miR-662
реклама

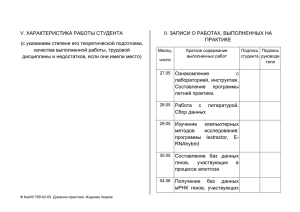
Микро-РНК и эволюционное становление человека современного типа Н.А. Колчанов, К.В. Гунбин, Д.А. Афонников, А.П. Деревянко Институт цитологии и генетки СО РАН Институт археологии и этнографии СО РАН Новосибирский государственный университет Национальный исследовательский центр «Курчатовский институт» Долина р. Ануй By A.P. Derevyanko and M.V. Shunkov, IAE SB RAS В 2007 году ученые Института археологии и этнографии СО РАН под руководством профессора Деревянко нашли фрагменты тел денисовских людей, новых эволюционных родичей H. sapiens. Денисова пещера возле реки Ануй. Алтайские горы Возраст культурных слоев в Денисовой пещере около 80 тысяч лет. А.П.Деревянко и М.В.Шунков, ИАЭ СО РАН Большое число артефактов, найденных в Денисовой пещере, свидетельствует о высоком уровне интеллекта и технических способностях H. denisova Что касается неандертальцев, то многочисленные археологические находки показывают, что они не были достаточно искусными, чтобы изготовлять сложные орудия или утонченные артефакты, особенно украшения. А.П.Деревянко и М.В.Шунков, ИАЭ СО РАН Секвенирование ДНК из фрагментов тела древнего человека было проведено S. Paabo в Институте эволюционной антропологии общества Макса Планка в Лейпциге. Денисовцы близки к современным людям и к ныне вымершим неандертальцам, но все же отличаются от тех и других. Геномы ряда представителей современных этнических групп содержат ~ 5% ДНК денисовцев, что является убедительным доказательством скрещиваний между предками человека и денисовцами во время их сосуществования. миРНК: эволюция H.denisova и H. neandartalensis ~0.6 MYA ~0.8 MYA ~6.2 MYA ~6.5 MYA ~16 -12 MYA H. neandarthalensis H. denisova человек шимпанзе горилла Эволюционное дерево гоминид и H. denisova/H.neandartalensis (вымерших родичей H. Sapiens), чьи геномы секвенированы к настоящему времени орангутан макака (аутгруппа) В отличие от большинства подходов, сосредоточенных на кодирующих участках генов, наш подход к анализу геномов человека, других гоминид, а также древних людей был обращен на регуляторные механизмы, управляющие экспрессией генов. Почему мы рассматриваем регуляторные механизмы, контролирующие экспрессию генов, а не кодирующие участки генов? Обширные данные, накопленные к настоящему времени, показывают, что: 1) самые быстрые и значительные эволюционные преобразования организмов связаны с изменениями в регуляторных системах и процессах и 2) мутации, оказывающие действие на регуляторные системы и процессы, могут иметь наиболее значительные фенотипические следствия в ходе эволюции. Созревание и функции миРНК Ген миРНК Транскрипция Дуплекс пре-миРНК DICER Пре-миРНК Расщепление пре-миРНК Пре-миРНК DROSHA Зрелая миРНК Ядро Пре-миРНК Подавление трансляции RISC Рибосома RISC Разрушение мРНК миРНК в комплексе с RISC белком может комплементарно взаимодействовать с мРНК-мишенями, подавляя их трансляцию или способствуя разрушению мРНК. Каждая миРНК в геноме человека имеет от десятков до сотни мРНК-мишеней, являясь кассетным регулятором экспрессии определенной группы генов. Поэтому мутации, вызывающие серьезные структурные повреждения даже в нескольких миРНК, могут сильно влиять на трансляцию больших групп мРНК-мишеней. Таким образом, даже одиночная мутация в миРНК может привести к множественным фенотипическим проявлениям. мРНК миРНК Трансгенные мыши: примеры множественных фенотипических проявлений, вызванных одиночными нуклеотидными заменами в генах пре-миРНК (PLoS ONE (2012) 7(1): e30737) миРНК mmu-miR-96 