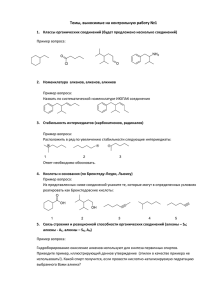
Саратовский государственный медицинский университет имени В.И. Разумовского Русецкая Наталья Юрьевна
реклама

1 Саратовский государственный медицинский университет имени В.И. Разумовского На правах рукописи Русецкая Наталья Юрьевна СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ЗАКОНОМЕРНОСТИ БИОЛОГИЧЕСКОГО ДЕЙСТВИЯ ХАЛЬКОГЕНОРГАНИЧЕСКИХ СОЕДИНЕНИЙ 03.01.04 – биохимия Диссертация на соискание ученой степени доктора биологических наук Научный консультант: доктор медицинских наук, профессор В.Б. Бородулин Саратов 2014 2 ОГЛАВЛЕНИЕ Список используемых сокращений……………………………………………….5 Введение……………………….…………………………………………………….7 Глава 1. Обзор литературы…..………………………………..…….…………14 1.1. Халькогены. Физико-химические свойства, биологическая активность, метаболизм в клетках эукариот и прокариот…………………………….……..14 1.2. Окислительный стресс: механизмы развития и предотвращения…….....30 1.3. Антитоксическая активность халькогенорганических соединений при интоксикации солями кадмия, свинца и ртути……………………………..…..56 1.4. Антибактериальная активность халькогенорганических соединений в отношении грамотрицательных и грамположительных микроорганизмов…………………………………………………………………..73 1.5. Гормоноподобное действие синтетических нестероидных соединений………………………………………………………………….…..…..90 Глава 2. Материалы и методы исследования……………………………......108 2.1. Халькогеноорганические препараты, использованные в исследовании……………………………………………………………………....108 2.2. Постановка эксперимента…………….……………………………….……..109 2.2.1. Подготовка препаратов к исследованиям……………………………..…109 2.2.2. Подготовка биологического материала…………………………………111 2.2.3. Изучение свободнорадикального окисления липидов и антиоксидантного статуса в тканях мышей……………………………………………………….….112 2.2.4. Изучение антитоксического действия халькогенорганических препаратов при подостром отравлении солями тяжелых металлов…………………..…..113 2.2.5. Изучение антибактериальной активности халькогенорганических соединений……………………………………………………………………...…115 2.2.6. Изучение гормоноподобного действия халькогенорганических соединений…………………………………………………………………….…..116 2.3. Методы исследования…………………………………………………..……117 2.3.1. Методы изучения свободнорадикального окисления липидов и антиоксидантной системы………………………………………………………..117 3 2.3.2. Методы исследования биохимических показателей плазмы крови…….121 2.3.3. Методы исследования гематологических показателей плазмы крови……………………………………………………………………………….123 2.3.4. Методы определения показателей углеводного обмена……………….126 2.3.5. Определение антибактериального действия халькогенорганических соединений по числу выросших клеточных колоний…………………..…….129 2.4. Квантово-химические расчеты………………………………………….….130 2.5. Компьютерное прогнозирование биологической активности с использованием компьютерной системы предсказания спектра биологической активности (PASS C&T)……………………………………………………....….131 2.6. Статистические параметры, использованные в работе …………..……...133 Глава 3. Антиоксидантная активность халькогенорганических соединений………………………………………………………………….…….134 3.1. Антиоксидантные показатели гемолизата эритроцитов, плазмы крови и тканей интактных мышей с различной оксидорезистентностью …………..…135 3.2. Концентрации продуктов перекисного окисления липидов в гемолизате эритроцитов, плазме крови и тканях мышей с различной оксидорезистентностью …………….……………………………………………137 3.3. Активность антиоксидантных ферментов в плазме крови и тканях гемолизате эритроцитов, мышей с различной оксидорезистентностью………………………………………..…………..……..140 3.4. Механизм антиоксидантного действия халькогенорганических соединений……………………………………………………………….………..147 Глава 4. Антитоксическое действие халькогенорганических соединений при интоксикации солями тяжелых металлов………………........………..151 4.1. Оценка функциональной активности органов и тканей белых крыс при использовании халькогенорганических препаратов в качестве антитоксикантов при интоксикации солями тяжелых металлов ………..………………………..153 4.2. Оценка антитоксической активности халькогенорганических препаратов по результатам гематологических показателей крови крыс..………….………….170 4 4.3. Механизм антитоксического действия халькогенорганических препаратов……………………………………………………………...………….184 Глава 5. Антибактериальная активность халькогенорганических соединений………………………………………………………………………..187 5.1. Антибактериальное действие халькогенорганических соединений на клинические штаммы Staphylococcus aureus …………………………..……….187 5.2. Антибактериальное действие халькогенорганических соединений на клинические штаммы Pseudomonas aeruginosa………………………..……….196 5.3. Антибактериальное действие халькогенорганических соединений на клинические штаммы Escherichia coli……………………………………..……205 5.4. Механизм антибактериального действия халькогенорганических соединений…………………………………………………………………..…….214 Глава 6. Элементы структурного подобия молекул как предпосылка сходного физиологического действия халькогенорганических соединений и глюкокортикоидов…………………………………………………………..….219 6.1. Компьютерное халькогенорганических прогнозирование соединений, биологической полученные с активности использованием компьютерной системы предсказания спектра биологической активности PASS………………………………………………………………………….……220 6.2. Квантовохимические расчеты, подтверждающие структурное сходство селеноорганического соединения ДАФС-25, его производных и глюкокортикоидов…………………………………………………………..……226 6.3. Влияние препарата ДАФС, его производных и преднизолона на показатели углеводного обмена экспериментальных животных………………………..…230 Заключение…………………………………………………………….…………236 Выводы…………………………………...…………………………….…………261 Список использованной литературы………………...……………………….263 5 Список используемых сокращений АлАТ – аланинаминотрансфераза АсАТ - аспартатаминотрансфераза АОЗ - антиоксидантная защита АОС - антиоксидантная система АФА - активные формы азота АФК - активные формы кислорода ГГТ - -глутамилтрансфераза ГК – глюкокортикоид ГКР – глюкокортикоидный рецептор ГПО (GPX) - глутатионпероксидаза ГР - глутаредоксин ДАФС-25, ДАФС(Se) – Диацетофенонилселенид, селенапентадион-1,5 ДАФС(Cl2) - 1,5-ди-(п-хлорфенил)-3-селенапентандион-1,5 ДАФС(F2) - 1,5-ди-(п-фторфенил)-3-селенапентандион-1,5 ДАФС(NO2)2 - 1,5-ди-(м-нитрофенил)-3-селенапентандион-1,5 ДАФС(S) – 1,5-дифенил-3-тиапентадион-1,5 ДАФС(Те) - 1,5-дифенил-3-теллурапентадион-1,5 ДК – диеновые конъюгаты ДМФА - диметилформамид КК (КФК) - креатинкиназа ЛДГ - лактатдегидрогеназа ЛИИ – лейкоцитарный индекс интоксикации ЛПНП - липопротеины низкой плотности МДА – малоновый диальдегид НАДФ - никотинамидадениндинуклеотидфосфат ПНЖК – полиненасыщенные жирные кислоты ПОЛ - перекисное окисление липидов ПР - прогестероновый рецептор СОД – супероксиддисмутаза СРО - свободно-радикальное окисление ТБК - тиобарбитуровая кислота ТР - тиоредоксин ТРР (TrxR, TXR) - тиоредоксинредуктаза УФ - ультрофиолетовый ФAД - флавинадениндинуклеотид ФАФС – фосфоаденозинфосфосульфат 1,5-дифенил-3- 6 ЦПЭ - цепь переноса электронов ЩФ - щелочная фосфатаза E.coli - Escherichia coli G–SH – восстановленный глутатион GS–SG – окисленный глутатион еNOS - эндотелиальная NO синтаза iNOS - индуцибельная NO синтаза nNOS - нейронная NO синтаза • NO - оксид азота Nox - НАДФН-оксидаза ONOO- - пероксинитрит ONOOCO2- - нитрозо-пероксокарбоксилат PASS - Prediction of Activity Spectra for Substances P. aeruginosa - Pseudomonas aeruginosa S. aureus - Staphylococcus aureus 7 Введение Актуальность исследования. Согласно данным Всемирной организации здравоохранения, количество и характер потребляемых продуктов питания являются основными факторами, определяющими здоровье человека [ВОЗ, 1993]. В «Основах государственной политики Российской Федерации в области здорового питания населения на период до 2020 г.» особое внимание уделяется профилактике заболеваний, обусловленных неполноценным и несбалансированным питанием [Распоряжение Правительства РФ №1873-р, 2010]. В этой связи значительную актуальность приобретает коррекция питания населения с целью снижения распространенности селенодефицитных состояний [Баранова Т.А., 2008; Ширшова Т.И. и др., 2011]. По современным данным, до 80% населения России имеет недостаточную обеспеченность селеном [Сенькевич О.А. и др., 2011; Ширшова Т.И. и др., 2011; Давыденко Н.И. и др., 2012]. В то же время проблема болезней, связанных с дефицитом селена, остается нерешенной до сих пор. В настоящее время для борьбы с селенодефицитом применяются биологически активные добавки, содержащие неорганический селен, главным образом, селенит натрия [Перепелкина Л.И. и др., 2012]. В то же время органические соединения селена по сравнению с неорганическими являются менее токсичными, более биодоступными и лучше усваиваемыми живыми организмами [Комзалова А.В. и др., 2012; Бикчантаев И.Т., Шакиров Ш.К., 2013; Фроловичев А.С., Трошин А.Н., 2013]. Поэтому научные разработки последних лет направлены на синтез и использование органических форм селена в целях (беломышечной профилактики болезни, некроза селенодефицита и жирового и ряда заболеваний перерождения печени, экссудативного диатеза, энцефаломаляции, расстройства сперматогенеза и др.) [Комзалова А.В. и др., 2012; Бикчантаев И.Т., Шакиров Ш.К., 2013; Фроловичев А.С., Трошин А.Н., 2013]. 8 Одним из перспективных селеноорганических соединений является диацетофенонилселенид (ДАФС-25) (производитель: ЗАО «Сульфат», г. Саратов), который позволяет нормализовать деятельность иммунной, антиоксидантной и детоксицирующей систем организма животных и птиц, приводит к увеличению яичной и мясной продукции [Древко Б.И. и др., 2001; Иванова Л.В., 2009]. ДАФС-25 в 40 раз менее токсичен селенита натрия, селен в нем находится в органической, более доступной для животных форме [Кулешов К.А., 2008]. Известно, что селен расположен в VI группе Периодической системы элементов Д.И. Менделеева, все элементв которой относятся к халькогенам, являются электронными аналогами и обладают сходными химическими свойствами, что, вероятно, будет обусловливать близость биологических свойств этих элементов и их соединений [Паперная Л. К., 2007]. В связи с этим несомненный интерес представляет обнаружение взаимосвязи между структурой и биологической активностью халькогенов (серы, селена, теллура), расположенных в VI группе Периодической системы элементов Д.И. Менделеева. систематизированные В данные настоящее о время биологической отсутствуют активности халькогенорганических соединений. Целью исследования явилось установление биологической активности халькогенорганических соединений в зависимости от их структуры, включая наличие гетероатома халькогена (серы, селена, теллура) и боковых заместителей (атомов хлора, фтора и нитрогрупп) в составе этих соединений. Задачи исследования: 1. Изучить влияние халькогенорганических соединений на антиоксидантный статус и процессы свободнорадикального окисления в тканях белых мышей. 2. Установить эффекты халькогенорганических соединений на отдельные стороны обмена веществ и функциональное состояние 9 тканей крыс при интоксикации сульфатом кадмия, нитратом свинца и нитратом ртути, а также оценить интоксикацию тяжелыми металлами и антитоксическое действие халькогенорганических соединений по гематологическим показателям крови крыс и лейкоцитарным индексам интоксикации. 3. Выявить способность халькогенорганических соединений подавлять рост клинических штаммов Staphylococcus aureus, Escherichia coli и Pseudomonas aeruginosa. 4. Установить влияние халькогенорганических соединений и глюкокортикоидных препаратов на отдельные стороны углеводного и белкового обмена мышей. 5. Провести компьютерное прогнозирование биологической активности халькогенорганических соединений при помощи системы PASS C&T, а также квантовохимические расчеты, выявляющие структурнофункциональные аналогии препарата ДАФС-25, его производных и ряда стероидных гормонов. 6. Установить структурно-функциональные закономерности, объясняющие разнообразные биологические свойства органических соединений, содержащих гетероатомы халькогенов (серы, селена и теллура), а также различные боковые заместители (атомы хлора, фтора и нитрогруппы). Научная новизна работы. В работе впервые установлена взаимосвязь между структурой органических соединений, содержащих гетероатомы халькогенов (серы, селена и теллура), различные боковые заместители (атомы хлора, фтора и нитрогруппы) и их биологической активностью. Впервые доказано, что нитро- и хлорсодержащие производные ДАФС-25 оказывают антиоксидантное действие, которое выражается в снижении концентрации продуктов перекисного окисления липидов и увеличении 10 активности ферментов супероксиддисмутазы, каталазы и глутатионпероксидазы в тканях и крови мышей. Впервые установлена антитоксическая активность нитро- и хлорсодержащих производных ДАФС-25, а также его серосодержащего аналога при интоксикации экспериментальных животных солями тяжелых металлов: сульфатом кадмия, нитратом свинца и нитратом ртути. Впервые показана зависимость антибактериальной активности соединений от их токсичности. В ряду селеноорганических соединений наиболее токсичный препарат - фторированный аналог ДАФС-25, а в ряду халькогенпроизводных ДАФС-25 - теллурорганическое соединение, которые подавляли рост колоний клинических штаммов Staphylococcus aureus, Escherichia coli и Pseudomonas aeruginosa даже в низких концентрациях 0,00010,001 мг/мл. В работе впервые проведен компьютерный анализ биологической активности селен-, серо и теллуроорганических соединений, а также квантовохимические расчеты, выявляющие структурно-функциональные аналогии препарата ДАФС-25 (1,5-дифенил-3-селенапентандион-1,5), его производных и ряда стероидных гормонов. Установлено сходное действие ДАФС-25 и глюкокортикоидных препаратов на отдельные стороны углеводного и белкового обменов экспериментальных животных. Основные положения, выносимые на защиту. 1. Халькогенорганические соединения в зависимости от наличия гетероатома халькогена (серы, селена и теллура) и боковых заместителей (атомы хлора, фтора и нитрогруппы) в различной степени оказывают влияние на антиоксидантный статус и процессы свободнорадикального окисления в тканях белых мышей. Селеноорганические соединения 1,5-дифенил-3- селенапентандион-1,5 (ДАФС-25); 1,5-ди-(м-нитрофенил)-3-селенапентандион1,5; 1,5-ди-(п-хлорфенил)-3-селенапентандион-1,5 антиоксидантные свойства. демонстрируют лучшие 11 2. Селеноорганические соединения 1,5-дифенил-3-селенапентандион-1,5; 1,5-ди-(м-нитрофенил)-3-селенапентандион-1,5; 1,5-ди-(п-хлорфенил)-3- селенапентандион-1,5 обладают низкой токсичностью и при предварительном введении per os крысам снижают интоксикацию солями тяжелых металлов, о чем свидетельствуют биохимические, гематологические показатели крови крыс и лейкоцитарные индексы интоксикации. Серосодержащий аналог препарата ДАФС-25 (1,5-дифенил-3-тиапентандион-1,5) оказывает аналогичное действие, тогда как теллурсодержащее соединение в силу своей собственной токсичности не проявляет антитоксической активности при интоксикации солями тяжелых металлов. 3. Наибольшей токсичностью отношении клинических штаммов Pseudomonas aeruginosa и антибактериальной активностью в Staphylococcus aureus, Escherichia coli и обладают соединения 1,5-ди-(п-фторфенил)-3- селенапентандиона-1,5 и 1,5-дифенил-3-теллуропентандиона-1,5 даже в низких концентрациях 0,0001-0,001 мг/мл. 4. Соединение ДАФС-25 оказывает стимулирующее действие на отдельные стороны углеводного и белкового обменов мышей подобно глюкокортикоидному препарату преднизолону. 5. Согласно квантово-химическим расчетам, молекула препарата ДАФС-25 (1,5-дифенил-3-селенапентандиона-1,5) наиболее близка к молекулам глюкокортикоидов преднизону и преднизолону как по размеру, так и по расстояниям между полярными реакционноспособными группами, которые потенциально могут участвовать в связывании с рецептором как посредством электростатических сил, так и за счет водородных связей. 6. В ряду халькогенорганических заместителей соединений, лишенных боковых (1,5-дифенил-3-тиапентандион-1,5;1,5-дифенил-3- селенапентандион-1,5 и 1,5-дифенил-3-теллурапентандион-1,5), а также в ряду ДАФС-25 содержащих (1,5-дифенил-3-селенапентандиона-1,5) боковые заместители и его производных, (1,5-ди-(м-нитрофенил)-3- селенапентандиона-1,5; 1,5-ди-(п-хлорфенил)-3-селенапентандиона-1,5 и 1,5- 12 ди-(п-фторфенил)-3-селенапентандиона-1,5), установлены следующие закономерности: снижаются антиоксидантная и антитоксическая активности, возрастает антибактериальное действие. Теоретическая и практическая значимость. Полученные в настоящей работе данные представляют существенный интерес для фундаментальной науки в понимании молекулярных механизмов реализации биологической активности органических соединений серы, селена и теллура. Экспериментально показана антитоксическая активность соединения 1,5дифенил-3-тиапентандиона-1,5, что можно связать с наличием в структуре этого соединения фенильных радикалов, лишенных боковых заместителей. Результаты исследований позволяют рекомендовать соединения 1,5-ди-(м-нитрофенил)-3- селенапентандион-1,5; 1,5-ди-(п-хлорфенил)-3-селенапентандион-1,5 и 1,5дифенил-3-тиапентандиона-1,5 для дальнейших исследований с перспективой использования в качестве протекторов для предотвращения отравления тяжелыми металлами на производстве и средств, повышающих окислительную резистентность организма, селенапентандиона-1,5 и а соединения 1,5-ди-(п-фторфенил)-3- 1,5-дифенил-3-теллуропентандиона-1,5 могут применяться при госпитальных инфекциях, либо в качестве бактериостатических средств в медицинской практике и ветеринарии. Получены свидетельство на полезную модель № 22478 от 21.11.2001 «Кювета для выращивания микроорганизмов» и патент № 016974 от 26.04.2007 «Средство для лечения и профилактики отравлений соединениями тяжелых металлов». Материалы диссертации используются в учебном процессе на кафедре биохимии ГБОУ ВПО «Саратовский ГМУ им. В.И. Разумовского» Минздрава России. Апробация диссертационной работы. Материалы диссертации докладывались на межрегиональной научнопрактической конференции с международным участием «Молодежь и наука: итоги и перспективы» (Саратов, 2007, 2008, 2009, 2010), Российской конференции, посвященной 80-летию со дня рождения Р.И. Лифшица «Актуальные проблемы теоретической и прикладной биохимии» 13 (Челябинск, 2009), 2-ой международной телеконференции «Фундаментальные науки и практика» (Томск, 2010), IХ межвузовской конференции с международным участием «Обмен веществ при адаптации и повреждении» (Ростов-на-Дону, 2010), VII международной научно-практической конференции «Ключевые вопросы в современной науке» (София, 2011), Международной научно-практической конференции «Современные проблемы отечественной медико-биологической и фармацевтической промышленности. Развитие инновационного и кадрового потенциала Пензенской области» (Пенза, 2011), V Международной научно-практической конференции «Актуальные проблемы биологии, нанотехнологий и медицины» (Ростов-на-Дону, 2013), Международной конференции «Фундаментальные и прикладные исследования в медицине» (Франция, Париж, 2013), Международной конференции «Фундаментальные и прикладные проблемы медицины и биологии» (ОАЭ, Дубай, 2013), Международной конференции «Фундаментальные исследования» (Израиль, Тель-Авив, 2013). Публикации. По теме диссертации опубликовано 39 работ, в том числе 13 статей в изданиях, рекомендованных ВАК, 1 монография; получены 2 патента на изобретения № 016974 от 26.04.2007 и №2011118470/15 от 06.05.2011 и свидетельство на полезную модель № 22478 от 21.11.2001. Общий объем публикаций 13,19 п.л., личный вклад 85%. Структура и объем диссертации. Диссертация изложена на 318 страницах машинописного текста и состоит из введения, обзора литературы, описания постановки экспериментов, методов исследования, результатов собственных исследований, заключения, выводов и списка литературы. Работа содержит 43 таблицы и иллюстрирована 19 рисунками. Список использованной литературы включает 528 наименований, в том числе 431 зарубежный источник. 14 1.1. Глава 1. Обзор литературы Халькогены. Физико-химические свойства, биологическая активность, метаболизм в клетках эукариот и прокариот Общая характеристика халькогенов Халькогены расположены в VI главной подгруппе (или 16-ой группе по новой номенклатуре ЮПАК) периодической системы элементов Д.И. Менделеева. К халькогенам относятся кислород (О), сера (S), селен (Se), теллур (Te) и полоний (Ро). Групповое название этих элементов - халькогены (термин "халькоген" происходит от греческих слов "chalkos"-медь и "genos"рожденный), то есть "рождающие медные руды", обусловлено тем, что в природе они встречаются чаще всего в форме соединений меди (сульфидов, оксидов, селенидов и т.д.) [Fischer W., 2001]. У атомов халькогенов одинаковое строение внешнего энергетического уровня — ns2nр4. Электронная конфигурация серы - 3s23р4 , селена - 3d10 4s2р4, теллура - 4d105s25р4. Следовательно, селен и теллур являются полными электронными аналогами, что определяет общность химических свойств этих элементов и их соединений. В отличие от теллура селен и сера являются неполными электронными аналогами, полная электронная аналогия между этими элементами наблюдается только в низшей и нулевой степенях окисления. К тому же для серы и селена в низшей степени окисления характерны очень близкие размеры ионных радиусов (0,184 нм и 0,198 нм соответственно). Этим объясняется сходство их химических и биологических свойств. Все халькогены в соединениях с водородом и металлами проявляют степень окисления -2, а в соединениях с кислородом и другими активными неметаллами — обычно +4 и +6. Такие значения степеней окисления следуют из электронного строения халькогенов [Гринвуд Н.Н., Эрншо А., 2008]. Некоторые свойства атомов халькогенов представлены в таблице 1. 15 Таблица 1. Сравнительные характеристики элементов VI группы, главной подгруппы (Гринвуд Н.Н., Эрншо А., 2008). Сравнительные характеристики 8Окислород 16Sсера 34Seселен 52Teтеллур 84Poполоний Электронное строение 1S22S22p4 3S23p4 3d104S24p4 4d105S25p4 4f145d106S26p4 Степень окисления Физические свойства -2, +2, +4,+6 -2, +2, +4 * ― только в оксиде F2O Редкий элемент, Самородная сера; сульфиды: содержится в В свободном состоянии ― в Редкий элемент, свинцовый блеск PbS, медный малом атмосфере (О2 ― кислород, содержится в Редкий блеск Сu2S, пирит FeS2, количестве в О3 ― озон), в связанном ― в малом количестве в радиоактивсероводород H2S; сульфаты: самородной составе Н2О, SiO2, в сложных самородной сере, ный элемент гипс CaSO4∙2H2O, горькая соль сере, соединениях сульфидных рудах MgSO4∙7H2O, белки сульфидных рудах О2 ― бесцветный газ без вкуса и запаха, умеренно растворим в воде, парамагнитен. O3 ― газ синего цвета, диамагнитен, сильный окислитель ρ = 9,3 г/см3, мягкий, S-ромбическая ― желтые Кристаллическое ρ = 6,3 г/см3, серебристохрупкие кристаллы без запаха, вещество с серебристобелый, нерастворима в воде, ρ = 2 металлическим белый, хрупкий радиоактивг/см3; S-пластическая ― блеском, ρ = 4,8 с металическим ный металл 3 коричнево-желтая, г/см , темно-серый, блеском резиноподобная полупроводник полупроводник 15 Нахождение в природе -1, -2, +2* 16 Продолжение таблицы Химические свойства По активности уступает только фтору, реагирует со всеми простыми веществами, И окислительные, и (исключение: галогены, Pt , восстановительные свойства Au, инертные газы) и со многими сложными веществами Применение Для получения серной Для получения серной и кислоты; азотной кислот; в сельском хозяйстве для выплавки чугуна и стали; для борьбы с вредителями; для резки и сварки металла; производство спичек; как окислитель ракетного вулканизация каучука; топлива; в производстве черного в органическом синтезе. пороха. В промышленности: из самородных руд. В лаборатории: окислением сероводорода, сульфидов Se + O2 → SeO2 Se + Cl2 → SeCl2 Слабо выраженные металические свойства Проявляет свойства металла Se + H2 → H2Se 16 Получение Фракционная перегонка жидкого воздуха; в лаборатории ― при термическом разложении CrO3, KNO3, RClO3, BaO2 Свойства типичного неметалла Из отходов цветной металлургии и сернокислой промышленности, особо чистые ― дистилляцией в вакууме и зонной плавкой Производство фотоэлементов и выпрямителей электрического тока В полупроводниковой технике Искусственно облучением висмута в ядерных реакторах 17 При переходе от кислорода к полонию размер атомов и их возможные координационные числа увеличиваются, а энергия ионизации (Еион) и электроотрицательность уменьшаются. По электроотрицательности кислород уступает лишь атому фтора, а атомы серы и селена также азоту, хлору, брому; кислород, сера и селен относятся к типичным неметаллам. У атома кислорода на 2р-подуровне два неспаренных электрона. Его электроны не могут разъединяться, поскольку отсутствует d-подуровень на внешнем (втором) уровне, т. е. отсутствуют свободные орбитали. Поэтому валентность кислорода всегда равна двум, а степень окисления -2 и +2 (например, в Н2О и ОF2). Таковы же валентность и степени окисления у атома серы в невозбужденном состоянии. При переходе в возбужденное состояние (что имеет место при подводе энергии, например при нагревании) у атома серы сначала разъединяются 3 р-, а затем 3 s-электроны. Число неспаренных электронов, а, следовательно, и валентность в первом случае равны четырем (например, в SO2), а во втором — шести (например, в SO3). Очевидно, четные валентности 2, 4, 6 свойственны аналогам серы — селену, теллуру и полонию, а их степени окисления могут быть равны -2, +2, +4 и +6. Se, Te и Po соединяются непосредственно с большинством простых веществ, хотя и труднее, чем О и S. Среди их соединений наиболее устойчивы селениды, теллуриды и полониды металлов, но их устойчивость ниже, чем у аналогичных оксидов и сульфидов. В подгруппе халькогенов сверху вниз с увеличением заряда атома закономерно изменяются свойства элементов: уменьшается их неметаллический характер и усиливаются металлические свойства. Так кислород ― типичный неметалл, а полоний ― металл (радиоактивен). Биологическая роль халькогенов (серы, селена и теллура) Биологическая роль серы исключительно велика. Сера является биогенным элементом, выполняющим важные функции: входит в состав серосодержащих аминокислот метионина, цистеина и цистина, витаминов 18 (тиамин, биотин, липоевая кислота) и кофермента А (КоА~SH), а также гормонов (инсулин, вазопрессин, окситоцин и некоторые другие). Сера участвует в формировании конформации белковой молекулы благодаря образованию дисульфидных связей, участвует в формировании активных центров ферментов за счет сульфгидрильной (–SH) группы остатка аминокислоты цистеина. Сера в составе метионина участвует в транспорте метильных групп в процессе синтеза лецитина [Petti A.A. et al, 2012]. Аминокислота цистеин в составе глутатиона участвует в окислительновосстановительных реакциях. Железосерные белки являются компонентом митохондриальной цепи переноса электронов и участвуют в процессе тканевого дыхания. Сера способна к образованию богатых энергией связей в макроэргических соединениях. Производное аминокислоты цистеина - биогенный амин таурин - участвует в синтезе парных желчных кислот. Серосодержащие соединение фосфоаденозинфосфосульфат (ФАФС) участвует в обезвреживании токсинов в печени [Jung Y.S. et al., 2013]. Сера является компонентом простетической группы структурного белка коллагена хондроитинсульфата [Erickson J.M., Messer T.M., 2013]. Хондроитинсульфат присутствует в коже, хрящах, ногтях, связках и клапанах миокарда. Важными серосодержащими метаболитами также являются гемоглобин, гепарин, цитохромы, фибриноген, иммуноглобулины и сульфолипиды. В медицинской практике применение серы основано на ее способности при взаимодействии с органическими веществами организма образовывать сульфиды и пентатионовую кислоту, от присутствия которых зависят кератолитические (растворяющие - от греч. keras - рог и lytikos растворяющий), противомикробные и противопаразитарные эффекты. Сера входит в состав мази Вилькинсона и других препаратов, применяемых для лечения чесотки [Утц С.Р. и др., 2011]. Очищенную и осажденную серу употребляют в мазях и присыпках для лечения некоторых кожных заболеваний (себорея, псориаз и других); в порошке - при глистных инвазиях (энтеробиоз); в растворах - для пиротерапии прогрессивного паралича и других. 19 Сера входит в состав многих других лекарственных фармпрепаратов седативного, нейролептического, противоопухолевого действия (тиопентал, тиопроперазин, тиоридазин и др.) [Костюченко С.И., 2013]. Соединения серы в виде сульфаниламидных препаратов (бисептол, сульфацил-натрия, сульгин и др.) обладают противомикробной активностью [Лекарь А.В. и др., 2011]. Биологическая роль теллура изучена недостаточно. Предположительно теллур (и ряд других элементов: золото, олово, таллий, германий, галлий) участвует в проводимости импульсов головного мозга млекопитающих и потому относится к группе брейн-элементов [Щербаков В.А., 1991]. Интересны данные В.А. Щербакова (1991) [Щербаков В.А., 1991.], который, изучая особенности геохимии Атлантического океана, пришел к выводу, что теллур входит в состав некоторых водорослей Атлантики. Лекарственные препараты, содержащие в качестве активных ингредиентов соединения теллура, используются для лечения гастритов и язвенной болезни желудка, а также злокачественных опухолей [Махаева Н.А., 2007]. Установлено, что избыток теллура ведет к замещению серы в тиоловых группах, что приводит к ингибированию ферментов. В организме соединения теллура восстанавливаются до элементарного теллура, который выделяется постепенно в виде различных производных, обладающих сильным чесночным запахом. Биологическая роль селена. Селен - жизненно важный микроэлемент с уникальными биологическими функциями и широким спектром биологического действия его соединений. Являясь природным антиоксидантом, селен участвует в защите организма от возникновения и развития кардиологических и некоторых онкологических заболеваний, в метаболизме тиреоидных гормонов, репродукции, выведению тяжелых металлов из организма, устойчивости к вирусным заболеваниям, поддержании иммунитета и пр. [Капитальчук М., 2008; Schrauser G.N., 2003]. 20 Заключение об антиоксидантной роли селена, впервые представленное английским биохимиком А. Диплоком в 1970 г. [Diplock A.T., 1970], до настоящего времени остается основополагающим в метаболических функциях соединений селена. Биологическая активность селена связана в большинстве случаев с экспрессией по крайней мере 30 селенопротеинов (таблица 2), закодированных в 25 генах человека [Reeves M.A., Hoffmann P.R., 2009]. Таблица 2. Селенопротеины млекопитающих и их функции [Риш М.А., 2003; Kryukov G.V. et al., 2003; Reeves M.A., Hoffmann P.R., 2009; Shchedrina V.A. et.al., 2010]. Селенопротеин Глутатиопероксидазы (ГПО) ГПО 1 ГПО2 ГПО3 ГПО4 ГПО5 ГПО6 Тиоредоксинредуктазы (ТРР) Предполагаемая функция Антиоксидант в цитозоле клеток Антиоксидант в ЖКТ Антиоксидант в межклеточном веществе и плазме Мембранный антиоксидант, структурный белок спермы (участие в апоптозе?) Неизвестна Гомолог ГПО1 Многие функции, активность дитиол-дисульфид оксидоредуктазы, обезвреживание пероксидов, восстановление тиоредоксина (контроль клеточного роста), поддержание редокс статуса транскрипционных факторов. Цитозольная (во многих клетках) Тестикулы Митохондриальная (во многих клетках) ТР1 ТР2 ТР3 Йодотирониндейодиназы Типы D1 и D2 Превращение тироксина (Т4) в биоактивный 3,5,3`-три-йодониронин (Т3) Типы D1 и D3 Превращение тироксина (Т4) в неактивный реверсивный рТ3 Селенопротеин Р Главный селенотранспортный белок Селенопротеин W Антиоксидант в сердечной и скелетных мышцах Селенофосфатсинтаза Синтез селенофосфата для синтеза селенопротеинов 15 кДа Селенопротеин Защита против рака H, I, K, M, N, O, R, S, T, Роли широко неизвестны V 21 Селенопротеины содержат селен в качестве остатка селеноцистеина, который полностью ионизирован при физиологическом значении рН и действует как очень эффективный редокс катализатор. Из более чем 30 описанных селенопротеинов шесть являются глутатионпероксидазами (ГПО), три – йодотиронин дейодиназы и три – тиоредоксинредуктазы (ТРР). Селенопротеин Р – количественно главный селенопротеин плазмы крови выполняет как антиоксидантную, так и транспортную функции [Kryukov G.V. et al., 2003; Reeves M.A., Hoffmann P.R., 2009; Shchedrina V.A. et.al., 2010]. Таким образом, селен может влиять на три основные области клеточной биохимии: антиоксидантную функцию, редокс-статус и метаболизм в щитовидной железе [Риш М.А., 2003; Patching S.G., Gardiner P.H.E., 1999; Shchrauser H.W., 2003; Beckett G.J., Arthur J.R., 2005]. Селенсодержащие ферменты и белки. Первым открытым у эукариот селенспецифическим белком оказался фермент глутатионпероксидаза (ГПО, GPX) эритроцитов, известная в настоящее время как глутатионпероксидаза 1 (ГПО1, GPX-I). Она катализирует реакцию: H2O2 + 2 GSH → GSSG+ 2 H2O. Главная детоксикации биологическая роль реакционноспособного ГПО-I, таким оксиданта - образом, перекиси состоит в водорода, образующейся в эритроцитах в ходе транспорта кислорода к тканям. [Mills G.C., 1957; Halliwel B., Gutteridge J.M.C., 1984; Frei B. et al, 1988]. Классическая ГПО1 - это белок с молекулярной массой около 85 кДа, образованный четырьмя одинаковыми субъединицами с молекулярной массой 21 кДа, организованными в комплекс квадратной формы. Каждая субъединица содержит 1 атом селена в составе селеноцистеина. Окрестности этого остатка представляют собой очень консервативную в межвидовом отношении последовательность, что подчеркивает важность остатка селеноцистеина в формировании активного центра фермента [Кулинский В.И., Колесниченко Л.С., 1993; Flohe L., 1982; Chaudiere J., Tappel A.L., 1983; Reddy C.C. et al., 1990]. 22 Секвенирование гена ГПО1 человека показало, что остаток селеноцистеина (Se-Cys) кодируется триплетом UGA, который в огромном большинстве других случаев означает остановку трансляции [Amberg R., et al., 1996]. Тем не менее, в случае ГПО и других селенопротеинов при этом происходит “перекодирование” триплета под действием специальной структуры в составе мРНК, обозначенной как SECYS. За счет этого происходит присоединение специфической тРНКuga, ацилированной остатком серина, и его последующее встраивание в полипептидную цепь, сопряженное с заменой ОНгруппы серина на SeH остатка селенофосфата. Процесс “перекодирования” опосредуется, как предполагают, особым SECYS-связывающим белковым фактором SBP. Необходимым условием для этого является также взаимодействие рибосомы со специфическим белком - фактором элонгации SelB и “защищающим тРНК” фактором SePF [Larsen P.R., Berry M.J., 1995]. В большинстве случаев синтез всех необходимых компонентов системы включения Se-Cys в белки (т.е. тРНКuga, мРНК селенопротеина, фактора элонгации Sel-B и др.) регулируется поступлением селена с рационом на транскрипционном уровне. Внутриклеточный и тканевой уровень ГПО1 также влияет на активность апоптотических путей, фосфорилирование протеинкиназ. Следует подчеркнуть, что гиперэкспрессия ГПО1 приводит к развитию инсулинорезистентности и ожирению. Накоплены экспериментальные данные о связи изменения экспрессии ГПО1 с этиологией рака, сердечно-сосудистых и аутоиммунных заболеваний, диабета и пр. В составе гранулоцитов человека недавно был идентифицирован селенопротеин, представляющий собой пероксидазу, для которой донором водорода является не глутатион, а различные органические восстановители, в частности, о-дианизидин. Белок состоит из 2 идентичных субъединиц по 15 кДа, и селен входит в их состав в виде селеноцистеина. Антиокислительные качества селена не ограничиваются реакцией катализа в активном центре глутатионпероксидазы. Селен может применять 23 свое основное химическое сходство в создании молекул, содержащих в составе серу. Серусодержащие аминокислоты, как метионин и цистин, легко образуют с селеном комплекс и при этом сохраняют антиокислительные качества с глутатионпероксидазой. Такие комплексы сохраняют не только структуру и функцию биологической мембраны, но и предотвращают окисление и инактивацию многих ферментов. Уникальным свойством глутатионпероксидазы является ее способность разрушать разнообразные гидроперекисные субстраты, в том числе гидроперекиси кортикостероидных гормонов, холестерола, тиамина, ДНК. Сопутствующая глутатионпероксидазе каталаза разрушает лишь перекись водорода, которая образуется как при нормальном протекании обмена веществ, так и в результате действия супероксиддисмутазы. Разрушая высокотоксичные гидроперекиси липидов, образующиеся в качестве нормальных промежуточных продуктов в обмене веществ тканей животных (например, при синтезе простагландинов из арахидоновой кислоты), глутатионпероксидаза предотвращает образование свободного гидроксилрадикала и синглетного кислорода, дезинтегрирующих клеточные и субклеточные мембраны и приводящие к гибели клетки [Kryukov G.V. et al., 2003; Reeves M.A., Hoffmann P.R., 2009; Shchedrina V.A. et.al., 2010]. Помимо глутатионпероксидазы селен входит в состав таких ферментов как: глутатионредуктаза, тиоредоксинредуктаза, йодтирониндейодиназа и др., играющих ключевую роль в обмене веществ. В 1991 г. было доказано, что один из важных ферментов, ответственных за обмен тиреоидных гормонов - 5’-йодтиронин дейодиназа щитовидной железы типа I является селеноэнзимом [Berry M.J. et al., 1991, Berry M.J. et al., 1991]. Молекулярная масса этого белка равна 77 кДа. В настоящее время установлено, что эта дейодиназа I входит в семейство близкородственных ферментов, содержащих в своем активном центре селеноцистеин. Роль селена в функционировании тканевых дейодиназ подчеркивает тесную связь обмена 24 этого микроэлемента с обменом йода [Contempre B. et al., 1992; Salvatore D. et al., 1995, 1996; Bates J.M. et al., 2000]. Тиоредоксинредуктазы (ТРР, TrxR) относятся к семейству пиридиновых оксидоредуктаз. Особенностью тиоредоксинредуктаз является наличие С-концевого цистеином селеноцистеинового формирующим Тиоредоксинредуктазы специфичностью: низкомолекулярные активный отличаются помимо остатка, вместе с соседним очень тиоредоксина, соединения, селеносульфидный широкой они окисленные сайт. субстратной восстанавливают гидроперекиси, многие являются ключевыми ферментами метаболизма селена [Kryukov G.V. et al., 2003; Reeves M.A., Hoffmann P.R., 2009; Shchedrina V.A., et.al., 2010]. Селензависимая тиоредоксинредуктаза (ТРР) была впервые выделена из культуры клеток аденокарциномы человека в 1996 г [Tamura T., Stadtman T.C.A., 1996]. ТРР представляет собой гомодимер, состоящий из двух субъединиц молекулярной массой 55-58 кДа. В состав активного центра фермента, наряду с Se-Cys, входит простетическая группа ФАД. Селенопротеин Р был открыт в 1977 г., а выделен в чистом виде и подробно изучен в 1987 году. Белок циркулирует в плазме крови; его биосинтез осуществляется, по-видимому, в ряде органов, включая легкие, почки, печень, сердечную мышцу и др. Молекулярная масса основной изоформы селенопротеина Р - 57 кДа, и в его составе - 10 или 11 атомов селена. Присутствующий в сыворотке крови крысы селенопротеин Р представлен основной (57 кДа) и дополнительной, “укороченной” с С-конца изоформами. Каждая из них, в свою очередь, подразделяется на 2-3 субфракции, различающиеся своими углеводными компонентами. Нормальная концентрация селенопротеина Р в плазме - 51±4 мкг/мл; при глубоком дефиците селена она падает ниже 5 мкг/мл. Биологической функцией селенопротеина Р, по современным представлениям, является, во-первых, защита организма от воздействия перекисей (окислительного стресса), осуществляемая по иному механизму, 25 нежели в случае глутатионпероксидаз. Во-вторых, данный белок выступает в роли агента, способствующего нейтрализации токсического действия тяжелых металлов (Pb, Hg) [Campbell J.W. et al., 2010]. Недавно был выделен, охарактеризован и полностью секвенирован селенопротеин W. Это небольшой белок (M=9,5 кДа), экспрессированный, главным образом, в мышечной ткани. Предполагается, что селенопротеин W, подобно селенопротеину Р, обладает функцией антиоксиданта, но не в кровотоке, а на тканевом уровне [Brown K.M., Arthur J.R., 2001]. Функции других селенопротеинов менее изучены. Известно, однако, что селенопротеин H играет роль редокс-зависимого регулятора транскрипции для генов глутатиона и детоксикации. Селенопротеин K также является антиоксидантом, преимущественно в кардиомиоцитах. Генетические дефекты селенопротеина S являются фактором риска кардиоваскулярных заболеваний, особенно у женщин [Kryukov G.V. et al., 2003; Reeves M.A., Hoffmann P.R., 2009; Shchedrina V.A. et.al., 2010]. Таким образом, биологическая роль селена связана с функционированием селенсодержащих белков и ферментов [Решетник Л.А., Парфенова Е.О., 2000; Brown K.M., Arthur J.R., 2001], необходимых для окислительно- восстановительных реакций, обезвреживания тяжелых металлов, репродукции, функционирования щитовидной железы, иммунной системы и т.д. Метаболизм соединений халькогенов в клетках эукариот Метаболические превращения халькогенов и их соединений в эукариотических клетках, в том числе в клетках млекопитающих и человека, напрямую зависят от их физико-химических свойств. Как уже отмечалось в начале главы, в группе халькогенов в направлении сера, селен, теллур возрастает атомный радиус и поляризация связей, но снижается электроотрицательность, а это означает, что селен и теллур образуют более слабые связи, чем сера. Причем теллур обладает более выраженными 26 металлическими свойствами среди всех халькогенов и является истинным металлоидом [Cunha R.L.O.R. et al., 2009]. Поляризованность связей «Углеродхалькоген» и их прочность снижаются в направлении: «C-S»>«C-Se»>«C-Te», особенно в случае алкил-производных [Cunha R.L.O.R. et al., 2009]. Реакционная способность атомов серы и селена во многом зависит от константы диссоциации (pKa) тиольных и селенольных групп. В отличие от гидроксильной группы, которая в биологических системах чрезвычайно редко бывает депротонирована, сульфгидрильная группа цистеина и селеногидрильная группа селеноцистеина часто оказываются в анионном состоянии в физиологических условиях благодаря низким значениям pKa. Эти низкие значения pKa при физиологических значениях рН усиливают реактивность не только цистеина и селеноцистеина, но и их тио- и селенопроизводных. Например, тиоэфиры в 100 раз более реактивны к амино- и тиолатным нуклеофилам, чем соответствующие оксоэфиры. А селенолат (RSe-) более нуклеофилен, чем его тиолатный аналог (RS-) и благодаря более низкому значению pKa селеноцистеин быстрее включается в биохимические процессы, чем цистеин [McGrath N.A., Raines R.T., 2011]. Следовательно, реакционная активность халькогенов возрастает в направлении от кислорода к теллуру. Есть мнение, что метаболические пути превращения селенита (Se(IV)) и теллурита (Te(IV)) очень близки и тесно связаны с метаболизмом серы [Ottosson L.-G. et al., 2010]. Например, в опытах на дрожжевых клетках показано образование теллур-метионина, который способен замещать в белках до 40% остатков метионина, а внедрение теллуро-цистеина в активный центр глутатионтрансферазы приводит к изменению ее активности [Liu X. et al., 2009]. Кроме этого введение в молекулу глутатиона остатка теллуроцистеина вызывало нарушение его восстанавливающей способности. Однако этих данных недостаточно, превращениями чтобы теллура [Ottosson L.-G., et al., 2010]. в проводить организмах аналогии высших с метаболическими эукариот и человека 27 Несмотря на то, что в последнее десятилетие активно изучается биохимия халькогенов, наибольший интерес у научного сообщества вызывают метаболические превращения селена [Mehdi Y. et al., 2013]. Селен поступает в организм человека в составе пищевых продуктов в органической форме селенометионина, селеноцистеина и селенометилселеноцистеина. Каждое из этих селенсодержащих соединений может метаболизироваться до селеноводорода/гидроселенид-аниона, который является необходимым субстратом для биосинтеза селенопротеинов [Sunde R.A., 2006; Zeng H. et al., 2013]. Организмы млекопитающих и человека лучше абсорбируют и аккумулируют органические формы селена, чем его неорганические соли [Finley J.W., 2006; Zeng H., 2009]. Органические соединения селена содержат селен в восстановленном состоянии (селенида Se(-II)), тогда как неорганические соли содержат селен в окисленных формах (селенита (IV) и селената (VI)). Селенат- и селенит-анионы, поступающие с пищей, быстро восстанавливаются под действием белка тиоредоксина до селеноводорода, находящегося при физиологических значениях рН, в основном, в виде гидроселенид-аниона (HSe-). Необходимым кофактором данного процесса является восстановленный глутатион (GSH). Причем предполагается, что в качестве интермедиата образуется селенодиглутатион (GS-Se-SG) [Тутельян В.А. и др., 2002; Weekley C.M. et al., 2013]. Некоторое количество образующегося селеноводорода присоединяется к особым селенсвязывающим белкам. Избыточные количества селеноводорода медленно подвергаются ферментативному метилированию с последовательным образованием метилгидроселенида, триметилселенония. Эти соединения диметилселенида Se и экскретируются с катиона мочой, а диметилселенид в больших количествах также и с потом. Строго определенное количество селена, входящего в состав пула селеноводорода, через стадию селенофосфата включается в высокоспецифический процесс синтеза 28 селенопротеинов. В состав этих белков селен входит у позвоночных исключительно в виде остатка селеноцистеина. Необходимо отметить, что поступивший с пищей селенометионин, как правило, превращается сначала в селеноцистеин, а последний метаболизируется с образованием селенида (H2Se) [Finley J.W., 2006]. Селенид/селеноводород может подвергаться метилированию или превращаться в селеносахара, которые выделяются с мочой [Ogra Y., Anan Y., 2012]. Некоторые биопроизводные селена (H2Se, CH3SeH и селенометионин) имеют большое значение для клетки, в том числе для регуляции клеточного цикла и апоптоза [Zeng H. et al., 2013]. На рисунке 1 представлены предполагаемые пути метаболизма биологически важных соединений селена (адаптировано на русс. яз. по Zeng H., 2009). Рис. 1. Предполагаемые пути метаболизма биологически важных соединений селена Следовательно, селенид является ключевой точкой двух метаболических путей превращения биоорганических и неорганических соединений селена. Первый путь отвечает за биосинтез селенопротеинов, который связан с котрансляционным включением селеноцистеина. Этот процесс основан на 29 каталитическом обмене гидроксильной группы остатка серина в серил-тРНК на селенольный остаток (-SеН) селеноцистеинил-тРНК под из селенофосфата действием с образованием селеноцистеинил-тРНК-синтазы [Sunde R.A., 2006]. Уникальность этого процесса объясняется тем, что аминокислота селеноцистеин кодируется кодоном УГА, который обычно выступает в роли стоп-кодона [Gladyshev V.N., Hatfield D.L., 1999; Hatfield D.L. et al., 2006]. Второй метаболический путь превращения селена связан с утилизацией избытка селена через реакции метилирования и образования селеносахаров, которые выводятся из организма с мочой [Kobayashi Y. et al., 2002; Sunde R.A., 2006; Ogra Y., Anan Y., 2012]. Таким образом, поступивший с пищей селен (в неорганической или биоорганической формах) подвергается последовательным метаболическим превращениям с образованием селенида и селенофосфата. Последний участвует в биосинтезе селеноцистеинил-тРНК, которая необходима для образования селенопротеинов и селеноферментов в разных органах и тканях. Селен в различных органах и тканях распределяется по-разному: 30% в печени, 30% в мышцах, 15% в почках, 10% в плазме крови и 15% в других органах и тканях [Mistry H.D. et al., 2012; Mehdi Y. et al., 2013]. Причем селенопротеины, содержащие селен в форме селеноцистеина, выполняют большое количество жизненно важных функций, включая антиоксидантную защиту, иммунный ответ, репродукцию и пр. [Mehdi Y. et al., 2013]. Представленные данные объясняют различия в эффективности органического и неорганического селена для человека. Поэтому, большинство авторов рекомендуют органическую форму селена как предпочтительную при снабжении организма селеном в профилактических целях [Гореликова Г.А. и др., 1997; Гмошинский И.В., Мазо В.К., 1999]. В связи с этим встает вопрос о необходимости эффективной профилактики и терапии селенодефицита с использованием низкотоксичных селеноорганических соединений. 30 В итоге многолетних исследований российскими учеными Б.И. Древко и А.Ф. Блинохватовым (1970-2000 гг.) (цит. по Т.Н. Родионовой, 2004) удалось синтезировать селенсодержащие органические вещества (диацетофенонилселенид и селенопиран), которые существенно отличаются от неорганических препаратов селена, свободны от недостатков. Они обладают значительно меньшей токсичностью, высокой липофильностью, что обеспечивает возможность их пролонгированного действия [Родионова Т.Н., 2004]. Селеноорганический позволяет нормализовать препарат диацетофенонилселенид деятельность иммунной, (ДАФС-25) антиоксидантной и детоксицирующей систем организма животных и птиц, приводит к увеличению яичной и мясной продукции. [Б.И. Древко и др., 2001]. ДАФС-25 в 40 раз менее токсичен селенита натрия, селен в нем находится в органической, более доступной для животных форме [Родионова Т.Н., 2004]. 1.2. Окислительный стресс: механизмы развития и предотвращения Антиоксидантная система организма (АОС) – это система, блокирующая образование высокоактивных свободных радикалов [Ratnam D.V. et al., 2006; Wagener F.A.D.T.G. et al., 2013], которые химически очень агрессивны и способны повреждать белки и ДНК а, главное, вызывать перекисное окисление липидов (ПОЛ) - процесс, ведущий к тяжелому повреждению клеточных мембран [Droge W., 2002; Jones D.P., 2008; Trachootham D. et al., 2008; Finkel T., 2011]. Более 50 лет назад Д. Харман в своей «Свободно-радикальной теории старения» описал активные формы кислорода (АФК), которые отвечают за повреждение клеточных структур и играют ключевую роль в процессе старения [Valko M. et al., 2007; Silva J.P., Coutinho O.P., 2010]. 31 В нормальных физиологических условиях выработка свободных радикалов и других активных форм кислорода и азота поддерживается равновесием с Gutteridge J.M.C., системой 1999]. антиоксидантной Однако, когда защиты развивается [Halliwell дисбаланс B., между окислителями и антиоксидантами в пользу окислителей можно говорить о развитии окислительного стресса [Sies H., 1997]. Это может произойти по двум причинам: либо из-за уменьшения содержания антиоксидантов (вследствие мутаций антиоксидантных ферментов или недостаточного поступления антиоксидантов с пищей), либо из-за чрезмерной выработки активных форм кислорода и азота (АФК и АФА), например, при увеличении содержания кислорода, присутствия токсинов, которые в ходе своих метаболических превращений вырабатывают АФК [Halliwell B., Gutteridge J.M.C., 1999]. Активные формы кислорода и азота Внутриклеточная аккумуляция АФК и АФА обусловлена как эндогенными, так и экзогенными факторами [Fleury C. et al., 2002]. К экзогенным факторам относятся сигаретный дым, загрязнение окружающей среды токсинами и излучение (УФ-, γ- и рентгеновское излечение), а к эндогенным факторам можно отнести воспалительный процесс (при участии нейтрофилов и макрофагов) и клеточный метаболизм, в первую очередь, митохондриальное окисление. Митохондрии восстанавливают до воды более 95% всего кислорода, находящегося в клетке, главным образом, благодаря функционированию цепи переноса электронов (ЦПЭ). Однако вследствие утечки небольшого количества электронов на кислород образуется супероксидный радикал, а из него и другие АФК, такие как перекись водорода и гидроксильный радикал [Cadenas E., Davies K.J., 2000; Somayajulu M. et al., 2005]. Действительно, митохондрии благодаря наличию переносчиков электронов могут «упускать» один электрон на кислород и превращать его в супероксид-анион. Такими переносчиками являются, например, цитохром b5 редуктаза и моноаминооксидаза (встроенные 32 во внешнюю мембрану митохондрии), глицерофосфатдегидрогеназа (расположенная на наружной поверхности внешней мембраны митохондрии), сукцинатдегидрогеназа (комплекс II ЦПЭ, расположенный во внутренней мембране митохондрии), аконитаза (фермент цикла Кребса, локализованный в матриксе) и α–кетоглутаратдегидрогеназный комплекс (связанный матриксной стороной внутренней мембраны митохондрии) с [Silva J.P., Coutinho O.P., 2010]. Тем не менее, основная часть АФК в миитохондриях вырабатывается в комплексах I и III ЦПЭ [Jezek P., Hlavata L., 2005]. Лучше описан вклад комплекса III (убихинол-цитохром с редуктаза) в генерацию супероксида. В этом комплексе перенос электрона от убихинона (UQH2) на цитохром с происходит посредством ряда реакций, известных как Q-цикл. Однако в ходе восстановление функционирования кислорода радикалом этого цикла, одноэлектронное (UQ•−) убисемихинон-аниона – нестабильного интермедиата Q-цикла – может приводить к формированию супероксида по обе стороны внутренней мембраны митохондрии [Andreyev A.Y. et al., 2005; Jezek P., Hlavata L., 2005]. Комплекс I (НАДН-дегидрогеназа, также называемая НАДН-убихинон оксидоредуктаза) – это первичный источник супероксида в мозге в нормальных условиях [Barja G., 1999; Turrens J.F., 2003;]. Этот комплекс окисляет НАДН, используя убихинон сопряженной с как акцептор протонной электронов в обратимой помпой, генерирующей реакции, трансмембранный потенциал. К настоящему времени замечено увеличение продукции АФК в этом комплексе в трех различных ситуациях: в ходе нормального функционирования ЦПЭ, в присутствии ротенона (ингибитора, который блокирует перенос электронов от комплекса I на убихинон) и в ходе обратного электронного транспорта (ряд реакций в ЦПЭ, которые позволяют электронам транспортироваться против градиента редокс потенциалов электронных переносчиков, от восстановленного убихинона к НАД+ вместо кислорода) [Silva J.P., Coutinho O.P., 2010]. 33 Однако основным генератором супероксидного радикала в клетках является НАДФН-оксидаза (NADPH-оксидаза - Nox) [Brown D.I., Griendling K.K., 2009; Jiang F. et al., 2011; Altenhofer S. et al., 2012]. Фермент НАДФН-оксидаза (Nox), активно продуцирующий супероксид, вслед за которым возникает вся гамма АФК, до недавнего времени считали сугубо специфическим «инструментом» фагоцитирующих клеток иммунной системы, объясняя необходимость продукции АФК защитой от патогенных микроорганизмов и вирусов [Griscavage J.M. et al., 1996]. Однако к настоящему моменту известно, что тканеспецифическая для ферментов экспрессия. этого Например, семейства Nox1 характерна преимущественно экспрессируется в толстой кишке и обнаруживается в ткани матки и простаты [Krause K.H., 2004; Schluter T. et al., 2010]; Nox2 — в нейтрофилах, моноцитах, макрофагах, эозинофилах [Pendyala S. et al., 2009]; кардиомиоцитах [Nabeebaccus A. et al., 2011]; Nox3 — исключительно в кортиевом органе и спиральных ганглиях внутреннего уха; Nox4 — в тканях почки, сердца, поджелудочной железы, поперечнополосатых и гладких мышцах, яичнике, яичках, эндотелии, остеокластах, фибробластах, астроцитах [Kleinschnitz C., 2010]; Nox5 — в лимфоидных тканях и яичке, преимущественно в сперматоцитах, а также в коронарных артериях [Guzik T.G. et al., 2008]; DUox1 — в ткани щитовидной железы и в эпителии респираторного тракта, DUox2 — в ткани щитовидной железы и в эпителии пищеварительного тракта [Griendling KK., 2006; Kawahara T. et al., 2007; Pendyala S., Natarajan V., 2010]. Механизмы рецептор-ассоциированной активации НАДФH-оксидазы наиболее изучены у фагоцитирующих клеток, у которых во время фагоцитоза наблюдается усиленная продукция АФК, получившая название «дыхательного или респираторного стимулирующими взрыва» [Yeligar S.M. et al., 2012]. Основными факторами механизмов активации НАДФH-оксидазы являются цитокины — трансформирующий фактор роста (TGF-1b), TNF-α и IL1b, пептидные факторы роста (PDGF, EGF, VEGF, bFGF и инсулин), агонисты G-протеинсвязанных рецепторов (GPCR — G-protein-coupled receptors) — 34 ангиотензин кислота, II, тромбин, эндотелин-1, серотонин, лизофосфотидиновая сфингозин-1-фосфат, гистамин, брадикинин; патоген- ассоциированные молекулярные структуры (РАМР) инфекционных агентов, активирующие Toll-подобные рецепторы (TLR). РАМР, в частности флагелин Pseudomonas aeruginosa, могут взаимодействовать с рецептором P2Y, что ведет к высвобождению АТФ, которая индуцирует НАДФН-оксидазу [Абатуров А. Е., 2009]. Активная НАДФH-оксидаза (Nox), осуществляя перенос одного электрона на молекулярный кислород, генерирует короткоживущий (с периодом полужизни в несколько 10-6 секунд) супероксидный радикал [Hong H. et al., 2006; Pamenter M.E. et al., 2012], который спонтанно или под влиянием супероксиддисмутазы (СОД) дисмутирует до перекиси водорода (H2O2) с последующим образованием гидроксильного радикала (ОН·) или кислорода и воды под действием каталазы и/или пероксидазы [Makino J. et al., 2012; Taverne Y.J.H.J. et al., 2013]. Другими источниками распространенный АФК являются ксантиноксидаза, широко в тканях млекопитающих фермент, который в ходе каталитического превращения гипоксантина в ксантин и в мочевую кислоту приводит к восстановлению молекулярного кислорода с образованием супероксида на первом этапе и перекиси водорода на втором [Valko M. et al., 2004]. Дополнительные источники клеточных АФК – это клетки нейтрофилы, эозинофилы и частично макрофаги (Conner E.M., Grisham M.B., 1996), метаболизм цитохрома Р450 [Moon Y.J. et al., 2006] и пероксисомы, которые, главным образом, вырабатывают перекись водорода [Gupta M. et al., 1997]. Химия и биохимия АФК и АФА Большинство внутриклеточных активных форм АФК и АФА включают радикальные (O2• –, •OH, •NO и др.) и нерадикальные формы (O2, ONOO–, H2O2, HOCl, O3), которые могут быть опасны для клеток [Butterfield D.A. et al., 1999, Silva J.P., Coutinho O.P., 2010]. 35 Свободные радикалы могут определяться как молекулы или молекулярные фрагменты, способные существовать независимо и имеющие один или более неспаренных электронов. Присутствие этих неспаренных электронов определяет их реактивность, т.