нейромодуляторы
реклама

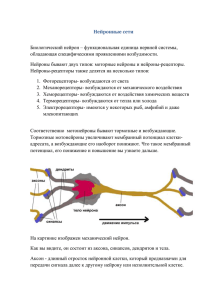
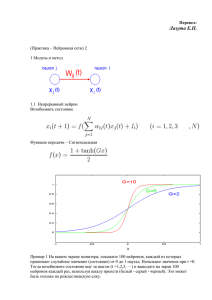
Нейробиология Нервная ткань Нейробиологический подход к исследованию нервной системы человека Диффузная нервная система примитивного кишечнополостного животного – гидры. Нервная система плоского червя - планарии Нервная система человека Нейроны, их отростки, процессы, протекающие в нейронах, остаются практически неизменными в ходе эволюции как у примитивных животных (например, членистоногих, рыб, амфибий, рептилий и др.), так и у человека. Открытие клеточного строения нервной системы Ками́лло Го́льджи (Camillo Golgi) (1843 —1926) Итальянский врач и учёный, лауреат Нобелевской премии по физиологии и медицине в 1906 году «В знак признания трудов о структуре нервной системы» Сантья́го Рамо́н-и-Каха́ль (Santiago Ramón y Cajal) (1852 — 1934) Испанский врач и гистолог, лауреат Нобелевской премии по физиологии медицине в 1906 году «В знак признания трудов о структуре нервной системы» Рисунки Рамон-и-Кахаля. Слева — гистологическое строение двигательной зоны коры головного мозга человека. Справа — клетка Беца из двигательной зоны коры; импрегнация по методу Гольджи. (Cajal, 1911). Схемы Кахала, указывающие направление переноса сигналов в нервных клетках и в нервных сетях в соответствии с «законом динамической поляризации». А. Ганглий беспозвоночного. Б. Мозжечок. (Cajal, 1911) ♦ В ядре находятся хромосомы, содержащие ДНК – запись наследственной генетической информации. Эндоплазматическая сеть (эндоплазматический ретикулум) служит для синтеза, накопления и транспорта различных веществ. Глия Основные типы клеток глии: 1. Микроглия – участвует в образовании мозговых оболочек, выполняет фагоцитарную роль (уничтожение инородных тел и погибших клеток) 2. Олигодендроглия (олигодендроциты) – участвует в образовании миелиновых оболочек 3. Астроглия (астроциты) – обеспечивают нейронам механическую защиту, доставляют питательные вещества, удаляют шлаки, электрически изолируют нейроны друг от друга Миелиновые оболочки периферических нервных волокон образованы Шванновскими клетками Разные типы нейроглии. А. Протоплазматические астроциты. Б. Фиброзные астроциты. В. Микроглия. Г. Олигодендроциты. Олигодендроцит наматывает свою мембрану вокруг аксона, образуя многослойную миелиновую оболочку. В миелинизированном аксоне переход ионов через мембрану происходит только в разрывах между сегментами миелиновой оболочки - перехватах Ранвье. Миелиновая оболочка (My), образованная Шванновской клеткой (Schw.), вокруг аксона нейрона (A) Астроциты и олигодендроциты в белом веществе Нейрон Нейрон на препарате, окрашенном по Гольджи Фотография нейрона, полученная с помощью сканирующего электронного микроскопа, и рисунок типичного нейрона. Схема нейрона. 1 — дендрит; 2 — шипики; 3 — тело нейрона; 4 — дендрит; 5 — аксонный холмик; 6 — аксон; 7 — синаптическая щель; 8 — окончание аксона; 9 — постсинаптический нейрон. ♦ Внутреннее строение типичного нейрона. Микротрубочки обеспечивают структурную жесткость, а также транспортировку материалов, синтезируемых в теле клетки и предназначенных для использования в окончании аксона (внизу). В этом окончании находятся синаптические пузырьки, содержащие медиатор, а также пузырьки, выполняющие иные функции. На поверхности постсинаптического дендрита показаны предполагаемые места рецепторов для медиатора Примеры различных нейронов. Все имеют дендриты, тело и аксон Основные типы нейронов млекопитающих: Псевдоуниполярные клетки. Клетки, от тела которых отходит только один отросток. На самом деле при выходе из сомы этот отросток разделяется на два: аксон и дендрит. Расположены в сенсорных узлах. Биполярные клетки — это клетки, которые имеют один аксон и один дендрит. Они характерны периферической части некоторых сенсорных систем. Мультиполярные клетки имеют один аксон и множество дендритов. К такому типу нейронов принадлежит большинство нейронов ЦНС. Исходя из особенностей формы этих клеток их делят на веретенообразные, корзинчатые, звездчатые, пирамидные и др. Только в коре