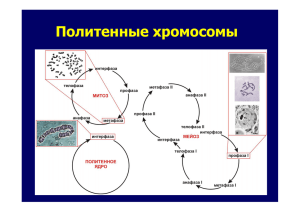
Диссертация - Институт цитологии и генетики СО РАН
реклама

РОССИЙСКАЯ АКАДЕМИЯ НАУК СИБИРСКОЕ ОТДЕЛЕНИЕ ИНСТИТУТ ЦИТОЛОГИИ И ГЕНЕТИКИ На правах рукописи УДК 575.1:575.17:575.224:595.773.4 ЗАХАРОВ ИЛЬЯ КУЗЬМИЧ МУТАЦИИ И МУТАЦИОННЫЙ ПРОЦЕСС В ПРИРОДНЫХ ПОПУЛЯЦИЯХ DROSOPHILA MELANOGASTER 03.00.15 - генетика ДИССЕРТАЦИЯ в виде научного доклада на соискание ученой степени доктора биологических наук НОВОСИБИРСК 1995 Работа выполнена в Институте цитологии и генетики Сибирского отделения РАН, г. Новосибирск Официальные оппоненты: доктор биологических наук, профессор Васильева Л.А., Институт цитологии и генетики СО РАН, г. Новосибирск доктор биологических наук, профессор Высоцкая Л.В., Новосибирский Государственный университет, г. Новосибирск доктор биологических наук, профессор Стегний В.Н., Институт биологии и биофизики Томского Государственного университета, г. Томск Ведущее учреждение - Санкт-Петербургский Государственный университет, г. Санкт-Петербург Защита состоится " __ " ____________ 1995 г. на ________ заседании диссертационного совета по защите диссертаций на соискание ученой степени доктора наук (Д-002.11.01) в Институте цитологии и генетики СО РАН в конференц-зале Института по адресу: 630090, г. Новосибирск, проспект академика Лаврентьева, 10. С диссертацией в виде научного доклада можно ознакомиться в библиотеке Института цитологии и генетики СО РАН. Диссертация в виде научного доклада разослана "___"____________ 1995 г. Ученый секретарь диссертационного совета, доктор биологических наук Груздев А.Д. ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ Актуальность проблемы. В основе эволюционного процесса лежит возникновение мутаций и их дальнейшая судьба, их роль в жизни популяции, вида. Оценка скорости мутационного процесса в природных популяциях представляет собой сложную и многофакторную задачу. Уровень существующей генетической изменчивости (концентрация) и частота возникновения de novo (мутабильность) могут быть определены при изучении разного типа мутаций: видимых, летальных, электрофоретических вариантов. Существующие различия по темпам мутирования между генами, между скоростями мутирования одних и тех же генов на разных генетических фонах и в зависимости от факторов среды еще более усложняет оценку потока генов, поставляемого мутационным процессом. Особый интерес для понимания закономерностей спонтанного мутационного процесса представляют случаи повышенной мутабильности, изучение которой велось разными авторами на дрозофиле в двух направлениях: 1) генетический анализ высокомутабильных локусов - как случаи специфической внутригенной мутабильности, так и взаимодействия в системе генотипа, где повышенная мутабильность в локусе определяется свойствами его самого, генами-мутаторами или внехромосомными агентами; 2) изучение случаев резкого повышения общей мутабильности в лабораторных линиях и искусственных популяциях, отводках из природных популяций и непосредственно в природных популяциях. Постоянные, длительные во времени, с широким географическим охватом наблюдения за генофондом природных популяций Drosophila melanogaster позволяют проследить глобальные и локальные изменения генофондов и выявить факторы, вызывающие эти изменения. Проводимые отечественными генетиками исследования, дали новое развитие идеи о существовании мутационных периодов и колебаниях мутабильности во времени [Berg, 1966, 1982; Голубовский и др., 1974]. Свое дальнейшее развитие эти исследования получили после открытия и возросшего интереса к мобильным генетическим элементам. Высокомутабильные аллели, выделенные из природы [Голубовский, 1977; Голубовский, Захаров, 1983; Захаров, 1984], были отнесены к инсерционным мутациям [Golubovsky et al., 1977; Голубовский, Беляева, 1985]. Исследования по генетической нестабильности позволили установить несомненную связь этого явления с транспозициями мобильных генетических элементов. Однако, как правило, в подавляющем большинстве исследований изучались искусственно ин1 дуцированные мутации и проводился детальный анализ на молекулярном уровне отдельных нестабильных локусов. Описано лишь несколько отдельных случаев исследований мобильных генетических элементов в природных нестабильных хромосомах. Значительная часть возникающих спонтанных мутаций, по-видимому, являются инсерционными. Следует подчеркнуть, что активация мобильных элементов индуцирует целый спектр генетических событий на генном, хромосомном, организменном и популяционном уровнях: изменение характера экспрессии и регуляции генов, нестабильность генов, увеличение мутабильности, мультигенные повреждения, хромосомные перестройки [McClintock, 1956, 1978; Green, 1977, 1986; Bregliano, Kidwell, 1983; Engels, 1983, 1989; Герасимова и др., 1984; Golubovsky, Kaidanov, 1994]. В последнее время получены новые данные о роли мобильных элементов в горизонтальном переносе генов [Kidwell, 1993; Clark et al., 1994]. Исследования популяций Drosophila melanogaster показали, что существуют глобальные изменения во времени в составе некоторых мобильных элементов [Anxolabehere et al., 1985, 1989; Kidwell, 1992]. Вспышки мутаций, генетическая нестабильность, активация мобильных элементов могут играть важную роль в эволюционном процессе. Особенности нашего подхода в рассмотрении явления нестабильности заключаются, во-первых, в изучении нестабильных мутаций, возникших и существующих в природных популяциях, во-вторых, в прослеживании изменения, происходящие с линиями на протяжении длительного времени (до 20 лет) их поддержания и наблюдения в лаборатории. Наличие клонов Х-хромосом с известными родословными позволяет описать не только статику, но и динамику этого процесса. Цели и задачи исследования. В работе поставлены следующие цели исследования: 1) Изучение динамики и выявление закономерностей формирования генетической изменчивости в природных популяциях Drosophila melanogaster в пространстве и во времени. 2) Изучение характера генетического полиморфизма и свойств мутаций, выделяемых из природных популяций в период вспышек мутаций. 3) Определение относительной роли нестабильных мутаций в формировании генофонда популяций. 2 Особенностью проводимых нами исследований является анализ феномена вспышек мутаций в природных популяциях Drosophila melanogaster, связанного с генетической нестабильностью. В соответствии с этим, в задачи работы входит: 1) Генетический мониторинг аллелофонда и динамики спонтанного мутационного процесса в природных популяциях. Изучение и анализ синхронных изменений концентрации мутаций и локус-специфической мутабильности в природных популяциях Drosophila melanogaster из смежных и географически удаленных регионов. 2) В период глобальной вспышки мутаций (начало в 1973 г.) мы выделили серии нестабильных аллелей гена singed, а в период локальной вспышки (начало в 1982 г.) - аллелей гена yellow. Это явилось базой для классификации нестабильных аллелей локусов Х-хромосомы - singed и yellow, выделенных из различных популяций в течение вспышек мутаций на основе двух главных параметров - оценки мутационных свойств и частот межаллельных переходов. 3) Получение комплекса генетических данных о правилах поведения выделенной из природы бимутантной системы sn49::Tn-clw в ряду поколений при длительном разведении линий в лаборатории. 4) Изучение влияния генетического окружения (хромосом, генов-мутаторов, цитотипа) на мутационные свойства нестабильных аллелей. 5) Сравнительный анализ мутационных свойств нестабильных мутаций в генеративных и в соматических клетках. Научная новизна. Особенностью проведенных исследований является комплексный подход и непрерывность изучения состава и динамики генофондов природных популяций Drosophila melanogaster. В наших работах были установлены следующие важные черты вспышек мутаций: (1) Вспышки мутаций могут быть как глобальными, так и локальными. При глобальной вспышке мутантные аллели одних и тех же генов появляются одновременно в географически удаленных популяциях D. melanogaster. Этот феномен получил название "моды на мутации" [Голубовский и др., 1974]. (2) Наблюдалось повторное проявление моды на мутации, например, для гена yellow первая вспышка 30-40-х годов носила глобальный характер, а "возвращение моды" в 80-е годы - локальный [Захаров, Голубовский, 1985, 1989; Захаров и др., 1995; Захаров, Скибицкий, 1995]. (3) Изучена динамика вспышек мутаций. Длительность вспышки - 7-11 лет. (4) Мутации, определяющие вспышку, в своем большинстве не3 стабильны и, на основании комплекса их генетических свойств, могут быть отнесены к разряду инсерционных мутаций [Golubovsky et al., 1977; Голубовский, Захаров, 1982; Захаров, Голубовский, 1984а; Yurchenko et al., 1984; Захаров, Юрченко, 1985, 1989]. Проведен детальный генетический анализ серии нестабильных аллелей генов singed, yellow, lozenge, который включал определение частот и направлений мутирования различных нестабильных аллелей и их мутантных производных в ряду поколений, изучение влияния генетических и эпигенетических факторов на мутационные свойства нестабильных аллелей в половых и соматических клетках, показана аллелеспецифичность нестабильности и комплексный характер контроля ее мутабильности [Захаров, 1984а; Захаров, Голубовский, 1984, 1985; Захаров, Юрченко, 1985; Голубовский и др., 1986, 1987; Захаров и др., 1995; Вайсман, Захаров, 1995а; Захаров, Скибицкий, 1995]. Впервые исследовался случай совместного мутирования двух генов (бимутантная нестабильная система sn49::Tn-clw), выделенных из природной популяции [Голубовский, Захаров, 1979; Golubovsky, Zakharov, 1980a; Захаров, Юрченко, 1982, 1983, 1984; Yurchenko et al., 1984]; проведен генетический анализ гомологичной рекомбинации нестабильных аллелей гена singed в гомо- и гетероаллельных комбинациях, показано влияние структуры гомологичного локуса на частоту мутирования нестабильного аллеля [Юрченко и др., 1985 а, б; Юрченко, Захаров, 1990]; изучено взаимодействие нестабильных аллелей, связанных с различными мобильными элементами, в системе генотипа [Голубовский и др., 1986]. Причины, индуцирующие синхронную мутабильность в одном и том же гене в удаленных популяциях, должны отвечать следующим требованиям: они должны постоянно присутствовать в природных популяциях; они специфичны к определенным локусам-мишеням, индуцируя в них генетическую нестабильность; они меняют со временем интенсивность и направленность своего действия. Было показано, что нестабильное состояние присуще и аллелям генов, выделенных из природных популяций в которых вспышек мутаций не наблюдалось [Захаров и др., 1995]. Полный перечень установленных событий в экспериментах, связанных со вспышками мутаций, указывает на комплексный характер феномена генетической изменчивости и сложность факторов ее формирующих. Апробация результатов. Материалы диссертации были доложены, представлены тезисами докладов или стендовых сообщений: на 3-6-м Всесоюзных совещаниях по проблемам биологии и 4 генетики дрозофилы - Харьков-1979, Минск-1985, Львов-1987 и Одесса-1989; на 1-3 школах-семинарах по генетике и селекции животных - Улан-Уде-1981, Новосибирск1985 и Бийск-1989; на 45-м симпозиуме в Колд-Спринг Харборе "Подвижные генетические элементы", Колд-Спринг Харбор, США, 1981; на 4-м и 5-м съездах Всесоюзного общества генетиков и селекционеров им. Н.