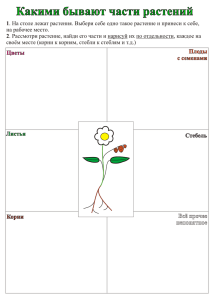
Лекция 1. Структура растительной клетки
реклама

Лекция 1. Структура растительной клетки Методы изучения растительной клетки Световая микроскопия Электронная микроскопия Метод замораживания-скалывания Дифференциальное центрифугирование Метод культуры клеток Клетка является основной структурной и функциональной единицей живых организмов. Клетки эмбриональных (неспециализированных) тканей животных и растений в общем плане строения очень сходны. Именно это обстоятельство в свое время явилось причиной для появления и развития клеточной теории. Морфологические различия проявляются уже в дифференцированных клетках специализированных тканей растений и животных. Особенности строения растительной клетки, как и растения в целом, связаны с образом жизни и способом питания. Большинство растений ведет относительно неподвижный (прикрепленный) образ жизни. Специфика питания растений состоит в том, что вода и питательные вещества: органические и неорганические, находятся вокруг в рассеянном виде и растению приходится их поглощать путем диффузии. Кроме того, зеленые растения на свету осуществляют автотрофный способ питания. Благодаря этому, эволюционно сложились некоторые специфические особенности строения и роста растительных клеток. К ним относятся: прочная полисахаридная клеточная стенка, окружающая клетку и составляющая жесткий каркас; пластидная система, возникшая в связи с автотрофным типом питания; вакуолярная система, которая в зрелых клетках обычно представлена крупной центральной вакуолью, занимающей до 95% объема клетки и играющей важную роль в поддержании тургорного давления; особый тип роста клеток путем растяжения (за счет увеличения объема вакуоли); тотипотентность, то есть возможность регенерации полного растения из дифференцированной растительной клетки; есть еще одна деталь, отличающая растительные клетки от клеток животных: у растений при делении клеток не выражены центриоли. Строение клетки в самом общем виде известно вам еще из курса общей биологии и при подготовке к вступительным экзаменам вы достаточно хорошо штудировали эту тему. Эта тема в разных аспектах рассматривается и в соответствующих университетских курсах (например, зоология беспозвоночных, низшие растения). Кроме того, более детальное знакомство с клеткой на высоком уровне предстоит в курсе "цитология". Нам же важно акцентировать внимание на специфических особенностях строения растительной клетки, причем преимущественно клетки высшего растения. При самом поверхностном рассмотрении структуры типичной растительной клетки в ее составе обнаруживаются три основных компонента: (1) клеточная стенка, (2) вакуоль, занимающая в зрелых клетках центральное положение и заполняющая практически весь их объем и (3) протопласт, оттесняемый вакуолью к периферии в виде постенного слоя. Именно эти компоненты обнаруживаются на малом увеличении светового микроскопа. Причем клеточная оболочка и вакуоль являются продуктами жизнедеятельности протопласта. Живое тело клетки ≈ протопласт состоит из органоидов, погруженных в гиалоплазму. К организмам клетки относятся: ядро, пластиды, митохондрии, диктиосомы, эндоплазматический ретикулум, микротельца и др. Гиалоплазма с органеллами за вычетом ядра составляет цитоплазму клетки. Для выражения размеров субклеточных структур используются определенные меры длины: микрометр и нанометр. Микрометр в системе единиц измерения СИ величина, равная 10-6 м. Говоря другими словами, микрометр (аббревиатура мкм) составляет 1/1000000 долю метра и 1/1000 долю миллиметра. 1 мкм = 10-6 м. Старое название этой меры микрон. Нанометр в той же системе представляет миллионную долю миллиметра 1 нм = 10-9 м и тысячную долю микрометра. Размеры и форма растительных клеток варьируются в широком диапазоне. В типичном случае размеры клеток высшего растения колеблются в пределах 10 - 300 мкм. Правда, встречаются клетки - гиганты, например, клетки сочной мякоти плодов цитрусовых составляют в поперечнике несколько миллиметров или чрезвычайно длинные лубяные волокна у крапивы достигают 80 мм длины при микроскопической толщине. По форме различают изодиаметрические клетки, у которых линейные размеры во всех направлениях равны или отличаются незначительно (то есть длина, ширина и высота этих клеток сопоставимы). Такие клетки называют паренхимными (паренхима). Сильно вытянутые клетки, у которых длина во много раз (иногда в сотни и тысячи) превышает высоту и ширину, называют прозенхимными (прозенхима). Методы изучения растительной клетки Для изучения клеток разработано и применяется множество методов, возможности которых определяют уровень наших знаний в этой области. Успехи в изучении биологии клетки, включая наиболее выдающиеся достижения последних лет, как правило, связаны с применением новых методов. Поэтому для более полного понимания клеточной биологии необходимо иметь хотя бы некоторое представление о соответствующих методах исследования клетки. Световая микроскопия Самым древним и, вместе с тем, наиболее распространенным методом изучения клетки является микроскопия. Можно сказать, что и начало изучения клетки было положено изобретением светового оптического микроскопа. Невооруженный человеческий глаз имеет разрешающую способность около 1/10 мм. Это означает, что если вы смотрите на две линии, которые находятся друг от друга на расстоянии меньше 0,1 мм, они сливаются в одну. Чтобы различить структуры, расположенные более тесно, применяют оптические приборы, например, микроскоп. Но возможности светового микроскопа не безграничны. Предел разрешения светового микроскопа задается длиной световой волны, то есть оптический микроскоп может быть использован только для изучения таких структур, минимальные размеры которых сопоставимы с длиной волны светового излучения. Лучший световой микроскоп имеет разрешающую способность около 0.2 мкм (или 200 нм), то есть примерно в 500 раз улучшает человеческий глаз. Теоретически построить световой микроскоп с большим разрешением невозможно. Многие компоненты клетки близки по своей оптической плотности и без специальной обработки практически не видны в обычный световой микроскоп. Для того, чтобы сделать их видимыми, используют различные красители, обладающие определенной избирательностью. В начале XIX в. Возникла потребность в красителях для окрашивания текстильных тканей, что в свою очередь вызвало ускоренное развитие органической химии. Оказалось, что некоторые из этих красителей окрашивают и биологические ткани и, что было уж совсем неожиданно, часто предпочтительно связываются с определенными компонентами клетки. Использование таких избирательных красителей дает возможность более тонко исследовать внутреннее строение клетки. Приведем лишь несколько примеров: краситель гематоксилин окрашивает некоторые компоненты ядра в синий или фиолетовый цвет; после обработки последовательно флороглюцином и затем соляной кислотой одревесневшие оболочки клеток становятся вишнево красными; краситель судан III окращивает опробковевшие клеточные оболочки в розовый цвет; слабый раствор йода в йодистом калии окрашивает крахмальные зерна в синий цвет. Для проведения микроскопических исследований большую часть тканей перед окраской фиксируют. После фиксации клетки становятся проницаемыми для красителей, а структура клетки стабилизируется. Одним из наиболее распространенных фиксаторов в ботанике является этиловый спирт. Фиксация и окрашивание не единственные процедуры, используемые для приготовления препаратов. Толщина большинства тканей слишком велика, чтобы их сразу можно было наблюдать при высоком разрешении. Поэтому выполняют тонкие срезы на микротоме. В этом приборе использован принцип хлеборезки. Для растительных тканей изготавливают чуть более толстые срезы, чем для животных, поскольку клетки растений обычно крупнее. Толщина срезов растительных тканей для световой микроскопии около 10 мкм - 20 мкм. Некоторые ткани слишком мягкие, чтобы из них сразу же можно было получить срезы. Поэтому после фиксации их заливают в расплавленный парафин или специальную смолу, которые пропитывают всю ткань. После охлаждения образуется твердый блок, который затем режется на микротоме. Правда, для растительных тканей заливка применяется значительно реже, чем для животных. Это объясняется тем, что растительные клетки имеют прочные клеточные стенки, составляющие каркас ткани. Особенно прочны одревесневшие оболочки. Однако заливка может нарушить структуру клетки, поэтому применяют еще и другой метод, где эта опасность уменьшена ≈ быстрое замораживание. Здесь можно обойтись без фиксации и заливки. Замороженную ткань режут на специальном микротоме (криотоме). Замороженные срезы, приготовленные таким способом, имеют явное преимущество, поскольку в них лучше сохраняются особенности естественной структуры. Однако их труднее готовить, а присутствие кристаллов льда все же нарушает некоторые детали. Микроскопистов всегда беспокоила возможность потери и искажения некоторых компонентов клетки в процессе фиксации и окраски. Поэтому полученные результаты проверяют другими методами. Весьма заманчивой представлялась возможность исследовать под микроскопом живые клетки, но так, чтобы более отчетливо проявились детали их строения. Такую возможность дают особые оптические системы: фазово-контрастный и интерференционный микроскопы. Хорошо известно, что световые волны, подобно волнам воды, могут интерферировать друг с другом, увеличивая или уменьшая амплитуду результирующих волн. В обычном микроскопе, проходя через отдельные компоненты клетки, световые волны меняют свою фазу, хотя человеческий глаз этих различий не улавливает. Но за счет интерференции можно преобразовать волны, и тогда разные компоненты клетки можно отличить друг от друга под микроскопом, не прибегая к окрашиванию. В этих микроскопах используют 2 пучка световых волн, которые взаимодействуют (налагаются) друг на друга, усиливая или уменьшая амплитуду волн, поступающих в глаз от разных компонентов клетки. Электронная микроскопия Возможности светового микроскопа, как уже было сказано, ограничиваются длиной волны видимого света. Его максимальная разрешающая способность составляет примерно 0.2 мкм. Большой шаг вперед был сделан в микроскопии в 20-х годах нашего века, когда было обнаружено, что соответствующим образом подобранные электромагнитные поля можно использовать подобно линзам для фокусирования пучков электронов. Длина волны электрона значительно меньше, чет длина волны видимого света, и если вместо света использовать электроны, то предел разрешения микроскопа может быть заметно снижен. На основе всего этого был создан микроскоп, в котором вместо света используется пучок электронов. Первый электронный микроскоп сконструировали в 1931 г. Кнолл и Руска в Германии. Прошло, однако, много лет, прежде чем появилась возможность изучать при помощи этого микроскопа срезы тканей. Лишь в 50-е годы были разработаны методы изготовления срезов, обладающих необходимыми качествами. С этого времени началась новая эра микроскопии, и в науку буквально хлынул поток информации о тонком строении клеток (ультраструктуре клеток). Сложности электронной микроскопии состоят в том, что для исследования биологических образцов необходима специальная обработка препаратов. Первая трудность заключается в том, что электроны обладают очень ограниченной проникающей способностью, поэтому следует изготавливать ультратонкие срезы, толщиной 50 - 100 нм. Для того, чтобы получить столь тонкие срезы, ткани сперва пропитывают смолой: смола полимеризуется и формирует твердый пластмассовый блок. Затем с помощью острого стеклянного или алмазного ножа срезы нарезают на специальном микротоме. Есть еще одна трудность: при прохождении через биологическую ткань электронов не получается контрастного изображения. Для того, чтобы получить контраст, тонкие срезы биологических образцов пропитывают солями тяжелых металлов. Существует два основных типа электронных микроскопов. В трансмиссионном (просвечивающем) микроскопе пучок электронов, проходя сквозь специально подготовленный образец, оставляет его изображение на экране. Разрешающая способность современного трансмиссионного электронного микроскопа почти в 400 раз больше светового. Эти микроскопы имеют разрешающую способность около 0,5 нм (для сравнения: диаметр атома водорода около 0,1 нм). Несмотря на столь высокое разрешение, просвечивающие электронные микроскопы имеют крупные недостатки: Первое: приходится работать с фиксированными материалами; изображение на экране получается двумерным (плоским); при обработке тяжелыми металлами разрушаются и видоизменяются некоторые клеточные структуры. Трехмерное (объемное) изображение получают с помощью сканирующего электронного микроскопа (ЭМ). Здесь луч не проходит через образец, а отражается от его поверхности. Исследуемый образец фиксируют и высушивают, после чего покрывают тонким слоем металла ≈ операция называется оттенением (образец оттеняют). В сканирующем ЭМ сфокусированный электронный пучок направляется на образец (образец сканируют). В результате металлическая поверхность образца испускает вторичные электроны слабой энергии. Они регистрируются и преобразуются в изображение на телевизионном экране. Максимальное разрешение сканирующего микроскопа невелико, около 10 нм, но зато изображение получается объемным. Метод замораживания-скалывания Принципиально новые возможности электронной микроскопии открылись сравнительно недавно, после разработки метода "замораживания - скалывания". С помощью этого метода исследуются тончайшие детали строения клетки, при этом получается объемное изображение в трансмиссионном электронном микроскопе. При обычном замораживании в клетках образуются кристаллики льда, которые заметно искажают их структуру. Во избежание этого клетки замораживают очень быстро при температуре жидкого азота (- 196 С). При таком мгновенном замораживании кристаллы льда не успевают образоваться, и клетка не испытывает деформаций. Замороженный блок раскалывают лезвием ножа (отсюда и название метода). Затем, обычно в вакуумной камере, избыток льда удаляют возгонкой. Эта операция называется травлением. После травления более резко обозначается рельеф в плоскости скола. Полученный образец оттеняется, то есть на поверхность образца напыляется тонкий слой тяжелых металлов. Однако весь фокус состоит в том, что напыление производится под углом к поверхности образца. Это очень важный момент. Появляется эффект тени, изображение выглядит объемным. В трансмиссионном микроскопе электронный луч способен проникнуть только через очень тонкие срезы. Обычная толщина оттененных образцов чрезмерно велика, поэтому органическую материю, подстилающую слой металла, необходимо растворить. В результате остается тонкая металлическая реплика (или отпечаток) с поверхности образца. Реплику и используют в трансмиссионном микроскопе. Этот метод предоставил, например, уникальную возможность наблюдать внутреннее строение мембран клетки. Дифференциальное центрифугирование Помимо микроскопии, другим основным и широко распространенным методом изучения клеток является дифференциальное центрифугирование или фракционирование. Принцип метода состоит в том, что при центрифугировании развивается центробежная сила, под воздействием которой взвешенные частицы оседают на дно центрифужной пробирки. После того, как в начале 40-х годов начали использовать ультрацентрифугу, разделение клеточных компонентов стало вполне реальным. Прежде, чем подвергнуть клетки центрифугированию, их необходимо разрушить разрушить жесткий каркас клеточных оболочек. Для этого используют различные методы: ультразвуковую вибрацию, продавливание через маленькие отверстия или самое обычное измельчение растительных тканей пестиком в фарфоровой ступе. При осторожном применении методов разрушения можно сохранить некоторые органеллы целыми. При высокоскоростном центрифугировании крупные компоненты клетки (например, ядра) быстро оседают (седиментируют), при относительно низких скоростях и образуют осадок на дне центрифужной пробирки. При более высоких скоростях в осадок выпадают более мелкие компоненты, такие как хлоропласты и митохондрии. То есть при центрифугировании компоненты клетки распадаются на фракции: крупные и мелкие, поэтому второе название метода ≈ фракционирование. При этом, чем выше скорость и длительность цетрифугирования, тем мельче полученная фракция. Скорость седиментации (осаждения) компонентов выражается с помощью коэффициента седиментации, обозначаемого S. Этапы дифференциального центрифугирования: низкая скорость (ядра, цитоскелет), средняя скорость (хлоропласты), высокая скорость (митохондрии, ризосомы, микротельца), очень высокая скорость (рибосомы). Фракционированные клеточные экстракты, называемые также бесклеточными системами, широко используются для изучения внутриклеточных процессов. Только работая с бесклеточными экстрактами, можно установить детальный молекулярный механизм биологических процессов. Так, использование именно этого метода принесло триумфальный успех в изучении биосинтеза белка. Ну и вообще, чистые фракции внутриклеточных структур можно подвергать любым видам анализа. Метод культуры клеток Клетки животных, выделенные в культуру (то есть помещенные на питательную среду), погибают после определенного числа делений, поэтому считаются трудным и неудобным объектом для культивирования. Другое дело клетки растений, способные делиться неограниченное число раз. Метод культуры клеток облегчает изучение механизмов клеточной дифференциации у растений. На питательной среде клетки растений образуют однородную недифференцированную клеточную массу ≈ каллус. Каллус обрабатывают гормонами. Под влиянием гормонов клетки каллуса могут давать начало разным органам. Лекция 2. Клеточная оболочка Химический состав Одревеснение клеточных оболочек Опробковение и кутинизация клеточных оболочек Ослизнение клеточных оболочек Минерализация клеточных оболочек Структурная организация клеточной оболочки Формирование и рост клеточных оболочек Плазмодесмы Поры Функции и эволюция клеточной стенки Наличие клеточной оболочки более чем все другие признаки отличает растения от животных. Например, простейшие жгутиковые, покрытые оболочкой, причисляют к примитивным растениям (Protophyta), а голые жгутиковые к примитивным животным (Protozoa). У низших растений голыми являются лишь репродуктивные клетки, а клетки, составляющие вегетативное тело, имеют клеточные стенки. У высших растений клеточной стенкой обладают даже гаметы: как яйцеклетка, так и пыльцевая трубка. Клеточная оболочка окружает собственно клетку со всех сторон и служит связующим звеном между ней и соседними клетками, обеспечивая, таким образом, единство и целостность всего растительного организма. В жестких оболочках растительных клеток образуются каналы, в которых располагаются тончайшие тяжи цитоплазмы - плазмодесмы. Благодаря этому, осуществляются межклеточные взаимодействия. Иными словами, у растений клеточные оболочки призваны обеспечивать те функции, которые у животных выполняют скелет, кожа и система кровообращения (т.е. опорную, защитную и транспортную.) Не удивительно поэтому, что в ходе эволюции у растений возникли весьма разнообразные по структуре и химическому составу типы клеточных стенок. Собственно говоря, растительные клетки во многом различают и классифицируют именно по форме и природе клеточных стенок. Оболочка, как правило, бесцветна и прозрачна. Она легко пропускает солнечный свет. Оболочки соседних клеток как бы сцементированы межклеточными веществами, образующими так называемую срединную пластинку. Вследствие этого соседние клетки оказываются отделёнными друг от друга стенкой, образованной двумя оболочками и срединной пластинкой. Это и даёт основание называть оболочку также клеточной стенкой. Клеточная стенка растительных клеток состоит, главным образом, из полисахаридов. Оболочку эмбриональных тканей и клеток, растущих растяжением, называют первичной. В этот период оболочка достаточно эластична. После прекращения роста клетки изнутри на первичную клеточную стенку начинают откладываться новые слои и образуется вторичная клеточная стенка, придающая клетке жёсткость и прочность. Химический состав Все компоненты, входящие в состав клеточной стенки, можно разделить на 4 группы: Структурные компоненты, представленные целлюлозой у большинства автотрофных растений, хитином (грибы), глюканом (дрожжи), манналом и ксиланом (водоросли). Компоненты матрикса, т.е. основного вещества, наполнителя оболочки - гемицеллюлозы, белки, липиды. Компоненты, инкрустирующие клеточную стенку, (т.е. откладывающиеся и выстилающие её изнутри) - лигнин и суберин. Компоненты, адкрустирующие стенку, т.е. откладывающиеся на её поверхности, - кутин, воск. Основной структурный компонент оболочки - целлюлоза представлена длинными неразветвленными полимерными молекулами, состоящими из 1000-11000 остатков -D глюкозы, соединённых между собой гликозидными связями. Наличие гликозидных связей создаёт возможность образования поперечных стивок. Благодаря этому, длинные и тонкие молекулы целлюлозы объединяются в элементарные фибриллы или мицеллы. Каждая мицелла состоит из 60-100 параллельно расположенных цепей целлюлозы. Мицеллы сотнями группируются в мицеллярные ряды и составляют микрофибриллы диаметром 10-25 нм. Целлюлоза обладает кристаллическими свойствами благодаря упорядоченному расположению мицелл в микрофибриллах. Микрофибриллы, в свою очередь, перевиваются между собой как пряди в канате и объединяются в макрофибриллы. Макрофибриллы имеют толщину около 0,5 мкм и могут достигать в длину 4 мкм. Они так же прочны, как равная им по толщине стальная проволока. Целлюлоза относится к индиферентным веществам: она не обладает ни кислыми, ни щелочными свойствами. По отношению к тепловым воздействиям она достаточно стойка и может быть нагрета без разложения до температуры 200 градусов. Многие из важных свойств целлюлозы обусловлены её высокой стойкостью отношению к ферментам и химическим реагентам. Она не растворима в воде, в спирте, в эфире и в других нейтральных растворителях; не растворяется и в кислотах и щелочах. Если целлюлоза и растворяется при известных условиях под действием и щелочей, то это растворение сопровождается сильным разрушением её организации. Без разрушения мицелл целлюлоза может быть растворена в реактиве Швейцера (аммиачном растворе окиси меди), при этом целлюлоза превращается в гель с беспорядочным расположением молекул. Такой гель более гигроскопичен и легче окрашивается, чем природная целлюлоза. При кратковременной и осторожной обработке целлюлозы серной кислотой образуется амилоид, представляющий по некоторым свойствам нечто промежуточное между клетчаткой и крахмалом: по консистенции амилоид подобен клейстеру. Приготовление "пергаментной бумаги", не пропускающей масла, основано именно на этом принципе. Листы бумаги погружаются на очень короткое время в серную кислоту, затем отмываются и высушиваются, при этом тонкий верхний слой бумаги превращается в непромокаемый для жиров амилоид. Действием азотной кислоты могут быть получены нитроцеллюлозы. Нитроцеллюлозы находят обширное применение в технике: они используются для изготовления шёлка, пороха, лаков, красок пластмасс. Целлюлоза, пожалуй, самый распространенный вид органических макромолекул на Земле. Она составляет основу питания многих гетеротрофных организмов. Целлюлоза разрушается действием определенных ферментов, содержащихся в теле некоторых низших растений, грибов, животных. Например, грибы - домовой гриб и трутовики, используя клетчатку живых деревьев или построек, разрушают целлюлозу. Термиты способны переваривать целлюлозу, благодаря наличию в их пищеварительном тракте простейших, энзимы которых перерабатывают клетчатку в вещества, усваиваемые термитами. В пищеварительном аппарате крупных травоядных животных содержится целая микрофлора организмов, подготавливающих клетчатку к усвоению. Микрофибриллы оболочки погружены в амморфный пластичный гель - матрикс. Матрикс является наполнителем оболочки. В состав матрикса входят полисахариды, называемые гемицеллюлозами и пектиновыми веществами. На долю гемицеллюлоз приходится около 3040 % сухого веса клетчатых стенок. По строению гемицеллюлозы напоминают целлюлозу и крахмал (это тоже полимеры); их цепи состоят из остатков гексоз либо пентоз, связанных кислородными мостиками. Но если молекулы целлюлозы насчитывают в своём составе от 1000 до 10000 мономеров, то цепочки гемицеллюлоз состоят из 150-300 молекул мономеров. Они значительно короче. Именно поэтому гемицеллюлозы нередко называют полуклетчатками. Основными гексозами гемицеллюлоз являются D-глюкоза и D-галактоза; пентозами - L-ксилоза и L-арабиноза. По отношению и химическим агентам гемицеллюлозы гораздо менее стойки, чем целлюлоза: они растворяются в слабых щелочах без подогревания; гидролизуются с образованием сахаров в слабых растворах кислот; растворяются полуклетчатки и в глицерине при температуре 300 градусов. Гемицеллюлозы в теле растений играют: механическую роль, участвуя наряду с целлюлозой и другими веществами в построении клетчатых стенок. роль запасных веществ, отлагающихся, а затем расходующихся. При этом функцию запасного материала несут преимущественно гексозы; а гемицеллюлозы с механической функцией обычно состоят из пектоз. В качестве запасных питательных веществ гемицеллюлозы отлагаются также в семенах многих растений, особенно у однодольных, например, в клеточных стенках эндосперма многих пальм (в частности финиковой пальмы). Пектиновые вещества имеют довольно сложный химический состав и строение. Химики определяют их, как кальциевые-магниевые соли полимерной -D галактуроновой кислоты, соединённые -1,4-связями, т.е. мономером является галактуроновая кислота. Характерная особенность: пектиновые вещества сильно набухают в воде, а некоторые даже в ней растворяются. Легко они разрушаются и под действием щелочей и кислот. Все клеточные стенки на ранней стадии развития почти нацело состоят из пектиновых веществ. Межклеточное вещество срединной пластинки, как бы цементирующее оболочки смежных стенок, состоит обычно из пектиновых веществ, главным образом из пектата кальция. Пектиновые вещества, хотя и в небольших количествах, имеются в основной толщине и взрослых клеток. В состав матрикса клеточных стенок помимо углеводных компонентов входит также структурный белок, называемый экстенсином. Он является гликопротеином, углеводная часть которого представлена остатками сахара арабинозы. Одревеснение клеточных оболочек Сильному метаморфозу состава и структуры подвергаются оболочки при одревеснении, опробковении, ослизнении и минерализации. Одревеснение состоит в том, что часть целлюлозной толщи стенки пропитывается лигнином. Ароматическое вещество лигнин является основным инкрустирующим веществом клеточной стенки. Это полимер с неразветлённой молекулой, состоящей из ароматических компонентов. Мономерами лигнина могут быть конифериловый, синаповый и другие спирты. Интесивная лигнификация клеточных стенок начинается после прекращения роста клетки. Отношение между целлюлозой и лигнином в одревесневших слоях оболочки было признано аналогичным конструкции железобетонных сооружений. Лигнин, подобно бетонной массе, заполняет промежутки ячеек сетки; при этом арматура и заполнение образуют монолитное целое. Одревеснение широко распространено в растительном царстве, за исключением низших растений и мохообразных. У папоротникообразных одревеснение - явление довольно обычное; а у голосеменных и покрытосеменных распространено повсеместно; здесь одревеснение встречается во всех органах и в разнообразных тканях. Понятно, что особенно сильному одревеснению подвержены клетки деревьев и кустарников. Так, древесина хвойных содержит 27-30 % лигнина, а древесина лиственных - 18-27 %. Одревеснение понижает пластичность клеточных стенок, закрепляет их форму. Однако клетки с одревесневшими стенками могут оставаться живыми десятки лет. Лигнин обладает и консервирующими свойствами и потому действует как антисептик, придавая тканям повышенную стойкость по отношению к разрушительному действию грибов и бактерий. При разложении в почве растительных остатков относительное содержание лигнина в них повышается. Несомненно, высокая стойкость лигнина играет большую роль при торфо- и углеобразовании. Кроме того, разрушение и конденсация лигнина в почве служит одним из факторов образования гумуса. Опробковение и кутинизация клеточных оболочек Весьма распространено в растительном мире наличие в толще клеточных оболочек, либо на их поверхности веществ, называемых кутинами, суберинами и спорополленинами. Общим для них являются следующие черты. Все они - высокополимерные вещества, обязательным компонентом которых являются насыщенные и ненасыщенные, жирные кислоты и жиры. От жиров, встречающихся в полости клетки, в протопласте, они отличаются нерастворимостью в ряде реактивов. Эти вещества стойки даже по отношению к концентрированной серной кислоте. Суберины, кутипы и спорополленины почти непроницаемы для воды, воздуха и других газов. Эти свойства объясняют нам значение суберинов, кутипов и спорополленинов для растений и их локализацию в растениях. Они находятся в оболочках перефирических (покровных) тканей и защищают органы растения от излишней потери воды. Суберины. Клеточные оболочки, содержащие суберины, называют опробковевшими. Суберин отлагается внутри клеточной оболочки и поэтому относится к инкрустирующим веществам. Обычно суберин составляет пластинку, находящуюся в так называемом вторичном слое клеточной стенки, так называемые субериновые пластинки. Кутины - это адкрустирующие гидрофобные вещества, покрывающие поверхность эпидермальных клеток растенеия в виде пленки - кутикулы. Обычно кутикулярная пленка многослойна. У папоротников кутикула отсутствует или слабо развита; не выражена кутикула и в корнях растений. Спорополленины имеются в наружных оболочках спор, в том числе пыльцевых зерён голосеменных и покрытосеменных растений. Спорополленины настолько стойки к разрушающим факторам, что пыльцевые зёрна со всеми тонкостями структуры их оболочек прекрасно и надолго сохраняются в почве и в торфяных отложениях и могут быть использованы для особого споро-пыльцевого анализа, позволяющего восстановить картину растительности былых эпох. Ослизнение клеточных оболочек При ослизнении клеточных оболочек образуются слизи и камеди. Те и другие представляют собой высокомолекулярные углеводы, состоящие большей частью из пентоз и их производных. Они нерастворимы в спирте, эфире, а в воде сильно набухают. Резкой границы между ними не установлено. Обычно их различают по консистенции в набухшем состоянии: камеди клейки и могут вытягиваться в нити, слизи же сильно расплываются и в нити не тянутся. В сухом состоянии камеди и слизи очень тверды и хрупки, и лишь при смачивании водой они переходят в тягучее желеобразное состояние. Ослизение бывает или нормальным явлением в жизни растения, или же происходит как патологический процесс. В некоторых случаях слизи и камеди появляются в клеточных оболочках путём отложения на ранее образовавшиеся слои клеток, иногда они образуются из имеющегося уже материала стенок в результате химического метаморфоза - "слизистого перерождения". При патологическом ослизнении - камедетечении (или гуммозе) - в камедь превращаются и стенки, и содержание клеток; процесс этот захватывает большие участки тканей. Гуммоз часто наблюдается у вишен, слив, персиков, у многих акаций и астрагалов. Значение ослизнения клеточных стенок во многих случаях очевидно. Например, ослизненные наружные слои клеток кожицы семян, набухая весной, входят в соприкосновение с почвой. Слизь, благодаря клейкости, закрепляет семена на влажном месте и, поглощая воду из почвы, улучшает водный режим проростка, передавая ему воду и защищая от высыхания. Слизь может использоваться как запасное, питательное вещество. Минерализация клеточных оболочек В более поздней стадии развития оболочки содержат минеральные вещества, причём в некоторых случаях в весьма значительных количествах. Эти вещества могут отлагаться и в толще оболочки и на её внутренней и наружной поверхности, или же в особых выростах клеточных стенок. По структуре эти отложения могут быть аморфными и кристаллическими. Наиболее распространены отложения кремнезёма и солей извести. Богаты кремнезёмом клетки кожицы стеблей и листьев хвощей, злаков, осок. Окремнение стенок свойственно и многим растениям из двудольных, особенно из семейства мареновых. Окремнению подвергаются жгучие волоски у крапивы двудольной. Кальций встречается в клеточных оболочках в виде углекислой, щавелевокислой и пектиновокислой извести. Широко распространено наличие кальция в срединной пластинке клеточных стенок. Структурная организация клеточной оболочки Клеточная стенка, как мы видели, построена из немногих основных компонентов. Применение химических методов анализа позволило выявить, что: соседние цепи целлюлозы в микро - и макрофибриллах связаны водородными связями; молекулы гемицеллюлозы прикреплены к поверхности целлюлозных микрофибрилл также водородными связями; некоторые молекулы гемицеллюлозы связаны с молекулами кислого пектина через молекулы нейтрального пектина; сами пектиновые полимеры сшиваются между собой ионами кальция (Ca); гликопротеины вероятно присоединены к молекулам пектина; и, наконец, существуют ковалентные связи между лигнином и целлюлозой. Таким образом, согласно этой модели, клеточную стенку можно рассматривать как единую гигантскую макромолекулу. Формирование и рост клеточных оболочек Даже молодые меристематические клетки снабжены первичной оболочкой. Новая оболочка формируется в процессе деления клетки в заключительной стадии митоза телофазе. Заключительная в телофазе стадия - процесс деления цитоплазмы на дочерние клетки называется цитокинезом. В ранней телофазе между двумя дочерними ядрами формируется бочкообразная система волокон, называемая фрагмопластом. Волокна фрагмопласта, так же как и волокна митотического веретена состоят из микротрубочек. В световом микроскопе видно, как в экваториальной плоскости фрагмопласта появляются мелкие капли, которые затем сливаются, образуя клеточную пластинку. Клеточная пластинка растёт центробежно до тех пор, пока не достигнет оболочки делящейся клетки. С помощью электронного микроскопа было установлено, что сливающиеся капельки - это пузырьки, отрывающиеся от аппарата Гольджи. Они в основном содержат пектиновые вещества, из которых формируется срединная пластинка, а мембраны пузырьков участвуют в построении плазматической мембраны по обеим сторонам пластинки. В это время из фрагментов трубчатого эндоплазматического ретикулума образуются плазмодесмы. После образования срединной пластинки протопласт дочерних клеток откладывает на нее первичную оболочку. Слой целлюлозы, который откладывается во время роста клетки, называется первичной клеточной оболочкой. