ГЕНЕРАТОРНЫЕ МОДЕЛИ УПРАВЛЕНИЯ ЭНЕРГЕТИЧЕСКИМ
реклама

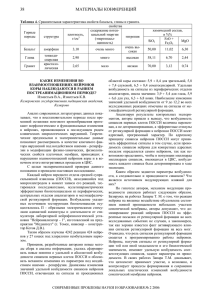
УДК 615.47 ГЕНЕРАТОРНЫЕ МОДЕЛИ УПРАВЛЕНИЯ ЭНЕРГЕТИЧЕСКИМ СОСТОЯНИЕМ ПРОЕКЦИОННЫХ ЗОН Р.А. Крупчатников В работе рассматриваются виды взаимодействия внутренних структур организма с поверхностными проекционными зонами с учетом импульсного характера управления органами со стороны ретикулярных нейронов Ключевые слова: генератор, импульсная активность, ретикулярные нейроны, проекционная зона Теоретической основой для использования прогностической и диагностической информации, снимаемой с проекционных зон человека, включая внемеридианные и меридианные биологически активные точки (БАТ), служит современная рефлексология, основу которой составляет учение о строении и функционировании многоуровневой рефлекторной системы, включающей терминальные, промежуточные и центральные рефлекторные кольца холинергического (парасимпатического) и андренергического (симпатического) вида. Морфологическую основу рефлексологии связывают с такими понятиями, как рецепторное поле, рефлексогенная зона, проекционные точки (зоны), акупунктурные точки, рефлекторные дуги и кольца и др. в нашей работе будем использовать определение этих понятий так, как это сформулировано в работе [1]. Многочисленными исследованиями было установлено, что проекционные зоны (ПЗ) и, в частности, биологически активные точки (БАТ) находятся на поверхности кожи человека, имеют связи с соответствующими внутренними органами. Точные анатомические пути прохождения информации от органов до их проекционных зон на совместном этапе развития медицинской науки установить не удалось, однако обобщение многочисленных результатов исследований отечественных и зарубежных ученых позволяет построить обобщенную информационную модель связи органа со «своими» рефлексогенными зонами. К центральной составляющей этой модели, учитывая современные Крупчатников Роман Анатольевич – ЮЗГУ, канд. с.-х. наук, доцент, тел. (4712) 58-70-98 анатомические и морфологические представления, будем относить андренергическую зону ретикулярной формации, выполняющую интегрирующую и активирующую функции. Как анатомическое образование ретикулярная формация (РФ) в центральной нервной системе занимает достаточной большой участок, начиная от спинного мозга до промежуточного. Спинальную зону РФ, с филогенетической точки зрения, относят к более древней в структурной организации РФ. В ней, как и в РФ головного мозга, выделяют три зоны: соматическую, парасимпатическую и симпатическую. Такое разделение позволяет осуществлять автономную активацию эффекторов соматического и парасимпатического вида на спинальном уровне. В свою очередь каждая из этих зон разбивается на подзоны, а последние на микрозоны. Микрозоны характеризуются тем, что их ретикулярные нейроны имеют примерно одинаковые пороги срабатывания. Микрозоны ретикулярной формации, состоящие в основе своей из ретикулярных нейронов (РН), ориентированных на «свои» органы, стимулируются сигналами Х * cj поступающими из афферентного канала связи. Каждый сигнал определенной модальности может запустить только свои группы РН. Условием возбуждения РН является превышение входным сигналом Х * некоторого порога Spн. cj Каждая модальность афферентных сигналов в микрозоне имеет свою группу РН с примерно одинаковыми порогами Spн [2]. Если интенсивность пришедшего сигнала больше пороговой, то соответствующие РН на один входной импульс генерируют до 100 и более квантов 1 импульсов своей активности в соответствии с выражением fрн=Kp fc, где fрн — частота генерации РН; fс — частота стимула из >>1 — афферентного канала; Кр коэффициент генерации. Считается, что нейроны РФ с различным значением Spн генерируют импульсы различной частоты в соответствии с выражением fрн=К/ Spн, то есть нейроны с низким порогом (менее чувствительные) генерируют импульсы с большой частотой. Причем высокочувствительные нейроны (с высоким порогом