«Горизонтальный перенос (генов, наборов/фрагментов-их, др.) в различных контекстах (у прокариот и эукариот)»
реклама
 в различных контекстах (у прокариот и эукариот)»")
«Горизонтальный перенос (генов,
наборов/фрагментов-их, др.) в различных
контекстах (у прокариот и эукариот)»
ДЕЙЧМАН А.М.
Лаборатория экспериментальной диагностики и биотерапии
опухолей НИИ ЭДиТО, РОНЦ им.Н.Н.Блохина РАМН
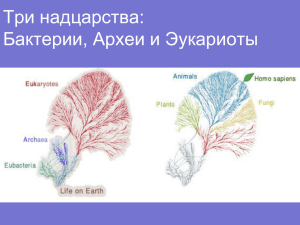
I.
Прокариоты. а) на ~3 млрд.л. «старше»; б) в совершенстве освоен
взаимообмен
именно
генов;
(~4-6
тыс.
генов).
Линн Маргулис: един/огромн/невероят-полиморфн вид. Гены преальфапротеобактер и пре-цианобактер → Яд (симбиогенез).
Прокар-ты представл подавляющ больш генетич, метаболич
(обеспеч. геобиохимич. циклы) и экосистемн разнообразия
биосферы. (2-5 кг; клет: на порядок > хозяйских). Внехромосомн
факторы наследст: плазмидами, вставочн посл-ми и транспоз-ми.
Перенос: Десятк/сотен генов (не менее 5-15 % генома у свободноживущих бакт/архей, и до 8,3% – у патогенных бактерий) (табл.).
«Недавние» переносы: неск сотн млн.л. (~4 гена за миллион лет). На самом
деле перенос мб много бол масштабн (древние акты переноса выявить
сложнее…). Возм не мен 80% генов (на том/ином этапе эволюц).
Мнение: без гориз-переноса жизнь даже за 4 млрд. лет могла не успеть
развитися до человека (вертикальный способ: дарвиновская дивергенция).
И.П.Павлов: «Дабы не обольщаться, следует помнить, что у
микробов есть собственные интересы».
Максимальные переносы – у свободноживущих бактерий (с широкими
экологическими ареалами).
Минимальные – у патогенных бактер из узких экол-ниш (потери генов).
Переносы в осн специфичны (особ «недавние»; «древн» – дивергируют, и
станов общ-достоянием бол-таксона).
Реже всего переносятся гены транскрипц, трансляц, репликац (их белков
продукты «обязаны» вписаться/функционир в сложн белк-комплексах).
Чаще всего переносятся гены, связанные с метаболизм, транспорт-путями и
передач-сигналов.
В составе приобр-сегментов ДНК-бакт часто обнаруживаются профаги,
плазмиды, гены белков рекомбинации.
(Шестаков С.В.):
Cравнительн геномика дает информац о степ сходства гомологич генов в разных
геномах, но не дает информац о том, какие конкр-виды организм были первич
донорами, (особ, – в случ перенос меж представ разных царств; зд. бакт/архей).
Превалир-причина даже адаптив-переноса не ясна: а) то ли завоев новой эколниша, то ли сначал просто реализ факт контакта клеток. Доп-осложнение: цепоч
промежут-хозяев.
Методом «молекуляр-часов» (напр., у геномов E.coli и Salmonella): возмож не тол
приобретен, но и потери генов («правил оптимизац размера генома»). Учет:
геологич-эпох, экол-кризисов и регионал-динамики биоты.
Сам перенос «состоит-в»/«ведет-к»:
1. обмене плазмидами (вкл полов-«пилями»; с ген устойч к лекар-препаратам, др.).
Плазмиды (разл-видов)
могут рекомбинир др с друг и геномами свободно
контактир свободн и принадлежащ различ живот-хозяевам бактерий. Примитив
ретроны бактер обладают обратной транскриптазой. Плазмиды: обыч содерж не
более 200 генов (включ мобил-«гены-адаптации»).
2. ведет к: мутациям множ-генов у бактер-штаммов; плазмид (R, включающих факторы
переноса и устойчивости; F, Ent, JR, pKMR, др.) при лек-устойч (вкл множественн) к
препаратам с разл.-химич-структур. Мутир гены: ДНК-гираз/топоизомераз (регул.
суперскрученность ДНК), метилаз, рибосомал.-белков (L4/L22), 23S-рРНК, лактамаз,
трансферазы групп ААС/APH/ANT (присоединяют к аминогликозидам уксусн/фосфорн
кислоты и аденин), др.