И. Вавилова - Кишинев-1981 и Москва-1987; На 5-м и 6-м Всесоюзных симпозиумах "Молекулярные механизмы генетических процессов" - Москва, 1983 и 1987; на 2-й и 4-й Всесоюзных конференциях "Экологическая генетика растений и животных", Кишинев - 1984 и 1991; на 3 Всесоюзном совещании "Фенетика популяций", Саратов, 1985; на 1-й и 2-й Всесоюзных конференциях по проблемам эволюции, Москва - 1985 и 1988; на Всесоюзном совещании "Экология популяций", Новосибирск, 1988; на 12-м Европейском конгрессе по исследованиям на дрозофиле, Майнц (ФРГ), 1991; на 1-й Всесоюзной конференции по генетике насекомых, Москва, 1991; на научной конференции "Оценка последствий радиационного воздействия ядерных взрывов на Семипалатинском испытательном полигоне и антропогенного загрязнения окружающей среды на население Алтайского края", Барнаул, 1993; на 1-м съезде Вавиловского общества генетиков и селекционеров, Саратов, 1994.; на отчетных сессиях Института цитологии и генетики СО РАН, Новосибирск, 1988, 1991 и 1994. Объем и структура работы. В качестве диссертации представляется совокупность из 44 опубликованных работ. Основные результаты и вытекающие из них выводы изложены в виде научного доклада. Фактический материал получен автором самостоятельно и в коллективных исследованиях с соавторами опубликованных работ: М.Д. Голубовским, Н.Н. Юрченко, А.В. Иванниковым, Е.Э. Скибицким, О.А. Соколовой, Н.Я. Вайсман, М.А. Волошиной и другими, всем им автор выражает глубокую и искреннюю признательность. Работа выполнена в лаборатории генетики популяций ИЦиГ СО РАН. Работа поддержана ГНТП "Приоритетные направления генетики". МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ Эксперименты проводились в следующих направлениях: (а) Сбор и генетический анализ выборок Drosophila melanogaster из природных популяций, представляющих экологически сходные и контрастные регионы на терри5 тории бывшего СССР - южные районы европейской и азиатской части (юг Западной Сибири, Дальний Восток) России, Украина, Кавказ, Средняя Азия. Особое внимание уделено анализу популяции Умань, которая находится под наблюдением с 30-х годов. Все мухи анализировались под бинокулярным микроскопом для установления фенотипических отклонений. Всех самцов с аномалиями затем индивидуально скрещивали с самками со сцепленными Х-хромосомами для выделения сцепленных с полом видимых мутаций. (б) Одновременная оценка концентрации и частоты возникновения мутаций Ххромосомы возможна с использованием инбридинга оплодотворенных в природе самок. Эти методы дают возможность изучать изменения в концентрации и мутабильности отдельных сцепленных с полом генов и сравнивать удаленные популяции, следить за составом и динамикой генофондов в популяциях во времени без вовлечения в скрещивания лабораторных линий. (в) Особенностью и основой наших исследований нестабильных мутаций является семейный анализ, который, во-первых, позволяет проследить генеалогию межаллельных переходов и установить факты мутирования в исследуемой Х-хромосоме, вовторых, наряду с суммарной оценкой "частота мутирования" аллеля, мы получаем значения возникновения мутации de novo в семьях, а также можем установить стадию на которой произошло мутационное событие. (г) Популяционный и генетический анализ серий нестабильных аллелей Ххромосомы из природы включал, во-первых, изучение состава и динамики генофондов популяций Drosophila melanogaster, во-вторых, классификацию нестабильных мутаций генов singed и yellow и других генов Х-хромосомы, выделенных из природной популяции в период вспышки мутабильности. (д) Выделение из популяций, генетический анализ, изучение свойств и взаимодействия сцепленных с полом мутаций и генов-мутаторов в системе генотипа проводили с использованием маркерных генов и линий по принятым схемам скрещиваний. Подробное описание материалов и методов, детали и особенности генетических экспериментов описаны в соответствующих разделах опубликованных работ. 6 РЕЗУЛЬТАТЫ Глава 1. ДИНАМИКА И ОСОБЕННОСТИ ВСПЫШЕК МУТАЦИЙ В ПРИРОДНЫХ ПОПУЛЯЦИЯХ DROSOPHILA MELANOGASTER Колебания концентрации мутаций и неравномерность темпа спонтанного мутационного процесса в природных популяциях и в лабораторных линиях различных видов дрозофил отмечались целым рядом исследователей. Однако, непрерывных, длительных по времени, с широким географическим охватом исследований популяционной динамики генного состава и динамики темпов (скорости) спонтанного мутационного процесса в популяционной генетике, по существу, нет. Благодаря преемственности в работе нескольких поколений советских генетиков-популяционистов, возникла уникальная возможность свести воедино данные, полученные за более чем полувековой период, при изучении природных популяций Drosophila melanogaster на обширной территории СССР. Результатом этих исследований стало обнаружение и описание целого ряда новых популяционно-генетических феноменов: глобальные и локальные вспышки мутаций или "моды на мутации", возвращение моды на мутации, выделение из популяций и генетический анализ серий нестабильных аллелей разных генов. Начиная с 1937 года, одновременно по всему ареалу вида в географически удаленных популяциях Drosophila melanogaster на территории СССР в повышенной концентрации распространились сцепленные с полом мутации yellow body и в меньшей степени наблюдалась повышенная активность и в других локусах Х-хромосомы white eyes и singed bristles [Берг и др., 1941; Гершензон, 1941; Дусеева, 1948] (cм. рис. 1 и 2). В двух популяциях - Воронежа и Душанбе, Н.Д. Дусеева (1948) обнаружила повышенную мутабильность по локусу white. По данным Р.Л. Берг (1965) превышение концентрации и мутабильности наблюдалось и для генов singed и forked bristles, хотя и в меньшей степени, чем для гена yellow. Возможно, что мутационная активность по генам white и singed географически имела мозаичную картину распространения. Как оказалось, наблюдаемая в этот период повышенная концентрация мутаций в популяциях, сопровождалась (или была следствием) повышенной мутационной активностью нормальных аллелей этих генов, особенно уеllow+ [Дусеева, 1948; Гершензон, 1980]. Интерес исследователей в 30-40-е годы был сфокусирован на описании наблюдаемого феномена и на определении частоты возникновения мутаций, которая была на порядок выше обычной и достигала, например, для yellow-мутаций значений 0.02-0.08%. Исследования периода 1937-1946 годов ограничились лишь изучением 7 мутационных свойств нормальных аллелей и, к сожалению, остались неизученными мутабильные свойства самих мутантных аллелей. Поэтому, исследования первого случая вспышки мутаций в природных популяциях оставили открытым вопрос о генетических (в частности, мутационных) свойствах выделенных из природы аллелей мутаций yellow, white, singed и forked. По характеру географического распространения мода на мутацию yellow в период 1937-1946 годах носила глобальный характер. Всего было исследовано 35 популяций Европейской и Азиатской части СССР. В среднем, при частотах возникновения yellow-мутаций 0.38% и 0.06% концентрация yellow-мутаций достигала значений 0.06% и 0.17% [Дусеева, 1948; Берг, 1948]. Если за период наблюдения (1937-1945 годы) разница между частотами yellow-мутаций в популяциях и частотами их возникновения (мутабильностью) была незначительной, то в 1946 году для двух исследованных популяций Р.Л. Берг было отмечено резкое различие по этим двум параметрам: в популяции Тирасполя значение мутабильности равное 0.023% было на порядок ниже, чем значение концентрации - 0.233%, в популяции Умани концентрация оставалась без изменения - на уровне 0.3%, при резком падении мутабильности по сравнению с 1937 годом (см. рис. 1). Период вспышки мутаций 30-40-х годов в популяциях продолжался, по меньшей мере, десятилетие. После 1946 года исследования природных популяций дрозофил на территории Советского Союза были прерваны до 1957 г. 1.1. Возвращение моды на мутацию yellow После периода вспышки мутаций 30-40-х годов концентрация и частота возникновения yellow-мутаций были фоновыми и не превышали уровня 0.04% [Берг, 1961, 1965]. Замечательное исключение представляет популяция Drosophila melanogaster Умани, где, начиная с 1982 года, нами было обнаружено резкое возрастание концентрации yellow-Х-хромосом и прослежена год за годом динамика вспышки (Табл. 1) [Захаров, Голубовский, 1985, 1989; Голубовский и др., 1987; Захаров и др., 1995; Захаров, Скибицкий, 1995]. Только в популяции Умань в период с 1982 по 1991 годы с повышенной частотой встречались мутантные yellow-самцы - средняя частота за весь период 0.9% (N=11139), гетерозиготные по yellow-мутации самки - средняя частота за весь период 1.7% (N=3217), и оплодотворенные в природе мутантными yellow-самцами самки - средняя частота за весь период 0.8% (N=2609). Концентрация 8 yellow X-хромосом в популяции Умань за весь период в среднем составила 0.9% (N=20182). Средняя концентрация yellow-X-хромосом в 30 раз превышает среднюю частоту возникновения yellow-мутаций, которая равнялась 0.03% (N=198210). Динамика концентрации и мутабильности yellow показана на рис. 1. Хотя мы и говорим о "возвращении моды на мутацию", по своей сущности, два пика вспышек по yellow различаются по целому ряду свойств (Табл. 2). Как уже было отмечено, в отличие от глобального в 30-40-х годах распространения yellow-мутаций, в 80-х наблюдалась лишь локальная вспышка по yellow, которая была продолжительна по времени, но зарегистрирована только в популяции Умань. Здесь следует отметить, что в период с 1981 по 1993 годы выборки Drosophila melanogaster брались по всему ареалу вида из 36 различных популяций Европейской и Азиатской части СССР. Отметим еще одну разницу двух периодов –уровень концентрации yellow-Xхромосом в 80-х годах в 30 раз превышал уровень частоты возникновения мутации, тогда как в 30-40-х годах эти параметры были близки, за исключением последнего 1946, года наблюдения [Берг, 1948] (см. рис. 1). Экспрессия yellow-аллелей носит тканеспецифический характер. В соответствии с их фенотипическим проявлением выделяют два типа аллелей: yellow-1 - все кутикулярные структуры - тело, аристы, крылья, макро- и микрохеты имеют желтый цвет, и yellow-2 - тело желтое, а остальные структуры практически неотличимы от окраски дикого типа [Lindsley, Zimm, 1992]. По данным С.М. Гершензона (1941) в популяции Китаево (Украина) в 1937 году возникали в основном мутации типа y1. В исследованиях Н.Д. Дусеевой (1948), вновь возникавшие аллели yellow были как y1-, так и y2-типов. Согласно нашим данным, в период вспышки 1982-1991 годов были выделены из природы и вновь возникали исключительно мутации y2-типа [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров, Скибицкий, 1994, 1995]. 