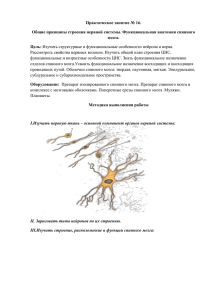
Раздел III. Нервная и гормональная регуляция функций Занятие № 10. Функции спинного мозга, ствола головного мозга и мозжечка 1. Морфофункциональная организация спинного мозга. Спинной мозг — наиболее древнее образование центральной нервной системы; Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде передних корешков. Спинной мозг человека имеет 31—33 сегмента: 8 шейных (СI— CVIII), 12 грудных (ТI—TXII), 5 поясничных (LI—LV), S крестцовых (SI—SV), 1—3 копчиковых (CoI— СоIII). Морфологических границ между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным и определяется зоной распределения в нем волокон заднего корешка и зоной клеток, которые образуют выход передних корешков. Каждый сегмент через свои корешки иннервирует три метамера тела и получает информацию также от трех метамеров тела. В итоге перекрытия каждый метамер тела иннервируется тремя сегментами и передает сигналы в три сегмента спинного мозга. Спинной мозг человека имеет два утолщения: шейное и поясничное — в них содержится большее число нейронов, чем в остальных его участках. 2. Нейронная организация сегментов спинного мозга. Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных двух передних и двух задних рогов в шейном, поясничном и крестцовом отделах. Серое вещество распределено на ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет, помимо названных, еще и боковые рога. Спинной мозг человека содержит около 13 млн. нейронов, из них 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы: 1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; 2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения; 3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков; 4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами. Начиная с I грудного сегмента спинного мозга и до первых поясничных сегментов, в боковых рогах серого вещества располагаются нейроны симпатического, а в крестцовых — парасимпатического отдела автономной (вегетативной) нервной системы. Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами экстра- и интрамуральных ганглиев симпатического и парасимпатического отделов автономной нервной системы. 1 Первая группа афферентных входов спинного мозга образована чувствительными волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов. Эта группа рецепторов образует начало так называемой проприоцептивной чувствительности. Проприоцептивные волокна по толщине и скорости проведения возбуждения делятся на 3 группы. Волокна каждой группы имеют свои пороги возникновения возбуждения. Вторая группа афферентных входов спинного мозга начинается от кожных рецепторов: болевых, температурных, тактильных, давления — и представляет собой кожную рецептирующую систему. Третья группа афферентных входов спинного мозга представлена рецептирующими входами от висцеральных органов; это висцеро-рецептивная система. Интернейроны. Эти промежуточные нейроны, генерирующие импульсы с частотй до 1000 в секунду, являются фоновоактивными и имеют на своих дендритах до 500 синапсов. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. Очень важной функцией интернейронов является торможение активности нейронов, что обеспечивает сохранение направленности пути возбуждения. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцыантагонисты. Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду). Функции задних и передних корешков сегментов спинного мозга. Закон БеллаМажанди. В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем». Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, и их волокна иннервируют всю скелетную мускулатуру. Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом. Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга. Нейроны парасимпатического отдела автономной системы локализуются в сакральном отделе спинного мозга и являются фоновоактивными. В случаях раздражения и поражения задних корешков спинного мозга наблюдаются «стреляющие», опоясывающие боли на уровне метамера пораженного сегмента, снижение чувствительности всех видов, утрата или снижение рефлексов, вызываемых с метамера тела, который передает информацию в пораженный корешок. В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой 2 чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх. Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность. В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние — эфферентными, двигательными, центробежными (закон Белла— Мажанди). 3. Альфа- и гамма-мотонейроны спинного мозга, их функции. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чем меньше количественно мотонейронная единица), тем более дифференцированные, точные движения выполняет мышца. Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул. Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы; в этом случае развивается максимальное сокращение мышцы. Мотонейроны спинного мозга функционально делят на α- и γ-нейроны. αМотонейроны образуют прямые связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена, имеют до 20 000 синапсов на своих дендритах и характеризуются низкой частотой импульсации (10—20 в секунду), γМотонейроны, иннервирующие интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны. Сокращение интрафузального мышечного волокна не приводит к сокращению мышцы, но повышает частоту разрядов импульсов, идущих от рецепторов волокна в спинной мозг. Эти нейроны обладают высокой частотой импульсации (до 200 в секунду). 4. Нейроны боковых рогов сегментов спинного мозга, их функции. Симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков; Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду). Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными. В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней. 3 Симпатический отдел по своим основным функциям является трофическим. Он обеспечивает усиление окислительных процессов, усиление дыхания, учащение деятельности сердца, т.е. приспосабливает организм к условиям интенсивной деятельности. В связи с этим тонус симпатической нервной системы преобладает днем. Парасимпатический отдел выполняет охранительную роль (сужение зрачка, бронхов, снижение частоты сердечных сокращений, опорожнение полостных органов), ее тонус преобладает ночью («царство вагуса»). Медиатором в симпатических нервных окончаниях является норадреналин. Медиатором парасимпатических нервных окончаний - ацетилхолин. 5. Восходящие проводящие пути спинного мозга, их функции. Пучки Голля (тонкий пучок) и Бурдаха (клиновидный пучок) проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. В задних канатиках. В боковых: Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем в соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование соответствующей чувствительности. Нисходящие проводящие пути спинного мозга, их функции. Кортикоспинальные (пирамидные) пути идут от пирамидных и экстрапирамидных нейронов коры к альфамотонейронам двигательных центров спинного мозга, осуществляют координацию произвольных движений. Руброспинальный путь (Монаков) проводит сигналы от красного ядра среднего мозга к мотонейронам мышц сгибателей, регулирует тонус соответствующих мышц при изменениях положения тела. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга (в первую очередь, от ядра Дейтерса) к мотонейронам мышц-разгибателей; участвует в поддержании позы и равновесия тела. 6. Классификация спинномозговых рефлексов, их характеристика. 1) Соматосенсорные: рефлексы с рецепторов кожи носят характер, зависящий от силы раздражения, вида раздражаемого рецептора, но чаще всего конечная реакция выглядит в виде усиления сокращения мышц-сгибателей. 2) Соматовисцеральные: имеют свои пути; они начинаются от различных рецепторов, входят в спинной мозг через задние корешки, задние рога, далее в боковые рога, нейроны которых через передний корешок посылают аксоны не непосредственно к органам, а к ганглию симпатического или парасимпатического отдела автономной нервной системы; автономные (вегетативные) рефлексы обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов; эти рефлексы отличаются большим латентным периодом (ЛП) двумя фазами реакции: первая — ранняя — возникает с ЛП 7—9 мс и реализуется ограниченным числом сегментов, вторая — поздняя — возникает с большим ЛП — до 21 мс и вовлекает в реакцию практически все сегменты спинного мозга; поздний 4 компонент вегетативного рефлекса обусловлен вовлечением в него вегетативных центров головного мозга; сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение; в основе реализации произвольного движения лежит γ-афферентная рефлекторная система. В нее входят пирамидная кора, экстрапирамидная система, α- и γ-мотонейроны спинного мозга, экстра- и интрафузальные волокна мышечного веретена. 3) Висцеросоматические: рефлексы возникают при стимуляции афферентных нервов внутренних органов и характеризуются появлением двигательных реакций мышц грудной клетки и брюшной стенки, мышц разгибателей спины. 4) Миотатические рефлексы: рефлексы на растяжение мышцы; быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и двигательной реакции; например, легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра и разгибание голени; дуга этого рефлекса следующая: мышечные рецепторы четырехглавой мышцы бедра —> спинальный ганглий — > задние корешки —> задние рога III поясничного сегмента —> мотонейроны передних рогов того же сегмента —> экстрафузальные волокна четырехглавой мышцы бедра; реализация этого рефлекса была бы невозможна, если бы одновременно с сокращением мышц-разгибателей не расслаблялись мышцы- сгибатели; рефлекс на растяжение свойствен всем мышцам, но у мышц- разгибателей, они хорошо выражены и легко вызываются. 