к. им нужен другой электрон, чтобы заполнить орбиталь и стать стабильными [Wilcox J.K. et al., 2004; Valko M. Et al., 2006]. Интересно, что молекулярный кислород имеет уникальную электронную конфигурацию и потому рассматривается как активная форма. Принимая один электрон, молекулярный кислород формирует радикал супероксид-аниона, который становится высокоактивным свободным радикалом и утрачивает способность проходить сквозь липидные мембраны, поэтому он остается в компартментах, в которых образовался [Nordberg J., Arner E.S., 2001]. Радикал супероксид-аниона рассматривается как первичный АФК, из которого потом образуются вторичные формы, а также как участник многих метаболических процессов (ЦПЭ в митохондриях, флавин-зависимая ксантиноксидаза, липоксигеназа, циклооксигеназа и НАДФН-оксидаза) (рис. 2). В присутствии супероксиддисмутазы два супероксидных радикала могут легко трансформироваться в перекись водорода и молекулярный кислород [Spiteller G., 2001]. Синглетный кислород также очень активная АФК, которая индуцирует различные генотоксичные, канцерогенные и мутагенные эффекты посредством действия на полиненасыщенные жирные кислоты и ДНК [Cui K. et al., 2004]. Перекись водорода – это не свободный радикал. Тем не менее, он очень важен с точки зрения его способности к проникновению через биологические мембраны. Он играет роль в формировании более активных АФК, например, образование HOCl (гипохлорита) под действием миелопероксидазы (фермент, присутствующий в фагосомах нейтрофилов) и, что более важно, формирование гидроксильного радикала посредством окисления переходными металлами в реакции Фентона [Nordberg J., Arner E.S., 2001; Wilcox J.K. et al., 2004]. Перекись водорода обезвреживается по крайней мере тремя антиоксидантными 36 ферментными системами, а именно каталазой, глутатионпероксидазой и пероксиредоксинами [Nordberg J., Arner E.S., 2001] (рис. 2). Рис. 2. Общая схема образования и утилизации различных активных форм кислорода и азота. Пунктирные линии означают ингибирование. Супероксидный радикал (О2·-), образованный в процессе митохондриального окисления или под действием ферментов НАДФ-оксидазы, цитохром Р450монооксигеназы, миелопероксидазы, эндотелиальной синтазы оксида азота и пр., может дисмутировать с участием супероксиддисмутазы (СОД) до перекиси водорода (H2O2), которая затем разрушается ферментами каталазой и пероксидазой до воды и кислорода. Гидроксильный радикал (ОН·) формируется благодаря реакциям Фентона (1) и Хабер-Вейса (2), а также из пероксинитритной кислоты (HOONO). Оксид азота (NO) образуется из аргинина под действием синтазы NO. Супероксидный радикал (О2·-) также может взаимодействовать с NO, образуя пероксинитрит-анион (ONOO-), который способствует еще большему синтезу О2·- синтазой NO, и создавая тем самым «порочный круг» продукции АФК [Taverne Y.J.H.J. et al., 2013]. Гидроксильный радикал (•OH) высокоактивен, способен разрушать все биомолекулы: углеводы, липиды, белки и ДНК [Halliwell B., 2001]. Поскольку время полужизни этого радикала менее 1 наносекунды, он действует in vivo в 37 непосредственной близости от места своего образования [Valko M. et al., 2006]. Гидроксильный радикал может генерироваться несколькими путями, включая ионизирующее излучение и фотолитическое разложение алкилгидроперекисей, но главный путь образования •OH in vivo связан с реакцией Фентона (каталитическое разложение перекиси водорода при участии металлов, главным образом, железа и меди) [Nordberg J., Arner E.S., 2001; Valko et al., 2004]. Супероксид также играет важную роль в этой реакции за счет реутилизации ионов металлов. Переходные металлы выполняют важную функцию в формировании различных свободных радикалов, в том числе гидроксильного радикала. Редокс-состояние клетки тесно связано с железом (и иногда с медью). Эти переходные металлы могут освобождаться из белков, таких как ферритин, и из [4Fe-4S]-центров различных ферментов семейства дегидратаз-лиаз за счет реакции с супероксидом [Levi S. et al., 1996; Nordberg J., Arner E.S., 2001]. Показано, что ряд агентов – супероксид-анион-радикал [Aust S.D. et al., 1985; Biemond P. et al., 1986; Bolann B.J., Ulvik R.J., 1987; Monteiro H.P., Winterbourn C.C., 1988; Samokyszyn V.M. et al., 1988], аскорбиновая кислота [Halliwell B., Gutteridge J.M.C., 1986], полигидроксипиримидины [Monteiro H.P., Winterbourn C.C., 1989] – способны мобилизовать из молекулы ферритина ионы Fe2+, которые вовлекаются в цитотоксические процессы через реакцию Фентона [Костюк В.А., Потапович А.И., 2004]. Общепринято, что в условиях in vivo •OH образуется, главным образом, в результате катализируемой железом реакции Хабера-Вейса (2) [Фридович И., 1979; Костюк В.А., Потапович А.И., 2004; Halliwell B., Gutteridge J.M.C., 1986.; Cadenas E., элементарных 1989], которая процессов: представляет реакции Фентона трехвалентного железа супероксидом (О2-•) (1b): H2O2 + Fe2+ →•OH + -OH + Fe3+ (1а) Fe3+ + О2•- → Fe2+ + O2 (1b) ___________________________________ H2O2 + О2•- → •OH + -OH + O2 (2) собой (1а) совокупность двух и восстановления 38 • Оксид азота ( NO) синтезируется из аргинина с участием фермента NO синтазы, имеющей 3 изоформы: нейронная NO синтаза (nNOS), эндотелиальная NO синтаза (еNOS) и индуцибельная NO синтаза (iNOS) [Beal M.F., 2002; Silva J.P., Coutinho O.P., 2010]. Подобно супероксид-аниону, •NO небыстро реагирует с большинством биомолекул, несмотря на наличие неспаренного электрона. С другой стороны, он легко реагирует с другими свободными радикалами (например, пероксильным и алкильным радикалами), генерируя при этом менее реактивные молекулы [Rubbo H. et al., 2000] и функционируя, таким образом, как «защитник» от свободных радикалов [Nordberg J., Arner E.S., 2001]. Кроме того, оксид азота также участвует в передаче нервных импульсов, регуляции релаксации сосудов и в воспалительных процессах [Beal M.F., 2002; Antunes et al., 2005]. Однако он может реагировать с супероксид-анионом, производя пероксинитрит (ONOO-). Это соединение способно наподобие гидроксильных радикалов вызывать окисление липидов и белков, быстро реагировать с СО2, образуя нитрозо-пероксокарбоксилат (ONOOCO2-) и затем пероксоазотистую кислоту (ONOOH,) и разлагаться до •OH и •NO2 [Radi R. et al., 2001]. В живых системах могут образовываться и другие АФК, например, пероксильные радикалы (ROO•). Простейший пероксидный радикал НОО• образуется протонированием супероксида и обычно называется либо гидропероксидным либо пергидроксидным радикалом. Показано его участие в инициации липопероксидации (Valko M. et al., 2007). Окислительное повреждение биомолекул Благодаря своей высокой активности, АФК и АФА способны вызывать повреждение всех биомолекул (белки, липиды и нуклеиновые кислоты) и оказывать токсическое, мутагенное и канцерогенное действие [Kohen R., Nyska A., 2002; Silva J.P., Coutinho O.P., 2010]. 39 Окислительное повреждение белков. Прямое окисление боковых радикалов аминокислот в белках (особенно пролина, аргинина, лизина и треонина) под действием АФК сопровождается образованием реактивных карбонильных производных [Stadtman E.R., 2004; Dalle-Donne I. et al., 2006]. Образование этих производных в белке может изменять конформацию и/или даже вызывать фрагментацию полипептидной цепи, таким образом способствуя частичной или полной инактивации белков. Кроме того, это может привести к потере ферментативной активности, усиленной протеолитической деградации и изменению клеточных функций, таких как выработка энергии, формирование мембранных потенциалов и пр. [Fagan J., et al., 1999; Kohen R., Nyska A., 2002; Dalle-Donne I. et al., 2006;]. Повреждение белков нелетально для клетки, однако, оно может привести к развитию ряда заболеваний, включая болезнь Альцгеймера, болезнь Паркинсона, ревматоидный артрит, амиотрофный латеральный склероз, а также сопровождает процесс старения [Stadtman E.R., 2004]. Кроме того, можно привести еще два примера болезней, связанных с карбонилированием белков – это респираторный дистресс синдром и воспалительные болезни кишечника [Dalle-Donne I. et al., 2006]. Перекисное окисление липидов. Полиненасыщенные жирные кислоты (ПНЖК) благодаря наличию нескольких двойных связей чрезвычайно чувствительны радикальному окислению [Niki E. et al., 2005]. В к свободно- клеточных мембранах млекопитающих преобладают арахидоновая и линоленовая кислоты и они способны подвергаться перекисному окислению липидов (ПОЛ). Однако линоленовая кислота содержиться в большем количестве в мембранах [Spiteller G., 2001]. ПОЛ запускается радикалами (например, гидроксильными радикалами, образованными в реакции Фентона), способными удалить атом водорода от ПНЖК (LH), приводя к образованию липидных радикалов (L·), которые затем преобразуются в липоперекиси (LOOH). Терминация ПОЛ происходит за счет 40 взаимодействия двух радикалов с образованием конечных нерадикальных продуктов или вследствие разрушения субстрата [Girotti A.W., 1998; Lehuédé J. et al., 1999]. Образованные липоперекиси могут быстро деградировать до различных продуктов, например, малонового диальдегида (МДА), С3-С10 прямо- цепочечных альдегидов и α,β-ненасыщенных альдегидов [Meagher E.A., FitzGerald G.A, 2000; Lovell M.A., Markesbery W.R., 2007]. Кроме того, липопероксидация также приводит к образованию изопростанов и нейропростанов, которые являются маркерами перекисного повреждения ПНЖК в мозге и цереброспинальной жидкости [Lovell M.A., Markesbery W.R., 2007]. Для изучения интенсивности ПОЛ определяют содержание продуктов ПОЛ и ПНЖК. Наиболее широко используемый метод оценки интенсивности ПОЛ – это определение концентрации ТБК-активных продуктов (ТБК тиобарбитуровая кислота), которые отражают образование МДА [McCall M.R., Frei B., 1999; Meagher E.A., FitzGerald G.A., 2000]. Некоторые продукты ПОЛ способны оказывать цитотоксическое, мутагенное и генотоксичное действие [McCall M.R., Frei B., 1999]. Например, МДА и акролеин способны повреждать ДНК либо реагируя напрямую с азотистыми основаниями, либо формируя более активные бифункциональные производные, которые формируют экзоциклические аддукты ДНК с пятью- или шестью-членными кольцами, связанными с основаниями ДНК. Более того, реакции ПОЛ также вовлечены в развитие атеросклероза путем окисления липопротеинов низкой плотности (ЛПНП), которые затем поглощаются фагоцитами через их скэвенджер-рецепторы. Эти фагоциты постепенно накапливаются в субэндотелиальном пространстве, где они стимулируют образование атеросклеротических бляшек [Young I.S., McEneny J., 2001]. Окислительное повреждение ДНК. Повреждение ДНК происходит двумя разными путями: окислительной модификацией и фрагментацией ДНК эндонуклеазами (в процессе 41 апоптотической гибели клетки). Окислительное повреждение ДНК может повлечь за собой нарушение запуска транскрипции и сигнальных путей, ошибки репликации и геномную нестабильность. Все эти изменения могут привести к развитию рака и нейродегенеративных заболеваний, а кроме того, они связаны с нормальным процессом старения [Bjelland S., Seeberg E., 2003; Cooke M.S. et al., 2003; Silva J.P., Coutinho O.P., 2010]. Таким образом, окислительный стресс может вызвать повреждение всех биомолекул и неизбежно привести к гибели клетки и развитию некоторых патологий, таких как атеросклероз [Uemura M. et al., 2002], нейродегенеративные заболевания [Halliwell B., 2001; Jang J.H. et al., 2004], ишемическое поражение [Aikawa R. et al., 1997; He A. et al., 2007] и др. Обезвреживание АФК Инактивация АФК осуществляется антиоксидантной системой (АОС), включающей в себя биологически активные неферментативные антиокислители и антиоксидантные ферменты [Ratnam D.V. et al., 2006; Wagener F.A.D.T.G. et al., 2013]. К биологическим неферментным антиоксидантам относятся церулоплазмин, трансферрин, биофлавоноиды; витамины (токоферол, аскорбинат, ретинол, рибофлавин, никотиновая кислота); ионы металлов (селен, медь, цинк, марганец); аминокислоты (метионин, цистеин, триптофан, тирозин, фенилаланин, гистидин, пролин) и глутатион [Belozerskaya T.A., Gessler N.N., 2007; Bose K.S. et al., 2012; Koppula S. et al., 2012; Mates J.M. et al., 2012]. Основными ферментами антиокислительной системы являются супероксиддисмутаза, дисмутирующая супероксидный анион; глутатионпероксидаза, водорода, а также глутатионпероксидаза; каталаза, разрушающая инактивирующая перекиси глутатионредуктаза, система перекись липидов водорода; и перекись фосфолипид-гидропероксид- тиоредоксин-пероксиредоксина и др. [Arthur J.R., 2000; Brigelius-Flohe R. et al., 2006; Dehghan G. et al., 2013; Hanschmann E.M. et al., 2013; Zhang Q. et al., 2013]. 42 Антиоксидантные ферменты Супероксиддисмутаза (СОД, SOD). Впервые супероксиддисмутазная активность была выявлена Мак-Кордом и Фридовичем в 1969 году [McCord J.M., Fridovich I., 1969]. К настоящему моменту известно о трех изоформах СОД (КФ 1.15.1.1) [Zelko I.N. et al., 2002; Rhee S.G., 2006]. СОД1 – это главная внутриклеточная изоформа СОД (цитозольная Cu/ZnСОД). СОД1 существует как 32 кДа гомодимер и локализована главным образом в цитозоле, а также в небольшом количестве в межмембранном пространстве митохондрий [Crapo J.D. et al., 1992.; Okado-Matsumoto A., Fridovich I., 2001; Sturtz L.A. et al., 2001.]. СОД2 – это митохондриальный марганец-содержащий фермент (MnСОД), который представляет собой гомотетрамер с молекулярной массой 96 кДа и локализован в митохондриальном матриксе [Weisiger R.A., Fridovich I., 1973; Fridovich I., Freeman B., 1986]. СОД2 синтезируется в цитоплазме, а затем с помощью специального сигнального пептида доставляется в митохондрии, где фермент вовлекается в процесс дисмутации супероксида, образованного ферментами дыхательной цепи [Fukai T., Ushio-Fukai M., 2011]. СОД3 – секреторный внеклеточный Cu/Zn-содержащий фермент – это главный СОД во внеклеточной среде. СОД3 – гомотетрамер с молекулярной массой 135 кДа, имеющий два связанных дисульфидными мостиками димера. СОД3 локализуется во внеклеточном матриксе, на поверхности клеток и в небольшом количестве в плазме и внеклеточных жидкостях. На долю тканевой СОД3 приходится до 90-99% от общего количества СОД организма. [Marklund S.L., 1984; Marklund S.L., 1984.]. Тканевая СОД3 обнаруживается в кровеносных сосудах, легких, почках, матке и в меньшем количестве в сердце [Marklund S.L., 1984; Folz R.J., Crapo J.D., 1994; Stralin P. et al., 1995; Ookawara T. et al., 1998]. Механизм действия СОД. Редокс-активные металлы, такие как медь, цинк и марганец являются кофакторами СОД [Buettner G.R., 2011]. Супероксид может выступать в роли окислителя и/или восстановителя в зависимости от 43 наличия металлов. Супероксид взаимодействует с металл-содержащим активным центром СОД альтернативно как восстановитель, а затем как окислитель, образуя молекулярный кислород и перекись водорода [Zhang H.J. et al., 2002; Venkataraman et al., 2004; Venkataraman S. et al., 2005; Buettner G.R. et al., 2006; Connor K.M. et al., 2007]: O2•- + O2•- + 2H+ → Н2О2 + О2. Содержание изоферментов супероксиддисмутазы в отдельных органах и тканях экспериментальных животных существенно различается. Наиболее высокий уровень Cu/Zn-СОД выявлен в печени (до 350 мкг/г массы ткани), почках (185 мкг/г массы ткани) и поджелудочной железе (165 мкг/г массы ткани). В сердце, мозге и эритроцитах содержание этого изофермента составляет около 60-70 мкг/г массы ткани (клеток) [Костюк В.А., Потапович А.И., 2004]. Наименьший уровень супероксиддисмутазной активности выявлен в мышечной и жировой ткани [Grankvist K. et al., 1981; Mavelli I. et al., 1982]. Активность Mn-СОД снижается в ряду: сердце, почки, печень, мозг, поджелудочная железа, мышечная и жировая ткань. В эритроцитах Mn-СОД отсутствует [Grankvist K. et al., 1981]. Пероксид водорода, образующийся в процессе диспропорционирования супероксидного анион-радикала, может разрушаться каталазой (КФ 1.11.1.6), глутатионпероксидазой (КФ 1.11.1.9) и системой глутаредоксин-тиоредоксинтиоредоксинредуктазы, которые превращают перекись водорода в кислород и воду [Maritim A.C. et al., 2003; Matough F.A. et al., 2012; Takahashi M., 2012]. Каталаза. Каталаза катализирует разложение пероксида водорода с образованием молекулярного кислорода и воды: 2 H2O2 → 2 H2O + O2 Наибольшая каталазная активность выявлена в печени, крови (эритроциты) и почках. В головном мозге каталаза практически отсутствует [Grankvist K. et al., 1981; Mavelli I. et al., 1982]. Каталитическая активность этого фермента чрезвычайно высокая. Одна молекула каталазы может разложить за секунду 44000 молекул пероксида водорода [Уайт А. др., 1981]. В присутствии фермента реакция почти не требует энергии активации и ее скорость определяется скоростью диффузии субстрата к активному центру 44 фермента [Уайт А. др., 1981; Aebi H., 1984]. Однако каталаза характеризуется большой величиной Кm, и эффективное разложение пероксида водорода происходит только при его высокой концентрации [Aebi H., 1984]. Поэтому в клетке максимальная каталазная активность характерна для мест с высокой продукцией H2O2, например в пероксисомах [Delrio L.A. et al., 1992]. Защита каталазы от высоких концентраций собственного субстрата, по-видимому, обеспечивается никотинамидадениндинуклеотидфосфатом (НАДФН). Каждая субъединица тетрамерной молекулы каталазы способна связывать молекулу восстановленного НАДФН, который препятствует образованию неактивной формы фермента под действием H2O2 [Костюк В.А., Потапович А.И., 2004; Kirkman H.N. et al., 1987]. Глутатионпероксидаза (ГПО, GPX). Основную роль в детоксикации пероксида водорода в клетке играет ГПО [Lubos E. et al., 2011]. Впервые глутатион-зависимый фермент, защищающий в эритроцитах гемоглобин от повреждающего действия пероксида водорода, был выявлен Милсом в 1957 году [Mills G.C., 1957]. В настоящее время установлено, что молекула "классической" глутатионпероксидазы (ГПО1) представляет собой тетрамер, состоящий из идентичных субъединиц. Молекулярный вес всей молекулы около 85 кДа, а одной субъединицы – 21 кДа [Flohe L., 1982; Chaudiere J., Tappel A.L., 1983; Reddy C.C. et al., 1990]. Каждая субъединица фермента содержит один атом селена в форме селеноцистеина [Rotruck J. et al., 1973; Chaudiere J., Tappel A.L., 1983; Tappel A.L., 1984]. В качестве донора водорода ГПО1 использует только восстановленный глутатион, однако фермент обладает широкой субстратной специфичностью в отношении восстанавливаемого субстрата и, кроме пероксида водорода, способен катализировать двухэлектронное восстановление различных органических гидропероксидов, в том числе и гидропероксидов свободных полиненасыщенных жирных кислот [Little C., O'Brien P.J., 1968; Flohe L., 1982]: 2GSH + H2O2 → GSSG + 2H2O 2GSH + ROOH → GSSG + ROH + H2O 45 К настоящему моменту описаны по крайней мере 6 изоферментов ГПО у млекопитающих [Ufer C., Wang C.C., 2011]. ГПО 1-4 мышей и человека и ГПО6 человека содержат в активном центре селеноцистеин и потому относятся к группе селенопротеинов. Биосинтез селенопротеинов чрезвычайно зависит от наличия селеноцистеин-специфичной тРНК. Цитозольный фермент (ГПО1) вырабатывается всеми клетками млекопитающих и является одним из наиболее распространенных членов семейства ГПО. Это семейство также включает эпителий-специфичный фермент, который высоко экспрессируется в кишечнике (ГПО2). Существует интересный синергизм между ГПО1 и ГПО2, т.к. они очень похожи с ферментативной точки зрения, но различаются по экспрессии генов. ГПО2 в клетках пищеварительного тракта обеспечивает необходимую защиту от патогенов и воспаления кишечника [Chu F.F. et al., 2004; Esworthy R.S. et al., 2001, 2005.]. Кроме того, ГПО2 может также вырабатываться в других тканях в ответ на стрессорное воздействие, например, в легких в ответ на сигаретный дым и в клетках рака груди. Таким образом, ГПО2 осуществляет выдающуюся функцию в антиоксидантной и клеточной защите в некоторых тканях в дополнение ГПО1. Внеклеточная ГПО3 – это секреторный гликопротеин, который является вторым наиболее важным селенопротеином плазмы крови после селенопротеина Р. Фосфолипид-гидропероксид-ГПО (ГПО4) обнаружена в клетках всех типов и может специфично восстанавливать гидроперекиси фосфолипидов в клеточных мембранах и включаться в регуляцию апоптоза и созревание спермы [Гоголева И.В., Громова О.А., 2009]. ГПО4, главным образом, предотвращает липопероксидацию, препятствуя действию липоксигеназ и циклооксигеназы. Благодаря уникальным ферментативным свойствам активность ГПО4 может определяться с использованием фосфолипидных гидропероксидных субстратов [Weitzel F. et al., 1990.]. Важно заметить, что ГПО4 имеет стабильно высокий 46 уровень экспрессии даже при недостатке селена, что означает строгую зависимость организма от этого селенопротеина. Снижение активности ГПО4 в клеточной культуре вызывает гибель клетки по апоптотическому механизму [Imai H., Nakagawa Y., 2003.]. Активность ГПО в различных тканях организма уменьшается в ряду: печень > эритроциты > почки > желудок > сердце = легкие = мозг > плазма > мышцы [Tappel A.L., 1974; Grankvist K. et al., 1981; Zhang L. et al., 1989]. Около 75 % глутатионпероксидазной активности определяется в цитоплазме клеток, остальное количество фермента локализовано в митохондриях [Tappel A.L., 1974; Stults F.H. et al., 1977; Mbemba F. et al., 1985]. Показано, что существуют возрастные и суточные изменения активности ГПО в различных тканях экспериментальных животных [Zhang L. et al., 1989; Lapenna D. et al., 1992]. В период снижения активности ГПО в сердце отмечена активация процессов перекисного окисления липидов [Lapenna D. et al., 1992]. Уровень ГПО в тканях чрезвычайно чувствителен к алиментарному поступлению селена в организм. Содержание животных в течение нескольких недель на Se-дефицитной диете приводит к резкому снижению глутатионпероксидазной активности [Tappel A.L., 1974; Takahashi K. et al., 1986; Reddy C.C. et al., 1990] и пропорциональному уменьшению тканевого уровня соответствующего иммунореактивного белка [Tappel A.L., 1974; Takahashi K. et al., 1986]. Степень снижения активности фермента неодинакова в различных тканях, которые по чувствительности к дефициту располагаются следующим образом: селена плазма >печень >почки >сердце = легкие > эритроциты [Tappel A.L., 1974]. При однократном внутрижелудочном введении селена в форме селенита или селенометионина животным, содержавшимся на Se-дефицитной диете, нормальный уровень активности фермента в плазме, почках, желудке и печени восстанавливался в течение 48 часов [Костюк В.А., Потапович А.И., 2004; Tappel A.L., 1974; Takahashi K. et al., 1986]. 47 Система глутаредоксин – тиоредоксин - тиоредоксинредуктаза. Глутаредоксины (ГР, GRX). Глутаредоксины (КФ 1.20.4.1) – GSH-зависимые оксидоредуктазы с низкой молекулярной массой (9-14 кДa), относящиеся по своей структуре к суперсемейству тиоредоксинов и играющие важную роль в клеточных редокс-зависимых процессах [Fernandes A.P., Holmgren A., 2004]. В ГР-зависимой системе перенос электронов происходит от НАДФНзависимой глутатионредуктазы на окисленный глутатион с образованием GSH (1), который, в свою очередь, восстанавливает окисленный глутаредоксин (2): GSSG + НАДФН → 2 GSH + НАДФ+ (1) глутатионредуктаза 2 GSH + ГР-S2 → GSSG + ГР-(SH)2 (2) Глутаредоксины функционально сопряжены с глутатионредуктазой и с соотношением GSH/GSSG. Соотношение GSH/GSSG является индикатором клеточного редокс-статуса и важной детерминантой редокс-потенциала, который коррелирует с биологическим статусом клетки [Schafer F.Q., Buettner G.R., 2001]. У млекопитающих и человека обнаружены три изоформы ГР (GRX): цитозольная ГР1 и две митохондриальные – ГР2 и ГР5 [Lundberg M. et al., 2001; Wingert R.A. et al., 2005]. ГР1 – белок с молекулярной массой 12 kДa, локализован в основном в цитозоле (хотя обнаружен и в ядре) [Lysell J. et al., 2003], был открыт как GSHзависимый донор для рибонуклеозидредуктазы [Калинина Е.В. и др., 2008]. Изоформа ГР2 (14 кДa) обнаружена в двух сплайсинговых формах – ГР2α и ГР2β, локализованных, соответственно, в митохондриях и в ядре [Lundberg M. et al., 2001]. Катализируя обратимое S-глутатионилирование митохондриального сопровождается комплекса образованием I, функциональная O2•–, ГР2 активность регулирует уровень которого АФК в митохондриях [Taylor E.R. et al., 2003]. Кроме того, ГР2 катализирует обратимое глутатионилирование белков внутренней мембраны митохондрий 48 в зависимости от изменения соотношения GSH/GSSG [Калинина Е.В. и др., 2008]. Установлено, что ГР2 может подавлять развитие апоптоза посредством предотвращения выхода цитохрома с из митохондрий и окисления кардиолипина [Lillig C.H. et al., 2004; Enoksson M. et al., 2005]. Изоформа ГР5 человека имеет высокую степень гомологии с ГР5 дрожжей, в связи с чем эти белки получили одинаковое название [Wingert R.A. et al., 2005]. ГР5 человека содержит N-концевой участок аминокислотной последовательности, обеспечивающий его транслокацию в митохондрии. ГР2 и ГР5, локализованные в митохондриях кардиомиоцитов, могут вносить определенный вклад в развитие и течение сердечно-сосудистых заболеваний, поскольку участвуют в поддерживании гомеостаза железа [Lillig C.H. et al., 2004; Wingert R.A. et al., 2005; Molina-Navarro M.M. et al., 2006]. Следует отметить, что глутаредоксин играет важную роль в процессах формирования лекарственной резистентности опухолевых клеток к препаратам с прооксидантным эффектом, усиливая систему антиоксидантной защиты [Kalinina E.V. et al., 2001; Chen Y. et al., 2002]. Подобной активностью обладает также и тиоредоксин. Оба фермента функционально взаимосвязаны и дополняют друг друга, участвуя в контроле клеточного редокс-баланса. Установлено существование перекреста путей регуляции (cross-talk) между этими двумя системами в клетках млекопитающих [Casagrande S. et al., 2002]. Тиоредоксин (TP, TRX). Тиоредоксин (КФ 1.8.4.8) – полифункциональный низкомолекулярный белок, имеющий в своей структуре активный дитиол/дисульфидный участок и обладающий оксидоредуктазной активностью [Powis G., Montfort W.R., 2001]. Основными изоформами тиоредоксина (ТР) являются цитозольный ТР1 и митохондриальный ТР2. Обе изоформы тиоредоксина вносят заметный вклад в антиоксидантную защиту клеток, что определяется не только их способностью восстанавливать каталитическую активность пероксиредоксинов и глутатионпероксидаз, разлагающих гидроперекиси и H2O2 [Rhee S.G. et al., 2001], но и способностью 49 ТР1 непосредственно восстанавливать H2O2 и GSSG [Nordberg J., Arner E.S.J., 2001]. Кроме того, обе изоформы могут играть роль «ловушки» •OH радикалов [Das K.C., Das C.K., 2000]. Известно, что ТР1 служит кофактором ряда ферментов [Arner E.S.J., Holmgren A., 2000], таких, например, как пероксиредоксины [Rhee S.G. et al., 2001], рибонуклеотидредуктазы и метионинсульфоксидредуктазы. Кроме того, ТР1 участвует в репарации ДНК [Powis G., Montfort W.R., 2001]. ТР в окисленной форме является субстратом для тиоредоксинредуктазы. Тиоредоксинредуктаза (ТРР, TRXR). ТРР (КФ1.8.1.9, систематическое название «тиоредоксин:НАДФH-оксидоредуктаза») является НАДФН- зависимым гомодимером оксидоредуктазы (с одним ФАД на субъединицу), которая восстанавливает активный центр дисульфида в окисленном ТР [Калинина Е. В. и др., 2008; Arner E.S.J., Holmgren A., 2000]. Выявлены и охарактеризованы три изоформы ТРР млекопитающих: цитоплазматическая (ТРР1), митохондриальная (ТРР2) и ТРР, катализирующая восстановление как ТР, тиоредоксинглутатионредуктазой так и (ТРГР), GSSG высокое и называемая содержание которой обнаружено в семенниках [Miranda-Vizuete A. et al., 1999]. Все три изоформы обладают одинаковой структурой домена и характеризуются наличием селеноцистеина в С-концевом активном центре (Gly–Cys–Sec–Gly–COOH). Механизм катализа ТРР млекопитающих включает процесс восстановления ТР с участием НАДФН, сходный с аналогичным процессом для глутатионредуктазы и липоамиддегидрогеназы [Калинина Е. В. и др., 2008]. Вначале осуществляется перенос электрона с НАДФН через ФAД на дисульфид в активном центре, образованном остатками цистеина в положении 59 и 64 в Nконцевом домене, который идентичен аналогичному домену в глутатионредуктазе. Затем электроны переносятся от образовавшегося дитиола одной субъединицы в димерном ферменте на селенилсульфид в С-концевой последовательности другой субъединицы. Такой перенос электронов на другую субъединицу может иметь место и при восстановлении GSSG 50 глутатионредуктазой [Zhong L. et al., 2000]. Восстановленный С-концевой селеноцистеин участвует в восстановлении многочисленных субстратов ТРР1 у млекопитающих. Перекиси, включая непосредственно гидроперекиси липидов и H2O2, могут восстанавливаться ТРР1 [Калинина Е. В. и др., 2008]. Посредством этого механизма ТРР1 функционирует как альтернативный путь ферментативной детоксикации гидроперекисей липидов. Однако, высокое значение Km ТРР1 для H2O2 (2,5 мM) делает значимой роль фермента только при повышении уровня H2O2 [Калинина Е.В. и др., 2008]. ТРР1 восстанавливает также различные низкомолекулярные соединения, включая антибактериальные полипептиды, цистин, аллоксан и витамин К [Калинина Е.В. и др., 2008]. ТРР1 восстанавливает многие Se-содержащие соединения, в том числе селенит [Kumar S. et al., 1992] и Se-содержащий активный центр глутатионпероксидазы в плазме [Bjornstedt M. et al., 1994]. Для активной инкорпорации Se в Se-содержащие белки необходим переход неорганического Se, например, в форме селенита (SeO32–), в селенид (HSe−). Это восстановление может катализироваться ТРР1 с участием или без ТР1 [Kumar S. et al., 1992], либо включает образование селенодиглутатиона (GS-Se-SG) как интермедиата. Образование GS-Se-SG требует присутствия GSH, уровень которого поддерживается глутатионредуктазой, которая может и непосредственно восстанавливать GS-Se-SG. Однако ТРР1 также высоко эффективна в восстановлении GS-Se-SG. Таким образом, ТРР1 играет существенную роль в метаболизме Se-содержащих соединений и является важным звеном, связывающим метаболизм Se и антиоксидантные процессы [Калинина Е.В. и др, 2008]. Помимо этого, следует подчеркнуть важность Se для ферментативной активности ТРР [Zhong L. et al., 2000]. Предполагается, что наличие Se снижает потенциал восстановления остатков Cys, повышает эффективность работы ферментов при низких значениях рН и расширяет спектр восстанавливаемых 51 соединений [Меньшикова Е.Б. и др., 2006]. Отмечается, что активность ТРР снижается при дефиците Se у животных и в клеточных линиях человека. В то же время, добавление Se к культурам опухолевых клеток человека в несколько раз повышает активность ТРР в отличие от других Se-содержащих ферментов, включая глутатионпероксидазу [Калинина Е.В. и др, 2008]. Таким образом, функционирование антиоксидантной системы млекопитающих и человека напрямую зависит от активности селензависимых ферментов глутатионпероксидазы и тиоредоксинредуктазы. Опыты на млекопитающих, содержавшихся на селенодефицитном рационе, показали, что активность этих ферментов строго зависит от адекватного поступления селена в организм животных [Костюк В.А., Потапович А.И., 2004; Tappel A.L., 1974; Takahashi K. et al., 1986]. Антиоксидантная активность органических соединений селена, серы и теллура Учитывая, что селенодефицитными, большинство встает вопрос регионов об России эффективной являются профилактике селенодефицита и связанных с ним заболеваний. В этой связи на протяжении последних двух десятилетий разрабатываются методы химического синтеза различных по структуре селеноорганических соединений, которые служат не только формой введения микроэлемента в организм, но также проявляют широкий спектр биологический активности и в первую очередь антиоксидантную активность. Основными группами синтетических селеноорганических соединений являются селенопираны [Галочкин В.А. и др., 1998], селеноксантены и селенсодержащие арилалифатические дикетоны. Все указанные препараты содержат селен в двухвалентной форме, что способствует включению его в метаболизм организма [Levander O., Burk R.F., 1998]. 52 Особого внимания заслуживают селенсодержащие арилалифатические дикетоны. Простейшим представителем этого ряда соединений является 1,5дифенил-3-селенапентандион-1,5 (ДАФС-25): Синтез препарата произведен в НИИ химии Саратовского государственного университета [Древко Б.И. и др., 1996]. После биологических испытаний 1,5-дифенил-3-селенапентандион-1,5 был внедрен в ветеринарную практику и в настоящее время производится в промышленных масштабах. Содержание селена в препарате составляет 25%. Торговое название препарата диацетофенонилселенид (ДАФС-25). Препарат относится к 3 классу токсичности (умеренно токсичные вещества) по классификации Hodge @ Sterner. Среднесмертельная доза ДАФС (LD50), установленная при введении препарата белым крысам, составила 385 мг/кг живого веса. В сравнении с селенитом натрия, для которого интервал среднесмертельных доз для разных видов животных составляет от 0,59 до 18 мг/кг живой массы (10 мг/кг для белых крыс), ДАФС-25 вещество с менее выраженными токсическими свойствами. Коэффициент кумуляции - 5,24. По классификации кумулятивного действия препарат можно отнести к веществам со слабо выраженными кумулятивными свойствами, т.е. к 5 классу. Препарат ДАФС-25 прошел успешные испытания в ветеринарии и в настоящее время применяется для профилактики беломышечной болезни, токсической дистрофии печени, экссудативного диатеза [Древко Б.И. и др., 1996, 2001]. Антиокислительная активность селенсодержащих арилалифатических дикетонов подтверждена в исследования in vitro и in vivo [Кирова Ю.И., 2004]. Полученные данные антиоксидантную показывают, активность in что vitro, препарат что ДАФС-25 выражалось в проявлял снижении 53 концентрации первичных (диеновые конъюгаты) и вторичных (малоновый диальдегид) продуктов свободно-радикального окисления липидов. Антиокислительный эффект препарата ДАФС-25 был установлен в гомогенатах сердца, поджелудочной железы, легких и почек. Указанные органы характеризуются низкой оксидорезистентностью. Предполагаемый механизм действия можно представить непосредственным взаимодействием этого препарата с активными кислородными и липидными метаболитами (О 3, О2-., Н2О2, ROOH) и образованием соответствующего моноселенооксида. Серосодержащий аналог ДАФС-25 (1,5-дифенил-3-тиапентандион-1,5) in vitro проявлял прооксидантную активность в гомогенатах сердца, мозга и в плазме крови: Восстановительная способность атома серы выражена в меньшей степени, чем у селена, что обусловлено строением электронных оболочек атома и меньшим атомным радиусом. По данным проведенных исследований in vivo [Кирова Ю.И., 2004] препарат ДАФС и его серный аналог проявляли выраженную антиоксидантную активность в гомогенатах печени и эритроцитах, то есть в клетках с высокой оксидорезистентностью. Однако прооксидантную серосодержащий активность в аналог ДАФС-25 гомогенатах in легких, vivo сердца, проявлял мозга, поджелудочной железы и в плазме крови, то есть в тканях с низкой оксидорезистентностью. Приведенные выше результаты показывают, что селеноорганическое соединение ДАФС-25 обладало большей антиоксидантной активностью in vivo и in vitro, чем его серосодержащий аналог [Кирова Ю.И., 2004]. 54 Структура халькогенсодержащих арилалифатических дикетонов позволяет рассматривать их как потенциальные антиоксиданты, которые возможно способны взаимодействовать с гидроперекисями липидов и органическими радикалами. Однако нельзя исключить и опосредованное оксидопротекторное действие селенсодержащих арилалифатических дикетонов in vivo через аппарат селензависимых антиоксидантных ферментов. Как известно, основу антиоксидантной системы (АОС) клетки составляют высокоэффективные ферменты-антиоксиданты, содержащие в структуре активного центра селеноцистеин, принимающий непосредственное участие в ферментативном обезвреживании активированных кислородных метаболитов [Кирова Ю.И., 2004]. Как было описано выше, основными ферментами антиоксидантной защиты являются глутатионпероксидазы (ГПО 1–6). Функцией глутатионпероксидаз является поддержание стабильной внутриклеточной концентрации восстановленного глутатиона. Наиболее изученным является цитозольная глутатион-пероксидаза (ГПО1). Доказано, что ГПО1 играет основную протективную роль при развитии окислительного стресса. В эксперименте показана линейная зависимость между активностью ГПО1 и выживаемостью мышей в условиях выраженного окислительного стресса. Кроме того, активность ГПО1 более зависима от содержания селена по сравнению с другими ферментами, а потому активность ГПО1 в эритроцитах является простым и чувствительным показателем селенового статуса организма [Гоголева И.В., Громова О.А., 2009]. Анализ зарубежной литературы [Rossato J.I. et al., 2002] показал, что не только селеноорганическое соединение бис-[S-4-изопропил-2-фенил оксазолин]диселенид, но и его серопроизводные обладают потенциальными антиоксидантными свойствами, тогда как теллурорганический аналог не проявляет антиоксидантную активность. По другим данным селенорганический препарат эбселен (2-фенил-1,2бензисоселеназол-3-(2Н)-он) и его теллурорганический аналог обладают ГПО- 55 подобной активностью благодаря своей способности реагировать с пероксинитритом [Masumoto H. et al., 1996; Klotz L.-O. et al., 2003]. Эбселен Образующиеся в результате этого взаимодействия селеноксид или теллуроксид могут восстанавливаться тиолами (но не GSH), например, Znметаллотионеином [Jacob C. et al., 2000]. Помимо этого окисленная форма эбселена (селеноксид) может также восстанавливаться тиоредоксинредуктазой млекопитающих при участии НАДФН [Arteel G.E. et al., 1999; Klotz L.-O. et al., 2003]. Однако окислительно-восстановительные халькогенорганических антиоксидантное или соединений и прооксидантное свойства их способность действие напрямую оказывать зависят от особенностей их строения. Например, к настоящему времени известно о группе теллуорганических соединений, проявляющих прооксидантные свойства и потому обладающих мощной противораковой [Doering M. et al., 2010; Jamier V. et al., 2010; Coriat R. et al., 2011; Marut W.K. et al., 2012;] и антибактериальной активностью [Borsetti F. et al., 2007; Calderon I.L. et al., 2009; Sandoval J.M. et al., 2011]. В заключение халькогенорганических следует отметить, соединений что зависят антиоксидантные от их строения, свойства физико- химических свойств и, главное, от наличия атома халькогена (серы, селена или теллура). При этом селеноорганические соединения обладают намного большей антиоксидантной активностью именно благодаря наличию атома селена [Кирова Ю.И., 2004; Inglot A.D. et al., 1996]. Можно предположить, что селеноорганические соединения являются донором селена, необходимого для синтеза селенопротеинов, в том числе главных антиоксидантных ферментов глутатионпероксидазы и тиоредоксинредуктазы. Эти селен-зависимые 56 ферменты участвуют в разложении гидроперекисей липидов и перекиси водорода, подавляют процессы ПОЛ, что и обусловливает их антиоксидантные свойства в живых организмах. В этой связи адекватное поступление селена с пищей или в составе биодобавок необходимо для нормального функционирования антиоксидантной системы организма. При этом органические формы селена предпочтительнее, поскольку они менее токсичны и более биодоступны по сравнению с неорганическими соединениями селена, такими как селенит и селенат натрия [Гмошинский И.В., Мазо В.К., 1999]. В настоящее время одним из перспективных селеноорганических соединений является препарат ДАФС-25, который применяется в ветеринарии и оказывает антиоксидантное, антитоксическое, гепатопротекторное и другие действия [Древко Б.И. и др., 1996, 2001]. Более углубленное изучение биологической активности этого препарата возможно позволит рекомендовать его в качестве биологически активной добавки. 1.3. Антитоксическая активность халькогенорганических соединений при интоксикации солями кадмия, свинца и ртути Токсические эффекты тяжелых металлов Загрязнение окружающей среды тяжёлыми металлами, постоянная интоксикация человека в условиях неблагоприятной экологической обстановки делает актуальным вопрос о поиске эффективных антитоксикантов при отравлении солями тяжёлых металлов как животных, так и человека [Satarug S., et al., 2010]. Тяжелые металлы рассматриваются Всемирной Организацией Здравоохранения как наиболее опасные для здоровья людей токсиканты [Satarug S., 2012; Nair A.R. et al, 2013]. Токсичность ионов непереходных металлов Pb2+, Hg2+, СН3Нg+ и Cd2+ обусловлена образованием комплексов с аминокислотами и другими биомолекулами, содержащими концевые тиогруппы. Другой важный механизм токсического действия заключается в 57 вытеснении эссенциальных металлов из металлсодержащих комплексов, приводящем к потере последними биологической активности [Исидоров В.А., 1999]. Еще одним аспектом токсического действия тяжелых металлов является их способность активировать процессы перекисного окисления липидов (ПОЛ). Ведущую роль в обезвреживании продуктов ПОЛ играет антиоксидантная система защиты организма. Главными антиоксидантами и антитоксикантами являются селено-зависимые ферменты и селенопротеины. В условиях селенодефицита и неблагоприятной экологической обстановки встает вопрос о необходимости применения селенсодержащих препаратов, оказывающих гепатопротекторное, нефропротекторное и антитоксическое действие при отравлении различными ксенобиотиками [Саноцкий И.В., 2010; Кильметова И.Р. и др., 2012], в том числе тяжелыми металлами. В настоящее время наиболее распространенными токсикантами окружающей среды являются соли ртути, кадмия и свинца [Agency for Toxic Substances and Disease Registry, 1999, 2007, 2008; Fowler B. A., 2009; Matovic V. et al, 2011; Sakamoto M. et al., 2011; Bernhot R.A., 2012; Ciesielski T. et al., 2012; Flora G. et al., 2012.; National Toxicology Program, 2012], которые при поступлении в организм млекопитающих стимулируют окислительный стресс и конкурируют с эссенциальными металлами (цинк, марганец, медь, железо, кальций и др.), что вызывает нарушение функций многих белков и ферментов. Поэтому интоксикация тяжелыми металлами в конечном итоге способствует развитию сердечно-сосудистых заболеваний, неврологических нарушений, поражений печени и почек, а также вызывает канцерогенез [Sears M.E., 2013]. Информация, касающаяся молекулярных механизмов токсичности тяжелых металлов, довольно фрагментарная, тем не менее, есть данные, демонстрирующие, что отравление тяжелыми металлами окислительный стресс на клеточном уровне [Cuypers A. et al., 2010]. вызывает 58 Роль тяжелых металлов в развитии окислительного стресса Тяжелые металлы при поступлении в организм вызывают ряд клеточных ответов, в которых ключевую роль играет редокс-статус [Luparello C. et al., 2011]. Первый возможный механизм развития окислительного стресса при интоксикации тяжелыми металлами связан с образованием свободных радикалов. Поскольку тяжелые металлы сами не участвует в реакции Фентона, они не способны напрямую вырабатывать АФК. Однако они запускают окислительный стресс опосредованно за счет вытеснения из белков и ферментов редокс-активных металлов (железа и меди) [Thevenod F., 2009; Cuypers A. et al., 2010;]. Действительно, тяжелые металлы вытесняют железо из белков, которое затем участвует в реакции Фентона, приводя к образованию высокоактивного гидроксильного радикала (Cuypers A. et al., 2010). Кроме этого, тяжелые металлы, в частности, кадмий, в связанной или несвязанной формах, участвуют в повреждении митохондрий [Cannino G. et al., 2009; Thevenod F., 2009]. Этому может способствовать ингибирование кадмием комплекса III митохондриальной ЦПЭ, в результате чего образуются нестабильные семихиноны, которые передают электрон на молекулярный кислород с образованием супероксид-аниона [Wang Y., et al., 2004], а из него и других свободных радикалов [Stinson L.J., et al., 2003]. Активные формы кислорода и азота запускают свободно-радикальное окисление, сопровождающееся повреждением макромолекул (белков, липидов и ДНК) и всех клеточных структур, что в конечном итоге приводит к гибели клетки по апоптотическому пути [Wang L., et al., 2009]. Ключевым моментом запуска апоптоза является выход цитохрома с из митохондриальной мембраны [Lillig et al., 2004; Enoksson et al., 2005]. Причинами этого являются: 1) металл-индуцированное окислительное повреждение клеточных структур, в том числе комплексов I, III и IV митохондрий [Mastrocola R., et al., 2005]; 59 2) деполяризация митохондриальной мембраны [Yang L.Y., et al., 2009; Gobe G.and Crane D., 2010] вследствие выхода ионов кальция из ЭПР под действием тяжелых металлов, в частности кадмия. Есть данные подтверждающие, что тяжелые металлы могут транспортироваться через кальциевые каналы, блокируя их [Marchetti C., 2013]. Второй механизм прооксидантного действия тяжелых металлов связан с подавлением антиоксидантной защиты организма. Например, в печеночных клетках крыс тяжелые металлы, в том числе кадмий, ингибируют цитозольную Cu/Zn-СОД [Ikediobi C.O. et al., 2004], ГПО и ГР [Waisberg M. et al., 2003; Ikediobi C.O. et al., 2004; Lopez E., et al., 2006]. Механизм ингибирования Cu/ZnСОД основан, скорее всего, на вытеснении эссенциальных ионов меди и цинка из активного центра фермента тяжелыми металлами [Ikediobi C.O. et al., 2004], а ингибирование ГПО и ГР - на связывании тяжелых металлов с восстановленным глутатионом (GSH) в активном центре этих ферментов [Cuypers A. et al., 2010]. Кроме доставляются того, в при печень, энтеральном где поступлении взаимодействуют с тяжелые металлы металлотионеинами, глутатионом и другими белками или пептидами [Thevenod F., 2009]. Металлотионеины - это группа низкомолекулярных белков (6-8 кДа), в которых на долю остатков цистеина приходится до 30% [Hamer D.H., 1986]. Металлотионеины синтезируются в ответ на поступление в организм тяжелых металлов и могут оказывать двоякое действие [Andrews G.K., 2000]. С одной стороны, металлотионеины связывают тяжелые металлы, обезвреживая их и удаляя из клеточного пространства. С другой стороны, металлотионенины используют свои сульфгидрильные группы для обезвреживания активных форм кислорода (АФК), которые вырабатываются в ответ на окислительный стресс, вызванный тяжелыми металлами [Hart B.A. et al., 2001; Cuypers A. et al., 2010]. Причем аффинность тяжелых металлов к металлотионеинам снижается в направлении: Hg2+>Cu+>Cd2+>Zn2+ [Varotto L. et al., 2013]. 60 Таким образом, тяжелые металлы конкурируют с АФК за связывание с сульфгидрильными группами металлотионеинов, восстановленного глутатиона и других тиол-богатых молекул. После связывания тяжелых металлов с сульфгидрильными группами восстановленного глутатиона [Cuypers A., et al., 2010], он утрачивает свои антиоксидантные свойства и это неизбежно ведет к снижению активности главного антиоксидантного фермента ГПО. Следовательно, интоксикация тяжелыми металлами приводит, с одной стороны, к усилению выработки АФК, запуску свободно-радикального окисления, а с другой стороны, к подавлению активности антиоксидантной защиты организма, следствием чего является нарушение редокс-баланса в клетках, окислительное повреждение клеточных структур и развитие патологических процессов в различных органах и тканях (печени, почках, сердечно-сосудистой и нервной системах, поджелудочной железе и пр.). Роль тяжелых металлов в развитии патологических процессов Развитие окислительного стресса обусловливает металл-индуцированные генотоксичность и мутагенность. Тяжелые металлы (кадмий, ртуть, свинец) стимулируют выработку АФК (гидроксильного, пероксидного и супероксидного радикалов) и АФА (оксида азота, пероксинитрита и Sнитрозотиолов) в системах in vivo и in vitro. Генерация свободных радикалов сопровождается значительным повреждением белков, липидов и ДНК [Koedrith P., Seo Y.R., 2011], что может стать причиной гибели клетки по пути апоптоза или при нарушении апоптотического механизма привести к канцерогенезу. Главными мишенями для действия как АФК, так и ионов тяжелых металлов являются сульфгидрильные группы, связывание или окисление которых приводит к инактивации белков и ферментов. На рисунке 3 показано участие тяжелых металлов в развитии апоптоза и канцерогенеза. 61 61 Рис. 3. Роль тяжелых металлов в развитии апоптоза и канцерогенеза Примечание: ↑СРО - повышение свободно-радикального окисления; ↓АОЗ - снижение антиоксидантной защиты; ↓САТФ - снижение концентрации АТФ 62 Одним из возможных механизмов развития канцерогенеза при участии тяжелых металлов является их действие на цинк-фингерные белки, в которых ионы тяжелых металлов конкурируют с ионами цинка за связывание с сульфгидрильными группами. Тяжелые металлы могут вытеснять ионы цинка и, встраиваясь на их место, менять конформацию и активность этих белков [Sunderman F.W.Jr., Barber A.M., 1988; Hartwig A., 2001; Beyersmann D., Hartwig A., 2008]. В цинк-фингерных белках часть молекулы, отвечающая за взаимодействие с ДНК, стабилизируется связыванием иона Zn2+ с двумя имидазольными азотами и двумя сульфгидрильными группами. В отсутствии цинка конформация «цинковых пальцев» изменяется, и они утачивают способность связываться с ДНК. Экспериментальные данные демонстрируют, что ионы кадмия и свинца не только могут конкурировать с цинком за присоединение к активному центру «цинковых пальцев», но даже имеют преимущества перед цинком благодаря большей величине константы связывания. Такая замена ионов приводит к потере способности «цинковых пальцев» контактировать с ДНК, а, значит, регулировать экспрессию генов [Petering D.H. et al., 2000; Gong P. et al., 2000; WitkiewiczKucharczyk A., Bal W., 2005]. Кроме того, сама ДНК очень чувствительна к повреждающему действию как внешних (УФ-излучение, химические и биологические токсины), так и внутренних факторов, главным образом окислительному повреждению. При этом некоторые канцерогенные металлы (As(III)/As(V), Cd(II), Cr(VI), Ni(II), Hg(II) и Pb(II)) в низких концентрациях способны подавлять репарацию поврежденной ДНК (Beyersmann D., Hartwig A., 2008). Повреждение ДНК тяжелыми металлами может стать причиной развития некоторых патологий, включая злокачественные новообразования. Другим возможным механизмом токсического действия тяжелых металлов является подавление роста клетки путем изменения экспрессии факторов роста. 63 Например, некоторые металлы способны запускать МАР(mitogen-activated protein)-киназные пути, включающие активацию ядерных транскрипционных факторов (AP-1, NF-κB, p53, NFAT и HIF-1), которые в свою очередь управляют экспрессией цитопротективных генов, необходимых для репарации ДНК, иммунного ответа, остановки клеточного цикла и запуска апоптоза. Тяжелые металлы и АФК, вероятно, взаимодействуют с тиольными группами фосфатаз разных типов (в частности серин/треонин-фосфатаз и фосфолипидфосфатаз), которые при окислении образуют дисульфидные мостики [Thannickal V.J., Fanburg B.L., 2000; Poli G. et al., 2004]. Модифицированные таким образом ферменты утрачивают свою способность включаться в различные регуляторные каскады и активировать траксрипционные факторы, названные выше. Некоторые тяжелые металлы ингибируют экспрессию опухолевых супрессоров р53 и р16. Поэтому тяжелые металлы могут увеличивать пролиферацию обеспечивать клетки адаптацию путем подавления клетки к апоптотических интоксикации процессов тяжелыми и металлами, способствуя таким образом канцерогенезу [Koedrith P., Seo Y.R., 2011]. Кроме того, тяжелые металлы влияют на экспрессию генов, участвующих в стрессорном ответе, например, белков теплового шока [Goering P.L., et al., 2000; Young J. C. et al., 2004) и регуляции клеточного цикла (Jeong E. M. et al., 2004]. Белки теплового шока (Hsp) обычно синтезируются в ответ на какое-либо стрессвоздействие (повышение температуры, интоксикацию и пр.), запускают реакции иммунной защиты и активируют многие сигнальные белки (рецепторы стероидных гормонов, киназы клеточного цикла и пр.) [Wang Z. et al., 2009; Fu D. et al., 2011]. Однако в работах разных авторов есть сведения как о подавлении экспрессии мРНК HSP27, HSP40, HSP60, HSP70 и HSP90 при интоксикация тяжелыми металлами [Cannino G. et al., 2009; Chen X. et al., 2012], так и об усилении экспрессии этих шаперонов [Ivanina A.V. et al., 2008]. 64 В любом случае при интоксикации тяжелыми металлами функции шаперонов будут нарушены, поскольку для их активности необходимы значительные затраты АТФ [Hartl F.-U. et al., 1994]. Вместе с тем ранее отмечалось, что деполяризации токсическое действие митохондриальной металлов мембраны, выражается ингибировании также в комплексов митохондриальной ЦПЭ и нарушении окислительного фосфорилирования, что в конечном итоге приводит к выходу цитохрома с из митохондрий и запуску апоптоза. Разные формы клеточной гибели, связанной с токсичностью тяжелых металлов, являются дозо-зависимыми и включают некроз, апоптоз-подобную гибель клетки, а также аутофагию, показанную на различных типах клеток [Templeton D.M., Liu Y., 2010]. Субмикромолярные концентрации тяжелых металлов, в частности кадмия, приводят к развитию или задержке апоптоза; средние концентрации около 10 мкМ вызывают различные виды апоптотической гибели клетки и очень высокие концентрации (более 50 мкМ) приводят к некрозу [Templeton D.M., Liu Y., 2010]. Таким образом, поступление и биоаккумуляция тяжелых металлов в клетках млекопитающих считаются потенциальными факторами риска в патогенезе рака [Verougstraetea V. et al., 2003; Fasinu P.S., Orisakwe O.E., 2013; Nair A.R. et al., 2013], поскольку повреждение ДНК, нарушение ее репарации и задержка апоптоза являются главными причинами канцерогенеза [Waisberg M. et al., 2003; Schwerdtle T. et al., 2010; Templeton D.M., Liu Y., 2010; Nemmiche S. et al., 2011] в различных органах, в первую очередь, в печени и почках [Mandel J.S. et al., 1995; Waalkes M.P., Misra R.R., 1996; Thijssen S. et al., 2007; Shimada H. et al., 2008; Fowler B.A., 2009; Reyes J.L. et al., 2013]. Тем не менее, в подавляющем большинстве случаев происходит отравление малыми или средними дозами тяжелых металлов. При поступлении в организм через пищеварительный тракт (с загрязненной пищей и/или водой), через кожу 65 или с вдыхаемым воздухом тяжелые металлы доставляются в печень и почки, поэтому именно эти органы являются основными мишенями при интоксикации тяжелыми металлами [Abdel-Monem N. et al., 2013; Reyes J.L. et al., 2013]. Токсическое повреждение почек тяжелыми металлами выражается в развитии глюкозурии, аминоацидурии и фосфатурии вследствие повреждения гломерулярных клеток [Hu H., 1991; Nolan C.V., Shaikh Z.A., 1992], а хроническая интоксикация приводит к артериальной гипертензии и почечной дисфункции [Clarkson T.W., 1997; Wang L. et al., 2009; Reyes J.L. et al., 2013]. Важными биохимическим маркерами нефротоксичности являются азотсодержащие метаболиты креатинин и мочевина, экскреция которых нарушается при повреждении почек [Брин В.Б. и др., 2011; 2012], а также альбумины, концентрация которых в плазме крови снижается вследствие их выведения с мочой [Брин В.Б. и др., 2011; 2012]. Печень также является мишенью при отравлении тяжелыми металлами, поскольку она выполняет функцию детоксикации различных веществ, поступающих в организм [Abdel-Monem N. et al., 2013]. Тяжелые металлы запускают окислительный стресс, вырабатывая АФК, которые способствуют перекисному повреждению мембран гепатоцитов [Watanabe M. et al., 2003]. Биохимическими маркерами токсического поражения печени являются увеличение активности аминотрансфераз [Reus I.S. et al., 2003; Jadhav S. H. et al., 2007; Sobutskii M.P. et al., 2007; Wadaan M.A.M., 2009] и уменьшение концентрации альбуминов плазмы крови, чей синтез происходит в печени [Dabak D. et al., 2009; Abdel-Monem N. et al., 2013]. Помимо вышеперечисленных патологий, отравление солями тяжелых металлов может привести к развитию остеопороза и остеомаляции [Pounds J.G., 1984; Fullmer C.S., 1991.; Fullmer C.S., 1997; Satarug S., Moore M.R., 2004; Dalton T.P. et al., 2005; Abouhamed M., et al., 2007; Johri N. et al., 2010], воспалению дыхательных путей (Kirschvink N. et al., 2006), сердечно-сосудистым заболеваниям 66 [Messner B., Bernhard D., 2010], диабету [Schwartz G.G. et al., 2003], неврологическим нарушениям [Mates J.M. et al., 2010] и пр. Как было отмечено ранее, развитие патологий при интоксикации тяжелыми металлами основано на нарушении редокс-баланса на клеточном уровне, а также на переводе сульфгидрильных групп в дисульфидное состояние, что в свою очередь приводит к инактивации многих важных белков и ферментов. Учитывая значительное загрязнение окружающей среды тяжелыми металлами и их токсическое воздействие на живые организмы, встает вопрос о необходимости поиска препаратов, способных предотвращать интоксикацию тяжелыми металлами, проявлять антиоксидантные свойства и оказывать гепато- и нефропротекторное действие. Таким широким спектром биологической активности обладают соединения селена, как природные в форме селеноцистеина, так и синтетические: неорганические, например, селенит натрия, и органические, например, эбселен, ДАФС-25 и др. Антитоксическая активность органических соединений селена, серы и теллура Давно известно об антиоксидантной активности селена как составной части ГПО, обезвреживающей перекись водорода и липоперекиси в тканях и клетках [Mills G.C., 1957; Lubos E. et al., 2011]. Селензависимая ГПО при участии восстановленного глутатиона обезвреживает токсичный пероксид водорода до воды, предотвращая тем самым окислительное повреждение биомолекул. Установлена прямая зависимость между поступлением селена в организм и активностью ГПО [Костюк В.А., Потапович А.И., 2004; Tappel A.L., 1974; Takahashi K. et al., 1986]. Однако антиоксидантная активность селена и его соединений напрямую связана и с его антитоксической активностью. Ведущую роль как в антиоксидантном, так и антитоксическом действии селена играет 67 соотношение GSH/GSSG, поскольку от него зависит редокс статус клетки. Поступление селена в организм усиливает синтез ГПО и GSH, который наряду с другими тиолсодержащими молекулами является мишенью для связывания тяжелых металлов, о чем говорилось выше. Восстановленный глутатион и селен совместно участвуют в гомеостазе тиолсодержащих белков, таких как металлотионеины и цинк-фингерные белки. Изначально цинк-фингерные белки представлялись как группа транскрипционных факторов. Теперь известно, что «цинковые пальцы» это довольно обширная группа белков у эукариот, участвующая в различных клеточных процессах [Laity J.H. et al., 2001]. Около 3% генов генома человека кодируют белки с цинкфингерными доменами [Maret W., 2003], среди них не только транскрипционные факторы [Youn B.W. et al., 2001], но также и белки, отвечающие за стабильность генома и участвующие в репарации ДНК и контроле клеточного цикла [Laity J.H. et al., 2001]. Предположительно многие из этих белков регулируются окислением цинк-связывающих остатков цистеина, что ведет к потере ДНК-связывающей способности, выходу ионов цинка и образованию дисульфидных мостиков [Wilco D.E. et al., 2001]. В присутствии GSH происходит восстановление сульфгидрильных групп, благодаря чему они возвращают способность связывать ионы цинка и формировать «цинковый палец», а, значит, восстанавливают способность взаимодействовать с ДНК и регулировать экспрессию генов. Таким образом, поступление селена в организм усиливает синтез ГПО и восстановленного глутатиона. Последний не только связывает тяжелые металлы, но также восстанавливает активность «цинковых пальцев», регулирующих экспрессию генов, репарацию ДНК и клеточный цикл. Кроме того, превентивное введение соединений селена при кадмиевой интоксикации сопровождается одновременно и стабилизацией редокс-состояния клетки [Zhou Y.J. et al., 2009] и усилением синтеза шаперонов, необходимых для нормального фолдинга белков [Manikandan R. et al., 2011; Chen X. et al., 2012]. 