1.2. Вспышка мутаций разных генов, вызывающих сходные аномалии - abnormal abdomen В период между двумя вспышками по yellow-локусу были выявлены еще два мутационных периода: "abnormal abdomen" и "singed". В 1968 году во всех изучаемых популяциях в необычно высоких концентрациях распространилась аномалия, нарушающая структуру тергитов и стернитов брюшка, сходная с известной мутацией abnormal abdomen [Berg, 1972, 1973, 1974; Голубовский и др., 1974]. Динамика распространения данной аномалии показана на рис. 3. Увели9 чение концентрации аномальных мух произошло практически одновременно и во всех исследуемых популяциях. Однако, в отличие от моногенных, с известной локализацией мутаций yellow и singed, гены, определяющие признак abnormal abdomen, могут быть как сцеплены с полом, так и локализованы в аутосомах, признак может иметь полигенный характер наследования и в своем проявлении характеризуется неполной пенетрантностью и изменчивой экспрессивностью. Концентрация аномальных самок и самцов в период вспышки 1968-1975 гг. достигала значений нескольких десятков процентов, что свидетельствует о высоком уровне abnormal abdomen в популяциях по сравнению с фоновым уровнем. Анализ генетической изменчивости природных популяций показывает, что многие из мутаций характеризуются сложной картиной проявления и взаимодействия, обладают неполной пенетрантностью и изменчивой экспрессивностью, плейотропным действием, связаны с хромосомными мутациями (инверсиями) [Захаров, 1985; Захаров, Юрченко, 1988; Захаров, Иванников, 1991; Захаров и др., 1989, 1991, 1993; Вайсман, Захаров, 1995б; Weisman et al., 1995; Юрченко, Захаров, 1995; Юрченко и др., 1995]. Повышенная концентрация мутации в популяции может быть следствием действия целого ряда причин: результатом отбора за счет преимущества носителей мутантного гена (признака), она может поддерживаться за счет повышенной частоты общего или локус-специфического мутирования. Если повышение концентрации в случае с abnormal abdomen явилось следствием повышенной концентрации фенотипически сходных мутаций, относящихся к разным генам, в результате возросшей мутабильности, то подобного рода явление можно соотнести с явлением, известным под названием "гетерогения сходных фенов". Инъекция ДНК-содержащих онкогенных вирусов в полярную плазму эмбрионов D. melanogaster приводит в потомстве к массовому появлению полудоминантных гетерогенных глазных мутаций типа Lobe и гетерогенных наследственных опухолей [Gazarian et al., 1987]. Другой пример - линия claret у D. ananassae несет экстрахромосомный фактор, который активирует ретротранспозон tom, в составе последнего предполагаются энхансеры, действующие в цисположении и специфичные для измененной (усиленной или эктопической) экспрессии генов [Hastie, 1986; Hinton, 1984, 1988; Awasaki et al., 1994]. В результате возникают гетерогенные мутации Om, приводящие к уменьшению размеров глаз и локализованные примерно в 20 локусах. Сам по себе tom-элемент способен встраиваться в разные локусы генома, но выражение на уровне фенотипа получают в основном полудоми10 нантные глазные мутации. Аналогичным образом можно представить, что вспышка мутаций типа abnormal abdomen явилась индикатором ответа на возможную вирусную инфекцию или активацию мобильных элементов в популяциях Drosophila melanogaster. Таблица 1. Концентрация и частота возникновения de novo мутаций yellow в популяции Умань и в других природных популяциях Drosophila melanogaster [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров и др., 1995; Захаров, Скибицкий, 1995] Концентрация Умань Все другие популяции* Год Частота возникновения Умань Все другие популяции N** % N % N % N % 1980 1241 0.08 2674 0 - - 17144 0 1981 1660 0.12 9105 0.01 4800 0 35808 0.003 1982 342 3.22 5596 0 2929 0 31991 0.001 1983 2174 0.78 11569 0 4648 0 30570 0 1984 3691 0.49 7286 0.01 27141 0.011 42139 0 1985 720 2.50 3617 0 13083 0 61725 0 1986 2879 0.87 8409 0 20305 0.005 86142 0.001 1987 2258 1.73 7234 0 34718 0.112 63295 0 1988 1738 1.04 5530 0 32219 0.016 123639 0 1989 2047 0.88 1216 0 21946 0.027 12117 0 1990 1531 0.46 3123 0 16076 0 24123 0.004 1991 1142 0.35 - - 20345 0 - - 1992 - - 5283 0 - - 105903 0.001 1993 885 0 882 0 10195 0 882 0 * Ежегодно исследовалось несколько популяций из различных регионов. За период 1981-1993 гг. исследовалось 36 популяций. Например, кроме популяции Умань, в 1986 г. были изучены популяции: Чернобыль, Ужгород, Брест, Одесса, Запорожье и Самарканд, в 1990 г. - Запорожье, Магарач, Минеральные Воды, Хорог, Душанбе. 11 ** N - общее количество изученных Х-хромосом. 12 Êî í öåí òðàöèÿ (%) 3 0.08 2 0.04 1 0 1930 1940 1950 1960 1970 Ä àòà 1980 1990 ×àñòî òà âî çí èêí î âåí èÿ (%) 0.12 0.00 2000 Риc. 1. Концентрация и частота возникновения yellow-мутаций в природных популяциях Drosophila melanogaster на террито- рии СССР в период 1931-1993 гг. Данные по популяции Умань приведены отдельно. Обозначения: - частота возникновения yellow-мутаций в популяциях; - концентрация yellow-мутаций в популяциях; - частота возникновения yellow-мутаций в популяции Умань; - концентрация yellow-мутаций в популяции Умань. Данные по годам были получены: 1931-1935 - [Дубинин и др.,1934 а, б; Дубинин,1937]; 1937 - [Оленов, Хармац,1938; Гершензон,1941; Берг,1948]; 1938 - [Берг,1948; Дусеева,1948]; 1939 - [Гершензон,1941; Берг,1948; Дусеева,1948]; 1940 1943 - [Дусеева, 1948]; 1941 - [Берг,1948]; 1945 - [Берг,1948; Дусеева, 1948]; 1946-1958, 1957-1964 - [Берг,1948, 1961; Berg, 1965]; 1963-1984 - [Захаров, Голубовский,1985; Голубовский и др.,1987]; 1972-1993 - [Захаров и др.,1995]; 1985-1993 - [Захаров, Скибицкий, 1995; Захаров и др., 1995]. В период с 1947 по 1956 годы популяции не исследовались. С 1966 по 1971 годы популяции исследовались Р.Л. Берг и качественно известно, что содержание yellow-мутаций в популяциях не превышало фонового уровня. Êî í öåí òðàöèÿ (%) 0.06 0.04 0.04 0.02 0.02 0.00 0.00 2000 1930 1940 1950 1960 1970 1980 1990 ×àñòî òà âî çí èêí î âåí èÿ (%) 0.06 Ä àòà Рис. 2. Концентрация и частота возникновения сцепленной с полом мутации singed bristles в природных популяциях Drosophila melanogaster на территории СССР в период 1931-1993 года. Обозначения: - частота возникновения singed; - концентрация singed. Данные по годам были получены следующими авторами: 1931-1935 - [Дубинин и др., 1934 а, б; 1937]; 1937 - [Оленов, Хармац, 1938; Гершензон, 1941; Berg, 1974]; 1938 - [Berg, 1974]; 1939 - [Гершензон, 1941; Berg, 1974]; 1940, 1945, 1946, 1957-1964 - [Berg, 1965]; 1960 - 1973 - [Berg, 1974 a, b]; 1974 - 1983 - [Захаров, 1984; Захаров, Юрченко, 1985]; 1984 - 1993 [Захаров и др., 1995; Захаров и др., неопубликованные данные]. Êî í öåí òðàöèÿ (%) 50 40 30 20 10 0 1930 1940 1950 1960 1970 1980 1990 Ä àòà Рис. 3. Концентрация аномалии abnormal abdomen в природных популяциях Drosophila melanogaster на территории СССР в 1931-1993 гг. Обозначения: - концентрация abnormal abdomen среди самок; - концентрация abnormal abdomen среди самцов. Данные по годам были получены следующими исследователями: 1933-1935 - [Дубинин и др., 1934 а, б; 1937]; 19371940, 1945, 1946, 1957, 1960-1971 - [Berg, 1972]; 1972, 1973 - [Berg, 1973, 1974]; 1973-1993 - [Голубовский и др., 1974; Захаров и др., неопубликованные данные]. Примечание. В отличие от объективно регистрируемых мутаций yellow и singed при сравнении результатов, полученных разными авторами, при количественной оценке признака abnormal abdomen в популяциях, в силу вариабельности его фенотипической экспрессии, следует учитывать субъективность оценок исследователя при проведении границы между нормой и аномальным признаком. Таблица 2. Сравнительная характеристика мутационных периодов в природных популяциях Drosophila melanogaster Евразии в 1931-1994 гг. Годы Длительность Мутация, тип аллеля Характер наследования Характер распространения Концентрация Частота возникновения Отношение концентрации и частоты возникновения 1937-1946 Не менее 10 лет Аллели как yellow-2-, так и yellow1-типа. Другие гены: singed, forked, white. Сцепленная с полом рецессивная мутация 1982-1991 10 лет Только аллели типа yellow-2 Повсеместное распространение yellow-мутаций в популяциях Евразии. Мутация white была обнаружена только в 2 из 35 популяций. 0.05-0.25% Локальная вспышка, проявилась только в популяции Умань (Украина) 0.04-0.15% Концентрация в популяции и частота возникновения мутации de novo примерно одного уровня. Генетическая не- Генетическая нестабильность алстабильность лелей не изучалась. Проверялось ревертирование только некоторых аллелей [Гершензон, 1941]. 16 Сцепленная с полом рецессивная мутация В отдельные годы до 3.2% В среднем 0.03% Концентрация в популяции превышает мутабильность и концентрация на порядок величин выше, чем в 3040-х годах. Серии нестабильных аллелей yellow-2. Зарегистрированы супернестабильные аллели, мутирующие с частотой до 11% [Использованы данные авторов, перечисленных в подписях под рис. 1-3]. 1968-1975 8 лет Аномалии, сходные с мутациями типа abnormal abdomen 1973-1979 7 лет singed bristles, фенотипически и мутационно различающиеся аллели Рецессивные и полудоминантные мутации, мутации с неполной пенетрантностью и изменчивой экспрессивностью, обусловленные разными генами, локализованными в различных хромосомах. Повсеместное распространение в популяциях D. melanogaster Евразии Сцепленная с полом рецессивная мутация Самки - от 10 до 60%. Самцы - от 10 до 30%. Не оценивалась. Обнаружены в различных популяциях Евразии. Скорее всего, следует отнести к мозаичному характеру распространения по ареалу вида. 0.03-0.05% 0.01% Чрезвычайно высокая концентрация, возможно, в результате повышения мутабильности фенотипически сходных мутаций разных локусов. Похоже на явление "гетерогения сходных фенов". Не исключено сильное влияние селективных факторов. Не исследована. Концентрация и частоты возникновения близки. Множественные серии нестабильных аллелей singed. Супермутабильные аллели. Плеяды нестабильных аллелей. 17 Таблица 3. Спектр и разнообразие аллелей гена singed bristles в зависимости от направлений и частот мутирования, выделенных из природных популяций Drosophila melanogaster в 1973-1990 гг. [Golubovsky et al., 1977; Захаров, 1984б; Захаров, Голубовский, 1984а; Захаров, Юрченко, 1985]. Класс A1 Аллель* sns-U36 sns-G78 A2 A3 sns-K3-2 sns-V49 sns-N88 Происхождение, год Умань, 1979 Геленджик, 1978 Краснодар, 1978 Владивосток, 1975 Нальчик, 1988 Направление мутирования Частота snf 3x10-3 sn+ 1x10-3 snm 7x10-4 snm 1x10-3 snf 1x10-4 sn+ 7x10-5 sn+ 8x10-4 snf 7x10-5 sn+ 3x10-3 snm 3x10-4 snm 3x10-4 sn+ 6x10-4 мутирования sns-T77-27 Ташкент, 1973 sn+ 1x10-2 ** sns-11 sns-T77-27 sn+ 2x10-1 ** A5 sns-90719 Магарач, 1990 стабильный*** B1 snm-T63-15 Ташкент, 1973 snf 1x10-2 sn+ 3x10-3 sns 2x10-3 A4 18 Таблица 3, Класс B2 Аллель* snm-K33-13 snm-90667 B3 snm-TB76 snm-Z3195 Происхождение, год Краснодар, 1974 Магарач, 1990 Тбилиси, 1976 Запорожье, 1986 Направление мутирования продолжение Частота мутирования sn+ 1x10-3.** snf 1x10-3 sn+ 3x10-4 snf 1x10-4 sn+ 2x10-3 sns 9x10-4 sn+ 5x10-4 sns 1x10-4 1x10-3 ** B4 snm-K88-9 Краснодар, 1974 sns B5 snm-Z514 Запорожье, 1987 стабильный C1 snf-D163 Душанбе, 1981 sn+ 1x10-1 * Обозначение фенотипического проявления аллелей singed: sns - strong, snm moderate, snf - faint, sn+ - дикий тип, щетинки и волоски практически неотличимы от нормального фенотипа. ** Данные по частоте мутирования этих аллелей взяты из работы [Golubovsky et al., 1977]. *** Здесь и в табл. 4 аллель обозначен как "стабильный" только в том случае, если его стабильность установлена (оценена его частота мутирования на множестве не менее 10 тыс.). 19 1.3. Повышенная частота мутаций гена singed в природных популяциях Drosophila melanogaster в 1973-1979 гг. В 1973-1979 годы в природных популяциях Drosophila melanogaster в повышенной концентрации встречалась другая сцепленная с полом мутация - singed bristles [Berg, 1974; Ivanov, 1974; Иванов, Голубовский, 1977; Захаров, Голубовский, 1984; Захаров, 1984] (рис. 2). Повышенная концентрация мутаций гена singed наблюдалась на фоне повышенной мутабильности нормальных аллелей и вспышка продолжалась до 1979 года. В этот период были выделены нестабильные аллели гена singed, которые при изучении их мутационных свойств дали начало сериям фенотипически различающихся аллелей данного локуса (Табл. 3). В большинстве своем аллели singed из природы оказались генетически нестабильными [Ivanov, 1974; Иванов, Голубовский, 1977; Golubovsky et al., 1977; Голубовский, 1977; Захаров, 1984; Захаров, Голубовский, 1984а; Захаров, Юрченко, 1985]. В работе М.