя как идеальные. Согласно закону Дальтона общее давление в
смеси газов определяется суммой давления всех компонентов смеси:
Р = Р, + Р2 + Р3 + ....+ Рп.
Висцеральные функции
345
Этапы дыхания
Дыхание представляет собой сложный многоэтапный процесс
доставки кислорода к тканям, окисления органических веществ с
высвобождением энергии и выведения образовавшейся углекислоты
из организма. Дыхательная функция — одна из основных и непос­
редственно обеспечивает само существование живого организма.
Выделяют несколько этапов дыхания.
• Внешнее дыхание — газообмен между атмосферным и альвеоляр­
ным воздухом.
• Газообмен между альвеолярным воздухом и кровью.
• Транспорт газов кровью.
• Газообмен между кровью и клетками тканей организма.
• Внутреннее, или тканевое, дыхание.
Первый этап — внеш нее дыхание
Газовые среды
К газовым средам относятся: атмосферный воздух, выдыхаемый
воздух и альвеолярный воздух (табл. 5.3).
Таблица 5.3. Процентное содержание газов
20,94
16,3
14,5
М
о 2,%
О
Вдыхаемый воздух
Выдыхаемый воздух
Альвеолярный воздух
и
Газ
м 2,%
0,03
4,0
5,5
79,03
79,7
80
Каждый газ в смеси оказывает давление в соответствии с его про­
центным содержанием. В табл. 5.4 приведены величины парциальных
давлений газов в атмосферном и альвеолярном воздухе.
Таблица 5.4. Парциальное давление газов
Среда
р02
рС 02
Атмосферный воздух
760 мм рт. ст.
Альвеолярный воздух
760 —47 (давление водяных
паров) = 713 мм рт. ст.
В тканях
(760 х 21)/100 =
= 159 мм рт. ст.
(713 х 14)/100 =
= 100 мм рт. ст.
(760 х 0,03)/Ю0 =
= 0 мм рт. ст.
(713 х 5,6)/100 =
= 40 мм рт. ст.
40 мм рт. ст.
46 мм рт. ст.
346
Глава 5
Обмен газов между атмосферным и альвеолярным воздухом про­
исходит на основе только физических закономерностей — разности
парциальных давлений газов.
Свойства альвеолярного воздуха.
• Объем и газовый состав альвеолярного воздуха в норме постоянны.
• Газовый состав альвеолярного воздуха отличается от состава
атмосферного.
• Постоянство газового состава альвеолярного воздуха регулирует­
ся содержанием С 0 9.
Следует отметить, что в выдыхаемом воздухе количество углекис­
лого газа меньше, чем в альвеолярном. Это связано с тем, что при
выдохе к альвеолярному воздуху присоединяется воздух мертвого
пространства, содержащий небольшое количество углекислого газа.
Дыхательные объемы
При спокойном вдохе в легкие поступает 500 см3 воздуха — дыха­
тельный объем. Минутный объем дыхания — объем воздуха, проходящий
через легкие за минуту, — в среднем равен 8 л. При форсирован­
ном вдохе в легкие входят еще 1500 см3 воздуха — дополнительный
объем (резервный объем вдоха). При форсированном выдохе выдыха­
ется 1500 см3 воздуха — резервный объем (резервный объем выдоха).
Указанные объемы легочной вентиляции составляют в целом жизнен­
ную емкость легких (рис. 5.3.1). Функциональная остаточная емкость —
объем воздуха в легких после спокойного выдоха. Максимальная
вентиляция легких — объем воздуха, который проходит через легкие
за одну минуту при максимальной частоте и глубине дыхательных
движений, — 150—200 л/мин. Дыхательные объемы измеряются мето­
дом спирометрии с помощью спирометров. Около 1000 см3 воздуха,
содержащегося в легких и воздухоносных путях, — остаточный
воздух — нельзя выдохнуть. Его определяют методом разведения.
Испытуемые после глубокого выдоха в течение 5—10 мин дышат Зл
смеси, содержащей 1 л аргона. После выдоха до 3 л определяют разведение смеси и содержание в ней аргона.
Биомеханика внешнего дыхания
Трахея человека имеет диаметр 15—30 мм и на уровне V грудного
позвонка делится на два бронха. Каждое последовательное деле­
ние бронхиального дерева образует 16 генераций бронхов, которые
относятся к проводящей зоне легкого и создают не участвующее в
газообмене анатомическое мертвое пространство объемом 150—180 мл.
347
Висцеральные функции
If
спирограмма
Рис. 5.3.1. Легочные объемы и емкости: 1 —дыхательный объем; 2 — резерв­
ный объем вдоха; 3 — резервный объем выдоха; 4 — остаточный объем;
5 — резерв вдоха; 6 — функциональная остаточная емкость; 7 — жизненная
емкость легких; 8 — общая емкость легких
Последующие три генерации бронхиол составляю т переходную
зону. Последние четыре генерации (20—23) образуются альвеоляр­
ными ходами и альвеолярными мешочками, которые переходят в
отдельные альвеолы. Вентиляция воздухоносных путей осущест­
вляется при внешнем дыхании конвективным путем, а начиная с
20-й генерации — путем диффузии. Кроме анатомического, в легких
выделяют физиологическое (функциональное) мертвое пространство, к
которому относят объем альвеол, где имеется вентиляция, но нет
перфузии кровью.
Под влиянием нервных импульсов из дыхательного центра наруж­
ные межреберные мышцы сокращаются. При косом расположении
этих Мышц плечо поворота и момент силы у верхнего ребра меньше,
чем .у - нижнего. По этой причине ребра поднимаются, и поперечноег^-т'акже переднезаднее сечения грудной клетки увеличивают­
ся.'Э нергия мышц затрачивается на эластическое сопротивление
грудной клетки, эластическое сопротивление легких, вязкое сопро­
тивление перемещаемых тканей, аэродинамическое сопротивление
Глава 5
348
дыхательных путей, тяжесть перемещаемой грудной клетки и верх­
них конечностей. При сокращении диафрагмы последняя несколько
уплощается, и грудная клетка увеличивается в нижнем направлении.
Это приводит к увеличению объема плевральной полости с последу­
ющим уменьшением давления внутри нее. В результате этого воздух
в легких расширяется, а давление его становится ниже атмосферного.
Вследствие перепада давления между атмосферой и легким воздух
пассивно заполняет альвеолы (рис. 5.3.2). В плевральных полостях
грудной клетки имеется отрицательное давление. У новорожденного
легкие заполняют всю грудную клетку. При этом в легких имеет
место давление слегка ниже атмосферного. В онтогенезе грудная
клетка растет быстрее ткани легких. Однако легкие все время под­
Ребра
Межреберные
мышцы
Диафрагма
р пх Уп = Рлх vn
Рис. 5.3.2. Размеры грудной клетки, объемы легких и потоки воздуха в
состоянии вдоха (А) и выдоха (Б). Во время вдоха увеличение объема плев­
ральной полости ^сопровождается уменьшением плеврального давления Р,
что приводит к снижению давления внутри легкого и поступлению в него
воздуха (А). Во время выдоха имеют место противоположные процессы (Б)
Висцеральные функции
3 49
жимаются к грудной стенке атмосферным воздухом. В результате
даже при выдохе легкие растянуты. Растяжение легких обусловлено
эластической тягой легочной ткани. Разница между атмосферным
давлением и эластической тягой легких создает отрицательное дав­
ление в плевральной полости:
р
атм
- р
эл .тяги
= р
отр.
Величина отрицательного давления в плевральной полости застав­
ляет легкие следовать за движениями грудной клетки (рис. 5.3.3).
Величину отрицательного давления в плевральной полости можно
измерить, если в плевральную полость ввести иглу, связанную с
манометром. При вдохе оно составляет — 9 мм рт. ст., при выдохе —
6 мм рт. ст. Во время вдоха общая скорость воздушного потока, про­
ходящего через дыхательные пути (Q), связана с перепадом давления
(АР) и сопротивлением дыхательных путей ( R) по формуле:
Q = AP/R,
где
.
— Ральвеолярное
^ ДР = Ратмосферное
Следует отметить, что сопротивление воздухоносных путей умень­
шается с увеличением объема легкого при вдохе. Поскольку число
бронхов каждого последующего уровня значительно увеличивается,
то основное сопротивление воздушному потоку создается в верхних
дыхательных путях. Активный контроль сопротивления дыхательных
путей регулируется ВНС: симпатические влияния на гладкие мышцы
бронхов вызывают их расширение, а парасимпатические — сужение.
При вдохе с закрытым носом и ртом (опыт Мюллера) отрицательное
давление в плевральной полости возрастает до — 53—63 мм рт. ст.
В плевральной полости в норме содержится только межплевральная
жидкость и нет воздуха. Под влиянием силы отрицательного давле­
ния в плевральной полости легкие при вдохе пассивно следуют за
несколько опережающим расширением объема грудной клетки. Это
создает динамическую «засасывающую» силу. При введении воздуха
в плевральную полость легкие под влиянием силы пластической тяги
спадаются (пневмоторакс). При каждом вдохе в альвеолы за вычетом
воздуха так называемого вредного пространства — ротовой полости,
трахеи и крупных бронхов — поступает 500 — 170 = 340 см3 воздуха.
Число альвеол в легких человека — приблизительно 300 млн. При
наличии в альвеолах 1000 см3 остаточного и 1500 см3 резервного возду-
Глава 5
350
к
0,25
-
11
вдох
2
13 41
выдох
*
к
Т Ч.
.....1
-3-
-5-
1г
Рис. 5.3.3. Изменение объема легких (V, литры), альвеолярного (Рал) и
плеврального (Р мм. рт. ст.) давления в дыхательном цикле
ха в альвеолах содержится около 3000 см3 воздуха. При каждом вдохе
обменивается около 1/5 альвеолярного воздуха. Растяжению альвеол
легких препятствуют находящиеся в клетках альвеолярного эпителия
сурфактанты, поверхностно-активные вещества, которые снижают
Висцеральные функции
351
поверхностное натяжение альвеол. По своему составу сурфактанты
легких представляют смесь белков и липидов. Сурфактанты стаби­
лизируют состояние альвеол. При вдохе они предохраняют альвеолы
от перерастяжения благодаря тому, что молекулы сурфактантов рас­
полагаются далеко друг от друга, что сопровождается повышением
величины поверхностного натяжения.
При выдохе имеет место та же последовательность перечислен­
ных процессов, но с обратным знаком. Силами, обеспечивающи­
ми спокойный выдох, являются: вес грудной клетки, эластическая
тяга легких, давление органов брюшной полости, эластическая тяга
растянутых во время вдоха хрящей. При активном форсированном
выдохе (физическая нагрузка, выдох под водой) к перечисленным
силам присоединяется сокращение внутренних межреберных мышц
и мышц брюшного пресса. При выдохе сурфактанты предохраняют
альвеолы от спадения: их молекулы располагаются близко друг к
другу, в результате чего величина поверхностного натяжения сни­
жается. Среди факторов эластичности легкого выделяют характер
скрепления альвеол друг с другом. Так, при сжатии альвеол большого
размера наблюдается растяжение связанных с ними альвеол меньше­
го размера. Подобные взаимные влияния обеспечивают поддержание
просвета мелких бронхов, препятствуя их спадению.
Защитные функции дыхательных путей:
• в дыхательных путях холодный воздух согревается на I—2 °С;
• вдыхаемый воздух увлажняется, для чего в сутки через слизистые
выделяется до 500 мл воды;
• поступающий воздух очищается в процессе его турбулентного
движения в носоглотке;
• рефлекторные защитные реакции — кашель, чихание, остановка
дыхания при попадании воды в нос.
Второй этап — газообмен м еж ду альвеолярным воздухом
и кровью
Газообмен в легких обеспечивается: непрерывной вентиляцией
альвеол с целью поддержания постоянства состава альвеолярного
воздуха; диффузией газов через альвеолярно-капиллярный барьер;
перфузией кровью капилляров легких в соответствии с их венти­
ляцией. На рис. 5.3.4 приведены величины парциального давления
и напряжения газов в альвеолярном воздухе, артериальной и веноз­
ной части легочного капилляра, определяющие процессы диффу-
Глава 5
3 52
альвеола
х
\ /
клетки
ткани
v
Рис. 5.3.4. Д иффузия газов между альвеолярным воздухом и кровью
легочного капилляра, а также между кровью тканевого капилляра и клет­
ками ткани на основе разницы парциальных давлений и парциальных
напряжений
зии. Локальный кровоток и локальная вентиляция связаны друг
с другом: в участках с пониженным кровотоком возникает умень­
шение просвета бронхиол с последующим снижением вентиляции,
в участках с пониженной вентиляцией кровоток снижается в резуль­
тате уменьшения просвета сосудов, вызванного гипоксией и гиперкапнией. Вертикальный градиент легочной перфузии более выражен,
чем вертикальный градиент легочной вентиляции. Эта закономер­
ность получила название «вентиляционно-перфузионное отноше­
ние». Суммарная площадь альвеолярно-капиллярного барьера, через
который идет газообмен, составляет 70—80 м2 при толщине от 0,3
до 2,0 мкм. Количество крови в легочных капиллярах составляет
Висцеральные функции
353
100—150 мл при общем ее количестве, одновременно находящемся в
малом круге кровообращения, до 500 мл. Сурфактанты регулируют
скорость абсорбции кислорода на границе раздела «газ—жидкость»
и интенсивность процессов испарения воды с поверхности альвеол.
Газообмен через барьер обусловлен физическими, химическими и
физиологическими факторами.
Физическое растворение газов в жидкой среде
Газ, окружающий жидкость, способен растворяться в ней. Коли­
чество газа, физически растворенного в жидкости, пропорционально
парциальному давлению этого газа над жидкостью, температуре и
объему жидкости (закон Генри). Все это создает напряжение газа в
жидкости. 0 2 и С 0 2 находятся в крови в состоянии простого физи­
ческого растворения. В артериальной крови 0 2 содержится 0,3 об.%
(0,3 см3 на 100 см3 крови), СО, — 3,0об.%.
Химическая связь газов с кровью
И.М. Сеченов с помощью ртутного абсорбциометра показал, что
наряду с простым физическим растворением 0 2 и С 0 2 находятся
в крови в химически связанном состоянии. В артериальной крови
в связанном состоянии 0 2 — 18,7 об.%, С 0 2 — 50 об.%. В венозной
крови 0 2 — 12об.%, С 0 2 — 57 об.%. Обмен газов между кровью и
альвеолами осуществляется на основе диффузии газов благодаря
разности их парциального давления.
Скорость диффузии 0 2 в венозную кровь при разности его парци­
ального давления в 1 мм составляет 25—60 мл/мин; при разности пар­
циального давления в 60 мм — 3600 мл/мин; средняя потребность в 0 2 в
покое — 300 мл/мин. Диффузия С 0 2 из крови в альвеолы в 25 раз выше.
Следует отметить, что на распределение крови в легком значительно
влияет гравитационный фактор. У человека в вертикальном положе­
нии уровень легочного кровотока убывает в направлении сверху вниз.
По этой же причине меньше всего кровоснабжаются верхушки легкого.
Третий этап - транспорт газов кровью
Транспорт кислорода кровью осуществляется двумя путями:
1) кислород переносится в форме простого физического растворения
в крови; 2) в связанной с гемоглобином форме. При соединении кис­
лорода с гемоглобином образуется оксигемоглобин (НЬ + 0 2 -» Н Ь 02).
1 г гемоглобина переносит 1,36 см3 0 2, что составляет кислородную
Глава 5
354
емкость гемоглобина. В 100 мл крови содержится 16—17 г гемоглобина.
Следовательно, 100 мл крови способны перенести 18—20 мл кисло­
рода, а 1 л — 180—200 мл, что составляет кислородную емкость крови.
Процесс соединения гемоглобина с кислородом в легких называется
оксигенацией и протекает без изменения степени окисления железа.
Прочность молекулярных связей гемоглобина с кислородом зависит
от р 0 7 в крови. Эта связь прочна при высоком р 0 2 и ослабляется при
его уменьшении. Кривая диссоциации оксигемоглобина определяет зави­
симость процентного содержания оксигемоглобина от величины р 0 2
(рис. 5.3.5). Кривая демонстрирует, что при парциальном напряжении
кислорода в крови, равном 60 мм рт. ст., кровь практически полно­
стью насыщена кислородом. При парциальном напряжении кислоро­
да в 40 мм рт. ст. (что наблюдается в капиллярах мышц в состоянии
Рп ,мм. рт. СТ.
2
Рис. 5.3.5. Кривая диссоциации оксигемоглобина (1) и ее изменения при
увеличении (2) и уменьшении (3) содержания углекислого газа, темпера­
туры, концентрации ионов водорода и количества 2,3-дифосфоглицерата
эритроцитов
Висцеральные функции
3 55
покоя) кровь содержит 65—70% оксигемоглобина, т.е. она отдает
тканям только 30% кислорода. В капиллярах работающих мышц
при парциальном напряжении кислорода в 20 мм рт. ст. — насыще­
ние гемоглобина кислородом составляет 25%, т.е. 75—80% кислорода
уходит в ткань. Этому способствует выделяющаяся здесь двуокись
углерода, которая сдвигает кривую диссоциации оксигемоглобина
вправо. В работающих мышцах накапливается молочная кислота,
что ведет к диссоциации оксигемоглобина практически полностью.
Образующийся при окислении глюкозы метаболит 2,3-дифосфоглицерат также снижает сродство гемоглобина к кислороду.
В легких кривая диссоциации оксигемоглобина сдвигается влево,
и связь кислорода с гемоглобином становится прочной при низких
значениях его парциального напряжения.
Транспорт углекислоты кровью. Двуокись углерода переносится в
форме простого физического растворения в плазме крови (3 об.%) и
в форме химических соединений: в плазме крови — с бикарбонатами
КНСОэ и N aH C 03(50 об.%), в эритроцитах — с гемоглобином (в форме
карбогемоглобина) (5 об.%) и с бикарбонатом калия — К Н С 03.
В клетках тканей и тканевой жидкости наибольшее р С 0 2. Здесь
оно составляет 60 мм рт. ст. В притекающей артериальной крови
р С 0 2 равно 40 мм рт. ст. Благодаря этому градиенту С 0 2 движется
из тканей в капилляры. В результате р С 0 2 возрастает, и в венозной
крови оно составляет 46—48 мм рт. ст. Под влиянием высокого пар­
циального напряжения часть С 0 2 физически растворяется в плазме
крови. Из плазмы венозной крови С 0 2 легко проникает в эритро­
циты, где соединяется с водой, образуя нестойкую угольную кис­
лоту. Процесс катализируется ферментом карбонгидразой (ускоряет
реакцию примерно в 15 000 раз). Этот фермент в плазме отсутствует.
Таким образом, эритроциты переносят в растворенном виде в 3 раза
больше С 0 2, чем плазма. Оксигемоглобин, соединенный с ионом К+
(К Н Ь02), при высокой концентрации С 0 2 легко отдает 0 2 тканям
(эффект Холдейна), соединяется с угольной кислотой, превращаясь
в гемоглобиновую кислоту (ННЬ). Эффект Холдейна основывается на
механизме, сходном с механизмом эффекта Бора. ННЬ соединяется
с С 0 2, образуя карбогемоглобин, который переносит около 15% С 0 2.
Освободившиеся ионы калия связываются с ионами гидрокарбоната,
образуя бикарбонат калия. Ионы Н С 03~ поступают в плазму крови
и соединяются с ионами Na+, образуя N aH C 03. Выход Н С 03 ком­
пенсируется поступлением в эритроциты ионов С1_. Таким образом,
повышение р 0 2 в легком облегчает освобождение углекислоты из
Глава 5
3 56
Б
а
Эритроцит
Плазма
Клетки
Эритроцит
Плазма
Альвеоп.
Рис. 5 .3 .6. Динамика процессов, происходящих в плазме и эритроцитах
при газообмене в тканях и легких
химически связанного состояния. Поступление кислорода в ткани
способствует увеличению связывания углекислоты кровью.
В легочных капиллярах процесс идет в обратном направлении.
Часть С 0 2 диффундирует в альвеолярный газ. Этому способствует
более низкое, чем в плазме, р С 0 2 в альвеолах, а также усиление
кислотных свойств гемоглобина при его оксигенации. В ходе дегид­
ратации угольной кислоты в эритроцитах под влиянием карбоангидразы оксигемоглобин вытесняет ионы К+ из гидрокарбонатов. Ионы
Н С 03_ поступают из плазмы в эритроциты, а ионы С1_ — в плазму
крови (рис. 5.3.6).
Кривая диссоциации С 0 2 в крови. Содержание С 0 2 в крови
определяется величиной ее парциального напряжения. Зависимость
содержания углекислоты в крови от ее парциального напряжения
описывается кривой содержания углекислоты (рис. 5.3.7).
Этап четвертый — газообмен между кровью и клетками тканей
организма
Р С 0 2 в тканях равно 60 мм рт. ст. Р 0 2 в тканях практически сни­
жается до нуля, чему, как указывалось выше, способствуют кислые
продукты метаболизма — С 0 2 и молочная кислота (см. рис. 5.3.4).
Переходу кислорода в ткани способствует раскрытие дополнитель-
Висцеральные функции
3 57
р С 0 2ММ рт. СТ.
—
— —
Артериальная кровь
Венозная кровь
Рис. 5.3.7. Кривая диссоциации углекислого газа в крови. Линия свя­
зывания углекислого газа, соединяющая точки а (артериальная кровь)
и b (венозная кровь), отражает фактический газообмен
ных капилляров под влиянием продуктов обмена веществ, в частнос­
ти гистамина. Переход кислорода в ткани усиливается при повыше­
нии температуры тканей. Количество кислорода, отдаваемое тканям,
обозначается как коэффициент утилизации кислорода. В покое он
равен 40%, при работе — 60%. Следует отметить, что в мышцах во
время сокращения увеличивается потребность в кислороде, однако
при этом ухудшаются условия его доставки, поскольку сдавливаются
мышечные капилляры. Следует отметить специальный механизм,
обеспечивающий аэробные метаболические процессы в ритмически
сокращающемся миокарде. Это кратковременное депо кислорода в
миоглобине. Сродство миоглобина к кислороду выше, чем у гемогло­
бина. Уже при напряжении миоглобина около 4 мм рт. ст. 50% мио­
глобина переходит в оксимиоглобин, а при напряжении 40 мм рт. ст.
миоглобин насыщен кислородом полностью. При давлении кислоро­
да в легких 100 мм рт. ст. весь гемоглобин и миоглобин насыщаются
кислородом. При снижении р 0 2 в ткани до 10 мм рт. ст. оксигемоглобин отдает до 90% кислорода, а оксимиоглобин удерживает до 80%.
Глава 5
358
Этого запаса кислорода достаточно, чтобы поддерживать окисли­
тельные процессы 3—4 с.
Регуляция дыхания
Дыхательный центр
Участие различных отделов ЦНС было продемонстрировано в экс­
периментах Ламсдена (1924) на животных (рис. 5.3.8). Показано, что
1
2
3
4
5
Рис. 5.3.8. Влияние перерезок в различных участках центральной нервной
системы на характер внешнего дыхания: 1 — верхний край среднего
мозга; 2 — верхний край моста; 3 — верхний край продолговатого мозга;
4 — верхний край спинного мозга; 5 — разрез между нижними шейными
и верхними грудными сегментами спинного мозга. Отделы дыхательно­
го центра: а — корковый; б — пневмотаксический; в — апноэтический;
г — инспираторный и экспираторный; д — шейный моторный; е — спи­
нальный моторный
Висцеральные функции
3 59
дыхательный центр охватывает практически все отделы головного
мозга. Ритмическое дыхание хорошо осуществляется даже у бульбар­
ных животных. Нейроны, контролирующие дыхательные движения,
расположены в nucleus tractus solitarius и nucleus retroambiguus.
При перерезке спинного мозга на уровне верхних шейных сегмен­
тов дыхание прекращается.
При перерезках по верхнему краю продолговатого мозга дыхание
становится ритмическим, но судорожным. В продолговатом мозге нахо­
дятся инспираторный и экспираторный компоненты дыхательного центра.
При перерезках по верхнему краю варолиева моста дыхание рит­
мично, с длинными паузами и задерживается на выдохе. Здесь рас­
полагается так называемый апноэтический дыхательный центр.
При перерезках по верхнему краю среднего мозга дыхание регу­
лярное, плавное. Здесь находится так называемый пневмотаксический
центр, направляющий дыхание.
Гипоталамус и ретикулярная формация определяют участие дыха­
ния в эмоциональных и вегетативных реакциях, а мозжечок приуро­
чивает дыхание к движениям.
Кора головного мозга, особенно ее префронтальные отделы,
регулирует дыхание в соответствии с поведенческой деятельностью
субъектов, при реакциях различной биологической и социальной
значимости. Установлено, что чем выше расположен соответствую­
щий отдел дыхательного центра, тем сложнее проследить его связь с
внешним дыханием. Кроме того, вышерасположенный отдел влияет
на расположенный ниже, обеспечивая его гибкость и приспособляе­
мость к дыхательным запросам организма.
Автоматия дыхательного центра
Нейроны дыхательного центра работают ритмически. Одновремен­
ная регистрация дыхательных движений животного и импульсной
активности нервных клеток дыхательного центра ствола мозга выявила
несколько классов нейронов, которые активируются в разные моменты
актов вдоха и выдоха (рис. 5.3.9). По времени активизации выделяют:
«ранние» инспираторные и экспираторные нейроны, активирующиеся
в начале вдоха и выдоха; «поздние» инспираторные и экспираторные
нейроны — в конце вдоха и выдоха; «полные» инспираторные и экс­
пираторные нейроны — на протяжении акта вдоха и выдоха; «инспираторно-экспираторные» нейроны — в конце вдоха и в начале выдоха;
«экспираторно-инспираторные» нейроны — в конце выдоха и в начале
вдоха, и, наконец, «непрерывные» нейроны, работающие без пауз, но с
Глава 5
360
1
’
IIIIIII
III 1111
3
Mill
lull II
И
11III
1III llllll
1II
II1
1
1
МММ1
И11111 1
4
-
5
1lllllllll
1111II rt
6
llllllllll
1
П 1111111
IIII 1
1111111 N 11II1111
llllllllrtlllllll
II1
7
8
llllll
Mil IIII
1
III ГПMil
9 IIIIIII
IIIIIII
10 1
I I 1 IIII -III1 lr II
II
1
1 1 II
II
11
1 II
A
IIIIIII
llllll 1
1
II II II
1
Б
Р и с. 5.3.9. Активность различных типов нейронов дыхательного цен­
тра продолговатого мозга во время вдоха и выдоха: 1 — пневмограмма;
2 — «ранние» инспираторные нейроны; 3 — «поздние» инспираторные
нейроны; 4 — полные инспираторные нейроны; 5 — «ранние» экспира­
торные нейроны; 6 — «поздние» экспираторные нейроны; 7 — полные
экспираторные нейроны; 8 — «инспираторно-экспираторные» нейроны;
9 — «экспираторно-инспираторные» нейроны; 10 — фоновые нейроны
увеличением частоты разрядов во время вдоха и выдоха. Дыхательные
нейроны связаны между собой прямыми и обратными активационны­
ми и тормозными связями. Следует отметить, что деятельность ней­
ронов дыхательного центра находится под влиянием многочисленных
внешних и внутренних факторов: боль, эмоции, атмосферное давление,
температура, кровяное давление, тонус мышц.
Гуморальные влияния на центр дыхания
Фредерик в опытах с перекрестным кровообращением у двух собак,
когда артериальная кровь из туловища одного животного поступала
I
Висцеральные функции
361
Рис. 5.3.10. Опыт Фредерика с перекрестным кровообращением у собак
в голову другой собаки, а венозная кровь из головы этой собаки — в
туловище первой, показал, что при асфиксии одного животного у вто­
рого наблюдается одышка (рис. 5.3.10). Результаты опыта показывают,
что дыхательный центр возбуждается либо недостатком кислорода,
либо избытком углекислого газа. Ведущим фактором, изменяю­
щим деятельность дыхательного центра, выступает двуокись углеро­
да, которая действует как на периферические, так и на центральные
хеморецепторы. Ионы водорода с трудом проникают через мембраны
клеток, поэтому действующим началом являются ионы водорода,
образующиеся в межклеточной жидкости ствола мозга при прохожде­
нии в нее окиси углерода через гематоэнцефалический барьер.
Рефлекторные влияния на центр дыхания
Нейроны дыхательного центра получают информацию от большо­
го числа механорецепторов дыхательных путей. Различают три типа
механорецепторов:
• медленно адаптирующиеся рецепторы растяжения легких, распо­
ложенные в гладких мышцах трахеи и бронхов и активирующиеся
при вдохе;
3 62
Глава 5
• ирритационные быстроадаптирующиеся рецепторы слизистой
оболочки трахеи и бронхов, активирующиеся при резких измене­
ниях объема легких;
• юкстакапиллярные J-рецепторы интерстиции легких и бронхов
вблизи капилляров, активирующиеся при повышении давления
в малом круге кровообращения.
Вдох начинается с того, что нейроны пула I («ранние» инспираторные нейроны) посылают импульсы к мышцам вдоха и активиру­
ют нейроны пула II («поздние» инспираторные нейроны), которые
посылают импульсы к нейронам пула III. Нейроны пула III тормозят
нейроны пула I. Афферентная импульсация от хеморецепторов пос­
тупает к нейронам пула I, а от механорецепторов легких — к нейро­
нам пула II. Нейроны пула IV (пневмотаксический центр) получают
импульсы от коры и модулируют деятельность всех нейронов про­
долговатого мозга (рис. 5.3.11).
Кроме того, по мере заполнения поступающим воздухом аль­
веол все больше нарастает частота импульсаций в отходящих от
рецепторов растяжения афферентных волокнах блуждающего нерва.
Импульсации блуждающих нервов поступают к возбужденным нако­
пившейся в крови двуокисью углерода инспираторным нейронам.
При достижении частоты импульсаций 80—100 имп./с в одиночном
афферентном волокне блуждающего нерва эта частота становится
пессимальной для инспираторных нейронов, и они переходят в
состояние пессимума. В результате вдох прекращается. Происходит
пассивный выдох — спадается грудная клетка и поднимается купол
диафрагмы.
К этому процессу подключаются и активные механизмы за счет
возбуждения экспираторных нейронов вентральной области про­
долговатого мозга. В специальных экспериментах было показа­
но, что искусственное раздувание легких вызывает рефлекторное
торможение вдоха, а резкое уменьшение объема легких приводит
к активации выдоха. Эти реакции получили название рефлекса
Геринга—Брейера, в котором афферентным звеном являются волок­
на блуждающего нерва. После перерезки блуждающего нерва
дыхание становится глубоким и редким. Исследования показа­
ли, что у людей для осуществления нормального дыхания реф­
лекс Геринга—Брейера не имеет большого значения. Тем не менее,
при усиленном дыхании он ограничивает глубину дыхательных
движений.
Висцеральные функции
3 63
4
Рис. 5.3.11. Нервно-гуморальная регуляция дыхания: 1 — афферентные
волокна блуждающего нерва; 2 — диафрагмальный нерв; 3 — межреберные
нервы; 4 — межреберные мышцы; 5 — диафрагма; 6 — корковые влияния;
7 — спинной мозг; 8 — продолговатый мозг; 9 — мост; МР — механорецеп­
торы легких; ХР — хеморецепторы сосудов
Дыхательный центр и контроль механизма дыхания
В дыхательном акте принципиальное значение имеет вопрос о том,
как организм контролирует дыхательную потребность и как оценивает
ее удовлетворение. Обычно выдох в нормальных условиях происходит
еще задолго до того, как поступивший в легкие воздух и содержащийся
в нем кислород переходят в кровь и поступают к тканям, т.е. в услови­
364
Глава 5
ях, когда на дыхательный центр еще продолжает действовать накопив­
шаяся в результате метаболизма двуокись углерода. В этом процессе
проявляется общий принцип сенсорного насыщения. Афферентация о
поступившем в альвеолы воздухе является гарантией прекращения
акта вдоха, так как дыхательная потребность организма при поступ­
лении воздуха в альвеолы обязательно будет обеспечена. Дыхательный
акт — циклический процесс. По мере израсходования поступившего
при вдохе кислорода и дальнейшего накопления в тканях и крови дву­
окиси углерода начинается следующий вдох: процессы ритмического
дыхания последовательно осуществляются всю жизнь индивида.
В структуре дыхательного центра обнаружен контролирую ­
щий аппарат — акцептор результата действия (В.А. Полянцев,
Ю.И. Фельдшеров). Этот аппарат работает следующим образом.
Как только инспираторные нейроны посылают команды к испол­
нительным органам по коллатералям своих аксонов, «копии команд»
распространяются к специальной группе интернейронов, связанных
между собой циклическими соотношениями. Возбуждение в этих
нейронных кругах продолжает циркулировать до поступления афферентации от рецепторов растяжения легких. Эти нейроны все время
оценивают характер данной афферентации. Если характер поступаю­
щей к ним от рецепторов растяжения бронхиол и альвеол афферента­
ции соответствует ранее сформированной дыхательной потребности,
дыхательный акт успешно завершается и формируется следующий
дыхательный цикл. Если характер поступающей обратной афферента­
ции от альвеол и бронхиол не соответствует дыхательной потребности,
инспираторные нейроны посылают при следующем вдохе измененную
команду, дополнительно направленную на удовлетворение дыхатель­
ной потребности (рис. 5.3.12). Кроме того, к центрам дыхания поступа­
ют тонические влияния от самых разных периферических рецепторов
через ретикулярную формацию. В случаях произвольного сокращения
дыхательной мускулатуры импульсы из моторных отделов коры мозга
поступают к спинальным мотонейронам по пирамидному тракту,
минуя бульбарные центры автоматии дыхания.
Функциональная система, определяющая
оптимальный для метаболизма уровень газов
в организме
Саморегуляция дыхания осуществляется деятельностью спе­
циальной функциональной системы, которая включает несколь-
Висцеральные функции
365
Результаты
действия
Обратная
афференция
Рис. 5 .3 .1 2 . Схематическое изображение стадии афферентного си н те­
за, принятия решения и оценки результата в структуре дыхательного
центра: инспираторный нейрон (А) обрабатывает поступающую к нему
афферентацию от альвеол, дыхательных мышц, болевых и температур­
ных воздействий и высших психических влияний (афферентный синтез).
Это приводит к формированию инспираторной команды. Одновременно
по коллатералям возбуждается группа вставочных интернейронов
(а, б, в), связанны хциклическимивзаимоотнош ениями (акцепторрезуль­
тата действия). К вставочным интернейронам поступает сигнализация
(обратная афферентация) от альвеол и бронхиол, которая сравнивается
и оценивается ими
3 66
Глава 5
ко подсистем саморегуляции со своими приспособительными для
организма результатами. Ведущим, конечным результатом деятель­
ности данной ф ункциональной системы является поддержание
оптимального для метаболизма тканей организма соотнош ения
газов: С 0 2/ 0 2. По конечному приспособительному результату дан­
ная функциональная система называется функциональной систе­
мой, определяющей оптимальный для метаболизма уровень газов в
организме (рис. 5.3.13).
Результат деятельности функциональной системы дыхания
Результат деятельности данной функциональной системы явля­
ется многопараметренным. Он включает р 0 2 и р С 0 2. С последним
показателем тесно связана реакция тканей и крови (pH). Все эти три
показателя тесно взаимосвязаны.
Исходя из принципа мультипараметрического взаимодействия функ­
циональная система дыхания не направлена своей деятельностью на
один конечный показатель, а постоянно определяет динамическое
оптимальное перераспределение газовых показателей р 0 2, р С 0 2 и pH
в организме.
Ф ункциональная система дыхания, как и многие другие функци­
ональные системы гомеостатического уровня, имеет предконечный,
тесно связанный с конечным, результат — уровень р 0 2, р С 0 2 и pH
в крови. Это защитное, предохранительное для организма свойство
функциональной системы дыхания — поддержание показателей р 0 2,
р С 0 2 и pH в крови — надежная гарантия устойчивого метаболизма
в тканях.
Показатели газового гомеостазиса находятся в тесной связи с дру­
гими показателями внутренней среды, в частности с температурой,
уровнем кровяного, осмотического давления и др. Взаимодействие
различных показателей внутренней среды, обеспечиваемое деятель­
ностью различных специфических функциональных систем, также
строится по мультипараметрическому принципу взаимодействия,
определяя тем самым динамическое состояние гомеостазиса в
целом.
Показатели газового гомеостазиса в их мультипараметрическом
взаимосодействии — пример одной из жестких динамических конс­
тант организма. При серьезном смещении этих показателей (напри­
мер, при отсутствии поступления кислорода из окружающей среды —
асфиксии) живой организм не может существовать более 2—3 мин.
Поэтому функциональная система дыхания постоянно и напряженно
Висцеральные функции
367
Поведение в экстремальных условиях
Хемо-рецепторы сосудов
Рис. 5.3.13. Общая схема функциональной системы, определяющей опти­
мальный для метаболизма уровень газов в организме
368
Глава 5
работает по обеспечению оптимального для метаболизма организма
уровня газовых показателей. Ведущим параметром метаболического
процесса, обеспечивающего нормальную жизнедеятельность тканей,
является уровень в крови кислорода. Для нормальной жизнедеятель­
ности тканей у человека в покое средняя потребность в кислороде
составляет около 250—500 мл/мин. Этот показатель существенно
изменяется в процессе жизнедеятельности организма при движени­
ях, тяжелой мышечной работе, гормональных и других изменениях
и составляет в каждом случае динамическую потребность организма
в кислороде. Потребность в кислороде и необходимость удаления
избытков накапливающейся в результате обмена веществ в тканях
двуокиси углерода — ведущие факторы стимуляции рецепторов газо­
вых показателей данной функциональной системы — хеморецепто­
ров. Соотношение р 0 2 и р С 0 2 в тканях, венозной и артериальной
крови и в альвеолярном воздухе динамически изменяется.
Хеморецепторы газовых показателей
Хеморецепторы, включенные в деятельность функциональной
системы дыхания, специфичны. Они реагируют только на изменения
газовых показателей крови и тканей (рис. 5.3.14). Периферические хемо­
рецепторы газовых показателей располагаются в сосудистой стенке
практически любых тканей (В.Н. Черниговский, Б.И. Ткаченко). На
это указывают опыты с перфузией изолированных органов, сохра­
нивших с организмом нервные связи, растворами с повышенным
содержанием двуокиси углерода или со сниженным содержанием
кислорода. В этом случае наблюдается отчетливая дыхательная реак­
ция экспериментальных животных; гиперпноэ в первом случае и
гипопноэ, вплоть до апноэ, во втором. Значительное количество
хеморецепторов газовых показателей крови находится в дуге аорты.
Снижение рО, в артериальной крови ниже 50—60 мм рт. ст. сопро­
вождается увеличением вентиляции легких уже через 3—5 с. Наиболее
изученные хеморецепторы — каротидные тельца в области бифурка­
ции общей сонной артерии на внутреннюю и внешнюю. Каротидные
тельца представлены крупными эпителиоидными клетками I типа и
мелкими интерстициальными клетками II типа. С клетками 1типа
контактируют окончания афферентных волокон синусного нерва
(нерв Геринга), который является веточкой языкоглоточного нерва.
Каротидные тельца обильно васкуляризированы. Наличие в синокаротидной области хеморецепторов газовых показателей снова дока­
зывают эксперименты с перфузией гипоксическими и гиперкапни-
369
Висцеральные функции
35
70
105
140
рО мм рт. ст.
Рис. 5.3.14. Зависимость частоты разрядов сосудистого хеморецептора от
напряжения кислорода (рО,) в артериальной крови (А). Зависимость час­
тоты разрядов сосудистого хеморецептора от напряжения углекислого газа
(р С 02) в артериальной крови (Б)
ческими растворами изолированной и сохранившей с организмом
только нервные связи синокаротидной области.
Особую группу составляют центральные хеморецепторы газовых
показателей. Наличие в ЦНС таких рецепторов доказывает уже опи­
санный классический опыт Фредерика с перекрестным кровообра­
щением. Современные исследования показывают, что центральные
хеморецепторы газовых показателей располагаются у вентральной
поверхности продолговатого мозга на глубине около 0,2 мм. Кроме
того, нейроны, чувствительные к изменению уровня двуокиси угле­
рода в крови, обнаружены в ретикулярной формации продолговатого
и среднего мозга (E.J1. Голубева). Центральные хеморецепторные
3 70
Глава 5
нейроны обильно васкуляризированы. Центральные хеморецепторы
называют хеморецепторными триггерными зонами. Это название
говорит о пусковой роли таких рецепторов в саморегуляции процес­
сов дыхания. Хеморецепторы синокаротидных телец преимуществен­
но реагируют на снижение в крови уровня кислорода. Центральные
хеморецепторы преимущественно реагируют на изменения в спин­
номозговой жидкости pH и р С 0 2.
Общее свойство всех хеморецепторов газового гомеостазиса —
большой прирост импульсаций при более выраженных изменениях
соответствующего параметра газового гомеостазиса. К тому же чувс­
твительность периферических хеморецепторов находится под кон­
тролем BHC: парасимпатические нервы снижают, а симпатические
повышают чувствительность хеморецепторов газовых показателей к
изменению газового гомеостазиса.
Информация о состоянии периферических и центральных рецеп­
торов газовых показателей, возбужденных непосредственно гумо­
ральным путем или опосредованно — нервным, наряду с влия­
ниям и высших отделов дыхательного центра распространяется к
исполнительным механизмам дыхательного центра, в результате чего
в деятельность функциональной системы дыхания включаются ее
внутреннее и внешнее звенья саморегуляции.
Исполнительные механизмы функциональной системы
Для обеспечения своего жизненно важного для организма резуль­
тата функциональная система дыхания включает внутреннее, вне­
шнее и поведенческое звенья саморегуляции.
Внутреннее звено саморегуляции функциональной системы дыха­
ния осуществляется за счет внутренних, генетически детерминиро­
ванных вегетативных механизмов.
Внешнее звено связано с потреблением кислорода из окружаю­
щего воздуха и выделением в окружающую среду двуокиси углерода.
Внешнее звено саморегуляции функциональной системы дыхания
тесно связывает организм с окружающей средой, факторы которой,
в частности газовый состав, можно рассматривать как неотъемлемую
часть живого организма.
Поведенческое звено саморегуляции функциональной системы
дыхания проявляется, как правило, только в экстремальных условиях
жизнедеятельности, связанных с уменьшением содержания кислоро­
да в окружающей среде или накоплением в ней двуокиси углерода.
Результаты поведения направлены в этом случае на обеспечение
Висцеральные функции
371
нормального для жизнедеятельности организма газового состава
окружающего воздуха.
В нормальных условиях существования при содержании в окру­
жающей среде кислорода около 21% и двуокиси углерода 0,03% само­
регуляция дыхания осуществляется относительно пассивно за счет
деятельности внешнего звена; потребления кислорода из достаточно
обеспеченной кислородом окружающей среды и свободного выделе­
ния в нее двуокиси углерода.
Внутреннее звено саморегуляции функциональной системы дыхания
Внутреннее звено саморегуляции функциональной системы дыха­
ния активно включается у человека при длительной произвольной
задержке дыхания или в экспериментальных условиях у животных
при гипоксии или асфиксии.
Поскольку критическая ситуация гипоксии, и особенно асфик­
сии, требует максимальной мобилизации организма, внутреннее
звено саморегуляции функциональной системы дыхания представ­
лено деятельностью многочисленных органов.
Во внутреннее звено саморегуляции функциональной системы
дыхания включается деятельность сердца: изменяются ЧСС и сила
сердцебиений, систолический и минутный объем. Как следствие
изменений сердечной деятельности изменяются скорость крово­
тока и величина кровяного давления. Меняются свойства крови:
количество эритроцитов, гемоглобина, кислородная емкость крови,
сродство гемоглобина к кислороду и т.д. Активируется эритропоэз.
Накапливающаяся в организме при гипоксии и асфиксии двуокись
углерода активно выделяется почками и секретами пищеварительных
желез, а также путем потоотделения. При этом активно включаются
гормональные механизмы.
Этот процесс определяет накопление в организме биологически
активных веществ, прямо влияющих на интенсивность метаболи­
ческих процессов. Опыты показывают, что пережившие глубокую
асфиксию и возвращенные к жизни путем искусственного дыхания
животные приобретают устойчивость даже к газовой эмболии.
С другой стороны, установлено, что адаптация к периодическому дейс­
твию гипоксии обладает антиаритмическим эффектом (Ф.З. Меерсон).
По-видимому, при гипоксии в организме появляются специальные
факторы устойчивости организма. Возможно, с помощью этих факторов
индийские йоги путем специальных гипоксических упражнений при­
обретают способность к длительной остановке дыхания.
3 72
Глава 5
Внешнее звено саморегуляции функциональной системы дыхания
Это звено связано с потреблением кислорода из окружающей
среды и выделением в нее двуокиси углерода. Эти процессы осу­
ществляются с помощью легочного аппарата. Как правило, функци­
ональная система дыхания в нормальных условиях существования
определяет свою деятельность за счет этого звена саморегуляции и
включает внутреннее и поведенческое звенья только в экстремальной
ситуации.
Внешнее звено саморегуляции имеет свой полезный приспособи­
тельный результат — сохранение постоянного состава альвеолярного
воздуха. В поддержании этого полезного для организма показателя
особо важная роль принадлежит двуокиси углерода. Показано, что
только изменение содержания двуокиси углерода в альвеолярном
воздухе приводит к существенным изменениям легочной вентиля­
ции. Изменений легочной вентиляции не наблюдается при умень­
шении в альвеолярном воздухе уровня кислорода, если при этом
сохраняется неизменный уровень двуокиси углерода.
Регуляция поступления 0 2 в организм и выделение С 0 2 осущест­
вляется за счет изменения глубины и частоты дыхания. Этот процесс
определяют и показатели жизненной емкости легких.
Процессы саморегуляции вдоха и выдоха лучше всего прослежи­
ваются на примере первого вдоха новорожденного.
Как известно, плод, получая кислород через плацентарное кро­
вообращение от организма матери и выделяя в кровь матери двуо­
кись углерода, активно не использует внешнее звено саморегуляции
дыхания. Его внешней газовой средой является организм матери, с
которым он сохраняет общий кровоток. После перевязки пуповины
в организме новорожденного, как следствие, происходит снижение
уровня кислорода и накопление двуокиси углерода. Ведущим факто­
ром первого вдоха является накопление двуокиси углерода. Двуокись
углерода, накопившаяся в крови, начинает действовать на перифери­
ческие рецепторы тканей и сосудов. Воспринимаемый центральными
хеморецепторами повышенный уровень двуокиси углерода и сопутс­
твующее ему снижение уровня pH (ацидоз) — причина возбуждения
инспираторных нейронов, расположенных в дорсальном дыхатель­
ном ядре продолговатого мозга, около задвижки. Инспираторные
нейроны дорсального дыхательного ядра связаны своими аксонами
с мотонейронами спинного мозга, иннервирующими диафрагму и
наружные межреберные мышцы.
Висцеральные функции
373
Системная деятельность дыхательного центра
Инспираторные нейроны в их совокупности на разных уровнях
дыхательного центра постоянно получают информацию от хеморе­
цепторов о дыхательной потребности организма. С другой стороны,
как указывалось выше, на них действуют дополнительные факторы,
связанные с состоянием организма: температура, афферентация о
деятельности дыхательных и других мышц, импульсация от альвеол,
эмоциональное состояние — например, боль, высшие психические
влияния, связанные с социальной деятельностью человека (пение,
чтение лекций и др.).
Все это определяет процессы афферентного синтеза в функ­
циональной системе дыхания. В результате афферентного синтеза
инспираторные нейроны дыхательного центра на основе молекуляр­
ных процессов «принимают решение» взять потребное количество
воздуха. Это «решение» в форме нервных импульсаций адресуется
дыхательному мышечному аппарату.
Таким образом, в функциональной системе дыхания на основе
нервных и гуморальных кодов все время оценивается дыхательная
потребность. С помощью аппарата акцептора результата действия она
постоянно сопоставляется с количеством и качеством поступившего
в легкие воздуха. Эти процессы осуществляются бессознательно.
Дыхательный акт автоматизирован. Однако в любых случаях изме­
нения деятельности этого механизма (например, при поступлении
в организм воздуха, по объему не соответствующего дыхательной
потребности) включаются механизмы сознательной деятельности
человека, в частности, поведение.
Поведенческое звено саморегуляции функциональной системы дыхания
Это звено включается, как правило, только в так называемых экс­
тремальных ситуациях, когда организм по каким-либо причинам не
может самостоятельно, за счет внутренних механизмов саморегуля­
ции, длительно оптимально удовлетворять свои дыхательные потреб­
ности. Примером является нахождение большого количества людей в
ограниченном пространстве. По мере того как содержание кислорода
в окружающем воздухе уменьшается, а содержание двуокиси углерода
увеличивается, у находящихся в этих условиях людей автоматически
возникает эмоциональный дискомфорт. В этом случае формируется
мотивация по устранению экстремальной ситуации или избеганию
ее. На основе дыхательной мотивации формируются специальные
374
Глава 5
действия, результат которых обеспечивает адекватное поступление
кислорода и выделение двуокиси углерода из организма.
Компенсация функций в деятельности функциональной
системы дыхания
На примере деятельности функциональной системы дыхания
отчетливо проявляются основные принципы системной компенсации
нарушенных функций, столь важные для клинической практики.
Нарушения могут затрагивать различные стороны деятельности фун­
кциональной системы дыхания. Однако во всех случаях компенсаторные
процессы направляются полезным приспособительным результатом фун­
кциональной системы — значением газовых показателей в организме.
В первую очередь, это могут быть нарушения функций рецепторов
газового гомеостазиса (например, при хирургической замене проте­
зом восходящей дуги аорты). В этом случае компенсаторная функция
в значительной степени ложится на другие хеморецепторные зоны:
синокаротидную и центральную, состояние которых необходимо
оценить еще до операции.
Компенсаторные процессы могут затрагивать и механизмы рас­
пространения афферентации от периферических хеморецепторов в
дыхательный центр (например, при хирургических операциях, когда
перерезаются некоторые афферентные нервы). В этом случае компен­
саторные функции берут на себя другие хеморецепторы. При этом
может усилиться значение прямых гуморальных влияний на дыха­
тельный центр. С другой стороны, при повреждении гуморального
звена саморегуляции компенсаторная роль в значительно большей
степени ложится на нервное звено.
Компенсация может затронуть сами структуры дыхательного цен­
тра, особенно связанные с приуроченностью дыхательной функции к
эмоциональным состояниям, движениям, процессам пения и т.д.
Наиболее подвержены компенсации исполнительные звенья само­
регуляции процессов дыхания. Во внутреннем звене саморегуляции
имеется возможность огромного выбора различных органов, обес­
печивающих жизнедеятельность в условиях гипоксии и асфиксии.
В любом случае при удалении или повреждении того или иного орга­
на необходимо учитывать, на какие другие исполнительные органы
и процессы ляжет дополнительная нагрузка.
Выраженное влияние на деятельность внутреннего звена само­
регуляции оказывают повреждения внешнего звена. Например, при
Висцеральные функции
3 75
удалении одного легкого нагрузка ложится не только на оставше­
еся интактным второе легкое, но и на сердце, связывающие кис­
лород системы крови, деятельность выделительных органов и т.д.
(П.К. Анохин и Е Л . Голубева).
Индивидуальные избирательные перестройки во внутреннем и
внешнем звеньях саморегуляции функциональной системы дыхания
имеют место в экстремальных поведенческих ситуациях (например,
при монотонной работе человека в неудобных позах с мелкими
деталями и т.п.). Все это необходимо учитывать при диагностике
состояния здоровья в условиях работы человека на промышленных
предприятиях, при занятии спортом и т.д. Часто эти индивидуаль­
ные изменения в деятельности разных звеньев функциональной
системы дыхания обусловлены эмоциональным перенапряжением
(стрессом).
Дыхание при пониженном атмосферном давлении
В условиях пониженного атмосферного давления в результате
низкого р 0 2 во вдыхаемом воздухе развивается гипоксия. Гипоксии
бывают экзогенные и эндогенные. Экзогенная гипоксия развива­
ется в результате измененных газовых факторов внешней среды.
Эндогенная гипоксия возникает при различных функциональных и
патологических состояниях организма.
Так, при подъеме на 2,5—5,0 км наступает увеличение вентиля­
ции легких, вызванное стимуляцией каротидных хеморецепторов.
Наблюдаются повышение АД и увеличение ЧСС. Все эти реакции
направлены на усиление снабжения тканей кислородом. Увеличение
вентиляции легких на высоте приводит к снижению р С 0 7 в альвео­
лярном воздухе — гипокапнии — при котором уменьшается стиму­
ляция хеморецепторов, особенно центральных. Это может вызвать
ограничение дыхания.
На высоте 4—5 км может развиться высотная (горная) болезнь,
которая характеризуется слабостью, цианозом, снижением ЧСС и
АД, головной болью, уменьшением глубины дыхания.
На высоте свыше 7 км могут наступить опасные для жизни нару­
шения дыхания, кровообращения и потеря сознания. Особенно
большую опасность представляет быстрое развитие гипоксии, при
которой потеря сознания может наступить внезапно. Нормализация
дыхания на высоте достигается применением газовой смеси чистого
кислорода с 5% двуокисью углерода — карбогена.
3 76
Глава 5
Длительное пребывание в условиях низкого атмосферного давле­
ния (например, жизнь в горной местности) сопровождается акклима­
тизацией к кислородному голоданию, которая характеризуется:
• увеличением количества эритроцитов в крови в результате усиле­
ния эритропоэза;
• увеличением содержания гемоглобина в крови и, следовательно,
повышением кислородной емкости крови;
• увеличением вентиляции легких;
• ускорением диссоциации оксигемоглобина в тканевых капилля­
рах в результате сдвига кривой диссоциации вправо из-за увели­
чения содержания в эритроцитах 2,3-глицерофосфата;
• повышением плотности кровеносных капилляров в тканях, уве­
личением их длины и извилистости;
• повышением устойчивости клеток, особенно нервных, к гипок­
сии;
• гипоксией, вызывающей вазодилатацию сосудов мозга с последу­
ющим увеличением мозгового кровообращения.
Дыхание при повышенном атмосферном давлении
Под повышенным давлением воздуха человеку приходится нахо­
диться во время водолазных и кессонных работ. При погружении под
воду с каждыми 10 м глубины давление воды на поверхность тела
увеличивается на 1 атм, следовательно, на глубине 90 м на человека
действует давление около 10 атм.
При погружении под воду в водолазных костюмах человек может
дышать только воздухом под соответствующим погружению повы­
шенным давлением. Так, на глубине 100 м водолаз дышит газовой
смесью под давлением более 10 атм. В этих условиях увеличивает­
ся количество растворенных в крови кислорода и особенно азота.
Поэтому при погружении на большие глубины для дыхания при­
меняются гелиево-кислородные смеси. Гелий почти нерастворим в
крови. Кислород добавляют к гелию в такой концентрации, чтобы его
парциальное давление на глубине (т.е. при повышенном давлении)
было близким к тому, которое имеется в обычных условиях. Дыхание
чистым кислородом возможно только на глубине до 20 м из-за его
токсического действия, вызывающего мышечные судороги.
После работ на больших глубинах специального внимания требует
переход человека от высокого давления к нормальному. Если водолаз
всего лишь час проработал на глубине 160 м, то обратный путь зани­
мает 3 суток — столько времени он должен провести в барокамере.
Висцеральные функции
377
В настоящее время мировым достижением является погружение
на глубину 500 м с длительностью проживания в глубоководном ком­
плексе 15 сут при давлении 50 атм. Дыхание при этом осуществлялось
гелиевой смесью, в которой кислорода было всего 2%.
При всплытии на поверхность с закрытой голосовой щелью воз­
можны вздутие легких и разрывы альвеол (баротравма).
При быстром подъеме водолаза (быстрая декомпрессия) раство­
ренные в крови и тканях газы не успевают выделиться из организ­
ма и образуют пузырьки. Кислород и углекислый газ представля­
ют меньшую опасность, так как они быстро связываются с кровью
и тканями. Особую опасность представляет образование пузырьков
азота, которые разносятся кровью и закупоривают мелкие сосуды
(газовая эмболия), что сопряжено с большой опасностью для жизни.
Состояние, возникающее при быстрой декомпрессии, называется кес­
сонной болезнью. Она характеризуется болью в мышцах, головокруже­
нием, рвотой, одышкой, потерей сознания, в тяжелых случаях могут
возникать параличи. При появлении признаков кессонной болезни
необходимо немедленно вновь подвергнуть пострадавшего действию
высокого давления (такого, с которого он начинал подъем), чтобы
вызвать растворение пузырьков азота, а затем постепенно производить
декомпрессию. Декомпрессионный синдром может проявиться при
резкой разгерметизации кабины пилота самолета, летящего на высотах
7000 м и более. В этом случае используются кислородные маски.