68 Последнее обеспечивает восстановление конформации и активности белков и ферментов, необходимых для нормальной жизнедеятельности клетки. Немаловажный вклад в антитоксическое действие селена вносит селенопротеин Р, который является главной транспортной формой селена в плазме крови. Селенопротеин Р содержит 10 остатков селеноцистеина, 17 остатков цистеина и 28 остатков гистидина. Боковые радикалы этих аминокислотных остатков также являются мишенями для связывания тяжелых металлов [Suzuki K.T. et al, 1998]. Причем селеногидрильные группы обладают большим сродством к тяжелым металлам, в частности к ртути, чем сульфгидриные группы. Поэтому селенопротеин Р обладает большим сродством к ртути, чем другие белки. Исследование китайских ученых доказывает антитоксическое действие селена при отравлении ртутью [Chen C. et al., 2006]. У людей с повышенным содержанием ртути в организме отмечались типичные симптомы отравления ртутью: расстройство пищеварения, тремор, потеря веса, нарушение сна и памяти. Молярное соотношение селен/ртуть было близко к 1. Эти данные подтверждают задержку селена в организме людей с отравлением ртутью. У лиц контрольной группы концентрация ртути в сыворотке крови была низкой. Небольшая часть ртути была связана с альбуминами сыворотки крови, а основная часть ртути связывалась с селенопротеином Р и ГПО. Причем связывание ртути с селенопротеином Р увеличивалось при повышении концентрации ртути в сыворотке крови [Chen C. et al., 2006]. Неорганическая ртуть, поступая в организм животных, птиц и человека превращается в высоко токсичную органическую метилртуть. Препараты селена являются эффективными антитоксикантами при отравлении метилртутью беременных самок крыс, способствуют увеличению постнатальной выживаемости потомства и благоприятно влияют на репродуктивную функцию самок крыс при отравлении метилртутью [Beyrouty P., Chan H.M., 2006]. Токсичность метилртути зависит от содержания селена в тканях и от соотношения Hg/Se [Ralston N.V. et al, 69 2007, 2008; Scheuhammer A.M. et al., 2008]. Установлено, что пищевой селенометионин ингибировал аккумуляцию ртути у рыб, а ртуть в составе пищи рыб, напротив, усиливала аккумуляцию селена [Deng D.F. et al., 2008]. У птиц, питающихся рыбой, отмечено совместное накопление ртути и селена. Причем биоаккумуляция метилртути значительно превышает биоаккумуляцию неорганической ртути. Применение селенита натрия вызывает снижение биоаккумуляции неорганической ртути, но не снижает биоаккумуляцию метилртути [Cabanero A.I. et al., 2005]. Таким образом, селенит натрия оказывает антитоксическое действие при отравлении ртутью при молярном соотношении Se/Hg ≥ 1 [Liao C.Y. et al., 2007; Peterson S.A., 2009]. Метилртуть является одним из наиболее опасных токсикантов окружающей среды, который накапливается преимущественно в мозге животных и человека [Bakir F. et al., 1973; Castoldi A.F. et al., 2008]. Причем главной мишенью этого токсиканта является митохондриальная цепь переноса электронов [Allen J.W. et al., 2001; Shanker G. et al., 2004; Yin Z. et al., 2011]. Нарушение функции окислительного стресса и митохондрий тесно связано с развитием подавлением антиоксидантной системы организма. Причем метилртуть избирательно подавляет экспрессию мРНК главного антиоксидантного фермента ГПО1, тогда как экспрессия мРНК другого фермента ТРР, напротив, усиливается. Однако механизм подобного действия остается неясным [Usuki F. et al., 2011]. Кроме того, метилртуть имеет высокую аффинность к селеногидрильным и сульфгидрильным группам, а также к селенидам. Ранее сообщалось, что селенит натрия способен взаимодействовать с метилртутью и предотвращать ее токсическое действие на организм [Usuki F. et al., 2011]. Однако селенит натрия обладает собственной токсичностью и его применение возможно только в небольших дозах [Гмошинский И.В., Мазо В.К., 1999]. 70 Наиболее перспективными антиоксидантами и антитоксикантами при отравлении солями тяжелых металлов являются органические соединения селена. На протяжении последних тридцати лет изучается биологическая активность селеноорганического препарата эбселена (2-фенил-1,2-бензисоселеназол-3(2Н)он), который обладает ГПО- и ТРР-подобной активностями [Muller A. et al., 1984; de Freitas A.S. et al., 2010] и способен напрямую инактивировать свободные радикалы, образующиеся при интоксикации тяжелыми металлами [Masumoto K. et al., 1996.; Yin Z. et al., 2011]. Помимо эбселена антитоксическое действие оказывает природное селеноорганическое соединение селенонеин (2-селенил- Nα,Nα,Nα-триметил-Lгистидин), которое обладает не только высокой антирадикальной активностью, но также обезвреживает токсичную метилртуть [Yamashita Y., Yamashita M., 2010; Yamashita Y. et al., 2010, 2011]. Селенонеин, обнаруженный в крови морских рыб, может существовать в двух таутомерных формах: мономерных селенольной и селеноксо-формах и димерной окисленной форме [Yamashita Y., Yamashita M., 2010]. Причем в димерной окисленной форме селенонеин может восстанавливаться в клетках при участии GSH до мономерной формы, которая проявляет выраженные антиоксидантные свойства [Yamashita Y. et al., 2010]. Кроме того, селенонеин снижает аккумуляцию неорганической ртути и метилртути в культурах клеток рыб и человека. По мнению авторов [Yamashita Y. et al., 2011; Yamashita M. et al., 2013], биоинактивация метилртути происходит в три стадии: 1) образование комплекса «селенонеин-метилртуть»; 2) реакция деметилирования метилртути в составе этого комплекса и 3) образование селенида ртути, который затем выводится из организма [Yamashita M. et al., 2013]. Вышеизложенное позволяет сделать вывод, что органические препараты селена обладают значительной антитоксической активностью при отравлении как неорганической ртутью, так и органической метилртутью. 71 Наряду с различными селеноорганическими препаратами, изучаемыми за рубежом, эффективным антитоксическим препаратом в России является соединение ДАФС-25. Селеноорганический препарат является 1,5-дифенил-3селенапентандион-1,5 (ДАФС-25) оказывает антитоксическое действие при разовом (выживаемость 80%) и многократном введении (выживаемость 100%) в организм беспородных мышей арсенита натрия [Кирова Ю.И., 2004]. Токсичное действие арсенита связано с окислением сульфгидрильных групп в составе ферментов, что приводит к острому отравлению [Rabbani G.H. et al., 2003]. Антитоксический эффект ДАФС-25 in vivo связан с наличием в его молекуле атома селена в степени окисления +2, благодаря которому, вероятно, происходит восстановление арсенита натрия до низкотоксичного As0 [Кирова Ю.И., 2004]. Наравне с ртутью и мышьяком чрезвычайно опасными поллютантами считаются свинец и кадмий, внесенные ВОЗ в список токсических веществ I класса опасности. Проникая в организм, свинец и кадмий оказывают токсическое воздействие практически на все органы и ткани, но особенно на почки, печень [Брин В.Б. и др., 2011; 2012] и мозг [Li W.-H., et al., 2013], поскольку именно в этих органах аккумулируются большинство тяжелых металлов. Интоксикация тяжелыми металлами сопровождается поражением печени и подтверждается увеличением активности АлАТ, снижением коэффициента де Ритиса и значительным повышением концентрации общего билирубина в плазме крови [Брин В.Б. и др., 2011]. Кроме того, накопление ионов тяжелых металлов в клетках приводит к усилению свободно-радикальных процессов [Коваль Ю.И., Бокова Т.И., 2010]. Вместе с тем применение антиоксидантных препаратов способно снизить биоаккумуляцию тяжелых металлов и их токсическое воздействие на организм. Поскольку препарат ДАФС обладает антиоксидантной активностью, он также является эффективным антитоксикантом при отравлении мышей солями свинца и цинка [Макарова Е.С., Павленко Г.И., 2011]. Как известно, на 72 молекулярном уровне свинец и цинк действуют как тиоловые яды. В механизме их действия ведущая роль принадлежит процессам сульфгидрильными группами, которые обуславливают ферментов, их взаимодействия активность с многих участвующих в различных метаболических и регуляторных процессах. Результаты, полученные авторами, свидетельствуют о достоверном снижении содержания SH-групп в печени крыс при интоксикации солями свинца и цинка. Добавление в корм селеносодержащего препарата ДАФС-25 положительно сказывалось на выработке тиоловых групп: на всём протяжении опыта авторы наблюдали нормализацию данного показателя и у животных при воздействии одного свинца, и у животных, получавших комбинацию металлов. Причем у крыс, получавших в качестве антидота ДАФС-25, все биохимические показатели, характеризующие функциональное состояние печени (активность ферментов АлАТ, АсАТ, щелочной фосфатазы, концентрация билирубина, общего белка), и в какой-то степени почек (концентрация креатинина и мочевины), находились в пределах контрольных величин и достоверно от них не отличались [Макарова Е.С., Павленко Г.И., 2011]. Таким образом, необходимо подчеркнуть взаимосвязь между установленной антиоксидантной природой селеноорганических соединений и их антитоксической активностью. Основу механизмов их биологической активности составляет инактивация токсичных активных форм кислорода (АФК), возникающих при любом стресс-воздействии на организм, а также инактивация тяжелого металла либо за счет связывания с соединением селена, либо за счет взаимодействия с сульф/селеногидрильными группами глутатиона или белков тканей организма. Наравне с селеносодержащими соединениями эффективными антидотами при отравлении солями тяжелых металлов могут выступать также соединения серы, которые способны связывать тяжелые металлы, например, ртуть, своими сульфгидрильными группами [Go M.L. et al., 2005; Peng H. et al., 2012]. Однако селеногидрильная группа селеноцистеина в активном центре селенопротеинов при 73 физиологическом значении рН обладает большей редокс-чувствительностью по сравнению с сульфгидрильной группой остатка цистеина [Emmert S.W. et al., 2010]. Следовательно, соединения селена являются более эффективными антиоксидантами и антитоксикантами при отравлении солями тяжелых металлов. К сожалению, информации о потенциальной антитоксической активности соединений теллура крайне мало, но есть сведения о возможном участии теллура в образовании аминокислоты теллур-метионина, которая способна связывать атомы тяжелых металлов, в частности кадмия. На этом основана методология применения наночастиц теллурида кадмия в качестве квантум-дот для диагностики онкологических заболеваний [Ba L.A. et al., 2010]. Важно отметить, что в ряду сера → селен → теллур наибольшей токсичностью для живых организмов обладает теллур (и его производные: теллуроксианион, теллурит и теллурат) [Chasteen T.G. et al., 2009]. Таким образом, проведенный анализ литературы позволяет сделать вывод, что наиболее эффективными антитоксикантами при отравлении солями тяжелых металлов являются органические соединения селена и серы. 1.4. Антибактериальная активность халькогенорганических соединений в отношении грамотрицательных и грамположительных микроорганизмов Бактериальная мульти-резистентность становится серьезной проблемой во всем мире. Для решения этой проблемы научное сообщество и фармацевтическая промышленность предпринимают немалые усилия для открытия новых соединений с антибактериальными свойствами. Однако, за последние 40 лет были созданы только два новых антибиотика: оксазолидинон линезолида и липопептид даптомицина [Ford C.W. et al., 2001; LaPlante K., Rybak M., 2004]. Несмотря на это, штаммы, устойчивые к этим новым антибиотикам, появляются постоянно, делая ситуацию критической. 74 Ранее сообщалось, что некоторые антибиотики могут использовать в качестве синергистов генетически модифицированные бактериофаги или металлоорганические соединения [Banin E. et al., 2008; Lu T.K., Collins J.J., 2009; Hemaiswarya S., Doble M., 2010]. В 2007 году был предложен общий механизм токсичности антибиотиков, основанный на образовании активных форм кислорода (АФК) [Kohanski M. et al., 2007]. С другой стороны, еще в 1932 году Александр Флеминг сообщил об антибактериальных свойствах теллурита (TeO32-) и пенициллина [Fleming A., 1932] и с тех пор теллурит используется для выделения теллурит-резистентных штаммов Escherichia coli O157, Proteus spp., и других бактерий [Chapman P. et al., 1991]. В течение последних лет авторов интересовали вопросы молекулярных механизмов токсичности соединений халькогенов, которые были основаны на выработке АФК [Pérez J.M. et al., 2007], повреждении ферментов [Calderon I. et al., 2009; Castro M. et al., 2009], окислении глутатиона [Turner R.J. et al., 2001] или усилении липопероксидации [Perez J. et al., 2008]. В настоящее время во избежание появления мульти-резистентности у бактерий, предлагается использовать соединения с разными видами токсичности. В этой связи считается перспективным совместное применение современных антибиотиков и соединений халькогенов (теллура и селена) [Molina-Quiroz R.C. et al., 2012]. За последние 20 лет были синтезированы органические соединения селена и теллура, многие из которых показали значительную антимикробную активность. Антибактериальное действие селенорганических соединений На протяжении антибактериальная последних активность двух нескольких десятилетий классов обнаружена селенорганических 75 соединений: 4Н-селенопиранов и солей селенопирилия, селеноксантенов, селеноциклогексанов, бензоселенохроменов и др. 4Н-селенопираны (I) и соли селенопирилия (II) проявляли выраженную антимикробную активность: R R1 Ar ArR1-p R R2 Se Ar + Ar Se A- I Ar 2 p-R Ar II 1 + SeA ArR2-p III-V 2 I: Ar= Ph, R=Ph, R =H, R =H; II: Ar=Ph, R=Ph, R1=CH3, R2=H; III: Ar=Ph, R=C6H4OCH3-p, R1=H, R2=H; IV: Ar=C6H4OCH3-p, R=Ph, R1=H, R2=H; V: Ar=Ph, R=H, R1=CH3, R2=CH3. Например, 4Н-селенопираны в концентрации 0,7-12 мкг/мл обладали высокой антибактериальной активностью в отношении стафилококков и грибов рода Кандида [Древко Б.И., 1997]. Соли селенопирилия в концентрации 0,78-1,56 мкг/мл проявляли противостафилококковую и антигрибковую активность. Причем замена аниона Cl-, BrF4-, ClO4- на HCOO- и C6H4(OH)COO- вело к снижению антибактериальной активности. При этом хлориды селенопирилия были эффективными антимикробными агентами по отношению к стафилококкам, резистентных к антибиотикам [Древко Б.И., 1997]. Следовательно, 4Н-селенопираны и соли селенопирилия проявляли значительную противостафилококковую [Древко Б.И., 1997]. и противогрибковую активность 76 В работе [Ратушная Е.В., 2001] доказана антибактериальная активность пергидроселеноксантена и солей селенопирилия, представленных на рисунке 4: Cl OCH3 + Se Cl H3CO Se II OCH3 I F Se O H2O Se III IV Рис. 4. Соединение I - Хлорид 2,4,6-трипараметоксифенилселенопирилия, соединение II - 9-парахлорфенилокгагидроселеноксантен, и 9-парафторфенилоктагидроселеноксантен соединение III - соединение 4 - пергидроселеноксантен. Из четырех представленных выше селенорганических соединений наибольшей антибактериальной активностью на клетки E.coli НВ-101, выросших на 1% агаре из жидкой питательной среды, обладали соединения I - хлорид 2,4,6трипараметоксифенилселенопирилия и IV – пергидроселеноксантен в концентрациях 10-2, 10-3, 10-4, 10-5 М. При этом наибольшей антибактериальной активностью обладало соединение IV. 77 Кроме того, доказана антибактериальная активность этих препаратов против St. aureus 209P. Наибольшей антимикробной активностью обладало соединение I во всех изученных концентрациях. Соединения IV > II >III проявляли меньшую, но достоверную активность против золотистого стафилококка. Таким образом, наибольшей антибактериальной активностью обладали соединение IV в концентрации 10-6 М в отношении E. coli и соединение I в концентрации 10-5 М в отношении St. aureus 209P [Ратушная Е.В., 2001]. Исследование антибактериальной активности на примере культуры Escherichia coli показало, что селеноорганические соединения трифторацетат 2,4,6-три-(п-метоксифенил)-селенопирилия, 2,6-дифенил-4-(п-метоксифенил)-4Нселенопиран и 2,4,6-трифенилселенациклогексан оказывают значительное влияние на процессы ее жизнедеятельности [Меркулова Е.П., 2012]: C6H4OCH3 p C6H4OCH3 p p H3COC6H4 Se+ C6H4OCH3 p CF3COO- C6H5 Трифторацетат 2,4,6-три- Se C6H5 2,6-дифенил-4-(п-метоксифенил)- (п-метоксифенил)-селенопирилия 4Н-селенопиран C6H5 C6H5 Se C6H5 2,4,6-трифенилселенациклогексан Действие соединения трифторацетат 2,4,6-три-(п-метоксифенил)- селенопирилия выражается наибольшим угнетением роста клеток при обработке концентрациями 1·10-3, 1·10-2 и 1·10-1 мг/мл. Соединения 2,6-дифенил-4-(п- 78 метоксифенил)-4Н-селенопиран и 2,4,6-трифенилселенациклогексан значительно подавляли рост культуры E. сoli концентрациях от 1·10-4 - 1·10-1 мг/мл [Меркулова Е.П., 2012]. В последние годы за рубежом широко изучается биологическая активность селеноорганического соединения эбселен (2-фенил-1,2-бензисоселеназол-3-(2Н)он). Установлена антиоксиданитная, противоопухолевая, противовоспалительная, антиатеросклеротическая, цитопротекторная и антибактериальная активность производных эбселена [Narajji C. et al., 2007]. Эбселен проявляет антибактериальную активность в низких концентрациях (MIC=2,0-32,0 мкг/мл) против широкого спектра грамположительных и грамотрицательных бактерий, а также различных грибов и вирусов [Młochowski J. et al., 2007]. Напрмер, установлена антибактериальная активность 8 производных эбселена (3 бензизоселеназола, 1 бензизоселенозалон и 4 дисарилдиселенида) против 258 штаммов Сandida albicans и 127-8b штамма Saccharomyces cerevisiae. Эбселен и 2-ацетил-1,2-бензизоселеназол-3(2Н)-он вызывали значительное подавление роста 127-8b штамма Saccharomyces cerevisiae, хотя менее заметное угнетение роста эти соединения оказывали на рост штаммов Сandida albicans. Бензизоселенозалоны значительно сокращали рост грамотрицательных штаммов Escherichia coli K-12 ROW и грамположительных штаммов Staphylococcus aureus 209P (Oxford) [Bień M. et al., 1999]. Причем эбселен проявлял бактерицидную активность в концентрации MIC=1,56 мкг/мл против 90% метициллин-резистентных штаммов S. aureus. Интересно заметить, что серный аналог эбселена не обладал антибактериальной активностью [Nozawa R., et al., 1989]. Кроме того, обнаружена активность аналогов эбселена против вируса герпеса, вируса энцефаломиокардита и грамположительных бактерий Bacillus in vitro [Wójtowicz H. et al., 2004], тогда как в отношении грамотрицательных 79 бактерий (E. coli, P. aeruginosa, K. pneumoniae) их активность была заметно слабее. Ряд производных эбселена были эффективны против грибов A. niger [Pietka-Ottlik M. et al., 2008]. Установлено, антибактериальной что некоторые активностью, аналоги чем эбселена стандартные обладали большей химиотерапевтические препараты. Например, производные нового класса селеноорганических препаратов 5,7-диарил-4,4-диметил-4,5,6,7-тетрагидропиридино[3,4-d]-1,2,3-селенадиазолов были более эффективны против Escherichia coli, Staphylococcus aureus, betaHeamolytic streptococcus, Vibreo cholerae, Salmonella typhii и Shigella flexneri, чем стандартный антибиотик ципрофлоксацин. Кроме того, одно из соединений этого класса проявляло большую активность против Aspergillus flavus и Mucor, чем флюконазол [Gopalakrishnan M. et al., 2008]. Антибактериальная активность эбселена и некоторых других его аналогов против грамположительных и грамотрицательных бактерий обусловлена их способностью взаимодействовать с тиольными группами макромолекул [Młochowski J., 2007], а фунгицидная активность основана на способности эбселена ингибировать мембранный фермент Н+-АТФаза дрожжей [Chan G. et al., 2007]. Следует отметить, что сравнительное исследование антибактериальной активности серо- и селеноорганических соединений свидетельствует о значительно большей противомикробной активности органических соединений селена, поскольку селеносодержащие соединения были в 30-75 раз более токсичнее, чем их серные аналоги и поэтому их биологическая активность против ряда бактерий была также выше. В частности, некоторые селеноорганические соединения подавляли рост патогенных грибов растений в дозах значительно ниже токсичных и были эффективнее кетоконазола [Nozawa R., 1989; Deidda D. et al., 1997]. 80 Таким образом, селеноорганические соединения являются перспективными антибактериальными препаратами, проявляющими высокую биологическую активность против антибиотикорезистентных штаммов микроорганизмов и обладающие большей эффективностью, чем стандартные химиотерапевтические препараты. Антибактериальная активность теллуроорганических соединений Сравнительно недавно появилась информация об антибактериальной активности некоторых теллурорагнических соединений [Soni D. et al., 2005]. Гетероциклические производные дийодида дибензоилтеллура показали антибактериальную активность против грамположительных (Bacillus subtilis, Staphylococcus aureus) и грамотрицательных (Escherichia coli и Salmonella typhi) бактерий, однако активности у исходного препарата не было обнаружено. Также сообщалось об антимикробной активности некоторых теллуроорганических и селеноорганических соединений, полученных из альфа-бромо-4- метилацетофенона. Причем теллур-содержащие аналоги этого соединения показали более высокую активность, чем селеноорганические. Комплексы теллуроорганического трихлорида с тетраметилтиомочевиной, бензамидазолом, триэтиламином, гамма-пиколином и оксидом три-фейларзина проявляли антибактериальную активность против Pseudomonas aeruginosa и Bacillus subtilis [Soni D. et al., 2005]. Как ранее сообщалось, теллуроорганические соединения обладали антимикробной активностью в отношении E. coli, тогда как бактерии P. aeruginosa не были чувствительны к этим соединениям. Карбоксилатные производные проявляли меньшую активность, чем исходные соединения. Фенацил и нафтацил (3-метил-4-гидроксифенил) дихлорид теллуроорганические соединения (соединения 2 и 5 соответственно) проявляли значительную активность против грамположительных и грамотрицательных бактерий [Soni D. et al., 2005]: 81 Среди исследованных соединений максимальной активностью против грамположительных бактерий обладало соединение 2, тогда как соединение 5 было активно против всех бактерий. Нафтацил и стирилацил (п-метоксифенил) дихлорид теллуроорганические соединения (соединения 4 и 6) проявляли хорошую активность против всех грамотрицательных бактерий. Однако максимальную активность они проявляли против P. aeruginosa. Фенацил и нафтацил (п-гидроксифенил) дихлорид теллуроорганические соединения (соединения 1 и 3) показали более выраженную активность против и грамположительных, и грамотрицательных бактерий. Ранее сообщалось, что теллурорганические соединения, у которых атом теллура расположен у алифатического атома углерода, обладали лучшей активностью, чем соединения, имеющие атом теллура у ароматического атома [Soni D. et al., 2005]. Это, вероятно, обусловлено более легким разрывом связи «С-Те» у алифатического атома углерода и последующим освобождением атома теллура в виде неорганического теллура (IV), который значительно токсичнее 82 своей органической формы [Садеков И.Д., Минкин В.И., 1995; Meinerz D.F., et al., 2013]. Неорганические соединения теллура и селена также проявляли высокую антибактериальную активность против грамположительных и грамотрицательных бактерий, поскольку оксианионы теллурита (ТеО32-), селената (SeO42-) и селенита (SeO32-) хорошо растворимы в воде и очень токсичны для живых организмов [Choudhury H.G. et al., 2011]. Метаболизм халькогенов в бактериальных клетках Учитывая, что в ряду халькогенов – сера, селен, теллур – только сера и селен являются эссенциальными элементами, их метаболизм в бактериальных клетках происходит различными путями. Сера является незаменимым для всех живых организмов элементом, который участвует в синтезе серосодержащих аминокислот цистеина и метионина, огромного числа белков, а также входит в состав многих коферментов, необходимых для метаболических превращений в клетке [Kopriva S. et al., 2012]. Важной особенностью серы, входящей в состав различных соединений, является способность быстро менять валентность. Сера находится в природе в форме неорганического сульфата, который поступает в живые организмы, восстанавливается и встраивается в органическую молекулу. Поэтому большинство биоорганических соединений содержат серу в органической сульфидной или тиольной формах. Однако многоклеточные организмы и большинство микроорганизмов, ведущих паразитический образ жизни, утратили способность восстанавливать сульфат. Только растения, грибы и бактерии сохранили способность ассимилировать сульфат и, тем самым, обеспечивать биогеохимические циклы серы [Kopriva S. et al., 2012]. Метаболические превращения серы включают образование цистеина, метионина и S-аденозилметионина, а также других серосодержащих метаболитов, 83 например, глутатиона, ацетил-КоА жизнедеятельности клетки аденозилметионина как и вещества донора др. Некоторые синтезируются метильной важные при группы, участии как, для S- например, фосфатидилхолин – фосфолипид, входящий в состав биологических мембран. Метаболические пути ассимиляции серы также связаны с образованием многих белков, в том числе железосерных белков – компонентов митохондриальной цепи переноса электронов. Причем нарушение синтеза метионина на уровне экспрессии соответствующих генов приводит к задержке клеточного цикла. Ранее было показано, что двойная делеция генов MET31 и MET32, отвечающих за синтез метионина у дрожжей, коррелирует со способностью клетки бороться с окислительным стрессом [Petti A.A., et al. 2011]. Кроме того, установлено, что транскрипционные факторы, отвечающие за синтез метионина, более тесно связаны с метаболическим циклом дрожжей, особенно с липидным обменом [Hickman M.J. et al., 2011], чем транскрипционные факторы других аминокислот [Tu B.P., et al. 2005; Murray D.B. et al., 2007]. Совсем недавно было открыто влияние регуляторов синтеза метионина на различные метаболические фосфатидилхолина, дезоксирибонуклеотидов, процессы, кардиолипина, в частности, пиримидинов, на биосинтез пуринов, жирных кислот, триптофана, НАД, а также на метаболизм глутатиона и азотистый обмен [Petti A.A. et al., 2012]. И, наконец, необходимо отметить участие метионил-тРНК в процессе трансляции. Метионин кодируется старт-кодоном АУГ и запускает биосинтез любого белка в дрожжевой клетке, поэтому недостаточное поступление серы в клетку может вызвать не только нарушение метаболических процессов, но и привести к остановке клеточного цикла и роста клетки [Petti A.A. et al., 2012]. Большое значение в метаболических процессах у бактерий имеет аминокислота цистеин благодаря химическим свойствам серы. Сульфид является 84 мощным нуклеофильным агентом, участвующим в реакциях с участием активных групп (например, КоА), в образовании дисульфидных мостиков, а также в транспорте электронов в составе железосерных белков митохондриальной цепи переноса электронов [Hell R., Wirtz M., 2011]. Благодаря значительной биологической роли серы в бактериальных клетках серосодержащие соединения не проявляют антибактериальной активности. Кроме того, отмечается существенное понижение энергий связей «Углерод-Халькоген» в ряду S, Se, Te, поэтому в органических соединениях халькогенов наибольшей энергией обладает связь «С-S», а, следовательно, сера практически не освобождается из своих органических соединений [Садеков И.Д., Минкин В.И., 1995]. Вместе с тем, связь «С-Те» разрывается легче, чем связи, образованные другими халькогенами и происходит с освобождением атома теллура в неорганического теллура (IV), который значительно токсичнее виде своей органической формы [Садеков И.Д., Минкин В.И., 1995; Meinerz D.F. et al., 2013]. Поскольку энергия связи «C-Se» имеет средние значения между связями «C-S» и «С-Те», освобождение атома селена происходит сложнее, чем из соответствующих соединений теллура, но легче, чем из серосодержащих соединений. Освободившиеся атомы халькогенов превращаются в соответствующие анионы, обладающие высокой токсичностью и способные метаболизироваться в живых клетках. Оксианионы теллурита (ТеО32-), селената (SeO42-) и селенита (SeO32-) хорошо растворимы в воде и очень токсичны для живых организмов, тогда как элементарные теллур (Те0) и селен (Se0) нерастворимы в воде и почти нетоксичны [Choudhury H.G. et al., 2011]. Грамотрицательные бактерии Escherichia coli и Pseudomonas syringae способны обезвреживать токсичные для них оксианионы теллурита, селената и селенита либо за счет реакции трансметилирования с S-аденозилметионином в 85 качестве донора метильной группы, либо за счет восстановления до элементарных теллура/селена. Реакция трансметилирования с S-аденозилметионином катализируется бактериальной неорганические тиопуринметилтрансферазой, и органические которая соединения селена может метилировать (селеноцистеин) до диметилселенида и диметилдиселенида [Ranjard L. et al., 2002]. Аналогичным образом может происходить метилирование теллурита до диметилтеллурида, токсичность которого намного меньше, чем у теллурита [Cournoyer B. et al., 1998; Choudhury H.G. et al., 2011]. Причем фермент бактериальная тиопуринметилтрансфераза принадлежит к группе метилтрансфераз, которые имееются и у эукариот (человека, мышей, крыс и т.д.), а, значит, подобным образом происходит метаболизм халькогенов и у млекопитающих [Otterness D.M., Weinshilboum R.M., 1987; Szumlanski C. et al., 1996]. Для ряда микроорганизмов установлен механизм восстановления теллурита до элементарного теллура: 1) Micrococcus lactilyticus восстанавливают неорганические окианионы, в том числе теллурит, с участием молекулярного водорода [Woolfolk C.A., Whitely H.R., 1962]; 2) различные виды Rhodobacter spp. утилизируют теллурит и другие оксианионы с помощью железосерных белков, выполняющих функцию терминальных акцепторов электронов [Moore M. D., Kaplan S., 1992], 3) нитрат-редуктазы E. coli обладают теллурит-редуктазной активностью [Avazeri C. et al., 1997]. Вместе с тем известно, что агар Байрд-Паркера содержит теллурит (ТеО32-) и используется в качестве дифференциальной среды для выделения золотистого стафилококка (Staphylococcus aureus) из пищевых продуктов [Baird-Parker A., 1962]. 86 Ранее сообщалось, что в клетках S. аureus, E. coli и Pseudomonas соединения теллура, в том числе теллурит, могут взаимодействовать с сульфгидрильными группами цистеина, сокращая уровень тиольных групп и таким образом снижая токсичность теллурита [Albeck A. et al., 1998; Turner R. et al., 1999; Lithgow J. K. et al., 2004; Tremaroli V. et al., 2007]. Кроме бактериальной того, установлено, клетке ведет к что снижение остановке содержания роста, цистеина нарушению в синтеза митохондриальных железосерных белков, что в свою очередь обусловливает образование значительного количества АФК и запуск окислительного стресса в клетке [van de Waterbeemd B. et al., 2013]. Это вероятно, обусловлено участием цистеина в синтезе большого количества функционально важных пептидов, белков и ферментов, например, железосерных белков и цитохромов дыхательной цепи, глутатиона, а также ферментов фумаразы и аконитазы [Fuentes D.E. et al., 2007]. Цистеин-содержащие молекулы, такие как глутатион и тиоредоксин, играют важную роль в антиоксидантной защите бактериальной клетки в том числе и от теллурит-опосредованного окислительного стресса. Однако есть данные, что тиолы способны, напротив, усиливать токсичность теллурита [Turner M.S. et al., 2007]. Например, S. аureus имеет повышенное количество тиольных групп, которые вырабатывают значительное количество АФК в процессе внутриклеточного восстановления теллурита. Предполагается, что этот механизм лежит в основе токсичности не только соединений теллура [Tremaroli V. et al., 2007], но и соединений селена, в частности селенита [Kessi J., Hanselmann K.W., 2004]. Действительно, теллурит в клетках P. Pseudoalcaligenes KF707 способствует образованию АФК, окислению тиольных групп, а также структурным и функциональным модификациям дыхательной цепи [Tomaso G.D. et al., 2002; Tremaroli V. et al., 2007]. Восстановление и детоксикация теллурита/селенита во 87 многих бактериях связаны с дыхательной цепью и тиольными соединениями [Zannoni D. et al., 2008]. Относительно недавно была установлена роль дыхательной цепи в восстановлении телурита до элементарного теллура в клетках некоторых теллурит-резистентных грамотрицательных бактерий (Pseudomonas, Agrobacterium, Erwinia и Escherichia). В пользу этого свидетельствует отложение черных гранул элементарного теллура вблизи от периплазматических или цитоплазматических мембран, содержащих терминальные мембран-связанные оксидазы [Тrutko S. M., et al. 2000]. Причем цитоплазматическая аккумуляция элементарного теллура характерна для стационарной фазы роста, хотя функциональные и структурные изменения дыхательной цепи уже очевидны в начале экспоненциальной фазы роста [Tomaso G. D. et al., 2002]. Кроме этого, также сообщалось о специфической активации цитохрома с в клетках P. aeruginosa, при которой наблюдалось снижение уровня теллура в клетках. Это подтверждает, что восстановление теллурита и молекулярного кислорода осуществляется одними и теми же восстанавливающими эквивалентами дыхательной цепи. В клеточных мембранах P. pseudoalcaligenes KF707, выращенных на среде с теллуритом, обнаружено снижение активности цитохром с-оксидазы на 50%; компонентов цитохромов b и с на 50% и 75% соответственно [Tomaso G. D. et al., 2002]. Есть мнение, что эти эффекты теллурита на дыхательную цепь бактерий связаны с его влиянием на определенные «дыхательные» гены [Tomaso G. D. et al., 2002]. В клетках Geobacillus stearothermophilus V идентифицированы 3 гена, которые кодируют ферменты, участвующие в метаболизме цистеина и экспрессия которых определяет теллурит-резистентность у E. coli. При инкубации с теллуритом в бактериальных клетках происходит усиление экспрессии этих генов, 88 что в свою очередь, приводит к синтезу цистеин-содержащих антиоксидантов, препятствующих развитию окислительного стресса [Fuentes D.E. et al., 2007]. Кроме того, установлено, что в отсутствии теллурита в бактериальных клетках теллурит-резистентных P. Pseudoalcaligenes определяется большее количество глутатиона и разветвленных аминокислот (валина и лейцина), чем в клетках дикого типа. Подобное наблюдение было сделано также при паракватиндуцированном окислительном стрессе в клетках Escherichia coli [Tweeddale H. et al., 1999]. При инкубировании клеток P. Pseudoalcaligenes Т5 с теллуритом обнаружили снижение содержания глутамата, аспартата, глицина, гистидина и триптофана. Глутамат и аспартат – это гликогенные аминокислоты, которые могут превращаться в метаболиты цикла Кребса и использоваться в качестве энергетических субстратов. С другой стороны, глицин и глутамат – аминокислоты, участвующие в синтезе глутатиона. Авторы предполагают, что увеличенное содержание глутатиона в клетках Т5 помогает защитить тиольные соединения от окислительного действия теллурита [Tremaroli V. et al., 2009]. Вероятно, теллурит может реагировать с сульфгидрильной группой тиольных соединений, в том числе глутатиона (RSH) с образованием теллурэфирных дисульфидных связей (RS-Te-SR) в активном центре соответствующего белка или фермента, активность которого вследствие этого снижается [Turner R. et al., 1999]. Следовательно, бактериальные клетки способны переводить токсичные формы теллурита и селенита в малотоксичные элементарные формы селена и теллура с участием сульфгидрильных групп тиолсодержащих соединений. Однако этот процесс сопровождается нарушением функций дыхательной цепи и повышенной выработкой АФК на фоне снижения содержания тиольных групп цистеина, что вызывает тяжелые нарушения в бактериальной клетке, остановку ее роста и размножения. 89 Различия в антимикробной активности халькогенсодержащих соединений в отношении грамотрицательных и грамположительных бактерий, вероятно, связаны также с особенностями строения их клеточных стенок. Структурные различия в мембранах грамположительных и грамотрицательных бактерий обусловливают различие в проходимости различных веществ, и это может объяснить неэффективность большинства других синтетических и натуральных антимикробных агентов против Escherichia coli и Pseudomonas aeruginosa. Наружная мембрана этих бактерий состоит из липополисахарид-липопротеиновых комплексов и недоступна для большинства антибиотиков, включая пенициллин и другие препараты, такие как олиго-3-аминопиридин и др. У грамположительных бактерий над пептидогликаном и тейхоевыми кислотами располагается лишь аморфная капсула полисахаридной (крахмалоподобной) природы (рисунок 5). Рис. 5. Различия в строении клеточных стенок грамотрицательных и грамотрицательных бактерий. Транспорт питательных веществ (сахаров, аминокислот) и антибиотиков внутрь бактериальной клетки осуществляется через воронкообразные белковые структуры (пориновые каналы), встроенные в липополисахаридный слой. Гидрофобные соединения (среди антибиотиков к таковым относятся хинолоны, макролиды и тетрациклины) способны диффундировать через 90 липополисахаридный слой. Сравнительно высокомолекулярные антибиотики, такие как гликопептиды, природные пенициллины с трудом проникают через пориновые каналы грамотрицательных бактерий, чем и объясняется природная устойчивость этих микроорганизмов к перечисленным препаратам [Сидоренко С.В., 2000]. В заключение можно предположить, что высокая антибактериальная активность различных селено- и теллуроорганических соединений обусловлена, в первую очередь, их физико-химическими свойствами, а именно низкой молекулярной массой и гидрофобностью молекулы, благодаря чему эти соединения могут легко проникать через бактериальную мембрану, метаболизироваться бектериальными ферментами с освобождением атомов селена и теллура соответственно, взаимодействовать с сульфгидрильными группами белков и глутатиона, понижая тем самым их количество. Последнее приводит к повышенной выработке АФК, окислительному повреждению внутриклеточных структур, остановке роста и размножения бактериальных клеток. 1.5. Гормоноподобное действие синтетических нестероидных соединений Уже на протяжении более полувека изучается потенциальная гормоноподобная активность различных соединений стероидной и нестероидной природы, действие которых направлено либо на усиление, либо на торможение активности природных гормонов. Хорошо известно гормоноподобное действие ряда нестероидных препаратов, обладающих эстрогенной, андрогенной и глюкокортикоидной активностью [Biggadikea K., et al. 2009.; Caboni L., et al. 2012.]. Половые гормоны - производные трех углеводородов: андростана, прегнана и эстрана - можно разделить на две группы: женские и мужские [Харкевич Д.А., 2006.]. 91 Известны три естественных эстрогенных гормона: эстрадиол, эстрон (фолликулин) и эстриол: Эстрадиол-17β Эстрон Эстриол Эстрогенной активностью обладают многие производные естественных эстрогенных гормонов (их эфиры и ацетиленовые производные). Известен целый ряд синтетических веществ, обладающих эстрогенной активностью, по химическому строению резко отличающихся от естественных гормонов: Диэтилстильбэстрол Диенэстрол Бензэстрол (октэстрол) Диместрол Эти синтетические Метилстильбол Хлортрианизен (тейс) вещества воспроизводят действие натуральных эстрогенных гормонов и находят в тех или иных странах применение в качестве лекарственных веществ [Харкевич Д.А., 2006.]. Препараты с эстрогенным действием имеют широкое применение. Их используют при недоразвитии женских половых органов (инфантилизм), отсутствии менструаций, дисменореях, в некоторых случаях функциональных маточных кровотечений, при бесплодии, 92 слабости родовой деятельности, переношенной беременности, при расстройствах в преклимактерическом и климактерическом периоде [Харкевич Д.А., 2006.]. Эстрогенными свойствами, хотя и менее выраженными, обладают и гормоноподобные соединения растительного происхождения — фитоэстрогены [Никитин А.И., 2002.]. Фитоэстрогены содержатся в растениях и грибах в чистом виде или в качестве предшественников соединений с эстрогенной активностью. Установлено [Ayres D.C., 1990; Barrett J. R., 1996.], что их физиологическое значение для самих растений заключается в регуляции процессов роста и размножения, защите растений от вредного действия ультрафиолетового излучения и поражения грибами и другими паразитами, контроле действия других биологически активных соединений и т.п. Многие из веществ этого класса обладают также антивирусным и бактерицидным свойствами. Биохимический анализ показал, что фитоэстрогены по структуре обладают определенным сходством с эндогенными эстрогенами животных и имеют близкую с ними молекулярную массу. Указанные свойства позволяют им “узнавать” эстрогенные рецепторы и связываться с ними. Это было доказано в опытах in vitro на примере энтеролактона и энтеродиола, которые активно реагировали с эстрогенными рецепторами цитозоля матки овец и крыс [Coward L. et. al., 1993; Shutt D.A. et al., 1972] и рецепторами к эстрогенам клеток рака молочной железы человека [Martin P.M. et al., 1972], что позволило рассматривать фитоэстрогены в качестве соединений, обладающих способностью мимикрировать или блокировать действие эндогенных эстрогенов. Характер действия фитоэстрогенов в этих случаях зависит от их способности не только быть подходящим “ключом” к “замку-рецептору”, но и способностью “ключа” “открывать замок”, стимулируя в клетке специфический синтез. В последнем случае фитоэстрогены проявляют эстрогенные свойства. При отсутствии такой способности фитоэстрогены могут выступать в качестве антиэстрогенных агентов. Следовательно, фитоэстрогены обладают 93 потенциальной способностью модифицировать механизмы, регулирующие половой цикл и репродуктивный процесс у человека и животных. Гормоноподобная активность фитоэстрогенов может быть связана, как полагают некоторые авторы, и с другими механизмами. Выявлена способность фитоэстрогенов стимулировать образование в печени глобулинов, связывающих половые стероиды [Aldercreutz H. et al., 1993], и таким путем модулировать биологическую активность эндогенных половых гормонов. Установлено, что ряд изофлавоноидов и лигнанов, влияя на активность ферментов ароматазной системы, могут ингибировать переход андростендиона в эстрон, а последнего – в 17-бета эстрадиол [Henderson B.E. et al., 1988]. Показано также, что изофлавоноид генистейн ингибирует активность тирозинспецифической протеинкиназы, “работающей на уровне рецепторов”, к ряду ростовых факторов: эпидермальному, инсулиноподобному, тромбо- и моноцитарному, играющих важную роль в регуляции процессов пролиферации и трансформации клеток [Shutt D.A. et al., 1972.]. Из коры надпочечников выделено большое число гормонов, получивших название кортикостероидов. В зависимости от той роли, которую играют эти вещества в организме, их разделяют на две основные группы: минералокортикоиды и глюкокортикоиды. Минералокортикоиды, основными представителями которых являются альдостерон и дезоксикортикостерон (Δ4-прегнен-21-ол-3,20-дион), играют большую роль, как показывает само название, в регуляции минерального обмена [Харкевич Д.А., 2006.]. Они способствуют задержке в организме воды и натрия, повышают выделение из организма калия, усиливают явления анаболизма (синтез белка), усиливают воспалительные процессы (повышают «воспалительный потенциал» организма, т. е. способность отвечать на определенной силы раздражения воспалительной реакцией определенной интенсивности), в больших дозах вызывают гипертонию, склероз почек, артриты, отеки. 94 Глюкокортикоиды, к которым относится гидрокортизон и кортизон, в меньшей степени влияют на минеральный обмен и в большей — на углеводный и белковый обмены [Харкевич Д.А., 2006.]. Они вызывают усиление катаболизма (повышают распад белков и усиливают образование глюкозы). Глюкокортикоиды обладают противовоспалительным действием (снижают «воспалительный потенциал»), уменьшают количество лимфоцитов и эозинофилов в крови и вызывают инволюцию вилочковой железы и лимфатической системы, а также снижают резистентность организма к инфекциям. Со стороны нервной системы они вызывают эйфорию и бессонницу. Очень большое значение в медицине приобрели противовоспалительные кортикостероиды — кортизон (Δ4-прегнен-17α,21-диол-3,11,20-трион) и гидрокортизон (Δ4-прегнен-11β,17α,21-триол-3,20-дион). Наряду с указанными веществами применяются и их синтетические аналоги, многие из которых оказались значительно более активными и вызывающими меньше побочных явлений, чем естественные гормоны. К таким веществам относятся преднизон (Δ1дегидрокортизон) и преднизолон (Δ1-дегидрогидрокортизон), в 5 раз более активные по сравнению с естественными гормонами. Еще более активным оказался 9α-фторпреднизолон, в 15 раз превосходящий по активности кортизон. Наиболее активным из известных в настоящее время препаратов является 9αфтор-16α-метилпреднизолон (дексаметазон, декадрон), противовоспалительная активность которого в 190 раз выше, чем кортизона [Харкевич Д.А., 2006.]. Кортизон и его аналоги находят широкое применение в медицине как средства, снижающие патологически повышенную реактивность, в частности, как мощные противовоспалительные агенты. Эти препараты применяют при ревматизме, инфекционном полиартрите, бронхиальной астме, красной волчанке и при многих других кожных заболеваниях, а также заболеваниях глаз и т. п. Важным является применение кортизона и его аналогов при остром лейкозе, когда они в комбинации со специфическими средствами дают длительные ремиссии. 95 Следует заметить, что применение кортизона может сопровождаться целым рядом побочных явлений. К ним относятся появление отеков, отложение жира в подкожной клетчатке лица (лунообразное лицо), шеи, затылка и области ключиц, гипергликемия и другие нарушения обмена, возбуждение, бессонница, функциональные психоневрозы, половые расстройства, развитие хрупкости костей (остеопороз), наклонность к развитию язвы желудка, нарушение роста в молодом возрасте. Все эти явления проходят при отмене препарата [Харкевич Д.А., 2006.]. Таким образом, суммируя вышеизложенное, можно высказать предложение о двух основных механизмах действия гормоноподобных веществ: 1 – взаимодействие вещества с рецептором, приводящее к его активированию или блокированию, сопровождающегося дальнейшим изменением (усилением или ослаблением) внутриклеточной передачи сигнала, 2 – изменение активности ферментов, участвующих в метаболизме (синтезе или обезвреживании) природных гормонов. Для подтверждения первого механизма действия гормоноподобных веществ необходимо познакомится со структурой рецепторов половых и корковых гормонов. Исследования последних лет показали, что андрогеновые рецепторы (АР) принадлежат к суперсемейству ядерных рецепторов (ЯР), транксрипционные факторы которых регулируют транскрипцию генов при связывании лиганда. У ядерных рецепторов выделяют следующие общие черты организации: вариабельный функциональный домен активации (ФДА-1), ДНК-связывающий домен (ДСД), шарнирный участок, отвечающий за ядерную сигнализацию, Сконцевой лиганд-связывающий домен (ЛСД), включающий 12 спиральных структур, которые замыкают центральный лиганд-связывающий карман (ЛСК), а также второй функциональный домен активации (ФДА-2), который локализован у 96 С-конца лиганд-связывающего домена и является посредником лиганд-зависимой трансактивации [Caboni L. et al., 2012.]. Андрогеновые рецепторы активируются эндогенным гормоном тестостероном и его более мощным метаболитом дигидрокситестостероном (ДГТ) за счет взаимодействия с лиганд-связывающим карманом. Связывание этих эндогенных модуляторов вызывает перестройку 12-й спирали в составе рецептора, сопровождающуюся образованием структурированной гидрофобной поверхности, способной связывать агонист и образовывать второй функциональный домен для дальнейшей активации ядерного рецептора. Благодаря образованию второго функционального домена происходит «переключение» и дальнейшая транскрипционная активация ядерного рецептора [Caboni L. et al., 2012.]. Недавно открыта группа диарилгидразидов – антиандрогенных соединений нестероидной природы, не взаимодействующих с лиганд-связывающим карманом андрогенового рецептора, но способных вытеснять природные коактиваторы из этих рецепторов: Соединение 1 (MDG173) Соединение 2 (MDG15) Благодаря способности диарилгидразидов оказывать такое антиандрогенное действие, они являются потенциальными противоопухолевыми препаратами для лечения рака простаты. Авторы создали трехмерную фармакофорную модель, содержащую общие черты строения между коактиваторами андрогеновых 97 рецепторов и двумя ароматическими фрагментами диарилгидразидов, которая представлена на рисунке 6: Рис. 6. Виртуальный скрининг и идентификация диарилгидразидных моделей. А. Использовался ряд коактиваторных пептидов, сокристаллизованных в бороздке второго функционального домена активации (изображены темносерым цветом), на рисунке представлен коактиваторный фрагмент FxxLF из PDB (базы данных белков). Показаны только остатки Лиз-720 и Глу- 897. В. Трехмерная фармакофорная модель содержала общие черты строения между коактиваторами андрогеновых рецепторов и двумя ароматическими фрагментами. Фармокофоры использовали скрининговую базу данных соединений и оценку предполагаемого взаимодействия с участком второго функционального домена активации. C, D. Соединения 1 (MDG173) и 2 (MDG15) в оптимизированной форме совмещали аминокислотами. в функциональном домене активации с ключевыми 98 Согласно полученным результатам [Caboni L. et al., 2012], диарилгидразиды демонстрируют полный антагонизм к андрогеновым рецепторам, селективность к эстрогеновым, глюкокортикоидным рецепторам α и β и частичный антагонизм к рецепторам прогестерона. Обращает на себя внимание сходство в строении диарилгидразидов и синтетических веществ, обладающих эстрогенной активностью (диэтилстильбэстрол, диенэстрол, синэстрол, бензэстрол), описанных выше, а именно наличие фенильных колец и других ароматических заместителей. Исследования, проведенные за последние 5 лет, позволили детально изучить строение глюкокортикоидных рецепторов и открыть потенциальные агонисты и антагонисты для них. Совсем недавно был открыт флутиказон фуроат (соединение 1), который очень эффективно за одно применение устранял симптомы сезонного и постоянного аллергического ринитов [Biggadikea K. et al., 2009]: Соединение 1 (Флутиказон фуроат). Ранее сообщалось о рентгено-структурных моделях лиганд-связывающего домена глюкокортикоидного рецептора, способного связывать кортикостероидные агонисты дексаметазон, флютиказон пропионат и флютиказон фуроат. Белковая структура лиганд-связывающего домена глюкокортикоидного рецептора рыбы гардемарина [Arterbery A.S. et al., 2011] представлена на рисунке 7. Флютиказон фуроат полностью подходит к рецептору с частично видимым эфиром фуроата, аккуратно занимая липофильный карман 17-й α-спирали на левой стороне и 99 образуя водородные связи (Арг-611 и Глн-570) с кето-группой А-кольца, определяя окончание связывающего кармана на правой стороне (рис. 7А.). А. В. Рис. 7. Белковая структура лиганд-связывающего домена глюкокортикоидного рецептора рыбы гардемарина. А. Модель полной гомологии лиганд-связывающего домена глюкокортикоидного рецептора 1 при связывании кортизола (красный). В. Позиция 204 обнаружена в лиганд-связывающем участке на расстоянии 4 Ǻ от предполагаемого связанного лиганда. Однако особый интерес соединений нестероидной природы представляет потенциальная возможность взаимодействовать с лиганд-связывающим доменом глюкокортикоидного рецептора. В последние годы уделяется повышенный интерес новым нестероидным лигандам глюкокортикоидного рецептора [Schäcke H. et al., 2008; Hudson A.R. et 100 al., 2008; Mohler M.L. et al., 2008]. Эти исследования поднимают на новый уровень изучение глюкокортикоидов, которые проявляют трансрепрессивную активность, связанную с противовоспалительным действием через трансактивацию нежелательных эффектов ГК. Авторы [Biggadikea K. et al., 2009] недавно сообщили о группе нестероидных агонистов глюкокортикоидных рецепторов, внедрив в лигандсвязывающий домен аминопиразольный фрагмент (соединение 2) и показав чрезвычайную селективность к глюкокортикоидному рецептору через другие рецепторы стероидных гормонов: Соединение 2 Ограничение амида аминопиразола соединения 3 увеличивает его способность быть агонистом к глюкокортикоидному рецептору: Соединение 3 На рисунке 8 приведена кристаллическая структура лиганд-связывающего домена глюкокортикоидного рецептора для ряда синтетических агонистов. 101 Рис. 8. А. Кристаллическая структура лиганд-связывающего домена глюкокортикоидного рецептора для флютиказон фуроата (соединение 1); В. кристаллическая структура лиганд-связывающего домена глюкокортикоидного рецептора для деацетилкортивазола (соединение 5); С. Модель индазол (соединение 3)-глюкокортикоидного рецептора, показывающая доступ к новому «мета»-каналу (показан красной стрелкой); D. Аланинамидный фрагмент (соединение 6), располагающийся внутри «мета»канала. Попытки приспособить эти производные пиразола и индазола в лигандсвязывающий карман глюкокортикоидного рецептора оказались неудачными. Однако эти производные имеют структурные сходства со стероидным агонистом ГК кортивазолом (соединение 4) и хорошо подходят к модели связывающего участка ГР, который был создан для этого удлиненного кортикостероида: 102 Соединение 4 (кортивазол) Для размещения кортивазола в лиганд-связывающем домене необходимы два аминокислотных остатка (Арг-611 и Глн-570), благодаря которым происходит существенное перемещение арилпиразольного фрагмента от мета-позиции фенильного кольца и его встраивание в новый канал (new channel) рецептора. Это расширение лиганд-связывающего домена и формирование «мета»канала также подтверждено кристаллической структурой с низким внутренним разрешением для аналога кортивазола деацетилкортивазола (соединение 5): Соединение 5 (деацетилкортивазол) При появлении деацетилкортивазола, фенильного кольца пиразола в структуре лиганд-связывающий домен вынужден открыться в его направлении и создать новый канал, прилегающий к N-фенильной группе (рис. 8В). На основании опытов in vitro авторы установили, что соединение 3 показывает хорошее связывание с ГР и демонстрирует мощную активность агониста ГР в опытах трансактивации и трансрепрессии in vitro, а также показывает хорошую селективность к рецептору прогестерона. 103 Соединение 8, D-аланинамид, обладало в 10 раз большей активностью, чем незамещенный аналог 3 в исследовании трансрепрессии ядерного фактора (NFkB). Вместе с аналогом 3 и обычными ГК, соединение 8 демонстрирует полную эффективность (~106% ответ относительно стандартного дексаметазона, 100%) в этом исследовании трансрепрессии и в исследовании трансактивации вируса рака груди у мышей (130% ответ): Эта поразительная способность делает нестероидный агонист 8 сравнимым с наиболее эффективными ГК, такими как флютиказон фуроат (соединение 1). Интересно, что введение аланинамидного заместителя увеличивает активность агониста прогестеронового рецептора (ПР) и соединение 8 демонстрирует низкую селективность к ГР/ПР относительно соединения 3, лишенного этого заместителя. Это увеличение ПР-активности на введение мета-амидного заместителя является примечательным наблюдением и показывает, что сходный «мета»-канал также должен открываться и в ПР. Чтобы доказать эту гипотезу авторы [Biggadikea K. et al., 2009] изучили урезанные соединения, в которых сохранялся D-аланинамидный фрагмент. Одно такое соединение - бис-трифтор-метиловый аналог (соединение 11) показало в опытах глюкокортикоидный in vitro препарат активность, преднизолон превосходящую и полностью главный сохранило трансрепрессивную и трансактивационную активности. Аланинамидный радикал соединения 11 образует водородные связи также как и полиамид 9. Таким образом, водородные связи образуются с участием Лиз- 104 667 через водяной мостик и карбонильный остаток Глн-570. Водородная связь также образуется между ОН-группой и Асн-564: Соединение 12 сохраняет активность агониста ГКР, соответствующей средней активности между соединениями 8 и 9. Несмотря на очень разные фармакологические профили, соединения 9 и 10 индуцировали прирост пептидного участка ядерного рецептора, сходного с таковым для классического стероида флютиказона пропионата. Основываясь на этом, авторы сообщают об открытии новых мощных нестероидных лигандов глюкокортикоидного рецептора на основе индазола, который занимает ранее неизвестный участок рецептора [Biggadikea K. et al., 2009]. Лиганд-связывающий домен прогестеронового рецептора (ПР) имеет свои особенности [Petit-Topin I. et al., 2009] (рисунок 9). Он содержит 11 α-спиралей (H1, H3-H12) и 2 короткие β-складки, уложенные в 3 слоя. Внутри лигандсвязывающей полости остатки Глн-725 и Арг-766 (рис. 9 В) закрепляют С3-кетогруппу прогестерона и его аналогов водородными связями и контактируют с 105 молекулой воды. Остаток Асн-719 связан водородной связью с 17-β-гидроксигруппой левоноргестрела через молекулу воды (рис. 9 В). Рис. 9. Кристаллическая структура лигандсвязывающего домена прогестеронового рецептора в комплексе с левоноргестрелом. А. Диаграмма, показывающая взаимодействия между лиганд-связывающим доменом прогестеронового рецептора и левоноргестрелом. Водородные связи обозначены как непрерывные черные стрелки, а ван дер Ваальсовые взаимодействия - как пунктирные черные линии соответственно. W – обозначает молекулу воды. В. Структура лигандсвязывающего домена прогестеронового рецептора, связанная с левоноргестрелом (серый), наложенным на прогестерон (голубой). В структуре лиганд-связывающего домена ПР 19-метильная группа прогестерона контактирует с остатком Мет-759. Вследствие отсутствия 19 метильной группы в молекуле левоноргестрела остаток Мет759 участвует в образовании некоторых связей со стероидным скелетом. Благодаря 13-этильной группе левоноргестрела, Мет-909 (Н12 спираль) связывает ротамер отличный от такового у прогестерона и норетиндрона. Следовательно, в присутствии левоноргестрела Мет-909 образует связи с Гли-722 и Лей-726 (Н3 спираль) и с Мет-759 (Н5 спираль) и образует только одну связь с 13-мельной группой прогестерона. Остатки Асн-719, Мет759 и Мет-909 могут внести критический 106 вклад в высокую левоноргестрел-индуцированную активацию ПР. Важно заметить, что, остаток Мет-909 (спираль 12) играет ключевую роль в активации ПР как тестостероном так и прогестероном особенности [Petit-Topin I. et. al. 2009]. Несколько карману аминокислотных гидрофобность и остатков придают стабилизируют лиганд-связывающему положение левоноргестрела посредством ряда ван дер Ваальсовых взаимодействий. Наложение структуры лиганд-связывающего домена ПР с левоноргестрелом поверх лиганд-связывающего домена ПР с прогестероном показывает главные различия во взаимодействиях стероидов с лиганд-связывающим доменом. И, наконец, второй возможный механизм действия гормоноподобных веществ заключается в изменении активности ферментов (путем активации или ингибирования), участвующих в метаболизме (синтезе или обезвреживании) природных гормонов. Одним из ферментов, участвующих в метаболизме глюкокортикоидов является β-гидроксистероид-дегидрогеназа 1 типа, которая превращает кортизон в свою активную форму. Результаты, полученные в опытах на мышах линии KKAy с модельным сахарным диабетом 2 типа, показали, что препарат BVT116429, возможно, ингибировал фермент β-гидроксистероид-дегидрогеназу 1 типа [Sundbom M., et al., 2008]. Многочисленные исследования подтверждают, что ГК уменьшают уровень адипонектина у животных и людей. Поэтому можно предположить, что увеличение концентрации адипонектина, обнаруженное у мышей леченых препаратом BVT116429, есть следствие снижения концентрации глюкокортикоидов в результате ингибирования их синтеза данным препаратом [Sundbom M. et al., 2008]. Таким образом, учитывая разработки последних пяти лет, можно сделать вывод о возможности взаимодействия различных по строению нестероидных соединений с лиганд-связывающими доменами рецепторов стероидных гормонов 107 (андрогенов, эстрогенов и глюкокортикоидов). Дальнейшие исследования в этой области помогут нестероидной выявить природы, потенциальные обладающие гормоноподобные анаболическим, вещества андроген/эстроген- модулирующим, противовоспалительным и др. действиями, что позволит правильно оценить их биологическую активность и корректно использовать в медицине, ветеринарии, животноводстве, птицеводстве и других областях народного хозяйства. Например, применяемый в настоящее время в ветеринарии селеноорганический препарат диацетофенонилселенид (ДАФС-25) способствует увеличению яичной и мясной продукции сельскохозяйственных животных и птиц. Подобный анаболический эффект, вероятно, обусловлен особенностями строения этого соединения и его способностью взаимодействовать с рецепторами стероидных гормонов. Однако ДАФС-25 наряду с другими селеноорганическими соединениями обладает также антиоксидантной и антитоксической активностью. Вместе с тем в современной научной литературе отсутствуют сведения о сравнительной биологической активности органических соединений серы, селена и теллура с учетом их свойств. Настоящее исследование ставит своей целью изучить биологическую активность халькогенорганических соединений в зависимости от их структуры, включая наличие гетероатома халькогена (серы, селена, теллура) и боковых заместителей (атомов хлора, фтора и нитрогрупп) в составе этих соединений. 