Д. Голубовского и Е.С. Беляевой (1985), методом гибридизации in situ на препаратах хромосом слюнных желез было показано, что в цитологическом районе локализации singed в sn-Х-хромосомах из природы выявляются различные типы мобильных элементов - часть аллелей связана с внедрением P-элементов, часть с внедрением мдг3. Повышенная концентрация мутаций singed в природных популяциях, скорее всего, явилась следствием повышенного мутирования в локусе singed, а не результатом действия отбора. Об этом свидетельствует, во-первых, тот факт, что из 19 выделенных из природы аллелей 12 были зарегистрированы в момент их возникновения, во-вторых, аллели singed характеризовались высокой мутабильностью и обладали свойством многократного мутирования, сохраняющегося на протяжении десятилетий содержания линий в лаборатории. Возникающие при этом фенотипически нормальные аллели (неотличимые от дикого типа) оказывались в такой же степени нестабильными, как и аллели с мутантным выражением признака. В-третьих, аллели singed с фенотипически сходным проявлением могли существенно различаться по спектру и частотам мутирования как в половых, так и в соматических клетках, что говорит в пользу их независимого происхождения. Обычно размножение мутантных Х-хромосом и генетический анализ сцепленных с полом мутаций удобно проводить в скрещиваниях анализируемых самцов с самками лабораторных тестерных линий (со сцепленными Х-хромосомами, Basc и др.). Для некоторых аллелей гена singed это еще и вынужденная мера, так как гомозиготные самки оказываются стерильными. Это ограничение снимается для yellow-X20 хромосом, генетический анализ мутирования которых можно проводить на основе анализа по потомству yellow-самок, не прибегая к скрещиваниям с лабораторными тестерными линиями. Отметим, что именно в наших работах было показано, что повышенная мутабильность в локусе singed не связана с межлинейными скрещиваниями мух из природы с лабораторными линиями, а наблюдается на природном генетическом фоне [Захаров, 1987]. Это было подтверждено и для случая повышения концентрации мутаций по гену yellow в популяции Умань - нестабильное состояние yellow2аллелей наблюдается и в генотипе самок на природном цитотипе и генотипе [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров, Скибицкий, 1995]. Глава 2. ГЕНЕТИЧЕСКИЕ СВОЙСТВА АЛЛЕЛЕЙ ГЕНОВ ВЫДЕЛЕННЫХ ИЗ ПРИРОДНЫХ ПОПУЛЯЦИЙ Х-ХРОМОСОМЫ, DROSOPHILA MELANOGASTER В ПЕРИОД ВСПЫШКИ МУТАЦИЙ После вспышки мутаций в 30-40-х годах, концентрация мутаций yellow в природных популяциях на протяжении почти 4 последующих десятилетий не превышала фоновых и находилась на уровне порядка 10-5. В 1982 году в природной популяции Drosophila melanogaster Умани (Украина) нами было отмечено повышение концентрации мутации yellow [Захаров, Голубовский, 1985, 1989; Голубовский и др. 1987]. Распространение получили только аллели типа yellow-2. Концентрация yellow-2- мутаций с 1982 года по 1991 год в популяции Умань поддерживалась на уровне порядка 10-3-10-2 (Табл. 1). В отдельные годы концентрация yellow-2-Х-хромосом достигала значений 3%. Мутантные самцы и гетерозиготные самки по yellow-2хромосоме в отдельные годы составляли 4%. Основным свойством части выделенных yellow-2-аллелей является их генетическая нестабильность (Табл. 4). Частота мутирования yellow-2 в таких линиях могла колебаться от 10-4 до 10-2. Более того, получаемые в результате мутирования нормальные производные - у+, также могли оказаться нестабильными. Это свойство наблюдалось для аллелей, выделенных на протяжении всего десятилетнего периода вспышки мутаций, а с учетом времени развития Drosophila - на протяжении смены сотен поколений, и нестабильное состояние сохраняется столь продолжительное время как для выделяемых из природы аллелей, так и для содержащихся в лаборатории и вновь возникших в лабораторных условиях мутантных и ревертантных аллелей природных Х-хромосом. Подчеркнем, что мутационная активность ограничилась только одной популяцией - Уманью, и одним локусом - yellow. 21 Выделенные из исследованных в этот период популяций Drosophila melanogaster мутантные аллели других генов могли быть как стабильными, например, некоторые аллели dusky, lozenge, rudimentary singed и vermilion, так и нестабильными dusky, miniatute и singed (Табл. 5) [Захаров и др., 1995; Захаров, Скибицкий, 1994, 1995]. Само по себе присутствие в популяции нестабильных аллелей оказывается не достаточным для поддержания мутации в повышенной концентрации. Об этом свидетельствуют и данные сравнительного анализа соотношения между мутабильностью и концентрацией мутаций двух периодов мутабильности. Если в 30-40-х годах концентрация yellow-мутаций составляла 0,05-0,25% и частота возникновения de novo равнялась 0,04-0,15%, то вспышка 80-х годов имела совершенно другую количественную характеристику. Концентрация yellow-мутаций в популяции Умань в среднем составила 0,9% и в отдельные годы достигала значений 3,2%, что значительно превышало средний уровень частоты возникновения yellow в этот период в популяции - 0,03%. Следовательно, в популяции Drosophila melanogaster Умани должны были сложиться специфические условия, во-первых, для активации мутабильности в локусе yellow и, во-вторых, для сохранения мутантных аллелей и поддержания мутации в повышенной концентрации на протяжении целого десятилетия. Известно, что нестабильное состояние генов, в том числе и yellow, обусловлено процессами внедрения-вырезания мобильного элемента (а в некоторых случаях в гене могут быть локусы-мишени для различных типов мобильных элементов), поэтому полученные нами данные говорят о том, что в популяции должны были возникнуть и существовать длительное время генетические и (или) эпигенетические факторы, инициирующие подвижность мобильных элементов, которые и позволяют поддерживать в популяции инсерционные мутации в высокой концентрации. Таким образом, выявлена и прослежена динамика локального повышения локус-специфической мутабильности и концентрации yellow-мутаций в природной популяции Drosophila melanogaster. Длительность и систематичность генетикопопуляционных исследований популяции Drosophila melanogaster Умани позволяют использовать ее в качестве контрольной. Полученные результаты важны для понимания закономерностей спонтанного мутационного процесса и роли в нем инсерционных мутаций, которые являются значительным компонентом генома. Таблица 4. Группы множественных yellow-2-аллелей, выделенных из природной популяции Drosophila melanogaster Умани в 1983 и в 1990 годах, различающихся по характеру нестабильности исходных аллелей и их мутантных производных [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров, Скибицкий, 1995]. 22 Класс Аллели yellow-2- Мутирование от y2 к y+ частота аллели от y+ к y2 I y2-стабильные 0 II y2U1-5 2x10-3 y+(1-5) y2U23-10 7x10-3 y+(23-10) 1x10-4 y2U1-4 1x10-3 y+(1-4) 2x10-3 y2U1-6 6x10-4 y+691 стабильный y2U23-11 8x10-4 y+(23-11) 4x10-4 y2U23-4 4x10-4 y+(23-4) 3x10-3 y2U23-3 3x10-4 y+(23-3)* 3x10-2 y2U23-8 1x10-4 y+(23-8)* 1.1x10-1 III IV * Частота мутирования Возникшие ревертанты у+-аллели в группе IV, оказываются более нестабильными, чем их исходные у2-аллели. 23 стабильный Таблица 5. Частота мутирования аллелей генов dusky (dy), miniature (m), rudimentary (r), singed (sn), vermilion (v) и yellow (y), выделенных из природных популяций Drosophila melanogaster в 1986-1990 гг. [Захаров и др., 1991, 1995; Захаров, Скибицкий, 1995; Юрченко, Захаров, 1995]. Аллель dy1-155 Всего изучено самцовпотомков Частота мутирования (x10-4) 3 9801 3,1 3, 15, 21 8585 45 Число исключительных потомков в семьях Происхождение Направление мутирования Изучено семей Алма-Ата, dy dy+ 133 dy dy+ 92 dy dy+ 106 * 9 10232 8,8 dy dy+ 103 0 9516 0 1530 388 504 0 0 0 1988 dyZ7-1 Запорожье, 1990 dyZ7-2 Запорожье, 1990 dy90388 Запорожье, lz544 lz562 lz563 1990 Умань, 1985 Умань, 1988 Умань, 1988 m1-47a Умань, 1990 m m+ 102 21, 25 9039 51 r 1-157 Умань, 1989 r r+ 139 0 9372 0 snmZ Запорожье, snm sn + ** 9012 3.3 9012 1.1 1986 ** ** ** snmsnstrong 24 Таблица 5, продолжение Аллель Происхождение Направление мутирования Число ис- Всего Изу- ключи- изучено Частота чено тельных самцов- мутиросемей потомков потом-ков вания (x10-4) в семьях sns-90719 Магарач, 1990 sns sn + 107 0 12508 0 snm90667 Магарач, snm sn + 92 9333 3.2 1990 snmsnslight 1, 2 1 1.0 sn+35 snm90667 sn+ sn 108 0 10065 0 sn+56 v90042 v90088 v90225 v90719 snm90667 Умань, 1990 Умань, 1990 Умань, 1990 Магарач, 1990 sn+ sn v v+ v v+ v v+ v v+ 120 0 ** ** ** ** 10164 11351 9562 9884 13030 0 0 0 0 0 y1-90302 Запорожье, y1 y+ 125 1, 2, 5, 17 11265 22 90 8368 97 12508 0 1990 (y dy)1-156 Умань, 1986 y dy y+dy+ 2, 3, 8, 9, 18, 41 (y+dy+)1-156 (y dy)1-156 y+dy+ y dy 189 0 * В линии зарегистрировано 2 случая возникновения мутации "карликовые крылья" (2 самца), которые дали потомство, но впоследствии линии утеряны. ** Массовые скрещивания. 25 Исходя из совокупности генетических данных, еще до открытия мобильных элементов дрозофилы, был постулирован инсерционный механизм возникновения в природе серии нестабильных аллелей [Green, 1977; Golubovsky et al., 1977]. Нами установлено, что нестабильное состояние аллелей генов характерно для природных популяций D. melanogaster, хотя это и не всегда связано со вспышками мутаций, но вспышки мутаций, как правило, связаны с нестабильностью. Нестабильное состояние выделенных из природы аллелей генов мы рассматриваем в связи с их инсерционной природой [Golubovsky, Zakharov, 1980 a, b; Голубовский, Захаров, 1982; Захаров, Голубовский, 1984 б; Захаров, Юрченко, 1982, 1985, 1988, 1989; Юрченко, Захаров, 1990, 1995; Захаров и др., 1995]. Выделенные в результате проведенных исследований серии нестабильных аллелей генов singed и yellow, аллелей других сцепленных с полом генов являются базой для дальнейших молекулярно-генетических исследований по выяснению структуры и молекулярных механизмов инсерционых мутаций из природы. Глава 3. ГЕНЕТИЧЕСКИЕ СВОЙСТВА БИМУТАНТНОЙ СИСТЕМЫ ИЗ ПРИРОДНОЙ ПОПУЛЯЦИИ DROSOPHILA MELANOGASTER Из популяции Drosophila melanogaster Дальнего Востока была выделена бимутантная система двух тесно сцепленных генов singed и club wing. После того, как было показано наличие мобильных свойств у гена club wing данная система получила обозначение sn49::Tn-clw [Голубовский, Захаров, 1979; Golubovsky, Zakharov, 1980; Yurchenko et al., 1984]. В данной системе мутационные переходы упорядочены в четыре кластера, аллели которых различались фенотипическим проявлением нестабильных singed-производных, направлениями и частотами их мутационных переходов и характером проявления club wing-мутации (Табл. 6, рис. 4). Проведенные исследования нестабильной системы sn49::Tn-clw с помощью метода блот-гибридизации и полимеразной цепной реакции [Юрченко и др., в печати] выявили перестройки внутри первого интрона гена singed-49. В первом интроне у одного из аллельных производных нестабильной системы sn49::Tn-clw найдена инсерционная последовательность с нижним пределом длины 4 тпн. Используя ПЦР-праймеры к этому интрону, был обнаружен полиморфизм длин полученных фрагментов у различных аллельных производных. Принимая во внимание результаты рестрикционного анализа, есть достаточно оснований говорить о том, что в состав Tn-clw может включена последовательность hobo, которая и определяет его мобильные свойства. 