Слайд 1 - Институт экологии Волжского бассейна РАН
реклама

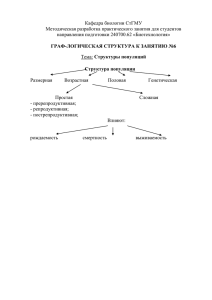
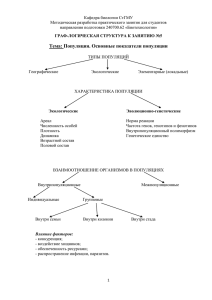
Тефлоновые популяции клональных видов Кутлунина Н.А. 1, Князев М.С. 2, Беляев А.Ю 3 1 Уральский федеральный университет имени первого Президента России Б.Н. Ельцина 2 Ботанический сад УрО РАН 3 Институт экологии растений и животных РАН Многие виды растения демонстрируют замечательную способность к распространению исключительно или преимущественно за счёт вегетативного размножения клубнями, луковицами, бульбами, фрагментами побегов и др., при отсутствии явных приспособлений к переносу на дальние расстояния таких пропагул Описан случай расселения не способного давать семена гибридного микровида опунции на расстояние 42 км (Грант, 1984). Тщательными и масштабными генетическими исследованиями (Pfeiffer et al., 2012) показано, что гусиный лук лопатчатый Gagea spathacea (Hayne)Salisb., распространённый от Дании до Западного Кавказа стерилен, размножается только мелкими дочерними луковичками и, по существу, представляет один клон Нами показано, что гладиолус тонкий Gladiolus tenuis Bieb. восточнее Волги весь стерилен, размножается только дочерними клубне-луковицами и представляет один клон Клон Gagea spathaceae Клон стерильного гладиолуса тонкого Изофореграмма G. tenuis: 2-8, 26-32 – Тормосино; 10-17, 34-41 – Тормосино; 19-24, 43-47 – Морское; 1, 9,18, 25, 33,42 - Петровское Изофореграмма G. tenuis: 1-13 – Дубиновка, 14 – Петровское; 15-27 – Рысаево; 28 – Петровское; 29-48 - Губерля ISSR-маркирование, праймеры М3 и М12 Последовательность образцов: 1- Ledder, 2 – 6 – «Рысаево»; 7 – 12 – «Губерля» Клон Gagea spathaceae Клон стерильного гладиолуса тонкого Столь значительные ареалы, освоенные клонами косвенно свидетельствуют о длительности их существования. По всей видимости, возраст клонов соизмерим с возрастом «обычного», размножающегося семенами амфимиктического вида, имеющего сопоставимый ареал. Можно, даже, предполагать, что время, необходимое для освоения такого ареала заметно больше, чем потребовалось бы для вида, распространяющегося семенами Таким образом, клоны демонстрируют поразительную устойчивость во времени и в экологическом градиенте Столь значительные ареалы, освоенные клонами косвенно свидетельствуют о длительности их существования. По всей видимости, возраст клонов соизмерим с возрастом «обычного», размножающегося семенами амфимиктического вида, имеющего соизмеримый ареал. Можно, даже, предполагать, что время, необходимое для освоения такого ареала заметно больше, чем потребовалось бы для вида, распространяющегося семенами Клоны демонстрируют поразительную устойчивость во времени и в экологическом градиенте Очевидно, что рассматриваемые клоны пережили различные контрастные варианты палеоклимата (например, на протяжении всего голоцена или, даже, части позднего плейстоцена). По нашим наблюдениям, клон гладиолуса тонкого восточнее Волги освоил достаточно разнообразные местообитания – примерно те же, что свойственны его типичному варианту с нормальным семенным размножением. Таким образом, потенциал экологической адаптации (в данном случае) реализуется на уровне одного генотипа, а не на уровне полиморфной популяции. Cardamine trifida (Lam. ex. Poiret) B.M. Jones короткокорневищноклубнеобразующий травянистый многолетник, весенний эфемероид. Еще один пример Для C. trifida характерно преимущественно вегетативное размножение – подземными клубеньками листового происхождения (Беркутенко, 1999). Почти все исследованные популяции были умеренно полиморфны - в них отмечалось по 4-12 генотипов, однако в долине р. Белая все образцы выборок принадлежали единственному стерильному клону C. trifida на Урале Долина р.Чусовой 1 – Левинская 2 – Георгиевская Долина р. Сим 3 – Берда 4 – Сим-Куряк 5 – Гребни Долина р. Белой 6 – Дуги 7 – Ашкарка 8 – Шулемовка 9 – Маячная 10 – Тимирово 11 – Сатра Белоречинский клон на протяжении до 250 км реки По результатам популяционно-генетических исследований сердечника трёхраздельного было опубликовано две большие статьи. Князев М.С., Кутлунина Н.А., Зимницкая С.