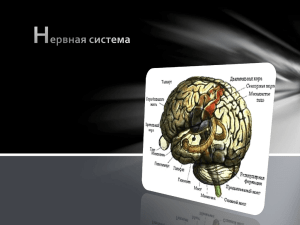
ИЖЕВСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ КАФЕДРА ГИСТОЛОГИИ. ЭМБРИОЛОГИИ И ЦИТОЛОГИИ ОБЩАЯ ГИСТОЛОГИЯ ИЖЕВСК–2002 УДК ББК Составители: докт.мед.наук Г.В.Шумихина, докт.мед.наук Ю.Г.Васильев, доц.А.А.Соловьев, канд.мед.наук В.М.Кузнецова, С.А.Соболевский, С.В.Кутявина, И.В.Титова, Т.Г.Глушкова Рецензент: доктор мед.наук, профессор каф. мед.биологии ИГМА Н.Н.Чучкова Общая гистология:Учебно-методическое Ю.Г.Васильев, А.А.Соловьев и др.–Ижевск, 2002.– Иллюстрации: доктор мед.наук Ю.Г.Васильев пособие с. /Сост.Г.В.Шумихина, Данное методическое пособие составлено согласно программе по гистологии, цитологии и эмбриологии для студентов высших учебных заведение ВУНМЦ МЗ РФ (Москва, 1997). Пособие предназначено для студентов медицинских вузов всех факультетов. Приведены современные представления о микроанатомической, гистологической и клеточной организации тканей человека. Пособие изложено в лаконичной форме, сопровождается вопросами для самоконтроля, клиническими примерами, иллюстрациями. Издание подготовлено сотрудниками кафедры гистологии, эмбриологии и цитологии Ижевской государственной медицинской академии. Предназначено для студентов лечебного, педиатрического, стоматологического факультетов. УДК ББК Г.В.Шумихина, Ю.Г.Васильев, А.А.Соловьев и др., составления, 2002. ВВЕДЕНИЕ В ТКАНИ Ткань – это возникшая в процессе эволюции (филогенезе) система из взаимодействующих между собой и нередко общих по происхождению гистологических элементов (клеток и их производных), обладающая собственной особенностью строения и специфическими функциями. Ткани возникли в ходе эволюции у многоклеточных организмов на определённых этапах филогенеза. Первые признаки примитивных тканей можно обнаружить у таких представителей животного мира, как губки и кишечно-полостные. В процессе индивидуального развития (онтогенеза), в значительной мере повторяющего филогенез, их источниками являются эмбриональные зачатки. Теория дивергентного развития тканей; развития тканей в фило- и онтогенезе (Н.Г. Хлопин), предполагает, что ткани возникли в результате дивергенции (расхождения признаков), в ходе которой однотипные клетки тканевого зачатка постепенно приобретают по мере развития всё более выраженные различия в структуре и функции, приспосабливаясь к новым условиям существования. Иными словами, тканевые элементы эволюционных и эмбриональных зачатков тканей, попадая в разные условия (окружение), дают большое разнообразие морфофункциональных типов вследствие приспособления их строения к новым условиям функционирования. Причины эволюции тканей описывает теория параллельных рядов тканевой эволюции (А.А. Заварзин), согласно которой ткани, выполняющие сходные функции, имеют сходное строение. В ходе филогенеза одинаковые ткани возникали параллельно в разных эволюционных ветвях животного мира, т.е. совершенно разные филогенетические типы первоначальных тканей, попадая в сходные условия существования внешней или внутренней среды, давали сходные морфофункциональные типы тканей. Эти типы возникают в филогенезе независимо друг от друга, т.е. параллельно, у абсолютно разных групп животных при стечении одинаковых обстоятельств эволюции. Эти две взаимодополняющие друг друга теории объединены в единую эволюционнную концепцию тканей (А.А. Браун и П.П. Михайлов), согласно которой сходные тканевые структуры в различных ветвях филогенетического древа возникали параллельно в ходе дивергентного развития. С теориями эволюции и происхождения тканей тесно связана их классификация. Существуют 2 основных принципа классификации тканей: 1.Гистогенетическая классификация основывается на происхождении тканей в процессах онто- и филогенеза из разных зачатков. Она логически связана с теорией дивергентного развития Н.Г. Хлопина и частоошибочно носит его имя. Наличие общих свойств у тканей, развившихся из одного эмбрионального зачатка, позволяет объединять их в единый тканевой тип. Выделяют ткани: а) эктодермального типа, б) энтодермального типа, в) нейрального типа, г) мезенхимального типа, д) мезодермального типа. 2. Морфофункциональная классификация, наиболее распространенная среди гистологов в настоящее время, объединяет ткани в четыре группы по признакам сходства их строения и (или) выполняемой функции. Различают: а) эпителиальные, б) соединительные (ткани внутренней среды), в) мышечные и г) нервную. Каждая морфофункциональная группа может включать в себя ряд подгрупп. Эту классификацию обычно связывают с именем А.А. Заварзина, на примере эволюции тканей показавшем тесную взаимосвязь строения и выполняемой функции. Генетическая и морфофункциональная классификации тканей не универсальны и дополняют друг друга, поэтому часто при характеристике тканей указывают на их происхождение, например: эктодермальный эпителий, мышечная ткань мезенхимального типа. На этом принципе построена классификация эпителиальных тканей по Н.Г. Хлопину, который онтогенетически в данной морфофункциональной группе выделяет: эпидермальные эпителии; энтеродермальные эпителии; целонефродермальные эпителии; эпендимоглиальные эпителии и эпителии ангиодермального типа. Принципы структурной организации тканей. Некоторые ткани состоят преимущественно из клеток (эпителиальная, нервная, гладкая и сердечная мышечные ткани). В тканях внутренней среды (кровь, соединительные, скелетные ткани) помимо клеток хорошо выражено межклеточное вещество. Основным компонентом скелетной мышечной ткани являются мышечные волокна. Эти разнообразные структурнофункциональные составляющие тканей в гистологии называются гистологические элементы и подразделяются на 2 основных типа : 1. Гистологические элементы клеточного типа обычно являются живыми структурами с собственным метаболизмом, ограниченные плазматической мембраной, и представляют собой клетки и их производные, возникшие в результате специализации. К ним относятся: а) Клетки – главные элементы тканей, определяющие их основные свойства; б) Постклеточные структуры, в которых утеряны важнейшие для клеток признаки (ядро, органоиды), например: эритроциты, роговые чешуйки эпидермиса, а также тромбоциты, вообще являющиеся частями клеток; в) Симпласты – структуры, образованные в результате слияния отдельных клеток в единую цитоплазматическую массу с множеством ядер и общей плазмолеммой, например: волокно скелетной мышечной ткани, остеокласт; г) Синцитии – структуры, состоящие из клеток, объединенных в единую сеть цитоплазматическими мостиками вследствие неполного разделения, например: сперматогенные клетки на стадиях размножения роста и созревания. 2. Гистологические элементы неклеточного типа представлены веществами и структурами, которые вырабатываются клетками и выделяются за пределы плазмолеммы, объединенными под общим названием «межклеточное вещество» (тканевой матрикс). Межклеточное вещество обычно включает в себя следующие разновидности: а) Аморфное (основное) вещество – представлено бесструктурным скоплением органических (гликопротеины, гликозоаминогликаны, протеогликаны) и неорганических (соли) веществ, находящихся между клетками ткани в жидком, гелеобразном или твердом, иногда кристаллизованном состоянии (основное вещество костной ткани); б) Волокна– состоят из фибриллярных белков (эластин, различные виды коллагена), часто образующих в аморфном веществе пучки разной толщины, взаимодействующие с клеточными элементами тканей. Среди них различают: 1) коллагеновые, 2) ретикулярные и 3) эластические волокна . Фибриллярные белки участвуют также в формировании капсул клеток (хрящи, кости) и базальных мембран (эпителии). Клеточные популяции. У человека более 120 типов клеток, которые можно идентифицировать на этапах их дифференцировки. Тканевые признаки клеток базируются на наличии или отсутствии межклеточных контактов, взаимоотношениями с межклеточным веществом и структурными элементами других тканей. Специфику клеток каждой разновидности тканей определяют размеры, форма, специальные структуры поверхностей, органоиды, ферменты и другие параметры. Тканевые признаки трудно идентифицировать у родоначальных (стволовых) клеток. В ходе дифференцировки клетки приобретают не только специфичные для каждого дифферона структурно-функциональные признаки, но и особый спектр рецепторов к регуляторам их жизненной активности (гормонам, медиаторам, факторам роста, кейлонам, цитокинам и другим). Указанные факторы носят системообразующий характер и определяют специфику жизнедеятельности того или иного вида тканей. Сообщества клеток, входящих в ткани, принято называть клеточными популяциями. В широком понимании клеточные популяции – это совокупность клеток организма или ткани, сходных между собой по какому-либо признаку. Например, по способности к самообновлению путём деления выделяют 4 категории клеточных популяций (по Леблону): 1. Эмбриональная (быстро делящаяся клеточная популяция) – все клетки популяции активно делятся, специализированные элементы отсутствуют. 2. Стабильная клеточная популяция – долгоживущие, активно функционирующие клетки, которые вследствие крайней специализации утратили способность к делению. Например, нейроны, кардиомиоциты. 3. Растущая (лабильная) клеточная популяция – специализированные клетки которой способны делиться в определённых условиях. Например, эпителии почки, печени. 4. Обновляющаяся популяция состоит из постоянно и быстро делящихся клеток и их специализированных функционирующих потомков, продолжительность жизни которых ограничена. Например, эпителии кишечника, форменные элементы крови. В узком смысле клеточная популяция – это однородная группа клеток (клеточный тип), сходных по строению, функции и происсхождению, а также по уровеню дифференцировки . Например, популяция стволовых клеток крови. К особому типу клеточных популяций относят клон – группу идентичных клеток, происходящих от одной родоначальной клетки-предшественницы. Понятие клон как самое узкое толкование клеточной популяции часто используется в иммунологии, например, клон Тлимфоцитов. Детерминация и дифференцировка клеток, дифферон. Развитие тканей в фило- и эмбриогенезе связано с процессами детерминации и дифференцировки их клеток. Детерминация – это процесс, определяющий направление развития клеток, тканей. В ходе детерминации клетки получают возможность развиваться в определённом направлении (т.е. происходит ограничение их потенций). На молекулярно-биологическом уровне этот механизм осуществляется поэтапным блокированием части клеточного генома и уменьшением числа разрешённых к экспрессии генов. Ступенчатое, согласованное с программой развития организма, ограничение возможных путей развития вследствие детерминации называется коммитированием. Детерминация клеток и тканей в организме, как правило, необратима. Дифференцировка. В ходе дифференцировки происходит постепенное формирование морфологических и функциональных признаков специализации клеток тканей (образование клеточных типов). Дифференцировка направлена на создание в многоклеточном организме нескольких структурно-функциональных типов клеток. У +э ,8с разным уровнем дифференцировки. Поэтому клеточные популяции ткани можно рассматривать как совокупность клеточных форм (видов клеток), находящихся на разных этапах своего развития, от наименее дифференцированных (стволовых), до зрелых, наиболее дифференцированных. Такой гистогенетический ряд развивающихся клеток одинакового происхождения, но находящихся на разных этапах дифференцировки, в гистологии принято называть диффероном. Многие ткани содержат не один, а несколько клеточных дифферонов, которые взаимодействуют друг с другом. Поэтому ткань нельзя рассматривать как систему клеток одного типа, сходных по строению, функции и происхождению. В составе дифферона последовательно (по степени дифференцировки) различают следующие клеточные популяции: а) стволовые клетки - наименее дифференцированные клетки данной ткани, способные делиться и являющиеся источником развития других её клеток; б) полустволовые клетки-предшественники имеют ограничения в способности формировать различные типы клеток, вследствие коммитирования, но способны к активному размножению; в) клетки-бласты, вступившие в дифференцировку но сохраняющие способность к делению; г) созревающие клетки, заканчивающие дифференцировку; д)зрелые (дифференцированные) клетки. Последние заканчивают гистогенетический ряд, способность к делению у них, как правило, исчезает, в ткани они активно функционируют. Можно выделить также популяцию закончивших активное функционирование (старых) клеток. Уровень специализации клеток в популяциях дифферона возрастает от стволовых до зрелых клеток. При этом происходят изменения состава и активности ферментов, органоидов клеток. Для гистогенетических рядов дифферона характерен принцип необратимости дифференцировки, т.е. в нормальных условиях переход от более дифференцированного состояния к менее дифференцированному невозможен. Это свойство дифферона часто нарушается при патологических состояниях (злокачественные опухоли, неоплазии). Наличие в тканях малодифференцированных клеток, способных к митотическому делению, обеспечивает способность ткани к самообновлению и восстановлению (регенерации). Такую, имеющуюся в ткани совокупность клеток, способных к делению, называют камбием. Камбиальные элементы – это популяции стволовых, полустволовых клеток-предшественников, а также бластных клеток данной ткани, деление которых поддерживает необходимое число ее клеток и восполняет убыль популяции зрелых элементов. В тех тканях, в которых не происходит обновления клеток путем их деления, камбий отсутствует. По распределению камбиальных элементов ткани различают несколько разновидностей камбия: *Локализованный камбий – его элементы сосредоточены в конкретных участках ткани, например, в многослойном эпителии камбий локализован в базальном слое; *Диффузный камбий – его элементы рассеяны в ткани, например, в гладкой мышечной ткани камбиальные элементы рассредоточены среди дифференцированных миоцитов; *Вынесенный камбий – его элементы лежат за пределами ткани и по мере дифференцировки включаются в состав ткани, например кровь как ткань содержит только дифференцированные элементы, элементы камбия находятся в органах кроветворения. Регенерация тканей. Регенерация ткани – процесс, обеспечивающий её обновление в ходе нормальной жизнедеятельности (физиологическая регенерация) или восстановление после повреждения (репаративная регенерация). Хотя полноценная регенерация ткани включает обновление (восстановление) ее клеток и их производных, включая межклеточное вещество, основную роль в регенерации тканей играют клетки, так как именно они служат источником всех остальных компонентов тканей. Поэтому возможность регенерации ткани определяется способностью ее клеток к делению и дифференцировке или уровнем внутриклеточной регенерации. Хорошо регенерируют те ткани, которые имеют камбиальные элементы или представляют собой обновляющиеся или растущие леблоновские клеточные популяции. Активность деления (пролиферации) клеток каждой ткани при регенерации контролируется факторами роста, гормонами, цитокинами, кейлонами, а также характером функциональных нагрузок. Необходимо отличать тканевую и клеточную регенерацию путем деления клеток от внутриклеточной регенерации, которую следует понимать как процесс непрерывного обновления или восстановления структурных компонентов клетки после их повреждения. Внутриклеточная регенерация универсальна, то есть свойственна всем клеткам тканей организма человека. В тех тканях, которые являются стабильными клеточными популяциями и в которых отсутствуют камбиальные элементы (нервная, сердечная мышечная ткань), данный тип регенерации является единственно возможным способом обновления и восстановления их структуры и функции. Ткани в процессе жизнедеятельности могут подвергаться гипертрофии и атрофии. Гипертрофия ткани – увеличение ее объема, массы и функциональной активности, - обычно является следствием а) гипертрофии ее отдельных клеток (при неизменном их числе) вследствие усиленной внутриклеточной регенерации в условиях преобладания анаболитических процессов над катаболическими; б) гиперплазии – увеличении числа ее клеток путем активации клеточного деления (пролиферации) и (или) в результате ускорения дифференцировки новообразующихся клеток; в) сочетания обоих процессов. Атрофия ткани – снижение ее объема, массы и функциональной активности вследствие а) атрофии ее отдельных клеток вследствие преобладания процессов катаболизма, б) гибели части ее клеток, в) резкого уменьшения скорости деления и дифференцировки клеток. Межтканевые и межклеточные отношения. Ткань поддерживает постоянство своей структурно-функциональной организации (гомеостаз) как единого целого только при условии постоянного влияния гистологических элементов друг на друга (внутритканевые взаимодействия), а также одних тканей на другие (межтканевые взаимодействия). Эти влияния можно рассматривать как процессы взаимного узнавания элементов, образования контактов и обмена информацией между ними. При этом формируются самые различные структурно-пространственные объединения. Клетки в ткани могут находиться на расстоянии и взаимодействовать друг с другом через межклеточное вещество (соединительные ткани), соприкасаться отростками, иногда достигающими значительной длины (нервная ткань), или образовывать плотно контактирующие клеточные пласты (эпителий). Совокупность тканей, объединенных в единое структурное целое соединительной тканью, координированное функционирование которого обеспечивается нервными и гуморальными факторами, образует органы и системы органов целого организма. Для образования ткани необходимо, чтобы клетки объединились и были связаны между собой в клеточные ансамбли. Способность клеток избирательно прикрепляться друг к другу или к компонентам межклеточного вещества осуществляется с помощью процессов узнавания и агдезии, которые являются необходимым условием поддержания тканевой структуры. Реакции узнавания и агдезии происходят вследствие взаимодействия макромолекул специфических мембранных гликопротеидов, получивших название молекул агдезии. Прикрепление происходит с помощью особых субклеточных структур: а) точечных агдезионных контактов (прикрепление клеток к межклеточному веществу), б) межклеточных контактов (прикрепление клеток друг к другу). В их состав входят особые трансмембранные белки и гликопротеиды – кадгерины, иммуноглобулины, интегрины и коннексины, а также белки, осуществляющие прикрепление этих структур к компонентам клеточного матрикса, – актинин, винкулин, талин. Кроме того, на поверхности клеток находятся агдезивные рецепторы и соответствующие им лиганды, обеспечивающие специфическое взаимное распознавание элементов ткани. К агдезионным белкам межклеточного матрикса относят фибронектин и витронектин. Межклеточные контакты - специализированные структуры клеток, с помощью которых они механически скрепляются между собой, а также создают барьеры и каналы проницаемости для межклеточной коммуникации. Различают: 1) агдезионные клеточные контакты, выполняющие функцию межклеточного сцепления (промежуточный контакт, десмосома, полудесмасома), 2) замыкающие контакты, функция которых - образование барьера, задерживающего даже малые молекулы (плотный контакт), 3) проводящие (коммуникационные) контакты, функция которых состоит в передаче сигналов от клетки к клетке (щелевой контакт, синапс). Регуляция жизнедеятельности тканей. Гуморальные факторы, обеспечивающие межклеточное взаимодействие в тканях и их метаболизм, включают в себя разнообразные клеточные метаболиты, гормоны, медиаторы, а также цитокины и кейлоны. Цитокины являются наиболее универсальным классом внутри- и межтканевых регуляторных веществ. Они представляют собой гликопротеиды, которые в очень низких концентрациях оказывают влияние на реакции клеточного роста, пролиферации и дифференцировки. Действие цитокинов обусловлено наличием рецепторов к ним на плазмолемме клеток-мишеней. Эти вещества переносятся кровью и обладают дистантным (эндокринным) действием, а также распространяются по межклеточному веществу и действуют локально (ауто – или паракринно). Важнейшими цитокинами являются интерлейкины (ИЛ), факторы роста, колониестимулирующие факторы (КСФ), фактор некроза опухоли (ФНО), интерферон. Клетки различных тканей обладают большим количеством рецепторов к разнообразным цитокинам ( от 10 до 10000 на клетку), эффекты которых нередко взаимо перекрываются, что обеспечивает высокую надёжность функционирования этой системы внутриклеточной регуляции. Кейлоны представляют собой факторы, вырабатываемые дифференцированными клетками данной ткани и угнетающие деление её малодифференцированных камбиальных элементов. Благодаря продукции кейлонов осуществляется поддержание относительного постоянства числа клеток в зрелой ткани. При повреждении ткани и убыли её зрелых клеток снижение продукции кейлонов вызывает усиленную пролиферацию клеток, приводящую к регенерации ткани. Межтканевые отношения. Ткани в организме существуют не изолированно, а в постоянном взаимодействии с другими тканями, что способствует поддержанию их нормальной функциональной организации. Это так называемые индуктивные взаимодействия, утрата которых, например, при культивировании тканей in vitro в оптимальных условиях вызывает изменения морфологии и потерю ряда функций, характерных для этих тканей in vivo. Межтканевые взаимодействия осуществляются посредством локальных метаболитов и дистантных гуморальных факторов, включающих в себя гормоны, нейромедиаторы и другие информационные молекулы. Взаимодействие тканей, образующих органы на уровне целостного организма, контролируются эндокринной, нервной и иммунной системами. Межтканевые отношения определяют структуру и функцию органа, обеспечивают оптимальные уровни физиологической и репаративной регенерации. 1.ТЕМА: ЭПИТЕЛИАЛЬНЫЕ ТКАНИ. ЖЕЛЕЗЫ. Цели занятия: Научиться: 1.Характеризовать основные морфофункциональные и гистогенетические особенности эпителиальных тканей. 2.Сопоставлять микроскопические, ультрамикроскопические и гистохимические особенности различных видов эпителиальных тканей с выполняемой ими функцией. Объяснять механизм секреторного процесса в железистых эпителиальных клетках. 3. Определять эпителиальную ткань на микроскопическом уровне, идентифицировать различные виды покровного и железистого эпителия. 4.Научиться определять тип экзокринных желез по их строению и характеру выделяемого секрета. Эпителиальные ткани, или эпителии (от греч. epi – над и thele – сосок, тонкая кожица) – часто выступают как пограничные ткани, располагаясь на границе с внешней средой, покрывают поверхность тела, выстилают его полости, слизистые оболочки внутренних органов и образуют большинство желез. В связи с этим различают два вида эпителиев: I. Покровные эпителии (образуют разнообразные выстилки в виде пластов). II. Железистые эпителии (образуют железы). Общие морфологические признаки эпителия как ткани: 1.Эпителиоциты располагаются плотно друг к другу. 2.Между клетками практически нет межклеточного вещества. 3.Между клетками находятся межклеточные контакты. 4.Эпителии часто занимают пограничное положение (обычно между тканями внутренней среды и внешней средой). 5.Для эпителиоцитов характерна полярность клеток. Различают апикальный и базальный полюсы, последний обращен к базальной мембране. Многослойным эпителиям свойственна вертикальная анизоморфность – неодинаковые морфологические свойства клеток различных слоев эпителиального пласта. 6.Эпителиоциты располагаются на базальной мембране – особом неклеточном образовании, которое создает основу для эпителия, обеспечивает барьерную и трофическую функции. 7.В эпителии отсутствуют сосуды; питание осуществляется путем диффузии веществ через базальную мембрану из сосудов соединительной ткани. 8.Для большинства эпителиев характерна высокая способность к регенерации – физиологической и репаративной, которая осуществляется благодаря камбию. Морфологические особенности клеток составляющих эпителиальную ткань варьируют в широких пределах, различаясь как в разных типах эпителиев, так и между отдельными клетками в пределах одного типа. Эти особенности тесно связаны с функцией клеток и их положением в эпителиальном пласте. Форма эпителиоцитов служит важным классификационным признаком, как отдельных клеток, так и эпителиальных пластов в целом. Выделяют плоские, кубические и призматические клетки. Ядро эпителиоцитов может иметь различную форму, которая обычно соответствует форме клетки: в плоских– оно дисковидное, в кубических – сферическое, в цилиндрических – эллипсоидное. В большинстве клеток ядро сравнительно светлое, содержит хорошо заметное крупное ядрышко, однако в оровевающих эпителиях по мере дифференцировки клеток оно уменьшается, уплотняется или лизируется – подвергается кариопикнозу, кариорексису или кариолизису. Цитоплазма эпителиоцитов содержит все органеллы общего значения, а в некоторых клетках – также органеллы специального значения, обеспечивающие выполнение специфических функций данных клеток. В клетках железистого эпителия хорошо развит синтетический аппарат. В связи с полярностью клеток органеллы распределены в их цитоплазме неравномерно. Цитоскелет эпителиоцитов хорошо развит, представлен микротрубочками, микрофиламентами (диаметром до 4нм) и промежуточными филаментами (диаметром 810 нм). Последние в эпителиоцитах особенно многочисленны и называются тонофиламентами, которые при фиксации склеиваются, образуя крупные агрегаты, выявляемые под световым микроскопом и описанные под названием тонофибрилл. Цитокератины – белки, образующие тонофиламенты, которые специфичны для клеток эпителиальных тканей. Идентифицировано около 30 различных форм цитокератинов, причем выработка каждого вида цитокератина кодируется особым геном. Для конкретного вида эпителия (а в многослойных эпителиях – для каждого слоя) характерен определенный набор цитокератинов, экспрессию которых рассматривают как маркер дифференцировки эпителиальных клеток. Изменения нормальной экспрессии цитокератинов могут указывать на нарушения дифференцировки клеток и в ряде случаев служить важным диагностическим признаком их злокачественного перерождения. Поверхности эпителиоцита (латеральная, базальная, апикальная) обладают отчетливой структурно-функциональной специализацией, которая особенно хорошо выявляется в однослойном эпителии, в том числе в железистом эпителии. Латеральная поверхность эпителиоцитов обеспечивает взаимодействие клеток за счет межклеточных контактов, которые обуславливают механическую связь эпителиоцитов друг с другом – это плотные контакты, десмосомы, интердигитации, а также химическую (метаболическую, ионную и электрическую) связь между эпителиоцитами – это щелевые контакты. Базальная поверхность эпителиоцитов прилежит к базальной мембране, к которой она прикреплена с помощью полудесмосом. В функциональном плане базальная и латеральная (до уровня плотных соединений) части плазмолеммы эпителиоцита в совокупности образует единый комплекс, мембранные белки которого служат: а) рецепторами, воспринимающими различные сигнальные молекулы, б) переносчиками питательных веществ, поступающих из сосудов подлежащей соединительной ткани, в) ионными насосами и др. Базальная мембрана (БМ) связывает эпителий и подлежащую соединительную ткань и образована компонентами, которые вырабатываются этими тканями, БМ поддерживает нормальную архитектонику, дифференцировку и поляризацию эпителия; обеспечивает избирательную фильтрацию питательных веществ. На светооптическом уровне на препаратах она имеет вид тонкой полоски, плохо окрашивается гематоксилином и эозином. На ультраструктурном уровне в базальной мембране выделяют три слоя (в направлении от эпителия): 1) светлая пластинка, которая соединяется с полудесмосомами эпителиоцитов, содержит гликопротеины (ламинин) и протеогликаны (гепарансульфат), 2) плотная пластинка содержит коллаген IV, V, VII типов, имеет фибриллярную структуру. Тонкие якорные филаменты пересекают светлую и плотную пластинки, переходя в 3) ретикулярную пластинку, где якорные филаменты связываются с коллагеновыми (коллаген I и II типов) фибриллами соединительной ткани. В физиологических условиях базальная мембрана препятствует росту эпителия в сторону соединительной ткани, что нарушается при злокачественном росте, когда раковые клетки прорастают сквозь базальную мембрану в подлежащую соединительную ткань (инвазивный рост опухоли). Специфичные признаки эпителиев. Базальная исчерченность эпителиоцитов – термин, используемый для описания базального отдела некоторых клеток (например, в канальцах почки и части выводных протоков слюнных желез). На базальной поверхности много пальцевидных впячиваний плазмолеммы вглубь клетки. В цитоплазме базальной части клеток вокруг впячиваний плазмолеммы много митохондрий, которые обеспечивают энергозависимый процесс вывода молекул, ионов за пределы клетки. Апикальная поверхность эпителиоцитов может быть относительно гладкой или образует выпячивания. У некоторых эпителиоцитов на ней имеются специальные органеллы – микроворсинки и реснички. Микроворсинки максимально развиты в эпителиоцитах, участвующих в процессах всасывания (например, в тонкой кишке или канальцах проксимального отдела нефрона), где их совокупность называется щеточной (исчерченной) каемкой. Микрореснички – подвижные структуры, содержащие комплексы микротрубочек. Источники развития эпителиев. Эпителии развиваются из всех трех зародышевых листков, начиная с 3 – 4 недели эмбрионального развития человека. В зависимости от эмбрионального источника различают эпителий эктодермального, мезодермального и энтодермального происхождения. Классификация эпителиальной ткани I. Покровные эпителии. Морфологическая классификация покровного эпителия учитывает количество слоев клеток (одно- и многослойные), рядность однослойного эпителия (одно- и многорядные), форму клеток (для многослойных – поверхностного слоя): Однослойные эпителии 1.