Что приобретают бактерии при горизонтальном переносе
какие отрабат-ся отдельн (и сочетан) механизмов преодол-резистент;
касается анти-/ксенобиотиков, др. субстанц):
а) β-лактамаза может: i) расщеплять гетероциклы β-лактамного-кольца
цефалоспоринов); ii) модифицир активн уч-ки в аминогликозид,
левомицетина, др.);
возможны эфф-ты:
б) уменьш-я проницаемости кл-стенки, подавл-я транспорта, измен-я
рецепторов (лежит в основе устойчивости к тетрациклину);
в) изменения структуры: рРНК (метилир-ся, – в случ. исп.
аминогликозидов, макролидов; ферментов (РНК-синтетазы – в случ.
рифампицина; и др.)
г) приобретен-я уч-в антифаг-активности в отд-локус бакт (уникал-спейсеров, располож между повт-crRNAs в т.н. CRISPR-локусе). (~ РНК-интерфер)
д) формирован-я нов-метаболич-путей;
е) усил эф-т выведения антибиотиков/ксенобиотиков разл-ферм-ми т.н.
«помпы» (транспортерн, каналообразующ, и их связывающ-белками; при
множеств.-лек.-устойч. выводятся сразу несколько вещ-в).
Далеко не все известно про механизм мутаций у прокариот (и эукариот).
Классификация антибиотиков (химически разных)
С бета-лактамным гетероцикл-кольцом (пенициллины, цефалоспорины,
карбапенемы, монобактамы); (наибол «проблемны»)
2. С макроциклическим лактонным кольцом — макролиды (эритромицин и др.) и
азалиды (азитромицин);
3. С четырьмя конденсированных шестичленными циклами (тетрациклин и др.);
4. Производные диоксиаминофенилпропана (левомицетин);
5. С аминосахарами — аминогликозиды (гентамицин и др.);
6. С циклическими полипептидами (полимиксины);
7. Гликопептиды (ванкомицин);
8. Линкозамиды (линкомицин);
9. Фузидиевая кислота;
10. Другие (фузафунжин).
1.
Супербактерии и их фаги «страшнее СПИДа» {множ.-уст-ть практич. ко всем
ан-тибиотикам; имеют ген β-лактамазы NDM-1 (New Delhi Metallo-betalactamase) и др.; касается, в частности, E.coli, нормал-микрофлоры}. Скорость.
Внехромосомные
факторы
наследственности
бактерий:
плазмиды,
вставочные элементы, ДНК/РНК-транспозоны.
Применение антибиотиков (в итоге) развивается:
Устойчивость (с различной скоростью у разных бактерий) может возникать
быстро (к стрептомицину, эритромицину, рифампицину, др.),
постепенно (неомицину, ампициллину, оксациллину, гентамицину, др.),
медленно (нистатину, фурацилину, фуразолину, фуразолидону, др.).
Антибиотикорезистентность
(одному/неск.
бакт.-штам):
бывает
природной и приобретенной. Устойчив-ть обусл специфич.-измен-ми разн
генов, и некот. штам способ (разн-мех-ми) вызывать апоптоз (у разн кл-к: Мф,
Т-кл, эпител-х, эндотел-х).
Чувствительность к некоторым из них (например, ампициллину – в случае
Enterococcus faecalis) не меняется десятками лет.
Сохранение устойчивости, – ресурсо-затратный процесс (через сотни
поколений в геноме могут исчезать функционально не востребованы
последовательности).
Преодоление устойчивости (практические задачи): медицинские, – вне/внутри-больничных инфекций {смертность детей, напр. от стафилококкового
сепсиса (гнойно-воспалит. заболевания) до сих пор снижена недостаточно), др;
(туберкулез: ~50 млн. человек на Земле инфицированы мультирезистентными
штаммами
Mycobacterium
tuberculosis);
ветеринарии,
растениеводстве, с/х и др.
От прокариот – эукариотам.
Эукариотическая одноклеточная диатомовая водоросль Phaeodactylum содержит
сотни (587) генов прокариот: цианобактерий, протеобактерий, архей и др. (5,6%
генома; это гены белков: рецепторных/сигнальных/светочувствительных, др..
Более половины этих генов есть у другой диатомеи (Thalassiosira). Диатомеи
появились 180 млн. л. назад (юрский период). Средняя скорость переноса ~3 гена
(у E.coli 4 гена) за 1 млн. лет.
Массовый перенос, связыв-т с симбиогенезом (зд., гетеротроф-клеток с
одноклеточ-красной водорослью) и образов-м. эукариотич-клетки.