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат и структурный белок - гликопротеин. Первичные оболочки могут и лигнифицироваться, хотя, как правило, лигнин им не свойственен. Однако наиболее характерную часть первичной оболочки составляет пектиновый компонент. Он придаёт оболочке пластичность, позволяет ей растягиваться, по мере удлинения органов: корня, стебля, листа. Пектиновые вещества способны сильно набухать, поэтому первичные оболочки содержат много воды (60-90%). В целом, на долю гемицеллюлоз и пектиновых веществ, приходится 50-60 % сухого веса первичной оболочки, содержание целлюлозы не превышает 30 %, структурный белок занимает до 10%, лигнин, как правило, отсутствует. Вторичное утолщение оболочек обычно начинается после прекращения роста клеток. При этом толщина оболочки увеличивается, а объём, запасаемый полостью клетки, сокращается. Во многих случаях по завершении вторичного утолщения протопласты клеток отмирают, но клетки продолжают функционировать, выполняя главным образом механическую и опорную функцию. В соответствие с механической функцией строение и химический состав вторичной оболочки сильно отличается от первичной. В ней значительно меньше воды и преобладают плотно сомкнутые микрофибриллы целлюлозы (40-50 %), в первичной оболочке - они расположены рыхло. Много во вторичных оболочках и лигнина - 25-30%, он придаёт оболочкам дополнительную жёсткость и прочность; гемицеллюлозы составлют 20-30% и практически нет пектиновых веществ. Итак: лицо первичной оболочки составляют пектиновые вещества, в то время как вторичной - лигнин. Вторичная оболочка далеко не всегда откладывается равномерно. У некоторых специализированных водопроводящих клеток она имеет вид колец или спиральных лент. Такие клетки сохраняют способность к продольному растяжению и после отмирания. Плазмодесмы Протопласты соседних клеток связаны между собой тонкими нитями цитоплазмы плазмодесмами. Эти структуры присущи только растительным клеткам. В нормальном состоянии плазмодесмы невидимы в световой микроскоп, однако, если стимулировать набухание оболочки плазмодесмы, становятся заметными, поэтому выявлены и описаны они были уже достаточно давно. Хотя детали строения плазмодесм изучены сравнительно недавно с помощью электронного микроскопа. Под электронным микроскопом плазмодесмы выглядят как узкие каналы (диаметром от 30 до 60 нм), выстланные плазматической мембраной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера - десмотрубочка, которая сообщается с эндоплазматическим ретикулом обеих смежных клеток. Десмотрубка напоминает цитоплазматические микротрубочки или жгутики простейших. Она состоит из 11 спирально расположенных белковых субъедениц. Вокруг десмотрубки локализируется цитоплазма, которая во многих типах плазмодесм непосредственно не соединяется с цитоплазмой клеток. В плазмодесмах обнаружена АТФ - азная активность. Наличие плазмодесм обеспечивает непрерывность цитоплазмы клеток, составляющих органы и ткани. Такая непрерывная система называется симпласт. Кроме того, за счёт плазмодесм обеспечивается единство эндоплазматической сети, переходящей из клетки в клетку. Единая эндоплазматическая сеть получила название эндопласт. Таким образом, выделяется три непрерывных компартмента в растительных тканях - это: единая цитоплазма - симпласт, непрерывный ретикулум - эндопласт и соприкасающиеся между собой клеточные стенки, вместе с межклетниками составляют непрерывную систему - апопласт. Размещаются плазмодесмы в стенке либо группами, либо равномерно разбросаны по всей стенке. Поры Внутренне утолщение клеточной стенки не бывает вполне равномерным. Сформировавшаяся оболочка имеет более толстые и менее утолщенные участки. Даже в тех случаях, когда стенки, в общем, имеют равномерную толщину, в них, при детальном рассмотрении обнаруживаются небольшие углубления. Эти места, в которых оболочка очень тонка, и называются порами. Таким образом, поры у растений - это не сквозные многочисленные отверстия, как это понимается в общеупотребительном смысле. У растений порой называют любое неутолщенное место оболочки. Для обозначения сквозных отверстий у растений используется другое название перфорации. Поры в 2-х сосединх клетках располагаются одна против другой, образуя так называемую пару пор. У клеток с мощной вторичной оболочкой поры в разрезе имеют вид радиальных каналов. На поперечном срезе эти каналы могут иметь разную форму: чаще округлую, реже щелевидную (эллиптическую или крестообразную). Округлые поры обычно формируются в паренхимных клетках, щелевидные - в прозенхимных. По форме порового канала обычно различают поры 2-х типов: простые и окаймлённые. Простые поры имеют достаточно ровный канал, с одинаковым диаметром на всём протяжении. У окаймлённых пор голосеменных растений на первичной оболочке образуется линзовидное утолщение - торус, а выросты вторичных оболочек как бы нависают над торусом. Окаймлённые поры характерны для водопроводящих элементов древесины. Эти элементы имеют вид длинных труб разного диаметра. По этим трубкам, как по капиллярам, поднимается вода. Понятно, что давление воды в смежных клетках неодинаково. В этом случае торус смещается и прижимается к выступам вторичной оболочки клеток с меньшим давлением. Обычно к порам приурочены и плазмодесменные канальцы. Нередко через одну пору проходят десятки плазмодесм. В любом случае, поры, как и плазмодесмы, облегчают диффузию веществ, растворённых в воде, из одной клетки в другую. Сквозные отверстия клеточных стенок - перфорации особенно характерны для водопроводящих поперечных перегородок водопроводящих члеников сосудов. Как правило, в этих перегородках образуются одна, две или несколько крупных перфораций. Многочисленные мелкие перфорации имеются в так называемых ситовидных трубках, по этим трубкам также передвигается вода с органическими веществами, но сверху вниз, от листьев к корням. Функции и эволюция клеточной стенки Являясь продуктом метаболической деятельности протопласта клеточная стенка выполняет ряд функций: Она защищает клеточное содержимое от повреждений и инфекций (защитная функция); Клеточная стенка поддерживает форму и определяет размер клетки; стенка играет скелетную (опорную) роль, которая особенно возрастает у наземных растений; Она имеет большое значение в росте и дифференцировании клетки; Стенка участвует в ионном обмене и поглощении клеткой веществ; Единый апопласт способствует перемещению веществ из клетки в клетку внеклеточным путём (проводящая функция); Структура клеточных стенок предохраняет клетки от избыточной потери воды (покровная функция). Примитивные клетки были окружены слизистым чехлом, состоявшим из пектиновых веществ, как и фрагмопласт, возникающий при митотическом делении в клетках современных растений. Совершенствование защитной функции клеточной оболочки привело к появлению в её составе гемицеллюлозных компонентов. Форма клетки могла поддерживаться кремниевым и карбонатным наружным чехлом, сохранившимся, как вы знаете из предыдущего курса, у некоторых водорослей. По предположению Фрей-Висслинга первичный слизистый углеводный чехол мог быть предшественником матрикса клеточной стенки. С возникновением автотрофного способа питания в оболочках клеток в качестве структурного компонента появилась целлюлоза. Выход растений на сушу поставил клеточную стенку перед необходимостью выполнять функцию опоры тела в воздухе. Именно целлюлоза оказалась наиболее оптимальным материалом (в меру прочным и в то же время эластичным) в динамичной и переменчивой среде, где подземным органам растений пришлось испытывать более сильные нагрузки. Выход на сушу и увеличение размеров растительных организмов привели также к необходимости снабжения клеток водой. Именно с развитием у наземных растений сосудов, проводящих воду, связывают появление в клеточных стенках лигнина. Лигнин не обнаружен у ископаемых океанских и современных водных растений. МЕЖКЛЕТОЧНЫЙ ПЕРЕНОС ВОДЫ И ИОНОВ ПО ПЛАЗМОДЕСМАМ В РАСТИТЕЛЬНЫХ ТКАНЯХ. РЕАКЦИЯ НА ДИНИТРОФЕНОЛ И НИСТАТИН Великанов Г.А., Ценцевицкий А.Н. Казанский институт биохимии и биофизики КНЦ РАН, 420503 Казань Объектом исследования-были корни 6-дневных проростков пшеницы Московская-35. Эффективный коэффициент самодиффузии (КСД) молекул воды в направлении по радиусу корня определяли методам ЯМР с импульсным градиентом магнитного поля путем снятия зависимости относительной амплитуды стимулированного эха от времени наблюдения (Д) при Д>300 мсек, когда релаксационное затухание намагниченности становится одноэкспоненциальным. Об изменениях водопроницаемости плазмодесм под влиянием динитрофенола (ДНФ) или нистатина судили по изменению КСД в присутствии в апопласте корня не проникающих в клетку парамагнитных ионов марганца. В такой ситуации "наблюдаемый" диффузионный пробег молекул воды локализуется в пределах симпласта, а регистрируемый КСД оказывается зависимым от водопроницаемости плазмодесм. В параллельных опытах одноэлектродным методом фиксации тока при цитоплазматической локализации кончика микроэлектрода регистрировали мембранный потенциал (МП) и входное электрическое сопротивление (Rвх) эпидермальных клеток корня. Последнее складывается из параллельно включенных сопротивления мембраны (Rм) и входного сопротивления симпласта, т.е. сопротивления растеканию тока через плазмодесмы. КСД регистрировали как в присутствии, так и в отсутствие парамагнетика в апопласте корня. Сделано заключение, что ДНФ увеличивает сопротивление межклеточному переносу воды и ионов по плазмодесмам за счет уменьшения их апертуры. Обсуждается роль энергетического обеспечения актомиозинового сфинктера в таком уменьшении апертуры транспортных каналов в плазмодесмах. Нистатин (10-4 М) увеличивает Rвx при одновременном уменьшении Rm, что свидетельствует о доминирующем увеличении электрического сопротивления плазмодесм в результате действия этого антибиотика. Водопроницаемость плазмодесм при этом не изменялась. Предполагается, что нистатин оказывает влияние на состояние двойного электрического слоя в каналах плазмодесм (за счет воздействия на ионный гомеостаз клеток), не изменяя их апертуру. Это подтверждают наши опыты по прямому определению КСД воды внутри вакуолярного и цитоплазматического симпластов, осуществленному путем анализа неэкспоненциальной формы диффузионного затухания эха. В этом случае остаются фиксированными временные интервалы между радиочастотными импульсами и выявляется "гетерогенность" КСД. Два КСД, получившиеся в результате разложения диффузионного затухания на экспоненты, несут информацию о водопроницаемости двух транспортных каналов в плазмодесмах. Разницы с контрольным (без нистатина) вариантом опыта не было выявлено. Лекция 3. Плазмалемма Эндоплазматический ретикулум Аппарат Гольджи Эндомембранная концепция Вакуолярная система Состав вакуолярного сока Топопласт Функции вакуолей Пластидная система Симбиотическая гипотеза происхождения митохондрий и хлоропластов Митохондрии Рибосомы Полисомы Лизосомы Микротела Липидные капли Микротрубочки Микрофиламенты Основное вещество ≈ гиалоплазма Эргастические вещества или включения К клеточной стенке изнутри примыкает клеточная мембрана, называемая в ботанической литературе плазмалеммой. В голых клетках плазмалемма осуществляет контакт с внешней средой, в растительных клетках, одетых клеточной стенкой, ≈ контакт между стенкой и внутренними частями протоплазмы. Толщина плазмалеммы около 7,5 - 10 нм. Это на 2 - 3 нм превышает толщину внутренних цитоплазматических мембран за счет большей величины светлого (липидного) промежутка. Таким образом, плазмалемма является самой толстой мембраной клетки. Плазмалемма содержит 40 - 50 % липидов от массы мембраны. Особенностью липидного состава плазмалеммы является высокое содержание в ней стеринов. Плазмалемма обладает хорошей проницаемостью для воды. Предполагается существование в липидном слое молекулярных пор диаметром до 0,4 нм, пропускающих воду. Сегодня большинство ученых разделяют жидкостно-мозаичную модель строения плазмалеммы, предложенную в 1972 году Сингером (Singer) и Николсоном (Nicolson). Согласно этой модели липиды образуют двойной слой (бислой), на наружной и внутренней поверхности которого как айсберги плавают глобулы белка. Отдельные белковые частицы пронзают бислой насквозь и, по-видимому, работают как ионные насосы. Глобулярные белковые частицы (7 - 9 нм) занимают до 16 % площади мембраны. Интересно отметить, что жидкостно-мозаичная модель строения плазмалеммы получила блестящее подтверждение благодаря методу замораживания-скалывания. Белковые компоненты плазмалеммы в комплексе с липидами обеспечивают выполнение ряда важных функций мембраны. Прежде всего это белковые системы транспорта веществ через мембрану. К ним относятся хорошо известные ионные насосы. Крупные молекулы проходят через мембрану только в соответствие со своей растворимостью в липидах. Этим обеспечивается высокая избирательная способность плазмалеммы. Белковые системы плазмалеммы осуществляют синтез полисахаридов. Рецепторная функция плазмалеммы интенсивно изучается. Показано, что в плазмалемме локализованы рецепторы растительных гормонов. В наружном слое плазмалеммы локализованы ферментные комплексы, участвующие в синтезе клеточной стенки. Плазмалемма выполняет функцию механической защиты у клеток, лишенных клеточных стенок. Эндоплазматический ретикулум Эндоплазматический ретикулум (ЭПР) ≈ это сложная внутриклеточная мембранная система неопределенной протяженности. В разрезе под электронным микроскопом ЭПР выглядит как две элементарные мембраны с узким прозрачным просветом между ними. Возможный объем полостей ретикулума составляет до 16 % объема цитоплазмы. В клетках различают ретикулум 2-х типов: шероховатый (или гранулярный) ЭПР несет на своей поверхности многочисленные рибосомы. Рибосомы объединены в комплексы ≈ полисомы, которые являются основным местом синтеза белка в клетках. гладкий (агранулярный) ЭПР обычно имеет трубчатую форму и функционирует, по видимому, как коммуникационная система клетки, кроме того, в нем синтезируются липофильные соединения. Ядерная оболочка и плазмодесмы также относятся к системе ЭПР. Аппарат Гольджи С плазматической мембраной и ЭПР генетически тесно связана мембранная система аппарата Гольджи. Существование этой структуры было впервые доказано в 1898 г. итальянским ученым Камило Гольджи. В растительных и животных клетках аппарат Гольджи устроен одинаково и представлен образованиями трех типов: диктиосомами, везикулами (пузырьками) и межцистерными образованиями. Уплощенные цистерны - диктиосомы расположены пачками по 5 - 8 штук. Толщина мембран, ограничивающих цистерны, составляет 7 - 8 нм. Отличительной особенностью диктиосом является наличие у них полярности: один из полюсов называется регенерационным. Мембраны диктиосом здесь сходны с мембранами гладкого эндоплазматического ретикулума, а сами цистерны упакованы достаточно плотно. Другой полюс называется секреторным. От него отщепляются секреторные пузырьки, а мембраны по толщине и свойствам напоминают плазмалемму, то есть при переходе от регенерационного полюса к секреторному мембраны как бы созревают (утолщаются). В мембранах диктиосом обнаружен ряд ферментных систем, осуществляющих синтез разнообразных веществ, в том числе полисахаридов, структурных белков и слизей, входящих в состав матрикса клеточной стенки. Синтезированные вещества заключаются в мембрану и в форме секреторных пузырьков транспортируются к плазмалемме. Сливаясь с плазмалеммой, пузырьки выносят секретируемые продукты из протопласта клетки, а мембрана везикул становится при этом плазматической мембраной. Таким образом, мембраны аппарата Гольджи и плазмалемма сходны как в онтогенетическом, так и в функциональном отношении. Аппарат Гольджи является связующим звеном между мембранами эндоплазматической сети и плазмалеммой. Кроме того, для растительных клеток очень важно функционирование аппарата Гольджи в создании клеточной стенки на протяжении всего ее онтогенеза (индивидуального развития). Эндомембранная концепция Мембраны ≈ динамические, подвижные структуры, которые постоянно изменяют свою форму и площадь. На подвижности клеточных мембран основана концепция эндомембранной системы. Согласно этой концепции, внутренние мембраны цитоплазмы (за исключением мембран митохондрий и пластид) представляют собой единое целое и берут начало от эндоплазматического ретикулума. Новые цистерны диктиосом образуются из ЭПР через стадию промежуточных пузырьков, а секреторные пузырьки, отделяющиеся от диктиосом, в конечном итоге способствуют формированию плазматической мембраны. Таким образом, ЭПР и диктиосомы образуют функциональное целое, в котором диктиосомы играют роль промежуточных структур в процессе преобразования мембран. Вспомним также, что и плазмодесмы являются частями единой эндомембранной системы, а ядерную оболочку можно рассматривать как специализированную часть ЭПР. Наружная мембрана ядерной оболочки в некоторых местах объединяется с ЭПР. Вакуоли, лизосомы и микротельца непосредственно связаны с ЭПР и являются его производными. Важно отметить, что даже в тканях, клетки которых слабо растут и делятся, постоянно происходит обновление мембранных компонентов. Переход мембран из одного компонента в другой получил название тока мембран (membrane flow), например, ЭПР АГ (аппарат Гольджи) ПЛ (плазмалемма). Ток мембран обеспечивает функциональную непрерывность и целостность мембранной системы. Вакуолярная система Вакуоль является еще одним организмом, типичным для растительных клеток. Морфология вакуолярной системы очень разнообразна ≈ от мелких многочисленных пузырьков в меристематических клетках до крупной центральной вакуоли, занимающей до 90% объема в зрелых клетках. Комплексное использование разнообразных методов: электронно-микроскопических и биохимических, позволило швейцарскому цитологу Ф. Матилю (Matile) разработать схему образования вакуолярной системы в растительных клетках. Первичные элементы вакуолярной системы в виде небольших пузырьков ≈ провакуолей обнаруживаются уже в меристематических клетках. Современная электронная микроскопия позволяет проследить, что провакуоли могут возникать как расширения цистерн эндоплазматической сети, которые затем от нее отщепляются. По мере роста клетки провакуоли сливаются друг с другом и вакуоль увеличивается в размерах. При этом формируется вакуолярная мембрана ≈ топопласт. Топопласт является производным мембран эндоплазматического ретикулума. Возникший тонопласт может образовывать инвагинации, что приводит к включению в вакуоль цитоплазматического материала. Пузырьки ≈ производные аппарата Гольджи ≈ не сливаются с мембраной вакуоли, а попадают в вакуоль в результате инкапсуляции их топопластом. Затем в полости вакуоли эти мембраны лизируются. В последнее время появилось много электронно - микроскопических доказательств существования у растения автофагии (самопожирания). Участок цитоплазмы с различными компонентами окружается мембраной эндоплазматической сети. При этом возникает особая автофаговая вакуоль. Внутри вакуоли происходит переваривание ≈ лизис содержимого. Таким образом, образовавшаяся в результате автофагии вакуоль идентична лизосоме. Состав вакуолярного сока Вакуолярное содержимое ≈ клеточный сок ≈ представляет собой водный раствор самых разнообразных веществ. Он содержит: минеральные ионы; вещества первичного обмена: органические кислоты и их соли, углеводы, пектиновые соединения, белки, а также вещества вторичного происхождения ≈ фенолы, танины, флавоноиды, пигменты, алкалоиды. Состав и консистенция клеточного сока значительно отличается от свойств протопласта. Клеточный сок обычно имеет слабокислую реакцию РН = 5.0 -6.5. Из органических кислот в клеточном соке наиболее часто встречаются лимонная, яблочная, янтарная и щавелевая. Особенно много этих кислот в клеточном соке незрелых плодов. Алкалоиды ≈ обширная группа природных азотсодержащих соединений основного характера. Они относятся преимущественно к гетероциклическим соединениям с азотом в кольце. Алкалоиды имеют горький вкус. Часто алкалоиды обладают сильным фармакологическим действием. В настоящее время из растений выделено свыше 5000 алкалоидов. Обычно концентрация алкалоидов в растениях невелика. Уже при содержании 1 - 3% растения считаются богатыми алкалоидами. Многие алкалоиды сильные яды, другие обладают наркотическим или тонизирующим эффектом. Это обусловило их широкое применение в медицине и промышленности. Так в медицинской практике нашли применение более 80 алкалоидов. С использованием алкалоидов связано производство тонизирующих напитков: чая, кофе, какао; а также табачная промышленность. Ряд алкалоидов применяют в сельском хозяйстве как инсектициды. Биологические функции алкалоидов в растениях еще окончательно не выяснены. Их считают своеобразными стимуляторами и регуляторами биохимических процессов. Несомненна в некоторых случаях защитная функция алкалоидов у ядовитых растений, предохраняющая их от поедания. Танины (дубильные вещества) ≈ это высокомолекулярные фенольные соединения, способные осаждать белки и алкалоиды. Дубильные вещества обладают вяжущим вкусом. В природе немало растений, содержащих дубильные вещества. Особенно много их в двудольных растениях. Лекарственное сырье, содержащее дубильные вещества, отличается бактерицидными свойствами. Танины используются также при отравлении тяжелыми металлами и растительными ядами ≈ алкалоидами. Обладающие антисептическими свойствами дубильные вещества защищают растения от инфекции. Гликозиды ≈ сложные органические вещества, в состав которых входит какой - либо сахар и несахаристая часть ≈ агликон. Агликонами могут быть: спирты, альдегиды, фенолы и другие вещества. Гликозиды играют в растениях весьма разнообразную роль. Некоторые исследователи определяют их как одну из форм отложения сахаров и считают их запасными питательными веществами. В пользу этого положения свидетельствует тот факт, что гликозиды легко расщепляются ферментами в присутствии воды. Другие приписывают гликозидам защитное действие, предохраняющее растение от заболеваний и поедания. Кроме того, гликозиды весьма активные биологические вещества. Они участвуют в процессах обмена, например, в построении аминокислот. Широкое применение нашли гликозиды и в медицине, особенно гликозиды сердечной группы, стимулирующие сердечную деятельность. У растений, как известно, запасные белки откладываются в вакуолях. Существует два типа запасных белковых соединений: растворимые альбумины плотные белковые комплексы фитина и глобулинов. Обычно белки откладываются в особых запасающих вакуолях, получивших название алейроновых зерен. В вакуолях часто накапливаются пигменты. Они относятся к группе гликозидов. Голубой, фиолетовый, пурпурный, темно-красный и пунцовый цвета придают растениям пигменты из группы антоцианов. В отличие от большинства других пигментов, антоцианы легко растворяются в воде и содержатся в клеточном соке. Они определяют красную и голубую окраску многих овощей, фруктов и цветов. Антоцианы окрашивают осенние листья в яркокрасный цвет. Они образуются в холодную солнечную погоду, когда в листьях прекращается синтез хлорофилла. При высоком содержании некоторых веществ в вакуолях могут образовываться кристаллы. Особенно часто встречаются кристаллы оксалата кальция, имеющие различную форму. Топопласт Мембрана, окружающая вакуоль, была названа известным генетиком Гуго Де Фризом ≈ топопластом. Общая толщина топопласта несколько меньше, чем у плазмалеммы (до 8 нм), но больше, чем у мембран эндоплазматической сети. Топопласт беднее стеролами и богаче фосфолипидами по сравнению с плазмалеммой. Высокое содержание фосфолипидов придает топопласту большую эластичность. Это имеет для вакуолей огромное значение. Топопласт способен выдерживать значительное давление клеточного сока, растягиваться и спадаться при изменении объема вакуоли. Функции вакуолей Вакуоли выполняют две основные функции ≈ 1) накопление запасных веществ и отбросов, т.е. они совмещают в себе функции склада и свалки внутри клетки; 2) поддержание тургора растительной клетки. О запасных веществах я уже говорил, приводя характеристику химического состава клеточного сока. С особенностями химического состава тесно связана и вторая функция вакуолей. Дело в том, что концентрация солей и сахаров в вакуолях обычно выше, чем в цитоплазме и окружающей среде. Поэтому вода свободно (по градиенту концентрации) поступает в вакуоли в результате осмоса. Поступающая вода оказывает давление на протопласт и клеточные стенки, вызывая напряженное состояние клетки ≈ тургор. Тургор позволяет сохранять форму мягким и сочным необревесневшим органам. Увеличение размера клетки происходит за счет роста вакуоли. Потеря тургора в результате обезвоживания вызывает увядание растения. Обезвоживание происходит не только при длительной нехватке воды. Если погрузить клетку в гипертонический раствор соли или сахара (концентрация которого выше, чем концентрация клеточного сока), то начинается осмотический выход воды из вакуоли. При этом тургор исчезает, протопласт сморщивается, отходит от клеточных стенок, наступает плазмолиз клетки. Пластидная система Для растительных клеток типична еще одна система органоидов, которой нет в клетках животных ≈ это пластидная система. Обычно пластиды ≈ крупные тельца, хорошо видимые в световой микроскоп. Каждая пластида окружена собственной оболочкой, состоящей из 2-х элементарных мембран. Внутри пластиды различают мембранную систему и основное достаточно гомогенное вещество ≈ строму. Зрелые пластиды обычно классифицируют по окраске, обусловленной пигментным составом. Выделяют три типа пластид: бесцветные лейкопласты; зеленые хлоропласты; окрашенные в другие цвета (обычно желтый, оранжевый, красный) ≈ хромопласты. Пластиды каждого типа имеют свое строение и выполняют специфические функции. Однако возможны переходы одного типа в другой, своеобразные взаимопревращения пластид. Это и дает основания объединить их в единую систему. Такая система - (совокупность) пластид клетки называется пластидом. Пластиды всех типов образуются из пропластид. Пропластиды бесцветные тельца, несколько крупнее митохондрий. В больших количествах они встречаются в эмбриональных (меристематических) клетках. Пропластиды имеют гомогенную строму и неразвитую мембранную систему, имеются лишь небольшие инвагинации (впячивания) внутренней мембраны. Пластиды зрелых клеток, сохранившие структуру пропластид, называют лейкопластами. Как и пропластиды, лейкопласты бесцветны и не содержат в строме пигментов. При изготовлении препаратов лейкопласты легко повреждаются и расплываются; кроме того, они обладают показателем преломления почти таким же, как у цитоплазмы, поэтому их бывает трудно обнаружить в световой микроскоп. Лейкопласты могут иметь разнообразную форму: эллипсоидную, амебоидную, гантелевидную, но чаще всего они шаровидны. Лейкопласты содержатся в клетках бесцветных органов и тканей; сравнительно богаты ими семена и подземные органы. Нередко лейкопласты выполняют запасающую функцию. Например, они обладают способностью образовывать крахмал из притекающей к ним глюкозы. Обычно крахмал, отлагающийся в лейкопластах, имеет вид зерен и называется вторичным. Первичный же крахмал образуется в хлоропластах в процессе фотосинтеза. Лейкопласты, накапливающие крахмал, называют амилопластами. Встречаются и лейкопласты запасающие белки. Наименее распространены лейкопласты, заполненные жиром. Хлоропласты ≈ пластиды высших растений, в которых протекает фотосинтез. Зеленую окраску этим пластидам придает пигмент хлорофилл (точнее хлорофиллы разных типов), однако кроме хлорофилла в хлоропластах содержатся пигменты, относящиеся к группе каротиноидов: оранжевый ≈ каротин и желтый ≈ ксантофилл. Правда, на свету они маскируются хлорофиллом. Хлоропласты растений обычно имеют форму диска (двояковыпуклой линзы) средним диаметром 4 - 5 мкм. Находятся они в паренхимных клетках мякоти листьев и других зеленых частей высших растений. Число их в клетке, как правило, составляет 25 - 50 шт. Общая численность хлоропластов в растении громадна. Только в одном квадратном мм листа их около 500000. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских вытянутых пузырьков и каналов. Эти внутренние мембраны стромы называются тилакоидами или ламеллами. Считается, что тилакоиды образуют единую систему. Как правило, тилакоиды собраны в стопки, так называемые граны, напоминающие столбики монет. Тилакоиды отдельных гран связаны между собой тилакоидами стромы (или межгранными тилакоидами). Пигменты хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Именно здесь, на мембранах ламелл, происходят световые реакции фотосинтеза ≈ поглощение хлорофиллом световых лучей и превращение энергии света в энергию возбужденных электронов. Темновые реакции фотосинтеза протекают вне гран, с помощью ферментов стромы. Первое органическое вещество ≈ глюкоза, образующаяся в процессе фотосинтеза, подвергается большому числу перестроек и дает начало всему многообразию органических веществ, синтезируемых в растении и составляющих его тело. Ряд этих превращений происходит тут же, в строме хлоропласта, где имеются ферменты для образования сахаров, крахмала, жиров, а также все необходимое для синтеза белков. Сахара могут переходить в другие структуры клетки, либо здесь же, прямо в хлоропластах, из них синтезируется крахмал. Этот крахмал откладывается в виде зерен и называется первичным ассимиляционным крахмалом. Вторичный крахмал, как мы уже знаем, откладывается в амилопластах. Хлоропласты способны перемещаться по клетке. На слабом свету они концентрируются под стенкой, обращенной к свету. При этом они обращаются к свету своей большей поверхностью. Если свет слишком интенсивен, они поворачиваются к нему ребром и выстраиваются вдоль стенок параллельных лучам света. Такие перемещения хлоропластов называются фототаксисом. Хлоропласты обладают известной автономией в системе клетки ≈ это полуавтономные органеллы в известном отношении напоминающие бактерий. Например, рибосомы как бактерий, так и хлоропластов на одну треть меньше рибосом эукариот. Синтез белка на рибосомах бактерий и хлоропластов подавляется антибиотиком хлорамфениколом, не оказывающим подобного действия в клетках эукариот. Кроме того, и бактерии, и хлоропласты имеют один или несколько нуклеоидов, содержащих тяжи ДНК. ДНК пластид и бактерий организованы сходным образом, а именно: не окружена мембраной, имеет кольцевую форму. И, наконец, хлоропласты способны размножаться простым делением с помощью перетяжки. Генетические код пластидной ДНК в настоящее время изучается в нескольких лабораториях. Однако, уже сейчас ясно, что образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируется хромосомой ДНК. Малопонятным остается взаимодействие хромосомной ДНК с ДНК хлоропластов. Симбиотическая гипотеза происхождения митохондрий и хлоропластов На основании сходства бактерий с митохондриями и хлоропластами эукариотических клеток можно предположить, что митохондрии и хлоропласты произошли от бактерий, которые нашли себе "убежище" в более крупных гетеротрофных клетках эукариот. Бактерии имели возможность использовать молекулярный кислород для окисления питательных веществ и использовать энергию света. Более крупные клетки ≈ хозяева использовали эти полезные свойства и имели с такими помощниками явное преимущество перед своими современниками. Все ныне живущие эукариоты, за малым исключением, содержат митохондрии, а все автотрофные эукариоты содержат также хлоропласты. По-видимому, они были приобретены в результате независимых случаев симбиоза. Более крупные клетки эукариот защищали свои симбиотические органеллы от неблагоприятных воздействий. Этиопласты образуются у растений, выращиваемых в темноте, они имеются, например, у проростков, расположенных в почве, до их выхода на дневную поверхность. Этиопласты занимают промежуточное положение между пропластидами и настоящими хлоропластами. Для них характерно хорошо развитое проламеллярное тело с кристаллической структурой. На свету этиопласты тут же превращаются в зрелые хлоропласты. Хромопласты ≈ это пигментированные окрашенные пластиды, но в отличие от хлоропластов, они не содержат хлорофиллов, а синтезируют и накапливают каротиноиды. Каротиноиды придают этим пластидам желтую, оранжевую и красную окраску. При этом каротиноиды синтезируются не на поверхности внутренних мембран, а в строме хромопластов. Как правило, каротиноиды растворены в жирных маслах пластоглобул. Внутренняя мембранная система у хромопластов либо не развита, либо деградировала. Форма хромопластов весьма разнообразна. Они придают яркую окраску лепесткам цветов, зрелым плодам. Это имеет явное приспособительное значение. Хромопласты обычно возникают из хлоропластов, реже из лейкопластов. По целому ряду признаков их можно назвать стареющими пластидами. Старение хлоропластов происходит, например, при созревании фруктов. Массовое старение хлоропластов наблюдается при пожелтении листьев осенью. Митохондрии Как и хлоропласты, митохондрии окружены двумя элементарными мембранами, каждая толщиной 5 - 6 нм. Внутренняя мембрана образует множество складок и выступов, называемых кристами. Кристы значительно увеличивают внутреннюю поверхность митохондрии. Внутреннее содержимое митохондрий называется матриксом. Митохондрии обычно мельче, чем пластиды, имеют около половины (0,5 мкм) в диаметре и очень разнообразны по форме и величине. Они могут быть округлыми, вытянутыми, гантелевидными, неправильной формы. В митохондриях осуществляется процесс дыхания, в результате которого органические молекулы расщепляются с высвобождением энергии. Энергия идет на восстановление АТФ (АДФ АТФ). АТФ ≈ основной резерв энергии всех эукариотических клеток. Поскольку в митохондриях накапливается энергия, их называют энергетическими станциями клетки. Большинство растительных клеток содержит сотни и тысячи митохондрий, хотя их количество заметно варьирует и определяется потребностью клетки в АТФ. С помощью прерывистой съемки можно увидеть, что митохондрии находятся в постоянном движении. Они поворачиваются, изгибаются, перемещаются из одной части клетки в другую, а, кроме того, сливаются друг с другом и делятся простым делением. Митохондрии обычно собираются и накапливаются там, где нужна энергия. Митохондрии, подобно пластидам, являются полуавтономными органеллами. Они содержат компоненты, необходимые для синтеза собственных белков. Рибосомы Основной функцией рибосом является трансляция, то есть синтез белков. На фотографиях, полученных с помощью электронного микроскопа, они выглядят округлыми тельцами диаметром 20 - 30 нм. Рибосомы содержат примерно равные количества РНК и белка. Каждая рибосома состоит из 2-х субъединиц неравных размеров, формы и строения. Субъединицы рибосом обозначают по величине коэффициентов седиментации (то есть осаждения при центрифугировании). В цитоплазме локализованы 80 S рибосомы, состоящие из 40 S и 60 S субъединиц. В хлоропластах содержатся 70 S рибосомы, в митохондриях 80 S, но отличающиеся от цитоплазматических. По-видимому, малая субъединица располагается поверх большой так, что между частицами сохраняется пространство ("туннель"). Туннель используется для размещения м - РНК во время белкового синтеза. Полисомы Во время синтеза белка одну молекулу м - РНК могут транспортировать несколько рибосом. Рибосомы, связанные с одной молекулой м - РНК, образуют полирибосому или полисому. Полисомы могут находиться в свободном состоянии в цитоплазме, либо могут быть связаны с мембранами эндоплазматической сети, или с наружной мембраной ядерной оболочки. Размер полисом определяется длиной молекул м - РНК. Лизосомы Лизосомы были открыты в клетках печени животных и затем обнаружены у растений. Эти органоиды диаметром около 1 мкм ограничены одинарной мембраной и содержат набор гидролитических ферментов. Мембрана лизосом полностью предотвращает выход ферментов из органоидов. Мембраны способствуют также поддержанию оптимальных условий для действия ферментов в лизосоме ≈ формируют кислую среду. Лизосомы формируются в специализированных участках гладкого эндоплазматического ретикулума. Лизосомы осуществляют: деградацию (разрушение) участков цитоплазмы собственной клетки и гидролиз запасных веществ. В растительных клетках определение лизосом затруднено, поскольку лизосомные функции выполняет вакуолярная система. Многие исследователи склонны даже не различать эти органоиды и считают, что специализированные вакуоли по переваривающей активности сравнимы с лизосомами животных. Микротела Микротела у растений были выявлены совсем недавно в 1958 г с помощью электронного микроскопа. Это тельца округлой формы 0,2 - 1,5 мкм в поперечнике, ограниченные элементарной мембраной. В некоторых микротелах обнаруживается белковый кристаллоид, представляющий собой гексагонально расположенные трубочки диаметром около 6 нм. Число микротел в различных клетках неодинаково, но чаще чуть меньше или равно количеству митохондрий. Предполагается, что микротела являются производными эндоплазматического ретикулума. В клетках растений выявлены 2 основных типа микротел с идентичной структурой, но выполняющие различные физиологические функции: 1) пероксисомы и 2) глиоксисомы. Пероксисомы многочисленны в клетках листьев, где они тесно связаны с хлоропластами. В них происходят реакции светового дыхания ≈ поглощение О2 и выделение СО2 на свету, то есть процесс, противоположный световым реакциям фотосинтеза. Глиоксисомы возникают при прорастании семян и участвуют в превращении жирных масел эндосперма в сахара. Липидные капли Липидные капли ≈ это структуры сферической формы, содержащие липиды, размером около 0,5 мкм. Похожие, но более мелкие капли пластоглобулы встречаются в пластидах. Липидные капли первоначально принимали за органеллы и называли сферосомами (за идеально округлую форму). Считалось, что они окружены двуслойной или однослойной мембраной. Однако последние данные показывают, что у липидных капель нет мембраны, но они могут быть покрыты белком. Микротрубочки Микротрубочки обнаружены практически во всех эукариотических клетках, это тонкие цилиндрические структуры диаметром около 24 нм. Длина их варьируется. Каждая микротрубочка состоит из субъединиц белка ≈ тубулина. Субъединицы образуют 13 продольных нитей, окружающих центральную полость. Микротрубочки представляют собой динамические структуры, они регулярно разрушаются и образуются вновь на определенных стадиях клеточного цикла. У микротрубочек много функций. Одна из наиболее важных - это участие в формировании клеточной оболочки. По-видимому, микротрубочки контролируют и ориентируют упаковку целлюлозных микрофибрилл. Микрофиламенты Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Они представляют собой длинные нити толщиной 5 - 7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов играют ведущую роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом. Основное вещество ≈ гиалоплазма Еще недавно основное вещество клетки считали гомогенным и бесструктурным гелем. Однако последние исследования показали, что основное вещество обладает сложной структурой. Под электронным микроскопом было обнаружено, что основное вещество представляет собой трехмерную решетку, построенную из тонких (диаметром 3 - 6 нм) тяжей, заполняющих всю клетку. Другие компоненты цитоплазмы, в том числе микротрубочки и микрофиламенты, подвешены на этой микротрабекулярной решетке. Микротрабекулярная решетка делит клетку на 2 фазы: богатые белком тяжи решетки и богатое водой пространство между тяжами. Вместе с водой решетка имеет консистенцию геля. Микротрабекулярная решетка осуществляет связь между отдельными частями клетки и направляет внутриклеточный транспорт. Эргастические вещества или включения Эргастические вещества ≈ это пассивные продукты метаболизма: запасные вещества или отходы. Обычно они представлены в форме разнообразных кристаллоподобных включений. К образованию включений приводит избыточное накопление некоторых веществ, по тем или иным причинам выключаемых из обмена и выпадающих в осадок. К эргастическим веществам относятся крахмальные зерна, кристаллы, зерна белка, липидные капли, смолы и др. Лекция 4. Общие представления о тканях растений. Классификация тканей Классификация тканей Образовательные ткани Основные ткани Ассимиляционные (фотосинтезирующие) ткани Вентиляционная ткань (Аэренхима) Запасающие ткани Выделительные ткани Выделительные ткани внешней секреции Выделительные ткани внутренней секреции Клетки у большинства растительных организмов слагаются в достаточно однородные комплексы, называемые тканями. Классификация тканей может быть проведена по двум основным принципам: морфологическому и физиологическому. Морфологический принцип кладет в основу классификации происхождение и строение ткани, а физиологический ≈ выполняемую функцию. Чисто морфологический принцип при классификации тканей использовал Де Бари, но по видимому подобные классификации всегда не вполне приемлемы, поскольку редкая ткань состоит из одинаковых по форме и происхождению клеток. Наиболее последовательной и разработанной является физиологическая классификация тканей. Основные принципы этой классификации были предложены Швенденером и его учеником Габерландтом в конце XIX века и почти в неизменном виде дошли до наших дней. Однако принятие лишь одного принципа ≈ функционального, приводит к большим противоречиям и затруднениям в силу ряда причин: Большинство растительных тканей многофункционально, то есть одна и та же ткань может выполнять несколько функций (например, проведения и укрепления). Кроме того, одна ткань может состоять из разнородных элементов, выполняющих разные функции. Такую ткань называют сложной. Часто ткань с возрастом меняет функции или сохраняет лишь некоторые из первоначальных. Например, древесина в начале своего существования выполняет преимущественно водопроводящую функцию, а потом ее теряет и служит только для укрепления растения. Клетки, сходные по строению и функции, то есть относящиеся к одной ткани, могут быть рассеяны поодиночке среди клеток других тканей и разобщены между собой. Такие разбросанные клетки-вкрапления называют идиобластами. Примером могут служить толстостенные опорные клетки в листьях растений. Кроме того, многие анатомы в качестве тканей выделяют элементы, не состоящие из клеток, например, системы межклеточных пространств. Таким образом, классификация лишь по преобладающей функции в значительной степени условна. И все же, несмотря на трудность классификации в теле растения реально существуют хорошо различимые ткани, обладающие целым характерных признаков. Знание тканей составляет основу анатомии растений, без которой немыслим анатомический анализ любого органа. Тканями называют устойчивые, то есть закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций. Строение растений усложнялось в процессе эволюции в течение многих миллионов лет, увеличивалось и разнообразие тканей. Так, у бактерий и примитивных водорослей все клетки одинаковы; у наиболее сложных бурых водорослей насчитывают не менее 10 типов клеток. У мхов их уже около 20, у папоротникообразных ≈ примерно 40, а у покрытосеменных - около 80 различных типов клеток. Все ткани делятся на 2 большие группы. В первую группу входят эмбриональные ткани, состоящие из интенсивно делящихся клеток. Они носят название образовательных тканей или меристем. Образовательные ткани в процессе роста и дифференцировки дают начало "взрослым" специализированным тканям. Все взрослые ткани входят во вторую группу и именуются постоянными. Обычно постоянные ткани классифицируют, как мы уже знаем, в зависимости от выполняемой функции. Ткани удобно рассмотреть в следующей системе. Классификация тканей Образовательные ткани (меристемы): верхушечные (апикальные); боковые (латеральные); первичные (прокамбий, перицикл) вторичные (камбий, феллоген) вставочные (интеркалярные); раневые (травматические). Постоянные ткани Основная паренхима ассимиляционные ткани; запасающие ткани; аэренхима; Всасывающие ткани ризодерма; веламен; всасывающий слой в зародышах злаков; гаустории паразитных растений; Покрывные ткани первичная (эпидерма); вторичная (перидерма); третичная (корка, ритидом); Ткани, регулирующие прохождение веществ эндодерма; экзодерма; Выделительные ткани наружные: железистые волоски (трихомы) и выросты (эмергенцы), нектарники, гидаторы; внутренние: выделительные клетки, многоклеточные вместилища выделений, смоляные каналы, млечники; Механические ткани (опорные, скелетные, арматурные) колленхима склеренхима: волокна и склереиды; Проводящие ткани ксилема (древесина); флоэма (луб). Образовательные ткани В отличие от типичных животных, высшие растения растут и образуют новые клетки в течение всей жизни, хотя обычно с некоторыми перерывами. У многолетних растений средних широт, например, перерывы обусловлены наступлением зимнего периода покоя. Другой особенностью растений является локализованность их роста: они растут в определенных участках своего тела. Эти участки называют точками роста или зонами роста. Основной функцией зон роста является активное деление и порождение новых клеток. Возникающие из меристем клетки растут и дифференцируются в постоянные ткани. Форма клеток меристем разнообразна: но чаще всего они паренхимны и имеют очертания почти правильных изодиаметрических многогранников. Образовательные ткани состоят из плотно сомкнутых клеток, как правило, без межклетников. Хотя это правило полностью справедливо лишь в отношении меристем стеблей, меристемах же корней нередко имеются хорошо различимые межклетники. Полость каждой клетки заполнена густой цитоплазмой и сравнительно крупным ядром. Здесь имеется в виду отношение размеров ядра к размерам всей клетки. Дело в том, что в постоянных тканях объем клетки заметно увеличивается за счет растяжения, а объем ядра остается неизменным. Практически все органоиды в меристематических клетках находятся в стадии становления. Обычные меристематические клетки способны делиться определенное, ограниченное число раз, после чего превращаются в постоянные ткани. Но существуют клетки, способные к неограниченному делению, на протяжении всей жизни растения; такие клетки называют инициальными или инициалями. Тело зародыша в развивающемся семени состоит на первых стадиях сплошь из промеристемы. Но в скором времени на двух противоположных полюсах, в точках роста ≈ на кончиках зародышевого корешка и стебля из инициальных клеток формируются верхушечные (апикальные) меристемы. Вообще следует отметить, что по положению в теле растений различают: верхушечные (апикальные), боковые (латеральные) и вставочные (интеркалярные) меристемы. Боковые (латеральные: лат. латералис - боковой) меристемы в осевых органах: стеблях и корнях образуют цилиндрические слои. На поперечных срезах они имеют вид колец. В одном случае боковые меристемы формируются под апексами (верхушками) в тесной связи с апикальными меристемами. Такие меристемы относят к первичным. Первичными латеральными меристемами являются прокамбий и перицикл. Другие боковые меристемы появляются значительно позже, при уже развитых постоянных тканях и на этом основании их считают вторичными, хотя разграничение на первичные и вторичные меристемы в значительной мере условно. К вторичным боковым меристемам относятся камбий и пробковый камбий или феллоген. В наиболее типичном случае вторичные меристемы возникают из постоянной ткани путем ее обратного преобразования в меристему. Вставочные (интеркалярные) меристемы расположены обычно в основании междоузлий. Эти меристемы можно назвать остаточными, поскольку они происходят от верхушечных меристем, но их преобразование в постоянные ткани задерживается по сравнению с соседними участками стебля. В интеркалярные меристемы обычно вкраплены дифференцированные элементы, например, они пронизаны проводящими пучками и в них нет инициальных клеток. Этим вставочные меристемы отличаются от боковых и верхушечных. Можно сказать, что вставочные меристемы имеют временный характер и, в конце концов, превращаются в постоянные ткани. Вставочные меристемы хорошо выражены у основания молодых листьев, поэтому и листовая пластинка нарастает своеобразно ≈ основанием, а не верхушкой, как стебель. Хотя и стебли некоторых растений имеют дополнительный интеркалярный рост за счет междоузлий. Особенно это характерно для злаков. Особую группу составляют раневые или травматические меристемы, как уже видно из названия, появляются при залечивании поврежденных тканей и органов. Обычно на месте повреждения со временем формируется защитная пробка. Есть еще одна особенность, свойственная меристематическим клеткам: обычно во время роста оболочки соседних клеток растягиваются синхронно (согласовано). Этим обеспечивается сохранность плазмодесм между клетками. Поскольку протоплазмы соседних клеток, связанные плазмодесмами, образуют единую живую систему ≈ симпласт, то и подобный согласованный рост, при котором оболочки соседних клеток не сдвигаются друг относительно друга, носит название симпластического. Иногда, однако, наблюдается так называемый интрузивный рост, при котором клетка внедряется между соседними, как бы расталкивая их. Понятно, что при этом оболочки скользят одна по другой и плазмодесменные связи рвутся. Основные ткани Согласно уже очень старой концепции Сакса (Sacks, 1868) тело растения состоит из трех систем тканей. Наибольший объем в растении занимают основные ткани. Сакс называл их мякотью. Снаружи они защищены покрывными тканями, а изнутри пронизаны проводящими и механическими пучками. Образующие их клетки отличаются обычно паренхимным строением, из-за этого основные ткани нередко называют основной паренхимой. Основная ткань, в противоположность другим очень богата межклеточными пространствами. Нередко межклетники во много раз превышают размеры самих клеток. Основные ткани являются преимущественно питающими тканями, хотя по своему существу могут выполнять многие функции, начиная от ассимиляционной, кончая выделительной. В зависимости от специализации, основную паренхиму делят на ассимиляционную, вентиляционную, всасывающую, запасающую и выделительную. Ассимиляционные (фотосинтезирующие) ткани Ткани, основной функцией которых является работа ассимиляции, то есть фотосинтез, объединяют в систему ассимиляционных тканей. У высших растений они имеют обычно зеленую окраску. Поэтому могут быть названы зеленой паренхимой или хлорофиллоносной паренхимой (короче ≈ хлоренхимой). Ассимиляционная ткань устроена достаточно просто и состоит из однородных тонкостенных клеток. Хлоропласты в клетках хлоренхимы обычно расположены в один ряд в постенном слое цитоплазмы. Центральная часть полости клетки занята крупной вакуолью. Доступ углекислоты к клеткам хлоренхимы облегчается тем, что в ней имеется развитая система межклетников, сообщающаяся с атмосферой. Наличие межклетников является характернейшей особенностью ассимиляционной ткани. Межклетники обеспечивают газообмен с окружающей средой. В соответствии с тем, что работа ассимиляции происходит за счет солнечной энергии, хлоренхима располагается в местах, наиболее доступных свету: она находится в надземной части растений непосредственно под кожицей листьев и стеблей. Нередко хлоренхима дифференцирована на столбчатую (палисадную) и губчатую ткань. Палисадная ткань обычно состоит из удлиненных клеток цилиндрической формы, расположенных перпендикулярно к поверхности органа. Межклеточники в палисадной ткани развиты слабо. Палисадная ткань содержит большое количество хлоропластов, здесь происходят световые реакции фотосинтеза. Губчатая ткань построена из округлых или неопределенной формы клеток, образующих рыхлую сложную сетчатую систему. Межклетники хорошо развиты. Здесь протекает газообмен и темновая стадия фотосинтеза. Вентиляционная ткань (Аэренхима) Практически во всех тканях имеются межклетники, образующие единую систему. Межклетники через проходные отверстия или устьица сообщаются с атмосферой. Однако газовый состав в межклетниках сильно отличается от газового состава атмосферы. Если в растениях образуется ткань с очень большими межклетниками, нередко превышающими размеры самих клеток, и если вентиляционная функция такой ткани выступает на первое место, то ее называют аэренхимой. Как правило, в состав аэренхимы входят механические клетки, придающие этой рыхлой ткани дополнительную прочность. Особенно развита аэренхима у водных и болотных растений, в условиях, где затруднен нормальный газообмен. Запасающие ткани Поглощенные растением синтезированные вещества могут откладываться в виде запасов. К накоплению запасных веществ в той или иной мере способны все живые клетки, но в том случае, когда запасающая функция гипертрофированна (выступает на первое место), говорят о запасающих тканях. Запасы могут храниться длительное время, например, зимние запасы крахмала в корнях и клубнях двулетних и многолетних растений. В других случаях запасы почти непрерывно потребляются и вновь пополняются в период вегетации. В строении запасающих клеток встречается большое разнообразие. Некоторые запасающие ткани приспособлены к накоплению воды. Их называют водоносными. Как правило, клетки водоносных тканей чрезвычайно гигроскопичны (т.е. способны быстро впитывать влагу) и имеют тонкие оболочки. Наличие водоносных тканей характерно для некоторых ксерофитов ≈ растений, приспособленных переносить длительный засушливый период. Чаще всего водоносные ткани встречаются в листьях. Они являются резервуарами влаги. При подсыхании растения водоносные клетки передают воду главным образом ассимиляционным тканям. Мощно развитая водоносная ткань встречается у суккулентов ≈ растений с мясистыми сочными листьями и стеблями: агавы, алоэ, кактусы, молочаи. Развитой водоносной тканью снабжены клубневидные вздутия стеблей многих эпифитных орхидных. Однако подлинными рекордсменами в запасании влаги являются сфагновые мхи. Они могут накапливать такое количество влаги, которое в 40 - 50 раз превышает их сухой вес. Такая высокая гигроскопичность сфагнов связана с особенностями их анатомического строения. В листьях сфагновых мхов имеются две группы клеток: особые гиалиновые клетки, имеющие большие или многочисленные поры и накапливающие воду и обычные хлорофиллоносные клетки. Гиалиновые клетки занимают большую часть объема. Нередко крупные водоносные клетки имеют спиральные или кольчатые утолщения оболочек, придающих клеткам дополнительную прочность и предохраняющие их от слияния. Значительно шире распространены ткани, запасающие пластические вещества. Обычно запасы накапливаются в полостях клеток, реже в их оболочках. Наиболее распространенные запасные вещества: сахар, инулин, аминокислоты, белки, крахмал. В клеточных стенках обычно откладываются гемицеллюлозы. В этих случаях стенки чрезвычайно сильно утолщаются. В качестве примеров можно назвать запасающие ткани эндосперма семян кофе и финиковой пальмы. Выделительные ткани В процессе обмена веществ у растений, помимо продуктов потребляемых самим растением, образуется еще ряд отбросов или побочных продуктов, для питания растения никакого значения не имеющих; они могут быть отнесены к группе выделений. У высших растений происходит выделение капельно-жидкой воды и растворов различных веществ, а в некоторых случаях вещества затвердевают и выкристаллизовываются внутри тела растения или на его поверхности. В связи с этим различают выделительные ткани внутренней и внешней секреции. Выделительная деятельность растений очень разнообразна, поэтому изучение выделительных тканей имеет ряд трудностей. Во-первых, эти ткани сильно различаются по строению и размещению в теле растения. Во-вторых, растения выделяют очень разнообразные в химическом отношении вещества, причем одинаковые вещества могут вырабатываться разными видами выделительной ткани и наоборот. И, наконец, остается неясным значение выделяемых веществ для самих растений. Многие "отбросные" вещества в процессе приспособительной эволюции получили дополнительную вторичную функцию, например, имеют горький вкус или ядовиты и предохраняют растения от поедания или служат для дезинфекции как некоторые смолы. Выделительные ткани внешней секреции 1. Гидатоды ≈ устройства, служащие для выделения воды. У многих растений, особенно у тропических, разные органы (главным образом листья), выделяют воду в виде капель. Это явление называется гуттацией. Гуттация происходит особенно интенсивно в условиях, затрудняющих транспирацию ≈ испарение воды листьями. Такие условия создаются, например, в прохладные безветренные ночи при влажном воздухе. В ранние утренние часы после таких ночей можно наблюдать капельки воды на листьях растений ≈ растения гуттируют. Гуттационные капли обычно легко отличаются от росы. Они располагаются большей частью под верхушкой листа или на кончиках зубцов его пластинки. Положение капель гуттационной воды показывает и положение органов ее выделяющих ≈ гидатод. Гидатоды наиболее простого типа представляют собой видоизмененные клетки кожицы или многоклеточные волоски. Они пространственно не связаны с тканями, проводящими воду. Наиболее сложный тип представляют гидатоды, у которых кожица участвует в формировании специальных выводных отверстий для воды, в виде водяных устьиц или водяных щелей. В отличие от обычных устьиц, водяные устьица лишены подвижности, а щели постоянно открыты. Гидатода снабжена либо одним крупным устьицем с широкой щелью, как у первоцветов и аконитов, либо имеется группа мелких устьиц обычно с узкими щелями, как у многих толстянок, сложноцветных, зонтичных. 2. Весьма разнообразны и широко распространены наружные эпидермальные железки. У многих растений кожица листьев и стеблей обладает высокоспециализированными многоклеточными железистыми волосками. Эти волоски имеют обычно многоклеточную ножку и округлую одноклеточную головку (как у первоцветов, пеларгоний). Эфирные масла заполняют пространство между целлюлозной оболочкой и кутикулой. Достаточно часто встречаются и многоклеточные головки. Железистый волосок может быть и ветвистым, как у некоторых бегоний или хмеля. Очень своеобразны жгучие волоски крапивы. Они имеют многоклеточное основание, переходящее в крупную клетку с хрупкими кремниевыми стенками. Едкая жидкость наполняет эту клетку под большим давлением. При легком прикосновении верхушка клеткиампулы обламывается, оставляя острые края. Волосок впивается в кожу, впрыскивая сок. 3. Особый тип желез наружной секреции представляют нектарники. Нектарники ≈ органы выделения сахаристой жидкости нектара. Обычно они находятся в цветке. Нектар служит средством привлечения животных, чаще всего насекомых, которые производят перекрестное опыление растений. 