108 Глава 2. Материалы и методы исследования 2.1. Халькогеноорганические препараты, использованные в исследовании Исследуемые соединения впервые были синтезированы в Саратовском военном институте биологической руководством д.х.н., и химической безопасности под профессора Б.И. Древко. Соединения являются структурными аналогами (халькогенсодержащие арилалифатические дикетоны). Структурные формулы препаратов представлены на рисунке 10. 1,5-дифенил-3-селенапентандион-1,5 (ДАФС-25, ДАФС(Se), соединение 1) 1,5-ди-(п-фторфенил)-3-селенапентандион-1,5 (ДАФС(F2), соединение 3) 1,5-ди-(п-хлорфенил)-3- селенапентандион-1,5 (ДАФС(Cl2), соединение 2) 1,5-ди-(м-нитрофенил)-3-селенапентан дион-1,5 (ДАФС(NO2)2, соединение 4) 1,5-дифенил-3-тиапентандион-1,5 1,5-дифенил-3-теллуропентандион-1,5 (ДАФС(S), соединение 5) (ДАФС(Те), соединение 6) Рис. 10. Химическое строение халькогенорганических соединений (1-6). 109 2.2. Постановка эксперимента Исследования были выполнены на белых беспородных крысах и мышах. Контрольные и экспериментальные группы формировали из самцов. Возраст животных составлял 3-4 месяца. Масса мышей 203 г и крыс 20025 г. Всего было составлено 12 контрольных групп и 29 экспериментальных групп. Каждая контрольная группа включала по 12 животных, а экспериментальные по 10 особей. Общее количество животных, использованных в работе, составило 460 особей (таблица 3). Таблица 3. Дизайн эксперимента Серии эксперимента 1 2 Экспериментальные исследования Антитоксическое действие при подостром отравлении солями тяжелых металлов (Pb(NO3)2, Hg(NO3)2, CdSO4) Антиоксидантная активность Антимикробная активность 3 Количество особей 300 крыс 80 мышей Клинические штаммы Staphylococcus aureus (n=60), Escherichia coli (n=60), Pseudomonas aeruginosa Определение показателей углеводного обмена 4 80 мышей Гормоноподобное действие: Квантово-химические расчеты проводили гидридным методом теории функционала плотности (DFT) Расчеты вероятной биологической активности PASS 2.2.1. Подготовка препаратов к исследованиям Для изучения биологической активности халькогенорганических соединений экспериментальным животным вводили per os растворы препаратов в растительном масле на протяжении 14 дней. Суточную дозу препаратов 110 рассчитывали из соотношения 200 мкг Se/кг живого веса [Всемирная организация здравоохранения. Женева, 1989]. В таблице 4 приведены дозы препаратов в расчете на среднюю массу экспериментальных животных (крыс и мышей). Таблица 4. Схема использования препаратов в исследованиях in vivo. Характеристика препаратов Исследование антитоксического действия при подостром отравлении солями тяжелых металлов (Pb(NO3)2, Hg(NO3)2, CdSO4) суточная доза препарата для крыс M, г/моль № препарат халькоген Исследование антиоксидантной активности и показателей углеводного обмена суточная доза препарата для мышей мкг/кг мкг/кг мкг/200 г мкг/20 г Se (S, Те) препарат препарат препарат I 317 Se=79 200 800 160 16 II 385 Se=79 200 975 195 19,5 III 353 Se=79 200 900 180 18 IV 407 Se=79 200 1000 200 20 V* 270 S=32 80 680 136 13,6 VI * 436 Те=128 320 1088 218 21,8 * количество вещества атомарной S в препарате V и Те в препарате VI эквивалентно содержанию Se в препаратах I, II, III и IV (S(Те)=Se). Растворы препаратов в растительном масле готовили ex tempore при нагревании до 60ºС на водяной бане из расчета суточных доз, указанных в таблице 4. Мышам вводили per os суточную дозу препарата утром натощак в объеме 10 мкл, крысам - в объеме 50 мкл с помощью микродозатора («Gilson», Франция). Все работы с лабораторными животными проводили согласно принципам гуманного отношения к животным в соответствии с «Международными рекомендациями по проведению медико-биологических исследований с использованием животных», «Правилами проведения работ с использованием 111 экспериментальных животных» и «Правилами лабораторной практики в Российской Федерации» (приказ МЗ РФ №267 от 19.06.2003). 2.2.2. Подготовка биологического материала Приготовление гемолизата эритроцитов и супернатанта плазмы крови Забор крови проводили из хвостовой вены в объеме 2,0 мл. В качестве антикоагулянта использовали раствор гепарина (0,6 мл гепарина в 12 мл 0,9% NаCl) в соотношении стабилизатор/кровь 1:5. С помощью центрифугирования проводили отделение плазмы крови от форменных элементов. Форменные элементы крови суспендировали в десятикратном объеме физиологического раствора. Затем проводили центрифугирование при 3000 об/мин в течение 15 минут. Надосадочную жидкость удаляли. Процедуру повторяли трижды. Для приготовления гемолизата эритроцитов к 50 мкл отмытых эритроцитов добавляли 10 мл дистиллированной воды (разведение 1:200) [Карпищенко А.И., 1999; Артюхов В.Г., Наквасина М.А., 2000.]. Супернатант плазмы крови готовили путем добавления к 50 мкл плазмы 10 мл физиологического раствора (0,9% NаCl) (разведение 1:200). В цельной крови определяли гематологические показатели (содержание гемоглобина, количество эритроциты, тромбоцитов, лейкоцитов, лейкоцитарную формулу и гематокрит), а также актиность фермента глутатионпероксидазы. В плазме крови проводили определение биохимических показателей (активность γ-глутамилтрансферазы, щелочной фосфатазы, аминотрансфераз, лактатдегидрогеназы, креатинкиназы, α-амилазы, содержание мочевины, креатинина, глюкозы, общего белка, альбумина и общего холестерина). В супернатанте плазмы крови и гемолизате эритроцитов определяли показатели свободнорадикального окисления липидов (концентрацию диеновых 112 конъюгатов и малонового диальдегида) и антиоксидантного статуса (активность супероксиддисмутазы и каталазы). Подготовка гомогенатов тканей В исследованиях были использованы гомогенаты мозга, печени, почек и легких мышей. Ткани отмывали от крови в физиологическом растворе, обсушивали фильтровальной бумагой и проводили взвешивание на аналитических весах с точностью до 0,0001 г. Навески тканей механически гомогенизировали с применением в качестве абразива стеклянной крошки. Гомогенизацию проводили в течение 1 минуты в фарфоровой ступке при 0°С до получения однородной массы [Кирова Ю.И., 2004]. Объем полученного гомогената доводили до 5,0 мл 0,01 М фосфатного буфера (рН=7,4). В гомогенатах тканей определяли показатели свободнорадикального окисления липидов (концентрацию диеновых конъюгатов и малонового диальдегида) и антиоксидантного статуса (активность супероксиддисмутазы и каталазы). 2.2.3. Изучение свободнорадикального окисления липидов и антиоксидантного статуса в тканях мышей Животных (белых беспородных мышей, самцов) разделили на 6 групп: 1 контрольную и 5 экспериментальных. Мышам контрольной группы (n=12) вводили per os 10 мкл растительного масла, а мышам экспериментальных групп (n=10) вводили per os 10 мкл раствора препаратов 1-4 и 6 соответственно. Эксперимент проводили в течение 14 дней. В гемолизате эритроцитов, плазме крови, гомогенатах тканей (мозга, печени, почек и легких) определяли содержание диеновых конъюгатов, малонового диальдегида, активности супероксиддисмутады и каталазы; в цельной крови проводилось определение активности глутатионпероксидазы. 113 2.2.4. Изучение антитоксического действия халькогенорганических препаратов при подостром отравлении солями тяжелых металлов Токсичность изучаемых солей тяжелых металлов (Pb(NO3)2, Hg(NO3)2, CdSO4) исследовалась в сравнительном плане путем учета степени и характера их действия на организм экспериментальных животных (белые беспородные крысы) в подострых экспериментах. Дозировки тяжелых металлов расчитывались с учетом полулетальных доз (LD50), представленных в таблице 5 [Филов В.А., 1988]. Таблица 5. Расчет суточной дозы солей тяжелых металлов Соли тяжелых LD50 (per os для крыс), суточная доза для металлов мг/кг крыс, мг (в 50 мкл) Hg(NO3)2 26 5,2 Pb(NO3)2 93 18,6 CdSO4 47 9.4 В данной серии экспериментов животных разделили на 10 контрольных и 18 экспериментальных групп (таблицы 6 и 7). Таблица 6. Характеристика контрольных групп животных Группа Количество крыс Условия эксперимента 1 12 50 мкл растительного масла 2 12 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 3 12 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 4 12 50 мкл раствора CdSO4 с дозой 47 мг/кг 5 12 50 мкл соединения 1 с дозой 800 мкг/кг 6 12 50 мкл соединения 2 с дозой 975 мкг/кг 7 12 50 мкл соединения 3 с дозой 900 мкг/кг 8 12 50 мкл соединения 4 с дозой 1000 мкг/кг 9 12 50 мкл соединения 5 с дозой 680 мкг/кг 10 12 50 кл соединения 6 с дозой 1088 мкг/кг 114 Таблица 7. Характеристика экспериментальных групп животных Группа* 1 Условия эксперимента Через час 50 мкл соединения 1 с 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг дозой 800 мкг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 2 50 мкл раствора CdSO4 с дозой 47 мг/кг 3 4 5 50 мкл соединения 2 с дозой 975 мкг/кг 6 7 8 50 мкл соединения 3 с дозой 900 мкг/кг 9 10 11 50 мкл соединения 4 с дозой 1000 мкг/кг 12 13 14 50 мкл соединения 5 с дозой 680 мкг/кг 15 16 17 50 мкл соединения 6 с дозой 1088 мкг/кг 18 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 50 мкл раствора CdSO4 с дозой 47 мг/кг 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 50 мкл раствора CdSO4 с дозой 47 мг/кг 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 50 мкл раствора CdSO4 с дозой 47 мг/кг 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 50 мкл раствора CdSO4 с дозой 47 мг/кг 50 мкл раствора Hg(NO3)2 с дозой 26 мг/кг 50 мкл раствора Pb(NO3)2 с дозой 93 мг/кг 50 мкл раствора CdSO4 с дозой 47 мг/кг * - количество крыс в каждой экспериментальной группе 10 Исследуемые вещества вводились перорально с помощью с помощью микродозатора («Gilson», Франция) утром в течение 14 дней. Оценка состояния здоровья животных производилась с использованием комплекса тестов, интегрально отражающих функциональное состояние организма: поведение, общее состояние животных, динамика массы тела, 115 количество эритроцитов, тромбоцитов, лейкоцитов, расчет лейкоцитарных индексов интоксикации, содержание гемоглобина, гематокрит, активность ферментов щелочной (-глутамилтрансферазы, фосфатазы, аланинаминотрансферазы, аспартатаминотрансферазы, лактатдегидрогеназы, амилазы), белково-образующая функция печени (альбумин, общий белок), состояние мочевыделительной системы (креатинин, мочевина), концентрация глюкозы как главного метаболита углеводного обмена и холестерина как метаболита липидного обмена. Антитоксическое действие халькогенорганических препаратов оценивалось по нормализации всех параметров токсичности. 2.2.5. Изучение антибактериальной активности халькогенорганических соединений Определение антибактериального действия халькогенорганических соединений на бактериальные клетки клинических штаммов кишечной палочки (Escherichia coli), синегнойной палочки (Pseudomonas aeruginosa) и золотистого стафилококка (Staphylococcus aureus) проводили в бактериологической лаборатории Саратовского научно-исследовательского института травматологии и ортопедии. В исследовании использовали антибиотикорезистентные штаммы микроорганизмов (n=180), выделенные из раневого отделяемого пациентов с посттравматическими и послеоперационными осложнениями, находившихся на лечении в ФГБУ «Саратовский научно-исследовательский институт травматологии и ортопедии» Минздрава России: Escherichia coli (n=60), Staphylococcus aureus (n=60), Pseudomonas aeruginosa (n=60). Микроорганизмы идентифицировали на микробиологическом анализаторе BD BBL™ Crystal™ AutoReader (Becton Dickinson, США) с применением панелей CrystalTM 116 Enteric/Nonfermenter ID Kit (Becton Dickinson, США), CrystalTM Gram-Positive ID Kit (Becton Dickinson, США). Изучаемые штаммы микроорганизмов тестировали в отношении профильных антибиотиков в соответствии с МУК 4.2.1890-04 «Определение чувствительности микроорганизмов к антибактериальным препаратам». В исследовании использовали питательную среду Mueller-Hinton-Agar (Becton Dickinson, США) и сенси-диски с антибиотиками (Becton Dickinson, США). Для подтверждения метициллинрезистентности штаммов S. aureus использовали набор «MeReSa Agar Base, MRSA AlertTM» (Hi Media, Индия). Антибактериальную активность халькогенорганических соединений оценивали по снижению количества бактериальных колоний, выросших на твердой питательной среде после инкубации с растворами исследованых соединений в концентрациях 0,0001-1 мг/мл в течение 30, 60, 90, 120 и 150 минут. 2.2.6. Изучение гормоноподобного действия халькогенорганических соединений Эксперимент проводили на белых беспородных мышах (самцах). Контрольной группе мышей (n=12) вводили per os растительное масло в объеме 10 мкл. Пяти экспериментальным группам мышей вводили per os халькогенорганические соединения 1-4 и 6 с дозами, описанными в таблице 4 и в объеме 10 мкл. Шестой экспериментальной группе мышей вводили per os 8 мкл раствора преднизолона в концентрации 0,5 мг/кг. Эта доза преднизолона применяется в ветеринарии для подавления воспаления у мелких домашних животных [Пульняшенко П.Р., 2004]. Экспериментальные группы животных включали 10 мышей. Эксперимент проводили в течение 14 дней. Для исследования использовали цельную кровь, плазму крови и печень мышей. Приготовление гомогената печени. Навеску печени отмывали в ледяном физиологическом растворе (0,9 % NaCl), высушивали фильтровальной бумагой и 117 растирали в ледяной ступке с тремя объемами 0,25М сахарозы в течение 1 минуты, затем добавляли еще 7 объемов сахарозы. После перемешивания гомогената, проводили центрифугирование в течение 10 минут при 3000 оборотах в минуту. Для изучения гормоноподобного действия халькогенорганических соединений проводили определение некоторых показателей углеводного обмена: концентрацию глюкозы, пирувата, активность ферментов аланинаминотрансферазы (АлАТ) и аспартатаминотрансферазы (АсАТ) в плазме крови, концентрацию гликогена в печени и цельной крови, а также активность ферментов глюконеогенеза глюкозо-6-фосфатазы и фруктозо-1,6-бисфосфатазы в печени. 2.3. Методы исследования 2.3.1. Методы изучения свободнорадикального окисления липидов и антиоксидантной системы Определение содержания малонового диальдегида При 100°С в кислой среде малоновый диальдегид реагирует с 2тиобарбитуровой кислотой, образуя окрашенный триметиновый комплекс, с максимумом поглощения при 532 нм. Молярный коэффициент экстинкции этого комплекса - = 1,56·105см-1·М-1 [Стальная, Гаришвили, 1977]. Подготовленный биологический материал (гемолизат эритроцитов, супернатант плазмы крови, гомогенаты мозга, печени, почек и легких) в буферном растворе по 2,0 мл помещают в центрифужные пробирки и осаждают белок добавлением 1 мл 17% раствора трихлоруксусной кислоты. Образующийся осадок отделяют центрифугированием в течение 10 минут при 400 g (центрифуга ЦУМ – 1). Надосадочную жидкость по 2 мл переносят в 118 пробирки, добавляют по 1 мл 0,8% раствора тиобарбитуровой кислоты и помещают пробы на 10 минут в кипящую водяную баню. В качестве контроля используют пробы, содержащие вместо надосадочной жидкости буферный раствор. После развития розовой окраски пробы охлаждают до комнатной температуры и измеряют оптическую плотность при 532 нм на фотоэлектроколиримерта КФК-3 (Россия). Количество малонового диальдегида рассчитывают, используя указанную выше величину молярного коэффициента экстинкции, и полученный результат выражают в ммоль/л плазмы крови и гемолизата эритроцитов или в ммоль/г тканей (мозга, печени, почек, легких). Определение содержания диеновых коньюгатов В ходе перекисного окисления на стадии образования свободных радикалов в молекулах полиненасыщенных высших жирных кислот возникает система сопряженных двойных связей, что сопровождается появлением нового максимума в спектре поглощения - = 233 нм, = 2,210-5М-1см-1 [Стальная, 1977]. Подготовленный биологический материал (гемолизат эритроцитов, супернатант плазмы крови, гомогенаты мозга, печени, почек и легких) в буферном растворе по 2,0 мл помещают в центрифужные пробирки, добавляют 4 мл гептанизопропаноловой смеси и встряхивают 10 минут. Затем в пробу добавляют 1 мл раствора HCL и 2,0 мл гептана, интенсивно встряхивают. После центрифугирования в течение 10 минут при 400 g (центрифуга ЦУМ – 1), отбирают гептановый слой, измеряют оптическую плотность при 233 нм в кварцевых кюветах против контрольной пробы на спектрофотометре «SPECORD UV – VIS» (Германия). В качестве контроля используют пробы, содержащие вместо биологического материала буферный раствор. Содержание диеновых коньюгатов в пробе расчитывают, используя величину молярного коэффициента экстинкции при 233 нм для сопряженных 119 диенов полиненасыщенных высших жирных кислот, равного 2,2105см-1М-1. Полученный результат выражают в ммоль/л плазмы крови и гемолизата эритроцитов или в ммоль/г тканей (мозга, печени, почек, легких) Определение активности глутатионпероксидазы Глутатионпероксидаза катализирует окисление восстановленного глутатиона гидропероксидом кумина. В присутствии глутатионредуктазы и НАДФН происходит превращение окисленного глутатиона в восстановленный с одновременным окислением НАДФН в НАДФ+. Уменьшение оптической плотности определяют при длине волны 340 нм [Paglia D.E. and Valentine W.N.,1967]. Определение активности глутатионпероксидазы в цельной крови проводят с использованием наборов реактивов фирмы “Randox” (Великобритания): реагент 1 – глутатион (4 ммоль/л), глутатионредуктаза (≥0,5 ед/л), НАДФН (0,34 ммоль/л), фосфатный буфер (0,05 моль/л, рН 7,2), ЭДТА (4,3 ммоль/л), реагент 2 – гидропероксид кумина (0,18 ммоль/л), реагент 3 – растворитель. Для определения соедржания гемоглобина использовали гемоглобиновый реагент, который содержал: фосфат калия (1,03 ммоль/л), ферроцианид калия (0,61 ммоль/л), цианид калия (0,77 ммоль/л). В пробирку к 1 мл реагента 3 (растворителя) добавить 50 мкл гепаринизированной крови и после 5 минут инкубации добавить 1 мл гемоглобинового реагента (разведенного дистилированной водой в отношении 1:4). 20 мкл раствора из этой пробирки использовать для определения. Не позднее, чем через 20 минут провести определение активности глутатионпероксидазы. Для этого в кювету анализатора поместить 1 мл реагента 1 (должен быть заранее прогрет при 37ºС), добавить 40 мкл реагента 2 и 20 мкл «подготовленной» крови из пробирки. 120 Определение проводить на полуавтоматическом анализаторе “Hospitex screen master plus” (Швейцария) с использование заданной программы. Активность глутатионпероксидазы выражали в единицах на грамм гемоглобина (ед/г Hb). Определение активности супероксиддисмутазы СОД осуществляет катализ реакции рекомбинации супероксидных анионов (О-2). В результате реакции образуется перекись водорода (Н2О2) и кислород (О2). Принцип определения активности энзима основан на реакции восстановления формазанов из солей тетразолия в слабощелочной среде. Об активности СОД судили по степени торможения восстановления НСТ в присутствии НАДН и ФМС [Дубинина Е.Е. и др., 1983]. Готовят контрольную пробу – 3 мл 0,15 М фосфатного буфера (рН 7,8); стандартную пробу – 1 мл 0,15 М фосфатного буфера (рН 7,8) 2 мл реагента N1; опытную пробу – 1 мл 0,15 М фосфатного буфера (рН 7,8), добавляют 0,05 мл супернатанта гемолизата эритроцитов или плазмы крови. Общий объем каждой пробы – 3 мл. После перемешивания пробы помещаются в кюветы с l =1,0 см. На фотоэлектроколиримерта КФК-3 (Россия) при длине волны 540 нм измеряются экстинции в обеих пробах против контрольной пробы. Затем добавляют 0,1 мл реагента N2, содержимое кюветы перемешивают и после инкубации в течение 10 мин при Т=37°С производят повторное измерение экстинциий опытной и стандартной проб. Активность СОД выражают в условных единицах. За единицу активности принимается количество фермента, необходимое для снижения оптической плотности в процессе восстановления НСТ в опытной пробе на 50%. Активность энзима определяли в пересчете на 1 мл гемолизаты эритроцитов и плазмы крови или на 1 г ткани (мозга, печени, почек и легких). 121 Определение активности каталазы Метод определения активности каталазы основан на определении скорости утилизации перекиси водорода в реакционной смеси, в которую вносится биологический материал, содержащий фермент. Об интенсивности утилизации перекиси водорода судят по скорости снижения экстинкции при длине волны 260 нм, при которой перекись водорода имеет максимум светопоглощения [Карпищенко А.И.. 1999]. К 2,5 мл 0,3% перекиси водорода добавляют 200 мкл биологического материала (гемолизата эритроцитов, плазмы или гомогената тканей). Реакцию проводят 10 минут при комнатной температуре, после чего останавливают внесением 0,3 мл 50% раствора ТХУ. Параллельно ставят контрольную пробу, в которую ТХУ вносят до добавления биологического материала. После центрифугирования в течение 10 минут при 3000 оборотах в минуту супернатант опытных и контрольных проб фотометрируют при длине волны 260 нм в кювете (l = 1,0 см) против 5% раствора ТХУ. Измерение оптической плотности контрольной и экспериментальных проб проводили с помощью фотоэлектроколиримерта КФК-3 (Россия). Активность каталазы выражали в ммоль/мин на мл плазмы крови и эритроцитарной взвеси или на 1 г гомогената ткани. 2.3.2. Методы исследования биохимических показателей плазмы крови В работе использовали набор жидких реагентов, готовых к определению показателей в плазме крови. Производитель ЗАО «Диакон» (Россия). Методы являются унифицированными. Активность -глутамилтрансферазы в плазме крови определяли колориметрическим метод по Зейц/Персин (Szasz). Активность фермента выражали в мккат/л. 122 Активность щелочной фосфатазы в плазме крови определяли с помощью кинетического УФ – анализа в соответствии с рекомендациями DGKC (Германское Общество Клинической Химии). Активность фермента выражали в мккат/л. Активность аминотрансфераз и (аланинаминотрансферазы аспартатаминотрансферазы) в плазме крови определяли оптимизированным УФ анализом, разработанным в соответствии с рекомендациями IFCC (Международная Федерация Клинической Химии) без пиридоксальфосфата и без холостой пробы. Активность ферментов выражали в в мккат/л. Активность лактатдегидрогеназы в плазме крови определяли с помощью УФ-метода, оптимизированный в соответствии с требованиями DGKC (Германское Общество Клинической Химии). Активность фермента выражали в мккат/л. Активность креатинкиназы в плазме крови определяли оптимизированного УФ-анализа, разработанного в с помощью соответствии с рекомендациями DGKC (Германское Общество Клинической Химии) и IFCC (Международная Федерация Клинической Химии). Активность фермента выражали в мккат/л. Активность -амилазы в плазме крови определяли колориметрическим ферментативным методом. Активность фермента выражали в мккат/л. Определение концентрации креатинина в плазме крови проводили кинетическим методом без депротеинирования, основанном на цветной реакции Яффе (Jaffe). Концентрацию креатинина выражали в мкмоль/л. Определение концентрации мочевины в плазме крови проводили уреазным методом по конечной точке в плазме крови. Концентрацию мочевины выражали в в ммоль/л. Определение концентрации глюкозы в плазме крови проводили глюкозооксидазным методом. Концентрацию глюкозы выражали в в ммоль/л. 123 Определение концентрации общего белка в плазме крови проводили биуретовым методом. Концентрацию общего белка выражали в г/л. Определение концентрации альбумина в плазме крови проводили колориметрическим методом с применением бромкрезолового зеленого. Концентрацию альбумина выражали в г/л. Определение концентрации общего холестерина в плазме крови проводили энзиматическим колориметрическим методом. Концентрацию общего холестерина выражали в в ммоль/л. В работе использовали набор реагентов для определения концентрации общего холестерина в плазме крови энзиматическим колориметрическим методом. Производитель ЗАО «Витал Диагностикс СПб» (Россия). Определение биохимических показателей крови проводили с помощью полуавтоматического биохимического анализатора «HOSPITEX Screen master», оборудованного термостатом, фотометром и микропроцессором. 2.3.3. Методы исследования гематологических показателей плазмы крови Определение количества гемоглобина Гемоглобин при взаимодействии железосинеродистым калием (красная кровяная соль) окисляется в метгемоглобин, образующий с ацетонциангидрином окрашенный гемоглобинцианид, интенсивность окраски которого пропорциональна содержанию гемоглобина [Зупанец И.А. и др., 2005]. К 20 мкл крови прибавляют 5 мл трансформирующего раствора, хорошо перемешивают, оставляют стоять 20 минут, после чего измеряют на фотоэлектроколориметре при длине волны 500-560 нм в кювете (l=1 см) против трансформирующего раствора. Стандартный раствор колориметрируют без 124 обработки. Измерение оптической плотности проводят с помощью фотоэлектроколиримерта КФК-3 (Россия). Концентрацию гемоглобина выражают в г/л. Определение количества эритроцитов В сухую пробирку помещают 4 мл раствора Гайема (3% раствор хлорида натрия). Капиллярной пипеткой набирают 0,02 мл крови и преносят в пробирку с раствором Гайема. Пипетку тщательно промывают в верхнем слое жидкости. Содержимое пробирки перемешивают. Каплю разведенной крови отбирают концом круглой стеклянной палочки и наносят на край камеры с сеткой Горяева (с притертым до цветных колец Ньютона покровным стеклом). Через одну минуту после заполнения камеры приступают к подсчету эритроцитов. Диафрагма микроскопа должна быть прикрытой, увеличение малым. Подсчет ведется в 5 больших квадратах, расположенных по диагонали, каждый из которых разделен на 16 маленьких. Количество эритроцитов в 1мм 3 (1 мкл) периферической крови, т.е. с учетом разведения в 200 раз, равняется А50200= А10000. Практически количество эритроцитов, подсчитанное в 80 малых квадратах, умножают на 10000 [Зупанец И.А. и др., 2005]. Количественное определение гематокрита Определение гематокрита проводится прямым методом. Общий объем эритроцитов определяется в крови, смешанной с антикоагулянтами (раствор гепарина) [Зупанец И.А. и др., 2005]. Определение проводят в центрифужной пробирке с делениями или капилляре Панченкова. В качестве стандартного условия для получения надежных гематокритных данных принимается центрифугирование при 3000 об/мин в течение 30 мин. 125 Определение количества тромбоцитов [Зупанец И.А. и др., 2005] Кровь смешивают с 14% раствором сернокислого магния. Последующие приготовление и фиксация аналогичны таковым при приготовлении обычного мазка, лишь окрашивание мазка продолжается в течение 2-3 часов. В мазке под микроскопом с иммерсионной системой и окошечком по Фонио сосчитывают 1000 эритроцитов и все тромбоциты, встретившиеся во время подсчета. Расчет тромбоцитов проводят на общее количество эритроцитов, содержащееся в 1 мкл крови. Определение количества лейкоцитов В пробирку наливают 0,4 мл раствора Тюрка (жидкость Тюрка содержит уксусную кислоту для разрушения эритроцитов и метиленовую синь для окраски ядер лейкоцитов). Капиллярной пипеткой набирают из свежей капли 0,02 мл крови. Кровь помещают в пробирку с реактивом и ополаскивают пипетку. Смесь хорошо перемешивают. При этом кровь разводят в 20 раз. Концом круглой стеклянной палочки отбирают каплю разведенной крови и наносят на край шлифованного стекла камеры. Подсчет ведут в 100 больших нерасчерченных квадратах, собранных вместе по четыре. Используют малое увеличение. Количество лейкоцитов в 1 мкл периферической крови, т.е. с учетом разведения в 20 раз равняется (А10)/420=А50. Практически подсчитанное количество лейкоцитов (при разведении в 20 раз) умножают на 50 [Зупанец И.А. и др., 2005]. Определение лейкоцитарной формулы Лейкоцитарную формулу подсчитывают в окрашенных мазках. Для достаточно точного ее вычисления необходимо посмотреть не менее 200 лейкоцитов [Зупанец И.А. и др., 2005]. 126 Каплю крови наносят на обезжиренное предметное стекло в 1,5-5 см от его края. Ребром шлифованного предметного стекла со срезанными углами проводят по предметному стеклу, равномерно распределяя кровь. Мазок высушивают на воздухе и фиксируют в метиловом спирте в течение 3-5 минут. Затем мазок красят по Романовскому-Гимза в течение 30 минут, после чего излишки краски смывают водой и мазок высушивают. Изучение мазка проводят под микроскопом. Для достаточно точного ее вычисления необходимо просмотреть не менее 200 лейкоцитов по 50 клеток в начале и конце мазка, по его верхнему и нижнему краям. Чтобы получить процентное содержание в крови данного вида лейкоцитов, необходимо количество клеток в каждой графе разделить на 2, т. к. было подсчитано 200 клеток. 2.3.4. Методы определения показателей углеводного обмена Для изучения гормоноподобного действия халькогенорганических соединений проводили определение некоторых показателей углеводного обмена: концентрацию глюкозы в плазме крови, пирувата в цельной крови, гликогена в печени и цельной крови, активность ферментов глюконеогенеза глюкозо-6фосфатазы и фруктозо-1,6-бисфосфатазы в печени. Кроме того, в плазме крови также определяли активность ферментов аланинаминотрансферазы (АлАТ) и аспартатаминотрансферазы (АсАТ), поскольку в реакциях, катализируемых этими ферментами, образуются субстраты глюконеогенеза пируват и оксалоацетат соответственно. Определение концентрации пирувата Метод Умбрайта основан на взаимодействии пировиноградной кислоты с 2,4-динитрофенилгидразином в кислой среде с образованием динитрофенилгидразона пировиноградной кислоты [Бабаскин Б.Н., 1976]. 2,4- 127 В центрифужную пробирку (опыт) вносят 0,7 мл дистиллированной воды, 0,3 мл крови и 1 мл 10%-го раствора трихлоруксусной кислоты. Содержимое перемешивают и через 2-3 минуты центрифугируют при 1500 об./мин в течение 15 минут. Надосадочную жидкость полностью сливают в сухую обыкновенную пробирку, добавляют к ней 0,4 мл 0,1%-го раствора 2,4-динитрофенилгидразина, перемешивают и пробу помещают в темное место на 20 минут при комнатной температуре. Затем в пробирку приливают 1 мл 12%-го раствора гидроксида натрия, перемешивают и через 5 минут колориметрируют против контроля в кюветах на 5 мм с синим светофильтром (длина волны 440 нм). Контрольную пробу готовят так же, как и опытную, но вместо крови берут 0,3 мл дистиллированной Н2О, не центрифугируя. Измерение оптической плотности проводят с помощью фотоэлектроколиримерта КФК-3 (Россия). Концентрацию пирувата выражают в моль/л. Определение концентрации гликогена Органические вещества, мешающие определению гликогена, разрушеются кипячением в концентрированном растворе гидроксида калия. Гликоген при этом не разрушается. Гликоген осаждают спиртом и гидролизуют в кислой среде до глюкозы, количество которой и определяют глюкозооксидазным методом [Покровский А.А., 1969]. В центрифужную пробирку наливают 3 мл 30% раствора КОН, к которому добавляют 0,1 мл цельной крови или навеску печени массой 25-100 мг. Пробирку помещают на 20 минут в кипящую водяную баню. После этого пробирку охлаждают, прибавляют 3 мл 96% этилового спирта, размешивают и ставят на кипящую водяную баню до закипания спирта. После охлаждения пробирку центрифугируют 15 минут при 3000 оборотов в минуту, сливают надосадочную жидкость, а осадок растворяют, добавляя 1 мл воды и 1 мл спирта. Опять помещают в кипящую водяную баню. Снова центрифугируют, надосадочную 128 жидкость сливают, а осадок растворяют, прибавляя 2,5 мл дистиллированной воды. В полученном растворе определяют концентрацию глюкозы глюкозооксидазным методом как описано в разделе 2.3.2. Учитывая, что молекулярная масса свободной глюкозы 180, а глюкозы в составе гликогена - 162, для определения концентрации гликогена полученный результат умножают на коэффициент 0,9. Определение активности глюкозо-6-фосфатазы Метод основан на измерении количества неорганического фосфора, образующегося при дефосфорилировании глюкозо-6-фосфата в присутствии глюкозо-6-фосфатазы [Уварова Е. В., 2005]. В опытную пробу помещают 1,5 мл 0,05 М ацетатного буфера (рН 5,0), 0,5 мл 0,06 М глюкозо-6-фосфата и 0,1 мл гомогената печени. Полученную смесь инкубируют 10 минут при 37°С; по истечении этого времени в пробу добавляют 3 мл 5% ТХУ. После перемешивания пробирку центрифугируют 10 минут при 37°С. Параллельно ставят контрольную пробу, в которую ТХУ вносят до добавления гомогената. В надосадочной жидкости опытной и контрольной проб определяют содержание неорганического фосфата по методу Фиске-Субароу (Северин Е.С., Соловьева Г.А., 1989). Для этого в пробирку помещают 2 мл центрифугата (опыт/контроль), 1 мл 1% раствора аскорбиновой кислоты и 1 мл 1% раствора молибдата аммония. Реакцию проводяти 10 минут в темноте. Затем измеряют оптическую плотность опытной и контрольной проб при длине волны 625 нм на фотоэлектроколиримерте КФК-3 (Россия). Параллельно ставят пробу со стандартным раствором КН2РО4 с концентрацией 1 мкмоль/л. Активность глюкозо-6-фосфатазы выражают в мкмоль/(л · час). 129 Определение активности фруктозо-1,6-бисфосфатазы Метод основан на измерении количества неорганического фосфора, образующегося при дефосфорилировании фруктозо-1,6-бисфосфата в присутствии фруктозо-1,6-бисфосфатазы [Уварова Е. В., 2005]. В пробу вносят 1,4 мл глицинового буфера, 0,1 мл 0,15 М MgCl 2, 0,5 мл 0,06 М раствора фруктозо-1,6-дифосфата и 0,1 мл гомогената печени. Полученную смесь инкубируют 10 минут при 37°С; по истечении этого времени в пробирку добавляют 3 мл 5% ТХУ. После перемешивания пробу центрифугируют 10 минут при 37°С. Параллельно ставят контрольную пробу, в которую ТХУ вносят до добавления гомогената. В надосадочной жидкости опытной и контрольной проб определяли содержание неорганического фосфата по методу Фиске-Субароу (см. выше). Активность фруктозо-1,6-бисфосфатазы выражают в мкмоль/(л · час). Определение концентрации глюкозы и аминотрансфераз в плазме крови проволили как описано в разделе 2.3.2. 2.3.5. Определение антибактериального действия халькогенорганических соединений по числу выросших клеточных колоний Эксперимент по изучению антибактериального действия халькогенорганических соединений по числу выросших клеточных колоний проводили на клинических штаммах кишечной палочки (E. coli), синегнойной палочки (Ps. aeruginosa) и золотистого стафилококка (St. aureus). Суспензию бактерий готовили по стандарту мутности ГИСК им. Л.А. Тарасевича, путём последовательных разведений до конечной концентрации бактерий - 3·105 клеток в 1 мл. 1 мг соединения растворяли в 100 мкл ДМФА (диметилформамида), добавляли 900 мкл 0,9%-ного NaCl - проба 1 (1 мг/мл). В качестве контроля 130 использовали 1 мл ДМФА +9 мл 0.9%-ного NaCl. Затем из пробы 1 отбирали 100 мкл, добавляли 900 мкл из контрольной пробирки, получая пробу 2 (0,1 мг/мл). Из пробы 2 отбирали 100 мкл содержимого, добавляли 900 мкл из контроля, получая пробу 3 (0,01 мг/мл). Из пробы 3 отбирали 100 мкл раствора, добавляли 900 мкл из контроля, получая пробу 4 (0,001 мг/мл). Из пробы 4 отбирали 100 мкл раствора, добавляли 900 мкл из контроля, получая пробу 5 (0,0001 мг/мл). В пробирки с разведениями препарата добавляли по 100 мкл конечной суспензии (3·105 КОЕ/мл) микроорганизмов, встряхивали и инкубировали в течение 30, 60, 90, 120, 150 мин при комнатной температуре. В качестве контроля использовали такие же количества бактериальной взвеси, разведенные в аналогичных пропорциях с контролем (ДМФА с 0,9%-ным NaCl), а также выдержанные в течение тех же промежутков времени. После этого бактериальные взвеси из каждой пробирки в количестве 100 мкл высевали на чашки Петри с твердой питательной средой (мясо-пептонный агар) (Agar nutrient (Becton Dickinson, США)), которые затем помещали в термостат на 24 часа при 37ºС. Подсчет колоний производили на следующий день. 2.4. Квантово-химические расчеты Квантовохимические расчеты проводили гидридным методом теории функционала плотности (DFT) [Кон В., 2002] по программам пакета Gaussian 98 W. В рамках SCF использован гибридный функционал B3LYP, сочетающий трехпараметровый обменный функционал Беке [Becke A.D., 1988, 1993] и корреляционный функционал LYP [Lee C. et al., 1988]. Взят базисный набор 6311G(d,p) [Ditchfield R. et al., 1971]. В гармоническом приближении рассчитывались частоты колебаний. Все равновесные структуры без мнимых частот отвечают точкам минимумов на поверхностях потенциальной энергии. Условия сходимости по RMS-матрице плотности составляло 1∙10-8 в течение 200 131 итерационных циклов, по MAX-матрице плотности - 1∙10-6 а.е. Начальная геометрия генерировалась по программам HypeChem [HypeChem (TM), Hypercube, Inc., Gainesville, Florida 32601, U.S.A.] и оптимизировалась методом РМ3 [Stewart J.J.P., 1989; 1989]. Размер молекул оценивался как радиус (а0) сферической полости, занимаемой молекулой растворенного вещества в континууме растворителя. Величина а0 обычно используется для описания сольватационных эффектов методом Онзагера. 2.5. Компьютерное прогнозирование биологической активности с использованием компьютерной системы предсказания спектра биологической активности (PASS C&T) Изучение спектров биологической активности халькогенорганических соединений в зависимости от их химической структуры проводилось с использованием компьютерной системы Prediction of Activity Spectra for Substances (PASS C&T). Интерпретация полученных результатов осуществлялась с помощью программы «Pharma Expert». Данные компьютерные программы разработаны сотрудниками Лаборатории структурно-функционального конструирования лекарств НИИ Биомедицинской химии им. В.Н. Ореховича РАМН профессором В.В. Поройковым, к. ф.-м. н. Д.А. Филимоновым (Мякишева Ю.В., 2009; Поройков В.В. и др., 2009). Список, прогнозируемых PASS (версия 2008 г.) биологических активностей, включает около 4000 наименований, из них 140 наименований, характеризующих блокирование эндогенных биорегуляторов; 71 наименование, характеризующее активирование эндогенных биорегуляторов; 5 наименований, характеризующих блокирование высвобождения эндогенных биорегуляторов; 9 наименований, характеризующих стимулирование высвобождения эндогенных биорегуляторов; 9 наименований, характеризующих блокирование обратного захвата эндогенных 132 биорегуляторов; 2219 наименований, характеризующих ингибирование ферментов; 41 наименование, характеризующее активацию ферментов; 268 наименований, характеризующих блокирование рецепторов; 121 наименование, характеризующее активацию рецепторов; 28 наименований, характеризующих блокирование ионных каналов; 5 наименований, характеризующих активацию ионных каналов; 28 наименований, характеризующих блокирование транспортеров; 128 наименований, характеризующих вещества в качестве субстратов ферментов метаболизма 2; 24 наименования, характеризующих ингибирование ферментов метаболизма; 24 наименования, характеризующих индукцию ферментов ингибирование белков, метаболизма; не 28 наименований, являющихся ферментами; характеризующих 8 наименований, характеризующих ингибирование экспрессии транскрипционных факторов; 2 наименования, характеризующих усиление экспрессии транскрипционных факторов; 72 наименования, характеризующих блокирование биологических процессов; 40 наименований, характеризующих стимулирование биологических процессов; 66 инфекционные наименований, агенты; фармакотерапевтическое характеризующих 261 действие; воздействие наименование, 389 наименований, веществ на характеризующее характеризующих побочное/токсическое действие веществ. Результат прогноза спектра биологической активности представляется в PASS в виде упорядоченного списка названий соответствующих активностей и вероятностей Pa «быть активным» и Pi «быть неактивным» для прогнозируемого соединения, имеющие значения от 0 до 1. Поскольку эти вероятности рассчитываются независимо, их сумма не равна единице. Упорядочение выполняется по убыванию разности Pa–Pi, соответственно более вероятные виды активности находятся в начале спрогнозированного спектра. Величины Pa и Pi интерпретируются как оценки меры принадлежности вещества к классам активных и неактивных соединений либо как оценки ошибок первого и второго 133 рода. Чем больше для конкретной активности величина Pa и чем меньше величина Pi, тем больше шанс обнаружить данную активность в эксперименте (Поройков В.В. и др., 2009). 2.6. Статистические параметры, использованные в работе Статистическую обработку экспериментальных данных проводили с использованием программы Microsoft Excel на персональном компьютере. Проверяли гипотезы о виде распределений (критерий Шапиро-Уилкса). Большинство данных не удовлетворяют закону нормального распределения случайной величины, поэтому для сравнения значений использовали Т-критерий Манна-Уитни, на основании которого оценивали уровень доверительной вероятности 1-α (комплементарная ей величина α называется уровнем значимости). Критический уровень доверительной вероятности при проверке статистических гипотез принимали равным 1-α=0.95 (критический уровень значимости α=0,05), что соответствует показателю достоверности p [Гланц С., 1998]. Для вычисления Т-критерия Манна-Уитни данные контрольной и опытной групп объединяли и упорядочивали по возрастанию. Ранг 1 присваивали наименьшему из всех значений, ранг 2 – следующему и так далее. Наибольший ранг присваивали самому большому среди значений в обеих группах. Если значения совпадали, им присваивают один и тот же средний ранг. Для меньшей группы вычисляли Т – сумму рангов ее членов. Полученное значение Т сравнивали с критическими значениями. Если Т меньше или равно первому из них либо больше или равно второму, то нулевая гипотеза отвергалась (различия статистически значимы). U-критерий Манна-Уитни вычисляли по формуле: U = T – nм (nм + 1) / 2, где nм – численность меньшей из групп [Гланц С., 1998]. 134 Глава 3. Антиоксидантная активность халькогенорганических соединений Антиоксидантная система организма человека и животных направлена на обезвреживание активных форм кислорода и азота, главным из которых является супероксид-анион, способный превращаться в перекись водорода под действием фермента спероксиддисмутазы (СОД). Образованная в результате этой реакции перекись водорода также является токсичной формой кислорода и обезвреживается ферментами глутатионпероксидазой (ГПО) и каталазой. Помимо названных ферментов к антиоксидантам относятся другие ферменты, белки, пептиды, пигменты, витамины, гормоны и микроэлементы (селен, цинк, медь). Однако наиболее важными внутриклеточными антиоксидантами являются ферменты: спероксиддисмутаза (СОД), каталаза и глутатионпероксидаза (ГПО). СОД в своем активном центе может содержать ионы цинка, меди или марганца. Каталаза и ГПО селензависимым - гемопротеины. ферментом, в Кроме активном того, центре ГПО также которого является содержится восстановленный глутатион, который и является донором протонов для обезвреживания перекисей. Недостаток селена в рационе питания может привести к снижению активности ГПО, нарушению синтеза ряда селензависимых ферментов и белков и к снижению общей антиоксидантной защиты организма, следствием чего может явиться развитие таких тяжелых патологий как атеросклероз, ишемическая болезнь сердца, онкологические заболевания, бесплодие и многие другие. Учитывая широкое распространение селенодефицитных состояний, как в России, так и за рубежом, встает вопрос о профилактике селенодефицита. В этой связи в настоящее время проводится синтез и изучение биологической активности селеноорганических соединений. Причем предпочтение отдается органическим соединениям селена, поскольку они менее токсичны и лучше усваиваются в 135 желудочно-кишечном тракте сельско-хозяйственных животных и человека. Одним из перспективных соединений является диацетофенонилселенид (ДАФС-25), который применяется в птицеводстве и животноводстве для выживаемости молодняка, увеличения яйценоскости у кур и пр. В связи с этим нами исследовалась антиоксидантная активность соединения ДАФС, его хлор-, фтор- и нитропроизводных, а также теллурсодержащего аналога ДАФС-25. 3.1. Антиоксидантные показатели гемолизата эритроцитов, плазмы крови и тканей интактных мышей с различной оксидорезистентностью В нашей работе проводилось изучение содержания диеновых конъюгатов (ДК) и малонового диальдегида (МДА), активности антиоксидантных ферментов каталазы и СОД в плазме крови, гемолизате эритроцитов и гомогенатах тканей (мозга, печени, почек и легких), а также активности ГПО в цельной крови мышей. Выбранные ткани отличаются по активности антиоксидантной защиты. Согласно литературным данным, наиболее активно свободнорадикальное окисление липидов осуществляется в селезенке, сердце, легких, почках и мозге. Относительно низкая активность процессов пероксидации выявлена в эритроцитах, печени и плазме [Кирова Ю.И., 2004; Костюк В.А., Потапович А.И., 2004]. Следовательно, нами выбраны 3 органа с низкой оксидорезистентностью (легкие, почки и мозг) и 3 - с высокой оксидорезистентностью (эритроциты, печень и плазма крови). Антиоксидантные показатели у интактных мышей, представленные в таблице 8, подтверждают это. 136 Наибольшая концентрация продуктов ПОЛ – ДК и МДА – отмечается в мозге и легких, и наименьшая – в гемолизате эритроцитов и плазме крови. Однако содержание продуктов ПОЛ в печени больше, чем в почках. Таблица 8. Антиоксидантные показатели у мышей контрольной группы (n=12) Исследуемый показатель ДК, ммоль/л МДА, ммоль/л Каталаза, ммоль/ (мин·л) СОД, усл.ед Эритроциты 1,64± 0,06 0,84± 0,05 Плазма крови 0,65± 0,01 0,08± 0,02 Гомогенаты тканей мозг легкие 6,88± 11,59± 0,32 0,92 7,97± 3,88± 0,18 0,14 печень 4,19± 0,47 3,36± 0,03 0,31± 0,03 0,13± 0,01 0,04± 0,003 0,03± 0,002 0,01± 0,001 0,04± 0,01 1291,67 ±89,5 1121,31 ± 112,5 1436,2± 84,3 895,33± 83,5 3121,76± 142,2 1806,83 ± 71,9 почки 3,35± 0,19 2,03± 0,23 Активность антиоксидантного фермента каталазы максимальна в гемолизате эритроцитов и плазме крови. Активность каталазы в печени и почках сопоставима. В мозге и легких активность каталазы заметно ниже (таблица 8). Максимальная активность СОД обнаружена в легких, поскольку в легких повышена возможность реализации свободнорадикальных реакций. В отличие от других органов, респираторная ткань непосредственно подвергается действию кислорода – инициатора окисления (Бессонова Л.О. и др., 2008). Также высокая активность СОД отмечается в почках. Активность СОД в оксидорезистентных эритроцитах, плазме крови и печени сопоставима. Минимальная активность СОД выявлена в мозге интактных животных. Полученные данные демонстрируют высокую оксидорезистентность в эритроцитах и плазме крови и слабую – в легких и мозге. Низкая оксидорезистентность и одновременно значительная интенсивность липопероксидации в клетках мозга вызвана относительно высоким по сравнению с другими тканями и органами содержанием липидов, подвергающихся окислению [Северин Е.С., 2004], а в легких – высокой концентрацией кислорода, 137 индуцирующего свободно-радикальные реакции [Бессонова Л.О. и др., 2008]. Различная устойчивость эритроцитов, клеток печени и почек к окислительному стрессу связана с наличием внутриклеточных антиоксидантов и, в первую очередь, ферментов СОД, каталазы и ГПО. Сравнительно высокая антиоксидантная активность плазмы крови объясняется содержанием таких антиоксидантов как церулоплазмин, трансферрин, витамины Е и С, каротиноиды, эстрогены и др. [Теселкин Ю.О., 2003]. Литературные антиоксидантов данные свидетельствуют, способствует снижению что применение содержания различных продуктов ПОЛ и нарастанию активности антиоксидантных ферментов каталазы, СОД и ГПО [Костюк В.А., Потапович А.И., 2004]. 3.2. Концентрации продуктов перекисного окисления липидов в гемолизате эритроцитов, плазме крови и тканях мышей с различной оксидорезистентностью Диеновые конъюгаты и малоновый диальдегид являются продуктами ПОЛ, указывающими на интенсивность окислительного повреждения мембранных структур клетки [Тарасов Н.И. и др., 2002]. Действие антиоксидантов различной природы приводит к снижению концентрации этих продуктов. В нашем эксперименте концентрация диеновых конъюгатов достоверно снижалась по сравнению с контролем только в органах с низкой оксидорезистентностью – легких и/или мозге. В таблице 9 представлены результаты концентраций диеновых конъюгатов (ммоль/л) в гомогенатах тканей, гемолизате эритроцитов и плазме крови мышей, получавших per os халькогенорганические соединения. Например, пероральное селеноорганических соединений введение ДАФС-25 белым и его беспородным мышам галогенопроизводных 138 (соединения 1, 2 и 3) приводило к снижению концентрации диеновых конъюгатов в легких на 47,6%, 29,1%, 45,5% соответственно, а действие селеноорганического соединения 4 (нитропроизводное ДАФС-25) сопровождалось снижением концентрации диеновых конъюгатов в мозге на 24,3%. Теллурорганическое соединение вызывало снижение содержания диеновых конъюгатов в мозге и легких мышей на 26,6% и 66,0% соответственно (таблица 9). Таблица 9. Концентрация диеновых конъюгатов (ммоль/л) в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших per os халькогенорганические соединения Исследуемая ткань Контроль (n=12) Гемолизат 1,64± эритроцитов 0,06 Плазма крови 0,65± 0,01 Гомогенаты тканей печени 4,19± 0,47 мозга 6,88± 0,32 легких 11,59± 0,92 почек 3,35± 0,19 Халькогенорганические соединения ДАФС(Se) (n=10) 1,70 ± 0,03 ДАФС(Cl2) (n=10) 1,74± 0,05 ДАФС(F2) (n=10) 1,59± 0,03 ДАФС(NO2)2 (n=10) 1,74± 0,08 ДАФС(Те) (n=10) 1,72± 0,11 0,69 ± 0,04 0,71± 0,03 0,64± 0,04 0,73± 0,04 0,69± 0,04 4,69± 0,24 8,67± 0,66* 6,76± 0,36* 4,87± 0,26 5,96± 0,43* 6,63± 0,42 6,55± 0,3 6,1± 0,28 5,21± 0,27* 5,05± 0,2* 6,07± 0,33* 8,22± 0,86* 6,32± 0,41* 13,13± 0,17 3,94± 0,24* 4,29± 0,42 4,29± 0,64 3,59± 0,32 4,51± 0,18* 3,98± 0,22* Примечание: * - р<0,05 - здесь и далее достоверность различий по сравнению с контролем Однако обращает на себя внимание и увеличение концентрации диеновых конъюгатов в гомогенатах печени и/или почек при пероральном введении мышам селеноорганических соединений 2 (на 106,9%), 3 (на 61,3%) и 4 (на 34,3%), а также теллуроорганического соединения (на 42,2% в печени и 18,8% в почках). 139 Подобное действие исследованных соединений на печень и почки можно объяснить участием данных органов в метаболизме и экскреции этих гидрофобных ксенобиотиков (таблица 9). Малоновый диальдегид как конечный продукт ПОЛ является более информативным показателем окислительных процессов в клетках. Таблица 10 содержит результаты концентраций малонового диальдегида в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших per os халькогенорганические соединения. Таблица 10. Концентрация малонового диальдегида (ммоль/л) в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших per os халькогенорганические соединения Халькогенорганические соединения Исследуемая Контроль ткань (n=12) ДАФС(Se) (n=10) Гемолизат 0,84± эритроцитов 0,05 Плазма крови 0,08± 0,02 Гомогенаты тканей печени 3,36± 0,03 мозга 7,97± 0,18 легких 3,88± 0,14 почек 2,03± 0,23 0,42± 0,03* ДАФС(Cl2) (n=10) 0,19± 0,06* ДАФС(F2) (n=10) 0,67± 0,04 ДАФС(NO2)2 (n=10) 0,31± 0,02* ДАФС(Те) (n=10) 0,74± 0,05 0,05 ± 0,005* 0,09± 0,01 0,16± 0,01* 0,03± 0,002* 0,03± 0,006* 2,92± 0,32 3,76± 0,33 3,8± 0,44 3,69± 0,43 1,89± 0,16* 4,66± 0,2* 5,17± 0,48* 6,88± 0,09* 4,07± 0,19* 3,35± 0,42* 2,06± 0,17* 1,41± 0,29* 2,92± 0,11* 3,01± 0,22* 3,35± 0,45 1,61± 0,12* 1,57± 0,11 1,9± 0,04 1,74± 0,28 1,73± 0,16 * - р<0,05 Все исследованные соединения при введении мышам per os способствовали снижению концентрации МДА во всех исследованных тканях. Антиоксидантная активность соединения 1 (ДАФС-25) проявлялась снижением содержания МДА в 140 гемолизате эритроцитов (на 49,7%), плазме крови (на 41,7%), мозге (на 41,5%), легких (на 46,9%) и почках (на 20,7%). Действие соединения 2 in vivo приводило к снижению концентрации МДА в гемолизате эритроцитов (на 77,7%), в мозге (на 35,1%) и легких (на 63,7%), а соединения 3 – только в мозге (на 13,7%) и легких (на 24,7%). Антиоксидантное действие соединения 4 было сопоставимо с действием препарата ДАФС-25 (соединение 1), поскольку его пероральное введение сопровождалось снижением концентрации МДА в гемолизате эритроцитов (на 63,4%), плазме крови (на 59,5%), мозге (48,9%) и легких (22,4%). Теллурсодержащий аналог препарата ДАФС-25 способствовал снижению содержания МДА только в плазме крови (на 59,5%), печени (на 43,8%) и мозге (на 57,9%). Согласно полученным результатам, соединение 3 обладало наименьшей антиоксидантной активностью в тканях мышей, а кроме того, действие этого соединения сопровождалось увеличением концентрации МДА в плазме крови на 90,5%. 3.3. Активность антиоксидантных ферментов в гемолизате эритроцитов, плазме крови и тканях мышей с различной оксидорезистентностью СОД является ключевым антиоксидантным ферментом, который катализирует дисмутацию супероксидного радикала в перекись водорода и молекулярный кислород, защищая таким образом клетки, ткани и биомолекулы от воздействия свободных радикалов. Результаты активности СОД в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших соединения, представлены в таблице 11. per os халькогенорганические 141 В нашем эксперименте наиболее выраженное повышение активности СОД в плазме крови, гемолизате эритроцитов и гомогенатах тканей наблюдалось у мышей, получавших per os селеноорганические соединения 1-4. Таблица 11. Активность супероксиддисмутазы (в условных единицах) в гемолизате эритроцитов, плазме крови и гомогенатах тканей у мышей, получавших per os халькогенорганические соединения Исследуемая ткань Гемолизат (n=12) ДАФС(Se) (n=10) 1291,67± эритроцитов 89,5 Плазма крови 1121,31± 112,5 Гомогенаты тканей печени 1436,16± 84,3 мозга 895,33± 83,5 легких 3121,76± 142,2 почек Халькогенорганические соединения Контроль 1806,83± 71,9 34116,67 ± 1325,6* ДАФС(Cl2) (n=10) 10599,35± 625,4* ДАФС(F2) (n=10) 7262,33± 748,3* ДАФС(NO2)2 (n=10) 18333,33± 1102,2* ДАФС(Те) (n=10) 15797,33± 679,6* 1642,45± 72,7* 546,67± 35,67* 1746,33± 244,5* 2954,67± 153,6* 508,33± 12,4* 7891,0 ± 484,5* 3650,1± 161,7* 3100,29± 172,2* 3422,86± 145,6* 5619,37± 164,7* 3290,33± 278,5* 766,33± 63,5 1943,37± 105,6* 1591,87± 46,7* 1720,11± 173,4* 4835,57± 66,4* 5035,12± 266,7* 4489,93± 271,6* 4511,67± 263,4* 2081,73± 68,8* 3455,56 ± 148,4* 6377,19± 273,8* 2350,67± 175,4 3551,67± 172,3* 1422,86± 28,8* * - р<0,05 Селеноорганический препарат ДАФС-25 (соединение 1) вызывал наибольший эффект, что выражалось в увеличении активности СОД в гемолизате эритроцитов (на 2541,3%), плазме крови (на 46,5%), печени (на 449,5%), мозге (на 267,5%), легких (на 54,9%) и почках (на 91,2%). Соединение 4 демонстрировало сходный эффект на активность СОД, активность которой повышалась в гемолизате эритроцитов (на 1319,4%), плазме крови (на 163,6%), печени (на 138,4%), мозге (на 77,8%), легких (на 44,5%) и почках (на 96,6%). 142 Фторсодержащее производное препарата ДАФС-25 (соединение 3) оказывало заметно меньшее действие на активность СОД in vivo, а именно активность СОД возрастала в гемолизате эритроцитов (на 462,2%), плазме крови (на 55,8%), печени (на 115,9%), мозге (на 117,1%) и легких (на 43,8%) (таблица 11). Действие хлорсодержащего производного препарата ДАФС-25 (соединение 2) характеризовалось наименьшим стимулирующим действием на активность СОД среди изученных селеноорганических соединений. Так, активность СОД при пероральном введении мышам этого препарата увеличивалась в гемолизате эритроцитов (на 720,6%), в печени (на 154,2%), легких (на 61,3%) и почках (на 252,9%), тогда как в плазме крови, напротив, наблюдалось достоверное снижение активности СОД на 51,2% по сравнению с контролем. Пероральное введение мышам теллурорганического аналога препарата ДАФС-25 сопровождалось увеличением активности СОД только в гемолизате эритроцитов (на 1123%), печени (на 291,3%) и мозге (на 92,1%), а в плазме крови, легких и почках активность СОД, наоборот, снижалась на 54,7%, 33,3% и 21,3% соответственно. Следовательно, селеноорганический препарат ДАФС-25 и его производные, содержащие у фенильных радикалов атомы хлора, фтора и аминогруппы, оказывали выраженное стимулирующее действие на активность одного из главных антиоксидантных ферментов СОД, который обезвреживает высокоактивный супероксид-анион до перекиси водорода и кислорода. Образованная под действием СОД Н2О2 относится к АФК и может оказывать повреждающее действие на все компоненты клетки. Поэтому для нормальной обезвреживание жизнедеятельности перекиси глутатионпероксидазой. клетки водорода необходимо ферментами своевременное каталазой и 143 Таблица 12 демонстрирует результаты активности каталазы в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших per os халькогенорганические соединения. Таблица 12. Активность каталазы в гемолизате эритроцитов, плазме крови и гомогенатах тканей мышей, получавших per os халькогенорганические соединения Исследуемая Контроль Халькогенорганические соединения ткань (n=12) ДАФС(Se) (n=10) ДАФС(Cl2) (n=10) ДАФС(F2) (n=10) ДАФС(NO2)2 (n=10) ДАФС(Те) (n=10) Гемолизат 0,31± эритроцитов 0,03 0,84± 0,03* 0,47± 0,05* 1,43± 0,41* 0,97± 0,07* 0,83± 0,04* Плазма крови 0,13± 0,13± 0,01 0,27± 0,02* 0,28± 0,04* 0,32± 0,03* 0,22± 0,02* 0,06± 0,003* 0,08± 0,01* 0,24± 0,03* 0,17± 0,01* 0,13± 0,02* 0,08± 0,01* 0,07± 0,005* 0,08± 0,01* 0,04± 0,003* 0,05± 0,007* 0,04± 0,008* 0,03± 0,01* 0,05± 0,005* 0,05± 0,01* 0,05± 0,005* 0,05± 0,01 0,12± 0,01* 0,09± 0,02* 0,07± 0,01* 0,05± 0,003 0,01 Гомогенаты тканей печени 0,04± 0,003 мозга 0,03± 0,002 легких 0,01± 0,001 почек 0,04± 0,01 * - р<0,05 Наибольшее увеличение активности каталазы в тканях мышей вызывало пероральное введение хлор-, фтор- и нитропроизводных препарата ДАФС-25 (соединения 2, 3 и 4) (таблица 12). Причем фторсодержащий аналог ДАФС-25 оказывал максимальный эффект, что выражалось в увеличении активности каталазы в гемолизате эритроцитов (на 461,3%), плазме крови (на 115,4%), печени (на 500,0%), мозге (на 166,7%), легких (на 400,0%) и почках (на 125,0%). Действие нитропроизводного препарата ДАФС-25 (соединение 4) in vivo характеризовалось повышением активности каталазы в гемолизате эритроцитов (на 212,9%), плазме крови (на 146,2%), печени (на 325%), мозге (на 33,33%), легких (на 400) и почках 144 (на 75%). Хлорсодержащее производное препарата ДАФС-25 (соединение 2) при пероральном введении мышам оказывало стимулирующее действие на активность каталазы, что выражалось в достоверном увеличении активности этого фермента в гемолизате эритроцитов (на 51,6%), плазме крови (на 107,7%), печени (на 100%), мозге (на 133,3%), легких (на 200%) и почках (на 200%). У мышей, получавших per os препарат ДАФС-25 (соединение 1), отмечалось повышение активности каталазы в гемолизате эритроцитов (на 170,9%), печени (на 50%), мозге (на 166,7%) и легких (на 300%). Действие препарата ДАФС и его теллурсодержащего аналога были сопоставимы. Так, пероральное введение теллурорганического соединения сопровождалось увеличением активности каталазы в гемолизате эритроцитов (на 167,7%), плазме крови (на 69,2%), печени (на 225%), мозге (на 66,7%) и легких (на 400%). Главным антиоксидантным ферментом, участвующим в разрушении токсичной перекиси водорода, является селензависимая глутатионпероксидаза, результаты активности которой представлены в таблице 13. Таблица 13. Активность глутатионпероксидазы (ГПО) в цельной крови Исследуемый Халькогенорганические соединения Контроль показатель (n=12) ГПО, 365,31 601,47± 555,14± 524,8± 726,52± 449,36± ед/ г Hb ±13,4 42,3* 27,8* 16,7* 33,5* 17,6* ДАФС(Se) ДАФС(Cl2) (n=10) (n=10) ДАФС(F2) ДАФС(NO2)2 (n=10) (n=10) ДАФС(Те) (n=10) * - р<0,05 Согласно полученным результатам, достоверное увеличение активности ГПО в цельной крови мышей, получавших per os все исследуемые в этой серии экспериментов соединения на 64,7%, 51,9%, 43,7%, 98,9% и 23,0% соответственно. Значительное повышение активности ГПО при действии in vivo нитропроизводного препарата ДАФС-25 (соединение 4) можно объяснить более 145 легким разрывом связи «С-Se», благодаря чему селен быстрее освобождается из этого соединения и вступает в синтез селенопротеинов и ферментов, главным из которых является ГПО. Разрыву связи «С-Se» способствуют карбонильные группы в составе исследованных соединений, которые обладают отрицательным индуктивным эффектом и способны оттягивать на себя электроны, приводя к разрыхлению этой связи и освобождению атома селена из молекулы. Кроме того, нитрогруппа в составе соединения 4 также способна вызывать разрыхление связи «C-Se», т.