БИОфизика ВОДНЫХ ЭКОСИСТЕМ8 (2.6Mб, ppt)
реклама
")
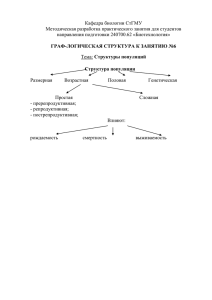
Дискретная модель популяции ветвистоусых ракообразных Блок-схема модели Популяция разбивается на 4 группы: неполовозрелые самки, партеногенетические самки, гаметогенетические самки самцы. Каждая группа описывается численностью рачков (Nx), входящих в нее. Модель разработана для лабораторной популяции рачков температура - 26oC Слив фотопериод - 16 свет: 8 темнота объем экспериментальной камеры – 400 мл скорость протока - 1200 мл в сутки Подача корма Каждый i-ый рачок, в момент времени j, описывается: весом wi,j возрастом ti,j принадлежностью к определенной группе gi,j способом размножения (для самок) набором дискретных переменных Ключевые события в жизни i-го животного: Наступление половозрелости Pi,jmaturity Переход в другой репродуктивный класс Pi,jchange, Pi,jGP Выбор способа размножения (партеногенетичекий Pi,jP или гаметогенетический Pi,jG) Отрождение потомства Pi,jbirth Смертность Pi,jdeath Наступление половозрелости Происходит при достижении рачком размера 0.7 мм. Дискретная переменная «описывающая» этот процесс принимает значение 1 (процесс происходит), когда размер рачка становится больше 0.7 мм. . Ti ti , j , maturity Pi , X Wi , j 1 W0.7 Wi , j W0.7 1, X Ti , g i , X 1 Выбор способа размножения Зависимость смены способа размножения от концентрации пищи и концентрации продуктов жизнедеятельности в среде – экспериментальные данные Gamogenesis efficiency, % 100 80 60 40 20 Food concentration x 1000 cells/ml 0 100 200 300 400 500 y = 72 + 0.13*Метаболиты - 0.7*Пища + 0.0015*Метаболиты * Пища Выбор способа размножения PiG , X 1, PS X PR , Pimaturity 1, g i , X 1 ,X PiP, X 1, PS X PR , Pimaturity 1, g i , X 1 ,X Сравнение функции, описывающей зависимость доли самок сменивших способ размножения от концентрации пищи и метаболитов в среде, со случайным числом в диапазоне от 0 до 1. Для всех самок у которых наступила половозрелость. Наступление смерти рачка t max t min td X j real ( S1 X j real ) Продолжительность жизни Максимальная длительность жизни в благоприятных условиях 13 дней. Смертность в первые 2 суток практически не наблюдается. Предполагаем, что максимальная длительность жизни зависит от концентрации пищи следующим образом: 13 12 11 10 9 8 7 6 5 4 3 2 1 0 0 2 4 Пищевая обеспеченность 6 Зависимость доли выживших рачков от их возраста. % 100 tmin tmax возраст рачка Функция описывающая вероятность смерти рачка Вероятность умереть 1 0 2 Pideath 1, ,X tmax FiD , X PR , g i , X 2,3,4 ось времени Отрождение кладки Доля самок Di в отрождаемом i-ой самкой потомстве. 1 0 Di 1, Di 1 ti , j Доля в потомстве самок и самцов: • В начале жизни в кладках Возраст только самки рачка tmax • К концу жизни ti , j вероятность самцовых / t max , ti , j кладок вырастает. Wi , j W1 Ei , W0.34 Весь вес набранный самкой после достижения размера 1 gi, j 3 мм идет на отрождение потомства Отрождение кладки Отрождение кладки связано с линькой рачка. Считается, что период линьки (Dex) величина устойчивая к изменению условий среды обитания и для M.macrocopa принимает значение 1-1.5 суток (в модели 28 часов). Pibirth , X 1, ti , j / DEX N , g i , X 2,3 После каждой линьки, все отрожденное потомство, в соответствии со значениями Di и Ei, переходит в группы самцов и/или неполовозрелых самок. Рост Wi , j 1 Wi , j dWi , j Pi ,birth Ei W0.34 Динамика веса рачка j 1. Экспериментальные наблюдения показывают, что моина от размера 0.34 мм (размер отражаемых партеногенетической самкой рачков) до размера 1 мм (размер после достижения которого самка