7. Нервные центры продолговатого мозга, их функции. I. Жизненно – важные: - Дыхательный (экспираторные и инспираторных отделы=формирование выдоха и вдоха) - Сердечно-сосудистый (контролирует тонус сосуда, работу сердца, влияние на АД) II. Защитные: - Слезоотделения - Кашля - Чихания - Мигания - Рвоты III. Пищеварительная: - Слюноотделения (вегетативные) Соматические: - Сосания - Жевания - Глотания IV. Постуральные (поддержание позы): - Статические (поддержание равновесия при отсутствии движения) - Статокинетические (поддержание равновесия во время движения) 8. Роль продолговатого мозга в рефлексах регуляции позы. Соматическими являются статические рефлексы продолговатого мозга, относящиеся к познотоническим или рефлексам позы. Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальные тракты. Рефлексы возникают 5 тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности 9. Нервные центры и ядра варолиевого моста, их функции. Нервные центры: 1.Центры поддержания позы: а)статическая б)статокинетическая 2. Пневмотаксический центр : - координирует фазу дых.цикла тормозят вдох и способствует выдоху. Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка. 1) Ядра VIII пары черепно-мозговых нервов, вестибулярное ядро — отвечает за первичный анализ вестибулярных раздражителей. 2) VII пара — лицевой нерв. иннервирует мимические мышцы лица, подъязычную и подчелюстную слюнные железы, передает информацию от вкусовых рецепторов передней части языка. 3) V пара — тройничный нерв. Двигательное ядро иннервирует жевательные мышцы, мышцы небной занавески и мышцы, напрягающие барабанную перепонку. Чувствительное ядро получает афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, надкостницы костей черепа, конъюктивы глазного яблока. Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга. Она влияет на кору больших полушарий, активируя ее и вызывая пробуждение. Аксоны ретикулярной формации идут в мозжечок и спинной мозг. 10.Функции ядер нижнего и верхнего двухолмия. Верхние бугры четверохолмия являются первичными зрительными центрами. К ним подходят пути от нейронов сетчатки глаза. От них сигналы идут к таламусу, а по нисходящему тектоспинальному пути – к мотонейронам спинного мозга. В верхнем двухолмии происходит первичный анализ зрительной информации. Например, определение положения источника света, направление его движения. В них также формируются зрительные ориентировочные рефлексы (поворот головы в сторону источника света). Нижние бугры четверохолмия являются первичными слуховыми центрами. К ним идут сигналы от фонорецепторов уха, а от них – к таламусу. От них к мотонейронам также идут пути в составе тектоспинального тракта. В нижних буграх осуществляется первичный анализ слуховых сигналов, а за счет связей с мотонейронами формируются ориентировочные рефлексы на звуковые раздражители. 6 11.Функции красного ядра среднего мозга. Расположены в верхней части ножки мозга. К нему идут нервные пути от коры полушарий, подкорковых ядер, мозжечка. От него идет руброспинальный тракт к мотонейронам сгибателей спинного и ретикулярной формации продолговатого мозга. Участвует в регуляции тонуса мышц антагонистов разгибателей и сгибателей. Влияет на альфа и гамма-мотонейроны спинного мозга. Вызывает возбуждение мотонейронов мышц сгибателей, а через тормозные клетки Реншоу тормозят мотонейроны мышц разгибателей. Антагонистом красного ядра является вестибулярное ядро Дейтерса. Функции черной субстанции среднего мозга. Располагается в ножках мозга, участвует в регуляции актов жевания, глотания и их последовательности, а также в координации мелких и точных движений пальцев рук. Нейроны этого ядра синтезируют дофамин, поставляемый к базальным ядрам головного мозга. Он играет важную роль в контроле сложных двигательных актов. Поражение черного вещества приводит к дегенерации дофаминергечиских волокон, проецирующихся в полосатое тело, нарушению тонких движений пальцев рук, развитию мышечной ригидности и тремору (болезнь Паркинсона). Принимает участие в пищевом поведении, регулирует пластический тонус, эмоциональное поведение. 12.Функции ретикулярной формации ствола мозга, их характеристика. 1. Соматодвигательный контроль (активация скелетной мускулатуры), может быть прямым через ретикулоспинальный путь и непрямым через мозжечок, оливы, бугорки четверохолмия, красное ядро, черное вещество, полосатое тело, ядра таламуса и соматомоторные зоны коры. 2. Соматочувствительный контроль, т.е. снижение уровней соматосенсорной информации — «медленная боль», модификация восприятия различных видов сенсорной чувствительности (слуха, зрения, вестибуляции, обоняния). 3. Висцеромоторный контроль состояния сердечно-сосудистой, дыхательной систем, активности гладкой мускулатуры различных внутренних органов. 