26 Плеяды нестабильных аллелей гена singed, по существу, можно рассматривать как своеобразный модельный аналог выделенных из природных популяций серии нестабильных аллелей. Если многообразие нестабильных аллелей, выделенных из разных популяций, в разные годы имеет, безусловно, различную и независимую историю происхождения, то аллели плеяд - это реализованная в процессе размножения потенциальная возможность одного исходного нестабильного аллеля, где в эксперименте прослежена родословная совокупности вновь появившихся в результате мутационных событий аллелей и изучены их свойства. Из природных популяций были выделены фенотипически различающиеся аллели - snstrong, snmoderate и snfaint. Учитывались различия в спектре, направлениях и частотах мутирования. В результате анализа, в соответствии с разнообразием фенотипического выражения исходных аллелей и разнообразием по спектру и направлениям мутирования, мутационного репертуара мутаций серии нестабильных аллелей подразделяются на классы мутаций (Табл. 3). Несмотря на высокие частоты мутирования, направления изменений определенной мутации оказалось ограничено и, таким образом, предсказуемо. Межаллельные переходы внутри плеяд X-хромосомы sn49 также носят устойчивый, регулярный и упорядоченный характер (Табл. 6). При анализе мутирования аллеля sn49 и его мутантных производных, были получены данные, подтверждающие ранее сделанные выводы о характере и некоторых свойствах высокомутабильных генов. Во-первых, были выделены фенотипически различающиеся аллели - snmoderate, snstrong и snextreme, а также генетически нестабильные sn+-аллели. Имеется и ограничение - не было обнаружено аллеля snfaint. Вовторых, были получены аллели, отличающиеся между собой по спектру, направлениям и частотам мутирования. Например, аллель sns49 мутировал к sn+ и обратно с частотой 0,3%, наряду с этим были выделены практически стабильные линии sn+ и sns. С частотой 3х10-4 sns мутировал к snm, который оказался супермутабильным - мутирование в половых клетках достигало значений нескольких процентов. В третьих, зарегистрировано "включение" повышенного мутирования и в соматических клетках 27 Таблица 6. Частота мутирования sn49 и его производных. Четыре плеяды [Голубовский, Захаров, 1979а; Golubovsky, Zakharov, 1980a; Захаров, Юрченко, 1982, 1983, 1984, 1985; Юрченко и др., 1984; Yurchenko et al., 1984]. Плеяда Исходный аллель Направление мутирования Изучено хромосом Частота мутирования sns1 sn+1 19392 2.7x10-3 sn+1 sns1 9743 8.2x10-4 12 sns1 snm2 19392 2.6x10-4 2 snm2 sn+2 7959 2.1x10-2 1 sns2 1.2x10-2 sn+2a sns2 11752 8.9x10-3 sns2 sn+2 5592 1.1x10-2 snm2 5.7x10-3 23 sn+2 snex3 11752 5.1x10-4 3 snex3 sns3 7880 1.6x10-3 sn+3 9.9x10-3 sns3 sn+3 2845 2.8x10-3 sn+3 snex3 5570 1.9x10-2 sns3 34 sn+3 sn+4 clw 4 sn+4 clw sns4 clw+ 1.2x10-2 * 1874 2.2x10-2 * Линия была выделена Н.Н. Юрченко из культуры с высокой пенетрантностью clw (самец №53) после 3-х лет содержания линии sn+3 clw в лаборатории. Мутационные свойства плеяды 4 были изучены Н.Н. Юрченко и Е.У. Болоболовой (неопубликованные данные). 28 Рис. 4. Четыре плеяды бимутантной нестабильной системы sn49::Tn-clw. Обозначения. Схематически изображены нормальный (sn+) и мутантные фенотипы щетинок (snmoderate, snstrong и snextreme), нормальный и мутантный фенотип крыльев (club wing). Появление соматических мозаиков обозначено как mos. Нижние индексы аллелей указывают на принадлежность к плеяде; фенотипически одинаковые sn-аллели, отличающиеся по своим генетическим свойствам, обозначены буквами a и b. Стрелками указаны направления регулярных межаллельных переходов внутри() и межплеядные переходы (). Ссылки на работы и значения частот мутирования по гену singed для некоторых аллелей приведены в Табл. 6. (в плеяде 4). В четвертых, мутирование носило регулярный характер - аллелеспецифические переходы осуществляются на протяжении длительного времени разведения линий. Следует подчеркнуть уникальность sn49-Х-хромосомы – сопряженное мутирование двух мутаций - singed bristles и club wing. Последняя - мутация с неполной пенетрантностью и изменчивой экспрессивностью. С учетом всех свойств, полученное разнообразие вновь возникших аллелей из sn49-Х-хромосомы, распадается на 4 плеяды (рис. 4). Каждая плеяда представляет собой связанную единым происхождением группу аллелей, подчиненных своим внутренним закономерностям мутирования sn и clw, и их экспрессии. В плеяде 1 аллели имеют частоту мутирования в генеративных клетках не выше порядка 10-3, частоту соматических мозаиков - 10-4 [Голубовский, Захаров, 1979а; Golubovsky, Zakharov, 1980]. В плеяде 2 частота мутирования порядка 10-2. Одновременно произошло увеличение частоты появления соматических мозаиков - до 2х10-3 [Захаров, Юрченко, 1982, 1984]. Основателем плеяды 3 является аллель snextreme. Частота перехода от плеяды 2 к плеяде 3 составила 5х10-4 и сопоставима с частотой перехода от плеяды 1 к плеяде 3х10-4 [Юрченко и др., 1984; Yurchenko et al., 1984]. Мутирование внутри плеяды 3 происходит с частотой 3х10-2, не наблюдалось мутирования к промежуточному по фенотипу состоянию snm. В плеядах 1 и 2 мутация clw проявляется только в сочетании с мутацией sns, тогда как в плеяде 3 мутация clw проявляется совместно с snex и sn+. Аллели плеяды 4 характеризуются: 1) высокой частотой мутирования в половых клетках - мутирование от sn+ к sns идет с частотой 2х102; 2) высоким уровнем соматических мозаиков - 3% (sns на фоне sn+-щетинок); 3) высоким уровнем стерильности (90%) самцов sn+. Необычайные мутабильные и другие генетические свойства аллеля sn49 позволили отнести его к инсерционным мутациям [Голубовский, Захаров, 1979а; Захаров, Голубовский, 1980; Golubovsky, Zakharov, 1980]. Предложена следующая реконструкция возникновения sn49-Х-хромосомы. По-видимому, определенный мобильный генетический элемент, внедрившись в район гена clw, захватив часть генетического материала локуса, переместился затем в район гена singed. Так возник транспозон sn::Tn-clw. В результате, два гена в норме независимые, после реконструкции перешли под контроль одного МЭ и обрели способность к совместному проявлению и мутированию. Как подтверждение этого можно рассматривать следующий ряд полученных результатов. 1) Мутация clw в своем проявлении зависит от температуры, от до30 зы Y-хромосом, обнаружен отцовский эффект на проявление мутации - все это в совокупности указывает на сходство с явлением, сопутствующими эффекту положения [Захаров, Голубовский, 1980]. 2) Появление нового, "транспозонного" аллеля clwq, летального по своему действию [Юрченко и др., 1984]. 3) Зависимость мутирования snаллелей от P-M-цитотипа [Захаров, 1984а]. 4) Получены данные о миграции Tn-clw из локуса sn в другой участок Х-хромосомы, находящийся на расстоянии примерно 30 ед. карты от локуса sn [Юрченко и др., 1984]. Показано влияние транспозона на рекомбинацию в соседних с геном-хозяином участках [Юрченко и др., 1985 а, б]. Результаты изучения генетических свойств sn49-X-хромосомы позволили выдвинуть следующие положения: (1) существование внутриплеядных мутационных изменений есть результат внутрилокусных транспозиций мобильного элемента; (2) редкие межплеядные переходы являются результатом мутационных изменений в самой бимутантной системе sn49::Tn-clw. Эти предположения были сделаны в порядке объяснения увеличенной частоты гомологичной рекомбинации, опосредованной этим транспозоном [Юрченко и др., 1985 а, б]. Таким образом, мутационный "репертуар" возникших нестабильных транспозонных singed-аллелей изучен в связи с гомологичной рекомбинацией [Юрченко и др., 1985 а, б; Юрченко, Захаров, 1990]. Глава 4. ВЛИЯНИЕ ГЕНЕТИЧЕСКИХ И ЭПИГЕНЕТИЧЕСКИХ ФАКТОРОВ НА МУТИРОВАНИЕ НЕСТАБИЛЬНЫХ АЛЛЕЛЕЙ Аллель sn49, находясь в одной дозе у самок (в гетерозиготном состоянии), мутирует в 1,5 раза чаще, чем в одной дозе у самцов (в гемизиготном состоянии). Для аллеля sn49m(1-23) частота мутирования в 4 раза выше у самцов по сравнению с частотой мутирования у самок [Захаров, Юрченко, 1982]. Эти результаты свидетельствовали о том, что на мутирование изучаемых нестабильных мутаций может оказывать влияние генетическое окружение и было выявлено разнообразие контроля мутирования нестабильных аллелей [Голубовский, Захаров, 1979 б; Захаров, Голубовский, 1980; Захаров, Юрченко, 1983 а, б; Захаров, 1984 б; Голубовский и др., 1986]. 4.1. Роль P- и M-цитотипов в контроле мутирования нестабильных аллелей гена singed у Drosophila melanogaster Хромосомы с аллелем sn49m(1-23) и его мутантные производные sns и sn+ переводили в цитоплазму линий C(1)DX; y f; cn bw (M-цитотип) и C(1)DX; y f; 2 (Pцитотип). Было показано , что в линии Р-цитотипа супермутабильность резко снижа31 ется - частота мутирования аллелей sns, snm и sn+ в генеративных клетках Р-цитотипа снижается на 1-2 порядка величин и не превышает 10-3 [Захаров, 1984, а]. Последующее возвращение исследуемых хромосом в цитотип стандартной лабораторной линии C(1)DX,ywf снимает репрессию мутирования и частота мутирования возвращается к значению величин порядка 10-2. В линии М-цитотипа C(1)DX; y f; cn bw мутабильность sns, snm и sn+ такая же, как в цитоплазматическом окружении, свойственной линии C(1)DX,ywf. Частота соматических мозаиков в линиях sns и sn+ в Р-цитотипе снижалась на порядок величин - до 10-4, тогда как в линиях snm разницы в частоте появления соматических мозаиков в цитотипе линий C(1)DX,ywf и в Р- и М-цитотипах не обнаружено, и она сохранялась на уровне порядка 10-3. В работе М.А. Волошиной и М.Д. Голубовского (1986) было показано, что если цитоплазма линии с Р-факторами подавляет транспозиции не только Р, но и других МЭ, то эффект Р-содержащих хромосом аллелеспецифичен и может быть нейтральным, активирующим или ингибирующим. Нами были изучены темпы мутирования мутантных и нормальных аллельных производных нестабильной системы sn49::Tnclw (нестабильных в половых и соматических клетка) в различных гибридных комбинациях с линией 2, которая имеет Р-цитотип и содержит активные Р-элементы в своих хромосомах [Голубовский и др., 1986]. Оказалось, что во всех случаях добавление Р-хромосом, безотносительно от фона цитоплазмы, ингибировало нестабильность разных sn-аллелей, связанных, как мы полагаем, с внедрением последовательности Tn-clw. С повышением доли Р-хромосом у гибридов степень подавления мутирования sn-аллелей возрастала. Предполагается, что мутационные события связаны с внутри- и межлокусной транспозицией Tn-clw и что оба мобильных элемента, могут конкурировать за один и тот же клеточный фактор, который активирует процесс транспозиции . 