головного мозга насчитывается до 60 вариантов форм тел нейронов. Различные типы нейронов: псевдоуниполярные, биполярные, безаксонные и мультиполярные Мембранные потенциалы. Потенциал покоя. Мембрана нейрона и ассоциированные с ней трансмембранные белки Нервная клетка и ионы. А- — органические анионы. Концентрации ионов в цитоплазме нейрона (слева) и в межклеточной жидкости (справа) Принцип работы натрий-калиевого насоса Ионный канал Диффузия и полупроницаемые мембраны Потенциал покоя создается преимущественно калиевым током Равновесный потенциал для ионов калия на мембране нейрона ♦ Потенциал покоя создается преимущественно выходящим калиевым током. Разность концентраций ионов поддерживается калий-натриевым насосом Возбуждение нейрона. Проведение возбуждения. Внутриклеточная регистрация мембранного потенциала покоя и потенциала действия в гигантском аксоне кальмара. А- стеклянная микропипетка диаметром 100 мкм заполнена морской водой и введена в рассеченный аксон кальмара (толщина аксона около 1 мм). B. Потенциал действия, регистрируемый микропипеткой. Внизу – отметки времени 2 мс. По Hodgkin and Huxley (1939). А.Л.Ходжкин и А.Хаксли удостоены Нобелевской премии по физиологии и медицине, 1963, за создание математической модели генерации и распространения потенциала действия. Потенциал действия развивается по закону «все или ничего» и достигает приблизительно +40 - +55 мВ (в цитоплазме относительно внеклеточной среды), т.е. амплитуда составляет порядка 100 - 125 мВ Строение периферического нерва Синапс Синаптические контакты на нейроне гиппокампа (микрофотография с применением иммунохимических красителей) Строение синапса Электронная микрофотография синапса Принцип работы простейшего ионотропного синаптического рецептора. Такой рецептор состоит из двух частей – узнающего центра и ионного канала. Принцип работы синаптического рецептора. Четыре рисунка слева направо показывают последовательность событий при активации синапса. Крайний справа рисунок демонстрирует блокирование рецептора искусственно введенным фармакологическим агентом – антагонистом данного типа рецепторов. Деполяризующие синапсы необходимы для генерации нервных импульсов, и поэтому потенциалы, возникающие в таких синапсах, были названы возбуждающими постсинаптическими потенциалами (ВПСП). Деполяризующий ток преимущественно связан со входом ионов Na+. Ток может быть также связан с открытием каналов либо для выхода катионов К+, либо для входа анионов Cl-. Эти ионные потоки приводят к удержанию мембранного потенциала на уровне покоя или к некоторой гиперполяризаци мембраны. Поскольку эти потенциалы препятствуют деполяризации мембраны и, следовательно, генерации нервных импульсов, их называют тормозными постсинаптическими потенциалами (ТПСП). Синаптические потенциалы представляют собой градуальные реакции. Структура н-холинорецептора (вертикальный срез) и его трехмерная реконструкция На каждом нейроне заканчиваются тысячи или десятки тысяч синапсов, как возбудительных, так и тормозных. Нейрон интегрирует это огромное количество входных воздействий и посылает единственный поток выходных сигналов по своему аксону. Схема мотонейрона спинного мозга кошки. Видны взаимоотношения между возбуждающими (изображены светлым) и тормозными (заштрихованы) синапсами, а также между этими синапсами и участком генерации потенциала действия. Ионотропный рецептор (рецептор и канал составляют единое целое) Метаботропный рецептор (активация рецептора ведет к синтезу вторичного посредника внутри клетки, который, в свою очередь, открывает ионные каналы и запускает другие процессы в клетке) Сравнение ионотропного рецептора (слева), состоящего из 5-7 субъединиц, образующих ионный канал, и метаботропного рецептора, состоящего из одной субъединицы, но требующего сопряжения с G-белком Ионотропные рецепторы обеспечивают очень быструю реакцию нейрона (порядка нескольких миллисекунд), однако способны непосредственно влиять лишь на потенциал мембраны нейрона, поэтому участвуют в быстрой передаче сигналов. Метаботропные рецепторы действуют значительно