начинает отрождать молодь) при благоприятных условиях вырастает за 2 суток (48 часов): , 48 real 0,01096 X real / * S X , j 2 j sh dWi , j Wi , j W1 Рост 2. Вес взрослого рачка увеличивается согласно Мальтузианскому закону, где скорость роста определяется по формуле Моно. dWi Wi dt max S KS S Wi Wi j j 1 * exp shag * dW Wi j 1 * exp shag * 1 , Прирост биомассы за один шаг времени real W X real T / * S X , j 2 j sh dWi , j sh i , j , W e 1 i, j Wi , j W1 Wi , j W1 0,0003 0,00025 0,0002 0,00015 , 0,0001 0,00005 0 0 0,01 0,02 0,03 0,04 0,05 Концентрация пищи Взрослая самка Ювенильная самка Взрослая самка 0,06 Жизненный цикл рачка Параметры были получены в экспериментах с одиночными животными или взяты из литературы √ √ √ √ NP j N Jj i 1 i 1 change N Jj 1 N Jj Di Ei Pibirth P i, j ,j N Jj NG j NP j i 1 i 1 i 1 N Jj NG j NG j i 1 i 1 i 1 GP death N Pj 1 N Pj PiP, j Pichange P P i, j i, j ,j GP death N Gj 1 N Gj PiG, j Pichange P P i, j i, j ,j N Pj NM j i 1 i 1 M birth death NM N ( 1 D ) E P P i, j j 1 j i i i, j Калибровка модели (a) 1000 500 800 400 600 300 400 200 200 100 (b) Population, animals Population, animals 600 0 0 0 3 6 9 0 12 2 4 8 10 12 14 Time, days Time, days (d) Population, animals 800 600 400 200 0 Population, animals 2000 1000 1500 1000 500 0 0 1 2 3 4 Time, days 5 6 (e) 2500 (c) 1200 Population, animals 6 7 2000 1500 1000 500 0 0 1 2 3 4 Time, days 5 6 7 0 1 2 3 4 5 6 Time, days Динамика численности популяции M.macrocopa при разных уровнях пищевой обеспеченности: экспериментальные и модельные данные 7 Динамика численности возрастных и половых групп в популяции M.macrocopa Эксперимент Модель 1 c 800 c c 2 4500 b The number of eggs b c d d 3 12000 b 10000 c d d 1 2 600 3000 a 8000 a a 400 6000 1500 4000 200 2000 0 0 0 0.1 0.5 PC 1 2 0 0 0.1 0.5 PC 1 2 0 0.1 0.5 PC Влияние коэффициента характеризующего накопление продуктов жизнедеятельности в среде на количество образованных диапаузирующих яиц. Вывод • Действие продуктов жизнедеятельности направлено на формирование максимально возможного количества покоящихся яиц – целевая функция. Верификация модели Независимый эксперимент: Условия культивирования получены в модельных экспериментах и затем проведен лабораторный эксперимент: • Разные объемы культивирования – 800 и 200 мл. • Одинаковое общее количество корма • Модель показала разницу в начале смены способа размножения в 1.5-2 суток. Culture volume - 200 ml 200 150 100 50 0 0 1 2 3 4 5 6 Time, days Culture volume - 800 ml 160 Population, animals Population, animals 250 120 80 40 0 0 1 Gamogenetic females 2 3 Time, days Males 4 5 6 Total Вывод • Продукты жизнедеятельности накапливаются в среде в процессе развития популяции • Действие продуктов жизнедеятельности носит концентрационный характер. Анализ модели на чувствительность Типичный результат моделирования Численность популяции Покоящиеся яйца Биомасса и концентрация пищи Параметры жизненного цикла проранжированные по чувствительности биомассы к их изменению 2,56988 dT Time interval required for animal to reach body length of 1mm with optimal food requirement 2,032205 Y Efficiency of food consumption 1,970899 Dmax Coefficient determining the maximal age of animals of opposite sex 0,615071 td Difference between maximal and minimal animal age 0,522666 max 0,22459 S1 Half-saturation total food quantity for td 0,217217 tmin Minimal age when an animal can die 0,202289 Ks Half-saturation food concentration for max 0,179115 S2 Half-saturation food quantity for dT 0,103815 0,079248 Dex Maximum specific growth rate of a mature animal Age when females start