Однорядные: Плоский Кубический Призматический 2.Многорядный: Призматический реснитчатый Многослойные эпителии 1. Плоские Ороговевающий: Неороговевающий 2. Переходный. Железистые эпителии С экзокринной секрецией (многоклеточные, одноклеточные железы. С эндокринной секрецией. Связь морфологических признаков эпителиев с их функциональными особенностями служит ярким примером неразрывного единства структуры и функции тканей. Так, эпителии, выполняющие преимущественно защитную функцию, обладающие устойчивостью к действию механических, химических и микробных факторов, обычно имеют значительную толщину и являются многослойными. Чем выше нагрузки, тем толще эпителий и более значительно его ороговение. Другой стратегией защиты эпителия от микробов, частиц пыли или действия агрессивной среды служит выделение на его поверхность постоянно обновляемого протективного слоя слизи. Эпителии, обеспечивающие функцию активного всасывания, напротив, однослойные, на апикальной поверхности которых находятся микроворсинки, увеличивающие площадь поверхности. Функциональная классификация подразделяет эпителий только по функциональным признакам (всасывательный, осморегулирующий, мерцательный, железистый эпителий и другие типы). Строение разных видов покровного эпителия. Однослойные эпителии – все клетки располагаются на базальной мембране, причем ядра клеток однорядных эпителиев находятся на одном уровне, а ядра клеток многорядного эпителия находятся на разных уровнях, что создает эффект многорядности (и ложное впечатление многослойности). 1. Однослойный плоский эпителий образован уплощенными клетками полигональной формы с утолщением в области расположения дисковидного ядра. На свободной поверхности клетки имеются единичные микроворсинки. Примером такого типа является эпителий (мезотелий), покрывающий легкое (висцеральная плевра) и эпителий выстилающий изнутри грудную полость (париетальная плевра), а также париетальный и висцеральный листки брюшины, околосердечная сумка. 2. Однослойный кубический эпителий образован клетками, содержащими ядро сферической формы. Такой эпителий встречается в фолликулах щитовидной железы, в мелких протоках поджелудочной железы и желчных протоках, в почечных канальцах. 3. Однослойный призматический (цилиндрический) эпителий (рис.1) образован клетками с резко выраженной полярностью. Ядро эллипсовидной формы лежит вдоль длинной оси клетки и смещено к их базальной части, хорошо развитые органеллы неравномерно распределены по цитоплазме. На апикальной поверхности находятся микроворсинки, щеточная каемка. Этот вид эпителия характерен для среднего отдела пищеварительного канала и выстилает внутреннюю поверхность тонкой и толстой кишки, желудка, желчного пузыря, ряда крупных протоков поджелудочной железы и желчных протоков печени. Для этого вида эпителия характерны функции секреции и (или) всасывания. В эпителии тонкой кишки встречаются два основных типа дифференцированных клеток – призматические каемчатые, обеспечивающие пристеночное пищеварение, и бокаловидные, вырабатывающие слизь. Такое неодинаковое строение и функции клеток в однослойном эпителии называется горизонтальной анизоморфностью. 4. Многорядный реснитчатый (мерцательный) эпителий воздухоносных путей (рис.2) образован клетками нескольких типов: 1) низкие вставочные (базальные), 2) высокие вставочные (промежуточные), 3) реснитчатые (мерцательные), 4) бокаловидные. Низкие вставочные клетки являются камбиальными, своим широким основанием они прилежат к базальной мембране, а узкой апикальной частью не доходят до просвета. Бокаловидные клетки вырабатывают слизь, которая покрывает поверхность эпителия, перемещаясь по ней благодаря биению ресничек мерцательных клеток. Апикальные части этих клеток граничат с просветом органа. Многослойные эпителии – эпителии, в которых лишь клетки, образующие базальный слой, располагаются на базальной мембране. Клетки, входящие в состав остальных слоев, утрачивают с ней связь. Многослойным эпителиям свойственна вертикальная анизоморфность – неодинаковые морфологические свойства клеток различных слоев эпителиального пласта. В основу классификации многослойных эпителиев положена форма клеток поверхностного слоя. Поддержание целостности многослойных эпителиев обеспечивается регенерацией. Эпителиоциты непрерывно делятся в самом глубоком базальном слое за счет стволовых клеток, далее идет смещение в вышележащие слои. После дифференцировки происходит дегенерация и слущивание клеток с поверхности пласта. Процессы пролиферации и дифференцировки эпителиальных клеток регулируются рядом биологически активных веществ, часть которых выделяется клетками подлежащей соединительной ткани. Наиболее важным из них являются цитокины, в частности эпидермальный фактор роста; оказывают влияние гормоны, медиаторы и другие факторы. Дифференцировка эпителиоцитов сопровождатся изменением экспрессии синтезируемых ими цитокератинов, образующих промежуточные филаменты. Многослойные плоские эпителии в зависимости от наличия или отсутствия рогового слоя подразделяются на ороговевающие и неороговевающие. 1. Многослойный плоский ороговевающий эпителий (рис.3) образует наружный слой кожи - эпидермис, и покрывает некоторые участки слизистой оболочки полости рта. Он состоит из пяти слоев: Базальный слой (1) образован клетками кубической или призматической формы, лежащими на базальной мембране. Они способны к митотическому делению, поэтому за счет них происходит смена вышележащих слоев эпителия. Шиповатый слой (2) образован крупными клетками неправильной формы. В глубоких слоях могут встречаться делящиеся клетки. В базальном и шиповатом слоях хорошо развиты тонофибриллы (пучки тонофиламентов), а между клетками десмосомальные, плотные, щелевидные контакты. Зернистый слой (3) состоит из уплощенных клеток, в цитоплазме которых содержатся зерна кератогиалина – фибриллярного белка, который в процессе ороговения превращается в элеидин и кератин. Блестящий слой (4) выражен только в эпителии толстой кожи, покрывающей ладони и подошвы. Он представляет собой зону перехода от живых клеток зернистого слоя к чешуйкам рогового слоя, не обладающим признаками живых клеток. На гистологических препаратах он имеет вид узкой оксифильной гомогенной полоски и состоит из уплощенных клеток. В блестящем слое завершаются процессы ороговения, который заключается в превращении живых эпителиальных клеток в роговые чешуйки – механически прочные и химически устойчивые постклеточные структуры, образующие в совокупности роговой слой эпителия, который выполняет защитные функции. Хотя собственно формирование роговых чешуек происходит в наружных отделах зернистого слоя или в блестящем слое, синтез веществ, обеспечивающих ороговение, осуществляется уже в шиповатом слое. Роговой слой (5)наиболее поверхностный и имеет максимальную толщину в эпидермисе кожи в области ладоней и подошв. Он образован плоскими роговыми чешуйками с резко утолщенной плазмолеммой. Клетки не содержат ядра и органелл и заполнены сетью из толстых пучков кератиновых филаментов, погруженных в плотный матрикс. Роговые чешуйки в течение определенного времени сохраняют связи друг с другом и удерживаются в составе пластов благодаря частично сохраненным десмосомам, а также взаимному проникновению бороздок и гребешков, образующих ряды на поверхности соседних чешуек. В наружных частях рогового слоя десмосомы разрушаются, и роговые чешуйки слущиваются с поверхности эпителия. Большинство клеток многослойного ороговевающего эпителия относится к кератиноцитам. Дифферон кератиноцита включает в себя клетки всех слоев этого эпителия: базального, шиповатого, зернистого, блестящего, рогового. Кроме кератиноцитов в пласте присутствуют в небольшом количестве меланоциты и макрофаги. 2. Многослойный плоский неороговевающий эпителий покрывает поверхность роговицы глаза, слизистой оболочки полости рта, пищевода, влагалища. Он образован тремя слоями: 1) Базальный слой аналогичен по строению и функции соответствующему слою ороговевающего эпителия. 2) Шиповатый слой образован крупными полигональными клетками, которые по мере приближения к поверхностному слою уплощаются. Их цитоплазма заполняется многочисленными тонофиламентами, которые располагаются диффузно. В более наружных клетках этого слоя накапливается кератогиалин в виде мелких округлых гранул. 3) Поверхностный слой нерезко отделен от шиповатого. Содержание органелл снижено по сравнению с таковым в клетках шиповатого слоя, плазмолемма утолщена, ядро с плохо различимыми гранулами хроматина (пикнотическое). При десквамации клетки этого слоя постоянно удаляются с поверхности эпителия. Благодаря доступности и легкости получения материала многослойный плоский эпителий слизистой оболочки полости рта является удобным объектом для цитологических исследований. Клетки получают методом соскоба, мазка или отпечатка. Далее переносят на предметное стекло и готовят постоянный или временный цитологический препарат. Наибольшее распространение получило диагностическое цитологическое исследование этого эпителия с целью выявления генетического пола индивидуума; нарушения нормального течения процесса дифференцировки эпителия при развитии воспалительных, предопухолевых или опухолевых процессов ротовой полости. Клетки этого эпителия исследуют с целью определения уровня адаптации организма, влияния некоторых биологически активных веществ. Для этого, в частности, можно использовать метод прижизненного исследования с анализом микроэлектрофореза клеток, усовершенствованный на кафедре гистологии ИГМА. 3. Переходный эпителий (рис.4) – особый вид многослойного эпителия, который выстилает большую часть мочевыводящих путей. Он образован тремя слоями: 1) Базальный слой образован мелкими клетками, имеющими на срезе треугольную форму и своим широким основанием прилежат к базальной мембране. 2) Промежуточный слой состоит из удлиненных клеток, более узкой частью направленных к базальному слою и черепицеобразно накладывающихся друг на друга. 3) Поверхностный слой образован крупными одноядерными полиплоидными или двуядерными клетками, которые в наибольшей степени изменяют свою форму при растяжении эпителия (от округлой до плоской). Этому способствует формирование в апикальной части цитоплазмы этих клеток в состоянии покоя многочисленных инвагинаций плазмолеммы и особых дисковидных пузырьков – резервов плазмолеммы, которые встраиваются в нее по мере растяжения органа и клеток. Регенерация покровных эпителиев. Покровный эпителий, занимая пограничное положение, постоянно испытывает влияние внешней среды, поэтому эпителиальные клетки быстро изнашиваются и погибают. Восстановление эпителия – физиологическая регенерация – происходит путем митотического деления клеток. В однослойном эпителии большинство клеток способны к делению, а в многослойном такой способностью обладают только клетки базального и частично шиповатого слоев. Высокая способность эпителия к физиологической регенерации служит основой для быстрого восстановления его в патологических условиях - репаративная регенерация. Гистогенетическая классификация покровных эпителиев (по Н.Г. Хлопину) выделяет 5 основных типов эпителия, развивающихся в эмбриогенезе из различных тканевых зачатков: 1. 2. 3. 4. 5. 6. Гистогенетический тип эпителия Эпидермальный 7. Энтеродермальный Целонефродермальный Ангиодермальный Эпендимоглиальный Эмбриональные зачатки – источники развития эпителия эктодерма, прехордальная пластинка кишечная энтодерма целомическая выстилка, нефротом ангиобласт нервная трубка 1) Эпидермальный тип эпителия образуется из эктодермы, имеет многослойное или многорядное строение, приспособлен к выполнению прежде всего барьерной и защитной функции. 2) Энтеродермальный тип эпителия развивается из энтодермы, является по строению однослойным цилиндрическим, осуществляет процессы всасывания веществ. 3) Целонефродермальный тип эпителия имеет мезодермальное происхождение, по строению он однослойный, плоский или призматический, выполняет главным образом барьерную или экскреторную функцию. 4) Ангиодермальный тип включает в себя эндотелиальные клетки, имеющие мезенхимное происхождение. 5) Эпендимоглиальный тип представлен специальным видом ткани нейрального происхождения, выстилающим полости мозга и имеющий строение сходное с эпителием. Приведенная классификация не является общепринятой. Железистые эпителии. Железистые эпителиальные клетки могут располагаться одинаково, но чаще формируют железы. Железы, состоящие из железистого эпителия, выполняют секреторную функцию, вырабатывая и выделяя разнообразные вещества. Клетки железистого эпителия - гландулоциты или железистые клетки, процесс секреции в них протекает циклически, называется секреторным циклом и включает в себя пять стадий: 1. Фаза поглощения исходных веществ, служащих субстратами для синтеза секреторного продукта, обеспечивается высокой активностью транспортных механизмов, связанных с плазмолеммой базального полюса клетки, через который указанные вещества поступают из крови. 2. Фаза синтеза секрета связана с процессами транскрипции и трансляции, деятельностью грЭПС и агрЭПС, комплекса Гольджи. 3. Фаза созревания секрета связана с уменьшением воды в секрете и пополнением секрета новыми молекулами Гольджи 4. Фаза накопления синтезируемого продукта в цитоплазме железистых клеток обычно проявляется нарастанием содержания секреторных гранул. 5. Фаза выведения секрета может осуществляться несколькими путями (рис.5): мерокриновый - без нарушения целостности клетки, апокриновый – с разрушением апикальной части цитоплазмы, голокриновый - с полным нарушением целостности клетки. Строение гландулоцитов. Гландулоциты расположены на базальной мембране. Форма их весьма разнообразна и меняется в зависимости от фазы секреции. Для гландулоцитов характерна хорошо выраженная полярная дифференцировка, которая обусловлена направленностью секреторных процессов от базальной к апикальной части клеток (при внешней секреции). В связи с этим плазмолемма имеет различное строение на апикальных (микроворсинки), базальных (базальная мембрана) и латеральных (межклеточные контакты) поверхностях клеток. В апикальных отделах клеток обычно присутствуют секреторные гранулы, размер и строение которых зависят от химического состава секрета. В клетках, которые вырабатывают секреты белкового характера (например: пищеварительные ферменты), хорошо развита гранулярная эндоплазматическая сеть. В клетках, синтезируемых небелковые секреты (липиды, стероиды), выражена агранулярная эндоплазматическая сеть. Комплекс Гольджи хорошо развит и участвует в секреторном цикле. Митохондрии многочисленны, накапливаются в местах наибольшей активности клеток. В ряде желез в цитоплазме гландулоцитов находятся внутриклеточные секреторные канальцы – глубокие впячивания цитолеммы, стенки которых покрыты микроворсинками (например, в париетальных клетках желез желудка). Железы делятся на две группы: железы внутренней секреции, или эндокринные, и железы внешней секреции, или экзокринные. Эндокринные железы (железы внутренней секреции) продуцируют гормоны – вещества, обладающие высокой биологической активностью, которые выводятся из клетки через базальный полюс. Выводные протоки в таких железах отсутствуют, секрет в последующем поступает через капилляры в кровь. Экзокринные железы вырабатывают секреты, выделяющиеся во внешнюю среду, состоят из концевых отделов и выводных протоков. 1) концевые (секреторные) отделы (рис.6) состоят из железистых клеток, которые продуцируют секрет. В некоторых железах, образованных эпителиями эпидермального типа (например, потовых, молочных, слюнных), концевые отделы помимо железистых клеток содержат миоэпителиальные клетки – видоизмененные эпителиоциты с развитым сократительным аппаратом. Миоэпителиальные клетки своими отростками охватывают снаружи железистые клетки и, сокращаясь, способствуют выделению секрета из концевого отдела. 2) выводные протоки связывают концевые отделы с покровными эпителиями и обеспечивают выделение синтезированных веществ на поверхность тела или в полость органов. Разделение на концевые отделы и выводные протоки затруднено в некоторых железах (например, желудка, матки), так как все участки этих простых желез способны к секреции. Классификация экзокринных желез. I. Морфологическая классификация экзокринных желез основана на структурном анализе их концевых отделов и выводных протоков. В зависимости от формы секреторного (концевого) отдела различают альвеолярные, трубчатые и смешанные (альвеолярно-трубчатые) железы; В зависимости от ветвления секреторного отдела различают разветвленные и неразветвленные железы. Ветвление выводного протока определяет деление желез на простые (проток не ветвится) и сложные (проток ветвится). II. По химическому составу вырабатываемого секрета различают серозные (белковые), слизистые, смешанные (белково-слизистые), липидные и др. железы. III. По механизму (способу) выведения секрета экзокринные железы делят на апокриновые (молочная железа), голокриновые (сальная железа) и мерокриновые (большинство желез). Примеры классификации желез. Классификационная характеристика сальной железы кожи: 1) простая альвеолярная железа с разветвленными концевыми отделами, 2) липидная – по химическому составу секрета, 3) голокриновая – по способу выведения секрета. Характеристика лактирующей (вырабатывающей секрет) молочной железы: 1) сложная разветвленная альвеолярно-трубчатая железа, 2) со смешанным секретом, 3) апокриновая. Регенерация. В железах в связи с их секреторной деятельностью постоянно происходят процессы физиологической регенерации. В мерокриновых и апокриновых железах, в которых находятся долгоживущие клетки, восстановление исходного состояния гландулоцитов после выделения из них секрета происходит путем внутриклеточной регенерации. В голокриновых железах восстановление осуществляется за счет размножения камбиальных (стволовых) клеток. Вновь образовавшиеся из них клетки затем путем дифференцировки превращаются в железистые клетки – это клеточная регенерация. Вопросы для самоконтроля: 1. Какие общие закономерности строения имеет покровный эпителий? 2. Из каких зародышевых листков образуются в эмбриогенезе различные виды эпителия? 3. Морфологическая классификация покровных эпителиев. 4. Назовите виды, опишите структуру и функции межклеточных контактов. 5. Назовите виды многослойного эпителия. Дайте характеристику дифферона кератиноцита. 6. В чем заключается отличие многослойного и многорядного эпителиев? 7. Назовите виды однослойных эпителиев. 8. В чем сущность физиологической регенерации? 9. Каковы основные фазы секреторного процесса? 10. Из каких отделов состоят экзокринные железы? В чем их отличие? 11. Назовите способы выведения секреторных веществ из гландулоцитов. 12. Какие органоиды и включения наиболее характерны для цитоплазмы гландулоцитов при выработке белкового секрета, липидного секрета? 13. На каких особенностях строения и функционирования экзокринных желез основана их классификация? Приведите примеры. 14. В чем принципиальные отличия в строении и функции экзокринных и эндокринных желез. Ситуационные задачи 1.В эксперименте у зародыша нарушен морфогенез энтодермы. На развитии эпителия какой системы ( и какого отдела) это скажется? 2.Одной из функций кишечника является всасывание. Какой вид эпителия соответствует этой функции? 3.На небольшом участке кожи удалены все слои эпидермиса. Как осуществляется регенерация? 4.На электронограмме эпителиоцитов в апикальном конце видны вакуоли, заполненные секретом и окруженные мембраной, которая соприкасается с апикальной плазмолеммой, целостность которой не нарушена. Какой тип сереции зарегистрирован в данном случае? 5.Какие клетки способствуют выведению секрета из концевых отделов слюнных желез? 2.ТЕМА: ТКАНИ ВНУТРЕННЕЙ СРЕДЫ В ходе эмбрионального развития человек вначале является одноклеточным организмом (зигота), затем многоклеточным бестканевым (бластула, гаструла). Формирование тканей (гистогенез) происходит параллельно с развитием органов (органогенез). Наиболее рано включаются в жизнедеятельность эмбриона эпителиальные ткани и ткани, образующиеся из мезенхимы. Мезенхимное происхождение имеют несколько разновидностей тканей (кровь, соединительные, хрящевые, костные ткани), которые в соответствии с принятой в настоящее время классификацией входят в группу соединительных тканей. Указанный термин не в полной мере отражает предназначение этих тканей. Трудно ставить знак равенства между волокнистой соединительной тканью, которая действительно соединяет отдельные части органа в единое целое и кровью или кроветворными тканями имеющим другую функциональную направленность. Более оправдано название предложенное исследователем тканей академиком А.А. Заварзиным «Ткани внутренней среды». Эти ткани А.А. Заварзин подразделял на трофические (кровь, миелоидная, лимфоидная ткани); опорно-трофические (рыхлая соединительная, ретикулярная ткани); механические (плотные соединительные, хрящевые, костные ткани). Название «Ткани внутренней среды» отражает их участие в поддержании постоянства внутренней среды человека (гомеостаз). Многие параметры внутренней среды организма не только постоянны (динамическое равновесие), но и являются жизнеопределяющими. Представляет интерес, что стволовые клетки и для крови и для соединительной ткани образуются в красном костном мозге. Это подчеркивает единство тканей внутренней среды. Характерно содружественное участие этих тканей в воспалении, иммунных реакциях. Классификация тканей внутренней среды человека Ткани внутренней среды I. Кроветворные ткани и кровь II. Эндотелий. III. Соединительные ткани: 1. Волокнистые соединительные ткани: а) Рыхлая волокнистая соединительная ткань. б) Плотные волокнистые соединительные ткани: *оформленная * неоформленная. 2. Специализированные соединительные ткани: * Жировая. * Ретикулярная. * Слизистая. Мезенхима. Источником всех тканей внутренней среды организма человека в эмбриогенезе является мезенхима. Она же является продуцентом соединительной основы внезародышевых органов (желточного мешка, хориона, амниона, аллантоиса). Мезенхимные клетки могут быть у зародыша человека в виде редко и плотно расположенных клеточных скоплений. Они способны изменять свою форму. В большинстве своем это многоотросчатые клетки с крупным овальным ядром. В ходе преобразований и последующей дифференцировки мезенхимные клетки образуют стволовые клетки для органов кроветворения, для эндотелия кровеносных сосудов, для волокнистой соединительной, ретикулярной, скелетных и жировой тканей. После рождения именно эти клетки обеспечивают обновление указанных тканей. 2.1 ТЕМА: КРОВЬ. КРОВЕТВОРЕНИЕ. ЛИМФА. Цели занятия: 1.Выявить морфофункциональную характеристику крови как ткани. 2.Научиться различать в мазках крови форменные элементы. 3.Научиться подсчитывать гемограмму и лейкоцитарную формулу крови. Кровь - своеобразная ткань внутренней среды с жидким межклеточным веществом (плазмой), в которой находятся разнообразные клетки и постклеточные элементы. Эта ткань имеет мезенхимное происхождение. Кровь–часть сложной системы (система крови), которая включает в себя органы кроветворения (гемопоэза) и иммуногенеза и с этой системой связаны органы, синтезирующие белки плазмы и органы, обеспечивающие нервную и гуморальную регуляцию качественного и количественного состава крови и структуры, обеспечивающие разрушение старых клеток крови. Общий объем крови у человека составляет 6-8% от массы его тела. В среднем – 4 - 6 л. До 1 л крови находится в депо, преимущественно в селезенке. Нахождение крови в жидком состоянии и циркуляция в сосудистой системе обеспечивает выполнение кровью ее функций: 1. транспортная - наиболее универсальная функция, связанная с переносом различных веществ: -перенос газов в растворенном (углекислый газ) и связанном (кислород) состоянии (дыхательная функция). - перенос питательных веществ от места всасывания и запасания к другим тканям (трофическая функция). - перенос метаболитов от тканей (метаболическая) и их выделение из организма. В почках образуется моча как фильтрат плазмы крови. - перенос гормонов, других биологически активных веществ (регуляторная) - распределение тепла между органами (терморегуляторная). 2. гомеостатическая –обеспечение постоянства внутренней среды (кислотно-щелочное, осмотическое равновесие, водный баланс тканевых жидкостей) 3. защитная – нейтрализация антигенов специфическими и неспецифическими механизмами. Кровь как ткань включает в себя форменные элементы (клетки и постклеточные структуры) и плазму (межклеточное вещество). Соотношение этих двух компонентов различно в разные возрастные периоды и при разных физиологических состояниях и называется гематокритом (греч. «разделяю кровь»). Форменные элементы крови. К форменным элементам крови относятся эритроциты, лейкоциты, тромбоциты. Концентрацию форменных элементов при лабораторном анализе определяют в расчете на 1 мкл или 1 л. Совокупность результатов образует гемограмму. Гемограмма взрослого человека в норме (средние показатели): эритроциты- 4-5,5 х 106/мкл (4-5,5 х 1012/л) лейкоциты- 4-8 х 103/мкл (4-8 х 109/л) тромбоциты- 200-400 х 103/мкл (200 –400 х 109/л) Эритроциты (греч. «красные клетки») – самые многочисленные клетки крови, утратившие в процессе дифференцировки ядро и практически все органеллы. Большинство эритроцитов (75-85 %) имеют форму двояковогнутого диска. Благодаря этому достигается: 1. Большая площадь поверхности (в 1,5 раза больше, чем у клетки сферической формы такого же объема). 2. Оптимальное диффузное расстояние между цитолеммой и наиболее удаленной частью цитоплазмы, что необходимо для эффективного газообмена. 