mmu-miR717 Замены A>T A>G Фенотипические проявления Потеря слуха (Lewis et al., 2009) Истощение, поведение, биохимия крови, общий анализ крови, липиды крови, прочие свойства крови, строение тела, вес тела, рост и размеры, кости, мозг, характер дыхания, рак, сердечно-сосудистые проблемы, поражения клеток и тканей, уши, эндокринная система, желчный пузырь, иммунная система, пищевые предпочтения, почки, печень, мышцы, дыхание, синхронизация ощущений, селезенка (Kunej et al., 2010; Zorc et al., 2012) Поиск генов пре-миРНК в геноме H. denisova Шаг 1. ENSEMBL-выравнивания геномов шести видов приматов. Картирование 1523 экспериментально верифицированных человеческих пре-миРНК из miRBase на выравнивании. Отобраны эволюционно консервативные гены человеческих пре-миРНК (ортологи по крайней мере в одном виде приматов помимо человека). Человек Шаг 1 Шаг 2 Pongo Macaca Gorilla Pan H.d X H.s H.d Шаг 2. Полученный набор эволюционно консервативных генов миРНК человека и их положения на хромосомах Homo sapiens были использованы для идентификации ортологов пре-миРНК в геноме H. denisova. Шаг 3. Мы исключили из анализа все пре-миРНК с низким качеством секвенирования и неуникальные пре-миРНК (более 1 копии на геном), чтобы быть уверенными в функциональной значимости пре-миРНК у древних людей. H.d Всего в геноме H. denisova было найдено 1419 миРНК – ортологов человека Все эти миРНК подразделяются на две группы: • нейтрально эволюционировавшие миРНК (большинство рассмотренных миРНК); • миРНК с не нейтральными заменами, нарушающими их структуру и/или функцию. Всего было выявлено 29 таких пре-миРНК. Из них для 11 имеется экспериментальная информация по мишеням их ортологов у человека. miR-609 A ---ug g U gu aaug cuc gcuguuccuAGGGUGUU CUCUCAUCUCUg cuau g ||| ||||||||||||||||| |||||||||||| |||| gag cgacaaggaucccacaa gggaguagagau gaua g ccaag a c # -aauu Замены в этой пре-миРНК найдены на участке зрелой миРНК, взаимодействующем с мРНК-мишенью miR-532 c a cucuccuc U A A CC uggca g cuugcuuu CA GCCUUG GUGU GGA GU u | |||||||| || |||||| |||| ||| || c c gagcgaaa GU CGGAAC CACA CCU ca u u c ----aaAC U C C CC uuaau G # G # Замены в этой пре-миРНК найдены в позициях, важных для процессинга и созревания Идентификация мРНК-мишеней для ортологов миРНК человека из генома H. denisova Экспериментально доказанные мРНК-мишени (H.s.) Humans H. Denisova Потенциальные мРНК-мишени (H.d) Общее число аннотированных миРНК – 543; количество мРНК-мишеней для этих миРНК - 8124 (т.е. в среднем 16 мРНК-мишеней на 1 миРНК); Количество аннотированных – мРНК-мишеней для 11 НЕНЕЙТРАЛЬНО эволюционировавших миРНК– 1102 (т.е. в среднем 100 мРНк на 1 миРНК, что в 6 раз выше чем для общего пула миРНК). Поэтому мы называем НЕНЕЙТРАЛЬНО эволюционировавшие миРНК кассетными МЕГАРЕГУЛЯТОРАМИ экспрессии генов. starBase ( stores information about tens of thousands of miRNA-targets in Human mRNAs. The mRNA/miRNA interactions were confirmed by: (1) CLIP-seq experiment(s) and (2) computer simulations Информация о числе потенциальных мРНК-мишеней для миРНК H. denisova была получена путем экстраполяции экспериментально установленных сайтов мРНК/миРНК взаимодействий, описанных в базе данных starBase для генома человека. Для вычисления регуляторного потенциала ненейтрально эволюционирующих миРНК использовались экспериментальные данные о тканеспецифичной экспрессии 18026 человеческих мРНК в 131 тканях, содержащиеся в Gene Expression Barcode Database (http://rafalab.jhsph.edu/barcode) Для вычисления регуляторного потенциала ненейтрально эволюционирующих