5.4. СИСТЕМНЫЕ МЕХАНИЗМЫ ГОЛОДА,
АППЕТИТА И НАСЫЩЕНИЯ
Голод и насыщение как эмоции
Каждый человек ежедневно испытывает чувство голода и, после
сильной пищевой эмоции, связанной с приемом пищи, — чувство
насыщения.
Чувство голода — одно из ярких эмоциональных ощущений, с
которым человек появляется на свет. Много забот и беспокойства
доставляют новорожденные уже в самом начале жизни своим мате­
рям, когда сильным криком и движениями стремятся привлечь их
внимание к испытываемому ими чувству голода. Наоборот, после
кормления новорожденные успокаиваются.
Ощущения голода всегда эмоционально неприятны. К ним отно­
сятся неприятное ощущение тяжести в эпигастральной области,
3 78
Глава 5
чувство боли в области желудка («голодные боли»), тошнотное состо­
яние, головная боль, слабость и т.д.
В отличие от этого, прием пищи сопровождается положительными
эмоциональными ощущениями удовольствия или даже наслаждения.
Эмоции голода и насыщения находятся на страже процессов
метаболизма как основных процессов жизни. Эмоциональное ощу­
щение голода — своеобразный сигнал о потребности организма
в пище. Эмоциональный сигнал голода, будучи во всех случаях
неприятным ощущением, позволяет живым существам быстро и
надежно, без детализации, оценивать возникающую потребность
в питательных веществах и на этой основе осуществлять поиск и
потребление пищи. В этом стимулирующем действии эмоции голода
большой биологический смысл. Отрицательная эмоция голода сти­
мулирует субъектов к настойчивому действию по удовлетворению
их ведущей метаболической пищевой потребности.
Эмоциональный сигнал голода позволяет живым существам оце­
нивать потребность в пище без учета количества недостающих бел­
ков, жиров и углеводов и т.д. В этом есть положительные и отрица­
тельные стороны. Эмоциональный сигнал голода без детализации
различных параметров пищевой потребности позволяет быстро оце­
нить эту потребность и быстро организовать пищедобывательную
деятельность. С другой стороны, стимулируемые к приему пищи
общим эмоциональным сигналом голода живые существа, и в том
числе человек, принимают избыточное количество питательных
веществ, которые порой и не требуются для метаболических нужд и
создают только неоправданную дополнительную нагрузку на пище­
варительный аппарат.
Эмоциональное ощущение голода субъективно весьма специфич­
но. Его нельзя спутать с чувством жажды, страха, полового возбуж­
дения и т.д.
Эмоции насыщения выступают в качестве сигнала, прекращаю­
щего прием пищи. Эмоциональное ощущение насыщения позволяет
субъектам довольно быстро оценивать количество и качество пищи
и быстро завершать ее прием.
Основное биологическое назначение эмоций голода и насыщения
заключается, таким образом, в том, чтобы своевременно информиро­
вать организм о возникшей пищевой потребности, быстро построить
необходимое пищедобывательное поведение и быстро осуществить
прием пищи. В этой быстроте оценки пищевой потребности и ее
удовлетворения заложен большой эволюционный приспособитель­
Висцеральные функции
379
ный смысл. Включение эмоционального ощущения между моментом
приема пищи и истинным удовлетворением питательных нужд орга­
низма — пластической и энергетической утилизацией вновь приня­
тых питательных веществ в тканях организма — позволило живым
существам использовать этот довольно значительный интервал вре­
мени на другие формы приспособительного поведения.
5.4.1. Функциональная система питания
Состояние голода, пищевого аппетита, пищедобывательное пове­
дение и пищевое насыщение определяются деятельностью единой
функциональной системы, которая в собирательном смысле может
быть названа функциональной системой питания. Это сложная
многозвеньевая функциональная система, включающая несколько
подсистем. Конечным приспособительным результатом функцио­
нальной системы питания является уровень питательных веществ
в организме, обеспечивающий нормальное течение метаболических
процессов. Этот показатель поддерживается деятельностью как внут­
реннего, так и внешнего звеньев саморегуляции функциональной
системы питания. Внутреннее — вегетативное звено. Внешнее звено
включает формирование пищевой мотивации, пищевой аппетит и
пищедобывательное поведение, направленное на прием потребных
пищевых веществ и их обработку в верхних отделах пищеваритель­
ного тракта. В функциональной системе питания имеется еще одно
звено, деятельность которого направлена на формирование и удале­
ние каловых масс из организма (рис. 5.4.1).
Процессы питания у человека в значительной степени определя­
ются социальными факторами, привычками, воспитанием, нормами
поведения и др. Однако во всех случаях побудительная сила процесса
питания заключается в пищевой потребности и ее удовлетворении.
Пищевая потребность
Пищевая потребность — физиологический, материальный про­
цесс; обусловленное метаболическими реакциями снижение уровня
питательных веществ в организме.
Пищевая потребность — мультипараметрический показатель,
определенное соотношение основных питательных веществ в орга­
низме: белков, жиров и углеводов. Пищевая потребность может
быть избирательной, обусловленной выраженной недостачей одного
какого-либо вещества, либо общей при снижении уровня всех или
нескольких веществ.
380
Глава 5
Прием
пиши
Ротовая
полость
Пищевод
Желудок
Двенадцати­
перстная киш ка
Тонкая
киш ка
Толстая
кишка
Дефекация
Рис. 5.4.1. Общая схема функциональной системы питания с соподчинен­
ными подсистемами: ЛГ —латеральный гипотоламус; ВМГ —вентромедиальный гипоталамус
Как правило, деятельность функциональной системы питания
направлена на поддержание оптимального для метаболизма уровня
питательных веществ в крови. В этом проявляется каскадный при­
способительный перестраховочный принцип обеспечения устойчи­
вости конечного метаболического результата функциональной сис­
темы питания за счет поддержания в крови на оптимальном уровне
предконечного результата, тесно связанного с конечным результатом.
С помощью поддержания предконечного результата на оптимальном
уровне в крови обеспечивается устойчивое течение метаболических
процессов в тканях.
Результат деятельности функциональной системы питания — под­
держание в крови оптимального уровня питательных веществ —
пример пластичной константы организма. Организм человека может
Висцеральные функции
381
нормально существовать при отсутствии приема пищи при свобод­
ном доступе к воде до 20 и даже 30 сут.
Восприятия пищевой потребности
Научные представления о механизмах восприятия организмом
потребности в пищевых веществах существенно изменялись с раз­
витием методических подходов в физиологии.
С открытием голодной периодической моторной деятельности
желудка считали, что ощущение голода, т.е. сигнал о пищевой потреб­
ности определяется периодическими сокращениями пустого желудка
(Карлсон). Все это легло в основу теории «пустого желудка», согласно
которой ведущей причиной ощущения голода является импульсация
от рецепторов желудка после эвакуации из него ранее принятых
веществ. Однако аутоэксперименты, проведенные С.В. Аничковым,
продемонстрировали, что ощущение голода не соответствует при­
ступам моторной деятельности желудка натощак. Ощущение голода
могло возникать в любые моменты голодной моторной периодичес­
кой деятельности желудка, включая периоды покоя.
Теория «пустого желудка» не нашла подтверждения в наблюде­
ниях хирургов, которые обнаружили, что после тотальной резекции
желудка пациенты после пищевой депривации по-прежнему сохра­
няли чувство голода. В экспериментах на собаках было убедительно
продемонстрировано, что после полной деафферентации Ж КТ при
двусторонней перерезке на шее обоих блуждающих нервов и пере­
резке спинного мозга на уровне нижних шейных сегментов, собаки,
несмотря на паралич задних конечностей, после пищевой деприва­
ции сохраняли выраженное стремление к приему пищи.
Как следствие этих экспериментов сложилась сформулированная
И.П. Павловым в 19П г. теория «голодной крови». Согласно теории
«голодной крови» причиной ощущения голода считают кровь с пони­
женным содержанием питательных веществ.
Теория «голодной крови» нашла достаточно убедительное под­
тверждение в следующих экспериментах. Если голодным собакам в
кровь вводили раствор глюкозы, то отдельные животные, у которых
введение глюкозы покрывало потребность организма в углеводах,
прекращали прием пищи. Наоборот, введение инсулина вызывало
у отдельных животных дополнительный прием пищи. Было уста­
новлено, что в возникновении состояния голода первоочередное
значение имеет не абсолютный уровень глюкозы в крови, а артериовенозная разница ее содержания. Показано, что состояние голода
3 82
Глава 5
формируется только тогда, когда артериовенозная разница содержа­
ния глюкозы стремится к нулю. Оказалось, что это состояние обыч­
но совпадает с началом приступа голодной моторной деятельности
пустого желудка.
Теория «голодной крови», несмотря на всю ее привлекательность
и перспективность в плане возможного выделения из крови факторов
голода и насыщения, не получила подтверждения в наблюдениях
на сросшихся близнецах, имеющих различные нервные системы и
общее кровообращение.
Оказалось, что кормление одной из девочек, что, естественно,
приводило к поступлению питательных веществ в кровь другой
девочки, никогда не приводило у нее к развитию насыщения. Другая
девочка продолжала длительное время обнаруживать потребность в
пище, несмотря на то, что ее кровь и ткани, казалось бы, получили
достаточное количество питательных веществ.
Сигнализация «пустого желудка». Значение афферентации, пос­
тупающей в ЦНС от пищеварительных органов, в формировании
состояния голода раскрывают эксперименты с гетерогенными анас­
томозами нервов (П.К. Анохин).
При сшивании центрального конца перерезанного на шее правого
блуждающего нерва с периферическим концом, например срединно­
го нерва, нервные волокна блуждающего нерва, прорастая в средин­
ный нерв, устанавливают функциональный контакт с поперечно­
полосатыми мышцами передней конечности.
При регистрации электромиограммы этих мышц видно, что их
электрическая активность в голодном состоянии животных резко
усиливается по частоте и амплитуде в периоды сокращения пустого
желудка и снижается в период покоя. Из этого следует, что элек­
трическая активность «вагусных» мышц отражает состояние ядра
блуждающего нерва у голодных животных. Характерно, что после
кормления животных электрическая активность «вагусных» мышц
снижается до минимального уровня.
Активность ядра блуждающего нерва в условиях голода и насы­
щения может быть прослежена на примере другого анастомоза: цен­
трального конца блуждающего нерва с нервом барабанной струны
(п. chorda tympani). В этом случае эфферентные волокна блуждающего
нерва, прорастая в ствол нерва барабанной струны, устанавливают
функциональный контакт с клетками подчелюстной слюнной желе­
зы. В результате этого подчелюстная слюнная железа в характере
слюноотделения начинает отражать активность ядра блуждающего
Висцеральные функции
383
нерва. У голодных животных слюноотделение «вагусной» слюнной
железы усиливается в периоды сокращения пустого желудка и ослаб­
ляется в период покоя. После кормления животных слюноотделение
«вагусной» слюнной железы резко снижается.
Таким образом, по характеру активности органов, получивших
в результате анастомозов нервов искусственную иннервацию блуж­
дающего нерва, оказалось возможным проследить состояние ядра
блуждающего нерва у голодных и накормленных животных. В состо­
янии голода при пустом желудке активность ядра блуждающего
нерва высокая и периодически усиливается в периоды моторной
деятельности пустого желудка. После приема пищи активность ядра
блуждающего нерва снижается.
Для выяснения роли Ж КТ в активации ядер блуждающего нерва
у голодных животных в условиях анастомоза правого блуждающего
нерва осуществляется функциональная механическая или химичес­
кая блокада второго (левого) блуждающего нерва, который помеща­
ется в кожную муфту на шее. Таким образом снижается активность
всех искусственно связанных с ядром блуждающего нерва органов.
При этом снижается электрическая активность «вагусной» попереч­
но-полосатой мышцы, а также снижается слюноотделение «вагусной»
слюнной железы.
Все это указывает на то, что повышенная активность ядер блуж­
дающих нервов у голодных животных в значительной степени опре­
деляется афферентными влияниями, поступающими в центры про­
долговатого мозга по интактному блуждающему нерву.
Эксперименты с прямой регистрацией электрической импульсной
активности афферентных волокон блуждающего нерва, отходящих от
желудка, показали, что при пустом желудке в афферентных волокнах
блуждающих нервов на фоне перерезанных эфферентных его воло­
кон регистрируется высокоамплитудная высокочастотная импуль­
сация. При кормлении животных или введении пищи через зонд
непосредственно в желудок импульсация в афферентных волокнах
блуждающих нервов подавляется. Примерно через 10 мин в связи с
началом желудочного сокоотделения происходят фазовые изменения
импульсной активности афферентных волокон блуждающего нерва.
Приведенные эксперименты свидетельствуют о том, что теория
«пустого желудка» не потеряла своего значения. У голодных живот­
ных в ядра блуждающих нервов продолговатого мозга от желудка
все время поступают афферентные сигналы. Причиной этой афферентации, с одной стороны, является тоническое сокращение стенок
384
Глава 5
желудка. В голодном состоянии желудок находится в сокращенном
состоянии и занимает минимальный объем в брюшной полости;
стенки желудка утолщены. При этом раздражаются механорецепто­
ры. Напротив, после приема пищи объем желудка резко увеличивает­
ся, его стенки утончаются. С другой стороны, сигнализация пустого
желудка определяется активностью его хеморецепторов, на которые
в голодном состоянии, в частности, активно действует желудочная
слизь, а возможно, и сам фактор «пустоты» желудка.
Возбуждение ядер блуждающего нерва под влиянием афферен­
тных импульсаций пустого желудка у голодных животных распро­
страняется к нейронам гипоталамической области (И.Г. Акмаеев).
Это, в свою очередь, активирует клетки передней доли гипофиза
за счет нейросекреторных процессов, включающих факторы роста
нервов (инсулиноподобные олигопептиды и др.). При этом усили­
вается активность тропных гормонов гипофиза. Гормоны гипофиза
поступают в кровь и способствуют депонированию питательных
веществ в печени, поперечно-полосатых мышцах, в жировой ткани.
Механизмы депонирования питательных веществ в организме
еще плохо изучены. У тучных людей процессы депонирования пита­
тельных веществ осуществляются значительно интенсивнее, чем
у худощавых, с чем связаны, в частности, избыточная масса тела
тучных людей и повышенное содержание у них депонированных
питательных веществ.
Системная информация о потребности
Теория функциональных систем исходит из того, что в передаче
информации о пищевой потребности в нервные центры участвуют
два фактора — импульсация от пустого желудка и «голодная кровь»,
но в определенной временной последовательности.
Первой во времени возникает афферентная импульсация из опус­
тевшего желудка по мере эвакуации из него ранее принятой пищи.
Эта афферентация активирует нейроны ядер блуждающих нервов в
продолговатом мозге и приводит к формированию голодной перио­
дической деятельности ЖКТ. Одновременно за счет распространения
возбуждения от нейронов ядер блуждающих нервов в продолговатом
мозге к нейронам гипоталамуса и активации тропных гормонов гипо­
физа усиливаются процессы депонирования питательных веществ из
крови. Под влиянием сигнализации из пустого желудка в крови сни­
жается уровень питательных веществ. В результате этого кровь при
наличии достаточного количества питательных веществ в депониро­
Висцеральные функции
385
ванном состоянии приобретает раздражающие свойства «голодной
крови». В «голодной крови» появляются специальные информаци­
онные молекулы — олигопептиды, такие как пентагастрин, окситоцин, грелин, мотилин, нейропептид YV и другие, которые несут
информацию о пищевой потребности к пищевым центрам головного
мозга. Введение этих веществ накормленным животным вызывает у
них пищевое поведение и дополнительный прием пищи.
Сигнализация, идущая от пустого желудка, как правило, не вос­
принимается субъективно. Однако появление факторов «голодной
крови» приводит, в свою очередь, к активации специальных (лим­
бических) мозговых структур, возбуждение которых начинает субъ­
ективно осознаваться в виде чувства голода. В этом случае субъекты
уже откладывают второстепенные дела и отправляются на поиск и
потребление пищи.
Рассмотренные механизмы свидетельствуют о том, что ощущение
голода формируется как предупредительный сигнал организма задол­
го до истинного расходования питательных веществ в организме.
Двойной механизм формирования голодных ощущений дела­
ет понятным, почему после удаления желудка человек продолжает
испытывать ощущение голода. В этом случае под влиянием пищевой
потребности в крови по-прежнему появляются информационные
молекулы голода, ведущие к формированию «голодной крови» и на
ее основе — к голодным ощущениям. С другой стороны, наличие
ощущения голода у одной из сросшихся девочек-близнецов при кор­
млении другой может быть объяснено тем, что при наличии сигна­
лизации от пустого желудка факторы «сытой крови» не оказывают
насыщающего действия на пищевые центры.
Центральные механизмы голода и насыщения
Совокупность нервных элементов, расположенных на разных
уровнях ЦНС и участвующих в различных сторонах деятельности
функциональной системы питания, в собирательном смысле обозна­
чается как «пищевой центр» (И.П. Павлов). «Пищевой центр, — как
писал И.П. Павлов, — дистанция порядочного размера». Здесь будут
рассмотрены только некоторые отделы пищевого центра, располо­
женные в области гипоталамуса и определяющие формирование
состояний голода, аппетита и насыщения.
Как показали специальные эксперименты, афферентная сигнали­
зация от пустого желудка и факторы «голодной крови» адресуются к
нейронам латерального гипоталамуса.
Глава 5
386
«Центр голода» латерального гипоталамуса. Нейроны латераль­
ного гипоталамуса составляют так называемый «центр голода»
(Б.К. Ананд, Дж. Бробек). Электрическое раздражение латеральных
отделов гипоталамуса через вживленные электроды у накормленных,
отказывающихся от еды животных приводит к дополнительному при­
ему пищи. Наоборот, двустороннее разрушение латеральных отделов
гипоталамуса приводит к тому, что голодные животные погибают от
истощения в окружении самой лакомой пищи, не притронувшись
к ней. Все это свидетельствует о том, что нейроны латерального
(рис. 5.4.2) гипоталамуса воспринимают сигналы пищевой потреб-
Раздражение
ВМГ
Рис. 5.4.2. Фронтальный срез структур мозга на уровне гипоталамуса.
Показано, что электрическое раздражение латерального гипоталамуса
(ЛГ) вызывает дополнительный прием пищи (Б). Раздражение вентромедиального гипоталамуса (ВМГ) приводит к отказу от приема пищи (А)
Висцеральные функции
387
ности и трансформируют их в ощущение голода. Разрушение этих
отделов головного мозга приводит к тому, что голодные животные не
ощущают пищевой потребности.
«Центр насыщения» вентромедиального гипоталамуса
По соседству с «центрами голода» латерального гипоталамуса
в вентромедиальных ядрах гипоталамуса располагаются «центры
насыщения». Электрическое раздражение «центра насыщения» через
вживленные электроды у голодных животных останавливает прием
пищи. Двустороннее электролитическое разрушение вентромеди­
альных отделов гипоталамуса приводит к тому, что животные с
трудом завершают прием пищи. У животных наблюдаются гиперфагия и булимия («волчий голод»). Такие животные становятся туч­
ными.
Реципрокность пищевых центров гипоталамуса
Между «центрами голода» латерального гипоталамуса и «центра­
ми насыщения» вентромедиального гипоталамуса существуют реципрокные отношения: при возбуждении «центра голода» латерального
гипоталамуса тормозится «центр насыщения» и наоборот. Эта реци­
прокность не абсолютная. При возбуждении нейронов латерального
гипоталамуса часть нейронов вентромедиального гипоталамуса воз­
буждается по опережающему типу, «ожидая» при этом поступления
соответствующей афферентации от принятой пищи.
Восходящие активирующие сигналы пищевых центров гипоталамуса
Возбуждение нейронов латерального гипоталамуса не ограничи­
вается влияниями на нейроны вентромедиального гипоталамуса.
Нейроны латерального гипоталамуса, благодаря их обширным свя­
зям с нейронами лимбико-ретикулярных структур головного мозга
и посредством этих образований — с корой больших полушарий,
оказывают на них восходящие активирующие влияния. В результате
этих влияний возникает эмоционально окрашенное ощущение голо­
да. На этой основе формируются пищевые мотивации — эмоциональ­
но окрашенные состояния, ведущие к приему пищи.
Пищевой аппетит
Как правило, под аппетитом понимают нерезко выраженный,
ослабленный голод. Это определение не совсем точно. Пищевой
аппетит — избирательное отношение испытывающего слабый голод
388
Глава 5
субъекта к определенному виду пищи. При сильном голоде избира­
тельное отношение к пище устраняется.
Избирательность пищевого аппетита определяется двумя момен­
тами: избирательностью пищевой потребности и опытом индивида.
Избирательность пищевой потребности может заключать в себе пот­
ребность в соленых, сладких и других веществах, чем и определяется
избирательное влечение субъекта именно к этим веществам. Особенно
избирателен, порой даже «капризен», аппетит беременных женщин.
Он определяется не только потребностями метаболизма матери, но и
быстрорастущим плодом. Вот почему аппетит беременной женщины
может быстро изменяться.
Опыт субъекта по удовлетворению пищевых потребностей, опреде­
ляющий избирательное отношение к пище, может быть генетически
детерминированным или индивидуально приобретенным и включать
национальные и семейные привычки к определенному виду пищи.
Как правило, функциональная система питания определяет удов­
летворение пищевой потребности субъектов преимущественно за счет
внешнего звена саморегуляции — приема пищи извне. Однако в опреде­
ленных условиях— при вынужденном или добровольном голодании —
она может функционировать за счет внутреннего звена саморегуля­
ции. Эти процессы определяют эндогенное питание организма.
Э ндогенное питание
Эндогенное питание осуществляется за счет внутреннего звена
саморегуляции функциональной системы, определяющей оптималь­
ный уровень питательных веществ в организме.
Эндогенное питание включает процессы расходования депо пита­
тельных веществ в организме. Как правило, голодающий организм
очень разумно осуществляет самоинъекции питательных веществ.
Каждое очередное поступление питательных веществ из тканей в
кровь приурочено к периоду голодной моторной деятельности желудка
(В.Н. Болдырев). Поступившие из ткани в кровь питательные вещества
нередко переходят снова в пищеварительный тракт и после обработки
ферментами в адекватной форме снова всасываются в кровь.
Второй механизм эндогенного питания — перераспределение пита­
тельных веществ внутри голодающего организма (И.П. Разенков).
При этом питательные вещества из органов, менее значимых для
выживания, например поперечно-полосатых мышц, поступают в
кровь и ЖКТ, а затем идут на построение наиболее значимых для
выживания органов, например сердца и головного мозга.
Висцеральные функции
389
Специальный механизм внутреннего звена саморегуляции фун­
кциональной системы питания при голодании направлен на изме­
нение и, как правило, снижение уровня метаболических процессов
в тканях.
Внутреннее звено саморегуляции функциональной системы пита­
ния включает гормональную регуляцию, осуществляемую гормонами
гипофиза, щитовидной, поджелудочной и других желез внутренней
секреции.
Все перечисленные механизмы внутреннего звена саморегуляции
функциональной системы питания определяют довольно длитель­
ный период относительно нормального функционирования организ­
ма без приема пищи извне — до 20—30 сут. Следует, однако, иметь в
виду, что сроки голодания индивидуальны: от 5—10 до 60 сут.
Возможность изменения при голодании интенсивности мета­
болических процессов в последнее время стала широко использо­
ваться в клинике лечебного голодания. Снижение интенсивности
метаболических процессов в тканях, особенно головного мозга,
оказывает нормализующее влияние на течение ряда невротических
заболеваний, навязчивых состояний, приводит к излечению от
нарушений деятельности сердца, артериальной гипертензии, язвен­
ных поражений Ж КТ и других, так называемых «функциональных»,
нарушений.
Э кзогенное питание
Экзогенное питание осуществляется за счет принимаемой извне
пищи. Как правило, уже через 3—4 ч после предыдущего приема
пищи человек испытывает ощущение голода и под его настойчивым
влиянием ищет и принимает очередную порцию пищи. Следует
иметь в виду, что при этом в организме еще достаточно питательных
веществ на 20—30 сут! Питание человека, таким образом, осущест­
вляется впрок.
Неоправданное питание впрок при наличии гарантированных
условий питания часто является причиной приобретения человеком
избыточной массы тела, ожирения и ведет к нарастанию распростра­
ненности и тяжести сердечно-сосудистых заболеваний и к сокраще­
нию жизни человека.
Внешнее звено саморегуляции функциональной системы питания
включает формирующуюся на основе метаболической пищевой пот­
ребности пищевую мотивацию, избирательный аппетит, пищедобы­
вательное поведение, прием пищи и процессы обработки принятой
390
Глава 5
пищи в пищеварительном тракте. При этом пища выступает в качес­
тве второго полезного приспособительного результата функциональ­
ной системы питания.
Все перечисленные процессы, составляющие внешнее звено само­
регуляции функциональной системы питания, определяют процессы
пищевого насыщения.
В механизме пищевого насыщения следует выделить две фазы:
сенсорное и метаболическое насыщение.
Сенсорное насыщение
Сенсорное (эмоциональное, первичное) насыщение определяет
быстрый процесс насыщения (насыщение за обеденным столом!).
Быстрота насыщения биологически оправданна. Быстрый прием
пищи животными в биологической среде, без ожидания, когда при­
нятая пища поступит в метаболические процессы, дает возможность
животным быстро спрятаться, чтобы не оказаться жертвой хищ­
ников.
Сенсорное насыщение связано с действием принятой пищи
на рецепторы ротовой полости, пищевода и особенно желудка.
Возникающая под воздействием на рецепторы этих органов пищи
афферентная импульсация устремляется, как показали специальные
физиологические опыты, к нейронам вентромедиального гипота­
ламуса. Возбуждение нейронов вентромедиального гипоталамуса в
процессе приема пищи все более затормаживает инициативно воз­
бужденные пищевой потребностью клетки латерального гипотала­
муса. Вследствие этого снижаются их восходящие влияния на другие
отделы головного мозга, исчезает пищевая мотивация и заканчива­
ется прием пищи. Одновременно возбуждение нейронов вентромеди­
ального ядра гипоталамуса путем гормональных, пока еще не совсем
изученных влияний, приводит к опорожнению депо питательных
веществ в организме и поступлению питательных веществ в кровь.
Кровь при этом теряет свои «голодные» свойства и раздражающее
действие на нейроны латерального гипоталамуса. При этом кровь
наводняется гуморальными факторами насыщения. Среди них обна­
ружены олигопептиды насыщения: холецистокинин, соматостатин,
бомбезин, глюкагон, лептин и др.
Из слизистой оболочки двенадцатиперстной кишки выделены оли­
гопептиды энтерина, которые также снижают аппетит (А.М. Уголев).
Механизм опорожнения депо питательных веществ под влиянием
нервной сигнализации от рецепторов верхних отделов пищевари­
Висцеральные функции
391
тельного тракта биологически также оправдан. Поступление при­
нятой пищи в желудок — надежная гарантия того, что принятые
вещества поступят в кровь и впоследствии к тканям.
Механизм сенсорного насыщения, разыгрывающийся на нервной
основе, очень хрупок. Вот почему для хорошего пищеварения вредно
отвлекаться во время приема пищи, читать газету, смотреть телеви­
зионные передачи, принимать алкогольные напитки и прочее. Все
это может привести к приему не соответствующего пищевой пот­
ребности количества пищи и расстройству нормальных процессов
пищеварения.
Механизм сенсорного насыщения заключает в себе некоторые
подходы направленного регулирования питания, особенно у тучных
людей. При частом, но небольшими порциями, потреблении пищи
можно «обмануть» пищевые центры вентромедиального гипоталамуса.
При этом выброс питательных веществ из депонированного состоя­
ния может превысить поступление питательных веществ в организм,
и тем самым могут быть созданы условия для уменьшения количества
питательных веществ, находящихся в депонированном состоянии, и
как следствие этого — снижения массы тела.
Сенсорное насыщение завершается метаболическим (вторичным,
обменным) насыщением.
М етаболическое насыщение
Метаболическое насыщение включает обработку принятой пищи
ферментами пищеварительного тракта, ее всасывание из пищева­
рительного тракта, поступление принятых питательных веществ в
кровь, восполнение нужд метаболизма и израсходованных при сен­
сорном насыщении депо питательных веществ в организме.
На этом цикл работы функциональной системы питания, деятель­
ность которой направлена на удовлетворение пищевой потребности,
заканчивается, с тем чтобы через несколько часов человек, стиму­
лируемый очередными голодными ощущениями, снова, несмотря на
имеющиеся в организме запасы питательных веществ, отправился на
прием очередной порции пищи.
5.5. ПИЩЕВАРЕНИЕ
Пищеварение включает комплекс механических и химических про­
цессов, направленных на обработку пищи, всасывание питательных
3 92
Глава 5
веществ, секрецию специальных ферментов в ротовой полости, желуд­
ке и кишечнике, выделение непереваренных компонентов пищи.
Внутриклеточное и пристеночное пищеварение
В зависимости от локализации процесса пищеварения его делят
на внутриклеточное и внеклеточное. Внутриклеточное пищеварение —
гидролиз пищевых веществ, которые попадают в энтероцит в резуль­
тате фагоцитоза и пиноцитоза. В организме человека внутрикле­
точное пищеварение может иметь место в лейкоцитах и в клетках
лимфо-ретикуло-гистиоцитарной системы.
Внеклеточное пищеварение подразделяют на дистантное (полостное)
и контактное (пристеночное, мембранное).
Дистантное (полостное) пищеварение осуществляется на значи­
тельном расстоянии от места образования ферментов. Ферменты в
составе пищеварительных секретов осуществляют гидролиз пищ е­
вых веществ в полостях ЖКТ.
Контактное (пристеночное, мембранное) пищеварение осущест­
вляется ферментами, фиксированными на клеточной мембране
(А.М. Уголев). Структуры, на которых фиксированы ферменты, пред­
ставлены в тонком отделе кишечника гликокаликсом. Первоначально
гидролиз пищевых веществ начинается в просвете тонкой киш ки под
влиянием ферментов поджелудочной железы. Затем образовавшиеся
олигомеры гидролизуются в зоне гликокаликса адсорбированными
здесь ферментами поджелудочной железы. Непосредственно у мем­
бран клеток кишечника гидролиз образовавшихся димеров произ­
водят фиксированные на ней собственно кишечные ферменты. Эти
ферменты синтезируются в энтероцитах и переносятся на мембраны
их микроворсинок.
Принципы регуляции процессов пищеварения
Деятельность пищеварительной системы регулируется нервными
и гуморальными механизмами. Нервная регуляция пищеваритель­
ных функций осуществляется симпатическими и парасимпатичес­
кими влияниями.
Секреция пищеварительных желез осуществляется условно-рефлекторно и безусловно-рефлекторно. Такие влияния особенно выра­
жены в верхней части пищеварительного тракта. По мере продви­
жения к дистальным отделам пищеварительного тракта участие
рефлекторных механизмов в регуляции пищеварительных функций
уменьшается. При этом повышается значение гуморальных меха­
Висцеральные функции
393
низмов. В тонком и толстом отделах кишечника особенно велика
роль локальных механизмов регуляции — местное механическое и
химическое раздражение повышает активность кишечника в месте
действия раздражителя. Таким образом, в пищеварительном тракте
существует градиент распределения нервных, гуморальных и мест­
ных регуляторных механизмов.
Местные механические и химические раздражители влияют на
ф ункции пищеварительного тракта посредством периферических
рефлексов и через гормоны пищеварительного тракта. Химическими
стимуляторами нервных окончаний в Ж КТ являю тся кислоты,
щелочи и продукты гидролиза пищевых веществ. Поступая в кровь,
эти вещества приносятся ее током к пищеварительным железам и
возбуждают их непосредственно или опосредованно. Объем крови,
поступающей в желудок, киш ечник, печень, поджелудочную железу
и селезенку, составляет около 30% ударного объема сердца.
Значительная роль в гуморальной регуляции деятельности орга­
нов пищеварения принадлежит гастроинтестинальным гормонам,
образующимся в эндокринных клетках слизистой оболочки желудка,
двенадцатиперстной киш ки, тощей киш ки, в поджелудочной железе.
Они оказывают влияние на моторику пищеварительного тракта, на
секрецию воды, электролитов и ферментов, на всасывание воды,
электролитов и питательных веществ, на функциональную актив­
ность эндокринных клеток ЖКТ. Кроме того, гастроинтестинальные
гормоны оказывают влияние на обмен веществ, эндокринные и сердечно-сосудистые функции, на ЦНС. Некоторые гастроинтестиналь­
ные пептиды обнаружены в различных структурах мозга.
По характеру влияний регуляторные механизмы можно раз­
делить на пусковые и корригирующие. Последние обеспечивают
приспособление объема и состава пищеварительных соков к коли­
честву и качеству пищевого содержимого желудка и киш ечника
(Г.Ф. Коротько).
Методы исследования функций
пищеварительного тракта
Острые (вивисекционные) методы. Имеют недостатки: применение
наркоза, неестественное раздражение нервов и т.д.
Хронические методы. В 1842 г. русский хирург В.А. Басов впер­
вые осуществил у собаки операцию наложения хронической фисту­
лы желудка, что позволило исследовать желудочную секрецию при
Глава 5
394
естественном кормлении животных. Однако желудочный сок, полу­
ченный при этом, содержит много примесей перевариваемой пищи.
Методы получения чистых пищеварительных соков
Выведение протоков пищеварительных желез наружу.
Операция эзофаготомии (И.П. Павлов и E.O. Шумова-Симановская).
В сочетании с фистулой желудка позволяет получать чистый желудоч­
ный сок.
Операция изолированных желудочков. По Клименсиевичу—
Гейденгайну — перерезается иннервация изолированного желудочка.
По И.П. Павлову — изолированный желудочек сохраняет с большим
желудочком нервные связи и адекватно отражает его работу («зерка­
ло» большого желудка).
Клинические методы
• Зондирование после пробного завтрака.
• Радиозондирование с помощью радиопилюль.
• Рентгеноскопия, рентгенография, рентгенокимография.
• Гастроскопия.
Для исследования слюны у человека используют специальные
капсулы-присоски.
Пищеварение в полости рта
Поступившая в рот пища раздражает рецепторы ротовой полос­
ти. Импульсы от вкусовых рецепторов по афферентным волокнам
тройничного, лицевого и языкоглоточного нервов поступают в соот­
ветствующие центры продолговатого мозга и другие центры ЦНС.
Несмотря на кратковременность пребывания пищи в полости рта
(в среднем 15 с), ее рецепторы оказывают значительные пусковые
влияния почти на весь пищеварительный аппарат.
В полости рта происходит механическая обработка пищи при
кусании и жевании, а также ее химическая обработка за счет секрета
слюнных желез, главными из которых являются пары околоушных,
подъязычных и подчелюстных.
Секреция слюны
После приема пищи слюноотделение начинается через латентный
период 1—3 с. Это связано с прохождением возбуждения от чувстви­
тельных рецепторов ротовой полости по афферентным волокнам .
язычных, языкоглоточных и блуждающих нервов в слюноотдели­
Висцеральные функции
3 95
тельный центр продолговатого мозга и по эфферентным волокнам
симпатических (из верхнего шейного узла) и парасимпатических
(п. chorda tympany) нервов к слюнным железам. У человека наблю­
дается постоянное слюноотделение, даже ночью. В течение суток
выделяется 0,5—2 л слюны, из них около 30% приходится на долю
околоушной железы. 95% всей слюны выделяются околоушными и
подчелюстной железами. Остальные 5% — подъязычной и железами
слизистой оболочки рта.
Каждая слю нная железа выделяет своеобразный секрет.
Околоушная — белковый, подъязычные и небные железы — слизистый.
Слюна на 99% состоит из воды. Плотный остаток (0,5—1,5%)
составляют аминокислоты, креатинин, аммиак, мочевая кислота,
мочевина. Слюнные железы являются также и органами выделения,
содержание в слюне мочевины, мочевой киелоты, аммиака, креатинина существенно повышается при нарушении функции почек.
Слюна содержит катионы натрия, калия, кальция, магния и мик­
роэлементы (железо, медь, никель и др.), а также анионы хлоридов,
бикарбонатов, фосфатов, сульфатов, иодидов, бромидов, фторидов.
Органические вещества представлены в основном белками — гло­
булинами. В слюне имеются и другие белки, в том числе слизистое
вещество муцин, а также некоторые регуляторные пептиды (паротин, фактор роста нервов, инсулиноподобный пептид), ферменты.
Паротин регулирует синтез белков, уровень кальция в крови (подоб­
но кальцитонину), усиливает сперматогенез, гемопоэз, повышает
проницаемость гистогематических барьеров.
В слюне обнаружено свыше 50 ферментов, которые относятся к
гидролазам, оксиредуктазам, трансферазам, липазам, изомеразам.
В небольших количествах в слюне обнаружены протеазы, пепти­
дазы, кислая и щелочная фосфатазы. В слюне содержится фермент
калликреин, который принимает участие в образовании кининов,
расширяющих кровеносные сосуды.
Основные ферменты слюны: а-ам илаза (птиалин) расщепляет
крахмал до мальтозы, а мальтаза расщепляет мальтозу до глюкозы.
На разные пищевые и отвергаемые вещества выделяется слюна
разного состава.
Показано, что механические, температурные и химические свойс­
тва пищи вызывают различную импульсацию и различную скорость
ее возникновения в язычном нерве (И.И. Лаптев). Сначала формиру­
ется импульсация на раздражения механорецепторов, затем терморе­
цепторов и, наконец, на раздражение хеморецепторов.
396
Глава 5
Достаточное количество выделяемой слюны при воздействии на
заднюю треть языка, а также твердое и мягкое небо определяет про­
цесс глотания. «Сухой комок застревает в горле».
На слюнные железы оказывают влияние гормоны, в частности
гипофиза, надпочечников, щитовидной и поджелудочной желез, а
также половые гормоны. В слюне, в свою очередь, отражается содер­
жание гормонов и олигопептидов в крови. Слюнные железы активно
включаются в пищевую условно-рефлекторную деятельность орга­
низма (И.П. Павлов).
Теории слюноотделения
Еще К. Людвиг показал, что слюноотделение — не простая филь­
трация жидкости, солей и биологически активных веществ из крови,
а сложный физиологический процесс.
Согласно гистологической теории разные клетки слюнных желез
выделяют слюну разного состава в зависимости от их различного
строения. Однако оказалось, что слюну разного состава выделяют
одни и те же клетки.
JI. Гейденгайн полагал, что различный состав слюны определяется ра з­
личным влиянием нервов. Парасимпатическая нервная система, по его
мнению, оказывает секреторные влияния и стимулирует выделение
серозной слюны. Симпатическая нервная система, по Гейденгайну,
оказывает трофическое влияние. Симпатические нервы сами по себе
не стимулируют слюноотделение, но на фоне парасимпатических
влияний стимулируют отделение слизистой слюны.
Отдельные авторы (В.П. Бабкин) считают, что характер слюноот­
деления определяется изменением кровообращения слюнных желез.
В частности, парасимпатические влияния расширяют сосуды слюн­
ных желез.
У. Кэннон и А. Розенблюд связывали различное слюноотделение
с различными медиаторами, выделяющимися в нейросекреторных
синапсах.
М еханизм слюноотделения
Возбуждение от рецепторов полости рта передается в ЦНС по
афферентным волокнам тройничного, лицевого, языкоглоточного и
блуждающего нервов. Импульсы достигают продолговатого мозга,
других его отделов, включая кору большого мозга. Основной центр
слюноотделения расположен в продолговатом мозге, сюда и в боко­
вые рога верхних грудных сегментов спинного мозга поступают
Висцеральные функции
397
импульсы из расположенных выше отделов мозга. Слюнные железы
иннервируются эфферентными парасимпатическими и симпатичес­
кими нервными волокнами.
Парасимпатическая иннервация слюнных желез. Парасимпатичес­
кая иннервация подчелюстной и подъязычной слюнных желез начина­
ется от верхнего слюноотделительного ядра продолговатого мозга.
Волокна преганглионарных нейронов в составе барабанной струны
доходят до ганглиев соответствующих желез. Здесь они переключают­
ся на постганглионарные нейроны и по их аксонам достигают гландулоцитов. Иннервирующие околоушные железы преганглионарные
волокна берут начало из нижнего слюноотделительного ядра продол­
говатого мозга, проходят в составе языкоглоточного нерва до ушного
узла. Здесь расположены вторые нейроны, по их аксонам в составе
височно-ушного нерва импульсы достигают слюнных желез.
Под влиянием ацетилхолина, высвобождаемого окончаниями
постганглионарных нейронов, происходят стимуляция М1-холинорецепторов и образование инозитолтрифосфата, что вызывает выде­
ление большого количества жидкой слюны с высокой концентрацией
электролитов и низкой концентрацией муцина.
Симпатическая иннервация слюнных ж елез осуществляется из
боковых рогов II—IV грудных сегментов спинного мозга, откуда
волокна преганглионарных нейронов следуют в верхний шейный
ганглий, где образуют контакты с постганглионарными нейронами.
Их аксоны достигают слюнных желез.
Норадреналин, высвобождаемый окончаниями постганглионар­
ных нейронов, стимулирует (32-адренорецепторы, увеличивает содер­
жание цАМФ, что способствует выделению небольшого объема гус­
той слюны, увеличению образования в железах ферментов и муцина.
Одновременное раздражение парасимпатических нервов усиливает
секреторный эффект. У человека симпатические нервы в наибольшей
мере повышают секрецию подчелюстных слюнных желез.
Торможение слюноотделения
Слюноотделение относится к числу легко тормозимых процес­
сов. Торможение слюноотделения вызывают болевые раздражения,
отрицательные эмоции, умственное напряжение, сон, дегидратация
организма.
Существуют отклонения от нормы: гипосалия, или сиалопения —
уменьшение выделения слюны (например, при лихорадке), или про­
тивоположное явление — сиалорея, или птиализм (наблюдается при
Глава 5
398
отравлении, например, солями ртути, мышьяка и является способом
удаления этих веществ). Обильное выделение слюны отмечается при
асфиксии, когда резкое повышение содержания углекислого газа в
крови повышает активность центра слюноотделения.
Глотание и моторная фукция пищевода
Глотательный цикл длится в среднем 1 с. Координированными
сокращениями мышц языка и щек пищевой комок, сформированный
в ротовой полости, перемещается к корню языка, что приводит к
раздражению рецепторов мягкого неба, корня языка и задней стенки
глотки. Возбуждение от этих рецепторов по языкоглоточным нервам
поступает в центр глотания, расположенный в продолговатом мозге,
от которого в составе тройничных, подъязычных, языкоглоточных и
блуждающих нервов идут эфферентные импульсы к мышцам стенок
полости рта, гортани, глотки и пищевода. Сокращение мышц, при­
поднимающих мягкое небо, обеспечивает закрытие входа в полость
носа, а поднятие гортани закрывает вход в дыхательные пути. Во
время акта глотания происходят сокращения пищевода, которые
имеют характер волн, возникающих в верхней части и распростра­
няющихся в сторону желудка.
Верхняя треть пищевода содержит поперечно-полосатые мышеч­
ные волокна, остальная часть — гладкую мускулатуру. Ниж ний
пищеводный сфинктер открывается в начале глотания, благодаря
рефлексу, который обеспечивается возбуждением вагуса, ВИП- и
NO-синтезирующим нейронам. В остальное время нижний сфинктер
остается закрытым, что препятствует агрессивному действию соля­
ной кислоты на слизистую оболочку пищевода.
Моторика пищевода регулируется в основном эфферентными
волокнами блуждающего и симпатического нервов и интрамураль­
ными нервными образованиями пищевода. Центр глотания распо­
ложен рядом с центром дыхания продолговатого мозга и находится с
ним в реципрокных отношениях.
Выделяют три фазы глотания: ротовая (произвольная), глоточная
(быстрая, короткая, непроизвольная), пищеводная (медленная, дли­
тельная непроизвольная). Две первые фазы глотания длятся около
1 с, продолжительность третьей фазы при глотании твердой пищи
8—9 с, жидкой — 1—2 с.
Сокращения пищевода имеют характер волны, возникающей в
верхней его трети. Такой тип сокращения называется перистальти­
ческим. Перед ним движется волна пониженного тонуса (релаксаци­
I
Висцеральные функции
399
онная волна). Скорость ее движения несколько больше, чем волны
сокращения. Перистальтика пищевода обеспечивает глотание и без
содействия ему силы тяжести (например, при горизонтальном поло­
жении тела и вниз головой, в условиях невесомости у космонавтов).
Пищеварение в желудке
Пищеварительные функции желудка — депонирование пищи,
механическая и химическая обработка и постепенная (порционная)
эвакуация содержимого в кишечник. В желудке пища набухает, раз­
жижается, ее компоненты растворяются и подвергаются гидролизу
ферментами слюны и желудочного сока. Карбогидразы слюны дейс­
твуют на углеводы пищи в центральной части содержимого желуд­
ка, куда еще не проник желудочный сок, прекращающий действие
карбогидраз. Ферменты кислого желудочного сока действуют на
белки пищи только в той части пищевого содержимого, которая
непосредственно контактирует со слизистой оболочкой желудка, и в
небольшом удалении от нее, и только в том случае, если желудочный
сок не был нейтрализован буферными свойствами пищи. Ш ирина
этой зоны зависит от количества и свойств как желудочного сока,
так и принятой пищи, т.е. вся масса пищи в желудке одновремен­
но не смешивается с соком, а по мере разжижения и химической
обработки пищи слой пищевой кашицы, прилегающий к слизистой
оболочке, движениями желудка перемещается в антральную часть
и затем эвакуируется в кишечник. Таким образом, пищеварение в
полости желудка осуществляется в некоторой степени за счет слюны,
но основное значение имеет секреторная и моторная деятельность
самого желудка.
Секреторная деятельность желудка
Желудочный сок
Желудочный сок вырабатывается железами желудка, расположен­
ными в его слизистой оболочке. В области свода желудка железы
содержат главные гландулоциты (главные клетки), которые проду­
цируют пепсиногены; париетальные гландулоциты (обкладочные
клетки), синтезирующие и выделяющие соляную кислоту; мукоциты
(добавочные клетки), выделяющие мукоидный секрет. Сок фундального отдела желудка содержит пепсины и много соляной кислоты.
Сок этого отдела желудка имеет ведущее значение в желудочном
пищеварении. Сок пилорического отдела содержит мало ферментов,
400
Глава 5
много слизи и мало соляной кислоты. За сутки у человека выделяется
2—4 л желудочного сока.
В состав желудочного сока входят органические вещества: пеп­
син, гастриксин, ренин, лизоцим, муцин, мукоиды, аминокислоты,
мочевина, мочевая кислота; неорганические вещества — соляная
кислота, хлориды, сульфаты, фосфаты, бикарбонаты, натрий, калий,
кальций, магний. Желудочный сок имеет кислую реакцию, его pH
равен 0,8 во время пика секреции, проглоченная пища увеличивает
pH до 1,8—4,0.
Главный ферментативный процесс в желудке заключается в
начальном расщеплении белков до альбумоз и пептонов. Основными
ферментами, которые гидролизуют белки, являю тся пепсины.
Главные гландулоциты желудочных желез синтезируют и выделяют
пепсиногены двух групп. Пепсиногены I группы (их 5) образуются в
своде желудка, а II группы (их 2) — в пилорической части желудка.
Пепсиногены I и II групп активируются соляной кислотой, и таким
образом образуется несколько пепсинов. Из пепсиногенов I группы
образуются пепсины, которые гидролизуют белки с максимальной
скоростью при pH 1,5—2,0. Из пепсиногенов II группы образуется
другой протеолитический фермент, близкий к пепсинам — гастрик­
син, гидролизующий белки при pH 3,2—3,5. Возможность пепсинов
активно функционировать при различных значениях pH обеспечи­
вает гидролиз белков в различных слоях химуса при разной кислот­
ности. Еще один фермент желудочного сока — ренин (химозин) —
в присутствии солей кальция створаживает молоко.
В желудочном соке содержится фермент липаза, расщепляющий
жиры до глицерина и жирных кислот. Она малоактивна и гидроли­
зует только эмульгированные жиры. Гидролиз углеводов в желудке
осуществляется в толще пищевого комка под влиянием ферментов
слюны.
Важной составной частью желудочного сока являются мукоиды
(желудочная слизь), которые покрывают слизистую оболочку желуд­
ка по всей поверхности и предохраняют ее от механических пов­
реждений и от самопереваривания. Вместе со слизью продуцируется
анион Н С 03_, в комплексе с которым и создается мукозо-бикарбонатный барьер, защищающий слизистую оболочку желудка от дейс­
твия соляной кислоты и ферментов. К числу мукоидов относится
гастромукопротеид (внутренний фактор Касла), который необходим
для всасывания витамина В)2 (внешний фактор Кастла), при взаимо­
действии с которым образуется антианемический фактор крови.
Висцеральные функции
401
Пепсиногены и липазы продуцируются главными клетками желуд­
ка, а соляная кислота и внутренний фактор Кастла — обкладочными
клетками слизистой оболочки желудка.
Ф ункции соляной кислоты:
• участвует в антибактериальном действии желудочного сока;
• вызывает денатурацию и набухание белков, что способствует их
последующему расщеплению пепсинами;
• активирует пепсиногены;
• создает кислую среду, которая необходима для действия пепсинов;
• участвует в регуляции моторной деятельности пищеварительного
тракта.
Образование соляной кислоты
Источником энергии для активного транспорта протонов из
обкладочных клеток служит АТФ, а катализирует этот процесс
(Н+-К+)-АТФ-аза. Этот фермент, локализованный в мембранах секре­
торных микроворсинок, осуществляет эквивалентный обмен ионов
Н+ на ионы К+. Ионы Н+ образуются в эквивалентных с Н С 03- коли­
чествах при диссоциации Н2С 0 3 и в очень небольшом количестве —
при диссоциации Н20. Ионы Н С 03“ перемещаются по градиенту
концентрации в кровь в обмен на ионы С1_. Когда этот процесс
достигает максимума, концентрация Н С 03_ в сыворотке заметно
повышается — явление, получившее название «щелочного периода».
Одновременно с ионами Н+ в просвет желудка против градиента кон­
центрации и электрического градиента путем активного транспорта
выводятся ионы С1_ (рис. 5.5.1).
Фазы желудочной секреции
Нервные, гуморальные факторы и паракринные механизмы тонко
регулируют секрецию желез желудка, обеспечивая определенное
количество сока, кислото- и ферментовыделение в зависимости от
количества и качества принятой пищи. Происходящие при этом
процессы делят на три фазы: сложнорефлекторную, гуморальную и
кишечную.
Сложнорефлекторная фаза
Отделение желудочного сока происходит при действии пищи
на внешние зрительные, обонятельные и слуховые рецепторы, что
может быть связано с обстановкой приема пищи (условно-рефлекторные раздражители).
Глава 5
402
Париетальная
Париетальная
клетка слизистой
желудка
НСО
Париетальная
клетка слизистой
желудка
¥
Ф-
■
Висцеральные функции
403
Ведущим фактором желудочного сокоотделения является также
раздражение пищей рецепторов полости рта и глотки (безусловно­
рефлекторные влияния).
Так как роль пускового влияния при данной фазе желудочной
секреции принадлежит нервным импульсам, ее называют сложно­
рефлекторной, или мозговой.
Механизмы пищеварения были в значительной степени раскры­
ты в исследованиях И.П. Павлова, удостоенного за это в 1904 г.
Нобелевской премии.
Доказательством первой фазы желудочной секреции служат опыты
на собаках с перерезанным пищеводом (эзофаготомия), проведенные
И.П. Павловым и Е.О. Шумовой-Симановской. При кормлении эзофаготомированных собак пища выпадает из пищевода и не попадает
в желудок, однако через 5—10 мин после начала мнимого кормления
начинает выделяться желудочный сок. Пусковые явления, обусловлен­
ные приемом пищи, стимулируют выделение секретов с высокой фер­
ментативной активностью, обеспечиваемой накопленными в железах
в период их относительного покоя ферментами (по И.П. Павлову —
«запальный сок»).
Рефлекторные нервные влияния передаются на желудок через
блуждающие нервы. После их перерезки ни мнимое кормление, ни
запах, ни вид пищи не вызывает секреции желудка.
В возбуждении желудочных желез в первой фазе желудочной сек­
реции принимает участие гастрин. После удаления пилорического
отдела желудка, где продуцируется гастрин, секреция угнетается.
Желудочный сок, выделяемый при действии пищи на внешние
рецепторы, назван И.П. Павловым «аппетитным».
Желудочная фаза
Усиление желудочной секреции во второй фазе вызывается дейс­
твием пищи на рецепторы желудка, а также нейрогуморальным
путем. Эта фаза определяется раздражающим влиянием пищи на
Рис. 5.5.1. Процессы транспорта ионов, участвующих в образовании соля­
ной кислоты в париетальных клетках желудка: А — процесс транспорта и
секреции ионов хлора; Б — процессы образования, транспорта и секреции
ионов водорода; В — образование соляной кислоты в желудке путем взаи­
модействия ионов хлора и водорода; КА — карбоангидраза
404
СО с
Глава 5
главные клетки желудка. Сокоотделение отмечается при механи­
ческом раздражении желудка. Механическое раздражение желудка,
его пилорической части, приводит к высвобождению из G -клеток
гастрина. Гастрин, поступая в кровь, действует на железы желудка
и вызывает желудочную секрецию. Стимулирующую роль в реали­
зации желудочной фазы секреции играет и гистамин, который пос­
тупает в кровь и действует на обкладочные клетки, продуцирующие
соляную кислоту.
И.П. Разенков показал, что кровь, взятая у накормленной собаки
на пике пищеварения и введенная в кровяное русло накормленной
собаки, вызывает у нее выделение желудочного сока. Именно поэто­
му вторую фазу называют также гуморальной.
Стимулируют желудочную секрецию гастрин, ацетилхолин, про­
дукты переваривания белков, экстрактивные вещества мяса и ово­
щей, секретин (выделяясь в слизистой оболочке кишечника, тормозит
секрецию соляной кислоты, но усиливает выделение пепсиногенов),
холштистокинин-панкреозимин (тормозит секрецию соляной кислоты
и усиливает секрецию пепсиногенов), бомбезин и другие вещества.
Тормозят желудочную секрецию продукты гидролиза жира, гормо­
ны (гастрон и энтерогастрон, ВИП, серотонин и другие вещества).
Кишечная фаза
Возбуждающие и тормозные влияния с двенадцатиперстной и
тощей киш ки на железы желудка осуществляются нервными и гумо­
ральными путями. Нервные влияния передаются с механо- и хемо­
рецепторов кишечника.
Под влиянием экстрактивных веществ в клетках двенадцатиперс­
тной кишки образуются энтерогастрин и энтерогастрон. Первый из
них стимулирует, а второй — тормозит желудочную секрецию.
Стимуляция желудочной секреции осуществляется за счет продук­
тов гидролиза питательных веществ, особенно белков. Эти вещества
могут возбуждать железы желудка через влияние гастрина и гистами­
на, а также непосредственно действовать на желудочные железы.
Торможение желудочной секреции осуществляется рядом веществ,
которые по убывающей силе тормозного действия располагаются в
следующем порядке: продукты гидролиза жира, полипептиды, ами­
нокислоты, продукты гидролиза крахмала, ионы водорода (pH ниже
3 оказывает сильное тормозное действие).
Характер желудочной секреции в ответ на разные виды пищи был
изучен И.П. Павловым (рис. 5.5.2). Оказалось, что наиболее интен-
Висцеральные функции
405
I —Сложнорефлекторная (мозговая) фаза
II —Желудочная фаза
III —Кишечная фаза
Рис. 5.5.2. Фазовые показатели желудочной секреции из изолированного,
по И.П. Павлову, желудочка у собаки при кормлении разными видами
пищи: А — белковая пища; Б — углеводная пища; В — жирная пища
сивным возбудителем желудочной секреции является белковая пища,
на которую секреция отвечает тремя фазами.
На углеводную пищу (хлеб) желудок отвечает секрецией меньшей
интенсивности, но со значительной продолжительностью (до 10 ч).
На хлеб отсутствует желудочная фаза секреции, так как в хлебе мало
химических возбудителей секреции (экстрактивных веществ, про­
дуктов гидролиза и др.).
Жиры молока тормозят секрецию желудка, поэтому на графике
слабо выражена 1 фаза секреции, максимальная секреторная реакция
наблюдается к концу 3-го часа в желудочной фазе.