4. И, наконец, самым редким типом выделительных органов являются переваривающие железки насекомоядных растений. Наружными переваривающими железками обладают, например, листья росянки. Заканчивая обзор наружных выделительных органов, необходимо отметить, что в целом, растительные выделения, в противоположность животным, относительно редко выводятся наружу. В этом состоит одна из характерных особенностей растений. Объясняется это отчасти тем, что выделяемые вещества образуются внутри клеток, одетых к тому же клеточными оболочками. Значительно чаще растительные выделения собираются внутри самого растения. Поэтому и внутренние выделительные ткани заметно разнообразнее. Перейдем к их рассмотрению. Выделительные ткани внутренней секреции Вместилища выделений весьма разнообразны по форме, величине и происхождению. В зависимости от способа их образования различают: 1) схизогенные; 2) рексигенные; 3) лизигенные вместилища. Схизогенные вместилища образуются путем расхождения оболочек клеток, первоначально тесно примыкавших друг к другу. Расхождение клеток происходит путем расщепления срединной пластинки. Рексигенные межклетники возникают путем разрыва целых участков тканей, а затем высыхания и отмирания клеток. Таким способом образуются крупные полости в междоузлиях стеблей многих злаков, зонтичных и др. Лизигенные вместилища появляются при растворении - лизисе клеток и их оболочек. Обычно это полости или карманы правильной сферической формы. Таковы, например, вместилища в наружной корке плодов апельсина и других цитрусовых. Наибольшие шаровидные полости, наполненные выделениями, встречаются преимущественно в листьях. Эти вместилища лежат чаще вблизи поверхности под кожицей, поэтому нередко просвечивают и видны невооруженным глазом, как светлые прозрачные точки. Подобные вместилища встречаются в листьях зверобоя, лавра, эвкалипта, мирта, магнолии и представляют собой идиобласты среди паренхимной ткани. Каналообразные выделительные устройства или ходы образуются преимущественно в стеблях и корнях, реже в листьях. Каналы по их содержимому называют: масляными, смоляными, слизевыми и камедевыми. Обычно полости выделительных ходов возникают схизогенным путем. Смоляные ходы имеются, например, у многих хвойных, зонтичных, сложноцветных. Типичный смоляной ход представляет собой длинный трубчатый межклетник, окруженный живыми клетками эпителия. Выстилающие эпителиальные клетки выделяют в полость канала экскреты в виде смол и эфирных масел. Расположение смоляных ходов сложное. Они проходят и в вертикальном и в горизонтальном направлениях, соединяясь в общую систему. Наиболее своеобразными трубчатыми каналами являются млечные сосуды или млечники. Содержащийся в млечниках млечный сок (или латекс) представляет собой эмульсию или суспензию, водный раствор, в котором во взвешенном состоянии находятся гидрофобные капельки разнообразных веществ и твердые частицы: смолы, камеди, каучук и др. Типичный млечный сок похож на молоко и обычно молочно-белого, как, например, у молочая, цвета. Реже молочный сок бывает желтого ≈ у некоторых маков или оранжевого (у чистотела) ≈ цвета. Помимо смол и каучуков латекс содержит сахара, белковые вещества, эфирные масла и алкалоиды, гликозиды. Так, в опии (затвердевшем млечном соке незрелых коробочек опийного мака) найдено до 20 алкалоидов, в том числе морфин, папаверин, кодеин. Наличие в млечниках продуктов ассимиляции, таких как белки, углеводы, жиры, заставляет многих исследователей считать млечники проводящей тканью, подобно ситовидным трубкам, тем более, что при наличии млечников ситовидные трубки немногочисленны. Более того, в проводящих пучках снотворного мака ситовидные трубки вообще отсутствуют. Однако наличие в млечниках конечных продуктов обмена веществ, таких как смолы, эфирные масла ≈ позволяет относить млечники к выделительной системе. Так что классификационная принадлежность млечников ≈ вопрос спорный. Сами млечники бывают двух видов: 1) членистые и 2) нечленистые. Нечленистый млечник представляет собой гигантскую многоядерную клетку с одной непрерывной вакуолью. Одновременно с развитием проростка клетка-млечник растет в длину, ветвится и, протискиваясь подобно гифам гриба-паразита, проникает в разные части тела растения. Нечленистые млечники имеют большинство молочайных, некоторые тутовые, бересклеты. Членистые млечники состоят из многих отдельных млечных клеток. Эти клетки в простейших случаях сообщаются друг с другом порами (как у кленовых, в чешуях луков). Но в большинстве случаев перегородки с порами имеются только в ранней стадии развития членистых млечников. Позже поперечные перегородки растворяются и исчезают полностью или частично. Членистые млечники характерны для растений семейств маковых, колокольчиковых, ароидных, банановых, некоторых сложноцветных и многих других. Вопрос о значении млечников для растений до сих пор не выяснен. Несомненна лишь побочная роль некоторых млечников в жизни растений, в том случае, когда латекс содержит горькие или ядовитые вещества и предохраняет растение от поедания. Если значение многих выделений в жизни растений не вполне понятно, то их значение в хозяйственной деятельности человека очевидно. Бальзамы, смолы, эфирные масла, каучук широко используются в промышленности и медицине. Например, ароматические летучие жидкости ≈ эфирные масла специфичны для каждого вида растений. Запахи розы, сирени, фиалки нельзя спутать. Эфирные масла применяются в технике, медицине, парфюмерной, кондитерской и других отраслях промышленности. В наибольшем количестве они употребляются для приготовления ликеров и водок, и используются как растворители смол для приготовления лаков. Эфирные масла при соприкосновении с воздухом затвердевают, превращаясь в смолы. Смеси смол и эфирных масел, имеющие вид густой сиропообразной жидкости и отличающиеся характерным запахом называются бальзамами. При поранении деревьев бальзамы вытекают, окончательно затвердевают и превращаются в смолы. В отличие от бальзамов, смолы практически не имеют запаха. Особенно ценны даммаровая смола, акароня и масличная смола, получаемые из тропических и субтропических деревьев. Смолы применяются при изготовлении лаков, мастики, смазочных масел, типографской краски. К числу наиболее широко распространенных и имеющих практическое значение относятся бальзамы хвойных деревьев. Главнейшим продуктом этих бальзамов является скипидар. "Канадский бальзам", широко применяющийся в микроскопии, в нашей стране получают из пихты сибирской. Народная медицина много веков применяла различные бальзамы для лечения ран. Выражение "бальзам на рану" вошло в обиходную речь. Знаменитый перуанский бальзам добывается из живицы тропического дерева Myroxylon balsamum. Отличными заживляющими свойствами обладают бальзамы сибирской пихты. И, наконец, до тех пор, пока не была разработана технология синтетического получения каучука, он добывался исключительно из растений. Впервые сырьевой каучук был обнаружен европейцами в бассейне р. Амазонки в XVII веке. Индейцы добывали его из тропического лесного дерева Hevea brasilensis, относящегося к семейству молочайных. Только в конце 19 века семена были перевезены в Европу, и оттуда доставлены на о. Цейлон и на Малайский полуостров. Ныне Малайя и Индонезия ≈ основные центры культивирования гевеи. В 30-х годах в нашей стране был своеобразный бум по поиску растений-каучуконосов, в целях избавиться от импорта каучука из-за границы. Впервые были найдены и введены в культуру травянистые каучуконосы, такие как кок-сагыз (Taraxacum kok-saghyz). Лекция 5. Покровные ткани Эпидерма Перидерма Пробка (феллема) Ритидом Механические ткани Колленхима Склеренхима Склереиды Организация тела растения с точки зрения строительно-механических принципов К этой категории относятся ткани, покрывающие органы растений. Они защищают внутренние ткани от вредных наружных воздействий и ослабляют действие факторов внешней среды, которые при высокой интенсивности могут быть опасными для растений. Особенно большое значение покровные ткани имеют для надземных органов, обитающих в переменчивой (динамичной) воздушной среде. Покровные ткани предохраняют органы растения от резких температурных колебаний (сильного нагревания или охлаждения), от чрезмерной потери воды, путем испарения, от повреждения каплями дождя, града, твердыми частицами атмосферной пыли, от проникновения внутрь растения болезнетворных организмов. Многие из этих моментов имеют значение и в отношении подземных и подводных органов. У корней роль первичной защитной покровной ткани играет экзодерма, а у стеблей и листьев ≈ эпидерма (кожица). На смену первичной покровной ткани у многих растений образуется вторичная покровная ткань ≈ пробка (феллема), она является частью комплекса тканей, называемого перидермой. Еще позже, во многих случаях образуется весьма сложный тканевый комплекс ≈ корка (третичная покровная ткань). Рассмотрим все по порядку. Эпидерма Покровные ткани не могут наглухо изолировать растение от внешней среды, растение находится в состоянии непрерывного обмена со средой. Поэтому второй, не менее важной, чем защитная, функцией эпидермы является регуляция газообмена и транспирации (естественного испарения воды живыми тканями). В процессе эволюции эпидерма возникла очень давно, в самом начале приспособления растений к условиям жизни на суше. Без нее не мыслимо существование высших сухопутных растений. Уже у ринии имелась вполне развитая эпидерма. Кроме типичных функций, характерных для покровной ткани, эпидерма может функционировать как всасывающая ткань, принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев. Таким образом, эпидерма многофункциональная ткань. В структурном отношении эпидерма - сложная ткань, поскольку в ее состав входит ряд морфологически различных элементов: основные клетки эпидермы; замыкающие и побочные клетки устьиц; трихомы (производные эпидермальных клеток в виде выростов и волосков). Основная ткань эпидермы состоит из живых плотно сомкнутых клеток, имеющих нередко извилистые стенки. За счет извилистости стенок увеличивается сила сцепления клеток и дополнительно повышается прочность ткани. Обычно основные клетки эпидермы прозрачны и не содержат хлоропластов, а если хлоропласты и имеются, то в очень незначительном количестве. Через прозрачные клетки основной ткани беспрепятственно проходят солнечные лучи. Оболочки клеток кожицы утолщены неравномерно: в каждой клетке наиболее толста наружная стенка, боковые стенки несколько тоньше, внутренние еще более тонки. Клетки эпидермы обычно покрыты тонкой пленкой ≈ кутикулой. Она представляет собой продукт жизнедеятельности цитоплазмы, клетки, которая выделяет через оболочку на ее поверхность жидкий кутин, затвердевающий в пленку. Обычно кутикула неоднородна в своем строении и многослойна. Кутикулярные слои нередко пропитаны воском, залегающим в виде включений и прослоек. Отложения воска на поверхности кутикулы разнообразны и имеют вид: а) мелких зерен, расположенных равномерным слоем; б) чешуек; в) тонких палочек, часто изогнутых и на конце закрученных, например, на стеблях сахарного тростника палочки воска достигают длины 0,1 мм. Восковой налет снижает интенсивность транспирации у листьев: в опытах листья эвкалипта, с которых осторожно удаляли воск, испаряли воды на 30% больше по сравнению с контрольными. Восковой покров может иметь и иное значение, особенно для растений теплых дождливых районов. Он делает поверхность органов несмачиваемой и с них легко и быстро стекает вода. У насекомоядных растений из рода Nepenthes цветы имеют вид урночек или колпачков. Их поверхность изнутри покрыта мелкими восковыми чешуйками, легко отделяющимися при нажиме лапок насекомых. Даже бескрылые осы, способные ползать по вертикальной поверхности стекла, садясь на окраину урны, не могут удержаться и неизбежно падают на дно. Эпидерма листьев и стеблей растений, растущих погруженными в воду, почти не имеет кутикулы и, тем более, воскового налета. Чем суше местообитание растения, тем кутикулярная пленка более отчетливо выражена. Иногда эпидерма состоит из нескольких слоев клетки. Предполагают, что в этом случае эпидерма выполняет водозапасающую функцию. Поскольку такая эпидерма отмечена преимущественно у тропических растений, произрастающих в условиях непостоянной обеспеченности водой, таких как фикусы, бегонии. В клетках эпидермы могут образовываться различные продукты жизнедеятельности протопласта. Особенно интересны так называемые цистолиты. Они формируются в гипертрофически разросшихся клетках эпидермы и представляют собой гроздьевидные кристаллы углекислой извести. Клетки с цистолитами представляют собой идиобласты. Другая особенность внешних стенок клеток эпидермы отдельных растений ≈ пропитывание их минеральными солями кальция и кремния. У осок кремний отлагается даже в кутикуле. В некоторых случаях клеточные оболочки приобретают настолько большую прочность, что хвощи, например, в кожице которых отлагается кремнезем, используют для полировки. Наличие непрерывного слоя кутикулы лишило бы растения возможности какого-либо газообмена со средой, что неизбежно привело бы его к гибели. Поэтому в процессе эволюции возникли специфические структуры ≈ устьица. Через них и осуществляется сообщение с внешней средой. Там, где нет кутикулы, например, у подводных растений, нет и устьиц. Чем толще кутикула, тем многочисленнее устьица. Через устьица проходит чрезвычайно интенсивная диффузия водяного пара, кислорода и углекислого газа. Каждое устьице состоит из пары замыкающих клеток и устьичной щели, которая представляет собой межклетник. Замыкающие клетки отличаются от окружающих их обычных эпидермальных клеток своей формой и наличием хлоропластов. Чаще всего замыкающие клетки имеют бобовидную форму. Кроме того, замыкающие клетки обычно имеют более мелкие размеры. Как правило, замыкающие клетки окружены так называемыми побочными клетками устьиц, отличающимися морфологически от основных клеток эпидермы. Побочные клетки функционально тесно связаны с замыкающими и составляют вместе устьичный аппарат (или устьичный комплекс). Побочные клетки устьиц весьма разнообразны по форме, для них даже разработана специальная классификация, а примитивные высшие растения вообще не имеют побочных клеток. Раскрывание и закрывание устьиц представляет чрезвычайно важное явление в жизни растений. Полностью механизм работы устьичного аппарата был выявлен совсем недавно, но уже со времен Швенденера известно, что основным фактором здесь является изменение тургора (осмотического давления) внутри замыкающих клеток. Раскрыванию устьиц, кроме того, способствует неравномерно утолщенные оболочки замыкающих клеток. Внутренние стенки, окаймляющие устьичную щель, более толстые, чем наружные. Поэтому при повышении давления в замыкающих клетках наружные стенки изгибаются сильнее и устьичная щель приоткрывается. Изменение тургорного давления в замыкающих клетках обусловлено изменением в них концентрации ионов калия. Ионы калия закачиваются в замыкающие клетки против градиента концентрации. На это требуется большое количество энергии, поэтому замыкающие клетки содержат многочисленные митохондрии. Углеводы, необходимые для активной деятельности митохондрий, синтезируются хлоропластами. При высокой концентрации калия вода всасывается в замыкающие клетки, их объем увеличивается и устьице открывается. Отток ионов калия и соответственно воды совершается пассивно. Резервуаром ионов калия служат побочные клетки. В движении устьиц особое значение имеет также и радиальная ориентация целлюлозных микрофибрилл в оболочках замыкающих клеток. Эти радиальные мицеллы позволяют замыкающим клеткам удлиняться и одновременно не дают им расширяться. В большинстве случаев устьица в значительно больших количествах расположены на нижней стороне листовых пластинок, чем на верхней. В этом случае устьица не подвержены прямому воздействию солнечных лучей и меньше нагреваются. Устьица на верхней стороне листа преобладают у травянистых растений, обитающих на сильно нагреваемых каменистых склонах. И, наконец, у водных растений, таких, как кувшинки, водной лилии, у которых листья расположены на поверхности воды, устьица находятся только на верхней стороне листа. Таким образом, количество устьиц и их локализация в значительной мере зависит от экологических условий. В среднем на 1мм2 поверхности листа насчитывается 100-300 устьиц. У огромного большинства высших растений клетки эпидермы образуют выросты ≈ трихомы или волоски (греч. трихос ≈ волосок). К трихомам относятся самые разнообразные выросты эпидермы. Некоторые из них действительно напоминают формой волоски, другие имеют вид сосочков, бугорков, крючочков, чешуек. Трихомы бывают железистые и кроющие. В железистых трихомах накапливаются экскреты, поэтому их относят к выделительной системе. Кроме того, различают волоски одноклеточные и многоклеточные, мертвые и живые. Мертвые волоски лишены протопласта, полости их заполнены воздухом, вследствие чего они кажутся белыми. Растение густо покрытое мертвыми волосками, имеет седой вид. Такие волоски лучше отражают солнечные лучи и этим уменьшают нагревание и испарение у растения. Форма волосков очень разнообразна и характерна для того или иного вида растения. Волоски бывают головчатые, звездчатые, крючковидные, чешуйчатые, ветвистые. Нередко трихомы защищают растения от насекомых. При этом, чем гуще опушено растение, тем реже насекомые посещают его и используют в качестве пищи и для откладки яиц. От трихомов следует отличать эмергенцы (лат. emergere - выдаваться) ≈ структуры, в образовании которых принимает участие не только эпидерма, но и глубже расположенные ткани. У некоторых растений, малин, роз, образуются эмергенцы, называемые шипами. В образовании шипов у шиповника, например, кроме эпидермы участвуют 2 ниже лежащих слоя. От настоящих колючек (метаморфозов органов) эмергенцы отличаются беспорядочным расположением. Перидерма У большинства многолетних растений осевые органы (корень и стебель) неуклонно нарастают в толщину, за счет деятельности вторичных меристем. Но первичная покровная ткань ≈ эпидерма не обладает меристематической активностью и не может следовать за утолщением органов. Под напором образующихся вторичных тканей она разрывается и слущивается. На смену ей приходит сложная вторичная покровная ткань ≈ перидерма. Начало образованию перидермы кладется формированием вторичной латеральной меристемы ≈ пробкового камбия или феллогена. Феллоген работает на две стороны. Наружу он откладывает слои пробки (или феллему), а внутрь органа ≈ живую паренхимную ткань ≈ феллодерму. На поперечном срезе органа феллоген выглядит в виде кольца плотно сомкнутых тонкостенных клеток, имеющих, как правило, прямоугольные очертания. Пробка (феллема) Клетки пробки образуют правильные радиальные ряды над производящими их клетками феллогена. Характерная особенность клеток феллемы ≈ суберинизация их оболочек. Жироподобное вещество суберин откладывается в виде сплошных пластинок, а сами целлюлозные оболочки, кроме этого, нередко одревесневают. Оболочки пробковых клеток могут иметь коричневый или желтый цвет. Поры в клетках пробки развиты слабо. В этом нет необходимости, поскольку живое содержимое в клетках пробки обычно рано отмирает и полости клеток заполняются воздухом. У некоторых растений в клетках пробки имеется зернистое содержимое, например, белое смолоподобное вещество бетулин у березы или церин в клетках дуба. Предполагают, что эти вещества обладают антисептическими свойствами. У многих растений, например, у берез, ежегодно образуются новые тонкостенные и толстостенные слои пробки, что-то вроде годичных колец. Именно поэтому береста легко расслаивается. Мощность феллемы сильно варьирует. Чрезвычайно мощную пробку имеют знаменитые пробковые дубы (бутылочная пробка). Пробка функционирует в качестве покровной (защитной) ткани. Особенно велика роль пробки у надземных органов. Пробка непроницаема для воды, поэтому предохраняет стволы и ветви от высыхания. Ритидом У большинства древесных пород на смену вторичной покровной ткани перидерме приходит третичная покровная ткань - корка или ритидом. Деятельность пробкового камбия не может продолжаться неограниченно долго. Обычно через определенный период времени его активность затухает и тогда в более глубоких слоях коры закладывается новый слой феллогена, производящий пробку. Так, в результате многократного заложения новых прослоек перидермы и образуется третичная покровная ткань - ритидом. Живые клетки, попавшие между этими прослойками, гибнут. Таким образом, корка представляет собой сложный тканевый комплекс, состоящий из чередующихся слоев пробки и других отмерших тканей. Мертвые ткани, составляющие корку, растрескиваются под напором нарастающих изнутри тканей, поэтому в отличие от гладкой пробки, корка характеризуется трещиноватостью. Особенно ярко граница между гладкой перидермой (берестой) и трещиноватой коркой выражена при основании стволов березы. Грубая, нередко толстая корка надежно предохраняет стволы деревьев от механических повреждений, резкой смены температур и даже лесных пожаров. Механические ткани Высшие растения ведут прикрепленный образ жизни, поэтому способность противостоять нагрузкам имеет для них особое значение. Некоторые растения вынуждены десятки и даже сотни лет оказывать сопротивление бурям, ливням, граду и снегу. Любая растительная клетка (за редким исключением) имеет наружный скелет в виде целлюлозной оболочки. Поэтому, весьма примечательно, сто в обеспечении прочности тела растения нет резкого разграничения между различными тканями. В то время как работа фотосинтеза может выполняться лишь специализированными клетками, снабженными хлоропластами, функцию укрепления растительного тела в той или иной мере выполняют все клетки и ткани, живые и мертвые. Еще раз повторю, что в обеспечении прочности растения принимают участие все клетки и ткани. Однако если для небольших многоклеточных организмов (особенно погруженных в воду) наличие тонких оболочек у каждой из клеток оказывается вполне достаточным для обеспечения прочности и поддержания формы, то для крупных наземных растений такая опорная система явно недостаточна и у них возникли специализированные механические ткани, которые даже после отмирания живого содержимого продолжают выполнять опорную функцию. Тело растения в целом представляется чем-то вроде железобетонной конструкции, где механические ткани соответствуют арматуре (тяжам проволоки и стержням), а остальные ткани играют роль наполнителя (бетона). Именно поэтому Ф.М. Раздорский считал более правильным называть механические ткани ≈ арматурными. Различают два основных типа механических тканей: колленхиму и склеренхиму. Колленхима Колленхима (греч. кола - клей), которую рассматривают как опорную ткань, состоит из толстостенных клеток. Она очень близка к паренхиме. Клетки обеих тканей содержат протопласты со всеми органеллами. Для той и другой ткани характерны первичные и нелигнифицированные оболочки клеток. Отличие между этими тканями состоит главным образом в том, что у колленхимы оболочки толще, а клетки прозенхимные вытянуты в длину и имеют скошенные концы. Кроме того, характернейшая особенность клеток колленхимы состоит в том, что их оболочки неравномерно утолщены, что придает этой ткани на поперечном срезе очень своеобразный вид. В зависимости от характера утолщения различают три основных типа колленхимы: 1) уголковую; 2) пластинчатую и 3) рыхлую. В клетках уголковой колленхимы оболочка сильно утолщается в углах, где соприкасается несколько клеток. Пластинчатая колленхима, менее распространенная, характеризуется тем, что утолщенные части оболочек расположены параллельными слоями, а сами клетки на поперечном срезе имеют форму прямоугольников. Сравнительно редко встречается рыхлая колленхима. Строение этой ткани усложняется тем, что клетки на ранних стадиях развития разъединяются в углах с образованием межклетника. Оболочки же утолщаются именно в местах примыкающих к межклетнику. Колленхима приспособлена прежде всего для выполнения функции опоры растущих листьев и стеблей. Колленхима появляется на ранних этапах развития побега. Ее оболочки пластичны и способны к растяжению. Поэтому она не препятствует удлинению органа. Если бы в это время возникали жесткие ткани, неспособные к растяжению, то и удлинение органов было бы невозможным. Кроме того, колленхима способна не только пассивно растягиваться, но и активно расти, благодаря сохранению в клетках живого содержимого. Одна из особенностей колленхимы состоит в том, что она выполняет свое назначение только в состоянии тургора. Если молодые побеги теряют воду, тонкие участки оболочек складываются "гармошкой" и побеги увядают, то есть теряют упругость, обвисают. Итак, колленхима живая ткань, состоящая из вытянутых клеток с неравномерно утолщенными стенками, способная растягиваться и выполняющая свои функции лишь в состоянии тургора клеток. Для колленхимы характерно периферическое положение в побеге. Она располагается либо непосредственно под эпидермой, либо на расстоянии одного или нескольких слоев от нее. В стеблях колленхима чаще образует сплошной кольцевой слой. Она встречается иногда и в форме тяжей в ребрах травянистых стеблей. Склеренхима Склеренхима ≈ это наиболее распространенный в растительном царстве тип механической ткани. Как и колленхима, она состоит из вытянутых прозенхимных заостренных на концах клеток, но в остальном заметно от нее отличается. Типичная склеренхима состоит из равномерно утолщенных, плотно сомкнутых клеток. Сформировавшись, клетки склеренхимы обычно теряют живое содержимое и их полости заполняются воздухом; клеточные стенки к этому времени в большинстве случаев одревесневают; сравнительно редко они остаются чисто целлюлозными. Таким образом, склеренхима выполняет свою функцию уже после отмирания протопластов. Материал клеточных стенок склеренхимы обладает высокой прочностью и упругостью. По прочности на разрыв он близок к строительной стали, а по упругости даже превышает ее. Склеренхима значительно превосходит сталь в способности противостоять динамическим (ударным) нагрузкам без деформации. Склеренхима имеется в вегетативных органах почти всех сосудистых растений. Ее нет или она слабо развита в погруженных в воду органах. Обычно склеренхимные клетки подразделяются на две группы: 1) волокна и 2) склереиды. Волокна представляют собой длинные узкие прозенхимные клетки, длина которых во много раз превышает ширину. Обычно волокна имеют толстые стенки и очень узкую полость. Прочность стенок повышается еще и оттого, что фибриллы целлюлозы проходят в них винтообразно, а направление витков в слоях чередуется. Волокна вторичного происхождения отмечаются специальными терминами. Наружу от камбия в лубе формируются так называемые лубяные волокна, а внутрь от камбия в древесине откладываются древесинные волокна или либриформ. Волокна могут размещаться в виде сплошного кольца, отдельными тяжами и даже поодиночке. В практике лубяные пучки называют техническим волокном. Лубяные волокна некоторых растений широко используются в промышленности. Прочность и сырьевое качество технического волокна зависят от степени сцепления лубяных клеток между собой и плотности соединения их в пучки, а также от длины волокна. Текстильные ткани из длинных тонких и извитых волокон имеют большую прочность. В производстве особенно ценны те редкие растения, у которых волокна не одревесневают, например, лен. Назову лишь наиболее известные волокнистые растения и изделия из них изготавливаемые: конопля (Cannabis sativa) ≈ канаты и веревки; джут (Corchorus capsularis) ≈ канаты, веревки и грубые ткани; кенаф (Hibiscus cannabinus) ≈ грубые ткани; лен (Linum usitatissimum) ≈ тканию; рами (Bochmeria nivea) ≈ ткани. Склереиды Склереиды не имеют форму волокон и сильно варьируют по форме. Обычно по форме клеток их и классифицируют. Наиболее распространенными являются изодиаметрические округлые склереиды, называемые брахисклереидами или каменистыми клетками. Брахисклереиды встречаются в скорлупе плодов лещины, желудя; в косточках плодов сливовых, грецкого ореха; в мякоти плодов груши, айвы; в кожуре семян кедровой сосны. Брахисклереиды встречаются и в подземных органах ≈ в коре корневищ пионов, в корнях хрена, клубнях георгина. Остеосклереиды имеют длинноцилиндрическую форму с расширениями на концах, напоминающими сочленовые головки костей. Астросклереиды или звездчатые склереиды имеют ветвистую форму. Чаще всего астросклереиды встречаются в листьях кожистой консистенции. Они придают листовым пластинках дополнительную прочность. Склереиды могут образовывать сплошные группы, тканевую массу, как в скорлупе плодов. Могут они встречаться и поодиночке, в виде идиобластов, как, например, в листьях. Организация тела растения с точки зрения строительно-механических принципов Прикрепленные к месту растения не имеют возможности укрываться от действия стихий. При этом растения нередко обладают очень крупными размерами и почти всегда ≈ огромной наружной поверхностью. Они подвергаются энергичному воздействию разнообразных механических нагрузок. К тому же многие нагрузки существуют десятки и сотни лет, то есть являются многократными. Поэтому растения обладают поразительной способностью противостоять различного рода механическим нагрузкам. Тонкая соломина поддерживает тяжелый колос и листья, раскачивается при порывах ветра и не ломается. Эта ржаная соломина долге время вызывала изумление ботаников. Ее сравнивали то с Эйфелевой башней, то с высокими дымовыми трубами. У соломины высота в 500 раз превышает диаметр при основании, а инженерные сооружения по этому показателю заметно проигрывают. Целесообразность строения растений с точки зрения механики пытался объяснить еще Галилей. Его интересовал вопрос, как меняются пропорции тела организмов при значительном изменении их размеров. Много внимания этим вопросам уделяли Грю и Гук. Однако лишь два века спустя в 1874г немецкий ботаник Швенденер подробно рассмотрел распределение механических тканей в теле растения с точки зрения инженерной теории сопротивления материалов. Было выдвинуто положение, утверждающее, что органы растений строятся в соответствии с принципами достижения прочности при экономной затрате материала. Рассмотрим это положение более подробно. Если стержень, испытываемый на прочность, положить на две опоры и нагрузить, то он прогнется. При этом его нижняя сторона будет растягиваться, то есть противодействовать разрыву. Наоборот, верхняя сторона будет противодействовать раздавливанию и сжиматься. Материал же находящийся в центре стержня останется в этом отношении нейтральным. Таким образом, с точки зрения инженерным расчетов материал целесообразно сосредоточить в верхней и нижней частях стержней, где он будет выполнять наибольшую нагрузку. В центре же с целью экономии употребить лишь в той мере, чтобы предотвратить сжатие конструкции в поперечном направлении. В соответствии с этими принципами инженеры установили наиболее экономичную и целесообразную конструкцию в виде двутавровой балки, применяемой для перекрытий. Швенденер показал, что в листьях растений механические ткани по расположению очень напоминают двутавровые балки. Стебель подвергается изгибам в различных направлениях и его можно сравнить с вертикальной трубой. В целях наибольшей экономии и прочности механические элементы должны быть отнесены к периферии такой конструкции. Действительно, в стеблях колленхима и склеренхима чаще всего располагаются непосредственно под эпидермой или близко к поверхности. Центр стебля обычно занят тонкостенной паренхимой или даже имеет обширную полость. Соломина злаков является типичной полой трубой. Корню, окруженному почвой, не грозит опасность изгиба и излома. Корень выполняет другую механическую задачу, он "заякоривает" растения в почве и противодействует напряжениям, стремящимся выдернуть его, то есть противодействует разрыву. В соответствие с этим, механические ткани размещаются в центре корня. Существенный вклад в рассматриваемую проблему внес отечественные ботаник В.Ф.