Факты гориз-переноса (генов) становятся много более редкими при переходе от
однокл-эукариот к многоклеточным (но их количество быстро растет).
Между митохондриями растений:
Мт растений активно участвуют в горизонтальном переносе. Приведено 40
случаев горизонтального переноса (2008) Мт-генов между растениями.
Характерно для паразитических или эпифитных (получают энергию и
питател.-вещ-ва за счет фотосинтеза; а влагу – из воздуха/осадков) растений. Необходим плотный (cognate) физический контакт (им-сист).
От бактерий – животным (и обратно):
1.Показано (2007): гены и фрагм/целые геномы паразитических бактерий Вольбахий
(Wolbuhia) могут встраиваться в хромосомы животных-хозяев. В геномах 4
видов насекомых (вкл яйцекл-осы; рис.) и 4 видов круглых червей (филярий)
обнаружены фрагменты генома Вольбахии, а в случае Drosophila ananassae
(рис.) – геном бактерии был встроен целиком. Многие гены Вольбахии, как
минимум, транскрибируются в геноме мух. А в геноме самой бакт-Вольбахии
есть гены, заимствован у эукариотич-хозяев.
Случаи гориз-переноса между животными
и другими клеточными организмами – редкие.
Переносятся, обычно, гены, имеющиеся
у вирусов, транспозонов и ретротранспозонов
(предложен термин «молекулярное одомашнивание»).
А нормал/мутантн популяц вирусов (полиморфн квазивиды) использ-т для собственсохранения / взаимовыгодн-кооперации (комплементации) кл-животного.
НО: часто ключев роль в эволюц млекопитающ играют изменен не в генах, а в
некодирующ-х (в том числе нов-образов) регуляторн посл-стях.
Коловратки (особый случай):
Горизонтал-обмен (генами), считают, зд. столь мощен, что «заменяет» половое
размножение в случ т.н. бделлоидных партеногенетических коловраток (~1500 видов;
однополые первично-полостные черви; размером от 40 нм до 1-2 мм; самые мелкие из
многоклеточн). При этом донорами генов оказывались не вирусы, а гены
симбиотич/паразитич взаимодействующ с ними бактерий, грибов, растений и
животных.
(Марков А.В.): Исключение: коловратки (Adineta vaga),
облад сотн/тысяч чуж-генов (в теломерн/центральн
участ хромосом); не извест ни для одного другого
животного.
Раб-гипотез (необыч образ жизни): перенос (как
пыль) между мелк-пересых лужами. При высых
образ разрывы хромос и поврежд мембр-кл-к
(заносит чужер-ДНК). Репарац наступ при увлажнен;
одновр происх включен чужер-фрагм-ДНК (репарац игр необыч роль).
Вывод: «Вероят, в эволюц многоклеточ гориз-перенос играет бол заметн роль, чем
известно на сегодняшн-день.
Пока прочтены десятки геномов многоклеточных, – трудно оценить реал-масштабы
явления. Когда он пойдет на сотни, как у прокариот, тогда, возможно, мы узнаем много
нового о гориз-переносе у высш-организмов».
(Шестаков С.В.):
Гориз.-перенос – главн. источн. инноваций, возникнов нов-генов, радикал меняющ
свойства и расшир адаптацион потенциал клеток (участв в видообразовании).
Считают:
Перенос генов завис от:
у прокар – плазмид, фагов, мобильных элементов (транспозонов/ретронов).
у высш-эукариот (редкий) – почти исключит от МГЭ (вирус; транспозон,
ретротранспоз):
а) Сист-приобр-иммунитета (Ig’s-сегмен) и плацента (транспоз-подобн ген Peg10).
б) Некот транспоз геномов риса / проса ведут себя как челночн / самосохран вирусы
с вторич-приобр инфекц-ю: они дивергир ~30-60 млн.л.н., накопил различ, но оказ-сь
практич идентич.
В ходе эволюц (вертикал): усил автономизация, барьеры, модиф систем деградац
"чужой" ДНК, репаратив-механизм (препят/огранич гор-перенос). Частота горизпереносов (генов) была наибол выс на ранних этапах, – и снижал с эволюц высш
эукар (и появлен эндогамных видов). (м: мб просто не представл как это возмож)
Чуж-ДНК в геноме обнар по косв-призн: 1. (ГЦ)-содерж (профаг, МГЭ, кодир/некодир
обл-х, т.н. "остров" патогенн, «хромосомн-территор-ми», др.; 2. част встреч опр кодон.