4. Нейроэндокринная трансдукция через влияние на нейромедиаторы, центры гипоталамуса и далее гипофиз. 5. Биоритмы через связи с гипоталамусом и шишковидной железой. 6. Различные функциональные состояния организма (сон, пробуждение, состояние сознания, поведение) осуществляются посредством многочисленных связей ядер ретикулярной формации со всеми частями ЦНС. 7. Координация работы разных центров ствола мозга, обеспечивающих сложные висцеральные рефлекторные ответы (чихание, кашель, рвота, зевота, жевание, сосание, глотание и др.). Восходящие и нисходящие влияния ретикулярной формации на другие структуры головного и спинного мозга. При восходящем влияние ретикулярной формации, повышается активность аналитикосинтетической деятельности, увеличивается скорость рефлексов, организм подготавливается к реакции на неожиданную ситуацию. Поэтому ретикулярная формация участвует в организации оборонительного, полового, пищеварительного поведения. С другой стороны, она может избирательно активировать или тормозить определенные системы мозга. В свою очередь кора больших полушарий, через нисходящие пути, может оказывать возбуждающее действие на ретикулярную формацию. 7 Нисходящие ретикулоспинальные пути идут от ретикулярной формации к нейронам спинного мозга. Поэтому она может оказывать нисходящие возбуждающие и тормозящие влияния на его нейроны. Например, ее гипоталамические и мезэнцефальные отделы повышают активность альфа-мотонейронов спинного мозга. В результате этого растет тонус скелетных мышц, усиливаются двигательные рефлексы. Тормозящее влияние ретикулярной формации на спинальные двигательные центры осуществляется через тормозные нейроны Реншоу. Это приводит к торможению спинальных рефлексов. 13.Морфофункциональная организация таламуса. Классификация ядер таламуса. Таламус - один из 2-х отделов промежуточного мозга, представляет собой большое парное скопление серого вещества на боковых стенках промежуточного мозга по бокам III желудочка, имеющее яйцевидную форму, причем передний его конец заострен (передний бугорок), а задний расширен (подушка). В нем происходит обработка почти всей информации, идущей от рецепторов к коре. Через него проходят сигналы от зрительных, слуховых, вкусовых, кожных, мышечных, висцеральных рецепторов, а также ядер ствола мозга, мозжечка, подкорковых. Сам он содержит около 120 ядер, делящихся на: - Специфические - Ассоциативные - Неспецифические Функции специфических, ассоциативных и неспецифических ядер таламуса. -Специфические ядра: делятся на переключающие (релейные) и ассоциативные. Латеральные коленчатые тела – зрение, медиальные – слух. Переключающие ядра состоят из нейронов, у которых мало дендритов и длинный аксон. С их помощью происходит переключение сигналов, идущих от нижележащих отделов ЦНС, на соответствующие соматосенсорные зоны коры, в которых находится представительство определенных рецепторов. Например, в латеральных коленчатых телах переключаются зрительные сигналы на затылочные доли коры. В переключающих ядрах выделяется наиболее важная информация. При нарушении функции этих ядер выключается восприятие соответствующих сигналов. Ассоциативные нейроны имеют большее количество отростков и синапсов. Это позволяет им воспринимать различные по характеру сигналы. Они получают эти сигналы от переключающих нейронов и осуществляют их первичный синтез. От них пути идут к ассоциативным зонам коры, в которых происходит высший синтез и формируются сложные ощущения. -Ассоциативные ядра таламуса: представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Участие в формировании эмоциональной и двигательной активности. Узнавание предметов и интеграция речевых и зрительных функций -Неспецифические относятся к переднему отделу ретикулярной формации ствола мозга и представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. При возбуждении неспецифических ядер в коре мозга развивается периодическая электрическая активность в виде веретен, что свидетельствует о переходе к сонному состоянию. Т.е. они обеспечивают 8 определенный уровень функционального активности коры. ретикулярное и срединное ядро (воспринимают сенсорную неспецифическую информацию и переключают вторичные проекционные поля коры) Кроме того, ядра таламуса участвуют в формировании безусловных двигательных рефлексов сосания, жевания, глотания. В таламусе находится подкорковый центр болевой чувствительности, в котором формируется общее ощущение боли, не имеющее определенной локализации и окраски 14.