к. она обладает отрицательным мезомерным эффектом и способна оттягивать на себя электронную плотность в сопряженной системе [Тюкавкина Н.А., 2004]. Таким образом, в разрыхление связи «C-Se» соединения 4 вносят вклад как карбонильные группы, так и нитрогруппы, поэтому, можно предположить, что атом селена из этого соединения будет освобождаться быстрее, чем из других селеноорганических соединений, что подтверждается более высокой активностью ГПО при действии этого соединения. Теллурорганическое соединение вызывало наименьшее активирующее действие на активность ГПО в цельной крови экспериментальных мышей. Подводя итог полученным результатам, нельзя не отметить, что исследованные халькогенорганические препараты - ДАФС-25 (соединение 1), его хлор-, фтор-, нитросодержащие аналоги (соединения 2, 3 и 4 соответственно), а также препараты, в которых атом селена замещен на атомы серы и теллура (соединения 5 и 6 соответственно) - наиболее эффективно оказывали антиоксидантное действие на органы мышей с низкой оксидорезистентностью, главным образом, на легкие и мозг. Более слабый антиоксидантный эффект наблюдался в высокооксидорезистентных гемолизате эритроцитов и плазме. Наименьшая обнаружена в антиоксидантная клетках печени активность и почек. исследованных Подобную соединений неэффективность антиоксидантов в клетках этих органов можно объяснить их участием в 146 метаболизме и экскреции ксенобиотиков. В печени происходит детоксикация гидрофобных веществ, а почки участвуют в их выведении. Селеноорганические соединения 1, 2, 3 и 4 оказывали наименьшее антиоксидантное действие на печень, а теллуроорганическое соединение 6 – на почки. Таким образом, антиоксидантное действие исследованных соединений напрямую зависело от их общего биологического эффекта in vivo. Из изученных соединений наибольший антиоксидантный эффект продемонстрировало соединение 1 (ДАФС-25), поскольку при его применении происходило одновременное снижение концентрации продуктов увеличение активности ферментов каталазы, СОД и ГПО в ПОЛ и гемолизате эритроцитов, плазме крови и гомогенатах тканей. Высокой антиоксидантной активностью обладало соединение 4 (нитропроизводное ДАФС-25), которое значительно снижало реакции ПОЛ в мозге, легких, эритроцитах и плазме крови. Соединение 2 (хлорсодержащий аналог ДАФС-25) наибольшую антиоксидантную активность оказывал на эритроциты и легкие. Соединение 3 (фторсодержащее производное ДАФС-25) способствовало снижению концентрации продуктов ПОЛ в мозге и легких. В печени, напротив, наблюдалось нарастание концентрации продуктов ПОЛ (ДК). Данное соединение оказывало слабый суммарный антиоксидантный эффект, что, безусловно, связано с токсичностью самого препарата, обусловленной наличием двух атомов фтора в пара-положении фенильных заместителей. Теллуроорганическое соединение также оказывало токсический эффект, но за счет наличия в молекуле атома теллура. Как упоминалось выше, в ряду сера>селен>теллур нарастает токсичность, поэтому данное соединение гораздо токсичнее селенсодержащего аналога – соединения 1. Этим объясняется наименьшее антиоксидантное действие данного соединения в эритроцитах и 147 почках. Теллуроорганическое соединение обладало достоверной антиоксидантной активностью лишь в органах с низкой оксидорезистентностью – в легких и мозге. Установленные ранее [Кирова Ю.И., 2004] эффекты серосодержащего аналога ДАФС-25 (соединение 5) на показатели свободнорадикального окисления в тканях мышей с различной оксидорезистентностью позволяют нам сделать вывод о сходной антиоксидантной активности этого серосодержащего соединения с действием селеноорганических соединений 2 и 4. Подводя итог представленным выше данным, можно заключить, что антиоксидантная активность изученных халькогенорганических соединений убывала в ряду: ДАФС(Se) > ДАФС(NO2)2) ≥ ДАФС(S) ≥ДАФС(Сl2) > ДАФС(F2) > ДАФС(Те). Очевидно, что наличие боковых радикалов в фенильных заместителях, а также замена атома селена на серу или теллур снижает антиоксидантные свойства халькогенорганических соединений. 3.4. Механизм антиоксидантного действия халькогенорганических соединений Предположительный механизм антиоксидантного действия изученных халькогенорганических соединений имеет 3 аспекта. Во-первых, атом селена может освобождаться из молекулы соединения и использоваться для синтеза селензависимых белков и ферментов, в т.ч. и ГПО, которые будут разрушать токсичные перекиси, подавлять реакции свободно-радикального окисления и, тем самым, препятствовать ПОЛ. Освобождение атома селена, в свою очередь, возможно благодаря разрыхлению связи «C-Se» в молекулах селеноорганических соединений 1-4. Этому способствуют карбонильные группы в составе данных соединений, которые обладают отрицательным индуктивным эффектом и 148 способны оттягивать на себя электроны, приводя к разрыхлению связи «C-Se» и освобождению атома селена из молекулы. Галогены (атомы хлора и фтора) в составе соединений 2 и 3 соответственно имеют отрицательный индуктивный эффект и, обладая неподеленной парой электронов, оказывают положительный мезомерный эффект, который по величине больше отрицательного индуктивного эффекта. Следовательно, атомы хлора и фтора в составе препаратов 2 и 3 не вносят вклада в разрыхление связи C-Se [Тюкавкина Н.А., 2004]. Кроме того, галогены обладают окислительными свойствами, что обусловливает токсичность их соединений и, как следствие, более слабый антиоксидантный эффект по сравнению с соединением 1. Нитрогруппа в составе препарата 4 также способна вызывать разрыхление связи «C-Se», т.к. она обладает отрицательным мезомерным эффектом и способна оттягивать на себя электронную плотность в сопряженной системе [Тюкавкина Н.А., 2004]. Таким образом, в разрыхление связи «C-Se» соединения 4 вносят вклад как карбонильные группы, так и нитрогруппы, поэтому, как мы предполагаем атом селена из соединения 4 будет освобождаться быстрее, чем из остальных халькогенорганических соединений 1, 2, 3, 5 и 6. Это предположение подтверждается максимальной активностью селензависимой ГПО у мышей, получавших перорально соединение 4 по сравнению с другими экспериментальными группами животных (таблица 13). Активность ГПО в цельной крови у мышей, получавших перорально соединение 4, увеличивалась на 98,9% по сравнению с контролем, что заметно выше по сравнению с другими исследованными препаратами (соединение 1 – активность ГПО возрастала на 64,6%, соединение 2 – на 51,9%, соединение 3 на 43,7% по сравнению с контролем, теллурорганическое соединение 6 – изменения активности ГПО статистически недостоверны). Отсутствие повышения активности ГПО в цельной крови у мышей, получавших per os препарат 6, подтверждает предположение, что 149 селеноорганические препараты являются источником селена для синтеза селенсодержащих белков и ферментов, в т.ч. ГПО. Помимо этого предположения о механизме антиоксидантного действия халькогенорганических соединений есть версия [Кирова Ю.И., 2004], что атом селена в составе исследованных препаратов способен нейтрализовать активные формы кислорода (АФК). Во всех изученных соединениях присутствует халькоген (селен или теллур) в степени окисления +2, что определяет восстановительную природу соединения и, следовательно, его активное участие в процессах неферментативного перекисного окисления биологических молекул. Подобный восстановительный центр присутствует в молекуле диалкилсульфидов, для которых в настоящее время доказана способность к эффективному разложению гидроперекисей липидов, что предотвращает образование высокореакционных алкоксильных (RO˙) и перекисных радикалов (RO2˙): O + ROOH R-S-R → R-S-R + диалкилсульфид органическая монотиооксид гидроперекись ROH оксипроизводное органической молекулы Учитывая общность в строении диалкилсульфидов и халькогенсодержащих арилалифатических дикетонов, можно предположить сходный характер их химических свойств и, прежде всего, способности разлагать гидроперекиси. Кроме того, селен характеризуется меньшей величиной относительной электроотрицательности по сравнению с серой и, следовательно, более эффективен при восстановлении гидроперекисей [Кирова Ю.И., 2004]. Наконец, третий предполагаемый механизм действия халькогенорганических соединений связан с их фенильными радикалами. Максимальной антиоксидантной активностью обладало соединение 1, особенностью которого являлось отсутствие боковых радикалов у фенильных заместителей. Можно предположить, что фенильные радикалы в составе 150 соединения 1 подвергались гидроксилированию при участии цитохрома Р450 в ЭПР печени с образованием антиоксидантной фенонильных активностью групп, наравне с обладающих другими выраженной гидрофобными антиоксидантами, такими как токоферол, филлохинон, убихинон и т.д. [Burton G.M., Traber M.G., 1990.; Olson J.A., 1992]: Следовательно, в этой серии экспериментов из всех исследованных соединений максимальной антиоксидантной активностью обладали соединения 1 и 4 благодаря способности освобождать атом селена из соединения и/или способности образовывать фенонильные радикалы при окислении в гепатоцитах. 151 Глава 4. Антитоксическое действие халькогенорганических соединений при интоксикации солями тяжелых металлов В настоящее время значительно ухудшилась экологическая обстановка вследствие загрязнения окружающей среды токсическими веществами, главными из которых являются соли тяжелых металлов. Тяжелые металлы попадают в окружающую среду с промышленными и бытовыми отходами, загрязняют почву, аккумулируются в тканях растений и животных и, наконец, в составе загрязненного воздуха, воды и пищи попадают в организм человека [Satarug S. et al., 2010]. Наиболее распространенными и токсичными являются соли ртути, свинца и кадмия. Согласно классификации металлов по их токсичности ртуть и кадмий относятся к особо токсичным металлам, свинец – к умеренно токсичным металлам. Главный токсический эффект ионов металлов Pb2+, Hg2+ и Cd2+ заключается в образовании прочных комплексов с аминокислотами и другими биомолекулами, содержащими концевые тиогруппы (-SH) [Matovic V. et al, 2011; Thevenod F., 2009]. Другой важный механизм токсического действия ртути и свинца заключается в вытеснении эссенциальных металлов из металлсодержащих комплексов, приводящем к потере последними биологической активности [Ikediobi C.O. et al., 2004]. Так происходит дезактивация участвующих в синтезе гема ферментов карбоангидразы и аминолевулинатдегидрогеназы в результате замены содержащегося в них иона Zn2+ на Hg2+ или на Рb2+ [Исидоров В.А., 1999]. Особенно опасны с точки зрения ингибирования синтеза гема ионы Рb2+. Помимо уже упомянутых ферментов - карбоангидразы и аминолевулинатдегидрогеназы - мишенью этих ионов служит феррохелатаза. Кроме того, ионы свинца активируют фермент гемоксигеназу, разлагающий гем. 152 Таким же действием обладают ионы кобальта и кадмия. Потеря организмом животного гема приводит к дефициту гемоглобина и развитию анемии. Токсический эффект тяжелых металлов связан также с нарушением синтеза различных форм цитохрома Р-450 монооксигеназ гладкого эндоплазматического ретикулума, содержащих в качестве простетической группы гем. Поскольку цитохром Р-450 отвечает за окисление ксенобиотиков, делающее возможным их последующую конъюгацию и выведение из организма, нарушение этой системы приводит к накоплению органических токсикантов в тканях и органах. Тяжелые металлы ингибируют также и некоторые звенья второй фазы детоксикации, в ходе которой, собственно, и осуществляется конъюгация подвергшегося окислению ксенобиотика с остатком глюкуроновой или серной кислоты. Цитохромы Р-450 участвуют в метаболизме не только ксенобиотиков, но и эндогенных биологически активных веществ - кортикоидных и тиреоидных гормонов, андрогенов, катехоламинов, витаминов группы D, холестерина и т. д. Поэтому нарушение их синтеза или снижение активности может вызвать глубокие нарушения самых различных процессов в организмах животных [Исидоров В.А., 1999]. Еще одним важным аспектом токсичности тяжелых металлов является активация перекисного и свободнорадикального окисления. В результате такой активации повреждаются некоторые белки, нуклеиновые кислоты, липиды, а также биомембраны. Частично повреждающий эффект объясняется ингибированием металлами ферментов, защищающих организм от накопления в нем Н2О2. Пероксид водорода является предшественником высокоактивного в реакциях окисления свободного радикала гидроксила [Koedrith P., Seo Y.R., 2011]. Таким образом, основными молекулярными и клеточными мишенями для ионов тяжелых металлов служат: гемсодержащие белки и ферменты; 153 ферменты, участвующие в процессах конъюгации; системы пероксидного и свободнорадикального окисления липидов и белков, а также системы антиоксидантной и антипероксидной защиты; ферменты транспорта электронов и синтеза АТФ. Следовательно, биологическая активность металлов связана с их способностью связываться с белками, блокировать многие ферментные системы, повреждать клеточные мембраны, повышать проницаемость барьеров, что в итоге ведет к токсическим изменениям в клетках и органах. Соединения металлов, хорошо растворимые в воде и биологических жидкостях, легче проникают через биологические барьеры и вызывают нарушения в организме. Любой металл, поступивший в организм, довольно быстро проникает в кровь и затем во внутренние органы. Обычно органами максимального накопления металлов в организме является печень, почки и костная система. Из внутренних органов металлы медленно, приблизительно в течение 1 месяца, выделяются из организма, преимущественно почками или кишечником. Для оценки тяжести отравления солями тяжелых металлов и антитоксической активности халькогенорганических препаратов определяли биохимические, гематологические показатели крови крыс, а также проводили расчет лейкоцитарных индексов интоксикации. 4.1. Оценка функциональной активности органов и тканей белых крыс при использовании халькогенорганических препаратов в качестве антитоксикантов при интоксикации солями тяжелых металлов Используя биохимический анализ крови, можно оценить функциональное состояние почек, печени и других органов при интоксикации организма солями тяжелых металлов, а также проследить нормализацию биохимических показателей 154 крови при использовании халькогенорганических препаратов в качестве антитоксикантов. В нашем эксперименте в качестве антитоксикантов при интоксикации организма солями кадмия, свинца и ртути использовали халькогенорганические соединения: препарат диацетофенонилселенид – ДАФС-25 (соединение 1), его хлоро- (соединение 2), фторо- (соединение 3), нитропроизводные (соединение 4), а также аналоги препарата ДАФС, содержащие атомы серы (соединение 5) и теллура (соединение 6) вместо атома селена. В плазме крови экспериментальных животных определяли концентрацию основных метаболитов (глюкозы, общего белка, альбумина, креатинина, мочевины и холестерина) и активность ферментов: аспартатаминотрансферазы (АсАТ), аланинамнотрансферазы (АлАТ), амилазы, щелочной фосфатазы, γ-глутамилтрансферазы (ГГТ) и лактатдегидрогеназы (ЛДГ). Биохимические показатели крови экспериментальных животных при введении per os халькогенорганических препаратов in vivo представлены в таблице 14. Халькогенорганические препараты не оказывали значительного токсического действия на организм экспериментальных животных. Однако можно отметить увеличение концентрации глюкозы сыворотки крови на 41,2% у крыс, получавших препарат ДАФС (соединение 1) и уменьшение содержания глюкозы в плазме крови у крыс, получавших соединения 4 и 6 на 41,2% и 35,3% соответственно по сравнению с контролем (р<0,05). Достоверное увеличение содержания общего белка в плазме крови у крыс, принимавших препараты 3, 4, 5 и 6, было незначительным. Обращает внимание увеличение в плазме крови содержания мочевины и креатинина. Например, концентрация креатинина возрастала у крыс, принимавших почти все соединения на 22,0% (соединение 1), 36,5% (соединение 3), 62,9% (соединение 4), 56,6% (соединение 5) и на 87,7% (соединение 6); содержание мочевины также увеличивалось на 51,3% (соединение 155 2), 38,5% (соединение 3), 48,7% (соединение 5) и на 56,4% (соединение 6) соответственно по сравнению с контролем. Достоверное увеличение концентрации холестерина в сыворотке крови на 46,2% наблюдалось только у крыс, получавших препарат ДАФС. Кроме того, следует отметить увеличение активности почти всех исследованных ферментов плазмы крови у крыс, принимавших халькогенорганические препараты. Активность АлАТ в плазме крови достоверно возрастала у крыс на 140,5% (соединение 1), 21,9% (соединение 3) и 31,8% (соединение 6), тогда как применение соединения 2, напротив, приводило к незначительному снижению активности этого фермента на 24.8%. Также наблюдалось увеличение активности АсАТ у крыс, получавших соединения 2, 3, 4 и 6 на 37%, 51,1%, 70,9% и 80,8% соответственно по сравнению с контролем. Активность щелочной фосфатазы также возрастала на 37,3% (соединение 2), 32,2% (соединение 3) и 42,4% (соединение 4) соответственно. Активность амилазы достоверно увеличивалась лишь у крыс, принимавших препараты 2 и 6 на 31,9% и 22,5% соответственно. Увеличение активности ЛДГ отмечалось у крыс, принимавших соединения 1, 2, 3, 4 и 6 на 20,9%, 140,4%, 32,1%, 39,7% и 32,2% соответственно по сравнению с контролем. Активность печеночного фермента ГГТ достоверно снижалась на 28,3% лишь у крыс, получавших теллуроорганическое соединение (таблица 14). Приведенные результаты свидетельствуют о незначительном токсическом воздействии халькогенорганических соединений на организм экспериментальных животных, который выражался, главным образом, в воздействии на почки и печень крыс. Из исследованных соединений наибольший токсический эффект оказывал теллуроорганический препарат, а наименьший – серосодержащий аналог ДАФС. 156 Таблица 14. Биохимические показатели крови экспериментальных животных при введении per os халькогенорганических препаратов Показатель Единицы Контроль измерения (n=12) Халькогенорганические соединения 1 (n=10) 2 (n=10) 3 (n=10) 4 (n=10) 5 (n=10) 6 (n=10) Глюкоза ммоль/л 3,4±0,3 4,8±0,1* 2,9±0,2 2,8±0,2 2,0±0,2* 2,9±0,3 2,2±0,2* Общий г/л 69,3±3,4 68,1±4,5 70,4±3,7 78,6±3,8* 75,9±3,6* 76,3±3,2* 76,9±3,5* альбумин г/л 32,5±2,4 30,7±1,7 33,5±1,3 32,4±2,4 31,6±1,8 29,9±2,1 29,6±1,6 креатинин мкмоль/л 35,9±2,2 43,8±2,3* 39,5±1,5 49±3,9* 58,5±3,7* 56,2±3,2* 67,4±5,8* мочевина ммоль/л 3,9±0,3 4,7±0,3 5,9±0,3* 5,4±0,2* 4,9±0,3 5,8±0,3* 6,1±0,3* холестерин ммоль/л 1,3±0,1 1,9±0,2* 1,3±0,54 1,5±0,09 1,4±0,1 1,4±0,1 1,3±0,1 АлАТ Ед/л 112,8±6,7 271,3±11,3* 84,8±4,6* 88,1±6,9* 148,7±9,6* 121,4±8,7 118,7±7,7 АсАТ Ед/л 98,3±2,8 123,7±12,2 134,7±6,7* 148,5±9,9* 168,1±9,3* 106,3±6,7 177,7±11,5* амилаза Ед/л 1397,2± 54,5 1568,3±79,5 1843,2±78,5* 1323,7±52,5 1643,4±78,8 1461,3±110,7 1711,1±122,7* Щелочная Ед/л 5,9±0,3 5,9±0,4 8,1±0,3* 7,8±0,3* 8,4±0,6* 5,6±0,3 5,5±0,2 ГГТ Ед/л 14,5±0,9 13,4±0,5 14,9±0,7 16,1±3,4 16,8±1,8 13,8±1,4 10,4±1,6* ЛДГ Ед/л 269,1±13,5 325,3±30,5* 377,7±12,8* 355,4±21,2* 375,7±28,9* 266,7±18,9 355,7±20,5* белок * - р<0,05 156 фосфатаза 157 Таблица 15 содержит биохимические показатели крови экспериментальных животных при подостром отравлении солями тяжелых металлов. Соли тяжелых металлов оказывали выраженное токсическое действие на белых крыс, что подтверждается увеличением концентрации креатинина и мочевины в плазме крови (нефротоксическое действие), а также изменением активности всех исследованных ферментов Концентрация креатинина плазмы в крови крови (гепатотоксическое значительно действие). повышалась у всех экспериментальных животных, получавших соли кадмия, свинца и ртути на 117,3%, 62,4% и 91,6% соответственно. Нефротоксическое действие солей кадмия, свинца и ртути подтверждалось увеличением концентрации мочевины на 48,7%, 94,9%, 33,3% по сравнению с контрольной группой животных. Отравление крыс сульфатом кадмия сопровождалось увеличением концентрации холестерина на 26,9%, тогда как отравление нитратами свинца и ртути, напротив, приводило к достоверному снижению содержания холестерина в плазме крови на 30,8% и 23,1% соответственно. При отравлении нитратом свинца наблюдалось снижение концентрации глюкозы на 26,5% по сравнению с контролем. Содержание общего белка в плазме крови крыс при отравлении солями тяжелых металлов изменялось незначительно, а концентрация альбумина не изменялась по сравнению с контролем (таблица 15). Токсическое действие исследованных солей тяжелых металлов на крыс также подтверждалось изменением активности ферментов плазмы крови. Например, при отравлении сульфатом кадмия достоверно увеличивалась активность ферментов АсАТ (на 78,3%), амилазы (на 26,5%), ГГТ (на 41,4%) и ЛДГ (на 36,9%). При отравлении нитратом свинца было выявлено снижение активности ферментов АлАТ (на 47,8%) и ГГТ (на 46,9%), а также увеличение активности АсАТ (на 57,4%), амилазы (на 28,1%) и щелочной фосфатазы (на 33,9%). 158 Таблица 15. Биохимические показатели крови экспериментальных животных при подостром отравлении солями тяжелых металлов Показатель Единицы измерения Контроль (n=12) CdSO4 (n=10) Pb(NO3)2 (n=10) Hg(NO3)2 (n=10) ммоль/л 3,4±0,3 2,9±0,2 2,5±0,2* 3,5±0,15 Общий белок г/л 69,3±3,4 78,5±6,7 76,2±4,2 76,4±4,3 альбумин г/л 32,5±2,4 36,6±2,8 31,4±3,1 31,3±2,7 креатинин мкмоль/л 35,9±2,2 78,3±4,2* 58,3±4,9* 68,8±4,8* мочевина ммоль/л 3,9±0,3 5,8±0,3* 7,6±0,4* 5,2±0,3* холестерин ммоль/л 1,3±0,1 1,7±0,1* 0,9±0,1* 1,0±0,1* АлАТ Ед/л 112,8±6,7 115,4±17,6 58,9±6,1* 121,2±8,4 АсАТ Ед/л 98,3±2,8 175,3±13,6 * 154,7±15,2* 133,3±11,4* амилаза Ед/л 1397,2± 54,5 1767,4±78,8* 1789,2±112,3* 1328,5±57,8 Щелочная Ед/л 5,9±0,3 6,7±0,4 7,9±0,5* 6,3±0,5 ГГТ Ед/л 14,5±0,9 20,5±1,4* 7,7±0,2* 18,7±1,8 ЛДГ Ед/л 269,1±13,5 368,3±12,3* 296,2±15,6 259,4±11,6 фосфатаза * - р<0,05 158 Глюкоза 159 Отравление крыс нитратом ртути сопровождалось увеличением активности только фермента АсАТ на 35,6% по сравнению с контролем (таблица 15). Несмотря на то, что все использованные соли тяжелых металлов обладали как нефротоксическим, так и гепатотоксическим действием, наибольший сдвиг в биохимических показателях крови крыс наблюдался при ведении per os солей свинца и кадмия. Результаты, представленные в таблице 16, показывают, что при использовании халькогенорганических препаратов в качестве антитоксикантов нормализовались некоторые биохимические показатели крови у крыс при подостром отравлении сульфатом кадмия. CdSO4 оказывал значительное нефротоксическое действие, что выражалось в увеличении концентрации креатинина в плазме крови на 118,1% по сравнению с контролем. Все исследованные препараты оказывали нефропротекторное действие, т.к. при их использовании в качестве антитоксикантов нормализовалась концентрация креатинина в плазме крови экспериментальных животных. Однако в результате антитоксического действия всех исследованных препаратов концентрация креатинина осталась повышенной на 42,1% (соединение 1), 39,3% (соединение 2), 44,0% (соединение 3), 40,7% (соединение 4), 69,1% (соединение 5) и на 35,4% (соединение 6) по сравнению с контролем. При этом концентрация мочевины осталась повышенной при действии препаратов 4 (на 46,2%), 5 (на 34,6%) и 6 (на 41,0%) по сравнению с контролем. Таким образом, наиболее эффективное нефропротекторное действие при отравлении сульфатом кадмия оказывали препараты 1, 2 и 3. 160 Таблица 16. Биохимические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении сульфатом кадмия Показатель Единицы Контроль CdSO4 измерения (n=12) (n=10) Халькогенорганические соединения + CdSO4 Глюкоза ммоль/л 3,4±0,3 2,9±0,2* 1 + CdSO4 (n=10) 2,5±0,3* 2 + CdSO4 (n=10) 1,2±0,1* 3 + CdSO4 (n=10) 3,4±0,2 4 +CdSO4 (n=10) 2,1±0,2* 5 + CdSO4 (n=10) 3,1±0,1 6 + CdSO4 (n=10) 1,6±0,3* Общий г/л 69,3±3,4 78±6,7 72,5±2,9 72,2±4,3 73±3,8 78,2±5,7* 81,3±6,7 74,5±3,7 альбумин г/л 32,5±2,4 36,6±2,8 27,2±2,7* 26±2,2* 25,5±1,6 31,4±1,8 29,2±2,1 31,6±2,4 креатинин мкмоль/л 35,9±2,2 78±4,2* 51±4,2* 50,2±2,9* 51,7±3,6* 50,5±3,6* 60,7±4,3* 48,6±3,7* мочевина ммоль/л 3,9±0,3 5,8±0,3* 4,6±0,6 4,4±0,4 5,3±0,5 5,7±0,3* 5,25±0,2* 5,5±0,2* холестерин ммоль/л 1,3±0,1 1,7±0,1* 1,12±0,1 0,9±0,04* 0,88±0,02* 1,2±0,1 1,2±0,1 1,3±0,2 АлАТ Ед/л 112,8±6,7 115±17,6 132,2±18,3 45,6±4,9* 177,5±8,8* 87,9±6,6* 89,9±10,5* 107,1±11,5 АсАТ Ед/л 98,3±2,8 175±13,6 * 267,1±12,7* 176,4±14,5* 332,1±20,5* 188,4±12,6* 184,3±9,9* 173,2±10,6* амилаза Ед/л 1397,2± 1767± 1696,5± 861,3± 1267,7± 1389,2± 1518,4± 1369,7± 54,5 78,8* 216,3* 52,5* 109,9 128,9 118,6 98,7 Ед/л 5,9±0,3 6,7±0,4 9,3±0,8* 9,6±1,7* 15,6±1,8* 6,7±0,4 7,3±0,5* 5,8±0,4 ГГТ Ед/л 14,5±0,9 20,5±1,4* 5,3±0,2* 26,2±2,6* 10,3±1,1* 9,4±0,1* 8,4±0,9* 6,3±0,1* ЛДГ Ед/л 269,1± 368± 476,3± 423,5± 478,9± 569,6± 577± 595± 13,5 12,3* 49,5* 125,6* 28,9* 22,5* 27,8* 39,4* белок фосфатаза * - р<0,05 160 Щел, 161 Однако превентивное введение per os исследованных соединений 1-6 при интоксикации сульфатом кадмия также наблюдалось снижение концентрации глюкозы в плазме крови на 26,5% (соединение 1), 64,7% (соединение 2), 32,3% (соединение 4) и 52,9% (соединение 6), а также снижение содержания холестерина на 32,8% (соединение 2) и 34,3% (соединение 3) по сравнению с контролем. Вместе с тем содержание в плазме крови общего белка и альбумина изменялось незначительно. Приведенные выше данные свидетельствуют в пользу токсического поражения печени при совестном воздействии на организм экспериментальных животных сульфата кадмия и препаратов 2, 3 и 4. Гепатотоксический эффект при совместном использовании халькогенорганических препаратов и сульфата кадмия также подтверждается изменением активности всех исследованных ферментов в плазме крови (таблица 16). Например, активность АлАТ в плазме крови достоверно снижалась на 59,6% (соединение 2), 22,1% (соединение 4) и 20,3% (соединение 5), а при действии препарата 3, напротив, увеличивалась на 57,4% по сравнению с контролем. Активность АсАТ также достоверно возрастала на 171,6% (соединение 1), 79,0% (соединение 2), 237,7% (соединение 3), 91,3% (соединение), 87,2% (соединение 5) и 75,9% (соединение 6) по сравнению с контролем. Поражение печени также подтверждается увеличением активности щелочной фосфатазы в плазме крови на 57,6% (соединение 1), 62,7% (соединение 2), 164,4% (соединение 3) и 23,7% (соединение 5), а также увеличением активности ЛДГ на 76,9% (соединение 1), 57,4% (соединение 2), 77,9% (соединение 3), 111,7% (соединение 4), 114,6% (соединение 5) и 121,2% (соединение 6) по сравнению с контролем. Активность ГГТ в этом эксперименте в большинстве случаев снижалась на 36,6% (соединение 1), 28,9% (соединение 3), 35,2% (соединение 4), 42,1% (соединение 5) и 56,6% (соединение 6) по сравнению с контролем и лишь при предварительном введении препарата 2 активность ГГТ увеличивалась на 79,3%. Активность амилазы в 162 плазме крови достоверно возрастала на 21,4% при предварительном введении препарата ДАФС и снижалась на 38,4% при действии соединения 2. Изменение активности печеночных ферментов (ГГТ, АсАТ, ЛДГ и щелочной фосфатазы) в плазме крови может быть результатом поражения паренхимы печени вследствие совместного действия халькогенорганических препаратов и сульфата кадмия, поскольку атомы серы, селена и теллура могут образовывать нерастворимые соединения (селениды, сульфиды и теллуриды кадмия соответственно), которые откладываются в гепатоцитах, оказывая долгосрочное токсическое действие. Значительное увеличение активности щелочной фосфатазы, вероятно, является следствием совместного токсического действия сульфата кадмия и препарата 3. Как известно, фтор является самым электроотрицательным элементом и мощным окислителем. Поэтому возможно участие атомов фтора в образовании токсичных фторидов кадмия, оказывающих гепатотоксическое действие. Помимо этого, халькогенорганические препараты могут оказывать гепатотоксический эффект вследствие гидрофобных свойств своих молекул. Таким образом, из изученных халькогенорганических соединений наилучшими антитоксическими свойствами при отравлении сульфатом кадмия обладали препараты 2 и 4, т.к. при их использовании при подостром отравлении сульфатом кадмия нормализовалась концентрация креатинина в плазме крови и не наблюдалось значительного изменения активности печеночных ферментов, меньшую активность проявляло соединение 1 (ДАФС-25), а препараты 3, 5 и 6 демонстрировали незначительное антитоксическое действие при подостром отравлении сульфатом кадмия (таблица 16). Таблица 17 демонстрирует результаты антитоксической активности халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении Pb(NO3)2. Нитрат свинца оказывал значительное токсическое действие на организм экспериментальных животных, поражая, главным образом, почки и 163 печень. О нефротоксическом действии соли свинца свидетельствует увеличение в крови концентрации креатинина в плазме крови на 62,4% и мочевины на 94,9%. Гепатотоксическое действие характеризуется снижением активности фермента ГГТ на 46,9% и увеличением активности щелочной фосфатазы на 33,9% по сравнению с контролем. Пять из шести исследованных халькогенорганических препаратов (кроме соединения 6) оказывали нефропротекторное действие при подостром отравлении крыс нитратом свинца. Наиболее выраженными антитоксическими свойствами обладало соединение ДАФС, поскольку при его предварительном введении нормализовалась концентрация креатинина и мочевины в крови. Антитоксический эффект препаратов 2, 4, 5 и 3 был менее выраженным, т.к. при превентивном введении этих препаратов нормализовалась только концентрация мочевины в крови, а концентрация креатинина в плазме крови оставалась повышенной на 40,9% (соединение 2), 42,9% (соединение 4), 38,1% (соединение 5) и 45,4% (соединение 3) соответственно по сравнению с контрольной группой (таблица 17). Превентивное введение теллурсодержащего соединения 6 при отравлении солью свинца сопровождалось увеличением концентрации креатинина на 51,5% и мочевины – на 58,9% по сравнению с контролем. Все исследованные препараты не были эффективными гепатопротекторами при подостром отравлении нитратом свинца, поскольку применение препаратов 1, 2, 4 и 5 сопровождалось снижением активности фермента ГГТ на 61,4%, 44,8%, 50,3% и 42,1% соответственно по сравнению с контролем, а использование препаратов 3 и 6 характеризовалось значительным увеличением активности печеночных ферментов в крови экспериментальных животных. Активность щелочной фосфатазы возрастала на 144,1% (соединение 3) и 103,4% (соединение 6) соответственно и активность АлАТ увеличивалась на 119,2% (соединение 6) по сравнению с контролем (р<0,05). 164 Таблица 17. Биохимические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении нитратом свинца Показатель Контроль (n=10) Pb(NO3)2 (n=10) Глюкоза Общий белок альбумин креатинин мочевина холестерин АлАТ ммоль/л г/л 3,4±0,3 69,3±3,4 г/л мкмоль/л ммоль/л ммоль/л Ед/л АсАТ Ед/л амилаза Ед/л ЩФ ГГТ ЛДГ Ед/л Ед/л Ед/л 32,5±2,4 35,9±2,2 3,9±0,3 1,3±0,1 112,8± 6,7 98,3± 2,8 1397,2± 54,5 5,9±0,3 14,5±0,9 269,1± 13,5 * - р<0,05 2,5±0,2* 76,2±4,2* 1+Pb(NO3)2 (n=10) 1,7±0,1* 71,4±3,5 Халькогенорганические соединения + Pb(NO3)2 2 +Pb(NO3)2 3+Pb(NO3)2 4+Pb(NO3)2 5 +Pb(NO3)2 6+Pb(NO3)2 (n=10) (n=10) (n=10) (n=10) (n=10) 1,2±0,1* 2,5±0,2* 1,1±0,1* 2,2±0,2* 1,1±0,1* 75,3±4,3 75,7±3,8 75,3±4,3 72,5±6,8 78,6±5,3* 31,4±3,1 58,3±4,9* 7,6±0,4* 0,9±0,1* 58,9± 6,1* 154,7± 15,2* 1789,2± 112* 7,9±0,5* 7,7±0,2* 296± 15,6 26,7±1,7* 44,7±3,8* 4,7±0,2 1,3±0,1 133,6± 14,7* 169,3± 9,85* 1816,6± 76,3* 5,5±0,3 5,6±0,5* 413,5± 38,4* 30,6±1,9 50,6±4,2* 4,7±0,2 1,2±0,1 92,6± 13,3* 126,7± 6,67* 1242,4± 93,7* 7,2±0,3* 8,2±0,3* 346,4± 25,9* 31,4±2,2 52,2±3,4* 3,8±0,2 1,4±0,1 128,2± 10,6* 198,2± 12,6* 1847,8± 138,9* 14,4±0,7* 14,2±1,8 461,3± 49,6* 30,4±1,7 51,3±3,6* 4,1±0,2 1,8±0,2* 126,6± 11,4* 199,4± 15,6* 1366,7± 82,3 7,2±0,2* 7,2±0,5* 468,4± 35,6* 29,8±1,7 49,8±3,7* 4,9±0,2 1,3±0,1 161,2± 11,5* 168,3± 9,7* 1446,7± 55,6 7,1±0,3* 8,4±0,6* 373,6± 21,4* 30,2±2,1 54,4±3,3* 6,2±0,3* 1,5±0,1 247,2± 23,6* 234,2± 13,7* 1623,5± 38,9* 12,3±0,9* 13,6±1, 653,7± 38,9* 164 Единицы измерения 165 Кроме того, отмечалось увеличение активности АсАТ и ЛДГ почти во всех исследованных группах животных при отравлении нитратом свинца. Активность АсАТ увеличивалась на 72,2% (соединение 1), 28,9% (соединение 2), 101,6% (соединение 3), 102,9% (соединение 4), 71,2% (соединение 5) и 138,3% (соединение 6) по сравнению с контролем. Активность ЛДГ увеличивалась на 53,5% (соединение 1), 28,6% (соединение 2), 71,4% (соединение 3), 73,9% (соединение 4), 38,7% (соединение 5) и 142,8% (соединение 6) соответственно по сравнению с контролем (таблица 17). Кроме того, о поражении печени экспериментальных животных в данном эксперименте свидетельствует снижение концентрации глюкозы в сыворотке крови на 50,0% (соединение 1), 64,7% (соединение 2), 26,5% (соединение 3), 67,7% (соединение 4), 35,3% (соединение 5) и 67,7% (соединение 6) по сравнению с контролем. Изменения концентрации общего белка, альбумина и холестерина в сыворотке крови были незначительны. Следовательно, изученные халькогенорганические препараты обладали гепатотоксическим действием при отравлении солью свинца. Причем наибольший токсический эффект оказывали препараты 3 и 6. Вероятно, их токсичность была обусловлена наличием в молекулах атомов фтора и теллура соответственно. Таким образом, наибольшей антитоксической активностью при подостром отравлении крыс нитратом свинца обладало соединение 1 (ДАФС-25), тогда как препараты 2, 4 и 5 оказывали менее выраженное антитоксическое действие; соединение 3 демонстрировало незначительный антитоксический эффект, а теллурорганический препарат 6 не оказывал антитоксического действия. Результаты антитоксической активности халькогенорганических соединений при интоксикации нитратом ртути представлены в таблице 18. Из литературных данных известно о нефротоксическом действии солей ртути. 166 Таблица 18. Биохимические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении нитратом ртути Показатель Контроль (n=12) Глюкоза Общий белок альбумин креатинин мочевина холестерин АлАТ АсАТ ммоль/л г/л 3,4±0,3 69,3±3,4 3,5±0,2 76,2±4,3* 1+ Hg(NO3)2 (n=10) 2,9±0,2 54,8±4,6* г/л мкмоль/л ммоль/л ммоль/л Ед/л Ед/л 32,5±2,4 35,9±2,2 3,9±0,3 1,3±0,1 112,8±6,7 98,3±2,8 31,3±2,7 68,8±4,8* 5,2±0,3* 1,0±0,1* 121±8,4 133,3±11,4* 25,6±0,2* 41,8±3,5* 2,9±0,2* 0,9±0,1* 87,9±6,7* 82,2±6,3* амилаза Ед/л ЩФ ГГТ ЛДГ Ед/л Ед/л Ед/л 1397,2± 54,5 5,9±0,3 14,5±0,9 269,4± 13,5 1328,2± 57,8 6,3±0,5 18,7±1,8 259,4± 11,6 783,3± 36,7* 6,2±0,4 7,4±0,3* 395,6± 25,6* * - р<0,05 Hg(NO3)2 (n=10) Халькогенорганические соединения + Hg(NO3)2 2+ 3+ 4+ 5+ Hg(NO3)2 Hg(NO3)2 Hg(NO3)2 Hg(NO3)2 (n=10) (n=10) (n=10) (n=10) 2,2±0,1* 3,2±0,5 1,5±0,2* 3,1±0,2 79,3±4,6* 75,8±6,5 84,8±6,2* 62,7±3,7 6+ Hg(NO3)2 (n=10) 4,1±0,3 69,6±2,9 31,3±2,4 69,6±3,7* 6,1±0,3* 0,9±0,04* 81,8±8,3* 153,1±13,3 * 1582,3± 78,9* 7,6±0,4* 12,6±0,9 779,2± 45,6* 34,4±2,9 60,7±4,6* 5,1±0,3 0,5±0,04* 87,3±6,3* 154,5±10,7 * 1039,3± 58,9* 4,7±0,3* 3,8±0,3* 214,4± 15,6* 31,9±1,9 66,3±4,4* 3,6±0,3 0,8±0,02* 127,8±8,9* 203,4±12,6 * 1155,6± 88,7* 14,7±0,6* 12,7±0,8 631,3± 35,7* 33,9±2,3 79,6±4,9* 5,4±0,3* 0,9±0,1* 127,3±13,3* 166,5±9,8* 1767,7± 105,6* 5,8±0,3 13,6±0,9 798,4± 37,9* 31,3±2,9 42,5±2,7* 4,5±0,3 1,1±0,1* 102,1±12,4 129,3±10,9 * 1201,2± 78,8* 5,1±0,3* 8,2±0,6* 306,3± 27,6* 166 Единицы измерения 167 В нашем эксперименте введение per os раствора нитрата ртути сопровождалось увеличением концентрации креатинина в крови на 91,6% и мочевины – на 33,3%. Из исследованных халькогенорганических соединений только два препарата (1 и 5) оказывали значительное антитоксическое действие при подостром отравлении нитратом ртути. Остальные препараты были не эффективны (таблица 18). Максимальный нефропротекрорный эффект при отравлении солью ртути продемонстрировали соединения 1 и 5, поскольку при их предварительном введении per os практически нормализовалась концентрация креатинина и мочевины в крови (можно отметить лишь незначительные колебания уровня этих метаболитов в крови по сравнению с контролем). Применение серосодержащего соединения 5 в качестве антитоксиканта при отравлении солью ртути сопровождалось снижением активности фермента печени ГГТ на 44,8%. Колебания остальных биохимических показателей сыворотки крови были минимальны. Вместе с тем, применение соединения 1 сопровождалось изменением активности ряда ферментов: активность амилазы и ГГТ снижалась на 43,9% и 48,9% соответственно, а активность ЛДГ возрастала на 47,0% по сравнению с контролем. Изменение концентрации остальных показателей крови были незначительны (таблица 18). Применение препаратов 2 и 4 сопровождалось увеличением концентрации креатинина на 93,9% и 121,7% соответственно, увеличением концентрации мочевины на 56,4 % и 38,5%, а также увеличением активности ферментов: активность ЛДГ увеличивалась на 189,6% и 196,7% соответственно по сравнению с контролем; активность АсАТ возрастала на 55,7% (соединение 2) и 69,4% (соединение 4) соответственно; активность АлАТ увеличивалась на 27,5% (соединение 2) и 42,9% (соединение 4). Использование препаратов 2 и 4 также сопровождалось незначительным увеличением активности амилазы на 13,2% и 26,5% по сравнению с контрольной группой. Помимо вышеназванного, можно отметить также достоверное снижение концентрации 168 глюкозы на 35,3% (соединение 2) и 55,9% (соединение 4) и концентрации холестерина – на 30,8% (соединения 2 и 4) по сравнению с контролем. Наибольшие изменения биохимических показателей крови у крыс в этом эксперименте вызывали препараты 3 и 6: концентрация креатинина возрастала на 84,7% и 69,1% соответственно, концентрация холестерина, наоборот, уменьшалась на 38,5% и 61,5% соответственно, активность АсАТ увеличивалась на 134,4,5% и 57,2% соответственно по сравнению с контролем (таблица 18). Кроме того, применение препарата 3 в качестве антитоксиканта при отравлении солью ртути приводило к увеличению активности ферментов щелочной фосфатазы на 149,2% и ЛДГ – на 134,6%, а использование соединения 6 сопровождалось уменьшением активности амилазы на 25,6% и ГГТ – на 73,8% по сравнению с контролем (р<0,05). Следует обратить внимание на снижение активности ГГТ при предварительном ведении per os препаратов 1, 5 и 6 на 48,9%, 44,8% и 73,8% соответственно по сравнению с контрольной группой. Как уже говорилось выше, атомы халькогенов (серы, селена и теллура) могут образовывать нерастворимые соединения с солями тяжелых металлов (кадмия, свинца и ртути) – сульфиды, селениды и теллуриды соответственно, которые, накапливаясь в печени, могут оказывать гепатотоксическое действие. Кроме того, известно, что токсичность халькогенов возрастает в направлении сера > селен > теллур, что подтверждается характерным снижением активности ГГТ: 44,2% > 48,1% > 72,7% соответственно. Следовательно, соединение 6 обладало минимальной антитоксической активностью вследствие собственной токсичности. Таким образом, лучшими антитоксическими свойствами при подостром отравлении нитратом ртути обладали соединения 1 и 5. Остальные халькогенорганические препараты были неэффективны при отравлении солью ртути. 169 Оценивая результаты антитоксической активности халькогенорганических соединений при отравлении солями тяжелых металлов, был сделан вывод об эффективности четырех из шести соединений: 1, 2, 4 и 5. При этом препараты 1 и 5 эффективно предотвращали токсическое действие солей свинца и ртути на организм экспериментальных животных. Соединения 2 и 4 оказывали нефропротекторное действие при отравлении солями кадмия и свинца. Антитоксические свойства исследованных соединений убывали в направлении: 1 > 4 > 5 > 2 > 3 > 6. Соединения 3 и 6 не были эффективными антитоксикантами и оказывали гепатотоксическое действие. Токсичность этих препаратов обусловлена наличием в их структурах атомов фтора и теллура соответственно. Фтор – самый электроотрицательный элемент и самый мощный окислитель. Кроме того, атом фтора, вероятно, мог участвовать в образовании токсичных фторидов металлов, которые благодаря своей нерастворимости осаждались в гепатоцитах и оказывали токсическое действие на печень. Атом теллура более токсичен, теллуроорганического чем соединения сера в и печени селен, поэтому приводила к аккумуляция значительному токсическому эффекту. Гепатотоксическое действие соединений 3 и 6 подтверждалось изменением активности печеночных ферментов: увеличением активности АсАТ, АлАТ, ЛДГ, щелочной фосфатазы и снижением активности ГГТ. Препарат 2 обладал сравнительно слабой антитоксической активностью, которая заключалась в нормализации концентрации креатинина в крови при отравлении солями свинца и кадмия; антитоксическое действие этого препарата уступало действию соединений 1, 4 и 5. Таким образом, свидетельствовал экспериментальных о проведенный биохимический анализ токсическом действии солей животных и антитоксической об халькогенорганических препаратов 1, 2, 4 и 5. тяжелых крови металлов на активности 170 4.2. Оценка антитоксической активности халькогенорганических препаратов по результатам гематологических показателей крови крыс Токсическое действие тяжелых металлов на организм приводит к инактивации ряда ферментов, нарушению метаболических процессов, усилению пероксидации липидов, накоплению токсичных продуктов (мочевина, креатинин, активные формы кислорода, продукты ПОЛ). Все перечисленные явления приводят к развитию экзогенной и эндогенной интоксикации. Развитию интоксикации противостоят детоксицирующие системы организма, обеспечивающие химическое превращение токсичных веществ в менее токсичные или растворимые вещества, поддающиеся выведению из организма (окисление, гидролиз, метилирование, восстановление и образование парных соединений и другие процессы). Эти процессы происходят с участием ферментных систем организма, обеспечивающих обезвреживание и метаболические превращения ядовитых веществ. Яды, поступающие в кровь, через воротную вену попадают в печень, являющуюся мощным детоксицирующим барьером, и частично в ней нейтрализуются. Интоксикация развивается при поступлении в организм токсичных веществ в количестве, превышающем возможности детоксицирующих систем или при их неполноценности. Предварительное введение в организм экспериментального животного антитоксиканта (например, халькогенорганического препарата) позволяет обезвредить ион тяжелого металла, активировать ферменты антиоксидантной защиты и, как следствие, снизить тяжесть интоксикации. В клинической практике для оценки эндотоксикации используют различные лейкоцитарные индексы интоксикации (ЛИИ). Одним из показателей, характеризующих степень выраженности эндогенной интоксикации, считается лейкоцитарный индекс интоксикации, который 171 рассчитывается по формуле Я.Я. Кальф-Калифа и отражает остроту воспаления в организме и реакцию на эндогенную интоксикацию [Кальф-Калиф Я.Я., 1941]. Формула для расчета ЛИИ по формуле Я.Я. Кальф-Калифа имеет следующий вид: (4 • миел. + 3 • юн. +2 пал. + 1 сегм.) (пл.+1) ЛИИ = -----------------------------------------------------------------, (мон. + лимф.) (эоз. +1) где: миел. - миелоциты, юн. - юные, пал. - палочкоядерные, сегм. - сегментоядерные, пл. - плазматические клетки, лимф. - лимфоциты, мон. - моноциты, эоз. - эозинофилы. Нормальные значения ЛИИ для крыс - от 0,04 до 0,06. Повышение ЛИИ связано с исчезновением эозинофилов, увеличением количества сегментоядерных форм лейкоцитов, плазматических клеток и снижением числа лимфоцитов при воспалительных процессах и отравлении солями тяжелых металлов. Лейкоцитарный индекс Кальф-Калифа учитывает ряд предпосылок, основанных на функциональной принадлежности различных групп клеток, входящих в лейкоцитарную формулу. Другой вариант расчета лейкоцитарного индекса интоксикации был предложен В.К. Островским [Островский В.К. и др., 2006, 2007]: сегм. + пал. + юн. + миел. + пл. ЛИИ = ------------------------------------------мон. + лимф. + эоз. Нормальные значения данного ЛИИ для крыс 0,23 – 0,3. 172 ЛИИ является наиболее простым, доступным и достаточно информативным показателем и отражает степень воспалительного ответа организма на действие экзо- и эндотоксинов. Кроме того, при отравлении солями тяжелых металлов и при эндогенной интоксикации отмечается снижение концентрации гемоглобина, увеличение количества лейкоцитов и нейтрофилов, уменьшение количества эозинофилов и тромбоцитов [Зупанец И.А., 2005]. Поэтому для оценки тяжести отравления солями тяжелых металлов и антитоксической активности халькогенорганических соединений целесообразно определять гематокрит, общее содержание клеток крови (лейкоцитов, эритроцитов и тромбоцитов), количество отдельных форм лейкоцитов крови, а также концентрацию гемоглобина, поскольку известно об ингибировании процесса синтеза гема солями тяжелых металлов. Таблица 19 содержит результаты гематологических показателей крови экспериментальных животных при введении per os халькогенорганических препаратов. Исследуемые халькогенорганические соединения благодаря своей гидрофобной структуре оказывали некоторое токсическое действие на организм экспериментальных животных. Наиболее токсичным являлся препарат 6, т.к. при его поступлении в организм per os наблюдался лейкоцитоз - значительное увеличение в крови числа лейкоцитов на 83,3%, а также увеличение ЛИИ по формуле Я.Я. Кальф-Калифа на 500%, а по формуле В.К. Островского – на 162,5% (р<0,05). Пероральное введение препарата 3 также характеризовалось признаками токсикоза, поскольку наблюдалось незначительное увеличение в крови числа эритроцитов, а также значительное увеличение ЛИИ по формуле Я.Я. КальфКалифа на 325,0% и по формуле В.К Островского – на 112,5%. Токсичность соединения 4 подтверждалась увеличением ЛИИ по формуле Я.Я. Кальф-Калифа на 375,0%. Однако ЛИИ по формуле В.К. Островского увеличивался лишь на 37,5% (р<0,05). 173 Таблица 19. Гематологические показатели крови экспериментальных животных при введении per os халькогенорганических препаратов Показатель Единицы Контроль измерения (n=12) Халькогенорганические препараты 1 (n=10) 2 (n=10) 3 (n=10) 4 (n=10) 5 (n=10) 6 (n=10) 10^3/uL 10,2±0,2 13,7±0,6 12,3±1,1* 8,8±0,5* 11,2±1,1 11,8±1,1 18,7±1,1* эритроциты 10^6/uL 7,1±0,1 8,2±0,4* 7,2±0,3 8,3±0,5* 7,2±0,4 7,8±0,5* 7,7±0,5* Гемоглобин g/L 123,8±6,3 129,4±6,8 120±8,7 132±5,7 119±9,4 127±9,8 128±9,7 гематокрит % 37,1±2,1 38,2±3,2 36,0±2,9 41,3±3,6 41,2 ± 4,3 36,4±3,4 37,2±3,3 тромбоциты 10^3/uL 720,5±49,5 660±39,7 740±33,7 908±68,7* 835±50,5* 735±42,6 807±69,8 Пал/ядерные % 0,14±0,05 0,11±0,04 0,14±0,01 0,25±0,02* 0,75±0,06* 0±0 * 1,3±0,2 * Сегм/ядерные % 16,3±0,2 25,2±1,2* 27,8±1,9* 34,3±4,7* 23,2±1,7 * 18,6±1,6 37,0±2,8* Моноциты % 7,2±0,8 3,1±0,1* 4,2±0,5* 1,1±0,1* 13,1±0,8 * 5,7±0,5* 6,7±0,7 Лимфоциты % 69,6±3,7 63,3±7,9 63,2±4,6 62,6±4,8 60,2±3,8 70,5±4,2 52,2±2,6* Эозинофилы % 4,9±0,8 5,7±0,8 5,4±0,4 1,8±0,2* 0,8±0,1* 4,1±0,3 1,6±0,4* ЛИИ К-К - 0,04±0,01 0,06±0,016 0,06±0,01* 0,18±0,03* 0,19±0,03* 0,05±0,01 0,24±0,04* ЛИИ Остр - 0,24±0,05 0,37±0,05* 0,39±0,04* 0,51±0,12* 0,33±0,05* 0,24±0,05 0,63±0,06* * - р<0,05 Обозначения: Пал/ядерные – палочкоядерные, Сегм/ядерные – сегментоядерные, ЛИИ К-К - ЛИИ по формуле Я.Я. Кальф-Калифа, ЛИИ Остр – ЛИИ по формуле В.К. Островского 173 лейкоциты 174 Такая значительная разница результатов ЛИИ объясняется различиями в расчете (см. формулы выше). Менее токсичным оказалось соединение 2, т.к. его применение сопровождалось увеличением ЛИИ по формуле В.К. Островского на 50,0% (р<0,05), а ЛИИ по формуле Я.Я. Кальф-Калифа – на 58,3% (таблица 19). Еще менее токсичным было соединение 1, поскольку при его введении per os лишь незначительно увеличивалось число эритроцитов (на 15,5%), также наблюдалось увеличение ЛИИ по формуле В.К. Островского на 54,2% (р<0,05), тогда как ЛИИ по формуле Я.Я. Кальф-Калифа соответствовал норме (таблица 20). Остальные гематологические показатели крови при введении соединения 1 (ДАФС-25) также соответствовали норме. Серный аналог препарата ДАФС (соединение 5) не оказывал токсического действия, поскольку при его использовании все исследуемые гематологические показатели крови соответствовали норме (таблица 19). Таким образом, халькогенорганические соединения по степени их токсичности можно расположить в следующей последовательности: 6 > 3 > 4 > 2 > 1 > 5. Соли кадмия, свинца и ртути оказывали выраженное токсическое действие на организм экспериментальных животных, что видно из результатов, представленных в таблице 20. Наибольший сдвиг в гематологических показателях крови наблюдался при отравлении солью свинца, поскольку наблюдалось сокращение числа эритроцитов на 14,4%, уменьшение концентрации гемоглобина на 24,2% и снижение гематокрита на 21,2% (р<0,05). ЛИИ по формуле Я.Я. КальфКалифа, напротив, значительно увеличивался в14 раз по сравнению с контролем, ЛИИ по формуле В.К. Островского возрастал на 383,3%, что свидетельствовало о тяжелой интоксикации организма экспериментального животного. 175 Таблица 20. Гематологические показатели крови экспериментальных животных при подостром отравлении солями тяжелых металлов Показатель Единицы Контроль (n=12) CdSO4 (n=10) Pb(NO3)2 (n=10) Hg(NO3)2 (n=10) измерения 10^3/uL 10,2±0,2 13,7±0,9* 11,4±0,9 14,4±1,3* эритроциты 10^6/uL 7,1±0,1 7,9±0,6 6,1±0,5* 6,8±0,4 Гемоглобин g/L 123,8±6,3 142,4±11,3 93,9±6,7* 117,3±7,7 гематокрит % 37,1±2,1 42,5±2,8* 29,2±1,6* 37,3±1,7 тромбоциты 10^3/uL 720,5±49,5 806,6±32,2* 910,4±72,6 * 873,9±53,7* Пал/ядерные % 0,14±0,05 0,17±0,01 0,13±0,06 0,14±0,03 Сегм/ядерные % 16,3±0,2 40,7±2,6 * 54,3±3,4 * 28,0±2,7* Моноциты % 7,2±0,8 6,7±0,5 8,1±0,5* 7,7±0,6 Лимфоциты % 69,6±3,7 50,0±2,9* 36,8±2,7* 57,5±2,8* Эозинофилы % 4,9±0,8 3,0±0,4* 0,7±0,1* 4,7±0,3 ЛИИ К-К - 0,04±0,01 0,18±0,02* 0,6±0,14 * 0,08±0,01 * ЛИИ Остр - 0,24±0,05 0,67±0,08* 1,16±0,17* 0,43±0,05* * - р<0,05 Обозначения как в таблице 19 175 лейкоциты 176 Отравление сульфатом кадмия также сопровождалось значительными изменениями в картине крови: содержание лейкоцитов увеличивалось на 34,3%, ЛИИ по формуле Я.Я. Кальф-Калифа возрастал на 350,0% и ЛИИ по формуле В.К. Островского – на 179,2% (р<0,05). Меньшее токсическое действие оказывал нитрат ртути. Однако отмечалось увеличение числа лейкоцитов на 41,2%. ЛИИ по формуле Я.Я. Кальф-Калифа увеличивался на 100,0% и ЛИИ по формуле В.К. Островского – на 79,2% (р<0,05). Следовательно, токсичность исследованных солей тяжелых металлов снижается в следующем направлении: Pb(NO3)2 > CdSO4 > Hg(NO3)2 (таблица 20). Согласно результатам таблицы 21, лучшими антитоксическими свойствами при отравлении сульфатом кадмия обладало соединение 1 (ДАФС-25), поскольку при его превентивном введении per os большинство гематологических показателей крови соответствовали норме, за исключением незначительного снижения числа тромбоцитов на 20,2% (р<0,05). Менее эффективными антитоксикантами являлись соединения 4 и 5. Однако, при их использовании ЛИИ по формуле Я.Я. Кальф-Калифа оставался повышенным на 50,0% и 75,0% соответственно (р<0,05) и число лейкоцитов было повышено на 36,3% и 34,3% (р<0,05) соответственно по сравнению с контролем. Кроме того, применение препарата 4 в качестве антитоксиканта при отравлении солью кадмия сопровождалось увеличением числа тромбоцитов на 35,4% (р<0,05). Остальные гематологические показатели крови, включая ЛИИ по формуле В.К. Островского, соответствовали контрольным значениям (таблица 21). Остальные халькогенорганические соединения не оказывали выраженного антитоксического действия. Предварительное введение per os соединения 2 при интоксикации сульфатом кадмия, с одной стороны, характеризовалось нормализацией ЛИИ по формуле В.К. Островского, а, с другой, значительным увеличением ЛИИ по формуле Я.Я. Кальф-Калифа на 500,0% (р<0,05). 177 Таблица 21. Гематологические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении сульфатом кадмия Показатель Единицы Контроль измерения (n=12) Халькогенорганические соединения + CdSO4 1+CdSO4 2+CdSO4 3+CdSO4 4+CdSO4 5+CdSO4 6+CdSO4 (n=10) (n=10) (n=10) (n=10) (n=10) (n=10) 10^3/uL 10,2±0,2 11,2±1,6 9,7±0,7 20,6±1,8 * 13,9±1,2* 13,7±1,6* 9,7±1,2 эритроциты 10^6/uL 7,1±0,1 7,6±0,4 6,0±0,4 4,2±0,4* 7,8±0,4 6,7±0,6 7,5±0,3 Гемоглобин g/L 123,8±6,3 118,1±9,9 87,3±6,3* 74,1±4,7 * 132,6±15,6 116,3±7,7 121,3± 7,6 гематокрит % 37,1±2,1 36,42±3,2 27,9±4,3 24,8±2,6* 40,4±4,6 36,3±3,1 35,8±3,5 тромбоциты 10^3/uL 720,5±49,5 574,9± 606,6± 215,3± 975,6± 695,2± 541,7± 51,7* 31,2* 19,3 * 63,6* 41,8 23,7* 0,14±0,05 0,11±0,05 3,83±0,31* 0,86±0,05* 0,14±0,03 1,83±0,35* 0,86±0,01* Сегм/ядерные % 16,3±0,2 18,0±1,4 11,3±1,2* 31,5±3,3 * 18,9±1,2 19,4±2,8 31,6±3,1 * Моноциты % 7,2±0,8 1,9±0,3* 2,1±0,3* 7,1±0,4 3,3±0,4* 6,3±0,5 1,1±0,2 * Лимфоциты % 69,6±3,7 77,9±6,5 81,9±6,4* 58,2±3,5* 75,0±6,2 67,3±5,2 65,0±3,7 Эозинофилы % 4,9±0,8 2,8±0,3* 0,1±0,04* 3,1±0,4* 3,0±0,3* 3,4±0,3* 1,1±0,3* ЛИИ К-К - 0,04±0,01 0,06±0,01 0,24±0,02* 0,12±0,04* 0,06±0,01* 0,07±0,01 * 0,26±0,05* ЛИИ Остр - 0,24±0,05 0,23±0,02 0,19±0,02 0,49±0,07* 0,25±0,02 0,29±0,03* 0,49±0,08* Пал/ядерные % * - р<0,05 Обозначения как в таблице 19 177 лейкоциты 178 Кроме того, применение препарата 2 сопровождалось отклонениями ряда гематологических показателей крови экспериментальных животных: концентрация гемоглобина снижалась на 29,5%, число тромбоцитов уменьшалось на 15,8% (р<0,05) по сравнению с контролем. Соединения 3 и 6 не были эффективны при подостром отравлении сульфатом кадмия, т.к. при их превентивном введении оставались повышенные ЛИИ. При использовании препарата 3 ЛИИ по формуле Я.Я. Кальф-Калифа увеличивался на 200,0% и по формуле В.К. Островского – на 104,2% (р<0,05). Кроме того, отмечалось значительное увеличение числа лейкоцитов на 102,0%, уменьшение числа эритроцитов на 40,9%, снижение концентрации гемоглобина на 40,2%, сокращение величины гематокрита на 33,2% и уменьшение числа тромбоцитов на 70,1% (р<0,05) по сравнению с контрольной группой. Применение препарата 6 характеризовалось значительным увеличением ЛИИ по формуле Я.Я. Кальф-Калифа на 550,0% и ЛИИ по формуле В.К. Островского – на 104,2%. Помимо этого, отмечалось снижение числа тромбоцитов на 24,8% по сравнению с контролем (р<0,05). Следовательно, наиболее эффективным соединением при подостром отравлении сульфатом кадмия был препарат 1. Менее выраженное антитоксическое действие оказывали соединения 4 и 5. Препараты 2, 3 и 6 не обладали антитоксической активностью при отравлении сульфатом кадмия (таблица 21). Результаты, представленные в таблице 22, демонстрируют, что из исследованных халькогенорганических соединений эффективными антитоксикантами при подостром отравлении нитратом свинца являлись препараты 1, 4 и 5. Предварительное введение препаратов 4 и 5 при интоксикации нитратом свинца сопровождалось нормализацией всех исследованных гематологических показателей крови экспериментальных животных. 179 Таблица 22. Гематологические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении нитратом свинца Показатель Единицы измерения Контроль (n=12) 10^3/uL 10,2±0,2 эритроциты 10^6/uL 7,1±0,1 6,4±0,4 7,7±0,5 7,4±0,8 7,4±0,5 7,1±0,6 7,3±0,4 Гемоглобин g/L 123,8±6,3 96,4±10,5 * 126,7±11,9 98,5±11,3* 133,2±12,4 129,4±8,3 116,5±12,3 гематокрит % 37,1±2,1 31,1±5,5 41,5±6,4 33,8±5,3 39,7±5,3 36,6±4,5 36,4±5,6 тромбоциты 10^3/uL 720,5± 630,8± 43,4* 728,2± 38,8 685,3± 42,5 702,5± 38,2 731,8± 42,3 694,8± 40,5 0,14±0,05 0±0* 1,17±0,02* 1,17±0,21* 0,14±0,05 0±0* 0±0* Сегм/ядерные % 16,3±0,2 25,7±1,8* 21,4±2,1* 44,4±3,6* 17,8±1,6 21,7±2,1 * 39,2±3,2* Моноциты % 7,2±0,8 5,1±0,1* 21,3±2,3* 6,2±0,5 1,2±0,2* 4,2±0,3* 4,2±0,4* Лимфоциты % 69,6±3,7 63,1±5,2 75,2±3,3 48,2±4,2* 74,8±4,3 69,3±5,9 55,6±6,1* Эозинофилы % 4,9±0,8 6,8±0,8* 0±0* 3,2±0,4* 6,7±0,5 * 5,7±0,5* 1,8±0,2 * ЛИИ К-К - 0,04±0,01 0,05±0,01 0,25±0,05* 0,22±0,03 * 0,03±0,01 0,04±0,01 0,22±0,03 * ЛИИ Остр - 0,24±0,05 0,32±0,05 0,24±0,06 0,78±0,06* 0,22±0,03 0,28±0,06 0,63±0,6* 49,5 Пал/ядерные % * - р<0,05 Обозначения как в таблице 19 179 лейкоциты Халькогенорганические соединения + Pb(NO3)2 1+Pb(NO3)2 2+Pb(NO3)2 3+Pb(NO3)2 4+Pb(NO3)2 5+Pb(NO3)2 6+Pb(NO3)2 (n=10) (n=10) (n=10) (n=10) (n=10) (n=10) 13,2±1,2 * 11,4±0,9 15,6±2,2* 8,7±0,9* 9,8±1,1 16,2±1,6* 180 При использовании соединения 1 в качестве антитоксиканта отмечалось увеличение числа лейкоцитов на 29,4% и уменьшение концентрации гемоглобина на 22,1% (р<0,05) по сравнению с контролем. Остальные гематологические показатели, включая ЛИИ, соответствовали контрольным значениям. Применение препарата 2 как антитоксиканта при отравлении солью свинца было не эффективно, т.к. ЛИИ по формуле Я.Я. Кальф-Калифа был значительно увеличен на 525,0% (р<0,05). Остальные показатели, включая ЛИИ по формуле В.