амелиорация кодонов/посл-й (за сч мутац/рекомб +...). 3. положен гена в разл филогендревах. 4. др. Ядер-геном эукар: следы бактер/архейн генов происхож-я (Мт/Хп→Яд).
Перенос: эукариот → прокариот, как у бакт Вольбахии (соотв ГЧОС-сист, далее).
Перенос в симбиотч/паразитич сист-х треб-т плот-физ-конт и челн-вект.
Примеры переноса:
1. У археи (Pyrococcus) донором эукар-гена термоуст Аа-тРНК-синт-зы мб морск-жив
полихеты.
2. У патоген-бакт (риккетсий/хламидий) обнаруж ~20 генов белков эукар-хозяев
(транспортирующ АТФ/АДФ, протеазы, ингибиторы им-отв, поверх-рецепторы, др.).
3. У Риккетс (возб-тифа): неск генов, гомологич таков у животных-соврем-хозяев.
4. У Хламид (возб хрон-инф-забол-глаз, трахомы), – гены растен (прежних хозяев).
5. Хламид↔Риккетс: перенос ген-патоген одной экол-ниши/организ-млекопит.
Важно: перенос фрагмент-генов, т.е.
отд-доменов,
рекомбинир с хоз-генами.
Образ-ся друг/нов гены и псевдог (с нов локал/узнаван/сигнал-свойств, др).
Пример: для >40 одинак белков (эукар/бакт/архей), – появлен сравнит «нов/чуж»
доменов, не успевш «усредн-ся» и ответств за бел-бел/ДНК-бел взаим-я и регулятфункц.
Партнеры: филоген близк / отдален организмы. Перекомбинации доменов. (рис.).
Важно: Сами мех-мы переноса/рекомбинац также явл
объектами эволюц-процесса. В периоды глобал-гео-биосферн-климатич-экол перемен: диапаз вект-сист (вирус,
плазмид, МГЭ) - меня-ся, а рекомбинац акт-ть - усилив.
Дейст-т т.н. «волны переноса» в услов повышен-я
плотности контактир популяций многокомпонентнсообществ.
Ранн-этап эволюц: общ-генное "коммунал-хозяйство", активн-горизонт обмен
генами. Клетки автономизировались внутри таксономич линий.
У прокариот показано, а для эукариот предполагают: картин эвол-связ – не стол
ветвящ древо, скол своег рода мицелий с переплет сетью гориз-связ самых разнообр /
/ неожид направлений (в зав от клет-контакт в общ/перекрыв экол-нишах).
Поэтому геномы прокариот и эукариот мозаичны.
Массив-генн переносы происх внутри/между царств. При усложнии: развив полов
размнож и репродукт изоляц, перен генов станов бол редким явл-ем (Но: слиян двух
гамет в диплоид-кл, последующ рекомбинац, – в некот смысл это также специф.-вид
гориз-переноса). Мног животн (но не коловратк), утрачив полов размнож и переход к
партеногенезу (таких случ не мало), оказыв-сь склонн к быстр вымир, растворению в
диверг-виды, – но не образ-ю беспол-семейств/отрядов.
(Гор-перенос по Кунину Е.В.):
Иссл экзон/интр-струк совр-эукар показ:
1. масс-потер генов (у однокл-грибов);
2. возник-е бол числ нов-генов (у живот).
3. у некот параз-бактер > 80% генов безвозвр потеряно за последн 200-300 млн. лет
(корот по эвол-меркам период).
Связь потери генов паразитир-бакт-ми (п.3) с переносом их в геномы многокл-эукар
(п.2, животн) остается неясной. Но массов-потери генов явл характер черт эвол-эукар
(по кр м в некот линиях).
Луи Долло (1893 г.) сформул закон «the Law of Irreversible Evolution»: «Сложн биолсистемы, потер/изменивш у организм за эвол-значим срок, не могут снова появит в
своем исходном виде» (но: возм ~ парал происхожд).
«Вопрос о перенос (имен) генов межд эукар-ми остает открытым. И даже доминир
т.зрен. (сейчас): этот процесс не явл-ся существен для эволюц эукариот» (!?).
(по Колчанову Н.А.; рис / схема ↓↓↓↓):
У эукариот (с их малочисл-популяц) име-ся две разновидн гориз-переноса:
а) меж Ядр/(Мт; + ХП у растен), одной клетки (внутриклеточный);
б) меж Ядр разных клеток (одного/разных организмов).