Высвобождение в двенадцатиперстной кишке секретина и холецистокинина под влиянием содержимого тормозит секрецию соляной кис­
лоты, но усиливает секрецию пепсиногенов. В обеспечении торможения
желудочной секреции участвуют и другие кишечные гормоны из группы
406
Глава 5
гастронов и глюкагон, а также серотонин. Рефлекторное высвобожде­
ние АДГ задней доли гипофиза тормозит секрецию желудка.
Регуляция желудочной секреции
Регуляция секреции обкладочных клеток
Непосредственная стимуляция обкладочных клеток и секреция
ими соляной кислоты осуществляются холинергическими волокнами
блуждающих нервов, медиатор которых — ацетилхолин — возбуждает
МЗ-холинорецепторы базолатеральных мембран гландулоцитов.
Эффекты ацетилхолина и его аналогов блокируются атропином.
Велика роль в эффектах ацетилхолина ионов кальция. Непрямая сти­
муляция обкладочных клеток блуждающими нервами опосредуется
также гастрином и гистамином.
Гастрин высвобождается из G -клеток, основная часть которых
находится в слизистой оболочке пилорической части желудка. После
хирургического удаления пилорической части желудка желудоч­
ная секреция резко снижается. Высвобождение гастрина усилива­
ется импульсами блуждающего нерва, а также местным механи­
ческим и химическим раздражением пилорической части желудка.
Химическими стимуляторами G -клеток являются продукты перева­
ривания белков — пептиды и некоторые аминокислоты, экстрактив­
ные вещества мяса и овощей. Если pH в антральной части желудка
понижается до 3,0 и меньше, что связано с повышением секреции
соляной кислоты железами желудка, то высвобождение гастрина
уменьшается. При pH 1,0 выделение гастрина прекращается, и сек­
реция желудочных желез резко понижается. Таким образом, гастрин
принимает участие в саморегуляции желудочной секреции в зависи­
мости от величины pH антрального содержимого.
Носителем физиологической активности гастрина является
С-терминальный тетрапептид. Он же входит в состав синтетического
аналога гастрина — пентагастрина, получившего применение в кли­
нической практике.
К стимуляторам обкладочных клеток желудочных желез относит­
ся гистамин, образующийся в тучных клетках слизистой оболочки
желудка. Высвобождение гистамина обеспечивается гастрином и
рядом других факторов. Гистамин стимулирует в основном парие­
тальные гландулоциты, влияя на Н2-рецепторы их мембран и вызы­
вая выделение большого количества сока высокой кислотности, но
бедного пепсином.
Висцеральные функции
407
Желудочную секрецию возбуждают также всосавшиеся в кровь
продукты переваривания белков: экстрактивные вещества мяса и
овощей; секретин, образующийся в слизистой оболочке киш ечни­
ка (тормозит секрецию соляной кислоты, но усиливает секрецию
пепсиногенов); холецистокинин-панкреозимин (усиливает секрецию
пепсиногенов, тормозит секрецию соляной кислоты); бомбезин и
другие вещества.
Торможение секреции соляной кислоты вызывают секретин,
холецистокинин, глюкагон, желудочный ингибирующий петид,
ВИП, нейротензин, соматостатин, тиролиберин, энтерогастрон,
вазопрессин, кальцитонин, окситоцин, простагландин Е2, бульбогастрон, кологастрон, серотонин. Высвобождение некоторых из
них в соответствующих эндокринных клетках слизистой оболочки
киш ечника определяется свойствами кишечного химуса. В част­
ности, торможение желудочной секреции жирной пищей в большей
мере обусловлено влиянием на железы желудка холецистокинина.
Повышенная кислотность дуоденального содержимого нервным
путем и через дуоденальные гормоны тормозит выделение соляной
кислоты железами желудка.
Регуляция секреции главных клеток
Стимуляторами главных клеток и секреции ими пепсиногена
являются холинергические волокна блуждающих нервов, гастрин,
гистамин, симпатические волокна, оканчивающиеся на р-адренорецепторах, секретин. Усиление секреции пепсиногена главными
клетками желудочных желез осуществляется посредством несколь­
ких механизмов. Среди них — увеличение переноса ионов кальция в
клетку и стимуляция Na+-K+-ATO-a3bi; усиление внутриклеточного
перемещения гранул зимогена; активация мембранной фосфорилазы, что усиливает экструзию через апикальные мембраны; активация
системы цГМФ и цАМФ.
Гистамин и гастрин влияют на секрецию пепсиногена опосре­
дованно — повышая секрецию соляной кислоты. Снижение pH
содержимого желудка через местный холинергический механизм
усиливает секрецию главных клеток. Описано и прямое стимулирую­
щее влияние гастрина на главные клетки. В высоких дозах гистамин
тормозит их секрецию. Холецистокинин, секретин и адреномиметики непосредственно стимулируют секрецию главных клеток, но
тормозят секрецию обкладочных. Тормозят секрецию пепсиногена
соматостатин, кинины , простагландины.
408
Глава 5
Защита слизистой оболочки желудка от агрессивного действия
желудочного сока обеспечивается слоем слизи и секрецией гид­
рокарбонатных ионов. Простагландины PGE2 и PGI2 усиливают
секрецию клетками слизистой оболочки желудка гидрокарбонатных
ионов. Противовоспалительные препараты, блокирующие циклооксигеназу 1, уменьшают продукцию простагландинов и таким образом
способствуют язвообразованию.
Моторная функция желудка
После приема пищи тонические влияния блуждающих нервов на
желудок снижаются. Желудочная стенка расслабляется.
В желудке наблюдается два основных вида движений — перис­
тальтические и тонические. Перистальтические движения, способс­
твующие перемещению пищи, осуществляются за счет сокращения
циркулярных мышц желудка. Эти движения начинаются на большой
кривизне в участке, примыкающем к пищеводу, где находится кар­
диальный водитель ритма. Наряду с сокращением циркуляторных
мышц перистальтическая волна формируется сокращением про­
дольных мышц перед перемещаемой порцией химуса. Большая часть
перистальтических волн гасится в пилорическом отделе желудка.
Некоторые из них распространяются по пилорическому отделу с
увеличивающейся амплитудой за счет активности второго водителя
ритма, локализованного в пилорическом отделе желудка, что приво­
дит к выраженным перистальтическим сокращениям этого отдела,
повышению давления и переходу части содержимого желудка в две­
надцатиперстную кишку.
Тонические сокращения желудка возникают за счет изменения
тонуса мышц, что приводит к уменьшению объема желудка и повы­
шению давления в нем. Тонические сокращения способствуют пере­
мешиванию содержимого желудка и пропитыванию его желудочным
соком, что значительно облегчает ферментативное переваривание
пищевой кашицы.
Регуляция моторики желудка
Регуляция моторной деятельности желудка осуществляется
нервными и гуморальными механизмами. Влияния, поступающие по
волокнам блуждающих нервов, усиливают моторику желудка: увели­
чивают силу и ритм сокращений, ускоряют эвакуацию желудочного
содержимого. Влияния, которые идут по симпатическим нервам,
уменьшают силу и замедляют ритм сокращений желудка.
Висцеральные функции
409
В регуляции моторной деятельности желудка принимают участие
гастроинтестинальные гормоны. Усиливают моторику желудка: гаст­
рин, мотилин (образуется в слизистой оболочке двенадцатиперстной
киш ки при повышении pH ее содержимого), серотонин, инсулин.
Торможение моторики желудка вызывают: секретин, холецистокинин-панкреозимин, бульбогастрон, энтерогастрон, ВИП и др.
После приема пищи в желудке усиливается кровоснабжение,
выделяется слизь, вырабатывается специальный иммунитет к мета­
болитам, что в норме препятствует перевариванию соляной кислотой
желудочной стенки.
Переход химуса из желудка в двенадцатиперстную кишку
В голодном состоянии субъектов при пустом желудке пилоричес­
кий сфинктер открыт. Он закрывается под влиянием поступления
и обработки пищи в ротовой полости. Последующим открытиям
пилорического сфинктера способствует выделяющаяся в желудке
соляная кислота. При этом порция кислого желудочного содержи­
мого поступает в двенадцатиперстную кишку. Под действием соля­
ной кислоты, жиров и углеводов на рецепторы двенадцатиперстной
киш ки пилорический сфинктер снова закрывается. Его закрытию
способствует также выделяющийся в клетках двенадцатиперстной
киш ки энтерогастрон. Поступившая в двенадцатиперстную киш ку
порция желудочного содержимого затем нейтрализуется соком под­
желудочной железы, желчью, и реакция в двенадцатиперстной кишке
снова становится нейтральной или слабощелочной. При этом соля­
ная кислота из желудка снова способствует открытию пилорического
сфинктера, и в двенадцатиперстную киш ку поступает очередная
порция желудочного содержимого и т.д. (рис. 5.5.3).
Время, необходимое для опорожнения 50% содержимого желудка,
варьирует от 10—20 мин для воды до 1—4 ч для твердой пищи.
Таким образом, рефлекторные влияния на пилорический сфинк­
тер осуществляются с двух сторон: из желудка и двенадцатиперстной
кишки.
На скорость перехода пищи из желудка в двенадцатиперстную
киш ку влияют следующие факторы.
• Наличие жира в двенадцатиперстной кишке тормозит открытие
пилорического сфинктера.
• Открытие сфинктера определяет характер принятой пищи (угле­
водистая пища эвакуируется быстрее, чем белковая, жирная пища
задерживается в желудке на 8—10 ч).
410
Глава 5
$4
• Осмотическое давление пище­
вого комка в желудке. Гипертонические растворы задерживают
эвакуацию пищи из желудка.
• Степень наполнения желудка и
двенадцатиперстной кишки.
• Работа
препилорического
сфинктера.
• Консистенция пищи — содер­
жимое желудка переходит в
двенадцатиперстную кишку,
когда его консистенция стано­
вится жидкой или полужидкой.
Жидкости начинают переходить
в кишку сразу же после поступ­
ления их в желудок.
Рис. 5.5.3. Д инамика основных
этапов в механизме перехода
пищи из желудка в двенадцати­
перстную кишку: А — состоя­
ние сокращ енного пилоричес­
кого сфинктера; Б — состояние
расслабленного пилорическо­
го сфинктера; В — повторное
закрытие пилорического сф ин­
ктера. 1 — возбуждение аф фе­
рентного интрамурального ней­
рона межмышечного (интраму­
рального) сплетения желудка;
2 — тормозящий вставочный ней­
рон межмышечного сплетения;
3 — эфферентный нейрон меж­
мышечного сплетения; 4 — аф­
ферентный нейрон межмышеч­
ного сплетения двенадцатиперс­
тной кишки; 5 — вставочный
возбуждающий нейрон межмы­
шечного сплетения двенадцати­
перстной кишки
Висцеральные функции
411
• Гастроинтестинальные гормоны — секретин, холецистокининпанкреозимин — тормозят моторику желудка и скорость эвакуа­
ции его содержимого.
• Энтерогастральный рефлекс — проявляется в торможении мотор­
ной активности желудка при поступлении химуса в двенадцати­
перстную кишку.
Экскреторная функция желудка
Через желудочную стенку и ее просвет в желудок из крови выде­
ляются мочевина и ряд ионов и белковых веществ.
Пищеварение в двенадцатиперстной кишке
Двенадцатиперстная кишка — основная арена пищеварительного
процесса, который в значительной степени определяется секретом
поджелудочной железы и печени (желчи).
Поджелудочный сок
Бесцветная жидкость имеет щелочную среду — pH равен 7,8—8,4
за счет высокой концентрации бикарбонатов (150 ммоль/л), которые
нейтрализуют химус, поступающий из желудка. За сутки вырабаты­
вается 1,5—2,5 л сока.
Поджелудочный сок богат ферментами, которые секретируются
в ацинозных панкреоцитах, преимущественно в форме неактив­
ных предшественников. Амилаза, липаза и нуклеазы секретируются
поджелудочной железой в активном состоянии, а протеазы — в виде
проферментов (зимогенов). Ферменты поджелудочного сока перева­
ривают все виды питательных веществ.
Трипсиноген поджелудочного сока в двенадцатиперстной кишке
под действием фермента энтерокиназы превращается в трипсин.
Энтерокиназа (энтеропептидаза) была открыта в лаборатории
И.П. Павлова. Последующую активацию трипсиногена вызывает
трипсин. Активация состоит в отщеплении от трипсиногена гек­
сапептида под действием энтерокиназы и образовании трипсина
при pH 6,8—8,0. Процесс ускоряется в присутствии ионов каль­
ция. Энтерокиназа выделяется дуоденальной слизистой оболочкой в
зимогенной форме и активируется дуоденазой.
Трипсин и активирующий химотрипсин (а также панкреатопептидаза, или эластаза) расщепляют преимущественно внутренние пеп­
тидные связи белков. Химотрипсин синтезируется в виде химотрипсиногена и активируется трипсином.
В поджелудочной железе синтезируются прокарбоксипептидазы
А и Б, проэластаза и профосфолипаза, которые активируются трип­
сином, превращаясь в активные формы — карбоксипептидазы А и Б,
эластазы и фосфолипазы.
Трипсин, химотрипсин и эластазы — эндопептидазы, расщеп­
ляющ ие пептидные связи внутри цепи, а карбоксипептидазы
А и Б — экзопептидазы, так как они отщепляют аминокислоты на
карбоксильном конце белковой молекулы (рис. 5.5.4).
Сок поджелудочной железы богат а -амилазой, которая продуциру­
ется в активной форме и расщепляет полисахариды (крахмал и гли­
коген) до ди- и моносахаридов. Дисахариды (мальтоза) расщепляются
малыпазой до моносахаридов (глюкозы). Производные нуклеиновых
кислот расщепляются при действии рибонуклеазы (РНК) и дезоксири­
бонуклеазы (ДНК).
Панкреатическая липаза продуцируется в активной форме и рас­
щепляет жиры до моноглицеридов и жирных кислот. На липиды
действуют также фосфолипазаА (для ее активации необходим трип­
син) и эстераза. Ферментативный гидролиз жиров усиливается в при­
сутствии колипаз, желчи (солей желчных кислот) и ионов кальция.
Фазы панкреатической секреции
Секреция поджелудочной железы начинается через 3—5 мин после
приема пищи. В голодном состоянии сок поджелудочной железы
секретируется в периоды голодной моторной деятельности ЖКТ.
Количество и качество секреции зависят от количества и качества
принятой пищи. На хлеб пик секреции поджелудочной железы при­
ходится на 1-й час, на мясо — на 2-й час, на молоко — на 3-й час.
Как и секреция желудка, секреция поджелудочной железы осу­
ществляется в три фазы: сложнорефлекторную, желудочную и кишеч­
ную. Выделяют также четвертую — циркуляторную фазу секреции.
При этом работа поджелудочной железы гуморально корригирует­
ся всосавшимися нутриентами. Их влияние осуществляется либо
Рис. 5.5.4. Основные этапы расщепления пептидных связей в молекулах
белка под влиянием ферментов желудка, двенадцатиперстной кишки
и тонкого кишечника: А — этап расщепления белков, протекающий
в желудке и двенадцатиперстной кишке; Б — этап гидролиза олигопепти­
дов в тонком кишечнике
Висцеральные функции
413
ЭНДОПЕПТИДАЗЫ
(в желудке и 12-перстной кишке)
[п е п с и н
NH
| п е п с и н | [ т р и п с и н ~ | | ХИМОТРИПСИН | [ " т р и п с и н ] | П Е П С И Н 11 ХИМОТРИПСИН |
СО ' 4фенОф
™
ОО*;ргОС/© .Х/ Е|Э
*“©Оф
е/нООс
оон
*
\
✓
\
/
ч
/
/
/
/
\
/
/
\
nhCQS ф
снОф
ен©LJ0I ОО® 0 0 0"ф
™
:ООф
ен ООсоон
ЭКЗОПЕПТИДАЗЫ
(в тонком кишечнике)
АМИНОПЕПТИДАЗЫ
КАРБОКСИПЕПТИДАЗЫ
ДИПЕПТИДАЗЫ
00
//
NH,
б б
соон
о р
\\
ноос
NH,
фен
6
НООС
оо
NH,
о
соон
414
Глава 5
непосредственно на панкреациты, либо через нервные центры ВНС
и регуляторные пептиды.
Механизм и регуляция пищеварительной секреции поджелудоч­
ной железы
Секреция поджелудочной железы регулируется нервными и гумо­
ральными механизмами.
Нервная регуляция. Подобно желудку, эфферентные парасимпати­
ческие влияния через блуждающий нерв усиливают, а через симпати­
ческие волокна — тормозят ее секреторную активность. И.П. Павлов
установил, что раздражение блуждающего нерва вызывает выделе­
ние поджелудочного сока. Холинергические волокна блуждающих
нервов посредством ацетилхолина действуют на М-холинорецепторы панкреацитов. Затем высвобождение ионов кальция и комплекс
гуанилатциклаза—цГМФ в роли вторичных мессенджеров стимули­
руют секрецию клетками поджелудочной железы ферментов и бикар­
бонатов. Хирургическая и фармакологическая ваготомия существен­
но снижают секрецию поджелудочной железы.
Симпатические волокна, иннервирующие поджелудочную железу,
через посредство а-адренорецепторов тормозят поджелудочную секре­
цию. С другой стороны, снижение секреции симпатической системой
обеспечивается путем сужения кровеносных сосудов через их а-адренорецепторы. Эффект торможения секреции вызывают многие воздейс­
твия — боль, сон, эмоциональный стресс, умственная работа и др.
Гуморальная регуляция
В 1902 г. У. Бейлис и Э. Старлинг при введении в кровь собаки
настоя слизистой оболочки двенадцатиперстной киш ки на соля­
ной кислоте получили обильную секрецию поджелудочной железы
натощак. Из слизистой оболочки двенадцатиперстной киш ки был
выделен ведущий фактор стимуляции панкреатической секреции —
гормон секретин. Он вырабатывается в клетках двенадцатиперстной
киш ки в неактивной форме просекретина и становится активным
под влиянием соляной кислоты, поступающей в двенадцатиперс­
тную кишку из желудка. Поступающий в кровь секретин активирует
пищеварительную секрецию поджелудочной железы, богатую бикар­
бонатами, но бедную ферментами. Секретин разрушается в печени,
но от портальной вены отходит прямая коллатераль к поджелудочной
железе, благодаря чему сохраняется активирующее действие секрети­
на на секрецию поджелудочной железы.
Висцеральные функции
415
Холецистокинин (панкреозимин) усиливает секрецию поджелудочной
железы, высвобождаясь из ССК-клеток слизистой тонкого кишеч­
ника. Он стимулирует образование сока, богатого ферментами.
Выделение холецистокинина стимулирует жирная пища. Трипсин в
полости тонкого кишечника подавляет секрецию холецистокинина
по механизму обратной связи.
Химоденин стимулирует секрецию химотрипсиногена.
Секрецию поджелудочной железы усиливают также гастрин, серо­
тонин, инсулин, бомбезин, соли желчных кислот. Тормозят — глюкагон, соматостатин, вазопрессин, вещество П, АКТГ, энкефалин,
кальцитонин, желудочный ингибирующий пептид и панкреатичес­
кий полипептид. ВИП может как возбуждать, так и тормозить сек­
рецию панкреатического сока.
Саморегуляция секреции поджелудочной железы
Секреция зависит от объема и состава дуоденального содержи­
мого: чем больше объем, тем сильнее торможение секреции. При
повышении содержания протонов в составе химуса происходит
выделение пептидов, которые усиливают секрецию поджелудочной
железы.
Секреция поджелудочной железы также зависит от поступа­
ющих в кровь продуктов гидролиза пищевых веществ и баланса
гастроинтестинальных гормонов. Например, выработка инсулина
увеличивается при повышении концентрации глюкозы в крови и
действии на (3-клетки гастрина и секретина. Инсулин, в свою оче­
редь, стимулирует выработку амилазы, трипсиногена и химотрип­
синогена. Образование глюкагона возрастает при снижении в крови
содержания глюкозы, а также при действии на 5-клетки поджелу­
дочной железы холецистокинина. Глюкагон уменьшает выработку
ферментов поджелудочного сока.
Защитное значение могут иметь еще два тормозных механизма —
повышение активности панкреатических ферментов в крови и повы­
шение давления секрета в протоках поджелудочной железы.
Секреция поджелудочной железы подвергается как сроч­
ной, так и долговременной адаптации, в зависимости от режи­
ма питания. Реализация адаптации осуществляется центральными
и периферическими рефлекторными, гормональными и паракринными механизмами, а также всосавшимися в кровь нутриентами.
Глава 5
416
Функции печени
Пищеварительные функции печени связаны с секретируемой ею
желчью. Желчь секретируется постоянно, а выделяется в двенадцати­
перстную кишку во время голодных моторных сокращений и через
5—10 мин после приема пищи. У человека за сутки образуется около
500—1500 мл желчи (в среднем 10,5 мл на 1 кг массы тела).
Ф ункции желчи в пищеварении
• Эмульгирует жиры в двенадцатиперстной кишке, увеличивая
поверхность, на которой осуществляется их гидролиз липазой.
• Способствует омылению жирных кислот и их всасыванию.
• Нейтрализует соляную кислоту желудочного сока.
• Стимулирует секрецию поджелудочной железы.
• Повышает активность ферментов (панкреатических и кишеч­
ных), особенно липаз.
• Инактивирует пепсины.
• Способствует всасыванию жирорастворимых витаминов, холес­
терина, аминокислот и солей кальция.
• Участвует в пристеночном пищеварении, облегчая фиксацию на
энтероцитах ферментов.
• Усиливает моторную и секреторную функции тонкой кишки.
• Оказывает бактерицидное действие.
• Активирует холецистокинин и тем самым усиливает моторику
желчного пузыря.
Состав желчи
Желчь является не только секретом, но и экскретом. Вместе с
желчью из организма выводятся различные эндогенные и экзогенные
вещества. В желчи содержатся также белки, аминокислоты, витами­
ны и другие вещества. Желчь обладает небольшой ферментативной
активностью; pH печеночной желчи равен 7,3-8,0.
Качественное своеобразие желчи определяют находящиеся в ней
желчные кислоты, пигменты и холестерин.
Основное количество желчных кислот и их солей содержится в желчи
в виде соединений с гликоколом (80%) и таурином (20%). При употреб­
лении пищи, богатой углеводами, в желчи увеличивается содержание
гликохолевых кислот, при высокобелковой диете — таурохолевых.
Желчные пигменты являются экскретируемыми печенью продуктами
распада гемоглобина и других производных порфиринов. Основным
Висцеральные функции
417
желчным пигментом является билирубин — пигмент красно-желтого
цвета, придающий печеночной желчи характерную окраску.
Другой пигмент — биливердин, придающий печени зеленоватую
окраску, содержится в следовых количествах. Он образуется при окис­
лении билирубина (например, при забрасывании желчи в желудок).
В желчи содержится комплексное липопротеиновое соединение, в
состав которого входят фосфолипиды, желчные кислоты, холестерин,
белок и билирубин. Это соединение играет важную роль в транспорте
липидов из печени в кишечник и принимает участие в печеночно­
кишечном кругообороте и общем метаболизме организма.
Всасывание желчных кислот. Из тонкой киш ки всасывается в кровь
около 85—90% желчных кислот, выделившихся в киш ку в составе
желчи. Всосавшиеся желчные кислоты кровотоком воротной вены
вновь приносятся в печень и включаются в состав желчи. Остальные
Ю—15% желчных кислот выводятся из организма в основном с калом
(значительное количество их связывается с непереваренными волок­
нами пищи). Эта потеря желчных кислот восполняется их синтезом
в гепатоцитах.
Ж елчеобразование (холерез)
У человека за сутки образуется около 500—1500 мл желчи. Процесс
образования желчи идет непрерывно, а желчевыделение — поступле­
ние желчи в двенадцатиперстную киш ку — осуществляется периоди­
чески, в основном в связи с приемом пищи. Натощак желчь скапли­
вается в желчном пузыре и выделяется в двенадцатиперстную киш ку
при периодической моторной деятельности ЖКТ. Принято различать
печеночную и пузырную желчь, их состав несколько отличается. При
прохождении желчи по желчевыводящим путям и при нахождении
в желчном пузыре за счет всасывания воды и минеральных солей
происходит концентрирование желчи в 4-10 раз, к ней добавляется
муцин, увеличивается ее плотность и снижается pH (6-7) вследствие
образования желчных кислот и всасывания бикарбонатов.
Акт еды и различные виды принятой пищи усиливают желчеоб­
разование. Образование желчи изменяется при раздражении рецеп­
торов ЖКТ и внутренних органов, а также условно-рефлекторно.
Гуморальными стимуляторами желчеобразования являются сама
желчь, секретин, глюкагон, гастрин, холецистокинин-панкреозимин.
Раздражение блуждающих нервов, введение желчных кислот и
высокое содержание в пище полноценных белков усиливают желче­
образование и выделение с ней органических компонентов.
418
Глава 5
Ж елчевыделение (холекинез)
Во время пищеварения за счет сокращения желчного пузыря дав­
ление в нем резко увеличивается, и обеспечивается выход желчи в
двенадцатиперстную киш ку через открывающийся сфинктер Одди.
Сильными возбудителями желчевыделения являются молоко, яич­
ный желток, сульфат магния, жиры. Через 3—6 ч после приема пищи
происходит снижение желчевыделения, желчь опять начинает скап­
ливаться в желчном пузыре.
Рефлекторные механизмы желчевыделения
Открытие сфинктера общего желчного протока осуществляется
при раздражении пищей рецепторов ротовой полости и желудка,
особенно при механическом раздражении желудка и поступлении в
него жирной пищи.
Эти влияния осуществляются через блуждающие нервы. Слабое
раздражение веточек блуждающего нерва, иннервирующих желчный
пузырь, расслабляет сфинктер общего желчного протока и вызывает
сокращение желчного пузыря. При сильном раздражении сфинктер
сокращается и одновременно расслабляется желчный пузырь.
Симпатическая нервная система тормозит сокращения желчного
пузыря. Это влияние опосредовано а 2-адренорецепторами.
Гуморальные механизмы желчевыделения
Желчевыделение усиливается при переливании голодному живот­
ному «сытой крови». Ведущим гуморальным фактором, стимулиру­
ющим желчевыделение, является холецистокинин-панкреозимин,
образующийся в клетках слизистой двенадцатиперстной киш ки под
влиянием соляной кислоты и жирных кислот. Сокращения желчного
пузыря и усиление желчевыделения вызывают также гастрин, секре­
тин, бомбезин, вещество П.
Тормозят сокращение желчного пузыря активация симпатической
нервной системы, а также глюкагон, кальцитонин, ВИП, панкреати­
ческий полипептид.
Непищеварительные функции печени
Барьерная функция, инактивация токсичных веществ
Печень обезвреживает многие токсичные соединения. Среди них
вещества, поступившие с пищей, образованные микрофлорой в
кишечнике, лекарственные вещества, инфекционные агенты. Печень
участвует в детоксикации ряда гидрофобных соединений, которые
Висцеральные функции
419
могут быть образованы в процессе метаболизма (билирубин, стероид­
ные гормоны) или поступать из пищеварительного тракта (например,
антибиотик хлорамфеникол). Это происходит путем ферментативно­
го окисления, метилирования, ацетилирования, гидролиза (первая
фаза) и последующей конъюгации с рядом веществ — глюкуроновой, серной и уксусной кислотами, глицином, таурином и другими
веществами (вторая фаза). Таким образом конъюгаты становятся
гидрофильными и могут выделяться почками с мочой или печенью
в составе желчи с калом.
Некоторые вещества без изменений выводятся в составе желчи и
мочи, особенно растворимые конъюгаты. И нактивация токсичного
аммиака происходит за счет образования малотоксичной мочевины
и креатинина. Инфекционные агенты обезвреживаются в основном
путем их фагоцитоза и лизиса.
Инактивация гормонов и аминов. Печень принимает участие в инакти­
вации ряда гормонов — глюкокортикоидов, альдостерона, андрогенов,
■эстрогена, инсулина, глюкагона, ряда гастроинтестинальных гормонов
и биогенных аминов — гистамина, серотонина, катехоламинов.
Экскреторная функция
Выражается в элиминации из крови и выделении в составе желчи
большого числа веществ, обычно трансформированных в печени, т.е.
она участвует в обеспечении гомеостаза.
Обмен белков. Печень играет большую роль в обмене белков: в
ней синтезируются белки крови (весь фибриноген, 95% альбуминов,
85% глобулинов), происходят дезаминирование и переаминирование
аминокислот, образование мочевины, глутамина, креатина, ф акто­
ров свертывающей и противосвертывающей систем крови (I, II, V,
VII, IX, X, XII, X III, антитромбин, антиплазмин). Желчные кислоты
влияют на транспортные свойства белков крови.
Обмен липидов. Печень участвует в обмене липидов, а именно, в
их гидролизе и всасывании, синтезе триглицеридов, фосфолипидов,
холестерина, желчных кислот, липопротеидов, ацетоновых тел, окис­
лении триглицеридов. Обмен углеводов — еще одна функция печени.
Она принимает участие в процессах гликогенеза, гликогенолиза, во
включении в обмен глюкозы, галактозы и фруктозы, в образовании
глюкуроновой кислоты.
Эритрокинетика. Установлена роль печени в разрушении эритро­
цитов, деградации гема и последующем пигментном обмене через
образование билирубина.
Глава 5
4 20
Обмен витаминов. Печень обеспечивает обмен витаминов, особен­
но жирорастворимых — A, D, Е, К, всасывание которых в кишечнике
происходит с участием желчи. Ряд витаминов депонируется в печени
и высвобождается по мере их метаболической потребности (A, D, К,
С, РР).
Депонирование микроэлементов и электролитов
В печени депонируются микроэлементы (железо, медь, марганец,
кобальт, молибден и др.) и электролиты.
Иммунопоэз и иммунологическая реакция
Печень участвует в иммунопоэзе и иммунологических реакциях.
Кишечно-печеночная циркуляция желчных кислот
Желчные кислоты важны не только для гидролиза и всасывания
липидов, но и для других процессов. Они являются регуляторами
холереза и выделения в составе желчи холестерина, желчных пигмен­
тов; определяют активность печеночных цитоферментов, влияют на
транспортную активность энтероцитов, регулируют пролиферацию,
передвижение и отторжение энтероцитов от кишечных ворсинок.
Регуляторное влияние желчи распространяется на секрецию желудка,
поджелудочной железы и тонкой киш ки, эвакуаторную деятельность
гастродуоденального комплекса, моторику кишечника, реактивность
органов пищеварения к нейротрансмиттерам, регуляторным пепти­
дам и аминам.
Пищеварение в тонкой кишке
Секреция тонкой кишки
Тонкий кишечник взрослого человека имеет длину около 2 м.
Главная его функция — завершение расщепления пищи и всасывание
расщепленных веществ, воды, электролитов и витаминов.
Кишечный сок имеет щелочную реакцию. Он представляет собой
мутную вязкую жидкость и является продуктом деятельности кишеч­
ных желез всей слизистой оболочки тонкой киш ки. За сутки у чело­
века выделяется до 2,5 л сока тонкой кишки.
В верхней части двенадцатиперстной киш ки заложены бруннеровы
(дуоденальные) железы. Сок бруннеровых желез — густая бесцветная
жидкость слабощелочной реакции, обладающая небольшой протеолитической, амилолитической и липолитической активностью.
Висцеральные функции
421
Секреторной способностью обладают либеркюновы железы (кишеч­
ные крипты).
В гранулярной эндоплазматической сети бокаловидных клеток
образуются белковые компоненты секрета, в комплексе Гольджи
(пластинчатом комплексе) — мукополисахариды. Секрет этих клеток
обладает ферментативной активностью, в том числе и протеолитической.
Секрет энтероцитов содержит гидролитические ферменты. В крип­
тах заложены также аргентаффинные клетки, выполняющие эндок­
ринные функции.
Кишечный эпителий секретирует многие вещества в полость тон­
кой киш ки, ряд веществ транспортируется из крови. Находящиеся в
кишке вещества активно и пассивно переносятся из ее полости и с
поверхности слизистой оболочки в кровь и лимфу. Полное обновле­
ние кишечного эпителия происходит каждые 3—6 сут.
Состав киш ечного сока
В состав кишечного сока входят неорганические вещества (около
10 г/л) — хлориды, гидрокарбонаты и фосфаты натрия, калия, каль­
ция; pH сока — 7,2—7,5, при усилении секреции pH повышается до 8,6.
Органические вещества в составе жидкой части сока представлены
слизью, белками, аминокислотами, мочевиной и другими продукта­
ми обмена веществ.
Слизь образует защитный слой, предотвращающий чрезмерное
механическое и химическое воздействие химуса на слизистую обо­
лочку кишки. В слизи высока активность ферментов, гидролизую­
щих питательные вещества.
В слизистой оболочке тонкой киш ки происходит непрерывная
смена слоя клеток поверхностного эпителия. Они образуются в крип­
тах, затем продвигаются по ворсинкам и слущиваются с их верхушек —
морфокинетическая (или морфонекротическая) секреция. Полное
обновление этих клеток у человека совершается за 1,4—бсут, т.е. за
I ч слущивается около 2% клеток. Столь высокий темп образования
клеток и их отторжения обеспечивает достаточно большое их коли­
чество в кишечном соке (у человека за сутки отторгается около 250
эпителиоцитов).
Ферменты кишечного сока
Основная часть ферментов синтезируется в слизистой оболоч­
ке киш ки, но некоторое их количество экскретируется из крови.
4 22
Глава 5
В кишечном соке более 20 различных ферментов. Основные из
них: энтерокиназа, несколько специфических пептидаз (аминополипептидаза и дипептидаза), щелочная фосфатаза, нуклеазы, липа­
за, фосфолипаза, амилаза, мальтаза, инвертаза, лактаза, сахараза,
дуоденаза. Для большинства кишечных ферментов характерен проксимодистальный градиент — снижение их содержания и активности в
тонкой кишке в направлении толстой кишки.
Моторная деятельность тонкого кишечника
Моторика тонкой киш ки обеспечивает перемешивание ее содер­
жимого (химуса) с пищеварительными секретами, продвижение
химуса по кишке, смену его слоя у слизистой оболочки, повышение
внутрикишечного давления (способствующего фильтрации раство­
ров из полости киш ки в кровь и лимфу) и продвижение химуса по
градиенту давления. Следовательно, моторика тонкой киш ки участ­
вует в процессах гидролиза и всасывания, содействует им.
Типы сокращений тонкой кишки
Движение тонкой киш ки происходит в результате координирован­
ных сокращений продольного и циркулярного слоев гладких мышц.
Принято различать несколько типов сокращений тонкой кишки.
Ритмическая сегментация обеспечивается преимущественно сокра­
щениями циркулярного слоя мышц. При этом содержимое кишки
делится на части. Следующим сокращением образуется новый сег­
мент киш ки, содержимое которого состоит из двух частей бывше­
го сегмента. Данными сокращениями достигаются перемешивание
химуса и повышение давления в каждом сегменте.
Маятникообразные сокращения обеспечиваются продольными мыш­
цами и некоторым участием в сокращении циркулярных мышц. При
этом химус перемещается вперед-назад и происходит слабое посту­
пательное движение его в направлении толстой кишки. В верхних
отделах тонкой кишки человека частота ритмических сокращений
составляет 9—12, в нижних — 6—8 сокращений в минуту.
Перистальтическая волна , состоящая из перехвата и расширения
тонкой киш ки, продвигает химус в направлении толстой киш ки.
Одновременно по длине киш ки продвигается несколько перисталь­
тических волн. Перистальтическая волна продвигается по кишке
со скоростью 0,1—0,3 см/с, в проксимальных отделах скорость боль­
ше, чем в дистальных. Скорость стремительной (пропульсивной)
волны — 7—21 см/с.
Висцеральные функции
423
При антиперисталыпических сокращениях волна движется в обрат­
ном, оральном направлении. Это характерно для рвоты.
Тонические сокращения могут продвигать содержимое киш ки с очень
малой скоростью или вообще не продвигать его. Тонические сокра­
щения суживают просвет киш ки на большом ее протяжении.
Исходное (базальное) давление в полости тонкой киш ки состав­
ляет 5—14 см вод.ст. Монофазные волны повышают внутрикишечное
давление в течение 8 с до 30-90 см вод.ст. Медленный компонент
сокращений длится от 1 мин до нескольких минут и повышает дав­
ление не столь значительно.
Регуляция моторики тонкой кишки
Моторика тонкой кишки регулируется миогенными, нервными и
гуморальными механизмами. Фазная сократительная деятельность
стенки киш ки реализуется нейронами мезентериального нервного
сплетения, обладающими ритмической фоновой активностью. Кроме
них, имеются два «датчика» ритма кишечных сокращений: первый —
у места впадения в двенадцатиперстную кишку общего желчного
протока, второй — в подвздошной кишке. Эти «датчики» и ганглии
энтерального нервного сплетения контролируются нервными и гумо­
ральными механизмами.
Нервная регуляция. Ведущее значение в регуляции моторики тонкой
кишки имеет интрамуральная нервная система (метасимпатическая
система). Интрамуральные нейроны обеспечивают координирован­
ные сокращения кишки. На интрамуральные механизмы регуляции
оказывают влияния экстрамуральные симпатические и парасимпа­
тические нервные механизмы, а также гуморальные факторы.
Парасимпатические влияния преимущественно усиливают, симпа­
тические — тормозят моторику тонкой кишки. Моторная функция кон­
тролируется центрами спинного и продолговатого мозга, гипоталамуса,
лимбической системы, коры большого мозга: раздражение ядер пере­
дних и средних отделов гипоталамуса преимущественно возбуждает, а
заднего — тормозит моторику желудка, тонкой и толстой кишки.
Акт еды кратковременно тормозит, а затем усиливает кишечную
моторику. В дальнейшем она зависит от физических и химических
свойств химуса: усиливают ее грубые виды пищи и жиры.
Гуморальная регуляция. Усиливают моторику тонкой кишки вазопрессин, окситоцин, брадикинин, серотонин, вещество П, гистамин, гастрин, мотилин, холецистокинин-панкреозимин, щелочи, кислоты,
соли. Тормозят — секретин, ВИП, гастроингибирующий пептид.
4 24
Глава 5
Эвакуация кишечного химуса в толстую киш ку
Из тонкой киш ки химус через илеоцекальный сфинктер (баугиниева заслонка) порционно переходит в толстую кишку. Сфинктер
имеет сложное строение; он играет роль клапана, который сужен­
ной частью обращен в просвет слепой кишки; здесь же сосредо­
точены циркулярные мышцы, образующие собственно сфинктер.
Их расслаблению и раскрытию илеоцекального прохода способс­
твуют сокращения продольных мышц тонкой и толстой киш ки.
При наполнении слепой киш ки и ее растяжении сфинктер плотно
закрывается, и содержимое толстой киш ки в норме в тонкую кишку
не переходит.
Вне пищеварения илеоцекальный сфинктер закрыт. Через 1—4 мин
после приема пищи каждые 0,5—1 мин он открывается и химус неболь­
шими порциями (до 15 мл) поступает в толстую кишку. Раскрытие
сфинктера происходит рефлекторно: перистальтическая волна тонкой
киш ки, повышая давление в ней, расслабляет его и пилорический
сфинктер (бисфинктерный рефлекс). Повышение давления в толстой
кишке увеличивает тонус илеоцекального сфинктера и тормозит пос­
тупление в нее содержимого тонкой кишки.
Пищеварение в толстой кишке
Пища почти полностью переваривается и всасывается в тонкой
кишке. Небольшое количество веществ пищи, в том числе клет­
чатка и пектин, пищеварительные соки в составе химуса подверга­
ются гидролизу в толстой кишке, которая имеет длину около 1,3 м.
Гидролиз осуществляется ферментами химуса, микроорганизмов и
сока толстой кишки. Толстая кишка выполняет функцию резервуара
для содержимого кишечника, а также функцию всасывания воды и
электролитов. За сутки у здорового человека из тонкой в толстую
киш ку переходит 0,5—4 л химуса. За счет всасывания в толстой кишке
объем содержимого может уменьшиться до 100—200 мл.
Значение микрофлоры кишечника заключается в том, что она
участвует в конечном разложении остатков непереваренной пищи.
Микрофлора участвует в инактивации и расщеплении ферментов и
других биологически активных веществ. Нормальная микрофлора
подавляет патогенные микроорганизмы и предупреждает инфици­
рование. Ферменты бактерий расщепляют волокна клетчатки, не
переваренные в тонкой кишке. Кишечная флора синтезирует вита­
мин К и витамины группы В, а также другие вещества, необходимые
организму. С участием микрофлоры кишечника в организме про­
Висцеральные функции
4 25
исходит обмен белков, фосфолипидов, желчных и жирных кислот,
билирубина и холестерина.
Состав сока и функции толстой кишки
Сок состоит из жидкой и плотной частей, имеет щелочную реак­
цию (pH 8,5—9,0). Плотную часть сока составляют слизистые комоч­
ки из отторгнутых кишечных эпителиоцитов и слизи, секретируемой
бокаловидными клетками.
Сок толстой киш ки в небольшом количестве выделяется вне раз­
дражения киш ки. Ее местное механическое раздражение увеличива­
ет секрецию в 8—10 раз.
В толстом кишечнике осуществляются процессы:
• сгущения содержимого вследствие всасывания воды;
• брожения за счет действия микрофлоры.
Железы слизистой оболочки толстой киш ки выделяют небольшое
количество сока, богатого слизистыми веществами, но бедного фер­
ментами. В соке толстой киш ки в небольшом количестве содержатся
катепсин, пептидазы, липаза, амилаза и нуклеазы.
Весь процесс пищеварения у взрослого человека длится 1—3 сут,
из них наибольшее время приходится на пребывание остатков пищи
в толстой кишке.
Моторная деятельность толстой киш ки и ее регуляция
Моторика толстой киш ки осуществляет накопление содержимо­
го, его продвижение, всасывание из него ряда веществ, в основном
воды (до 6 л/сут), формирование каловых масс и их удаление (дефе­
кация).
Различают следующие виды сокращений толстой кишки:
• тонические сокращения;
• маятникообразные сокращения;
• ритмическая сегментация;
• перистальтические сокращения;
• антиперистальтические сокращения (способствуют всасыванию
воды и формированию каловых масс);
• пропульсивные сокращения (обеспечивают продвижение содер­
жимого киш ки в каудальном направлении).
Регуляция моторной деятельности толстой кишки осуществляется
как нервным (за счет ВНС), так и гуморальным путем.
Парасимпатический отдел ВНС усиливает моторику толсто­
го киш ечника (иннервация блуждающими и тазовыми нервами).
426
Глава 5
Симпатические нервы проходят в составе чревных нервов и тормозят
моторику кишки. Метасимпатический отдел ВНС реализует саморе­
гуляцию движений кишки.
Моторику тормозят серотонин, адреналин, глюкагон; усиливает —
раздражение механорецепторов прямой кишки.
Всасывание
Всасывание — процесс транспорта питательных веществ из ЖКТ
во внутреннюю среду организма — в кровь и лимфу. Всосавшиеся
вещества разносятся по организму и включаются в клеточный мета­
болизм.
Всасывание в различных отделах пищеварительного тракта
Всасывание в полости рта. В полости рта пища не расщепляется до
мономеров и находится кратковременно, поэтому всасывание здесь
ничтожно мало. Однако некоторые лекарственные средства всасыва­
ются достаточно быстро, что делает возможным их применение под
язык (сублингвально).
Всасывание в желудке бывает незначительным; в больших коли­
чествах всасываются лишь некоторые аминокислоты, глюкоза, вода
и растворенные в ней минеральные соли. Хорошо всасываются рас­
творы этилового спирта (алкоголь).
Основное всасывание нутриентов, воды, электролитов происхо­
дит в тонкой кишке и сопряжено с гидролизом питательных веществ.
Всасывание зависит от величины поверхности, на которой оно осу­
ществляется. На 1 мм2 слизистой оболочки киш ки человека при­
ходится 30—40 ворсинок, а каждый энтероцит имеет около 1700—
4000 микроворсинок, следовательно, на 1 мм2 поверхности кишечного
эпителия приходятся 50—100 млн микроворсинок. Таким образом,
общая площадь поверхности Ж КТ составляет около 100 м2.
У взрослого человека число всасывающих кишечных клеток
составляет Ю10, а соматических клеток — 1015. Из этого следует,
что одна кишечная клетка обеспечивает питательными веществами
около 100 000 других клеток организма человека.
В толстой кишке происходит в основном всасывание воды и фор­
мирование каловых масс. В небольших количествах в толстой кишке
могут всасываться глюкоза, аминокислоты, а также другие легко
всасывающиеся вещества. На этом основано применение так назы­
ваемых питательных клизм, т.е. введение легкоусвояемых веществ в
прямую кишку.
Висцеральные функции
427
В верхних отделах кишечника глюкоза всасывается быстрее воды.
В нижних частях кишечника вода всасывается быстрее хлористого
натрия.
Всасывание макромолекул
Различные вещества всасываются посредством разных механиз­
мов. Транспорт макромолекул и их агрегатов осуществляется путем
фагоцитоза и пиноцитоза. Эти механизмы объединены под названи­
ем эндоцитоза. С эндоцитозом связано внутриклеточное пищеваре­
ние. Ряд веществ попадает в клетку путем эндоцитоза, транспорти­
руется в везикуле через клетку и выделяется из нее в межклеточное
пространство путем экзоцитоза. Такой транспорт веществ назван
трансцитозом. Он не имеет существенного значения во всасывании
нутриентов, но важен в переносе веществ иммунной защиты, вита­
минов и ферментов из кишечника в кровь. У новорожденных детей
трансцитоз важен для транспорта многофункциональных белков
материнского молока.
Некоторое количество веществ может транспортироваться по меж­
клеточным пространствам. Такой транспорт называется персорбцией. Посредством персорбции переносится некоторое количество воды
и электролитов и меньшее количество других веществ, в том числе
белков (антител, аллергенов, ферментов и др.), и даже бактерий.
Всасывание микромолекул
Всасывание микромолекул — основных продуктов гидролиза
питательных веществ в ЖКТ, а также электролитов осуществля­
ется тремя видами транспорта: пассивным, облегченной диффузией и
активным. Пассивный транспорт включает в себя диффузию, осмос
и фильтрацию. Движущей силой диффузии является концентраци­
онный градиент частиц растворенного вещества. Разновидностью
диффузии является осмос, при котором перемещение происходит в
соответствии с концентрационным градиентом растворителя. Под
фильтрацией понимают процесс переноса раствора через пористую
мембрану под действием гидростатического давления.
Облегченная диффузия, как и простая диффузия, осуществляется
без затраты энергии по градиенту концентрации, но с помощью осо­
бых мембранных переносчиков.
Активный транспорт — перенос веществ через мембраны против
электрохимического или концентрационного градиента с затратой
энергии и при участии специальных транспортных систем: мембран­
428
Глава 5
ных транспортных каналов, мобильных переносчиков, конформационных переносчиков.
Эти механизмы переносят один или несколько, но ограниченное
число типов веществ. Часто транспорт веществ сопряжен с перемеще­
нием другого вещества, движение которого по градиенту концентрации
служит источником энергии для сопрягаемого транспорта. В такой роли
используются ионные градиенты, особенно градиент Na+.
В тонкой кишке № +-зависимым является всасывание глюко­
зы, галактозы, свободных аминокислот, дипептидов и трипептидов,
солей желчных кислот, билирубина и ряда других веществ. № +-зависимый транспорт осуществляется также через специальные каналы и
мобильными переносчиками. Распространены № +-зависимые пере­
носчики на апикальных мембранах, Na+-Hacocbi — на базолатераль­
ных мембранах энтероцитов (рис. 5.5.5).
В тонкой кишке существует и Ка+-независимый транспорт многих
мономеров нутриентов.
Полость
кишечника
белок-переносчик
Рис. 5.5.5. Основные этапы всасывания глюкозы в тонком кишечнике:
А — поступление молекул глюкозы внутрь энтероцита; Б — высвобожде­
ние молекул глюкозы и ионов натрия белком-переносчиком и поступление
в межклеточное пространство; В —поступление глюкозы и натрия из меж­
клеточного пространства в кровеносные сосуды
Висцеральные функции
4 29
Градиент Na+ и К+ между вне- и внутриклеточными жидкостями
обеспечивается путем активного транспорта. Переносчики в клет­
ках связаны с деятельностью ионных насосов, которые используют
энергию АТФ с помощью нескольких транспортных АТФ-аз. Наиболее
важна в процессах всасывания Na+, К+-АТФ-аза. Она участвует в
обеспечении энергией № +-зависимого транспорта.
Роль внутрикишечного давления во всасывании
Повышение внутрикишечного давления до 8—10 мм рт. ст. в 2 раза
увеличивает скорость всасывания из тонкой киш ки раствора хлорида
натрия. Это указывает на значение фильтрации во всасывании и на
роль кишечной моторики в этом процессе.
Роль моторики кишечника во всасывании
Моторика тонкой киш ки обеспечивает не только внутрикишечное
давление, но и периодическую смену пристеночного слоя химуса, что
важно для гидролиза и всасывания его продуктов.
Скорость всасывания из тонкой киш ки в большой мере зависит от
уровня ее кровоснабжения. В свою очередь, оно увеличивается при
наличии в тонкой кишке продуктов, подлежащих всасыванию.
Сокращение ворсинок и микроворсинок
Большое значение для всасывания имеют движения ворсинок
слизистой оболочки тонкой киш ки и микроворсинок энтероцитов, имеющих специальные сократительные элементы. Сокращения
ворсинок выдавливают из сжимающейся полости лимфатических
сосудов лимфу с всосавшимися в нее веществами. Наличие кла­
панов препятствует возврату лимфы в сосуд при последующем
расслаблении ворсинки и обеспечивает присасывающее действие
центрального лимфатического сосуда. Сокращения микроворсинок
усиливают эндоцитоз и, возможно, являю тся одним из его меха­
низмов.
Натощак ворсинки сокращаются редко и слабо, при наличии в
кишке химуса сокращения ворсинок усиливаются и учащаются.
Механическое раздражение основания ворсинок в эксперименте
усиливает их сокращения, тот же эффект наблюдается под влиянием
химических компонентов пищи, особенно продуктов ее гидролиза —
пептидов, некоторых аминокислот, глюкозы, а также экстрактивных
веществ пищи. В реализации этих воздействий определенная роль
отводится метасимпатической нервной системе.
430
Глава 5
Установлено, что кровь сытых животных, перелитая голодным,
вызывает у них усиление движения ворсинок.
Всасывание различных веществ
Всасывание воды и минеральных солей
Вода поступает в Ж КТ в составе пищи и выпиваемых жидкос­
тей (2—2,5 л), секретов пищеварительных желез (6—7 л), выводится
же с калом в сутки 100—150 мл воды. Все остальное количество
воды всасывается из пищеварительного тракта в кровь, небольшое
количество — в лимфу. Всасывание воды начинается в желудке, но
наиболее интенсивно оно происходит в тонкой и особенно толстой
кишке — за сутки около 8 л.
Всасывание некоторого количества воды происходит по осмотичес­
кому градиенту, но возможно и при отсутствии разности осмотичес­
кого давления. Абсорбция воды из изотонических и гипертонических
растворов требует затраты энергии. Активно всасываемые эпителиоцитами растворенные вещества «тянут» за собой воду. Энергия,
освобождаемая в тонкой кишке при гликолизе и окислительных про­
цессах, повышает всасывание воды. Решающая роль в переносе воды
принадлежит ионам, особенно натрия и хлора. Ингибитор натриевого
насоса оуабаин подавляет всасывание воды.
Всасывание воды сопряжено также с транспортом сахаров и ами­
нокислот. При подавлении их всасывания флорицином всасывание
воды замедляется.
Выключение из пищеварения желчи замедляет всасывание воды
из тонкой кишки. Ваготомия также замедляет всасывание воды. На
процесс всасывания воды оказывают влияние гормоны: АКТГ усили­
вает всасывание воды и хлоридов, не влияя на всасывание глюкозы,
Т4 повышает всасывание воды, глюкозы и липидов. Гастрин, секре­
тин, холецистокинин-панкреозимин, бомбезин, серотонин и ВИП
ослабляют всасывание воды.
Натрий интенсивно всасывается в тонкой и подвздошной кишке.
Ионы Na+ переносятся из полости тонкой киш ки в кровь через
кишечные эпителиоциты и по межклеточным каналам. Поступление
ионов Na+ в эпителиоцит происходит пассивно по электрохимичес­
кому градиенту. Из эпителиоцитов через их латеральные и базальные
мембраны ионы Na+ активно транспортируются в межклеточную
жидкость, кровь и лимфу. По межклеточным каналам транспорт
ионов Na+ осуществляется пассивно по градиенту концентрации.
Висцеральные функции
431
В толстой кишке всасывание Na+ не зависит от наличия сахаров и
аминокислот, а в тонкой кишке — зависит от этих веществ. В тонкой
кишке перенос ионов Na+ и С1~ сопряжен. В толстой кишке идет обмен
всасывающихся ионов Na+ на ионы К+. При снижении содержания в
организме натрия его всасывание в кишечнике резко увеличивается.
Всасывание ионов Na+ усиливают гормоны гипофиза и надпочечников,
угнетают — гастрин, секретин и холецистокинин-панкреозимин.
Всасывание ионов калия происходит в основном в тонкой кишке с
помощью пассивного транспорта по электрохимическому градиенту.
Всасывание ионов хлора происходит в желудке, а наиболее активно —
в подвздошной кишке по механизму активного и пассивного транс­
порта. Транспорт ионов С1~ сопряжен с транспортом ионов Na+.
Всасывание аминокислот
Белки всасываются в основном в кишечнике после их гидролиза
до аминокислот. Расщепление белков начинается в желудке после
денатурации соляной кислотой и превращения пепсиногенов в пеп­
сины (рис. 5.5.6).
Всасывание аминокислот из полости киш ки в ее эпителиоциты
осуществляется активно с участием переносчика и с затратой энер­
гии АТФ. В апикальной мембране эпителиоцитов функционирует
пять видов переносчиков аминокислот. Из эпителиоцитов амино­
кислоты по механизму облегченной диффузии транспортируются в
межклеточную жидкость и в кровь.
Интенсивность всасывания аминокислот зависит от возраста
(более интенсивно оно у молодых людей), уровня белкового обмена
в организме, содержания в крови свободных аминокислот, нервных
и гуморальных влияний.
Всасывание углеводов
Углеводы всасываются только в виде моносахаридов. С наибольшей
скоростью всасываются гексозы (глюкоза, галактоза и др.), пентозы
всасываются медленнее. Всасывание глюкозы и галактозы — резуль­
тат их активного транспорта через апикальные мембраны кишечных
эпителиоцитов. Транспорт глюкозы и других моносахаридов активи­
руется транспортом ионов Na+ через апикальные мембраны по меха­
низму совместного транспорта (симпорта). Глюкоза аккумулируется в
кишечных эпителиоцитах. Дальнейший транспорт глюкозы из них в
межклеточную жидкость и кровь через базальные и латеральные мем­
браны происходит пассивно по градиенту концентрации.
Глава 5
4 32
Белковая молекула
Пепсины
Карбопептидазы
Полипептиды
Б
Протеазы
OQOOOO ООООО
Олигопептиды
ООО ООО ©ОО
Трипептиды
ОО
Дипептиды
ОО
ОО
Висцеральные функции
433
Всасывание глюкозы усиливают парасимпатические влияния,
гормоны — глюкокортикоиды, Т4, инсулин, некоторые аминокисло­
ты. Несколько замедляет этот процесс гистамин. Значительно тор­
мозят всасывание глюкозы соматостатин, активация симпатической
нервной системы и ингибиторы тканевого дыхания.
Всасывание продуктов гидролиза жиров
Среднее потребление жиров с пищей составляет 60—100 г/сут.
Основные превращения веществ в организме происходят в водной
среде, а липиды и часть продуктов их гидролиза в воде нераство­
римы. Поэтому всасывание липидов связано с их сложными биохи­
мическими превращениями. Наиболее активно они всасываются в
двенадцатиперстной кишке и проксимальной части тощей кишки.
Скорость всасывания различных липидов в кишечнике зависит от
степени их эмульгирования и гидролиза. При этом для оптималь­
ного гидролиза жиров требуется их предварительное эмульгирова­
ние желчью, при котором размер липидных частиц уменьшается до
1—2мкм. При эмульгировании значительно увеличивается площадь
поверхности капелек липидов, что облегчает доступ гидролитичес­
ких ферментов, необходимых для расщепления жиров.
Липазы выделяются в полости рта, в желудке и поджелудочной
железе. Около 10—30% жира пищи подвергается гидролизу в желудке,
в то время как остальные 70—90% — в двенадцатиперстной кишке и
в начальных отделах тонкого кишечника.