К. Островского, соответствовали данным контрольной группы (таблица 22). Препараты 3 и 6 не обладали антитоксической активностью при подостром отравлении нитратом свинца, поскольку применение этих препаратов не сопровождалось нормализацией лейкоцитарной формулы. При использовании соединения 3 в качестве антитоксиканта ЛИИ по формуле Я.Я. Кальф-Калифа увеличивался на 450,0%, а ЛИИ по формуле В.К. Островского – на 225,0% (р<0,05). Кроме того, отмечалось увеличение числа лейкоцитов на 52,9% по сравнению с контролем. Применение соединения 6 в качестве антитоксиканта сопровождалось увеличением ЛИИ по формуле Я.Я. Кальф-Калифа на 450,0% и ЛИИ по формуле В.К. Островского – на 162,5%, а также увеличением числа лейкоцитов на 58,8% (р<0,05) по сравнению с данными контрольной группы. Следовательно, подострое отравление солью свинца могли эффективно предотвращать соединения 5, 4 и 1. Препараты 2, 3 и 6 не оказывали антитоксического действия при интоксикациии Pb(NO3)2. Таблица 23 экспериментальных представляет животных при гематологические использовании показатели крови халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении нитратом ртути. Наиболее эффективным антитоксическим препаратом при подостром отравлении нитратом ртути оказался препарат 5, поскольку все исследованные гематологические показатели крови соответствовали контрольным значениям 181 (таблица 23). Можно только отметить незначительное увеличение числа эритроцитов на 18,3% по сравнению с контролем (р<0,05). При использовании остальных халькогенорганических соединений как антитоксикантов наблюдалось значительное отклонение основных гематологических показателей крови экспериментальных животных. Тем не менее, применение соединений 1 и 2 характеризовалось нормализацией ЛИИ, что подтверждало их антитоксическую активность. Однако следует отметить снижение числа клеток крови: лейкоцитов на 21,6%, эритроцитов на 42,3%, тромбоцитов на 19,4%, а также уменьшение концентрации гемоглобина на 47,5% и гематокрита на 44,2% (р<0,05) по сравнению с контролем при использовании препарата 1 в качестве антитоксиканта. Предварительное введение per os соединения 2 характеризовалось уменьшением числа лейкоцитов на 34,3%, эритроцитов на 21,1%, снижением концентрации гемоглобина на 32,7% и гематокрита на 25,1% по сравнению с контролем (р<0,05). Соединение 4 оказалось неэффективным при отравлении нитратом ртути, поскольку при его использовании в качестве антитоксиканта наблюдалось достоверное снижение ЛИИ по формуле В.К. Островского на 50,0% по сравнению с контролем. Кроме того, отмечалось увеличение числа лейкоцитов на 103,9%, эритроцитов на 47,9%, концентрации гемоглобина на 35,9% и величины гематокрита на 43,7% (р<0,05) по сравнению с данными контрольной группы (таблица 23). Использование препаратов 3 и 6 в качестве антитоксикантов при отравлении солью ртути сопровождалось значительным увеличением ЛИИ по формуле Я.Я. Кальф-Калифа на 275,0% (соединение 3) и в 44 раза (соединение 6) соответственно и увеличением ЛИИ по формуле В.К. Островского на 145,8% (соединение 3) и 641,7% (соединение 6) соответственно по сравнению с контролем. Кроме того, использование соединений 3 и 6 в данном эксперименте сопровождалось увеличением соответственно (р<0,05). числа лейкоцитов на 107,8% и 168,6% 182 Таблица 23. Гематологические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении нитратом ртути Показатель Единицы Контроль измерения (n=12) Халькогенорганические соединения + Hg(NO3)2 1+Hg(NO3)2 2+Hg(NO3)2 3+Hg(NO3)2 4+Hg(NO3)2 5+Hg(NO3)2 6+Hg(NO3)2 (n=10) (n=10) (n=10) (n=10) (n=10) (n=10) лейкоциты 10^3/uL 10,2±0,2 8,0±0,8* 6,7±0,6* 21,2±1,7* 20,8±1,8* 9,1±0,7 27,4±1,9* эритроциты 10^6/uL 7,1±0,1 4,1±0,4* 5,6±0,3* 7,1±0,6 10,5±0,7* 8,4±0,8* 6,4±0,36 Гемоглобин g/L 123,8±6,3 65,0±6,3* 83,3±7,9* 113,4±12,4 168,2±14,3* 132,4±9,9 103,5±11,1 гематокрит % 37,1±2,1 20,7±2,5* 27,8±1,9* 37,3±2,8 53,3±4,3* 37,8±2,6 тромбоциты 10^3/uL 720,5± 580,8± 656,4± 551,2± 664,4± 771,4± 1032,4± 49,5 29,3* 33,6* 26,4* 26,7 43,9 52,7 * 33,1±3,3 % 0,14±0,05 0,86±0,33* 0±0* 0±0* 0±0* 0±0 * 0±0* Сегм/ядерные % 16,3±0,2 17,4±1,9 22,0±1,8* 37,0±2,3* 9,8±1,1* 19,7±1,7 64,8±3,5 * Моноциты % 7,2±0,8 5,2±0,4* 3,2±0,3* 4,8±0,4* 6,2±0,5* 5,9±0,3 * 0,9±0,1* Лимфоциты % 69,6±3,7 70,8±5,6 71,0±6,5 55,2±4,1* 82,9±6,2* 70,2±3,6 35,7±3,1* Эозинофилы % 4,9±0,8 6,17±0,4* 4,2±0,3 3,2±0,02 * 1,1±0,2* 5,1±0,4 0±0 * ЛИИ К-К - 0,04±0,01 0,04±0,005 0,06±0,01 * 0,15±0,02* 0,06±0,005* 0,04±0,001 1,76±0,12* ЛИИ Остр - 0,24±0,05 0,22±0,03 0,28±0,03 0,59±0,01* 0,11±0,02* 0,23±0,03 1,78±0,2* * - р<0,05 Обозначения как в таблице 19 182 Пал/ядерные 183 Следовательно, наиболее халькогенорганическими препаратами эффективными при антитоксическими отравлении нитратом ртути являлись соединения ДАФС-25 (соединение 1), его серосодержащий аналог (соединение 5) и хлорсодержащее производное (соединение 2). Остальные соединения не оказывали антитоксического действия при подостром отравлении солью ртути. Таким образом, гематологические показатели крови экспериментальных животных при использовании халькогенорганических препаратов в качестве антитоксикантов при подостром отравлении солями кадмия, свинца и ртути подтверждают выраженную антитоксическую активность соединений ДАФС25 (соединение 1), его серосодержащий аналог (соединение 5) и ниросодержащее производное (соединение 4) при отравлении сульфатом кадмия и нитратом свинца и соединений 1, 5 и 2 при отравлении нитратом ртути. Суммируя биохимические и гематологические показатели крови крыс при подостром отравлении солями тяжелых металлов, можно сделать вывод о значительной антитоксической активности соединений 1, 4 и 5. Причем антитоксическая активность этих соединений при отравлении сульфатом кадмия убывала в направлении 1 > 4 > 5. Антитоксический эффект этих соединений при отравлении нитратом свинца уменьшался в направлении 4 > 5 > 1. Наконец, при подостром отравлении нитратом ртути были эффективны препараты 1 и 5. В заключение следует отметить изменение антитоксической активности исследованных халькогенорганических соединений в направлении: ДАФС(Se) > ДАФС(S) > ДАФС(NO2)2) > ДАФС(Cl2) > ДАФС(F2) > ДАФС(Те). 184 4.3. Механизм антитоксического действия халькогенорганических препаратов Токсическое действие солей тяжелых металлов обусловлено взаимодействием иона металла с сульфгидрильными группами белков и ферментов, связыванием с атомами азота пуринов и пиримидинов в составе нуклеиновых кислот [Фомина Н.Ю., 2002], ингибированием ряда ферментов, а также формированием активных форм кислорода, которые способны повреждать мембранные структуры клетки [Koedrith P., Seo Y.R., 2011]. В этой связи антитоксическое действие халькогенорганических соединений обусловлено, с одной стороны, связыванием ионов тяжелых металлов, а, с другой, инактивацией радикалов кислорода, образованных под действием тяжелых металлов. Поэтому серо- и селеноорганические соединения обладали выраженной антитоксической активностью. Атом серы в составе сернистого аналога препарата ДАФС-25, вероятно, являлся мишенью для связывания ионов тяжелых металлов, предотвращая их взаимодействие с сульфгидрильными группами белков и ферментов: Кроме того, атом серы менее токсичен по сравнению с атомами селена и теллура, поэтому сероорганический препарат не оказывал значительного гепатотоксического действия. Тем не менее, наилучшими антитоксическими свойствами обладало соединение 1 (ДАФС-25), поскольку этот препарат, вероятно, может поставлять атом селена для образования селеноцистеина, с последующим его включением в состав селенопротеинов и селеноферментов. Главной мишенью для связывания ионов тяжелых металлов является селенопротеин Р плазмы крови, 185 который с помощью своих селеногидрильных групп способен связывать ионы тяжелых металлов, защищая сульфгидрильные группы белков и ферментов: Селенсодержащие антиоксидантные ферменты (глутатионпероксидазы, формиатдегидратаза, пероксидаза нейтрофилов, тиоредоксинредуктаза, йодтирониндейодиназы I,II,III, глицинредуктаза) обезвреживают радикалы, образованные ионами металлов. Кроме того, как предполагается, атом селена в составе молекулы ДАФС сам может связывать активные формы кислорода, предотвращая, тем самым, их токсическое действие на клетки [Кирова Ю.И., 2004]: O R-Sе-R Среди + ROOH изученных антитоксической → R-Sе-R селеноорганических активностью обладало + ROH соединений наименьшей фторсодержащее производное препарата ДАФС-25 (соединение 3). Как известно, фтор является самым мощным окислителем, что предопределило высокую токсичность этого соединения и слабое антитоксическое действие. Лишь теллурорганический препарат 6 не был эффективным антитоксикантом, поскольку атом теллура не включался в состав белков и ферментов. Кроме того, в ряду халькогенов O > S > Se > Te > Po убывают неметаллические свойства и возрастают металлические свойства и токсичность. Поэтому атом теллура в составе теллурорганического соединения 6 проявлял металлические свойства, не связывал ионы тяжелых металлов и не оказывал антитоксического действия при отравлении солями тяжелых металлов. Помимо этого данный препарат, являясь самым токсичным из исследованных 186 соединений, сам оказывал токсический эффект на организм экспериментальных животных. Вышесказанное позволяет сделать вывод, что, антитоксическая активность халькогенорганических соединений зависит от антиоксидантных свойств этих соединений. Соединения 1, 5, 4 и 2 обладали как антиоксидантной, так и антитоксической активностью. А препараты 3 и 6, напротив, проявляли слабые антиоксидантные свойства, обладали собственной токсичностью и поэтому не оказывали антитоксического действия при отравлении солями тяжелых металлов. 187 Глава 5. Антибактериальная активность халькогенорганических соединений 5.1. Антибактериальное действие халькогенорганических соединений на клинические штаммы Staphylococcus aureus Повсеместно увеличивается число инфекций, вызванных патогенными резистентными бактериями. Преобладающими патогенами являются метициллинрезистентные стафилококки (MRSA), мультирезистентные штаммы Pseudomonas aeruginosa, ванкомицин-резистентные энтерококки (VRE) и др. Инфекции, вызванные устойчивыми штаммами, приводят к росту затрат на лечение и высокой летальности в случае назначения неадекватной терапии [Попов Т.В., 2006]. Низкая активность лекарственных препаратов (или полное ее отсутствие) к бактериям обусловлена развитием лекарственной устойчивости к синтетическим антибиотикам или препаратам, полученным из лекарственных растений. В связи с этим, синтез и изучение биологической активности новых антимикробных агентов, к которым бы не развивалась лекарственная устойчивость, является чрезвычайно актуальным в современной биологии и медицине. В настоящее время активно изучается биохимия и фармакология селеноорганических соединений. На протяжении последнего десятилетия установлена противоопухолевая, противовирусная, антимикробная, антиоксидантная, антитоксическая активность органических соединений селена [Holl V. et al, 2000.; Lindenblatt N. et al., 2003;. Sakurai T. et al., 2006]. Например, селеноорганический препарат эбселен обладает антимикробной активностью против грамположительных и грамотрицательных бактерий при низкой концентрации (MIC = 2-32 мкг/мл) [Wójtowicz H. et al., 2004; Chan G. et al., 2007; Pietka-Ottlik M. et al., 2008]. Кроме того, имеются сведения об антимикробной теллуроорганических соединений в отношении грамотрицательных бактерий [Soni D. et al., 2005]. активности грамположительных и 188 В связи с этим значительный интерес представляет изучение антибактериальной активности халькогенорганических соединений – аналогов диацетофенонилселенида (ДАФС-25). В этой главе представлены результаты антимикробного действия халькогенорганических препаратов на клинические штаммы грамположительных (Staphylococcus aureus) и грамотрицательных (Escherichia coli, Pseudomonas aeruginosa) бактерий. В исследовании использовали препарат ДАФС (препарат 1), его хлор- (препарат 2), фтор- (препарат 3) и нитро(препарат 4) производные, а также теллурсодержащий аналог препарата ДАФС25 (препарат 6). Согласно литературным данным [Садеков И.Д., Минкин В.И., 1995], серосодержащие соединения не проявляют антибактериальной активности вследствие высокой прочности связи «С-S», а, следовательно, сера практически не освобождается из своих органических соединений и потому не оказывает антимикробного действия. Кроме того, сравнительное исследование антибактериальной активности серо- и селеноорганических соединений свидетельствует о значительно большей противомикробной активности органических соединений селена, поскольку селеносодержащие соединения в 30-75 раз более токсичны, чем их серные аналоги и поэтому их биологическая активность против ряда бактерий также выше [Nozawa R., 1989]. В связи с этим в данной серии экспериментов не проводилось исследование антибактериальной активности серосодержащего аналога препарата ДАФС (соединение 5). Проведенные исследования по изучению антибактериального действия препарата ДАФС на клинические штаммы золотистого стафилококка, представленные в таблице 24, демонстрируют незначительную антимикробную активность этого препарата. В минимальной незначительно, но концентрации достоверно 0,001 сокращал мг/мл рост препарат колоний ДАФС-25 золотистого стафилококка только при краткосрочной инкубации 30 минут на 17,7% 189 (р<0,05), а при инкубации 150 минут наблюдалось, наоборот, увеличение роста колоний S. aureus на 20,3%. При 30-минутной инкубации препарата 1 в концентрации 0,01 мг/мл с клиническими штаммами золотистого стафилококка отмечалось достоверное (р<0,05), но незначительное уменьшения числа бактериальных колоний на 20,1%, тогда как 150-минутная инкубация приводила, напротив, к повышению числа бактериальных колоний на 24,5% (таблица 24). Таблица 24. Антибактериальное действие соединения 1 на клинические штаммы Staphylococcus aureus Условия эксперимента Время воздействия, мин 30 60 90 120 150 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (ДМФА и (n=10) физ. р-р) 915,3 ± 794,4± 95,5 676,4± 73,2* 731,3± 753,4± 105,6 95,4* 63,3* 842,4 ± 722,2± 237,3 761,3± 158,6 657,7± 891,6± 89,5 235,2 141,2 906,5 ± 609,7± 753,5± 87,3 713,4± 786,3± 150,4 197,5* 227,6 176,4 820,2 ± 139,6 824,5 ± 205,4 710,4± 149,2 595,6± 203,7 718,3± 170,4 925,6± 241,4 938,2± 176,5 1026,6± 236,7* 868,7± 131,3 991,4± 80,3* * - р<0,05 , ** - р< 0,001 Препарат ДАФС-25 в концентрации 0,1 мг/мл при краткосрочной инкубации 30 минут вызывал достоверное подавление роста колоний золотистого стафилококка на 26,1%. Более длительная инкубация этого соединения с бактериальными клетками не приводила к изменению их роста по сравнению с контролем. Наконец, в максимальной концентрации 1 мг/мл и при инкубации 90 минут соединение 1 сокращало число колоний S. aureus на 32,8%. Инкубация этого соединения в других временных интервалах в данной концентрации не 190 сопровождалась достоверным изменением числа бактериальных колоний (таблица 24). Повышение числа бактериальных колоний золотистого стафилококка при длительной инкубации 150 минут с соединением 1 в низких концентрациях 0,01 и 0,001 мг/мл, вероятно, связано с антиоксидантной активностью этого соединения и/или освобождением атома селена, необходимого для доказывают, что жизнедеятельности бактериальных клеток. Результаты, представленные в таблице 25, хлорированный аналог препарата ДАФС-25 (препарат 2) обладал большим антибактериальным действием по сравнению с препаратом ДАФС-25 по отношению к клиническим штаммам золотистого стафилококка. Препарат 2 эффективно снижал рост бактерий золотистого стафилококка в концентрации 0,001 мг/мл при инкубации 60, 90, 120, 150 минут на 31,2%, 52,2%, 70,6% и 91,5% (р<0,05) соответственно. Препарат 2 также проявлял антибактериальную активность в концентрации 0,01 мг/мл, что сопровождалось уменьшением роста бактериальных колоний St. aureus при инкубации 60-120 минут: на 41,7%, 78,4%, 95,3%, а при инкубации 150 минут препарат 2 полностью подавлял рост бактериальных клеток (таблица 25). Обнаружено достоверное снижение роста колоний золотистого стафилококка при инкубации 60-150 минут с препаратом 2 в концентрации 0,1 мг/мл. При этом отмечалось уменьшение роста колоний на 54,4% (60 минут), 59,6% (90 минут), 86,3% (120 минут). При инкубации 150 минут препарат 2 в концентрации 0,1 мг/мл полностью подавлял рост бактерий. Препарат 2 эффективно подавлял рост колоний золотистого стафилококка в концентрации 1 мг/мл при инкубации от 30 до 150 минут: на 52,2% (30 минут), 46,9% (60 минут), 87,9% (90 минут), 99,6% (120 минут) соответственно (р<0,05). При максимальной инкубации 150 минут препарат 2 полностью подавлял рост бактерий золотистого стафилококка. 191 Таблица 25. Антибактериальное действие соединения 2 на клинические штаммы Staphylococcus aureus Время воздействия, мин Условия эксперимента 30 60 90 120 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (n=10) (ДМФА и физ. р-р) 713,4± 341,3± 954,4± 217,6 565,6± 609,3± 152,2 139,3* 110,5 215,4 781,2± 414,4± 356,7± 161,7* 455,6± 537,5± 154,4 146,6** 115,4* 153,4* 787,6± 95,6± 318,6± 170,2± 376,7± 136,6 61,4** 167,7** 70,6** 207,2* 684,7± Нет роста 94,3± 65,6** 32,2± 201,2± 166,7 12,4** 91,3** 719,8± Нет роста 125,6 * - Р<0,05 , ** - Р< 0,001 Нет роста 150 При минимальной инкубации 30 минут Нет роста и при 61,3± 21,4** более низких концентрациях препарата 2 от 0,1 мг/мл до 0,001 мг/мл антимикробной активности препарата не наблюдалось (таблица 25). Таким образом, препарат 2 – хлорированный аналог препарата ДАФС – оказывал достоверно большее антибактериальное действие на клинические штаммы золотистого стафилококка, чем собственно препарат ДАФС-25. Причем антибактериальное действие возрастало по мере увеличения концентрации препарата и времени инкубации. Препарат 2 был эффективен уже при краткосрочном воздействии 30 минут в концентрации 1 мг/мл. С увеличением времени инкубации увеличивался и антибактериальный эффект, достигая максимума при времени инкубации 150 минут, когда полностью подавлялся рост бактерий в концентрациях от 1 мг/мл до 0,01 мг/мл (р<0,05). Таблица 26 содержит результаты антибактериального действия соединения 3 на клинические штаммы Staphylococcus aureus. Фторсодержащий аналог препарата ДАФС-25 (препарат 3) проявлял выраженную антибактериальную активность в широком диапазоне концентраций (от 1 до 192 0,0001 мг/мл) при разном времени инкубации (30-150 минут) в отношении клинических штаммов золотистого стафилококка (таблица 26). В минимальной концентрации 0,0001 мг/мл препарат 3 подавлял рост колоний золотистого стафилококка при инкубации 30-150 минут на 25,4%, 33,6%, 61,4%, 66,4%, 81,7% соответственно по сравнению с контролем. Таблица 26. Антибактериальное действие соединения 3 на клинические штаммы Staphylococcus aureus Условия эксперимента Количество колоний на твердых питательных средах Контроль (n=10) (ДМФА и физ. р-р) Время воздействия, мин 959,9± 42,5 60 866,4 ± 60,3 90 891,2 ± 189,7 120 875,6 ± 170,4 150 904,3± 43,5 * - р<0,05 , ** - р< 0,001 30 Опытные группы, концентрация вещества, мг/мл 1 (n=10) 0,1 (n=10) 0,01 0,001 0,0001 (n=10) (n=10) (n=10) 78,8± 74,4± 24,4** 14,5** 21,3± 3,3± 10,5** 1,1** Нет роста Нет роста 260,4 ± 59,5** 196,6± 94,3* 123,3± 49,7** Нет роста Нет роста 11,2± 9,3** Нет роста Нет роста Нет роста 563,4± 13,5** 432,4± 58,5** 211,2± 76,7** 114,4± 96,7** 13,2± 3,3** 715,6± 204,4* 575,6± 185,8* 344,4± 83,6* 294,2± 176,4* 163,3± 76,2** В концентрации 0,001 мг/мл препарат 3 проявлял более выраженные антибактериальные свойства, снижая рост колоний золотистого стафилококка на 41,3% (30 минут), 50,1% (60 минут), 76,3% (90 минут), 86,9% (120 минут) и на 98,6% (150 минут) соответственно (р<0,001). При более высокой концентрации 0,01 мг/мл препарат 3 значительно подавлял рост колоний S. aureus при разном времени инкубации на 72,9% (30 минут), 77,4% (60 минут), 86,2% (90 минут), 98,7% (120 минут), а при максимальном времени инкубации 150 минут препарат 3 полностью подавлял рост клеток золотистого стафилококка (р<0,001) (таблица 26). 193 Препарат 3 как антибактериальный агент был наиболее эффективен в отношении клинических штаммов золотистого стафилококка в высоких концентрациях 1 и 0,1 мг/мл. В этих концентрациях препарата 3 при инкубации 90-120 минут не наблюдалось роста бактериальных колоний, а при инкубации 30 и 60 минут отмечалось значительное уменьшение числа клеточных колоний на 92,3% (30 минут), 99,7% (60 минут) в концентрации 0,1 мг/мл и на 91,9% (30 минут), 97,6% (60 минут) в максимальной концентрации 1 мг/мл (таблица 26). Следовательно, фторсодержащий аналог препарата ДАФС-25 (препарат 3) оказывал значительное антимикробное действие во всех исследованных концентрациях и краткосрочной инкубации. Антибактериальная активность препарата 3 возрастала с увеличением концентрации препарата и времени инкубации. При этом полное подавление роста колоний золотистого стафилококка отмечалось при концентрациях 1 и 0,1 мг/мл и при длительной инкубации 90-120 минут (таблица 26). Особый интерес представлял препарат 4 – нитропроизводное препарата ДАФС-25, поскольку наличие двух нитрогрупп в мета-положении фенильных радикалов соединения 4 позволяет проводить аналогию с нитрофуранами. Однако селеноорганическое соединение обладало гораздо меньшей антибактериальной активностью, что следует из данных, представленных в таблице 27. Например, препарат 4 обладал антибактериальной активностью только в высоких концентрациях 0,1 и 1 мг/мл, тогда как в малых концентрациях 0,01 и 0,001 мг/мл антибактериальное действие этого препарата было незначительным или недостоверным. Препарат 4 в концентрации 0,1 мг/мл значительно подавлял рост клинических штаммов золотистого стафилококка на 53,3% (30 минут), 69,3% (60 минут), 64,7% (90 минут), 83,9% (120 минут) соответственно (р<0,001). При максимальном времени инкубации 150 минут рост бактериальных клеток не наблюдался. Максимальная концентрация препарата 4 (1 мг/мл) была еще более эффективна в отношении штаммов золотистого стафилококка. При краткосрочной инкубации 30 и 60 минут число бактериальных колоний уменьшалось на 90,6% и 92,8% 194 соответственно (р<0,001). Увеличение времени инкубации до 90, 120 и 150 минут приводило к полному подавлению роста бактериальных клеток. Таблица 27. Антибактериальное действие соединения 4 на клинические штаммы Staphylococcus aureus Время воздействия, мин Условия эксперимента 30 60 90 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (n=10) (ДМФА и физ. р-р) 974,4 ± 92,3± 455,1± 891,3± 1125,6± 90,5 31,1** 62,5** 62,2* 105,5* 956,3 ± 69,7± 294,2± 695,4± 1017,4± 91,2 61,3** 69,5** 137,5* 107,5 960,5 ± Нет 339,8± 903,3± 905,2± 99,3 роста 141,5** 149,4 54,1 942,4 ± Нет 132,5 роста 150 1023,3 ± Нет 148,4 роста * - р<0,05 , ** - р< 0,001 120 151,2± 91,5** Нет роста 876,2± 71,5 976,1± 210,1 1138,2± 87,5 797,6± 213,5 Следовательно, препарат 4 проявлял антибактериальную активность в отношении клинических штаммов золотистого стафилококка лишь в высоких концентрациях 0,1 и 1 мг/мл (таблица 27). Согласно результатам, представленным в таблице 28, теллуроорганический аналог препарата ДАФС-25 (препарат 6) обладал достаточно высокой антибактериальной активностью. В высоких концентрациях этот препарат (1 и 0,1 мг/мл) приводил к полному подавлению роста бактериальных клеток золотистого стафилококка. В концентрации 0,01 мг/мл препарат 6 значительно сокращал число бактериальных колоний на 81,6% при инкубации 30 минут, на 94,4% при инкубации 60 минут и на 94,1% при 90-минутной инкубации (р<0,001). При увеличении времени инкубации до 120 и 150 минут роста бактериальных клеток не наблюдалось. 195 Таблица 28. Антибактериальное действие соединения 6 на клинические штаммы Staphylococcus aureus Условия эксперимента Время воздействия, мин 30 60 90 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 972,3 ± Нет 14,2± 179,4± 379,7± 620,1± 140,1 роста 11,1** 36,4** 93,3** 55,6** 983,3 ± Нет Нет 55,6± 197,4± 674,2± 86,5 роста роста 40,1** 84,5** 164,5** 977,2 ± Нет Нет 58,7± 111,4± 724,2± 86,5 роста роста 46,4** 26,2** 182,3* Нет роста Нет роста Нет роста 50,1± 42,2** 515,6± 271,4* 789,9 ± Нет 84,5 роста * - р<0,05 , ** - р< 0,001 Нет роста Нет роста 16,4± 11,1** 422,4± 168,2** 120 869,4 ± 68,5 150 Препарат 6 был эффективен и при низких концентрациях. Например, при концентрации этого препарата 0,001 мг/мл число бактерий уменьшалось по мере увеличения времени инкубации: на 61,0% (30 минут), 79,9% (60 минут), 88,6% (90 минут), 94,3% (120 минут) и на 97,9% при инкубации 150 минут соответственно по сравнению с контролем (р<0,001). При минимальной концентрации 0,0001 мг/мл препарат 6 достоверно подавлял рост клинических штаммов золотистого стафилококка на 36,2% (30 минут), 31,4% (60 минут), 25,9% (90 минут), 40,7% (120 минут) и на 46,5% при инкубации 150 минут (таблица 28). Следовательно, препарат 6 эффективно подавлял рост клинических штаммов золотистого стафилококка в широком диапазоне концентраций 0,0001-1 мг/мл и при разных режимах инкубации 30-150 минут. При этом антибактериальная активность этого препарата была максимальна в концентрации 0,01-1 мг/мл и возрастала с увеличением времени инкубации. Из исследованных халькогенорганических соединений наибольшей антимикробной активностью в отношении клинических штаммов золотистого 196 стафилококка обладал теллурорганический препарат 6. Высокая антибактериальная активность этого соединения обусловлена токсичностью атома теллура в его составе. Выраженное антимикробное действие также оказывал селеноорганический препарат 3, токсичность которого связана с наличием двух атомов фтора у фенильных заместителей. Хлорсодержащий аналог препарата ДАФС-25 (соединение 2) также эффективно подавлял рост колоний золотистого стафилококка, хотя его активность была заметно слабее, чем у соединений 5 и 3. Соединение 4, которое содержало две нитрогруппы в мета-положении фенильных радикалов оказывало менее выраженный антимикробный эффект по сравнению с соединениями 5, 3 и 2. И, наконец, препарат ДАФС-25 обладал наименьшей антибактериальной активностью из изученных халькогенорганических соединений. Очевидно, что наличие боковых групп у фенильных радикалов препарата ДАФС-25 приводило к увеличению токсичности и, следовательно, антимикробной активности этих соединений. В целом антибактериальная активность исследованных халькогенорганических соединений на клинические штаммы S. aureus убывала в направлении: ДАФС(Те) > ДАФС(F2) > ДАФС(Cl2) > ДАФС(NO2)2) > ДАФС(Se) > ДАФС(S). 5.2. Антибактериальное действие халькогенорганических соединений на клинические штаммы Pseudomonas aeruginosa Препарат ДАФС-25 (соединение 1) оказывал антимикробное действие на клинические штаммы Pseudomonas aeruginosa (Ps. aeruginosa), что выражалось в уменьшении числа колоний по сравнению с контролем (результаты представлены в таблице 29). Причем данный препарат достоверно снижал число колоний синегнойной палочки на 27,03% (р<0,05) даже при низкой концентрации 0,001 мг/мл и при минимальной инкубации 30 минут. При увеличении времени инкубации до 90 и 150 минут наблюдалось снижение 197 роста бактерий на 31,33% и 12,39% (р<0,05). Однако препарат 1 не был эффективен в минимальной концентрации 0,0001 мг/мл. Таблица 29. Антибактериальное действие соединения 1 на клинические штаммы Pseudomonas aeruginosa Время воздействия, мин Условия Количество колоний на твердых питательных средах экспери Контроль Опытные группы, концентрация вещества, мг/мл (n=10) мента 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 30 1332,4± 985,6± 1021,4± 976,3± 972,4± 1332,4± 235,3 102,2* 119,8 87,8* 24,6* 235,3 60 1156,3± 180,3 1059,2± 201,4 971,1± 195,3 966,4± 95,5 983,3± 154,4 1156,3± 180,3 90 983,3 ± 123,3 676,1± 128,4* 880,3± 54,7* 828,8± 93,4 675,2± 203,3* 983,3 ± 123,3 120 912,5 ± 151,2 389,4±16 4,3** 657,4± 206,6* 882,3± 139,4 915,6± 76,2 912,5 ± 151,2 150 880,4 ± 65,6 158,3± 74,5** 476,2± 220,8* 693,4± 83,4* 771,2± 134,4* 880,4 ± 65,6 * - р<0,05 , ** - р< 0,001 Препарат ДАФС-25 в концентрации 0,01 мг/мл эффективно снижал рост колоний на 26,7% при инкубации 30 минут и на 21,3% при инкубации 150 минут (р<0,05). Данная концентрация препарата ДАФС была неэффективна при инкубации 60, 90 и 120 минут. Антибактериальная активность препарата ДАФС-25 в концентрации 0,1 мг/мл была достоверна при инкубации 90, 120, 150 минут, что выражалось в уменьшении числа клеток на 10,5%, 27,9% и 45,9% соответственно по сравнению с контролем (р<0,05). Таким образом, из полученных результатов можно отметить закономерность – антибактериальная активность препарата ДАФС-25 на клинические штаммы Ps. aeruginosa возрастала с увеличением концентрации 198 препарата и с увеличением времени инкубации. При этом максимальная антимикробная активность препарата ДАФС-25 наблюдалась в концентрации 1 мг/мл при инкубации 150 минут. Кроме того, следует отметить, что соединение 1 оказывало больший суммарный антибактериальный эффект на клинические штаммы золотистого стафилококка, чем на синегнойную палочку. Согласно результатам, представленным в таблице 30, препарат 2 также проявлял антимикробную активность в отношении клинических штаммов синегнойной палочки в широком диапазоне концентраций и времени инкубации. Достоверную антибактериальную активность препарат 2 проявлял в концентрации 0,001 мг/мл, подавляя рост бактериальных колоний на 30,1% (60 минут), 66,3% (90 минут), 61,2% (120 минут), 71,4% (150 минут) соответственно по сравнению с контролем (р<0,05). Более выраженную антибактериальную активность препарат 2 проявлял в концентрации 0,01 мг/мл. Число колоний синегнойной палочки достоверно снижалось по сравнению с контролем при инкубации 30, 60, 90, 120 и 150 минут на 17,6%, 24,8%, 83,9%, 79,1%, 97,9% соответственно (р<0,05). В концентрации 0,1 мг/мл препарат 2 эффективно уменьшал число колоний Ps. aeruginosa при инкубации от 30 до 150 минут на 22,5%, 41,1%, 75,2%, 92,9%, 97,8% (р<0,05) соответственно по сравнению с контролем (таблица 30). Наибольшую антимикробную активность препарат 2 проявлял в максимальной концентрации 1 мг/мл. В зависимости от времени инкубации рост колоний снижался на 40,0% (30 минут), 61,1% (60 минут), 88,9% (90 минут), 98,3% (120 минут), а при инкубации 150 минут препарат 2 полностью подавлял рост бактериальных клеток синегнойной палочки (р<0,001). Следовательно, препарат 2 обладал значительным антибактериальным действием в отношении клинических штаммов синегнойной палочки при инкубации от 60 до 150 минут в концентрациях от 1 до 0,001 мг/мл. Следует 199 отметить увеличение антибактериального действия препарата 2 с увеличением времени инкубации и концентрации препарата. Таблица 30. Антибактериальное действие соединения 2 на клинические штаммы Pseudomonas aeruginosa Условия эксперимента Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (n=10) (n=10) (ДМФА и Время воздействия, мин физ. р-р) 30 60 90 120 939,8±107, 4 900,6 ± 57,7 1067,8±13 8,5 900,6±62,5 869,4± 76,5 * - р<0,05 , ** - р< 0,001 150 563,3± 728,4± 99,6* 158,3* 350,3± 530,2± 52,5** 169,5** 118,2± 265,4± 89,4** 137,8** 15,6± 64,4± 55,6** 5,2** Нет роста 19,8± 9,2** 774,5± 122,3* 677,8± 192,3* 171,2± 136,2** 188,9± 153,4** 18,2± 14,6** 750,2±248,5 629,4±258,5* 360,4±127,5** 349,2± 203,6* 249,4±188,6** Сравнивая антибактериальное действие препарата 2 на золотистый стафилококк и синегнойную палочку, можно отметить больший антимикробный эффект препарата 2 при минимальном времени инкубации 30 минут в отношении синегнойной палочки. Однако, при максимальном времени инкубации 150 минут отмечалось практически полное подавление роста колоний золотистого стафилококка во всех исследованных концентрациях препарата 2. Таблица 31 содержит результаты антибактериального действия соединения 3 на клинические штаммы Pseudomonas aeruginosa. Препарат 3 в низких концентрациях 0,0001 и 0,001 мг/мл проявлял антибактериальную активность против клинических штаммов синегнойной палочки при инкубации 200 60-150 минут, а при краткосрочной инкубации 30 минут не наблюдалось достоверного снижения числа бактериальных колоний. Таблица 31. Антибактериальное действие соединения 3 на клинические штаммы Pseudomonas aeruginosa Условия эксперимента Время воздействия, мин 30 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 829,6± 183,5± 197,2± 606,3± 729,4± 688,8± 54,4 141,2** 91,4** 103,5* 166,7 203,4 60 985,6 ± 98,5 25,2± 13,1** 77,8± 34,3** 389,4± 148,5** 693,3± 154,5* 675,6± 190,4* 90 849,2 ± 62,5 Нет роста 15,2± 4,4** 181,2± 64,4** 433,3± 107,3* 348,4± 91,2** 120 805,3 ± 89,5 Нет роста 7,6± 3,3** 73,4± 55,6** 173,3± 50,2** 159,7± 82,5** 150 829,5 ± 51 Нет роста Нет роста 20,4± 8,2** 166,2± 74,6** 104,5± 65,6** * - р<0,05 , ** - р< 0,001 В минимальной концентрации 0,0001 мг/мл препарат 3 достоверно подавлял рост синегнойной палочки на 31,5% (60 минут) (р<0,05), 59,0% (90 минут), 80,3% (120 минут), 87,5% (150 минут) соответственно по сравнению с контролем (р<0,001). В концентрации 0,001 мг/мл препарат 3 также эффективно снижал число бактериальных колоний на 29,6% (60 минут), 48,9% (90 минут) (р<0,05), 78,5% (120 минут), 79,9% (150 минут) соответственно (р<0,001). При более высокой концентрации 0,01 мг/мл препарат 3 оказывал антибактериальную активность при инкубации от 30 до 150 минут, подавляя рост колоний на 26,9% (30 минут) (р<0,05), 60,5% (60 минут), 66,9% (90 минут), 90,9% (120 минут) и на 97,6% (150 минут) (р<0,001) соответственно по 201 сравнению с контролем (таблица 31). При высоких концентрациях препарата 3 (1 и 0,1 мг/мл) наблюдался максимальный антимикробный эффект. Например, при краткосрочной инкубации 30 минут препарат 3 в концентрации 0,1 мг/мл снижал рост бактерий синегнойной палочки на 76,24%, а при увеличении времени инкубации до 60, 90, 120 и 150 минут антибактериальный эффект возрастал до 92,2%, 98,2%, 99,1% соответственно, а при максимальной инкубации 150 минут рост бактерий подавлялся полностью (р<0,001). В максимальной концентрации препарат 3 (1 мг/мл) оказывал наиболее выраженную антибактериальную активность, снижая рост бактерий на 77,9% при 30-минутной инкубации и на 97,5% при инкубации 60 минут (р<0,001). При увеличении времени инкубации до 90, 120 и 150 минут наблюдалось полное подавление роста бактерий синегнойной палочки (р<0,001) (таблица 31). Таким образом, можно сделать вывод о высокой антибактериальной активности препарата 3 в отношении клинических штаммов Ps. aeruginosa в концентрациях от 0,0001 до 1 мг/мл и при инкубации 60-150 минут. Препарат 3 был наиболее эффективен в высоких концентрациях 1 и 0,1 мг/мл и при длительной инкубации 90-120 минут. При этом рост бактерий подавлялся полностью. Однако при краткосрочной инкубации (30 минут) препарат 3 в низких концентрациях 0,001 и 0,0001 мг/мл не обладал достоверной актимикробной активностью по отношению к клиническим штаммам синегнойной палочки. Кроме того, можно заключить, что препарат 3 в малых концентрациях и при минимальном времени инкубации более эффективен против клинических штаммов золотистого стафилококка, чем синегнойной палочки. Результаты антибактериального действия препарата 4 на клинические штаммы Pseudomonas aeruginosa представлены в таблице 32. Клинические штаммы синегнойной палочки были более чувствительны к действию препарата 4, чем золотистый стафилококк, т.к. была обнаружена антибактериальная активность этого препарата в концентрации 0,01 мг/мл. В этой концентрации препарат 4 подавлял рост бактериальных клеток на 34,0% 202 (30 минут), 38,0% (60 минут), 51,2% (90 минут), 35,8% (120 минут) (р<0,001) и на 57,3% (150 минут) соответственно (р<0,05) по сравнению с контролем. Таблица 32. Антибактериальное действие препарата 4 на клинические штаммы Pseudomonas aeruginosa Время воздействия, мин Условия эксперимента Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (ДМФА и (n=10) физ. р-р) 30 1002,2 ± 83,3± 298,2± 661,2± 975,4± 70,3 55,5** 46,5** 109,7** 140,4 60 1034,4 ± 76,2± 278,9± 641,3± 1016,4± 138,7 52,1** 83,4** 128,5** 65,5 90 947,3 ± 9,2± 170,1± 462,5± 180,1* 894,4± 94,5 5,3** 46,5** 50,2 120 977,6 ± Нет 74,5± 627,2± 762,4± 138,5 роста 47,4** 136,4** 125,5* 150 885,2 ± Нет 6,2± 378,2± 259,8* 803,2± 86,5 роста 3,1** 162,5 * - р<0,05 , ** - р< 0,001 Препарат 4 в концентрации 0,1 мг/мл обладал более выраженным антибактериальным действием, что выражалось в уменьшении числа бактериальных колоний на 70,3% (30 минут), 73,1% (60 минут), 82,0% (90 минут), 92,4% (120 минут) и на 99,3% (150 минут) (р<0,001) соответственно по сравнению с контролем. Максимальную антибактериальную активность препарат 4 проявлял в концентрации 1 мл/мл. При инкубации 30, 60 и 90 минут число бактериальных колоний уменьшалось на 91,7%, 92,7% и 99,1% соответственно (р<0,001) по сравнению с контролем. При инкубации 120 и 150 минут рост бактерий подавлялся полностью (р<0,001). Таким образом, препарат 4 оказывал значительное антибактериальное действие на клинические штаммы синегнойной палочки в концентрациях от 0,001 мг/мл до 1 мг/мл. Однако в низкой концентрации (0,001 мг/мл) препарат 4 203 достоверно (р<0,05), но незначительно (на 22,0%) подавлял рост бактериальных колоний лишь при длительной инкубации 120 минут, а при остальных режимах инкубации не наблюдалось достоверного антибактериального эффекта препарата 4 (таблица 32). Антибактериальная активность теллурорганического препарата 6 в отношении клинических штаммов синегнойной палочки была значительно ниже по сравнению с действием на клинические штаммы золотистого стафилококка. Согласно данным таблицы 33, этот препарат не оказывал антимикробного действия в минимальной концентрации 0,0001 мг/мл, а также не вызывал достоверного и значимого снижения роста бактерий в концентрации 0,001 мг/мл при инкубации 30 и 60 минут. Увеличение времени инкубации при данной концентрации препарата приводило к уменьшению числа бактериальных колоний на 29,7% (90 минут), 27,7% (120 минут) и на 26,1% (150 минут) соответственно по сравнению с контролем (р<0,05). Таблица 33. Антибактериальное действие препарата 6 на клинические штаммы Pseudomonas aeruginosa Время воздействия, мин Условия Количество колоний на твердых питательных средах экспери- Контроль Опытные группы, концентрация вещества, мг/мл (n=10) мента 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 30 907,4± 34,3± 74,4± 378,4± 729,4± 1041,4± 80,2 14,2** 40,1** 52,6** 157,5* 207,5 60 862,4 ± 103,5 Нет роста 98,7± 87,6** 321,2± 76,7** 785,4± 115,6 976,3± 100,2 90 880,3 ± 90,2 Нет роста 20,2± 12,1** 319,9± 98,4** 619,2± 206,5* 1082,3± 150,4 120 892,2 ± 98,4 Нет роста Нет роста 270,2± 129,7** 645,3± 135,6* 1066,7± 135,6* 150 879,9 ± 61,5 Нет роста Нет роста 114,3± 56,2** 650,1± 207,5* 947,4± 272,3 * - р<0,05 , ** - р< 0,001 204 В концентрации 0,01 мг/мл препарат 6 оказывал значительное антибактериальное действие при всех режимах инкубации 30-150 минут. При этом число бактериальных колоний уменьшалось по мере увеличения времени инкубации: на 58,3% (30 минут), 62,8% (60 минут), 63,8% (90 минут), 69,7% (120 минут) и на 87,0% (150 минут) соответственно (р<0,001). Более выраженный антибактериальный эффект наблюдался при использовании препарата 6 в концентрации 0,1 мг/мл. Инкубация (30- 90 минут) синегнойной палочки с данным препаратом приводила к подавлению роста бактерий на 91,8%, 88,6%, 97,7% соответственно по сравнению с контролем (р<0,001). Дальнейшее увеличение времени инкубации до 120 и 150 минут полностью подавляло рост колоний Ps. aeruginosa. Максимальная антибактериальная активность препарата 6 наблюдалась в высокой концентрации 1 мг/мл, т.к. в данной концентрации при короткой инкубации 30 минут рост колоний подавлялся на 96,3%, а при увеличении времени инкубации до 60, 90, 120 и 150 минут роста бактериальных колоний не наблюдалось (р<0,001) (таблица 33). Следовательно, препарат 6 в концентрациях 0,01-1 мг/мл оказывал выраженное антибактериальное действие на клинические штаммы aeruginosa. Меньшие концентрации препарата не обладали Ps. значимой антибактериальной активностью. Подводя итог вышеизложенному, можно заключить, что селеноорганические соединения 1, 2 и 3, а также теллурорганический препарат 5 обладали большей суммарной антимикробной активностью на клинические штаммы золотистого стафилококка, чем на синегнойную палочку. Подобные различия в антибактериальном действии можно объяснить особенностями строения бактериальных стенок грамположительных и грамотрицательных бактерий. Кроме того, максимальной антимикробной активностью на клинические штаммы Ps. aeruginosa обладал препарат 3 (фторсодержащий аналог ДАФС-25), тогда как теллурорганическое соединение 5 оказывало сравнительно меньший суммарный антибактериальный эффект. 205 Селеноорганический препарат ДАФС-25, лишенный боковых заместителей у фенильных радикалов, оказывал наименьшее антимикробное действие на клинические штаммы синегнойной палочки по сравнению с другими исследованными соединениями. 5.3. Антибактериальное действие халькогенорганических соединений на клинические штаммы Escherichia coli Селеноорганический препарат ДАФС-25 (соединение 1) проявлял антибактериальную активность на клинические штаммы Escherichia coli (E. coli) только в максимальной концентрации 1 мг/мл (таблица 34). Таблица 34. Антибактериальное действие соединения 1 на клинические штаммы Escherichia coli 60 90 821,2 ± 118,2 465,4± 156,7* 685,8± 230,4 929,9± 152,6 762,3± 210,3 905,5± 155,5 120 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 937,6± 1005,6± 937,5± 973,3± 773,3± 971,3± 52,3 131,5 37,8 202,4 91,2* 131,7 882,4 ± 611,3± 895,6± 789,2± 892,4± 903,4± 163,3 110,4* 41,3 169,7 111,6 168,4 804,7 ± 291,5 118,6± 76,4** 677,8± 259,6 724,3± 222,4 854,5± 191,2 1073± 188,3 150 Время воздействия, мин 30 Условия эксперимента 943,3 ± 87,8 114,4± 59,5** 362,3± 86,5** 778,3± 204,4 823,2± 131,4 860,1± 54,8 * - р<0,05 ,** - р< 0,001 Причем антибактериальный эффект возрастал по мере увеличения времени инкубации. При инкубации 60 минут препарат 1 подавлял рост 206 колоний кишечной палочки на 30,7% (р<0,05), при инкубации 90 минут – на 43,4% (р<0,05), при инкубации 120 минут – на 85,3% (р<0,05) и при инкубации 150 минут – на 87,9% соответственно по сравнению с контролем (р<0,001). При максимальной инкубации препарат ДАФС был также эффективен в концентрации 0,1 мг/ мл и вызывал достоверное уменьшение роста колоний кишечной палочки на 61,6% (р<0,001) по сравнению с контролем. Достоверная антибактериальная активность также наблюдалась при минимальной инкубации 30 минут в концентрации препарата ДАФС 0,001 мг/мл – число колоний сокращалось на 17,5% (таблица 34). Следовательно, клинические штаммы E. coli чувствительны только к высокой концентрации селеноорганического препарата ДАФС-25. Причем эта чувствительность возрастает по мере увеличения времени инкубации и составляет максимум при инкубации 120 и 150 минут в концентрации 1 мг/мл. Сравнительно большей антибактериальной активностью на клинические штаммы кишечной палочки обладало соединение 2 (хлорсодержащий аналог препарата ДАФС-25), что следует из результатов таблицы 35. Таблица 35. Антибактериальное действие соединения 2 на клинические штаммы Escherichia coli Условия эксперимента Время воздействия, мин 30 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (n=10) (ДМФА и физ. р-р) 896,7 ± 225,6± 564,4±108,2* 836,3±96,5 926,7±105,5 48,5 50,6** * 60 908,9± 135,5 276,7± 134,5** 458,2±95,5** 604,3±234,5* 765,6±92,3* 90 982,3± 120,5 787,4± 62,5 702,3±89,5 98,4± 24,3** Нет роста 480,3±249,2* 676,7±112,5* 694,4±192,5 * * * 192,3±125,6* 464,5±158,5* 604,3±156,2 * 169,2±78,4** 371,2± 347,4± 213,4** 110,5** 120 150 Нет роста * - Р<0,05 ,** - Р< 0,001 207 Это соединение оказывало антибактериальной действие в концентрациях от 1 до 0,001 мг/мл и при инкубации 30-150 минут. Минимальная концентрация 0,001 мг/мл препарата 2 эффективно подавляла рост колоний кишечной палочки при инкубации в течение 60-150 минут на 15,8% (60 минут), 29,3% (90 минут), 23,3% (120 минут) и 50,6% (150 минут) (р<0,05). В концентрации 0,01 мг/мл препарат 2 достоверно уменьшал число клеточных колоний кишечной палочки на 33,5% (60 минут), 33,1% (90 минут), 41,0% (120 минут), 47,2% (150 минут) соответственно по сравнению с контролем (р<0,05). Антибактериальная активность препарата 2 в концентрации 0,1 мг/мл выражалась в подавлении роста колоний E. coli на 37,1% (30 минут), 49,6% (60 минут), 51,1% (90 минут), 75,6% (120 минут), 75,9% (150 минут) соответственно (р<0,05). Наконец, в максимальной концентрации препарата 2 (1 мг/мл) наблюдалась наибольшая антимикробная активность в отношении кишечной палочки. При этом уменьшалось число колоний на 74, 9% (30 минут), 69,6% (60 минут), 90,0% (90 минут). При инкубации кишечной палочки с препаратом 2 в концентрации 1 мг/мл в течение 120 и 150 минут рост колоний подавлялся полностью (р<0,001) (таблица 35). Полученные результаты демонстрируют высокую антибактериальную активность препарата 2 в отношении клинических штаммов кишечной палочки. Однако полное подавление роста бактерий наблюдалось лишь в максимальной концентрации препарата 1 мг/мл и при длительной инкубации 120 и 150 минут. При краткосрочной инкубации (30 минут) препарат 2 был эффективен только в высоких концентрациях 1 и 0,1 мг/мл. Тем не менее, препарат 2 оказывал достоверное антибактериальное действие во всех исследованных концентрациях при времени инкубации от 60 до 150 минут (таблица 35). Кроме того, следует отметить сравнительно большую антимикробную активность препарата 2 в концентрации 1 мг/мл в отношении кишечной палочки по сравнению с действием этого препарата на штаммы золотистого 208 стафилококка и синегнойной палочки. Однако в низкой концентрации 0,001 мг/мл препарат 2 был более эффективен против штаммов золотистого стафилококка при инкубации от 60 до 150 минут. Также обращает на себя внимание полное подавление роста колоний золотистого стафилококка препаратом 2 в концентрациях от 1 до 0,01 мг/мл при длительной инкубации 150 минут. Следовательно, препарат 2 как антимикробный агент наиболее эффективен против клинических штаммов золотистого стафилококка. Значительно большая антибактериальная активность препарата 2 (хлорсодержащего аналога препарата ДАФС) по сравнению с препаратом 1 (ДАФС-25), очевидно, связана с наличием в его структуре двух атомов хлора. Фторсодержащее производное препарата ДАФС-25 (соединение 3) благодаря наличию двух атомов фтора в своей структуре обладало большей суммарной антимикробной активностью на клинические штаммы кишечной палочки по сравнению с соединениями 1 и 2. Препарат 3 эффективно подавлял рост бактериальных клеток во всех исследованных концентрациях (0,0001-1 мг/мл) и при разном времени инкубации 30-150 минут, что демонстрирует таблица 36. Значительное уменьшение числа клеточных колоний наблюдалось при инкубации клеток кишечной палочки с препаратом 3 в минимальной концентрации 0,0001 мг/мл: на 19,9% (30 мнут) (р<0,05), 35,8% (60 минут), 50,4% (90 минут), 92,1% (120 минут), 93,6% (150 минут) соответственно по сравнению с контролем (р<0,001). Препарат 3 в низкой концентрации 0,001 мг/мл также был высоко эффективен против кишечной палочки, уменьшая число клеточных колоний на 41,1% (30 минут) (р<0,001), 50,2% (60 минут) (р<0,05), 77,6% (90 минут), 89,7% (120 минут), 99,2% (150 минут) (р<0,001) соответственно. Препарат 3 в концентрациях 0,01-1 мг/мл проявлял более выраженную антибактериальную активность, значительно снижая число выросших колоний. Например, препарат 3 в концентрации 0,01 мг/мл подавлял рост колоний 209 кишечной палочки на 62,9% при инкубации 30 минут, на 77,2% при инкубации 60 минут и на 94,6% при 90-мнутной инкубации (р<0,001). При увеличении времени инкубации до 120 и 150 минут рост бактериальных клеток был полностью ингибирован (р<0,001) (таблица 36). Таблица 36. Антибактериальное действие соединения 3 на клинические штаммы Escherichia coli Время воздействия, мин Условия Количество колоний на твердых питательных средах экспери- Контроль Опытные группы, концентрация вещества, мг/мл (n=10) мента 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 30 967,8± 200,6± 222,3± 358,4± 570,3± 775,6± 44,5 155,5** 131,5** 193,5** 280,2** 207,5* 60 896,7 ± 28,5± 78,6± 204,3± 446,4± 575,6± 96,2 21,2** 62,4** 162,5** 269,5* 197,5** 90 1003,4 ± Нет роста Нет 54,2± 225,7± 498,2± 97,3 роста 43,1** 137,5** 210,1** 120 877,2 ± Нет роста Нет Нет роста 90,2± 69,2± 82,5 роста 56,6** 25,2** 150 893,4 ± Нет роста Нет Нет роста Нет роста 57,3± 79,5 роста 36,3** * - р<0,05 ,** - р< 0,001 Наибольший антибактериальный эффект препарата 3 наблюдался в высоких концентрациях (0,1 и 1 мг/мл). При этом при 30-минутной инкубации штаммов кишечной палочки с препаратом 3 в концентрации 0,1 мг/мл отмечалось уменьшение числа бактериальных колоний на 77,0%, а при инкубации 60 минут – на 91,3% (р<0,001) соответственно по сравнению с контролем. Увеличение времени инкубации до 90, 120 и 150 минут приводило к полному подавлению роста бактериальных клеток. Наконец, препарат 3 в концентрации 1 мг/мл подавлял рост колоний кишечной палочки на 79,3% при 30-минутной инкубации и на 96,9% при 60минутной инкубации. При более длительной инкубации (90, 120, 150 минут) клинических штаммов E. coli рост клеток подавлялся полностью (таблица 36). 210 Следовательно, препарат 3 оказывал выраженное антибактериальное действие на клинические штаммы кишечной палочки во всех исследованных концентрациях и при разном времени инкубации, достигая максимума в концентрациях 0,01-1 мг/мл и при длительной инкубации от 90 минут и более. Кроме того, можно отметить сходную антибактериальную активность препарата 3 в отношении клинических штаммов золотистого стафилококка и кишечной палочки. Однако препарат 3 во всех концентрациях при минимальной инкубации 30 минут был более эффективен против штаммов золотистого стафилококка, а в минимальной концентрации 0,0001 мг/мл и при длительной инкубации (120 и 150 минут) этот препарат оказывал большее противомикробное действие на штаммы кишечной палочки. Менее выраженное антибактериальное действие препарат 3 оказывал на клинические штаммы синегнойной палочки, т.к. низкие концентрации этого препарата были неэффективны при инкубации 30 минут. Согласно данным таблицы 37, нитропроизводное препарата ДАФС-25 (соединение 4) в концентрациях 0,01-1 мг/мл достоверно подавляло рост клинических штаммов кишечной палочки. В концентрации 0,01 мг/мл этот препарат снижал рост клеточных колоний на 39,5% (30 минут), 50,1% (60 минут), 64,8% (90 минут), 79,4% (120 минут) и на 96,4% (150 минут) соответственно по сравнению с контролем (р<0,001). Более выраженный антибактериальный эффект препарата 4 проявлял в концентрации 0,1 мг/мл. При этом число бактериальных колоний уменьшалось по мере увеличения времени инкубации: на 56,3% (30 минут), 71,6% (60 минут), 85,1% (90 минут), 90,9% (120 минут) и на 92,5% при инкубации 150 минут по сравнению с контролем (р<0,001). В максимальной концентрации 1 мг/мл препарат 4 оказывал наибольшее антибактериальное действие. В зависимости от времени инкубации рост бактерий подавлялся на 75,9% (30 минут) и на 93,2% (60 минут) (р<0,001) соответственно. При инкубации 120 и 150 минут роста бактериальных колоний не наблюдалось (р<0,001). 211 Таблица 37. Антибактериальное действие соединения 4 на клинические штаммы Escherichia coli Условия эксперимента Время воздействия, мин 30 Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 (n=10) 0,1 (n=10) 0,01 (n=10) 0,001 (n=10) (ДМФА и физ. р-р) 1081,2 ± 261,3± 472,4± 654,2± 1020,4± 95,5 46,5** 65,5** 132,5** 49,2 60 971,3 ± 63,4 66,4± 32,2** 276,3± 30,1** 485,6± 198,5* 980,7± 99,9 90 1046,3 ± 153,5 50,1± 41,4** 156,2± 65,7** 368,2± 124,5** 977,2± 151,5 120 898,7 ± 130,1 Нет роста 82,6± 64,5** 185,4± 90,5** 779,4± 126,5 830,3 ± Нет 128,5 роста * - р<0,05 ,** - р< 0,001 62,3± 35,2** 30,1± 11,1** 615,1± 118,5* 150 Во всех перечисленных случаях число бактериальных клеток уменьшалось прямо пропорционально повышению концентрации препарата и увеличению времени инкубации (таблица 37). В низкой концентрации 0,001 мг/мл препарат 4 достоверно, но незначительно (на 25,9%) подавлял рост бактерий кишечной палочки лишь при максимальной инкубации 150 минут. Сокращение времени инкубации (до 120 минут и менее) не приводило к достоверному изменению числа бактериальных колоний по сравнению с контролем (таблица 37). Следовательно, препарат 4 в концентрации 0,01 мг/мл был более эффективен против штаммов кишечной палочки, в меньшей степени - против штаммов синегнойной палочки и, наконец, не эффективен против штаммов золотистого стафилококка. Однако максимальная концентрация препарата 4 (1 мг/мл) оказывала более значительное антибактериальное действие на клинические штаммы синегнойной палочки и золотистого стафилококка. При 212 этом антибактериальная активность возрастала по мере увеличения времени инкубации. Следует также отметить меньшую антибактериальную активность препарата 4 по сравнению с препаратами 2 и 3. Тем не менее, несмотря на низкую антибактериальную активность препарата 4 в малых концентрациях необходимо отметить, что активность препарата 4 гораздо выше, чем у препарата 1 – ДАФС-25, что, безусловно, зависело от наличия в структуре препарата 4 двух нитрогрупп. Вероятно, нитрогруппы в составе препарата 4 подвергаются окислению под действием бактериальных ферментов, примерно также как это происходит с нитрогруппами нитрофуранов [Глыбочко П.В. и др., 2010], но в значительно меньшей степени, т.к. антибактериальная активность нитрофуранов на несколько порядков выше, чем у препарата 4. Наконец, теллурорганический антибактериальную активность на препарат 6 клинические оказывал штаммы выраженную E. coli в концентрациях 0,001-1 мг/мл. В минимальной концентрации 0,0001 мг/мл препарат 6 не вызывал значительного подавления роста клеток кишечной палочки (таблица 38). При низкой концентрации 0,001 мг/мл препарат 6 эффективно подавлял рост бактериальных колоний: на 37,9% (30 минут), 39,4% (60 минут), 58,9% (90 минут), 57,9% (120 минут) и на 78,4% (150 минут) соответственно по сравнению с контролем (р<0,001). В концентрации 0,01 мг/мл препарат 6 оказывал сравнительно большее антибактериальное действие, уменьшая число бактериальных колоний на 57,5% (30 минут), 66,3% (60 минут), 71,6% (90 минут), 80,8% (120 минут), 92,8% (150 минут) соответственно (р<0,001). Наибольший антибактериальный эффект наблюдался в максимальных концентрациях препарата 6 (1 и 0,1 мг/мл). Препарат 6 в концентрации 0,1 мг/мл практически полностью подавлял рост штаммов кишечной палочки, уменьшая число бактериальных колоний на 92,6% при 30-минутной инкубации (р<0,001). Более длительные периоды инкубации приводили к полному подавлению роста бактерий (р<0,001) (таблица 38). 213 Максимальная концентрация препарата 6 (1 мг/мл) характеризовалась полным отсутствием роста бактериальных клеток кишечной палочки при всех режимах инкубации (р<0,001). Таблица 38. Антибактериальное действие соединения 6 на клинические штаммы Escherichia coli Время воздействия, мин Условия эксперимента Количество колоний на твердых питательных средах Контроль Опытные группы, концентрация вещества, мг/мл (n=10) 1 0,1 0,01 0,001 0,0001 (ДМФА и (n=10) (n=10) (n=10) (n=10) (n=10) физ. р-р) 30 906,8 ± Нет 67,4± 385,4± 562,2± 872,2± 86,3 роста 49,3** 43,2** 54,5** 26,1 60 903,4 ± Нет Нет 304,2± 547,5± 776,7± 72,2 роста роста 90,5** 67,8** 163,5* 90 938,3 ± Нет Нет 266,4± 385,4± 713,2± 89,9 роста роста 134,5** 86,9** 154,1* 120 926,4 ± Нет Нет 178,2± 390,1± 724,2± 103,3 роста роста 60,5** 133,5** 194,5* 150 936,2 ± Нет Нет 67,3± 202,2± 729,3± 94,1 роста роста 48,4** 55,5** 214,5 * - р<0,05 ,** - р< 0,001 В заключение следует отметить, что препарат 6 обладал наибольшей антибактериальной активностью в отношении клинических штаммов золотистого стафилококка, подавляя рост бактерий уже при концентрации 0,0001 мг/мл. Однако в данной концентрации препарат 6 не оказывал достоверного и значимого антибактериального действия на клинические штаммы E. coli и Ps. aeruginosa. Менее чувствительными к препарату 6 оказались клинические штаммы кишечной палочки. Рост этих бактерий снижался при концентрации препарата 0,001 мг/мл, а при больших концентрациях отмечалось более выраженное антибактериальное действие этого препарата. Наконец, наименее чувствительными к действию препарата 6 оказались клинические штаммы синегнойной палочки, т.к. значительный 214 антибактериальный эффект этого препарата наблюдался в концентрациях 0,01 мг/мл и выше. 5.4. Механизм антибактериального действия халькогенорганических соединений Согласно полученным результатам суммарная антибактериальная активность исследованных халькогенорганических соединений на клинические штаммы грамположительных (Staphylococcus aureus) и грамотрицательных (Escherichia coli и Pseudomonas aeruginosa) бактерий снижалась в направлении: ДАФС(Те) > ДАФС(F2) > ДАФС(Cl2) > ДАФС(NO2)2) > ДАФС(Se) > ДАФС(S). Достаточно высокие (теллуроорганический антибактериальные аналог препарата свойства ДАФС) по препарата 6 сравнению с селеноорганическими соединениями, вероятно, обусловлены особенностями его строения. Теллур, также как сера и селен является халькогеном и обладает сходными химическими и, как можно предположить, биологическими свойствами. Известно, что у халькогенов в направлении кислород → сера → селен → теллур → полоний усиливаются металлические свойства и возрастает токсичность, достигая максимума у полония, который является радиоактивным металлом. Согласно литературным данным, отмечается существенное понижение энергий связей «Углерод-Халькоген» в ряду S, Se, Te, поэтому в органических соединениях халькогенов наибольшей энергией обладает связь «С-S», а, следовательно, сера практически не освобождается из своих органических соединений [Садеков И.Д., Минкин В.И., 1995]. Вместе с тем, связь «С-Те» разрывается легче, чем связи, образованные другими халькогенами и происходит освобождением атома теллура в виде неорганического теллура (IV), который значительно токсичнее своей органической формы [Садеков И.Д., Минкин В.