Опасн переносы: наруш кодиров-е транскрипц.-фактор, – ключ-генов глобал/локальн генн-сетей, вызывающ системн-мутации.
Важно: сложность некодир-регуляторн-област генома бол чем на порядок превыш
таков у кодирующ исполнит-компоненты эукар: одной мутации в гене соответств-ло
~13 мутац в некодирующ-части. Так образ, мишенями и регуляторных и эволпреобразован явл-ся, в перв очер, некодир-регуляторные системы организмов.
Цит. про СПИД: ...«ВИЧ облад слабой способ-ю к переносу (бактер, риккетс,
простейш, кровососущ и гельминтами). Подоб заболев редко выход на уров пандем.
Этом вирусу "помог" гор-перенос в чел-попул (гомосекс-связи, перенос с засохш
кров на шприц наркомана)). {это упрощение: ВИЧ, скорее, также использ дополнитспособ гор-переноса: до своей упаков в оболоч он, не искл, мож перехват предназч не
для него ген-информацию (нукл-вектор), вырабат прежд всего при норм-имм ответе
(др. кл) и переносим прежд всег клетк имм-сист (АПК/Т-хелп; или их частиц типа
экзосом), и, прежде всего, в ходе норм-специф-имм-ответа (далее)}.
Место горизонтал-переноса (упрощ)...: Генн-сети (из работ Колчанова Н.А.):
Экзосомы, слущив микр/мульт-везикулы (SMV),
апоптотические тельца (AB).
Многообразие видов белков экзосом
Внеклет-везикулы, секретируем из различ ствол кл (кост-моз/стромы/сердц/др.),
способны, в частности (и с участ липос / рост-фактор / цитокин):
1. защитить клетки от повреждений (апоптоза)
2. стимулир: пролиферацию клеток / васкуляризацию поражен-тканей., др.процессы.
II. Горизонтальный перенос в контексте 2-х гипотетических
механизмов (схематично) у эукариот:
1. Образов олигонуклеотидн (мини / микро-сателлит-подобных, – и сенс / антисенс-уникальных) послед-тей, т.н. олиго-НЭ. Детали опустим.
Образуются олиго-НЭ из эпитопов белка. Эпитопы – линейн / конформац
(длиной до 5-10 аминокисл). Место действия: а) внутренняя мембрана
митохондрий, Мт (у растений также Хп / Тлк); б) с участ специал-самоорганизующ-ся структуры, – «ретранслосомы» (в кот могут быть необходим
активности: протеазн, полимеразн, лигазн, эндо/экзо-нуклеазн, др.).
Механизм назыв-ся вариабельная Поэпитопная Обратная Трансляции
(вПОТ-механизм) отдельн эпитопа. И т.д. (есть в опубл-работ, и на сайте
www.amdeich-var-reverse-translation.ru). Данн мех-м пост оказ давлен.
2. Перенос содержащих олиго-НЭ (не все известно) в: а) вектор-подобных
{вирус/(ретро)-транспозон-подобных} последов-тей; б) информосомподобных частиц (как экзосома, др. микро/мульти-везикулярные частицы;
активны внутри/между тканями); в) с белк, РНП; г) с липидами {подобн
микроРНК/липиды-выс-плотн-(HDL)}. Изв устойч в крови/плазме ДНП-комплексы.
Олиго-НЭ посл-ти могут (конспективно):
i). взаимод-ть со следующ множеством отдел / перекрывающ,
постоян / динамич-возникающ олигонукл-сайтов в геноме, в
ассоц с ним белках (и в новосинтез.-РНК):
1. рестрикции (тысячи);
2.
«ломкими сайтами» (много);
3.
«горячими точками» (много);
4.
модификации ДНК / нуклеосомн-белков: метилирования (ДНК-по-CpG / гистонов /
/ др.-белков); ацетилирования, фосфорилирования, убиквитинирования
нуклеосомных белков (оч-много);
5.
мутационными сайтами (много);
6.
начала репликации Яд / Мт-ДНК (ориджны=ori; тысячи/несколько);
7.
транскрипционными сайтами (промоторных, энхансерных, экзонных / интронных, 5’ / 3’-концевых областей, др.) (много);
8.
повторяющихся последовательностей (мини / микро-сателлитными, др.
представленными в некодирующей/кодирующей областях);
9.