4.2. Мутирование аллелей singed в соматических клетках Большинство выделенных singed-аллелей характеризовались нестабильным состоянием только в генеративных клетках. Однако для некоторых singed-аллелей было свойственно повышенное мутирование и в соматических клетках [Захаров, Голубовский, 1984 а]. Говоря о генеративных клетках, мы имеем в виду мутирование в зародышевом пути, хотя мутирование может происходить как при митотических делениях - в случаях пучкового характера появления исключительного потомства в семьях, так и при мейотическом делении - в случаях появления исключительного потомства в се32 мьях в единичных экземплярах. Отметим свойство "включения" нестабильного состояния в соматических клетках. Например, для аллеля sn49 частота появления соматических мозаиков равнялась 4х10-4. Однако возникновение супермутабильного аллеля sn49m(1-23) в плеяде 3 сопровождалось резким усилением мутаторной активности и в соматических клетках: в отдельных случаях в линиях с sn49m(1-23)-аллелем и его мутантных производных частота соматических мозаиков достигала значений 0,7%, а в плеяде 4 частота появления соматических мозаиков составила 3%. Следует подчеркнуть, что в наших работах впервые показано влияние цитотипа на мутирование в соматических клетках [Захаров, 1984а; Голубовский и др., 1986]. Глава 5. ГЕНЫ-МУТАТОРЫ ИЗ ПРИРОДНЫХ ПОПУЛЯЦИЙ DROSOPHILA MELANOGASTER Рассматривая высокомутабильные гены, мы показали, что их мутационные свойства аллелеспецифичны и обусловлены, прежде всего, свойствами самих генов [Захаров, 1984; Захаров, Голубовский, 1985; Голубовский и др., 1987]. Примером множественной мутабильности в природе служит Х-хромосома из популяции Запорожье с нестабильным аллелем singed-mZ [Юрченко, Захаров, 1995]. Мутация snmZ обнаружена в потомстве оплодотворенной в природе самки, которая была гетерозиготна по мутации Notch. В дальнейшем в XZ-хромосоме наблюдались одновременные множественные мутационные события в локусах yellow, white, vermilion, garnet и др., возникали летальные мутации, которые сопровождались инверсиями и реинверсиями с точками разрывов в области локализации инсерционных мутаций [Юрченко и др., 1995]. Особый интерес представляют гены-мутаторы, которые, не имея собственного фенотипического проявления, могут оказывать влияние на скорость мутирования других генов. 5.1. Фактор Mos - специфический ген-мутатор для нестабильных, нормальных по фенотипу аллелей singed+ Выделенный из природы доминантный ген Mos локализован в хромосоме 3 в районе центромеры - 45-50 ед. карты, и вызывает у гетерозигот Mos/+ частичную редукцию дорзоцентральных и скутеллярных щетинок [Голубовский, Захаров, 1979; Вайсман, Захаров, 1995а]. Действие гена Mos проявляется в виде повышения частоты мутирования в соматических клетках для нестабильных, нормальных по фенотипу ал33 лелей гена singed. На фоне действия Mos мутабильность sn+49 возрастает в 70 раз, для sn+10m мутабильность увеличивается в 6 раз. Два супермутабильных аллеля sn+SR1 и sn+SR2 (нормальные по фенотипу ревертанты аллеля sn77), отличаются необычайно высокой мутабильностью в соматических клетках - частота мозаиков 10 и 20%, соответственно. Но даже и для этих аллелей заметно действие Mos. Mos является специфическим геном-мутатором не только в соматических клетках. Для аллеля sn+10m исключительные потомки возникают в 10% семей (при числе потомков в каждой семье до 70), тогда как на фоне Mos - в 40% семей. В половых клетках в присутствии Mos увеличивается и общая частота мутирования аллеля sn+10m . Поскольку нестабильность генов в большинстве случаев объясняется инсерциями в них мобильных элементов, то ген Mos в генотипе с нестабильными аллелями singed, возможно, выполняет роль регулятора подвижности гипотетического мобильного элемента [Голубовский, Захаров, 1979]. Исходя из ранее обнаруженных генетических свойств Mos, нам представилось интересным рассмотреть взаимодействие его с другими генами, контролирующими формирование кутикулярных структур - щетинок и волосков на теле мухи, изучить влияние гена Mos в цис- и транс-гетерозиготе на частоту мутаций гена multi wing hairs (mwh) в соматических клетках, используемого в тест-системах для определения темпа индуцированного мутагенеза. Анализ взаимодействия генов Mos и mwh в цис- и транс-гетерозиготах показал, что ген Mos оказывает существенное влияние на мутабильность (рекомбиногенез) в соматических клетках mwh-хромосомы. В цис-гетерозиготе mwh Mos/+ + наблюдалось повышение частоты малых мутантных клонов на крыльях мух. Редукция щетинок в результате действия гена Mos была высокой: у mwh/Mos - 78%, у mwh +/+ Mos - 85% и у mwh Mos/mwh + - 98% [Вайсман, Захаров, 1995а]. 5.2. Гены-мутаторы типа MR-факторов В природных популяциях Drosophila melanogaster Восточной Европы, средней Азии и юга Западной Сибири в период с 1977 по 1992 годы были изучены свойства и распространенность MR-факторов [Иванников, Захаров, 1992; Иванников и др., 1995]. Оказалось, что частоты MR-хромосом в популяциях Евразии не отличаются от таковых в Северной Америке и составляют 18-40% хромосом 2. 34 В природных популяциях Drosophila melanogaster широко распространены две мутаторные системы: MR (male recombination) и P-M [Thompson, Woodruff, 1978; Engels, 1989]. Рекомбинация хромосом в геноме самцов является одним из характерных свойств обеих мутаторных систем. Две данные системы имеют много общих черт и иногда рассматриваются как единая, хотя между ними есть и отличия. Так, например, при гибридизации Drosophila melanogaster из природы некоторые хромосомы с высокой частотой рекомбинируют в геноме самцов, но не индуцируют стерильности у самок, то есть могут быть отнесены только к MR-системе [Green, 1984]. Мутаторным агентом P-M-системы на молекулярном уровне является мобильный генетический элемент Р. Присутствие его полноразмерных копий в геноме определяет цитотип индивида: P-цитотип в присутствии Р-элемента или М-цитотип - в его отсутствие. Выделяют также промежуточный цитотип - Q. По критериям Р-М-системы цитотипы индивидов в популяциях Drosophila melanogaster североамериканского и евразийского континентов существенно отличаются [Anxolabehere et al., 1984]. В североамериканских популяциях в высоких концентрациях присутствуют полноразмерные активные Р-элементы, поэтому индивиды этих популяций имеют преимущественно Р- или Qцитотип. В Европе и Азии распространены М- и Q -цитотипы. Это вызвано тем, что Рэлементы в евразийских популяциях представлены в большинстве делетированными неактивными копиями. Следовательно, в глобальном масштабе MR и Р-М мутаторные системы проявляются независимо друг от друга и MR-активность в популяциях Евразии должна определяться другим (другими) элементом. Гены мутаторы типа MR-фактор могут быть причиной повышенного возникновения широкого спектра мутаций, связанных с процессами разрыва-воссоединения хромосом и, тем самым, повышать значения общей мутабильности. Не исключено, что некоторые мутаторы могут обладать направленным, локус-специфичным действием. Известно, что летальная мутация l(2)gl является одной из распространенных мутаций в природных популяциях, высокая концентрация которой поддерживается, возможно, за счет преимущества гетерозигот [Golubovsky, 1978; Golubovsky et al., 1991]. Для мутатора MR h-12 была показана способность индуцировать мутации в локусе l(2)gl [Green, Shepherd, 1979], и высокая концентрация в популяции может быть следствием активности генов-мутаторов. Следовательно, гены-мутаторы могут высту- пать в качестве усилителей общей и локус-специфической мутабильности. Таким образом нами установлено, что наряду с определенной аллелеспецифичностью нестабильных аллелей (спектр, направления и частоты мутирования, 35 характер мутирования в половых и соматических клетках), наблюдается комплексный характер контроля мутабильности нестабильных аллелей (зависимость мутирования от генного окружения, локуса-гомолога, цитотипа, генов-мутаторов). ОБСУЖДЕНИЕ Представленные в настоящей работе совокупные данные по более чем полувековому слежению за генетической изменчивостью и мутационным процессом в природных популяциях Drosophila melanogaster позволяют сделать вывод, что вспышки мутаций - достаточно регулярное событие в жизни вида. Популяционная феноменология данного явления такова: (1) вспышки могут затрагивать отдельные гены или группы генов со сходным фенотипическим проявлением; (2) вспышки могут быть локальными или глобальными, когда в короткое время, практически синхронно, они затрагивают географически удаленные популяции; (3) определенная вспышка (или мода на мутации) продолжается 7-11 лет; (4) возможно повторение моды на мутации определенного гена (генов). Скорость мутирования в популяции должна рассматриваться как некий особенный "фенотип" популяции и как один из важнейших признаков она, несомненно, находится под контролем естественного отбора, его стабилизирующей формы. Вполне разумно ожидать, что с эволюционной точки зрения, скорость мутирования должна быть минимизирована естественным отбором. В случае же выхода скорости мутирования из-под контроля естественного отбора, как, видимо, наблюдается при вспышках мутаций, естественный отбор должен быть направлен на подчинение мутационного давления. Что мы и наблюдаем в реальных ситуациях: после вспышки мутабильности наступает период ее затухания. Тот факт, что в природных популяциях существуют периоды высокой мутаторной активности, может говорить о том, что в популяциях не исключены процессы, аналогичные гибридному дисгенезу, в основе которых лежат сходные явления - активация мобильных генетических элементов. И то явление, которое мы определяем термином "спонтанная мутабильность", может означать лишь то, что нам, к сожалению, неизвестны истинные факторы и причины, определяющие эту мутабильность. Остаются во многом неизвестны и причины вспышек мутабильности в природных популяциях. Вспышки мутабильности сопровождаются переходом определенных генов в нестабильное состояние. Этот вывод наиболее отчетливо демонстрируется в случае 36 глобальной вспышки по генам yellow и singed. Впервые из природы удалось выделить серии мутантных нестабильных аллелей на протяжении всего периода вспышек и провести генетический анализ нестабильности. Аллели различались как по фенотипу, так и по характеристикам мутирования в генеративных и соматических клетках. Хотя локус singed является горячей точкой для внедрения Р-элементов, глобальная вспышка singed оказалась связана с активацией не только Р, но и других элементов [Голубовский, Беляева, 1985]. У наиболее генетически изученного нами аллеля sn49 в первом интроне гена singed была найдена крупная инсерционная последовательность не-P-элементной природы - в составе транспозона обнаружена последовательность hobo-элемента [Юрченко и др., в печати]. Различные алели yellow, также как и singed, связаны с внедрением различных типов мобильных элементов. Например, аллель y76d28 связан с внедрением Рэлемента в локус yellow [Geyer et al., 1988]. Проанализированные в работе [Biessmann, 1985] все аллели y2-типа были связаны с внедрением мобильного элемента gypsy и большинство y2-аллелей локализуются на 0.9 тпн дистальнее от места картирования аллелей y1-типа. Другие аллели y1-типа либо содержат элемент hobo, либо другие, неидентифицированные транспозируемые элементы. Алели типа y2, связанные с внедрением мобильного элемента gypsy, супрессируются супрессором Su(Hw). Однако, по нашим данным ни один из аллелей типа y2, выделенных во время вспышки мутабильности, не супрессировался данным супрессором [Захаров, Скибицкий, неопубликованные данные]. Логично предположить, что вспышка yellow вызвана инсерциями другого, нежели gypsy, мобильного элемента. Стабильные и нестабильные мутации, возникающие в период вспышки, могут быть связаны с активацией разных мобильных элементов, которые способны к сайтспецифическому встраиванию. Подвижные генетические элементы являются неотъемлемым составным компонентом генофонда природных популяций. Известно, что мутации в одном и том же локусе могут вызываться различными типами МЭ. Однако, имея гомологичные сайты внедрения, МЭ различной структуры могут обладать и различной степенью сродства к сайтам-мишеням. Это, в свою очередь, может определять набор состояний, получаемых при мутировании, обуславать различие преимущественных направлений и значений частот мутирования [Голубовский, Захаров, 1982]. Предстоит понять, каким образом в природе происходит активация мобильных элементов? Какие процессы запускают активацию, как происходит синхронизация этих процессов в географически удаленных популяциях, почему она носит волнооб37 