Раздорский. Прежде всего Раздорский показал, что рассмотренные нами принципы распределения механических тканей в различных органах растения реализуются не столь однозначно. Дело в том, что растения повергаются воздействию механических нагрузок 2-х категорий, предъявляющих к ним совершенно противоположные требования, нагрузкам: 1) статическим и 2) динамическим. К первым относится воздействие силы тяжести, собственного веса. Иногда статические нагрузки заметно увеличиваются, например, при оседании снега в ветвях крон. К механическим факторам динамического (ударного) рода можно отнести порывы ветра, удары капель дождя и града. Схема, предложенная Швенедером, в большей мере соответствует противодействию статическим нагрузкам. Однако, с точки зрения динамики, как показал Раздорский, органы растения должны работать подобно пружинам изгиба, способным всякий раз возвращаться в исходное состояние после снятия нагрузки. Поэтому ствол дерева, раскачивающийся под тяжестью кроны, имеет конструкцию не полой жесткой трубы, а сплошной упругой пружины. Кроме того, "инженерные требования" растений заметно меняются в ходе онтогенеза. У молодого проростка отчетливо проявляется периферическая тенденция развития механических тканей. В этой стадии растение тянется к свету и его задача возможно быстрее достигнуть максимальной высоты с возможной экономией материала. Но, выбравшись к свету, растение начинает ветвиться и подвергается более сильному воздействию ветра и других динамических факторов. Соответственно меняется и распределение механических тканей: в большей степени укрепляется центр. Раздорский же впервые обратил внимание на сходство растений с так называемыми комплексными сооружениями, к разряду которых главным образом относятся железобетонные конструкции. В органах растения бетону соответствует основная масса мягких и тонкостенных тканей, а каркасу (арматуре) ≈ механические тяжи. Но растительные конструкции стоят на более высокой ступени, чем технические сооружения. Бетон разрушается уже при незначительном удлинении в 0,01%. Поэтому в железобетоне высокие механические качества железа используются далеко не полностью. В растительных конструкциях основная масса способна сильно деформироваться, ткани заполнения могут разрушиться лишь после разрыва тяжей арматуры. Поэтому в органах растений сопротивление арматуры используется полностью. Кроме того, железо обладает свойством текучести, то есть железный прут либо вытягивается, либо пучится при сжатии, без возврата в исходное состояние. Растительная арматура текучестью не обладает. Однако природа всегда многообразнее, чем наши суждения о ней. Некоторые растения строятся как бы наперекор инженерным принципам. Например, южно-африканские клейнни (Kleinia - из семейства сложноцветных) имеют стебли тонкие у основания и утолщающиеся на вершине. В результате ветви отламываются при малейшем ветре. Отломившиеся побеги служат для вегетативного размножения. Лекция 6. Проводящие ткани Ксилема (древесина) Трахеальные элементы Флоэма (луб) Проводящие пучки Возникновение покровной ткани ≈ эпидермы было важным, но не единственным требованием при переходе растений к обитанию в наземных условиях. Растения столкнулись с проблемой обеспечения водного режима. Решение этой проблемы привело к появлению проводящих тканей, специально приспособленных к выполнению функции водопроведения. Дело в том, что проведение воды из одной живой клетки в другую осуществляется крайне медленно и требует определенных энергетических затрат. Рациональнее всего такая организация водопроводящей системы, при которой она состоит из мертвых клеток. Однако мертвая клетка может сохраняться среди живых тургоресцентных клеток только в том случае, если она будет достаточно механически укреплена. На суше тело растения оказалось расчлененным на две части: надземную и подземную (корень и побег), обеспечивающие почвенное и воздушное питание. Соответственно возникли и две проводящие ткани, по которым вещества передвигаются в 2-х противоположных направлениях. Восходящий ток воды и минеральных солей из корней в стебли и листья обслуживается ксилемой; Нисходящий ток пластических веществ из листьев обеспечивается флоэмой. Однако резкого разграничения между флоэмой и ксилемой в плане функционирования нет. Так весной при возобновлении роста и развития почек ксилема доставляет в них запасные вещества. Это так называемая "пасока" богатая сахарами. По флоэме же часть пластических веществ движется вверх в созревающие плоды и семена. Ксилема и флоэма имеют ряд общих особенностей: прежде всего, проводящие элементы и ксилемы и флоэмы имеют удлиненную форму и вытянуты по направлению тока веществ; поперечные стенки проводящих клеток имеют поры или сквозные отверстия ≈ перфорации и практически не препятствуют продвижению жидкости; в рабочем состоянии проводящие элементы лишены живого содержимого ≈ протопласта, который затруднял бы передвижение веществ, или имеют особый проницаемый протопласт. ксилема и флоэма представляют собой сложные ткани, поскольку в их состав входят разнородные элементы: проводящие, механические, запасающие. и, наконец, обычно ксилема и флоэма пространственно сопряжены между собой в так называемые проводящие пучки. Ксилема (древесина) Как сложная ткань ксилема состоит из нескольких элементов: трахеальные элементы ≈ выполняют проводящую функцию; древесные волокна (или волокна либриформа) ≈ обеспечивают опорную (мех) функцию; паренхимные клетки ≈ осуществляют запасание и передвижение пластических веществ. Трахеальные элементы Трахеальные элементы представляют собой наиболее высокоспециализированные клетки ксилемы. Как правило, они вытянуты в длину и в зрелом состоянии мертвы. Для них характерны лигнифицированные оболочки со вторичными утолщениями и порами. Процесс отложения вторичной оболочки и пропитывание ее лигнином осуществляется еще в живой клетке. При этом на первых этапах развития растения все клетки растущих частей удлиняются (вытягиваются). Однако такое удлинение было бы невозможным при сплошной жесткой оболочке. В этом отношении у высших растений выработалось оптимальное приспособление: вторичная оболочка не одевает клетку сплошь, а расположена кольцами или спиралью. Подобные кольчатые и спиральные утолщения позволяют молодым трахеальным элементам вытягиваться в длину и в то же время препятствуют из сдавливанию. Кроме того, этот тип вторичного утолщения является чрезвычайно экономичным. При всех достоинствах кольчатых и спиральных элементов как путей для проведения воды, их механическая прочность оставляет желать лучшего. Поэтому, как только у молодого растения заканчиваются ростовые процессы, связанные с удлинением клеток, в ксилеме начинают формироваться трахеальные элементы со сплошной вторичной одревесневшей оболочкой. Когда формирование сплошной оболочки заканчивается, клетки очень быстро отмирают. Однако и в этом случае оболочка водопроводящего элемента не может быть совершенно сплошной. Обычно в ней имеются многочисленные углубления в виде пор. Поэтому зрелые водопровдящие элементы называют точечно-поровыми. Итак, в процессе онтогенеза (индивидуального развития растения) наблюдается ряд взаимопревращения трахеальных элементов: кольчатые, спиральные, сетчатые, лестничные, точечно-поровые. Различают два типа проводящих элементов: 1) трахеиды и 2) членики сосудов. Отличаются они главным образом тем, что членики сосудов имеют сквозные отверстия ≈ перфорации, в то время как трахеиды являются неперфорированными элементами. Поэтому по сосудам растворы продвигаются значительно легче, чем по трахеидам. Трахеиды ≈ это основной водопроводящий элемент высших растений с момента их появления на суше и по сей день. Трахеиды имеют замкнутую со всех сторон первичную оболочку. Поэтому вода по трахеидам должна проходить через окаймленные поры, просачиваясь через первичные оболочки и склеивающий их межклеточный слой. Понятно, что подобная структура не является оптимальной для быстрой подачи воды. Поэтому в процессе эволюции в ряде групп высших растений возник новый более совершенный проводящий элемент ≈ членик сосуда. Для члеников сосудов характерно наличие в концах клеток перфораций. Соединяясь между собой перфорациями, сотни и тысячи члеников образуют сосуды ≈ длинные сквозные трубки, по которым вода перемещается практически беспрепятственно. Водопроводящие элементы, обладающие достаточно толстой оболочкой, инкрустированной лигнином, играют в теле растения также и механическую роль. Поэтому специализация трахеальных элементов в процессе эволюции шла в направлении разделения механической и проводящей функций. Примитивная древесина большинства хвощей, папоротников, голосеменных имеет гомогенную (однородную) ксилему и состоит исключительно из трахеид и небольшого количества древесинной паренхимы. В такой древесине узкопросветные толстостенные трахеиды выполняют преимущественно механическую функцию, а широкопросветные и тонкостенные ≈ водопроводящую. Более совершенная гетерогенная древесина покрытосеменных состоит из сосудов, трахеид, волокон - либриформа и запасающей паренхимы. Членики сосудов у цветковых растений морфологически довольно разнообразны и образуют четкий эволюционный ряд. Этот эволюционный ряд, построенный на работах выдающегося американского ботаника И. Бэйли, помещается во всех учебниках, как одна из ярких и бесспорных демонстраций эволюционных преобразований. Эволюционный ряд члеников сосудов начинается с длинных трахеид, имеющих лестничную поровость и косые заостренные концы. Постепенно клетки укорачиваются, становятся более широкими, а их концевые стенки все менее наклонными и в конце концов оказываются поперечными. В наиболее примитивных члениках лестничная перфорационная пластинка имеет множество перегородок. В процессе эволюции они сокращаются и исчезают совсем, так что остается одна сквозная крупная перфорация. Характерно, что с появлением сосудов цветковых растений не исчезли трахеиды, поскольку совершенный водопроводящий аппарат не всегда является преимуществом. Так, в условиях высокой влажности и затенения способность быстро проводить воду не имеет особого значения. У таких растений процент трахеид и примитивных члеников сосудов с лестничной перфорацией заметно выше, чем у растений, произрастающих в засушливых условиях. Получается, что соотношение проводящих элементов четко обусловлено экологическими условиями и определяет водный баланс растения. Мы уже говорили, что сочетание в одном клеточном элементе ≈ трахеиде ≈ и способности проводить воду и механической прочности было важным эволюционным приспособлением. Появление сосудов, с их все увеличивающейся полостью и относительно тонкими стенками, несколько ослабило бы механическую роль ксилемы, если сосуды остались бы единственным элементом этой ткани. В связи с этим наметился второй эволюционный ряд специализации в сторону повышения механической прочности, и трахеиды, через промежуточную стадию волокнистой трахеиды превратились в волокна либриформа. При этом происходило утолщение оболочек клеток, сужение полостей, все большая редукция окаймления пор. Поры у волокон либриформа стали узкие, щелевидные. Вместе с тем, сократилось и количество пор. Паренхимные клетки, входящие в состав древесины, запасают крахмал, жиры и многие другие эргастические вещества. В ходе онтогенеза у растения из первичной латеральной меристемы прокамбия появляется первичная ксилема. У некоторых растений со временем начинает работать вторичная боковая меристема ≈ камбий, дающая начало вторичной ксилеме. Обычно первичная ксилема отчетливо подразделяется на два структурных типа: 1) протоксилему и 2) метаксилему. Протоксилема возникает несколько раньше. Это первая стадия развития первичной ксилемы. Обычно протоксилема содержит только трахеальные элементы, погруженные в паренхиму, причем с кольчатыми утолщениями оболочек: в это время клетки еще растягиваются. Метаксилема более сложная по составу ткань. В ней помимо трахеальных элементов, могут содержаться и волокна, а утолщения оболочек более мощные. Граница между этими возрастными стадиями проводится весьма приблизительно. Флоэма (луб) Флоэма ≈ ткань сосудистых растений, проводящая пластические вещества, образованные в результате процесса фотосинтеза в зеленых клетках листа, от кроны вниз, к корням, а также вверх, к цветам и плодам. Подобно ксилеме, флоэма состоит из нескольких типов клеток и по времени своего возникновения может подразделяться на первичную и вторичную. Первичная флоэма возникает из прокамбия. Вторичная флоэма происходит из камбия. Первичная и вторичная флоэма содержит одинаковые типы клеток: ситовидные элементы, которые представлены либо ситовидными клетками, либо члениками ситовидных трубок с клетками спутницами ≈ обеспечивают дальний транспорт пластических веществ; склеренхимные клетки 2-х типов: волокна и склереиды ≈ несут опорную функцию; разнообразные паренхимные клетки запасают и транспортируют пластические вещества в радиальном направлении. Так же как и элементы ксилемы, элементы флоэмы в процессе эволюции высших растений претерпели целый ряд значительных морфологических и функциональных изменений. Однако надо подчеркнуть, что если водопроводящие элементы явились новым приобретением у первых высших растений, осуществивших выход на сушу, то элементы, аналогичные флоэме, уже имелись у некоторых водорослей, и, таким образом, в эволюционном плане флоэма значительно старше, чем ксилема. Более того, можно сказать, что основной физиологический признак клеток флоэмы ≈ их способность проводить растворенные органические вещества ≈ в той или иной степени присущ любой живой клетке. И, следовательно, механизмы, обеспечивающие эту способность, возникли на самых первых этапах развития жизни на Земле. Само собой разумеется, что эта способность была значительно развита и улучшена. Рассматривая эволюцию флоэмы, нужно отметить, что тонкие целлюлозные оболочки ее клеток несравненно хуже сохраняются в ископаемом состоянии, чем одревесневшие клетки ксилемы. Менее долговечна флоэма и в онтогенезе растения. Обычно новые слои флоэмы достаточно быстро сплющиваются и погибают под напором нарастающих внутренних тканей. В силу плохой сохранности наши сведения о последовательности этапов эволюции флоэмы гораздо более отрывочны, чем для ксилемы. Но даже по имеющимся данным можно утверждать, что у самых первых наземных растений ≈ риниофитов ≈ имелись клетки вытянутой формы подобные флоэме. Уже эти клетки имели на стенках скопления своеобразно устроенных отверстий, образующих ситечко, или ситовидное поле, по более строгой терминологии. Итак, межклеточный контакт ситовидных элементов осуществляется при посредстве ситовидных полей. Ситовидные поля ≈ это участки оболочки, пронизанные многочисленными отверстиями ≈ перфорациями. По сути, это скопление плазмодесм, тончайших тяжей цитоплазмы, пронизывающих клеточную оболочку и связывающих протопласты соседних клеток. В ситовидных пластинках с крупными отверстиями плазмодесмы сливаются в единый цитоплазменный пояс, образуя гигантскую плазмодесму. В классическом случае каждое отверстие ситовидного поля облицовано особым углеводом ≈ каллозой, которая покрывает цитоплазменный тяж чехлом. Кроме того, у многих растений каллоза окружает небольшим валиком отверстия канала. Чем старше ситовидный элемент, тем больше в нем накапливается каллозы. По мере того, как ситовидный элемент переходит в состояние покоя, каллоза закупоривает перфорации и образует подушечку над отверстиями ситовидных полей. Детальное исследование показало, что: 1) клетки, находящиеся в функционально активном состоянии, содержат очень мало каллозы или не содержат ее вообще; 2) каллоза исключительно быстро синтезируется в ответ на поранения и при окончании функционирования ситовидного элемента. Различают два типа ситовидных элементов. Более примитивные элементы присущи споровым и голосеменным растениям. Они называются ситовидными клетками. Ситовидные клетки заметно вытянуты в длину и обладают заостренными концами, ситовидные поля рассеяны по боковым стенкам. Для цветковых растений характерны ситовидные трубки, состоящие из члеников и своеобразных сопровождающих клеток ≈ клеток-спутниц. Членики ситовидных трубок имеют крупные, хорошо выраженные перфорации, располагающиеся на их концах, конечные стенки становятся практически поперечными или слегка скошены. Таким образом, последовательность эволюционных изменений в ситовидных элементах во многих отношениях сходна с эволюцией члеников сосудов ксилемы. Специализация сопровождалась сокращением числа ситовидных полей, увеличением размеров перфораций и уменьшением угла наклона концевых стенок. Чрезвычайно важным признаком организации ситовидных элементов является отсутствие в этих клетках клеточного ядра при весьма высокой жизненной активности. Ситовидные элементы в этом отношении представляют собой уникальное явление среди всех остальных клеток. Впрочем, полное отсутствие ядра отмечено только у цветковых растений, тогда как ситовидные элементы голосеменных и других высших растений в большинстве случаев сохраняют ядро, но, как правило, в полуразрушенном состоянии. Такие сморщенные ядра почти всегда лишены ядрышка и называются некротическими. В противоположность трахеальным ситовидные элементы в зрелом состоянии сохраняют живые протопласты. Внутренняя организация протопласта также не лишена своеобразия. Как правило, в зрелых ситовидных элементах рибосомы полностью отсутствуют, аппарат Гольджи отмечается только на первых стадиях их формирования. Напротив, хорошо развиты митохондрии, пластиды, эндоплазматическая сеть. Вакуоль в этих клетках также не выражена, поэтому цитоплазма находится в разжиженном состоянии. Ситовидные элементы обычно формируют своеобразные белковые включения, которые прежде назывались "слизистыми тельцами". Теперь их принято называть флоэмным белком или просто Ф-белком. Этот белок имеет вид нитей трубчатой фоэмы со спиральным расположением субъединиц, по своей структуре он напоминает белок мышц животных. Предполагают, что Ф-белок вместе с раневой каллозой участвует в закупорке перфораций при повреждении ткани, предотвращая тем самым вытекание содержимого из ситовидных трубок. Кроме того, высказывается предположение, что Ф-белок, обладающий сократительной реакцией, гонит вещества вдоль по ситовидной клетке. Отсутствие или полная некротизация ядра в ситовидных элементах находится в разительном противоречии с интенсивной физиологической активностью этих клеток. Поэтому широко распространено мнение, что управление всей жизнедеятельностью ситовидных элементов возложено на ядра соседних клеток. Так, у цветковых растений членики и связанные с ними клетки-спутники возникают из одной материнской клетки в результате ее митотического деления. Таким образом, эти клетки являются сестринскими и имеют общее происхождение. С каждым ситовидным элементом может быть связана одна или несколько клеток-спутниц. Предположение о, своего рода, "дистанционном" управлении деятельностью ситовидных элементов со стороны ядра клетки-спутницы широко распространено, но надо признать, что оно не имеет прямого экспериментального подтверждения. Оно скорее основано на убеждении большинства биологов о решающем значении ядра в жизнедеятельности клетки. Предполагают, что клетки-спутницы регулируют флоэмный транспорт. Кстати и механизм флоэмного транспорта представляет сам по себе большую загадку и сложную проблему. Если движение воды по ксилеме понятно и хорошо объясняется с позиций гидродинамики (вода поднимается по трубкам и капиллярам), то движение веществ по живым клеткам флоэмы невозможно объяснить простой диффузией. Дело в том, что растворы сахаров и аминокислот передвигаются по флоэме со значительно более высокой скоростью (около 1 метра в час). Для объяснения механизмов флоэмного транспорта выдвигалось множество гипотез. Одна из них ≈ гипотеза тока под давлением, предложенная еще в 1927 г. немецким физиологом Эрнстом Мюнхом, удовлетворительно объясняет экспериментальные данные и в настоящее время поддерживается многими учеными. Это объяснение является самым простым, поскольку оно основывается исключительно на осмосе, как на движущей силе транспорта ассимилятов. В кратком изложении эта гипотеза утверждает, что ассимиляты транспортируются к месту потребления по градиенту тургорного давления, то есть по градиенту концентрации. Градиент концентрации создается путем активной загрузки ситовидных трубок сахарозой, при этом вода за счет осмоса свободно поступает в эти клетки. Метаболическая энергия, необходимая для загрузки и разгрузки, поставляется клетками-спутницами. Следовательно, флоэма является единой системой, состоящей из проводящих элементов вдоль всего пути перемещения сахаров от их источников к местам потребления. Паренхимные клетки являются постоянными компонентами флоэмы и выполняют типичную запасающую функцию. Кроме того, паренхимные клетки, расположенные рядом с ситовидными элементами, могут участвовать в загрузке и разгрузке проводящих элементов. Проводящие пучки Обычно проводящие ткани ≈ ксилема и флоэма ≈ образуют в теле растения тяжи, называемые проводящими пучками. Нередко к проводящим пучкам примыкает механическая ткань-склеренхима, такие комбинированные пучки называются сосудисто-волокнистыми или армированными. По способности ко вторичному утолщению проводящие пучки делят на открытые и закрытые. В открытых пучках закладывается камбий и они открыты для дальнейшего роста (разрастание в толщину); закрытые пучки, не имеющие камбия, закрыты для вторичного утолщения. Неполные проводящие пучки содержат или только ксилему или только флоэму. Обычно они очень малы по размерам и встречаются сравнительно редко. Наиболее распространенными являются полные пучки, включающие в свой состав обе проводящих ткани, и флоэму и ксилему. Полные пучки весьма разнообразны по конструкции. 1) в коллатеральных пучках флоэма залегает над ксилемой: 2) биколлатеральные пучки обладают дополнительным солем флоэмы: 3) в концентрических пучках один компонент окружает другой: в амфивазальных ксилема окружает флоэму в амфикрибральных, наоборот, флоэма окружает ксилему. Лекция 7. Дифференциация тела растения в связи с выходом на сушу. Возникновение органов Строение тела растения. Основные органы Корень Строение молодого корневого окончания Первичное строение корня Вторичное строение корня Корневые системы В 1859 году канадский геолог Джеймс Досон в девонских отложениях на полуострове Гаспе в Канаде обнаружил остатки удивительно примитивного высшего растения не похожего ни на одно из известных науке в то время. Его вильчато разветвленные стебли были лишены листьев, а проводящая система представляла собой типичную протостелу, а это, как мы узнаем, был самый примитивный тип организации проводящей системы. Досон дал своему растению название Psilophyton princeps, что можно перевести на русский язык как "голорос первичный". Позднее в Шотландии были найдены и другие, еще более примитивные растения: риния, хорнеофит и куксония. Эти древние растения были выделены в особый отдел, для обозначения которого прежде употреблялось название "псилофиты" (Psilophyta), но в настоящее время многие ученые предпочитают название "риниофиты" (Rhiniophyta). Остатки самых древних сухопутных растений были найдены в породах раннедевонского возраста, образовавшихся более 415 млн. лет назад. Породы состоят из окаменелых слоев торфа, который подвергался периодическим затоплениям. Вертикально стоящие растения заносились слоями песка прямо на месте произрастания. Благодаря тому, что в процессе захоронения растения эти пропитывались раствором кремнекислоты, сохранилась не только их внешняя форма, но и тончайшие детали внутреннего строения. Так что, мы имеем возможность воссоздать морфологическую эволюцию высших растений с большой достоверностью. Надо отметить, что уже и до этого сушу населяли различные виды бактерий, водорослей и грибов. Но эти организмы в своей жизнедеятельности были полностью зависимы от наличия влаги в окружающей среде. Если наступала засуха ≈ они впадали в состояние анабиоза. Такие растения можно назвать пойкилогидрическими, по аналогии с пойкилотермными (холоднокровными) животными, неспособными регулировать температуру своего тела. Высшие растения приобрели способность регулировать водный режим своего тела независимо от содержания влаги в почве и в воздухе. Поэтому их можно назвать гомойогидрическими. Уже самые примитивные из известных нам высших растений были дифференцированы на элементарные органы, хотя и напоминали больше по внешнему виду своих водорослевых предков. Выход на сушу потребовал приспособления к совершенно новым условиям. Тело растения оказалось разделенным на две части: надземную и подземную. Одни ответвления, называемые теломами, поднимались вертикально, другие прижимались к поверхности почвы, поглощая воду и минеральные соли. Последняя задача была не слишком трудной, учитывая то, что риниофиты обитали в заболоченных переувлажненных местах и на мелководьях. Корневищеподобные веточки риниофитов получили название ≈ ризомоидов, они были покрыты волосовидными ризоидами и явились прототипом корней. В ходе дальнейшей эволюции, по мере освоения более сухих почв, происходила более глубокая специализация подземных органов. Увеличивалась их поглощающая способность, благодаря обильному ветвлению и возникновению волосконосного слоя. Защиту апикальной меристемы в плотной почве обеспечило возникновение чехлика. Боковые ответвления корня стали закладываться изнутри (эндогенно) и не мешали продвижению в почве. Образование настоящих корней у высших растений было крупным эволюционным событием. Одновременно с обособлением корней, надземная часть ≈ телома риниофитов, преобразовалась в побег, расчлененный на стебли и листья. В качестве листьев дифференцировались выросты на поверхности телома либо его ответвления. Образование листьев как аппаратов транспирации и фотосинтеза явилось важным приспособлением к надземному образу жизни. Это был новый прогрессивный признак общей структуры растительных организмов. Морфогенез листьев шел двумя путями, приводившими к формированию или микрофиллов (малых листьев) или макрофиллов (крупных листьев) (микрофильная и макрофильная линии эволюции). Микрофиллы возникали как выросты (энанции) на поверхности оси телома. Макрофиллы ≈ это своеобразные плосковетки, видоизмененные и в результате специализации уплощенные ответвления телома (то есть возникли в результате уплощения боковых веточек и целых систем ветвления). В процессе эволюции совершенствовалась и система ветвления телома. У самых примитивных высших растений телом ветвился вильчато (дихотомически), причем ветвление было равнодихотомическим. Путем неравномерного роста двух развилок, из которых одна растет сильнее (этот процесс был назван перевершиниванием) выработались более совершенные системы ветвления ≈ неравнодихотомическая, моноподиальная и симподиальная. При моноподиальном ветвлении более развитая главная ось совершенно выпрямляется. Но главная особенность ветвления этого типа заключается в том, что боковые ветви закладываются под верхушкой главной оси. При симподиальном ветвлении боковая ветвь перевершинивает главную, сдвигает ее в сторону и занимает ее место и так далее. Открытие древнейших растений заставило пересмотреть принципы организации тела растения. Строение тела растения. Основные органы Основной план строения тела растения в морфологии истолковывался по-разному. Ранее принималось, что тело растения состоит из нескольких "основных частей" или органов, ≈ корня, стебля, листа, цветка, семяпочек, волосков. Позднее число этих основных органов было сведено к трем (так называемая "железная триада" органов (кстати, она вошла во все школьные учебники) ≈ корень, стебель и лист. В настоящее время стебель и его придаточные органы рассматриваются как единое целое ≈ побег. Вопрос об эволюционном происхождении органов растения решался длительное время. Одни ученые считали для надземных органов первичным стебель, другие ≈ лист. И только открытие псилофитов позволило вполне однозначно утверждать, что у растений основных вегетативных органов два: корень и побег. Таким образом, строение тела высшего растения можно представить так: Корень Мы начинаем изучение органов растения именно с корня не случайно. Дело в том, что корни растений, в сравнение с побегом, существуют в более инертной, менее пластичной среде ≈ в почве, поэтому имеют и более простое строение; в то время, как побег развивается в нестабильной, динамичной воздушной среде. С другой стороны, о строении и функциях корня и о различных его модификациях мы знаем гораздо меньше, чем о стебле и листе и это, в первую очередь, связано с техническими трудностями, возникающими при изучении подземных органов. Что же такое корень с анатомо-морфологической точки зрения? Какими признаками его можно охарактеризовать? Первый (1) признак я уже называл: корень ≈ это один из двух основных вегетативных органов; 2) второе: как правило, корень обладает радиальной симметрией, причем характер симметрии более резко выражен в первичном строении; 3) корень ≈ открытая система роста, он, как и стебель, продолжительное время нарастает в длину за счет деятельности апикальной (верхушечной) меристемы, но 4) в отличие от стебля точка роста корня прикрыта чехликом, поэтому апикальная меристема работает на две стороны: наращивая корень и пополняя чехлик; 5) как и стебель, корень способен ветвиться, но боковые корни эндогенны, они закладываются внутри корня в особом слое - перицикле, в то время, как боковые выросты стебля имеют экзогенное происхождение; 6) корень совершенно безлистен, на нем никогда не развиваются листья, ни простые, ни метаморфизированные; 7) корень имеет особый всасывающий слой, состоящий из корневых волосков; 8) корень имеет особую опробковевшую внутреннюю кору ≈ эндодерму; 9) проводящие и механические ткани в корне сосредоточены в центре, а в стебле ≈ вынесены на периферию; 10) на поперечном срезе через корень первичного строения большую часть его объема занимает первичная кора, а меньшую часть стела (центральный осевой цилиндр); в стебле же все наоборот; 11) корень нарастает только верхушкой, в то время как для побега характерен еще и вставочный (интеркалярный) рост. Сразу следует оговориться, что ни один из этих признаков не выдерживается с абсолютной строгостью: то есть, встречаются, например, корни, не имеющие лучевой симметрии, или корни без чехлика; в некоторых случаях боковые корни развиваются экзогенно. И даже безлистность корня далеко не абсолютна, так у некоторых эпифитных орхидей корни принимают внешний облик листьев и фотосинтезируют, правда, сохраняя при этом ряд характерных для корней анатомоморфологических черт. Вместе с тем, иногда у органов побегового происхождения развиваются структуры присущие корню. Например, листовые черешки хохлатки полой покрыты волосками ничем не отличающимися от корневых. Отсутствие резких анатомо-морфологических различий между корнем и побегом становится понятным, если вспомнить их общее происхождение от примитивных осевых органов древнейших сухопутных растений. Корень можно охарактеризовать как орган и с позиции функциональной. В типичном случае функциями корней являются 1) всасывание воды с раствором минеральных веществ из почвы и подача их в стебель (функция питания); 2) закрепление, заякоривание растения в почве; кроме того, 3) в корнях синтезируются многие специфические аминокислоты, гормоны, алкалоиды, откладываются запасные вещества; 4) корни большинства растений взаимодействуют с грибами (микориза). Типичные корни имеют некоторые дополнительные физиологические особенности, отличающие их от стебля: 1) корень положительно (+) геотропичен, то есть растем вертикально вниз под действием силы тяжести; 2) отрицательно (-) фототропичен, это проявляется в том, что корни уклоняются от падающих лучей света в противоположную сторону и 3) положительно (+) гидротропичен, то есть, ориентирует свой рост в почве в сторону большей влажности. Переходя к изучению строения корня, прежде всего, рассмотрим строение молодого