рекомбинации (гомологичной/негомологичной; др.);
10. негативной регуляции трансляции (в мРНК) ;
11. интерференции РНК (микроРНК / мРНК-мишень);
12. редактирования РНК (вкл экзон/интрон дуплексы);
13. сплайсинга мРНК / РНК-(др.);
14. транспозиции / ретротранспозиции (за счет вз-я с концев-повторяющ-посл-ми);
15. посадки праймеров репликации (ядДНК / мтДНК);
16. посадки праймеров фрагментов Оказаки (десятки миллионов) реплицир-ДНК;
17. теломерных / субтеломерных, центромерных / субцентромерных,
эухроматиновых / гетерохроматиновых, кодирующей / некодирующей,
нуклеосомных / межнуклеосомных областей;
18. релаксирования / скручивания / суперскручивания ДНК; 19. др.
ii). Это может активировать / модифицировать (количественно /
/ качественно) следующие процессы:
1. эндонуклеазный надрез;
2. различные виды рекомбинации (гомологичной/негомологичной; др.);
3. Транспозицию / ретротранспозицию;
4. различные виды мутаций (транзиции/трансверсии/делеции/вставки;)
5. репликацию (которую можно приостановить по каждому фрагменту
Оказаки);
6. репарацию (различные ее виды);
7. транскрипцию;
8. трансляцию;
9. расширения / сжатия повторяющихся последовательностей;
10. эксперессию генов / генома;
11. синтез лидирующей / отстающей нитей ДНК (Яд / Мт);
12. взаимодействие с вирусами (РНК / ДНК);
13. взаимодействие с бактер / внутрикл патогенами (их поверхностн / др.белками, ДНК / РНК);
14. интерференцию РНК;
15. редактирование РНК;
16. точечн / блочн мутацию ДНК;
17. соотношение гетеро- / эухроматиновых областей;
18. обмен нуклеиновыми последовательностями между клетками;
19. др.
iii) В случае онкологии такими активирован / модифицирован могут
оказаться некотор из в.н. и дополнительн процессы:
1. усиления пролиферации;
2. усиления / ослабления экспрессии, соответственно, онкогенов /
/ антионкогенов;
3. усиления / ослабления, соответственно, гипо / гипер-метилирования ДНК
по CpG-сайтам внутри / поблизости / отдаленно по отношению к генаммишеням;
4. инициация / усиление метастазирования (инвазивности, др. опух-хар-ки);
5. Рекомбинации / транспозиции / репарации / мутации/ др. процессы;
6. усиления микро-(в-осн-3-нукл) / минисателлитной и (ретро)транспозоновой нестабильности ядДНК (в отношении транскрипцфакторов, белков / ферментов / их-комплексов при репликации,
репарации, др.). Касается раков: прямой / толстой кишки, жедудка,
молочной железы, легких, мочевого пузыря, простаты, яичника, почки;
некоторых менингиом, лейкозов, миелом, др.;
7. усиления микро-/минисателлитной нестабильности мтДНК. Касается
раков: прямой кишки, желудка, печени, поджелудочной железы, легких,
молочной железы, почки, яичника, пищевода, простаты, лейкоза.
8. изменение соотношения и активности гетеро / эухроматина;
9. и т.д.
Горизонт перенос 2-х видов (эукар):
1. между клеточн-органеллами: Мт→Яд {мт-псевдог чел заним десят
млн.п.н.; [наиб част перено-ся фрагм в 8-9 п.н.; (Малярчук Б.А.)]}; коротк мтРНК
мог регул экспрес мт-/ядерн-бел генов (вкл транскрипц-факт), др. {У растен, –
переносы, в том числе: а) крупн-фрагм-тов в неск.-тыс.п.н., и б) асимметр
(предпочт в Яд), между всеми Мт/Яд/Хп-органеллами}
2. между клетками:
а) одного организма: {показан переносы: i) РНК, при поиске т.н. «иммунРНК» («супер-антигена»), от Мкрф к Лфц (Карр Я., 1978), др.; эти исслед приостанов по разн прич; ii) мРНК регуляторн-белков от трофоцита к ооциту домашнмухи (Корочкин Л.И., 2002), др. (оба случ треб тесн-физич-контакт); iii) экзосом
{разл-видов селект-РНК, в том числ челночно-курсир между разн кл-ми; белк-связРНК / шаперонов / бел-факторов / адгезии / MHC / некот-др.-бел; липидов (бол
десятка видов); ионов-Ме; возм ДНК?; экзосомы (по бел-кам) происх из болкрупн-эндосомн пузырьков, еще не закисл и не преврат/захват в лизосомы}; iv)
гипотетический (в отнош ретропозон-подобн-ВНП с вариантом олиго-НЭ внутри),
также треб физ-контакт (как и в перв 2-х случ; см. схему имм-варианта).