3. Способность к обратимой деформации при прохождении через капилляры, некоторые участки которых почти в 2 раза меньше в диаметре, чем эритроцит. Поддержание формы эритроцитов обеспечивается осмотическим равновесием (работой ионных насосов плазмолеммы), элементами цитоскелета. Нарушение структуры того или иного компонента приводит к изменению формы эритроцита, ====что может свидетельствовать о старении клетки или патологии. Наличие в крови эритроцитов различной формы называется пойкилоцитоз (от греч. разнообразный). Могут встречаться сфероциты, эхиноциты (с выростами), куполообразные, гребневидные клетки и другие формы. Размеры эритроцитов также могут варьировать. Это явление отражает понятие анизоцитоза (от греч.неравные клетки). Различают макроциты (более 9 мкм), нормоциты (7-8 мкм), микроциты (менее 6 мкм). Форма эритроцитов, как уже говорилось, поддерживается в основном благодаря взаимодействию мембранных компонентов и цитоскелета. Плазмолемма эритроцита является наиболее изученной из биологических мембран. Ее толщина составляет 20 нм. На ее поверхности содержится огромное количество рецепторов (более 300) к иммуноглобулинам, компонентам комплемента, белкам плазмы, гормонам, биологически активным веществам. Также на поверхности мембраны находятся антигены Rh и детерминаты группы крови. Среди многочисленных белков мембраны особое значение имеют интегральные белки гликофорин (содержит аглютиногены, обусловливающие группу крови) и белок полосы 3 (участвует в транспорте анионов хлора и гидрокарбоната, глюкозы). Изменение его конфигурации является маркером старения эритроцита. Узнавание такого белка макрофагами селезенки приводит к уничтожению старого эритроцита). С трансмембранными белками связаны элементы цитоскелета. Главный компонент – белок спектрин. Он состоит из двух цепей (альфа и бета). Цепи скручены и образуют гибкую сеть на внутренней поверхности мембраны. Между собой цепи связаны с помощью актина и белка полосы 4.1. Белок полосы 4.1 также связывает спектрин с гликофорином. С белком полосы 3 связь образует анкирин. В покое цепи спектрина скручены равномерно. При деформации они в одном участке раскручиваются, а в другом скручиваются еще сильнее. Благодаря такому устройству цитоскелета, эритроцит обладает гибкостью и способен обратимо деформироваться в мелких сосудах. Цитоплазма эритроцитов оксифильна и обладает высокой электронной плотностью. На 66% она состоит из воды, 33% приходится на гемоглобин, 1% составляет белки (в основном ферменты, их более140), липиды, глюкоза, АТФ. Гемоглобин располагается в виде гранул 4-5 нм. В разные периоды онтогенеза в эритроцитах можно наблюдать различные типы гемоглобина: 1.Эмбриональный гемоглобин обнаруживается у 19-дневного зародыша и сохраняется в течение 3-6 месяцев эмбриогенеза. 2.Фетальный гемоглобин составляет 95% гемоглобина плода и сохраняется после рождения до 8 месяцев, при этом его количество снижается. 3.Дефинитивный (окончательный) гемоглобин сменяет фетальный и составляет 98% от гемоглобина эритроцитов взрослого. Типы гемоглобина различаются строением цепей глобина (белковой части) и уровнем сродства к кислороду. Зрелые эритроциты не содержат ядра, синтетического и секреторного аппарата, митохондрий. Содержат отдельные лизосомы и элементы цитоскелета. Для них характерен низкий уровень обмена веществ, что и обеспечивает длительность жизни (100- 120 дней). Энергию получают путем гликолиза и прямого окисления глюкозы (пентозофосфатный шунт). Свои функции эритроциты выполняют в сосудистом русле, которое в норме не покидают. Функции эритроцитов: 1.Перенос газов (преимущественно кислорода) с помощью гемоглобина. 2.Перенос других веществ на своей поверхности (иммуноглобулинов, гормонов, биологически активных веществ и др.) Кроме зрелых эритроцитов в периферической крови можно встретить молодые формы – ретикулоциты. Созревание последних происходит в течение двух суток после выхода из красного костного мозга и заключается в завершении синтеза гемоглобина и разрушении остатков органелл, которые выявляются в ретикулоцитах при специфической окраске. В норме ретикулоциты составляют до 1% всех эритроцитов. Лейкоциты (от греч. белые клетки) в отличие от эритроцитов многие лейкоциты лишь циркулируют в крови и доставляются к месту функционирования. Свои функции большинство лейкоцитов выполняют в тканях, мигрируя через стенки микрососудов, двигаясь с образованием ложноножек (псевдоподий). Лейкоциты - это полиморфная группа клеток. Их классификация основана на ряде признаков, основным из которых является присутствие в цитоплазме специфических гранул. Лейкоциты (100%- 4-8 х 109/л) Гранулоциты Эозинофил Нейтрофи ы лы Процентное 1-5 65-75 содержание от юные-0,5 общего числа палочкоя лейкоцитов дерные3,5 сегментоя дерные60-65 Диаметр клетки 12-17 10-12 (мкм) Время пребывания 3-8ч 8-12 ч в кровотоке Продолжительност 8-14 суток 8 суток ь жизни Базофил ы 0-1 агранулоциты Моноцит лимфоци ы ты 6-8 20-45 10-12 18-20 4,5-10 6ч1сутки ? 8ч-4суток Часыгоды Часыгоды ? Концентрация лейкоцитов в крови – важный диагностический показатель. Имеет значение не только их общее число, но и количество клеток отдельных субпопуляций. Поэтому при цитологическом исследовании крови лейкоциты подсчитывают дифференцированно. Результаты выражаются в процентах от общего количества лейкоцитов. Процентное соотношение лейкоцитов называется лейкоцитарной формулой. Дифференцировка лейкоцитов производится на основе их морфологических различий. Нейтрофилы - самая многочисленная группа лейкоцитов (рис.7), представленная клетками с разным уровнем дифференцировки. На оптическом уровне их в основном отличают по форме ядра. Юные нейтрофилы – имеют ядро бобовидной формы со светлым содержимым. Палочкоядерные нейтрофилы– имеют ядро в виде палочки или подковы, с большим содержанием гетерохроматина. Сегментоядерные нейтрофилынаиболее зрелые формы. Они имеют дольчатое ядро, состоящее из 2-5 сегментов, соединенных перетяжками. У 3% нейтрофилов женщин выявляется дополнительный сегмент в виде барабанной палочки (тельце Барра). Считается, что это неактивная Ххромосома. В процессе дифференцировки нейтрофилов, кроме сегментации ядра происходит накопление гликогена, уменьшение органелл синтетического аппарата и накопление специфических гранул. Для зрелого нейтрофила характерна следующая структура цитоплазмы: *Содержат несколько митохондрий, но значительное количество гранул гликогена, т.к. энергия получается в основном путем гликолиза. Это объясняется тем, что клеткам приходится работать в поврежденных тканях бедных кислородом. *Минимальное количество органелл синтетического аппарата, поэтому после единственной вспышки активности нейтрофил погибает, т.к. теряет возможность восстановить органеллы. *Хорошо развитый цитоскелет, состоящий из 12-20 микротрубочек, многочисленных актиновых филаментов, расположенных преимущественно в периферической части цитоплазмы. В состоянии покоя актин находится в цитоплазме в виде отдельных глобул. При активизации клетки он быстро полимеризуется с образованием филаментов, участвующих в движении клетки с образованием псевдоподий. *Гранулы: а)первичные (азурофильные, неспецифичные) часто рассматриваются как лизосомы, но в отличие от них содержат большее количество антимикробных веществ. Гранулы содержат катепсин- антимикробное вещество с широким спектром действия; лизоцим - разрушает клеточную стенку бактерий; эластазу и коллагеназу, которые разрушают эластин и коллаген тканей нейтрофилов; пероксидазу -значительно усиливает активность антимикробных ферментов и т.д. б) вторичные (специфические). Их количество увеличивается по мере созревания. Они содержат адгезивные белки, обеспечивающие адгезию нейтрофилов на эндотелиоцитах, лактоферрин - связывает факторы роста бактерий, коллагеназу, лизоцим. Нейтрофилы передвигаются в тканях путем хемотаксиса, по градиенту концентрации веществ, образующихся при расщеплении микроорганизмов. Факторами хемотаксиса также могут служить вещества, выделенные активированными нейтрофилами и другими клетками (тучными, макрофагами). Функции нейтрофилов: 1)Повреждающее воздействие на микробы (катепсин, лизоцим и др.). 2)Фагоцитоз микроорганизмов, поврежденных клеток - одна из основных функций нейтрофилов, поэтому их еще называют микрофагами. 3) Участие в специфических иммунных реакциях. Вырабатывая биологически активные вещества (цитокины), нейтрофилы регулируют кооперацию клеток при иммунном ответе. Эозинофилы - из кровотока мигрируют в основном в рыхлую соединительную ткань органов, контактирующие с внешней средой (слизистая дыхательных, мочеполовых путей, кишечника). В рыхлой соединительной ткани эозинофилов в 200 раз больше, чем в крови. Эозинофил (рис.8) содержит палочковидное или сегментированное (обычно два сегмента с перемычкой) ядро, хорошо развитую гранулярную ЭПС, большое количество рибосом, полисом и гранул гликогена и несколько митохондрий. В цитоплазме содержатся гранулы двух типов: а) неспецифические (азурофильные) и б) специфические. Первые аналогичны лизосомам. По мере дифференцировки их количество уменьшается. Специфические - овоидной формы, содержат кристаллоид, образованный главным основным белком (обусловливает эозинофилию). Обладает мощным противогельминтозным, противопротозойным и антимикробным действием. В аморфном матриксе, расположенном по периферии от кристаллоида содержатся фермент гистаминаза, иннактивирующий гистамин, а также коллагеназа и другие биологически активные вещества. Эозинофилы способны к хемотаксису. Факторами хемотаксиса являются паразиты и продукты их жизнедеятельности, вещества, выделяемые лимфоцитами, макрофагами, тромбоцитами, эндотелиоцитами. Особенно эффективен гистамин тучных клеток. Функции эозинофилов: 1)Уничтожение микроорганизмов и в особенности паразитов (гельминтов и простейших) нефагоцитарным путем 2)Способны к фагоцитозу, но в меньшей степени, чем нейтрофилы. 3)Ограничивают область иммунной реакции, создавая препятствие распространению антигенам и медиаторам воспаления, разрушая их. Базофилы – клетки аналогичные тучным клеткам, но не идентичные им (различаются по соотношению и составу гранул). Это клетки со слабооксифильной цитоплазмой и S - образным трехдольным ядром ( рис.9). В цитоплазме содержатся все органеллы в умеренном количестве, липидные капли, гликоген и два типа гранул: 1) Азурофильные - аналогичны лизосомам. 2) Специфичные – плотные гранулы округлой или овальной формы, содеращие гепарин антикоагулянт, гистамин - вещество, расширяющее сосуды, увеличивающее их проницаемость, а также факторы хемотаксиса нейтрофилов и эозинофилов и другие факторы. Функции базофилов: выделяя локально в большом количестве биологически активные вещества гранул, привлекают ряд других клеток в защитные реакции организма (в первую очередь эозинофилов при аллергических и воспалительных реакциях). Моноциты. Фактически это незрелые клетки, находящиеся на пути из места образования (красный костный мозг) в место функционирования (органы и ткани). В органах и тканях происходит их окончательная дифференцировка. Совокупность всех потомков моноцитов крови в тканях называется системой мононуклеарных фагоцитов. К ней относятся: дендритные клетки кроветворных органов, макрофаги (гистиоциты) соединительной ткани, альвеолярные макрофаги легких, селезенки и красного костного мозга, макрофаги (клетки Купфера) печени, остеокласты костной ткани, перитонеальные макрофаги, макрофаги (клетки Хофбауэра) плаценты и микроглия нервной ткани. Моноцит (рис.10) имеет крупное, эксцентрично расположенное бледное ядро с выемкой, которая увеличивается по мере созревания клетки. В слабобазофильной цитоплазме содержится большое количество лизосом и вакуолей, рибосом и полирибосом, умеренное количество цистерн ЭПС и хорошо развитый аппарат Гольджи, цитоскелет, мелкие удлиненные митохондрии. Имеются центриоли. Основной функцией моноцитов является фагоцитоз и эндоцитоз. Кроме того, они участвуют в кооперации клеток при иммунном ответе, представляя антиген иммунокомпетентным клеткам, а также выделяют биологически активные вещества, регулирующие гемопоэз и хемотаксис других клеток крови. Лимфоциты – большая группа морфологически сходных, но функционально различных клеток. Эти клетки играют центральную роль в иммунологических реакциях. Способны выходить из крови в ткани, затем снова возвращаться в кровь через лимфу. Такое явление называется рециркуляцией. В крови находится лишь 2% лимфоцитов, 98% рассредоточено по другим органам и тканям. Это округлые клетки с крупным ядром, занимающим до 90% объема клетки. Цитоплазма слабобазофильна. Все органеллы представлены в умеренном количестве, кроме цитоскелета, который хорошо развит. По размерам лимфоциты делятся на малые (6-7мкм), их 80-90% от общего количества лимфоцитов крови; средние (8-9 мкм) - 10% и большие (10-18 мкм) – в норме в крови отсутствуют. Более существенна функциональная классификация лимфоцитов. Согласно ей лимфоциты подразделяют на Т- и В-клетки. Они различаются: 1. Местом дифференцировки. Т-лимфоциты дифференцируются в тимусе. В-лимфоциты в лимфоидной ткани других органов. 2. Выполняемыми функциями. Обеспечивают различные типы иммунитета. Тлимфоциты - преимущественно клеточный, а В-лимфоциты - гуморальный иммунитет. Функционально Т- и В-клетки делят на субпопуляции. Среди Т-лимфоцитов выделяют Т х (хелперы)- активируют эффекторные клетки, Тк (киллеры) – эффекторные цитотоксические клетки, Тс (супрессоры)- подавляют иммунный ответ, Т-памяти. В - лимфоциты дифференцируются в плазматические клетки, вырабатывающие иммуноглобулины (антитела) и в клетки памяти, несущие информацию о встрече с какимлибо антигеном. 3. Содержанием в крови. Т - лимфоцитов – 70-80%, В-лимфоцитов - 10-20%. Тромбоциты - являются фрагментами цитоплазмы мегакариоцитов красного костного мозга, поэтому правильнее называть их кровянные пластинки.. В мазке крови агрегируются, поэтому выявляются там в виде скоплений. Тромбоциты - это овальные, двояковыпуклые тельца с небольшими отростками (рис.11). Внутреннее содержимое состоит из двух частей. Центральная часть – грануломер, содержит азурофильные зерна, наружная - гиаломер, имеет гомогенную консистенцию и бледно-голубую окраску. Плазмолемма покрыта слоем гликокаликса. Он состоит из многочисленных рецепторов, обусловливающих прикрепление тромбоцита к эндотелию (адгезию) и склеивание тромбоцитов друг с другом (агрегацию). Здесь есть рецепторы к адреналину, коллагену и другим веществам. Гиаломер представляет собой однородную, тонкозернистую структуру с микротрубочками и филаментами по периферии. Микротрубочки формируют краевое кольцо – жесткий каркас тромбоцита. Филаменты, представленные в основном актином, в виде коротких нитей располагаются по всей цитоплазме, в гиаломере они концентрируются между плазмолеммой и краевым кольцом и формируют подмембранный аппарат. Последний формирует выпячивания при движении и агрегации тромбоцитов. В грануломере содержатся 1-2 митохондрии, гранулы гликогена в виде агрегатов, единичные рибосомы и гранулы нескольких типов: 1. Азурофильные гранулы содержат вещества, участвующие в свертывании (фибронектин, фибриноген), фактор роста тромбоцитов и ряд других биологически активных веществ. 2. Гранулы с плотным матриксом, содержащим АТФ, ионы кальция, магния, гистамин, серотонин. 3. Гранулы, содержащие гидролитические ферменты, соответствуют лизосомам. Функции тромбоцитов: 1. Восстановление целостности сосудистой стенки при повреждении (первичный гемостаз). 2. Свертывание крови, в совместной реакции с эндотелием и плазмой крови (вторичный гемостаз) путем прилипания агрегатов тромбоцитов к месту повреждения. 4. Участие в иммунных реакциях ( вырабатывают факторы хемотаксиса клеток иммунной системы). Плазма крови. Клетки крови переносятся жидким межклеточным веществомплазмой. На 90% она состоит из воды, 9% составляют органические вещества, главным образом белки (более 200 видов: альбумины, глобулины, коагулянты и антикоагулянты, белки системы комплемента), 1%-неорганические вещества. РН составляет 7,36 (стабильность РН обеспечивает буферная система плазмы крови). Гемопоэз (кроветворение) - процесс образования крови, включает в себя эритропоэз (образование эритроцитов), гранулоцитопоэз (образование гранулярных лейкоцитов), моноцитопоэз (образование моноцитов), тромбоцитопоэз (образование кровяных пластинок), лимфоцитопоэз (образование лимфоцитов и иммуноцитов). Выделяют эмбриональный гемопоэз, обеспечивающий гистогенез крови (образование крови как ткани) и постэмбриональный гемопоэз–процесс физиологической регенерации крови. Эмбриональный гемопоэз включает три этапа: 1. Мезобластический. На этом этапе происходит образование первой генерации стволовых клеток крови (СКК). Процесс происходит интраваскулярно (внутри сосудов) в мезенхиме желточного мешка (внезародышевый провизорный орган) на 3-10 неделе внутриутробного периода. Из желточного мешка СКК мигрируют в другие кроветворные органы. 2. Гепатолиенальный (кроветворение в печени и селезенке) этап протекает, начиная с 5-6 недели, достигая максимальной активности на втором месяце, когда на кроветворение на 80% обеспечивается печенью, а на 20% селезенкой. В этих органах дифференцировка клеток крови из СКК протекает экстраваскулярно (вне сосуда). В печени образуются преимущественно эритроциты, гранулоциты, кровяные пластинки. В селезенке первоначально образуются все виды форменных элементов крови, а во второй половине внутриутробного развития начинает преобладать лимфоцитопоэз. 3. Медуллярный (тимо-медулло-лимфоидный)- образование форменных элементов крови в тимусе, лимфоидной ткани и красном костном мозге (ККМ), начинается на 10-ой неделе внутриутробного развития. В тимусе образуются Т-лимфоциты с последующим расселением их в лимфоидные органы. В красном костном мозге (ККМ) СКК дают начало всем форменным элементам, формируя кроветворные (гемопоэтические) островки. Совокупность СКК и гемопоэтических островков составляет паренхиму ККМ. Гемопоэз постепенно нарастает к рождению и ККМ становится центральным органом кроветворения. Кроветворной тканью ККМ является миелоидная ткань (от греч. красный мозг). Она содержит стволовые кроветворные клетки и является местом образования эритроцитов, гранулоцитов, моноцитов, тромбоцитов, В-лимфоцитов, предшественников Т-лимфоцитов, NK- клеток. Лимфоидная ткань располагается в органах иммунной системы (в тимусе, селезенке, лимфатических узлах, миндалинах, пейеровых бляшках, червеобразном отростке и многочисленных лимфоидных образованиях, имеющихся в стенках органов различных систем). В ней происходит образование Т- и В-лимфоцитов, которые взаимодействуя между собой, а также с макрофагами, дендритными и другими клетками, обеспечивают развитие и течение иммунных реакций. Регуляция гемопоэза осуществляется гемопоэтическими факторами роста (гемопоэтинами), которые вырабатываются стромальными элементами кроветворных органов. Они продуцируются ретикулярными клетками, эпителиальными клетками тимуса, макрофагами, Т-лимфоцитами, эндотелиальными клетками, а также клетками, расположенными вне кроветворных тканей (например, эритропоэтин вырабатывается клетками почек и печени). Гемопоэтины оказывают влияние в низких концентрациях, связываясь со специфическими рецепторами на плазмолемме развивающихся клеток крови. Каждый этап развития конкретной линии клеток требует присутствия определенной концентрации гемопоэтинов. Отдельный гемопоэтический фактор может оказывать влияние на один или несколько типов развивающихся клеток. Гемопоэтические классы клеток. (см. табл.1) В настоящее время доказано, что общим источником развития всех форменных элементов крови служат СКК, которые образуют самоподдерживающуюся популяцию полипотентных клеток. Это положение впервые сформулировано профессором А.А.Максимовым в унитарной теории кроветворения, которой противопоставляли ряд других теорий, допускавших развитие различных форменных элементов из двух, трех или большего числа родоначальных клеток (дуалистическая, полифилитическая теории). Полипотентные стволовые клетки образуют первый класс полипотентных клеток. Всего же на основании способности к самообновлению, клеточному делению и образованию форменных элементов различных типов выделяют шесть классов кроветворных клеток. Три первые класса объединяют в группу так называемых морфологически нераспознаваемых клеток, поскольку они фенотипически идентичны и похожи на малые лимфоциты. Их диаметр 8-10 мкм. Имеют круглую или неправильную форму, круглое крупное ядро с1-2 ядрышками. Цитоплазма узким ободком окружает ядро. Точная идентификация клеток может быть произведена только иммуноцитохимически по антигенам на клеточной поверхности. 1. Стволовые клетки крови (СКК). 2. Полустволовые клетки (их также называют колониеобразующие единицы (КОЕ)). Частично детерминированные, полипотентные клетки-предшественники, образующиеся при делении СКК. Их два типа: * клетки - предшественники для эритроцитов, гранулоцитов, моноцитов и тромбоцитов (КОЕ-ГЭММ) *клетки предшественники для лимфоцитов (КОЕ-Л) 3. Унипотентные (коммитированные) клетки – могут развиваться только в напрвлении определенного (одного) вида форменных элементов (за исключением КОЕГМ, которая дает два вида клеток). Эти клетки в отличии от клеток 1 и 2 классов являются поэтинчувствительными. Существуют следующие виды унипотентных клетокпредшественниц: *клетки, образующие гранулоциты (нейтрофилы - КОЕ-Г) и моноциты (КОЕ Мо). *клетки-предшественники для эритроцитов (КОЕ-Э). *для кровяных пластинок (КОЕ-Мег) *клетки – предшественники базофилов (КОЕ-Баз) *клетки – предшественники эозинофилов (КОЕ-Эо) *клетки – предшественники Т- и В- лимфоцитов (про В- лимфоциты, протимоциты) Следующие гемопоэтические классы клеток (4,5,6) объединяют в группу морфологически распознаваемых клеток- предшественников. 4. Бластные формы (бласты). Эти клетки обладают высокой митотической активностью, но не являются самоподдерживающейся популяцией. Бласты разных гемопоэтических рядов обладают минимальными различиями, но их можно идентифицировать при помощи гистологической окраски. Как правило, это крупные клетки с базофильной цитоплазмой, с крупным светлым ядром, содержащим 1-2 ядрышка. 5. Созревающие (дифференцирующиеся) клетки подвергаются структурной и функциональной дифференцировке, образуя соответствующий специфичный вид форменных элементов. В ходе дифференцировки клетки утрачивают способность к делению (исключение составляют моноциты и лимфоциты). 6. Зрелые (дифференцированные) форменные элементы переходят из ККМ в кровь и циркулируют в ней. Не способны к делению (исключение: моноциты и лимфоциты). Особенности морфологически распознаваемых клеток - предшественниц различных гемопоэтических рядов 4 класс Э Р И Т Р О Ц И Т О П О Э З 5 класс -Базофильный эритробласт 12-16 мкм, ядро с умеренно конденсированным хроматином и ядрышком. Цитоплазма базофильная, поскольку содержит большое количество полисом, активно синтезирующих гемоглобин. Активно делится. -Полихроматофильный эритробласт 10-12 мкм, компактное ядро, ядрышки отсутствуют, хроматин в виде глыбок. Цитоплазма окрашивается полихромно (воспринимает основные красители, вследствие присутствия полисом и кислые, благодаря наличию гемоглобина). Способность к делкнию сохраняется. -Оксифильный эритробласт (нормобласт) 8-10 мкм, ядро мелкое пикнотическое, цитоплазма оксифильная, богатая гемоглобином, органеллы почти полностью отсутствуют. Способность к делению утрачивается. Г -Промиелоцит 16-24 мкм, круглое светлое ядро с 1-2 Миелобл Р ядрышками. Слабобазафильная цитоплазма с А аст большим количеством послисом, цистерн Н гранулярной ЭПС. Накапливаются азурофильные У гранулы и появляются специфичные. Активно Л делятся. О -Миелоцит 10-16 мкм, бобовидное ядро с крупными Ц глыбками гетерохроматина, расположено И эксцентрично. В цитоплазме возрастает количество Т специфичеких гранул. Митотически активен. О -Метамиелоцит 8-12 мкм, с более плотным ядром, П форма которого изменяется до палочкоядерного и О начинает сегментироваться. Хроматин Э конденсируется. В цитоплазме уменьшается З количество гранулярной ЭПС, рибосом, комплекса Гольджи. Увеличивается количество элементов цитоскелета, обеспечивающего высокую подвижность клетки. Не способны к делению. М Моноблас Промоноцит 12-18 мкм, с крупным светлым ядром, О т содержащим 1-2 ядрышка. Цитоплазма базофильная, Н (моноцито содержит все органеллы в умеренном количестве. В О бласт) цитоплазме накапливаются азурофильные гранулы. Ц Способна делится. И Т О П О Э З Проэритр област 18-22 мкм с крупным светлым ядром, содержащи м 1-2 ядрышка, бледно базофильна я цитоплазм ой. 6 класс Ретикулоцит Эритроцит -Нейтрофил -Эозинофил -Базофил Моноцит Процесс включает 1. Умен 2. Выра гемог 3. Сниж дальн орган 4. Сниж к дел 5. Конд удале 6. Форм нутой 1.Уменьш 2. Сниже делению 3.Измене ядра. 4.Накопл 5. Изм нарастан клето 1.Увелич 2.снижен 3.накопл азурофил 4.измене округлог Л В- Про-В-лимфоцит- морфологически соответствует большому лимфоциту. В цитоплазме выявляется Ig И лимфоблас М. Незрелый В-лимфоцит-морфологически М т соответствует среднему лимфоциту. На поверхности мембраны экспрессируется Ig М. Ф Протимоцит- наиболее ранняя стадия развития Тлимфоцитов в тимусе после миграции из ККМ. О Морфологически соответствует большому Тлимфоциту. лимфобоас Ц Незрелый Тлимфоцитморфологически т соответствуют средним лимфоцитам. На их И поверхности экспрессируются разнообразные антиген - распознающие рецепторы, а также маркеры Т характерные для зрелых лимфоцитов. Зрелый Влимфоцитмалый лимфоцит. На поверхности мембраны экспрессирует ся Ig М совместно с Ig D. Зрелые Тлимфоциты – морфологичес ки соответствую т малому лимфоциту, на их поверхности отсутствуют маркеры незрелых Тлимфоцитов. Антиген развити 1.Миграц предшест костного органы иммуног 2.приобр рецептор 3.отбор необходи 4.миграц перифери кроветво зависимы Антиген О развити Осущест П антигено органах О сопровож пролифер Э завершае эффектор З плазмати клеток па Примечание: лимфоцитопоэз имеет некоторые особенности: образованные дифференцированные ф переходить в менее дифференцированные клетки- бласты, которые размножаются и специализирую иммуноциты. Т Мегакарио Мегакариоцит- полиплоидная клетка, достигающая Увеличен Тромбоцит Р бласт 20-50 мкм в диаметре. Имеет ядро состоящее из клетки – (кровяная О многих долей, связанных перемычками, содержание фрагмент пластинка) М ДНК в котором своответствует 4-128n. Хроматин накоплен Б конденсируется , ядрышки отсутсвуют. В цитоплазме О сформирована система мембран (демаркационных Ц каналов), являющихся границей будущих И тромбоцитов. Также присутствуют гранулы , Т характерные для тромбоцитов. Мегакариоцит О образует множество выростов -филоподий, которые П через поры эндотелия проникают в просвет О кровеносных синусов ККМ, где распадаются на Э отдельные тромбоциты. З Лимфа (гр. чистая влага, ключевая вода) - биологическая жидкость, образующаяся из интерстициальной (тканевой) жидкости. Проходя через систему лимфатических сосудов и узлов, обогащается форменными элементами, очищается и через грудной проток попадает в кровь. Объем лимфы составляет 1-2 литра. Также как и кровь, она состоит из плазмы и форменных элементов. Плазма лимфы аналогична плазме крови по составу и концентрации солей, но отличается от нее меньшим содержанием белков и имеет щелочную реакцию (рН-8,4-9,2). Форменные элементы составляют 2-20х109/л. Их количество значительно варьирует в течение суток и при различных воздействиях. В лимфе встречаются следующие клеточные элементы: 1. лимфоциты, составляют 90% 2. моноциты-5% 3. эозинофилы-2% 4. нейтрофилы-1% 5. другие клетки-2% Эритроциты в лимфе в норме отсутствуют. Есть тромбоциты, фибриноген и другие факторы свертывания, поэтому она способна свертываться. Вопросы для самоконтроля. 1.Что такое гемограмма и лейкоцитарная формула. Каковы они у здорового человека? 2.По каким признакам различают гранулоциты и агранулоциты? 3.Какие гранулоцитарные лейкоциты Вам известны? Их функциональное значение? 4.Какие агранулоциты Вам известны? Их функциональное значение? 5.