Морфофункциональная организация мозжечка. Мозжечок состоит из 2-х полушарий и червя между ними. Серое вещество образует кору и ядра. Белое образовано отростками нейронов. Мозжечок получает афферентные нервные импульсы от тактильных рецепторов, рецепторов вестибулярного аппарата, проприорецепторов мышц и сухожилий, а также двигательных зон коры. Эфферентные импульсы от мозжечка идут к красному ядру среднего мозга, ядру Дейтерса продолговатого мозга, к таламусу, а затем к моторным зонам КБП и подкорковым ядрам. Особенности морфофункциональной организации и связей мозжечка. Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка: -кора мозжечка построена достаточно однородно, имеет стереотипные связи, что создает условия для быстрой обработки информации; -Наиболее крупным и функционально значимым среди нейронов коры мозжечка является клетка Пуркинье. Она имеет большое количество входов и формирует единственный аксонный выход, коллатерали которого заканчиваются на ядерных структурах; -на клетки Пуркинье проецируются сенсорные раздражения практически всех видов: проприоцептивные, кожные, зрительные, слуховые, вестибулярные -выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом. В мозжечке анатомически и функционально выделяют старую, древнюю и новую части. Кора мозжечка имеет специфическое, нигде в ЦНС не встречающееся строение. Верхний (I) слой коры мозжечка — молекулярный — состоит из параллельных волокон, разветвлений дендритов и аксонов II и III слоев. В нижней части молекулярного слоя встречаются корзинчатые и звездчатые клетки, которые обеспечивают взаимодействие клеток Пуркинье. Средний (II) слой коры образован клетками Пуркинье, расположенными в один ряд и имеющими самую мощную в ЦНС дендритную систему. На дендритном поле одной клетки Пуркинье может быть до 60000 синапсов. Следовательно, эти клетки выполняют функцию сбора, обработки и передачи информации. Аксоны клеток Пуркинье являются единственным путем, по которому информация из коры мозжечка передается в ею ядра и ядра структуры большого мозга. Под II слоем коры, т. е. под клетками Пуркинье, лежит гранулярный (III) слой, состоящий из клеток-зерен, число которых достигает 10 млрд. Аксоны этих клеток поднимаются вверх, Тобразно делятся на поверхности коры, образуя дорожки контактов с клетками Пуркинье. Здесь же лежат клетки Гольджи. 9 15.Функции мозжечка. Регуляция позы и мышечного тонуса, поддержание равновесия; координация работы разных групп мышц при выполнении целенаправленных движений, команда к выполнению которых поступает от моторной коры; регуляция быстрого чередования двигательных актов и их координация с рефлексами поддержания позы; сенсомоторная интеграция и обеспечение точного выполнения целенаправленных движений; адаптационнотрофическая функция (регуляция висцеральных функций с целью вегетативного обеспечения физической нагрузки) III Ситуационные задачи: 1. С целью оценки рефлекторной функции нервной системы у людей разного возраста проводили исследование подошвенного рефлекса. При этом штриховое раздражение поверхности стопы у новорожденных сопровождалось тыльным сгибанием стопы, разгибанием пальцев ноги и их веерообразным расхождением, а у юношей и людей зрелого возраста – к подошвенному сгибанию стопы и пальцев. Есть ли у испытуемых отклонение от нормы? Чем объясняется такое различие в характере подошвенного рефлекса? В обоих случаях – норма. У детей до 3-4 месяцев жизни такая реакция связана с Недостаточным развитием головного мозга и системы кортикального пути. Волокна этого пути проводят тормозные импульсы, которые препятствуют возникновению Онтогенетически более старых сегментарных спинальных рефлексов. При нарушении этого пути поступление импульсов нарушается, что выражается в возникновении рефлекса Бабинского. У здоровых взрослых этот рефлекс отсутствует. 2. Известно, что уровень сознания у человека определяется состоянием нейронов коры полушарий большого мозга. В то же время для наркотизации пациента перед оперативным вмешательством часто используют наркотические вещества, которые не оказывают непосредственного влияния на корковые нейроны. Какая структура мозга должна быть мишенью для этих наркотических веществ? Ретикулярная формация, т.к. она чувствительна к наркотическим веществам 3. Пациент жалуется на шаткость походки. При обследовании обнаружены пониженный тонус мышц, асинергия при движениях рук, дрожание кистей, усиливающееся при целенаправленных движениях (интенционный тремор). Нарушение какой структуры мозга можно предположить у пациента? Перечислите ее функции. Нарушение функций мозжечка. Его функции: регулирование мышечного сокращения, координация произвольных и непроизвольных сокращений, синергия движений, тонус мышц разгибателей, равновесие. 1) регуляция позы и мышечного тонуса, 2) коррекция медленных и целенаправленных движений и их координация с рефлексами поддержания позы, 3) коррекция быстрых целенаправленных движений, регулируемых корой больших полушарий, 4) адаптационно-трофическая, 5) вегетативное обеспечение двигательных функций, 6) тормозная. 10