И., 1995; Meinerz D.F., et al., 2013]. Поскольку энергия связи «C-Se» имеет средние значения между связями «C-S» и «С-Те», освобождение атома 215 селена происходит сложнее, чем из соответствующих соединений теллура, но легче, чем из серосодержащих соединений. Освободившиеся атомы халькогенов превращаются в соответствующие оксианионы теллурита (ТеО32-), селената (SeO42-) и селенита (SeO32-), которые обладают высокой токсичностью [Choudhury H.G., et al., 2011]. Однако грамотрицательные бактерии родов Escherichia и Pseudomonas способны обезвреживать токсичные для них оксианионы теллурита, селената и селенита до элементарных теллура/селена, которые нетоксичны [Choudhury H.G., et al., 2011]. Этим можно объяснить меньшую антимикробную активность теллурсодержащего препарата 6 в отношении клинических штаммов E. coli и P. aeruginosa в сравнении с S. aureus. Следовательно, можно предположить, что высокая антибактериальная активность препарата 6 обусловлена токсичностью атома теллура в его составе. Антибактериальная активность препарата 6 достаточно высока и сопоставима с активностью препарата 3 (фторсодержащего аналога препарата ДАФС) в отношении клинических штаммов золотистого стафилококка. Однако препарат 3 был самым эффективным среди исследованных в отношении клинических штаммов E. coli и P. aeruginosa даже в низких концентрациях 0,0001-0,001 мг/мл и при инкубации 30-150 минут. Выраженной антибактериальной активностью также обладал препарат 2 (хлорсодержащий аналог препарата ДАФС), который эффективно подавлял рост клинических штаммов золотистого стафилококка уже в низкой концентрации 0,001 мг/мл. Меньшую антибактериальную активность препарат 2 проявлял в отношении клинических штаммов кишечной палочки. Клинические штаммы синегнойной палочки были чувствительны только к высоким концентрациям препарата 2. Антибактериальная активность препаратов 2 и 3 объясняется наличием в их структуре галогенов – атомов фтора и хлора соответственно. Антимикробная активность препарата 3 сильнее, чем у препарата связано, в первую очередь, с наличием в структуре 2. Это препарата 3 216 (фторсодержащего аналога препарата ДАФС) двух атомов фтора. Известно, что фтор самый электроотрицательный элемент и самый мощный окислитель, поэтому благодаря наличию двух атомов фтора препарат 3 становится самым токсичным из всех исследованных халькогенорганических препаратов, проявляя максимальную антибактериальную активность. Окислительные свойства хлора значительно слабее, чем у фтора. Кроме того, хлор, являясь важным биогенным элементом, участвует во многих жизненно важных процессах, поэтому хлорсодержащий аналог препарата ДАФС (препарат 2) обладал меньшей антимикробной активностью по сравнению с препаратом 3. Кроме того, его активность слабее, чем у препарата 5 (теллурорганического соединения), но гораздо выше, чем у препаратов 1 и 4. Сравнительно низкая антибактериальная активность препаратов 1 и 4, вероятно, связана с их низкой токсичностью и высокими антиоксидантными и антитоксическими свойствами. Однако препарат 4 проявлял большую антибактериальную активность по сравнению с препаратом 1 (ДАФС) в отношении клинических штаммов золотистого стафилококка, кишечной и синегнойной палочки. При этом данный препарат в концентрации 0,01 мг/мл более эффективно подавлял рост клинических штаммов кишечной палочки, в меньшей степени - синегнойной палочки и не был эффективен против штаммов золотистого стафилококка. Особый интерес представляет наличие в структуре препарата 4 двух нитрогрупп, что позволяет проводить аналогию с антибактериальным действием нитрофуранов, хотя нитрофураны проявляют широкий спектр антибактериальной активности уже при концентрации 0,5-50 мкг/мл. Установлено, что сами по себе нитрофураны антибактериальной активностью не обладают, но приобретают ее при восстановлении флавинзависимыми редуктазами, т.е. для проявления как терапевтического, так и побочного действия нитрофуранов необходимо их одноэлектронное восстановление ферментами с локализованными в простейших микробах, нитроредуктазной или тканях активностью, организма. 217 Промежуточные продукты последовательных одно- или двухэлектронных этапов восстановления являются высоко реакционноспособными, особенно анион нитрорадикала, антибактериальную, благодаря которому противоопухолевую и нитрофураны генотоксическую приобретают активность [Глыбочко П.В. и др., 2010]. В заключение следует отметить большую чувствительность клинических штаммов золотистого стафилококка к исследованным халькогенорганическим соединениям по сравнению со штаммами грамотрицательных бактерий (кишечной и синегнойной палочек). При этом чувствительность клинических штаммов кишечной и синегнойной палочки к исследованным соединениям была сопоставима. Различия в антимикробной активности халькогенсодержащих соединений в отношении грамотрицательных и грамположительных бактерий, вероятно, связаны также с особенностями строения их клеточных стенок, которые обусловливают различие в проходимости разнообразных веществ. Этим можно объяснить неэффективность большинства синтетических и натуральных антимикробных агентов против грамотрицательных бактерий, таких как Escherichia coli и Pseudomonas aeruginosa. Наружная мембрана этих бактерий состоит из липополисахарид-липопротеиновых комплексов и недоступна для большинства антибиотиков, включая пенициллин и другие препараты [Soni D. et al, 2005]. Транспорт питательных веществ (сахаров, аминокислот) и антибиотиков внутрь бактериальной клетки осуществляется через воронкообразные белковые структуры (пориновые каналы), встроенные в липополисахаридный слой (см. рис. 9). Гидрофобные соединения (среди антибиотиков к таковым относятся хинолоны, макролиды и тетрациклины) способны диффундировать через липополисахаридный слой. Сравнительно высокомолекулярные антибиотики, такие как гликопептиды, природные пенициллины с трудом проникают через пориновые каналы грамотрицательных бактерий, чем и объясняется природная 218 устойчивость этих микроорганизмов к перечисленным препаратам [Сидоренко С.В., 2000]. У грамположительных бактерий над пептидогликаном и тейхоевыми кислотами располагается лишь аморфная капсула полисахаридной (крахмалоподобной) природы. Следовательно, активности предположительный исследованных механизм халькогенорганических антибактериальной соединений, которые являются низкомолекулярными гидрофобными соединениями, заключается в их способности легко проникать через мембраны бактерий, метаболизироваться бектериальными ферментами с освобождением атомов селена и теллура в форме оксианионов. Последнее приводит к повышенной выработке АФК, окислительному повреждению внутриклеточных структур, остановке роста и размножения бактериальных клеток. 219 Глава 6. Элементы структурного подобия молекул как предпосылка сходного физиологического действия халькогенорганических соединений и глюкокортикоидов Исследуемые в нашей работе халькогенорганические соединения в зависимости от наличия боковых радикалов и гетероатома халькогена (серы, селена или теллура) демонстрировали антиоксидантную, антитоксическую и антимикробную активности. Наибольшей антиоксидантной активностью обладал препарат ДАФС-25 (соединение 1), в меньшей степени - соединения 2 и 4. Эти препараты снижали концентрацию продуктов ПОЛ и приводили к увеличению активности антиоксидантных ферментов каталазы, СОД и ГПО в крови и тканях мышей. Препарат ДАФС также был эффективным антитоксикантом при подостром отравлении животных солями кадмия, ртути и свинца, меньшее антитоксическое действие оказывали соединения 2, 4 и 5. Соединения 3 и 6, являясь токсичными по своей природе, не оказывали антитоксического действия. По этой же причине именно эти соединения оказывали выраженное антибактериальное действие на клинические штаммы грамположительных и грамотрицательных бактерий, тогда как препараты 1 и 5 ввиду своей низкой токсичности не обладали выраженной антибактериальной активностью. Таким образом, родоначальник всех исследованных в нашей работе соединений селеноорганический препарат диацетофенонилселенид (ДАФС-25) обладал широким спектром биологической активности. Полученные нами результаты также демонстрируют гипергликемию, гиперхолестеринемию, увеличение активности ферментов АлАТ и ЛДГ, лейкоцитоз (35%) и незначительное увеличение числа эритроцитов (16%) в крови экспериментальных животных. Помимо этого, проведенные ранее исследования установили, что ДАФС25 увеличивает сохранность поголовья молодняка и взрослых животных, приводит к увеличению яичной и мясной продукции, благодаря чему этот 220 препарат применяется в настоящее время в животноводстве и птицеводстве ряда регионов России [Древко Б.И., 2001]. Биологическое действие этого препарата как иммуностимулятора, антиоксиданта, антитоксиканта и нестероидного анаболика связано с особенностями его строения, а именно с наличием в его структуре 1,5дифенилпентандиона-1,5. Вероятно, пространственная структура этого соединения сходна с пространственной структурой стероидных гормонов (половых гормонов и глюкокортикоидов). Поэтому на следующем этапе исследования мы предприняли попытку определить весь спектр биологической активности соединения ДАФС-25 и его производных с использованием современной версии компьютерной системы предсказания спектра биологической активности PASS. Нас, в первую очередь, интересовало подтверждение уже полученных нами и ранее описанных результатов по антиоксидантной, антитоксической, антибактериальной, радиопротекторной активности, а также подтверждение гормоноподобного действия соединения ДАФС и его производных. 6.1. Компьютерное прогнозирование биологической активности халькогенорганических соединений, полученные с использованием компьютерной системы предсказания спектра биологической активности PASS С использованием современной версии компьютерной системы предсказания спектра биологической активности PASS обнаружены: - для соединения 1 (ДАФС-25): 16 дескрипторов, из них 2 новых; 2175 возможных активностей при Pa > Pi. - для соединения 2 (ДАФС(Cl2)2): 20 дескрипторов, из них 2 новых; 1823 возможных активностей при Pa > Pi. - для соединения 3 (ДАФС(F2)2): 20 дескрипторов, из них 2 новых; 1722 возможных активностей при Pa > Pi. 221 - для соединения 4 (ДАФС(NO2)2): 24 дескриптора, из них 2 новых; 1462 возможной активности при Pa > Pi. - для соединения 5 (ДАФС(S)): 16 дескрипторов, из них 0 новых; 2209 возможных активностей при Pa > Pi. - для соединения 6 (ДАФС(Те)): 16 дескрипторов, из них 3 новых; 2267 возможных активностей при Pa > Pi. Выдаваемые результаты прогноза помимо названий активности включают в себя оценки вероятностей наличия (Pa) и отсутствия каждой активности (Pi), имеющие значения от 0 до 1. Поскольку эти вероятности рассчитываются независимо, их сумма не равна единице. В некоторых случаях вещество прогнозируется одновременно как агонист и антагонист (стимулятор и блокатор, активатор и ингибитор) по отношению к одним и тем же рецепторам (ферментам и т.п.). Это означает, что система не может дифференцировать внутреннюю активность вещества, а лишь указывает на его тропность (способность к связыванию) с данным рецептором (ферментом). Результаты предполагаемой халькогенорганических соединений, биологической полученные с активности использованием компьютерной программы PASS, лишь отчасти совпадали с полученными нами результатами. предполагаемой Представленные биологической в таблицах активности 39, 40 и 41 демонстрируют результаты высокую вероятность радиопротекторного, гепатопротекторного и антибактериального действия всех исследованных нами соединений (Ра > 0,7), что соответствует описанным ранее результатам. Однако, несмотря на полученные нами результаты по превентивному антитоксическому действию соединений селена при подостром отравлениями солями кадмия, ртути и свинца, потенциальная способность (Ра) предотвращать отравление тяжелыми металлами для всех соединений была невелика (Ра<0,7). 222 Обозначения: Ра – потенциальная активность, Pi –отсутствие активности 221 Таблица 39. Компьютерное прогнозирование антиоксидантной активности халькогенорганических соединений, полученные с использованием современной версии компьютерной системы предсказания спектра биологической активности PASS Потенциальные Исследованные препараты активности, ДАФС ДАФС-Cl ДАФС-F ДАФС-NO2 ДАФС-S ДАФС-Te механизмы Pa Pi Pa Pi Pa Pi Pa Pi Pa Pi Pa Pi действия и эффекты Ингибитор ПОЛ 0,831 0,002 0,743 0,003 0,750 0,003 0,735 0,003 0,813 0,002 Агонист 0,857 0,024 0,874 0,019 0,781 0,045 0,447 0,124 0,786 0,043 0,879 0,018 целостности мембраны Обезвреживание 0,567 0,020 0,421 0,092 0,421 0,092 0,331 0,184 0,629 0,006 0,587 0,013 АФК Обезвреживание 0,251 0,101 0,225 0,115 0,364 0,036 0,272 0,080 свободных радикалов Антиоксидант 0,417 0,030 0,325 0,047 0,268 0,065 0,306 0,052 0,184 0,119 0,385 0,034 Ингибитор СОД 0,773 0,008 0,551 0,025 0,451 0,043 0,888 0,004 0,852 0,005 0,833 0,005 Активатор СОД 0,443 0,017 0,429 0,021 0,315 0,093 0,298 0,109 0,394 0,035 0,468 0,011 Ингибитор 0,529 0,034 0,206 0,109 0,206 0,109 0,190 0,119 0,606 0,022 0,581 0,026 пероксидазы Активатор ГПО 0,370 0,004 0,204 0,023 0,204 0,023 0,171 0,034 0,300 0,008 0,407 0,004 Активатор каталазы 0,162 0,038 0,161 0,039 0,142 0,074 0,201 0,010 0,132 0,099 0,153 0,052 223 Таблица 40. Компьютерное прогнозирование действия халькогенорганических соединений на активность ферментов углеводного обмена, полученные с использованием современной версии компьютерной системы предсказания спектра биологической активности PASS ДАФС Pa Pi 2 3 0,672 0,004 0,620 0,009 ДАФС-Cl Pa Pi 4 5 0,490 0,013 0,468 0,027 Исследованные препараты ДАФС-F ДАФС-NO2 Pa Pi Pa Pi 6 7 8 9 0,490 0,013 0,390 0,026 0,468 0,027 0,320 0,061 ДАФС-S Pa Pi 10 11 0,635 0,005 0,546 0,016 ДАФС-Te Pa Pi 12 13 0,704 0,004 0,654 0,007 0,503 0,020 0,617 0,011 0,373 0,048 0,718 0,006 0,485 0,023 0,544 0,016 0,365 0,019 0,276 0,032 0,221 0,044 0,309 0,026 0,355 0,020 0,407 0,015 0,229 0,048 0,115 0,097 0,115 0,097 - - 0,266 0,041 0,269 0,041 0,213 0,038 0,139 0,077 0,139 0,077 0,231 0,033 - - 0,100 0,013 0,207 0,032 0,147 0,062 0,147 0,062 0,124 0,059 0,175 0,045 0,223 0,027 0,201 0,008 0,141 0,029 0,225 0,005 0,190 0,010 0,167 0,016 0,218 0,006 0,217 0,194 0,025 0,044 0,138 0,120 0,058 0,104 0,123 0,120 0,068 0,104 0,099 0,211 0,091 0,037 0,173 0,206 0,039 0,038 0,242 0,213 0,019 0,036 0,860 0,017 0,780 0,040 0,780 0,040 0,659 0,085 0,824 0,026 0,876 0,013 Обозначения: Ра – потенциальная активность, Pi –отсутствие активности 222 Потенциальные активности, механизмы действия и эффекты 1 Ингибитор транскетолазы Ингибитор глюконолактоназы Ингибитор пируватдекарбоксилазы Ингибитор пируватдегидрогеназы Ингибитор гликогенсинтазы Ингибитор глицеральдегид3-фосфат-дегидрогеназы (фосфорилирующей) Ингибитор фруктозо-2,6бифосфат-2-фосфатазы Ингибитор фосфоенолпируваткарбоксикиназы (ATP) Ингибитор АлАТ Ингибитор пируваткарбоксилазы Ингибитор тестостерон-17дегидрогеназы (NADP+) 224 Таблица 41. Компьютерное прогнозирование биологической активности халькогенорганических соединений, полученные с использованием современной версии компьютерной системы предсказания спектра биологической активности PASS ДАФС Pa Pi 2 3 0,116 0,053 0,857 0,002 ДАФС-Cl Pa Pi 4 5 0,110 0,072 0,801 0,003 Исследованные препараты ДАФС-F ДАФС-NO2 Pa Pi Pa Pi 6 7 8 9 0,710 0,007 0,620 0,027 0,806 0,003 0,770 0,003 0,643 0,080 0,665 0,065 0,627 0,084 0,618 0,090 0,580 0,115 0,669 0,059 0,609 0,034 0,497 0,103 0,577 0,050 0,425 0,164 0,604 0,036 0,630 0,025 0,560 0,233 0,734 0,038 0,110 0,018 0,416 0,186 0,746 0,109 0,142 0,015 0,416 0,181 0,617 0,109 0,145 0,070 0,395 0,183 0,521 0,125 0,143 0,129 0,625 0,499 0,692 0,020 0,061 0,033 0,591 0,514 0,752 0,028 0,022 0,013 0,554 0,078 0,572 0,068 0,515 0,106 0,322 0,304 0,572 0,068 0,598 0,055 0,869 0,018 0,906 0,019 0,710 0,007 0,620 0,027 - - 0,887 0,012 0,857 0,857 0,857 0,006 0,002 0,006 0,874 0,801 0,874 0,018 0,003 0,018 0,782 0,186 0,782 0,032 0,172 0,032 0,625 0,625 0,625 0,030 0,023 0,030 0,424 0,314 0,502 0,045 0,063 0,050 0,292 0,879 0,879 0,104 0,002 0,005 0,671 0,306 0,016 0,022 0,248 0,233 0,173 0,059 0,233 0,059 0,228 0,194 0,198 0,095 0,375 0,338 0,060 0,015 0,336 0,324 0,085 0,018 Обозначения: Ра – потенциальная активность, Pi –отсутствие активности ДАФС-S Pa Pi 10 11 0,114 0,058 0,892 0,003 ДАФС-Te Pa Pi 12 13 0,120 0,044 0,879 0,002 223 Потенциальные активности, механизмы действия и эффекты 1 Анаболический эффект Антибактериальная активность Ингибитор проницаемости мембраны Цитопротекторный эффект Стимулятор лейкопоэза Иммуностимулятор Гиперхолестеринемическ ий эффект Гипергликемический эффект Антиостеопоротический эффект Радиопротектор Гепатопротектор Хемопротективный эффект Антитоксический эффект Антидот, тяжелые металлы 225 По результатам расчета программы PASS вероятность потенциальной антиоксидантной активности, которая складывается из способности увеличивать активность антиоксидантных ферментов каталазы, СОД и ГПО, и подавлять пероксидацию липидов клеточных мембран была также мала для всех исследованных соединений (Ра<0,7). Предполагаемая способность соединений оказывать гормоноподобное действие за счет ингибирования или активирования ферментов углеводного обмена для всех соединений была низкая (Ра<0,7). Однако обращает на себя внимание потенциальное антиостеопоротическое действие соединений 1, 2, 3 и 6 (Ра> 0,7), что, вероятно, связано с наличием в их структуре 1,5-дифенилпентандиона-1,5, способного, как мы предполагаем, в оптимизированной форме взаимодействовать с рецепторами клеток. Несоответствие потенциальной полученных биологической нами активности, результатов с рассчитанной результатами компьютерной программой PASS, вероятно, связано с наличием в структуре исследованных соединений новых дескрипторов. Так, для соединений 1, 2, 3 и 4 обнаружены два новых дескриптора, для соединения 6 – три и лишь серосодержащий аналог ДАФС (соединение 5) не содержал новых дескрипторов. Учитывая наличие в исследованных халькогенорганических соединениях двух и более десктрипторов, результаты предполагаемой биологической активности этих соединений следует считать приблизительными [Филимонов Д.А., Поройков В.В., 2006]. В связи с вышеизложенным на следующем этапе данного исследования мы провели квантовохимические расчеты, подтверждающие структурное сходство селеноорганического соединения ДАФС-25, его производных и глюкокортикоидов. 226 6.2. Квантовохимические расчеты, подтверждающие структурное сходство селеноорганического соединения ДАФС-25, его производных и глюкокортикоидов Автор благодарит докт. хим. наук, профессора Панкратова А.Н. за содействие в проведении квантово-химических расчетов. В предсказательных целях нами дана квантовохимическая трактовка структурного сходства препарата ДАФС-25 и глюкокортикоидов (преднизона и преднизолона). Не имея информации о механизме и энергетике взаимодействия глюкокортикоидов и аналогичных по физиологическому действию веществ с рецепторами живых организмов, можно, тем не менее, полагать, что взаимная комплементарность молекул эффектора и рецептора в значительной степени определяется принципами топологического и электростатического соответствия. Чтобы молекула эффектора должным образом встраивались в структуру рецептора, необходимо ее геометрическое соответствие полости последнего. На рисунке 11 представлены химическое строение гормонального препарата преднизолон и селеноорганического соединения ДАФС-25 в оптимизированной форме: CH2OH Se H3C O CO OH H3C OO O O O O O H3C CH2OH OH H3C O ДАФС-25 в оптимизированной форме Преднизолон Рис. препарата 11. Черты сходства гормонального преднизолон селеноорганического соединения ДАФС-25 в оптимизированной форме и 227 Молекулярному распознаванию, взаимодействию биологически активных веществ с рецепторами биосубстратов предшествует отбор химических соединений по электростатическому механизму. В качестве дескрипторов геометрии нами выбраны величина а0, характеризующая размер молекулы в целом, молярный объем Vm, а также расстояния между атомами углерода, связанными с кислородными атомами (l1), и между самими атомами кислорода (l2). Величины l1 и l2 в значительной степени характеризуют топологию (форму) молекулы, во всяком случае, дают прямую информацию о расстоянии между полярными реакционноспособными карбонильными группами (или между группами С=О и С-ОН в молекуле преднизолона), участие которых в связывании эффектора с рецептором наиболее вероятно. Как интересная характеристика электрических свойств и геометрии молекул, мера их электростатического (ион-дипольного, диполь-дипольного) взаимодействия с другими молекулярными системами использован дипольный момент (μ). Наряду с препаратом ДАФС-25 нами рассмотрены его аналоги, содержащие вместо селена группу СН2 и атом серы. На рис. 12-16 приведены названия, нумерация и общий вид молекул указанных соединений, а также преднизона и преднизолона в в оптимизированной оптимизированной форме. Рис. 12. Молекула конформации 1,5-дифенилпентадион-1,5 228 Рис. 13. Молекула 1,5-дифенил-3-тиапентадиона-1,5 в оптимизированной конформации Рис. 14. Молекула 1,5-дифенил-3-селенпентадиона-1,5 (препарат ДАФС-25) в оптимизированной конформации Рис. 15. Молекула преднизона в оптимизированной конформации Рис. 16. Молекула преднизолона в оптимизированной конформации 229 В таблице 42 приведены результаты квантово-химических расчетов для молекул 1,5-дикарбонильных соединений и кортикостероидов. Как видно из всех рассмотренных нами 1,5-дикарбонилов молекула препарата ДАФС-25 наиболее близка (и достаточно близка; особенно это касается величин l1 и l2) к молекулам глюкокортикоидов преднизона и преднизолона как по размеру (а0, Vm), так и по расстояниям (l1 и l2) между полярными реакционноспособными группами С=О, С-ОН, которые потенциально могут участвовать в связывании с рецептором как посредством электростатических сил, так и за счет водородных связей или более глубокого химического взаимодействия. Таблица 42. Значения а0, Vm, l1, l2 и μ для молекул 1,5-дикарбонильных соединений и кортикостероидов, рассчитанные на уровне теории B3LYP/6311G(d,p) а0, Å Vm, cm3·mol-1 4,90 160,790 5,069 6,504 0,162 5,09 183,491 3,413 3,386 0,882 5,45 229,866 5,267 7,021 3,711 Преднизон * 5,70 266,403 5,162(4,777) 6,504(6120) Преднизолон * 5,84 287,321 5,037(4,816) 6,825(5,816) 4,198 Соединение 1,5-дифенилпента- l1, Å l2, Å μ, D дион-1,5 1,5-дифенил-3тиапентадион-1,5 1,5-дифенил-3селенпентадион-1,5 4,743 *Cначала приводится расстояние между двумя атомами углерода или кислорода, входящими в состав экзоциклических групп С=О или С-ОН, а в скобках – расстояние между соответствующими атомами экзоциклической и внециклической групп Применительно к стероидным гормонам преднизону и преднизолону в качестве величин l1 и l2, близких к таковым в молекуле ДАФС-25, выступают расстояния между атомами, входящими в состав экзоциклических групп С=О или С-ОН. 230 Как молекулу ДАФС-25, так и системы преднизон и преднизолон отличают высокие значения дипольных моментов, причем величины моментов диполей для молекул ДАФС-25, преднизон и преднизолон сопоставимы между собой и заметно отличны от таковых для 1,5-дифенилпентадион-1,5 и 1,5-дифенил-3-тиапентадион-1,5 (таблица 42). Молекула 1,5-дифенилпентадион-1,5 имеет значения l1 и l2, близкие к таковым для преднизона и преднизолона, однако их подобие молекулам глюкокортикоидов не удовлетворяет другим геометрическим (а0, Vm) и электростатическому (μ) критериям. Таким образом, налицо как геометрические (а0,Vm, l1, l2 и μ), так и электростатические (μ) предпосылки сходного характера биологической активности препарата ДАФС-25 и глюкокортикоидов. Квантовохимический прогноз, основанный на установлении электроннотопологического подобия молекул препарата ДАФС-25 и кортикостероидов, подтвержден нами экспериментально. 6.3. Влияние препарата ДАФС, его производных и преднизолона на показатели углеводного обмена экспериментальных животных Учитывая полученное нами подтверждение структурного подобия препарата ДАФС и глюкокортикоидов, на заключительном этапе нами было проведено изучение влияния селеноорганического препарата 1,5-дифенил-3селенпентадиона-1,5 (ДАФС-25), его производных и гормонального препарата преднизолона на показатели углеводного обмена у самцов белых беспородных мышей. Известно, что преднизолон обладает выраженным дозозависимым действием на метаболизм углеводов, белков и жиров; он стимулирует глюконеогенез, способствует захвату аминокислот печенью и почками и повышает активность ферментов глюконеогенеза. В печени преднизолон усиливает депонирование гликогена, стимулируя активность 231 гликогенсинтетазы [Lecocq F.R. et al., 1964]. Кроме того, данный гормон подавляет захват глюкозы жировыми клетками, что приводит к активации липолиза [Fain J.N., Czech M.P., 1975]. На примере другого синтетического глюкокортикоида – дексаметазона – показано, что его пероральное введение индуцирует экспрессию гена киназы пируватдегидрогеназы [Qi D. et al., 2004], следствием чего является снижение активности этого мультиферментного комплекса, замедление процесса окисления глюкозы и накопление пирувата и глюкозы в цитозоле клеток. Кроме того, дексаметазон уменьшает фосфорилирование фермента гликогенсинтазы, что приводит к восстановлению ее активности. Избыток глюкозы (при наличии активной гликогенсинтазы) используется для синтеза гликогена в печени, скелетных мышцах и миокарде [Qi D. et al., 2004; Puthanveetil P. et al., 2008]. Также следует отметить, что глюкортикоиды увеличивают экспрессию генов ферментов глюконеогенеза глюкозо-6-фосфатазы - и фосфоенолпируваткарбоксикиназы [Yabaluri N., Bashyam M.D., 2010], что влечет за собой усиление реакций глюконеогенеза в печени, выход свободной глюкозы в кровь и развитие гипергликемии. Субстратами для реакций глюконеогенеза могут служить аминокислоты, образующиеся в процессе катаболизма тканевых белков [Wilcke J.R., Davis L.E., 1982; Jin J.Y., Jusko W.J., 2009]. Гипергликемия усиливается за счет снижения поступления глюкозы в клетки периферических тканей [Baxter J.D., 1976; Andrews R.C., Walker B.R., 1999; Vila G. et a., 2010]. С целью изучения влияния препарата ДАФС-25 и его производных на углеводный обмен глюконеогенеза проводилось определение глюкозо-6-фосфатазы и активности ферментов фруктозо-1,6-бисфосфатазы в гомогенате печени, определение концентрации гликогена в печени и крови, а также определение содержания глюкозы в плазме крови и пирувата в крови белых мышей. Поскольку основными субстратами глюконеогенеза являются пируват и оксалоацетет, нами также оценивалась активность аминотрансфераз (АлАТ и АсАТ) в плазме крови. 232 Таблица 43 содержит результаты некоторых показателей углеводного обмена контрольной и экспериментальных групп животных. Таблица 43. Влияние препарата ДАФС, его производных и преднизолона на показатели обмена углеводоd b ,tkrjd экспериментальных животных Показатели углеводного обмена Глюкозо-6фосфатаза Фруктозо1,6-бисфосфатаза гликоген печени гликоген крови глюкоза Единицы Контроль ПредниСеленоорганические соединения измерезолон (n=10) (n=12) ния (n=8) 1 2 3 4 мкмоль/ 11,2 ± 1,1 25,5 17,9 ± 5,6 ± 6,5 ± 5,7 (л∙час) ±2,6* 2,4* 0,7* 0,7* ±0,9 * мкмоль/ 3,0 ± 0,4 6,9 ± 0,7* 4,4 ± 2,3 ± 3,8 ± 1,8 ± (л∙час) 0,6* 0,5 0,5 0,4* пируват ммоль/л АлАТ мккат/л АсАТ мккат/л ммоль/г ткани ммоль/л ммоль/л 3,6 ± 0,3 6,2 ± 0,4* 5,3 ± 0,3* 3,9 ± 0,5 6,1 ± 0,3* 5,3 ± 0,4* 3,4 ± 0,3 6,8 ± 0,3* 4,8 ± 0,2* 10,8 ± 1,0 5,7 ± 0,5* 6,6 ± 0,3 * 112,8 ± 6,5 283,7 ± 271,3 ± 11,2* 15,5 * 174,7 ± 7,2 191,7 ± 183,0 ± 13,5 8,8 4,1 ± 0,6 4,2 ± 0,6 2,9 ± 0,3 7,9 ± 0,5* 84,8 ± 4,9* 134,7 ± 6,7* 1,9 ± 0,2* 5,5 ± 0,2 * 2,8 ± 0,3 7,5 ± 0,8* 88,1 ± 6,4 * 148,5 ± 9,3 1,9 ± 0,2* 4,5 ± 0,4 2,0 ± 0,2* 8,1 ± 0,8* 148,7 ± 8,7* 168 ± 11,5 * - р <0,05 Полученные результаты демонстрируют стимулирующее действие гормонального препарата преднизолон и селеноорганического соединения ДАФС-25 (соединение 1) на показатели углеводного обмена экспериментальных животных. Активность одного из ключевых ферментов глюконеогенеза глюкозо-6-фосфатазы печени достоверно увеличивалась у мышей обеих экспериментальных групп: на 128,9% (преднизолон) и 60,7% (ДАФС) соответственно по сравнению с контролем (р<0,05). Однако активность данного фермента у мышей, получавших хлор-, фтор- и 233 нитропроизводные препарата ДАФС-25 (соединения 2, 3 и 4), снижалась на 50%, 41,9% и 49,1% соответственно по сравнению с контролем. Активность другого фермента глюконеогенеза – фруктозо-1,6- бисфосфатазы – достоверно увеличивалась у мышей, получавших per os ДАФС25 (на 46,7%), а действие преднизолона приводило к увеличению активности фермента на 130%. Применение нитропроизводного препарата ДАФС (соединение 4) сопровождалось снижением активности этого фермента на 40%. Субстраты глюконеогенеза - пируват и оксалоацетат - образуются в процессе обратимых реакций, катализируемых аминотрансферазами, из аланина и аспартата соответственно. Активность АлАТ в плазме крови также увеличивалась на 151,5% (преднизолон), 140,5% (ДАФС-25) и 31,8% (ДАФС(NO2)2) соответственно (р<0,05). Однако введение per os галогенопроизводных препарата ДАФС-25 (соединений 2 и 3) сопровождалось падением активности АлАТ на 24,8% и 21,9% соответственно. Следует отметить, что активность другой аминотрансферазы АсАТ оставалась неизменной почти у всех экспериментальных животных (только у мышей, получавших per os препарат 2, активность АсАТ снижалась на 22,9%). Известно, что любые состояния, требующие срочной мобилизации компонентов белка для покрытия энергетических нужд организма (недостаточное или несбалансированное питание, все виды стресса и т.п.), связаны с адаптивным, гормонально-стимулируемым биосинтезом аланин- и аспартатаминотрансфераз, участвующих в глюконеогенезе (Браунштейн, 1983). Активность АлАТ в клетках печени значительно выше активности АсАТ, поэтому в сыворотке крови мышей обеих экспериментальных групп активность АлАТ увеличивалась на фоне неизменной активности АсАТ. Значительное увеличение активности АлАТ сопровождалось достоверным снижением концентрации пирувата в плазме крови на 47,2% (преднизолон), 38,9% (препарат 1 - ДАФС), 26,9% (препарат 2), 30,6% (препарат 3) и 25,0% (препарат 4) соответственно по сравнению с контролем. 234 Это явление можно объяснить усилением распада тканевых белков, обратимым трансаминированием пирувата до аланина при участии фермента АлАТ, включением пирувата/аланина в глюкозо-аланиловый цикл, а в дальнейшем и в процесс новообразования глюкозы в печени. Следует также отметить, что концентрация глюкозы в плазме крови увеличивалась только у мышей первой и второй экспериментальных групп на 101,0% (преднизолон) и 40,6% (ДАФС) соответственно по сравнению с контролем. У мышей, получавших per os препарат 4, концентрация глюкозы снижалась на 41,2% (р<0,05). Гормоноподобное действие препарата ДАФС-25 также подтверждалось увеличением содержания гликогена в печени и крови. Причем содержание гликогена в печени возрастало только у мышей, получавших преднизолон (на 72,2%) и ДАФС-25 (на 47,2%) соответственно. При этом применение селеноорганических препаратов 3 и 4 сопровождалось снижением содержания гликогена в печени на 47,2% (р<0,05). Концентрация гликогена крови возрастала на 53,3% (преднизолон), 32,0% (ДАФС) и 41,0% (препарат 3) соответственно по сравнению с контролем. Согласно полученным результатам, селеноорганический препарат ДАФС25 (соединение 1) оказывало стимулирующее действие на отдельные стороны углеводного и белкового обменов экспериментальных животных подобно глюкокортикоидному препарату преднизолону, что выражалось в увеличении активности ферментов глюкозо-6-фосфатазы печени и аланинаминотрансферазы сыворотки крови, в увеличении концентрации глюкозы и снижению содержания пирувата в сыворотке крови, а также в увеличении содержание глюкокортикоидный углеводного обмена, гликогена препарат чем в вызывал органическое печени более и крови. выраженное соединение селена. При этом усиление Согласно квантовохимическим расчетам только молекула препарата ДАФС-25 (1,5дифенил-3-селенапентандиона-1,5) наиболее близка к молекулам глюкокортикоидов преднизона и преднизолона как по размеру, так и по 235 расстояниям между полярными реакционноспособными группами С=О, С-ОН, которые потенциально могут участвовать в связывании с рецептором как посредством электростатических сил, так и за счет водородных связей или ван дер Ваальсовых взаимодействий. Учитывая приведенные выше результаты квантовохимических электростатические расчетов, отражающих предпосылки сходного геометрические характера и биологической активности препарата ДАФС-25 и глюкокортикоидов, можно предположить, что диацетофенонилселенид в оптимизированной конформации способен взаимодействовать с рецепторами глюкокортикоидов и через них оказывать гормоноподобное действие на клетки экспериментальных животных. Селеноорганические соединения 2, 3 и 4 - производные препарата ДАФС25 - подобного эффекта не оказывали, вероятно, это связано с наличием боковых групп, которые могут препятствовать взаимодействию этих соединений с глюкокортикоидными рецепторами. В заключение необходимо подчеркнуть, что наличие в структуре препарата ДАФС-25 атома селена и 1,5-дифенилпентадиона-1,5 вносит решающий вклад в биологическую активность этого соединения и определяет его гормоноподобное действие. Результаты проведенного исследования демонстрируют важность изучения биологической активности кормовых добавок и биологически активных веществ, применяемых в ветеринарии и медицине, поскольку дальнейшие исследования в этой области помогут выявить потенциальные гормоноподобные вещества нестероидной природы, обладающие анаболическим, андроген/эстроген-модулирующим и противовоспалительным действием, что позволит правильно оценить их биологическую активность и корректно использовать в медицине, ветеринарии, птицеводстве и других областях народного хозяйства. животноводстве, 236 Заключение В современных условиях здоровье человека во многом определяется качеством продуктов питания, содержащих все необходимые микроэлементы, в том числе и селен. Учитывая, что до 80% населения России имеет недостаточную обеспеченность селеном [Сенькевич О.А. и др., 2011; Ширшова Т.И. и др., 2011; Давыденко Н.И. и др., 2012], в настоящее время проводится синтез и использование органических форм селена для профилактики селенодефицита и ряда заболеваний (беломышечной болезни, некроза и жирового перерождения печени, экссудативного диатеза, энцефаломаляции, расстройства сперматогенеза и др.) [Комзалова А.В. и др., 2012; Бикчантаев И.Т., Шакиров Ш.К., 2013; Фроловичев А.С., Трошин А.Н., 2013]. Интерес к селену и его соединениям связан с перспективами их использования в медицине, ветеринарии и сельском хозяйстве. Однако не меньший интерес вызывает изучение биологической активности и других халькогенов – серы и теллура. Известно, что халькогены (сера, селен и теллур) расположены в VI группе Периодической системы элементов Д.И. Менделеева, являются электронными аналогами, обладают сходными химическими свойствами, что обусловливает близость биологических свойств этих элементов и их соединений [Паперная Л. К., 2007]. В нашей работе антибактериальное соединений – и изучалось антиоксидантное, гормоноподобное производных диацетофенонилселенид (ДАФС-25) действие антитоксическое, халькогенорганических селеноорганического (производитель: г. Саратов), который применяется в животноводстве и ЗАО препарата «Сульфат», птицеводстве ряда регионов России и который позволяет нормализовать деятельность иммунной, антиоксидантной и детоксицирующей систем организма животных и птиц, приводит к увеличению яичной и мясной продукции [Древко Б.И. и др., 2001; Иванова Л.В., 2009]. Исследованные в нашей работе соединения можно разделить на две группы: 237 1) производные 1,5-дифенил-3-селенапентандион-1,5 (ДАФС(Se) – соединение 1), содержащие боковые радикалы у фенильных заместителей - 1,5-ди-(п-хлорфенил)-3-селенапентандион-1,5 соединение (ДАФС(Cl2), селенапентандион-1,5 2), 1,5-ди-(п-фторфенил)-33), (ДАФС(F2),соединение нитрофенил)-3-селенапентандион-1,5 1,5-ди-(м- (ДАФС(NO2)2), соединение 4); 2) производные 1,5-дифенил-3-селенапентандион-1,5 (ДАФС(Se), соединение 1), лишенные боковых радикалов, но содержащие атомы серы и теллура вместо атома селена - 1,5-дифенил-3тиапентандион-1,5 (ДАФС(S), соединение 5) и 1,5-дифенил-3теллуропентандион-1,5 (ДАФС(Те), соединение 6). В результате проведенных исследований удалось установить зависимость биологической активности халькогенорганических соединений от наличия в составе соединений боковых радикалов и гетероатома халькогена (S, Se, Te). Например, в ряду халькогенорганических соединений ДАФС(Se) и его производных, содержащих боковые заместители ДАФС(NO2)2, ДАФС(Cl2), и ДАФС(F2) установлены следующие свойства: снижаются антиоксидантная и антитоксическая активности, возрастают токсичность и антибактериальная активность. Такие же закономерности антиоксидантной и антитоксической активности обнаружены заместителей, но и в ряду соединений, лишенных боковых содержащие атомы серы и теллура вместо атома селена: ДАФС(Se), ДАФС(S), и ДАФС(Те), тогда как токсичность и антибактериальная активность возрастали в направлении ДАФС(S) → ДАФС(Se) → ДАФС(Те). Проведенные нами халькогенорганические исследования соединения доказали, обладали в что разной изученные степени и антиоксидантной и антитоксической активностью, а те соединения, которые обладали собственной токсичностью, оказывали выраженное антимикробное 238 действие в отношении клинических штаммов грамположительных и грамотрицательных микроорганизмов. Исследованные халькогенорганические препараты ДАФС-25 - (соединение 1), его хлор-, фтор-, нитросодержащие аналоги (соединения 2, 3 и 4 соответственно), а также препараты, в которых атом селена замещен на атомы серы и теллура (соединения 5 и 6 соответственно) - наиболее эффективно оказывали антиоксидантное действие на ткани мышей с низкой оксидорезистентностью, главным образом, на легкие и мозг. Более слабый антиоксидантный эффект наблюдался в высокооксидорезистентных гемолизате эритроцитов и плазме крови. Наименьшая антиоксидантная активность исследованных соединений обнаружена в печени и почках. Подобную неэффективность антиоксидантов в клетках этих органов можно объяснить их участием в метаболизме и экскреции ксенобиотиков. В печени происходит детоксикация гидрофобных веществ, а почки участвуют в их выведении. Селеноорганические соединения 1, 2, 3 и 4 оказывали наименьшее антиоксидантное действие на печень, а теллуроорганическое соединение 6 – на почки. Таким образом, антиоксидантное действие исследованных соединений напрямую зависело от их общего биологического эффекта in vivo. Из изученных соединений наибольший антиоксидантный эффект продемонстрировало соединение 1 (ДАФС-25), поскольку при его применении происходило одновременное снижение концентрации продуктов ПОЛ и увеличение активности ферментов каталазы, СОД и ГПО в органах и крови. Высокой антиоксидантной активностью обладало соединение 4 (нитропроизводное ДАФС-25), которое значительно снижало реакции ПОЛ в мозге, легких, эритроцитах и плазме крови. Соединение 2 (хлорсодержащий аналог ДАФС-25) антиоксидантную активность оказывало на эритроциты и легкие. наибольшую 239 Соединение 3 (фторсодержащее производное ДАФС-25) - самое токсичное из изученных препаратов – способствовало снижению концентрации продуктов ПОЛ в мозге и легких. В печени и плазме, напротив, наблюдалось нарастание концентрации продуктов ПОЛ (МДА и ДК). Данное соединение оказывало наименьший суммарный антиоксидантный эффект, что, безусловно, связано с токсичностью самого препарата, обусловленной наличием двух атомов фтора в пара-положении фенильных заместителей. Теллуроорганическое соединение 6 также вызывало токсический эффект, но за счет наличия в молекуле атома теллура. Как упоминалось выше, в ряду сера → селен → теллур нарастает токсичность, поэтому соединение 6 гораздо токсичнее селенсодержащего аналога – соединения 1. Соединение 6 обладало достоверной антиоксидантной активностью лишь в органах с низкой оксидорезистентностью – в легких и мозге. Как сообщалось ранее, серосодержащее соединение ДАФС(S) имеет сходную антиоксидантную активность с селеноорганическими соединениями 2 и 4. Поэтому, подводя итог представленным выше данным, можно заключить, что антиоксидантная активность изученных халькогенорганических соединений убывала в ряду: ДАФС(Se) > ДАФС(NO2)2) ≥ ДАФС(S) ≥ ДАФС(Cl2) > ДАФС(Те) > ДАФС(F2). Очевидно, что наличие боковых радикалов в фенильных заместителях, а также замена атома селена на теллур снижает антиоксидантные свойства халькогенорганических соединений. Предположительный механизм действия изученных халькогенорганических соединений имеет 3 аспекта. Во-первых, атом селена (теллура) может освобождаться из молекулы соединения и использоваться для синтеза селензависимых белков и ферментов, в т.ч. и ГПО, которые будут подавлять реакции свободно-радикального окисления и, тем самым, препятствовать ПОЛ. Освобождение атома селена, в свою очередь, возможно благодаря разрыхлению связи C-Se в молекуле соединения. Этому способствуют карбонильные группы в составе всех исследованных соединений, 240 которые обладают отрицательным индуктивным эффектом и способны оттягивать на себя электроны, приводя к разрыхлению связи C-Se и освобождению атома селена из молекулы. Галогены (атомы хлора и фтора) в составе соединений 2 и 3 соответственно имеют отрицательный индуктивный эффект и, обладая неподеленной парой электронов, оказывают положительный мезомерный эффект, который по величине больше отрицательного индуктивного эффекта. Следовательно, атомы хлора и фтора в составе препаратов 2 и 3 не вносят вклада в разрыхление связи C-Se [Тюкавкина Н.А., 2004]. галогены обладают окислительными свойствами, что Кроме того, обусловливает токсичность их соединений и, как следствие, более слабый антиоксидантный эффект по сравнению с соединением 1. Нитрогруппа в составе препарата 4 также способна вызывать разрыхление связи C-Se, т.к. она обладает отрицательным мезомерным эффектом и способна оттягивать на себя электронную плотность в сопряженной системе [Тюкавкина Н.А., 2004]. Таким образом, в разрыхление связи C-Se соединения 4 вносят вклад как карбонильные группы, так и нитрогруппы, поэтому, как мы предполагаем атом селена из соединения 4 будет освобождаться быстрее, чем из остальных халькогенорганических соединений 1, 2, 3 и 5. Это предположение подтверждается максимальной активностью селензависимой ГПО в цельной крови у мышей, получавших перорально соединение 4 по сравнению с другими экспериментальными группами животных. Активность ГПО, у мышей, получавших перорально соединение 4, увеличивалась на 98,9% по сравнению с контролем, что заметно выше по сравнению с другими исследованными препаратами (соединение 1 – активность ГПО возрастала на 64,6%, соединение 2 – на 51,9%, соединение 3 на 43,7% по сравнению с контролем, соединение 6 – изменения активности ГПО статистически недостоверны). Отсутствие повышения активности ГПО у мышей, получавших per os препарат 6, подтверждает предположение, что селеноорганические препараты являются 241 источником селена для синтеза селенсодержащих белков и ферментов, в том числе ГПО. Помимо этого предположения о механизме антиоксидантного действия халькогенорганических соединений есть версия, что атом селена в составе исследованных препаратов способен нейтрализовать активные формы кислорода (АФК). Во всех изученных соединениях присутствует халькоген (селен или теллур) в степени окисления +2, что определяет восстановительную природу соединения и, следовательно, его активное участие в процессах неферментативного перекисного окисления биологических молекул. Подобный восстановительный центр присутствует в молекуле диалкилсульфидов, для которых в настоящее время доказана способность к эффективному разложению гидроперекисей липидов, что предотвращает образование высокореакционных алкоксильных (RO˙) и перекисных радикалов (RO2˙): Учитывая общность в строении диалкилсульфидов и халькогенсодержащих арилалифатических дикетонов, можно предположить сходный характер их химических свойств и, прежде всего, способности разлагать гидроперекиси. Кроме того, селен характеризуется меньшей величиной относительной электроотрицательности по сравнению с серой и, следовательно, более эффективен при восстановлении гидроперекисей [Кирова, 2004]. Наконец, третий предположительный механизм действия халькогенорганических соединений связан с их фенильными радикалами. Максимальной антиоксидантной активностью обладало соединение 1, особенностью которого являлось отсутствие боковых радикалов у фенильных заместителей. Можно предположить, что фенильные радикалы в составе соединения 1 подвергались гидроксилированию при участии цитохрома Р 450 в ЭПР печени с образованием фенонильных групп, обладающих выраженной 242 антиксидантной активностью наравне с другими гидрофобными антиоксидантами, такими как токоферол, филлохинон, убихинон и т.д. [Кирова, 2004; Burton G.M., Traber M.G., 1990; Olson J.A., 1992]: Образование активных форм кислорода усиливается при отравлении солями тяжелых металлов, поскольку тяжелые металлы связываются с тиогруппами (-SH) биомолекул (например, глутатионом). Токсическое действие солей тяжелых металлов обусловлено взаимодействием иона металла с сульфгидрильными группами белков и ферментов, связыванием с атомами азота пуринов и пиримидинов в составе нуклеиновых кислот, ингибированием ряда ферментов, а также формированием активных форм кислорода, которые способны повреждать мембранные структуры клетки. В этой связи антитоксическое действие халькогенорганических соединений при подостром отравлении крыс солями тяжелых металлов (нитрат свинца, нитрат ртути, сульфат кадмия) обусловлено, с одной стороны, связыванием ионов тяжелых металлов, а, с другой, инактивацией радикалов кислорода, образованных под действием тяжелых металлов. Поэтому использованные в нашей работе серо- и селеноорганические соединения обладали выраженной антитоксической активностью. Как известно [Брин В.Б. и др., 2010, 2011, 2012], соли тяжелых металлов оказывают выраженное нефротоксическое действие на организмы животных, которое заключается в увеличении концентрации креатинина и мочевины в плазме крови. Гепатотоксическое действие тяжелых металлов [Брин В.Б. и др., 2011; Рукавишников В.С., 2011] выражается изменением активности ферментов плазмы крови аспартатаминотрансферазы (АсАТ), аланинамнотрансферазы (АлАТ), щелочной фосфатазы и γ-глутамилтрансферазы (ГГТ). 243 Несмотря на то, что все использованные соли тяжелых металлов обладали как нефротоксическим, так и гепатотоксическим действием, наибольший сдвиг в показателях функциональной активности тканей крыс наблюдался при введении per os нитрата свинца и сульфата кадмия. Из изученных халькогенорганических соединений наилучшими антитоксическими свойствами при подостром отравлении крыс сульфатом кадмия, нитратом ртути и нитратом свинца обладали препараты 1, 2, 4 и 5, поскольку при их использовании нормализовалась концентрация креатинина в крови и активность печеночных ферментов. При этом препараты ДАФС(Se) и ДАФС(S) эффективно предотвращали токсическое действие солей свинца и ртути на организм экспериментальных животных. Соединения 2 и 4 оказывали нефропротекторное действие при отравлении солями кадмия и свинца. Механизм антитоксического действия может быть связан со способностью атомов серы и селена в составе исследованных соединений связывать ионы тяжелых металлов, предотвращая их взаимодействие с сульфгидрильными группами белков и ферментов. И, наконец, следует подчеркнуть, что наилучшими антитоксическими свойствами обладало соединение ДАФС, поскольку этот препарат, вероятно, может поставлять атом селена для образования селеноцистеина, с последующим их включением в состав селенопротеинов и селеноферментов. Селеногидрильные группы селенопротеинов способны связывать ионы тяжелых металлов, защищая сульфгидрильные группы белков и ферментов: В заключение следует отметить изменение антитоксической активности исследованных халькогенорганических соединений в направлении: ДАФС(Se)> ДАФС(S)> ДАФС(NO2)2)>ДАФС(Cl2)> ДАФС(F2)> ДАФС(Те). 244 Вышесказанное позволяет сделать вывод, что антитоксическая активность халькогенорганических соединений зависит от антиоксидантных свойств этих соединений. Соединения 1 и 4 обладали как антиоксидантной, так и антитоксической активностью. А препараты 3 и 6 (ДАФС(F2) и ДАФС(Те)), напротив, обладали собственной токсичностью и поэтому не оказывали антитоксического действия при отравлении солями тяжелых металлов. Токсичность этих препаратов (ДАФС(F2) и ДАФС(Те)) обусловлена наличием в их структурах атомов фтора и теллура соответственно. Фтор – самый электроотрицательный элемент и самый мощный окислитель. Кроме того, атом фтора, вероятно, мог участвовать в образовании токсичных фторидов металлов, которые благодаря своей нерастворимости осаждались в гепатоцитах и оказывали токсическое действие на печень. Атом теллура более токсичен, чем сера и селен, поэтому аккумуляция теллуроорганического соединения в печени приводила к значительному токсическому эффекту. Именно по этой причине соединения ДАФС(F2) и ДАФС(Те) обладали выраженной антимикробной активностью в отношении клинических штаммов грамотрицательных и грамположительных бактерий. Высокая антибактериальная активность препарата 6 (ДАФС(Те)) обусловлена токсичностью атома теллура в его составе и сопоставима с активностью препарата 3 (фторсодержащего аналога препарата ДАФС) в отношении клинических штаммов золотистого стафилококка. Однако препарат 3 был самым эффективным среди исследованных в отношении клинических штаммов E. coli и P. aeruginosa. Выраженной антибактериальной активностью также обладал препарат 2 (хлорсодержащий аналог препарата ДАФС), который эффективно подавлял рост клинических штаммов золотистого стафилококка уже в низкой концентрации 0,001 мг/мл. Меньшую антибактериальную активность препарат 2 проявлял в отношении клинических штаммов кишечной палочки. Клинические штаммы синегнойной палочки были чувствительны только к высоким концентрациям препарата 2. 245 Антибактериальная активность препаратов 2 и 3 объясняется наличием в их структуре галогенов – фтора и хлора соответственно. Антимикробная активность препарата 3 сильнее, чем у препарата 2. Это связано, в первую очередь, с наличием в структуре препарата 3 (фторсодержащего аналога препарата ДАФС) двух атомов фтора. Известно, что фтор самый электроотрицательный элемент и самый мощный окислитель, поэтому благодаря наличию двух атомов фтора препарат 3 приобретает прооксидантные свойства и, как следствие, становится самым токсичным из всех исследованных халькогенорганических препаратов, проявляя максимальную антибактериальную активность. Окислительные свойства хлора значительно слабее, чем у фтора. Кроме того, хлор, являясь важным биогенным элементом, участвует во многих жизненно важных процессах (водно-минеральном обмене, обезвреживании аммиака, переваривании белков в желудке, активации амилолитических ферментов и т.д.). Поэтому хлорсодержащий аналог препарата ДАФС (препарат 2) обладал меньшей антимикробной активностью по сравнению с препаратом 3. Кроме того, его активность слабее, чем у препарата 5 (теллурорганического соединения), но гораздо выше, чем у препаратов 1 и 4. Сравнительно низкая антибактериальная активность препаратов 1 и 4, вероятно, связана с их низкой токсичностью и высокими антиоксидантными и антитоксическими свойствами. Однако препарат 4 проявлял большую антибактериальную активность по сравнению с препаратом 1 (ДАФС) в отношении клинических штаммов золотистого стафилококка, кишечной и синегнойной палочки. При этом данный препарат в концентрации 0,01 мг/мл более эффективно подавлял рост клинических штаммов кишечной палочки, в меньшей степени - синегнойной палочки и не был эффективен против штаммов золотистого стафилококка. Особый интерес представляет наличие в структуре препарата 4 двух нитрогрупп, что позволяет проводить аналогию с антибактериальным 246 действием нитрофуранов, хотя нитрофураны проявляют широкий спектр антибактериальной активности уже при концентрации 0,5-50 мкг/мл. Установлено [Гасанова З.М., 1966], что сами по себе нитрофураны антибактериальной активностью не обладают, но приобретают ее при восстановлении флавинзависимыми редуктазами, т.е. для проявления как терапевтического, так и побочного действия нитрофуранов необходимо их одноэлектронное восстановление ферментами с нитроредуктазной активностью, локализованными в микробах, простейших или тканях организма [Кирпичев В.И., 1972; 1973]. Промежуточные продукты последовательных одно- или двухэлектронных этапов восстановления являются высоко реакционноспособными, особенно анион нитрорадикала, благодаря которому нитрофураны приобретают антибактериальную, противоопухолевую и генотоксическую активность [Глыбочко П.В. и др., 2010]. Кроме того, следует отметить большую чувствительность клинических штаммов золотистого стафилококка к исследованным халькогенорганическим соединениям по сравнению со штаммами грамотрицательных бактерий (кишечной и синегнойной палочки). При этом чувствительность клинических штаммов кишечной и синегнойной палочки к исследованным соединениям была сопоставима. Особенности грамотрицательных строения бактерий мембран грамположительных обусловливают различие и в проходимости разнообразных веществ, этим можно объяснить неэффективность большинства синтетических и натуральных антимикробных агентов против грамотрицательных бактерий, таких как Escherichia coli и Pseudomonas aeruginosa (см. рис. 9). Наружная мембрана этих бактерий состоит из липополисахарид-липопротеиновых комплексов и недоступна для большинства антибиотиков, включая пенициллин и другие препараты [Soni D. et al., 2005]. Исследованные в данной работе халькогенорганические соединения являются низкомолекулярными гидрофобными соединениями, которые, 247 вероятно, легко проникали через липополисахаридный слой внешней мембраны грамотрицательных бактерий. Таким образом, главной функцией селена и селенсодержащих соединений является их участие в синтезе селенопротеинов и селеноферментов, защищающих клетки от действия активных форм кислорода, ионов тяжелых металлов и других токсикантов. Главным антиоксидантным селено-зависимым ферментом, как уже было обозначено выше, является ГПО, способная разлагать токсичную перекись водорода до двух молекул воды. Однако исследования последних лет показывают [Jiang Y., 2012; Sartoretto J.L., 2012; Koziel R. et al., 2013; Shibata A. et al., 2013], что перекись водорода является важной регуляторной молекулой и участвует в передаче рецептор-опосредованного сигнала в клетки. Основная часть перекиси водорода в клетках образуется в процессе дисмутации супероксид-анион радикала (O2− ∙). Последний образуется в процессе функционирования дыхательной цепи переноса электронов (цпэ) путем одноэлектронного восстановления кислорода ферментами комплексов I и III цпэ, а также глицерол-3-фосфатдегидрогеназы. Большая часть супероксида обезвреживается с образованием перекиси водорода под действием Mnзависимой супероксиддисмутазы (Mn-СОД) в матриксе митохондрий или под действием Cu/Zn-СОД в цитозоле. Другим важным источником этой АФК является НАДФН-оксидаза (Nox), участвующая в работе фагоцитирующих иммунных клеток за счет выработки большого количества супероксида, необходимого для уничтожения поглощенных патогенных клеток. НАДФН-оксидаза обнаружена на внутренней поверхности плазматической мембраны нефагоцитирующих клеток, где она связана с цитоплазматическими доменами рецепторов. Известны 7 белков НАДФН-оксидазы: Nox1-Nox5 и двойные оксидазы 1 и 2. При связывании специфического лиганда с рецептором НАДФН-оксидаза активируется и вырабатывает супероксид, который затем превращается в пероксид водорода 248 под действием СОД. Недавние исследования показали (DeYulia G.J. et al., 2005), что перекись водорода может вырабатываться непосредственно при образовании лиганд-рецепторного комплекса. Образованный супероксид и/или перекись водорода вызывают активацию киназной активности комплекса, который запускает серию реакций фосфорилирования белков, специфичных для этого рецептора. Например, связывание инсулина с рецептором инициирует каскадный механизм передачи сигнала в клетку. Участниками этого процесса являются субстрат инсулинового рецептора (IRS-ИРС), протеин-тирозин-фосфатаза (PTPПТФ) и протеинкиназа В (серин/треонинкиназа Akt), а также транскрипционный фактор О1а (Fox) и его пероксисомальный пролифераторактивированный рецепторный гамма коактиватор (PGC)-1α. На рисунке 17 представлена схема потенциальной активности соединений селена на компоненты инсулинового каскада передачи сигнала. При связывании инсулина со своим рецептором на плазматической мембране адипоцита инсулин стимулирует кратковременный «взрыв» активных форм кислорода (супероксида и перекиси водорода). Это происходит благодаря тому, что инсулин активирует НАД(Ф)Н оксидазу (Nox) 4 для выработки супероксида, который затем превращается в перекись водорода. Это небольшое количество перекиси водорода служит вторичным посредником, который уменьшает активность фосфатаз с редокс-чувствительными остатками цистеина и таким образом увеличивает фосфорилирование компонентов сигнального каскада инсулина. Небольшое увеличение концентрации перекиси водорода (0,1-1 мкмоль/л) стимулирует торможение клеточного цикла, старение и/или апоптоз, тогда как миллимолярные концентрации перекиси водорода могут вызвать мощное окислительное повреждение и некротическую гибель клетки. Поверхностные клеточные рецепторы, вырабатывающие АФК после активации, включаются эпидермальным фактором роста, фактором роста 249 тромбоцитов, инсулиноподобным фактором роста, фактором роста сосудов и различными цитокинами (рис. 17). Рис. 17. Схема потенциальной активности селена на компоненты инсулинового каскада передачи сигнала. Селенопротеины и низкомолекулярные соединения селена могут вмешиваться в различные стадии инсулин-индуцированной трансдукции сигнала, в конечном счете приводя к разлаживанию углеводного метаболизма (адаптировано на русский язык по Steinbrenner H. et al., 2011). Обозначения: ИР – инсулиновый рецептор; ИРС – субстрат инсулинового рецептора, Nox4 – НАДФН-оксидаза; PI3K – фосфоинозитид-3-киназа; PIP3 – фосфатидилинозитол-3,4,5-трисфосфат; Akt – протеинкиназа В (серин/треонинкиназа); GLUT4 – белок-транспортер глюкозы; РТЕN – протеинфосфатаза с двойной специфичностью; РТР-1В – протеин тирозинфосфатаза; DBE – деветвящий фермент; FoxO1a – «вилкоголовый» бокс; PGC-1a – пероксисомальный пролифератор-активированный рецепторный гамма коактиватор. 250 АФК регулируют аутофагию, проникновение кальция в митохондрии и освобождение кальция из эндоплазматического ретикулума, который необходим для проведения кальциевого сигнала к митохондриям. АФК, образованные в митохондриальной цпэ, выходят в цитоплазму, где они могут реагировать с молекулами-мишенями, чтобы запустить клеточный ответ. Благодаря такому участию перекиси водорода в жизни клетки, можно ее назвать новым вторичным посредником. Концентрация перекиси водорода зависит от активности ГПО в различных органах и тканях. Вместе с тем, синтез ГПО напрямую зависит от поступления в организм селена и от метаболизма главного селен-транспортного белка селенопротеина Р (М=57кД, в его составе 10 или 11 атомов селена), который синтезируется, главным образом, в печени и обеспечивает периферические ткани селеном [Steinbrenner H. et. al., 2011]. Селенопротеин Р представляет собой биомаркер селенового статуса, поскольку его концентрация в плазме повышается в ответ на различные пищевые формы и дозы селена. У человека промотор селенопротеина Р состоит из связывающего участка для транксрипционного фактора FoxO 1a, локализованного в непосредственной близости от связывающего участка для ядерного фактора 4α (HNF-4α) гепатоцитов. Этот участок обнаружен в промоторах селенопротеина Р человека, крыс, мышей; он обусловливает высокий уровень экспрессии селенопротеина Р в печени и гормональную регуляцию транскрипции печеночного селенопротеина Р. Транксрипционный фактор FoxO 1a также участвует в регуляции экспрессии генов ферментов глюконеогенеза и гликогенолиза (рис. 26). Оба транскрипционных фактора (FoxO 1a и HNF-4α) коактивируются PGC1α (пероксисомный пролифератор-активированный рецепторный гамма коактиватор), который действует как «молекулярный переключатель» в ответ на гормоны, такие как инсулин, глюкагон и глюкокортикоиды. Хорошо известно контролирующее влияние этих гормонов на продукцию глюкозы печенью и уровень глюкозы крови. Инсулин ингибирует транксрипцию гена селенопротеина Р через ось PI3K/Akt/FoxO 1a, тогда как PGC-1α-индукция 251 глюкокортикоида дексаметазона значительно увеличивает уровни мРНК селенопротеина Р в культуре гепатоцитов крысы [Speckmann B. et al., 2008]. Введение глюкокортикоидного препарата дексаметазона per os приводит к перераспределению селена у мышей, вызывая уменьшение печеночной ГПО в пользу повышения уровня селена в составе селенопротеина Р в плазме крови. Следовательно, комплекс между FoxO1а и его коактиватором PGC-1α играет чрезвычайно важную роль для транскрипционной регуляции как ферментов глюконеогенеза – глюкозо-6-фосфатазы (Г6Фаза) и фосфоенолпируваткарбоксикиназы (ФЕПКК) [Yoon J.C. et al., 2001; Puigserver P., and et al. 2003], так и селенопротеина Р [Speckmann B. and et al., 2008], обеспечивая гипотетическую связь между метаболизмом селена и углеводным обменом. Можно предположить, что селенопротеин Р и низкомолекулярные соединения селена могут затрагивать инсулин-индуцированные сигнальные пути, регулирующие углеводный и липидный обмены. Кроме того, следует отметить, что повышение печеночного PGC-1α приводит не только к развитию гипергликемии, но и к нарушению гомеостаза селена. Например, антигипергликемическое средство метформин широко применяется для лечения сахарного диабета 2 типа, т.