миРНК использовались экспериментальные данные о тканеспецифичной экспрессии 20 936 мРНК человека в 2200 структурах мозга, содержащиеся в Allen Human Brain Atlas: (http://human.brain-map.org) / По всем тканям, описанным в этих базах данных, была проведена оценка величины регуляторного потенциала ненейтрально эволюционировавших миРНК миРНК RISC Рибосома Регуляторный потенциал взаимодействий миРНК/мРНК для всех не нейтрально эволюционировавших миРНК в k-й ткани: Jk real Lk Ekj* Jk E j 1 j* k – уровень j-й мРНК-мишени, регулируемой некоторой не нейтрально эволюционировавшей миРНК, в k-й ткани, оцененный согласно Gene Expression Barcode Database. - общее число мРНК, регулируемых не нейтрально эволюционировавшими миРНК, в k-й ткани. simulated P( L k Lk ) real 100 000 симуляций случайных пулов мРНК ПРИМЕР 1: Тестирование вклада ненейтрально эволюционирующих миРНК в регуляцию экспрессии мРНК в энторинальной коре человека Данные по тканеспецифической экспрессии мРНК взяты из Gene Expression Barcode Database. Lreal k Lsimulated k В случае энторинальной коры p=0.00008. Вклад ненейтрально эволюционирующих миРНК в регуляцию экспрессии мРНК в энторинальной коре существенно выше случайного уровня для этой ткани. ПРИМЕР 2: Тестирование вклада ненейтрально эволюционирующих миРНК в регуляцию экспрессии мРНК в миндалинах человека Данные по тканеспецифической экспрессии мРНК взяты из Gene Expression Barcode Database. Lreal k Lsimulated k В случае миндалины p=0.5408. Вклад ненейтрально эволюционирующих миРНК в регуляцию экспрессии мРНК в миндалинах не отличается от случайного уровня для этой ткани. Gene Expression Barcode Database: объединение результатов по линиям человека и денисовца. Отобранные ткани-мишени с повышенным регуляторным потенциалом мРНК/миРНК взаимодействий, опосредованным ненейтрально эволюционирующими миРНК Ткань/орган (Barcode 2.0) • • • • • • • • • • • • dorsal root ganglia hippocampus entorhinal cortex superior frontal gyrus cerebellum cervix fallopian tube epithelium myometrium ovary breast lobular cells cecum head and neck epithelial cells Вероятность наблюдения суммы величин экспрессии (p) 2.00E-05 6.00E-05 8.00E-05 0.00032 0.00054 4.00E-05 2.00E-05 0.00014 0.00014 0.0007 6.00E-05 4.00E-05 Gene Expression Barcode Database Примеры структур человеческого мозга, ответственных за сложные интеллектуальные и познавательные функции, которые характеризуются повышенным потенциалом регуляторных взаимодействий, определяемым ненейтрально эволюционировавшими миРНК HIPPOCAMPUS (p = 0.00006) играет важную роль в консолидации информации из оперативной памяти в долговременную и в ориентировании; ENTORHINAL CORTEX (p = 0.00008) отвечает за память и ориентирование; UPPER FRONTAL GYRUS (p=0.00032) ответственна за самоосознание; СEREBELLUS (p=0.00054) ответственен за координацию движений. Allen Human Brain Atlas: найдено 48 структур-мишеней мозга с повышенным регуляторным потенциалом взаимодействий мРНК с ненейтрально эволюционировавшими миРНК Макроструктура Wernicke's area Primary visual association areas Limbic system Primary motor area Primary somatosensory and Primary somatosensory association areas Orbitofrontal cortex Major association cortex Middle frontal gyrus Superior frontal gyrus Precuneus Primary visual cortex ID микроструктуры (Allen Human Brain Atlas) Микроструктура (Allen Human Brain Atlas) Вероятность наблюдения суммы величин экспрессии (p) 4142, 4143 4135, 4136 4149, 4150, 4151 4178 4193, 4194 4158, 4159, 4160 4200, 4201 4244, 4245 4280 4223, 4224, 4230, 4231 4288 4258 4073, 