to hatch males Moulting period Параметры жизненного цикла проранжированные по чувствительности количества покоящихся яиц к их изменению 3,603146 dT Time interval required for animal to reach body length of 1mm with optimal food requirement 1,951064 Y Efficiency of food consumption 1,790306 Dmax 1,204909 Age when females start to hatch males 1,047398 td Difference between maximal and minimal animal age 0,924898 S2 Half saturation total food quantity for dT 0,552588 S1 Half-saturation total food quantity for td 0,457869 max Maximum specific growth rate of an animal 0,327513 Dex Moulting period 0,283851 tmin Minimal age when an animal can die 0,19618 Ks Half-saturation food concentration for max Coefficient determining the maximal age of animals of opposite sex Структура и биомасса частично контролируются различными параметрами….. Покоящиеся Биомасса яйца dT dT Y Y Dmax Dmax td max td S1 S2 tmin S1 Ks max S2 Dex tmin Dex Ks Чувствительность популяционных характеристик к варьированию параметров жизненного цикла Ранг характеристики Total relative variability of the characteristic, for all parameters Variability of characteristic (%) / Variability of i parameter (%) Популяционная характеристика 4 1 2 11,05 16,48 14,64 Njuv Nmale Juvenile females Males Npart 5 7 6 9,99 8,54 Ngam Parthenog Gametoge netic enetic females females 8,72 3 12,34 Ntot B Neggs Total Biomass Resting eggs Структура популяции более вариабельна чем общая численность популяции. Одна и та же биомасса может быть образована популяцией с различной структурой, что приведет к разному количеству образованных покоящихся яиц и повлияет на исход конкуренции. Анализ сосуществования видов с разными стратегиями размножения. Один цикл Два цикла A B 1 – вид без диапаузы 2а – могут оба, но мало яиц для реактивации 2б – могут оба, достаточно яиц для реактивации 3 – вид с диапаузой Maximum specific growth rate of species without diapause, h-1 Исход конкуренции двух видов зоопланктона с разными репродуктивными стратегиями (один вид с диапаузой, другой без) в пространстве максимальных скоростей роста этих видов 0,04 0,028 1 2a 2b 0,016 3 0,004 0,004 0,016 0,028 0,04 Maximum specific growth rate of species with diapause, h-1 Влияние начальной численности и пищевой обеспеченности на исход конкуренции 0,60 20 100 0,64 0,45 0,40 0,29 0,30 0,20 0,10 0,170,15 0,09 0,20 0,10 Food low medium 0,17 0,090,09 0,00 high 0,58 0,60 0,56 0,50 0,70 Specific area, % Specific area, % 0,70 Initial numbers of 5 population with diapause 0,50 0,46 0,46 0,40 0,30 0,20 0,10 0,27 0,21 0,17 0,28 0,17 0,18 0,090,09 0,05 0,00 1 2a 2b 3 The outcome of scecies coexistence, type 1 2a 2b 3 The outcome of species coexistence, type Вариация начальной численности или пищевой обеспеченности слабо влияет на площадь областей полного доминирования, но существенно на площади областей сосуществования. Вывод Структура популяции и ее интегральные характеристики (численность или биомасса) могут контролироваться разными параметрами жизненного цикла Структура популяции более чувствительна к изменениям жизненного цикла чем ее биомасса. Прямая конкуренция и конкуренция видов размножающихся циклически будет частично контролироваться разными параметрами жизненного цикла. Полезная информация для будущих моделей и исследований Так как получена информация о параметрах жизненного цикла наиболее важных с точки зрения регуляции популяционной динамики и структуры, то можно: сфокусироваться на исследовании этих параметров обращать меньше внимания на «не важные» параметры (даже если они считаются важными экспериментаторами)