В результате действия в полости киш ки панкреатической липазы
из триглицеридов образуются диглицериды, затем моноглицериды
и жирные кислоты, хорошо растворимые в растворах солей жел­
чных кислот. Кишечная липаза завершает гидролиз липидов. Из
моноглицеридов и жирных кислот с участием солей желчных кис­
лот, фосфолипидов и холестерина образуются мельчайшие мицел­
лы (диаметр их около 20—100 нм). Снаружи мицелл, контактируя
с водной средой кишечника, расположены полярные гидрофиль­
ные компоненты, включающие желчные кислоты, моноглицериды и
Рис. 5.5.6. Динамика последовательных процессов расщепления белко­
вых молекул под влиянием ферментов желудочно-киш ечного тракта:
А — процесс расщепления белковых ферментов до полипептидов; Б — про­
цесс расщепления полипептидов
Глава 5
434
фосфолипиды. Внутри мицелл находятся неполярные гидрофобные
соединения (производные холестерина, жирорастворимые витамины
и др.) (рис. 5.5.7).
Таким образом, через апикальные мембраны мицеллы переходят
в кишечные эпителиоциты. Желчные кислоты мицелл остаются в
полости киш ки и всасываются в подвздошной киш ке по механизму
активного транспорта.
В кишечных эпителиоцитах происходит ресинтез триглицеридов.
Из них, а также холестерина, фосфолипидов и глобулинов образуются
хиломикроны — мельчайшие жировые частицы, заключенные в тон­
чайшую белковую оболочку. Хиломикроны покидают эпителиоциты,
переходя в соединительные пространства ворсинок, откуда проникают
в центральный лимфатический сосуд ворсинки, чему в большой мере
содействуют ее сокращения. Основное количество липидов всасыва­
ется в лимфу, поэтому через 3—4 ч после приема пищи лимфатические
сосуды наполнены большим количеством лимфы, напоминающей
молоко и называемой потому млечным соком. Небольшое количество
хиломикронов поступает и в кровеносные сосуды ворсинок.
Парасимпатические влияния усиливают, а симпатические —
замедляют всасывание жиров. Усиливают всасывание жиров гормо­
ны коры надпочечников, щитовидной железы и гипофиза, а также
секретин и холецистокинин-панкреозимин.
Микрофлора пищеварительного тракта
Пищеварительный тракт заселен микроорганизмами, которые
составляют единую динамичную экологическую систему и выпол­
няют ряд важных функций.
Топография микрофлоры
Содержимое Ж КТ у новорожденного стерильно, но в течение
первых недель жизни его заселяют попавшие через рот анаэробные
микроорганизмы. Для каждого отдела пищеварительного тракта
Рис. 5.5.7. Основные этапы гидролиза и всасывания продуктов гидролиза
жиров в тонком кишечнике: А — последовательность основных процессов
образования мицелл; Б — основные этапы образования хиломикронов в
энтероцитах; В — поступление свободных жирных кислот и хиломикро­
нов из энтероцитов в кровеносные и лимфатические сосуды (Ж К — жир­
ная кислота; МГ — моноглицериды; СЖ — свободная жирная кислота;
ДГ —диглицериды; ТГ —триглицериды)
Энтероцит
Б
436
Глава 5
характерны свои количество и набор микроорганизмов. Их число в
полости рта, несмотря на бактерицидные свойства слюны, состав­
ляет 10 ООО 000—100 000 000 клеток на 1 мл ротовой жидкости. Так
как содержимое желудка имеет низкое значение pH, обычно оно
бывает стерильным, но может содержать до 1000 микроорганизмов
на 1 мл содержимого. По той же причине в двенадцатиперстной и
начальной части тощей киш ки микрофлора также бедна или отсутс­
твует. В содержимом подвздошной киш ки число микроорганизмов
составляет 1 000 000 на 1 мл содержимого. В толстой кишке число бак­
терий максимальное: 1 г фекалий здорового человека содержит 10 млрд
и более микроорганизмов.
Эубиоз
Особого внимания в механизме обеспечения эубиоза заслужи­
вают взаимоотношения между различными видами бактерий. Этот
механизм играет роль в формировании достаточно специфичной
микрофлоры сапрофитов и защите от патогенных бактерий. Данный
вопрос был объектом научных интересов И.И. Мечникова, который
предлагал заменить патогенную флору кишечника ацидофильны­
ми бактериями. Он полагал, что включение в диету специальных
молочнокислых смесей может избавить человека от многих болезней
и существенно продлить жизнь.
Важный экзогенный фактор, влияющий на состав микрофлоры, —
питание. Его постоянство, сбалансированность и адекватность
важны в стабилизации эубиоза человека. Вегетарианская диета уве­
личивает количество энтерококков и эубактерий. Избыточный прием
животных белков и жиров повышает в составе микрофлоры число
клостридий, бактероидов. При избытке в рационе животных жиров
увеличивается число бактероидов и уменьшается число бифидобак­
терий и энтерококков, недостаток жиров вызывает обратные изме­
нения микрофлоры. Молочная диета (лактоза) повышает количество
бифидобактерий. Включение в рацион человека нефизиологических
компонентов (например, белков одноклеточных), сублимированных
продуктов неблагоприятно влияет на микрофлору кишечника.
Ф изиологическая роль микрофлоры
Нормальная микрофлора выполняет ряд важнейших функций.
• Исключительно важно ее участие в формировании иммуноби­
ологической реактивности организма: она предохраняет мак­
роорганизм от внедрения и размножения в нем патогенных
Висцеральные функции
437
микроорганизмов. Нарушение нормальной микрофлоры при
заболевании или в результате длительного введения антибак­
териальных препаратов нередко влечет за собой осложнения,
вызываемые бурным размножением в киш ечнике дрожжей, ста­
филококка, протея и т.п.
• Кишечная микрофлорасинтезируетвитамины Ки группы В, учеловека микробный синтез частично покрывает потребность в них.
• Ферменты бактерий расщепляют целлюлозу, гемицеллюлозу и
пектины, не переваренные в тонкой кишке.
• М икроорганизмы утилизируют непереваренные питательные
вещества, образуя при этом ряд веществ, которые всасываются из
кишечника и включаются в общий обмен веществ. Микрофлора
существенно влияет на печеночно-кишечную циркуляцию ком­
понентов желчи и через них — на деятельность печени. С участи­
ем микрофлоры кишечника происходит обмен белков, фосфоли­
пидов, желчных и жирных кислот, билирубина, холестерина.
• М икрофлора участвует в инактивировании ферментов энте­
рокиназы, щелочной фосфатазы, трипсина, амилазы. В ряде
случаев инактивирующие эффекты микрофлоры сочетаются с
таковыми протеолитических ферментов пищеварительных желез.
Микроорганизмы принимают участие в разложении желчных
кислот, ряда органических веществ с образованием органических
кислот, их аммонийных солей, аминов и др.
Функциональная система, обеспечивающая
акт дефекации
Выделение каловых масс из киш ечника (дефекация) — важный
физиологический процесс. С калом выделяются непереваренные
продукты, такие как растительная клетчатка, кератины и некоторые
коллагены; холестерин, соли и бактерии, слизь, продукты изменения
пигментов желчи, придающие калу характерный цвет. При нару­
шении пищеварительных процессов или снижении усвоения пита­
тельных веществ в пищеварительном тракте в кале обнаруживаются
различные количества белков, жиров и углеводов. Калообразование
зависит не только от характера питания человека, но и от его мышеч­
ной и эмоциональной активности. С медицинской точки зрения кал,
его консистенция, химический состав имеют важное диагностичес­
кое значение.
438
Глава 5
Регулируемым, полезным для организма результатом данной фун­
кциональной системы служит содержание определенного количества
каловых масс в прямой кишке.
Ведущим параметром этого результата является прежде всего
механическое раздражение рецепторов растяжения, в большом
количестве представленных в стенке прямой киш ки. Увеличение
калонаполнения приводит к нарастающей стимуляции рецепторов
растяжения прямой кишки. Иногда же процесс дефекации опреде­
ляется раздражением каловыми массами хеморецепторов слизистой
оболочки сигмовидной и прямой кишки. Удержание каловых масс
в прямой кишке и ее опорожнение определяют, в первую очередь,
сфинктеры прямой кишки: внутренний, гладкомышечный, и наруж­
ный, образованный поперечно-полосатой мускулатурой. Внутренний
сфинктер получает тонические влияния от симпатических нейронов
(L,, L2) опосредованно через а-адренорецепторы, а наружный сфин­
ктер иннервируется срамным нервом.
При отсутствии значительного раздражения прямой киш ки кало­
выми массами оба сфинктера находятся в состоянии непрерывного
тонического сокращения.
Ф ункциональная система, определяющая акт дефекации, имеет
внутреннее звено саморегуляции, обеспечивающее формирование
кала в прямой и толстой кишке. Внешнее звено саморегуляции
данной функциональной системы определяет опорожнение прямой
кишки. Внутреннее звено саморегуляции функциональной систе­
мы, определяющей акт дефекации, включает процессы всасывания
остаточных питательных веществ в толстом кишечнике и склеивание
непереваренных остатков пищи слизью, поступающей из верхних
отделов пищеварительного тракта. Значительное место в процессах
саморегуляции формирования каловых масс занимают механизмы
всасывания и выделения воды в толстом кишечнике.
Ф ормирование позыва к деф екации
Возбуждение рецепторов прямой кишки усиливается по мере уве­
личения интенсивности раздражения при увеличении поступления
каловых масс в прямую кишку. Импульсации от этих рецепторов
распространяются по чувствительным волокнам, идущим в составе
задних корешков к спинальному центру дефекации, расположенному
на уровне III—IV поясничных и крестцовых сегментов. За счет этого
повышается тонус симпатических влияний, идущих по подчревным
нервам к внутреннему сфинктеру прямой кишки, и снижается тонус
Висцеральные функции
439
парасимпатических влияний тазовых нервов. Под влиянием симпати­
ческих воздействий внутренний сфинктер прямой кишки закрывает­
ся. Снижение парасимпатического тонуса расслабляет прямую кишку.
В результате создаются условия для заполнения прямой киш ки кало­
выми массами. При достижении определенной критической степени
растяжения прямой киш ки возбуждение из спинального центра дефе­
кации начинает распространяться в восходящем направлении, возбуж­
дая гипоталамические и лимбические структуры головного мозга. На
этой основе формируется эмоционально окрашенный позыв к дефекации. При этом возбуждения подкорковых центров распространяются
до коры головного мозга. За счет возбуждения корковых нейронов и
на основе оценки обстановки и индивидуального опыта формируется
специфичное для каждого вида животных и человека ритуальное пове­
дение, заканчивающееся актом дефекации (рис. 5.5.8).
Акт дефекации возникает на основе принципа доминанты, когда
наиболее значительное в биологическом отношении возбуждение
завладевает большинством синапсов и нейронов головного мозга.
Акт деф екации
Сформированное на основе позыва к дефекации возбуждение,
охватывающее и механизмы произвольной регуляции функций —
пирамидные клетки коры больших полушарий, особенно в области
передней центральной извилины, — распространяется в нисходящем
направлении на спинальные центры дефекации. Вследствие этого
снижается тонус подчревных нервов и повышается тонус парасим­
патических тазовых нервов. В результате расслабляется внутрен­
ний сфинктер прямой киш ки. За счет возбуждения крестцовых
парасимпатических нейронов происходит сокращение мускулатуры
дистальных отделов толстого кишечника и прямой кишки. За счет
произвольных влияний через срамные нервы расслабляется наруж­
ный сфинктер прямой киш ки и каловые массы перистальтическими
движениями толстой и прямой киш ки выделяются наружу.
Акту дефекации способствуют так называемое натуживание,
сокращение мышц брюшного пресса, задержка дыхания, напряже­
ние диафрагмы и мышц тазового дна, а также мышцы, поднимающей
заднепроходный сфинктер. При дефекации происходит опорожнение
не только прямой киш ки, но и за счет активации парасимпатических
влияний — толстого кишечника.
Процесс дефекации регулируется произвольно за счет нисходящих
корковых влияний на подкорковые центры. Произвольная регуляция
440
Глава 5
А
Гипоталамический
центр дефекации
Афферентные
заднекореш ковые нервы
Нисходящая толстая киш ка]
Сигмовидная толстая киш ка
П рямая киш ка
|
и сакральные сегменты
Внутренний сф инкер
Б
П ирамидные
Гипоталамический
дефекации
мезентериальныи
Подчревные
нервы
С рамны е
нервы
Висцеральные функции
441
дефекации формируется у человека в процессе индивидуального
обучения. У ряда животных ритуальное поведение, обеспечивающее
акты мочеиспускания и дефекации, передается по наследству.
Спинальные центры удерживают сфинктеры прямой кишки вне
дефекации в тоническом напряжении. Вышележащие центры дефе­
кации, особенно лимбические и корковые, определяют формирова­
ние позыва к дефекации и ее произвольный характер.
Масса кала взрослого человека составляет около 60—80 г/сут. При
диарее масса кала может увеличиваться до 200 г/сут. Около четверти
массы кала составляют твердые вещества, а одна треть представлена
бактериями толстого кишечника.
Нарушение деф екации
Перерезка спинного мозга выше поясничной области — места,
где расположены спинальные центры дефекации — приводит к тому,
что дефекация становится непроизвольной. В то же время после
перерезки спинного мозга ниже спинального центра дефекации или
при его разрушении происходит паралич сфинктеров прямой кишки.
Заднепроходное отверстие начинает зиять («зияющий анус»), удер­
живание кала при этом становится невозможным.
Непроизвольное расслабление анальных сфинктеров и дефека­
ция могут происходить при сильном отрицательном эмоциональном
возбуждении — например, при страхе (так называемая «медвежья
болезнь»). Дефекацию стимулируют некоторые гормоны — например,
тиреоидин и питуитрин.
Под влиянием ряда токсических веществ наблюдается учащение
позывов к дефекации, возникают поносы. При некоторых заболе­
ваниях происходит извращение нормального соотношения между
позывом и полноценным актом испражнения, возникают так назы­
ваемые ложные позывы — тенезмы.
Психические влияния, особенно длительные эмоциональные
переживания неприятного характера, подавляющие позыв к дефе-
Рис. 5.5.8. Взаимодействие афферентного и эфферентного нейронального
звена в механизмах акта дефекации: А — афферентная интероцептивная
сигнализация о степени растяжения дистальных отделов толстого кишеч­
ника; Б — эфферентные пути, контролирующие моторную деятельность
толстого кишечника и анальных сфинктеров
442
Глава 5
кации, могут существенно расстроить последующее опорожнение
прямой киш ки. Длительные и часто повторяющиеся задержки
позывов к дефекации могут привести к извращению вегетативной
регуляции внутреннего анального сфинктера и развитию запоров.
Поэтому для нормальной дефекации очень важно регулярное, при­
уроченное к одному и тому же времени, опорожнение кишечника,
желательно в утреннее время и сразу же после ощущения позыва
к дефекации.
Поступление пищи в желудок нередко вызывает сокращение пря­
мой киш ки и приводит к позыву и дефекации. Эта реакция чаще
наблюдается у детей и может быть использована для формирования
систематической, приуроченной к месту и времени, дефекации.
5.6. ВЫДЕЛЕНИЕ
Системные механизмы мочеобразования
Процессы выделения из организма различных веществ осущест­
вляются через почки, потовые железы, желудочно-кишечный тракт
(ЖКТ) и легкие. Они направлены на поддержание многих жизненно
важных констант организма: осмотического давления, pH, парциаль­
ного давления дыхательных газов, уровня питательных веществ и, в
частности глюкозы, температуры, артериального давления. Однако
ведущая роль выделительных процессов заключается в выведении
из организма продуктов тканевого метаболизма. Известно, что окис­
ление жиров и углеводов в клетках организма заканчивается обра­
зованием С 0 2 и Н20 , которые могут быть удалены через легкие,
желудочно-кишечный тракт и потовые железы. Белковый обмен
заканчивается образованием ряда азотистых продуктов, таких как
мочевина и мочевая кислота, высокая концентрация которых обла­
дает токсическим действием на метаболические процессы в организ­
ме. Количество продуктов белкового обмена в крови определяется
по величине остаточного азота, которую определяют после осаж­
дения белков плазмы крови. Остаточный азот в норме равняется
200—400 мг/л. Он включает азот мочевины (наибольшая часть, при­
близительно 70%), азот креатинина, креатина, мочевой кислоты,
аминокислот, аммиака, индикана и других веществ.
Комплекс физиологических процессов, направленных на подде­
ржание низкой концентрации продуктов белкового обмена в крови,
443
Висцеральные функции
представляет собой функциональную систему, работающую по при­
нципу саморегуляции (рис. 5.6.1).
Увеличение концентрации продуктов белкового обмена крови
приводит к активации нервных и гуморальных механизмов, усили­
вающих процессы выделения. Основная роль в выделительных про­
цессах принадлежит мочеобразованию и мочевыделению.
Кроме продуктов белкового метаболизма, из организма удаляются
вода и соли, которые постоянно замещаются на новые молекулы,
поступающие при приеме пищи и воды.
Процессы выделения из организма воды и солей играют важную
роль в регуляции осмотического давления крови. Удаление воды и
солей из организма осуществляется главным образом с помощью
мочеобразования. Большое значение имеет потоотделение и выделе­
ние воды и солей через желудочно-кишечный тракт. Кроме того, при
дыхании вода испаряется через легкие.
цнс
Сосудистые
хеморецепторы
Прием белковой пищи
Мочеобразован ие
и мочевыведение
Q
-О
Выделение через
потовые железы
Выделение через
желудочно-кишечный
________ тракт
Э
//
/
Рис. 5.6.1. Функциональная система поддержания концентрации продук­
тов белкового метаболизма (М) в крови
444
Глава 5
Функционально значимые этапы мочевыделения
Процесс образования мочи из крови не может происходить сразу,
так как уловить в крови невысокие концентрации продуктов метабо­
лизма и выделить их из крови в ходе какого-то одного процесса для
организма невозможно. Поэтому процесс образования мочи пред­
ставляет собой непрерывный конвейер, состоящий из нескольких
этапов (рис. 5.6.2).
Структурные единицы почек — нефроны, начинаются капсулой,
продолжаются извитыми канальцами и заканчиваются впадением в
собирательную трубочку (рис. 5.6.3). В каждой почке свыше 1 млн.
нефронов. На каждом участке нефрона происходит определенный
этап мочеобразования.
,еСс**оче^ азова»«**,
---------Осмотическое
сгущение мочи в петлях
нефронов
\
/
Обязательная
реабсорбция и
секреция
в проксимальных
извитых канальцах
Факультативные
реабсорбция
и секреция
в дистальных извитых
канальцах
\
О кончательное
концентрирование
мочи
в собирательных
трубочках
Клубочковая
фильтрация
\
\
1
М очевыделение
/
ч
М очеиспускание
Рис. 5.6.2. Взаимодействие этапов мочеобразования и их включение в фун­
кциональные системы гомеостатического уровня
I Le Qsr
,
» I
a -
I 4 )a v u -C A V
-
„
/
^ . Але* К L;--Ci ^
5.
!'С Н к Л -< Л
^ ' - А<й
%ё м . / ! &
U
**“ " W
ji
»
.... ^ у ч <
t
i ' - t •' < д г
13 ® V е*
<S "tOCe t- f П К C ^ t i ^ t X - A r <"
.
V* t f t t 4 x £ * t t < к ,
В исцеральны е ф ункц ии ^ C t o f u J v G i £•. 7 £ i «.& a
Приносящий сосуд
U
... , „ ол,
1 t c i i ^ t r c t . : _ Cocjcofci
? / г
______4 4 5
Выносящий сосуд
i^ o J
Капсула
^иизагельнан
реабсорбация ]P‘*fO*^y
Д. Фильтрация
100%
\ЗМа+ К+ СГ Н20
Гаминокислоты
витамины
глюкоза
,1 1 l i t
«. %
Факультативная
реабсорбция и секреция
Дистальный
извитой каналец
Проксимальный
С извитой каналец
h
o£®'u''?<8 f
, o«>
15%
:
? I*
NH3 H+ ПАГ
Обязательная
секреция
Н ,0
К NH3 н
Na?
Н2С* - -
^ q ^___- - -►
2
Н20 < - --
Н20 < - - - '
Na
N a+
№
н2о^—
н2о<—
Собирательная
трубочка
Петля
нефрона
(у\щ щ лхсоС
Вторичная моча
Лоханка
Рис. 5.6.3. Процессы мочеобразования, транспорт веществ и количество
мочи (%) при прохождении ее по нефрону
Процесс образования мочи начинается клубочковой фильтрацией,
результатом, которой становится разделение крови на форменные
элементы и белки со стороны крови и жидкую часть плазмы со сто­
роны мочи. Форменные элементы и белки плазмы крови, а также
не профильтровавшаяся в нефроны жидкая часть плазмы уходят от
нефронов по венам. Профильтровавшаяся из первичных капилля­
ров в нефроны жидкая часть плазмы называется первичной мочой.
У человека за сутки образуется в обеих почках 180 л первичной мочи.
Далее происходит уменьшение количества мочи и сгущение ее.
Осуществляется это за счет реабсорбции — обратного всасывания воды и
446
Глава 5
большинства растворенных в ней веществ во вторичные капилляры. При
этом продукты метаболизма остаются ji моче. Сначала происходит обяза­
тельная (облигатная) реабсорбция в диетальмых извитых канальцах нефронов. В ходе ее обратно в кровь поступает до 70% воды и ионов, а полезные
для организма вещества всасываются полностью (см. рис. 5.6.3).
Затем происходит осмотическое сгущение мочи в петлях нефронов.
Результатом этого становится повышение концентрации ионов Na+ в
месте поворота петель нефронов, что необходимо для происходящего
затем окончательного концентрирования мочи в собирательных трубоч­
ках. Из петель нефронов реабсорбируется около 15% воды и ионов Na+.
В нроксимальньв* извитых канальцах происходит факультативная
(необязательная) реабсорбция, в ходе которой в кровь всасывается до
10% воды и ионов Na+. Если в проксимальных извитых канальцах
величина реабсорбции практически ни от чего не зависит и никогда
не меняется, то в дистальных извитых канальцах реабсорбция может
увеличиваться или уменьшаться, в зависимости от потребностей
организма. Гормоны, которые изменяют интенсивность мочеобразования, действуют в этой части нефронов.
В собирательных трубочках происходит окончательная концентра­
ция мочи, в ходе которой факультативной реабсорбции подвергается
около 4% воды. Факультативная и обязательная реабсорбция сочета­
ется с секрецией из крови в мочу дополнительных и некоторых новых
продуктов метаболизма.
В результате реабсорбции и секреции количество первичной мочи
уменьшается в 100 раз. Образуется 1,8 л концентрированной вто­
ричной мочи, которая затем по мочеточникам поступает в'мочевой
пузырь, где и накапливается. Удаление мочи из мочевого пузыря
происходит во время мочеиспускания.
Клубочковая фильтрация
Результатом клубочковой фильтрации является образование из
крови первичной мочи. Образование первичной мочи происходит в
капсуле каждого нефрона. Ж идкая часть крови поступает из капил­
ляров клубочков в капсулы нефронов по многочисленным порам
под действием гидростатического давления крови. Вместе с водой в
капсулы профильтровываются низкомолекулярные вещества: соли,
мочевина, мочевая кислота, глюкоза, аминокислоты, витамины,
микроэлементы, инсулин, креатинин, индикан, уробилин и другие
пигменты. Вещества, имеющие молекулярный вес более 70 тыс. уг­
леродных единиц, не могут пройти через поры между кровеносным
447
Висцеральные функции
капилляром и полостью капсул. Поэтому белки плазмы крови не
проходят в первичную мочу и притягивают к себе воду, уменьша­
ют её фильтрацию. Кроме того, фильтрации препятствует гидро­
статическое давление первичной мочи в капсулах. Следовательно,
эффективное фильтрационное давление равно гидростатическому дав­
лению крови (70 мм рт.ст.) за вычетом онкотического давления крови
(30 мм рт.ст.) и гидростатического давления первичной мочи: 70 —
(30 + 20) = 20 мм рт.ст. (рис. 5.6.4).
Таким образом, состав первичной мочи по содержанию неоргани­
ческих и органических веществ (за исключением крупномолекуляр­
ных) полностью соответствует плазме крови.
Инулин и креатинин не реабсорбируются обратно в кровь, и поэтому
по их концентрации в конечной моче можно судить об интенсивности
Клетки крови
Кровь
Белки крови
Гидростатическое
давление мочи
давление крови
н2о
Ионы
Аминокислоты
Глюкоза
Мочевина
Первичная моча
Рис. 5.6.4. Факторы образования первичной мочи в капсуле нефрона
фильтрации, чем и пользуются в клинических и экспериментальных
исследованиях.
Клубочковая фильтрация за счет саморегуляторных механизмов
обеспечивает постоянное количество первичной мочи. Механизмы
саморегуляции направлены на сохранение параметров, определя­
ющих эффективное фильтрационное давление. Гидростатическое
давление крови в капиллярах клубочков сохраняется постоянным
при изменении системного давления в пределах от 70 до 180 мм рт.ст.
Сохранение постоянного кровяного давления в капиллярах обуслов­
лено сокращением или расслаблением прекапиллярных сфинктеров
при изменении системного давления крови. Происходит миогенная
регуляция сохранения постоянства кровотока без участия нервной
системы и гормонов. Миогенная регуляция хорошо выражена в кор­
ковых нефронах и отсутствует в юкстамедуллярных нефронах, распо­
лагающихся на границе коркового и мозгового вещества почки.
Онкотическое давление крови является жесткой константой орга­
низма. Поэтому в условиях нормы онкотическое давление не изменяет
скорость образования и количество образующейся первичной мочи.
Постоянство гидростатического и онкотического давления крови
определяет неизменность гидростатического давления первичной мочи и,
следовательно, величины эффективного фильтрационного давления.
Если же силы, способствующие образованию мочи, возрастают
(увеличение гидростатического или снижение онкотического давле­
ния крови), то это приводит к увеличению гидростатического давле­
ния первичной мочи и, как следствие, к сохранению на постоянном
уровне скорости клубочковой фильтрации.
Обязательная реабсорбция и секреция в проксимальных
извитых канальцах
Результатом этого этапа является уменьшение на 70% количества пер­
вичной мочи, полное обратное всасывание в кровь полезных для метабо­
лизма веществ и выделение из крови в мочу продуктов метаболизма.
В проксимальных извитых канальцах реабсорбируется 70% воды
и солей. Реабсорбция катионов солей (Na+, К+, Са2+, M g-') проис
ходит активно против градиента концентрации, с использованием
энергии АТФ. Отрицательно заряженные анионы притягиваются
положительно заряженными катионами и поступают из мочи в кровь
пассивно (С1“ и_НС03 — вслед за Na+ и К+; S 0 42", Н Р 0 42- — вслед за
Са2+ и Mg2+). Вода всасывается из первичной мочи в кровь пассивно
вслед за солями по осмотическому градиенту.
ааЛ
k;t (
. Cfc©jvC>c-ЫА tlu^St>cVfe-f
^ .p Y ^ J y .c<'-.и O ii& C c u . , A
(-<
-'<■С&л*
сдих . ,' i f CCj' - s’ о O'©> Э
Висцеральные функции
~1
4 ° .°
a_________________________ 4 4 9
Основной ион, определяющий осмотическое давление, и, следо­
вательно, реабсорбцию воды, — Na+ входит в эпителиальные клетки
пассивно, по градиенту концентрации, а затем выбрасывается актив­
но с другой стороны клетки № +,К+-АТФ-азой. Суммарно на весь
переход Na+ из мочи в кровь затрачивается небольшое количество
энергии, так как разность потенциалов между мочой и кровью равна
только 1 мВ. Это связано с особенностью заряда мембран эпители­
альной клетки. Обращенная в сторону канальца нефрона апикальная
мембрана имеет заряд 69 мВ, а обращенная в сторону кровеносного
капилляра базальная мембрана имеет заряд 70 мВ.
Ионы К+ реабсорбируются активно на апикальной мембране и затем
выходят в кровь за счет диффузии. Механизмы реабсорбции Са2+, Mg2+,
S042-, Н Р042 сходны с механизмами реабсорбции Na+, К+ и СГ.
В проксимальных извитых канальцах полностью обратно всасываются
в кровь глюкоза, аминокислоты, низкомолекулярные белки, витамины,
микроэлементы (см. рис. 5.6.4). Всасывание этих веществ в кровь происхо­
дит в большинстве случаев с помощью облегченной диффузии или актив­
но с затратой энергии макроэргических фосфатов. Облегченная диффузия
заключается в переносе через апикальную мембрану в цитоплазму почеч­
ной эпителиальной клетки веществ совместно с ионами Na+. Из эпите­
лиальной клетки в кровь вещества поступают через базальную мембрану
с помощью диффузии по градиенту концентрации. Реабсорбция этих
веществ может осуществляться пассивно через апикальную и базальную
мембраны эпителиальных клеток при увеличении концентрации этих
веществ в моче после реабсорбции воды из канальцев нефрона.
При определенной концентрации вещества в крови, которая назы­
вается порогом выведения, вещества, названные пороговыми, не могут
полностью реабсорбироваться, и часть профильтровавшихся веществ
оказывается в конечной моче. К пороговым веществам относится
глюкоза, которая в норме (3,8—7,1 ммоль/л в крови) фильтруется, а
затем полностью реабсорбируется. При увеличении ее концентрации в
крови выше значения 7,1 ммоль/л часть глюкозы не успевает реабсор­
бироваться. Нереабсорбировавшаяся глюкоза выделяется с мочой из
организма. Выделение глюкозы с мочой называется глюкозурией.
Реабсорбция в проксимальных извитых канальцах сочетается с сек­
рецией некоторых веществ из крови в мочу. Секреция необходима для
удаления из организма с мочой высокомолекулярных продуктов обмена
веществ, которые не смогли профильтроваться из крови в первичную мочу.
Эпителиальные клетки активно секретируют из крови холин, парааминогиппуровую кислоту, видоизмененные молекулы лекарственных веществ.
450
Глава 5
Кроме того, эпителиальные клетки поглощают из первичной мочи
глютамин и с помощью фермента глютаминазы расщепляют его на
глютаминовую кислоту и аммиак. Затем аммиак выделяется в мочу
и выносится из организма в виде аммонийных солей. Таким обра­
зом, азот распавшегося в организме белка выделяется с мочевиной
и мочевой кислотой за счет фильтрации и в виде аммиака за счет
секреции.
В эпителиальных клетках ферментом карбоангидразой расщеп­
ляется угольная кислота Н 2С 0 3. Ионы Н С 03“ всасываются в кровь
за счет электростатического притяжения их Na+ и К+, что способс­
твует щелочной реакции крови. Ионы Н+ секретируются в мочу и,
соединяясь с профильтровавшимися молекулами Na2H P 0 4, удаля­
ются с мочой в виде NaH2P 0 4. Удаление ионов Н+ из крови с мочой
предотвращает закисление организма. Этим же объясняется кислая
реакция конечной мочи (pH = 4,5—6,5).
Если на входе в проксимальный извитой каналец первичная моча
практически не отличается по составу от жидкой части крови, то на
выходе из этой части нефрона состав мочи становится специфичес­
ким. Пороговые вещества (глюкоза, аминокислоты) перешли обратно
в кровь. К моче добавились высокомолекулярные продукты обмена,
аммиак и ионы Н+, которые сделали ее реакцию кислой, в отличие от
слабощелочной реакции крови. Кроме того, общее количество мочи
значительно уменьшилось.
Постоянство результата обязательной реабсорбции и секреции
в этой части нефронов определяется постоянством количества пер­
вичной мочи, постоянством почечного кровотока и неизменностью
активности ферментов почечного эпителия.
О смотическое сгущ ение мочи в петлях нефронов
В результате прохождения мочи по петлям нефронов создается
высокое осмотическое давление в жидкости, окружающей место
поворота петель нефронов. Это необходимо для окончательного
концентрирования мочи в собирательных трубочках (рис. 5.6.5).
Нисходящее и восходящее колена петель нефронов работают как
единое целое. В восходящем колене происходит активная реабсор­
бция Na+ за счет энергии АТФ, синтезируемой большим количест­
вом расположенных здесь митохондрий. При этом эпителиальные
клетки восходящего колена не пропускают воду (см. рис. 5.6.5А). За
счет выхода Na+ повышается осмотическое давление в жидкости,
окружающей петли нефронов, и из нисходящего колена пассивно
Висцеральные функции
451
Д истальный
извитой каналец
_____>.
..................
Н20
i---
■■-►
N a+
N a+ N a+
н2о<—
N a+ N a+ N a +
N a+ N a+ N a+ N a+
<—
N a+ N a+ N a+ N a+ N a+
300 м оем
~Л
200 моем
*
300 моем
Na+
Н20 * - "
Na+ Na+ Na+
Н20
Na+ Na+ Na+ Na+
H20 < - . .
Н.0
Собирательная
--
трубочка
Na+ Na+ Na+ Na+ Na+ H20 < - - -
1200mocm
1200моем
I
Лоханка
Вторичная моча
Рис. 5.6.5. Реабсорбция воды и ионов Na+ в петле нефрона и окончательное
сгущение мочи в собирательной трубочке
Глава 5
452
выходит вода. Ионы Na+ притягивают к себе воду. Стенки нисходя­
щего отдела нефронов состоят из плоских эпителиальных клеток,
бедных митохондриями и ферментами, поэтому реабсорбция Na+ в
этом отделе не происходит. Место соединения эпителиальных клеток
представляет собой мембрану с порами, через которые по осмотичес­
кому градиенту вода выходит из мочи в межклеточную жидкость и
кровь (см. рис. 5.6.5Б).
Выход воды приводит к увеличению концентрации Na+ в
моче и окружающей петли нефронов жидкости от 0,9% до 3,6%.
Изотоническая моча превращается в гипертоническую. Осмотическое
давление увеличивается от 300 до 1200 мосм/л. Поднимаясь по вос­
ходящему колену петель нефронов, моча теряет ионы Na+ и снова
становится изотонической, а при выходе из петель нефронов даже
гипотонической (0,6% NaCl). Осмотическое давление уменьшается до
200 мосм/л (см. рис.5.6.5В).
Механизм реабсорбции в петлях нефронов известен как противоточно-поворотный множительный механизм (рис.5.6.6). В нисходящем
и восходящем коленах моча течет в разные стороны, то есть имеется
310 моем
300 моем
Умножение
еденичных
механизмов
реабсорбции
Противото
1
бООмОсм
300 моем
610 моем
600 моем
Н20
1200 моем
Поворот
Рис. 5.6.6. Противоточно-поворотный механизм реабсорбции воды и
ионов Na+ в петле нефрона
Висцеральные функции
453
её противоток. Переходя из нисходящего в восходящее колено, моча
делает поворот. На каждом горизонтальном уровне петель нефронов
разница между концентрациями Na+ внутри и снаружи петли нефрона невысокая. Поэтому единичный механизм реабсорбции Na+ и
воды на каждом горизонтальном уровне требует небольшой затра­
ты энергии. При движении мочи по петлям нефронов единичный
механизм реабсорбции умножается, что способствует реабсорбции
значительного количества воды и солей.
Постоянство обязательной реабсорбции в петлях нефронов опре­
деляется одинаковым количеством мочи, поступающей в петлю, пос­
тоянством почечного кровотока и постоянством активности Na+,K+АТФ-азы, реабсорбирующей Na+.
Факультативная реабсорбция и секреция в дистальных
извитых канальцах
Результатом факультативной реабсорбции и секреции в дисталь­
ных извитых канальцах является изменение количество и качество
мочи в зависимости от потребностей организма.
В дистальных извитых канальцах реабсорбируется в среднем 10%
Na+ и воды. Реабсорбция регулируется гормоном надпочечников альдостероном, который активирует № +,К+-АТФ-азу и усиливает реаб­
сорбцию ионов Na+. За ионами Na+ пассивно реабсорбируется вода.
Натрийуретический гормон может ингибировать № +,К+-АТФ-азу, в
результате чего реабсорбция Na+ и воды уменьшается.
По сравнению с проксимальными извитыми канальцами в дис­
тальных извитых канальцах реабсорбция Na+ затруднена и требует
больших энергетических затрат. Это связано с тем, что базальная
часть мембраны эпителия имеет потенциал 70 мВ, а апикальная —
только 20 мВ. Поэтому переход через эпителиальную клетку прохо­
дит против электростатического градиента 50 мВ.
В дистальных извитых канальцах может изменяться активность
реабсорбции ионов К+. При избытке ионов К+ в крови направление
транспорта его может изменяться на противоположное, и он может
пассивно выделяться из крови в мочу.
Реабсорбция других ионов также может регулироваться в этом
участке нефрона. Конкретные механизмы реабсорбции этих веществ
принципиально не отличаются от механизмов реабсорбции их в
проксимальных извитых канальцах.
Кроме ионов К+, в дистальных извитых канальцах секретируется
аммиак, ионы Н+ и парааминогиппуровая кислота.
454
Глава 5
Окончательное концентрирование мочи
в собирательных трубочках
Результатом этого этапа становится выделение в почечную лохан­
ку большего или меньшего количества разбавленной или концентри­
рованной мочи, в зависимости от потребностей организма.
Благодаря работе нескольких петель нефрона, окружающих каж­
дую собирательную трубочку вокруг них создается высокая кон­
центрация ионов Na+ и других солей (см. рис. 5.6.5). Выход воды из
собирательных трубочек происходит пассивно по осмотическому
градиенту. Вода проходит сквозь поры, находящиеся в межклеточном
пространстве.
Реабсорбция воды в собирательных трубочках происходит факуль­
тативно. Антидиуретический гормон (вазопрессин) задней доли
гипофиза активирует белки аквапорины в собирательных трубочках.
Это приводит к усилению проницаемости собирательных трубочек
для воды. Усиление реабсорбции воды сопровождается увеличе­
нием концентрации в моче различных веществ: солей, мочевины,
мочевой кислоты, аммиака. Максимальное концентрирование мочи
происходит при выходе мочи из собирательных трубочек в почечную
лоханку.
В собирательных трубочках происходит диффузия части моче­
вины и других азотосодержащих веществ из конечной мочи в меж­
клеточную жидкость почки. Мочевина и другие вещества пассивно
проникают в петли нефрона, с током мочи проходят по дистальным
извитым канальцам и возвращаются в собирательные трубочки. Так
происходит кругооборот мочевины между собирательными трубоч­
ками и петлями нефрона.
Особенности регуляции, реабсорбции и секреции
в различных отделах нефронов
Процессы облигатной реабсорбции и секреции в проксималь­
ных извитых канальцах и петлях нефронов генетически детерми­
нированы и не поддаются существенным изменениям. Постоянство
количественного и качественного состава мочи после выхода из
проксимальной части нефронов связано в основном с постоянством
количества образующейся первичной мочи в капсулах нефронов и
саморегуляторными механизмами этого образования.
Процессы же реабсорбции и секреции в дистальных извитых
канальцах и собирательных трубочках изменчивы, регулируемы,
факультативны. В них отсутствуют механизмы поддержания коли­
Висцеральные функции
4 55
чественного и качественного состава мочи. За счет внешней регуля­
ции, преимущественно гормональной, изменяется количественный
и качественный состав мочи, что способствует поддержанию ряда
гомеостатических констант организма.
Нервно-гормональная регуляция
мочеобразования
Нервные и гормональные факторы избирательно регулируют
фильтрацию, реабсорбцию и секрецию при образовании мочи в
почках. Эффективность регуляции реабсорбции выше, чем ф иль­
трации. Так, например, при изменении фильтрации на 10% коли­
чество конечной мочи изменится только на 0,1%. При изменении
реабсорбции на 10% диурез изменится в 10 раз. Поэтому большая
часть механизмов регуляции направлена на изменение факульта­
тивной реабсорбции.
Реабсорбция изменяется с помощью изменения функции почечного
эпителия, осуществляющего этот процесс. Ведущая роль в регуляции
реабсорбции принадлежит гормонам. Альдостерон и вазопрессин умень­
шают диурез, а натрийуретический гормон усиливает его (рис.5.6.7).
Важная роль в регуляции мочеобразования принадлежит ренинангиотензиновой системе (рис.5.6.8). При уменьшении осмотического
или артериального давления юкстамедуллярные нефроны, находящие­
ся на границе коркового и мозгового вещества почки (см. рис. 5.6.8А),
вырабатывают гормон ренин, который выделяется в кровь почечных
капилляров. Отсутствие миогенной регуляции постоянства крово­
тока в юкстамедуллярных нефронах способствует постоянной оцен­
ке уровня системного давления крови. В плазме крови находится
белок ангиотензиноген, синтезированный печенью и биологически
неактивный. Ренин способствует превращению ангиотензиногена
в ангиотензин I, обладающий слабой биологической активностью.
Проходя через легкие с током крови, ангиотензин I превращается в
ангиотензин II с помощью вещества конвертина, синтезируемого в
легочной ткани (см. рис. 5.6.8Б). Ангиотензин II вызывает ряд физио­
логических эффектов в организме. Он суживает артериолы, повышая
артериальное давление до нормального уровня. Ангиотензин II уси­
ливает секрецию альдостерона корой надпочечников. Это сопровож­
дается увеличением реабсорбции натрия, уменьшением диуреза и
нормализацией осмотического давления (см. рис. 5.6.8В). Кроме того,
ангиотензин II возбуждает нейроны гипоталамуса, способствуя фор­
мированию мотивации жажды и пищедобывательного поведения,
Глава 5
4 56
ф и льтраци я
Низкое
артериальное
давление
Обязательная
реабсорбация
Г Недостаток 4
^ВОДЫ В K pO B jlj
Na+K+ С1'Н20
юо%
______________
Ьминокислоты (Альдостерон)
• витамины
глюкоза
Факультативная
Na+ СГ Н90 реабсорбция и секреция
И
Ш-
? П
Na+
N i+K+N^Н,0*
- H+ - Na+
н2о+-н,о+--
1
д
Диурез-I
Концентрация Т
мочевины
NH , Н+ ПАГ
Н2°— '
Н20 + - Na+
Гипоталамус
1
Задняя доля
гипофиза
tit ^ '
NH , Н+ ПАГ
Н ,0
Обязательная н2о+----%_
секреция
н20+
/
^
Na+
ii
Диурез I
Концентрация
мочевины
Висцеральные функции
457
что также направлено на нормализацию осмотического давления
крови и артериального давления.
Изменение объема первичной мочи имеет меньшее значение в
регуляции диуреза. Изменение скорости фильтрации в основном
связано с изменением гидростатического давления крови в почечных
клубочках (рис.5.6.9А). Решающая роль в регуляции давления при­
надлежит симпатической нервной системе, которая определяет тонус
почечных артериол. Действие симпатической нервной системы неод­
нозначно. При сужении выносящих из клубочков сосудов давление
увеличивается, что увеличивает фильтрацию (см. рис. 5.6.9Б). При
сужении приносящих в клубочки сосудов давление крови в капил­
лярах уменьшается, что уменьшает количество первичной мочи (см.
рис. 5.6.9В). Симпатической нервной системе принадлежит ведущая
роль в уменьшении диуреза при острой кровопотере и болевом шоке,
которые сопровождаются уменьшением почечного кровотока и оста­
новкой образования мочи, что называется болевой анурией.
Кроме того, различные гормоны и биологически активные вещес­
тва опосредованно могут влиять на почечный диурез. Действие неко­
торых из этих веществ обусловлено изменением почечного кровотока.
К ним относятся сосудосуживающие гормоны: адреналин, серото­
нин, вазопрессин, простагландины и сосудорасширяющие гормоны:
гистамин, кинины, ацетилхолин, простагландины.
Ко второй группе относятся гормоны, опосредованно влияю ­
щие на реабсорбцию и секрецию. К ним относятся секретируемые
в эпифизе аденогломерулотропин и мелатонин, секретируемый в
гипоталамусе кортиколиберин, секретируемый в гипофизе аденокортикотропный гормон и секретируемый в почках медулин.
К третьей группе относятся гормоны, действие которых может
сопровождаться изменением диуреза. Усиленное выделение гормонов
щитовидной железы, соматотропного гормона, половых гормонов
усиливает метаболизм белков. Увеличение концентрации продуктов
обмена белков сопровождается повышением диуреза. Действие глю-
Рис. 5.6.7. Гормональная регуляция мочеобразования: А - уменьшение
диуреза и увеличение факультативной реабсорбции при действии на
нефрон альдостерона или вазопрессина; Б —увеличение диуреза и умень­
шение факультативной реабсорбции при действии на нефрон Na-уретического гормона
Глава 5
ДИК
од*
АД \
х'
Юг пик * f
пн
пл
ф\
Ангиотензиноген
Ренин
Ангиотензин I
Конвертин —*
Ангиотензин II
Альдостерон
Реабсорбция
Na+ |
459
Висцеральные функции
кагона, адреналина и тироксина сопровождается увеличением кон­
центрации глюкозы в крови, что вызывает глюкозурию и увеличение
диуреза. Гормон паращитовидной железы кальцитонин уменьшает
концентрацию кальция в крови, в частности, за счет выделения его
с мочой, что сопровождается увеличением диуреза.
Невыделительные функции почек
Почки обладают выраженной метаболической активностью, прямо
не связанной с их выделительной функцией. В клетках почечного эпи­
телия происходит окисление белков, жиров и углеводов, что влияет на
обмен этих веществ во всем организме. Поступление этих веществ в
клетки происходит как из почечных кровеносных капилляров, так и
из первичной мочи, если эти вещества попадают в нее при фильтра­
ции в капсуле нефрона. Почечный эпителий осуществляет частичное
обезвреживание токсических продуктов обмена веществ в организме и
ядовитых веществ, попавших в организм из внешней среды.
Почки секретируют ряд гормонов. Ренин принимает участие
в работе ренин-агиотензиновой системы. Эритрогенин усиливает
эритропоэз. К инины вызывают уменьшение тонуса периферических
сосудов и уменьшение системного давления крови. Простагландины
регулируют распределение крови между корковым и мозговым вещес­
твом почки. Урокиназа принимает участие в фибринолизе кровяного
сгустка после свертывания крови. Серотонин обладает сосудосужувающим эффектом. Витамин D3 увеличивает реабсорбцию из мочи
ионов К+,Са2+ и Н Р 0 42", при этом Са2+ увеличивает минерализацию
костей.
Мочевыделение
Конечная моча выделяется из собирательных трубочек в почечную
лоханку. Гидростатическое давление мочи в ней увеличивается, и
Рис. 5.6.8. Роль ренин-ангиотензиновой системы в нормализации арте­
риального и осмотического давления крови. А — секреция ренина в юкстагломерулярных (ЮГ) клетках юкстамедуллярных нефронов почек при
уменьшении артериального и осмотического давления крови (АД, ОД);
Б — образование ангиотензина II; В — нормализация АД и ОД крови с
помощью ангиотензана II (К — капсула нефрона; П И К — проксимальный
извитой каналец; ПН — петля нефрона; ДИ К — дистальный извитой кана­
лец; СТ — собирательная трубочка; ПЛ — почечная лоханка)
460
А
Онкотическое
давление крови
(ОД)
t
Гидростатическое
давление мочи
(ДМ)
Первичная моча
и
Первичная моча
Диурез I
Спазм
ДМ
ОД
Первичная моча
f I
Диурез Т
Висцеральные функции
461
часть ее поступает в начальный отдел мочеточника. Установлено, что
при вскрытой брюшной полости моча перемещается от почек к моче­
вому пузырю с помощью медленных перистальтических сокращений
мочеточников. С помощью контрастной рентгенографии показано,
что передвижение мочи по мочеточникам в интактном организме
человека происходит за счет специального механизма, отличного от
перистальтики. Столбик мочи около 10 мл перемещается по моче­
точникам с ускорением. В начале мочеточника скорость небольшая,
а в конце высокая. Механизм увеличения скорости движения мочи
по мочеточникам связывается с величиной забрюшинного давления,
ускоряющего продвижение мочи. Частота выделения порции мочи по
мочеточникам зависит от гидростатического давления мочи в почеч­
ных лоханках и начальных отделах мочеточников. За счет механизмов
местной саморегуляции ускорение или замедление мочевыделения
обеспечивает постоянство гидростатического давления в почечных
лоханках.
Мочеточники входят в мочевой пузырь под острым углом. Это
препятствует обратному току мочи из мочевого пузыря в мочеточни­
ки при наполнении мочевого пузыря.
Диурез и состав мочи
Диурез — это количество мочи, выделившейся из организма за
определенный промежуток времени. Суточное выделение мочи
составляет в среднем 1,8 л, или 70% от принятой за сутки воды. Эта
величина может существенно изменяться при изменении характера
питания, количества выпитой жидкости, при физическом или эмо­
циональном напряжении и изменении условий окружающей среды.
При голодании и во время сна диурез уменьшается, а при охлажде­
нии организма — увеличивается.
М инимальный объем мочи, необходимый для выделения почками
из организма продуктов метаболизма, составляет 500 мл/сут. В связи
Рис. 5.6.9. Нервная регуляция образования первичной мочи: А — при
нормальном тонусе симпатической нервной системы; Б — при умеренном
увеличении тонуса симпатической нервной системы или уменьшении
онкотического давления крови при дефиците белков в пище; В — при
значительном увеличении тонуса симпатической нервной системы при
болевом шоке
462
Глава 5
с этим минимальное количество принятой воды за сутки не должно
быть меньше 800 мл.
Увеличение количества выделяющейся мочи называется полиурией, уменьшение — олигоурией, прекращение выделения мочи — ану­
рией. В норме дневной диурез в 3—4 раза выше ночного. Увеличение
ночного диуреза называется никтурией.
В зависимости от осмотического давления мочи различают опре­
деленные виды изменения диуреза. Осмотический диурез харак­
теризуется большим количеством мочи с высокой концентрацией
осмотически активных веществ. Антидиурез характеризуется малым
объемом мочи с высокой концентрацией веществ. Водным диурезом
называется выделение большого количества мочи с низкой концен­
трацией веществ.
С мочой из организма выводятся продукты окисления белка,
содержащие азот: мочевина, мочевая кислота, аммиак, пуриновые
основания, креатинин, индикан. Выводится также определенная
часть продуктов гниения белков в кишечнике — индол, скатол,
фенол. В моче содержатся органические соединения: соли щавелевой
кислоты, молочная кислота, кетоновые тела. Содержащиеся в моче
пигменты (уробилин, стеркобилин) определяют её желтый цвет.
С мочой выделяется хлористый натрий, хлористый калий, сернокис­
лые и фосфорнокислые соли. Ионы водорода определяют кислую
реакцию мочи (pH = 4,5 —6,5).
В моче отсутствуют углеводы, белки и аминокислоты. Однако эти
молекулы могут кратковременно появляться в моче при физическом
или эмоциональном напряжении организма.
Качественный состав мочи и диурез зависят от скорости накоп­
ления продуктов обмена веществ в крови. Активная реакция мочи
может изменяться в зависимости от качества потребляемой пищи.
При питании растительной пищей, богатой щелочами, реакция мочи
сдвигается в щелочную сторону. При питании мясной пищей или
при интенсивной физической работе наблюдается увеличение кис­
лотности мочи за счет молочной и фосфорной кислот, образующихся
в скелетных мышцах.
М очеиспускание
Результатом мочеиспускания является опорожнение мочевого
пузыря, вследствие чего из организма во внешнюю среду удаляются
продукты обмена веществ. Мочеиспускание состоит из двух этапов:
Висцеральные функции
463
накопление мочи в мочевом пузыре и выделение мочи из организма —
собственно мочеиспускание.
Выделению мочи из мочевого пузыря в мочеиспускательный канал
препятствуют сфинктеры мочевого пузыря и мочеиспускательного
канала. Первый из них имеет гладкомышечное строение, а второй
состоит из поперечнополосатой мускулатуры. Сфинктеры находятся
в тоническом состоянии благодаря возбуждению, приходящему к
ним по симпатическим и соматическим нервам (рис.5.6.10А).
Моча спускается из обеих почек по двум мочеточникам порци­
ями. Увеличение объема мочи в мочевом пузыре приводит к рас­
тяжению гладких мышц его стенок. За счет пластического тонуса
гладких мышц заполнение пузыря мочой происходит без увеличения
тонуса его стенок. Затем растяжение пузыря достигает критической
величины, мышцы начинают сокращаться, что увеличивает гидро­
статическое давление в нем до 15 мм рт.ст. Обычно мочеиспускание
происходит при наполнении мочевого пузыря до 150—300 мл. Чем
выше скорость наполнения мочевого пузыря, тем при меньшем его
объеме начинается мочеиспускание. Это связано с быстрым растя­
жением гладких мышц его стенки, что является для них адекватным
раздражением.
Причиной мочеиспускания является раздражение механорецеп­
торов мочевого пузыря при его наполнении. Сила раздражения
увеличивается при кратковременных сокращениях мышц мочевого
пузыря вследствие их растяжения. Определенная роль принадле­
жит также хеморецепторам мочевого пузыря и мочеточников, кото­
рые возбуждаются высокой концентрацией некоторых веществ в
моче. Возбуждение передается по афферентным парасимпатическим
волокнам в спинной мозг, где происходит торможение симпатичес­
ких и соматических нейронов, обеспечивающих до этого сокраще­
ние сфинктеров мочевого пузыря и мочеиспускательного канала
(см. рис.5.6.10Б).
Одновременно возбуждение достигает сакрального отдела спин­
ного мозга, где находится центр мочеиспускания. Возбуждение
парасимпатических нейронов этого центра передается к мочевому
пузырю по тазовому нерву п. pelvicus, в результате чего расслабляется
сфинктер мочевого пузыря, и сокращение мышц мочевого пузы­
ря приводит к поступлению мочи в мочеиспускательный канал.
Одновременно исчезает возбуждение, проходящее по соматическим
срамным нервам п. pudendus, что расслабляет сфинктер мочеиспус-
Глава 5
464
Спинной мозг
Мочеточники
Поясничные
сегменты L \, L-2
Мочевой пузырь
Крестцовые
сегменты
Внутренний
сфинктеры сжаты
Наружный
Срамной нерв
Б
Мочеиспускательный канал
Тормозной нейрон
Тормозной нейрон
Рецепторы
Сокращение
мочевого пузыря
Расслабление
сфинктеров
Мочеиспускание
Висцеральные функции
465
кательного канала и моча выливается из мочевого пузыря по моче­
испускательному каналу (см. рис.5.6.10В). После мочеиспускания
около 50 мл мочи остается в мочевом пузыре. Таким образом, за счет
рефлекторных механизмов происходит саморегуляция количества
мочи в мочевом пузыре.
Системные механизмы мочеиспускания
Рефлекторные механизмы мочеиспускания входят составной час­
тью в функциональную систему мочеиспускания, системообразую­
щим фактором которой является объем мочевого пузыря от 50 до
300 мл (рис. 5.6.11).
Раздражение механо-, а иногда хеморецепторов мочевого пузыря
вызывает возбуждение, которое через спинной мозг и подкорковые
центры распространяется к коре больших полушарий головного
мозга. При этом возникает биологическая мотивация — позыв к
мочеиспусканию. Кора головного мозга контролирует возбуждение
в спинальном центре мочеиспускания через пирамидальный тракт.
В процессе индивидуального развития и обучения у ребенка выра­
батываются условно-рефлекторные механизмы регуляции мочеис­
пускания. В дальнейшем при позывах к мочеиспусканию на стадии
афферентного синтеза происходит оценка окружающей обстановки.
В соответствии с этическими нормами поведения окружающие фак­
торы оцениваются как запрещающие или санкционирующие акт
мочеиспускания.
Сознательная регуляция необходимости опорожнения мочево­
го пузыря сводится к временной задержке акта мочеиспускания и
поведению, направленному на поиск адекватной обстановки для
мочеиспускания в соответствии с общепринятыми нормами пове­
дения человека в социальном обществе. Последующее нахождение
общепринятого места для мочеиспускания сознательно оценивается
центрами головного мозга, и тормозящее влияние на спинальные
центры мочеиспускания устраняется. За счет непроизвольных меха­
низмов увеличивается тонус парасимпатической нервной системы,
Рис. 5.6.10. Рефлекторные механизмы мочеиспускания: А — нервная регу­
ляция сфинктеров мочевого пузыря при его наполнении; Б — афферент­
ные механизмы перед началом опорожнения мочевого пузыря; В — эф ф е­
рентные механизмы опорожнения мочевого пузыря
466
Глава 5
Рис. 5.6.11. Функциональная система мочевыделения
что приводит к расслаблению сфинктера мочевого пузыря и увеличе­
нию тонуса стенок мочевого пузыря. Уменьшение тонуса сфинктера
мочеиспускательного канала приводит к акту мочеиспускания. При
различных вариантах нарушений связи головного и спинного мозга
сознательная регуляция мочеиспускания утрачивается. При этом
механизмы спинальной регуляции мочеиспускания могут работать
самостоятельно, и по мере накопления мочевого пузыря акт мочеис­
пускания происходит непроизвольно.