4074 4012, 4013, 4014, 4015 4098, 4099 middle temporal gyrus superior temporal gyrus inferior temporal gyrus planum polare lingual gyrus fusiform gyrus occipito-temporal gyrus parahippocampal gyrus Head of Caudate Nucleus Cingulate gyrus Putamen Dentate gyrus paracentral lobule precentral gyrus supraparietal lobule 0.00016 0.00048 0.00074 0.00148 0.00112 0.00154 0.00638 0.0007 0.00082 0.00124 0.00218 0.0027 0.00054 0.00376 0.0006 4087, 4088, 4089, 4090 postcentral gyrus 0.00132 4901 4113, 4114 4030, 4031 4023, 4024 4120, 4121 4186, 4187 inferior rostral gyrus angular gyrus middle frontal gyrus superior frontal gyrus precuneus cuneus 0.00194 0.0011 0.00024 0.0006 0.00042 0.00042 Примеры структур человеческого мозга, которые характеризуются повышенным потенциалом регуляторных взаимодействий с ненейтрально эволюционировавшими миРНК 1. Wernicke's area - один из двух основных участков коры головного мозга, связанных с речью ( участвует в понимании письменной и устной речи). 2. Major association cortex играет важную роль в абстрактном мышлении*) 3. Superior frontal gyrus участвует в самосознании и в координации работы сенсорных органов. 4. Middle frontal gyrus контролирует осознанные движения глаз, участвует в самосознании личности, обуславливает такие качества как проницательность и дальновидность. 5. Precuneus участвует в работе временной памяти, обрабатывает визуальнопространственную информацию, участвует в самосознании. 6. Limbic system контролирует питание, воспроизведение, борьбу и бегство, чувства, и память. Она включает в себя области, энторинальной коры. 7. Orbitofrontal cortex представляет собой основной центр обработки обонятельной информации. ---------------------------------------------------------------------------------------------------------*) Ширина этой зоны в мозге Эйнштейна (и ряда других гениев) на 15% (и более) превышает контрольные величины. Найден ряд структур мозга, выполняющих важнейшие перцептивные, когнитивные и интеллектуальные функции, которые характеризуются достоверно высоким уровнем регуляторного потенциала, определяемого их взаимодействием с мРНК-мишенями миРНК RISC Рибосома Allen Human Brain Atlas: эволюционная ветвь H. neanderthalensis Структуры мозга, характеризующиеся увеличенным потенциалом регуляторных взаимодействий миРНК/мРНК, определяемые ненейтрально эволюционировавшими миРНК неандартальца орангутан горилла шимпанзе человек denisovian neanderthal НЕ НАЙДЕНЫ! Филогенетическое дерево шимпанзе, макаки и человека, построенное на основе последовательностей пре-микроРНК МАКАКА Ожидаемая длина человеческой ветви Выявлено резкое увеличение скорости эволюции миРНК в человеческой ветви. Реальная длина этой ветви в шесть раз больше, чем можно ожидать из скоростей эволюции остальных двух видов ШИМПАНЗЕ Реальная длина человеческой ветви [Hu HY et al., MicroRNA expression and regulation in human, chimpanzee, and macaque brains. PLoS Genet. 2011 Oct;7(10):e1002327. 2011 Oct 13] Скорость фиксации однонуклеотидных замен в генах ненейтрально эволюционирующих миРНК более чем на порядок превышает скорость эволюции ортологичных генов миРНК приматов миРНК Homo sapiens miR-1178 miR-1252 miR-1269a miR-1321 miR-146a miR-532 miR-608 miR-609 miR-662 miR-671 miR-943 Предковый нуклеотид реконструирован ный по геномам гоминид Hominida: 37G Hominida:13G Hominida:79A Hominida:55G Hominida:60G Hominida:79G; 82G Hominida:37G 72A Hominida:17G макака: 33U; 67G Hominida:87C Hominida:12G; 17AG; 27G Усредненная скорость фиксации мутаций в год от точки дивергенции вида Замены в линиях Homo sapiens/ Homo denisova Пре-миРНК Homo sapiens Пре-миРНК Homo denisova Пре-миРНК гориллы Пре-миРНК шимпанзе Пре-миРНК орангутана H.s.:37A H.s.:13A H.s.:79G H.d.:55U H.s.:60C H.s.:79A; H.s.:82A H.s.:37C; H.d.:72G H.d.:17A H.d.:33A; H.d.:67A H.d.:87U H.s.:12A; H.s.:17-; H.s.:27A -9 6.8*10-10 1.86*10-9 1.3*10-8 1.19*10 1.9*10-8 30!! 