Sрн) способны вырабатывать некоторую фоновую частоту при заторможенных режимах (например, во сне). Стимуляция множества РН сигналами даже одной модальности приводит к генерации широкого спектра частот, поскольку стимулирующие сигналы могут иметь разную интенсивность (например, со зрительного анализатора), а тем более широкий спектр частот формируется сигналами разной модальности, обладающими различной энергетикой. Пороговая величина РН представляет собой управляемый переменный параметр, на который влияют сигналы нервной стимуляции и гормонов эндокринной системы. Гормональная стимуляция характеризуется длительным поддержанием Sрн на высоком уровне чувствительности. В целом ретикулярная формация, как совокупность РН, может рассматриваться как многоканальный генератор, излучающий спонтанные импульсы, формируемые под влиянием: внешних воздействий (Z*ст); афферентных сигналов от интерорецепторов экстерорецепторов с органов Хсi; поверхности тела Yэi; гормональных и кортикальных сигналов из управляющих зон коры головного мозга Yмi.; сигналов, поступающих от сопряженных (функционально-зависимых) с контролируемыми органами систем Ycoi. Учитывая, что контроль и управление органами осуществляется посредством импульсной активности рецепторов и ретикулярных нейронов, одну из возможных информационных моделей можно построить как генераторную схему (рис. 1), в которой орган Оi; будем рассматривать как управляемый многоканальный генератор, вырабатывающий импульсную последовательность Xci интенсивностью (амплитудой) bio с частотой, определяемой модальностью передающих нейронов и управляющей последовательностью Хпi со стороны парасимпатических колец. Эта импульсная последовательность с частотой fo и интенсивностью bio поступает на вход ретикулярных нейронов, которые схемотехнически можно представить в виде последовательно соединенных порогового элемента ПЭ1с и умножителя частоты (УЧ1с). Учитывая, что симпатическая микрозона ретикулярной формации МРФсi., реализует связи с другими структурами ВНС, включая органные проекционные зоны ПЗij, в схему МРФ включены: функциональный узел сопряжения ФУСс; схема формирования обратной связи замыкающая центральное СФОСсi, рефлекторное кольцо симпатического типа, формирующее порог срабатывания Sпнi эффекторной клетки ЭК органа Оi; второй пороговый элемент ПЭ2с и второй умножитель частоты УЧ2с, обеспечивающие связь МРФсi с соответствующей проекционной зоной ПЗij и настроенные на частоту работы симпатического канала, обеспечивающего эффекторные клетки ЭК проекционной зоны; вторая схема формирования обратной связи СФОСс2, замыкающая центральное рефлекторное кольцо симпатического типа от МРФсi и ПЗij. Микрозона МРФсi через собственные каналы связи и каналы связи РФс активно взаимодействует со своими терминальными рефлекторными кольцами (ТКс), центральными нервными структурами (ЦСс), сопряженно работающими органами (ОСс) и гуморальной системой (Гс). Кроме этого, существует связь Fкп с соответствующими микрозонами парасимпатической ретикулярной формации МРФпi, которая (рисунок) имеет структуру МРФсi. В эту схему введена также самостоятельная система управления проекционной зоной СУПЗ. Известно, что пороги Sрн1 и Spн2 пороговых элементов есть динамические величины, определяемые совокупностью корректирующих сигналов, поступающих со стороны взаимодействующих с МРФсi элементов, то есть S рн1 = f1 ( Fкт , Fкц , Fко , Fкг , Fкп ) , S рн 2 = f 2 ( Fкт , Fкц , Fко , Fкг , Fкп ) , Учитывая, что спектр частот, вырабатываемых рецепторами даже одной модальности, может быть достаточно широк, и он должен восприниматься соответствующими ретикулярными 2 нейронами для его трансформации и передачи в другие цепи всей системы управления, введем понятие рабочего диапазона αip, воспринимаемых МРФi, настроенными на работу с органом Оi рецепторами различных модальностей р. То есть f нip ≤ α ip ≤ f вip , где f нip и f вip – соответственно нижние