При отсутствии патологии поведение является ведущим механиз­
мом регуляции начала мочеиспускания. Меньшее значение имеют
внутренние механизмы регуляции объема мочевого пузыря, завися­
щие от изменения интенсивности мочеобразования в почках. Кроме
того, показано, что при длительном отсутствии мочеиспускания воз­
можно частичное всасывание содержимого мочевого пузыря в кровь
и незначительное концентрирование мочи.
Висцеральные функции
467
Внешние и внутренние механизмы регуляции в конечном итоге
определяют изменение объема мочевого пузыря в пределах физиоло­
гической нормы.
Потоотделение, выделение через легкие
и пищеварительный тракт
Наряду с мочеиспусканием осмотическое давление крови опре­
деляется выделительными процессами через потовые железы, желудочно-кишечный тракт и легкие. Через легкие удаляется 0,5 л воды
в сутки. Атмосферный воздух обычно не полностью насыщен водя­
ными парами. При вдыхании в легкие воздух насыщается водяными
парами за счет испарения воды с поверхности альвеол и выдыхается
из легких, имея 100% влажность. С выдыхаемым воздухом также
могут удаляться пары алкоголя и средств для наркоза: закиси азота,
эфира, хлороформа.
Через потовые железы удаляется до 0,4 л воды в сутки. Одновременно
с потом удаляются соли и продукты метаболизма. По качественному
составу пот сходен с мочой и содержит мочевину, мочевую кислоту,
аммиак, определяющий его неприятный запах. Водородные ионы
создают кислую реакцию пота. Однако концентрация растворенных
в поте веществ значительно ниже, чем в моче, за счет отсутствия в
потовых железах механизма реабсорбции и сгущения.
Потовые железы располагаются во всех областях кожной поверх­
ности тела, но больше всего их в подмышечных впадинах, ладонях и
ступнях. Потоотделение усиливается при возбуждении симпатичес­
кой нервной системы.
Известно, что в нейроорганных синапсах симпатической нервной
системы выделяется норадреналин. Особенностью иннервации пото­
вых желез является то, что постганглионарные волокна симпатичес­
кой нервной системы выделяют ацетилхолин, что имеет определен­
ное фармакологическое и клиническое значение.
Небольшое количество воды (0,1л/сут) выделяется из прямой
кишки при дефекации. Выделительная функция желудочно-кишечного тракта (ЖКТ) не сводится только к удалению непереваренных
продуктов пищи и воды. При прохождении химуса, а затем фекалий
по пищеварительному тракту в полость ЖКТ выделяются из крови
мочевина, мочевая кислота и другие продукты обмена. Выделительная
функция эпителиальных клеток ЖКТ начинается в желудке и про­
должается в тонком и толстом кишечнике. С желчью, образуемой в
печени, удаляются продукты расщепления гемоглобина: билирубин
Глава 5
468
и биливердин, которые превращаются в кишечнике в стеркобилин и
уробилин. Большая часть этих пигментов удаляется с калом, опреде­
ляя его коричневый цвет. Меньшая часть всасывается в кровь и уда­
ляется через почки с мочой, определяя желтый цвет мочи.
5.7. РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГО ОБМЕНА
Функциональная система,
поддерживающая постоянство осмотического
давления крови
Регуляторные механизмы, определяющие осмотическое давление
в тканях организма, объединяются в динамическую саморегулирую­
щуюся организацию, все составные элементы которой содействуют
поддержания осмотического давления крови на постоянном уровне
7,6-8,1 атм., 768-818 кПа, 295—310 мосм/л (рис. 5.7.1). Составной час­
тью осмотического давления является онкотическое давление, равное
0,03—0,04 атм., 23—30 мм рт.ст., 3,0—4,0кП а, 1,3—1,8 мосм/л.
Осмотическое и онкотическое давление имеют существенное зна­
чение для жизнедеятельности органов, тканей и всего организма.
Величины осмотического и онкотического давления удерживаются
в организме в пределах узкого диапазона, что обеспечивает неиз­
менность метаболических процессов в тканях. Отклонение этих
параметров от нормального уровня опасно для жизнедеятельности
организма.
Осмотическое и онкотическое давление как результат
функциональной системы
Результатом деятельности функциональной системы является
определенное соотношение воды и растворенных в ней веществ в
организме.
Осмотическое давление — сила, вызывающая переход воды через
полупроницаемую мембрану в сторону большей концентрации рас-
Рис. 5.7.1. Функциональная система, поддерживающая оптимальное для
метаболизма осмотическое давление крови: А — при повышении осмоти­
ческого давления; Б — при уменьшении осмотического давления
469
Висцеральные функции
Прием Н 2 О
Дегидратация
коллоидов
Мочеобразование |
Потоотделение
а
I
-о
Испарение
при дыхании
о
Выделение воды
через пищеварительный тракт
Выход крови из
депо______
/ /
ядра гипота
Прием солей
Центр солевого
аппетита
Гидратация
коллоидов
Мочеобразование |
Потоотделение
t
Испарение
при дыхании
Выделение воды ,
через пищевари- |
тельный тракт
Передние
ядра
гипоталамуса
Депонирование
_____ крови_____
О
-►о
/
Осмои волюморецепторы
творенного в воде вещества. Осмотическое давление имеется только
на границе двух жидких сред^разделенных полупроницаемой мемб­
раной, через которую может осуществляться движение воды \
к
рая не пропускает растворенные в воде вещества. Движение го д а по
осмотическому градиенту от меньшей концентрации растворенного'хЗ
вещества к большей называется осмосом. Разница давлений может
играть существенную роль в обмене воды между желудочно-кишеч­
ным трактом и кровью, между кровью и межклеточной жидкостью,
между межклеточной и внутриклеточной жидкостями, между кровью
и мочой. Осмотическое и онкотическое давление не связаны с пар­
циальным давлением дыхательных газов и гидростатическим дав­
лением крови и межтканевой жидкости, что обусловлено различной
физической природой этих давлений.
Современной единицей измерения осмотического давления био­
логических жидкостей является миллиосмоль. 1 осмоль (осм) соот­
ветствует осмотическому давлению, создаваемому 1 грамм-молекулой
вещества, растворенного в 1 л воды. Осмотическое давление 1 осм/л
соответствует давлению 22,4 атм. 1 осмоль равняется 1000 мйллиосмолей (моем).
В первую очередь величину осмотического давления исследуют
в артериальной и венозной крови, что обусловлено важностью этих
параметров для величины давления в тканях и простотой взятия
крови на анализ.
Осмотическое давление артериальной крови равно 300 моем.
Большая часть этого давления обусловлена гидратными оболочка­
ми вокруг катионов (140 моем) и анионов (130 моем). Ионы натрия,
хлора и бикарбонатов играют основную роль в создании этой части
давления. Меньшая часть давления (30 моем) обусловлена неэлектро­
литами (глюкозой, мочевиной, холестерином, белками).
За счет поступления в кровь продуктов метаболизма венозная
кровь имеет более высокое осмотическое давление. Его величина
колеблются от 310 до 315 мосм/л в покое, а при напряжении организ­
ма увеличиваеться до 330 мосм/л.
Онкотическое давление определяет силу, с которой белки плазмы
крови притягивают к себе воду. В качестве единиц измерения онкотйческого давления чаще используют мм рт.ст. Величина онкотического давления (1,3—1,8 мосм/л или 25—30 мм Рт.ст.Гнамного меньше,
чем величина осмотического давления. Несмотря на это, онкотичес­
кое давление играет большую роль в постоянном обмене воды между
кровью и тканями и в процессе мочеобразования в почках.
Висцеральные функции
471
Величина осмотического давления может изменяться при изме­
нении интенсивности метаболизма, при усиленной работе органов,
при изменении количества и состава пищи, при изменении условий
окружающей среды. Кровь может стать гипер- или гипоосмотической.
Изменение величины осмотического давления крови возбуждает
осморецепторы,' Имеющиеся в различных органах и тканях организма.
Скопление этих рецепторов имеется в языке, кишечнике, селезенке
и левом предсердии. Наличие таких рецепторов подтверждено экс­
периментально. Так, было обнаружено, что при пропускании гипер­
тонического раствора через селезенку экспериментальные животные
увеличивают прием воды.
Одновременно с изменением осмотического давления может изме­
ниться объем циркулирующей крови, что возбуждает волюморецепторы
(рецепторы объема) сосудов. Основное скопление волюморецепторов,
называемых также барорецепторами, находится в дуге аорты и синокаротидной области. В венозной части сосудистого русла основное место
расположения волюморецепторов находится в правом предсердии.
От осмо- и волюморецепторов возбуждение передается по аффе­
рентным нервным волокнам в центральную нервную систему (ЦНС).
Передача информации об изменении осмотического давления проис­
ходит также гуморальным путем при протекании крови через мозг.
П оведенческие механизмы регуляции осмотического
давления
Ж аж да и солевая мотивация
Ведущая роль в регуляции осмотического давления принадлежит
ядрам переднего гипоталамуса. Возбуждение нейронов этих ядер зави­
сит от афферентного притока информации от осмо- и волюморецеп­
торов, а также от осмотического состояния притекающей к гипотала­
мусу крови. В супраоптическом и паравентрикулярном ядрах переднего
гипоталамуса обнаружены нейроны, чувствительные к осмотическому
давлению крови, протекающей в гипоталамусе по капиллярам. Такие
нейроны были названы центральными осморецепторами. Особенность
их строения — наличие в цитоплазме вакуоли, размер которой изме­
няется при изменении осмотического давления крови (рис. 5.7.2).
При изоосмолярном состоянии крови вакуоли имеют средний
размер, и нейроны генерируют потенциалы действия с оптимальной
частотой. При протекании по капиллярам гиперосмотической крови
размер вакуолей уменьшается, так как вода выходит из вакуолей в
4 72
Глава 5
Кора
£
Подкор­
ковые
центры
Ретикулярная
формация
&
Нейрон-осморецептор
гипоталамуса
Вакуоль
(Н 2 О, ионы)
Т
-ОэЛ
.
Рис. 5.7.2. Динамика работы центральных осморецепторов при изменении
осмотического давления крови: А —при нормальном осмотическом давле­
нии; Б — при увеличении осмотического давления; В — при уменьшении
осмотического давления
Висцеральные функции
4 73
кровь. Это приводит к уменьшению частоты импульсов, возникаю­
щих в нейронах. В случае протекания около нейронов гипоосмотической крови растворенные в вакуолях вещества притягивают к себе
воду из плазмы крови, и размер вакуолей увеличивается. Частота
генерируемых при этом потенциалов действия увеличивается.
От центральных осморецепторов возбуждение проводится к дру­
гим ядрам гипоталамуса, подкорковым нейронам в лимбической
системе и ретикулярной формации мозга, эпифизу и доходит до коры
больших полушарий.
Внутренние исполнительные механизмы регуляции осмотического
давления зависят, прежде всего, от секреции вазопрессина в супраоптическом и паравентрикулярном ядрах гипоталамуса. Немаловажно
и изменение интенсивности секреции аденогломерулотропина и мела­
тонина в эпифизе (рис. 5.7.3).
При недостатке жидкости в организме и увеличении осмотическо­
го давления активация лимбических структур мозга и коры больших
полушарий приводит к формированию мотивации жажды , характе­
ризующейся эмоционально окрашенным желанием удовлетворить
питьевую потребность (см. рис. 5.7.1 А). При снижении осмотического
давления возбуждение структур мозга приводит к возникновению
солевой мотивации, направленной на восполнение недостатка солей
в организме (см. рис. 5.7.1 Б). Соответствующая мотивация определяет
внешнее звено регуляции постоянства осмотического давления крови
с помощью формирования поведения организма, направленного на
потребление воды или солей.
Одновременное уменьшение количества воды и солей в организме
приводит к появлению обеих мотиваций. Так, например, при работе
в горячих цехах обильное потоотделение сопровождается у рабочих
потерей не только воды, но и солей. В этих случаях для поддержания
постоянства осмотического давления рекомендуется пить минераль­
ную воду, содержащую определенный набор солей.
Возникновение жажды связано с сенсорными и метаболическими
механизмами. При уменьшении количества воды в организме рефлекторно уменьшается секреция пищеварительных желез и, в частности,
слюнных желез. Уменьшение слюноотделения вызывает сухость рото­
вой полости и глотки. Раздражение при этом осморецепторов языка и
слизистой глотки является ведущей причиной сенсорных механизмов
жажды. При этом осмотическое давление крови остается неизменным.
Дальнейшая потеря воды организмом приводит к изменению осмо­
тического давления крови, а в дальнейшем и тканей. Возбуждение
Глава 5
Передние ядра
N a -уретический
Супраоптическое,
паравентрикулярное
ядра
— ► — Возбуждение
__ Торможение
Висцеральные функции
4 75
осморецепторов сосудов, селезенки и тканей при их дегидратации
лежит в основе метаболических механизмов жажды.
Активация ренин-ангиотензиновой системы способствует возник­
новению жажды за счет возбуждения нейронов гипоталамуса высо­
кой концентрацией ангиотензина II в крови.
Сенсорные и метаболические механизмы жажды представляют
собой два последовательных этапа. Особенность сенсорного этапа —
возникновение в ходе его метаболических механизмов жажды. Уже
в начале возникновения сухости ротовой полости регуляторные
процессы в организме направлены на сохранение воды в организме.
Уменьшается диурез и потоотделение, увеличивается объем депони­
рованной крови. Вследствие этого происходит возбуждение осмо- и
волюморецепторов в различных частях организма. Следовательно,
так называемый метаболический компонент сенсорных механизмов
жажды возникает уже в начале появления дефицита воды в орга­
низме.
Исполнительные механизмы регуляции осмотического
давления
Внешним звеном регуляции осмотического давления является
поведение, направленное на потребление организмом воды и солей
из внешней среды. Удовлетворение жажды при приеме воды обус­
ловлено сенсорными и метаболическими механизмами. Сенсорные
механизмы удовлетворения связаны с устранением сухости в ротовой
полости при приеме воды и действием воды на осморецепторы желуд­
ка. Афферентное возбуждение от пищеварительного тракта тормозит
центры жажды в головном мозге.
После всасывания воды из пищеварительного тракта в кровь возни­
кает истинное или метаболическое насыщение, связанное с нормали­
зацией осмотического давления и количеством жидкости в организме.
Однако уже до всасывания воды из желудочно-кишечного тракта
осмотическое давление крови и количество циркулирующей крови
нормализуется за счет выхода крови из депо и перераспределения воды
Рис. 5.7.3. Нервно-гормональные механизмы регуляции осмотическо­
го давления крови и мочеобразования: А — афферентные процессы;
Б — эффекторные процессы (А К Т Г — аденокортикотропный гормон;
A ll —ангеотензин II)
476
Глава 5
между кровью и коллоидами тканей. Этот процесс называется метабо­
лическим компонентом сенсорного этапа удовлетворения жажды.
Внутренние механизмы регуляции связаны с изменением тонуса
вегетативной нервной системы и секрецией эндокринных желез.
Ведущая роль в регуляции осмотического давления принадлежит
гормонам вазопрессину, натрийуретическому гормону и ренин-ангиотензиновой системе.
Вазопрессин регулирует осмотическое давление крови при изме­
нении объема воды в крови. Выработанный нейронами гипота­
ламуса вазопрессин накапливается в задней доле гипофиза. При
недостатке в организме воды он выделяется в кровь и увеличивает
проницаемость для воды собирательных трубочек нефронов за счет
увеличения в них количества открытых водных каналов (аквапоринов). Реабсорбция воды снижает диурез и уменьшает потерю воды
организмом. Осмотическое давление уменьшается (рис. 5.7.4А). При
избытке воды в организме вазопрессин не секретируется, диурез уве­
личивается и осмотическое давление повышается (см. рис. 5.7.4Б).
Ренин-ангиотензиновая система регулирует осмотическое давление
крови при недостатке солей в организме (рис. 5.7.5А). Непосредственной
причиной усиления секреции ренина является снижение артериально­
го давления. Система увеличивает секрецию альдостерона, возбуждает
центр жажды в гипоталамусе и вызывает сужение сосудов, в результате
чего осмотическое и артериальное давление крови нормализуется.
Н атрийуретический гормон регулирует осмотическое дав­
ление крови при избытке солей в организме (см. рис. 5.7.5Б).
Непосредственной причиной секреции гормона является повышение
венозного давления крови. При увеличении выработки натрийуретического гормона снижается активность ренин-ангиотензиновой
системы. Гормон уменьшает факультативную реабсорбцию натрия в
нефроне и секрецию альдостерона, в результате чего осмотическое
давление крови нормализуется, и за счет увеличения диуреза веноз­
ное давление крови возвращается к исходному уровню.
С помощью вегетативной и гормональной регуляции происходит изме­
нение процессов всасывания воды и солей из пищеварительного трак­
та, мочеобразования, мочевыделения, потоотделения, выделения воды
через легкие и пищеварительный тракт. Изменяется состояние коллои­
дов в тканях, депонирование крови и скорость кровообращения.
Кроме того, в организме имеются местные механизмы саморегуля­
ции количества воды и солей в крови и тканях. При незначительном
изменении осмотического давления возможно поддержание давления
477
Висцеральные функции
Недостаток
воды
I
Осмотическое давление увеличивается
Б
Рис. 5.7.4. Роль вазопрессина в саморегуляции осмотического давления
крови: А — при увеличении осмотического давления; Б — при уменьшении
осмотического давления
Глава 5
4 78
Р е а б с о р б ц и я N a + Ь*
А н г и о т е н зи н о ге н
А
-►Ренин
А н ги о т е н зи н I
К о н в ер т и н
А н г и о т е н зи н I
О смотическое
давление крови
М асса
крови
Артериальное i
давление
’
А н ги о т е н зи н о ге н
-► Ренин
Осмотическое |
давление крови
А н г и о т е н зи н I
Г
К онверти н - - ■
4
Y
А н г и о т е н зи н II
Р е а б с о р б ц и я N a + {-► Диурез f
А л ьдостерон
Т о р м о ж е и и е ^^ Ч
Х ^ е н т р а ж аж ды ^/
Артериальное
давление I
N a-уретический гормон
Венозное
давление
Висцеральные функции
4 79
на постоянном уровне за счет изменения коллоидного состояния бел­
ков плазмы крови и перераспределения ионов между эритроцитами и
плазмой крови. Показано, что такая способность имеется у взятой на
анализ крови. Реальное значение этого механизма в живом организме
второстепенно по сравнению с другими механизмами внутреннего
звена регуляции осмотического давления.
Несмотря на многообразие, комплексность и важность внутренних
механизмов регуляции осмотического давления, их недостаточно для
поддержания жизнедеятельности организма в течение длительного
времени без приема воды. Известно, что в условиях отсутствия внешне­
го звена регуляции осмотического давления крови, то есть при отсутс­
твии приема воды человек может существовать только 2—3 суток.
Водный баланс организма
Вода в организме является той средой, в которой происходит
большая часть биохимических реакций и обменных процессов. Вода
также выполняет транспортную функцию. В ней происходит движе­
ние ионов, питательных веществ, биологически активных веществ,
в частности, гормонов и медиаторов, промежуточных и конечных
продуктов тканевого метаболизма.
Количество воды в организме имеет тенденцию к постоянному
уменьшению за счет мочевыделения, испарения при потоотделении и
дыхании и выделения фекалий из желудочно-кишечного тракта. Вместе
с водой из организма удаляются растворенные в ней вещества. Большое
значение имеет выведение с водой продуктов метаболизма. Кроме того,
с водой выводятся ионы и частично могут выходить полезные для орга­
низма вещества (глюкоза, аминокислоты, гормоны, витамины).
Все эти процессы могут изменять осмотическое давление в жид­
ких средах организма. Для поддержания постоянного уровня осмоти­
ческого давления необходимо сохранение водного баланса организма,
то есть сохранение равенства между количеством поступающей в
организм и выделяемой из организма воды.
Рис. 5.7.5. Роль ренин-ангиотензиновой системы, альдестерона и Na-ypeтического гормона в саморегуляции осмотического давления, объема
крови и артериального давления: А — при уменьшении осмотического и
артериального давления; Б — при увеличении осмотического и артериаль­
ного давления
Глава 5
480
В среднем за сутки в организм человека массой 70 кг поступает
с питьем и пищей 2,5 л воды (рис. 5.7.6). Кроме того, при окислении
питательных веществ в тканях организма образуется метаболическая
или конституционная вода в объеме 0,3 л/сут. Наибольшее количест­
во воды образуется из жиров (1,09 мл из 1 г) и меньшее из углеводов —
0,6 мл/г и белков — 0,44 мл/г. Поэтому на потребление воды влияет
характер питания. При углеводной или углеводно-жировой диете
потребность в воде снижается, а при белковой диете — повышается.
Следовательно, общее количество поступающей в организм воды
приблизительно равно 2,8 л/сут. Такое же количество воды выделя­
ется из организма с мочой, через легкие, с потом и калом. 8,0 л/сут
воды обменивается между кровью и пищеварительными железами.
Выделение
Прием
воды
волы
М оча 1,8 л
Выдыхаемый
воздух 0,5 л
Потоотделение 0,4 л
Д еф екация 0,1 л
Ж идкость 1,5 л
Вода с пищ ей 1 л
х"
'
Слю на 1,5 л
** - х
Ж елудочный сок 2,0 л
4
С ок поджелудочной железы 1,5 л
Ж елчь 0,5 л
"^
К иш ечный сок 2,5 л
„>
Рис. 5.7.6. Средний объем поступающей, циркулирующей и выделяющей­
ся воды из организма за сутки
Висцеральные функции
481
Следовательно, из пищеварительного тракта в кровь всасывается
Ю,5л/сут (2,5л/сут при приеме пищи и воды и 8л/сут при всасыва­
нии воды в кровь из пищеварительных соков).
Функциональная система объема крови
Ф ункциональная система, поддерживающая оптимальный для
метаболизма объем крови, — динамическая саморегулирующаяся
организация, все составные элементы которой содействуют подде­
ржанию объема крови на постоянном уровне: 4—6 л, 1/13 массы тела,
60,5—84,7 м л/кг (рис. 5.7.7). Эти параметры способствуют поддержа­
нию оптимального уровня метаболизма в организме. Изменение
объема крови воспринимается волюмо- и барорецепторами сосудов.
Информация об объеме крови нервным и гуморальным путем пере­
дается в нервный центр в гипоталамусе и доходит до коры больших
полушарий. Переработав полученную информацию, нервный центр
возвращает объем крови к исходному уровню с помощью поведен­
ческой, вегетативной и гормональной регуляции. Поведение связано
с приемом воды (см. рис. 5.7.7А) или воздержанием от излишнего ее
потребления (см. рис. 5.7.7Б). Вегетативная и гуморальная регуляции
направлены на перераспределение воды между кровью, внеклеточным
и внутриклеточным пространством, регионарное перераспределение
крови и ее депонирование, изменение мочеобразования, потоотделе­
ния, испарения при дыхании, выделение воды через пищеваритель­
ный тракт, изменение сосудистого тонуса и работы сердца.
Обмен воды м еж ду кровью и тканями организма
Между кровью тканевых капилляров и тканями происходит посто­
янный обмен воды, не поддающийся количественному учету. Обмен
воды между кровью и тканью определяется разностью гидростатичес­
кого и онкотического давления между кровью и межтканевой жид­
костью (рис. 5.7.8). Фильтрация воды из крови происходит в артери­
альной части тканевых капилляров. Гидростатическое давление крови
(40 мм рт.ст.) способствует, а онкотическое давление крови (30 мм рт.ст.)
препятствует выходу воды из крови. Вышедшая из крови вода поступа­
ет в тканевые клетки и частично (0,02%) в лимфатические капилляры.
В венозной части капилляров происходит осмос воды из ткани в кровь.
Сосудистое сопротивление уменьшает гидростатическое давление
крови с 40 до 20 мм рт.ст. по ходу капилляра. Онкотическое давление
(30 мм рт.ст.) способствует, а гидростатическое давление (20 мм рт.ст.)
препятствует вхождению воды обратно в кровь.
Глава 5
482
Прием воды
|
*■
Увеличение воды
в тканях
Гидратация коллоидов
О I Баро-,
Мочеобразование |
волюмо-.
Потоотделение
|
Испарение при
дыхании
t
рецеп­
торы
/ /
Выведение воды
через пищеваритель- 1
ный тракт
Делонирование
крови
Прием
воды
Возбуждение
центра жажды
Уменьшение воды
в тканях
Дегидратация
коллоидов
Мочеобразование |
Супраоптическое
и паравентрикулярное
ядра
гипоталамуса
Потоотделение
I
Испарение при
дыхании
ТУ
Выведение воды через
пищ еварительный I
________тракт_______ t
Выход крови из депо
Рис. 5.7.7. Функциональная система, поддерживающая оптимальный для мета­
болизма объем крови: А — при гипергидратации; Б — при гипогидратации.
Висцеральные функции
483
| лк
Oi
9
10
Cfra
' Д = 40
ОД = 30
тк
Рис. 5.7.8. Обмен воды между кровью, интерстициальной жидкостью и
лимфой: ТК - тканевые капилляры; ЛК — лимфатические капилляры:
ГД — гидростатическое давление крови; ОД — онкотическое давление
крови. (Давление указано в мм рт.ст.)
Обмен воды м еж ду внеклеточной и внутриклеточной
жидкостями тканей организма
Общее количество воды в организме человека равно 30—50 л,
что составляет 60% массы тела. Большая часть воды (40% массы
тела) находится в клетках. Внутриклеточная вода частично нахо­
дится в свободном состоянии, частично связана с коллоидами
цитоплазмы.
15% воды находится в межклеточных пространствах. В число
этих пространств входят также объемы внутриполостных жидкостей
(лимфа, спинномозговая жидкость, внутриглазная жидкость, перии эндолимфа, внутриплевральная и внутрибрюшная жидкости). 5%
воды входит в плазму крови (содержимое клеток крови относят к
внутриклеточной жидкости). Плазма крови, внутриполостные жид­
кости и межклеточная жидкость составляют внеклеточную жидкость
организма, масса которой равняется 20% массы тела.
Перераспределение воды между вне- и внутриклеточными про­
странствами зависит от действия осмотических сил. Если осмотичес­
кое давление жидкости какого-либо из этих пространств изменяется,
вода немедленно переходит из одного пространства в другое, и уста­
навливается новое осмотическое равновесие. Вода всегда перемеща­
ется в сторону более высокого осмотического давления. При этом
объем водных пространств изменяется (рис. 5.7.9).
Глава 5
484
н2о
+
ИОНЫ
Н.О + ионы
Н ,0 + ионы
А
Внутри­
клеточная
вода
40%
Вне­
клеточная
вода
15%
Е
Н ,0
Н ,0
Кровь
Ч> веса тела
ж
Г
Ионы
Ионы
Рис. 5.7.9. Объем внеклеточной и внутриклеточной жидкости при изме­
нении воды и ионов в крови: А — норма; Б — изотоническая гипергид­
ратация; В — гипотоническая гипергидратация; Г — гипертоническая
гипергидратация; Д — изотоническая дегидратация; Е — гипертоническая
дегидратация; Ж — гипотоническая дегидратация
Висцеральные функции
485
Если при избытке или недостатке воды и солей жидкости организма
остаются изотоническими, то изменяется объем только внеклеточного
пространства и, в частности, плазмы крови. При этом объем внутрик­
леточной воды не меняется, и функции клеток не нарушаются.
При изменении осмотического давления плазмы крови изменя­
ется объем как внеклеточной, так и внутриклеточной жидкости, что
может вызвать различные нарушения жизнедеятельности клеток.
Таким образом, решающий фактор для обмена воды между вне- и
внутриклеточными пространствами — разность осмотического давле­
ния в этих пространствах. В отличие от этого ведущая роль в обмене
воды между кровью и межклеточными пространствами принадлежит
разности гидростатического и онкотического давлений крови и меж­
клеточной жидкости в области тканевых капилляров.
5 .8. ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
Обмен веществ и энергии (метаболизм) — основное свойство
живых существ. В процессе обмена питательные вещества превра­
щаются в собственные компоненты тканей и конечные продукты
метаболизма. Эти процессы сопровождаются поглощением и выделе­
нием энергии. Обмен веществ определяет важный физиологический
процесс — формирование метаболических потребностей организма.
Обмен веществ
Обмен веществ в организме подробно описан в курсе биохимии.
Здесь будут рассмотрены только некоторые важнейшие физиологи­
ческие стороны обмена веществ. Питательные вещества, поступа­
ющие в организм, несут пластическую и энергетическую нагрузку.
Пластическую роль в организме, связанную с построением тканей,
играют в основном белки. Энергетическую роль играют углеводы, жиры
и, в незначительной степени, белки.
Потребности в питательных веществах варьируют в зависимости
от генетической конституции субъектов, размеров их тела, возраста,
пола, активности желез внутренней секреции, физической актив­
ности, репродуктивной функции и т.д. Для нормального обмена
веществ, кроме белков, жиров и углеводов, абсолютно необходимы
вода, минеральные соли, микроэлементы и витамины.
Средняя суточная потребность в белке — 80-100 г, углеводах —
500 г, жирах — 70—80 г.
486
Глава 5
При всех видоизменениях поступающих в организм веществ в
нормальном организме общее количество потребленных веществ
равно выделяемому.
Обмен углеводов
Обмен углеводов представляет собой систему непрерывных про­
цессов образования, транспорта, поддержания на оптимальном уров­
не концентрации в крови и распада углеводов. Углеводы человек
получает с пищей в виде сахаров, растительного полисахарида —
крахмала, животного полисахарида — гликогена, содержащегося
главным образом в мышцах и печени. Группу углеводов составляют
моносахариды — глюкоза, фруктоза, лактоза, пентоза и другие соеди­
нения; дисахариды, состоящие из двух моносахаридов (например,
сахароза, которая состоит из глюкозы и фруктозы); полисахариды, в
том числе крахмал, клетчатка, гиалуроновая кислота, гепарин.
В процессе переваривания в Ж КТ всех видов углеводов в конце
концов все они превращаются в глюкозу, содержание которой в
крови — жизненно важная константа организма. Углеводы служат
важнейшим источником энергии, а также необходимы для пласти­
ческих функций. При полном окислении 1 г углеводов образуется
4,1 ккал тепла. Концентрация глюкозы в крови в норме находится
в пределах 3,7—6,1 ммоль/л.
Глюкоза поступает в клетки через клеточную мембрану посредством
облегченной диффузии с помощью белка-переносчика. Инсулин усили­
вает транспорт глюкозы, особенно через мембраны клеток мышечной
ткани и печени. Поступившая в клетки глюкоза фосфорилируется
ферментами — глюкокиназой или гексакиназой. Под влиянием энзи­
мов цитоплазмы глюкоза превращается в пировиноградную кислоту
в процессе аэробного гликолиза. Далее пируват превращается в ацетилкоэнзим А, который далее поступает в митохондрии.
Молекулы АТФ, образовавшиеся в ходе гликолиза и окислительно­
го фосфорилирования, используются потом для обеспечения энерги­
ей всех внутренних метаболических реакций. В клетке молекула АТФ
быстро расходуется (в течение одной минуты после ее образования),
и запасания АТФ не происходит. Поэтому в организме синтезируется
креатинфосфат, а его расщепление сопровождается высвобождением
энергии для ресинтеза АТФ. Благодаря этому концентрация АТФ
поддерживается на относительно постоянном уровне. Это обеспечи­
вает «рывковую» мышечную активность.
Висцеральные функции
487
Функции углеводов
Углеводы относятся к быстро мобилизуемым энергетическим суб­
стратам, являются основным источником энергии при интенсивной
мышечной нагрузке и эмоциональном возбуждении. Особенно важны
углеводы как источник энергии для ЦНС, в частности, для головного
мозга. Многие клетки способны депонировать в небольших количес­
твах гранулы гликогена, более длительное депонирование гликогена
происходит в клетках печени и мышцах.
Углеводы могут также превращаться в липиды (липогенез). Их плас­
тическая функция включает участие углеводов в синтезе гликопро­
теинов, гликолипидов, промежуточные продукты окисления — пентозы — входят в состав нуклеотидов и нуклеиновых кислот. Глюкоза
необходима для синтеза некоторых аминокислот, синтеза и окисления
липидов, полисахаридов. Организм человека нуждается в одном из про­
изводных углеводов — аскорбиновой кислоте (витамин С), которая не
синтезируется в организме человека. Глюкоза участвует в поддержании
осмотического давления крови. При снижении уровня глюкозы в крови
возникает состояние гипогликемии, проявляющееся в виде общей сла­
бости и быстрой утомляемости. При снижении уровня глюкозы в крови
до 2,2—1,7 ммоль/л, или 40—30 мг% возникает гипогликемическая кома,
которая характеризуется потерей сознания, судорогами, бредовыми про­
явлениями со стороны психики, усиленным потоотделением. Глюкоза
поступает в кровь из кишечника и расходуется или захватывается
внутренними органами в разных количественных пропорциях: 12% —
головным мозгом, 9% — кишечником, 7% — мышцами.
Основные пути поступления глюкозы в сосудистое русло:
• поступление из Ж КТ после переваривания углеводов;
• поступление из печени в результате распада гликогена;
• образование из белков и липидов в результате глюконеогенеза,
например, при длительном голодании или недостатке углеводов
в пище.
Основные пути расходования глюкозы в организме:
• Тканевой метаболизм:
—аэробный гликолиз с участием кислорода;
—анаэробный гликолиз в условиях недостатка кислорода;
—пентозофосфатный путь.
• Депонирование в виде гликогена (гликогеногенез).
• Превращение в липиды (липогенез).
• Удаление с мочой (глюкозурия) в условиях повышения концент­
рации глюкозы в крови выше пороговой.
488
Глава 5
Регуляция обмена углеводов
Регуляция уровня глюкозы в крови осуществляется с помощью
функциональной системы саморегуляции, которая включает в себя
множество нервных и гуморальных механизмов. Главным нервным
центром регуляции уровня глюкозы в крови является гипоталамогипофизарная система, которая реализует свои эфферентные влия­
ния через симпатический и парасимпатический отделы ВНС и через
эндокринную систему.
Важная роль в регуляции уровня глюкозы принадлежит гормо­
нам. Система гормонов, повышающих уровень глюкозы в крови:
адреналин, глюкокортикоиды, глюкоген, АКТГ, СТГ. И только один
гормон — инсулин — снижает уровень глюкозы в крови за счет
увеличения транспорта глюкозы в клетку, особенно в мышечной
ткани и печени. Инсулин увеличивает процессы гликогеногенеза и
тормозит процессы гликогенолиза и глюконеогенеза.
Обмен липидов
Обмен липидов представляет собой непрерывные процессы обра­
зования, отложения, мобилизации, транспорта и распада жиров в
организме. Основными представителями класса липидов в организ­
ме являются нейтральные жиры (триглицериды) и их производные
(моно- и диглицериды), фосфолипиды, холестерол, а также жирорас­
творимые витамины (A, D, Е, К).
Жиры по химической природе являются производными высших
жирных, или карбоновых, кислот. Жирные кислоты характеризуются
нерастворимостью в воде и растворимостью в органических раство­
рителях, в том числе в бензине, спирте, эфире.
Нейтральные жиры депонируются в жировой ткани, в том числе
и в форме бурого жира.
Жирные кислоты в зависимости от способа соединения атомов угле­
водородной части молекулы липида подразделяются на две основные
группы — насыщенные и ненасыщенные кислоты. Общее количество
жиров в организме человека составляет 10—20% массы тела, в случае
патологического ожирения их количество достигает 50%. При этом
количество протоплазматического жира остается неизменным. Самые
известные липиды — это жиры и масла. Жирами называют триглицери­
ды, включающие насыщенные жирные кислоты, которые затвердевают
при комнатной температуре; масла включают ненасыщенные жирные
кислоты и при комнатной температуре остаются жидкими.
Висцеральные функции
489
Олеиновая, линолевая, линоленовая и арахидоновая кислоты
являются незаменимыми, не синтезируются в организме и поступа­
ют только с пищей.
Линолевая и линоленовая кислоты содержатся в основном в расти­
тельных жирах, арахидоновая — только в животных. Из арахидоновой
кислоты образуются простагландины, простациклины, тромбоксаны и
лейкотриены. При недостатке их поступления в организм могут возникать
различные нарушения (задержка роста, заболевания кожи, бесплодие).
Липиды представляют собой медленно мобилизуемый энерге­
тический субстрат. Значительный запас липидов в депо позволяет
поддерживать жизнедеятельность организма в течение более месяца в
условиях полного голодания. В процессе окисления жирные кислоты
превращаются в ацетилкоэнзим А, исходный продукт для цикла трикарбоновых кислот, который в дальнейшем расщепляется до двуоки­
си углерода и воды. При помощи ацетилкоэнзима А осуществляется
связь углеводного и жирового обмена.
Важна роль липидов в качестве пластического материала. Они
являются структурной основой биологических мембран. Они спо­
собствуют всасыванию в кишечнике жирорастворимых витаминов.
Подкожная жировая клетчатка выполняет функцию теплоизоля­
ции, поддерживая тепловой баланс организма, создавая одновре­
менно защитный механический футляр для многих органов и тка­
ней. Липиды поддерживают водный баланс организма. В процессе
окисления жиров образуется довольно много воды — 107 мл на 100 г
жира. Жиры принимают важное участие в регуляции половой фун­
кции мужчин и женщин, из них образуются стероидные гормоны и
различные биологически активные вещества. Окисление жиров дает
в 2 раза больше энергии, чем окисление углеводов, — 9,3 ккал/г.
Бурый жир представлен особой жировой тканью, располагающейся
в области шеи и верхней части спины у новорожденных и грудных
детей, у детенышей млекопитающих, рождающихся голыми, и у неко­
торых зимнеспящих взрослых животных (1—2% общей массы тела).
В небольшом количестве (0,1—0,2% общей массы тела) бурый жир
имеется и у взрослого человека. Особенностью состава бурого жира
является огромное количество митохондрий с красновато-бурыми
пигментами — цитохромами; в них происходят интенсивные процес­
сы окисления, не сопряженные с образованием АТФ. Важнейшую
роль в механизмах этого явления играет белок термогенин, составля­
ющий 10—15% общего белка митохондрий бурого жира.
490
Глава 5
Термогенин способствует обратной транслокации ионов Н+, что
снижает протонный потенциал до уровня, при котором невозможен
синтез АТФ. Энергия переноса ионов Н+ по градиенту концентра­
ции рассеивается в виде тепла. В этом случае не 30—40% энергии,
как в обычных митохондриях, а почти вся энергия окисления
превращается в тепло. Жирные кислоты активируют функции термогенина, вследствие чего усиливается разобщение окисления и
образования АТФ.
Продукция тепла бурым жиром (на единицу массы его ткани) в
20 раз и более превышает таковую обычной жировой ткани. Несмотря
на минимальное содержание бурого жира, в нем может генерироваться
1/3 всего образующегося в организме тепла. Бурому жиру принадлежит
важная роль в адаптации организма к низким температурам.
У новорожденных и зимнеспящих животных низкая функцио­
нальная активность организма и незрелость центральных и перифе­
рических механизмов терморегуляции не обеспечивают достаточную
теплопродукцию. По-видимому, роль дополнительного специфичес­
кого генератора тепла в этих случаях играет бурый жир. У взрослых
же необходимость в дополнительном источнике тепла отпадает, так
как теплопродукция обеспечивается иными, более совершенными,
механизмами. Следует отметить, что бурый жир является также
источником эндогенной воды.
Триглицериды
Основные пути поступления в кровь:
• поступление из кишечника в результате переваривания пищи и
последующего всасывания (экзогенные липиды);
• мобилизация из жировой ткани в процессе липолиза (эндогенные
липиды);
• поступление из печени в результате липогенеза (из белков, угле­
водов).
Основные пути расходования:
• катаболизм;
• депонирование в жировой ткани;
• превращение в глюкозу.
Основным энергетическим субстратом для использования в тка­
нях с быстрым обменом (мышцы, головной мозг) являются жирные
кислоты и кетоновые тела (ацетоуксусная кислота, ацетон и р-оксимасляная кислота) Для окисления кетоновых тел необходимы угле­
воды («жиры сгорают в пламени углеводов»).
Висцеральные функции
491
Фосфолипиды
Фосфолипиды участвуют в формировании плазматических кле­
точных мембран. В нервной системе находится большое количество
сфингомиелинов, участвующих в качестве электроизоляторов миелиновой оболочки. Фосфолипиды поступают с пищей (экзоген­
ные фосфолипиды), но большая часть их синтезируется в печени
(эндогенные фосфолипиды). Только гепатоциты способны выделять
фосфолипиды в кровь, поэтому печень — единственный орган, опре­
деляющий уровень фосфолипидов крови.
Холестерол, или холестерин, — представитель одного из классов сте­
роидов — стеринов, к которым также относятся стероидные гормоны,
витамин D и желчные кислоты. Вместе с фосфолипидами он входит в
состав клеточных мембран, обеспечивая мембране высокую устойчи­
вость. Экзогенный холестерин (его поступает около 400 мг/сут) входит
в состав пищевых продуктов, больше всего его содержится в молоке и
яичном желтке. Синтез эндогенного холестерина (около 1000 мг/сут)
происходит в организме. Из этого количества 500 мг/сут образуются
в печени. В печени образуются желчные кислоты, которые способс­
твуют пищеварению и всасыванию жиров. Холестерин используется
в синтезе стероидных гормонов (в коре надпочечников).
Транспорт холестерина. Холестерин переносится кровью в составе
ЛПВП, ЛПНП и ЛПОНП плотности.
ЛПНП — основной аккумулятор холестерина и «поставщик» его
тканям, в том числе эндотелию сосудистой стенки. Повышение
содержания фракции ЛПНП несет опасность развития атеросклероза
вследствие накопления этих липопротеидов в сосудистой стенке.
ЛПВП, напротив, способствуют удалению холестерина из клеток.
ЛПВП образуются в печени, выделяются в кровь и, циркулируя в сосу­
дистом русле, насыщаются извлекаемым из тканей холестеролом. В физио­
логических условиях проникновение холестерина из ЛПНП в стенки
сосудов за счет активности фибробластов идет постоянно, так же, как
извлечение холестерина из нее за счет ЛПВП. Поэтому для оценки транс­
портных механизмов липидов имеет значение не абсолютное содержание
холестерина в крови, а соотношение его количества в ЛПНП и ЛПВП. В
норме это соотношение близко к единице, что говорит о функциональ­
ном балансе процессов поглощения холестерина тканями и его удаления.
Регуляция липолиза и липогенеза
Гормоны надпочечников (адреналин, глюкокортикоиды), тиреоидные гормоны, СТГ усиливают липолиз, активируя катаболизм.
4 92
Глава 5
Когда уровень глюкозы в плазме повышается, жирные кислоты под
влиянием инсулина депонируются в жировой ткани. Хронический
стресс, сопровождаемый напряжением симпато-адреналовой систе­
мы, может приводить к истощению жировых депо и потере массы
тела. Напротив, дефицит инсулина, например при сахарном диабете,
сочетается с ожирением.
Обмен белков
В организме идут непрерывные процессы биосинтеза, обнов­
ления и разрушения белков. Белки, или протеины, представляют
собой высокомолекулярные органические соединения, построенные
из аминокислот, соединенных пептидными связями и образую­
щих цепи различной длины. Аминокислотные цепи, состоящие из
2—10 аминокислотных остатков, называются олигопептидами. Цепи,
содержащие от 10 до 50—100 аминокислотных остатков, называются
полипептидами. Важнейшие сложные белки организма человека
представлены гликопротеинами, липопротеинами, металлопротеинами, нуклеопротеинами, хромопротеинами.
Во взрослом организме количество распавшегося белка за сутки
должно быть равно количеству синтезированного.
Ф ункции белков в организме многообразны; главные из них:
• каталитическая — обеспечение белками-ферментами биохими­
ческих реакций;
• реализация функций движения с помощью сократительных бел­
ков актина и миозина;
• пластическая — биосинтез новых белков в структурах клетки
взамен утраченных;
• энергетическая — обеспечение энергией процессов анаболизма и
катаболизма;
• поддержание коллоидно-осмотического давления (альбумины
плазмы крови);
• участие в реакциях иммунитета (глобулин);
• источники синтеза гормонов, аминов и биологически активных
веществ.
Поступление полноценных белков в составе пищевых продуктов
животного и растительного происхождения, содержащих необходи­
мый набор аминокислот, — важнейшее условие нормального белкового
обмена и важный фактор поддержания здоровья человека (рис. 5.8.1).
Аминокислоты, содержащиеся в пищевых белках, подразделяют
на 2 группы: заменимые и незаменимые. Заменимые аминокислоты
493
Висцеральные функции
П оступление пищевых
белков
Выделение с мочей
Ж елудочнокиш ечный
тракт +
+ печень
Креатин ин
Пурины
пиримидины
Структурные
белки тканей
тела
Синтез гормонов,
нейротрансмиттеров
и ферментов
Дезаминирование
аминокислот в почках
Общий
метоболический
пул
NH4;N H 3
(сосудистое
русло)
, \
Моча
Инертные белки
(волосы, ногти)
Выделение
с калом
Рис. 5.8.1. Основные пути поддержания пула аминокислот в организме:
А — основные пути поступления аминокислот: из пищевых продуктов,
в результате метаболизма структурных белков тела и общего метаболи­
ческого пула; Б — главные пути расходования аминокислот. Расходование
идет по разным путям: с мочой, аминокислоты используются для образо­
вания креатинина, пуринов, пиримидинов, служат источником для био­
синтеза структурных белков, гормонов, нейротрасмиттеров и ферментов
в случае их недостаточного поступления с пищей могут синтезиро­
ваться в организме. Незаменимые аминокислоты не синтезируются
в организме.
494
Глава 5
Недостаток белков, содержащих незаменимые аминокислоты,
может быть причиной заболеваний человека и различных форм бел­
ковой недостаточности.
Белки не депонируются в органах и системах организма, единствен­
ным их источником является пища. В ЖКТ происходит гидролиз поли­
мерных белков на аминокислоты, которые после всасывания поступают
в кровь и далее к клеткам тканей организма. Система крови, таким обра­
зом, выступает накопителем фонда общего аминокислотного пула.
О
количестве распавшегося белка в организме можно судить по
количеству выделенного (с мочой, калом, потом) азота. Известно/что
в белках в среднем содержится 16% азота, или 6,25 г белка содержат
1 г азота. Если, например, за сутки выделилось 16 г азота, то коли­
чество распавшегося белка будет равно 16 х 6,25 = 98,80 г.
В здоровом организме количество выделенного азота равно коли­
честву поступившего в составе пищевых белков. В этом случае
говорят об азотном равновесии. Когда в организм поступает азота
больше, чем выделяется, говорят о положительном азотном балан­
се (состояние во время роста, беременности, выздоровления после
тяжелого заболевания).
В состояниях отрицательного азотного баланса количество выделя­
емого из организма азота превышает его поступление в организм. Это
наблюдается при голодании, тяжелых заболеваниях, старении.
Допустимые потери белка при отсутствии его в пище и достаточном
поступлении всех иных питательных веществ составляют 0,025—0,075 г
азота на 1 кг массы тела человека в сутки (коэффициент изнашивания,
по Рубнеру). Минимум белка, который должен поступить с пищей для
сохранения белкового баланса, составляет 60—75 г/сут.
При систематическом белковом голодании даже при условии пос­
тупления других питательных веществ в организме развиваются
процессы, приводящие к летальному исходу.
Скорость распада и обновления белков организма различна.
Небольшие периоды полураспада у белков в плазме крови и печени
(около 10 сут), белков мышц — около 180 сут.
Поддержание аминокислотного пула в крови и белков плазмы
крови осуществляется на основе механизмов саморегуляции, в кото­
рый включены нервные и гуморальные звенья.
Высшая регуляция белкового обмена, как и других видов обмена,
осуществляется гипоталамо-гипофизарной системой через симпати­
ческие и парасимпатические отделы ВНС и эндокринное звено.
Висцеральные функции
495
Гормональная регуляция осуществляется системой гормонов,
находящихся под контролем гипоталамо-гипофизарной системы.
Гормон роста влияет на синтез белков; инсулин ускоряет транс­
порт аминокислот в клетки; тестостерон увеличивает накопление
белков в организме; эстрогены тоже увеличивают накопление белков;
глюкокортикоиды усиливают катаболизм белков, увеличивают кон­
центрацию аминокислот в плазме крови; Т4 увеличивает метаболизм
во всех клетках и повышает интенсивность обмена белков.
Правило изодинамии Рубнера. Исходя из физических энергети­
ческих эквивалентов пищевых веществ, немецкий физиолог Рубнер
усмотрел возможность их замены в соответствии с их энергетической
ценностью (ккал). Однако это правило не универсально. Ж иры в
определенной степени возможно заменять углеводами в соответствии
с их калорической значимостью. Углеводы нельзя заменять жирами —
«жиры сгорают в пламени углеводов». Белки играют пластическую
роль в организме, поэтому их также нельзя заменять другими вещес­
твами.
Обмен энергии
В основе процессов обмена энергии лежат законы термодинамики.
С точки зрения термодинамики живые существа относятся к откры­
тым стационарным неравновесным системам. Они обмениваются с
окружающей средой веществом и энергией. С другой стороны, благо­
даря механизмам саморегуляции составляющих их функциональных
систем живые организмы удерживают свои основные функции и
энергетические затраты на них в определенных пределах.
Принцип устойчивого неравновесия живых систем (Э.С. Бауер) гласит:
живые системы никогда не бывают в равновесии с окружающей
средой и исполняют за счет своей внутренней энергии постоянную
работу против равновесия, требуемого законами физики и химии.
Первый закон термодинамики — закон сохранения и превращения
энергии (М.В. Ломоносов, А. Лавуазье) гласит: энергия не исчезает и
не образуется вновь, а только переходит из одной формы в другую —
энергия механической работы, кинетическая энергия и теплота могут
превращаться друг в друга.
Второй закон термодинамики (Л. Больцман): часть энергии организ­
ма способна к превращениям и совершению полезной работы. Другая
часть рассеивается в виде тепла, характеризуя меру термодинамичес­
кой неупорядоченности системы — энтропию.
496
Глава 5
Полагают, что в основе энергетических процессов жизнедеятель­
ности лежат сложные перемещения электронов (А. Сент-Дьерди).
Живые организмы не могут снова использовать выделяющуюся в
процессах жизнедеятельности тепловую энергию.
Свободная энергия поступает в организм в основном с питатель­
ными энергетическими веществами, а также с солнечной энергией.
Энергия, заключенная в поступающих в организм веществах, транс­
формируется в клетках организма в химическую, электрическую,
механическую и осмотическую энергию. Часть энергии выделяется
из организма в виде тепла.
Энергия солнечных лучей стимулирует синтез биологически
активных веществ в коже, сетчатке глаза, а также вызывает измене­
ния деятельности желез внутренней секреции.
Важнейший переносчик энергии в клетках — АТФ. Потенциальная
энергия, содержащаяся, например, в химических связях молекул
глюкозы, переходит в результате аэробного или анаэробного окисле­
ния в форму богатых энергией фосфатных связей АТФ.
Солнце — источник внеатомной электромагнитной энергии. При
столкновении фотонов (квантов света) с электронами, находящимися
на какой-либо орбите, за счет приобретения электронами дополнитель­
ной энергии этих квантов происходят увеличение внутренней энергии
атомов и переход их в возбужденное состояние. Коротковолновая часть
электромагнитного спектра, излучаемого Солнцем, которая включает
рентгеновские лучи, гамма-лучи, ультрафиолетовые лучи, обладает
высокой энергией и способна повреждать живую ткань. Видимый
свет, а также инфракрасное излучение обладают относительно низкой
энергией, но способны переводить электроны, которые находятся на
внешних орбитах, на более высокие энергетические уровни. Переходя
на низшую орбиту, электроны делают атом возбужденным, выделяют
энергию и могут вступать в различные химические реакции.
Энергозатраты у человека зависят от пола, возраста, массы, интен­
сивности выполняемой работы, времени суток, сезона и др. Они
меняются в связи с изменением окружающей температуры, влажнос­
ти, барометрического давления и пр.
Поступающая в организм энергия используется в процессах кле­
точного метаболизма, мышечной деятельности, секреторной деятель­
ности и в процессах нервного возбуждения.
Висцеральные функции
497
Ф изическая калориметрия
Физическая калориметрия — метод определения ценности пита­
тельных веществ. Калорическую ценность питательных веществ
определяют путем сжигания 1 г вещества в специальном калориметре
(бомба Бертло) при пропускании электрического тока. Калориметр
погружается в воду, измеряется разность температур воды до и после
сжигания веществ.
Жиры и углеводы сгорают в бомбе Бертло и в организме до воды
и углекислого газа. Белки в бомбе Бертло сгорают до конечных про­
дуктов — N2, СО, и Н20. Однако в организме белки окисляются до
мочевины, креатинина и мочевой кислоты. 1 г мочевины при сгора­
нии выделяет 1,3ккал. Поэтому при окислении 1 г белка в организме
тепла выделяется меньше, чем при окислении 1 г белка в бомбе Бертло.
Физиологическая энергетическая ценность белка в организме равна
4.1 ккал. Калорическая ценность 1 г жира — 9,3 ккал, 1 г углеводов —
4.1 ккал (рис. 5.8.2).
Указанные расчеты относительны. Трудно определить, сколько
питательного продукта усваивается в организме в каждом конкрет­
ном случае. К тому же определенное количество принимаемых пита­
тельных веществ аккумулируется в депонированном состоянии.
Для расчета энергетической ценности принимаемых веществ
необходимо количество белков, жиров и углеводов помножить на их
физиологическую ценность.
Химическая калориметрия
Основана на окислении пищевых веществ сильными окислите­
лями с известным количеством кислорода. После реакции избыток
кислорода оттитровывается иодометрически.
Физиологическая калориметрия
Физиологическая калориметрия служит для определения энер­
гозатрат организма. Когда не совершается физическая работа, вся
химическая энергия переходит в тепловую. Это позволяет по коли­
честву выделяемого организмом тепла судить об интенсивности
энергетического обмена.
Количество тепла, выделяемого организмом, рассчитывают мето­
дом прямой и непрямой калориметрии.
Глава 5
498
О кисление
Продукты окисления
Энергия
1 г белка
+ о2
---- ---------
N H 3, H 2S, н20, С02 +
4,1 ккал
1 г углеводов
+ 02
---- ---------
н2о + со2+
4,1 ккал
1 г ж ира
+ о2
------ ---------
н2о, со2+
9,3 ккал
100 г белка
-------------------------------
4 ,1 x 1 0 0 г
410 ккал
500 г углеводов
-------------------------------
4 ,1х 500 г
2050 ккал
9,3 х 100 г
930 ккал
Б
100 г жира
100 г белка
410 ккал
500 г углеводов
2050 ккал
100 г жира
930 ккал
итого за сутки
3390 ккал
Висцеральные функции
499
Прямая калориметрия
Основана на непосредственном и полном учете количества выде­
ленного организмом тепла с помощью специальных камер — биокало­
риметров. Первый калориметр для измерения энергозатрат у живот­
ных был изобретен в 1893 г. В.В. Пашутиным и А.А. Лихачевым.
Калориметры герметизированы и надежно теплоизолированы от
окружающей среды. Для расчета количества тепла, выделяемого
помещенным в калориметр животным или человеком, измеряют
изменения температуры жидкости, протекающей через камеру за
единицу времени.
Современные биокалориметры представляют собой специальные
костюмы, снабженные термочувствительными датчиками.
Непрямая калориметрия
Данный метод основан на газовом анализе. Выделяют непрямую
калориметрию с полным и неполным газовым анализом.
При полном газовом анализе энергозатраты рассчитывают за еди­
ницу времени по количеству поглощенного испытуемым кислорода
и выделенного углекислого газа. Отношение объема выделенного
углекислого газа к объему поглощенного к:ислорода называется дыхателъным коэффициентом.
Дыхательный коэффициент позволяет определить, за счет каких
веществ преимущественно осуществляется метаболизм у испыту­
емых. При дыхательном коэффициенте, равном 1, обмен веществ
преимущественно идет за счет окисления углеводов:
С 6Н 1 2 ° 6
+ 6 0 2 = 6С 02 + 6Н20
При дыхательном коэффициенте ниже 0,7 обмен веществ осу­
ществляется преимущественно за счет окисления жиров:
2С51Н580 6 + 14502 = Ю2С02 + 98Н20
Рис. 5.8.2. Показатели прихода энергии при среднестатистическом сбалан­
сированном поступлении основных пищевых продуктов: А — показатели
энергетической ценности основных пищевых продуктов; Б — сравнитель­
ные показатели прихода энергии при окислении белков, жиров и углеводов
в суточном рационе; В —расчет суммарного показателя суточного прихода
энергии при окислении белков, жиров и углеводов
1
500
Глава 5
Калорические эквиваленты 1 л кислорода при окислении в организме
различных веществ
Калорический эквивалент 1 л кислопода показывает, какое коли­
чество тепла выделяется при поглощении организмом 1 л кислорода.