20!! Местоположение гена, кодирующего миРНК-941 на выравнивании геномов высших приматов. Эта миРНК возникла из вставки фрагментов ДНК в геном предка денисовца и современного человека Вторичная структура премиРНК-941 эволюционно консервативна у человека и денисовца. Возникновение гена пре-миРНК-941 датируется 1-5 млн.л. назад Hu et al. Evolution of the human-specific microRNA miR-941. Nat Commun. 2012 Oct 23;3:1145 Somel M и сотр. провели экспериментальное и биоинформационное исследование экспрессии мРНК в мозгах человека и шимпанзе на разных стадиях пре- и постнатального онтогенеза (MicroRNA-driven developmental remodeling in the brain distinguishes humans from other primates. PLoS Biol. 2011 Dec;9(12):e1001214. 2011 Dec 6) RISC Ribosome ЧЕЛОВЕК: экспрессия мРНК наблюдается в первые 3 года после рождения Они нашли большую группу мРНК, регулируемых миРНК, экспрессирующихся в тканях мозга шимпанзе в последний месяц перед рождением. Экспрессия этих мРНК у человека длится три года после рождения. Именно эти три года важны для формирования человеческого мозга, обучения речи и овладения социальным общением. Somel и сотр. делают вывод, что существуют механизмы, направляющие реструктурирование мозга в ходе развития и направляемые микроРНК. Это отличает человека от других приматов. ШИМПАНЗЕ: экспрессия мРНК наблюдается в последний месяц беременности Модель видообразования, основанная на представлениях классической популяционной генетики: последовательная фиксация адаптивных мутаций, каждая из которых приводит к возникновению отдельного адаптивного фенотипического признака. В этом случае формирование адаптивного комплекса фенотипических характеристик, необходимых для адаптации к новой экологической нише, требует исключительно больших промежутков времени 100 P Фиксация адаптивных мутаций Последовательная фиксация адаптивных признаков T 0 Возникновение мутаций Модель видообразования, основанная на фиксации мегамутаций , т.е. мутаций в локусах кассетных регуляторов молекулярно-генетических систем, приводящих к одновременному изменению функции больших групп регулируемых генов и, как следствие, возникновению адаптивных комплексов фенотипических характеристик 100 Фиксация адаптивных мегамутаций 0 Возникнове ние мутаций Кассетная фиксация адаптивных признаков P мРНК КРИЗИС T миРНК Сетевая модель эволюционного возникновения человека современного типа (Stringer C. Evolution: What makes a modern human. Nature. 2012 485(7396):33-5.) Гибридизация между Ноmo sapiens и древними представателями рода Ноmo Неполное замещение аллелей при дивергенции линий древних людей от линии современного человека Reich D et al., Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania. Am J Hum Genet. 