А., Беляев А.Ю. Распространение и изменчивость Cardamine trifida (Brassicaceae) на Урале// Ботанический журнал. 2010. Т.95, №.12. С. 1718-1726. Кутлунина Н.А., Князев М.С., Зимницкая С.А., Беляев А.Ю. Генотипическое разнообразие и мужская стерильность Cardamine trifida (Brassicaceae) в разных частях ареала// //Turczeninovia, 2012, 15(2). С. 72-79. Однако, при обсуждении результатов был упущен один очевидный, но не тривиальный феномен, на который мы обратили внимание только при работе над статьёй по ещё одному клональному виду – Tulipa riparia C. trifida на Урале Как показывает распространение «белоречинского клона» C. trifida способен превосходно распространятся вдоль долины рек. На так ли? Тогда в других популяциях мы, вполне постоянно, должны отмечать клоны общие для многих или хотя бы еще одной популяции в долине той же реки (или, даже, в соседних речных долинах) Но мы этого не отметили – набор генотипов был уникален даже в соседних популяциях. ? Белоречинский клон на протяжении до 250 км реки Приведём результаты популяционно-генетических исследований ещё по одному клональному виду Tulipa riparia Это эндемик Южного Урала, облигатно триплоидный, а поэтому, в высокой степени, (но не на 100%) стерильный вид Генетический полиморфизм в популяциях (в том числе и изменчивость окраски цветков) свидетельствует, что изредка семенное размножение происходит, но при устойчивом сохранении триплоидности (возможно, в результате избирательных леталей гамет?) Популяционно-генетическая структура Tulipa riparia исследована в 4 популяциях долины р. Сим В 3 популяциях долины р. Юрюзань и в одной популяции долины р. Нугуш Пример исследования в популяции «Куряк» на небольшом левом притоке р. Сим – По серии трансект было взято более 60 проб. Обнаружено 6 генотипов (клонов), которые сложным образом чередовались, сменяя друг друга в пространстве популяции Несмотря на очевидные признаки способности тюльпана к миграциям вдоль речных долин – смотри, например, распространение генотипа «6» Генотипический состав изученных популяций был уникален, за исключением соседней пары популяций Илек-Янгантау, где один из клонов зарегистрирован повторно В 3 популяциях долины р. Юрюзань и в одной популяции долины р. Нугуш Сходные результаты получены раннее другими исследовательскими группами для Bistorta vivipara (Lundqvist, Andersson, 2001), Convallaria majalis (Vandepitte et al., 2010), Poa bulbosa L. (Novak, Welfley, 1997), где набор генотипов не повторялся даже у соседних популяций (однако, попыток объяснения этим особенностям перечисленные авторы не дают). Каков возможный механизм, препятствующий проникновению новых клонов на занятую территорию? Может быть сформулировано следующее правило: клоны способны распространятся, осваивая незанятые участки, однако их внедрение в популяции освоенные другими клонами затруднено Простейшее объяснение – на освоенной территории все приемлемые участки заняты (для видов речных долин, важное дополнение – заняты участки вне воздействия весенних половодьев – клон не просто должен адаптироваться к новому местообитанию, но распространится на не размываемый «безопасный участок» Может быть сформулировано следующее правило: клоны способны распространятся, осваивая незанятые участки, однако их внедрение в популяции освоенные другими клонами затруднено Возможно и более сложное объяснение как результат косвенной конкуренции (Charpentier et al, 2000; Honnay, Bossuyt, 2005; Honnay et al., 2006). Очевидно, что местные клоны вполне адаптированы к комплексу сопутствующих видов, например местному составу видов и штаммов фитопатогенных организмов, тогда как адвентивный клон адаптирован к несколько иному составу «спутников», а потому будет проигрывать конкуренцию с аборигенными генотипами уже на стадии начального освоения новой территории. Может быть сформулировано следующее правило: клоны способны распространятся, осваивая незанятые участки, однако их внедрение в популяции освоенные другими клонами затруднено Спасибо за внимание! Авторы благодарят РФФИ за финансовую поддержку (гранты №№ 10-04-00989, 10-04-96012урал, 13-04-01458 ).