и верхние частотные границы, в рамках которых срабатывает пороговый элемент ретикулярного нейрона. В соответствии с введенными обозначениями первый умножитель частоты УЧ1 МРФci вырабатывает последовательность импульсов с частотой f1, интенсивностью bi1 и длительностью tнi в соответствии с выражением ⎧X ci , если (bio ≥ Sрн1 ) & ([f 0 ± ∆f 0 ] ∈ α ip ); X*ci = ⎨ ⎩0, в противном случае, Учитывая, что контроль и управление органами осуществляется посредством импульсной активности рецепторов и ретикулярных нейронов, одну из возможных информационных моделей можно построить как генераторную схему (рисунок), в которой орган Оi; будем рассматривать как управляемый многоканальный генератор, вырабатывающий импульсную последовательность Xci интенсивностью (амплитудой) bio с частотой, определяемой модальностью передающих нейронов и управляющей последовательностью Xпi со стороны парасимпатических колец. Эта импульсная последовательность с частотой fо и интенсивностью bio поступает на вход ретикулярных нейронов, которые схемотехнически можно представить в виде последовательно соединенных порогового элемента ПЭ1с и умножителя частоты (УЧ1с). Учитывая, что симпатическая микрозона ретикулярной формации МРФсi, реализует связи с другими структурами ВНС, включая органные проекционные зоны ПЗij, в схему МРФсi включены: функциональный узел сопряжения ФУСс; схема формирования обратной связи замыкающая центральное СФОСc1, рефлекторное кольцо симпатического типа, формирующее порог срабатывания Sпн1 эффекторной клетки ЭК органа Оi; второй пороговый элемент ПЭ2с и второй умножитель частоты УЧ2с, обеспечивающие связь МРФсi с соответствующей проекционной зоной ПЗij и настроенные на частоту работы симпатического канала, обеспечивающего эффекторные клетки ЭК проекционной зоны; вторая схема формирования обратной связи СФОСс2 замыкающая центральное рефлекторное кольцо симпатического типа от МРФсi и ПЗij. Микрозона МРФсi через собственные каналы связи и каналы связи РФс активно взаимодействует со своими терминальными рефлекторными кольцами (ТКс), центральными нервными структурами (ЦСс), сопряженно работающими органами (ОСс) и гуморальной системой (Гс). Кроме этого, существует связь Fкп с соответствующими микрозонами парасимпатической ретикулярной формации МРФпi, которая (рисунок) имеет структуру МРФсi. В эту схему введена также самостоятельная система управления проекционной зоной СУПЗ. Известно, что пороги Sрн1 и Sрн2 пороговых элементов есть динамические величины, определяемые совокупностью корректирующих сигналов, поступающих со стороны взаимодействующих с МРФсi элементов, то есть Sрн1=f1(Fкт,Fкц, Fко,Fкг,Fкп), Sрн2=f2(Fкт,Fкц, Fко,Fкг,Fкп), Учитывая, что спектр частот, вырабатываемых рецепторами даже одной модальности, может быть достаточно широк, и он должен восприниматься соответствующими ретикулярными нейронами для его трансформации и передачи в другие цепи всей системы управления, введем понятие рабочего диапазона частот αip , воспринимаемых МРФi, настроенными на работу с органом Oi рецепторами различных модальностей р. То есть fнip≤αip≤fвip, где fнip и fвip - соответственно нижние и верхние частотные границы, в рамках которых срабатывает пороговый элемент ретикулярного нейрона. В соответствии с введенными обозначениями первый умножитель частоты УЧ1 МРФсi вырабатывает последовательность импульсов с частотой f1, интенсивностью bi1 и длительностью tнi в соответствии с выражением ⎧⎪ X ci , если (bio ≥ S рн1 ) & ([ f 0 ± ∆f 0 ]∈ α ip ); X ci* = ⎨ ⎪⎩0, в противном случае, где ∆fo - девиация частоты, вырабатываемой симпатическими рецепторами, ⎧⎪bij , если τ / f1 ≤ t ≤ (τ / f1 + tн1); X ci = ⎨ ⎪⎩0, в противном случае, 3 где τ=1,2,..n - номер отсчетов импульсной последовательности. Эти сигналы анализируются ФУСс и передаются: в параллельно работающие структуры; на выход в соответствующие проекционные зоны ПЗij и через СФОСС1 используются для управления работой эффекторных