медленнее (долее 100 мс), однако: - Реакция постсинаптического нейрона более длительная, так как вторичный посредник не сразу разрушается и продолжает действовать в цитоплазме еще некоторое время - Синтез вторичных посредников обеспечивает усиление сигнала (на одну молекулу медиатора синтезируются сотни молекул вторичного посредника) - Вторичные посредники способны влиять одновременно на множество процессов во всем нейроне, в том числе в его ядре. Соответственно, метаботропные рецепторы обеспечивают нейромодуляцию (настройку «режимов работы» нейронов). Как правило, к одному медиатору может существовать множество различных типы рецепторов, как ионотропных, так и метаботропных. Воздействие медиатора на нейрон определяется не только свойствами самого медиатора, но тем, какими рецепторами к этому медиатору обладает данный нейрон. В частности, многие медиаторы могут оказывать как тормозное, так и возбудительное действие на различные нейроны, принимать участие в передаче сигналов или осуществлять нейромодуляцию. Агонист – вещество, активирующее рецептор к данному медиатору Антагонист – вещество, блокирующее рецептор к данному медиатору или препятствующее действию медиатора Например, яд кураре – антагонист н-холинорецепторов к ацетилхолину в мышцах, блокирует нервно-мышечную передачу и вызывает временный паралич Вторичные посредники (вторичные мессенджеры) передают сигнал внутри нейрона от одной его части к другой: - цАМФ (циклический аденозинмонофосфат) - цГМФ (циклический гуанозинмонофосфат) - кальций (Ca2+) - окись азота (NO) – [может выступать как ретроградный мессенджер, т.е. передавать сигнал через синапс в обратном направлении] Примечание: кальций может входить в нейрон через ионотропный канал (например, НМДА-рецептор к глутамату), но действовать внутри клетки как вторичный посредник. Медиаторы нервной системы Химическая сигнализация в биологических системах Феромоны: сигнализация от одного организма к другим организмам (например, половые феромоны насекомых, запах которых привлекает самцов) Гормоны: сигнализация от одного органа к другим органам (например, адреналин вырабатывается надпочечниками и влияет на мозг, мышцы, сердце и другие органы) Нейромедиаторы (нейротрансмиттеры) и нейромодуляторы: сигнализация от одного нейрона к другим нейронам (глутамат, ацетилхолин, ГАМК и др.) Вторичные посредники (вторичные мессенджеры): сигнализация внутри одного нейрона Ранее считалось, что каждый нейрон синтезирует и выделяет только какой-либо один медиатор (принцип Дейла). По современным данным, как правило, в синаптических пузырьках нейрона содержится смесь нескольких медиаторов – одного «классического» и еще одного или нескольких нейропептидов. Поэтому названия ацетилхолинергический нейрон, норадренергический нейрон и т.п. следует понимать с учетом того, что вместе с «классическим» медиатором эти нейроны выделяют и нейропептиды (например, холинергические нейроны основания переднего мозга вместе с ацетилхолином выделяют нейропептид галанин). Медиаторы (нейротрансмиттеры) передают сигнал от одного нейрона к другому: - Глутамат – основной возбудительный медиатор в ЦНС - ГАМК (гамма-аминомасляная кислота) – основной тормозный медиатор в ЦНС - Глицин - тормозный медиатор в спинном мозге - Ацетилхолин – возбудительный медиатор (в т.ч. в нервномышечной передаче) и нейромодулятор - Норадреналин - нейромодулятор - Дофамин - нейромодулятор - Серотонин - нейромодулятор - Гистамин - нейромодулятор - Различные пептиды (десятки разных веществ!) нейромодуляторы Структура молекул некоторых медиаторных соединений Ацетилхолинергическая система Норадренергическая система Дофаминергическая система Серотонинергическая система Нервная сеть Нейрон Нервная сеть. Крупный нейрон с множеством дендритов получает информацию через синаптический контакт с другим нейроном (в левом верхнем углу). С помощью миелинизированного аксона образуется синаптический контакт с третьим нейроном (внизу). Поверхности нейронов изображены без клеток глии, которые окружают отросток, направленный к капилляру (справа вверху).