Какие особенности характерны для эритроцитов и кровяных пластинок? Ситуационные задачи. 1.В эксперименте в стенке желточного мешка на раннем этапе эмбриогенеза разрушена мезенхима. Каковы предполагаемые последствия такого нарушения? 2.В пробирку с кровью ввели мелкие пылевые частицы. В каких клетках крови они будут обнаружены? 3.По каким клеткам крови в судебной практике определяют пол преступника? 4.При нанесении татуировки подкожно вводят краску и она не разрушается. Рисунок сохраняется десятки лет. Какие из клеток крови, покидая сосуды поглощают краску? Назвать тканевую форму существования этих клеток. Назвать процесс поглощения красителя. 5.На месте преступления судмедэксперт обнаружил следы крови. После исследования им дано заключение, что преступление совершила женщина. Какие из клеток крови подвергнуты анализу и по какому морфологическому признаку клеток идентифицирован пол? 2. 2 ТЕМА: СОЕДИНИТЕЛЬНЫЕ ТКАНИ ВОЛОКНИСТЫЕ СОЕДИНИТЕЛЬНЫЕ ТКАНИ Цели занятия: 1. Обобщить изученный теоретический материал, выделяя наиболее значимые вопросы для будущей врачебной работы. 2. Приобрести навык гистологической диагностики различных видов соединительной ткани. 3. Научиться проводит диагностику типов клеток соединительной ткани по электронным фотограммам и схемам. Задания для самоподготовки: 4. Выпишите наиболее существенные признаки отличающие клетки РСТ друг от друга и запишите в виде логической или графической схемы алгоритм отличий дифференцированных клеток РСТ. 5. Студентам педиатрического факультета рекомендуем зарисовать схемы ультрамикроскопического строения клетки белого и бурого жира, выписать признаки отличий. Роль волокнистых соединительных тканей в организме. В соответствии с концепцией тканевого строения организма (А.А. Заварзин) 4 группы тканей обеспечивают выполнение 4 главных функций- задач: 1) Отделение наружной среды от внутренней среды (эпителий). 2) Формирование внутренней среды. 3) Сократимость (мышечные). 4) Возбудимость и передача импульсов (нервная ткань). Соединительные ткани являются тканями внутренней среды организма. Все ткани внутренней среды происходят из мезенхимы. Имея единый источник происхождения, они способны эффективно взаимодействовать друг с другом. Примером такого морфофункционального взаимодействия являются отношения между кровью и рыхлой соединительной тканью. Рыхлая соединительная ткань опосредует связи между кровью и другими видами тканей, формирует каркас органов. Плотная соединительная ткань формирует капсулы органов или связана со скелетными тканями. Классификация. Соединительные ткани: I. Волокнистые соединительные ткани (собственно соединительные) II. 1) Рыхлая волокнистая соединительная ткань (РВСТ). 2) Плотная волокнистая соединительная ткань (ПВСТ). Специализированные соединительные ткани 1) Ретикулярная. 2) Жировая. 3) Слизистая (у плода) Рыхлая волокнистая соединительная ткань. Название исходит из того, что структурные элементы (клетки, волокна) расположены рыхло. Рыхлая волокнистая соединительная ткань может быть названа тканью – посредником между кровью капилляров и другими тканями всех органов и систем человека. Эта универсальность определяет значение РВСТ и ее многочисленные функции. Принцип строения рыхлой волокнистой соединительной ткани 1.клетки 2.межклеточное вещество -коллагеновые и эластические волокна -аморфное вещество (гель) Клетки рыхлой волокнистой соединительной ткани. Среди клеток фибробласты (рис.12), макрофаги, адипоциты (жировые), плазматические клетки, тучные клетки, пигментные, адвентициальные. Около 60% мигрируют сюда из крови: макрофаги (производные моноцитов), плазмоциты (производные В-лимфоцитов), тучные клетки (лаброциты)– их предшественники развиваются в красном костном мозге. Исконно принадлежат рыхлой волокнистой соединительной ткани клетки дифферона фибробластов и производные стволовых клеток соединительной ткани (адипоциты, адвентициальные). Дифферон фибробластов включает стволовые и полустволовые клетки, поддерживающие эту популяцию клеток и клетки вступившие в дифференцировку: малодифференцированные (юные) фибробласты, зрелые (дифференцированные) фибробласты и конечные формы жизни этих клеток фиброциты. Стволовые и полустволовые клетки внешне сходны с лимфоцитами. Юные фибробласты проходят дифференцировку от клеток со слабо развитыми гранулярной ЭПС и митохондриями до умеренно развитых. Зрелые фибробласты распластаны, отростчатые, размером 40-50 мкм и более. Они содержат хорошо развитую гранулярную ЭПС и умеренно развитые митохондрии и комплекс Гольджи. В периферической зоне цитоплазмы фибробласты содержат небольшое количество миофибрилл, что позволяет им перемещаться в рыхлой волокнситой соединительной ткани. Значительную часть фибробластов человека по функции можно отнести к коллагенобластам. Они синтезируют на рибосомах гр. ЭПС белки, которые выходят за пределы фибробласта и здесь начинается образование волокон (фибриллогенез). Образование коллагеновых и эластических волокон и их свойства. На первом внутриклеточном этапе (рис.13) вначале в ядре происходит образование и-РНК, кодирующих синтез - цепей коллагена. Аминокислоты для синтеза коллагена поглощаются фибробластом путем эндоцитоза и транспортируются к гр.ЭПС, здесь происходит синтез полипептидных - цепей и их накопление в просвете каналов гр.ЭПС и сборка из трех - цепей тропоколлагена. Молекулы тропоколлагена переносятся в виде пузырьков в комплекс Гольджи, где происходит гликозилирование и упаковка молекул проколлагена в секреторные пузырьки. С помощью сократительного аппарата цитоскелета они перемещаются к поверхности клетки и выделяются экзоцитозом в межклеточное пространство. Здесь в межклеточной жидкости начинается второй этап фибриллогенеза: Ферменты проколлаген – пептидазы отщепляют регистрационные пептиды и переводят проколлаген в нерастворимую форму тропоколлаген. Происходит полимеризация тропоколлагена с образованием коллагеновых фибрилл толщиной 20 – 120 нм с типичной для коллагенов I, II, III типов ультраструктурной поперечной исчерченностью с периодичностью 64 нм, что связано с характером связей молекул тропоколлагена. Коллагеновые микрофибриллы объединяются за счет поперечных связей в волокна. Эти связи формируют гликозоаминогликаны и гликопротеины, которые секретируют фибробласт. Образованные коллагеновые волокна имеют диаметр от 1 до 20 мкм, прочны, мало растяжимы, могут объединяться в пучки. Типы коллагенов. Молекулы коллагенов состоят из трех спирально скрученных полипептидных - цепей, в которых преобладают аминокислоты глицин, пролин, лизин, гидрооксипролин, гидрооксилизин. Комбинации расположения молекул в - цепях приводят к появлению нескольких типов коллагена. В организме человека преобладают следующие типы коллагена: I,II,III,V – фибриллярные коллагены, IV тип – аморфные. Существуют и другие типы коллагенов. Распределение основных типов коллагена в организме человека (по Быкову В.Л., 1999г.) Тип коллагена I II III IV V РСТ, плотная соединительная ткань, роговица, волокнистый хрящ, коллагеновые волокна всех видов костной ткани и цемента. Гиалиновый, эластический хрящи и пульпозное ядро межпозвоночного диска. Ретикулярные волокна кроветворных органов. В стенке артерий и вен и в клапанах сердца, кишечнике, печени, легком, в базальной мембране вокруг гладкомышечных клеток. Аморфные компоненты коллагена. Базальные мембраны эпителиев, мышечные ткани ( кроме ГМК ), капсула хрусталика. Коллагеновые фибриллы и волокна базальных мембран, стенке кровеносных сосудов, в гладкой и исчерченной скелетной мускулатуры. Эластические волокна из белка эластина не имеют поперечной «сшивки» из гликозоаминогликанов и потому менее прочны, чем коллагеновые. Растяжимость эластических волокон связана с аминокислотным составом и характером взаимосвязей аминокислот в белковых молекулах. Растяжимость обеспечивает и аморфный эластин в осевой зоне эластического волокна. Разрушение коллагена и эластина может происходить внутриклеточно. С учетом ошибок в белковом синтезе внутриклеточно в фибробласте разрушается от 10 до 50% вновь образованного коллагена. Внеклеточное разрушение фибрилл обеспечивают ферменты, выделяемые фибробластом (коллагеназа, эластаза и другие). Нарушения фибриллогенеза лежат в основе системных заболеваний соединительной ткани – коллагенозов. Причина может быть связана с мутациями генов, кодирующих молекулы коллагена или эластина. Синтез коллагена может быть нарушен на фоне дефицита витамина С, который включен в биохимическую цепь синтеза. Внеклеточная сборка волокон может быть нарушена под влиянием токсинов микробов, иммунных реакций. Структурно- функциональные различия зрелых фибробластов. Наряду с приведенным выше описанием коллагенобласта в рыхлой волокнистой соединительной ткани есть два других вида фибробластов: миофибробласты и фиброкласты. В организме человека в течение жизни происходит изменение механических свойств стромы внутренних органов, кожи, сосудов. Это предполагает перестройку в сети коллагеновых и эластических волокон. Активная перестройка рыхлой волокнистой соединительной ткани происходит во время репаративной регенерации нарушенных структур. В указанных процессах участвуют фиброкласты, которые в своей цитоплазме наряду с аппаратом для синтеза фибриллярного белка содержат значительное число лизосом с ферментами коллагеназой, эластазой. Миофибробласты- результат гипертрофии сократительного аппарата (актиновые и миозиновые фибриллы), который заложен в любом виде фибробласта. Это направление в структурно-функциональной перестройке характерно для фибробластов матки в период беременности, для клеток окружающих края раны (контрактильная функция). Фиброциты- характерны для тех участков рыхлой волокнистой соединительной ткани, где нет образования или преобразования волокон. Эти клетки в значительной мере утрачивают способность к синтезу коллагена и эластина. Объем клеток меньше, чем у фибробластов, форма веретеновидная, слабо развиты органоиды цитоплазмы. Производные клеток крови и красного костного мозга в рыхлой волокнистой соединительной ткани. Макрофаг (Мф) - производное моноцита. Его функции не исчерпываются названием (именем) клетки. Современные представления о роли макрофагов значительно шире. Существенна роль макрофагов в иммунных реакциях. Мф расщепляет (процессирует) антиген. Тем самым он «размножает» антиген, выступая как антиген- представляющая клетка. Другая сторона участия Мф в иммунных реакциях– активация Т-лимфоцитов синтезируемым интерлейкином (ИЛ-1). Мф выделяют ростовые факторы, способные активировать фибробласты и ряд других типов клеток, что важно в процессе регенерации. Мф способны активировать воспаление через выработку простагландина Е2. Мф блокируют репликацию вирусов, вырабатывая интерферон. Основная фагоцитарная функция Мф основана: на способности к движению, хемотаксису, захвату разрушенных клеток и частиц, наличию антимикробных факторов и гидролитических ферментов в лизосомах (лизоцим, катионные белки, гидролазы и др.) Тучные клетки (тканевые базофилы) (рис.14) происходят в красном костном мозге. Ряд авторов являются сторонниками отождествления тучных клеток и базофилов крови, есть и противоположное мнение. Клетки локализованы в рыхлой волокнистой соединительной ткани вокруг сосудов микроциркуляторного русла. Их много в коже, в слизистой оболочке дыхательных путей и в пищеварительной системе. В среднем содержание тучных клеток в РСТ составляет 20%. Срок их жизни от нескольких недель до нескольких месяцев. Они способны перемещаться с помощью амебовидных движений. Основные функции тучных клеток обусловлены эффектами биологически активных веществ, которые содержатся в различных типах их цитоплазматических гранул. Наиболее многочисленны гранулы с гистамином или гепарином и дофамином. Гистамин способен увеличивать проницаемость капилляров, вызывать сокращения ГМК бронхов, повышает чувствительность к боли. Гепарин- антикоагулянт крови (связывает антитромбин III), уменьшает проницаемость межклеточного вещества рыхлой волокнистой соединительной ткани. Помимо этих типов гранул есть лизосомы с гидролитическими ферментами (протеазы, гидролазы и другие). Постоянная деятельность тучных клеток связана с обеспечением гомеостатической функции, которая реализуется через их воздействие на сосуды микроциркуляторного русла, проницаемость капилляров. Защитные функции связаны с синтезом гистамина, гепарина и дофамина как медиаторов воспаления, а также выделением хемотаксических факторов для нейтрофилов и эозинофилов. Регуляторные функции связаны с влиянием на другие типы клеток (крови, эндотелия), с помощью синтезируемых тучной клеткой цитокинов. Это способствует активации клеток. Вырабатываемые тучными клетками простагландины вызывают сокращение ГМК внутренних органов. Есть много данных о самоактивации тучных клеток в тех органах, функция которых повышена (гипертрофия). Ауторегуляторные факторы приводят к их миграции или митотическому делению. Участие в аллергических реакциях связано с наличием рецепторов в цитолемме к иммуноглобулинам класса Е (антителам) и с биологическими эффектами гистамина (см. выше). Отделение множества антител от поверхности тучной клетки приводит к последующему выходу гистамина и реализации аллергической реакции в виде расширения капилляров (крапивница или же анафилактический шок, сопровождаемый низким артериальным давлением). Спазм ГМК бронхов может приводить к гипоксии. Строение тучной клетки. Клетка с овальным ядром размером 10х22 мкм. Цитоплазма заполнена гранулами с различной плотностью и составом, диаметром от 0,3 до 1 мкм. Наиболее плотные гранулы с гепарином. Органоиды цитоплазмы развиты слабо. Плазмоциты являются иммунологическими активированными В-лимфоцитами. Ответственны за синтез иммуноглобулина (антител). Это овальные или округлые клетки диаметром 7- 10 мкм, имеют эксцентрично расположенное ядро, хорошо развитый аппарат Гольджи, максимальное развитие у этих клеток получает гранулярная ЭПС, что связано с синтезом белка- иммуноглобулина. Жизненный цикл плазматической клетки. По одним данным плазмоцит живет 2 – 3 суток, по другим до 30 суток. Плазмоциты способны синтезировать несколько типов иммуноглобулинов, которые являются гликопротеинами. Иммуноглобулины синтезируются не в виде смеси, а на каждый момент исследования плазмоциты синтезируют только один или два вида иммуноглобулинов ( из известных пяти классов иммуноглобулинов). За 1 секунду каждый плазмоцит синтезирует несколько тысяч молекул иммуноглобулинов (более 10 млн. молекул антител в час). Переключение на синтез нового класса иммуноглобулина занимает несколько часов и обеспечено необратимой рекомбинацией ДНК под влиянием цитокинов (к ним относятся: интерлейкины, факторы роста, интерферон и другие). Распределение плазматических клеток в организме человека. Наибольшая концентрация плазматических клеток характерна для лимфатических узлов (в центре фолликулов и тяжах мозгового вещества), белой пульпы селезенки, РСТ (особенно в РСТ эктодермальных желез). Патологические состояния, связанные с нарушениями развития или функций плазматических клеток. 1. Врожденный или приобретенный иммунодефецит, связанный с нарушением образования плазмоцитов. Их число в организме значительно снижено. Содержание иммуноглобулинов (антител) в крови и в межклеточной жидкости резко снижено. Указанные факторы приводят к развитию рецидивирующих инфекций. 2. Миеломная болезнь (плазмоцитома) достаточно часто встречающаяся злокачественная опухоль. При этом заболевании многократно возрастает число измененных плазмоцитов способных вырабатывать антитела. Возникает анемия, резорбция костей. Другие типы клеток рыхлой волокнистой соединительной ткани. Указанные выше клетки существенно определяют функции этой ткани, присутствуют в ней в значительных количествах (от 10 до 40%). Другие типы клеток находятся в рыхлой волокнистой соединительной ткани в небольшом числе, однако вносят важный вклад в деятельность организма. Адипоциты (жировые клетки) могут формировать крупные скопления в составе жировой ткани, а в рыхлой волокнистой соединительной ткани малочисленны. Подробно описаны в раздееле «Жировая ткань». Адвентициальные клетки– вытянутые веретеновидные клетки, часто локализуются вблизи кровеносных капилляров. Большинство из них имеют низкий уровень дифференцировки. Наиболее вероятно, что это предшественники малодифференцированных фибробластов, т.е. следующая фаза дифференцировки после стволовой клетки соединительной ткани (механоцитов). Структурные компоненты цитоплазмы развиты слабо, много свободных рибосом. Клетки способны к множественным митотическим делениям. Пигментные клетки. Ряд авторов считает (Быков В.Л.,1999г.), что в рыхлой волокнистой соединительной ткани нет клеток, способных к синтезу пигмента меланина (меланоциты), а есть лишь клетки способные захватывать и накапливать меланин – меланофоры. Это вытянутые или отростчатые клетки со слабо развитым синтетическим аппаратом, в их цитоплазме много зрелых гранул меланина (меланосом). Пигментных клеток (меланофоров) много в сосудистой оболочке сетчатки и в радужке глаза. Гранулоцитарные лейкоциты крови могут находиться в рыхлой волокнистой соединительной ткани при воспалении или при отсутствии его, поскольку здесь они заканчивают свой жизненный цикл. Участие РВСТ в защитных реакциях организма. Клетки рыхлой волокнистой соединительной ткани наряду с лейкоцитами крови участвуют в иммунных реакциях, воспалении. Иммунные реакции сопровождают антиген – представляющие клетки (специализированные макрофаги). В описании тучных клеток (см: выше) указана их роль в регуляции иммунитета и участие в аллергических реакциях. Воспаление – защитная реакция, направленная на борьбу с микроорганизмами, на отграничение воспалительного процесса. 1-ая фаза – фаза альтерации. Инициируют реакцию медиаторы воспаления из клеток крови. Значительную роль играет гистамин. С помощью гистамина, который выделяется через несколько минут после действия таких факторов, как токсины микробов, гидролитические ферменты разрушенных клеток и другие. Гистамин открывает «ворота» капилляров (приводит к увеличению пространств между эндотелиальными клетками) для выхода нейтрофилов из крови в РСТ. 2-ая фаза – фаза экссудации. Гистамин выделяет хемотоксические факторы, направляющие нейтрофилы в очаг воспаления. Значительный выход нейтрофилов происходит не только из капилляров, но и из посткапиллярных венул. Нейтрофилы создают вокруг очага воспаления лейкоцитарный вал через 12 – 24 часа после начала воспаления и вступают в борьбу с микробами используя гидролитические ферменты, катионные белки. В борьбе с микробами «победу одерживают» чаще нейтрофилы. Однако, потери имеют место с той и с другой стороны. На 2 сутки в «очистку» очага воспаления от «трупов» нейтрофилов и микробов включаются макрофаги, которые по хемотаксису мигрируют в очаг воспаления. Макрофаги заняты не только фагоцитозом, но и синтезем веществ, активирующих фибробласты (фактор роста фибробластов – ФРФ), а также вырабатывают простагландин Е 2, стимулирующий деятельность клеток в очаге воспаления. 3-я фаза – фаза пролиферации (фаза репарации). Под действием активирующих факторов на 3 сутки очаг воспаления окружают мигрировавшие сюда фибробласты, одновременно здесь происходит размножение фибробластов. Главная задача фибробластов: создать «забор» из коллагеновых волокон вокруг очага воспаления. Этот процесс наиболее активно идет с 3 по 7 сутки после начала воспаления. В итоге очаг воспаления или инородное тело (вызвавшее воспаление) отграничиваются от остальных частей органа. Внутри очага воспаления формируется грануляционная ткань – молодая рыхлая волокнистая соединительная ткань с высоким содержанием клеточных элементов. Аморфное (основное) вещество. Это гель, в образовании которого принимают участие кровеносные капилляры и клетки рыхлой волокнистой соединительной ткани. В состав геля входят гликозоаминогликаны, хондроитинсульфаты, липиды, протеогликаны, альбумины, глобулины крови, ферменты, минеральные вещества, вода. Схема транспорта жидкости в рыхлой волокнистой соединительной ткани. Аморфное вещество распределено в органах неравномерно. Его меньше рядом с кровеносными и лимфатическими сосудами и больше вблизи эпителиальной, мышечной и нервной ткани. Наличие протеогликанов способствует удержанию жидкости в РВСТ. Плотная волокнистая соединительная ткань. В зависимости от расположения пучков волокон эту ткань подразделяют на оформленную и неоформленную. Неоформленная волокнистая соединительная ткань. Имеет неупорядоченное расположение волокон, что характерно для капсул некоторых органов, надкостницы, надхрящницы, сетчатого слоя дермы кожи. В неоформленнолй волокнистой соединительной ткани пучки коллагеновых волокон разнонаправлены, разделены фиброцитами. Между пучками могут быть тонкие прослойки рыхлой волокнистой соединительной ткани с сосудами. Оформленная соединительная ткань встречается в виде связок, сухожилий, фиброзных мембран. Характерной чертой строения этой ткани является однонаправленное расположение пучков волокон, между которыми расположены фиброциты. Строение сухожилий. Наличие большого числа пучков коллагеновых волокон и прислоек соединительной ткани позволяет унифицировать деление сухожилия на его части– пучки. Пучки I порядка входят в состав пучков 2 порядка и разделены фиброцитами (сухожильные клетки). Пучки 2 порядка образуют основу сухожилия и включают несколько десятков пучков I порядка, которые отделены друг от друга прослойками рыхлой волокнистой соединительной ткани (эндотеноний). Крупное сухожилие может иметь пучки III, V, VI порядков, состоящие из нескольких пучков меньшего порядка. Снаружи сухожилие окружено пластиной РВСТ (перитеноний). Питание элементов сухожилия обеспечивают кровеносные капилляры, находящиеся в прослойках рыхлой волокнистой соединительной ткани. Сухожилия содержат механорецепторы. Специализированные соединительные ткани. Ретикулярная ткань (рис.15)– образует строму кроветворных органов как разновидность соединительной ткани, которая по строению наиболее близка к РСТ. Принцип строения совпадает : клетки, волокна, аморфное вещество. Ретикулярные клетки подобны фибробластам, способны образовывать коллаген III типа, из которого формируются ретикулярные волокна. Важной функцией ретикулярных клеток является участие в формировании среды окружения для кроветворных клеток вместе с макрофагами. Основные функции ретикулярных клеток. 1. Синтетическая – образование волокон и аморфного межклеточного вещества (гликозоаминогликаны и др.). 2. Регуляторная – для развивающихся клеток крови: синтез гемопоэтинов (цитокинов, факторов роста) для регуляции деления и дифференцировки клеток. 3. Трофическая – транспорт и распределение питательных веществ, поступающих из капилляров. Ретикулярные волокна. Образуют ретикулярные клетки, синтезируя коллаген III типа, гликопротеины и протеогликаны. Указанные вещества входят в состав волокон и влияют на их свойства. Волокна диаметром 0,1 – 0,2 мкм формируют тонкую сеть, обладают способностью к импрегнации солями серебра (аргирофилией). Волокна включают в себя много углеводных молекул и способны к небольшим растяжениям. Основное (аморфно)е вещество ретикулярной ткани. Содержит жидкость, диффундирующую из капилляров и вещества, выделяемые клетками. Ретикулярные клетки синтезируют и выделяют в межклеточное вещество гликопротеины, гликозоаминогликаны, а также вещества способствующие адгезии (связи) между кроветворными клетками и элементами стромы (фибронектин, гемонектин, ламинин). Макрофаги ретикулярной ткани взаимодействуют со всеми ее элементами. Основные функции макрофагов в ретикулярной ткани. 1. Фагоцитарная – макрофаги способствуют фагоцитозу разрушенных клеток. 2. Метаболическая – наиболее изучена в красном костном мозге (ККМ). Макрофаги ККМ накапливают железо и передают его развивающимся клеткам эритроцитарного ряда в виде комплекса железо – белок (ферритин). 3. Регуляторная – заключается в продукции цитокинов и факторов роста (ИЛ-1, КСФ, ФНО), которые влияют на гемопоэз, макрофаги способны индуцировать другие клетки (ретикулярные, фибробласты, Т-лимфоциты, эндотелиоциты) к синтезу гемопоэтинов. 4. В периферических лимфоидных образованиях макрофаги выступают как антиген – представляющие клетки. Жировая ткань. Основной объем в этой ткани занимают жировые клетки – адипоциты (рис.16); небольшой объем занят коллагеновыми и эластическими волокнами. Ткань составляет 15-20% - у мужчин и 20-25% - у женщин от обьема массы тела. Медицинские аспекты связаны с нарушением липидного обмена, ожирением, истощением и др. Жировая ткань содержит 80% энергоемких веществ тела. Установлено, что метаболизм адипоцитов высок и связан с непрерывным обменом липидов (липогенез – липолиз). В первые месяцы жизни у ребенка 2 вида жировой ткани: белая и бурая, а затем бурая жировая ткань подвергается атрофии Функции жировой ткани: 1. Энергетическая (трофическая, теплообразующая). При дефиците энергоемких веществ, происходит расщепление липидов (липолиз), что обеспечивает клетку веществами для энергетических (биохимических) процессов, часть энергии уходит в тепло. 2. Теплоизолирующая – топография жировой ткани в коже (гиподерма) является указанием на эту функцию. Прослойка жировой ткани в коже препятствует потере тепла. 3. Опорная и пластическая – окружая органы, сосудисто-нервные пучки жировая ткань препятствует их травматизации. Она создает амортизирующую прослойку под кожей подошвы и ладонных поверхностей кистей рук. 4. Регуляторная – через ферменты адипоцитов происходит регуляция липидного обмена. Здесь же депонируется часть женских половых гормонов (эстрогенов), а также витамины (А,Д,Е,К). Адипоциты вырабатывают гормон, регулирующий потребление пищи – лептин. Этот вид регуляции тесно связан с деятельностью пищевого центра (гипоталамус, кора больших полушарий мозга). В красном костном мозге жировые клетки входят в состав микроокружения кроветворных клеток и тем самым оказывают влияние на гемопоэз. Развитие. Белая жировая ткань развивается из стволовых клеток соединительной ткани. В ходе дифференцировки клетки превращаются в преадипоциты, в которых в последствии появляются ферменты для синтеза липидов. Клетки вступают в фазу синтеза и накопления липидов. Мелкие липидные капли сливаются и происходит увеличение объема крупной капли, происходит перестройка цитоскелета, оттеснение остальной цитоплазмы к периферии. Основную массу ткани составляют адипоциты (рис.16). Общее число адипоцитов взрослого человека составляет 20-30 · 109 . При ожирении число клеток может значительно возрастать. Адипоциты –крупные клетки диаметром от 25 до250 мкм, имеют округлую форму. Основной объем цитоплазмы заполнен липидными включениями в виде единой капли, остальная масса занимает небольшой объем с развитыми агранулярной ЭПС и аппаратом Гольджи. Плазмолемма содержит многочисленные рецепторы к гормонам и нейромедиаторам (норадреналину и др.). Размножение, дифференцировка адипоцитов происходит под воздействием: гормона роста гипофиза, тиреоидных гормонов и инсулиноподобного фактора роста –1. Отложение жиров тесно связано с участием ферментов липогенеза, с регуляторным действием гормона инсулина. Расщепление жиров обеспечивается ферментом липазой, которая находится под регуляторным действием нейромедиатора норадреналина, а также гипофизарных гормонов (роста, липотропного и других), гормонов коры надпочечников (глюкокортикоидов) и тиреоидных гормонов. С возрастом уменьшается чувствительность рецепторов адипоцитов к указанным регуляторам. Бурая жировая ткань характерна для новорожденных и детей первых месяцев жизни. В наибольшей степениэтот вид ткани ориентирован на термогенез. Адипоциты способны в сотни раз повышать активность окислительных процессов при охлаждении и увеличивать температутру в зоне метаболизма. Адипоциты заполнены множеством мелких капель липидов окруженных митохондриями. Митохондрии имеют окрашенные окислительные ферменты – цитохромы, которые и обусловили цвет ткани. Работа митохондрий направлена на термогенез, вследствие разобщения окисления и фосфорилирования (белок – термогенин). В итоге не происходит накопление (складирование) энергии в макроэргических соединениях (АТФ), а образуется значительное количество тепла. Главный регулятор адипоцитов - медиатор симпатических нервных окончаний норадреналин. Слизистая ткань. Присутствует у плода в пупочном канатике, препятствует сдавлению сосудов пупочного канатика, образованию петель, узлов. Является видоизмененной рыхлой волокнистой соединительной тканью, способной за счет высокой концентрации гиалуроновой кислоты в аморфном веществе связывать множество молекул воды, обеспечивать гидрофильность ткани. Клетки мукоциты сходны с фибробластами, образуют тонкие коллагеновые волокна, рыхло расположенные в аморфном веществе. Вопросы для самоконтроля: 1.