2011 89(4):516-28. Hammer MF et al., Genetic evidence for archaic admixture in Africa. Proc Natl Acad Sci U S A. 2011 108(37):15123-8. Eriksson A, Manica A. Effect of ancient population structure on the degree of polymorphism shared between modern human populations and ancient hominins. Proc Natl Acad Sci U S A. 2012 109(35):13956-60. Sánchez-Quinto F et al., North African populations carry the signature of admixture with Neandertals. PLoS One. 2012 7(10):e47765. Благодарю за внимание! Выражаем благодарность Российскому фонду фундаментальных исследований за финансовую поддержку работ. Предковый нуклеотид реконструирован ный по геномам гоминид Замены в линиях Homo sapiens/ Homo denisova Известные полиморфизмы Homo sapiens совпадающие с замененными нуклеотидами в пре-миРНК, с указанием частот в популяции человека согласно данным базы данных dbSNP. miR-1178 miR-1252 miR-1269a miR-146a miR-532 Hominida: 37G Hominida:13G Hominida:79A Hominida:60G Hominida:79G; 82G Hominida:12G; 17AG; 27G Hominida:37G 72A Hominida:55G H.s.:37A H.s.:13A H.s.:79G H.s.:60C H.s.:79A; H.s.:82A H.s.:12A; H.s.:17-; H.s.:27A H.s.:37C; H.d.:72G H.d.:55U 37A(96.1)/G(3.8):rs7311975 13A(99.2)/G(0.8):rs115256251 79G(61.7-74.2)/A(25.8-38.3):rs73239138 60C(23.1)/G(76.8):rs2910164 79A/G:rs456615; 82A/G:rs456617 Hominida:17G макака: 33U; 67G Hominida:87C H.d.:17A H.d.:33A; H.d.:67A H.d.:87U 33U(86.5)/A(13.5): rs74656628; 67G(92.6)/A(7.4):rs9745376 - miR-943 miR-608 miR-1321 miR-609 miR-662 miR-671 человек миРНК Homo sapiens денисовец Характеристики нуклеотидных замен, фиксировавшихся в 11 ненейтрально эволюционирующих миРНК, аннотированных в базе данных starBase 2.1 12A(88.5)/G(11.5): rs1077020; -/AG:rs3034718 37C(82.0)/G(18.0):rs4919510 - общий предок В ЛИНИИ ЧЕЛОВЕКА ИМЕЛО МЕСТО НЕПОЛНОЕ ВЫТЕСНЕНИЕ ПРЕДКОВОГО АЛЛЕЛЯ (Сравнение нуклеотидных замен, фиксировавшихся в ненейтрально эволюционирующих миРНК в линии человека, с полиморфизмами в популяциях человека, описанными в базе данных dbSNP) миРНК Homo sapiens Предковый нуклеотид реконструированн ый по геномам гоминид Замены в линиях Homo sapiens/ Homo denisova Известные полиморфизмы Homo sapiens совпадающие с замененными нуклеотидами в пре-миРНК, с указанием частот в популяции человека согласно данным базы данных dbSNP. miR-1178 miR-1252 miR-146a Hominids: 37G Hominids:13G Hominids:60G H.s.:37A H.s.:13A H.s.:60C 37A(96.1)/G(3.8):rs7311975 13A(99.2)/G(0.8):rs115256251 60C(23.1)/G(76.8):rs2910164 miR-1178 miR-1252 человек денисовец 37A 37G 37G miR-146a человек денисовец 13A 13G 13G человек денисовец 60C 60G 60G общий предок общий предок общий предок В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА 37 A(96.1) G(3.8) 13 A(99.2) G(0.8) 60 С(23.1) G(76.8) В ЛИНИИ ДЕНИСОВЦА ПРОИЗШЛА ФИКСАЦИЯ НОВОГО АЛЛЕЛЬНОГО ВАРИАНТА. ПРИ ЭТОМ ПО ДАННЫМ БАЗЫ dbSNP, В СОВРЕМЕННЫХ ПОПУЛЯЦИЯХ ЧЕЛОВЕКА «ДЕНИСОВСКИЙ АЛЛЕЛЬ» НЕ НАЙДЕН миРНК Homo sapiens Предковый нуклеотид реконструированн ый по геномам гоминид Замены в линиях Homo sapiens/ Homo denisova Известные полиморфизмы Homo sapiens совпадающие с замененными нуклеотидами в пре-миРНК, с указанием частот в популяции человека согласно данным базы данных dbSNP. miR-1321 miR-609 miR-671 Hominids:55G Hominids:17G Hominids:87C H.d.:55U H.d.:17A H.d.:87U - miR-1321 miR-609 человек денисовец 55G 55U 55G miR-671 человек денисовец 17G 17A 17G человек денисовец 87C 87U 87C общий предок общий предок общий предок В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА 55 U(~0) G(~100) 17 A(~0) G(~100) 87 U(~0) С(~100) Доказательство гибридизации человека и денисовца на основе сравнения нуклеотидных замен, фиксировавшихся в близко расположенных позициях миРНК, с полиморфизмами в популяциях человека миРНК Homo sapiens Предковый нуклеотид реконструированн ый по геномам гоминид miR-662 macaque: 33U; 67G Замены в линиях Homo sapiens/ Homo denisova H.d.:33A; H.d.:67A Известные полиморфизмы Homo sapiens совпадающие с замененными нуклеотидами в пре-миРНК, с указанием частот в популяции человека согласно данным базы данных dbSNP. 