корневого окончания. Строение молодого корневого окончания В отличие от воздушных стеблей корням приходится прокладывать путь в твердой почве, поэтому апикальная меристема корней защищена специальным образованием, называемым корневым чехликом (лат. название caliptra). Наружные клетки чехлика выделяют обильную слизь, которая облегчает продвижение в почве, и активно слущиваются, причем будучи живыми, сами по себе, а не только при трении о почву. Поскольку клетки чехлика постоянно слущиваются, необходимо их регулярное пополнение, поэтому, в отличие от побега, апикальная меристема корня откладывает клетки не только внутрь, но и наружу (пополняя чехлик), то есть работает в двух взаимно-противоположных направлениях. Средняя продольная (или осевая) часть чехлика называется колумеллой (сolumella), она состоит из клеток, содержащих большое количество крахмальных зерен. Предполагают, что именно эта зона определяет положительную геотропическую реакцию корня. Крахмальные зерна колумеллы являются статолитами. При изменении положения корня в пространстве статолиты под действием силы тяжести пассивно перемещаются и надавливают на клеточные стенки, вызывая характерное возбуждение. Природа этого возбуждения пока не ясна. Однако экспериментально показано, что растения, у которых корневой чехлик удален, геотропическую реакцию утрачивают. Корневой чехлик в общем более мощно развит у корней сильно и долго растущих. Слабо развитый чехлик имеют многие сапрофиты, верески, тонкие корни которых расположены в торфе или рыхлой почве. Вовсе лишены чехлика корни многих водных растений, в том числе рясок, водокраса. Помимо чехлика, в молодом корневом окончании традиционно выделяется еще 4 зоны, хорошо вам известные. Зона деления. Зона растяжения. Зона поглощения или всасывания. Зона проведения или окончательной дифференциации тканей. Кроме традиционно выделяемых поперечных зон в молодом корневом окончании хорошо различимы продольные слои - гистогены. Гистогенами называют меристематические тяжи (слои), из которых развиваются определенные постоянные ткани. Всего в кончике корня выделяется 3 таких гистогена: 1) внутренний тяж называется плерома, из него в дальнейшем образуется центральный осевой цилиндр; 2) наружный многоклеточный тяж периблема, который дает начало первичной коре корня и, наконец, 3) внешний однорядный слой клеток дерматоген, из которого развивается ризодерма (всасывающая ткань корня). плерома центральный осевой цилиндр (стела) периблема первичная кора дерматоген ризодерма Однако и гистогены не возникают на пустом месте. Каким же образом закладываются эти продольные тяжи? Вспомним, что меристематические клетки неодинаковы по своей функциональной активности, некоторые из них способны делиться определенное, конечное число раз, другие, называющиеся инициальными, способны к неограниченному делению. У голосеменных и покрытосеменных растений инициали представлены комплексом клеток, расположенных несколькими слоями. Например, у цветковых растений различают два основных типа функционирования инициалей. У двудольных растений инициали расположены тремя слоями (тремя этажами). Каждый слой насчитывает по нескольку клеток. Верхний слой инициальных клеток дает начало внутреннему гистогену ≈ плероме; из среднего слоя инициалей формируется периблема; нижний слой инициалей пополняет с одной стороны дерматоген, с другой стороны - чехлик (калиптру). Поэтому у двудольных растений этот слой клеток называется дерматокалиптроген. На препаратах у двудольных растений чехлик просматривается неявно, он как бы сливается с наружным слоем - дерматогеном. Второй тип работы инициалей свойственен большинству однодольных растений. Здесь также имеется 3 слоя инициалей. Верхний - дает начало плероме; средний слой ≈ образует и периблему и дерматоген (правильнее будет сказать, что наружный слой периблемы дифференцируется в дерматоген); а вот инициали нижнего слоя дают начало только чехлику (калиптре). Именно поэтому у однодольных нижний слой инициалей называется калиптрогеном, а корневой чехлик у однодольных резко обособлен от основного тела корня. Если и говорят, что кончик корня как наперстком прикрыт чехликом, то это справедливо лишь по отношению к однодольным растениям. Таким образом, формирование постоянных тканей в корне является сложным многоступенчатым процессом: инициали порождают гистогены, а те, в свою очередь, дают начало постоянным тканям. инициали гистогены постоянные ткани Гистогенез или развитие первичных постоянных тканей корня удобнее всего проследить на серии поперечных срезов через корень, проведенных на разных уровнях. Раньше всего, еще в зоне деления появляется первичная ткань ризодерма, она достигает полного развития в зоне поглощения. Ризодерма неоднородна в своем строении. Выделяются клетки ризодермы, образующие корневые волоски, они называются трихобластами и отличаются от остальных клеток формой и величиной. Другие клетки лишены этой способности и называются атрихобластами. У стрелолиста и элодеи трихобласты, например, более крупные, а у водокраса более мелкие, чем основные клетки ризодермы. У многих водных растений, корни которых находятся в воде или в иле, корневые волоски вообще не развиваются. Среди таких растений можно назвать сусак зонтичный, водяную сосенку, белую кувшинку. У некоторых других (у желтой кубышки, веха ядовитого, аира) корни в воде лишены волосков, а корни, проникающие в грунт, их имеют. С другой стороны, многие типичные сухопутные растения имеют громадное количество корневых волосков. Подсчитано, что у одной особи 4-месячного растения ржи примерно 14 млрд. корневых волосков с площадью поглощения 400м2 и суммарной длиной более 10 000 км. Еще одна интересная особенность, корневые волоски обычно весьма слабо развиты у видов с экзотрофной (наружной) микоризой (сосна, бук), то есть, живущих в симбиозе с почвенными грибами. Здесь функцию волосков частично выполняют грибы. В любом случае, в функциональном отношении ризодерма представляет собой одну из важнейших тканей растения. Именно благодаря появлению этой ткани растения смогли широко расселиться по суше, хотя, как мы видим, некоторые водные формы вторично ее утратили. Микроскопическое строение ризодермы соответствует выполняемым функциям. Прежде всего, корневые волоски, в отличие от трихомов, не отделяются клеточной стенкой и являются выростами клеток ризодермы. Волоски имеют тонкую оболочку, состоящую из целлюлозы и пектиновых веществ, наружные слои оболочки выделяют слизь. Клетки ризодермы отличаются большой активностью, они избирательно поглощают элементы почвенного питания, а это требует больших энергетических затрат. Поэтому в протопласте волосков имеется большое количество рибосом и особенно митохондрий. При образовании волоска происходит выпячивание наружной стенки клетки, ядро перемещается в вырост и передвигается к кончику волоска. Здесь же находятся многочисленные диктиосомы, активно наращивающие оболочку. Когда рост волоска закончен, ядро отходит от кончика и занимает центральное положение. У некоторых растений, например, у эпифитных папоротников из рода Drymoglossum на о. Ява, наблюдается регенерация волосков. При длительном недостатке воды в субстрате протопласт с ядром перемещается в основание волоска, а оставшийся кончик отделяется перегородкой, высыхает и отмирает. При выпадении дождя основание волоска вновь разрастается. В зоне проведения, когда волоски отмирают, клетки ризодермы становятся обычным защитным слоем. Первичное строение корня Если разрезать корень несколько выше в зоне проведения, то уже на малом увеличении микроскопа хорошо выделяется две зоны. Широкая первичная кора, занимающая большую часть поперечного сечения среза и сравнительно узкий центральный осевой цилиндр. Такое соотношение тканевых комплексов ≈ характерная особенность корня, в стебле все подругому: большую часть поперечного сечения стебля занимает стела. Наружный слой первичной коры называется экзодермой. Он приходит на смену отмирающему и разрушающемуся волосконосному слою, остатки которого можно обнаружить на некоторых препаратах. Экзодерма - это первичная покровная ткань корня. Клетки экзодермы плотно сомкнуты и обычно опробковевают. У большинства растений экзодерма однослойна, хотя у отдельных растений, например, у ириса-касатика, она многослойна. Под экзодермой залегает мощный слой клеток паренхимы первичной коры. Обычно это тонкостенные клетки многогранной или округлой формы, расположенные без особого порядка. Между паренхимными клетками всегда имеются межклетники. По межклетникам циркулируют газы, необходимые для дыхания клеток корня. У некоторых водных и болотных растений система межклетников развита настолько сильно, что первичная кора превращается в аэренхиму. Таким образом, мощность воздухоносной системы напрямую зависит от аэрации субстрата. В паренхимных клетках коры протекает энергичный метаболизм. Поэтому нередко в них можно обнаружить друзы кристаллов, как отходы метаболизма, или запасные вещества, например, крахмальные зерна. Внутренний слой коры представлен эндодермой. Это своеобразная внутренняя пограничная ткань, отделяющая первичную кору корня от центрального осевого цилиндра. Эндодерма сплошным кольцом плотно сомкнутых клеток окружает стелу. У однодольных растений клетки эндодермы имеют весьма своеобразные подкововидно-утолщенные оболочки, пропитанные суберином. Жироподобное вещество - суберин непроницаемо для воды и газов, поэтому эндодерма прекрасно выполняет свою пограничную функцию. Однако изоляция стелы от первичной коры не может быть сплошной, поэтому среди клеток эндодермы обнаруживаются отдельные клетки, не имеющие характерных утолщений оболочки. Они работают как своеобразные ворота, по которым вещества могут перемещаться из коры в стелу и обратно и называются пропускными клетками. Теперь самая сложная часть корня ≈ центральный осевой цилиндр или стела. Сложная, потому что развитие проводящих тканей необходимо рассмотреть в динамике. Из гистогена - плеромы образуется две первичные меристемы - перицикл и прокамбий. Обычно перицикл тонким слоем окружает прокамбий. Перицикл дает начало боковым корням, которые закладываются эндогенно и, пронзая мощный слой первичной коры, постепенно выходят наружу. На этом основании перицикл называют корнеродным слоем. Из прокамбия образуются все остальные ткани стелы корня. Наиболее важно рассмотреть особенности формирования проводящих тканей: флоэмы и ксилемы. Первой в корне закладывается флоэма. Закладывается она снаружи прокамбиального тяжа, как говорят, экзархно. Это означает, что на периферии тяжа группы клеток прокамбия превращаются в элементы флоэмы. Далее во флоэму превращаются все более внутренне расположенные слои прокамбия, т.е. ее развитие идет центростремительно. Ксилема по времени закладывается позднее и тоже снаружи прокамбиального тяжа, т.е. экзархно. Как и флоэма она развивается по направлению к центру прокамбиального тяжа. Однако в своем развитии ксилема обгоняет флоэму и первой достигает центра корня. В результате, в стеле корня формируется весьма своеобразная конструкция: многолучевая звезда ксилемы, между лучами которой располагаются массивы флоэмы. Звезда ксилемы может иметь разное количество лучей: два, и тогда ее называют диархной; четыре - тетрархная, пять - пентархная и т.д. Таким образом, обе проводящие ткани в корне закладываются и развиваются одинаково: экзархно и центростремительно. В стебле развитие проводящих тканей происходит подругому. Однако между однодольными и двудольными растениями на заключительной стадии формирования корня первичного строения наблюдаются существенные различия. Так, у однодольных растений весь прокамбий полностью (нацело) расходуется на образование проводящих тканей - флоэмы и ксилемы. Поэтому флоэма и ксилема плотно прилегают друг к другу, между ними не остается никакой меристематической прослойки, "никакого зазора". У двудольных же растений между лучами ксилемы и массивами флоэмы остается неизрасходованная прослойка прокамбия. Со временем у клеток этой прослойки повторно пробуждается меристематическая активность и прослойка начинает работать как вторичная меристема, т.е. превращается в камбий. Первичную структуру приобретают корни всех растений. У большинства однодольных растений и у папоротникообразных она сохраняется на протяжении всей жизни. Именно поэтому однодольные растения являются наиболее удобными объектами для изучения корня первичного строения. У двудольных же растений в корне происходят вторичные изменения. Вторичная структура развивается за счет функционирования вторичных меристем: камбия и феллогена (пробкового камбия). Вторичное строение корня Расположенная между лучами ксилемы и массивами флоэмы прослойка камбия начинает работать в две противоположные стороны, откладывая внутрь слои вторичной ксилемы, а наружу - слои вторичной флоэмы. При этом сам камбий все дальше отодвигается от центра корня. В конечном счете, корень вторичного строения имеет следующую последовательность расположения тканей, которая выдерживается с удивительной строгостью. В центре корня вторичного строения всегда располагается звезда первичной ксилемы, далее, по направлению к периферии, следуют слои вторичной ксилемы, затем камбий, вторичная флоэма и остатки первичной флоэмы. Вторичные изменения происходят и в покровах корня. Здесь на смену экзодерме приходит типичная вторичная покровная ткань - перидерма. Корневые системы Совокупность всех корней особи растения называют корневой системой. В состав корневой системы входят разные по значимости и по происхождению корни: Традиционно выделяют 3 вида корней: главный корень ≈ развивается непосредственно из зародышевого корешка семени. боковые корни, которые представляют собой ответвления главного или придаточных корней. Разнообразные боковые корни имеют одну общую особенность: они всегда закладываются в перицикле (именно поэтому перицикл называется корнеродным слоем). Сначала образуется меристематический бугорок, работают инициали, затем из клеток эндодермы, в которые упирается молодой корешок, формируется своеобразный кармашек. При росте корень давит на кармашек и прокладывает путь через первичную кору, действуя не только механически (расталкивая клетки), но и биохимически, выделяя особые ферменты, разрушающие клетки. При выходе бокового корешка в субстрат кармашек спадает. Эндогенное заложение боковых корней имеет чрезвычайно важное приспособительное значение. Если бы ветвление происходило в самом апексе, это бы затрудняло продвижение корня в почве. и, наконец, выделяют еще один тип ≈ придаточные корни. Придаточные корни весьма разнообразны. Если они возникают на стебле ≈ их называют стеблеродными придаточными корнями; они могут появляться также на листьях и на корнях. В последнем случае они именуются корнеродными. Если боковые корни закладываются исключительно в перицикле, то ткани из которых развиваются придаточные корни, весьма разнообразны. Обычно это меристемы: апикальные, камбий, феллоген или сердцевинные лучи. Классифицируют не только виды корней, но и корневые системы в целом. Наиболее широко (а вам еще со школы) известно разделение корневых систем на стержневые и мочковатые. У стержневых систем хорошо выражен главный корень; в мочковатых системах главный корень либо не выражен, либо по каким-то причинам утрачен. Такое деление корневых систем на 2 типа достаточно примитивно. При этом не учитываются причины развития определенных корневых систем, не учитывается также и происхождение корней. Кроме того, эта типология справедлива только по отношению к семенным растениям, поскольку только у семенных растений бывает настоящий главный корень. Дело в том, что среди высших растений существует обширная группа споровых растений. У высших споровых корневая система изначально образована только придаточными корнями, так как у них нет семени, а значит, и зародыша с зародышевым корешком. Такой тип корневой системы считается наиболее примитивным, а сама система называется первично гоморизной. Итак, первичная гоморизия характерна для всех споровых растений: папоротников, хвощей, плаунов. У семенных растений развиваются настоящие главный и боковые корни. В том случае, если хорошо выражены и те и другие, то есть и главный и боковые, корневая система называется аллоризной. Однако, у многих покрытосеменных, особенно у однодольных растений, главный корень из пророста отмирает очень быстро или вообще не развивается. В этом случае, корневая система составлена также только придаточными корнями, но называется она уже вторично гоморизной. Корни различаются не только по происхождению. В пределах одной и той же корневой системы всегда имеются корни, выполняющие разные функции. У большинства растений отчетливо различаются ростовые и сосущие окончания. Ростовые окончания обычно более мощные в сравнении с сосущими, они энергично нарастают и продвигаются вглубь почвы. У них всегда отчетливо выражена зона растяжения. Сосущие окончания развиваются как выросты на ростовых, они удлиняются медленно, их апикальные меристемы почти перестают работать. У древесных форм растений выделяют также толстые и прочные одревесневшие скелетные корни и нежные корневые мочки. Теперь, пожалуй, мы достаточно полно рассмотрели особенности строения типичных корней и корневых систем. Однако наше знакомство со строением и функциями корня было бы не полно, если бы мы не остановились на различных его структурных и функциональных видоизменениях. Нередко корни видоизменяются настолько сильно, что для выяснения из природы требуются специальные исследования. Такие резкие, наследственно закрепленные видоизменения называются метаморфозами. Лекция 8. Морфология побега. Системы побегов Гистологическое строение верхушки побега (апекса) Стебель Первичное строение стебля Вторичное утолщение стебля Стела и ее типы. Стелярная теория Напомню, что побег ≈ один из двух основных органов высших растений. Другим таким органом является корень. Различают побеги: 1) вегетативные и 2) генеративные или спороносные. Вегетативные побеги в типичном случае выполняют функцию воздушного питания; генеративные ≈ обеспечивают репродуктивную функцию, то есть размножение. Побег происходит из единого массива верхушечной меристемы и в этом плане является структурой по рангу подобной корню. Однако в сравнении с корнем побег имеет более сложное строение и расчленен на специализированные части. Вегетативный побег состоит из оси (стебля), имеющего обычно цилиндрическую форму, и листьев, плоских, в типичном случае, боковых органов, сидящих на оси. Облиственность побега является одним из самых существенных признаков, отличающих его от корня. Кроме того, обязательной принадлежностью побега являются почки ≈ зачатки новых побегов. Почки обеспечивают ветвление побега и образование за счет этого системы побегов. Характерно, что боковые ветви на побеге развиваются экзогенно. Главную функцию побега ≈ фотосинтез ≈ осуществляют листья. Стебли являются несущими органами. Они обеспечивают оптимальное размещение листьев в пространстве. Их главные функции механическая и проводящая. Стебли как бы выступают в роли посредников между корнями и листьями. Стебель имеет метамерное строение. Он состоит из узлов ≈ участков, к которым крепятся листья и междоузлий ≈ промежутков между узлами. В тех случаях, когда междоузлия не выражены и побег состоит из тесно сближенных узлов ≈ его называют укороченным. Укороченные побеги характерны, например, для лиственниц, сосен, берез (а также розеточные побеги ≈ образуются на 1-й год у травянистых двулетников). По происхождению принято выделять главный побег, который образуется непосредственно из верхушечной меристемы зародыша. До тех пор, пока сохраняется верхушечная или терминальная почка, главный побег способен к дальнейшему росту. Кроме верхушечной, на побеге образуются боковые (пазушные) почки. У семенных растений они находятся, как правило, непосредственно над узлами, в пазухах листьев, называемых кроющими. Пазушное положение почек имеет важное биологическое значение. С одной стороны, кроющий лист хорошо защищает молодую почку от механических повреждений и высыхания. С другой стороны, зеленый лист интенсивно снабжает почку ассимилянтами. Таким образом, каждый метамер типичного побега состоит из 1) узла с листом и пазушной почкой и 2) нижележащего междоузлия. Из боковых почек соответственно образуются боковые побеги, таким образом происходит экзогенное ветвление и формируется система побегов. В этой системе различают главный побег первого порядка; боковые побеги второго, третьего и т.д. порядков. Почка ≈ представляет собой зачаточный, еще не развернувшийся побег. Узлы на зачаточной оси в почке предельно сближены, поскольку междоузлия еще не успели вытянуться. Верхушка оси заканчивается конусом нарастания или апексом. Кроме того, на оси расположены друг над другом зачаточные листья разного возраста. Нижние, более развитые листовые зачатки прикрывают и защищают более молодые ≈ верхние. В пазухах почечных листочков нередко просматриваются зачатки будущих почек в виде бугорков. Таким образом, в почке заложен весь побег целиком. Появление почки в ходе эволюции было большим достижением для высших растений. Почка представляет собой замкнутую влажную камеру, которая обеспечивает надежную защиту зачаточным побегам в неблагоприятный период. Помимо чисто вегетативных, выделяют генеративные почки, которые заключают в себе зачаток соцветия или одиночный цветок, в последнем случае почку называют бутоном. У ряда растений развиты как бы смешанные вегетативно-генеративные почки, в которых заложены и вегетативные и генеративные структуры, то есть листья и цветы. У многих растений умеренных широт наружные листья почки видоизменены в почечные чешуи. Почки, обладающие такими чешуями, называют закрытыми. Открытые или голые почки не имеют специальных чешуй. Например, зимующие почки деревьев и кустарников закрыты чешуями, но летние почки тех же особей будут уже открытыми. Открытые почки характерны для многих растений влажных тропиков. Кроме нормальных, экзогенных по заложению пазушных почек, у растений часто образуются так называемые придаточные почки. Эти почки не имеют определенной правильности в расположении и возникают не в меристематической верхушке побега, а на взрослой, уже дифференцированной части органа, и притом эндогенно, из внутренних тканей. Придаточные почки могут образовываться и на стеблях, и на листьях, и даже на корнях. Однако по строению эти почки ничем не отличаются от обычных верхушечных и пазушных. Образуясь в большом количестве, придаточные почки обеспечивают интенсивное вегетативное возобновление и размножение. Поэтому они имеют большое биологическое значение. В частности, при помощи придаточных почек размножаются корнеотпрысковые растения. Примерами могут служить осина, осот, иван-чай. Корневые отпрыски ≈ это побеги, развившиеся из придаточных почек на корнях. Среди корнеотпрысковых растений ≈ ряд трудноискоренимых сорняков. Придаточные почки на листьях относительно редки. Все видели комнатное растение бриофиллюм, у которого почки сразу дают маленькие побеги с придаточными корешками, поэтому такие почки называют выводковыми. В сезонном климате умеренной зоны развертывание побегов из почек имеет периодический характер. Побеги, вырастающие из почек за один вегетационный период, называют годичными побегами или годичными приростами. У деревьев эти приросты хорошо различимы благодаря наличию почечных колец. У наших листопадных деревьев летом покрыты листьями только побеги текущего года, на годичных побегах прошлых лет ≈ листьев уже нет. По годичным почечным кольцам можно достаточно точно определить возраст ветви. У некоторых растений за один вегетационный период образуется несколько приростов, такие побеги, чтобы не путать их с годичными, было предложено называть элементарными. Таким образом, годичный побег может состоять из нескольких элементарных. Так, например, у дуба нередко в год образуется 2 элементарных побега: первый весной, второй ≈ в середине лета (так называемые "ивановы побеги"). Почки могут впадать на некоторое время в покой, в том случае, если покой обусловлен морозным периодом, их называют зимующими. Однако по функции все покоящиеся почки можно также назвать почками возобновления. Из этих почек после перерыва возобновляется рост новых побегов. В том случае, если боковые почки совсем не имеют периода ростового покоя и развертываются одновременно с ростом материнского побега, их можно назвать почками обогащения. Развивающиеся из таких почек побеги обогащения сильно увеличивают (обогащают) фотосинтетическую поверхность растения, а иногда и увеличивают семенную продуктивность, если побеги генеративные. Такие боковые побеги образуют одновозрастную с материнским побегом систему. Побеги обогащения характерны для большинства однолетних и многих многолетних трав (пример, Veronica longifolia). И, наконец, особую категорию составляют так называемые спящие почки, очень характерные для лиственных деревьев, кустарников и ряда многолетних трав. Эти почки не превращаются в нормальные побеги в течение многих лет, нередко они спят в течение всей жизни растения. Обычно спящие почки ежегодно нарастают, ровно настолько, насколько утолщается стебель, именно поэтому они не погребаются нарастающими тканями. Нередко спящие почки по мере нарастания многократно ветвятся и образуют густое скопление, целую систему. Стимулом для пробуждения спящих почек служит обычно гибель ствола. При порубке березы, например, из таких спящих почек образуется пневая поросль ≈ это один из способов вегетативного размножения. Особую роль спящие почки играют в жизни кустарников. От дерева кустарник отличается своей многоствольностью. Обычно у кустарников главный материнский стволик функционирует недолго - несколько лет. При затухании роста главного стволика пробуждаются спящие почки и из них формируются дочерние стволики, которые в росте обгоняют материнский. Таким образом, сама кустарниковая форма возникает в результате деятельности спящих почек. Итак, система побегов, как мы выяснили, формируется за счет верхушечных, пазушных и придаточных почек. Но даже при одинаковом способе заложения почек общий облик побегов может сильно различаться. Дело в том, что из этих, внешне одинаковых почек, могут развиваться сильные и слабые ветви. Различают три основных варианта расположения сильных ветвей: При акротонии (греч. акрос ≈ верхушка; тонос - сила, мощь) наиболее сильные ветви формируются ближе к верхушке побега. При мезотонном (греч. мезон - середина) варианте сильные ветви развиваются в средней части побега. А при базитонии (греч. базис - основание) ≈ в нижней (иногда даже из пазушных почек семядолей). Кроме того, побеги могут иметь различное направление роста. Вертикально ориентированные побеги называются ортотропными. Горизонтально растущие побеги получили название плагиотропных. Нередко у растений меняется направление роста побегов. Это явление получило название анизотропии. Например, приподнимающиеся или восходящие побеги у трав и кустарников. При ортотропном росте боковых ветвей у деревьев получается пирамидальная крона. Если плагиотропные побеги стелются по земле ≈ дерево принимает форму стланца. Стелющиеся кустарники называют шпалерными. Такую форму имеют многие полярные ивы. Мы рассматривали морфологические аспекты строения побега, перейдем к изучению особенностей его внутреннего строения, к анатомии. Гистологическое строение верхушки побега (апекса) Внутри почки заключена меристематическая верхушка побега ≈ его апекс. Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех постоянных тканей побега. Источником постоянного обновления апекса являются инициальные клетки, которые, как мы помним, способны к неограниченному делению. Апекс побега бугорчатый, в отличие от гладкого апекса корня. Бугорки представляют собой зачатки листьев ≈ так называемые листовые примордии. Эти выросты появляются экзогенно на поверхности апекса. Гладким остается лишь самый кончик апекса, который называют конусом нарастания, хотя по форме это обычно не конус, а параболоид. Листовые примордии закладываются в определенной последовательности, ритмично. Промежуток времени между появлением бугорков называется пластохроном. Например, у березы в период активного роста пластохрон равен 2-3 суткам. Раньше полагали, что все постоянные ткани побега формируются из одной единственной верхушечной инициальной клетки (теорию апикальной клетки). Несколько позже, в 60-е годы XIX века общепризнанной стала теория гистогенов, разработанная Ганштейном. Напомню, что гистогенами называют специализированные меристематические зоны. В корне такими зонами являются: дерматоген, периблема, плерома. Теорию гистогенов распространили и на побег однако впоследствии выяснилось, что в апексе побега отчетливо выделяется только два слоя. Возникла теория туники и корпуса. Эта теория была сформулирована немецким ботаником А. Шмидтом. Согласно этой теории верхушка побега состоит из двух массивов меристематических тканей или из двух зон ≈ туники и корпуса. Туника ≈ это периферическая меристема, которая обычно состоит из нескольких слоев клеток. Характерная особенность туники состоит в том, что составляющие ее клетки делятся только в одном направлении ≈ вдоль поверхности апекса. Под слоями туники находится корпус или сердцевина. В корпусе клетки делятся во всех направлениях, обеспечивая объемный рост апексов, но деления клеток происходят значительно реже, чем в тунике. Самый наружный слой туники становится протодермой и со временем преобразуется в эпидерму. Довольно рано в апексе обозначаются тяжи длинных и узких клеток, составляющих прокамбий, дающих начало проводящим пучкам. Протодерму и прокамбий можно назвать гистогенами, поскольку эти меристемы уже специализированы и дают начало строго определенным постоянным тканям. Остальная часть апекса, так называемая основная меристема, образует паренхимные и запасающие ткани. Стебель Стебель представляет собой ось побега. Он имеет метамерное строение и состоит из узлов и междоузлий. Нарастает стебель не только за счет верхушечного, но нередко и за счет вставочного роста. Этим стебель существенно отличается от корня. Стебли чрезвычайно разнообразны по форме, они могут быть четырехгранными, многогранными, плоскими в сечении, однако наиболее распространенная форма ≈ цилиндрическая. Основные функции стебля ≈ опорная и проводящая. Стебель морфологически и функционально связывает между собой основные органы питания ≈ корень и листья ≈ это орган-посредник. К этим основным функциям присоединяются многие другие, а иногда они гипертрофированны и выступают на первый план: это запасающая функция, защитная функция (когда стебель несет колючки), молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе. Стебли растений резко различаются по продолжительности жизни. Прежде всего выделяют однолетние и многолетние растения, травянистые и древесные жизненные формы. Многолетние стебли древесных растений называют стволами, у кустарников отдельные стебли именуются стволиками. Для стебля характерна сложная система меристем: верхушечных, боковых и вставочных. В результате деятельности первичных меристем складывается первичное строение стебля. У некоторых растений, например, у большинства однодольных, оно сохраняется на протяжении всей жизни. В том случае, если закладывается прослойка камбия, начинается вторичное утолщение стебля. Рассмотрим эти этапы более подробно. Первичное строение стебля В стебле первичного строения, как и в корне, различают центральный осевой цилиндр (стелу) и первичную кору (ПК). Но соотношение этих тканей совершенно иное: большую часть поперечного сечения занимает центральный цилиндр, имеющий нередко в центре крупную воздушную полость (ВП). Кроме того, в отличие от корня, первичная кора стебля покрыта типичной эпидермой с устьицами (в корне экзодерма). Нередко непосредственно под эпидермой залегают механические ткани (колленхима или склеренхима), вынесенные у стебля на периферию. Эти ткани встречаются в различных сочетаниях с хлорофиллоносной, выделительной и неспециализированной паренхимой. Граница между корой и стелой в стебле выражена значительно менее четко, чем в корне. Дело в том, что эндодерма в стебле не имеет столь характерных признаков как неравномерно утолщенные оболочки эндодермы корня. Правда, эндодерма стебля имеет свою особенность. Нередко в этом слое откладываются многочисленные крахмальные зерна, поэтому эндодерму называют также крахмалоносным влагалищем. Наиболее сложное строение имеет стела. Здесь же наблюдается и максимальное разнообразие различных вариантов строения. Это, в частности, явилось одной из причин создания особой стелярной теории, которую мы рассмотрим несколько позже. По характеру расположения проводящих тканей различают 1) стебли пучкового типа (у которых эти ткани закладываются в виде отдельных пучков) и 2) стебли непучкового типа: в этом случае проводящие ткани располагаются сплошным кольцом. Нам будет несколько удобнее и проще рассмотреть возникновение первичных тканей из прокамбия на примере отдельных пучков. Итак, прокамбий ≈ это первичная меристема, которая состоит из длинных клеток, вытянутых вдоль оси стебля. Так же как и в корне, по времени несколько раньше образуются элементы флоэмы, затем появляется ксилема. Но закладываются они совершенно по-другому, в иных местах проводящих пучков. Флоэма в пучках закладывается снаружи (экзархно) и развивается центростремительно. Ксилема же закладывается эндархно и развивается центробежно. Таким образом, в стебле эти ткани развиваются друг навстречу другу. В подавляющем большинстве случаев при первичном утолщении сильнее развивается сердцевина. Такое утолщение называют медуллярным (клубень картофеля). Значительно реже, при кортикальном утолщении, сильнее развивается первичная кора (как у некоторых кактусов). Вторичное утолщение стебля Если прокамбий всецело расходуется на образование первичных проводящих тканей ≈ вторичное утолщение не происходит, если же остается меристематически деятельная прослойка, то она со временем превращается в камбий и начинается вторичное утолщение. При вторичном утолщении клетки камбия активно работают, откладывая наружу слои вторичной флоэмы, а внутрь ≈ слои вторичной ксилемы. То есть в развитии тканей происходит полная инверсия (поворот на 180 ). Одновременно с началом проявления камбиальной активности происходят изменения и в покровных тканях. Эпидерма не выдерживает давления нарастающих тканей, распирающих ее изнутри, она разрывается и слущивается. Начинает свою работу феллоген (пробковый камбий). На смену эпидерме приходит вторичная покровная ткань ≈ перидерма. Стела и ее типы. Стелярная теория Уже в конце прошлого века в результате трудов фон Моля, де Бари и, главным образом, Сакса сложилось представление о том, что стебель растения состоит из трех систем тканей: покровные ткани, основная перенхима и погруженные в паренхиму проводящие пучки. Но с позиций такого членения трудно было описать и систематизировать разнообразные типы строения стебля, невозможно было сформулировать и четкую концепцию их эволюционного становления. Эти затруднения были преодолены стелярной теорией, сформулированной в основных чертах французским ботаником Ваном-Тигемом. Под стелой Ван-Тигель обозначил совокупность проводящих, арматурных и паренхимных тканей, ограниченных перициклом. Первоначально это чисто описательное понятие было введено для корня, впоследствии его распространили и на стебель. Изучение разных типов стелярной организации представляет большой интерес для понимания путей эволюции высших растений. Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу, и центральное ядро, состоящее из ксилемы, полностью окружено флоэмой. Такая стела называется протостелой. Протостела не только проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостелу имеют некоторые плауны и папоротники. Наиболее примитивной формой протостелы является гаплостела (греч. гаплос - простой), состоящая из центрального пучка первичной ксилемы, окруженного цилиндром флоэмы. Более продвинутой формой является актиностела (греч. актинос - луч), имеющая радиальные выступы ксилемы в виде лучей (на поперечном срезе имеет вид звезды). При переходе к актиностеле флоэма и ксилема имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ. В процессе эволюции протостела дала начало сифоностеле (от греч. сифон ≈ трубка). Сифоностела имеет трубчатое строение, в ней появляется сердцевина. Возникновение сифоностелы сделало возможным существование более крупных организмов. Ксилема, играющая еще и роль арматурной ткани, перемещается на периферию стебля, образование такой трубчатой конструкции делает стебель еще более прочным. Различные типы сифоностелы характерны для многих папоротников. Дальнейшее усложнение стелы связано с появлением у папоротников крупных листьев (макрофильная линия эволюции). Сифоностела как бы дробится на отдельные участки. В результате дробления возникает диктиостела (греч. диктион - сеть) папоротников и эустела (настоящая стела) семенных растений. Последнее звено в эволюции стелы стебля представляет атактостела (греч. а ≈ отрицание, тактос ≈ порядок). Она характерна для однодольных растений. От эустелы отличается отсутствием в проводящих пучках камбия. Сами же проводящие пучки как бы беспорядочно разбросаны по всему поперечному сечению среза. Особо следует отметить эволюцию стелы корня. По внешнему виду она сильно напоминает актиностелу побега. Но нужно иметь в виду, что она соответствует не отдельному пучку, а совокупности всех проводящих тканей органа. Лекция 9. Строение стебля древесного растения Древесина Луб Вторичное утолщение стебля однодольных Расположение камбия и его активность неодинаковы в разных группах растений. Все это разнообразие можно выстроить в единый морфологический ряд, отражающий постепенное затухание деятельность камбия и его полное исчезновение. 1. С самого начала камбий закладывается как непрерывный слой (в виде сплошного кольца) и затем длительно откладывает сплошные слои вторичных проводящих тканей. 2. В прокамбиальных пучках сначала возникает пучковый камбий и начинает работу. Через некоторое время появляются перемычки межпучкового камбия и камбий сливается в сплошное кольцо. Как и в первом случае, откладываются непрерывные слои вторичных тканей. 3. Третий вариант: в обособленных прокамбиальных пучках также возникает пучковый камбий и формирует пучки проводящих тканей. Межпучковый камбий откладывает исключительно паренхиму сердцевидных лучей. 4. Камбий вообще не образуется. Многолетние, длительное время утолщающиеся стебли деревьев и кустарников чаще всего развиваются по 1-му и 2-му типам; 3-й и 4-й - характерны для травянистых растений. Сам камбий состоит из удлиненных четырехгранных клеток, заостренных на концах. Характер заострения бывает разным: либо односторонним (как у долота); либо двусторонним (как у двускатной крыши). Средняя длина камбиальной клетки у двудольных ≈ 0,5 мм, а у голосеменных ≈ 3,5 мм. В процессе эволюции длина камбиальной клетки уменьшается. Оболочка камбия целлюлозная, с небольшим добавлением пектиновых веществ, довольно мягкая и пластичная. Клетка камбия одноядерна. Ядро занимает центральное положение. Клетки камбия делятся на две стороны в радиальном направлении, откладывая внутрь слои ксилемы, наружу ≈ слои флоэмы. Особенно энергично работает камбий весной. Деление камбиальных клеток не имеет строгой последовательности, хотя ксилемные элементы откладываются значительно чаще. Поэтому основная масса стебля состоит из древесины. Обычно при делении камбиальной клетки одна из дочерних клеток остается инициальной, а другая делится незначительное число раз, откладывая проводящие элементы. Процесс превращения клеток камбия в проводящие элементы идет постепенно. Именно поэтому камбий выглядит в виде многослойной, хотя и относительно узкой меристематической зоны. Камбиальное кольцо неуклонно отодвигается к периферии и подвергается растяжению, поэтому число клеток в кольце должно увеличиваться. Действительно, камбиальные клетки время от времени делятся в поперечном направлении. Увеличение числа клеток камбия в обхвате происходит двумя различными путями. В связи с этим различают неярусный и ярусный камбий. Неярусный камбий считается более примитивным. При этом способе клетки камбия сразу делятся наклонно, под углом, поэтому не образуют правильных рядов. В случае ярусного камбия клетки отшнуровываются правильными рядами. Кроме прозенхимных клеток, в состав камбия входят округлые короткие клетки ≈ материнские клетки сердцевидных лучей. В кольце камбия их намного меньше, чем прозенхимных клеток. Участки таких клеток располагаются на равных расстояниях друг от друга. В результате неравномерной работы камбия большую часть объема многолетних древесных стволов занимает древесина (нередко более 90 %). Все ткани, лежащие над камбием, включаются в зону коры. Сюда входит луб (нередко гетерогенный), паренхима вторичной коры, а также перидерма. Позднее, в результате многократного заложения перидерм, формируется третичная покровная ткань ≈ корка или ритодом. Корка также входит в состав коры. Характерно, что активный транспорт происходит в узкой зоне, прилегающей к камбию. Причем луб функционирует в течение одного вегетационного сезона, затем его клетки отмирают и сплющиваются. Проводящие элементы древесины активно функционируют несколько дольше, в течение ряда лет. Выведенные из активного транспорта проводящие элементы в дальнейшем выполняют важные функции ≈ опорную ≈ древесина, и защитную ≈ слои луба, входящие в состав коры. Древесина В составе древесины различают такни четырех физиолого-анатомических систем: 1) проводящей, 2) запасающей, 3) выделительной и 4) механической. Древесина голосеменных растений более однородна по составу. От древесины цветковых она отличается отсутствием сосудов и специализированной механической ткани. Эта древесина почти сплошь состоит из трахеид и называется гомогенной. Трахеиды расположены правильными радиальными рядами. Нередко радиальные ряды удваиваются, в соответствие с увеличением объема стебля. Трахеиды выполняют и проводящую и механическую функции. Причем функцию проведения выполняют преимущественно широкопросветные тонкостенные трахеиды, откладываемые камбием ранней весной, а механическую функцию обеспечивают поздние толстостенные трахеиды. Вместе с тем, древесина многих голосеменных обнаруживает черты высокой специализации. Об этом свидетельствуют, например, 1) сложно устроенные окаймленные поры с торусами (подобных структур нет у цветковых); 2) дифференциация трахеид на водопроводящие и механические; 3) гетерогенные сердцевидные лучи, состоящие из лучевых трахеид и лучевой паренхимы (у цветковых лучи гомогенные); 4) сложная система смоляных ходов. Все это свидетельствует о том, что эволюция хвойных шла по особому пути, независимо от эволюции покрытосеменных. Гетерогенные сердцевинные лучи состоят из коеток двух типов: лучевых трахеид и лучевой паренхимы. Клетки лучевой паренхимы богаты крахмальными зернами и выполняют запасающую функцию, а лучевые трахеиды проводят воду с минеральными солями в радиальном направлении. Древесина покрытосеменных (лиственных) растений имеет сложный гистологический состав и разнообразное строение. Однако главным ее признаком является наличие сосудов. Сосуды разных видов отличаются размерами, типом утолщения клеточных стенок и характером расположения. По расположению сосудов в годичном кольце различают древесину кольцесосудистую и рассеяннососудистую. В первом случае сосуды располагаются преимущественно в ранней древесине, в виде четко выраженных колец. В качестве примеров моно назвать древесину дуба, ясеня. Во втором случае сосуды равномерно расположены по всему годичному кольцу, хотя их размер в поздней древесине несколько уменьшается. Подобными древесинами обладает липа, тополь, клен, береза. Несмотря на наличие сосудов, трахеиды также являются немаловажным элементом в древесине покрытосеменных и, как правило, преобладают по численности и по массе. В некоторых случаях, у примитивных цветковых, древесина вообще изначально лишена сосудов. Другие виды утратили сосуды в процессе регрессивной эволюции, например, многие водные формы. В связи с этим целесообразно различать: 1) первичнобессосудистые растения, предки которых вообще не имели сосудов и 2) вторичнобессосудистые растения, по тем или иным причинам утратившие сосуды. В древесине происходят и существенные возрастные изменения. Нередко сосуды и трахеиды закупориваются тиллами. Это явление называется тиллозисом. Тиллы представляют собой выросты паренхимных клеток, которые через окаймленные поры внедряются в полости проводящих элементов. Тиллы имеют вид тонкостенных мешочков и часто содержат кристаллы углекислой извести, смолы, камеди. У многих древесных пород тиллы образуются в более старых слоях, примыкающих к центру ствола ≈ ядру. Эта часть древесины пропитывается смолами, дубильными и красящими веществами, поэтому хорошо выделяется на фоне более светлой поздней древесины. Подобные древесины называют ядровыми. Молодую древесину, лежащую около камбия, называют заболонью. Если заболонь не отличается по внешнему виду от центральной, старой древесины, то всю древесину именуют спелой. Механическая ткань древесины ≈ либриформ ≈ характерна также для цветковых растений. Это толстостенные, узкопросветные, заостренные на концах волокна, с немногочисленными узкими щелевидными порами. Последний признак является весьма надежным маркером. Дело в том, что иногда на поперечных срезах бывает трудно отличить толстостенные трахеиды от волокон либриформа. На продольных же срезах эти структуры различаются четко. Водопроводящие элементы имеют окаймленные поры, а механические волокна ≈ щелевидные. Сердцевина в зрелых стволах занимает очень небольшой объем в центре. Она состоит из паренхимных клеток, немного вытянутых по длине органа. Нередко сердцевина имеет довольно сложную дифференцировку. Так в примедуллярной зоне оболочки клетки несколько толще, чем в центральной части. Многие живые клетки сердцевины со временем отмирают, а клетки примедуллярной зоны остаются живыми дольше всех и содержат крахмал. Нередко отдельные живые клетки сохраняются и в центральной части, перемежаясь с мертвыми. Луб Наружу, в сторону периферии ствола камбий откладывает луб. Луб отличается от древесины 1) менее мощным годовым приростом, 2) сезонная периодичность в гистологическом строении выражена слабее, 3) в сравнении с древесиной луб имеет меньшую долговечность тканей. В лубе, как и в древесине, различают ткани четырех систем: 1) проводящие, 2) запасающие, 3) хранящие и 4) арматурные. В упрощенной схеме различают мягкий и твердый луб. К мягкому лубу относят элементы проводящей системы ≈ситовидные трубки и клетки спутники, если они имеются, кроме того, в эту же систему входят тонкостенные элементы запасающей и выделительной паренхимы. К твердому лубу относят волокна камбиформа и каменистые клетки. По характеру утолщений и многим другим признакам камбиформ является типичной склеренхимой. Следует подчеркнуть, что именно в клетках лубяной паренхимы обнаруживается исключительное разнообразие ценных для человека органических веществ: дубильные вещества, алкалоиды, гликозиды, эфирные масла и многое другое. В лубе, также как и в древесине, встречаются радиальные лучи, состоящие из паренхимных клеток. Обычно они располагаются строго напротив сердцевинных лучей и порождаются теми же клетками камбия. Функции лубяных лучей аналогичны функциям древесинных лучей. И, наконец, у многих растений в лубе имеются млечники. Вторичное утолщение стебля однодольных Наиболее характерная черта анатомического строения однодольных ≈ отсутствие камбия. Им свойственны проводящие пучки закрытого типа. Когда-то предки однодольных утратили камбиальное утолщение и уже не смогли восстановить его в прежней форме. В этом проявляется необратимость эволюционного процесса. В то же время для некоторых однодольных ≈ представителей семейства Лилейных ≈ характерны древовидные стволы. Подобными стволами обладают некоторые юкки, драцены. Однако, вторичное утолщение у однодольных происходит своеобразным, нетипичным образом. В периферической зоне стебля сохраняется слой клеток, обладающих меристематической активностью. Со временем клетки этого кольца начинают делиться, откладывая внутрь паренхиму и вторичные пучки закрытого типа, наружу откладываются только слои паренхимы. Лекция 10. Лист Строение пластинки типичного зеленого листа Строение нетипичных листьев Листья злаков Световые и теневые листья Листья ксерофитов и суккулентов Листья тенелюбов Морфология листьев Типичный лист представляет собой боковой вырост стебля и состоит из черешка и листовой пластинки. В отличие от стебля и корня, для строения которых характерна радиальная симметрия, в строении листовой пластинки наблюдается симметрия билатеральная, то есть двухсторонняя. В листе имеется верхняя ≈ брюшная и нижняя ≈ спинная сторона (лат. dorsum ≈ спина, venter ≈ брюхо). По своей физиологической роли ≈ лист ≈ орган фотосинтеза, газообмена и транспирации. В тканях листа совершается превращение неорганических веществ (СО2 и Н2О) в органические. Продукты фотосинтеза обычно не накапливаются в листе, а перетекают в другие органы растения. Основной, типичной категорией листьев являются зеленые, ассимилирующие (питающие) листья, которые называются трофофиллами. Вместе с тем, листья являются наиболее пластичными, изменчивыми в эволюционном отношении органами. Специализация листа шла и по другим направлениям. Поэтому у растений мы встречаем разнообразнейшие метаморфозы листа, связанные со сменой функций. Но об этом мы поговорим несколько позже. В связи с указанными функциями в листе хорошо представлены следующие ткани: питательная или ассимиляционная; покровная, регулирующая испарение воды и газообмен; проводящие такни - обеспечивают подведение почвенных растворов и отток продуктов ассимиляции; механические ткани, придающие листу прочность. Кроме этих четырех тканей, в листе могут встречаться группы клеток или отдельные клетки ≈ идиобласты: склереиды, млечники, места отложения отбросов ≈ минеральных солей и других специфических веществ. Листья возникают экзогенно (поверхностно), в виде бугорков вблизи от точки роста. Характерно, что в типичном случае листья не имеют верхушечного роста, а нарастают основанием, за счет интеркалярных меристем, причем в течение ограниченного времени. Эти признаки, однако, не являются абсолютными. Так, у папоротников, например, долго длится верхушечный рост их крупных листьев ≈ вай. У некоторых растений листья имеют цилиндрическое строение и радиально-симметричны. Примеры можно найти среди луков и ситников. Строение пластинки типичного зеленого листа Лист покрыт со всех сторон эпидермой (кожицей). В обычных, горизонтально ориентированных листьях верхний и нижний эпидермис различаются. В целом, кожица состоит из тесно сомкнутых паренхимных клеток с извилистыми очертаниями. Клетки нижнего эпидермиса обычно более мелкие и более извилистые. В верхнем эпидермисе в типичном случае сильнее развиты кутикулярные слои, однако опушение приурочено главным образом к нижней стороне листа. Здесь же, в нижнем эпидермисе обычно располагаются и устьица. Правда, у водных растений, с плавающими листьями, устьица расположены в верхнем эпидермисе. В верхнем же эпидермисе преобладают устьица у некоторых растений, произрастающих на перегреваемых каменистых склонах. Типичный эпидермис однослоен. Многослойные варианты связаны обычно с особыми экологическими условиями (как у Ficus). Под эпидермисом залегает основная зеленая хлорофиллоносная ткань ≈ мезофилл. У большинства растений хлоренхима дифференцирована на столбчатую (палисадную) и рыхлую (губчатую) ткани. В типичном случае к верхней кожице примыкает столбчатый мезофилл, к нижней ≈ губчатый. Нередко эти две ткани соединяются посредством особых собирательных (воронковидных) клеток. У многих растений: ветренниц, вейников, бамбуков столбчатая ткань замещается особыми ветвистыми клетками. На границе между палисадной и губчатой тканью располагаются мелкие ответвления проводящих пучков. Характерным элементом многих листьев являются склереиды, придающие нежным листовым пластинкам дополнительную прочность. Не менее свойственны листьям крупные клетки ≈ идиобласты с отбросами, например, кристаллами солей (как цистолиты у фикуса). В листьях они имеют особое значение. Так, у некоторых галофитов, растений, произрастающих на засоленных субстратах, единственный способ вывести избыток солей из организма ≈ опадание листьев. Проводящая система в листе представлена сосудисто-волокнистыми коллатеральными закрытыми пучками. Характерно расположение проводящих тканей: ксилема располагается в верхней части пучков, флоэма ≈ в нижней. Именно по этому признаку можно точно определить верх и низ листовой пластинки. Все другие признаки изменчивы, так, устьица могут находиться сверху и т. д. Строение нетипичных листьев Листья злаков Особое анатомическое строение имеют листья злаков. Эпидермис у злаков мощно развит, наружные стенки клеток нередко инкрустируются кремнеземом. Весьма своеобразны устьица злаков. Замыкающие клетки имеют вид прямоугольников с закругленными концами. Средняя часть каждой из замыкающих клеток очень толстостенна, концевые же участки ≈ тонкостенны. Здесь же имеются пузыревидные вздутия. При повышении тургора они увеличиваются в размерах и устьичная щель раздвигается. Устьица у злаков обычно располагаются продольными рядами по обеим сторонам листа. У многих ксерофитных злаков в кожице находится особый аппарат, играющий роль при свертывании и развертывании листьев. Пластинка листа таких злаков сильно ребриста. В ложбинках между ребрами в эпидермисе расположены особые пузыревидные клетки. Весьма своеобразно у злаков устроен также мезофилл. Здесь нет типичной палисадной и губчатой хлоренхимы. У многих злаков ассимиляционная ткань арсполагается вокруг проводящих пучков в виде обкладки из плотно сомкнутых, удлиненных клеток. У некоторых злаков клетки ассимиляционной ткани имеют складчатые стенки. Световые и теневые листья У деревьев и кустарников листья, находящиеся в условиях более благоприятного освещения, анатомически и морфологически отличаются от листьев, которые сильно затенены. Световые листья отличаются большей толщиной и жесткостью, кроме того, клетки кожицы имеют менее волнистые очертания и более толстостенны; число устьиц на единицу поверхности листа более значительно; палисадная ткань световых листьев развита мощнее. Листья ксерофитов и суккулентов Весьма своеобразное строение имеют листья растений, приспособившихся переносить длительный засушливый период - ксерофитов, обитающие в условиях дефицита влаги. Правда, приспособились к этому растения по-разному. Мясисто-сочные листовые суккуленты: алоэ, толстянки имеют особый водоносный слой в мякоти листа. Они способны быстро накапливать и экономно расходовать влагу. Примером настоящих ксерофитов являются листья фикуса, панцерий). Такие растения имеют: 1) очень мощную кутикулу; 2) многослойный эпидермис; 3) нередко на поверхности эпидермиса развиваются мертвые белые ветвистые волоски, хорошо отражающие солнечные лучи и предохраняющие растения от перегрева; 4) устьица обычно глубоко спрятаны в особые устьичные карманы. Листья тенелюбов Особую группу составляют теневые растения (тенелюбы). В эту группу входят многие представители, так называемого, таежного мелкотравья, образующие напочвенный покров в наших темнохвойных лесах: заячья кисличка, адокса мускатная. Многие теневые растения относятся к группе гигрофитов. Гигрофиты обитают в условиях постоянной влажности воздуха и почвы. Именно такие условия складываются под пологом густого темнохвойного леса. Структура теневых листьев приспособлена к возможно полному использованию слабого, рассеянного света. Ассимиляционная ткань имеет малую толщину. У таких растений нередко даже эпидермис богат хлорофиллом. У некоторых тропических бегоний палисадные клетки имеют конусовидную форму и работают как линзы, фокусируя лучи света. Все эти примеры лишний раз подтверждают огромную пластичность листьев. Морфология листьев При морфологическом изучении и описании листьев учитываются следующие признаки: характер листорасположения; положение листа на стебле; общая форма листовой пластинки; степень рассеченности листовой пластинки; форма основания листа; форма верхушки листа; характер края листа; типы жилкования листьев. Типичный лист делится на цилиндрический черенок и листовую пластинку, подобные листья называются черешковыми. Листья имеют основание, иногда переходящее во влагалище. Если черешок не выражен, как у некоторых злаков, листья называют сидячими. Листья делят на простые и сложные. Если у листа одна пластинка ≈ его называют простым. Если на одном черешке несколько пластинок ≈ лист называют сложным. Более подробно тему "Морфология листа" рассматривают на практических занятиях. Лекция 11. Метаморфозы органов Метаморфозы корня Метаморфозы побега Каудекс Корневище Подземные столоны и клубни Клубнелуковицы и луковицы Суккуленты Другие метаморфозы побегов Метаморфозами называют наследственно закрепленные видоизменения органов, связанные со сменой их основных функций. Метаморфозы вегетативных органов растений необычайно разнообразны. Метаморфозы корня Одним из наиболее интересных явлений в биологии корня является его взаимовыгодное сожительство с грибами. Это явление имеет специальное название ≈ микориза (что буквально означает грибокорень) и ему посвящена обширная литература. Микориза характерна для подавляющего большинства цветковых растений (вероятно, не менее 90 %). В силу столь широкого распространения микориза является скорее правилом, чем исключением, но, следуя установившейся традиции, мы рассматриваем ее все же как видоизменение корня. Исключением же, то есть растениями, не имеющими микоризы, являются водные растения и паразиты, а также некоторые однолетники: гречишные, крестоцветные, осоковые. С другой стороны, многие растения, такие как орхидные, особенно ведущие сапрофитный образ жизни, настолько тесно связаны с грибами, что даже не могут развиваться без "заражения" специфическим микоризным грибом. Микориза бывает двух разных типов. К первому типу относится эктомикориза (наружная микориза). В этом случае гифы гриба оплетают корни растения толстым чехлом (гифовой мантией) и проникают, кроме того, в межклетники (но не клетки коры). Под влиянием гормонов, выделяемых грибом, молодые корни обильно ветвятся и окончания их утолщаются. Эктомикориза характерна для многих деревьев умеренной зоны, в том числе для видов дуба, березы, ив, кленов, хвойных пород, тополя и других. Эктомикоризу образуют почти исключительно базидиомицеты, реже аскомицеты. Более широкое, почти универсальное распространение имеет эндомикориза (внутренняя микориза). Ее можно наблюдать, например, у яблони, груши, земляники, томатов, злаков, орхидных и многих других видов. Она характерна для большинства цветковых растений. При эндомикоризе грибной чехол вокруг корня не образуется, корневые волоски не отмирают, но гифы проникают гораздо глубже в ткани корня и внедряются в клетки паренхимы коры. Существует несколько различных типов эндомикоризы, причем некоторые из них сильно отличаются друг от друга. У представителей семейства вересковых, например, у вереска, грушанки, клюквы, черники и других, образуется особая эндомикориза, занимающая промежуточное положение между типичной эктомикоризой лесных деревьев и высокоспециализированной микоризой орхидных. Здесь гифы проникают в клетки коры, образуя в них плотную массу, но в то же время оплетают корни рыхлым чехлом. При этом клубни гиф в клетках коры впоследствии перевариваются растением - хозяином, что вообще характерно для эндомикоризы. С другой стороны, микоризный гриб при соответствующих условиях может, в свою очередь, стать настоящим паразитом. В образовании микоризы типа вересковых участвуют оомицеты и зигомицеты. Наиболее специализированный тип эндомикоризы можно наблюдать у орхидных. У орхидных нет гифовой мантии вокруг корней и мицелий почти целиком находится внутри корня. Гифы гриба в клетках коры образуют своеобразные клубки. Эти клубки впоследствии перевариваются растением-хозяином. Грибы, образующие микоризу орхидных, способны разлагать сложные органические вещества и снабжать корни продуктами их разложения. А это имеет особое значение, при сапрофитном способе питания. Грибной компонент эндомикоризы подобного типа почти всегда представлен оомицетами. Огромное значение микориза имеет в тропических дождевых лесах. Из-за сильного промывного режима (ежедневное выпадение осадков) эти леса практически лишены почвы (все питательные вещества из почвы вымываются). Перед растениями остро встает проблема питания. В то же время, свежей органики очень много: опавшие ветви, листья, плоды, семена. Но эта органика недоступна высшим растениям, и они вступают в тесный контакт с сапротрофными грибами. Таким образом, основным источником минеральных веществ в этих условиях является не почва, а почвенные грибы. Минеральные вещества поступают в корень непосредственно из гиф микоризных грибов, именно поэтому для растений дождевого леса характерна поверхностная корневая система. О том, насколько эффективно работает микориза, можно судить хотя бы по тому, что тропические дождевые леса являются самыми продуктивными на Земле сообществами, здесь развивается максимально возможная биомасса. Значительно меньше распространено сожительство корней растений с азотфиксирующими бактериями. Правда, биологическая значимость этого явления чрезвычайно высока. Кроме всем известных бобовых, корневые клубеньки отмечены и у представителей других семейств, например, у некоторых хвойных, у ольхи, у ряда казуариновых, крушиновых, лоховых. Подробно останавливаться на этом широко известном явлении мы не будем. Обычно в корнях любых растений откладывается определенное количество запасных питательных веществ, преимущественно углеводов: особенно крахмал и сахара; у других видов ≈ инулин. Но в некоторых случаях запасающая функция бывает гипертрофированна и выступает на первый план. При этом корни утолщаются, становятся мясистыми. Чаще всего встречается структура, имеющая название корнеплод. Она особенно характерна для двулетних растений: свекла, редька, брюква, турнепс, морковь, сельдерей и многие другие. Из экзотов можно назвать жень-шень, цикорий. Корнеплоды имеют сложную морфологическую природу, поскольку в образовании корнеплода принимает участие и корень и стебель (точнее подсемядольное колено ≈ гипокотиль). Но для того, чтобы установить: какая часть корнеплода представлена корнем, а какая стеблем ≈ необходимы специальные анатомические исследования. Утолщения в виде корневых шишек встречаются и у растений с мочковатой корневой системой, например, у георгина. Во многих случаях корни приспособлены для вегетативного размножения. У целого ряда многолетних растений из корневых придаточных почек развиваются надземные побеги. Эти побеги называются корневые отпрыски. К корнеотпрысковым растениям