Назовите известные Вам заболевания связанные с патологией соединительной ткани. 2.Как классифицируется собственная соединительная ткань? 3.Перечислить виды клеток в рыхлой волокнистой соединительной ткани? Их строение? 4.Перечислить специализированные соединительные ткани. Ситуационные задачи. 1.Какова реакция клеточных элементов соединительной ткани при попадании занозы под кожу? 2.При введении человеку живой вакцины вырабатывается специфический иммунитет. Какие клетки рыхлой волокнистой соединительной ткани участвуют в этом процессе? 3.Чем объяснить, что при укусе змеи, пчелы яд быстро проникает в кровь? 4.Спортсмен во время бега повредил сухожилие, в результате образовался соединительнотканный рубец. Каковы условия, которые необходимы для регенерации сухожилия? За счет каких структурных элементов произойдет восстановление сухожилия? 5.При недостатке витамина С в фибробластах нарушен синтез белка тропоколлагена. Назвать изменения происходящие в межклеточном веществе. 3.ТЕМА: ХРЯЩЕВЫЕ ТКАНИ Цели занятия: 1.Научиться определять разновидности хрящевых тканей в зависимости от структурных особенностей межклеточного вещества. 2.Знать гистофункциональные особенности разных видов хрящевой ткани. Общая характеристика и значение. В начальный период эмбрионального гистогенеза основу скелета составляют хрящевые ткани (рис.17). В последующем в скелете преобладают костные ткани, но роль хрящевых тканей незаменима в сочленениях, в росте костей в длину. Хрящевые ткани входят не только в скелет, но и образуют основу для воздухоносных путей (нос, гортань, трахея, крупные и средние бронхи), ушной раковины. У взрослого человека хрящи составляют приблизительно 2% от его массы. Основной объем в хрящевой ткани приходится на межклеточное вещество. Структурной основой которого, являются соединительнотканные волокна. Клетки (хондроциты) составляют не более 10% массы хряща. Аморфное вещество достаточно гидрофильно, что позволяет доставлять клеткам питательные вещества путем диффузии из капилляров надхрящницы. Для хрящевых тканей характерны: *относительно низкий уровень метаболизма *отсутствие сосудов *гидрофильность *прочность и эластичность. Классификация хрящевых тканей. По особенностям строения и биохимического состава межклеточного вещества выделяют: 1. Гиалиновую хрящевую ткань. 2. Эластическую хрящевую ткань. 3. Волокнистую хрящевую ткань. Гистогенез. В эмбриогенезе хрящи формирует мезенхима. Клетки мезенхимы размножаются, вступают в дифференцировку, округляются, образуют скопления хондробластов (хондрогенные островки). Хондробласты, вступая в дифференцировку, приобретают структуры и способность к образованию межклеточного вещества в виде коллагена (второго типа) и гликозоаминогликанов (для аморфного вещества). В окружении аморфного вещества синтетическая способность клеток снижается, они становятся хондроцитами. Рост хряща осуществляется двумя путями: внутренним ростом (интерстициальным) и аппозиционным. Первый путь обеспечен способностью хондроцитов к размножению и синтезу веществ. Аппозиционный рост обеспечивают хондробласты, внутреннего слоя надхрящницы. Аппозиционный рост у взрослого возможен только при травме хряща. Стимулируют рост хряща гормоны (СТГ, гормоны щитовидной железы и др.). Дифферон хрящевой ткани. Стволовые - полустволовые (прехондробласты) – хондробласты – молодые хондроциты – зрелые хондроциты. Клетки хрящевых тканей. Все виды хрящевых тканей имеют общие принципы строения. Это относится и к клеткам хондроцитам. Хондроциты – являются производными хондробластов и единственной популяцией клеток в хрящевой ткани. Расположены в лакунах. Хондроциты можно подразделить по степени зрелости на молодые и зрелые. Молодые сохраняют черты строения хондробластов. Они имеют продолговатую форму, развитую гранулярную ЭПС, крупный аппарат Гольджи, способны образовывать белки для коллагеновых и эластических волокон и сульфатированные гликозоаминогликаны, гликопротеины. Зрелые хондроциты имеют овальную или округлую форму. Синтетический аппарат развит в меньшей степени при сравнении с молодыми хондроцитами. В цитоплазме происходит накопление гликогена и липидов. Хондроциты способны к делению и образуют изогенные группы клеток, окруженные одной капсулой. В гиалиновом хряще изогенные группы могут содержать до 12 клеток, в эластическом и волокнистом хрящах – меньшее число клеток. Гиалиновая хрящевая ткань. Образует хрящевые пластинки роста в костях, основу воздухоносных путей, суставные поверхности. Клетки хряща хондроциты (описаны выше). Межклеточное вещество имеет три составляющих: 1. Коллагеновые волокна (20-25%). Их образует коллаген II типа. Ориентация волокон в соответствии с направлениями действующих сил. На срезе волокна образуют сетчатый рисунок, сложное переплетение. 2. Протеогликаны составляют 5-10% массы. Они включают в себя молекулы связывающие волокна и воду: сульфатированные гликозоаминогликаны, гликопротеины. Протеогликаны гиалинового хряща препятствуют его минерализации. 3. Интерстициальная вода (65-85%). Благодаря такому физическому свойству как несжимаемость, вода не только наполнитель хряща, но и амортизатор. Вода способствует эффективному обмену веществ в хряще, переносит соли, питательные вещества, метаболиты. Суставной хрящ. Эта разновидность гиалинового хряща имеет особенности строения и питания. Покрывая суставные поверхности, хрящ увлажнен синовиальной жидкостью, которая является основным источником питания его поверхностной зоны. В суставном хряще выделяют: поверхностную зону, которую можно назвать бесклеточной, среднюю (промежуточную) – содержащую колонки хрящевых клеток и глубокую зону, в которой хрящ взаимодействует с костью. Эластическая хрящевая ткань. Этот вид ткани необходим для тех участков органов, которые способны менять свой объем, форму и обладают обратимой деформацией. Хрящ входит в состав ушной раковины, наружного слухового прохода, евстахиевой трубы, надгортанника, гортани. Хондроциты – обладают структурой и функциями описанными выше. Межклеточное вещество в своей структурной основе содержит эластические и незначительное количество коллагеновых волокон, которые формируют густую сеть. Аморфное вещество эластического хряща сходно с таковым в гиалиновой хрящевой ткани. Волокнистая (коллагенововолокнистая) хрящевая ткань. Ее выявляют в межпозвоночных дисках, в симфизе (лонное сочленение), на месте прикрепления сухожилий к костям или гиалиновому хрящу. Хондроциты – расположены между пучками волокон поодиночке или в виде небольших изогенных групп. Хондроциты способны синтезировать не только типичный для хряща коллаген II типа, но в большей степени синтезирую коллаген I типа, характерный для волокнистой соединительной ткани. Межклеточное вещество образованно пучками волокон из коллагена I типа и аморфного вещества типичного для хрящевых тканей. Хрящ как орган. В хрящевой ткани нет клеток, способных обеспечить репаративную (посттравматическую) регенерацию, нет сосудов, обеспечивающих питание хряща, поэтому метаболизм хрящевых тканей неразрывно связан со структурами жизнеобеспечения. В подавляющем большинстве такой структурой является надхрящница. Она имеет две пластинки: наружная из плотной неоформленной соединительной ткани, внутренняя пластинка более тонкая, из рыхлой волокнистой соединительной ткани, содержит капиллярные сети и клетки – предшественницы хондробластов. Регенерация хряща. При травме хряща важным условием регенерации является сохранение клеток, способных к образованию хондробластов. Такие клетки находятся в надхрящнице. Активацию регенерации обеспечивают цитокины. Хондробласты включаются в воспроизводство коллагена (эластина) и гликопротеидов аморфного вещества. После формирования волокон и аморфного вещества часть хондробластов оказывается в окружении матрикса хряща и превращается в хондроциты. В регенерацию могут включатся стволовые клетки соединительной ткани. Регуляция хрящевой ткани. Хондроциты имеют рецепторы к соматотропному гормону (СТГ), тироксину, инсулину, глюкокортикоидам, эстрогенам, а также к цитокинам. Гормоны щитовидной железы (тироксин, трийодтиронин) ускоряют дифференцировку хондроцитов. Инсулин способен выступать как фактор роста хрящевой ткани. Глюкокортикоиды, напротив, угнетают синтез коллагена. Мужской половой гормон тестостерон стимулирует синтез несульфатированных гликозоаминогликанов. Это способствует удержанию воды в аморфном веществе хряща. Таким образом, хрящевая ткань находится под влиянием многочисленных регуляторов, влияющих на деятельность хондробластов и состояние межклеточного вещества. Вопросы для самоконтроля: 1.Назвать источник развития хрящевых тканей. 2.Как классифицируют хрящевые ткани? 3.За счет чего происходит рост хряща? 4.Перечислить компоненты межклеточного вещества. 5.Перечислить клетки хрящевых тканей, их строение. 6.Назвать факторы, влияющие на строение хрящевых тканей. Ситуационные задачи. 1.Под микроскопом рассмотрено три микропрепарата хрящевых тканей, окрашенных гематоксилином и эозином и орсеином. Какие волокна и в какой хрящевой ткани будут выявляться при этих способах окрашивания. 2.Из каких хрящевых тканей образованы хрящи гортани? 3.На микропрепаратах разных видов хрящевых тканей видно, что в изогенных группах разное количество хондроцитов: более 10 и от трех до пяти. Для каких видов хрящевой ткани это характерно? 4.У больного обнаружено отложение солей кальция в межклеточном веществе хряща. Какой хрящ (гиалиновый, эластический, волокнистый) подвержен обызвествлению? Какие участки (периферические или центральные) подвержены обызвествлению в первую очередь? 5.На электронограмме суставной поверхности в хондроцитах многочисленные видны многочисленные секреторные везикулы. Что может содержаться в везикулах? 4. ТЕМА:КОСТНЫЕ ТКАНИ Цели занятия: 1.Обобщить знания по развитию и строению костных тканей. Изучить в разделе «Волокнистая соединительная ткань» последовательность, механизмы и регуляцию образования волокон соединительной ткани. 2.Получить навыки микроскопической диагностики различных видов костной ткани и их клеток. Общая морфофункциональная характеристика, значение и классификация костных тканей. Костные ткани входят в состав скелета и составляют у взрослого человека 17-20% его массы. Кости определяют биологическую конституцию человека, входят в локомоторный аппарат, формируют зоны защиты для некоторых внутренних органов, являются минеральным депо. Основную массу (до 70%) в костных тканях составляют костные апатиты; около 30% составляет органический субстрат из коллагеновых волокон, которые минерализованы; 3-5% составляют клетки костной ткани. Деятельность клеток костной ткани находится под контролем регуляторных гормонов ( паратирин, кальциотонин и др.). Классификация костных тканей. Наибольший объем у взрослого человека составляет пластинчатая костная ткань, которая бывает компактная и губчатая. На поверхности пластинчатых костей в зоне прикрепления сухожилий, а также в швах черепа находится ретикулофиброзная костная ткань. Специфично строение костной основы зуба – дентина. Костную ткань формируют остеобласты. Остеобласты на начальном этапе деятельности сходны с фибробластами, поскольку образуют органический матрикс – остеоид из коллагеновых (преимущественно) волокон и аморфного вещества. Второй этап – минерализация. Происходит при участии ферментов остеобластов. В результате минерализации образуется костный апатит (гидрооксиапатит) в межклеточном веществе, в том числе и в волокнах. Структура и функции клеток костной ткани. Остеобласты, остеоциты, остеокласты включают в себя два дифферона клеток: остеоцитарный (остеобласты – остеоциты) и остеокластный. Остеобласты (рис.18) образуются из остеогенных стволовых клеток костного мозга. Эти клетки мигрируют в участки будущей кости или зоны ее перестройки, размножаются, трансформируются вначале в предшественников, а затем в остеобласты. Остеобласты способны к делению, располагаются группами, имеют неровную поверхность и короткие отростки, связывающие их с соседними клетками. Остеобласты обладают способностью к продукции белка, образуя преимущественно коллаген I типа (90%), а также гликопротеины матрикса (остеонектин, сиалопротеин, остеокальцин) и протеогликаны. Важное значение имеет продукция ферментов (щелочной фосфотазы и др.) и информационных молекул: факторов роста, цитокинов, морфогенетических белков. Синтез белков обусловил выраженное развитие гранулярной ЭПС, аппарата Гольджи, митохондрий. Покоящиеся (неактивные) остеобласты находятся на поверхности кости, веретеновидной формы с редуцированными органоидами. Участие остеобластов в минерализации костной ткани. Процесс минерализации начинается после образования межклеточных коллагеновых волокон – остеоида. Остеобласты, выделяя фермент щелочную фосфатазу, способствуют дефосфорилированию межклеточного субстрата, повышению концентрации фосфатных ионов. Характерно, что этот процесс связан с выведением из цитоплазмы остеобластов мелких матричных пузырьков с высоким содержанием фосфата кальция и щелочной фосфотазы. Пузырьки инициируют образование кристаллов гидроксиапатита. Наиболее активно этот процесс протекает в коллагеновых волокнах. Этому способствуют и некоторые протеогликаны, изменяющие некоторые молекулярные связи в волокнах. В результате 90-95% солей кальция включается в состав коллагеновых волокон и только 5%–10% содержится в остальной части костного матрикса. Регуляция и минерализация. Минерализация остеоида занимает у человека около двух недель. Дефицит витамина Д замедляет минерализацию. Гормон паращитовидной железы паратирин (паратгормон) препятствует минерализации. Угнетают развитие кости глюкокортикоиды, выделяемые корой надпочечников. Активируют минерализацию кальциотонин, тиреодные и половые гормоны. Возможны доброкачественные (остеомы) и злокачественные (остеосаркомы). В первом случае клетки сохраняют способность к выработке органического субстрата и его минерализации, но нарушены темпы и может быть изменена топография клеток. При злокачественном росте уровень дифференцировки клеток соответствует клеткампредшественникам остеобластов, отсутствует образование волокон. Клетки расположены группами, контактируя друг с другом. В цитоплазме много свободных рибосом, характерен полиморфизм клеток. Остеоциты – основной тип клеток костной ткани (рис.19). Эти клетки являются производными остеобластов. Говоря образно остеоциты – это остеобласты заключенную в камеру называемую костной полостью – лакуной. В ходе формирования кости остеобласт окружает себя органическим субстратом, который не нарушает питание остеобласта. Начавшаяся минерализация резко уменьшает приток питательных веществ и остеобласт выпускает сотни отростков, формируя трофические (питательные) каналы. Эти каналы не только позволяют найти клеткам «родники» питательных веществ (капилляры, аморфное вещество РСТ), но и установить структурно-функциональные связи между собой с помощью щелевидных (ионообменных) контактов. Образуется новое содружество клеток – сеть остеоцитов, тела которых расположены в костных лакунах. Строение и функции остеоцитов. Остеоциты запрограммированы на достижение оптимального обмена веществ в костной ткани. Это было бы невозможно сделать, если не создать в костях миллиарды тонких, пронизывающих кость каналов. По этим каналам можно «выкачивать» в кровь минералы и «закачивать» минералы в кость. Остеоциты являются «сборщиками урожая» с огромного минерального поля костной ткани. Главная характерная черта в строении остеоцитов наличие большого количества тонких выростов (до 300), отходящих от тела клетки. Эти выросты входят в костные каналы. Часть из них контактирует с отростками соседних клеток с помощью щелевидных контактов (обмен ионами и информационными молекулами) и десмосом. Остеокласты (рис.20)–производные моноцитов, которые в костной ткани сливаются в симпласты, достигая крупных размеров (до 100 мкм) и приобретая феномен многоядерности (до 50 ядер). В активированном остеокласте выражена полярность. Та часть клетки, которая прилежит к кости и разрушает ее, имеет многочисленные складки в цитолемме (гофрированный край). Микроворсинки постоянно меняют свою форму и размеры. В цитоплазме этого полюса много лизосом, митохондрий. Резорбция кости включает в себя две фазы: деминерализацию, растворение и фагоцитоз органического субстрата (волокон и др.). Остеокласт приспособлен для резорбции и осуществляет ее в несколько этапов. Первый этап – прикрепление к кости с помощью белков интегринов, витронектинов и др. Это обеспечивает герметизацию зоны прикрепления и препятствует выходу ферментов за пределы этой зоны. Второй этап – закисление и растворение минералов в участке разрушения путем накачивания ионов водорода с участием АТФаз мембран гофрированного края.Фермент карбоангидраза способствует образованию кислоты (Н2 СО3 ). Третий этап – растворение органического субстрата кости с помощью ферментов лизосом (гидролазы, коллагеназы и др.), которые остеокласт выводит экзоцитозом в зону разрушения. Регуляция остеокластов обеспечена общими и местными факторами. Общие факторы – гормоны паращитовидной (паратирин), щитовидной (кальциотонин) желез, яичника (эстрогены). Паратирин активирует, кальциотонин и эстрогены–угнетают активность остеокластов. К общим факторам следует отнести факторы роста и витамин Д3, который способствует увеличению числа остеокластов, активируя слияние моноцитов. Местные влияния могут быть со стороны клеток, окружающих остеокласт. Лимфоциты, макрофаги способны активировать остеокласты. Важную роль в активации играют пьезо – эффекты, возникающие в кости при ее растяжении. При этом изменяется электрический заряд кристаллов кости, что и является активирующим фактором. Общая характеристика ретикулофиброзной и пластинчатой костной ткани. Ретикулофиброзная костная ткань формируется у плодов человека как основа костей. У взрослых она представлена незначительно и находится в швах черепа в местах прикрепления сухожилий к костям. Основу этой ткани составляют беспорядочно расположенные пучки коллагеновые минерализованных волокон. В основном веществе этой ткани расположены костные полости с длинными анастомозирующими канальцами, в которых лежат костные клетки остеоциты с отростками. С поверхности участки кости покрыты надкостницей, из которой ретикулофиброзная костная ткань получает питательные вещества путем диффузии. Пластинчатая костная ткань – основной вид костной ткани во взрослом организме. Она состоит из костных пластинок, толщина и длина которых от нескольких десятков до сотен микрометров. Пластинки включают минерализованные тонкие коллагеновые волокна ориентированных в различных плоскостях аморфного вещества. В центральных частях пластинок фибриллы и волокна расположены продольно, по периферии тангенциально и поперечно. Пластинки могут расслаиваться, а фибриллы одной пластинки могут переходить в соседние, образуя единую волокнистую основу кости. Кроме того, костные пластинки пронизаны отдельными фибриллами и волокнами направленными перпендикулярно костным пластинкам. Волокна и фибриллы вплетаются в промежуточные слои между пластинками, все это обуславливает большую прочность пластинчатой костной ткани. В костных пластинках есть лакуны расположенные параллельно пластинкам, где лежат остеоциты. Их отростки, входящие в костные канальцы направлены перпендикулярно костным пластинкам. Из пластинчатой костной ткани построены компактное и губчатое вещество большинства плоских и трубчатых костей. В компактных и губчатых костях костные пластинки образуют комплексы – трабекулы. В губчатой кости трабекулы состоят из 2-3 параллельно расположенных пластинок, соединенных с другими пластинками. Трабекулы отграничивают полости заполненные красным костным мозгом. В компактной кости комплекс пластинок многократно массивнее. Характерны общие пластинки, пластинки остеонов и вставочные пластинки. В губчатой и компактной кости всегда присутствуют такие органные образования как кровеносные сосуды. Капилляры выступают в пластинчатой кости в роли трофической зоны, вокруг которой строятся костные пластинки. Вокруг капилляров тонкие прослойки соединительной ткани, которые участвуют в регенерации кости. Гистологическое строение трубчатой кости как органа. Кость как орган обладает сложной архитектоникой и тканевым составом. Ведущей тканью кости взрослого служит пластинчатая костная ткань, снаружи и со стороны костномозговой полости она покрыта соединительнотканными оболочками (надкостница, эндост). Кость содержит красный костный мозг, кровеносные и лимфатические сосуды и нервы. В кости, как в органе различают компактное (кортикальное) вещество кости и губчатое (трабекулярное) вещество, которые образованы пластинчатой костной тканью. Надкостница, или периост состоит из наружного (волокнистого) и внутреннего преимущественно клеточного слоев. Внутренний слой содержит остеогенные камбиальные клетки, преостеобласты, остеобласты. Камбиальные клетки веретеновидной формы, имеют небольшой объем цитоплазмы и умеренно развитый синтетический аппарат. Через надкостницу проходят питающие кость сосуды и нервы. Рыхлая соединительная ткань окружающая сосуды содержит камбиальные элементы для развития кости. Надкостница принимает участие в трофике костной ткани, развитии, росте и регенерации. Суставные поверхности эпифизов не имеют надкостницы и надхрящницы. Они покрыты разновидностью гиалинового хряща –суставным хрящом. Строение диафиза. Компактное вещество, образующее диафиз кости, состоит из костных пластинок, толщиной от 4-15мкм, расположенных в определенном порядке. В диафизе различают три слоя: наружный слой общих пластинок; средний слой, образованный концентрически наслоенными пластинками вокруг костных каналов с сосудами и называемый остеонным слоем; внутренний слой пластинок. Наружные общие пластинки не образуют полных колец вокруг диафиза кости, а прерываются. Внутренние общие пластинки хорошо развиты только там, где компактное вещество кости непосредственно граничит с костномозговой полостью. Там, где компактное вещество переходит в губчатое, его внутренние общие пластинки переходят в пластинки перекладин. В наружных общих пластинках залегают прободающие (фолькмановы) каналы, по которым из надкостницы внутрь кости входят сосуды. Из надкостницы в кость проникают коллагеновые волокна, их называют прободающими (шарпеевскими) волокнами. Они чаще разветвляются в слое наружных общих пластинок. Могут доходить и до среднего (остеонного) слоя, но в пластинки остеонов не входят. В среднем слое большинство костных пластинок располагаются в остеонах, а между остеонами лежат вставочные пластинки, направленные перпендикулярно пластинкам остеонов. Остеоны являются структурными единицами компактного вещества трубчатой кости. Они представляют собой цилиндрические образования, состоящие из костных пластинок, как бы вставленных друг в друга. В костных пластинках и между ними располагаются тела костных клеток и их отростки, проходящие в межклеточном веществе. Каждый остеон отграничен от соседнего остеона спайной линией, образованной основным веществом. В канале остеона проходят кровеносные сосуды с соединительной тканью и остеогенными клетками. Сосуды каналов остеонов сообщаются друг с другом и с сосудами костного мозга и надкостницы. На внутренней поверхности диафиза, граничащей с костномозговой полостью, пластинчатая костная ткань образует костные перекладины губчатого вещества кости. Эндост- оболочка, покрывающая кость со стороны костномозгового канала. В эндосте различают осмиофильную линию на наружном крае минерализованного вещества кости; остеоидный слой, состоящий из аморфного вещества, коллагеновых фибрилл и остеобластов, кровеносных капилляров и нервных окончаний; слоя чешуевидных клеток, нечетко отделяющих эндост от элементов костного мозга. Толщина эндоста превышает 12 мкм. Полость диафиза трубчатых костей заполнена костным мозгом (красным и желтым). Между эндостом и периостом есть микроциркуляция жидкости благодаря лакунарно–каналовидной системе кости. Васкуляризация костной ткани. Кровеносные сосуды образуют во внутреннем слое надкостницы густую сеть. Отсюда берут начало тонкие артериальные веточки, которые кровоснабжают остеоны, проникают в костный мозг через питательные отверстия и образуют питающую сеть капилляров, проходящую по остеонам. Иннервация костной ткани. В надкостнице миелиновые и безмиелиновые нервные волокна образуют сплетения. Часть волокон сопровождают кровеносные сосуды и проникают с ними через питательные отверстия в каналы остеонов и далее достигают костного мозга. Перестройка и обновление костной ткани. В течение всей жизни человека происходит перестройка и обновление костной ткани. Разрушаются первичные остеоны и одновременно появляются новые, как на месте разрушенных, так со стороны периоста. Под влиянием остеокластов, которые активизируются различными факторами, костные пластинки остеона разрушаются и на этом месте образуется полость. Этот процесс называется резорбцией костной ткани. В полости вокруг оставшегося сосуда появляются остеобласты, которые начинают строить новые пластинки, концентрически наслаивающиеся друг на друга. Так возникают вторичные генерации остеонов. Между остеонами располагаются остатки разрушенных остеонов прежних генераций (вставочные пластинки). Среди факторов, влияющих на перестройку костной ткани важную роль играет пьезоэлектрический эффект. Оказалось, что в костных пластинках при изгибах появляется разность потенциалов между вогнутой и выпуклой сторонами. Вогнутая сторона заряжена отрицательно, а выпуклая положительно. На отрицательно заряженной поверхности всегда отмечается активация остеобластов, в процессе аппозиционного новообразования костной ткани, а на положительно заряженной наблюдается ее резорбция с помощью остеокластов. Искусственное создание разности потенциалов дает этот же результат. Нулевой потенциал, отсутствие физической нагрузки на костную ткань (продолжительная иммобилизация и др.) обуславливает повышение функций остеокластов и выведение солей из кости. На структуру костной ткани и костей оказывают влияние витамины (С,Д,А), гормоны щитовидной железы, околощитовидной и др. Так, при недостатке витамина С подавляется образование коллагеновых волокон, уменьшена активность остеобластов. При дефиците витамина Д не происходит полной кальцификации органической матрицы кости, что ведет к размягчению костей. Витамин А поддерживает рост костей, но его избыток способствует усилению разрушения остеокластами метаэпифизарных хрящей- зоны роста кости в длину. Возрастные изменения. С возрастом увеличивается общая масса соединительнотканных образований, изменяется соотношение типов коллагена, гликозаминогликанов, больше становится сульфатированных соединений. В эндосте стареющей кости уменьшается популяция остеобластов, но возрастает активность остеокластов, что ведет к истончению компактного слоя и перестройке губчатого вещества кости. У взрослых полная смена образований кости зависит от ее размера и для бедра составляет 7-12 лет, для ребра 1 год. Перестройка кости происходит с участием ее разрушителей– остеокластов и «строителей»– остеобластов. У пожилых лиц, у 30%-80% женщин в менапаузе происходит выраженная декальцинация костей – остеопороз, при этом ухудшаются биохимические параметры костей. В мире остеопорозом болеет более 100 млн. человек. При длительном пребывании в постели потеря массы костной ткани достигает 30%. Развитие костной ткани в эмбриогенезе и в постнатальный период. У зародыша человека к началу органогенеза (3-5 недели) нет костной ткани. На месте будущих костей находятся остеогенные клетки или же хрящевые образования (гиалиновый хрящ). Это связано с тем, что развитие кости требует достаточного притока кислорода, а в этот период хорион (плацента) находится на стадии формирования и не создает условий для развития костной ткани. На 6 неделе эмбриогенеза эти условия создаются, и начинается развитие костной ткани, которое состоит из многочисленных этапов, включает как период эмбриогенеза, так и постнатальное развитие. Схема развития костной ткани (остеогенез). Развитие костной ткани человека. ↓ Источник развития: стволовые скелетогенные клетки Прямой остеогенез Непрямой остеогенез ↓ ↓ Формирование скелетогенного Формирование хондрогенного островка у зародыша: преобразование островка на месте будущей кос-ти. мезенхимных клеток в стволовые, а Преобразование мезенхим-ных клеток затем в остеобласты. У взрослого есть в стволовые, а затем в хондробласты источники для образования кости ↓ (стволовые клетки и предшественники Выработка хондробластами остеобластов). коллагеновых волокон, амор-фного ↓ вещества (гликоз-аминогликаны и Выработка остеобластами остеоида другие вещества). (коллагеновые волокна, аморфное ↓ органическое вещест-во). Формирование модели кости из ↓ гиалинового хряща. Синтез и выделение остеобластами ↓ фермента щелоч-ной фосфотазы и Разрушение хряща остеоклас-тами и пузырьков (кавеол) с минералами и развитие остеогенных островков с ферментами для минерализации остеобластами. волокон и матрикса. ↓ ↓ Выработка остеобластами осте-оида минерализация остеоида ↓ костным апатитом и образование Выделение остеобластами щелочной ретикулофиброзной костной ткани. фосфатазы и других веществ для минерализации ↓ Минерализация остеоида.