б) разных организмов (челночные векторы; в паразит / симбиотич систем,
др.). (разработана система Генетической Челночной Обратной Связи, ГЧОС-сист)
1. Возможный горизонтальный перенос между иммун-клетками
Треб физ-конт (рис.3): Чуж-АГ (эпит) → АПК (вПОТ: обр олиго-НЭ, кот встраив в
ретропоз-подобн-посл → CD4-хелп клетке (ВНП-передача) → НДП, низкодиф предшеств
в кост-мозг / тимус. Гориз / псевдогор-перенос (ВНП-передача).
Физ-конт не обяз: изв стабил внекл компл РНК/ДНК: 1. с бел: а) РНП: с разл микроРНК-чел; секреторно-циркул-х регуляторн РНК [срРНП (Ситников А.С.)]; б) изв стабил
ДНП; 2. с липид: РНК/липопротеин-выс-плотн (HDL). Все обнар-ся в сыворот / плазме
крови в свобод виде. 3. Кр того, перенос возм-н с участ: г) экзосом и др. везикулярнчастиц.
Интересн, показано: а) кл-костн мозга, взаим-я с
микровезик (cод-ми мРНК/miRNAs) повреж / стресс-ткан,
мог вступ в конт и стимул в таких ткан синтез необх
мРНК. С др стор, изв: б) инструм-м взаим ствол-х кл с кл
ниш-микроокруж (и кл-матрикс) также явл микро-РНК.
Важно: По мнен некот автор имм-сист мож участ в
контроле не тол «качества» (поверх-бел), но и оптимколич-ва клеток тканей: напр, когда «мало апоптоза»
в эт кл-х → опух, много → атрофия).
(Гипот «кл-двойников»).
Как вышли на кост-мозг:
вперв экзосомы иссл-сь в связи с дифференцир
ретикулоц-в в эритроциты. Оказал (конец 1990-х), экзосомы облад способ-ю регул-ть
имм-реакции организма (служ т.н. «конвейер-передатчик»); это один из видов межклкоммуникац с их участ. Экзосомы – посредники мног-биол-проц (в частн, апоптоза,
роста / метастаз рак-кл, ангиогенеза, метаболич реакц, и мн. др.).
2. Возможность горизонтального переноса в половых клетках:
Теор пангенезиса Ч.Дарвина и др. учен (наследов признаков 1868 г., в книге «Изменя домашн животн и культурн растений»). Субмикроскопич гранулы (микроскоп с 17
в.), — «геммулы». Близк гипотез наслед-ти выдвиг Гиппократ (5—4 в. до н.э.;
«экстракты из организма собир-ся в мужск / женск зародыш-элементах»), Джв.
Борелли (17 в.; ), Ж. Бюффон (18 в.). Двигаясь с током крови из сомы, «геммулы»
собир-ся в полов-элем-х. (→ теор. зарод.-плазмы А.Вейсмана).
Резул-ты иссл француз-учён (2010) позвол им выдвинуть на роль «геммул» Дарвина
т.н. микровезикулы (усил-но выдел-ся в норме и при многих / всех патологиях;
частично изготавл-ся только на экспорт; напомин липосомы в опыт ген-инженерии;
это мембр-пузырьк в 30 нм – 1-4 мкм; встреч во всех жидких средах организма и до
недавнего времени считавшиеся лишь побочным («мусорным») продуктом
жизнедеятельности клеток.
Предпосылки (некотор) переноса :
1) забарьерн органы (мозг / плацента-эмбрион / полов-кл) не явл абсолютно
непроницаем для экзоген / эндогенн агентов: вирусов (нукл-векторов), везикулярчастиц (типа экзосом) и нуклеопротеидн-комплексов (РНП, как в случ микроРНК;
ДНП) и кл-к (малых Лф, метастазир-кл), др.
Перенос в герминат-кл мб связ не тол с вектор (вирус/нукл-посл), но и с продуктами
их взаим-я с ядер/митохондр геномами.
2) в кажд ткани, извест, экспрес собств набор белк / эндог-ретровир-элементов; и
каждая ткан явл потенц плацдармом для предпочтител «посадки» определ-спектра
экзогенных ДНК/РНК-вирусов (включая ретровирусы).