33U(86.5)/A(13.5):rs74656628; 67G(92.6)/A(7.4):rs9745376 miR-662 человек денисовец 33U;67G 33A;67A 33U;67G общий предок В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА 33 A(13.5) 67 A(7.4) U(86.5) G(92.6) Следы гибридизации человека и денисовца в ненейтрально эволюционирующих миРНК человека Человек miR-662 33U 67G 33A 67A Денисовец 33U 67G 33A 67A 33A 67G 33U 67A 33U 67G 33A 67A Кроссинговер генов пре-миРНК-662 В современных популяциях человека представлены все четыре варианта продуктов кроссинговера генов премиРНК-662, что свидетельствует о гибридизации между денисовцем и человеком Доказательство гибридизации человека и денисовца на основе сравнения нуклеотидных замен, фиксировавшихся в близко расположенных позициях миРНК, с полиморфизмами в популяциях человека миРНК Homo sapiens Предковый нуклеотид реконструированн ый по геномам гоминид miR-608 Hominida: 37G 72A Замены в линиях Homo sapiens/ Homo denisova Известные полиморфизмы Homo sapiens совпадающие с замененными нуклеотидами в пре-миРНК, с указанием частот в популяции человека согласно данным базы данных dbSNP. H.s.:37C; H.d.:72G 37C(82.0)/G(18.0):rs4919510 miR-608 человек денисовец 37C;72A 37G;72G 37G;72A общий предок В ПОПУЛЯЦИЯХ ЧЕЛОВЕКА 37 С(~100) 72 G(18.0) G(~0) A(82.0) Следы гибридизации человека и денисовца в ненейтрально эволюционирующих миРНК человека Человек miR-608 37C 72A 37G 72G Денисовец 37C 72A 37G 72G 37G 72A 37C 72G 37C 72A 37G 72G Кроссинговер генов пре-миРНК-608 В современных популяциях человека представлены два варианта продуктов кроссинговера генов пре-миРНК-608 (из четырех), что свидетельствует о гибридизации между денисовцем и человеком с последующей утратой двух вариантов Анализ генов пре-миРНК, выявил неполное замещение аллелей при дивергенции линий древних людей от линии современного человека, а также гибридизацию между Ноmo sapiens и древними представителями рода Ноmo. Эти факты дают обоснование гипотезе о сильно пересекающихся генофондах современного человека и денисовца (сетевая эволюция) 11 генов пре-миРНК общий предок Случайный выбор мРНК-мишеней: численный эксперимент Мишени миРНКk и их уровни экспрессии Ткань i 3 10 1 5 5 Li(real)=31 1 5 Li(rand)=11 5 9 9 2 миРНК 7 2 миРНК 7 7 3 6 Случайные перестановки 1 3 10 Li(rand)=10 3 10 Случайные перестановки 7 Среднее значение 6 Li(rand)=11.66 1 5 5 3 1 3 10 1 5 5 9 Случайные перестановки Li(rand)=14 9 2 миРНК 7 2 миРНК 7 7 3 6 1 7 3 6 1 Экспрессия миРНК в тканях мишенях - необходимое условие реализации их регуляторного потенциала. Согласно экспериментальным данным, представленным в базе данных miRGator 3.0 (Cho S et. al. Nucleic Acids Res. 2013 41:D252-7) все 11 ненентрально эволюционирующих миРНК экспрессируются в вывяленных нами тканях-мишенях. миРНК miR-1178 miR-1252 miR-1269a miR-1321 miR-146a miR-532 miR-608 miR-609 miR-662 miR-671 miR-943 Орган / ткань с высоким уровнем экспрессии миРНК матка, печень, желудочно-кишечный тракт центральная нервная система, мозг обонятельный эпителий, В-клетки крови, эмбриональные стволовые клетки центральная нервная система, тимус, матка, селезенка, сердце, ооциты лимфоидная ткань, меланоциты центральная нервная система, обонятельный эпителий, почка мозг, селезенка, тимус, матка, молочные железы центральная нервная система, плацента, молочные железы матка, легкие обонятельный эпителий, жировая ткань, лимфоидная ткань, кератиноциты В-клетки крови, матка, кератиноциты их регуляторного потенциала. Согласно экспериментальным данным, представленным в базе данных miRGator 3.0 (Cho S et. al. Nucleic