Установлено, что при окислении в организме углеводов калоричес­
кий эквивалент 1 л кислорода равен 5 ккал, при окислении жиров —
4,6 ккал, при окислении белков — 4,85 ккал. Расчетным путем состав­
лены таблицы зависимости между величинами дыхательных коэф­
фициентов при окислении только жиров и углеводов и калорическим
эквивалентом 1 л кислорода.
Определенную трудность представляют расчеты энергозатрат при
дыхательном коэффициенте, равном 0,85, поскольку это могут быть
энергозатраты при окислении белков или при окислении смеси
жиров и углеводов. Это особенно важно для того, чтобы узнать,
какую величину калорийной ценности кислорода при этом исполь­
зовать — 4,85 ккал для белка или 4,6 для жиров и 5 для углеводов.
Для исключения этих ошибок определяют «чистый» дыхательный
коэффициент.
Для определения «чистого» дыхательного коэф фициента необ­
ходимо исклю чить из окислительных процессов белок. Для этого
определяют количество выделенного из организма с мочой азота.
В белке — 16% азота, т.е. 1 г азота содержится в 6,25 г белка. При
умножении количества азота, выделенного за сутки, на 6,25 г
получают количество белка, участвовавшего в суточных энерго­
затратах.
1 г белка при сгорании связывает 0,95 л 0 2 и выделяет 0,76 л
СО,. При умножении этих показателей на количество белка,
участвовавшего в суточных энергозатратах, определяют, сколько
0 2 пошло в сутки на окисление белков и сколько при этом выде­
лилось С 0 2. Эти цифры вычитают, соответственно, из общего
количества О, и СО-,, в результате чего оказывается возможным
определить «чистый» дыхательный коэф ф ициент как отношение
оставшегося С 0 2 к 0 2.
При неполном газовом анализе расход энергии определяют по коли­
честву кислорода, потребленного испытуемым за единицу времени.
Д ля этой цели используют спирометаболографы, поглощающие в
выдыхаемом воздухе С 0 2 и позволяющие определять только коли­
чество поглощенного испытуемыми 0 2.
Зная объем поглощенного за 1 мин кислорода (V 02) и усреднен­
ный калорический эквивалент кислорода, равный при употреблении
Висцеральные функции
501
смешанной пищи 4,8 ккал, энергообмен рассчитывают по следующей
формуле:
I = УР2 х КЭК = VO, х 4,8,
где КЭК — калорический эквивалент 1 л 0 2.
Как правило, неполный газовый анализ применяют для опреде­
ления энергозатрат при основном обмене. В этом случае при исклю­
чении из энергообмена белков дыхательный коэффициент соответс­
твует 0,85, т.е. только окисленной смеси углеводов и жиров.
Основной обмен
Основной обмен — показатель минимального расхода энергии,
необходимой для поддержания жизненно важных функций организ­
ма в стандартизованных условиях физиологического покоя в состо­
янии бодрствования. Величина основного обмена характеризуется
как исходный «базальный» уровень энергозатрат организма. Она
представляет собой один из важных параметров организма, наряду
с такими известными константами гомеостазиса, как АД, pH крови,
осмотическое давление и др.
В основной обмен включаются энергозатраты на поддержание
жизненно важных функций: деятельности сердца, дыхания, крово­
обращения, органов выделения, функционирования печени, почек,
желез внутренней секреции и других секреторных органов, которые
у живого организма практически невозможно исключить.
Основной обмен зависит от возраста, роста, массы тела, пола чело­
века и эколого-географических условий обитания.
В среднем у взрослых мужчин энергозатраты, идущие на основной
обмен, составляют 1 ккал на 1 кг массы тела в час. Зная массу тела,
легко вычислить приблизительную нормальную величину основного
обмена за сутки. У мужчины с массой тела 70 кг величина суточно­
го основного обмена будет равна 70 кг х 1 ккал х 24 ч = 1680 ккал.
У женщин величина основного обмена на 10% ниже в связи с мень­
шей массой тела.
Величину основного обмена определяет соотношение процессов
анаболизма и катаболизма. В детском возрасте процессы анаболизма
преобладают над процессами катаболизма, поэтому у детей величины
основного обмена выше по сравнению с взрослыми. Самый интенсивн ый основной обмен в расчете на 1 кг массы тела у новорожденных —
53 ккал/кг в час, у детей первого года жизни — 42 ккал/кг в час, у детей
7 лет — 1,8 ккал/кг в час, 12 лет — 1,3 ккал/кг в час (рис. 5.8.3).
Глава 5
5 02
16 лет
Взрослый, легкий труд
901 ккал
173 ккал
1728 ккал
2420 ккал
2000 ккал
8 лет
14 лет
Взрослая, легкий труд
614 к кал
144 ккал
440 ккал
1442 ккал
1850 ккал
2100 ккал
"1 Ежедневная активность (сидение, стояние, ходьба, двигательная
J активность, игровая деятельность, работа) — рабочий обмен
“1 С пецифическое динамическое действие пищ и — усиление под влиянием приема пищи
J интенсивности объема веществ
Основной обмен
Сумарные суточные энергозатраты
(общ ий обмен)
Рис. 5.8.3. Усредненные показатели различных видов обмена в зависимос­
ти от пола и возраста: А —динамика показаний обмена веществ у мужчин;
Б —динамика показаний составляющих обмена веществ у женщин
У пожилых людей интенсивность основного обмена снижена,
так как у них процессы катаболизма преобладают над процессами
анаболизма.
При сравнении величин основного обмена у людей, имеющих
примерно одинаковую массу тела, при расчете энерготрат на 1 кг
массы тела величины не очень различаются. При сравнении пока­
зателей основного обмена на 1 кг массы тела у разных животных
получаются существенные различия (табл. 5.5).
Висцеральные функции
5 03
Таблица 5.5. Сравнение показателей основного обмена у разных животных
Ж ивотное
Мышь
Гусь
Лошадь
М асса тела, Теплопродукция на 1 кг
кг
массы тела, ккал
0,018
3,5
441
654
66,7
11,3
На 1 м2 поверхности тела
в сутки, ккал
1188
967
948
Как видно из данных табл. 5.5, у разных видов животных эти вели­
чины примерно одинаковы при расчете величины основного обмена
на единицу поверхности тела.
Интенсивность основного обмена более тесно связана с раз­
мерами поверхности тела, чем с массой, что обусловлено прямой
зависимостью величины отдачи тепла от площади поверхности. Еще
в прошлом столетии немецкий физиолог М. Рубнер показал, что у
теплокровных организмов, имеющих разные размеры тела, с одного
квадратного метра поверхности тела в окружающую среду рассеи­
вается одинаковое количество тепла. На этом основании М. Рубнер
сформулировал закон поверхности тела , согласно которому энер­
гетические затраты теплокровного организма пропорциональны
площади поверхности его тела.
Закон имеет ряд ограничений:
• при одинаковой поверхности тела расход энергии зависит от эко­
логических условий существования и образа жизни животного.
Так, у волка величина основного обмена на единицу поверхности
тела больше, чем у собаки таких же размеров;
• у обитателей тропиков основной обмен ниже, чем у полярных
животных с одинаковой поверхностью тела.
Величина основного обмена зависит от возраста животных.
Величина основного обмена зависит от характера кожного покро­
ва (шерстяной покров, одежда у человека и др.), а также от специфики
организации протоплазмы клеток (теплокровные или холоднокров­
ные животные).
Величину основного обмена можно определить методом пря­
мой или непрямой калориметрии; как правило, используют метод
непрямой калориметрии. Процедуру определения основного обмена
проводят в стандартизированных, строго контролируемых услови­
ях: исследование проводят утром после сна, натощак (через 12—14 ч
после последнего приема пищи, в условиях температурного комфорта
504
Глава 5
(около 22 °С), в положении лежа, в условиях физического и психоэмо­
ционального расслабления, за 2—3 сут до процедуры ограничивают
прием белковой пищи.
Величина основного обмена характеризует состояние здоровья чело­
века. Известно, что при гиперфункции щитовидной железы основной
обмен существенно возрастает, а при гипофункции, наоборот, снижа­
ется. Поэтому в ситуации, когда определение уровня гормонов Т3 и Т4
затруднено, можно провести определение уровня основного обмена.
Для каждой возрастной группы людей установлены и приняты
в качестве стандартов определенные величины основного обмена.
Отклонение величины основного обмена от стандартного значения
не более чем на 10% считается нормальным.
Регуляция основного обмена
Главным центром регуляции основного обмена является гипоталамо-гипофизарная система, которая через симпатическую и парасимпа­
тическую нервную систему регулирует вегетативные функции организ­
ма, секрецию гормонов щитовидной железы, надпочечников и половых
гормонов, которые вносят существенный вклад в регуляцию основного
обмена, усиливая или ослабляя процессы анаболизма и катаболизма.
Симпатическая нервная система увеличивает, а парасимпати­
ческая — снижает уровень основного обмена. Повышают величину
основного обмена гормоны щитовидной железы, гипофиза и половые
гормоны.
Общий расход энергии человека
Общий расход энергии у человека склады вается из следую­
щих процессов: основного обмена, специфического динамического
действия пищ и, физической и умственной деятельности, терморе­
гуляции.
При мышечной рабате энергозатраты складываются из величи­
ны основного обмена и так называемой рабочей прибавки. Рабочая
прибавка — энергия, которую тратит организм в течение суток на
физическую и умственную деятельность. Сумма основного обмена
и рабочей прибавки составляет валовый энергетический обмен.
Расход энергии при мышечной нагрузке
Мышечная работа сопряжена со значительными затратами энергии,
с одной стороны, и увеличением теплопродукции — с другой. У спокой­
но лежащего человека расход энергии составляет примерно 35 ккал/м2.
Висцеральные функции
505
Если исследуемый слегка приподнимается и облокачивается на спинку
кровати, расход энергии повышается на 15%; если принимает сидячее
положение — на 42%; в положении стоя — на 70%. При спокойной нето­
ропливой ходьбе расход энергии увеличивается на 180%.
При кратковременных нагрузках энергия выделяется за счет окис­
ления углеводов. При длительных мышечных нагрузках в организме
расщепляются преимущественно жиры, обеспечивая 80% потребной
энергии. У тренированных спортсменов энергия мышечных сокра­
щений обеспечивается исключительно за счет окисления жиров.
У человека, занимающегося физическим трудом, энергетические
затраты возрастают пропорционально интенсивности труда.
Коэффициент физической активности
Согласно рекомендациям Всемирной организации здравоохра­
нения (ВОЗ) объективным физическим критерием, определяющим
адекватное расходование энергии для конкретных профессиональных
групп, является коэффициент физической активности (отношение общих
энергозатрат на все виды жизнедеятельности к величине основного
обмена по расходу энергии в состоянии покоя). Величины коэффици­
ента физической активности одинаковы для мужчин и женщин, но в
связи с меньшей массой тела и, соответственно, величиной основного
обмена у женщин, энергозатраты мужчин и женщин в группах с одним
и тем же коэффициентом физической активности различны.
Предельно допустимая по тяжести работа для человека, постоянно
выполняемая им в течение длительного времени, не должна превы­
шать по энергозатратам уровень основного обмена более чем в 3 раза.
Энергозатраты в зависимости от вида труда. По энергетическим
затратам все профессии разделены на несколько групп, каждая из
которых характеризуется своим суточным расходом энергии.
Группа 1 — работники преимущественно умственного труда: науч­
ные работники, студенты гуманитарных специальностей, операторы
ЭВМ, контролеры, педагоги, диспетчеры, работники пультов управле­
ния и др. Очень легкая физическая активность; коэффициент физи­
ческой активности — 1,4; расход энергии — 1800—2450 ккал/сут.
Группа II — работники, занятые легким физическим трудом: води­
тели трамваев, троллейбусов, работники конвейеров, весовщицы,
упаковщицы, работники сферы обслуживания, агрономы, меди­
цинские сестры, санитарки, продавцы промышленных товаров и др.
Легкая физическая активность; коэффициент физической активнос­
ти — 1,6; расход энергии — 2100—2800 ккал/сут.
5 06
Глава 5
Группа III — работники средней тяжести труда: слесари, налад­
чики, настройщики, станочники, буровики, водители экскаваторов
и бульдозеров, водители автобусов, врачи-хирурги, текстильщики,
обувщики, железнодорожники, водители угольных комбайнов, про­
давцы продовольственных товаров, водники, аппаратчики, метал­
лурги-доменщики, работники химических заводов и др. Средняя
физическая активность; коэффициент физической активности — 1,9;
расход энергии — 2500—3300 ккал/сут.
Группа IV — работники тяжелого физического труда: строительные
рабочие, помощники буровиков, проходчики, хлопкоробы, основная
масса сельскохозяйственных рабочих и механизаторов, доярки, ово­
щеводы, деревообработчики, металлурги, литейщики и др. Высокая
физическая активность; коэффициент физической активности — 2,2;
расход энергии — 2800—3850 ккал/сут.
Группа V — работники особо тяжелого труда (только мужчины):
механизаторы, сельскохозяйственные рабочие в посевной и убороч­
ный периоды, горнорабочие, вальщики леса, бетонщики, каменщи­
ки, землекопы, грузчики немеханизированного труда, оленеводы и
др. Очень высокая физическая активность; коэффициент физической
активности — 2,5; расход энергии — 3750—4200 ккал/сут (рис. 5.8.4).
Обмен веществ при психической деятельности
При психической деятельности, не сопровождаемой движени­
ями, обмен веществ по сравнению с покоем повышается не более
чем на 2—3%. При напряженной эмоциональной деятельности у
ораторов, лекторов, писателей, студентов во время экзаменов обмен
веществ возрастает на 11—20%. Выраженно повышается обмен
веществ при установке на будущую напряженную деятельность:
например, предстартовое состояние у спортсменов, у людей перед
ответственной работой — защ ита диссертации, ответственный
полет у летчиков и т.п.
Рис. 5.8.4. Усредненные показатели энергетических трат организма при
различных состояниях активности организма и видах трудовой деятель­
ности: А — энергозатраты в положении лежа; Б — энергозатраты в поло­
жении сидя; В — энергозатраты при медленной ходьбе; Г — энергозатраты
при быстрой ходьбе; Д — суточные энергозатраты монахов, художников;
Е — энергозатраты домохозяек; Ж — энергозатраты гребцов; 3 — энерго­
затраты каменщиков
Висцеральные функции
1265 ккал
1400 ккал
2300 ккал
Ж
4080 ккал
507
508
Глава 5
Специфически динамическое действие пищи
Возрастание энергозатрат, связанное с приемом пищи, обозначает­
ся как ее специфически динамическое действие и обусловлено аффе­
рентной импульсацией, поступающей в центр голода от рецепторов
ротовой полости и желудка. Уровень обмена веществ увеличивается
как при мнимом кормлении животных с перерезанным пищеводом,
так и при вкладывании пищи непосредственно в желудок.
Прием пищи приводит к возрастанию расхода энергии. При упот­
реблении белка через 1,5—2 ч энергозатраты возрастают на 30—40% и
продолжаются 7—8 ч.
При приеме углеводов энергозатраты возрастают на 4-6% и про­
должаются 2—3 ч.
Прием жиров незначительно увеличивает расход энергии.
Питание
Все живое для удовлетворения метаболических потребностей
нуждается в поступлении питательных веществ из внешней среды.
Питание необходимо для:
• построения и непрерывного обновления клеток и тканей;
• процессов роста и развития;
• поддержания основного обмена;
• покрытия энергозатрат.
Питание включает процессы поступления, переваривания, всасы­
вания в организме пищевых веществ и их утилизацию в метаболи­
ческих процессах.
В России основоположником учения о питании стал М.Н. ТПатерников, который на основе тщательных измерений энергозатрат у
лиц различных профессий научно обосновал нормы питания.
Сбалансированное питание
Сбалансированное питание строится на следующих постулатах.
• Оптимальное соответствие всех компонентов пищи физиологи­
ческим потребностям организма.
• Принимаемая пища с учетом ее усвояемости должна восполнять
энергозатраты человека с учетом основного обмена, специфичес­
кого динамического действия пищи и расхода энергии на выпол­
няемую работу.
• Баланс белков, жиров и углеводов. Среднее соотношение их
массы должно быть 1 : 2 : 4 ; энергетическая ценность 15 : 30 : 55%
соответственно.
Висцеральные функции
5 09
• Баланс белков с незаменимыми и заменимыми аминокислотами,
жиров с разной насыщенностью жирными кислотами, углеводов с
разным в них числом мономеров и наличием балластных веществ
в виде пищевых волокон (целлюлоза, пектин и др.).
• Наличие в рационе витаминов и минеральных веществ в их
соотношении с потребностями, энергозатратами, возрастом,
полом, климатическими условиями, временем года и другими
факторами.
Специальное сбалансированное адекватное питание предусмат­
ривается для детей, подростков, женщин (особенно беременных и
кормящих), лиц пожилого возраста, спортсменов и лиц, работающих
в специальных экстремальных условиях (А.А. Покровский).
А.М. Уголев предложил так называемую трофотропную адекватную
теорию питания. Согласно этой теории питание восполняет молеку­
лярный состав ферментного и изоферментного спектра пищевари­
тельного тракта, обеспечивающего полостное мембранное пищева­
рение. Фактором адекватной резорбции, по мнению А.М. Уголева,
должны быть и всасывающиеся из кишечника питательные вещества.
При ферментопатиях нужно особо тщательно подходить к выбо­
ру продуктов питания: например, при лактозной недостаточности
молоко в пищевом рационе является неадекватным видом пищи.
Автор отводит существенную роль в питании микроорганизмам
кишечника.
Общие принципы составления пищевых рационов
• Принимаемая пища по калорийности должна покрывать энерге­
тические расходы.
• Учет усвояемости пищи.
• Учет норм питания.
• Оптимальное соотношение белков, жиров и углеводов с учетом
критики правила Рубнера.
• Процентное распределение питательных веществ: завтрак — 30%,
обед — 40-50, ужин — 10—20%.
Энергетический баланс. Нормальный взрослый организм чело­
века характеризуется энергетическим балансом: поступление энергии
в организм равно ее расходу (рис. 5.8.5). Энергетический баланс
может быть положительным, когда поступление энергии в орга­
низм превышает ее расход, и отрицательным, когда расход энер­
гии превышает ее поступление в организм. Энергетический баланс
организма поддерживается механизмами саморегуляции соответс-
Глава 5
510
А
Приход
энергии
Расход
энергии
Рис. 5.8.5. Динамика энергетической сбалансированности прихода и рас­
хода энергии: А — энергетически сбалансированный обмен: приход энер­
гии равен расходу; Б — несбалансированный обмен: приход энергии
больше расхода; В — несбалансированный обмен: расход энергии больше
прихода
Висцеральные функции
511
твующей функциональной системы, определяющей оптимальный
уровень энергетических веществ в организме. Установлено, что при
пропуске кормления животные в очередной раз поглощают большее
количество пищи, в которой энергетическая ценность соответству­
ет недополученной при предыдущем кормлении; малокалорийная
пища съедается животными в большем количестве; потребление
пищи увеличивается после мышечной нагрузки; растущие организ­
мы также потребляют больше пищи.
Аппаратом, контролирующим поступление, содержание и расхо­
дование энергетических веществ в данной функциональной системе,
является находящийся на уровне гипоталамуса акцептор результата
действия. Характерно, что все процессы оценки энергетической
потребности и поступления энергетических веществ в организм осу­
ществляются у человека автоматически, на подсознательном уровне.
При этом не только автоматически калькулируется энергетическая
потребность и поступление питательных веществ в организм, но и
осуществляется избирательное всасывание принятых питательных
веществ из пищеварительного тракта в соответствии с истинной
метаболической потребностью тканей.
Депо питательных веществ
Установлено, что глюкоза депонируется в виде гликогена в печени
и поперечно-полосатых мышцах; жировые вещества — в жировой, а
белковые — в соединительной ткани.
Ритм опорожнения депо питательных веществ в основном опре­
деляется приемом пищи и регулируется гормонами гипофиза, эпи­
физа, поджелудочной и половых желез и стимулируемыми ими
ферментами.
Ведущая роль в деятельности депо питательных веществ при­
надлежит нейронам вентромедиального и супрахиазмальных ядер
гипоталамуса.
Количество депонированных питательных веществ регулируется
пищевой потребностью, с одной стороны, и уровнем энергозатрат
организма — с другой.
Вопрос об интимных механизмах регуляции депо питательных
веществ остается недостаточно изученным.
В случаях, когда в депо аккумулируется значительное количество
питательных веществ, масса тела увеличивается. При накоплении
жира наблюдаются тучность и ожирение. При истощении депо наблю­
дается потеря массы тела — кахексия.
512
Глава 5
Нарушение ритма работы депо питательных веществ определяется
изменением нормальных реципрокных отношений нейронов лате­
рального и вентромедиального отделов гипоталамуса, отвечающих
за процессы голода и насыщения.
С другой стороны, активность депо питательных веществ зависит
от интенсивности обменных процессов в тканях, желез внутренней
секреции и функций сигнальных молекул. В качестве молекул, сиг­
нализирующих о потребности в питательных веществах, выступают
олигопептиды: пентагастрин, мотилин, бомбезин. В механизмах
насыщения участвуют холецистокинин, соматостатин.
Сигнальную функцию выполняют стероидные гормоны, простагландины, прогестерон, а также лептин. Простагландины снижают
аппетит, при уменьшении их содержания в крови (это происходит у
тучных людей) усиливается голод.
Проблема ожирения
Проблема ожирения привлекает пристальное внимание врачей
и биологов. Исследования показали, что у людей от 50 до 59 лет с
массой тела на 15—24% больше нормы смертность возрастает на 17%.
Если масса тела превышает оптимальную на 25—34%, то показатель
смертности увеличивается более чем на 41%.
К показателям веса относится индекс массы тела, который равен
отношению массы тела (в килограммах) к росту (в метрах) в квадрате.
Рекомендуемое значение этого индекса составляет примерно 22 для
женщин и 24 для мужчин (рис. 5.8.6).
У тучных людей более выражен аппетит и выше интенсивность
процессов накопления питательных веществ в депо, поэтому кровь
приобретает «голодные» свойства значительно раньше, чем у здо­
ровых людей. С другой стороны, торможение центра голода после
приема пищи наступает позднее, поэтому снижается и поступление
питательных веществ из депо в кровь.
Для снижения массы тела могут быть использованы следующие пути:
• фармакологически понизить чрезмерную активность центра
голода в латеральном гипоталамусе;
• принимать пищу малыми порциями, но часто, стимулируя выброс
питательных веществ из депо;
• понизить интенсивность метаболизма в тканях;
• активировать мышечную деятельность;
• сбалансировать работу сигнальных молекул.
Висцеральные функции
513
Ож ирение III степе]
ИМТ, кг/м 2
40
38
37
36
Ожирение II степей и
#=
33
32
31
30
29 28 ,
27 ■
26 ■
25 ,
24 .
23 .
Ожирен ie I степей] i
22 .
21 .
20
-
19
1 9-24
2 5 -3 4
3 5 -4 4
4 5 -5 4
5 5 -6 4
>65
Возраст, годы
Рис. 5.8.6. Динамика изменения индекса массы тела (ИМТ): А — возрас­
тная динамика изменения индекса массы тела в норме; Б — показатели
изменения индекса массы тела при разных степенях ожирения
5.9. ТЕРМОРЕГУЛЯЦИЯ
Температура тела у человека и теплокровных животных — одна
из пластичных констант, допускающая отклонения от оптимального
уровня на определенное время. Температура тела определяет опти­
мальный уровень жизнедеятельности организма, обусловленный в
значительной степени процессами метаболизма тканей. Длительное
отклонение температуры тела в ту или другую сторону существенно
нарушает нормальные процессы жизнедеятельности.
514
Глава 5
По показателям значений температуры тела различают: нормотермию — оптимальный уровень температуры тела, обусловливающий
нормальное течение процессов метаболизма, гипотермию — понижение
температуры тела и гипертермию — повышение температуры тела. ^ , i
Гомойотермия (гомеотермия). Высшие животные и человек облада­
ют способностью поддерживать температуру тела в рамках оптималь­
ной для течения метаболических процессов. Это свойство получило
название гомойотермия («гомойос» — равный, «термос» — темпера­
тура). Живые существа, обладающие способностью поддерживать
постоянство температуры тела, называются гомойотермными или
теплокровными.
Гомойотермия у человека возникает через несколько месяцев
после рождения. Новорожденный ребенок не может самостоятельно
поддерживать температуру тела и может погибнуть при охлаждении.
Взрослый организм человека реагирует на изменение температуры
тела изменением теплопродукции или теплоотдачи. Эти процессы
стабилизируют его температуру тела.
Цойкилотермия. Животные, которые не способные поддерживать
постоянство температуры тела называются пойкилотермными («пойкилос» — разный) или холоднокровными. Температура их тела зависит
от температуры окружающей среды. К пойкилотермным относятся
земноводные и пресмыкающиеся животные.
При охлаждении пойкилотермных животных температура их
тела понижается. Их поведение во внешней среде прекращается.
Животные замирают. Уменьшение интенсивности метаболизма при
охлаждении называется гипобиозом и анабиозом. В этом состоянии
они могут пережить длительное охлаждение организма. При последу­
ющем увеличении окружающей температуры функции их организма
и поведение животных может быть восстановлено в полном объеме.
Гетеротермия. Промежуточная группа животных — гетеротермная
(«гетерос» — изменчивый). Такие животные имеют факультативную
гомойо- и пойкилотермию. Летучая мышь, например, в полете обладает
гомойотермией, а в вертикальном подвешенном состоянии в помеще­
нии — пойкилотермией. Зимнеспящие животные, например суслики и
медведи, во время зимней спячки становятся пойкилотермными.
В состоянии покоя у этих животных понижается температура
тела, что сохраняет энергетические запасы в организме. Понижение
температуры не должно приводить к замерзанию воды в организме.
Поэтому летучие мыши имеют пристанище в пещерах, суслик спят
в земляных норах и медведи ложатся на зимнюю спячку в берлоги.
Висцеральные функции
515
При переходе этих животных к целенаправленному поведению они
становятся гомойотермными. Увеличение интенсивности метаболиз­
ма увеличивает скорость двигательных реакций при их пищедобывательном поведении.
Функциональная система, поддерживающая
оптимальный уровень температуры тела
Ф ункциональная система, поддерживающая оптимальный уро­
вень температуры тела, является динамической саморегулирующейся
организацией, все составные элементы которой содействуют под­
держания постоянной температуры тела. Функциональная система
включает внутреннее относительно пассивное звено саморегуля­
ции и внешнее активное поведенческое звено регуляции (рис. 5.9.1).
Деятельность функциональной системы направлена главным обра­
зом на поддержание оптимальной температуры ядра тела.
___
Поведение
С окращ ение мыш ц
М ыш ечный тонус
М ыш ечная дрожь
Термо­
рецепторы
кожи
Л иполиз бурого жира
Термо­
рецепторы
сосулов
Клеточный метаболизм
X
Поведение
X
Конвекция
И злучение
Теплопроведение
П отоотделение
Отдача теплоты легкими
.
Отдача теплоты
х с мочой и калом /
Рис. 5.9.1. Функциональная система поддержания постоянства температу­
ры крови в организме человека (ПГ —передний гипоталамус; ЗГ —задний
гипоталамус)
i
.
Висцеральные функции
517
Конечным полезным для организма приспособительным резуль­
татом функциональной системы является такой уровень температуры
в тканях организма, который обеспечивает оптимальное течение
метаболических процессов в органах и тканях организма. В зависи­
мости от особенностей метаболизма уровень температуры в разных
тканях различен. Поэтому конечного результата функциональной
системы, поддерживающей оптимальный уровень температуры в
организме, мультипараметричен.
Температурная схема тела
Температура тела различна в отдельных его частях. Аксиллярная
температура в подмышечной впадине равна 36,6—36,8 °С, оральная
температура в ротовой полости температура и ректальная в прямой
кишке равна 36,8—37,2 °С. На пальцах рук температура равняется
28—32 °С. На пальцах ног температура равняется 24—28 °С.
Различают.температуру внутренних органов или ядра тела и тем­
пературу поверхности тела или его оболочки. Температура ядра тела
более высокая, чем температура оболочки тела. Температура ядра тела
поддерживается за счет метаболизма внутренних органов. Например,
температура венозной крови из печени равна 37,8—38,0 °С. Средней
температурой ядра тела принято считать температуру крови в правом
желудочке сердца — 37.5 °С.
В оболочке тела температура может изменяться на большую вели­
чину, чем температура ядра тела. При нагревании или охлаждении
организма температура кожи на руках и ногах может изменяться от
16 до 35 °С без неприятных ощущений.
Распределение температур по внутренним органам и поверхности
кожи у каждого субъекта имеет индивидуальный, но в нормальных
условиях относительно постоянный характер. Индивидуальное рас­
пределение температуры по поверхности кожи и разным органам
составляет температурную схему тела (рис. 5.9.1).
Температура тела зависит от температуры окружающей человека
среды, вызывая нагревание или охлаждение организма. Температура
тела зависит от функционального состояния организма. В тече-
Рис. 5.9.2. Температурная схема тела человека в зависимости от темпе­
ратуры окружающей среды: А — в условиях пониженной температуры;
Б —в условиях повышенной температуры
Глава 5
518
ние суток температура тела у человека изменяется на 0,5—0,7 °С,
Температура тела максимальна во второй половине дня и минималь­
на в середине ночи (рис. 5.9.2), что существенно зависит от динамики
секреции анаболических и катаболических гормонов, а также от
изменения тонуса вегетативной нервной системы.
У женщин выражены месячные циклы колебания температу­
ры тела. Ректальная температура снижается при менструации и
сохраняется на низком уровне первую фазу менструального цикла.
В середине менструального цикла происходит овуляция и темпера-
Бодрствование
Н очной сон
37,0
36.8
36,6
36,4
36,2
Г"
6
12
Время суток, часы
Рис. 5.9.3. Динамика колебаний ректальной температуры человека (t°°C)
в течение суток
Висцеральные функции
519
тура увеличивается на 0,5—1,0 °С. Увеличенная температура сохраня­
ется в течение второй фазы менструального цикла.
Температуру тела повышает прием пищи, что вызвано специфи­
ческим динамическим действием пищи. Температура тела увеличи­
вается за счет переваривания пищи в желудочно-кишечном тракте.
Повышение температуры тела происходит при физической работе,
а также при нервном возбуждении, особенно психоэмоциональном
стрессе. Температура тела изменяется у беременных и во время родов.
Температура тела увеличивается при инфекционных заболеваниях.
Терморецепторы
Выделяют три группы терморецепторов в соответствии с местом
их локализации: поверхностные терморецепторы, расположенные в
толще кожи, терморецепторы, локализованные в стенках кровенос­
ных сосудов и тканях организма, а также терморецепторы централь­
ной нервной системы, расположенные в гипоталамусе, ретикулярной
формации ствола мозга и в спинном мозге.
Кожные терморецепторы подразделяют на тепловые и холодовые.
Холодовые рецепторы располагаются в толще кожи, на глубине около
07Г7 мм. Тепловые рецепторы располагаются на глубине 0,3 мм.
Холодовые и тепловые рецепторы располагаются неравномерно
по кожной поверхности. Имеются индивидуальные зоны преиму­
щественной локализации тепловых и холодовых терморецепторов. У
людей, живущих в странах с холодным климатом, общее число точек
поверхности кожи, воспринимающих холод, значительно превышает
число точек, воспринимающих тепло.
При оптимальной для человека температуре окружающей среды
терморецепторы генерируют потенциалы действия со стационар­
ной частотой. При понижении окружающей температуры частота
импульсной активности холодовых рецепторов возрастает. При этом
снижается импульсная активность тепловых рецепторов. При воз­
растании окружающей температуры, наоборот, возрастает частота
импульсной активности тепловых рецепторов и снижается частота
возбуждений холодовых рецепторов (рис. 5.9.3).
Центры терморегуляции
Установлено, что ведущие центры, поддерживающие своей
деятельностью постоянство температуры тела, расположены в гипоталамической области. Перерезка ствола мозга по верхнему краю
передних бугров четверохолмия ниже гипоталамуса приводит к тому,
5 20
Глава 5
Рис. 5.9.4. Стационарная частота импульсной активности (имп/с) тепло­
вых и холодовых рецепторов (TP, ХР) кисти руки при сохранении кожной
температуры в течение десятков минут (t°°C —температура кожи в градусах
Цельсия)
что гомойотермные животные становятся пойкилотермными: темпе­
ратура их тела становится равной температуре окружающей среды.
Раздражение отделов гипоталамуса через вживленные электроды
приводит к изменению температуры тела экспериментальных живот­
ных. Разрушение ядер гипоталамуса также вызывало изменение
температуры тела.
В области задних ядер гипоталамуса находится центр теплопродук­
ции. Разрушение этого центра приводит к тому, что животные утра­
чивают способность поддерживать постоянство температуры тела в
условиях пониженной температуры окружающей среды. Температура
их тела в этих условиях начинает падать, и животные переходят в
состояние гипотермии.
Раздражение этих отделов гипоталамуса вызывает у животных
сужение поверхностных сосудов кожи, пилоэрекцию, появление
мышечной дрожи, увеличение секреции адреналина и кортикостеро­
идов надпочечниками.
■
Висцеральные функции
521
Нейроны центра теплопродукции возбуждаются при перфузии
через мозг охлажденной крови и при раздражении холодовых рецеп­
торов кожи при охлаждении организма.
В области передних ядер гипоталамуса располагается центр теп­
лоотдачи. Разрушение этих структур приводит к тому, что животные
утрачивают способность поддерживать постоянство температуры тела
в условиях высокой температуры окружающей среды. Температура их
тела в этих условиях начинает возрастать. Животные переходят в
состояние гипертермии. Гипертермия развивается у животных даже
при комнатной температуре.
Раздражение этих структур через вживленные электроды электри­
ческим током вызывает у животных одышку, расширение поверхнос­
тных сосудов кожи, падение температуры тела. Охлаждение организ­
мам животных не приводит к возникновению мышечной дрожи.
Нейроны центра теплоотдачи возбуждаются при перфузии через
мозг подогретой кров и при раздражении тепловых рецепторов кожи
при нагревании организма.
Между центрами теплоотдачи переднего гипоталамуса и теп­
лопродукции заднего гипоталамуса существуют реципрокны е
взаимоотнош ения (рис. 5.9.4). При усилении активности центра
теплопродукции тормозится возбуждение центра теплоотдачи и
наоборот.
При снижении температуры тела возбуждаются нейроны в центра
теплопродукции. При этом афферентная часть центра теплопродук­
ции тормозит эфферентную часть центра теплоотдачи.
При повышении температуры тела активируются нейроны центра
теплоотдачи. При этом афферентная часть центра теплоотдачи тор­
мозит эфферентную часть центра теплопродукции.
Теплопродукция и теплоотдача
Теплопродукция связана с биохимическими механизмами терморе­
гуляции. Она зависит от интенсивности метаболических процессов
в тканях. Интенсивность тканевого метаболизма и теплопродукции
зависит от ряда факторов.
• Генетически детерминированными особенностями субъекта:
его ростом, массой тела, общей величиной поверхности тела,
полом, активностью эндокринной системы, тонусом вегетатив­
ной нервной системы.
• Характером питания, что связано со специфическим динамичес­
ким действием пищи.
UUWJUjLT1- "ГОих^С — >
v-U 'i ( v
— > ^a^UM)4UCu~*~* KoU-
522
Глава 5
m tttm ttttttt
Тепло
tttmmtmttt
Холод
Висцеральные функции
523
• Интенсивностью мышечной работы. Более интенсивная мышеч­
ная работа увеличивает теплопродукцию. Существенным ф акто­
ром повышения теплопродукции в условиях понижения окружа­
ющей температуры является мышечная дрожь.
• Окружающей температурой. Теплопродукция увеличивается при
низких и снижается при высоких температурах.
• Психоэмоциональным состоянием субъекта. Состояние возбуж­
дения усиливает интенсивность теплопродукции и позволяет
переживать низкие температуры.
• Кислородным обеспечением организма. Недостаток кислорода
увеличивает теплопродукцию.
• Интенсивностью освещенности помещения. Как правило, в тем­
ноте теплопродукция снижается.
• Местом проживания на Земле. У жителей южных широт тепло­
продукция по сравнению с жителями северных широт понижена.
• Дополнительным процессом, увеличивающ им температуру
тела, может быть чихание, вызванное охлаждением организма.
Возникающие при этом интенсивные сокращения межреберных
мышц приводят к кратковременному нагреву грудной полости.
Теплоотдача зависит в основном от физических механизмов теп­
лообмена между организмом и внешней средой Теплоотдача опреде­
ляется следующими процессами.
• Около 70% тепла передается в окружающую среду с помощью
теплоизлучения — излучения электромагнитных волн в инф ­
ракрасном диапазоне электромагнитного спектра колебаний.
Интенсивность этого излучения регистрируют с помощью теп­
ловизоров, на экране которых возникают температурные схемы
тела, окрашенные в различные цвета в зависимости от темпера­
туры отдельных областей тела.
• Около 15% тепла отдается с помощью теплопроведения. Оно воз­
никает за счет непосредственной передачи кинетической энергии
Рис. 5.9.5. Реципрокное взаимоотношение между центром теплоотда­
чи в переднем гипоталамусе (ПГ) и центром теплопродукции в заднем
гипоталамусе (ЗГ): А —усиление теплоотдачи при нагревании организма;
Б —усиление теплопродукции при охлаждении организма (АО, ЭО —аффе­
рентные и эфферентные отделы центров теплопродукции и теплоотдачи;
ТН —тормозной нейрон)
5 24
Глава 5
колеблющихся молекул нагретого тела молекулам окружающей
среды при их соударении.
• Испарение воды с поверхности кожи и верхних дыхательных
путей происходит с затратой тепловой энергии при превращении
жидкости в пар. В покое около 14% тепла теряется организмом за
счет испарения.
• Конвекция или перемещение окружающей среды вокруг тела —
дополнительный фактор теплоотдачи. Охлаждение организма
быстрее происходит в ветреную погоду или при наличии сквоз­
няка в помещении. Охлаждение организма быстрее происходит в
реке, чем в неподвижной воде озера или бассейна.
Эффективность отдачи тепла прямо пропорциональна темпера­
туре организма и окружающих его предметов, площади поверхности
тела, скорости движения воздуха и обратно пропорциональна тепло­
изолирующим свойствам кожи, шерстного покрова, а у человека —
теплоизолирующим свойствам одежды.
Процессы теплоотдачи зависят от ряда факторов. Теплоизлучение
и теплопроведение прямо пропорциональны теплопроводности окру­
жающей среды. Так, на берегу реки или моря, где теплопроводность
воздуха выше, чем в помещении, теплоотдача осуществляется интен­
сивнее и ощущение окружающей высокой температуры понижается.
Потеря тепла организмом меньше на морозном, но сухом воздухе, чем
на влажном воздухе при температуре около О°С. Высокая влажность
воздуха при низкой температуре может привести к обморожению
конечностей.
Теплоотдача зависит от объема поверхности тела. Известно, что
многие животные на холоде сворачиваются в клубок, что уменьшает
открытую поверхность тела и, соответственно, потерю тепла. Человек
на холоде тоже пытается сжаться, втягивая голову в воротник паль­
то. Наоборот, в тепле животные распластываются по занимаемой
поверхности, стараясь занять больший объем для увеличения теп­
лоотдачи.
Физические процессы теплоотдачи зависят от свойств кожного
покрова. Шерстный покров кожи животных препятствует теплоотда­
че. Поэтому при нагревании животные (собаки, кошки, львы, тигры),
имеющие густой шерстный покров, усиливают теплоотдачу за счет
испарения воды с поверхности ротовой полости и языка при частом
дыхании через открытую пасть животного.
Процессы теплоотдачи изменяются в водной среде. Вода облада­
ет большей теплопроводностью, чем воздух. Поэтому температура
Висцеральные функции
525
холодной воды кажется ниже, чем такая же температура воздуха.
В воде исключается испарение.
Ф изиологические процессы теплоотдачи дополняют физические
механизмы тепловыделения. К физиологическим процессам относят­
ся: интенсивность внешнего дыхания, кожные сосудистые реакции,
согревание принятой холодной пищи, выделение тепла из организма
с калом и мочой и потоотделение (рис. 5.9.6). Дефекация и мочеиспус­
кание приводят к потере 1% тепла.
Испарение 1 мл воды с поверхности тела приводит к потере
600 кал. В горячих цехах при температуре воздуха до 50 °С человек
выделяет до 12 л/сут пота. За счет этого выделяется до 8000 ккал.
С медицинской точки зрения потоотделение имеет сущ ествен­
ное значение для поддержания оптимального уровня температуры
тела в условиях повышенной температуры окружающей среды,
Потоотделение,
Рис. 5.9.6. Потоотделение человека при работе: А — при умственной рабо­
те; Б — при механизированной физической работе; В — при немеханизи­
рованной физической работе средней интенсивности (t° С —температура
воздуха в градусах Цельсия)
5 26
Глава 5
особенно в жарких странах. Это исклю чительно важно для лиц,
которые на короткие сроки переезжают на работу из северных
стран в южные. Установлено, что все люди в разной степени
обладают способностью к усиленному потоотделению в условиях
повышенной температуры. Поэтому люди с пониженным пото­
отделением подвержены большему риску получения теплового
удара. С целью тренировки процессов потоотделения рекоменду­
ется систематически использовать разнообразные теплохолодовые
процедуры и, в первую очередь, бани.
В основе всех физических процессов теплоотдачи у человека лежат
физиологические процессы, связанные с сосудистыми реакциями. При
действии высокой температуры сосуды расширяются, при действии
низкой температуры они сужаются. Изменение просвета поверхнос­
тных сосудов кожи регулируется преимущественно симпатической
нервной системой (рис. 5.9.7).
Механизмы саморегуляции температуры тела
При повышении температуры внутренней среды активируются
соответствующие терморецепторы тканей и нейроны центра теплоот­
дачи переднего гипоталамуса. Это приводит к активации перифери­
ческих механизмов теплоотдачи: усилению отдачи тепла с помощью
механизмов физической теплоотдачи и торможению механизмов теп­
лопродукции. Благодаря этим процессам температура тела снижается
(см. рис. 5.9.1).
При снижении температуры внутренней среды за счет возбужде­
ния соответствующих терморецепторов тканей и действия охлажден­
ной крови на нейроны центра теплопродукции заднего гипоталамуса
происходит активация периферических механизмов теплопродукции
и торможение механизмов теплоотдачи. Благодаря этому температура
тела повышается.
Аналогичные механизмы включаются при температурных воз­
действиях на терморецепторы кожи. При действии на терморецеп­
торы кожи повышенной температуры нервным путем активируется
центр теплоотдачи переднего гипоталамуса, и благодаря включению
механизмов теплоотдачи температура тела понижается. При действии
на терморецепторы кожи пониженной температуры возбуждается
центр теплопродукции, и за счет механизмов теплопродукции тем­
пература тела повышается.
В деятельность функциональной системы включается и внешнее
поведенческое звено. Человек поддерживает постоянство температу-
Висцеральные функции
5 27
Рис. 5.9.7. Рефлекторные механизмы потоотделения
ры тела за счет одежды, жилища, обогрева или водно-воздушного
охлаждения.
В механизмах саморегуляции температуры тела участвует под­
кожная жировая клетчатка, обладающая малой теплопроводностью.
Слой подкожной жировой клетчатки увеличен у жителей север­
ных широт. В терморегуляции принимает участие расположенный в
области спины под лопатками бурый жир. Этот высококалорийный
жир способствует теплопродукции, особенно у грудных детей, у
5 28
Глава 5
которых еще не сформированы механизмы повышения температуры
тела за счет мышечной дрожи.
Еще одним механизмом саморегуляции температуры тела явля­
ется изменение толщины теплоизоляционного шерстного покрова
животных или перьевого птиц. Сокращения гладких мышц, рас­
полагающихся у корней кожных волос животных или перьев птиц,
приводит к увеличению толщины шерстного или перьевого покрова.
Это предохраняет животных и птиц от переохлаждения. С этим меха­
низмом тесно связана сезонная линька шерсти и перьев у животных
и птиц. У человека этот механизм приобрел рудиментарную форму
в виде так называемой «гусиной кожи» или мурашек. Для изменения
интенсивности теплоотдачи человек целенаправленно регулирует
толщину слоя теплоизолирующей одежды.
Локальная саморегуляция температуры тела
Разные отделы тела обладают локальной саморегуляцией темпе­
ратуры. Примером локальной саморегуляции температуры является
изменение размеров мошонки у мужчин. При низкой температуре
мошонка за счет сокращения мышц укорачивается. При высокой
температуре — расслабляется. Такой механизм предохраняет семен­
ники яичек от перегрева или охлаждения, оказывающих неблагопри­
ятное влияние на сперматогенез.
Примером локальной саморегуляции температуры тела является
изменение тонуса сосудов кожи. На холоде кровеносные сосуды кожи,
главным образом артериолы, сужаются. Раскрываются дополнитель­
ные артериовенозные анастомозы, и тем самым большее количество
нагретой крови поступает в сосуды брюшной полости. Это ведет к
ограничению теплоотдачи. Кроме того, прохладная венозная кровь,
возвращаясь внутрь тела по венулам, расположенным рядом с артериолами, захватывает большую долю тепла, отдаваемого артериальной
кровью. Такая система называется противоточным теплообменником.
Суммарным эффектом такой системы является снижение теплоот­
дачи. Одновременно происходит пилоэрекция, сопровождающаяся у
человека появлением «гусиной кожи» (рис. 5.9.8Б).
Рис. 5.9.8. Потоотделение, подкожный кровоток и пилоэрекция при измене­
нии температуры окружающее среды: А —при нагревании организма; Б —при
охлаждении организма (М —мышца, выпрямляющая кожный волос)
Висцеральные функции
5 29
530
Глава 5
При высокой окружающей температуре кровь минует подкожный
противоточный теплообменник. Подкожные капилляры наполняют­
ся кровью, и теплоотдача увеличивается (см. рис. 5.9.8А).
Гормональная терморегуляция
Саморегуляцию температуры тела определяет ряд гуморальных
факторов. В процессах теплопродукции принимают участие гормо­
ны гипофиза, щитовидной железы и надпочечников. Удаление этих
желез снижает устойчивость организма к воздействию высоких и
низких температур.
При низкой температуре окружающей среды в процессах само­
регуляции температуры тела преимущественно участвуют соматотропный, тиреотропный гормоны гипофиза, гормоны щитовидной
железы и адреналин. Адреналин усиливает окислительные процессы
в тканях, в частности в мышцах. Кроме того, он сужает кожные сосу­
ды и тем самым уменьшает теплоотдачу.
В процессах саморегуляции в^условиях повышенной температу­
ры окружающей среды снижается секреция тиреотропного гормона
гипофиза. В этом случае адреналин, взаимодействуя с (32-адренорецепторами артериол кожи и способствует их расширению, в резуль­
тате чего усиливается теплоотдача.
Показано, что у человека и приматов доминирую щ им медиа­
тором, регулирующим центральные механизмы терморегуляции
при холоде, является серотонин, который увеличивает теплопро­
дукцию.
Под влиянием микробной инвазии в мозге вырабатывается эндо­
генный пироген — интерлейкин-1. Интерлейкин-1 увеличивает воз­
будимость чувствительных к холоду нейронов гипоталамуса и умень­
шает импульсную активность нейронов, чувствительных к теплу.
В этих процессах промежуточная роль принадлежит метаболитам
арахидоновой кислоты — простагландинам группы Е, а также адренокортикотропному гормону гипофиза.
Условно-рефлекторная терморегуляция
Показано, что если у собаки сочетать действие условного сигнала,
например света, с введением теплой воды в желудок, то вырабаты­
вается условный рефлекс, когда в ответ на включение только одного
света без введения теплой воды в желудок изменяются процессы
теплоотдачи и теплопродукции.
Висцеральные функции
531
Отмечено, что у людей, работающих в условиях жарких цехов или
в холодильниках, привычная рабочая обстановка условно-рефлекторно изменяет процессы терморегуляции организма.
Динам ика температуры тела в различных условиях
Терморегуляция — динамический процесс постоянного взаимодеиствия процессов теплопродукции и теплоотдачи в зависимости от
состояния человека и температуры окружающей среды. Изменение
процессов терморегуляции происходит при приеме пищи, при дви­
гательной активности человека, при изменении температуры окру­
жающей среды.
Оптимальный уровень температуры тела у человека в легкой одеж­
де поддерживается механизмами саморегуляции при температуре
окружающей среды в пределах 18—20 °С, а у обнаженного человека —
при температуре 26—28 °С. Такая температура окружающей среды
получила название температуры комфорта.
Повышение температуры тела называется гипертермией.
Нагревание человека при температуре воздуха выше температуры
крови происходит за счет теплоизлучения и теплопроведения из
окружающей среды в тело человека. При повышении температуры
тела дыхание и сердцебиение учащаются. При длительных воздейс­
твиях это может привести к алкалозу и тепловому удару, сопровожда­
ющемуся потерей сознания. Некоторые животные, в частности, соба­
ки, за счет учащения дыхания усиливают испарение выделяющейся
воды с поверхности слизистой оболочки дыхательных путей.
Длительность процедуры нагревания организма различна в раз­
ных банях. В русской бане воздух максимально насыщен водяными
парами. У находящегося в бане человека потоотделение не сопро­
вождается испарением, что приводит к быстрому нагреву его тела
и требует охлаждения человека в бассейне или душе. Многократное
чередование нагревания и охлаждения имеет оздоровительное дейс­
твие на организм и повышает его устойчивость к неблагоприятным
воздействиям.
В финской сауне поддерживается относительно сухой горячий
воздух. Потоотделение сопровождается испарением, что приводит
к теплоотдаче. За счет этого человек может более длительное время
находиться в сауне, чем в бане.
Одним из условий оздоровительного действия бани является смена
тепловых и холодовых воздействий. Специальные наблюдения пока­
5 32
Глава 5
зали, что оба эти воздействия ведут к стрессорным состояниям. За
счет периодической смены высокой температуры парильного отделе­
ния бани на низкую температуру воды бассейна или реки поочередно
активируются механизмы теплоотдачи и теплопродукции. При этом
происходит тренировка механизмов терморегуляции. После адек­
ватных теплохолодовых процедур улучшаются защитные механизмы
поддержания постоянной температуры тела при охлаждении или
нагревании организма.
При снижении температуры ядра тела до 29—30 °С возникает гипо­
термия. Интенсивность метаболических процессов в тканях замед­
ляется. Факт снижения интенсивности метаболических процессов
при температурных воздействиях позволил использовать этот прием
при операциях на сердце. При этом сердце временно выключается
из кровообращения для оперирования его в «сухом состоянии».
При снижении температуры тела оказалось возможным значительно
увеличивать сроки остановки кровообращения за счет снижения
интенсивности метаболических процессов и перехода их на уровень
гипо- и анабиоза. Снижение температуры тела происходит с помо­
щью физической и химической гипотермии. В первом случае субъек­
та обкладывают льдом или погружают в холодную ванну. Процедура
физического охлаждения оказалась сложной из-за возникающего при
этом холодового стресса. Во втором случае центр теплопродукции
блокируют фармакологическими препаратами и уже на этом фоне
проводят охлаждение организма.
Теория гипотермии и гипобиоза еще требует серьезной разра­
ботки. Однако человечество под влиянием идей основоположника
учения об анабиозе В.М. Бахметьева мечтает о том времени, когда в
организме человека возможно будет произвольно, как маятник часов,
остановить метаболические процессы и снова запустить метаболизм
при необходимости. Такая задача особенно ценна для космонавтики.
Космические, особенно дальние, полеты не могут обеспечить боль­
шой груз продуктов питания для космонавтов. Поэтому временное
погружение их в состояние произвольно регулируемого анабиоза
является перспективной проблемой будущего.
Термоощ ущ ения
Термоощущения являются субъективным отражением в сознании
человека нагревания или охлаждения различных частей его тела.
Субъективность термоощущений зависит от теплопроводности окру­
жающей среды. Теплопроводность влажного воздуха больше, чем
Висцеральные функции
5 33
сухого. Поэтому дождливая погода при температуре 0—5 °С субъек­
тивно оценивается человеком как более неблагоприятная, чем сухая
зимняя погода при температуре от —10 до -20 °С. При комнатной
температуре прикосновение к металлическому предмету вызывает
чувство холода. Деревянный предмет при той же температуре оце­
нивается как теплый. Это обусловлено большей теплопроводностью
металла, чем дерева.
Тепловые ощущения зависят от величины потоотделения и испа­
рения с поверхности тела. Усиление потоотделения субъективно оце­
нивается как перегрев организма. Усиленное испарение сопровожда­
ется чувством холода.
Тепловые ощущения зависят от интенсивности конвекции. При
ветре, сквозняке, течении воды в реке человек оценивает окружа­
ющую среду как более холодную. Появление на теле мурашек или
мышечной дрожи воспринимается как охлаждение организма.
При резком изменении температуры поверхности тела частота
возбуждений терморецепторов кратковременно увеличивается до
100 имп/с (рис. 5.9.9). Максимальное изменение частоты возбуждения
рецепторов возникает при температуре кожи около 10 и 45 °С. При
быстром нагревании или охлаждении тела возбуждаются как теп­
ловые, так и холодовые рецепторы. Возбуждение рецепторов имеет
неспецифический характер и направлено на защиту организма от
перегрева или переохлаждения.
При этом возникаю т парадоксальные ощущения. При резком
нагревании тела возможно возникновение кратковременного ощу­
щения холода. Так, например, при погружении человека в ванну с
горячей водой первоначально возникает чувство холода, вызываю­
щего появление мурашек на теле. Это связано с кратковременным
возбуждением холодовых рецепторов, которые находятся в коже
поверхностно, и поэтому возбуждаются раньше тепловых рецеп­
торов. Затем возникает чувство жжения, вызванное раздражением
тепловых рецепторов. Далее термоощущения сменяются болевыми
ощущениями.
При резком охлаждении возможно возникновение ощущения
горячего. Так, например, при погружении человека в прорубь к
чувству холода примешивается кратковременное ощущение жжения,
связанное с неспецифическим возбуждением тепловых рецепторов.
Затем возникают болевые ощущения.
Выраженность неспецифических ощущений прямо зависит от
разности и скорости изменения температуры. Так, например, пара-
Глава 5
534
Быстрое охлаждение
2—5 и м п./с
100 и м п./с
Б
Быстрый нагрев
Быстрое охлаждение
Рис. 5.9.9. Неспецифическое кратковременное увеличение частоты воз­
буждений терморецепторов кожи кисти руки при резком изменении
кожной температуры: А — значения частоты нервных импульсов тепловых и холодовых терморецепторов (TP, ХР) при быстром нагревании и
охлаждении кожи; Б —динамика импульсации терморецепторов при быс­
тром нагреве или охлаждении кожи (t° С —температура кожи в градусах
Цельсия; t —время)
доксальные ощущения холода при погружении человека в горячую
ванну чаще возникают после его охлаждения на морозе.
Парадоксальные термоощущения возникают при заболеваниях,
сопровождающихся изменением температуры тела. В частности, при
инф екционных заболеваниях продукты метаболизма патогенных
Висцеральные функции
5 35
микроорганизмов возбуждают центр теплопродукции. Эти продукты
являются углеводно-белковыми молекулами и называются пирогенными веществами. Увеличение температуры крови с 37 до 41 °С уско­
ряет иммунные процессы защиты организма от воспаления. Однако
увеличение температуры ядра тела происходит за счет сужения сосу­
дов в конечностях тела, что уменьшает теплоотдачу от конечностей в
окружающую среду. Температура рук и ног понижается, что приводит
к возбуждению холодовых рецепторов. Для согревания конечностей
рефлекторно возникает мышечная дрожь. Больной человек оцени­
вает эти изменения как замерзание тела, несмотря на увеличение
температуры во внутренних органах.
При окончании заболевания продукты метаболизма м икро­
организмов прекращают действовать на центр терморегуляции.
Уменьшается тонус центра теплопродукции и увеличивается тонус
центра теплоотдачи. Периферические сосуды расширяются, обес­
печивая течение нагретой крови через охлажденные конечности.