разный характер, с чем связана сайт-специфичность встраивания мобильных элементов и связаны ли эти процессы с особенностью самих локусов-мишеней? Ответы могут быть лишь предварительными. Разберем ситуацию на примере локуса singed, в отношении которого есть наиболее подробные молекулярно-генетические и популяционные данные. Этот локус является мишенью для встраивания Р-элементов под действием мутаторов типа MR, а также в результате их активации в системе P-M гибридного дисгенеза [Brookfield, Mitchell, 1985; Gerasimova et al., 1985; Roiha et al., 1988; Engels, 1989; Eeken et al., 1991; Mogila et al., 1992]. Два факта находятся в согласии с этими данными. Вопервых, широкое распространение MR-факторов в исследованных популяциях [Иванников и др., 1995]. Во-вторых, наличие Р-инсерций среди выделенных из природы в период вспышки singed-аллелей. Однако следует учесть, что копии Р-ДНК в евразийских популяциях Drosophila melanogaster (в том числе исследованных нами) стали распространяться лишь в 60-х годах [Anxolabehere et al., 1985, 1988; Periquet et al., 1989; Kidwell, 1994]. Между тем повышение мутабильности sn отмечалось уже в 30-40-х годах (рис. 2). Кроме того, особи из исследованных природных популяций, где зафиксирована вспышка singed, относилась, по нашим данным, к М-цитотипу. Более того, эти особи несли в своих хромосомах делетированные варианты Р-элемента - названные КР, впервые найденные в линии из популяции Краснодара [Black et al., 1987; Jackson et al., 1988]. Они оказались распространенными по всему ареалу вида. Действие КР приводит к супрессии транспозиции полноразмерных копий, хотя присутствие КР-копий в геноме не всегда подавляет Р-опосредованную нестабильность [Ortori et al., 1994]. Следовательно, связь распространения копий P-элемента в конце 60-х годов в популяциях Евразии со вспышкой мутабильности singed 70-х годов существует, но вряд ли является прямой или единственной причиной вспышки мутаций в этом локусе. Еще более трудно объяснить другие типы вспышек, зафиксированных более 50 лет назад. Необходим, на наш взгляд, общебиологический подход для поисков факторов и пусковых механизмов, способных привести к установленным популяционногенетическим закономерностям вспышек мутаций. Все виды в биоценозах испытывают давление инфекционных агентов, и это взаимодействие по своей динамике во времени носит осциллирующий характер. Оно может вызывать локальную и глобальную синхронизацию изменений генофондов по- 38 пуляций, так как скорость распространения инфекционного агента несоизмеримо выше, нежели обычная миграция особей вида-хозяина [Эндрюс, 1969]. Вирусы могут выступать, во-первых, как непосредственные мутагенные факторы, во-вторых, как регуляторы подвижности мобильных элементов. Непрерывно происходящие в биоценозах взаимодействия геномов эукариотических организмов и геномов вируса могут приводить к активации мобильных элементов, что сопровождается множественными инсерционными мутациями и перестройками. Это было показано на разных системах у кукурузы и дрозофилы [McClintock, 1978, 1984; Mottinger et al., 1984; Peterson, 1985; Gershenson, 1986; Gazarian et al., 1987]. В данном контексте очень показательны данные, что локусспецифичные летальные мутации, возникающие в экспериментах под действием вирусных агентов, относятся к тем леталям, которые в разные годы имеют повышенную концентрацию в природных популяциях дрозофил [Александров, Голубовский, 1983]. Мобильный элемент gypsy, вызывающий спонтанные мутации, обладает свойствами и мобильного элемента и ретровируса, способного заражать через корм особей из чувствительных линий [Kim et al., 1994]. В некоторых популяциях дрозофил Европы обнаружены локальные изменения в концентрации разных вариантов рабдовируса "сигма", которые, как предполагается, могут коррелировать с изменениями в Р-М системе гибридного дисгенеза [Fleurie et al., 1992; Fleurie, Sperlich, 1992]. В случае пикорновирусов "iota" у Drosophila immigrans были обнаружены волны инфекций: в 1970-1971 гг. вирус "iota" повсеместно встречался в популяциях Франции, но в 1973 г. он практически исчез [Brun, Plus, 1980]. В ряде природных популяций и лабораторных линий Drosophila melanogaster был обнаружен пикорновирус дрозофилы подтипа C (DCV). Оказалось, что зараженность пикорновирусом ведет к увеличению общей и сайт-специфической мутабильности (Golubovsky, Plus, 1982). При анализе частоты возникновения летальных мутаций в Х-хромосоме линии Paris-Rennes, содержащей пикорновирус подтипа С, М.Д. Голубовским был найден тройной нестабильный мутант y2sc1waG. В ходе генетического анализа этой линии наблюдались множественные упорядоченные транспозиции мобильных элементов, в том числе Stalker и X [Georgiev et al., 1990; Mogila et al., 1992; Ладвищенко и др., 1990; Буфф и др., 1993]. С внедрением в локус yellow элемента Х, содержащего на одном конце копию дефектного Р-элемента, связано возникновение в линии y2sc1waG серии нестабильных мутантных аллелей типа y2 [Georgiev, Elagin, 1992; Елагин и др., 1992; Крачинска и др., 1992] 39 Таким образом, в спонтанном мутационном процессе большую роль играют факультативные генетические элементы генома, куда можно отнести разные классы мобильных элементов и вирусы [Temin, Engels, 1984; Smith, Corces, 1991]. Спонтанный мутационный процесс можно в этом смысле рассматривать как двухэтапный. Вначале происходит активация факультативных элементов генотипа, которые наиболее чувствительны к слабым "немутагенным" воздействиям среды, понимаемой в широком смысле. Возникают наследственные генотипические изменения, которые, в большинстве случаев, не выходят за рамки морфофизиологической нормы, как, например, транспозиционные взрывы [Gerasimova et al., 1985]. Такого рода изменения можно отнести к предмутационным. И лишь на втором этапе возникают классические генные или хромосомные мутации, опосредованные действием факультативных элементов генома [Golubovsky, Kaidanov, 1994]. Инсерционная природа мутагенеза расширяет наши представления о роли мутационного процесса. Многообразие мутаций может создаваться ускоренно и направленно. При этом наряду с генными мутациями, которым отдавалось предпочтение раньше при рассмотрении процессов микроэволюции, могут возникать и сложные перестройки генома. Пучковый характер возникновения новых мутаций служит своеобразным механизмом быстрого размножения новообразований. Их дальнейшую судьбу определяет отбор. Рассмотрим еще один важный аспект проблемы вспышек мутаций, а именно, имеют ли значение биологические свойства самих генов, определяющих вспышку. Не исключено, что фенотипически определяемые видимые мутации yellow body, singed bristles и abnormal abdomen выступают своеобразными индикаторами взаимодействия и реорганизации аллелофонда популяции с факторами окружающей среды по признакам компонентов общей приспособленности. Экспрессия yellow, singed и группы генов abnormal abdomen играет важную роль в развитии организма и не ограничивается признаками, обозначенными рамками исторически полученных названий. Это мутации с тканеспецифичным, широким спектром плейотропного действия на биохимическом, цитологическом и физиологическом уровнях. Отмечая, что вспышка мутаций типа abnoral abdomen формировалась за счет различных генов с одинаковым действием на фенотип [Berg, 1974], следует иметь ввиду, что их пенетрантрость и экспрессивность зависит от целого ряда факторов. Например, сцепленный с полом ген A53g в своем формировании мутантного фенотипа связан с действием системы генов-модификаторов. Первичный эффект на развитие 40 мутантного генотипа состоит в нарушении дифференцировки зрелых гистобластов в результате чего материал тергитов формируется неравномерно или утрачивается вовсе, приводя к аномалиям в формировании абдоминальной гиподермы. Вторичный эффект состоит в препятствии слиянию гистобластов. Пенетрантность и экспрессивность abnormal abdomen в сильной степени зависят от различных факторов внешней среды - температуры, влажности, плотности и др., при которых происходит развитие. Мутантный фенотип образуется в результате взаимодействия главного гена и систем модификаторов в сочетании с внешними факторами среды [Roseland, Schneiderman, 1979; Hillman, 1973]. Ген yellow у Drosophila melanogaster определяет не только тканеспецифическую пигментацию отдельных кутикулярных структур личинки и имаго, но и тесно связан с комплексом achaete-scute, который контролирует формирование и дифференцировку щетинок и волосков взрослой мухи [Biessmann, 1985; Campuzano et al., 1985; Mackay, Langley, 1990; Walter et al., 1991]. Ген singed у Drosophila melanogaster играет важную роль как в соматических, так и в половых клетках. Мутации по гену singed изменяют форму щетинок и волосков у имаго, аллели различаются по степени выраженности мутантного признака от сильного до слабого. Дополнительно к этому, некоторые singed-аллели в гомозиготном состоянии вызывают стерильность самок, хотя гемизиготные самцы фертильны [Bender, 1960; Lindsley, Zimm, 1992]. Оказалось, что его нормальное функционирование имеет ключевое значение в половом пути в течение овогенеза [Perrimon, Gans, 1983]. Наряду с другими генами - chickadee и quail, ген singed кодирует белки, гомологичные к актин-связывающему белку, который играет важную роль в формировании цитоскелета. Мутации по гену singed приводят к нарушению образования пучков из актиновых филламентов, которые играют решающую роль в перемещении цитоплазмы. Это, в свою очередь, приводит к нарушению морфогенеза щетинок и аномальному овогенезу [Cooley, Thewkauf, 1994]. В заключение мы должны с определенностью подчеркнуть, что открытие мобильных генетических элементов и наличие инсерционных мутаций расширило наше понимание механизмов формирования наследственной изменчивости и, делая основной упор в своей работе на изучение мутационного процесса в природных популяциях, на исследование нестабильных мутаций, мы ни в коей мере не принижаем и не пересматриваем ведущей, основной и творческой роли отбора в эволюции. 41 ВЫВОДЫ Исследован и обобщен характер генетической изменчивости, изучена динамика аллелофонда в природных популяциях Drosophila melanogaster на территории бывшего СССР. 1. Изучено явление неравномерности темпа мутационного процесса и периоды повышения концентрации мутаций отдельных генов и их аллелей в природных популяциях Drosophila melanogaster. а) Прослежена динамика повышения концентрации и мутабильности сцепленного с полом гена singed в 1973-1977 гг. в географически удаленных природных популяциях на территории бывшего СССР. б) Обнаружено и исследовано возвращение "моды" на мутацию - повышение концентрации и мутабильности по сцепленному с полом гену yellow. Первый период мутабильности был отмечен по всему ареалу вида отечественных популяций полвека назад - конец 30-х - первая половина 40-х годов, второй период имел локальный характер - начался в 1982 г. и продолжался до 1991 года. в) Обобщены и описаны границы периода повышения концентрации аномалии abnormal abdomen, наблюдавшейся в 1972-1979гг. 2. Из природных популяций были выделены и изучены серии множественных нестабильных аллелей сцепленных с полом генов, определяющих вспышки мутаций singed и yellow, и сопутствующих - dusky, miniature, vermilion и др. а) Проведена классификация выделенных аллелей генов singed и yellow в соответствии с характером и степенью их мутантного выражения, спектра и частот мутирования как их самих, так и их мутантных производных. Имеется определенная аллелеспецифичность в отношении свойств мутирования - как по частотам мутирования, так и по репертуару наблюдаемых межаллельных переходов. Межаллельные переходы носят устойчивый, регулярный и упорядоченный характер. "Переключение" на другой тип мутирования происходит на порядки величин реже, чем мутирование внутри типа. б) При изучении аллелей гена singed, выявлены аллели, проявляющие нестабильные свойства как в генеративных, так и в соматических клетках. 