Acids Res. 2013 41:D252-7), в мозге человека экспрессируются 10 из 11 ненентрально эволюционирующих миРНК. Экспрессия миРНК миРНК Орган / ткань в мозге miR-1178 + матка, печень, желудочно-кишечный тракт miR-1252 + центральная нервная система, мозг miR-1269a + обонятельный эпителий, В-клетки крови, эмбриональные стволовые клетки miR-1321 + центральная нервная система, тимус, матка, селезенка, сердце, ооциты miR-146a +/- лимфоидная ткань, меланоциты miR-532 + центральная нервная система, обонятельный эпителий, почка miR-608 + мозг, селезенка, тимус, матка, молочные железы miR-609 + центральная нервная система, плацента, молочные железы miR-662 - матка, легкие miR-671 + обонятельный эпителий, жировая ткань, лимфоидная ткань, кератиноциты miR-943 +/- В-клетки крови, матка, кератиноциты "+" экспрессия пре-миРНК четко отличается от фона, "+/-" экспрессия пре-миРНК слабо отличается от фона; "-" экспрессия пре-миРНК не отличается от фона. Gene Expression Barcode Database: общий пул – сетевая схема эволюции (11 миРНК) Отобранные ткани-мишени с повышенным регуляторным потенциалом мРНК/миРНК взаимодействий опосредованных ненейтрально эволюционирующими миРНК Ткань/орган (Barcode 2.0) Вероятность наблюдения суммы величин экспрессии (p) dorsal root ganglia hippocampus entorhinal cortex superior frontal gyrus cerebellum trigeminal ganglia postcentral gyrus cervix fallopian tube epithelium myometrium ovary vulva endometrial endothelial cell breast lobular cells cecum head and neck epithelial cells lung thyroid 2.00E-05 6.00E-05 8.00E-05 0.00032 0.00054 0.00096 0.00184 4.00E-05 2.00E-05 0.00014 0.00014 0.00044 0.0013 0.0007 6.00E-05 4.00E-05 0.00122 0.0018 В схеме сетевой эволюции было выявлено 18 тканеймишеней общий предок Второй вариант исследования регуляторного потенциала ненейтрально эволюционирующих миРНК: раздельное рассмотрение 7 и 5 генов пре-миРНК, фиксировавших мутации в линиях человека и денисовца, соответственно. общий предок Gene Expression Barcode Database: линия H. sapiens (7 миРНК) Отобранные ткани-мишени с повышенным регуляторным потенциалом мРНК/миРНК взаимодействий опосредованных ненейтрально эволюционирующими миРНК Ткань/орган (Barcode 2.0) Вероятность наблюдения суммы величин экспрессии (p) cerebellum hippocampus dorsal_root_ganglia posterior_cingulate entorhinal_cortex superior_frontal_gyrus primary_visual_cortex myometrium ovary fallopian_tube_epithelium cervix breast_lobular_cells cecum stomach neutrophils head_and_neck_epithelial_cells 4.00E-05 0.00022 0.00038 0.00056 0.00074 0.0008 0.001 8.00E-05 0.00034 0.00046 0.00044 0.00158 0.00136 0.0015 0.00092 0.00014 Gene Expression Barcode Database: линия H. denisova (5 миРНК) Отобранные ткани-мишени с повышенным регуляторным потенциалом мРНК/миРНК взаимодействий опосредованных ненейтрально эволюционирующими миРНК Ткань/орган (Barcode 2.0) Вероятность наблюдения суммы величин экспрессии (p) entorhinal_cortex hippocampus vagina 0.00058 0.0004 6.00E-05 В результате анализа были отобраны ткани-мишени миРНК, удовлетворяющие одному из двух условий: (1)Ткани были выявлены при анализе как общего пула пре-миРНК, так и пула пре-миРНК человека; (2)Ткани были выявлены при анализе как общего пула пре-миРНК, так и пула пре-миРНК денисовца; Такие ткани (структуры)-мишени представляют первостепенный интерес в связи с их возможной ролью в эволюции рода Homo (H.s и H.d). общий пул – 18 тканей-мишеней 6 10 денисовец 3 ткани-мишени 1 2 4 человек 16 тканей-мишеней Всего было отобрано 12 тканей-мишеней, удовлетворявших приведенным критериям