Например, вирус паротита («свинки») тропен к ткан слюн-желез/яичек; ЭБВ-вирус
(герпесвир-чел IV типа) тропен к В-лф и эпител-кл (за счет мол-CD21 – рецепт-фраг-
комплемента C3d); вирус геморр-лихорад тропен к эндотел-сосудов; вирусы грип
А/В, полиомиел (18 видов), – кардиотропны (миокардит); ВИЧ тропен, прежде всего
(за сч CD4-рец), к Мф, ДК, Т-хелп, нек др. Т.обр.: один вир мб тропен к неск-тканям, и
наоборот, отд-ткань мож абсорб-ть разл вирусы, т.е. существует, по кр мере,
огранич их перекрестн специфич-ть (напом соотн микроРНК / мРНК; фрактал ряд).
3) для переноса нукл-посл-й их необх интенсив нарабат-ть, – и некот акт-пролиферир
норм и опух клет делают это наибол «рьяно».
4) в организ всегда есть нескол тысяч потенц-малигнизир кл-к с бол пластич-геномом и
повыш пролиферативн потенциалом.
хххххххххххххххххххххххх
5) клетка может выполнять 2 программы (гипот): «в интересах» а) себя / организма, – и
б) биол-вида (т.е. иногда против организма). Не ясно для чего бόльш необх перенос
(даже корот-посл-тей)?: i) для внесения готовых / окончательн вариантов
(способных оказаться в «нужном»-сайте генома оплодотв-яйцекл / эмбрионе); или
ii) в качест инструмента регул воздей-я на уже имеющ-ся локусы генома (где уже
имею-ся мног необх компон-ты.; возмож оба вар-та).
6) Сущ противоб: а) разл-видов барьеров и б) пост давлен {(псев)-/гориз-перен} на
геном полов / сомат кл с помощ разн (вкл гип мех: вПОТ / ВНП-пер / ГЧОС-сист).
7) Возм природа консервати-ти посл-й.
Проц интеграц в эукар-геном: (вкл искус-трансген, введен в нукл-вектор или ретровир)
сложен и многоэтапен. На каж этап возм обрыв звена общ цепи процес интеграции.
Разл спос переноса на разл стад оплодотв / эмбриоген связ с разл их путям взаимод с клеточ компон (мембр, др.); эписома – одна из промеж-форм; интеграц мож
вовсе не состояться, оказ-ся по разн экспрессируемой и мозаично представл в
разн-органах / ткан, может «затухать» от поколения к поколению, и т.д.).
Соображение: перенос небол-посл-ми более предпочтит, т.к. их легче «вписать» /
«зарегулировать», т.е. взять под контроль и включить в общ программу генетичстроительства / регуляции в последующ онтогенезе.
{Далее следующие рисунки / схемы касаются темы горизонтального
переноса как в изложенном, так и в более широком / биосферном контесте
(на основе работ сайта: www.amdeich-var-reverse-translation.ru)}
Рис. 1 и 2 – два из 3-х авторских вариантов вПОТ-механизма.
Следующий слад – экспериментальные этапы и схематический вариант
«обратной трасляции» (rT-mechanism) по M.Nashimoto (2001).
Рис. 3 – горизонтальный перенос между клетками иммунной системы
(имеется выше ).
Схема 1 – формирование универсального генетического кода (УГК) в
хлоропластах/ /тилакодах, Хп/Тлк.
Схема 2 – связь редактирования РНК, полиморфизмов ДНК/РНК с
гипотетическим вПОТ-механизмом.
Схема 3 – связь различных механизмов регуляции/эволюции генома с
гипотетическим вПОТ-механизмом.
Схема 4 – функционирование системы Генетической Челночной Обратной
Связи (ГЧОС-системы) между разными организмами сообщества / группысообществ.
Рис. 1 и 2 – два из 3-х «авторских» вариантов вПОТ-механизма.
Экспериментальные этапы (a-c)
и
схематический вариант (a-h)
«обратной трасляции» (rT-mechanism) по M.Nashimoto (2001).
Схема 1 – формирование
универсального генетического
кода (УГК) в хлоропластах/
/тилакодах (Хп/Тлк).
Рис. 3 – горизонтальный
перенос между клетками
иммунной системы (имеется
выше).
Схема 2 – связь редактирования РНК, полиморфизмов в ДНК/РНК, – с гипотетическими вПОТ/ВНП-пердача-механизмом.
Схема 3 – связь различных механизмов регуляции/эволюции генома с
гипотетическим вПОТ/ВНП-передача-(ГЧОС-система)-механизмом.
Схема 4 – функционирование системы Генетической Челночной Обратной Связи
(ГЧОС-системы) между разными (фото-/нефотосинтезирующими; про-/эукариотическими) организмами сообщества / группы-сообществ.