3. Проведен генетический анализ выделенной из природы нестабильной генной конструкции sn::Tn-clw. Изучено в ряду поколений разнообразие вновь возникших нестабильных аллелей, в соответствии с их генетическими свойствами (экспрессия, ча- 42 стоты и направления мутирования) выделены их группы (плеяды) нестабильных аллелей. 4. Изучено влияние различных генетических и средовых факторов на мутационные свойства нестабильных аллелей. Исследовано действие доминантного специфического гена-мутатора Mos в отношении нестабильных, фенотипически нормальных аллелей singed+. Мутабильность в генеративных клетках в присутствии Mos возрастает более чем в 2 раза, а частота появления соматических мозаиков увеличивается для разных аллелей от 2 до 70 раз. Установлено влияние P- и M-цитотипов на гонадную генетическую нестабильность высокомутабильных аллелей singed. В P-цитотипе происходит резкое подавление мутационной активности нестабильных аллелей и частота мутирования не превышает порядка величин 10-3. В M цитотипе происходит возвращение частоты мутирования к исходному значению - 10-2. Впервые показано влияние цитотипа на мутирование в соматических клетках. Предполагается, что ген Mos может выступать в роли гена-регулятора подвижности мобильных элементов. 5. Показано широкое распространение в популяциях Drosophila melanogaster Евразии в большой концентрации генов-мутаторов типа MR, которые могут вызывать повышение скорости спонтанного мутирования (широкого спектра мутаций, связанных с разрывом хромосом). 6. Полученные данные рассматриваются в рамках концепции инсерционного мутагенеза у эукариот. Это дает возможность целенаправленно изучать и прогнозировать характер мутационных событий, вызванных внедрением и перемещением мобильных генетических элементов. Обоснование феномена неравномерности темпа мутационного процесса и колебаний концентраций мутаций в природных популяций имеет важное значение для теории эволюции. СПИСОК ОСНОВНЫХ ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ 1. Голубовский М.Д., Иванов Ю.Н., Захаров И.К., Берг Р.Л. Исследование синхронных и параллельных изменений генофондов в природных популяциях плодовых мух Drosophila melanogaster // Генетика. 1974. Т. 10. № 4. C. 72-83. 2. Голубовский М.Д., Захаров И.К. Совместные реверсии двух нестабильных мутаций в Х-хромосоме Drosophila melanogaster // Генетика. 1979а. Т. 15. № 9. С. 1599-1609. 43 3. Голубовский М.Д., Захаров И.К. Ген-мутатор, увеличивающий частоту ревертирования ряда нормальных по фенотипу нестабильных аллелей локуса singed у D. melanogaster // Генетика. 1979б. Т. 15. № 10. С. 1798-1808. 4. Захаров И.К., Голубовский М.Д. Влияние температуры и Y-хромосомы на проявление и частоту совместного мутирования двух нестабильных генов у Drosophila melanogaster//Генетика. 1980. Т. 16. ¹ 9. С. 1603-1612. 5. Golubovsky M.D., Zakharov I.K. Simultaneous reversions of two sex-linked unstable mutations //Drosophila Inform. Serv. 1980a. ¹ 55. P. 49-51. 6. Golubovsky M.D., Zakharov I.K. Unstable genes in natural Drosophila populations // In: Movable Genetic Elements. Abstracts XLV Cold Spring Harbor Symposium on Quantitative Biology, 1980b. CSH, New York. P. 15. 7. Голубовский М.Д., Захаров И.К. Инсерционный мутагенез в природных популяциях дрозофил // В кн.: Успехи теоретической и прикладной генетики. / Новосибирск: ИЦиГ СО АН СССР. 1982. С. 180-182. 8. Захаров И.К., Юрченко Н.Н. Генетическая нестабильность. Анализ супермутабильных аллелей гена singed у Drosophila melanogaster // Генетика. 1982. Т. 18. № 10. С. 1674-1682. 9. Захаров И.К., Юрченко Н.Н. Влияние замены участков Х-хромосомы на нестабильность в локусе singed в генеративных и соматических клетках у Drosophila melanogaster //Генетика. 1983а. Т. 19. № 7. С. 1137-1143. 10. Захаров И.К., Юрченко Н.Н. Транспозонный и цитоплазматический контроль мутирования локуса singed в природных популяциях Drosophila melanogaster // В кн.: Молекулярные механизмы генетических процессов. Тез. докл., Москва, 1983 / М.: Наука. 1983б. С. 27-28. 11. Захаров И.К. Роль Р- и М-цитотипов в контроле мутирования нестабильных аллелей гена singed у Drosophila melanogaster // Генетика. 1984а. Т. 20. № 1. С. 42-50. 12. Захаров И.К. Генетика природных популяций Drosophila melanogaster: колебание мутабильности и концентрации аллелей гена singed в природных популяциях // Генетика. 1984б. Т. 20. № 8. С. 1295-1304. 13. Захаров И.К., Голубовский М.Д. Серия нестабильных аллелей гена singed, выделенных из природных популяций Drosophila melanogaster: закономерности мутирования // Генетика. 1984а. Т. 20. № 7. С. 1117-1124. 14. Захаров И.К., Голубовский М.Д. Инсерционный мутагенез и генетическая структура природных популяций Drosophila melanogaster // В кн.: Макроэволюция. (1 Всесоюзн. конф. по пробл. эвол.) / М.: Наука. 1984б. С. 191-192. 15. Захаров И.К., Юрченко Н.Н. Совместное мутирование двух тесно сцепленных генов Х-хромосомы в супермутабильных линиях Drosophila melanogaster // Генетика. 1984. Т. 20. № 2. С. 266-273. 44 16. Юрченко Н.Н., Захаров И.К., Голубовский М.Д. Плеяды нестабильных аллелей гена singed у дрозофилы и их связь с транспозоном, маркированным видимой мутацией // Генетика. 1984. Т. 20. ¹ 6. С. 974-983. 17. Yurchenko N.N., Zakharov I.K., Golubovsky M.D. Unstable alleles of the singed locus in Drosophila melanogaster with reference to a transposon marked with a visible mutation //Mol. Gen. Genet. 1984. V. 194. P. 279-285. 18. Захаров И.К. Фенооблик природных популяций Drosophila melanogaster // В кн.: Фенетика популяций. (3 Всесоюзн. совещ.). Саратов, 1985 / Москва. 1985. С. 131. 19. Захаров И.К., Голубовский М.Д. Возвращение моды на мутацию yellow в природной популяции Drosophila melanogaster г. Умани // Генетика. 1985. Т. 21. № 8. С. 1298-1305. 20. Захаров И.К., Юрченко Н.Н. Генетика нестабильных аллелей гена singed у Drosophila melanogaster // В кн.: Молекулярные механизмы генетических процессов / М.: Наука. 1985. С. 5-12. 21. Юрченко Н.Н., Захаров И.К., Голубовский М.Д. Рекомбинация, опосредованная мобильным элементом у дрозофилы // Генетика. 1985а. Т. 21. № 7. С. 1125-1134. 22. Юрченко Н.Н., Захаров И.К., Голубовский М.Д. Влияние структуры гомологичного локуса на поведение транспозона у Drosophila melanogaster // Генетика. 1985б. Т. 21. № 8. С. 1290-1297. 23. Голубовский М.Д., Волошина М.А., Захаров И.К., Юрченко Н.Н. Взаимоотношение мобильных элементов P и mdg3 у Drosophila melanogaster: генетический аспект // Генетика. 1986. Т. 22. № 10. С. 2452-2458. 24. Голубовский М.Д., Захаров И.К., Соколова О.А. Анализ нестабильных аллелей гена yellow, выделенных из природной популяции дрозофил в период вспышки мутабильности // Генетика. 1987. Т. 23. № 9. С. 1595-1603. 25. Юрченко Н.Н., Захаров И.К. Возможная роль мобильных элементов в эволюции// В кн.: Проблемы макроэволюции / М.: Наука. 1988. С. 32-33. 26. Захаров И.К., Юрченко Н.Н. Мутационная изменчивость в природных популяциях Drosophila melanogaster // В кн.: Проблемы макроэволюции / М.: Наука. 1988. С. 56. 27. Захаров И.К., Юрченко Н.Н. О причинах мутационного процесса в природных популяциях Drosophila melanogaster // Известия СО АН CCCP, сер. Биол., вып. 2. 1989. С. 52. 28. Захаров И.К., Голубовский М.Д. Вспышки мутаций в природе и генетический анализ нестабильности в локусе yellow у Drosophila melanogaster // В кн.: Всесоюзн. совещ. по проблемам биологии и генетики дрозофилы. Одесса, 1989, Тез. докл. / Одесса. 1989. С. 36-37. 45 29. Захаров И.К., Юрченко Н.Н., Иванников А.В. Концентрация и спектр видимых мутаций в природных популяциях Drosophila melanogaster // В кн.: Всесоюзн. совещ. по проблемам биологии и генетики дрозофилы. Одесса, 1989, Тез. докл. / Одесса. 1989. С. 37. 30. Юрченко Н.Н., Захаров И.К. Модели рекомбинационных событий, опосредованных транспозоном у дрозофилы // В кн.: Молекулярные механизмы генетических процессов / М.: Наука. 1990. С. 46-55. 31. Захаров И.К., Иванников А.В. Генетический мониторинг природных популяций Drosophila melanogaster // В кн.: Экологическая генетика растений, животных, человека / Кишинев: Штиинца. 1991. С. 458. 32. Захаров И.К., Иванников А.И., Скибицкий Е.Э. и др. Генетическая изменчивость природных популяций Drosophila melanogaster // В кн.: 1 Всесоюзная конференция по генетике насекомых / М.: Наука. 1991. С. 44. 33. Golubovsky M.D., Sokolova K.B., Zakharov I.K. et al. Lethal alleles of l(2)gl antioncogene: wide distribution in nature and heterozygots advantage / In: 12th European Drosophila Res. Conf., Mainz, Late Abstract Book. 1991. P. 7 34. Иванников А.В., Захаров И.К. Пониженная MR-активность среди хромосом с летальной мутацией в природной популяции Drosophila melanogaster // Доклады Академии наук. 1992. Т. 324. № 2 . С. 436-438. 35. Захаров И.К., Баласов М.П., Беляева Е.С. и др. Мутации и мутагенез в популяциях Drosophila melanogaster Алтая. Оценка мутагенности продуктов питания // В кн.: Генетические эффекты антропогенных факторов среды / Новосибирск. ИЦиГ СО РАН. 1993. С. 44-61. 36. Захаров И.К., Скибицкий Е.Э. Динамика и свойства вспышек мутаций в природных популяциях Drosophila melanogaster // Генетика. 1994. Т. 30 (приложение). С. 54. 37. Иванников А.В., Голубовский М.Д., Коромыслов Ю.А., Захаров И.К. MRхромосомы в Евразийских популяциях Drosophila melanogaster // Генетика. 1995. Т. 31. № 2. С. 209-211. 38. Юрченко Н.Н., Захаров И.К. Мутабильная ХZ-хромосома, выделенная из природной популяции Drosophila melanogaster // Генетика. 1995. Т. 31. № 3. С. 422-426. 39. Вайсман Н.Я., Захаров И.К. Взаимодействие генов Mos и mwh, проявляющих свое действие в соматических клетках Drosophila melanogaster // Генетика. 1995а. Т. 31. № 3. С. 358-362. 40. Вайсман Н.Я., Захаров И.К. Генетический анализ мутантных хромосом 3 со сходным спектром фенотипического действия из географически удаленных популяций Drosophila melanogaster // Генетика. 1995б. Т. 31. № 7. С. 932-938. 46 41. Weisman N.Ya., Koryakov D.E., Zakharov I.K. New geographical allelic variants of the Drosophila melanogaster genes rotuned and roughened eye // Drosophila Inform. Serv. 1995. No. 76. P. 103-106. 42. Захаров И.К., Иванников А.В., Скибицкий Е.Э., Чурашов К.В., Юрченко Н.Н., Волошина М.А., Голубовский М.Д.. Генетические свойства аллелей генов Ххромосомы, выделенных из природных популяций Drosophila melanogaster в период вспышки мутаций // Доклады Академии наук. 1995. Т. 341. № 1. C. 126-129. 43. Захаров И.К., Скибицкий Е.Э. Генетика нестабильных аллелей генов Ххромосомы, выделенных в период вспышки уеllow-мутаций 1982-1991 годов в природной популяции Drosophila melanogaster Умани // Генетика. 1995. Т. 31. № 8. С. 1079-1084. 44. Юрченко Н.Н., Коряков Д.Е., Захаров И.К. Возникновение рецессивных летальных мутаций в производных от нестабильной XZ-хромосомы Drosophila melanogaster // Генетика. 1995. Т. 31. № 9. С. 1218-1224. 47 Подписано к печати 31.10.1995 Формат бумаги 60х90 1/16. Печ.л. 3, уч.изд.л. 2,1. Тираж 110 экз. Заказ 59. ___________________________________________________ Ротапринт Института цитологии и генетики СО РАН 630090, Новосибирск, пр. академика М.А. Лаврентьева, 10.