Усиливаются теплоизлучение и теплопроведение от рук и ног в
окружающую среду. Потоотделение с последующим испарением воды
усиливает процессы теплоотдачи. Нагрев конечностей, сопровожда­
ющийся потоотделением, субъективно воспринимается как нагрев
организма. Выздоравливающий человек оценивает подъем темпера­
туры тела как ухудшение самочувствия, несмотря на нормализацию
температуры ядра тела.
Глава 6
АНАЛИЗАТОРЫ
6.1. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ
ОРГАНИЗАЦИЯ АНАЛИЗАТОРОВ
Живой организм постоянно воспринимает воздействия внешней
и внутренней среды. Совокупность возбудимых структур перифе­
рической нервной системы и ЦНС, осуществляющих восприятие
и анализ воздействий окружающей и внутренней среды, получила
название анализаторов.
Все структуры, входящие в состав анализаторов, относятся к
афферентным, т.е. проводящим возбуждения от периферии в ЦНС.
Анализаторы «разлагают» внешний мир на элементы, а затем транс­
формируют раздражение в ощущение, после которого формируется
определенная тактика поведения организма. Клетки коры больших
полушарий осуществляют, по И.П. Павлову, высший анализ и синтез
воздействий внутренней и внешней среды.
Разнообразные раздражители внешнего мира, воздействуя на
живой организм, формируют в его ЦНС специфические реакции,
определяющие феномен психологического отражения. Существует
несколько уровней отражения. Это, во-первых, ощущение — отраже­
ние отдельных свойств раздражителя, во-вторых, восприятие — про­
цесс анализа и синтеза отдельных ощущений путем сопоставления
их с прежним опытом и, наконец, представление — создание образов
без непосредственного воздействия раздражителя на органы чувств.
Процессы психологического отражения обеспечиваются специаль­
ными аппаратами — анализаторами. Анализатор представляет собой
совокупность возбудимых структур периферической нервной сис­
темы и ЦНС, осуществляющих анализ внешних раздражителей с
последующим синтезом психического образа.
Анализаторы
5 37
Анализаторы в системной организации поведения
Анализаторы в системной организации физиологических функ­
ций — динамические самонастраивающиеся и саморегулирующиеся
организации. С системных позиций, при формировании поведения
афферентные потоки возбуждений всегда имеют конкретное фун­
кциональное значение. Они различаются не только по сенсорной
модальности афферентного возбуждения, но и по роли в механизмах
системной архитектоники поведенческих актов, в частности, в меха­
низмах афферентного синтеза и аппарата предвидения результатов
действия. В системной организации поведения различают следую­
щие потоки афферентных возбуждений: обстановочную афферентацию, пусковую афферентацию и обратную афферентацию.
Обстановочная афферентация
Участвует в формировании стадии афферентного синтеза. Она
складывается из воздействий на организм всей совокупности вне­
шних факторов, составляющих конкретную обстановку, на фоне
которой формируется поведенческий акт. По отношению к человеку
обстановочными раздражителями выступают многочисленные соци­
альные факторы: нравственные, культурные, бытовые и др.
Пусковая афферентация
Под пусковой афферентацией понимают афферентные возбуждения,
возникающие при воздействии на организм стимулов, непосредственно
развертывающих поведенческий акт. В качестве пусковых стимулов
могут выступать многочисленные условные раздражители и время.
Обратная афферентация
Афферентные возбуждения приносят в структуры головного
мозга информацию о возникшей в организме потребности и об
успешности достижения результатов целенаправленного поведения.
В функциональных системах организма обратная афферентация в
виде нервной импульсации от интерорецепторов и гуморальных
влияний на центры нервной системы постоянно сигнализирует о
степени достижения полезных для организма результатов, в частнос­
ти, об оптимальном уровне тех или иных показателей внутренней
среды организма. В любом поведенческом акте, который направлен
5 38
Глава 6
на получение какого-либо приспособительного, т.е. полезного для
организма, результата, обратная афферентация информирует об
этом результате совершенного действия, давая возможность орга­
низму в целом оценить степень успеха выполняемого им действия
и своевременно исправить ошибки. Например, у человека на основе
метаболических процессов и взаимодействия организма и внешней
среды может созреть намерение выпить стакан чая. Он протягивает
руку к стакану с чаем, берет его. Тактильное возбуждение ладони
поверхностью стакана, температурное, весовое, наконец, зритель­
ное раздражение от контакта руки со стаканом — все эти афферен­
тные раздражения в сумме дают обратную информацию о том, что
результат действия соответствует исходному намерению. К тому же
при осуществлении этого действия само продвижение руки к ста­
кану непрерывно регулируется проприоцептивной сигнализацией,
свидетельствующей о правильном и соответствующем распределе­
нии сокращенных мышц, о степени напряжения руки, о высоте ее
положения по отношению к намеченной цели и т.д.
Направляющая и результативная обратная афферентация
Обратная афферентация, возникающая при двигательном акте,
разделяется на две различные категории: направляющую движе­
ние и результативную афферентацию. Направляющая афферентация
представлена в основном импульсами от проприорецепторов мышц,
осуществляющих движение, результативная — всегда комплексная
и может включать афферентные потоки от фоторецепторов сетчат­
ки, тактильных, вкусовых, слуховых и прочих рецепторов. В свою
очередь, обратная результативная афферентация подразделяется на
поэтапную обратную афферентацию и санкционирующую обрат­
ную афферентацию, которая возникает при достижении субъектом
потребного результата, закрепляет наиболее успешную интеграцию
афферентных возбуждений и завершает функциональный системо­
квант поведения (например, «хочу пить» — «напился»).
Классические представления И.П. Павлова об анализаторе вклю­
чают в его состав три части: периферический отдел, проводнико­
вый отдел и центральный отдел. Периферический отдел состоит из
рецепторов, дополнительного аппарата и первичного афферентного
нейрона. Проводниковый отдел состоит из чувствительных нервов и
специфических афферентных путей спинного мозга и ствола мозга.
Центральный отдел представляет собой участки сенсорной коры
головного мозга, к которым поступают возбуждения конкретной
Анализаторы
5 39
сенсорной модальности. В функционировании анализаторов проявляютсяпротивоположные тенденции: с одной стороны, стремление
извлечь как можно больше информации, а с другой — не допустить
перегрузки ЦНС избытком ненужной информации. В поведенческом
акте информация, полученная анализаторами, используется не толь­
ко при подготовке к действию, но и в процессе выполнения самого
действия. Анализаторы при этом функционируют как следящие сис­
темы, настроенные на поиск и ожидание информации.
Периферический отдел анализатора
Основным компонентом периферического отдела являются рецеп­
торы, представляющие собой специализированные нервные оконча­
ния, которые преобразуют энергию внешнего раздражителя в энер­
гию электрического потенциала.
Классиф икация рецепторов
Классификация рецепторов осуществляется по ряду критериев. По
природе адекватного («родственного») раздражителя бывают механо-,
фото-, терморецепторы и др. По количеству эффективно действующих
раздражителей выделяют моно-, би-, три-, полимодальные рецепторы.
11о расположению в организме рецепторы делят на экстеро- и интерорецепторы. К первым относят рецепторы органов чувств, ко вторым —
рецепторы внутренних органов. По уровню чувствительности разли­
чают низко- и высокопороговые "рецепторы. По скорости адаптации
выделяют быстро адаптирующиеся (тельца Пачини), медленно адап­
тирующиеся (проприоцепторы) и неадаптирующиеся (вестибулярные)
рецепторы. Кроме того, рецепторы различаются по тем или иным видам
кодирования информации о действующем раздражителе. По механиз­
мам работы рецепторы бывают первичночувствующие и вторичночувствующие. Совокупность рецепторов, иннервируемых разветвлениями
одного чувствительного волокна, называют рецептивным полем.
Механизмы работы рецепторов
Процесс, обеспечивающий адекватный ответ сенсорного рецепто­
ра на раздражитель, называется трансдукцией.
В первичночувствующем рецепторе (например, в тельце Пачини) раз­
дражитель оказывает специфическое действие на мембрану нервного
окончания, что приводит к повышению ее проницаемости, прежде
всего, для ионов натрия. В участке действия раздражителя возника­
ет рецепторный потенциал, который вызывает быстрые изменения
540
Глава 6
в структуре белка тубулина, что приводит к изменению состояния
связанных с ним мембранных рецепторов и возникновению потен­
циала действия в сенсорном нейроне. Потенциал действия распро­
страняется по нервному волокну к структурам ЦНС (рис. 6.1).
Во вторичночувствующем рецепторе (например, в волосковых клет­
ках органа Корти) процессы специфического действия раздражителя
на мембрану, повышения ее проницаемости для ионов и возникно­
вения рецепторного потенциала протекают в специализированных
рецепторных клетках. Возбужденная рецепторная клетка выделяет
медиатор, который, воздействуя на постсинаптическую мембрану
чувствительного волокна, вызывает возникновение генераторного
потенциала с быстрыми изменениями структуры белка тубулина.
Изменения структуры тубулина в электрогенных участках сенсор­
ного нейрона приводят к изменению возбудимости клеточной мем­
браны и генерации потенциала действия, распространяющегося по
нервному волокну к структурам ЦНС (см. рис. 6.1).
Рис. 6.1. Процессы, протекающие в первичночувствующих и вторичночувствующих рецепторах: А. 1 — рецепторный потенциал, возникаю­
щий в инкапсулированном окончании (первичночувствующий рецептор);
2 — распространяющийся потенциал действия, возникший в чувстви­
тельном волокне; Б. 1 — рецепторный потенциал в специализированной
рецепторной клетке (вторичночувствующий рецептор); 2 — генераторный
постсинаптический потенциал, возникший в результате действия медиа­
тора М, который приводит к формированию распространяющегося потен­
циала действия (3) в чувствительном волокне
Анализаторы
541
Свойства рецепторов
• Специфичность рецепторов состоит в их высокой чувствительнос­
ти к адекватным раздражителям. Так, ощущение горького возни­
кает при раздражении рецепторов языка 0,000 005 г стрихнина.
В 1840 г. И. Мюллер сформулировал «закон специфической энергии
органов чувств». Согласно этому закону, качество ощущений не
зависит от характера раздражителя, а определяется всецело зало­
женной в чувствительных системах специфической энергией,
которая представляет собой прирожденное, неизменное «жиз­
ненное качество» организма.
• Рецепторы обладают способностью к кодированию, т.е. к преобра­
зованию и передаче информации в условной форме кода. Именно
благодаря кодированию информация сохраняется в структу­
рах головного мозга и используется в его работе. Существует
несколько видов физиологического кодирования:
—кодирование по меченой линии, когда проводящий путь состоит
из последовательности связанных сенсорно-специфических
нейронов, вплоть до участков в сенсорных областях коры; в
большинстве случаев кодирование по меченой линии является
основным видом кодирования сенсорной модальности раздра­
жителя;
—частотное кодирование, обеспечивающее передачу информации
о качестве раздражителя средней частотой следования импуль­
сов. Связь между изменением частоты и значением параметров
раздражителя имеет логарифмическую зависимость и описыва­
ется законом Вебера—Фехнера;
—интервальное кодирование, которое задается временными интер­
валами между импульсами (паттерн) при постоянной средней
частоте их генерации;
—режимное кодирование отражает временные параметры раздра­
жителя и представляет собой пачки из нескольких импульсов в
моменты включения, выключения, включения-выключения, а
также всего времени действия раздражителя;
—пространственное кодирование, дающее информацию о протяжен­
ности и конфигурации действующего раздражителя. Данный
вид кодирования достигается работой большого числа рецеп­
торов, каждый из которых посылает импульсы по одному из
множества параллельно идущих афферентных волокон.
• Одно из важных свойств рецепторов — адаптация к действию
раздражителя, выражающаяся в изменении импульсации рецеп­
Глава 6
542
тора на фоне продолжающегося воздействия на него раздражи­
теля. Рецепторы бывают быстро адаптирующимися, у которых
быстрый стимул вызывает короткий ответ, и медленно адапти­
рующимися, отвечающими на такой же раздражитель чередой
импульсов. Адаптация может проявляться также в виде сенси­
билизации, которая проявляется в снижении порогов и повы­
шении возбудимости рецепторов. Причинами адаптации слу­
жат изменения в околорецепторном пространстве, определенная
динамика физико-химических процессов, протекающих в самом
рецепторе, а также нисходящие регулирующие влияния нервных
центров. Показана роль симпатического отдела ВНС в настройке
рецепторов к адекватному раздражителю.
• Под функциональной мобильностью рецепторов понимается спо­
собность органа чувств реагировать на раздражение внешней
и внутренней среды большим или меньшим числом актив­
ных рецепторов. Уровень функциональной мобильности опре­
деляется условиями окружающей среды и состоянием целого
организма. Функциональная мобильность представляет собой
функциональный резерв реагирования рецепторов и изменя­
ется под воздействием гуморальных и центральных влияний.
Коэффициент функциональной мобильности рассчитывается
как отношение числа активных рецепторов к общему числу
рецепторов.
• Н адежность рецепторов обеспечивается функциональной избыточ­
ностью, которая состоит в дополнительных импульсах в ответе
рецептора, и морфологической избыточностью — дублированием
каналов связи от соседних рецепторов.
• Рецепторы образуют функциональные рецептивные поля, кото­
рые представляют собой совокупность рецепторов, активиру­
ющих один сенсорный нейрон. Клеточные элементы рецептив­
ного поля могут взаимодействовать друг с другом с помощью
латерального торможения. Так, после механического толчка,
нанесенного в область чувствительности отдельного механо­
рецептора кожи, рецепторы поля в течение короткого времени
оказываются полностью заторможенными и не отвечают на
действие пороговых раздражителей.
Проводниковый отдел анализаторов
Проводниковый отдел анализаторов состоит из периферических
чувствительных нервов и проводящих путей ЦНС. Среди проводящих
Анализаторы
543
путей можно выделить сенсорно-специфические пути через ядра тала­
муса, неспецифические пути через структуры ретикулярной формации
и ассоциативные пути, идущие к ассоциативной коре. В сенсорных и
неспецифических ядрах ствола мозга поступающая информация под­
вергается процессам фильтрации и дублирования, а также нисходяще­
го контроля со стороны расположенных выше отделов ЦНС.
Центральный отдел анализаторов
Центральный корковый отдел анализаторов состоит из ядра и
рассеянных элементов, которые образуют первичные, вторичные и
третичные зоны. Он обладает чрезвычайно сложной цитоархитек­
тоникой, включающей в себя соединение нейронов в виде колонок,
цилиндров, дисков. В центральных отделах анализаторов идут про­
цессы анализа сигнала путем декодирования поступившей информа­
ции с последующим синтезом психического образа и его распозна­
ния гностическими нейронами коры мозга. Различные проводящие
афферентные пути через возбуждение специальных подкорковых
структур в конечном счете приносят импульсы возбуждения в соот­
ветствующие области коры большого мозга — к высшему центрально­
му конечному звену любого анализатора. Вместе со специфическими
афферентными возбуждениями в кору поступают и неспецифические
восходящие возбуждения, которые формируются на уровне подкор­
ковых активирующих структур головного мозга — ретикулярной
формации, гипоталамуса и др. В свою очередь, возбуждения из коры
головного мозга адресуются лимбическим образованиям, формируя
тем самым эмоциональное восприятие раздражения.
Электрическое раздражение проекционных областей коры при
нейрохирургических операциях у человека сопровождается спе­
цифическими ощ ущ ениями. Например, при раздражении сома­
тосенсорной коры возникает чувство прикосновения к коже, при
раздражении зрительной коры возникают зрительные образы, при
раздражении слуховой коры появляется ощущение звучания. При
раздражении ассоциативных областей — лобной, височной, теменно-затылочной — возникают интегральные образы, нередко связан­
ные с прошлым жизненным опытом субъектов.
Свойства анализаторов
• Наличие скрытого периода ощущения. Это время, в течение кото­
рого возбуждение проходит все отделы анализатора. Скрытый
период может зависеть от силы раздражителя.
544
Глава 6
• Наличие абсолютного порога возникновения ощущения (порога
ощущения), что связано с пороговым значением силы раздраже­
ния. Слабые подпороговые раздражители не вызывают ощуще­
ния, какой бы сенсорной модальности они ни были.
• Наличие дифференциального порога возникновения ощущения (поро­
га различия), другими словами, факт изменения его различных
параметров. При определении дифференциального порога ощуще­
ния установлена зависимость, которую описывает закон Э. Вебера
(1831): величина, на которую требуется увеличить силу раздражите­
ля, чтобы вызвать едва заметное усиление ощущения, находится в
постоянном отношении к исходной величине этого раздражителя,
т.е. она составляет всегда определенную часть этой величины.
А /// = К,
где J — сила раздражителя; ДJ — прирост раздражения; К — посто­
янная величина.
Формула Г. Фехнера выражает закон Вебера: ощущение увеличи­
вается пропорционально логарифму силы раздражения (рис. 6.2.):
S = а х log R + b ,
где S — величина ощущения; R — величина раздражения; а и b —
постоянные величины.
Однако закон Вебера—Фехнера, принимая во внимание силу раз­
дражителя, не учитывал количественные изменения во времени
числа функционирующих рецепторов на раздражаемой площади.
Как установил П.Г. Снякин, величина константы порогового ощу­
щения зависит не только от пороговой силы раздражения и площади
раздражения, но и от количества функционирующих на ней в дан­
ный момент рецепторов (явление функциональной мобильности).
Под функциональной мобильностью понимается способность изменять
количество активных функциональных рецепторных единиц в ответ
на количественные и качественные колебания раздражений, посту­
пающих из внешней и внутренней среды организма. В этом случае
формула Э. Вебера приобретает следующий вид:
J х S х (Р —р) — const,
где J — пороговая сила раздражения; S — пороговая площадь раз­
дражения; Р — количество рецепторов на раздражаемой площади;
Анализаторы
5 45
LogS
П ороговое
значение
раздражителя
Сила раздражителя
в ф изических единицах
Рис. 6.2. Логарифмическая кривая зависимости величины ощущения от
силы раздражителя
р — количество нефункционирующих в данный момент рецепторов;
(Р -р ) — количество активных в данный момент рецепторов.
• Инерционность возникновения ощущения, связанная с динамикой
распространения процессов возбуждения в центральном отделе
анализатора. Сюда можно отнести явления одновременного и
последовательного контраста, которые состоят в сохранении
ощущений о различных параметрах раздражителя после его уст­
ранения. Так, зрительное ощущение при световом раздражении
сохраняется от 150 до 250 мс после его выключения.
• Активный характер восприятия достигается центральной настрой­
кой на раздражитель, что обеспечивается контролем поступаю­
щей информации на уровне сенсорных реле данного анализато­
ра. Например, уменьшение амплитуды потенциалов в наружном
коленчатом теле кошки, вызванных вспышками света, если
Глава 6
5 46
кошка видит мышь. У человека активный характер восприятия
имеет место при наличии внимания, мотивации или соответс­
твующей инструкции.
• Межанализаторные взаимодействия реализуются на различных
уровнях анализаторов. В частности, уровень функциональной
мобильности холодовых рецепторов повышается в условиях све­
товой адаптации в зрительном анализаторе. Особенно заметны
эти взаимодействия в условиях поражения одного анализатора,
что сопровождается повышением возбудимости различных зве­
ньев другого анализатора.
• Пластичность процессов анализа и синтеза, т.е. изменение уровня
функционирования анализатора для повышения эффективнос­
ти его деятельности. Пластичность процессов, протекающих в
центральном отделе анализаторов, лежит в основе явления сен­
сорного и сенсомоторного научения.
6 .2 . ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Зрительный анализатор представляет собой совокупность опти­
ческой системы, проводящих путей и нервных центров, обеспечива­
ющую восприятие, анализ и интеграцию зрительных раздражений.
Анатомически зрительный анализатор состоит из периферического
отдела, включающего фоторецепторный аппарат сетчатки глаза, зри­
тельный нерв и зрительный тракт, и центрального отдела, включа­
ющего подкорковые и стволовые центры (латеральное коленчатое
тело, подушка таламуса, верхнее двухолмие), зрительную лучистость
и зрительную область коры полушарий большого мозга (цитоархитектонические поля 17, 18 и 19). Структуры зрительного анализатора
связаны со многими образованиями головного мозга, ответственны­
ми за сложные формы поведенческой деятельности.
Оптическая система глаза
Периферическим отделом зрительного анализатора являю тся
фоторецепторы, расположенные в сетчатке глазного яблока. Глазное
яблоко имеет сферическую форму. На глазном яблоке принято выде­
лять ряд опознавательных точек и геометрических линий. Точка,
соответствующая центру роговицы, называется передним полюсом,
а соответствующая центру желтого пятна — задним полюсом глазного
яблока. Прямая линия, соединяющая оба полюса, называется зри­
Анализаторы
547
тельной осью глаза. Наибольшая окружность глаза во фронтальной
плоскости носит название экватора глаза, а окружности, мысленно
проводимые через оба полюса, называются меридианами. В образо­
вании стенок глазного яблока принимают участие три оболочки:
наружная — фиброзная, включающая роговицу и склеру, средняя —
сосудистая, в которой выделяют три части: радужку, ресничное
тело и собственно сосудистую оболочку, и внутренняя — сетчатка.
Ннутренние среды глазного яблока составляют водянистая влага
камер глаза, хрусталик и стекловидное тело. Роговица — передняя
прозрачная часть фиброзной оболочки глазного яблока. Она имеет
форму сегмента сферы со средним радиусом кривизны 8 мм. Толщина
роговицы на всей площади неодинакова и колеблется от 0,8—0,9 мм
и центре до 1,0—1,2 мм на периферии. Разница толщины роговицы в
центре и по периферии обусловливает несколько различную кривиз­
ну ее передней выпуклой и задней вогнутой поверхностей. Роговица
действует как сильная выпуклая линза. Ее преломляющая способ­
ность в 2,5 раза выше, чем у хрусталика. Важнейшая особенность
роговицы — отсутствие в норме в ней кровеносных сосудов, что и
обеспечивает ее прозрачность. Лишь по периферическому краю рого­
вицы вдоль лимба в поверхностных слоях стромы имеется сплетение
кровеносных капилляров, так называемая краевая петлистая сеть.
Источником чувствительной иннервации роговицы являются длин­
ные ресничные нервы — ветви носоресничного нерва, отходящего от
первой ветви тройничного нерва.
Склера — задний большой отдел наружной оболочки глазного
яблока — составляет 5/6 всей оболочки. К склере прикрепляются
глазные мышцы. Передняя часть склеры покрыта конъюнктивой. На
1аднем полюсе глазного яблока выходят волокна зрительного нерва.
Радужка составляет передний отдел сосудистой оболочки глаз­
ного яблока. Она представляет собой круглую диафрагму с расположенным в центре круглым отверстием — зрачком, отделяющую
переднюю камеру глазного яблока от задней. Диаметр радужки
у взрослых: горизонтальный — 12,5 мм, вертикальный — 12 мм,
толщина — в разных отделах 0,2—0,5 мм. Радужка не располагает­
ся строго во фронтальной плоскости, она несколько конусовидно
выпячена вперед вследствие того, что позади нее расположен хрус­
талик. Мышца, суживающая зрачок (сфинктер зрачка), состоит из
гладкомышечных клеток, расположенных циркулярно в переднем,
соединительнотканном, слое радужки вблизи ее зрачкового края.
Мышца, расширяющая зрачок (дилататор зрачка), состоит из ради­
Глава 6
548
ально расположенных миоэпителиальных клеток в заднем, пигмен­
том, слое радужки. Ф ункционально радужка представляет собой
диафрагму, регулирующую поток света в полость глаза. При этом
просвет зрачка регулируется рефлекторно как уровнем освещеннос­
ти, так и расстоянием до рассматриваемого предмета. Различия в
количестве пигментов определяют различия в цвете радужки.
Хрусталик — основная часть аккомодационного аппарата глазного
яблока. Он имеет вид двояковыпуклой прозрачной линзы непосто­
янной кривизны, расположен фронтально в полости глазного яблока
между радужкой и стекловидным телом. У взрослых экваториальный
диаметр хрусталика находится в пределах 9—10 мм, сагиттальный раз­
мер — 3,7—5 мм. Радиус кривизны передней поверхности при покое
аккомодации составляет 10 мм, задней — 6 мм. Хрусталик несколько
смещен медиально и вниз относительно оптической оси глаза.
Ресничное тело — утолщенная часть сосудистой оболочки глазно­
го яблока. В ресничном теле выделяют два главных слоя: наружный —
мышечный и внутренний — сосудистый. Мышечный слой образует
ресничная, или цилиарная, мышца. Ресничная мышца представля­
ет собой совокупность разнонаправленных групп гладкомышечных
клеток, которые начинаются от внутренней склеральной шпоры на
внутренней поверхности склеры вблизи лимба и теряются в сосудис­
той оболочке. Ресничная мышца является важной частью аккомода­
ционного аппарата глаза. При сокращении циркуляторных волокон
ресничные отростки приближаются к центру ресничного кружка, и
ресничные связки ослабляются, что ведет к увеличению выпуклос­
ти хрусталика. Одновременное сокращение меридиональных воло­
кон подтягивает заднюю часть сосудистой и ресничной оболочек,
уменьшая фокусное расстояние светового пучка. При расслаблении
вследствие эластичности ресничное тело принимает исходное поло­
жение, ресничные связки натягиваются, что приводит к уплощению
хрусталика. Чувствительная иннервация ресничного тела происходит
из глазного нерва — первой ветви тройничного нерва.
Расчет величины изображения предмета на сетчатке производят на
основании правила подобия треугольников, вершины которых лежат
в узловой точке (рис. 6.3).
X = J а/Ъ,
где X — величина изображения предмета на сетчатке; J — вели­
чина предмета; а — расстояние от узловой точки до сетчатки
Анализаторы
5 49
Рис. 6.3. Построение изображения на сетчатке, а - а ’ — главная оптическая
ось глаза; б — узловая точка; в — главный задний фокус; в' — главный
передний фокус; г—г' — главная плоскость линзы; д —д' — рассматриваемый
предмет; е—е' — его изображение на сетчатке
(у взрослого — около 17 мм, у новорожденного — 11 мм); b — рассто­
яние от предмета до роговицы глаза плюс расстояние от передней
поверхности роговицы до узловой точки (у взрослого — около 7 мм,
у новорожденного — 5,5 мм).
Оптическая система глаза обеспечивает построение изображения
на сетчатке и определяет остроту зрения человека, которая представ­
ляет собой способность глаза дифференцировать детали изображе­
ния. Расчет остроты зрения проводится по следующей формуле:
V = d/D,
где V — острота зрения; d — расстояние от испытуемого до таблицы,
м; D — расстояние, с которого данная строка правильно читается
нормальным глазом, м.
Угловой предел разрешающей способности глаза человека состав­
ляет 0,62'. За нормальную остроту зрения принимается одна угловая
минута, которая обозначается единицей.
5 50
Глава 6
Нормативные показатели:
• нормальная острота зрения — 1,0;
• пониженная острота зрения — 0,8 и ниже;
• повышенная острота зрения — 1,5-2,0.
Аккомодация — приспособление глаза к ясному видению объ­
ектов, находящихся на разных расстояниях (рис. 6.4). Ресничные
Рис. 6.4. Аккомодация хрусталика: А — ход лучей через оптическую систе­
му глаза при восприятии относительно удаленного предмета. Изображение
на сетчатке является уменьшенным, перевернутым и действительным;
Б — ход лучей через аккомодированную оптическую систему глаза, в кото­
рой хрусталик имеет меньшее фокусное расстояние. Изображение стро­
ится на сетчатке. Оно перевернутое, действительное, но имеет большие
размеры
Анализаторы
551
мышцы иннервируются парасимпатическими волокнами глазодви­
гательного нерва. Далекие предметы нормальный глаз рассматри­
вает без напряжения аккомодации, т.е. без сокращения ресничной
мышцы. Ближайшая точка ясного видения благодаря аккомодации
находится на расстоянии 10 см от глаза. С возрастом точка ясного
видения удаляется — от 6 см у детей до 100 см у лиц старше 60 лет.
Точка ясного видения изменяется при различных эмоциональных
состояниях человека.
Периферический отдел зрительного
анализатора
Сетчатка — периферическая часть зрительного анализатора. Она
обеспечивает зрительное восприятие за счет преобразования свето­
вой энергии в нервный импульс, передающийся по цепи нейронов
в корковый зрительный центр. Сетчатку можно рассматривать как
вынесенную на периферию часть головного мозга. В зрительной
части сетчатки особое клиническое значение имеют два анатоми­
ческих образования: диск зрительного нерва и пятно сетчатки. Диск
зрительного нерва представляет собой место соединения оптических
нервных волокон сетчатки и формирования зрительного нерва. Диск
располагается на глазном дне на 2,5—3 мм медиальнее заднего полюса
глаза и на 0,5—1 мм книзу от него. Форма диска круглая или слегка
овальная, вытянутая вертикально. Диаметр его равен 1,5—1,7 мм.
В центре диска имеется углубление, где проходят центральная арте­
рия сетчатки и сопровождающая ее вена. Область диска лишена све­
точувствительных элементов и физиологически представляет собой
слепое пятно. Второе важное образование — пятно сетчатки. Пятно
располагается с височной стороны от диска зрительного нерва на
расстоянии 4 мм. Оно имеет диаметр около 5,5 мм. В центре его рас­
полагается углубление — центральная ямка с ямочкой (foveola ) диа­
метром около 0,2 мм. Пятно составляет центральный отдел сетчатки.
Здесь нет кровеносных сосудов, фоторецепторы представлены только
колбочками, количество нервных волокон в этой зоне минимально.
Это зона максимальной остроты зрения. Только те изображения,
которые формируются здесь, воспринимаются мозгом ясно и четко.
Остальная зрительная часть сетчатки составляет периферический
отдел, где степень четкого видения убывает по направлению к зубча­
тому краю. За рецепторным слоем сетчатки лежит пигментный эпи­
телий, основная функция которого, поглощать лучи света, уменьшая
их рассеивание.
5 52
Глава 6
Фоторецепторы сетчатки представлены палочками и колбочками
(рис. 6.5). Палочки обеспечивают сумеречное черно-белое (скотопическое) зрение. Палочки в количестве 130 млн неравномерно
распределены по периферии сетчатки. Наибольшее их количество
Рис. 6.5. Распределение нейронов сетчатки. При воздействии цветных раз­
дражителей возбуждаются колбочки, плотно расположенные в центре сет­
чатки в области желтого пятна и обеспечивающие высокую остроту зрения
(1 и 2). При воздействии черно-белого раздражителя возбуждаются палоч­
ки, расположенные на периферии сетчатки и обеспечивающие широкое
поле зрения (3 и 4). К центру сетчатки на одну биполярную клетку кон­
вергирует все меньшее число рецепторов, а в самом центре конвергенция
отсутствует и одна колбочка возбуждает одну биполярную (5) и одну ган­
глиозную (6) клетку. На периферии сетчатки большая чувствительность
палочек обеспечивается широкой конвергенцией от 500 рецепторов на
одну ганглиозную клетку; 7 — оптический нерв
Анализаторы
5 53
находится на расстоянии в 20° от центра желтого пятна. Периферия
сетчатки отличается большой чувствительностью к слабому свету.
Это обусловлено тем обстоятельством, что до 500 палочек конвер­
гируют в этом участке сетчатки через биполярные клетки на одну
и ту же ганглиозную клетку. Кроме того, в палочках много больше
зрительного пигмента, чем в колбочках, что определяет их большую
чувствительность. В результате такой связи импульсы от многих
палочек суммируются и вызывают более интенсивную стимуляцию
этих клеток. При поглощении кванта света происходит изомериза­
ция хромоформной части зрительных рецепторов: 11-цис-ретиналь
превращается в трансретиналь. В результате происходят изменения
в белковой опсиновой части рецепторов, родопсин обесцвечивается
и переходит в метародопсин II. Метародопсин II взаимодействует
с другим белком — трансдуцином — и обменивает связанный с
ним в темноте ГДФ (гуанозиндифосфат) на ГТФ. Активированная
молекула трансдуцина, в свою очередь, активирует одну молекулу
фермента фосфодиэстеразы, которая разрушает молекулы цГМФ.
В результате в цитоплазме наружного сегмента рецептора падает
концентрация свободного цГМФ. Это, в свою очередь, приводит к
закрытию ионных каналов в плазматической мембране наружного
сегмента, которые были открыты в темноте и через которые внутрь
рецепторных клеток входили ионы Na+ и Са2+. Уменьшение входа
ионов Na+ приводит к гиперполяризации мембраны и возникно­
вению рецепторного потенциала. При действии света уменьшается
выделение глутамата из пре-синаптических окончаний фоторецеп­
торов, что приводит к гиперполяризации мембран ганглиозных
клеток. Таким образом, входящий ток, возникаю щий в темноте,
может называться темповым током; при освещении глаза он пре­
кращается.
Колбочки в количестве 7 млн находятся в области желтого пятна
и обеспечивают центральное зрение, которое характеризуется чет­
ким восприятием рассматриваемого предмета. Длинные и узкие
наружные сегменты колбочек в области желтого пятна обеспечива­
ют их максимально плотное расположение. Следует отметить, что
только несколько колбочек конвергируют на одной биполярной
клетке, а в центральной ямке одна колбочка связана с одним бипо­
лярным нейроном. Все это обеспечивает хорошую разрешающую
способность зрительной ямки в формировании четкого изображе­
ния. На рис. 6.6 показана острота зрения на различных участках
сетчатки.
Глава 6
554
70°
40"
10“
0*
10°
50°
Рис. 6.6. Острота зрения на различных участках сетчатки. По оси абс
цисс — расстояние от центра сетчатки в градусах
Ганглиозные клетки сетчатки выступают в роли детекторов,
среди которых выделяют детекторы включения-выключения, пря­
мой линии, дуги, края, движения, угла и цвета (рис. 6.7). В сетчатке
млекопитающих и человека различают Р- и М-ганглиозные клетки,
посылающие аксоны в латеральное коленчатое тело. Р-клетки неве­
лики, имеют небольшую скорость проведения возбуждения и их
преимущественно тонические ответы связаны с различением тон­
ких деталей предметов, а также зависят от длины световой волны.
М-клетки обладают быстрыми ответами на сложные стимулы и более
чувствительны к яркости света, чем Р-клетки.
Зрительные пигменты
В палочках сетчатки содержится пигмент родопсин, кото­
рый состоит из белковой части (опсин) и хромоформной части
(11-цис-ретиналь, или альдегид витамина А). Источником витами­
на А являются каротиноиды. При их недостатке развивается «кури­
ная слепота» — затруднение видимости в сумеречное время суток.
В палочках много больше зрительного пигмента, чем в колбочках.
Анализаторы
5 55
I
О
Э
Л
Центр
Периферия
Центр
Периферия
Рис. 6.7. Структура клеточной организации рецептивного поля с оп-центром и динамика ответа нейронов сетчатки на цветовые раздражители:
А — реакция нейрона красно-зеленой системы; Б — реакция нейрона
желто-фиолетовой системы. Отмечается антагонизм в реакциях нейронов
цветоспецифических рецептивных полей
Родопсин лучше всего поглощает световые волны длиной 500 нм.
В наружных сегментах колбочек содержится три типа зрительных
пигментов: чувствительные к синему цвету с максимумом поглоще­
ния световых волн длиной 420 нм, зеленому — с максимум поглоще­
ния 531 нм и красному — с максимумом поглощения 558 нм. Наличие
трех видов пигментов в колбочках с различными показателями пог­
лощения света лежит в основе трехкомпонентной теории цветоощу­
щения. Другая распространенная теория — оппонентных цветов —
основывается на существовании оппонентных цветов (красный и
зеленый, желтый и синий, фиолетовый и оранжевый). Колбочки
5 56
Глава 6
содержат пигменты иодопсин, эритролаб, хлоролаб, реагирующие
на цветовые раздражители. При этом каждая пара колбочек акти­
вируется двумя основными цветами: красный—зеленый, фиолето­
вый—желтый, оранжевый—голубой.
Проводниковый отдел зрительного анализатора
Проводниковый отдел зрительного анализатора состоит из опти­
ческого нерва, ядер верхних бугров четверохолмия и латерального
коленчатого тела.
У человека выделяют несколько уровней анализа зрительной инфор­
мации (рис. 6.8).
Первый уровень — сетчатка и наружное коленчатое тело, нейроны
которых выделяют сигналы из шума, подчеркивая контуры объекта,
его цвет и границы. Нейроны наружного коленчатого тела облада­
ют малыми простыми круглыми рецептивными полями. Наружное
коленчатое тело содержит ретинотопическую карту. В латеральных
коленчатых телах начинается бинокулярное взаимодействие от сет­
чатки правого и левого глаза. Взаимодействие возбуждений на ней­
ронах латерального коленчатого тела осуществляется на основе меха­
низма реципрокного торможения с участием тормозного медиатора
GABA. Здесь происходит взаимодействие афферентных сигналов,
приходящих от сетчатки, с эфферентными возбуждениями из зри­
тельной коры, таламуса и ретикулярной формации, что обеспечивает
процессы избирательного зрительного внимания.
Верхние бугорки содержат ретинотопическую карту. Нейроны
верхних бугорков четверохолмия преимущественно реагируют на дви­
жущиеся в определенном направлении световые стимулы и имеют
бинокулярные входы. Некоторые из них возбуждаются только в том
случае, когда стимул движется через их рецептивное поле в опре­
деленном направлении. В более глубоких слоях бугорков находятся
нервные клетки, которые возбуждаются в моменты, предшествующие
движениям глазных яблок.
Корковый отдел анализатора
Второй уровень анализа зрительной информации — стриарная
кора (17 поле). Стриарная кора содержит ретинотопическую карту,
в которой желтое пятно по сравнению с остальной сетчаткой имеет
преимущественное представительство. Нейроны этого уровня фор­
матируют все зрительное поле на отдельные квадранты с последу­
ющей оценкой положения объекта в поле зрения. Третий уровень —
Анализаторы
5 57
Рис. 6.8. Обработка информации в зрительном анализаторе: А — струк­
туры периферической и центральной нервной системы, участвующие в
организации процессов зрительного восприятия; Б — уровни обработки
сигнала: I — нейроны сетчатки и наружного коленчатого тела (выделение
углов, дуг, цвета и направления движений предмета); II — 17 поле зри­
тельной области коры мозга (построение изображения в том или ином
квадранте зрительного поля); III — 18 и 19 поля зрительной области коры
мозга (формирование инвариантного объемного изображения); IV — ниж ­
невисочная кора и заднетеменная кора (формирование образа, перемеща­
ющегося в определенном направлении, объемного цветного изображения
в конкретной зрительной среде)
престриарная кора (18 и 19 поля) с нейронами, создающими объемное
мобильное изображение, обладающее свойствами инвариантности,
т.е. узнаваемое в любом размере и положении. Четвертый уровень —
структуры нижневисочной и верхнетеменной коры, выполняющие
функции словаря и языка соответственно, процессов зрительного
5 58
Глава 6
опознания. В эти отделы коры поступает информация по P-пути от
Р-клеток сетчатки, обеспечивающая восприятие формы и цвета
объектов, а от М -клеток по М-пути — информация о перемещении
объектов в поле зрения. Нейроны этого уровня — «обучающиеся»
клетки, формирующие конкретное и абстрактное описание объекта.
Как было уже сказано, корковый отдел зрительного анализатора
находится в 17, 18, 19 полях затылочной коры, в составе которых
имеются «простые», «сложные» и «сверхсложные» нейроны, активи­
рующиеся в ответ на зрительные раздражители различной степени
сложности. Нейроны со сверхсложными полями в основном нахо­
дятся в полях 18 и 19 коры мозга. Нейроны с близкими свойствами
образуют глазодоминантные колонки со стороной в 1 мм и высотой в
несколько миллиметров. В основе формирования рецептивного поля
лежит принцип конвергенции к нейрону возбуждений из различных
отделов ЦНС, других элементов той же структуры, а также от рецеп­
торных образований.
Согласно классическим представлениям, ориентационная изби­
рательность корковых нейронов создается в связи с приходом к одно­
му и тому же нейрону импульсов от нервных элементов латерального
коленчатого тела, обладающих круговыми рецептивными полями.
В свою очередь, формирование «сложных» и «сверхсложных» клеток
определяется конвергенцией возбуждений от нейронов с «простыми»
рецептивными полями. Так, при световой стимуляции обоих глаз
активируются до 80% нейронов зрительной коры. В то же время при
восприятии движения световой полоски до 64% клеток были унимо­
дальны, 33 — бимодальны и только 3% — тримодальны.
Кроме того, в 18 поле зрительной области коры мозга, в отличие
от 17 и 19 полей, обнаружены нейроны с диффузионными рецеп­
тивными полями, имеющими большие размеры, но не обладающие
ориентационной и дирекционной избирательностью. Такие нейроны
отвечали за обнаружение различной степени освещенности.
Движения глаз и пространственное зрение
Существуют два механизма пространственного (глубинного) зре­
ния. Первый из них — монокулярный стереокинетический механизм,
который обеспечивает оценку расстояния до предмета на основе
механизма аккомодации при участии проприоцепторов циркулярной
мышцы хрусталика. При этом объемность изображения основана на
том, что при движении наблюдателя величина смещения на сетчатке
расположенных близко предметов больше, чем расположенных вдали.
Анализаторы_____________________________________________________ 5 59
Рис. 6.9. Конвергенция глазных яблок при рассматривании разноуда­
ленных предметов: А — предмет находится на относительном удалении;
Б — предмет находится на близком расстоянии от человека. По мере при­
ближения предмета происходит большее напряжение внутренних прямых
мышц глаз и растяжение наружных. Последнее приводит к активации
расположенных в мышцах мышечных веретен, импульсы от рецепторов
которых поступают в головной мозг и формируют представление о рассто­
янии до рассматриваемого предмета
Второй механизм — бинокулярный конвергентный (рис. 6.9). Он обеспе­
чивает оценку расстояния до предмета по степени конвергенции глаз11 ых яблок и уровню напряжен ия проприоцепторов наружных прямых
мышц глазного яблока. Изображения рассматриваемого предмета
попадают на соответствующие или корреспондирующие точки сет­
чатки правого и левого глаза. Объемность изображения основана на
бинокулярной диспарации глаз. Если фиксировать взгляд на одной
точке предмета, то другая точка того же предмета проецируется на
ином расстоянии от центра лучшего видения на сетчатке правого и
левого глаза.
560
Глава 6
Психофизика зрительного восприятия человека тесно связана с дви­
гательной активностью глазных яблок. Выделяют четыре основных
вида движений: саккады — прыжки взора длительностью 50 мс и
временем фиксации 200 мс, которые имеют место при разглядыва­
нии объекта; медленные следящие движения — при слежении за
перемещением объекта; конвергирующие движения — при фиксации
взгляда на объекте; вестибулярные движения — при повороте голо­
вы или туловища. Ядра глазодвигательных нервов, иннервирующие
мышцы глазного яблока, находятся также под корригирующим влия­
нием мозжечка, обеспечивающим более точное сведение зрительных
осей обоих глаз к одной и той же точке. В мозжечке вестибулярные
и зрительные сигналы взаимодействуют с сигналами, отражающими
положение головы и глаз.
Центры бинокулярного зрения находятся в верхних буграх четве­
рохолмия и в ретикулярной формации среднего мозга. При непод­
вижной позиции наблюдателя бинокулярный механизм осуществля­
ет оценку расстояния до объекта по степени напряжения наружных
прямых мышц глаза. В зрительных центрах происходит бинокуляр­
ное слияние двух изображений в одно (рис. 6.10).
6.3. СЛУХОВОЙ АНАЛИЗАТОР
Слуховой анализатор представляет собой совокупность органа
слуха с рецепторами, слухового нерва с проводящими путями и
участка слуховой коры, которые обеспечивают анализ звукового раз­
дражителя и синтез на его основе звукового образа. Периферическим
отделом анализатора являются механорецепторы, расположенные во
внутреннем ухе.
Рис. 6.10. Центральные механизмы построения зрительного образа:
А — при бинокулярном восприятии предмета изображение строится в зри­
тельных областях обоих полушарий коры; Б — при восприятии предмета
одним глазом его образ также строится в обоих полушариях: от височной
области сетчатки информация идет в то же полушарие, а от назальной —
в противоположное (правое) полушарие; В — при нарушении связей между
полушариями и оптическими трактами образ строится только в одном
(левом) полушарии
Анализаторы
561
*\
Глава 6
5 62
Периферический отдел слухового анализатора
Наружное ухо играет роль локатора, улавливающего звуковые
колебания воздушной среды, передающиеся на барабанную пере­
понку. Скорость звука в воздушной среде составляет 330 м/с. Эти
колебания посредством слуховых косточек среднего уха — молоточ­
ка, наковаленки и стремечка — усиливаются в 22 раза и передаются
на мембрану овального окна и столб жидкости в улитке. Внутреннее
ухо человека представляет собой сложный лабиринт соединенных
между собой каналов, расположенных внутри височной кости. Один
из каналов имеет форму улитки с завитком в два с половиной обо­
рота. Канал улитки имеет в длину 35 мм и с базальной стороны
заканчивается круглым и овальным отверстиями, выходящими в
полость среднего уха и закрытыми эластичными мембранами. По
мере удаления от среднего уха площадь поперечного сечения улитки
уменьшается в 6 раз. Барабанный и вестибулярный каналы улитки
на ее вершине соединяются и заполнены перилимфой со свойствами
плазмы крови. Средний канал замкнут и заполнен более вязкой эндо­
лимфой. В лестнице преддверия и барабанной лестнице находится
перилимфа, имеющая состав, сходный с составом спинномозговой
жидкости. В жидкости средней лестницы имеется много ионов калия
и мало ионов натрия, так что ее состав сходен с внутриклеточной
средой.
В среднем канале на основной мембране лежит орган Корти,
состоящий из одного ряда внутренних и трех рядов внешних рецеп­
торных волосковых клеток. 3500 внутренних и 1200 наружных
волосковых клеток равномерно распределены вдоль улитки и пос­
редством синаптических контактов соединены с волокнами бипо­
лярных клеток слухового нерва. Следует отметить, что до 90%
нервных волокон оканчиваются на внутренних и только 10% —
на наружных волосковых клетках.
Теории слуха
Резонансная теория Т. Гельмгольца исходит из того, что каждое
волокно основной мембраны резонирует со звуком определенной
частоты. Звуки низкой частоты вызывают колебания основной
мембраны ближе к верхушке улитки, а звуки высокой частоты —
ближе к основанию улитки. Подтверждением этой теории слу­
жат опыты Андреева, проведенные в лаборатории И.П. Павлова.
Разрушение основания улитки нарушало условные рефлексы на
Анализаторы
5 63
звуки высокой частоты, а разрушение верхушки улитки — на звуки
низкой частоты.
Теория «бегущей волны» Д. Бекеши. Согласно этой теории, высо­
кочастотные колебания звуков проходят по мембране лиш ь короткое
расстояние, а длинные волны распространяются по всей мембране.
Поскольку основная мембрана улитки расширяется от ее основания
к вершине, начальная часть мембраны служит высокочастотным
фильтром, а длинные волны распространяются по всей мембране
до геликотремы.
Механизм слухового восприятия
Звуковые колебания имеют несколько характеристик: период ( Т ),
частоту (/), амплитуду (Л) и дополнительные гармоники, создающие
тембр звука (рис. 6.11).
Рис. 6.11. Характеристики звукового колебания: Т — период колебания;
А — амплитуда колебания; f — частота колебания, где f = 1/Т; 1 — исход­
ное колебание; 2 — колебание той же частоты, но большей амплитуды;
3 — колебания той же частоты, но меньшей амплитуды; 4 — колебания
более высокой частоты; 5 — колебания той же амплитуды с меньшей
частотой, 6 — наличие дополнительных гармоник в сложном колебании,
определяющих специфический тембр звука
564
Глава 6
Механизм преобразования колебаний стремечка среднего уха в
импудьсацию волокон слухового нерва можно схематически пред­
ставить следующим образом: смещения стремечка и связанной с ним
мембраны овального окна сопровождаются изменениями давления
жидкости (перилимфы) в базальной части вестибулярного канала;
под действием этого приводится в движение рейснерова мембрана,
вязкая жидкость (эндолимфа) среднего канала улитки и базиляр­
ная мембрана, что, в свою очередь, сопровождается колебаниями
давления жидкости в тимпанальном канале улитки и смещения­
ми мембраны круглого окна. Разностное давление, возникающее
в базальной части улитки между вестибулярным и тимпанальным
каналами, распространяется по направлению к геликотреме, образуя
«бегущую волну» давления (рис. 6.12). Вследствие большой податли­
вости мембран, ограничивающих средний канал улитки, скорость
распространения бегущей волны давления значительно меньше ско­
рости распространения звука в жидкости со свойствами перилимфы.
Поскольку податливость среднего канала изменяется вдоль улитки,
скорость волны зависит еще и от расстояния от базальной части
улитки, сильно уменьшаясь по мере продвижения к геликотреме.
По этой же причине амплитуда колебания среднего канала улитки
достигает максимума в определенной точке, положение которой
зависит от частоты входного сигнала, а затем, при дальнейшем про-
Рис. 6.12. А — структура улитки человека; Б — в ответ на высокочастотный
раздражитель возникают колебания проксимальных участков базальной
мембраны и возбуждение находящихся там волосковых клеток; В — в ответ на
низкочастотный раздражитель возникают колебания дистальных участков
базальной мембраны и возбуждение находящихся там волосковых клеток
Анализаторы
5 65
движении, быстро затухает. Звук вызывает колебания улиткового
протока (средней лестницы) попеременно в сторону вестибулярной
и в сторону барабанной лестниц. Результатом такого движения ста­
новится смешение основной и покровной мембран кортиева органа
относительно друг друга. Так как цилии волосковых клеток плотно
соприкасаются с покровной мембраной, их сгибание вызывает воз­
буждение волосковых клеток. При этом в каждой волосковой клетке
возникает рецепторный потенциал, вызывающий высвобождение
медиатора. Сумма рецепторных потенциалов всех возбужденных кле­
ток называется микрофонным потенциалом улитки. Медиатор действует
возбуждающим образом на постсинаптическую мембрану афферен­
тного волокна биполярного нейрона спирального ганглия, что, в
конечном счете, приводит к возникновению потенциалов действия
в волокнах слухового нерва.
Описанный механизм получил название пространственного кодиро­
вания высоты звука. В свою очередь, кодирование интенсивности звука
осуществляется изменением количества импульсов в залпе волоско­
вых клеток. При этом в диапазоне звука 200-3000 Гц кодирование
обеспечивается не частотой импульсации, а активацией внутренних
и внешних волосковых клеток с различным порогом возбуждения.
Это происходит потому, что в данном звуковом диапазоне частота
импульсов должна нести информацию о высоте тона.
Проводниковый отдел слухового анализатора
Проводниковый отдел слухового анализатора представлен слухо­
вым нервом, по которому информация от рецепторов передается к
кохлеарным ядрам, нижним бугоркам четверохолмия и внутреннему
коленчатому телу. Здесь имеются нейроны, отвечающие на включение и
выключение звукового раздражителя, а также на изменение его интен­
сивности. Одна часть нейронов этих структур реагирует на чистые тоны
с малым скрытым периодом, другая реагирует на сложные частотномодулированные тоны, третья — только на разнообразные шумы. При
бинауральном восприятии нейроны верхних олив реагируют по-разному, в зависимости от того, какое ухо получило информацию раньше.
Корковый отдел анализатора
Корковый отдел анализатора находится в слуховой области коры,
нейроны которой обладают функциональной тонотопической органи­
зацией, т.е. имеют наименьшее значение порога возбуждения при опре­
деленных значениях интенсивности звука. Существуют различные
5 66
Глава 6
классы нейронов: реагирующие на начало звука, на окончание звука,
в течение всего времени его действия, на медленное усиление или
уменьшение амплитуды звука, на направление и изменение движения
источника сигнала, новизну и монотонность, а также на модуляции
громкости. В слуховой коре на расстоянии 0,2 мм располагаются ней­
роны с различием чувствительности в четыре октавы. В коре имеются
бинауральные нейроны, реагирующие на одновременное звуковое раз­
дражение правого и левого уха. Слуховая кора головного мозга, наряду
с сенсомоторной и зрительной, участвует в процессах восприятия и
смысловой оценки фразы (рис. 6.13).
Психофизика слухового восприятия человека имеет ту особен­
ность, что субъективная громкость звука зависит не только от его
интенсивности, но и от его частоты. Интенсивность звука, или уро­
вень звукового давления, выражается в белах, где 1 Бел — десятичный
Рис. 6.13. Динамика вовлечения структур головного мозга в восприятие
речи. Восприятие зрительного изображения текста, устного слова и так­
тильного шрифта Брайля осуществляется, соответственно, в зрительных
(1) и соматосенсорных (2) областях коры больших полушарий, импульсы
из которых направляются в угловую извилину (3). В нейронах угловой
извилины обрабатывается зрительная и соматическая информация, кото­
рая вместе с информацией из слуховой области коры (4) в случае устной
речи поступает в центр Вернике, расположенный в височной области коры
мозга (5). Центр Вернике осуществляет декодирование информации, что
обеспечивает понимание смысла слова или фразы
Анализаторы
567
логарифм действующей интенсивности звука к его пороговому зна­
чению. На практике используется 1 дБ, равный 0,1 Бела. Единицей
высоты тона служит мел. Тон с частотой 1000 Гц при интенсивности
60 дБ имеет высоту 1000 мелов. Громкость звука выражается в сонах.
I сон — громкость тона с частотой 1000 Гц и интенсивностью 40 дБ
над порогом ощущения. Человек слышит в диапазоне частот от
16 Гц до 20 кГц с максимальной слуховой чувствительностью в зоне
300—3000 Гц, где и расположено основное речевое поле. При этом
звуки с частотой 16 Гц и интенсивностью 70 дБ; с частотой 300 Гц
и интенсивностью 40 дБ; с частотой 20 кГц и интенсивностью 60 дБ
будут обладать равным порогом слышимости.
Определение остроты слуха основано на использовании результа­
тов исследования остроты слуха с помощью камертонов, что позво­
ляет оценить как качественное состояние слуха, т.е. состояние вос­
приятия высоких и низких тонов, так и количественное, т.е. степень
нарушения восприятия как тех, так и других тонов.
V = \00t/N,
где V — острота слуха, %; t — время восприятия испытуемым зву­
чания камертона, с; N — время звучания камертона у нормально
слышащего, с.
Звуки делят на два класса: тоны и шумы. Тоны имеют основную
частоту, определяющую их высоту, и обертоны — гармоники основных
частот, которые отличаются от основной в целое число раз. Шумы
состоят из частот, не находящихся в гармонических отношениях.
Белый шум содержит большую часть частот слышимого спектра.
Начиная с 35—40 лет у человека происходит повышение порогов слы­
шимости примерно на 80 Гц каждые полгода. Человек обладает бина­
уральным слухом и может оценить локализацию источника звука с
точностью до одного градуса. Физиологической основой бинаурально­
го слуха служит нейрональный механизм оценки временного различия
прихода звуковой волны в левое и правое ухо. В связи с тем, что звук
распространяется с конечной скоростью, он достигает более удаленно­
го уха несколько позже. При этом минимальное различие во времени
для левого и правого уха составляет 30 мкс, а разница в интенсивности
звука, которую способен выявить анализатор, составляет 1дБ. В отде­
льных нейронах верхней оливы возникает максимальное возбуждение,
если звук к одному уху приходит раньше и с большей силой, чем к
другому. Уже на уровне латеральной петли моста имеются волокна,
Глава 6
568
несущие сигналы от обоих ушей. В нижних бугорках четверохолмия
имеются два типа клеток, которые максимально возбуждаются, когда
акустические сигналы, приходящие к обоим ушам, различаются соот­
ветственно либо по времени, либо по интенсивности.
6.4. ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярный анализатор наряду, со зрительным и кинестетичес­
ким анализаторами, играет ведущую роль в пространственной ориен­
тации человека. Вестибулярный анализатор оценивает информацию
об ускорениях или о замедлениях, возникающих в процессе прямо­
линейного или вращательного движения тела, а также при изменении
положения головы в пространстве. Импульсы от вестибулорецепторов
вызывают перераспределение тонуса скелетной мускулатуры. Среди
вестибулярных реакций на первом месте находятся статические и
статокинетические реакции, обеспечивающие сохранение равновесия
при изменении положения тела и его частей или при возникающих
ускорениях во время перемещения тела в пространстве. В осуществле­
нии этих реакций участвуют также и проприорецепторы мышц.
Вестибулярный анализатор представляет собой совокупность
органа чувства с рецепторами, вестибулярного нерва с проводящими
путями и участка задней постцентральной извилины коры, которые
обеспечивают анализ вестибулярных раздражителей и синтез на его
основе ощущения равновесия и положения тела в пространстве.