Образование ретикулофиб-розной костной ткани. ↓ ↓ Преобразование ретикулофиброзной костной ткани в пластинчатую (преимущественно после рождения). Детализация развития костной ткани. Развитие костной ткани у эмбриона осуществляется двумя способами: непосредственно из мезенхимы (прямой остеогенез); и на месте ранее развившейся из мезенхимы хрящевой модели кости (непрямой остеогенез). Постэмбриональное развитие костной ткани происходит при физиологической регенерации. В процессе развития костной ткани образуется костный остеоцитарный дифферон: стволовые, полустволовые (преостеобласты), остеобласты, остеоциты. Прямой остеогенез характерен при формировании плоских костей ( например, костей черепа). Он наблюдается уже в первый месяц эмбриогенеза и включает три основные стадии: 1)формирование остеогенных островков; 2)дифференцировка клеток остеогенных островков и образование органического матрикса кости (остеоида); 3)обызвествление остеоида. 1.Формирование остеогенного островка происходит путем концентрации активно размножающихся клеток мезенхимы в участке развития будущей кости. 2.При второй стадии происходит дифференцировка клеток остеогенного островка и образование органического матрикса. Клетки мезенхимы внутри остеогенного островка прекращают делиться и начинают дифференцироваться в остеобласты. Остеобласты вырабатывают органический матрикс (остеоид), включающий коллагеновые фибриллы. В основном веществе появляются мукопротеиды (остеомукоид), цементирующие волокна в одну прочную массу. Постепенно клетки оказываются замурованными в межклеточном веществе, остеобласты теряют способность размножаться и превращаться в остеоциты. В то же время из окружающей мезенхимы образуются новые генерации остеобластов, которые наращивают кость снаружи (аппозиционный рост). 3.В третью стадию остеобласты выделяют фермент щелочную фосфатазу, которая расщепляет глицерофосфаты крови на углеводные соединения и фосфорную кислоту, последняя вступает в реакцию с солями кальция, который осаждается в основном веществе и в волокнах. В матриксе происходит дефосфорилирование. Одним из посредников кальцификации является остеонектин– гликопротеин, избирательно связывающий соли кальция и фосфора с коллагеном. В результате кальцификации образуются костные перекладины или балки. Формирование кости происходит, благодаря слиянию балок в единую сеть, промежутки которой заполнены волокнистой соединительной тканью с сосудами. Мезенхима вокруг формирующейся кости дает начало надкостнице. Сформировавшееся таким путем кость является ретикулофиброзной костной тканью и называется первичной губчатой костью. В дальнейшем эта кость в большинстве участков замещается пластинчатой костной тканью. Этот процесс иногда рассматривают как четвертую стадию остеогенеза. Развитие кости на месте ранее образованной хрящевой модели (непрямой остеогенез). Этот вид развития кости характерен для большинства костей скелета человека (длинные и короткие трубчатые кости, позвонки, кости таза). Первоначально формируется хрящевая модель будущей кости, которая служит основой для ее развития, а в дальнейшем она разрушается и замещается костью. Непрямой остеогенез начинается на втором месяце эмбрионального развития и заканчивается в среднем к 25 летнему возрасту. Непрямой остеогенез включает следующие стадии: 1)образование хрящевой модели кости; 2)образование перихондральной костной манжетки; 3)образование энхондральной кости в диафизе; 4)образование энхондральной кости в эпифизе; 5)формирование эпифизарной пластинки роста в хряще. Образование хрящевой модели кости происходит из мезенхимы в соответствии с закономерностями гистогенеза хряща. Модель хряща снаружи покрыта надхрящницей. Во вторую стадию в центре диафиза хряща, во внутреннем слое надхрящницы дифференцируются остеобласты, которые начинают продуцировать костное межклеточное вещество и формируют костную ткань поверх хряща под надхрящницей в виде манжетки (перихондральная кость). Перихондральная кость непрерывно утолщается и разрастается от центра диафиза к эпифизам. С момента появления в надхрящнице остеобластов, надхрящница заменяется надкостницей. Ретикулофиброзная костная ткань костной манжетки в дальнейшем заменится пластинчатой костной тканью. Образование костной манжетки нарушает питание хряща. Вследствиt этого в центре диафизарной части хрящевого зачатка возникают дистрофические изменения. Хондроциты вакуолизируются, их ядра подвергаются пикнозу, образуется так называемый пузырчатый хрящ с дегенеративными хондроцитами. Перихондральная костная манжетка увеличивается в длину, а зона деструкции хряща расширяется. Остеокласты формируют пути для врастающих кровеносных сосудов в образовавшиеся при деструкции хряща полости. На границе эпифиза и диафиза хондроциты собираются в колонки, так как рост неизмененных дистальных отделов хряща продолжается. В колонке хондроцитов идут два противоположно направленных процесса: размножение и рост хряща в его дистальном отделе и дистрофические изменения вблизи кости. В околокостной зоне между набухшими клетками хряща откладываются минеральные соли, которые придают базофилию межклеточному веществу. Кровеносные сосуды с окружающей мезенхимой, остеобластами, остеокластами врастают через отверстия в костной манжетке внутрь хряща и входят в соприкосновение с обызвествленным хрящом. Остеокласты выделяют ферменты, которые растворяют обызвествленное межклеточное вещество. Диафизарный хрящ разрушается, в нем возникают удлиненные пространства, в которых на поверхности оставшихся участков обызвествленного хряща остеобласты образуют костную ткань. Процесс отложения кости внутри хрящевого зачатка получил название энхондрального. При развитии энхондральной кости одновременно идет ее разрушение остеокластами и образование костно-мозговой полости, в которую врастает мезенхима и образуется строма костного мозга, в которой появляются стволовые клетки для развития костного мозга и кроветворных клеток. В это же время по периферии диафиза со стороны надкостницы нарастают новые перекладины костной ткани, образующиеся из надкостницы. Разрастаясь в длину по направлению к эпифизам и увеличиваясь в толщину, они образуют плотный слой кости. В дальнейшем в периостальной кости вокруг сосудов на месте разрушившейся грубоволокнистой кости начинают образовываться первичные остеоны. Они имеют широкий просвет, их границы слабо выражены. За образованием первой генерации остеонов со стороны периоста начинается развитие общих (генеральных) пластинок. Вслед за диафизом, центры окостенения появляются в эпифизах. Вначале в эпифизах идет дифференцировка хрящевых клеток, затем их гипертрофия, ухудшается питание, происходит дистрофия и кальцинация, затем отмечается процесс окостенения. Оссификация сопровождается врастанием в эпифизы сосудов. В промежуточной области между диафизом и эпифизом сохраняется хрящевая ткань в виде метаэпифизарного хряща, который является зоной роста костей в длину. В метаэпифизарном хряще различают пограничную зону, зону столбчатых клеток (размножение хондроцитов). Со временем в метаэпифизарной пластинке хряща процессы разрушения клеток начинают преобладать над процессом новообразования; хрящевая пластинка истончается и исчезает. Рост кости в длину прекращается. Рост трубчатых костей в толщину идет за счет периоста. Этот аппозиционный рост идет до окончания формирования кости. Количество остеонов после рождения невелико, но уже к 25 годам их число значительно увеличивается. Регенерация костной ткани. Физиологическая регенерация костных тканей и их обновление происходят медленно за счет остеогенных клеток надкостницы и остеогенных клеток в канале остеона. Посттравматическая регенерация (репаративная) протекает быстрее. Последовательность регенерации соответствует схеме остеогенеза. Процессу минерализации кости предшествует формирование органического субстрата (остеоида), в толще которого могут образоваться балки хряща (при нарушенном кровоснабжении). Оссификация в этом случае будет идти по типу непрямого остеогенеза (см. схему непрямого остеогенеза). 1. 2. 3. Вопросы для самоконтроля: Из какого источника развиваются хрящевые и костные ткани? Назовите функции надкостницы и надхрящницы? Из каких структурных элементов складываются хрящевая и костная ткани? 4. Что является структурно- функциональной единицей пластинчатой костной ткани? 5. Назовите структурные образования компактного вещества трубчатой кости? 6. Какие клетки костной ткани принимают участие в ее построении и разрушении? Укажите их строение. 7. Какие способы остеогенеза вам известны и какие стадии в них различают? 8. Как изменяются с возрастом хрящевые и костные ткани? Ситуационные задачи. 1. В костной ткани обнаружены многоядерные клетки, содержащие много лизосом. Как называются эти клетки, назовите их происхождение и функции. 2. В трубчатой кости, между остеонами расположены костные пластинки, не образующие остеонов. Откуда происходят эти пластинки? 3. В диете ребенка дефицит солей кальция. Как при этом будет развиваться костная ткань? 4. У ребенка, который страдает рахитом при осмотре выявлено искривление костей конечностей. Какой этап образования костной ткани нарушен? 5. С чем связана легкость переломов у пожилых и редкость их у детей? 5. ТЕМА: МЫШЕЧНЫЕ ТКАНИ Цели занятия. Научиться: 1.Определять на светооптическом уровне исчерченную, неисчерченную(гладкую),сердечную мышечные ткани. 2.Узнавать и анализировать отличия разных видов мышечной ткани. 3.Анализировать на электронно-оптическом уровне структуру мышечного волокна, гладкой мышечной клетки, кардиомиоцитов. Методические рекомендации к изучению темы: повторить общие вопросы раздела «Миология» из анатомии человека и ответить на следующие вопросы: 1.Что является источником формирования исчерченной мускулатуры органов опоры и движения? 2. Каково функциональное назначение скелетных мышц? 3. Принципы работы мышц. 4.Строение скелетной мышцы как органа. 5.Из каких мышц построены стенки полых внутренних органов? Имея разное происхождение и строение, мышечные ткани объединяет способность к сокращению. Сократительный аппарат занимает значительную часть в цитоплазме, в его составе присутствуют актиновые и миозиновые филаменты, которые формируют органеллы специального значения–миофибриллы. По морфофункциональному признаку различают: Скелетную или поперечно-полосатую или исчерченную мышечную ткань. Начало и прикрепление мышц находится на скелете. Мышцы являются произвольными, поскольку их сокращения подчиняются нашей воле. К этой группе мышц относят скелетные мышцы, мышцы языка, гортани и др. Сердечная мышечная ткань входит в состав мышечной стенки сердца. Иннервируется вегетативными нервами, сокращения не подчиняются нашей воле. Гладкая (неисчерченная) мышечная ткань характеризуется отсутствием исчерченности, а поскольку сокращения также не подчиняются нашей воле, то мышцы называют непроизвольными. Иннервация осуществляется вегетативной нервной системой. Из гладких мышц построены стенки внутренних органов, стенка сосудов. В зависимости от источников развития выделяют пять типов мышечной ткани: 1. Мезенхимного происхождения (гладкая мышечная ткань). 2.Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки (например, в потовых, слюнных железах). 3.Нейральное происхождение (из нервной трубки)–мышцы суживающие и расширяющие зрачок. 4.Целомическое происхождение (миоэпикардиальная пластинка) – сердечная мышечная ткань. 5. Из миотомов мезодермы– исчерченная мышечная ткань. Исчерченная (поперечно-полосатая) мышечная ткань Источником развития являются клетки миотомов миобласты. Различают головные, шейные, грудные, поясничные, крестцовые миотомы. Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов. Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие, с 3 недели внутриутробного развития мигрируют в мезенхиму и сливаясь, друг с другом образуют мышечные трубки (миотубы) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают. Гранулярная ЭПС значительно редуцируется. Такая многоядерная структура называется симпласт, а для мышечной ткани - миосимпласт. Часть миобластов дифференцируется в миосателлитоциты, которые располагаются на поверхности миосимпластов и принимают участие в регенерации мышечной ткани. Структурной единицей мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной (рис.21). Длина мышечных волокон колеблется от 1 до 40 мм, а толщина 0,1 мм. В мышечном волокне различают мембранный аппарат, фибриллярный (сократительный) аппарат, трофический аппарат (ядро, саркоплазма, цитоплазматические органеллы). Мембранный аппарат. Каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), плазмолемма образует впячивания (Т- трубочки). К плазмолемме снаружи прилежат миосателлитоциты. При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов. Фибриллярный аппарат. Исчерченные волокна можно разделить на составляющие их фибриллы (диаметром 1 мкм), названные миофибриллами. В мышечном волокне они ориентированы продольно. При рассматривании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками или Адисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными или I – дисками. В средней части диска А имеется более светлый участок Нзона (участок содержащий только толстые нити белка миозина). В области Н-зоны выделяется более темная М-линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z, которая построена из белковых фибриллярных молекул. В частности, большую роль играет альфа актинин. Z – линия соединена с соседними миофибриллами с помощью белка десмина и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечно-полосатой исчерченности мышечного волокна. Структурной единицей миофибриллы является саркомер (S) - это пучок миофиламентов заключенный между двумя Z линиями (рис.22). Принимая во внимание вышеуказанные обозначения можно структуру саркомера записать в виде формулы: S= Z1 + 1/2 I1 + А + 1/2 I2 + Z2 Под электронным микроскопом миофибриллы представляют агрегаты из толстых (меозиновых) филаментов ( диаметр 14 нм, длина 1500 нм, расстояние между ними 20-30 нм). Между толстыми филаментами располагаются тонкие филаменты ( диаметр 7-8 нм). Толстые филаменты (миозиновые нити) состоят из молекул белка миозина. Он является важнейшим сократительным белком мышцы. При непосредственном участии миозина химическая энергия трансформируется в механическую работу. Каждая миозиновая нить состоит из 300-400 молекул миозина. Молекула миозина – это гексамер, состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах глобулярные (шаровидные) головки. Между головкой и тяжелой цепью – шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити. Тяжелые и легкие цепи в молекуле миозина можно разделить обработкой мочевиной, гуанидинхлоридом и др. При мягкой обработке можно отделить только легкие цепи. Миозину свойственна АТФ-азная активность – высвобождающаяся энергия используется для мышечного сокращения. Тонкие нити (актиновые нити). Образованы тремя белками: актином, тропонином и тропомиозином. Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали. Толстые нити занимают центральную часть саркомера–А-диск, тонкие занимают I диски и частично входят между толстыми миофиламентами. Только толстые нити содержит Н-зона. При поступлении нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна. Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров, располагающихся в прилежащей рыхлой соединительной ткани. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном называется нервномышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон). В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие участки актина заблокированы тропомиозином. При высокой концентрации ионов кальция конформационные (пространственные) изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина. Плазмолемма миосимпласта образует пальцевидные впячивания (инвагинации) ориентированные поперечно по отношению к миосимпласту называемые Т-трубочки. К каждой Т-трубочке примыкают по две цистерны саркоплазматического ретикулума (гладкая ЭПС), образуя триаду: две цистерны и одна Т-трубочка. Са2+ концентрируется в цистернах ( там его концентрация в 800 раз больше, чем в саркоплазме). Механизм сокращения. При поступлении нервного импульса волна деполяризации доходит до цистерн саркоплазматического ретикулума, из них выделяются ионы кальция и концентрация кальция в саркоплазме резко возрастает. Са2+ диффундирует к тонким нитям (филаментам) саркомера, где связывается с тропонином и миозиновыми головками. Это приводит: 1.К изменению конформации (пространственного расположения) тропомиозина, что, в свою очередь, приводит к освобождению участков актина, необходимых для взаимодействия с миозиновыми головками. 2.Появлению АТФ-азной активности миозина. 3.Взаимодействию миозиновых головок с актином (актино- миозиновые «мостики»). Все это вместе взятое приводит к тому, что миозиновые головки «шагают» по актину, образуя в ходе перемещения новые связи актина и миозина, сближая две Z-линии. При сокращении уменьшаются только светлые диски. Расслабление. Са2+-АТФ-аза саркоплазматического ретикулума закачивает Са2+ из саркоплазмы в цистерны. В саркоплазме концентрация Са2+ становится низкой. Са2+тропомиозин закрывает миозин-связывающие участки тонких нитей и препятствует их взаимодействию с миозином. Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон изменяется натяжение соединительно-тканной капсулы веретена и соответственно изменяется тонус интрафузальных (расположенных под капсулой) мышечных волокон. Формируется нервный импульс. Классификация и типы мышечных волокон. Скелетные мышцы, состоящие из мышечных волокон отличаются по многим параметрам: цвету, диаметру, утомляемости, скорости сокращения и т.д. В каждой мышце присутствуют разные типы мышечных волокон. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные, которые преобладают и обуславливают собственно сократительную функцию мышцы и интрафузальные, входящие в состав проприоцепторов–нервномышечных веретен. По характеру сокращения мышечные волокона делят на фазные и тонические. Фазные способны осуществлять быстрые сокращения, но не могут длительно удерживать достигнутый уровень укорочения. Тонические –обеспечивают поддержание статического напряжения или тонуса. По биохимическим особенностям и цвету выделяют красные и белые мышечные волокна. Цвет мышцы обусловлен степенью васкуляризации. Кроме того, существует прямая корреляция между содержанием миоглобина и цветом мышцы. Характерной особенностью красных мышечных волокон является наличие многочисленных митохондрий, цепи которых располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше и они располагаются равномерно в саркоплазме мышечного волокна. Скорость сокращения определяется типом миозина. Высокую скорость сокращения обеспечивает быстрый миозин (для него характерна высокая активность АТФ-азы); меньшая скорость сокращения характерна для медленного миозина (характерна невысокая активность АТФ-азы). Следовательно, по активности АТФ-азы можно судить и о наборе миозинов. Тип окислительного обмена. Мышечные волокна используют два пути образования АТФ: *при анаэробном типе метаболизма из 1 молекулы глюкозы образуется 2 молекулы АТФ и молочная кислота. *при аэробном окислении из 1 молекулы глюкозы образуется 38 молекул АТФ и конечные продукты метаболизма: СО2 и Н2О. Идентификация мышечных волокон основана на выявлении активности фермента сукцинатдегидрогеназы (СДГ), которая является маркером для митохондрий и цикла Кребса. Активность этого фермента свидетельствует о напряженности энергетического метаболизма. Выделяют мышечные волокна А-типа (гликолитические) с низкой активностью СДГ, С-тип (оксидативные) с высокой активностью СДГ. Мышечные волокна В-типа занимают промежуточное положение. Переход мышечных волоко от А-типа в С-тип маркирует изменения от анаеробного гликолиза к метаболизму, зависящему от кислорода. Существует много и других классификаций. Факторами, определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное влияние, уровень васкуляризации, уровень двигательной активности и местоположение мышцы. Сердечная мышечная ткань находится в мышечной оболочке сердца (миокард) и в устьях связанных с ним крупных сосудов. Имеет клеточный тип строения и основным функциональным свойством служит способность к спонтанным ритмическим сокращениям. Развивается из миоэпикардиальной пластинки (висцеральный листок спланхнотома в шейном отделе), клетки которой размножаются митозом а потом дифференцируются. В клетках появляются миофиламенты, которые далее формируют миофибриллы. Сердечная мышечная ткань образована клетками, которые называются кардиомиоциты. Они расположены между элементами рыхлой волокнистой соединительной ткани, кровеносными капиллярами. Кардиомиоциты связаны друг с другом в области вставочных дисков. Последние являются комплексом контактов. На поперечном срезе вставочных дисков выявляют десмосомы и щелевидные контакты (нексусы). Выделяют рабочие (сократительные) кардиомиоциты, которые образуют цепочки клеток и обеспечивают силу сокращения сердечной мышцы. Клетки удлиненной формы с центрально расположенным ядром (рис.23). Вблизи ядра (или двух) комплекс Гольджи и гранулы гликогена. Между миофибриллами лежат многочисленные митохондрии. Имеются Т-трубочки и L-трубочки. Десмосомы обеспечивают механическое сцепление, которое препятствует расхождению кардиомиоцитов. Щелевидные контакты способствует передаче сокращения от одного кардиомиоцита к другому. Проводящие (атипичные) кардиомиоциты–среди них различают: 1.Водители ритма–это клетки небольших размеров, в саркоплазме мало гликогена, мало миофибрилл и они расположены по периферии. Клетки имеют хорошее кровоснабжение и иннервацию. Они воспринимают сигналы от нервных окончаний и способны автоматически генерировать сигналы обеспечивающие ритмические сокращения сердца. 2.Проводящие (переходные) кардиомиоциты проводят возбуждение от водителя ритма. Образуют длинные волокна. Миофибриллы в небольшом количестве, имеют спиральный ход, мелкие митохондрии, немного гликогена. 3.Волокна Пуркинье–являются самыми крупными клетками в мышечной ткани сердца с неупорядоченным расположением миофибрилл, множеством мелких митохондрий, много гликогена, нет Т-трубочек, клетки связаны между собой десмосомами и щелевидными контактами. Секреторные кардиомиоциты – находятся в, основном, в предсердиях, преимущественно в правом. Характеризуются отростчатой формой и слабым развитием сократительного аппарата. В саркоплазме, вблизи полюсов ядра-секреторные гранулы, содержащие атриопептин (гормон, регулирующий артериальное давление). Гормон вызывает потерю натрия и воды с мочой, расширение сосудов, снижение давления, угнетение секреции альдостерона, кортизола, вазопрессина. Сократительный аппарат рабочих кардиомиоцитов сходен со скелетными мышечными волокнами. Миофибриллы в кардиомиоците могут объединяться в комплексы, образуя единые сократительные структуры. В саркоплазме миофибриллы ориентированы продольно и располагаются преимущественно по периферии. Саркотубулярная система вцелом имеет сходство с исчерченными мышечными волокнами. Саркоплазматическая сеть развита слабее, не так активно накапливает Са2.. При расслаблении, ионы кальция выделяются в саркоплазму с низкой скоростью, что обеспечивает автоматизм и частые сокращения кардиомиоцитов. Т-трубочки широкие и образуют диады (одна Т-трубочка и одна цистерна сети), которые сходятся в области Zлинии. Энергетический аппарат-это митохондрии и включения. Неисчерченная (гладкая) мышечная ткань. Структурно-функциональной единицей данной ткани являются гладкие мышечные клетки ( ГМК), которые способны к гипертрофии и регенерации. Они образуют стенки внутренних полых органов, сосудов. Более крупные по размерам ГМК характерны для стенок полых внутренних органов, а меньших размеров–для стенки сосудов. Клетки контролируют моторику, величину просвета. Имеют веретенообразную форму, в центре палочковидной формы ядро. В ГМК отсутствует поперечно-полосатая исчерченность. ГМК окружены сарколеммой, которая снаружи покрыта базальной мембраной. Длина от 20 мкм до 1 мм. В саркоплазме у полюсов находится комплекс Гольджи, много митохондрий, рибосом, развит саркоплазматический ретикулум. Миофиламенты расположены вдоль продольной оси. В ГМК актиновые и миозиновые филаменты не формируют миофибрилл. Актиновые нити (тонкие филаменты) ориентированы по продольной оси ГМК. По количеству их больше и они прикрепляются к плотным тельцам, которые являются специальными сшивающими белками. Рядом с актиновыми нитями располагаются мономеры миозина (микромиозин). Обладая разной длиной они, значительно короче тонких нитей. Сокращение гладких мышечных клеток осуществляется при взаимодействии актиновых филаментов и миозина. Сигнал идущий по нервным волокнам обуславливает выделение медиатора, что изменяет состояние сарколеммы. Она образует колбовидные впячивания (кавеолы), где концентрируются ионы кальция. Сокращение ГМК индуцируется притоком ионов кальция в саркоплазму (кавеолы отшнуровываются и вместе с ионами кальция попадают в саркоплазму). Это приводит к полимеризации миозина и взаимодействию его с актином. Актиновые нити и плотные тельца сближаются, усилие передается на сарколемму и ГМК укорачивается. Миозин ГМК способен взаимодействовать с актином только после фосфорилирования его легких цепей особым ферментом–киназой легких цепей. После прекращения сигнала ионы кальция покидают кавеолы; миозин деполяризуется, теряет сродство к актину. В результате комплексы миофиламентов распадаются; сокращение прекращается. Особые типы гладких мышечных клеток. Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности. Окружают секреторные отделы и выводные протоки желез (слюнных, молочных, слезных). С железистыми клетками они связаны десмосомами. Сокращаясь, способствуют выделению секрета. В концевых (секреторных) отделах форма клеток отросчатая, звездчатая. Ядро в центре, в цитоплазме, преимущественно в отростках локализованы миофиламенты, которые образуют сократительный аппарат. В этих клетках есть и цитокератиновые промежуточные филаменты, что подчеркивает их сходство с эпителиоцитами. Мионейральные клетки–развиваются из клеток наружного слоя глазного бокала и образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок. По строению первая мышца сходна с ГМК мезенхимного происхождения. Мышца, расширяющая зрачок образована отростками клеток, располагающимися радиально, а ядросодержащая часть клетки находится между пигментным эпителием и стромой радужки. Миофибробласты относятся к рыхлой соединительной ткани и представляют собой видоизмененные фибробласты. Они проявляют свойства как фибробластов, так и ГМК (обладают выраженными сократительными свойствами). Как вариант этих клеток можно рассматривать миоидные клетки в составе стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника. При заживлении раны часть фибробластов синтезирует гладкомышечные актины и миозины. Эндокринные гладкие миоциты–это видоизмененные ГМК, представляющие основной компонент юкста-гломерулярного аппарата почек. Они находятся в стенке артериол почечного тельца, имеют хорошо развитый синтетический аппарат и редуцированный сократительный. Продуцируют фермент ренин, находящийся в гранулах и попадающий в кровь механизмом экзоцитоза. Вопросы для самоконтроля: 1.Как классифицируются мышечные ткани по морфо-функциональному признаку? по происхождению? 2.Что является структурно-функциональной единицей мышечной ткани? 3.Строение фибриллярного аппарата мышечных волокон. 4.Напишите формулу саркомера. 5.Строение мышечных волокон под световым и электронным микроскопом. 6.Механизм сокращения и расслабления мышечного волокна. 7.Как классифицируются мышечные волокна? Типы мышечных волокон? 8.Виды кардиомиоцитов в сердечной мышце, особенности их строения. 9.Строение ГМК. 10.Перечислить особые типы гладких мышечных клеток. Ситуационные задачи. 1. При обработке экспериментального материала (одна из скелетных мышц крысы) выявлено по активности СДГ три типа мышечных волокон: А, В, С. Какой тип метаболизма характерен для каждого из этих мышечных волокон? 2. Какие наследственные и ненаследственные заболевания мышц Вы знаете? 3. Как бы Вы обьяснили механизмы, происходящие при нарушении питания участка сердечной мышцы? 4.В судебном морге на медицинскую экспертизу представлены препараты исчерченной мышечной ткани. В одном препарате выявлены симпласты с ядрами расположенными по периферии, а в другом–клетки с центрально ориентированными ядрами. Каким органам принадлежат выявленные мышечные ткани? 5.У больного инфаркт миокарда, повреждена сердечная мышца. Какие клетки обеспечат восстановление дефекта в сердечной мышце? 6. ТЕМА: НЕРВНАЯ ТКАНЬ Цели занятия: 1. Обобщить изученный материал, выделяя наиболее важные аспекты темы. 2. Научиться узнавать на светооптическом уровне структуры нервной ткани (нейроны, нервные волокна, нейроглию).Анализировать их строение. 3. Анализировать на электронно-оптическом уровне структуру нейронов, синапсов, нервных волокон. Повторите вопросы из предшествующих тем: 1. Цитоскелет клетки. Значение микротрубочек в транспортных процессах в клетке. 2. Рецепторы клетки. 3. Облегченная диффузия и активный транспорт. Калий-натриевый насос. 4. Механизмы экзоцитоза. 5. Межклеточные контакты. 6. Строение и значение митохондрий, гранулярной эндоплазматической сети, пластинчатого комплекса. 7. Понятие о стабильной клеточной популяции. 8. Механизмы миграции клеток. 9. Макрофаги и макрофагическая система. Общая характеристика нервной ткани. Нервная ткань (рис.24) выполняет функции восприятия, проведения и передачи воздуждения, полученнного из внешней среды и внутренних органов, а также анализ, сохранение полученной информации, интеграцию органов и систем, взаимодействие организма с внешней средой. Основными структурными элементами нервной ткани являются нейроны с их отростками и нейроглия. Нейроглия в свою очередь делится на микроглию и макроглию. Макроглия центральной нервной системы подразделяется на следующие клетки: астроциты (волокнистые и протоплазматические), олигодендроциты и эпендимоциты. Макроглия периферической нервной системы включает в себя сателлитоциты и леммоциты. Нейроны, выполняют основные функции, приписываемые нервной ткани. Они состоят из тела (перикариона) и отростков. При этом последние могут многократно превышать объем тела нервной клетки. Среди отростков выделяют дендриты и аксон (нейрит). Дендриты сильно ветвятся, образуя дендритное дерево, и обычно короче аксона. От дендритов возбуждение направляется к телу нервной клетки. Они формируют постсинаптические структуры, воспринимающие возбуждение. Дендритов много, но может быть один. Аксон присутствует всегда, по одному на каждую нервную клетку. Он не ветвится или слабо ветвится в терминальных областях и заканчивается синаптическим бутоном, передающим возбуждение на другие клетки (пресинаптическая зона). Нейроны передают возбуждение с помощью специализированных контактов (синапсов). Вещество, обеспечивающее передачу возбуждения, называется медиатором. В каждом нейроне обычно обнаруживается один основной медиатор. Направление импульса Структурно-функциональная схема нейрона. аксон Классификация нейронов. Существует несколько классификаций нервных клеток. перикарион В последние годы предпринимаются попытки создать единую комплексную Терминаль аксона классификацию. Морфологическая классификация нейронов в соответствии с числом и типом отростков выделяет): мультиполярные - с множеством отростков, униполярные - с одним аксоном , холмик Аксонный Дендритное дерево ложно- (псевдо-) униполярные – дендрит и аксон отходят от нейрона в виде одного отростка, а затем разделяются . биполярные - с одним аксоном и одним дендритом . Морфологические классификации нейронов позволяют выделять и другие признаки клеток. Так, имеется классификация нервных клеток по размерам. Они делятся на гигантоклеточные, крупноклеточные, среднеклеточные, мелкоклеточные. По длине нейритов можно видеть длинноаксонные, среднеаксонные, короткоаксонные. По степени развития дендритного дерева мультиполярные нейроны бывают густоветвистые, средневетвистые и маловетвистые (изодендритные). Кроме этого можно подразделить нервные клетки по форме (звездчатые, округлые, грушевидные, пирамидные, веретеновидные, корзинчатые и т.д.), по медиатору характерному для них (адренэргические, холинэргические, глицинэргические, дофаминергические и т.д.). Мультиполярный нейрон Биполярный нейрон Псевдоуниполярный нейрон Униполярный нейрон Морфологические типы нейронов Функциональная классификация нейронов позволяет выделить среди них: Афферентные нейроны (воспринимают информацию и передают ее в нервные центры). Их можно рассматривать как клетки, осуществляющие центростремительную передачу возбуждения. Данные клетки могут рассматриваться как чувствительные нейроны. Но, если понимать выражение чувствительность, в смысле того, что принимается и анализируется ЦНС, то не все клетки этой группы имеют в ней представительство. Так, афферентные нейроны интрамуральных (внутриорганных) нервных узлов ограничивают передачу возбуждения другими нервными клетками узла. Типичными чувствительными являются ложноуниполярные и биполярные нейроны спинномозговых и черепно-мозговых узлов. Ассоциативные нейроны, осуществляющие взаимодействие между нейронами. Являются основной популяцией нейронов в центральной нервной системе и по численности составляют абсолютное большинство. Типичным для них является мультиполярность, средние или мелкие размеры перикарионов. Эфферентные (центробежные) нейроны генерируют нервный импульс и передают возбуждение другим нейронам или клеткам других видов тканей. Значительная часть этих клеток контролируют мышцы и, таким образом, являются двигательными, обеспечивая произвольные и непроизвольные сокращения. Другие клетки формируют окончания на телах секреторных клеток и регулируют инкреторные, секреторные и экскреторные функции. Некоторые из нейронов направляют аксоны к кровеносным капиллярам выделяют инкрет в кровь, являясь нейросекреторными. Среди функциональных типов нейронов выделяются возбуждающие, которые вызывают деполяризацию постсинаптических мембран клеток приводя к формированию так называемого потенциала действия, вызывающего стимулирующие эффекты. Тормозные нейроны приводят к гиперполяризации мембраны контролируемой клетки, снижая ее чувствительность к специфическим воздействиям. В этой связи, необходимо упомянуть о так называемых пейсмекерных нейронах. Эти клетки способны к автономному формированию возбуждения, независимо от внешних воздействий. Эти клетки характеризуются снижением мембранного потенциала покоя до порогового уровня даже без значительных внешних влияний или под действием возвратного возбуждения, возникающего в цепи нейронов. В последнем случае сигнал возвращается к возбуждающей клетке через цепь вставочных нейронов, что приводит к повторному формированию возбуждения по всей цепи. Часто клетка не может поддерживать постоянный потенциал покоя, и он опускается до минимального предела (порогового уровня) и происходит деполяризация мембраны с возбуждением нейрона. Регенераторные процессы. Важной особенностью основной популяции зрелых нейронов является способность лишь к внутриклеточной регенерации. Они являются стабильной популяцией клеток и в обычных условиях не делятся. Но имеются исключения. Так, доказана способность к делению у нервных клеток в эпителии обонятельного анализатора, в ганглиях (скоплениях нейронов периферической нервной системы), контролирующих звуковую активность певчих птиц (g.vocalae). Некоторые авторы приписывают подобные возможности части интрамуральных (внутриорганных) вегетативных ганглиев (во всяком случае доказана их способность к неполному амитозу и митозу с формированием многоядерных полиплоидных нейронов). Предполагают возможность деления нейронов при наследственно передающемся заболевании старческого слабоумия (болезни Альцгеймера) и даже деление нейронов в пределах субвентрикулярных зон (вблизи желудочков мозга). Но вцелом, деление нервных клеток, как уже упоминалось, невозможно. Невозможность деления нейронов необходима для сохранения стабильности важнейших функциональных и макроанатомических межнейронных взаимодействий, что, в частности, позволяет нам сохранять индивидуальные особенности личности. Невозможность клеточного деления, вовсе не предполагает неспособности нервных клеток к регенерации вообще. Нервные клетки на внутриклеточном уровне не только регенерируют, но характеризуются очень высокой физиологической внутриклеточной регенераторной способностью. Непрерывно происходит замещение старых органелл, белковых, липопротеидных и гликопротеидных комплексов (в частности, мембран), ресинтез ферментов, медиаторов и т.д. Это сопровождается высоким уровнем энергетического обмена, особенно в теле нейрона. В культуре нервной ткани нейроны активно делятся митозом. Строение нейрона. На светооптическом уровне при общих методах окрашивания тела нервных клеток имеют оксифильную цитоплазму, крупное ядро округлой или овальной формы. Ядро занимает центральное положение, но иногда смещается к одному из полюсов нейрона, что чаще всего связано с реактивными процессами. В ядре хорошо развито одно или несколько ядрышек. В части нейронов можно видеть два и более ядер (до 10-15). Это более характерно для вегетативных узлов, особенно встроенных в структуру внутренних органов (внутриорганные или интрамуральные ганглии). Такие многоядерные клетки по сути являются редуцированными проявлениями деления не завершающихся полноценным цитокинезом. Следует иметь в виду, что у некоторых разновидностей нервных клеток можно видеть полиплоидный набор хромосом, что связано с эндомитозом. Полиплоидия облегчает высокую метаболическую активность нейронов. Кариоплазма отличается преобладанием диффузного (слабо конденсированного) хроматина. Нейроны имеют высокое сродство к солям серебра (аргирофильность). Специфичными для нейрона структурами цитоплазмы на светооптическом уровне являются хроматофильное вещество цитоплазмы и нейрофибриллы. Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при окрашивании нервных клеток основными красителями (метиленовым синим, толуидиновым синим, гематоксилином и т.д.) в виде зернистости. Зернистость может быть в виде крупных глыбок неправильной формы, а также в виде сети или мелкой зернистости. Это зависит от типа нейрона (крупные нейроны обычно имеют более крупные глыбки) и от его функционального состояния. На электроннооптическом уровне хроматофильное вещество цитоплазмы есть не что иное, как скопления цистерн зернистой эндоплазматической сети. Эти органеллы отсутствуют в аксоне и в аксонном холмике, но имеются в начальных сегментах дендритов. На светооптическом уровне также тигроид не виден в начале аксона, но прослеживается в дендритах, что позволяет идентифицировать вид отростков. Процесс разрушения или распада глыбок базофильного вещества называется тигролизом и наблюдается при реактивных изменениях нейронов (например, при их повреждении) или при их дегенерации. Тигролиз нередко сопровождается вакуолизацией цитоплазмы, Уплощенные цистерны ЭПС разбухают, а цитоплазма приобретает вакуолярную орагнизацию. Нейрофибриллы, выявляют при импрегнации нейронов, в результате осаждения осадка серебра на структурах цитоскелета нейрона. Основой для выявления нейрофибрилл являются нейрофиламенты и нейротубулы, формирующие каркас нервной клетки. Кроме этого, в нейронах довольно часто можно видеть липидные включения (зерна липофусцина). Они характерны для старческого возраста и часто появляются при дистрофических процессах. Зерна липофусцина являются остаточными тельцами, возникающими в результате неполного переваривания. Их накопление может приводить к нарушению нормальных метаболических процессов в клетках и их гибели. У некоторых нейронов в норме обнаруживаются пигментные включения (например, с меланином), что обуславливает окрашивание нервных центров, содержащих подобные клетки (черная субстанция, голубоватое пятно). Электроннооптическое строение и некоторые цитофизиологические особенности тела нейрона и его отростков. Несмотря на крайнее морфологическое разнообразие нейронов, они имеют ряд общих черт построения. Ядра нейронов, особенно крупноклеточных, имеют округлую форму. Кариолемма часто формирует впячивания, что может значительно увеличивать площадь контакта поверхности ядра с цитоплазмой (нейроплазмой). Ядерная оболочка имеет большое количество ядерных пор, что указывает на активные процессы обмена, в том числе РНК и субъединицами рибосом. Кариоплазма в крупных нейронах в основном светлая, но особенно в мелких, можно видеть и повышенную склонность к импрегнации (осмиофильность). Данные особенности на светооптическом уровне проявляются в гипохромности или гиперхромности ядер (т.е., пониженной или повышенной склонности к окрашиванию ядерными красителями). Хорошо развит ядрышковый аппарат. В ядре обычно имеется 1-2 крупных умеренной плотности ядрышка, занимающих центральное положение. В мелких нервных клетках ядрышки мельче, их может быть до 3-6 и более. При реактивных проявлениях в клетке можно наблюдать смещение ядрышка и его распад. Матрикс цитоплазмы (нейроплазмы) гомогенный или мелкозернистый, слабой или умеренной электронной плотности. В нейроне сильно развита гранулярная эндоплазматическая сеть (ЭПС), представленная скоплениями или диффузно расположенными плоскими цистернами и трубочками. Как уже указывалось выше, ЭПС преобладает в теле, и может содержаться в начальных сегментах дендритов. За ней закреплено участие в процессах синтеза медиаторов и модуляторов, мембранных белков и т.д. Кроме связанных имеется и значительное число свободных полисом и рибосом. Митохондрии в нейронах многочисленны. Они имеют средние и крупные размеры (диаметр 1-3 мкм), могут быть овальной или нитчатой формы. Кристы имеют трабекулярное строение. Нейроны в энергетическом отношении крайне зависимы от аэробного фосфорилирования и во взрослом состоянии фактически не способы к анаэробному гликолизу. В связи с этим нервные клетки находятся в выраженной зависимости от поступления кислорода и глюкозы и при нарушении кровотока нервные клетки практически сразу прекращают свою жизнедеятельность. Момент прекращения кровотока в головном мозге означает начало клинической смерти. Практически сразу же начинаются процессы саморазрушения в нейронах, и прекращается их специфическая функциональная активность. Их мембраны деполяризуются. Митохондрии, ЭПС, ядерные оболочки набухают, а затем и разрушаются. Начинаются процессы аутолиза и перекисного окисления. Эти процессы могут носить обратимый или необратимый характер. При мгновенной смерти, при комнатной температуре, и нормальной температуре тела процессы саморазрушения в нейронах обратимы в течение 5-7 минут. Это и является сроком так называемой клинической смерти, когда возможно оживление организма. Необратимые изменения в нервной ткани приводят к переходу от клинической смерти к биологической. В случае продолжительного умирания, высокой внутренней или внешней температуре, особенно при сердечно-сосудистой и дыхательной недостаточности время необратимого разрушения нервных клеток значительно сокращается до 1-2 и менее минут. Наоборот, внезапная остановка сердечной деятельности при низких температурах окружающей среды и головного мозга может значительно удлинять время клинической смерти. В нейронах значительного развития достигает пластинчатый комплекс. Он может располагаться компактно или быть рассеян в цитоплазме тела нейрона. Нередко наблюдается поляризация этой органеллы. Это указывает на преимущественное направление транспортных потоков и процессов созревания секретируемых продуктов (медиаторов) от тела к аксону. Специфическими органеллами нейрона считают нейрофиламенты и нейротубулы. Нейрофиламенты представляют собой промежуточные филаменты диаметром 8-10 нм, образованные фибриллярными белками. Основной функцией этих элементов цитоскелета является опорная - для обеспечение стабильной формы нейрона. Подобную же роль играют тонкие микрофиламенты (поперечный диаметр 6-8 нм), содержащие белки актины. В отличие от подобных микрофиламентов в других тканях и клетках, они не соединяются с микромиозинами, что делает невозможным активные сократительные функции в зрелых нервных клетках. Нейротубулы по основным принципам своего строения фактически не отличаются от микротрубочек. Они, как и все микротрубочки имеют поперечный диаметр около 24 нм, кольца замыкают 13 молекул глобулярного белка тубулина. Микротрубки полярны. В нервной ткани они выполняют очень важную, если не сказать уникальную роль. Как и всюду они несут каркасную (опорную) функцию. В отличие от нейрофиламентов являются менее стабильными и обеспечивают процессы циклоза. Именно полярность микротрубки, в которой имеется отрицательно и положительно заряженные концы, позволяет контролировать диффузионно-транспортные потоки в аксоне (так называемый быстрый и медленный аксоток). Их подробное описание приведем несколько ниже. В цитоплазме тел нейронов часто встречаются лизосомы. Они участвуют в пластических процессах, осуществляя катаболизм (разрушение) старых органелл и структур. В результате переваривания образуются остаточные тельца. Часть из них содержит непереваренные остатки органелл, включающие липофусцин. Избыточное накопление липофусцина может приводить к дистрофическим процессам в нейроне, к нарушению его специфической активности и даже гибели. Такие явления характерны для старческих изменений и при различных патологических воздействиях. В теле нейронов можно видеть также транспортные пузырьки, часть из которых содержит медиаторы и модуляторы. Они окружены мембраной. Их размеры и строение зависят от содержания того или иного вещества. Особенности морфологии отростков нейронов. Дендриты короткие отростки, нередко сильно ветвятся. Их ветвления более выражены в терминальных областях. Дендриты в начальных сегментах содержат органеллы подобно телу нейрона. Хорошо развит цитоскелет. Аксон чаще всего длинный, слабо ветвится или не ветвится. Ветвления чаще всего наблюдаются в терминальных зонах. Уже в начальном сегменте аксона, в нем отсутствует гранулярная ЭПС. Микротрубочки и микрофиламенты располагаются упорядочено. В цитоплазме аксона видны митохондрии, транспортные пузырьки. Аксоны в основном миелинизированы и окружены отростками олигодендроцитов в центральной нервной системе, или леммоцитами в периферической нервной системе. Начальный сегмент аксона нередко расширен и имеет название аксонного холмика. Именно в зоне аксонного холмика происходит суммация поступающих в нервную клетку сигналов и если возбуждающие сигналы достаточной интенсивности, то в аксоне формируется потенциал действия и возбуждение направляется вдоль аксона, передаваясь на другие клетки (потенциал действия). От отростков нейронов, а нередко и от его тела, отходят небольшие выпячивания, которые имеют форму напоминающую шипики, откуда и получили название. Особенно развиты шипики на некоторых нервных клетках центральной нервной системы. Шипики являются постсинаптическими структурами и соответствуют зонам взаимодействия одних нервных клеток с другими. Они имеют элементы цитоскелета, митохондрии. Нередко видны уплощенные цистерны и электронно-плотное вещество мембраны. Аксоток (аксоплазматический транспорт веществ). Нервные волокна имеют своеобразный структурный аппарат– микротрубочки, по которым перемещаются вещества от тела клетки на периферию (антероградный аксоток) и от периферии к центру (ретроградный аксоток). Направление аксотока обеспечивает полярность микротрубочек. В нем участвует белок кинезин, взаимодействующий с тубулином микротрубочек, и осуществляющий транспорт с затратой энергии АТФ. Различают быстрый (со скоростью 100-1000 мм/сут.) и медленный (со скоростью 110 мм/сут.) аксоток. Быстрый аксоток - одинаков для различных волокон; требует значительной концентрации АТФ; происходит с участием транспортных пузырьков. Он осуществляет транспорт медиаторов и модуляторов. Медленный аксоток - за счет него от центра к периферии распространяются биологически активные вещества, а также составляющие компоненты мембран клеток и белков. За счет антероградного тока происходит дифференциация мышц. Это имеет большое биологическое значение. Есть мышцы фазные (мышцы конечностей) и тонические (поддерживают позу). В эксперименте установлено, что если перерезать нервы, иннервирующие эти мышцы, а затем произвести перекрест иннервации, то есть центральный конец нерва, иннервирующего фазные мышцы, подшить к нерву, иннервирующему тонические мышцы, то после прорастания нервов, фазные мышцы начинают выполнять функции тонических, а тонические - фазных. Меняется их структура, так как за счет аксотока обеспечивается трофическая функция двигательных нервов. За счет ретроградного тока нейротропные вещества поступают от периферии к центру, оказывая трофическое влияние на саму нервную клетку. За счет ретроградного тока в ЦНС могут поступать различные токсические вещества. Синапсы. Синапсы это специфические контакты нейронов обеспечивающие передачу возбуждения от одной нервной клетки к другой. В зависимости от способов передачи возбуждения (так называемого потенциала действия) выделяют химические (рис.25). и электрические синапсы. Эволюционно более древними и примитивными являются электрические синаптические контакты. Они по строению близки к щелевидным контактам (нексусам) и представляют собой ограниченные области контактов двух соседних нейронов, где расстояние между соседними мембранами сужено до 3-4 нм. В зоне этих сужений имеются белковые мембранные комплексы аналогичные коннексонам. Коннексоны представляют собой группу высокомолекулярных интегральных белков. В центральной зоне они формируют пору. При этом белки соседних мембран тесно взаимодействуют между собой, что приводит к тому, что пора, формируемая одной клеткой, переходит на соседнюю клетку. Через нее могут свободно диффундировать вода, ионы, низкомолекулярные органические вещества (в том числе и гормоноподобные). Считается, что обмен происходит в обе стороны, но имеются случаи, когда возбуждение передаются в одном направлении. Таким образом, возбуждение в таком синапсе в большинстве случаев идет в обе стороны и носит равновероятностный, диффузный характер. Такие контакты часто встречаются у низших беспозвоночных и хордовых. У млекопитающих электрические контакты имеют большое значение в процессе межнейронных взаимодействий в эмбриональном периоде развития. С дифференцировкой нервной ткани их число убывает. Подобный вид контактов у взрослых млекопитающих имеет место в ограниченных участках, например их можно видеть в мезэнцефалическом ядре тройничного нерва. Химические синапсы. Химические синапсы для передачи возбуждения от одной нервной клетки к другой используют специальные вещества – медиаторы, от чего и получили свое название. Кроме медиаторов ими используются и модуляторы. Модуляторы это специальные химические вещества, которые сами возбуждения не вызывают, но могут либо усиливать, либо ослаблять чувствительность к медиаторам (то есть модулировать пороговую чувствительность клетки к возбуждению). Согласно закону Дейля каждый нейрон содержит один специфичный для него медиатор и один или несколько модуляторов. Эти вещества накапливаются в синаптических пузырьках, которые отделены от матрикса цитоплазмы одной двухслойной мембраной. Форма, размеры, строение пузырьков зависит от содержащегося в них вещества. Любой из химических синапсов обеспечивает однонаправленную передачу возбуждения. В связи с чем в химическом синапсе выделяют следующие зоны: Пресинаптическая зона (пресинаптическое расширение, наиболее часто представляющее собой терминаль аксона) Синаптическая щель (размерыетром 20-50 нм) Постсинаптическая зона Пресинаптическая зона всегда содержит синаптические пузырьки, элементы цитоскелета (нейротубулы и нейрофиламенты), митохондрии. Часто мембрана, обеспечивающая передачу импульса имеет хорошо выраженное электронноплотное вещество. Оно представляет собой скопление мембранных белков, одной из функций которых является направленное выделение содержимого синаптических пузырьков в синаптическую щель. Таким образом, выделение медиаторов осуществляется лишь через строго определенные участки пресинаптической мембраны. Синаптическая щель изолируется от окружающего межклеточного вещества нейроглией (в центральной нервной системе отростками астроцитов), принимает медиаторы из пресинаптической зоны Постсинаптическая зона включает электронноплотное вещество постсинаптической мембраны, митохондрии. Электронноплотное вещество представлено комплексом мембранных белков, в том числе рецепторных (на гликокаликсе), белков ионных каналов, и ферментами расщепляющими медиаторы. В зависимости от того, какие структуры взаимодействуют в синапсе, можно выделить: аксо-дендритические (пресинаптическая структура аксон, постсинаптическая дендрит), аксо-аксональные (аксон с аксоном) аксо-соматические (аксон с телом нервной клетки). В нервной системе имеются и иные синаптические взаимодействия, в том числе дендро-дендритические, дендро-соматические и т.