относятся: осина, слива, вишня, сирень, вьюн, бадан, осот. Последние являются злостными и трудно искоренимыми сорняками. Разрезанные при обработке почвы небольшие кусочки коней легко приживаются и дают начало новым растениям. У многих лазящих растений, например, у плюща, на стебле образуются особые придаточные корни-прицепки, проникающие в трещины и неровности дерева, скалы, или стены и прочно удерживающие растения. Существуют даже корни-крючья, как у фикуса удушителя. Совершенно иную функцию несут так называемые контрактильные (сокращающиеся) или втягивающие корни. Они характерны для многих корневищных, луковичных и клубнелуковичных растений. Классическим примером растения с контрактильными корнями является крокус. У крокуса, кроме обычных корней, развиваются более длинные контрактильные корни, которые при сокращении втягивают клубнелуковицу в грунт. При этом контрактильные корни как бы спадаются, становятся поперечно-морщинистыми и по этому признаку легко отличаются от обычных корней. В особых случаях корни могут быть резервуарами для запасания воды. У многих тропических эпифитных орхидей (а эпифиты ≈ это растения, которые используют как субстрат для произрастания другие растения) наружная часть коры, которая называется веламен состоит из пустых крупных клеток, способных впитывать воду наподобие губки. Во время ливней эти клетки наполняются водой, которая в них хранится и по мере необходимости используется растением. По происхождению веламен ≈ это многослойная ризодерма. У некоторых паразитных растений, например, у представителей семейства гидноровых, корни изменили функции и превратились в присоски (гаустории). Гаустории внедряются в проводящие ткани растенияхозяина и оттягивают оттуда питательные вещества. Многие тропические древесные растения, главным образом мангровые (например, авицения), произрастающие на пресноводных тропических болотах, а также на мелководьях океанических побережий, развивают специальные вентиляционные или дыхательные корни ≈ пневматофоры. Они появляются на подземных боковых корнях и растут вертикально вверх, поднимаясь над водой или почвой. Примечательно, что для таких корней характерен отрицательных геотропизм. Те же мангровые растения образуют корни другого типа, которые называются ходульными. Это придаточные корни ≈ подпорки. Они появляются на стволах и ветвях и растут вниз, внедряются в субстрат и прочно удерживают растение, например, в мягком иле. Такие корни имеет широко распространенное растение мангровых зарослей ≈ ризофора. Но наиболее эффектны ходульные корни у фикуса-баньяна (Ficus benghalensis). Многочисленные придаточные корни баньяна растут вниз, укореняются и развивают собственную корневую систему. Благодаря этому одно дерево баньяна разрастается в целую рощу. Такие рощи могут занимать впечатляющие площади. Не менее интересны опорные досковидные корни, характерные для крупных деревьев тропического дождевого леса. По известным причинам деревья дождевого леса имеют поверхностную корневую систему, в то же время стволы деревьев первого яруса достигают огромных размеров. Корни обычного строения не могут заякоривать растение в почве (которой к тому же нет), удерживая этих гигантов во время частых штормов и ливней. Поэтому, у стелющихся по поверхности почвы корней таких деревьев развиваются особые вертикальные выросты, как доски прилегающие к стволу дерева. Сначала досковидные корни округлые в сечении, но затем происходит сильный односторонний вторичный рост. В тропическом дождевом лесу высота досковидных корней нередко превышает человеческий рост. Не менее своеобразны придаточные корни наших обычных болотных растений - они имеют "этажное" строение. Правда эти корни не имеют специального названия. Метаморфозы побега Побег является самым изменчивым органом растения. Для него характерны такие свойства как: многофункциональность; лабильность поведения; пластичность. Уже в первом приближении побеги делятся на два типа: 1) вегетативные и 2) генеративные. Наблюдается отчетливая смена форм роста и функций побега в процессе его биологического развития. Например: захват новой площади (плеть или корневище); усиленное питание (розеточная стадия); образование цветов и плодов (генеративная стадия). Рассмотрим основные типы специализированных и метаморфизированных органов побегового происхождения. Каудекс Развивается у многолетних трав и кустарников с хорошо развитым стержневым корнем. Это своеобразный многолетний орган побегового происхождения ≈ обычно одревесневшие нижние участки побегов, переходящие в деревянистый стержневой корень. Каудекс несет на себе многочисленные почки возобновления. Кроме того, каудекс обычно служит местом отложения запасных питательных веществ. Как правило, каудекс бывает подземным и достаточно редко ≈ надземным. Побеговое происхождение каудекса можно установить по листовым рубцам и закономерному расположению почек. От корневищ каудекс отличается способом отмирания. Постепенное отмирание идет от центра к периферии, при этом орган делится (растрескивается) продольно на отдельные участки ≈ партикулы. Соответственно, и процесс членения называется партикуляцией. В результате образуется структура, которая нередко называется: многоглавым корневищем, шишковатым корневищем, многоглавый стеблевый стержень, стеблекорень. Эти названия достаточно точно отражают облик каудекса, создают его образ. Следует отметить, что партикуляция характерна для старых (синильных) растений. Каудекс особенно выражен у полупустынных, пустынных и высокогорных растений. У некоторых видов каудексы достигают огромного размера и веса, например, у представителей рода Пангос до 15 кг. В систематическом отношении, каудексовых растений много среди бобовых (люцерна), зонтичных (бедренец), сложноцветных (одуванчик, полынь). Корневище Корневище или ризом (корнеподобный) ≈ это долговечный подземный побег, выполняющий функции вегетативного возобновления, размножения и нередко отложения запасов. Корневище обычно не несет зеленых листьев, но имеет четкую метамерную структуру и этим существенно отличается от корня. Узлы выделяются по листовым чешуйкам, рубцам или пазушным почкам. Как правило, на корневище формируются обильные придаточные корни, расположенные в узлах мочками или поодиночке. Нарастая верхушкой ≈ дистальной частью, корневище постепенно отмирает в старой ≈ проксимальной части. В зависимости от степени развития междоузлий корневищ растения делят на: длиннокорневищные; короткокорневищные; плотнодерновинные. Когда корневище ветвится, то образуется куртина надземных побегов, которые фактически принадлежат одной особи. Такие куртины называются клоном. Так что для многих растений характерна клональная форма роста. Существует два способа формирования корневищ. У некоторых растений первоначально весь побег надземный. Он несет и чешуевидные и зеленые розеточные листья. В дальнейшем листья отмирают, а стеблевая часть втягивается в почву, там утолщается за счет отложения запасных веществ и превращается в корневище. Таким образом, в структуре побега можно различить две фазы: надземную и подземную. В ходе онтогенеза побег претерпевает действительное превращение, метаморфоз в буквальном смысле. Такие корневища называют погружающимися или эпигеогенными ≈ надземнорожденными. Такая картина наблюдается при формировании корневищ: манжетки, гравилата, земляники, медуницы и других. У других растений корневище начинает фазу роста из почки, находящейся под землей. Такие корневища изначально подземного происхождения называются гипогеогенными. Наблюдаются у очень многих многолетних трав и кустарников: пырей, вороний глаз, купена, вероника длиннолистная и другие. В этом случае корневища тонки и служат больше для вегетативного размножения. Подземные столоны и клубни Клубни ≈ утолщения подземного побега как у картофеля, топинамбура. Клубневые утолщения начинают развиваться на концах подземных стеблей ≈ столонов. Столоны недолговечны и разрушаются обычно в течение вегетационного периода, этим они и отличаются от корневищ. В клубнях разрастаются преимущественно паренхимные клетки сердцевины. Проводящие ткани развиты очень слабо и заметны на границе сердцевины и коры. Снаружи клубень покрывается перидермой с толстым слоем пробки, помогающей переносить длительный зимний покой. Листья на клубне очень рано опадают, но оставляют рубцы, в виде так называемых глазков клубня. В каждом глазке находится по 2-3 пазушных почки, из которых прорастает только одна. Почки при благоприятных условиях легко прорастают, питаясь запасными веществами клубня и вырастают в самостоятельное растение. Таким образом, третья ведущая функция подземных побегов ≈ вегетативное возобновление и размножение. Некоторые виды растений образуют весьма своеобразные листовые клубни (например, тонколистный сердечник). Это видоизмененные листовые пластинки, сидящие на черешках корневищ. Эти листовые клубни имеют лопасти, перистое жилкование и даже ткань мезофилма, но бесхлорофильны и приспособлены для откладывания запасного крахмала. Клубнелуковицы и луковицы Клубнелуковица гладиолуса похожа по внешнему виду на луковицу. Однако на продольном срезе видно, что ее стеблевая часть сильно развита и превращена в клубень, содержащий запасные вещества. Снизу клубнелуковицы возникают многочисленные придаточные корни, образующие мочковатую систему. Среди них имеются и контрактильные (втягивающиеся) корни. Луковица представляет еще один вид сильно укороченного подземного побега. В противоположность клубню она имеет сравнительно небольшую стеблевую часть ≈ донце. К донцу прикреплены многочисленные сочные листья, налегающие друг на друга и получившие название луковичных чешуй. У огородного лука, например, мясистые чешуи снаружи покрыты защитными пленчатыми сухими чешуями, поэтому вся луковица такого типа называется пленчатой туникатной или концентрической. У лилий мясистые чешуи налегают друг на друга черепитчато, соответственно, и луковица называется черепитчатой. Сочные чешуи луковицы являются только низовыми питающими листьями побега. Верхние зеленые листья находятся в верхушечной почке донца. Все луковицы объединяются в две категории: с корневищами и без корневищ. Луковицы с корневищами обладают способностью размножаться откидышами: из донца луковицы отрастают корневища, которые удлиняются горизонтально в почве и на некотором расстоянии от материнской луковицы образуют новую луковицу ≈ откидыщ. Луковичка укореняется и через несколько лет может зацвести. К этой категории относятся некоторые тюльпаны и дикорастущие виды лука. Луковицы без корневищ знакомы всем, потому что служат обычным посадочным материалом в овощеводстве и цветоводстве. В пазухах сочных листьев закладываются дочерние луковицы (детки или зубки), которые развиваются до цветения. Много луковичек развивается, например, у чеснока. Запасание воды в паренхимных клетках сочных чешуй обеспечивается благодаря выработке этими клетками особых слизистых веществ, сильно набухающих в воде и удерживающих воду. В большинстве случаев луковичные ведут себя как эфемероиды. Их надземные побеги существуют весьма непродолжительное время: появляются рано весной и отмирают уже в начале лета. Растения как бы "убегают от засухи". Суккуленты Суккулентами называют растения, имеющие сочные, мясистые листья или стебли, которые служат своеобразными резервуарами для запасания влаги. Эту влагу суккуленты очень бережно и экономно расходуют на протяжении засушливого периода. Суккуленты разделяются на две большие группы: Стеблевые суккуленты - имеют мясистые стебли, листья же, как правило, превратились в колючки (для уменьшения транспирации). В качестве примеров стеблевых суккулентов можно назвать всем известные американские кактусы и очень похожие на них африканские молочаи. Листовые суккуленты - имеют толстые, мясистые листья. К ним относятся толстянковые (Crassulaceae): очиток, золотой корень; лилейные, амариллисовые, агавы, алоэ, гастерии, гавортии. Другие метаморфозы побегов Особый интересный случай метаморфоза почки в суккулентный орган ≈ кочан наблюдается у обычной культурной капусты. Как известно, капуста является двулетним растением. В первый год появляются розеточные, слегка суккулентные листья, затем почка резко увеличивается в размерах и превращается в кочан. На второй год после перезимовки капуста, как типичный двулетник, дает удлиненный цветоносный побег. У растений наблюдается большое разнообразие шипов и колючек, которые, к тому же, имеют разное происхождение. Например, у кактусов и барбарисов колючки представляют собой видоизмененные листья. Обычно такие колючки предназначены, в первую очередь, для уменьшения транспирации, защитная же функция в большинстве случаев является второстепенной. Другие растения (боярышник, дикая яблоня) имеют колючки побегового происхождения это видоизмененные укороченные побеги. Нередко они начинают развиваться как нормальные облиственные побеги, а затем одревесневают и утрачивают листья. Дальнейший шаг в недоразвитии листьев и передачи их функций зеленым стеблям приводит к формированию таких метаморфизированных органов, как филлокладии и кладодии. Филлокладии (греч. филлон - лист, кладос - ветвь) - это плоские листоподобные стебли и даже целые побеги. Наиболее изестным примером растений, имеющих метаморфозы подобного рода, являются иглицы (Ruscus). Эти растения произрастают в Крыму и на Кавказе; нередко они разводятся и в комнатных условиях. Весьма интересно, что на листопобобных побегах иглицы развиваются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях. Кроме того, филлокладии как и листья имеют ограниченный рост. Кладодиями же называют уплощенные стебли, которые в отличие от филлокладиев, сохранили способность к длительному росту. Это достаточно редкие видоизменения и встречаются, например, у австралийских мюленбекий. У многих лазящих растений (горохи, чины, тыквы и пр.) наблюдается видоизменение листьев в усики, которые обладают способностью закручиваться вокруг опоры. Стебель у таких растений обычно тонкий и слабый, неспособный сохранять вертикальное положение. Стелющиеся растения (земляника, костяника и др.) формируют особый тип побегов, служащих для вегетативного размножения, такие как плети и столоны. Их относят к категории надземно-ползучих растений. Лекция 12. Общие сведения о размножении растений Отдел Моховидные (Bryophyta) Важным аспектом раздела "Морфология растений" является изучение жизненных циклов их развития. Поэтому, прежде чем перейти к рассмотрению отдельных таксонов высших растений, необходимо освежить в памяти общие представления о размножении растений. Каждый организм, в том числе и растение, имеет способность к воспроизведению себе подобных. Именно этот процесс и обеспечивает длительное, как правило, существование вида. Воспроизведение организмов возможно благодаря их способности производить потомство. В том случае, если образование потомства приводит к увеличению численности данного вида, т.е. дочерних особей становится больше чем материнских, этот процесс можно назвать размножением. Выделяют два основных способа размножения растений: бесполое и половое. При бесполом размножении (в широком смысле) особь либо отделяет от себя неспециализированные вегетативные зачатки (почки, клубни, луковицы и пр.), и здесь мы имеем дело с вегетативным размножением. Нередко в качестве зачатков растение производит специализированные клетки, называемые спорами (греч. спора - посев). Спорами размножаются все низшие и многие высшие растения, за исключением голосеменных и цветковых, хотя и у последних образуются споры. Формирование спор на растении называют спороношением. Половое же размножение осуществляется путем слияния (копуляции) половых клеток гамет. Бесполое размножение Половое размножение (слияние гамет) Вегетативное С помощью спор У многих низших растений, например водорослей, споры возникают в результате обычного деления клетки - митоза, поэтому и называют их митоспорами. Кроме того, как правило, споры низших растений (зооспоры) имеют жгутики, с помощью которых передвигаются в воде. У всех высших растений, в отличие от низших, споры развиваются в специализированных многоклеточных органах - спорангиях и образуются они в результате редукционного деления клетки - мейоза. Другими словами, все без исключения высшие растения образуют мейоспоры. Как правило, споры высших растений не имеют жгутиков и окружены твердой оболочкой. В случае же полового процесса, при слиянии гаплоидных гамет образуется диплоидная зигота. Выделяют несколько основных типов полового процесса у растений. 1. Хологамия (греч. холос - целый, гамео - вступать в брак) - сливаются одноклеточные организмы. Такой способ характерен только для наиболее примитивных низших растений. У большинства же, как низших, так и высших растений гаметы образуются в специализированных органах - гаметангиях. Изогамия (греч. изо - равный) - в этом случае сливаются совершенно одинаковые по размеру и внешнему виду гаметы, одну из которых условно можно считать женской, другую - мужской. Гетерогамия (греч. гетерос - различный) - постепенно в процессе эволюции гаметы становятся разных размеров: женские более крупные, хотя они и не утрачивают подвижность; подвижные мужские гаметы - сперматозоиды имеют более мелкие размеры. Оогамия (греч. оон - яйцо) - на данной стадии крупные женские гаметы (яйцеклетки) утрачивают подвижность, однако более мелкие мужские гаметы - сперматозоиды все еще ее сохраняют. Необходимо отметить, что все без исключения высшие растения являются оогамными. 2. И, наконец, у семенных растений даже более мелкие мужские гаметы утрачивают подвижность. Такие неподвижные мужские гаметы называют спермиями. Отдел Моховидные (Bryophyta) Моховидные - это обширная группа растений (около 30000 видов), объединяемых нередко под общим названием - мхи. Однако в более точном, научном смысле мхами называют лишь представителей одного класса - настоящих или листостебельных мхов, поскольку кроме них отделе представлены еще 2 класса: антоцеротовые и печеночники. Среди высших растений моховидные образуют наиболее обособленную группу. Своеобразие этой группы настолько велико, что в ботанике выделилась особая наука бриология, в рамках которой занимаются исключительно изучением мхов. Для моховидных создаются совершенно особые определители и флоры, поскольку в стандартные общие флоры - они не вписываются. Моховидная наиболее примитивная и древняя группа высших растений. Их древность вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились основные линии развития моховидных, такие как печеночники, сфагновые и бриевые мхи. Моховидные представляют собой не только древнюю группу. Это еще и слепая ветвь в эволюции растений. Однако мхи заняли особое место в природе, особую нишу, и в настоящее время принимают заметное участие в сложении растительного покрова Земли. При этом они нередко оказывают существенное воздействие на среду обитания других растений и животных. Так, в условиях повышенного увлажнения в умеренных широтах северного и южного полушарий идет болотообразовательный процесс, при этом накапливаются значительные толщи торфяных отложений с преобладающим участием моховидных. Особенно характерно это для областей, где осадки преобладают над испарением. Например, заболоченность Западной Сибири по самым скромным оценкам составляет около 50% и эта территория по праву может считаться уникальным болотным регионом. Общеизвестно ландшафтное значение моховидных в тундровой зоне, там даже выделяется особая подзона моховых тундр. В достаточно больших количествах эти растения встречаются и в областях тропических. Так, в высокогорьях тропиков на высоте более 3000 м выделяется пояс мшистых лесов. Моховидные облепляют древесные растения настолько густо, что не видно даже стволов и ветвей. Все скрыто в сплошном зеленом футляре. Если говорить об условиях обитания моховидных, то следует отметить, что их обычно рассматривают как растения, связанные в своей жизни с избыточным увлажнением. Однако значение этой зависимости не следует преувеличивать. Да, бриофиты действительно в изобилии растут на болотах, в сырых лесах и тундрах. Но есть среди них и ксерофиты. Так, в некоторых пустынях Tortula покрывает до 90% поверхности. Стало быть, существуют не только моховые тундры, но и моховые пустыни! Моховидные практически вездесущи. Почти все моховидные обнаруживают, подобно лишайникам, поразительную устойчивость к длительному пересыханию. При увлажнении они снова оживают. Пожалуй, после лишайников моховидные являются самыми выносливыми растениями. В одних случаях моховидные играют большую роль в сложении растительного покрова, в других незначительную, но всегда они непременный компонент растительного мира любого региона. Никакое ботаническое изучение территории, даже пустынной, не будет полным без учета бриофлоры. Необыкновенно многообразны и субстраты, на которых селятся моховидные. Сравнительно немногие виды живут на поверхности почвы. Другие освоили скалы, камни, стволы, ветви, пни, валежины. И, наконец, еще одна уникальная особенность моховидных. В сравнении с другими высшими растениями - это растения космополиты, они имеют чрезвычайно широкие ареалы (область распространения вида). Причины: маленькие размеры тела, угнетенность полового процесса, повышенная толерантность (устойчивость), плюс дерновинная форма роста. Таким растениям легче отыскать подходящую экологическую нишу, которую можно назвать микронишей. Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития. Жизненный цикл большинства растений складывается из 2-х фаз развития, или ядерных фаз, последовательно сменяющих друг друга: растение с диплоидным набором хромосом спорофит, сменяется растением с гаплоидным набором хромосом - гаметофитом. У очень небольшого числа низших растений спорофит и гаметофит неразличимы. Но у большинства низших и у всех высших растений (в том числе и моховидных) гаметофит и спорофит совсем непохожи друг на друга. Если не знать жизненного цикла растения, то спорофазу и гаметофазу можно принять за совершенно разные виды и это не раз случалось в истории ботаники. Соотношение размеров и продолжительность жизни спорофита и гаметофита неодинаковы в разных группах растений. Но у большинства высших растений в жизненном цикле преобладает спорофаза. Под словом "преобладает" понимается такое положение, когда спорофит крупнее гаметофита и дольше живет. Собственно, все, что мы называем высшим растением: кочка осоки, цветущий куст, огромный ствол дерева с листьями, корнями, плодами и семенами - все это и есть спорофит. Таким образом, можно смело утверждать, что подавляющее большинство высших растений пошло в эволюции по пути преимущественного развития спорофазы - спорофита (осуществили диплоидную линию эволюции). Гаметофит занимает весьма незначительную часть жизненного цикла. Более того, наблюдается тенденция к неуклонному сокращению роли гаметофазы в жизненном цикле. Если у папоротников гаметофит - это крохотное, самостоятельно питающееся растение - заросток, то у покрытосеменных гаметофит имеет микроскопические размеры, состоит из нескольких клеток и живет внутри спорофита. Чем выше по эволюционной лестнице продвинулось растение, тем более редуцированный гаметофит оно имеет. Моховидные же представляют исключение. Они пошли в эволюции своей дорогой. У них все наоборот по сравнению с остальными высшими растениями, т.е. в жизненном цикле этих растений преобладает гаметофит. По сравнению с другими высшими растениями гаметофит значительно расширяет сферу своей физиологической деятельности: он не только обеспечивает половое размножение, но и выполняет основные вегетативные функции фотосинтез, водоснабжение и минеральное питание. Однако стебли и листья, развивающиеся на гаметофите моховидных не являются настоящими, поэтому стебли моховидных в бриологии принято называть каулидиями, а листья - филлидиями. Спорофит же у моховидных имеет меньшие размеры, лишен листьев и фактически низведен до роли спороносящего органа. Однако рассмотрим все по порядку. Из одноклеточной гаплоидной споры у моховидных развивается многоклеточная ветвистая зеленая нить - протонема. Она очень похожа на нитчатые зеленые водоросли. Однако в отличие от зеленых водорослей протонема имеет не прямые, а косые перегородки между клетками и хлоропласты вместо хроматофоров. На протонеме появляются многочисленные выводковые почки, из которых вырастают облиственные побеги гаметофоры. Их так называют потому, что на гаметофорах образуются половые органы - гаметангии: мужские - антеридии и женские - архегонии. Поскольку на протонеме мхов образуются многочисленные почки, большинству из них присущи групповые формы роста - плотные, компактные дерновинки. Это дает мхам возможность переносить неблагоприятные условия, быстрее поглощать и сохранять влагу. Компактное произрастание растений также обеспечивает половое размножение. Виды моховидных можно разделить на однодомные и двудомные растения. У однодомных - половые органы - антеридии и архегонии находятся на одном растении, у двудомных - на разных. Так, кукушкин лен является двудомным растением. Гаметангии располагаются обычно не поодиночке, а группами на верхушках облиственных гаметофоров, причем перемежаются с многочисленными стерильными нитями парафизами. Парафизы защищают гаметангии и предохраняют их от высыхания. Обычно скопления гаметангиев окружены специальными, сильно видоизмененными листовыми выростами. Антеридии имеют вид дисковых мешочков на ножке с однослойной оболочкой. Все их внутреннее содержимое заполнено сперматогенной тканью. Из каждой клетки этой ткани образуются подвижные двухжгутиковые сперматозоиды. Архегоний, в типичном случае, представляет собой колбовидное тельце, с расширенной нижней частью - брюшком, где помещается крупная яйцеклетка. В брюшке также размещается брюшная канальцевая клетка. Горловина архегония называется шейкой, в ней расположены шейковые канальцевые клетки. При созревании архегония брюшная и шейковые канальцевые клетки расслизняются и слизь немного выступает наружу. При этом внутри архегония образуется своеобразный слизистый канал, по которому сперматозоиды легко достигают яйцеклетки: слизь облегчает их продвижение. Оплодотворение происходит при наличии капельножидкой воды. Сперматозоиды подплывают к архегонию, проникают внутрь и, двигаясь по слизистому каналу, подходят к яйцеклетке. При слиянии гамет (сперматозоида и яйцеклетки) появляется диплоидная зигота и ее дальнейшее развитие происходит внутри архегония. Из зиготы формируется спорофит, который у мхов называется - спорогоном. Это очень яркое и меткое название. Действительно спорогон моховидных растений не имеет никакой иной функции кроме производства спор (гонит споры). Строение спорогона удобно рассмотреть на примере кукушкина льна обыкновенного (Politrichum commune). Он представляет собой коробочку на ножке, прикрытую волосистым колпачком. Интересно отметить, что спорогон в целом - диплоидное образование, в то время как сидящий на его вершине колпачок - гаплоидный. Как такое могло получиться? Когда зигота начинает делиться внутри архегония - она его разрывает и растущий спорогон поднимает верхнюю часть архегония вверх. Эта верхняя часть архегония впоследствии и преобразуется в колпачок. Сама коробочка состоит из нескольких основных структурных частей. В ее составе можно выделить урночку - своеобразную емкость для спор. Сверху урна прикрыта крышечкой, наподобии туристского котелка. Нижняя расширенная часть урны называется апофизой. В апофизе имеются хлоропласты, за счет которых спорогон может фотосинтезировать и, хотя бы частично, снабжает себя пластическими веществами. Отверстие урны, помимо крышечки, прикрыто тонкой белой пергаментообразной пленочкой - эпифрагмой. После отделения крышечки эпифрагма надежно прикрывает устье урны и препятствует преждевременному высыпанию спор. В центре разрезанной повдоль коробочки обнаруживается колонка, которая по периметру окружена полостью. В этой полости размещается так называемый споровый мешок. Он подвешен к внутренним стенкам урны на особых флорофиллоносных нитях, которые играют роль своеобразных аммортизаторов. Споры образуются в результате редукционного деления спорогенной ткани, заполняющей споровый мешок. Кукушкин лен обладает весьма тонким механизмом высеивания спор. Дело в том, что пленочка - эпифрагма крепится к краям зубцов так называемого перистома, окаймляющих устье урны. Клетки зубцов перистома имеют неравномерноутолщенные оболочки. Кроме того, перистом отличается удивительной гигроскопичностью, способностью впитывать влагу. Поэтому во влажную погоду зубцы перистома набухают, изгибаются и плотно прижимают эпифрагму к отверстию урны, препятствуя прорастанию спор внутри урны. В сухую погоду зубцы перистома расправляются (выпрямляются), теряя влагу, и эпифрагма слегка приподнимается над отверстием урны. В образовавшиеся между зубцами прорехи небольшими порциями высыпаются споры, когда коробочка раскачивается на ветру. При этом в сухую погоду споры разносятся на большое расстояние. Таким образом, развитие спорофита как самостоятельного организма у моховидных не происходит. Он прикреплен к гаметофиту и живет в значительной мере за его счет. Еще совсем недавно считалось, что спорофит мохообразных паразитирует на гаметофите. Однако это не вполне верно, поскольку спорофит, по крайней мере, в молодом возрасте фотосинтезирует, и, значит, о полном паразитизме говорить нельзя. Другие исследователи считают, что спорофит произрастает на гаметофите эпифитно. Правильнее будет сказать, что обе фазы образуют сложный двуединый организм, с главенствующей ролью гаметофита. Следует отметить, что размножение с помощью спор в большинстве групп моховидных не является главным. Для того, чтобы возник спорофит, необходимо оплодотворение. Для многих видов это почти непреодолимая трудность. Дело в том, что для этого необходима вода, в которой могут перемещаться сперматозоиды. И если антеридии и архегонии разделены пространством хотя бы в несколько сантиметров, то даже дождевой воды может не хватать, для реализации оплодотворения. А ведь нередко мужские и женские особи разделены сотнями и даже тысячами метров. Есть, например, очень интересный вид Tortula, у которого мужские особи отмечены только в Европе, а женские - исключительно в Северной Америке. В таких условиях оплодотворение просто нереально. И моховидные нашли выход в исключительно активном вегетативном размножении. Практически любая часть гаметофита - кусочки листьев, стеблей при благоприятных условиях может дать протонему, а через нее и новые растения. Большой кусочек слоевища или веточка могут развиваться в целое растение, минуя стадию протонемы. Моховидные в подавляющем большинстве многолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных колеблются от миллиметров до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода Fontinalis могут иметь стебли длиной до 60 см. и более. Такими же длинными являются и стебли некоторых мхов - эпифитов из рода Dawsonia, поселяющихся на стволах и ветвях деревьев. Это гиганты среди мохообразных. В силу многих причин гаплоидная линия эволюции, избранная моховидными, оказалась невыгодной. Остались отрезанными многие возможные пути эволюционного развития. Именно поэтому моховидные обречены оставаться мелкими растениями, кроме того, у них слабо развита водопроводящая и механическая система. В целом, структурные возможности гаплоидного гаметофита оказались значительно ниже. Моховидные - единственный отдел растительного царства в жизненном цикле которых безраздельно и полно господствует гаметофаза.