Периферический отдел анализатора
Периферическим отделом анализатора являются механорецепто­
ры, расположенные в перепончатом лабиринте. Вестибулярный орган
чувств представляет собой одну из частей перепончатого лабиринта,
который заполнен эндолимфой и погружен в другую жидкость —
перилимфу. Вестибулярный орган состоит из статолитового аппарата
и полукружных канальцев (рис. 6.14). Вестибулорецепторы расположе­
ны в двух статолитовых органах — утрикулусе (маточке) и саккулусе
Рис. 6.14. Схема строения лабиринта, расположения вестибулярных рецеп­
торов и их работы: А — лабиринт; 1 — купулы; 2 — макула утрикулюса;
3 — макула саккулуса; Б — макула; В — купула: 4 — клетки рецепторные;
5 — отолиты; 6 — желатинозная масса
569
Анализаторы
эндолимфа
торможение
В
вращение
возбуждение
5 70
Глава 6
(мешочке), а также в макулах трех полукружных каналов — горизон­
тальном, переднем вертикальном и заднем вертикальном. Внутри
утрикулуса и саккулуса находится сенсорный эпителий в виде пятен
(макул) с рецепторными волосковыми клетками. Каждая рецеп­
торная клетка имеет на своей поверхности, обращенной в просвет
лабиринта, ряд волосков — цилий, среди которых выделяется самый
длинный — киноцилия. Цилии погружены в желатинообразную отолитовую мембрану с кристаллами карбоната кальция — отолитами.
При наклоне головы под действием силы тяжести возникает смеще­
ние отолитовой мембраны, что приводит к возбуждению окончаний
вестибулярных афферентных волокон. Рецепторы вестибулярного
аппарата имеют постоянную фоновую активность. Смещение цилий
в разные стороны приводит либо к усилению, либо к уменьшению
разрядов в афферентном волокне (рис. 6.15). Адекватными раздра­
жителями для рецепторов саккулуса и утрикулуса являются сила
тяжести и линейные ускорения, которые возникают при прямоли­
нейном движении. Рецепторные клетки ампул полукружных каналов
находятся под желатинообразным образованием — купулой. При
’
1 I II I HHHHIIIIIIHi ni l l l l------ I I I
M i ll
Рис. 6.15. Зависимость формы импульсной активности волосковой клет­
ки вестибулярного рецептора от направления смещения стереоцилий:
1 — положение стереоцилий; 2 — рецепторный потенциал; 3 — разряд
афферентного волокна. При сгибании стереоцилий по направлению к
киноцилии волосковая клетка деполяризуется, и в афферентном волокне
возникают импульсы. При сгибании в сторону от киноцилии возникает
гиперполяризация волосковой клетки, и афферентное волокно не отвеча­
ет. (Kandel Е., Schwartz J.H. 1981.)
Анализаторы
571
Рис. 6.16. Смещение купул и динамика импульсной активности вести­
булярного нерва при кратковременных вращательных движениях тела
с различными скоростями: А — вращения в сагиттальной плоскости;
Б — вращения во фронтальной плоскости; В — вращения в горизонталь­
ной плоскости. Скорость V, меньше скорости У2
поворотах головы свободный конец купулы, окруженный эндолим­
фой перепончатого канала, отклоняется в противоположную сторону
(рис. 6.16). При этом смещаются цилии и происходит возбуждение
рецепторных клеток. Адекватным раздражителем для рецепторов
полукружных каналов являются движения с угловым ускорением
(вращения). Порог различения наклона головы составляет 1—2 гра­
дуса. Вестибулорецепторы не проявляют адаптации.
Проводниковый отдел анализатора
Проводниковый отдел вестибулярного анализатора включает в
себя первичные чувствительные волокна, заканчивающиеся в вес­
572
Глава 6
тибулярных ядрах Дейтерса, Швальбе, Роллера, Бехтерева, а также
проводящие пути в ЦНС. Нейроны ядер изменяют свою разрядную
деятельность при вращении тела и движениях конечностей. Из
вестибулярных ядер импульсы широким потоком распространя­
ются к мозжечку, ядрам глазодвигательного нерва, гипоталамуса,
структурам ретикулярной формации, а посредством вестибулоспинального пути — к у-мотонейронам мышц-разгибателей. Именно
эти связи лежат в основе вестибулярных рефлексов, обеспечиваю­
щих совместно с информацией от проприорецепторов поддержание
равновесия. Среди вестибулярных реакций на первом месте нахо­
дятся статические и статокинетические реакции, обеспечивающие
сохранение равновесия при изменении положения тела и его частей
или при возникающих ускорениях во время перемещения тела в
пространстве. В осуществлении этих реакций участвуют также и
проприорецепторы мышц. Ф ункциональные связи между выше­
указанны ми структурами обеспечивают не только поддержание
позы человека (сохранение равновесия), зрительного восприятия
(нистагм), но и координацию двигательных актов при выполнении
целенаправленной деятельности. Вторые афферентные нейроны от
вестибулярных ядер идут к таламусу, а от его вентрального постлатерального ядра поступают к задней постцентральной извилине
коры больших полушарий.
Корковый отдел анализатора
Корковый отдел анализатора (21—22 поля, по Бродману) имеет
нейроны, избирательно реагирующие на повороты тела в трех перпен­
дикулярных плоскостях. У человека активация данного участка коры
больших полушарий сопровождается осознанием положения тела в
пространстве, а также направления и скорости его перемещения.
Психофизика вестибулярного анализатора строится как на под­
держании равновесия, так и на обеспечении выполнения целе­
направленного движения. Поддержание равновесия и выполнение
движения осуществляются в условиях постоянно действующего
раздражителя — гравитации. При этом сохранение человеком равно­
весия возможно в том случае, если проекция центра тяжести нахо­
дится в пределах площади опоры, занимаемой стопами. Центром
тяжести называется точка, через которую проходит линия действия
силы тяжести при любом положении тела в пространстве. Само же
локальное движение выполняется с учетом равновесия тела. Чувство
равновесия создается комплексной деятельностью вестибулярно­
573
Анализаторы
го аппарата, афферентацией от проприодепторов мышц, связок,
суставов, работой тактильных рецепторов кожи и информацией,
поступающей от фоторецепторов глазных яблок. Вестибулярный
иистагм заключается в ритмических медленных движениях глаз в
сторону, противоположную движению тела, а затем — в быстрых
движениях в обратном направлении. Этот механизм обеспечивает
широкий обзор пространства при движении в нем наблюдателя.
Деятельность вестибулярного анализатора влияет не только на состо­
яние соматической мускулатуры, но через ВНС — на функции внут­
ренних органов, вызывая специфическую реакцию укачивания.
6.5. КОЖНЫЙ АНАЛИЗАТОР
Кожа — внешний покров тела человека. Как анатомо-физиологическая часть целостного организма кожа является органом, выполняющим
ряд очень важных физиологических функций: защитную, терморегу­
лирующую, секреторную, обменную и рецепторную. В коже осущест­
вляется синтез витамина Д. Площадь поверхности кожи достигает у
взрослого человека в среднем 1,6 м2. В области естественных отверстий
(рта, носа, заднего прохода, мочеиспускательного канала и влагалища)
кожа непосредственно переходит в слизистые оболочки. Почти на всем
своем протяжении кожный покров человека покрыт волосами.
Кожный анализатор представляет собой совокупность органа
чувства (кожи) с рецепторами, кожных нервов с центральными
проводящими путями и участка задней постцентральной извилины
коры, которая обеспечивает анализ раздражителей и синтез на его
основе соответствующего кожного ощущения.
Периферический отдел анализатора
Периферический отдел анализатора представляют разнообразные
рецепторы, расположенные в различных участках и слоях кожной
поверхности (табл. 6.1).
Различают следующие виды кожной чувствительности: 1) так­
тильную (чувство осязания и давления), 2) температурную (чувство
тепла и холода) и 3) болевую (температурная и болевая рецепция
рассматриваются в других главах учебника).
Тактильная рецепция включает в себя ощущения прикосновения
и давления, которые вызываются при воздействии на чувствительные
осязательные точки. Наибольшее число таких точек наблюдается
574
Глава 6
Таблица 6.1. Характеристики рецепторов кожи и проприоцепторов
Рецептор
Тельца
Пачини
Место
Кожа, сухо­
жилия,
слизистые
оболочки
Стимул
Ускорение
смещения
Ощущение
Фон
Прикосно­ Нет
вение, ви­
брация
250-300 Гц
Тельца
Мейснера
Кожа, сли­
зистые о бо­
лочки
Скорость
смещения
Прикосно­
вение, ви­
брация
3 0 -4 0 Гц
Кожа, сли­
зистые о бо­
лочки, связ­
ки, суставы
Тельца
Кожа, сли­
Меркеля
зистые о бо­
лочки
Аппарат
Волосистая
волосяного часть кожи
мешочка
Голые
Кожа, сли­
окончания зистые
оболочки,
суставы
Скорость
смещения,
положение
Прикосно­ Неболь­
вение, да­
шая
вление,
сокращение
Прикосно­
вение, дав­
ление
Тельца
Гольджи
Мышечные Мышцы
веретена
Скорость
смещения
Адаптация
Быстрая
Неболь­
шая
5 имп./с
Прикосно­ Нет
вение, виб­
рация 35 Гц
Терморецеп­ Тепло, хо­
Неболь­
ция
лод, пока­
шая
лывание,
боль
Растяжение Движение, Высокая
положение
Медленная
Быстрая
Медленная
на кончиках пальцев и губах. Ощущение легкого прикоснове­
ния возникает при механическом раздражении особых нервных
сплетений основания волоса — волосяных фолликулов. Повреждение
эпидермиса сопровождается нарушениями ощущения. Абсолютная
тактильная чувствительность колеблется от 50 мг до 10 г в различ­
ных частях кожи и появляется при вдавлении кожи на ладони на
0,01 мм. Пространственные пороги одновременного пространствен­
ного различения сильно отличаются в различных участках: от 0,5 мм
на слизистой языка до 65 мм на коже спины и связаны с размерами
рецептивных полей — от 0,5 мм2 до 3 см2 соответственно. Кожные
рецепторы реагируют также на изменение скорости и ускорения
действия тактильного раздражителя. Тактильные рецепторы в фун­
кциональном плане бывают тоническими и фазическими. Первые из
575
Анализаторы
них реагируют на постоянную деформацию, а вторые — на кратков­
ременные небольшие смещения кожи. Форма кожных рецептивных
нолей чаще всего округлая, на дистантных участках тела — эллип­
совидная. Рецептивные поля могут перекрываться между собой, что
шачительно увеличивает пространственные и дифференциальные
мороги тактильного различения.
Проводниковый отдел
Проводниковый отдел кожного анализатора состоит из перифе­
рических нервов, содержащих волокна А и С, афферентных путей и
переключающих ядер. Поступающие в спинной мозг импульсы от
рецепторов кожи распространяются двумя основными путями: лемп исковым и спиноталамическим. Лемнисковый путь представлен
нейронами с высокой скоростью проведения возбуждения, неболь­
шими рецептивными полями, коротким латентным периодом и
выраженной сенсорной специфичностью. Тела вторых нейронов рас­
положены в ядрах тонкого и клиновидного пучка, а после перекреста
на уровне олив направляются к специфическим ядрам таламуса. По
мере продвижения наверх размеры рецептивных полей нейронов
увеличиваются в десятки раз, а их ответы на раздражитель при­
обретают большую продолжительность. Спиноталамический путь
состоит из нейронов с меньшей скоростью проведения возбужде­
ния, достаточно широкими рецептивными полями и длительными
реакциями на раздражитель. Тела вторых нейронов находятся в
сером веществе и посылают свои аксоны в неспецифические ядра
таламуса. В ядрах вентро-базального таламуса переключается сиг­
нализация о прикосновении, давлении и движениях в суставах.
На нейронах интраламинарных ядер, задних и ретикулярных ядрах
таламуса наблюдается обширная мультисенсорная конвергенция
возбуждений различной сенсорной модальности, включая болевую.
Большинство нейронов вентро-базального ядра таламуса возбужда­
ется при механической стимуляции только их рецептивного поля;
каждый нейрон возбуждается только одним типом рецепторов —
например, медленно адаптирующимися рецепторами давления или
рецепторами волосяных луковиц. Интенсивность периферического
стимула в нейронах таламуса кодируется, как и в рецепторах, сред­
ней частотой разрядов нейронов.
В случаях болевого раздражения лемнисковая система передает
информацию о сенсорных качествах раздражителя, а спиноталами­
ческая — о характере эмоционального возбуждения.
576
Глава 6
Корковый отдел анализатора
Корковый отдел анализатора расположен в задней постцентральной извилине, где в соматосенсорных зонах SI и SII заканчиваются
таламо-кортикальные пути противоположной половины спинного
мозга. Зона SI имеет выраженную соматотопическую организацию,
при которой те или иные участки поверхности тела представлены
в коре пропорционально количеству расположенных там кожных
рецепторов (рис. 6.17 и 3.23, А). Пространственное представительство
тела в зоне SII билатерально. Функциональной единицей сомато­
сенсорной коры являются колонки, каждая из которых содержит
до 100 ООО нейронов. Нейроны одной колонки часто возбуждаются
рецепторами одного типа. В колонках обнаружены нейроны двух
Тактильные раздражения
Вызванные потенциалы
на поверхности
Рис. 6.17. Формирование соматосенсорного ощущения при действии раздра­
жителя на участки кожной поверхности кошки. Локализация рецептивных
полей на теле кошки, проекции участков тела на заднюю постцентральную
извилину мозга и вызванный потенциал в зоне соматосенсорного анализа­
тора. Размер проекции участка тела в соматосенсорной коре пропорциона­
лен силе раздражителя и его месту в формировании сенсорного образа
Анализаторы
577
иидов — простые нейроны, реакции которых аналогичны реакциям
соответствующих рецепторов, и сложные нейроны, у которых такая
закономерность не прослеживается. К сложным нейронам относятся
клетки, обладающие температурной чувствительностью. Первичные
и вторичные сенсомоторные области в симметричных отделах коры
мозга связаны между собой через мозолистые тела. В этих областях
выявлено соматотопическое различие нейронов. Так, нейроны поля
За, по Бродману, получают информацию от мышечных и суставных
рецепторов, нейроны поля 2 возбуждаются при сдавливании кожи и
практически не реагируют на легкое прикосновение. Нейроны поля
36 реагируют на легкое прикосновение к коже и сгибание отдельных
иолосков. Нейроны поля 1 реагируют как на легкое раздражение
кожи, так и на ее сдавливание. Тактильные и проприоцептивные
импульсы регулируются нисходящими кортикобульбарными влия­
ниями, которые начинаются в зоне SI и в двигательной коре.
Психофизика кожного ощущения у человека достаточно разно­
образна: осязание, восприятие тепла, холода, боли, зуда. Роль кож­
ного ощущения резко возрастает при нарушениях зрения и слуха.
У людей при этом развивается способность читать рельефный текст
и хорошо распознавать форму предметов. Значения температур в
различных участках поверхности составляют температурную схему
юла. Температурная схема тела очень индивидуальна и зависит от
генетических факторов, индивидуального воспитания, привычек,
условий жизни, профессии, экологии, закалки человека.
6.6. ОБОНЯТЕЛЬНЫЙ АНАЛИЗАТОР
Запахи проникают в эмоциональную жизнь человека, затрагивая
его сознание, привлекают, возбуждают или пугают его. Запахи могут
н известной степени управлять настроением и работоспособностью
пичности. Считается, что обонятельный анализатор является древисйшим в животном мире. Обонятельный и вкусовой анализаторы
функционируют сочетанно. Однако если для вкусовых рецепторов
раздражители являются полезными или вредными, то для обоня­
тельных — чаще выполняют сигнальное значение.
Обонятельный анализатор представляет собой совокупность органа
чувства — носа — с рецепторами, чувствительного нерва с обонятельным
трактом и участка коры, которые обеспечивают анализ раздражителя и
синтез на его основе ощущения или сложного обонятельного образа.
Глава 6
5 78
Периферический отдел анализатора
П ериф ерическим отделом анализатора являю тся рецепторы
обонятельной области, расположенной в верхней носовой ракови­
не. Рецепторы представляют собой погруженные в слизь 6—12 рес­
ничек, отходящих от дендритов биполярных клеток, на которые
воздействуют пахучие вещества. О бонятельная область человека
площадью 10 см2 содержит 10 млн рецепторов, которые постоян­
но замещаются. Рецептор обладает низкой фоновой импульсной
активностью и может реагировать при взаимодействии с несколь­
ким и пахучими веществами различны х классов. При этом отде­
льная рецепторная клетка возбуждается в ответ на единственную
молекулу вещества. Кодирование качества раздраж ителя обес­
печивается ответами многих рецепторов, а его интенсивности —
числом импульсов на выходе рецептора. Кроме того, слизистая
оболочка носа имеет свободные окончания тройничного, а слизис­
тая оболочка глотки — языкоглоточного и блуждающего нервов.
Следует отметить, что первичных обонятельных ощущений боль­
ше, чем вкусовых.
Проводниковый отдел анализатора
Проводниковая часть анализатора состоит из аксонов сенсор­
ных биполярных клеток, направляющихся в составе обонятельного
нерва в обонятельную луковицу. Каждое пахучее вещество вызыва­
ет в афферентных волокнах специфическое для данного вещества
пространственно-временное распределение импульсаций (рис. 6.18).
Обонятельная луковица — первичный центральный отдел анализа­
тора, она содержит нейроны второго порядка — митральные клет­
ки, на которых конвергируют импульсы от 100 волокон. На уровне
клеточных элементов обонятельной луковицы функционирует меха­
низм латерального торможения. Адаптация рецепторов обонятель­
ного анализатора наступает очень медленно.
Корковый отдел анализатора
Корковый отдел анализатора находится в префронтальной и орби­
тофронтальной коре, куда направляются аксоны митральных клеток,
составляющие латеральный обонятельный тракт. Латеральный тракт
также имеет связи с гипоталамусом, ретикулярной формацией, мин­
далиной и другими частями лимбической системы.
Психофизика восприятия запахов человеком имеет ту особенность,
что при действии очень малых концентраций веществ ощущение
Анализаторы
5 79
Иннервации
Зона иннервации
тройничного нерва
I
I
|
Обонятельная
луковица
Зона иннервации
языко-глоточного
нерва
Зона иннервации
блуждающего нерва
Рис. 6.18. А — схема воздухоносных путей с зонами иннервации языкогло­
точным, тройничным и блуждающим нервами; Б — динамика паттернов
реакций хеморецепторов на различные обонятельные раздражители
будет неспецифичным, но при повышении концентрации появляет­
ся возможность идентифицировать то или иное вещество. По этой
причине говорят о пороге выявления и пороге распознавания запаха.
Выделяют несколько основных запахов: камфарный, цветочный,
мускусный, мятный, эфирный, едкий, гнилостный. Ш ирокие связи
обонятельных путей с лимбической системой указывают на роль
>моционального возбуждения в восприятии запахов, что хорошо
известно на примере полового поведения.
582
Глава 6
Психофизика восприятия вкуса человеком состоит в том, что вкус
является полимодальным ощущением, куда входят температурные,
тактильные и обонятельные компоненты. Кончик языка чувствите­
лен к сладкому, средняя часть — к кислому, корень — к горькому,
край — к соленому и кислому. Адаптация к различным вкусовым
свойствам развивается в течение разного времени. Значение вкуса
заключается, во-первых, в оценке пригодности пищи; во-вторых, в
формировании слюноотделения в ротовой полости и сокоотделения
в желудке; в-третьих, в возникновении эмоционального возбужде­
ния, сопровождающего процесс пищеварения в ротовой полости.
Величина абсолютных порогов вкусовых ощущений может менять­
ся при различных биологических состояниях человека — голодание,
беременность, недостаток отдельных веществ в организме. Между
химическими свойствами веществ и их вкусом не выявлено какойлибо достоверной корреляции. Так, если фактором, определяющим
кислый вкус, служат ионы Н+, то остальные вкусовые ощущения
очень трудно связать с физическими или химическими свойс­
твами вещества. Вещества со сходным вкусом могут значительно
различаться по химической структуре, и наоборот. Например, не
только сахара, но и соли свинца имеют сладкий вкус, а некоторые
аминокислоты обладают в D -форме сладким вкусом, в L-форме —
горьким. Очевидно, в естественных условиях большинство вкусо­
вых ощущений — смешанные и определяются набором химических
раздражителей. Кроме того, восприятия вкуса широко варьируют в
зависимости от исходного мотивационного состояния организма.
Следует отметить, что вкус любого вещества можно имитировать,
смешав два или три вещества с различными вкусовыми характе­
ристиками.
Глава 7
ДВИГАТЕЛЬНЬЫЕ ФУНКЦИИ
7.1. ОРГАНИЗАЦИЯ ДВИХЖЕНИЙ
Биологическая роль движения
Поведение организмов за счет взаимодеййствия с окружающим
миром обеспечивает удовлетворение их ве;едущих потребностей.
В процессе эволюции двигательные реакции i существенно прогрес­
сировали: от движения протоплазматическиих ложноножек, плав­
ников, крыльев до сложных движений млекоюпитающих и человека,
включающих локомоцию, манипулирование, ь мимику, жестикулиро­
вание, речь, письмо и др.
С позиций теории функциональных систеюм движение выступает
как внешнее звено саморегуляции в ф у н к ц и о н а л ь н ы х системах пита­
ния, поддержания осмотического давления и тегерморегуляции. Вместе
с тем, двигательный акт имеет системную о р га н и за ц и ю , включаясь
в формирование общей архитектоники целекнаправленной деятель­
ности на стадии эфферентного синтеза, когда,а будущее действие уже
сформировано как центральный процесс, но о еще не реализуется в
работе мышц.
Эфферентный синтез
Стадия эфферентного синтеза формируетсгся под влиянием про­
цессов афферентного синтеза, принятия регешения и предвидения
полезного результата — стадии акцептора р езу л ьтато в действия. На
стадии эфферентного синтеза решаются задгцачи создания адекват­
ной программы действия и разработки спосоюбов ее реализации на
уровне исполнительных механизмов каждой i функциональной сис­
темы. Важнейшим компонентом исполнителыьных программ служат
разнообразные движения — установка позыы, перемещение тела в
пространстве, удержание частей тела в ф и к си р о ван н о м положении,
манипуляции. Сами движения сопровождаюются вегетативными и
эндокринными реакциями.
584
Глава 7
С физиологической точки зрения, эфферентный синтез есть вре­
менная организация в ЦНС процессов возбуждения и торможения,
адресующихся к мышцам, железам и другим тканям. Эфферентный
синтез заканчивается формированием общего эффекторного интег­
рала, который включает в себя соматические, вегетативные и эндок­
ринные компоненты целостной поведенческой деятельности.
Пластичность эф ф екторкы х функций
Различные физиологические функции — дыхание, кровообраще­
ние, пищеварение, выделение и другие — могут вовлекаться в качес­
тве исполнительных механизмов в разные функциональные системы.
Например, деятельность сердца поддерживает и должный уровень
кровяного давления, и постоянство газового состава крови, и уровень
осмотического давления. Выбор исполнительных механизмов конк­
ретной функциональной системы осуществляется при участии меха­
низмов памяти — опыта использования различных физиологических
функций в аналогичной ситуации в прошлом. Извлечение из памяти
нужной информации происходит при участии доминирующей моти­
вации и факторов обстановки. Процессы эфферентного синтеза
постоянно динамически перестраиваются под контролем обратной
афферентации от результатов поведенческой деятельности.
Мышечное чувство
Еще И.М. Сеченов указывал на то, что движение всегда имеет
единство «чувствования и действия». Мышечное чувство возникает
на основе импульсов, поступающих в ЦНС от рецепторов опорно­
двигательного аппарата. Эта импульсация дает возможность ЦНС
корригировать процесс выполнения движения. Импульсация от
мышц входит в состав обратной афферентации о параметрах этапных
и конечного результатов действия. Каждая динамическая программа
эфферентного синтеза оценивается аппаратом акцептора результатов
действия.
Общая характеристика мышечной активности
Мышечная активность обеспечивает позу, локомоцию — ходьбу,
бег, плавание; коммуникацию — письмо, речь, жестикуляцию, мими­
ку; манипулирование — предметную деятельность. Двигательные
функции реализуются тремя основными процессами: поддержанием
мышечного тонуса, формированием адекватной позы и выполнением
конкретного произвольного движения.
Двигательные функции
5 85
Мышечный тонус как основа движения
Мышечный тонус — состояние продолжительного поддержания
определенного уровня активности мышцы. Тонус мышцы определя­
ется активностью отдельных мышечных волокон при поступлении
к ним импульсов по двигательным волокнам от а-мотонейронов
спинного мозга.
Моторная единица
Каждый отдельный а-мотонейрон спинного мозга иннервирует
строго определенное число мышечных волокон. Структура «мото­
нейрон—мышечное волокно» получила название моторной единицы.
Количество мышечных волокон, объединенных в моторные едини­
цы, бывает различным (рис. 7.1). В наружных мышцах глаза один
мотонейрон иннервирует 3—4 мышечных волокна (иннервационный
индекс 1:3), а в широкой мышце спины — до 300 мышечных волокон
(иннервационный индекс 1:300). Чем точнее движения, совершаемые
мышцей, тем больше в ее составе моторных единиц. Моторные еди­
ницы бывают трех типов: S — медленные, устойчивые к утомлению и
состоящие из мышечных волокон I типа; FF — быстрые, утомляемые,
состоящие из мышечных волокон ПБ типа; FR — быстрые, устойчи­
вые к утомлению, состоящие из волокон IIA типа.
Мышечная единица
Совокупность мышечных волокон одной моторной единицы назы­
вается мышечной единицей. Волокна каждой мышечной единицы
относятся к одному и тому же гистохимическому типу: I, НБ, IIA.
Мотонейронный пул
Общее число мотонейронов, иннервирующих одну мышцу, состав­
ляет нейронный пул. Поддержание мышечного тонуса достигается
преимущественной активностью так называемых красных мышеч­
ных волокон, содержащих большое количество миоглобина и име­
ющих низкую скорость распространения возбуждения и развития
сокращения. У человека таких волокон больше среди мышц-разгибателей, поддерживающих вертикальное положение тела и работающих
против гравитационных сил.
В формировании мышечного тонуса принимают участие спе­
циальные рецепторы: мышечного веретена, сухожильные органы,
а также рецепторы, расположенные в суставной оболочке.
Глава 6
5 80
6.7. ВКУСОВОЙ АНАЛИЗАТОР
У человека вкус служит для апробирования, дегустации пищи,
определения качества продуктов питания, поддержания собствен­
ного гомеостазиса. Вкусовые рецепторы — своеобразные «ворота»
пищеварительной системы, определяющие реакции предпочтения и
отвергания.
Вкусовой анализатор представляет собой совокупность органа чувс­
тва — языка — с рецепторами, чувствительного нерва с промежуточны­
ми ядрами и участка коры, которые обеспечивают анализ раздражителя
и синтез на его основе ощущения или сложного вкусового образа.
Периферический отдел анализатора
Периферическим отделом анализатора являются вкусовые рецеп­
торы области, образующие вместе с базальными и опорными клет­
ками вкусовую почку, число которых у человека достигает 10 тысяч.
Каждая почка содержит около 50 рецепторных клеток. Вкусовые
почки располагаются в грибовидных, желобовидных и листовидных
сосочках. Грибовидные сосочки равномерно распределены по всей
поверхности языка, желобовидные — у корня, а листовидные — вдоль
задних краев языка. Количество функционирующих сосочков зави­
сит от состояния организма, а также от деятельности пищеваритель­
ных органов. Ответная реакция вкусового рецептора состоит в пачке
импульсов, которые возникают в первые 50 мс от начала раздражения
с последующим снижением активности до уровня исходного фона.
Одна рецепторная клетка, как правило, реагирует на вещества разного
химического качества, однако наиболее активная реакция возникает
только на раздражение одного химического качества. Конфигурация
импульсов в волокне несет информацию о химическом свойстве
вещества, а совокупность возбужденных волокон — о его концентра­
ции. При длительном действии вкусового раздражителя в рецепторах
возникает явление адаптации. Время жизни чувствительных клеток
составляет 10 сут, после чего образуются новые клетки.
Проводниковый отдел анализатора
Проводниковый отдел анализатора начинается от вкусовых почек
в виде 50 чувствительных волокон, которые вместе с тактильными,
температурными и болевыми волокнами языка входят в состав язы­
коглоточного и лицевого нервов (рис. 6.19). Тела клеток, принадле­
жащих вкусовым волокнам VII, IX и X черепных нервов, находятся
Анализаторы
581
Рис. 6.19. Схема языка человека с зонами иннервации. Динамика импуль­
сной активности чувствительных нервов, несущих информацию от рецеп­
торов языка: А — импульсная активность языкоглоточного нерва в ответ
на предъявление горького раздражителя; Б — импульсная активность
лицевого нерва в ответ на предъявление кислого, соленого и сладкого раз­
дражителей
в каменистом, коленчатом и узловатом ганглиях. Вкусовые волокна
входят в продолговатый мозг и в ядре солитарного тракта образуют
синапсы со вторыми нейронами. Отростки вторых нейронов после
частичного перекреста поступают в вентральные ядра таламуса.
Корковый отдел анализатора
Корковый отдел анализатора находится в постцентральной изви­
лине головного мозга в области проекции языка, куда поступают
импульсы от таламических ядер. Исключительная особенность вку­
сового пути заключается в том, что его волокна не перекрещиваются
с волокнами, идущими с противоположной стороны. При этом неко­
торые нейроны реагируют как на химическое, так и на тактильное и
температурное воздействие вещества.
Двигательные функции
5 87
Мышечные веретена
Мышечные веретена состоят из соединительнотканной капсулы
с заключенными в ней интрафузальными мышечными волокнами,
к которым и подходят отростки у-мотонейронов (рис. 7.2). Длина
мышечного веретена равна в среднем 10 мм, а диаметр — около
100 мкм. В веретенах имеются чувствительные окончания двух типов:
афференты группы 1а со скоростью проведения до 120 м/с и группы II
со скоростью до 70 м/с. Эти чувствительные окончания относятся
к рецепторам растяжения. Общее количество мышечных веретен в
мышце тем больше, чем тоньше и точнее осуществляемые движе­
ния. Мышечные веретена крепятся одним своим концом к мышеч­
ному сухожилию, а другим — к собственным фасциям мышцы,
т.е. параллельно рабочим (экстрафузальным) мышечным волокнам.
Двигательную функцию мышечных веретен обеспечивают также два
вида нервных волокон: статические у-эфференты, оканчивающиеся
на каждом волокне с ядерной сумкой и возбуждающиеся при стати­
ческих нагрузках, и динамические у-эфференты, оканчивающиеся
на волокнах с ядерной цепочкой, которые возбуждаются при дина­
мических нагрузках. При активации динамического у-мотонейрона
вторично возрастают динимические ответы афферентов группы 1а, а
при активации статического у-мотонейрона возрастают статические
ответы обеих групп — 1а и II.
Происхождение мышечного тонуса
Мышечный тонус поддерживается на основе принципа саморе­
гуляции: изменение мышечного тонуса вызывает процессы, направ-
Рис. 7.1. Типы моторных единиц и динамика их активности: А —моторная
единица ME-I имеет в своем составе небольшой низкочастотный нейрон
с тонким аксоном, низкой скоростью проведения возбуждения, с низкой
утомляемостью и с небольшим количеством иннервируемых мышечных
волокон (S-тип). Мышечные волокна такой моторной единицы развивают
малые мышечные усилия с низкой скоростью сокращения; В — моторная
единицаМЕ-ПБ состоит из крупных высокочастотных нейронов с толстым
аксоном, высокой скоростью проведения возбуждения, с низкой утомля­
емостью и с большим количеством иннервируемых волокон. Мышечные
волокна такой моторной единицы развивают большие мышечные усилия с
высокой скоростью сокращения (FF-тип); Б — моторная единица ME-IIA
занимает промежуточное положение (FR-тип)
5 88
Глава 7
Рис. 7.2. Взаимоотношение рецепторов мышцы с нейронами сегмента
спинного мозга
ленные на его восстановление. Основной механизм саморегуляции —
сегментарный аппарат спинного мозга. Тоническая активность ней­
ронов спинного мозга определяется как импульсацией, поступающей
к ним от кожных рецепторов и проприорецепторов мышц, так и нис­
ходящими центральными влияниями. Перерезка задних корешков
спинного мозга у спинальной лягуш ки на одной стороне приводит
к полному расслаблению мышц соответствующей лапки и ее прови­
санию (опыт Бронджеста).
При устранении нисходящих влияний (травматический разрыв
спинного мозга или его местная анестезия) также наблюдается выра­
женное изменение тонуса мышц — вялость или ригидность.
В сегментарном аппарате спинного мозга наряду с а-мотонейронами,
возбуждение которых вызывает сокращение экстрафузальных мышеч­
Двигательные функции
589
ных волокон, имеется группа у-мотонейронов, посылающая импульсы
к мышечным веретенам — собственным рецепторам мышцы.
При пассивном растяжении мышцы возбуждения от чувстви­
тельных окончаний у-афферентных волокон мышечного веретена
поступают через задние корешки в спинной мозг и переключаются
на а-мотонейроны передних рогов спинного мозга того же сегмен­
та. Это приводит к повышению возбудимости а-мотонейронов и
последующему усилению тонуса мышцы (рис. 7.3). При сокраще­
нии мышцы чувствительные окончания веретена перестают воз­
буждаться, вследствие чего прекращается поток импульсов, иду­
щих к а-мотонейронам, уровень их возбудимости падает, и тонус
мышцы снижается. С помощью этого механизма мышечные вере­
тена воспринимают длину мышцы и скорость ее изменения. При
большей скорости изменения длины мышечных волокон частота
импульсации в у-афферентных волокнах выше. Кроме мышечных
веретен, в саморегуляции мышечного тонуса принимают участие
рецепторы Гольджи, расположенные в сухожилиях мышц. Рецептор
имеет длину около 1 мм и диаметр до 100 мкм. От рецептора отходит
афферентрое волокно группы lb со скоростью проведения до 120 м/с.
Сухожильные рецепторы Гольджи реагируют на сокращение и на
растяжение мышцы, однако наибольшая реакция у них возникает
при мышечном сокращении. Число рецепторных структур в сухо­
жилиях мышц невелико. Рецепторы сухожилий последовательно
соединяются с экстрафузальными волокнами мышцы. Возбуждение
сухожильных рецепторов происходит как при пассивном растяже­
нии, так и при активном сокращении мышц. Возбуждение, идущее
от сухожильных рецепторов по у-афферентным волокнам, поступа­
ет к а-мотонейронам соответствующих сегментов спинного мозга,
которые иннервируют эти мышцы и их синергисты и вызывают (при
участии тормозных клеток Реншоу) их торможение. В то же время
часть импульсов от сухожильных рецепторов распространяется и к
а-мотонейронам мышц-антагонистов.
Рецепторы суставов
К рецепторам, участвующим в поддержании мышечного тонуса,
относятся рецепторы суставов, которые возбуждаются при растяже­
нии суставных сумок. Уровень активации этих рецепторов соответс­
твует углу поворота конечности в суставе. Диапазон углов смещения,
при котором возникает возбуждение суставных рецепторов, колеб­
лется от 2—3 до 30°.
591
Двигательные функции
Саморегуляция мышечного тонуса
Рассмотренные группы рецепторов участвуют в координации спи­
нальных непроизвольных механизмов саморегуляции длины мышц,
степени их напряжения и положения различных отделов конечностей
и туловища. Если наблюдается, например, хотя бы незначительное
пассивное сгибание ноги в коленном суставе, то происходит растя­
жение четырехглавой мышцы бедра с последующей активацией ее
мышечных веретен. Импульсы от веретен четырехглавой мышцы по
у-афферентным волокнам поступают к а-мотонейронам передних
рогов спинного мозга, активация которых приводит к сокращению
этой мышцы и прекращению сгибания ноги в коленном суставе.
При сильном разгибании ноги в коленном суставе возникают растяже­
ние мышцы-сгибателя и активация ее мышечных веретен. Импульсы от
веретен поступают к а-мотонейронам, иннервирующим мышцы-сгибатели, сокращение которых приводит к сгибанию ноги в коленном суставе.
Координация процессов саморегуляции мышечного тонуса на уровне
сегментов спинного мозга достигается возвратным торможением разряд­
ной деятельности мотонейронов при участии тормозных клеток Реншоу.
Активация самих клеток Реншоу происходит под влиянием импульсаций, поступающих к ним от у-афферентов мышечных веретен.
Поза
Поза — длительное, близкое к изометрическому, сокращение
групп мышц, которое обеспечивает оптимальное положение тулови-
Рис. 7.3. Работа сегментарного аппарата спинного мозга при различных состо­
яниях скелетной мышцы: А — состояние покоя; Б — пассивное растяжение
мышцы с активацией у-афферентных волокон; В — активация а-мотонейро­
нов и проведение возбуждений к мышце; Г — сокращение мышцы с активаци­
ей рецепторов сухожильного органа Гольджи; Д — возбуждение импульсами
от сухожильных рецепторов Гольджи вставочных тормозных нейронов и тор­
можение ими мотонейронов; Е —нисходящая команда к сокращению мышцы
по экстрапирамидной системе достигает у-мотонейрона, импульсы от кото­
рого приводят к сокращению интрафузальные волокна мышечного веретена;
Ж — возбуждения от рецепторов мышечного веретена достигают а-мотоней­
ронов спинного мозга. Возбуждения а-мотонейронов вызывают сокращение
мышцы, что сопровождается активацией рецепторов сухожильного органа
Гольджи. MB — мышечное веретено; СОГ — сухожильный орган Гольджи;
М — мышца
592
Глава 7
ща и конечностей в покое и при движении. В формировании позы
участвуют нейроны различных уровней ЦНС. Это, прежде всего,
структуры экстрапирамидной системы — мозжечок, базальные ядра,
средний мозг. Нейроны указанных структур получают импульсы
из ассоциативной коры и посылают их к у-мотонейронам передних
рогов спинного мозга. Импульсы от у-мотонейронов распространя­
ются к интрафузальным волокнам мышечных веретен, что приводит
к сокращению волокон и возбуждению чувствительных элементов
мышечных веретен. Импульсы от веретен поступают по у-афферентам к а-мотонейронам соответствующего сегмента, возбуждение
которых и вызывает сокращение экстрафузальных волокон мышцы.
Участие головного мозга в формировании позы
Участие структур головного мозга в формировании позы было пока­
зано в опытах с перерезками мозга на разных уровнях (Р. Магнус).
Средний мозг. Структуры среднего мозга участвуют в регуляции
позы путем формирования тонических реакций. Тонические мышеч­
ные реакции подразделяются на несколько видов.
Статические реакции включают два варианта. Во-первых, позные
реакции, возникающие при изменениях положения головы. Они
определяются импульсацией, поступающей к структурам среднего
мозга от рецепторов лабиринта, а также от проприорецепторов мышц
шеи и туловища. Во-вторых, выпрямительные реакции, направлен­
ные на восстановление позы в случаях ее нарушения. Они выра­
жаются выпрямлением головы и туловища. Выпрямление головы
определяется импульсацией, поступающей от рецепторов сетчатки
и мышц глаза, лабиринтов, кожи туловища. Выпрямление туловища
определяется импульсацией, поступающей от проприорецепторов
шеи и рецепторов кожи туловища.
Статокинетические реакции включают реакции на вращение тела,
определяемые импульсацией, поступающей от рецепторов полукруж­
ных каналов, и реакции на прямолинейное движение, определяемые
импульсацией, поступающей от рецепторов отолитового органа.
Децеребрационная ригидность. Указанные реакции отчетливо про­
являются у децеребрированных животных. Операция децеребрации
осуществляется путем перерезки ствола мозга между передними и
задними буграми четверохолмия. В результате перерезки наблюдает­
ся повышение тонуса мышц-разгибателей. Ж ивотных можно поста­
вить на ноги, но они не способны удерживать позу стояния и падают
при небольшом нарушении равновесия.
Двигательные функции
5 93
Состояние децеребрационной ригидности создается структурами
среднего мозга, особенно красными ядрами. Перерезки ниже красных
ядер устраняют децеребрационную ригидность. Пассивный подъем
головы у децеребрированной кошки с разрушенными лабиринтами
приводит к снижению тонуса разгибателей передних конечностей.
Пассивный наклон головы вызывает обратный эффект (рис. 7.4).
При отклонениях головы у децеребрированного животного направо
усиливается сокращение мышц-разгибателей правых конечностей
и наоборот. У децеребрированных животных двигательные позные
реакции определяются импульсами, возникающими в проприоцепторах шейных мышц.
У децеребрированных животных проявляются реакции восста­
новления исходного положения — выпрямление туловища. Если
децеребрированное животное с разрушенными лабиринтами держать
в воздухе, то оно не держит голову. Если же его положить на стол,
то голова животного занимает привычное положение. Это связано с
тем, что при соприкосновении головы с поверхностью стола раздра­
жаются кожные тактильные рецепторы соответствующей стороны
головы. Импульсы от этих рецепторов поступают в ЦНС и вызыва­
ют сокращение шейных мышц животного, вследствие чего голова
занимает устойчивое положение. У децеребрированных животных
наблюдается еще одна форма регуляции позы. Это так называемые
лифтные феномены, которые проявляются увеличением сокращения
мышц-разгибателей в случаях ускоренного перемещения всего тела
вниз и мышц-сгибателей при ускоренном движении вверх. Сюда же
входят компенсаторные движения глазных яблок и головы с целью
удержания зрительного изображения на сетчатке глаз при переме­
щении тела в пространстве.
Мозжечок получает импульсы от вестибулорецепторов, проприоцепторов мышц и связок и от коры большого мозга. Эфферентная
импульсация из мозжечка поступает в различные отделы головного
мозга. В системных механизмах поддержания позы и мышечного
тонуса ведущее место отводится червю мозжечка, получающему
информацию от мышечных проприорецепторов и кожных рецеп­
торов. Нейроны червя мозжечка посылают импульсацию к вести­
булярному ядру Дейтерса и ретикулярной формации ствола мозга.
Обратная афферентация о позе постоянно поступает в промежуточ­
ную часть мозжечка (пробковидные и шаровидные ядра, ядра шатра)
и сравнивается с запрограммированной нейронами коры большого
мозга моделью ожидаемого положения тела и его частей.
Двигательные функции
5 95
При поражениях коры мозжечка возникает общее снижение
мышечного тонуса в сочетании с повышенной утомляемостью.
Изолированное повреждение червя мозжечка сопровождается в
большинстве случаев увеличением мышечного тонуса (гипертонус).
Поражения мозжечка приводят к астазии — потере мышцами способ­
ности к слитному тоническому сокращению, в результате чего голова,
туловище и конечности дрожат и качаются.
Кроме того, в мозжечке выделяют три корково-ядерных зоны:
медиальную, промежуточную и латеральную (рис. 7.5). В частности,
промежуточная (околочервячная) зона проецируется на пробковид­
ные и шаровидные ядра, а через них на красное ядро, которое посы­
лает импульсы по руброспинальному тракту к нейронам спинного
мозга. Кроме того, промежуточная зона получает копию сигналов
от пирамидного тракта о готовящемся движении, что позволяет
мозжечку изменять положение тела для выполнения последующего
движения, а также обеспечивает взаимную коррекцию позных и
локальных движений в поведенческом акте.
Экстрапирамидная система. Базальные ядра конечного мозга, моз­
жечок, красное ядро и черное вещество среднего мозга составля­
ют так называемую экстрапирамидную систему. Экстрапирамидная
система определяет длительное тоническое возбуждение, одновре­
менную активацию или торможение больших мышечных групп,
контроль осевой мускулатуры туловища и проксимальных отделов
конечностей. Основными эфферентными путями экстрапирамидной системы являются красноядерно-покрышечно-преддверно-ретикулярные, луковично-ретикулярно-спинальные и моноаминергические пути. А ктивация красноядерно-спинномозговых влияний
приводит к возбуждению а - и у-мотонейронов сгибателей и тормо­
жению соответствующих мотонейронов разгибателей. Центробежные
моторные команды, идущие по вестибуло-спинномозговому пути,
вызывают обратные эффекты: активацию а - и у-мотонейронов раз­
гибателей и торможение соответствующих мотонейронов, иннерви-
Рис. 7.4. Позные реакции у децеребрированного животного с удаленными
лабиринтами: А — исходное состояние; Б — пассивный подъем головы
сопровождается снижением тонуса разгибателей задних лап и повышени­
ем тонуса разгибателей передних лап; В — пассивный наклон головы вниз
оказывает противоположный эффект (по Р. Шмидту, 1985)
596
Глава 7
Двигательные функции
5 97
рующих мышцы-сгибатели. Текто-спинальный тракт регулирует
контрлатеральные движения головы в ответ на зрительные, слуховые
и тактильные стимулы. Моноаминергические пути начинаются от
голубых ядер ствола мозга, содержащих норадреналин. Проекции
этих ядер на структуры спинного мозга вызывает торможение интер­
нейронов и мотонейронов спинного мозга.
Ретикулярная формация среднего и продолговатого мозга ока­
зывает сложные влияния на соматическую мускулатуру. Нейроны
ретикулярной формации продолговатого мозга возбуждают а - и
у-мотонейроны сгибателей и тормозят аналогичные мотонейроны
разгибателей. Нейроны ретикулярной формации моста мозга оказы­
вают возбуждающее влияние на моторные элементы спинного мозга.
Эти нисходящие влияния участвуют в поддержании тонуса мышц,
формировании позы и обеспечении равновесия.
Базальные ядра конечного мозга представляют собой подкорковое
звено между ассоциативными и двигательными областями коры
мозга. К ним относятся полосатое тело, состоящее из хвостатого ядра
и скорлупы, бледный шар и миндалина. Большая часть афферентных
сигналов от мозжечка поступает в полосатое тело. Эфферентные
волокна связывают базальные ядра с таламусом и крышей среднего
мозга. Установлено, что раздражение этих структур, как правило, не
вызывает какого-либо конкретного движения, но изменяет возбуди­
мость всех моторных нейронов на разных уровнях ЦНС.
Повреждения базальных ядер приводят к различным расстройс­
твам в моторной сфере. Наиболее известна болезнь Паркинсона, при
которой наблюдаются акинезия, ригидность, тремор покоя, маско­
образное неподвижное лицо. Акинезия представляет собой трудность
начала или завершения движения, ригидность — увеличение мышеч­
ного тонуса (восковая ригидность — пластическое изменение тонуса
мышц), тремор покоя — появление дрожаний головы и конечностей с
Рис. 7.5. Участие ядерных зон мозжечка в организации двигательных функ­
ций: А — изначально медиальная зона (1) мозжечка при участии ядер шатра
(2), ретикулярной формации (3) и вестибулярных ядер (4) обеспечивает фор­
мирование исходной позы тела; Б — затем промежуточная зона (1), вовлекая
пробковидное (2) и красное (3) ядро, формирует подготовку к движению
и дальнейшую коррекцию по мере его осуществления; В — наконец, лате­
ральная зона (1) при участии зубчатого ядра (2) и красного ядра (3) обеспе­
чивает осуществление самого быстрого локального движения
Глава 7
5 98
частотой 4 -7 Гц и более. Акинезию рассматривают как нарушение воз­
никновения импульсов в базальных ядрах, ригидность и тремор — как
повышенную активность ядер вследствие их растормаживания.
Медиаторами в различных нейронах базальных ядер являются
глутамат, дофамин, ГАМК, вещество П. Болезнь Паркинсона, воз­
можно, связана с разрушением путей, идущих от черного вещества к
полосатому телу. Симптомы заболевания часто снимаются введением
предшественника дофамина — L-дофы.
На рис. 7.6 показана последовательность вовлечения различных
структур головного мозга в формирование позы.
Произвольные движения
Участие головного м озга в формировании произвольных
движений
Началу произвольной двигательной реакции предшествуют про­
цессы афферентного синтеза и принятия решения, приводящие к
созданию общей цели поведения.
Ассоциативная кора мозга. Ведущая роль в этих процессах отво­
дится ассоциативным областям коры большого мозга — лобным и
теменным. Электрофизиологическим проявлением процессов пос­
тановки цели служит так называемая волна ожидания (Г. Уолтер).
Если человека обучить выполнять нажатие кнопки по световому
сигналу, следующему через 2 с после подачи звука, то в интервале
между двумя этими раздражителями (предупредительным — звуком
и пусковым — светом) в ассоциативных областях возникает мед­
ленное отрицательное колебание общей электрической активности.
Амплитуда волны ожидания отражает уровень внимания и степени
готовности субъекта к действию. Возбуждение, несущее информа­
цию от общей цели будущего действия из ассоциативных областей
коры, распространяется к базальным ядрам и мозжечку (рис. 7.7).
Рис. 7.6. Вовлечение различных отделов головного мозга в формирование
позы: А — активация ассоциативных областей коры (1), возбуждение из
которых поступает к мозжечку (2) и базальным ядрам (3); Б — активация
нейронов мозжечка и базальных ядер, возбуждения из которых поступают
к двигательным ядрам среднего мозга (4) и ретикулярной формации (5);
В — активация ядер среднего мозга и ретикулярной формации, возбужде­
ние из которых поступает к нейронам спинного мозга (6)
Двигательные функции
_
5 99
Глава 7
600
■
Шг
•••
Г
/1
—
Сгибание пальца
Двигательные функции
601
Базальные ядра. В базальных ядрах создаются первичные мотор­
ные команды, внешним проявлением которых служат моторные
потенциалы. Они возникают, в частности, в хвостатом ядре поло­
сатого тела за 60 мс до начала произвольного движения. Показано
участие нейронов скорлупы в медленных движениях руки в опре­
деленном направлении. При быстрых или медленных движениях,
но в противоположных направлениях, соответствующей активации
нейронов скорлупы не обнаружено. Начало активации нейронов
скорлупы, как правило, предшествует началу движения. Считают,
что базальные ядра формируют программы начала движения и его
окончания. По этой причине при поражении базальных ядер возни­
кают непроизвольные движения, нарушается кинематика начала и
конца движения (акинезия).
М озжечок. Вовлечение мозжечка в системные процессы локаль­
ных движений осуществляется в результате поступающей к нему
импульсации от ассоциативных зон коры большого мозга. Нейроны
мозжечка активируются через 30 мс после разряда корковых нейро­
нов — также задолго до начала движения. Показано, что импульсная
активность нейронов мозжечка на 50—60 мс опережает мышечную
активность руки обезьяны, выполняющей потягивание за рычаг
в ответ на световой сигнал. Скорость проведения возбуждения по
нервным волокнам внутри мозжечка, а также по его эфферентным
путям достаточно велика. Наличие значительного сенсорного при­
тока от рецепторов мышц и суставов позволяет мозжечку выступать
в качестве блока сравнения, осуществляющего общую регуляцию
моторной функции по принципу обратной связи. Вследствие этого
структуры мозжечка быстро исправляют отклонения в траектории
Рис. 7.7. Динамика электроэнцефалографических показателей произволь­
ной двигательной реакции: А — изолированное применение щелчка
и вспышек света сопровождается вызванными потенциалами в коре;
Б — вспышки света, останавливаемые испытуемым по инструкции нажа­
тии кнопки на фоне когнитивного негативного отклонения («волна ожи­
дания») в лобных отделах коры головного мозга человека в ответ на пре­
дупреждающий звуковой сигнал (по Г. Уолтеру, 1966); В — «потенциал
готовности», регистрирующийся на всей поверхности головы перед произ­
вольным сгибанием большого пальца правой руки; Г — «моторный потен­
циал» возникает в той же ситуации в участке двигательной области левой
прецентральной извилины (по Р. Шмидту, 1985)
6 02
Глава 7
движения. Двигательные программы мозжечка обеспечивают общую
траекторию движения путем объединения отдельных движений в
целостный двигательный акт. Именно поэтому при охлаждении моз­
жечка возникает распад целостного двигательного акта на отдельные
фрагменты.
Промежуточная (околочервячная) зона мозжечка (см. рис. 7.5) полу­
чает копию сигналов от пирамидного тракта о готовящемся движении,
что позволяет мозжечку изменять положение тела для выполнения
последующего движения, а также обеспечивает взаимную коррек­
цию позных и локальных движений в поведенческом акте. Благодаря
наличию значительного сенсорного притока от рецепторов мышц и
суставов мозжечок работает как аппарат сравнения, осуществляющий
общую регуляцию моторной функции по принципу обратной связи.
Вследствие этого мозжечок при участии промежуточной зоны быстро
исправляет отклонения в траектории движения, что обеспечивает
подготовку к движению с его последующей коррекцией.
Латеральная зона мозжечка, получающая информацию от всех
отделов коры мозга, посылает импульсы к зубчатому ядру и далее к
красному ядру среднего мозга. Выраженные тормозные процессы в
этом отделе мозжечка делают невозможным хранение информации
уже через 30 мс после ее поступления, что создает условия для обес­
печения быстрых двигательных актов, например, в спортивных играх
или игре на музыкальных инструментах. Таким образом, латераль­
ная зона принимает участие в формировании программы быстрых
фазических движений.
При органических поражениях мозжечка также происходит распад
сложных двигательных программ действия — наблюдаются различ­
ные виды атаксии — нарушение точности, скорости и направления
движения. Появляются асинергия (движения перестают быть плавны­
ми, согласованными) и интенционный тремор в начале движения,
направленного на определенный объект.
Таламус. Импульсы от мозжечка и базальных ядер поступают в
ядра таламуса. Активация ядер таламуса начинается также до появ­
ления двигательной реакции и до возбуждения мотонейронов дви­
гательной области коры мозга. В это время в таламусе наблюдаются
«потенциалы готовности», длительность которых составляет 800 мс
(см. рис. 7.7). Таламус, в свою очередь, посылает импульсы в мотор­
ные области коры мозга.
Двигательные области коры большого мозга. Основные про­
цессы, приводящие к формированию произвольного движения,
Двигательные функции
603
осуществляются в двигательной коре большого мозга, нейроны кото­
рой получают импульсы от мозжечка, базальных ядер и таламуса.
В двигательной коре за 40—50 мс до начала произвольного движения
возникает «моторный потенциал», что показано в специальных экс­
периментах на обезьянах, обученных тянуть рычаг в ответ на предъ­
явление светового сигнала. Считают, что в моторной коре имеется
представительство движений, совершаемых отдельными мышцами,
но не самих мышц. Установлен различный характер импульсной
активности одних и тех же нейронов при выполнении однотипного
движения в различных поведенческих ситуациях (рис. 7.8). Одни и те
же нейроны могут реагировать по-разному при выполнении живот­
ными различных движений в ответ на условный сигнал, подкрепля­
емый различными воздействиями.
Всякое изменение цели действия тут же приводит к изменению
общей моторной программы.
Показано, что соматотопическая организация двигательной коры
отражает вклад того или иного участка тела в сферу моторной
деятельности. Участки тела и конечностей, выполняющие разно­
образные и обширные двигательные функции, занимают особенно
большие территории. Пирамидные нейроны, выполняющие близкие
функции, находятся друг над другом, создавая вертикальные колон­
ки. Одна такая колонка контролирует работу нескольких мышц,
обеспечивая движение в одном суставе. В связи с этим представи­
тельство одной мышцы всегда множественно и вызывается раздра­
жением различных локусов одной области коры.
Двигательные колонки содержат несколько сотен пирамидных
клеток и имеют диаметр около 1 мм. Колонки активируют или
тормозят нейроны спинального моторного пула. Несколько мор­
фологических колонок объединяются в функциональную колонку,
а совокупность функционирующих колонок составляет моторное
представительство того или иного участка тела с мышцами и сус­
тавами. Представительство каждой отдельной мышцы в коре мно­
жественно. Это означает, что ее сокращение можно получить при
раздражении различных участков двигательной области коры. Было
установлено, что большие пирамидные клетки Беца, обладающие
высокой скоростью проведения возбуждения, разряжаются во время
движения, а малые пирамидные нейроны с меньшей скоростью про­
ведения возбуждения имеют постоянную импульсацию. Возможно,
что эти нейроны соответствуют фазическим и тоническим а-м ото­
нейронам спинного мозга.
604
Глава 7
0,5 кГ
- t t --------------- ffr
2 кГ
1 с.
\
Б ы стро
М ед л е н н о
•нтт
Двигательные функции
6 05
Двигательная программа может быть запущена искусственным
путем при электрическом раздражении либо клеток коры большого
мозга, либо проводящих путей. Работами X. Дельгадо установлено, что
вызванные таким образом движения хорошо скоординированы и выгля­
дят в достаточной степени естественно. Однако в этих ситуациях нельзя
добиться выполнения последовательных действий. В естественных же
условиях у субъектов происходят постоянный перебор и смена рабо­
чих программ, т.е. непрерывный динам