д. По строению выделяют сложные и простые синапсы. Простые синапсы содержат лишь одну поверхность взаимодействия между терминалью и посттерминальным (постсинаптическим) образованием. Сложные синапсы имеют несколько поверхностей взаимодействия. Они могут быть дивергентного, конвергентного, параллельного или эфаптического типа. Дивергентный синапс, это окончание, когда одно пресинаптическое расширение обеспечивает передачу возбуждения к нескольким постсинаптическим структурам. Конвергентный синапс предполагает одну постсинаптическую зону, к которой импульс передается от нескольких пресинаптических расширений. Параллельный синапс предполагает возможность передачи возбуждения от одной нервной структуры к близлежащей после ее стимуляции соседним претерминальным образованием. В центральной нервной системе преобладающими являются сложные синапсы. Функционально выделяют возбуждающие и тормозные синапсы. Возбуждающие приводят к деполяризации постсинаптической мембраны, что приводит к активации нервной клетки и возникновению в ней потенциала действия. Тормозные, наоборот, приводят к гиперполяризации мембраны, что снижает пороговую чувствительность нейрона к внешним влияниям. Синапсы могут быть симметричными (когда электронноплотное вещество хорошо выражено как на синаптической, так и постсинаптической мембранах) и асимметричными (слабее выявляется электронноплотное вещество пресинаптической мембраны). Симметричным синапсам часть авторов приписывает роль тормозных, а асимметричным – возбуждающих. По основному медиатору, содержащемуся в синаптических пузырьках, синапсы делятся на группы: 1. Холинергические (ацетилхолинергические). Возбуждающие и тормозные. 2. Адренергические (моноаминергические, норадренергические, дофаминергические). В основном, возбуждающие, но есть и тормозные. 3. Серотонинергические (иногда приписываются к предыдущей группе). Возбуждающие. 4. ГАМК-ергические (медиатор гаммааминомаслянная кислота). Тормозные. 5. Глутаматергические. Возбуждающие. 6. Аспартатергические. Возбуждающие. 7. Глицинергические (тормозные). 8. Пептидергические. Большая группа вешеств, в основном в ЦНС. Это может быть вазоинтерстициальный полипептид, вазопрессин, вещество Р (медиатор боли), нейропептид Y, окситоцин, бета-эндорфин и энкефалины (противоболевые), динорфин и т.д. В последние годы значительный интерес проявляется к роли закиси азота как к медиатору, в том числе и внесинаптической нервной передачи. В частности, значительная группа нейронов-водителей ритма (пейсмекеров) имеет NO-активность. В зависимости от типа медиатора синаптические пузырьки имеют разнообразную структуру. Все они отделены от гиалоплазмы одной двухслойной мембраной. Холинсодержащие пузырьки электронносветлые, диаметром 40-60 мкм. Адренсодержащие - с электронноплотной сердцевиной, светлой каемкой, диаметром 50-80 мкм. Глицинсодержащие и ГАМК-содержащие – имеют овальную форму. Пептидсодержащие – с электронноплотной сердцевиной, светлой каемкой, диаметром 90120 мкм. Механизм передачи возбуждения в химическом синапсе: Импульс, приходящий по афферентному волокну, вызывает возбуждение в синаптической бляшке и приводит к выделению медиатора через пресинаптическую мембрану. В периферических синапсах медиатором служит преимущественно ацетилхолин. В синаптическую щель поступает медиатор. На пресинаптической мембране имеются активные зоны, то есть участки пресинаптической мембраны, где больше всего происходит выброс ацетилхолина в синаптическую щель. Рядом расположены неактивные участки. На постсинаптической мембране имеются специальные холинорецепторы (для медиатора ацетилхолина) или адренорецепторы (для норадреналина). Виды секреции ацетилхолина: 1) квантовая секреция (вызванная) - с помощью импульса - является основным фактором, вызывающим деполяризацию постсинаптической мембраны и сокращение мышц. 2) неквантовая секреция - на нее приходится около 90% выделяемого медиатора, но, т.к. он поступает микродозами, не сопровождается деполяризацией постсинаптической мембраны, и не вызывает сокращения мышц. Этот механизм участвует в ауторегуляции секреции ацетилхолина и обеспечивает трофические воздействия на мышцу После выхода через активные зоны медиатор взаимодействует с холинорецепторами постсинаптической мембраны. Происходит изменение концентраций Na+ и К+, что приводит к возникновению потенциала действия. В постсинаптическом волокне медиатор холинэстераза разрушает ацетилхолин на ацетатную группу (утилизируется) и холин, который возвращается в постсинаптическую бляшку и участвует в ресинтезе медиатора. Особенности функционирования химического синапса: 1. Односторонняя передача возбуждения (от нерва на рабочий орган) – связано с тем, что рецепторы, воспринимающие медиатор, находятся на постсинаптической мембране. 2. “Синаптическая задержка” – время, необходимое на выделение медиатора в пресинаптической бляшке. 3. Следовые явления или синаптическое последействие – после прекращения раздражения продолжают поступать импульсы, т.е. продолжается потенциал действия (связано с тем, что необходимо время для разрушения медиатора холинэстеразой). 4. Суммация возбуждения – при действии подпорогового раздражителя выделяется небольшое количество медиатора, способное вызвать миниатюрный потенциал (т.е. локальное возбуждение), который не передается на постсинаптическую мембрану. Суммация может быть последовательная (временная) и пространственная. Последовательная - при частом поступлении по одному пути импульсов подпороговой величины происходит накопление достаточного количества медиатора и возникает возбуждение (потенциал действия), которое передается на постсинаптическую мембрану. Пространственная - при раздражении 2-х рецептивных полей, расположенных рядом повышается возбудимость и может возникнуть ответная реакиця. 5. Трансформация ритма раздражения в сторону увеличения или уменьшения количества импульсов на выходе – связано с низкой функциональной лабильностью синапса. 6. Высокая чувствительность к химическим веществам и специфичность (рецепторы обладают избирательной чувствительностью к медиаторам. 7. Быстрое утомление – связано с низкой функциональной лабильностью синапса и ограниченным наличием медиатора. 8. Способны изменять свою возбудимость в сторону повышения (синаптическая потенциация) или в сторону уменьшения (при длительном действии - десенситизация) Цитофизиологические и нейрофизиологические основы межнейронной передачи. Возбуждение в нейроне, как уже указывалось, связано со специфическими или (и) неспецифическими воздействиями на нервную клетку, приводящими к деполяризации мембраны нейрона. Потенциал покоя нервной клетки составляет 60-70 мВ, скорость деполяризации 1-2 мс, время реполяризации 1-2 мс. В момент деполяризации возбуждение достигает аксонального холмика, где оно может затормаживаться (если не достигает достаточной силы – пороговой чувствительности) или может распространяться далее по аксону к пресинаптической структуре (терминали аксона). Когда деполяризационная волна возбуждения достигает пресинаптического расширения, в нем открываются кальциевые каналы, в результате чего концентрация ионов кальция увеличивается. Это способствует связыванию синаптических пузырьков с пресинаптической мембраной. В данном процессе участвуют микротрубочки. Белок синаптических пузырьков – синаптобревин связывается с белками пресинаптических мембран (синтаксин и SNAP-25), после чего мембраны сливаются и содержимое пузырьков с медиатором выделяется в синаптическую щель. Медиатор взаимодейтсвует с рецепторами постсинаптических мембран. Часть из них, взаимодействуя с ионными каналами, открывает К/Na каналы и приводит к деполяризации мембран и возбуждению следующей нервной клетки. Другие – активируют процессы выделения натрия из клетки и гиперполяризации мембран нервных клеток. В последующем связь медиаторов с рецепторами разрывается. Медиатор либо метаболизируется, либо подвергается обратному всасыванию пресинаптическими мембранами, либо захватывается мембранами астроцитов с последующей передачей медиатора к нервным клеткам. Нейроглия. Впервые данный термин ввел Вирхов для описания клеток между нейронами. Занимают они около половины всего объема ткани. К ней относят нейроглию центральной нервной системы и периферической нервной системы. Выделяют макроглию и микроглию Среди макроглии центральной нервной системе имеются волокнистые (фиброзные) и протоплазматические астроциты, олигодендроциты, эпендимоциты (в том числе и танициты). В периферической нервной системе нейроглия представлена леммоцитами (шванновскими клетками) и сателлитоцитами. Астроциты. Astro – звездчатые, cites – клетки. В дословном переводе звездчатые клетки. Названы так из-за обилия отростков, отходящих от тела клетки. Многочисленные отростки ветвятся и окружают другие структуры мозга. Астроциты есть только в ЦНС и анализаторах – производных нервной трубки. Среди них встречаются волокнистые и протоплазматические астроциты. Терминали отростков обеих типов клеток имеют пуговичные расширения (ножки астроцитов), значительная часть из которых заканчивается в периваскулярном пространстве, окружая капилляры периваскулярными глиальными бухтами занимающими 80% обменной поверхности микрососудов. Волокнистые астроциты имеют многочисленные, длинные, тонкие, слабо или совсем не ветвящиеся отростки. В основном присутствуют в белом веществе мозга. Протоплазматические астроциты отличаются короткими, толстыми и сильно ветвящимися отростками. Имеются преимущественно в сером веществе мозга. Морфология астроцитов отличается крайним разнообразием и коррелирует с формой капиллярных петель и нейронных ансамблей в ЦНС. Клетки диффузно распределены в объеме мозга. Их отростки взаимопереплетаются между собой, так и другими элементами нейропиля. Астроциты занимают исключительное положение в ЦНС, располагаясь между телами нейронов, немиелинизированной и миелинизированной частями нервных отростков, синапсами, кровеносными сосудами, подэпендимными пространствами изолируя и в то же время структурно связывая их. Специфическим маркером астроцитов является так называемый глиальный фибриллярный кислый белок, из которого образуются промежуточные филаменты. Строение астроцитов. Клетки имеют относительно крупные светлые ядра, со слабо развитым ядрышковым аппаратом. Цитоплазма слабо оксифильная. В ней слабо развиты гладкая и гранулярная ЭПС, пластинчатый комплекс. Митохондрий мало, они небольших размеров. Цитоскелет развит умеренно в протоплазматических и хорошо - в волокнистых астроцитах. Между клетками значительное число щелевидных и десмосомоподобных контактов. В постнатальный период жизни человека астроциты способны к миграции, особенно в зоны повреждения и, как полагают, способны к пролиферации. Функции астроцитов: 1.Участвуют в гематоэнцефалическом и ликворогематическом барьерах. Астроциты своими ножками покрывают капилляры, поверхности мозга и участвуют в транспорте веществ от сосудов к нейронам и наоборот. Способы передавать метаболиты нейронам. 2. Обеспечивают ионный обмен, особенно ионов калия. При активации нейронов, их частая и длительная деполяризация может вести к значительному увеличению ионов калия, что может изменять мембранный потенциал нейронов, повышая их чувствительность к внешним воздействиям. Астроциты, захватывая избыточный калий, предотвращают перевозбуждение. Нарушения в этой функции могут вести к развитию состояний эпилепсии. 3. Изолируют рецепторные поверхности тел нейронов и синапсов. При этом клетки способны к ритмичным сокращениям (по некоторым авторам к набуханию) изменяя зоны изоляции. 4.Способны к захвату нейромедиаторов (глиоксиловой кислоты, гамма-аминомаслянной кислоты) из зон синаптической передачи. Под их влиянием изменяется активность самих астроцитов, что приводит к внесинаптической модуляции сигнала в соседних нервных клетках и их отростках. 5.Фагоцитоз погибших нейронов. На месте фагоцитированных нейронов в результате их гибели, глиальные рубцы. Это скопления гипертрофированных астроцитов, заменяющих собой рубцы в периферических органах. 6.Выделяют большое количество биологически активных веществ (факторы роста нервов, факторы роста фибробластов, ангиогенные факторы, эпидермальный фактор роста, интерлейкин–I, простагландины), контролирующих местные межклеточные метаболические и внесинаптические информационные взаимодействия. Факторы роста способны инициировать и ускорять рост отростков нейронов. Выделение интерлейкина–I, способность экспрессировать МНС-комплексы 1 и 2 классов, указывает на роль астроцитов в формировании специфических иммунных реакций и антигенпрезентирующую функцию. Выделение факторов роста фибробластов, компонентов межклеточного вещества (ламелин, фибронектин), простагландинов, антиваскулярных факторов позволяет астроцитам контролировать состояние местного кровотока в ЦНС. 7.В эмбриональном развитии человека предшественники астроцитов контролируют направление миграции астробластов, во всяком случае, в части зон головного мозга (мозжечок, гипоталамус), а также в зоне их отростков (зрительный нерв). В постнатальном развитии стабилизируют структуры ЦНС, ингибируя рост отростков нейронов и в то же время, предотвращая апоптозы (запрограммированную гибель) нейронов, предотвращая избыточное снижение их числа при повреждении. Олигодендроциты. Олигодендроциты (олигодендроглиоциты) центральной нервной системы относятся к миелинообразующим клеткам и участвуют в формировании нервных волокон. Подобную им функцию в периферической нервной системе играют леммоциты (шванновские клетки) периферической нервной системы. Тела по размерам как правило мельче астроцитов. Ядра мелкие, округлые, темноокрашенные. Ядрышки мелкие. Отростков мало. Они тонкие, не ветвятся или слабо ветвятся и заканчиваются вокруг аксонов и дендритов нервных клеток. Часть олигодендроцитов концентрируется в непосредственной близости к телам нервных клеток (сателлитные олигодендроциты). Терминальная зона каждого отростка участвует в формировании сегмента нервного волокна, то есть каждый олигодендроцит обеспечивает окружение сразу нескольких нервных волокон. Цитоплазма слабоксифильная и при общих методах окрашивания сливается с нейропилем. На электроннооптическом уровне–в цитополазме хорошо развиты органеллы по составу близкие к нейронам, но в отличие от них не имеют развитого цитоскелета. Леммоциты (шванновские клетки) периферической нервной системы. Окружают отростки нейронов в периферической нервной системе. При этом клетки имеют удлиненную форму, распластываясь на поверхности аксона, не имеют отростков и обеспечивают формирование глиальной оболочки в одном сегменте миелинового или безмиелинового нервного волокна. В области формирования корешков спинномозговых и черепно-мозговых нервов формируют скопления (глиальные пробки), предотвращая проникновение отростков ассоциативных нейронов ЦНС за ее пределы. Кроме леммоцитов в периферической нервной системе имеются сателлитные (мантийные) глиоциты в периферических нервных узлах вокруг тел нейронов, глиоциты нервных окончаний, конкретные морфологические особенности которых рассматриваются при изучении нервных окончаний и анатомии нервных узлов. Леммоциты характеризуются удлиненными, темноокрашенными ядрами, слабо развитыми митохондриями и синтетическим аппаратом (гранулярная, гладкая ЭПС, пластинчатый комплекс). Мантийные клетки уплощенной формы с несколькими отростками. Они распластаны на поверхности нейрона и формируют вокруг него глиальную капсулу. Ядра уплощены, темные, со слабо развитым ядрышковым аппаратом. Синтетический аппарат слабо развит. Клетки обеспечивают изолирующую, трофическую, опорную, защитную функции. Играют барьерную роль. Функции олигодендроглии и леммоцитов. 1. Формируют оболочки вокруг нервных клеток, обеспечивая барьерно-транспортные функции и изоляцию тел нейронов. 2. Образуют миелиновые оболочки, участвуя в проведении возбуждения в нервном отростке. Они изолируют отростки, ускоряя проведение возбуждения и предотвращая его затухание и распространение (ирритацию) на соседние отростки. 3. Механическая (опорная) функция. 4. Трофическая. 5. Участие в регенерации поврежденных нервных клеток. Повреждение стимулирует выделение олигодендроцитами большого количества биологически активных веществ, предотвращающих гибель нейронов и стимулирующих регенерацию. 6. Активация роста аксонов и направление роста их отростков при повреждении. 7. В составе нервных окончаний производные олигодендроглии способствуют рецепторным функциям. 8. Фагоцитируют остатки поврежденных осевых цилиндров и миелина при нарушении структуры аксона дистальнее места повреждения. Эпендимоциты. Это клетки низкопризматической формы. Они образуют непрерывный пласт, покрывающий полости мозга. Эпендимоциты тесно прилежат друг к другу, формируя плотные, щелевидные и десмосомальные контакты. Апикальная поверхность содержит реснички, которые у большинства клеток затем замещаются микроворсинками. Базальная поверхность имеет базальные впячивания (инвагинации), а также длинные тонкие отростки (от одного до нескольких), которые проникают до периваскулярных пространств микрососудов мозга. В цитоплазме эпендимоцитов обнаруживаются митохондрии, умеренно развитый синтетический аппарат, хорошо представлен цитоскелет, имеется значительное количество трофических и секреторных включений. Вариантом эпендимной глии являются танициты. Они выстилают сосудистые сплетения желудочков головного мозга, субкомиссуральный орган задней комиссуры. Активно участвуют в образовании ликвора (спинномозговой жидкости). Характеризуются тем, что базальная часть содержит тонкие длинные отростки. Функции эпендимоцитов: 1. Выстилают желудочки мозга, обеспечивая гемато-ликворный барьер и отделяя ликвор от структур мозговой ткани. 2. Образуют ликвор, т.е. обеспечивают секреторную функцию. 3. Контролируют ионный состав ликвора. 4. Направляют миграцию нейробластов в нервной трубке в эмбриональном периоде развития (предшественники таницитов). 5. Опорная функция. Микроглиоциты (нейральные макрофаги). Клетки небольших размеров, диффузно распределенные в центральной нервной системе, с многочисленными сильно ветвящимися отростками, способны к миграции. Ядра характеризуются грубыми глыбками гетерохроматина (выраженная конденсация хроматина). В цитоплазме обнаруживается много лизосом, гранул липофусцина. Умеренно развит белковосинтетический аппарат (гранулярная ЭПС, пластинчатый комплекс). Строение нервных волокон. Все нервные волокна подразделяются на миелиновые и безмиелиновые (рис.26): * Миелиновые (мякотные) - имеют миелиновую оболочку, перехваты Ранвье, имеющие важное значение для передачи возбуждения. Сама миелиновая оболочка является мощным биологическим изолятором. Через нее возбуждение не перескакивает с одного нервного волокна, на соседнее. Поэтому проходящий импульс неэффективен для соседних волокон; *Безмиелиновые нервные волокна - передача возбуждения в них происходит по поверхности нерва через изменение поверхностного заряда. Обычно нервный ствол содержит большое количество нервных волокон. Безмиелиновые волокна в нем находятся среди миелиновых. В периферической нервной системе нервные волокна окружают леммоциты. Один леммоцит связан с одним нервным волокном. В центральной нервной системе отростки нейронов окружают олигодендроциты. Каждый олигодендроцит участвует в формировании нескольких нервных волокон. В безмиелиновых нервных волокнах в состав волокна может входить несколько отростков нейронов (кабельный тип волокна) или один отросток; отростки могут переходит из одного волокна в другое. В нервном волокне выделяют осевой цилиндр (отросток нервной клетки), мезаксон, область плотного контакта впячивания (дубликатуры) глиоцита, цитоплазму глиоцита. Один отросток олигодендроцита или один леммоцит окружает участок нервного волокна, называемый сегментом. В миелиновом волокне мезаксон многократно оборачивается вокруг осевого цилиндра, формируя многократные витки мембраны – миелин. В миелиновом (мякотном) нервном волокне имеется только один осевой цилиндр. Зоны разрыхления миелина называются насечками (насечки Шмидта-Лантермана). Границы сегментов немиелинизированы и называются перехватами Ранвье. Эти области соответствуют контактам глиоцитов. Миелинизация волокон осуществляется путем удлинения и «наворачивания» мезаксона вокруг отростка нервной клетки (в периферической нервной системе) или удлинения и вращения отростка олигодендроцита вокруг осевого цилиндра в ЦНС. Классификация нервных волокон: 1. Тип А имеет подгруппы: А - обладают наибольшей скоростью проведения возбуждения - 70-120 м/с (соматические двигательные нервные волокна). А - скорость проведения составляет 40-70 м/с. Это соматические афферентные нервы и некоторые эфферентные соматические нервы. А - скорость проведения составляет 15-40 м/с - афферентные и эфферентные симпатические и парасимпатические нервы. А (дельта) - скорость проведения 5-18 м/с. По этой группе афферентных соматических нервов проводятся первичная (быстрая) боль. 2. Тип В - скорость проведения от 3 до 14 м/с - преганглионарные симпатические волокна, некоторые парасимпатические волокна, то есть это вегетативные нервы. 3. Тип С - скорость проведения 0,5-3 м/с. Постганглионарные вегетативные волокна (безмиелиновые). Проводят болевые импульсы медленной вторичной боли (от рецепторов пульпы зуба), а так как, нет ограничителей возникают сильные боли. Нейрогенез. Нервная ткань формируется из первичной эктодермы. На 15-17 сутки внутриутробного развития человека под индуцирующим влиянием хорды формируется нервная пластинка (скопление продольно лежащего клеточного материала). Такая индукция называется первичной. С 17 по 21 сутки формируется нервный желобок, а затем и трубка. К 25 суткам эмбриогенеза происходит отщепление нервной трубки от эктодермы и замыкание переднего и заднего отверстий (нейропоров). Индукция хордомезодермой нервной трубки осуществляется за счет сложного комплекса межклеточных взаимодействий. Среди них важную роль играют биологически активные факторы (серотонин, норадреналин) и биологически активные метаболиты. По бокам от нервного желобка располагаются структуры нервного гребня. На ранних сроках развития нервная трубка сформирована медулобластами (стволовые клетки нервной ткани центральной нервной системы). Из нервного гребня образуется ганглиозная пластинка состоящая из ганглиобластов (стволовые клетки нейронов и нейроглии периферической нервной системы. Медулобласты и ганглиобласты интенсивно иммигрируют, делятся и затем дифференцируются. В ранние сроки внутриутробного развития нервная трубка представляет собой пласт отростчатых клеток, лежащих в виде одного слоя, но в несколько рядов. Изнутри и снаружи они ограничены пограничными мембранами. На внутренней поверхности (прилежащей к полости нервной трубки) медулобласты делятся. В последующем нервная трубка формирует несколько слоев. Среди них можно выделить: Внутренняя пограничная мембрана. Отделяет полость нервной трубки от клеток. Эпендимный слой (вентрикулярный в области мозговых пузырей) представлен бластными клетками-предшественниками макроглии. В передних мозговых пузырях некоторые авторы выделяют субвентрикулярную зону, где происходит пролиферация нейробластов. Мантийный (плащевой) слой, содержит мигрирующие и дифференцирующиеся нейробласты и глиобласты. Маргинальный слой (краевая вуаль). Сформирована отростками глиобластов и нейробластов. В ней можно видеть тела отдельных клеток. Наружная пограничная мембрана. Диффероны нервной ткани центральной нервной системы. Из медулобластов дифференцируются эпендимобласты. Нейробласты и спонгиобласты. Назовем несколько гистоенетических рядов дифференцировки: 1. Медулобласт-нейробласт-молодой нейрон-зрелый нейрон. 2. Медулобласт-спонгиобласт (как вариант радиальный глиобласт) -астробластпротоплазматический или волокнистый астроцит. 3. Медулобласт-спонгиобласт-олигодендробласт-олигодендроцит. 4. Медулобаст-эпендимобласт-эпендимоцит или таницит. Микроглиоцит, в основной своей массе, по мнению большинства авторов формируются из моноцитов, проникающих в нервную трубку в первой половине внутриутробного развития. Для них дифферон выглядит следующим образом: стволовая клетка кровиполустволовая клетка крови (КОЕ ГЭММ) - КОЕ ГМ – КОЕ М – монобласт – промоноцит – моноцит – микроглиоцит покоя – активированный микроглиоцит. Диффероны нервной ткани в периферической нервной системе. 1.Ганглиобласт - нейробласт - молодой нейрон - зрелый нейрон. 2.Ганглиобласт- глиобласт- леммоцит (шванновская клетка) 3.Ганглиобласт- глиобласт- мантийный глиоцит (сателлитоцит). Механизмы нейрогенеза. В процессе внутриутробного развития нейробласты мигрируют в области анатомических закладок нервных центров. При этом они прекращают делиться. В ЦНС миграция нейробластов контролируется адгезивными межклеточными взаимодействиями (с помощью кадгеринов и интегринов радиальной глии), сигнальными молекулами межклеточного вещества (в том числе фибронектинами и ламининами). После того как нейробласты достигают области своей постоянной локализации, они начинают дифференцироваться и формировать отростки. Направление роста отростков также контролируется упомянутыми адгезивными молекулами (кадгерины, интегрины, сигнальные молекулы межклеточного вещества). Во внутриутробном развитии и после рождения происходит конкурентное взаимодействие между аналогичными нейронами нервных центров. При этом нервные клетки, не успевшие занять соответствующую зону, либо сформировать контакты, подвергаются апоптозу. В раннем развитии погибает от трети до половины нервных клеток. В последующем развитии вокруг нервных клеток формируется глиальное окружение и происходит миелинизация нервных волокон. Нервные клетки до полового созревания продолжают формировать отростки и синаптические контакты. Максимального развития нервная ткань достигает к 25-30 годам. С возрастом наблюдается гибель части нервных клеток и компенсаторная гипертрофия других. В нейронах может накапливаться липофусцин. Области с погибшими телами нервных клеток замещаются глиальными рубцам, образованными скоплением гипертрофированных астроцитов. Вопросы для самоконтроля: 1. 2. 3. 4. 5. Назовите структуры специфичные для нейронов. Специализированные органеллы нервной клетки. Классификации нейронов. Механизмы нейрогенеза. Классификация и строение синапсов. Их значение. 6. Классификация и строение нейроглии. 7. Сравнительная характеристика астроцитов и олигодендроцитов. 8. Строение глиоцитов периферической нервной системы. 9. Строение и значение микроглиоцитов. 10. Строение и значение миелиновых и безмиелиновых нервных волокон. 11. Механизмы проведения возбуждения в нервных волокнах. 12. Представление о миелинизации. 13. Постравматическая регенерация нейронов и нервных волокон. Ситуационные задачи. 1.В результате травмы у больного в месте перерыва нерва преждевременно возник грубый соединительнотканный рубец. Спрогнозируйте процесс регенерации нерва. 2.Как зависит от функционального состояния нейроцита гранулярная эндоплазматическая сеть? 3.Леммоцит является «футляром» для нескольких осевых цилиндров. О каком типе нервного волокна идет речь? 4.У зародыша в эксперименте удалена ганглиозная пластинка. Каковы нарушения дальнейшей дифференцировки нервной ткани? 5.В нервной клетке пять отростков. Назовите форму нейрона и число афферентных и эфферентных (аксонов) проводников импульса. СПИСОК ЛИТЕРАТУРЫ 1.Артишевский А.А., Леонтюк А.С., Слука Б.А. Гистология с техникой гистологических исследований.–Минск, «Вышейшая школа».–1999.–231с. 2..Гистология. Под ред. Афанасьева Ю.И., Юриной Н.А.–М.Медицина–1999.–744с. 3.Гистология. Под ред. проф. Улумбекова Э.Г., проф.Челышева Ю.А.–М. ГЭОТАР–МЕД– 2001.–670с. 4.Лабораторные занятия по курсу гистологии, цитологии и эмбриологии. Под ред.Афанасьева Ю.И., Яцковского А.Н.–М.Медицина–1999.–321с. 5.Леонтюк А.С., Слука Б.А.Основы возрастной гистологии .–Минск, «Вышейшая школа».–2000.–415с. СОДЕРЖАНИЕ ВВЕДЕНИЕ В ТКАНИ……………………………………………… 1.ЭПИТЕЛИАЛЬНЫЕ ТКАНИ………………………………………. 2.ТКАНИ ВНУТРЕННЕЙ СРЕДЫ…………………………………… 2.1. КРОВЬ.КРОВЕТВОРЕНИЕ. ЛИМФА……………………………… 2.2. СОЕДИНИТЕЛЬНЫЕ ТКАНИ……………………………………… 3.ХРЯЩЕВЫЕ ТКАНИ………………………………………………… 4.КОСТНЫЕ ТКАНИ………………………………………………….. 5.МЫШЕЧНЫЕ ТКАНИ………………………………………………. 6.НЕРВНЫЕ ТКАНИ……………………………………………………