клеток органа Oi , поддерживая вместе с сигналами Хпi, «равновесное» функционирование. Учитывая, что точное анатомическое строение канала передачи данных с входных РН микрозоны на выходные цепи, связанные с ПЗij, неизвестны, нами было сделано предположение, что в промежуточных cтруктурах возможно наличие нескольких пороговых элементов со своими умножителями частоты, что в обобщенном варианте представлено блоками ПЭ2с и УЧ2с. Такое предположение не нарушает общность рассуждений, поскольку если для каких-либо ПЗij идет «прямая» передача сигнала с УЧ1 в соответствующие проекционные зоны, то коэффициент передачи канала ПЭ2с – УЧ2с принимается равным единице. Для выходной части МРФсi по аналогии с входной частью имеем * ⎪⎧Yci , если (bio ≥ S рн 2 ) & ([ f1 ± ∆f1 ]∈ α ip ); Yci = ⎨ ⎪⎩0, в противном случае, где α ip* - диапазон «допустимых» частот для выходной группы ретикулярных нейронов, на который настроены эффекторные клетки проекционной зоны ПЗij X сi , X пi , Генераторная схема управления проекционными зонами ⎧⎪bi 2 , если τ / f 2 ≤ t ≤ (τ / f 2 + t н2 ); Yci = ⎨ ⎪⎩0, в противном случае, Информация о состоянии проекционных зон через системы обратной связи передается в ФУСс и ФУСп и через них частично влияет на режимы работы соответствующих пороговых элементов. Кроме этого, состояние ПЗij регулируется не только по информации со стороны Oi , но и со стороны других структур, например, центральных, терминальных и ганглионарных колец различного типа и уровня, что отражено обобщенным сигналом Fкпз, проходящим через систему управления проекционной зоны СУПЗ. Исходя из вышесказанного, обобщенный выходной сигнал можно представить как сумму сигналов: J 0 = Z ПЗ + ∑К ПЗr ⋅ YПЗ* ij r 4 где Zпз - эквивалентный сигнал с выхода СУПЗ; К ПЗ - коэффициент преобразования r импульсного потока в выходной сигнал природы r. Под природой r будем понимать различные типы измерительной информации (электродвижущая сила r=1, сопротивление r=2; геометрическая структура радужки глаза r=3; интенсивность потока инфракрасного излучения r=4; генерация информационной составляющей в КВЧ диапазоне r=5 и так далее). Сигнал YПЗ* ij определяется системой уравнений вида: YПЗ* = Yci ⋅ К ПСij , ij Здесь К ПСij - коэффициенты передачи сигналов с эффекторных клеток ПЗij до измерительной аппаратуры. В нормальном «равновесном» состоянии обмен информацией по всем каналам идет в некотором установившемся режиме, формируя сигнал J0 в некотором заданном интервале регулирования Jн ≤ J0≤ Jв,. Исходя из схемы (рисунок), выход за пределы этих границ может происходить при нарушении равновесия условий работы органов Оi„ когда исходный генератор переходит на другой режим работы под воздействием сигналов Fкт, Fкн, Fко, Fкг, Fкп и Fкпз. To, что канал передачи данных от Оi до эффекторных клеток «своей» проекционной зоны ПЗij настроен на свою полосу частот, позволяет сделать вывод, что доля сигнала от органа, вышедшего из состояния исходного равновесия, в интегральном сигнале J0 будет достаточно велика, что и создает предпосылки для решения соответствующих диагностических задач. Литература 1. Ананин, В. Ф. Рефлексология (теория и методы) [Текст]: монография / В. Ф. Ананин. М.: Издво РУДН и «Биомединформ», 1992. 168 с. 2. Кореневский Н.А., Крупчатников Р.А., Серегин С.П. Теоретические основы биофизики акупунктуры с приложениями в биологии, медицине и экологии на основе нечетких моделей [Текст]: монография / Курск.гос.тех.ун-т.-Курск: ОАО «ИПП «Курс», 2010.- 521с. Юго-Западный государственный университет (г. Курск) GENERATING MODELS OF MANAGEMENT OF THE POWER CONDITION OF PROJECTIVE ZONES R.A. Krupchatnikov In work kinds of interaction of internal structures of an organism with superficial projective zones taking into account pulse character of management of bodies from outside ретикулярных neurons are considered Key words: the generator, pulse activity, ретикулярные neurons, a projective zone 5