к. он подавляет выработку печенью глюкозы и повышает чувствительность к инсулину периферических тканей. Параллельно с глюконеогенезом метформин ослабляет биосинтез и секрецию селенопротеина Р in vitro, а также снижает биодоступность селена во внепеченочных тканях и за счет этого ослабляет экспрессию и активность селеноферментов in vivo [Steinbrenner H. et al., 2011]. Глюкокортикоидный препарат дексаметазон, напротив, приводит к увеличению секреции селенапротеина Р печенью и развитию гипергликемии. Гипергликемический эффект дексаметазона можно объяснить его индуцирующим действием на экспрессию гена киназы пируватдегидрогеназы [Qi D. et al., 2004], следствием чего является снижение активности этого мультиферментного комплекса, замедление процесса окисления глюкозы и 252 накопление пирувата и глюкозы в цитозоле клеток. Кроме того, дексаметазон уменьшает фосфорилирование фермента гликогенсинтазы, что приводит к восстановлению ее активности. Избыток глюкозы (при наличии активной гликогенсинтазы) используется для синтеза гликогена в печени, скелетных мышцах и миокарде [Qi D. et al., 2004; Puthanveetil P. et al., 2008]. Также следует отметить, что глюкортикоиды увеличивают экспрессию ферментов глюконеогенеза - глюкозо-6-фосфатазы и фосфоенолпируваткарбоксикиназы [Yabaluri N., Bashyam M.D., 2010], что влечет за собой усиление реакций глюконеогенеза в печени, выход свободной глюкозы в кровь и развитие гипергликемии. Глюкокортикоиды усиливают гипергликемию также за счет снижения поступления глюкозы в клетки периферических тканей [Baxter J.D., 1976; Andrews R.C., Walker B.R., 1999; Vila G. et al., 2010]. Подводя итог, можно заключить, что поступление селена с пищей и/или в составе синтетических селенсодержащих препаратов приведет в усилению синтеза селенопротеина Р и ферментов глюконеогенеза в печени. Действительно, исследования с применением различных неорганических и органических соединений селена, проведенные на различных животных и птицах (коровы, быки, свиноматки, овцы, утки, гуси и пр.), показывали увеличение концентрации глюкозы в крови [Кузнецов Ю.А., 2002; РодионоваТ.Н., 2004; Иванова И.В,. 2009; Поперечнева Т.Ю., 2009]. Наше исследование также не было исключением из правил. Однако достоверное увеличение глюкозы в плазме крови (40,6%) демонстрировал только препарат ДАФС-25. Действие всех остальных халькогенорганических соединений на мышей не приводили к значимому изменению этого показателя. Вместе с тем, применение препарата ДАФС-25 также приводило к изменению ряда других показателей углеводного обмена. Например, содержание гликогена в печени возрастало на 48,6%, в крови - на 32,0%. Кроме того, было обнаружено достоверное снижение концентрации пирувата в плазме крови (на 39,4%) на фоне увеличения активности аланинаминотрансферазы на 140,5%. Однако активность другой аминотрансферазы АсАТ оставалась неизменной по 253 сравнению с контролем. И, наконец, при изучении влияния халькогенорганических соединений на активность ферментов глюконеогенеза глюкозо-6-фосфатазу и фруктозо-1,6-бисфосфатазу было обнаружено достоверное увеличение активности только глюкозо-6-фосфатазы у мышей, получавших per os препарат ДАФС-25, тогда как активность фруктозо-1,6бисфосфатазы не изменялась по сравнению с контролем. Большинство из вышеперечисленных биохимических изменений свидетельствуют об усилении реакций глюконеогенеза в печени и снижении поступления глюкозы в клетки периферических тканях, что согласуется с литературными данными, приведенными выше. Как известно, соединения селена участвуют в синтезе ГПО, разрушающей перекись водорода, в результате чего снижается концентрация Н2О2, что приводит к активации фосфатаз (РТР-1В и РТЕN) и, как следствие, к дезорганизации внутриклеточной передачи инсулинового сигнала (рис. 26). Результатом этого является, с одной стороны, снижение концентрации инозитол-1,4,5-трисфосфата в цитоплазме клетки и нарушение включения транспортера глюкозы (GLUT4) в цитоплазматическую мембрану клеток жировой и мышечной тканей, а, с другой, изменение экспрессии генов ферментов углеводного обмена. Такое действие соединений селена сравнимо с действием природных и синтетических глюкокортикоидов, что позволяет рассматривать соединения селена как потенциальные гормоноподобные препараты. Однако полученные нами результаты показывают, что только соединение ДАФС-25 (соединение 1) способствовал достоверному повышению концентрации глюкозы в крови и гликогена в печени, а остальные соединения подобного действия не оказывали. Подробное изучение отечественной литературы, касающейся биологической активности органических и неорганических соединений селена, выявило интересные закономерности: органические соединения селена оказывают большее влияние на показатели углеводного и липидного обменов животных, чем неорганические. 254 В подавляющем большинстве исследований изучался препарат ДАФС-25 и/или другие формы селена: селенит натрия и селенопиран. Результаты по применению препарата ДАФС-25 [Кузнецов Ю.А., 2002; Родионова Т.Н., 2004; Иванова И.В,. 2009; Поперечнева Т.Ю., 2009] демонстрируют достоверное увеличение глюкозы от 26% [Родионова Т.Н., 2004] до 55% [Кузнецов Ю.А., 2002] по сравнению с контрольными группами животных. Во всех перечисленных выше работах также было показано увеличение в крови концентрации общих липидов, α- и β-липопротеинов, а также фосфолипидов. Кроме того, обращает на себя внимание значительное увеличение концентрации общего белка и его фракций в сыворотке крови животных, принимавших препарат ДАФС-25 от 50% [Родионова Т.Н., 2004] до 73% [Поперечнева Т.Ю., 2009]. Применение неорганического селенита натрия [Сайфульмулюков Э.Р., 2006] сопровождалось увеличение концентрации глюкозы (на 18,2%), общего белка (на 6,8%) и общих липидов (на 32%). Помимо этого в большинстве работ, проведенных на сельско- хозяйственных животных, обращало на себя внимание достоверное увеличение ежесуточного и общего прироста живой массы. Причем прирост живой массы животных, получавших селенит натрия колебался в пределах от 4% [Перунова Е.В., 2000; Сайфульмулюков Э.Р., 2006;] до 5,4% [Родионова Т.Н., 2004]; применение препарата ДАФС обнаружило прирост живой массы от 6% [Шперов А.С., 2009] до 10-13% [Перунова Е.В., 2000; Родионова Т.Н., 2004] у свиней; 12,4% у цыплят [Сотников Д.А., 2002] до 39,3% у кроликов [Родионова Т.Н., 2004]. В работах Перуновой Е.В., Брусковой О.Б., Сотникова Д.А., Родионовой Т.Н., и Шперова А.С. [Перунова Е.В., 2000; Брускова О.Б., 2002; Сотников Д.А., 2002; Родионова Т.Н., 2004; Шперов А.С., 2009], проводился сравнительный анализ влияния соединений селенита натрия и/или ДАФС-25 и/или селенопирана, которые показали достоверное увеличение прироста 255 живой массы в направлении селенопиран > ДАФС > селенит натрия по сравнению с контролем. Особый интерес вызвала работа Трошиной Т.А. [Трошина Т.А., 2010], посвященная фармакокоррекции селенодефицита у животных препаратом ДАФС-25 и его влияние на продуктивные качества, результаты которой показали более высокий уровень активности гипофизарно-тиреоидно- надпочечниковой системы сельскохозяйственных животных и пушных зверей и полученного от них молодняка, а также активацию анаболических процессов и увеличение андрогенного статуса у жеребцов, обуславливающих их интенсивный рост. Кроме того, полученные результаты показали повышение адаптационных возможностей организма, стимулирование роста, активацию глюконеогенеза, за счет которого организм обеспечивался глюкозой на 8590%. Таким образом, добавление в корма селеноорганических препаратов ДАФС-25 и селенопирана вызывали более значительное усиление роста животных по сравнению с селенатом натрия и контрольными животными. Все вышеизложенное позволяет высказать предположение о возможном гормоноподобном действии препарата ДАФС-25, которое согласуется и с полученными нами результатами (гипергликемия, гиперхолестеринемия, увеличение концентрации гликогена в печени и крови, а также повышение активности одного из ключевых ферментов глюконеогенеза глюкозо-6фосфатазы). Для выяснения и подтверждения такого гормоноподобного действия мы провели сравнение структуры селеноорганических соединений ДАФС-25 и селенопирана с известными стероидными гормонами и нестероидными препаратами, оказывающими гормоноподобное действие. Мы обнаружили, что соединение ДАФС-25 в оптимизированной форме имеет черты сходства с гормональным препаратом преднизолоном и кормовой добавкой рактопамином [Бородулин В.Б., Русецкая Н.Ю., 2013]. На рисунке 18 представлено химическое строение гормонального препарата преднизолон, 256 селеноорганического соединения ДАФС-25 и кормовой добавки рактопамина в оптимизированной форме: CH2OH H3C O CO HOCH2 OH O O H3C OH H3C H3C O O Преднизолон в оптимизированной Преднизолон форме O Se O Se OO ДАФС-25 в оптимизированной ДАФС-25 форме OH HO HO OH N H NH OH HO Рактопамин Рактопамин в оптимизированной форме Рис. 18. Химическое строение гормонального препарата преднизолон, селеноорганического соединения ДАФС-25 и кормовой добавки рактопамина в оптимизированной форме. 257 Фармакологически, рактопамин (запрещенный к применению в России) – это бета-адренергический агонист, принадлежащий к классу фенетаноламинов [Liu C.Y. et al., 1994; Mills S.E., 2001]. При использовании в качестве пищевой добавки рактопамин доставляется через кровь в мышечные клетки, где он связывается с бета-адренорецепторами, стимулируя синтез белка и приводя в конечном итоге к общему росту мышечной массы животных. Анаболический эффект ДАФС-25 и рактопамина, по нашему мнению, обусловлен определёнными пространственной структурой чертами сходства стероидных этих гормонов, препаратов в том с числе глюкокортикоидов. Кроме того, обращает на себя внимание структурное сходство селеноорганических гормоноподобными препаратов ДАФС-25 нестероидными и селенопирана веществами, с обладающими гормоноподобной активностью: Диацетофенонилселенид (ДАФС-25) Селенопиран (соединение 1) Метилстильбол Хлортрианизен (тейс) Вероятно, диацетофенонилселенид и селенопиран в оптимизированных конформациях способны взаимодействовать с рецепторами глюкокортикоидов и половых гормонов и через них оказывать гормоноподобное действие на клетки животных. 258 Анализ литературных данных подтверждает, что гормоноподобные вещества стероидной и нестероидной природы способны взаимодействовать с лиганд-связывающими доменами глюкокортикоидных, андрогеновых и/или прогестероновых рецепторов [Schäcke H., et al., 2008; Hudson A.R., et al., 2008; Mohler M.L., et al., 2008; Biggadikea K., et al., 2009; Petit-Topin I. et al., 2009; Caboni L., et al., 2012]. А, значит, такой способностью, скорее всего, обладает и селеноорганический препарат ДАФС. Так, например, в работе [Petit-Topin I. et al., 2009] рассматривались особенности строения лиганд-связывающего домена прогестеронового рецептора (ПР) и механизм присоединения к нему левоноргестрела. На рисунке 19 показаны взаимодействия между лигандсвязывающим доменом прогестеронового рецептора и левоноргестрелом. А. Рис. 19. Кристаллическая Б. структура лиганд-связывающего домена прогестеронового рецептора в комплексе с левоноргестрелом. А. Диаграмма, показывающая взаимодействия между лигандсвязывающим доменом прогестеронового рецептора и левоноргестрелом. Водородные связи и ван дер Ваальсовые взаимодействия обозначены как непрерывные черные стрелки и пунктирные черные линии соответственно. W – обозначает молекулу воды. В. Структура лигандсвязывающего домена прогестеронового рецептора, связанная с левоноргестрелом (серый), наложенным на прогестерон (голубой). Фиолетовым цветом обозначены остатки аминокислот, принадлежащие 3-й спирали, ярко-розовым цветом – 5-й спирали, зеленым цветом – 7-й спирали, красным цветом – 11-й спирали, сиреневым цветом – 12-й спирали. 259 Внутри лиганд-связывающей полости остатки Глн-725 (Q725) и Арг-766 (R766) (рис. 19 Б) закрепляют С3-кето-группу прогестерона и его аналогов водородными связями и контактируют с молекулой воды. Остаток Асн719(N719) связан водородной связью с 17- β-гидроксигруппой левоноргестрела через молекулу воды. Несколько аминокислотных остатков придают лигандсвязывающему карману гидрофобность и стабилизируют положение левоноргестрела посредством ряда ван дер Ваальсовых взаимодействий (рис. 19). Проводя аналогию между вышеописанными взаимодействиями левоноргестрела с лиганд-связывающим доменом прогестеронового рецептора и препаратом ДАФС-25 в оптимизированной форме, можно предположить, что за связывание с рецептором будут отвечать следующие аминокислотные остатки: В связи с вышеизложенным для подтверждения этой теории нами были проведены квантовохимические расчеты, подтверждающие структурное сходство селеноорганического соединения ДАФС-25, его производных и глюкокортикоидов. Согласно полученным результатам, из всех рассмотренных нами 1,5дикарбонилов молекула препарата ДАФС-25 была наиболее близка к молекулам глюкокортикоидов как по размеру (а0, Vm), так и по расстояниям (l1 и l2) между полярными реакционноспособными группами С=О, С-ОН, которые 260 потенциально могут участвовать в связывании с рецептором как посредством электростатических сил, так и за счет водородных связей или ван дер Ваальсовых взаимодействий. Таким образом, налицо как геометрические (а0,Vm, l1, l2 и μ), так и электростатические (μ) предпосылки сходного характера биологической активности препарата ДАФС-25 и глюкокортикоидов. Квантовохимический прогноз, основанный на установлении элктроннотопологического подобия молекул препарата ДАФС-25 и кортикостероидов, подтвержден нами экспериментально. Подводя итог, можно сделать вывод о возможности взаимодействия различных по строению нестероидных соединений с лиганд-связывающими доменами рецепторов стероидных гормонов (андрогенов, эстрогенов и глюкокортикоидов). Дальнейшие исследования в этой области помогут выявить потенциальные обладающие гормоноподобные анаболическим, вещества нестероидной природы, андроген/эстроген-модулирующим и противовоспалительным действием, что позволит правильно оценить их биологическую активность и корректно использовать в медицине, ветеринарии, животноводстве, птицеводстве и других областях народного хозяйства. 261 Выводы активность халькогенорганических 1. Антиоксидантная убывает в ряду: соединений ДАФС(Se)>ДАФС(NO2)2)≥ДАФС(S)≥ДАФС(Сl2)> ДАФС(F2)>ДАФС(Те). Наличие боковых радикалов у фенильных заместителей, а также замена атома селена на теллур в составе халькогенорганических соединений подавляет процессы свободнорадикального окисления липидов во всех исследованных тканях белых беспородных мышей: мозге ≥ легких > эритроцитах > плазме крови > печени ≥ почках. 2. Антитоксическая активность халькогенорганических соединений при интоксикации солями тяжелых металлов (CdSO4, Hg(NO2)2 и Pb(NO2)2) зависит от антиоксидантных свойств исследованных соединений и снижается в направлении: ДАФС(Se)>ДАФС(S)>ДАФС(NO2)2)> ДАФС(Cl2)>ДАФС(F2)> ДАФС(Те). 3. Препараты ДАФС(F2) и ДАФС(Те) значительно подавляют рост колоний клинических штаммов Staphylococcus aureus, Escherichia coli и Pseudomonas aeruginosa даже в низких концентрациях 0,0001-0,001 мг/мл, что соответствует результатам использованием компьютерного прогнозирования с программы PASS. Меньшую антибактериальную активность проявляет препарат ДАФС(Cl2). Наличие атомов хлора и фтора у фенильных заместителей, а также замена атома селена на теллур усиливают антибактериальную соединений, которая активность халькогенорганических убывает в ряду: ДАФС(F2)>ДАФС(Te)>ДАФС(Cl2)>ДАФС(NO2)2>ДАФС(Se)>ДАФС(S). 4. Препарат ДАФС-25 в отличие от других халькогенорганических соединений стимулирует глюконеогенез и гликогеногенез у экспериментальных животных подобно глюкокортикоидному препарату преднизолону. 5. Молекула препарата ДАФС-25 наиболее близка по структуре к глюкокортикоидам преднизону и преднизолону: размеру и расстояниям 262 между полярными реакционноспособными группами С=О, С-ОН, которые потенциально могут участвовать в связывании с рецептором. 6. В ряду халькогенорганических соединений, лишенных боковых заместителей, но содержащих гетероатомы халькогенов - ДАФС(Se), ДАФС(S), ДАФС(Te), а также в ряду производных ДАФС-25, содержащих боковые заместители - ДАФС(NO2)2, ДАФС(Cl2), ДАФС(F2), снижаются антиоксидантная и антитоксическая активности, возрастает антибактериальное действие. 7. Теоретически обоснован и экспериментально подтвержден новый подход к пониманию лиганд-рецепторного взаимодействия, в основу которого положено представление о структурно-функциональном подобии молекул, относящихся к различным классам органических соединений. 263 Список использованной литературы 1. Абатуров А. Е. Активированные кислородсодержащие метаболиты компонент системы неспецифической защиты респираторного тракта // Здоровье ребенка. - 2009. - №2.- С.120-125. 2. Артюхов В.Г., Наквасина М.А. Биологические мембраны: структурная организация, функции, модификация физико-химическими агентами: Учеб. пособие. – Воронеж: Издательство Воронежского государственного университета, 2000. - 296 с. 3. Бабаскин Б.Н. Модифицированный метод Умбрайта определения пировиноградной кислоты // Лаб. дело. - 1976. - №6. - С. 173. 4. Баранова Т.А. Гигиеническое и эпидемиологическое обоснование системы профилактики селенодефицитных состояний у населения Омской области. Дисс. … канд. мед. наук. Санкт-Петербург, 2008. - 134 с. 5. Бессонова Л.О., Верлан Н.В., Колесниченко Л.С. Роль системы глутатиона в антиоксидантной защите при сочетанной патологии гипоксического генеза // Сибирский медицинский журнал. – 2008. - № 6. - С. 19-21. 6. Бикчантаев И.Т., Шакиров Ш.К. Мясная продуктивность и экономическая эффективность использования препарата «Сел-Плекс» в рационах бычков на откорме // Ученые записки казанской государственной академии ветеринарной медицины им. Н.Э. Баумана. - 2013. - Т. 213. - С. 31-36. 7. Бородулин В.Б., Русецкая Н.Ю. Рактопамин, преднизолон и диацетофенонилселенид: структурное сходство и биологическая активность // Фундаментальные исследования. - 2013. - №4. - С. 1124-1127. 8. Браунштейн А.Е. Основные этапы изучения энзиматического переноса аминорупп // Ист. биол. иссл. - 1983. - С.16-50. 9. Брин В.Б., Кокаев Р.И., Бабаниязов Х.Х., Пронина Н.В. Профилактика ацизолом нефротоксического действия сулемы в хроническом эксперименте // Вестник новых медицинских технологий. – 2010. - Т. XVII. - №1. - С. 26-28. 264 10. Брин В.Б., Митциев А.К., Митциев К.Г. Способ коррекции гепатотоксического действия кадмия в эксперименте // Вестник новых медицинских технологий. – 2011. - Т. XVIII. - №4. - С. 209-211. 11. Брин В.Б., Митциев А.К., Митциев К.Г. Способ коррекции нефротоксического действия кадмия в эксперименте // Вестник новых медицинских технологий. – 2011. - Т. XVIII. - №2. - С. 194-195. 12. Брин В.Б., Митциев А.К., Митциев К.Г. Профилактика изменений структуры тканей сердца и почек при хроническом отравлении ацетатом свинца в эксперименте // Вестник новых медицинских технологий. – 2012. – Т. XIX. - №1. - С. 166-168. 13. Брускова О.Б. Влияние различных соединений селена на морфофункциональное состояние эндокринных желез, органов иммунной системы и скелетных мышц свиней. Дисс. … канд. биол. наук. Боровск. 2002. - 133 с. 14. Всемирная организация здравоохранения, Серия технических докладов, 797. Рацион, питание и предупреждение хронических заболеваний: доклад Исследовательской группы ВОЗ. Женева, 1993. - 208 с. 15. Всемирная организация здравоохранения Селен. Гигиенические критерии состояния окружающей среды. - 1989. - 270 с. 16. Галочкин В.А., Блинохватов А.Ф., Боряев Г.И., Колоскова Е.М. Селенопиран - новый высокоэффективный антиоксидант // 5-я Международная конференция «Биоантиоксидант». - М., 1998. 17. Гасанова З.М. Влияние фуразолидона на стафилококковую инфекцию в эксперименте: Автореф. … канд.мед. наук. Саратов, 1966. - 21 с. 18. Гланц С. Медико-биологическая статистика / Пер. с англ. Ю.А. Данилова; под ред. Н.Е. Бузикашвили, Д.В. Самойлова. М.: Практика, 1998. - 459 с. 19. Глыбочко П.В., Свистунов А.А., Бородулин В.Б., Русецкая Н.Ю. Нитрофураны: химическое Монография. Саратов: университета, 2010 – 199 с. строение Издательство и биологическая Саратовского активность. медицинского 265 20. Гмошинский И.В., Мазо В.К. Селен в питании: краткий обзор // Medicina Altera. - 1999. - № 4. - С. 18-22. 21. Гмошинский И.В., Мазо В.К., Тутельян В.А., Хотимченко С.А. Микроэлемент селен: роль в процессах жизнедеятельности // Экология моря. - 2000. - №54. - С. 5-19. 22. Гоголева И.В., Громова О.А. Селен. Итоги и перспективы применения в педиатрии // Практика педиатра. - март 2009. - С. 6-9. 23. Гореликова Г.А., Маюрникова Л.А., Позняковский В.М. Нутрицевтик селен: недостаточность в питании, меры профилактики // Вопр. питания. 1997. - №5. - С. 18-21. 24. Гринвуд Н.Н., Эрншо А. Химия элементов. В 2-х томах. М.: Бином, 2008. - Т. 2 - 666с. 25. Давыденко Н.И., Нестерова В.А., Карчевная А.И. Обоснование необходимости комплексного обогащения при разработке функциональных хлебобулочных изделий // Ползуновский вестник. - 2012. - №2/2. - С. 201-206. 26. Древко Б.И. Халькогенсодержащие гетероциклические соединения на основе 1,5-дикетонов. Синтез, свойства и некоторые закономерности реакций: Дисс….докт. хим. наук. - Саратов, 1997. - 362 с. 27. Фоменко Л. А. , Маркова Л. И., Древко Р.И., Родионова Т.Н., Ефремов В.И., Харченко В.Г. Средство для лечения и профилактики болезней, вызываемых недостаточностью селена в организме сельскохозяйственных животных и птиц. Пат. № 2051681 РФ // Бюл. изобрет. - 1996. - № 1. 28. Древко Б.И., Древко Р.И., Антипов В.А., Чернуха Б.А., Яковлев А.Н. Пат. 2171110 Россия. A61K33/04. Средство для лечения и профилактики инфекционных заболеваний и отравлений животных и птиц, повышающее их продуктивность и сохранность / Заявл. 26.05.1999, № 99111064/13; Опубл. 27.07.2001. - 8 с. // Изобретения. Полезные модели. - 2001. - Бюл. №21. 266 29. Дубинина Е.Е., Сальникова Л.А., Ефимова Л.Ф. Активность и изоферментный спектр супероксиддисмутазы эритроцитов и плазмы крови человека // Лаб. дело. – 1983. – №10. – С. 30-33. 30. Зайцев В.Г., Островский О.В., Закревский В.И. Связь между химическим строением и мишенью действия как основа классификации антиоксидантов прямого действия // Эксперим. клин. фармакол. – 2003.– Т.66. - № 4.– С.6670. 31. Зупанец И.А. Клиническая лабораторная диагностика: методы исследования: Учеб. пособие для студентов спец. «Фармация», «Клиническая фармация», «Лабораторная диагностика» вузов / И.А. Зупанец, С.В. Мисюрева, В.В. Прописнова и др.; Под ред. И.А. Зупанца. — 3-е изд., перераб. и доп. — Харьков: Изд-во НФаУ: Золотые страницы, 2005. — 200 с. 32. Иванова И. В. Влияние различного уровня селена в рационе бычков при откорме на их продуктивность и обмен веществ. Дисс. … канд. сель.хоз.наук. Дубровицы, 2009. - 100 с. 33. Исидоров В.А. Введение в химическую экотоксикологию: Учеб. пособие. – Спб: Химиздат, 1999. – 144 с. 34. Калинина Е. В., Чернов Н. Н, Саприн А. Н. Участие тио-, перокси- и глутаредоксинов в клеточных редокс-зависимых процессах // Успехи биологической химии. - 2008. - Т. 48. - С. 319-358. 35. Кальф-Калиф Я.Я. О лейкоцитарном индексе интоксикации и его практическом значении // Врачебное дело. — 1941. — №1. — С. 31–35. 36. Капитальчук М. Селен в природных водах и биогеохимической цепи «почва-растение» региона Украинской лесостепной и степной почвенных провинций Республики Молдова: Дисс. … докт. биол. наук. Кишинев, 2008. - 298 с. 37. Карпищенко А.И. Медицинские лабораторные технологии. Справочник / Под редакцией Карпищенко А.И. СПб.: Интермедика, 1999. В 2-х томах. Т.2. - С. 27-28. 267 38. Кильметова И.Р., Толмачев П.В., Струнин Б.П., Габдразанова С.Ф., Макара Н.С. Влияние композиции «ДАФС-25+Полизон» на функциональное состояние печени // Вестник ветеринарии. - 2012. - Т. 60. - №1. - С. 70-72. 39. Кирова Ю.И. Антиоксидантное и антитоксическое действие новых селеноорганических соединений: Дис. ... канд. биол. наук: Ростов н/Д, 2004. 186 c. 40. Кирпичев В.И. Влияние нитрофурановых препаратов на углеводный обмен золотистого стафилококка. Автореф. канд. дисс. Саратов, 1972. – 21 с. 41. Кирпичев В.И. Активность танскетолазы золотистого стафилококка, выращенного на среде с нитрофурановыми препаратами. В сб.: «Вопросы теоретической и прикладной микробиологии». – 1973. - Ростов-на-Дону. – С. 49-50. 42. Коваль Ю.И., Бокова Т.И. Влияние антиоксидантного статуса организма цыплят-бройлеорв на процессы детоксикации свинца и кадмия // Вестник НГАУ. - 2010. - Т. 14. - №2. - С. 35-40. 43. Комзалова А.В., Ошкина Л.Л., Трифонов Г.А. Влияние селенсодержащих препаратов на морфологические показатели крови быков-производителей // Нива Поволжья. - 2012. - №4. - С. 75-78. 44. Кон В. Электронная структура вещества – волновые функции и функционалы плотности // УФН. - 2002. - Т. 172. - №3. - С. 336-348. 45. Костюк В.А., Потапович А.И. Биорадикалы и биоантиоксиданты: Монография. – Мн.: БГУ, 2004. – 174 с. 46. Костюченко С.И. Тиоридазин: типичный антипсихотик с атипичным профилем побочных эффектов // Вестник ассоциации психиатров Украины. 2013. - № 2. - С. 58-63. 47. Кузнецов Ю.А. эффективность использования селеноорганического препарата ДАФС-25 в комбикормах-концентратах для высокопродуктивных коров. Дисс. …канд. сел.-хоз. наук. п. Дубровицы, 2002. - 123 с. 48. Кулешов К. А., Трифонов Г. А. Морфогенез желудка кур при применении препаратов селена // Ветеринария. - 2008. - № 11. - С. 54-57. 268 49. Кулинский В.И., Колесниченко Л.С. Структура, свойства, биологическая роль и регуляция глутатионпероксидазы // Успехи соврем. биол. —1993. — Т. 113, N 1. — C. 107—122. 50. Лекарь А.В., Ветрова Е.В., Борисенко Н.И., Яковишин Л.А., Гришковец В.И. Масс-спектрометрическое исследование молекулярного комплексообразования растительных гликозидов со стрептоцидом (сульфаниламидом) // Химия растительного сырья. - 2011. - № 2. - С. 103-106. 51. Макарова Е.С., Павленко Г.И. Влияние свинца и цинка на биохимические показатели белых крыс. Применение препарата ДАФС-25 при отравлении свинцом и цинком // Ветеринарная патология. - 2011. - № 4. - С. 117-120. 52. Махаева Н.А. Синтез функциональных органических халькогенидов, содержащих азот, фосфор, кремний, германий: Автореф. … канд. хим. наук. Иркутск, 2007. - 21 с. 53. Меньшикова Е.Б., Ланкин В.З., Зенков Н.К., Бондарь И.А., Круговых Н.Ф., Труфакин В.А. Окислительный стресс. Прооксиданты и антиоксиданты. Фирма «Слово», Москва, 2006. - 553с. 54. Меркулова Е.П. Биологическая активность селеноорганических соединений на основе гетероциклов: Автореф. … канд. биол. наук. Ростовна-Дону, 2012. - 24 с. 55. Никитин А.И. Гормоноподобные ксенобиотики и репродуктивная система // Проблемы репродукции. - 2002. - №2. - С. 5-24. 56. Островский В.К., Мащенко А.В., Янголенко Д.В., Макаров С.В. Показатели крови и лейкоцитарного индекса интоксикации в оценке тяжести и определении прогноза при воспалительных, гнойных и гнойно- деструктивных заболеваниях// Клин. лаб. диагностика. — 2006. — № 6. — С. 50–53. 57. Островский В.К., Машенко А.В., Макаров С.В. Оценка тяжести и прогноз гнойно-деструктивных заболеваний органов брюшной полости // Хирургия. 2007. - № 1. С. 33—37. 269 58. Паперная Л. К. Новые пути синтеза серо- и селенорганических соединений на основе халькогенильных радикалов, генерируемых в жидкой и газовой фазах. Дисс. … докт. хим. наук. Иркутск. 2007. - 329 с. 59. Перепелкина Л.И., Краснощекова Т.А., Ворсина Н.В. Физиологическое влияние добавок в рацион селена на рост и развитие телят в селенодефицитной провинции // Вестник алтайского государственного аграрного университета. - 2012. - Т. 88. - № 2. - С. 73-76. 60. Перунова Е.В. Физиолого-биохимические и продуктивные показатели свиней в зависимости от доз и способов введения в организм селена. Дисс. … канд. биол. наук. Пенза, 2000. - 141 с. 61. Покровский А.А. Биохимические методы исследования в клинике (справочник). М.: Медицина, 1969. - 544 с. 62. ПоперечневаТ.Ю. Приспособительные реакции организма теплокровных животных (крыс) на диацетофенонилселенид. Дисс. … канд. биол. наук. Саратов, 2009. - 119 с. 63. Попов Т.В. Нозокомиальные инфекции в отделении интенсивной терапии хирургического профиля. Дисс. …канд. мед. наук. Москва, 2006. - 146 с. 64. Пульняшенко глюкокортикоидов П.Р. в Принципы лечении мелких рационального домашних применения животных: Обзор литературы. - 2004. - 13 с. 65. Распоряжение Правительства Российской Федерации от 25 октября 2010 г. N 1873-р г. Москва // Российская газета. 3 ноября 2010. Федеральный выпуск № 5328. 66. Ратушная Е.В. Экологическая оценка новых классов селенорганических соединений: солей селенопирилия и октагидроселеноксантенов. Дисс. … канд. биол. наук. Саратов, 2001. - 126 с. 67. Решетник Л.А., Парфенова Е.О. Селен и здоровье человека (обзор литературы) // Экология моря. - 2000. - № 54. - С. 20-25. 270 68. Риш М.А. Наследственные биогеохимическая эволюция микроэлементозы таксонов биосферы // Техногенез (Тр. и Биогеохим. Лаборатории. Т. 24). М.: Наука, 2003. - С. 301-348. 69. РодионоваТ.Н. Фармакодинамика селенорганических препаратов и их применение в животноводстве. Дисс. …докт. биол. наук. Краснодар. 2004. – 296 с. 70. Рукавишников В.С., Шаяхметов С.Ф., Бодиенкова Г.М., Курчевенко С.И. Взаимосвязь между содержанием ртути в биосредах и изменением иммунологических показателей у работающих // Фундаментальные исследования. - 2011. - №1. - С. 165-170. 71. Руководство по контролю качества питьевой воды. Том 2. Гигиенические критерии и другая релевантная информация. Часть III. Неорганические компоненты, оказывающие влияние на здоровье. Всемирная организация здравоохранения. Женева, 1987. - 38 с. 72. Садеков И.Д., Минкин В.И. Специфика реакционной способности теллурорганических соединений // Успехи химии. - 1995. - Т. 64. - № 6. - С. 527-561. 73. Сайфульмулюков Э.Р. Токсикологическая оценка и фармакологическое обоснование применения препарата е-селен при интенсивном выращивании и откорме бычков. Дисс. … канд. вет. наук. Троицк, 2006. - 138 с. 74. Саноцкий И.В. Разработка новой селен-органической биологически активной пищевой добавки (БАД) как паллиативной меры защиты при химических воздействиях // Прикладная токсикология. - 2010. - Т. 1. - №2. С. 10-13. 75. Северин Е.С. Биохимия: Учебник / Под ред. Е.С. Северина. – 2-е изд., испр. – М.: ГЭЕТАР-МЕД, 2004. – 784 с. 76. Сенькевич О.А., Голубкина Н.А., Ковальский Ю.Г. Диагностика обеспеченности человека селеном и оценка степени его дефицита // Дальневосточный медицинский журнал. - 2011. - №4. - С. 78-80. 271 77. Сидоренко С.В. Место бактерий в живой природе // Инфекции и антимикробная терапия. - 2000. - Т.2. - №2. - С. 32-34. 78. Сотников Д.А. Продуктивность, морфофункциональное состояние внутренних органов и биохимические показатели крови мясных кур в зависимости от уровня и методов воздействия на них селенсодержащими соединениями. Дисс. … канд. сел.-хоз. наук. Пенза, 2002. - 105 с. 79. Стальная И.Д. Метод определения диеновой коньюгации ненасыщенных высших жирных кислот // Современные методы в биохимии / Под ред. В.Н. Ореховича. М.: Медицина, 1977. - С. 63-64. 80. Стальная И.Д., Гаришвили Т.Г. Метод определения малонового диальдегида с помощью тиобарбитуровой кислоты // Современные методы в биохимии / Под ред. В.Н. Ореховича. М.: Медицина, 1977. - С. 66-68. 81. Тарасов Н.И., Тепляков А.Т., Малахович Е.В., Федосова Н.Н., Калюжин В.В., Пушникова Е.Ю. антиоксидантной защиты Состояние перекисного крови у больных окисления инфарктом липидов, миокарда, отягощенным недостаточностью кровообращения // Тер. архив. - 2002. - №12. - C.12-15. 82. Теселкин Ю.О. Антиоксидантная активность плазмы крови как критерий оценки функционального состояния антиоксидантной системы организма и эффективности применения экзогенных антиоксидантов. Дис. … докт. биол. наук. Москва, 2003. - 272 с. 83. Трошина Т.А. Фармакокоррекция селенодефицита у животных препаратом дафс-25 и его влияние на продуктивные качества. Дисс. … докт. вет. наук. Ижевск, 2010. - 292 с. 84. Тутельян В.А., Княжев В.А., Хотимченко С.А., Голубкина Н.А., Кушлинский Н.Е., Соколов Я.А. Седен в организме человека: метаболизм, антиоксидантные свойства, роль в канцерогенезе. М.: Издательство РАМН, 2002. - 224 с. 85. Тюкавкина Н.А. Биоорганическая химия: Учебник Н.А. Тюкавкина, Ю.И. Бауков. – М.: Дрофа, 2004. – 544 с. для вузов / 272 86. Уайт А., Хендлер Ф., Смит Э., Хилл Р., Леман И. Основы биохимии: В 3-х томах. Т. 1. Пер. с англ. Под ред. Ю. А. Овчинникова. - Мир. М., 1981. – 534 с. 87. Уварова Е. В. Течение гликогеновой болезни у детей в условиях комплексной терапии: дис…. канд. мед.наук. - М., 2005. - С. 57-58. 88. Утц С.Р., Завьялов А.И., Марченко В.М. Клинико-эпидемиологические особенности чесотки и новые подходы к ее диагностике и лечению на современном этапе (Обзор) // Саратовский научно-медицинский журнал. 2011. - Т. 7. - № 3. - С. 636-641. 89. Филимонов Д.А., Поройков В.В. Прогноз спектра биологической активности органических соединений // Рос. хим. ж. (Ж. Рос. хим. об-ва им. Д.И. Менделеева). - 2006. - Т. L. - №2. - С. 66-75. 90. Филов В.А. Вредные химические вещества. Неорганические соединения I-IV групп. / Под ред. проф. В.А. Филова. Л.: Химия, 1988. - 512 с. 91. Фомина Н.Ю. (Русецкая Н.Ю.) Взаимодействие K2PdCl4 с синтетическими и природными нуклеиновыми кислотами / Фомина Н.Ю., Чаплыгина О.А., Шебалдова А.Д., Бородулин В.Б. // Журнал общей химии. – 2002. - Т. 72. - Вып. 5. – С. 755-760. 92. Фридович И. Радикалы кислорода, пероксид водорода и токсичность кислорода / Свободные радикалы в биологии. - М.: Мир, 1979. Т. 1. - С.272314. 93. Фроловичев А.С., Трошин А.Н. Применение диацетофенонил селенида в свиноводстве // Ветеринария Кубани. - 2013. - №3. - С. 9-11. 94. Харкевич Д.А. Фармакология: Учебник. – 9-е изд., перераб., доп. и испр. – М.: Гэотар-Медиа, 2006. – 736 c. 95. Ширшова Т.И., Голубкина Н.А., Бешлей И.В., Матистов Н.В. Селенодефицит и возможности его сокращения. Аккумулирующие свойства некоторых представителей рода Allium L. по отношению к селену // Известия Коми научного центра УрО РАН. - 2011. - Вып. 3. - №7. - С. 48-54. 273 96. Шперов А.С. Мясная продуктивность и качество мяса свиней при использовании в рационах селенорганических препаратов. Дисс. … канд. сел.-хоз. наук. Волгоград, 2009. - 164 с. 97. Щербаков В.А. Эликсир бессмертия – главный секрет Океана // Книга тайн. – М., 1991. 98. Abdel-Monem N., Abdel-Azeem A., El-Ashry E.H., Ghareeb D.A., Nabil- adam A. Pretreatment Hepatoprotective Effect of the Marine Fungus Derived from Sponge on Hepatic Toxicity Induced by Heavy Metals in Rats // BioMed Research International. V. 2013. Article ID 510879. - 15 p. http://dx.doi.org/10.1155/2013/510879. 99. Abouhamed M., Wolff N.A., Lee W.K., Smith C.P., Thevenod F. Knockdown of endosomal/lysosomal divalent metal transporter 1 by RNA interference prevents cadmium-metallothionein-1 cytotoxicity in renal proximal tubule cells // Am. J. Physiol. Renal. Physiol. - 2007. - V. 293. - P. F705–F712. 100. Aebi H. Catalase in Vitro // Methods in Enzymology. - 1984. - V. 105. P. 121-126. 101. Agency for Toxic Substances and Disease Registry. Toxicological Proile: Cadmium. 2008. http://www.atsdr.cdc.gov/toxproiles/tp.asp?id=48&tid=15.2-5. 102. Agency for Toxic Substances and Disease Registry, “Toxicological Proile: Lead,” 2007. http://www.atsdr.cdc.gov/ToxProiles/tp.asp?id=96&tid=22. 103. Agency for Toxic Substances and Disease Registry. Toxicological Proile: Mercury, US Department of Health and Human Services. Public Health Service, Atlanta, Ga, USA, 1999, http://www.atsdr.cdc.gov/ToxProiles/ TP.asp?id=115&tid=24. 104. Aikawa R., Komuro I., Yamazaki T., Zou Y.Z., Kudoh S., Tanaka M., Shiojima I., Hiroi Y., Yazaki Y. Oxidative stress activates extracellular signalregulated kinases through Src and ras in cultured cardiac myocytes of neonatal rats // J. Clin. Invest. - 1997. - V. 100. - P. 1813-1821. 274 105. Albeck A., Weitman H., Sredni B., Albeck M. Tellurium compounds: selective inhibition of cysteine proteases and model reaction with thiols // Inorg. Chem. 1998. - V. 37. - P. 1704–1712. 106. Aldercreutz H., Markkanen H., Watanabe S. Plasma concentration of phytoestrogens in Japanese men // Lancet. - 1993. - Vol. 342. - P. 1209–1210. 107. Allen J.W., El-Oqayli H., Aschner M., Syversen T., Sonnewald U. Methylmercury has a selective effect on mitochondria in cultured astrocytes in the presence of [U-13C]glutamate // Brain Res. - 2001. - V. 908. - P. 149–154. 108. Altenhofer S., Kleikers P.W., Radermacher K.A., Scheurer P., Hermans J.J., Schiffers P., Ho H., Wingler K., Schmidt H.H. The NOX toolbox: Validating the role of NADPH oxidases in physiology and disease // Cell. Mol. Life Sci. - 2012. V. 69. - P. 2327–2343. 109. Amberg R., Mizutani T., Wu X.Q., Gross-H.J. Selenocysteine synthesis in mammalia: an identity switch from tRNA(Ser) to tRNA(Sec) // J.Mol. Biol. - 1996. - V. 263. - N 1. - P. 8-19. 110. Andrews G. K. Regulation of metallothionein gene expression by oxidative stress and metal ions // Biochemical Pharmacology. - 2000. - V. 59(1). - V. 95– 104. 111. Andrews R.C., Walker B.R. Glucocorticoids and insulin resistance: old hormones, new targets // Clinical Science. - 1999. - V. 96. - P. 513–523. 112. Andreyev A.Y., Kushnareva Y.E., Starkov A.A. Mitochondrial metabolism of reactive oxygen species // Biochemistry Mosc. – 2005. - V. 70. - P. 200-214. 113. Antunes F., Nunes C., Laranjinha J., Cadenas E. Redox interactions of nitric oxide with dopamine and its derivatives // Toxicology. - 2005. - V. 208. - P. 207212. 114. Arai K., Maguchi S., Fujii S., Ishibashi H., Oikawa K., Taniguchi N. Glycation and inactivation of human Cu-Zn-superoxide dismutase // J. Biol. Chem. - 1987. V. 262. - P. 16969-16972. 115. Arner E.S.J., Holmgren A. Physiological functions of thioredoxin and thioredoxin reductase // Eur. J. Biochem. - 2000. - V. 267. - P. 6102–6109. 275 116. Arteel G. E., Briviba K. & Sies H. Function of thioredoxin reductase as a peroxynitrite reductase using selenocystine or ebselen // Chem. Res. Toxicol. 1999. - V. 12. - P. 264–269. 117. Arterbery A.S., Fergus D.J., Fogarty E.A., Mayberry J. Evolution of ligand specificity in vertebrate corticosteroid receptors // BMC Evolutionary biology. 2011. - Vol. 11. - P. 14-28. 118. Arthur J.R. The glutathione peroxidases // Cell. Mol. Life Sci. - 2000. - V. 57. P. 1825–1835. 119. Aust S.D., Morehouse L.A., Thomas C.E. Role of metals in oxygen radical reactions // J. Free Rad. Biol. Med. - 1985. - Vol. 1. - P.3-25. 120. Avazeri C., Turner R. J., Pommier J., Weiner J. H., Giordano G., Vermeglio A. Tellurite reductase activity of nitrate reductase is responsible for the basal resistance of Escherichia coli to tellurite // Microbiology. - 1997. - V. 143. - P. 1181-1189. 121. Ayres D.C., Loike J.D. Lignans. Chemical, biological and clinical properties. In: Chemistry &Pharmocology of Natural Products. Ed.: Phillipson J.D., Ayres D.C., Baxter H. Cambrige University Press. - 1990. - P. 402. 122. Ba L.A., Döring M., Jamier V., Jacob C. Tellurium: an element with great biological potency and potential // Org. Biomol. Chem. - 2010. - V. 8(19). - P. 4203-4216. doi: 10.1039/c0Ob00086h. 123. Baird-Parker A. An improved diagnostic and selective medium for isolating coagulase positive Staphylococci // J. Appl. Bacteriol. - 1962. - V. 25. - P. 12–19. 124. Bakir F., Damluji S.F., Amin-Zaki L., Murtadha M., Khalidi A., al-Rawi N.Y., Tikriti S., Dahahir H.I., Clarkson T.W., Smith J.C., Doherty R.A. Methylmercury poisoning in Iraq // Science. - 1973. - V. 181. - P. 230–241. 125. Banin E., Lozinski A., Brady K.M., Berenshtein E., Butterfield P.W. The potential of desferrioxamine-gallium as an anti-Pseudomonas therapeutic agent // Proc Natl Acad Sci USA. - 2008. - V. 105. - P. 16761–16766. 276 126. Barja G. Mitochondrial oxygen radical generation and leak: Sites of production in states 4 and 3, organ specificity, and relation to aging and longevity // J. Bioenerg. Biomembr. - 1999. - V. 31. - P. 347-366. 127. Barrett J. R. Phytoestrogens: friends or foes? // Environ. Health. Persp. - 1996. - V. 104. - P. 478–482. 128. Bates J.M., Spate V.L., Morris J.S., Germain D.L.St., Galton V.A. Effects of Selenium Deficiency on Tissue Selenium Content, Deiodinase Activity, and Thyroid Hormone Economy in the Rat during Development // Endocrinology. – 2000. - V. 141. - N 7. – P. 2490-2500. 129. Baxter J.D. Glucocorticoid hormone action // Pharmacol Ther. - 1976. - V. 2. P. 605–669. 130. Beal M.F. Oxidatively modified proteins in aging and disease // Free Radic. Biol. Med. - 2002. - V. 32. - P. 797-803. 131. Becke A.D. Density-Functional Exchange-Energy Approximation with Correct Asymptotic Behavior // Physical review A. - 1988. - V. 38. – N 6. - Р. 3098-3100. 132. Becke A.D. Density-Functional Thermochemistry. III. The role of exact exchange // J. Chem. Phys. - 1993. - V. 98. – N 7. - P. 5648-5652. 133. Beckett G.J., Arthur J.R. Selenium and endocrine systems // Journal of Endocrinology. - 2005. - V. 184. - Р. 455–465. 134. Behne D., Weiler H., Kyriakopolos A. Effects of selenium deficiency on testicular morphology and function in rats // J. Reprod. Fertil. - 1996. - V. 106. - N 2. - P. 291-297. 135. Belozerskaya T.A., Gessler N.N. Reactive oxygen species and the strategy of antioxidant defense in fungi: A review // Appl. Biochem. Microbiol. - 2007. - V. 43. - P. 506-515. 136. Bernhot R. A. Mercury toxicity and treatment: a review of the literature // Journal of Environmental and Public Health. - 2012. - V. 2012. Article ID 460508. 10 p. 277 137. Berry M.J., Kieffer J.D., Harney J.W. Selenocysteine confers the biochemical properties characteristic of the type I iodothyronine deiodinase // J. Biol. Chem. 1991. - V. 266. - P. 14155-14158. 138. Berry M.J., Banu L., Larsen P.R. Type I iodothyronine deiodinase is a selenocysteine-containing enzyme // Nature. – 1991. - N 31. – P.438-440. 139. Beyersmann D., Hartwig A. Carcinogenic metal compounds: Recent insight into molecular and cellular mechanisms // Arch. Toxicol. - 2008. - V. 82. - P. 493– 512. 140. Beyrouty P., Chan H.M. Co-consumption of selenium and vitamin E altered the reproductive and developmental toxicity of methylmercury in rats // Neurotoxicol. Teratol. - 2006. - V. 28. - N 1. - P. 49-58. 141. Bień M., Błaszczyk B., Kalinowska K., Młochowski J., Inglot A.D. Antifungal activity of 2-(4-chlorophenyl)-1,2-benzisoselenazol-3(2H)-one, the analog of Ebselen // Arch. Immunol. Ther. Exp. (Warsz). - 1999. - V. 47. - N 3. - P. 185-193. 142. Biemond P., Swaak A.J.G., van Eijk H.G., Koster J.F. Intraarticular ferritinbound iron in rheumatoid arthritis // Arthritis Rheum. - 1986. - V. 29. - P. 11871193. 143. Biggadike K., Bledsoe R.K., Coe D.M., Cooper T.W., House D., Iannone M.A., Macdonald S.J., Madauss K.P., McLay I.M., Shipley T.J., Taylor S.J., Tran T.B., Uings I.J., Weller V., Williams S.P. Design and x-ray crystal structures of high-potency nonsteroidal glucocorticoid agonists exploiting a novel binding site on the receptor // PNAS. - 2009. - V. 106. - N. 43. - P. 18114–18119. 144. Bjelland S., Seeberg E. Mutagenicity, toxicity and repair of DNA base damage induced by oxidation // Mutat. Res. - 2003. - V. 531. - P. 37-80. 145. Bjornstedt M., Xue J., Huang W., Akesson B., Holmgren A. The thioredoxin and glutaredoxin systems are efficient electron donors to human plasma glutathione peroxidase // J. Biol. Chem. - 1994. - V. 269. - P. 29382–29384. 146. Bolann B.J., Ulvik R.J. Release of iron from ferritin by xanthine oxidase. Role of the superoxide radical // Biochem. J. - 1987. - V. 243. - P. 55-59. 278 147. Borsetti F., Francia F., Turner R.J., Zannoni D. The Thiol:Disulfide Oxidoreductase DsbB Mediates the Oxidizing Effects of the Toxic Metalloid Tellurite (TeO32-) on the Plasma Membrane Redox System of the Facultative Phototroph Rhodobacter capsulatus // Journal of bacteriology. - 2007. – V. 189. N 3. - Р. 851–859. 148. Bose K.S., Vyas P., Singh M. Plasma non-enzymatic antioxidants-vitamin C, E, beta-carotenes, reduced glutathione levels and total antioxidant activity in oral sub mucous fibrosis // Eur. Rev. Med. Pharmacol. Sci. - 2012. - V. 16(4). - P. 530532. 149. Brigelius-Flohe R. Glutathione peroxidases and redox-regulated transcription factors // Biol. Chem. - 2006. - V. 387. - P. 1329–1335. 150. Brown D.I., Griendling K.K. Nox proteins in signal transduction // Free Radic. Biol. Med. - 2009. - V. 47. - P. 1239–1253. 151. Brown K.M., Arthur J.R. Selenium, selenoproteins and human health: a review // Public. Health. Nutr. – 2001. - N 4. – P. 593-599. 152. Buettner G.R. Superoxide Dismutase in Redox Biology: The roles of superoxide and hydrogen peroxide // Anticancer Agents Med. Chem. - 2011. V. 11(4). - P. 341–346. 153. Buettner G.R., Ng C.F., Wang W., Rodgers V.G.J., Schafer F.Q. A new paradigm: Manganese superoxide dismutase influences the production of H 2O2 in cells and thereby their biological state // Free Radic. Biol. Med. - 2006. - V. 41. P. 1338–1350. 154. Burton G.M., Traber M.G. Vitamin E - antioxidant activity, biokinetics, and bioavailability // Rev. Nutr. - 1990. - V. 10. - P. 357-382. 155. Butterfield D.A., Howard B., Yatin S., Koppal T., Drake J., Hensley K., Aksenov M., Aksenova M., Subramaniam R., Varadarajan S., Harris-White M.E., Pedigo N.W. Jr., Carney J.M. Elevated oxidative stress in models of normal brain aging and Alzheimer's disease // Life Sci. - 1999. - V. 65. - P. 1883-1892. 279 156. Cabañero A.I., Madrid Y., Cámara C. Effect of animal feed enriched with Se and clays on Hg bioaccumulation in chickens: in vivo experimental study // J. Agric. Food Chem. - 2005. - V. 53. - N 6. - P. 2125-2132. 157. Caboni L., Kinsella G.K., Blanco F., Fayne D. “True” Antiandrogens-Selective Non-Ligand-Binding Pocket Disruptors of Androgen Receptor−Coactivator Interactions: Novel Tools for Prostate Cancer // J. Med. Chem. - 2012. - V. 55. P. 1635−1644. 158. Cadenas E. Biochemistry of oxygen toxicity // Annu. Rev. Biochem. - 1989. V. 58. - P. 79-110. 159. Cadenas E., Davies K.J. Mitochondrial free radical generation, oxidative stress and aging // Free Radic. Biol. Med. - 2000. - V. 29. - P. 222-230. 160. Calderon I.L., Elıas A.O., Fuentes E.L., Pradenas G.A., Castro M.E., Arenas F.A., Perez J.M., Vasquez C.C. Tellurite-mediated disabling of [4Fe–4S] clusters of Escherichia coli dehydratases // Microbiology. - 2009. - V. 155. - Р. 1840–1846. DOI 10.1099/mic.0.026260-0. 161. Calvin H.I. Selective incorporation of selenium-75 into a polypeptide of the rat sperm tail (1) // J. Exp. Zool. - 1978. - V. 204. - N 3. - P. 445-452. 162. Campbell J.W., Waters M.N., Tarter A., Jackson J. Heavy metal and selenium concentrations in liver tissue from wild american alligator (Alligator Mississippiensis) livers near Charleston, South Carolina // Journal of Wildlife Diseases. – 2010. - 46(4). P. 1234–1241. 163. Cannino G., Ferruggia E., Luparello C., Rinaldi A.M. Cadmium and mitochondria // Mitochondrion. - 2009. - V. 9. - P. 377–384. 164. Casagrande S., Bonetto V., Fratelli M., Gianazza E., Eberini I., Massignan T., Salmona M., Chang G., Holmgren A., Ghezzi P. Glutathionylation of human thioredoxin: a possible crosstalk between the glutathione and thioredoxin systems // Proc. Natl. Acad. Sci. USA. - 2002. - V. 99. - P. 9745–9749. 165. Castoldi A.F., Onishchenko N., Johansson C., Coccini T., Roda E., Vahter M., Ceccatelli S., Manzo L. Neurodevelopmental toxicity of methylmercury: 280 Laboratory animal data and their contribution to human risk assessment // Regul. Toxicol. Pharmacol. - 2008. - V. 51. - P. 215–229. 166. Castro M., Molina R., Dıaz W.A., Pradenas G.A., Vasquez C.C. Expression of Aeromonas caviae ST pyruvate dehydrogenase complex components mediate tellurite resistance in Escherichia coli // Biochem. Biophys. Res. Commun. - 2009. - V. 380. - P. 148–152. 167. Ceballospicot I., Nicole A., Clement M., Bourre J.M., Sinet P.M. Age-related changes in antioxidant enzymes and lipid peroxidation in brains of control and transgenic mice overexpressing copper-zinc superoxide dismutase // Mutation Research. - 1976. - V. 275. - P.281-293. 168. Chan G., Hardej D., Santoro M., Lau-Cam C., Billack B. Evaluation of the antimicrobial activity of ebselen: role of the yeast plasma membrane H+-ATPase // J.Biochem. Mol.Toxicol. - 2007. - V. 21. - N 5. - P. 252-264. 169. Chapman P., Siddons C., Zadik P., Jewes L. An improved selective medium for the isolation of Escherichia coli O157 // J. Med. Microbiol. - 1991. - V. 35. P. 107–110. 170. Chasteen T.G., Fuentes D.E., Tantaleán J.C., Vásquez C.C. Tellurite: history, oxidative stress, and molecular mechanisms of resistance // FEMS Microbiol. Rev. - 2009. - V. 33(4). - P. 820-832. doi: 10.1111/j.1574-6976.2009.00177.x. 171. Chaudiere J., Tappel A.L. Purification and characterization of seleniumglutathione peroxidase from hamster liver // Arch. Biochem. Biophys. - 1983. V. 226. - P. 448-457. 172. Chen Y., Cai J., Murphy T.J., Jones D.P. Overexpressed human mitochondrial thioredoxin confers resistance to oxidant-induced apoptosis in human osteosarcoma cells // J. Biol. Chem. - 2002. - V. 277. - P. 33242–3324. 173. Chen C, Yu H, Zhao J, Li B, Qu L, Liu S, Zhang P, Chai Z. The Roles of Serum Selenium and Selenoproteins on Mercury Toxicity in Environmental and Occupational Exposure // Environmental Health Perspectives. - 2006. - V. 114. N. 2. - P. 297-301. 281 174. Chen X., Zhu Y.-H., Cheng X.-Y., Zhang Z.-W., Xu S.-W. The Protection of Selenium against Cadmium-Induced Cytotoxicity via the Heat Shock Protein Pathway in Chicken Splenic Lymphocytes // Molecules. - 2012. - V. 17. - P. 14565-14572; doi:10.3390/molecules171214565. 175. Choudhury H.G., Cameron A.D., Iwata S., Beis K. Structure and mechanism of the chalcogen-detoxifying protein TehB from Escherichia coli // Biochem. J. 2011. - V. 435. - P. 85–91. doi:10.1042/BJ20102014. 176. Chu F.F., Esworthy R.S., Chu P.G., Longmate J.A., Huycke M.M., Wilczynski S., and Doroshow J.H. Bacteria-induced intestinal cancer in mice with disrupted Gpx1 and Gpx2 genes // Cancer Res. - 2004. - V. 64. - P. 962–968. 177. Ciesielski T., Weuve J., Bellinger D. C. Cadmium exposure and neurodevelopmental outcomes in U.S. children // Environmental Health Perspectives. - 2012. - V. 120. - P. 758–763. 178. Clarkson T.W. The toxicology of mercury // Crit. Rev. Clin. Lab. Sci. - 1997. V. 34. - P. 369–403. 179. Conner E.M., Grisham M.B. Inflammation, free radicals and antioxidants // Nutrition. - 1996. - V. 12. - P. 274-277. 180. Connor K.M., Hempel N., Nelson K.K., Dabiri G., Gamarra A., Belarmino J., Van De Water L., Mian B.M., Melendez J.A. Manganese superoxide dismutase enhances the invasive and migratory activity of tumor cells // Cancer Res. - 2007. V. 67. - P. 10260–10267. 181. Contempré B., Duale N.L., Dumont J.E., Ngo B., Diplock A.T., Vanderpas J. Effect of selenium supplementation on thyroid hormone metabolism in an iodine and selenium deficient population. // Clin. Endocrinol. (Oxf). – 1992. – V. 36. N 6. – Р. 579-583. 182. Cooke M.S., Evans M.D., Dizdaroglu M., Lunec J. Oxidative DNA damage: mechanisms, mutation, and disease // FASEB J. 2003. V. 17. P. 1195-1214. 183. Coriat R., Marut W., Leconte M., Ba L.B., Vienne A., Chereau C., Alexandre J., Weill B., Doering M., Jacob C., Nicco C., Batteux F. The 282 organotelluride catalyst LAB027 prevents colon cancer growth in the mice // Cell Death and Disease. - 2011. - V. 2. - Р. 1-10, e191; doi:10.1038/cddis.2011.73. 184. Cournoyer B., Watanabe S., Vivian A. A tellurite-resistance genetic determinant from phytopathogenic Pseudomonads encodes a thiopurine methyltransferase: evidence of a widely-conserved family of methyltransferases // Biochim. Biophys. Acta. - 1998. - V. 1397. - P. 161–168. 185. Coward L., Barnes N., Setchell K., Barnes S. Genistein, diadzein and their beta-glycoside conjugates-antitumor isoflavones in soybean foods from American and Asian diets // J. Agric. Food Chem. - 1993. - V. 41. - P. 1961–1967. 186. Crapo J.D., Oury T., Rabouille C., Slot J.W., Chang L.Y. Copper,zinc superoxide dismutase is primarily a cytosolic protein in human cells // Proc. Natl. Acad. Sci. USA. - 1992. - V. 89. - P. 10405–10409. 187. Cui K., Luo X., Xu K., Ven M.M.R. Role of oxidative stress in neurodegeneration: recent developments in assay methods for oxidative stress and nutraceutical antioxidants // Prog. Neuropsychopharmacol. Biol. Psychiatry. 2004. - V. 28. - P. 771-799. 188. Cunha R.L.O.R., Gouvea I.E., Juliano L. A glimpse on biological activities of tellurium compounds // An Acad. Bras. Cienc. - 2009. - V. 81(3). - P. 393-407. 189. Cuypers A., Plusquin M., Remans T., Jozefczak M., Keunen E., Gielen H., Opdenakker K., Nair A.R., Munters E., Artois T.J. Cadmium stress: An oxidative challenge // Biometals. - 2010. - V. 23. - P. 927–940. 190. Dabak D., Gazuwa S. Y., Uborn G. A. Hepatoprotective: potential of calcium and magnesium against cadmium and lead induce hepatotoxicty in Wistar rats // Asian Journal of Biotechnology. - 2009. - V. 1(1). - P. 12–19. 191. Dalle-Donne I., Aldini G., Carini M., Colombo R., Rossi R., Milzani A. Protein carbonylation, cellular dysfunction, and disease progression // J. Cell Mol. Med. - 2006. - V. 10. - P. 389-406. 192. Dalton T.P., He L., Wang B., Miller M.L., Jin L., Stringer K.F., Chang X., Baxter C.S., Nebert D.W. Identification of mouse slc39a8 as the transporter 283 responsible for cadmium-induced toxicity in the testis // Proc. Natl. Acad. Sci. USA. - 2005. - V. 102. - P. 3401–3406. 193. Das K.C., Das C.K. Thioredoxin, a singlet oxygen quencher and hydroxyl radical scavenger: redox independent functions // Biochem. Biophys. Res. Commun. - 2000. - V. 277. - P. 443–447. 194. de Freitas A.S., Prestes A.S., Wagner C., Sudati J.H., Alves D., Porciúncula L.O., Kade I.J., Rocha J.B.T. Reduction of diphenyl diselenide and analogs by mammalian Thioredoxin Reductase is independent of their gluthathione peroxidase-like activity: A possible novel pathway for their antioxidant activity // Molecules. - 2010. - V. 15. - P. 7699–7714. 195. Dehghan G., Amjad L., Nosrati H. Enzymatic and non-enzymatic antioxidant responses of alfalfa leaves and roots under different salinity levels // Acta Biol. Hung. - 2013. - V. 64(2). - P. 207-217. doi: 10.1556/ABiol.64.2013.2.7. 196. Delrio L.A., Sandalio L.M., Palma J.M., Bueno P., Corpas F.J. Metabolism of oxygen radicals in peroxisomes and cellular implications // Free Radical Biology and Medicine. - 1992. - V. 13. - P.557-580. 197. Deng D.F., Teh F.C., Teh S.J. Effect of dietary methylmercury and selenomethionine on Sacramento splittail larvae // Sci. Total Environ. - 2008. - V. 407. – N 1. - P. 197-203. 198. DeYulia G.J., Carcamo J.M., Borquez-Ojeda O., Shelton C.C. Hydrogen peroxide generated extracellularly by receptor–ligand interaction facilitates cell signaling // Proc. Natl. Acad. Sci. USA. - 2005. - V. 102. - P. 5044–5049. 199. Deidda D., Lampis G., Maullu C., Pompei R., Isaia F., Lippolis V., Verani G. Antifungal, antibacterial, antiviral and cytotoxic activity of novel thio- and selenoazoles // Pharmacol. Res. - 1997. - V. 36. - N 3. - P. 193-197. 200. Ditchfield R., Hehre W.J., Pople J.A. A self-consistent molecular-orbital methods. IX. An extended Gaussian-type basis for molecular-orbital studies of organic molecules // J. Chem. Phys. - 1971. - V. 54. – N 2. - P. 724-728. 284 201. Diplock A.T. Recent studies on the interaction between vitamin E and selenium // Trace element metabolism in animals. Ed. by C.F. Mills. Edinburgh and L.: E. and S. Livingstone, 1970. - P. 190-203. 202. Doering M., Ba L.A., Lilienthal N. Synthesis and selective anticancer activity of organochalcogen based redox catalysts // J. Med. Chem. - 2010. - V. 14. - Р. 6954–6969. 203. Droge W. Free radicals in the physiological control of cell function // Physiol. Rev. - 2002. - V. 82. - P. 47–95. 204. Emmert S.W., Desai D., Aminb S., Richie J.P. Enhanced Nrf2-Dependent Induction of Glutathione in Mouse Embryonic Fibroblasts by Isoselenocyanate Analog of Sulforaphane // Bioorg. Med. Chem. Lett. - 2010. - V. 20(8). - P. 2675– 2679. doi:10.1016/j.bmcl.2010.01.044. 205. Enoksson M., Fernandes A.P., Prast S., Lillig C.H., Holmgren A., Orrenius S. Overexpression of glutaredoxin 2 attenuates apoptosis by preventing cytochrome c release // Biochem. Biophys. Res. Commun. 2005. V. 327. P. 774–779. 206. Erickson J.M., Messer T.M. Glucosamine and chondroitin sulfate treatment of hand osteoarthritis // J. Hand Surg. Am. - 2013. - V. 38(8). - P. 1638-40. doi: 10.1016/j.jhsa. 207. Esworthy R.S., Aranda R., Martin M.G., Doroshow J.H., Binder S.W., Chu F.F. Mice with combined disruption of Gpx1 and Gpx2 genes have colitis // Am. J. Physiol. Gastrointest. Liver Physiol. - 2001. - V. 281. - P. G848–G855. 208. Esworthy R.S., Yang L., Frankel P.H., Chu F.F. Epithelium-specific glutathione peroxidase, Gpx2, is involved in the prevention of intestinal inflammation in selenium-deficient mice // J. Nutr. - 2005. - V. 135. - P. 740–745. 209. Fagan J., Sleczka B.G., Sohar I. Quantitation of oxidative damage to tissue proteins // Int. J. Biochem. Cell Biol. - 1999. - V. 31. - P. 751-757. 210. Fain J.N., CzechM.P. Glucocorticoid effects on lipid mobilization and adipose tissue metabolism. In: Blashko H., editor. Adrenal gland. Vol 6, sect 7, Endocrinology. Handbook of physiology. Physiological Society; 1975. - P. 169–172. Washington, DC: American 285 211. Fasinu P.S., Orisakwe O.E. Heavy Metal Pollution in Sub-Saharan Africa and Possible Implications in Cancer Epidemiology // Asian Pacifc J. Cancer Prev. 2013. - V. 14 (6). - P. 3393-3402. DOI:http://dx.doi.org/10.7314/APJCP.2013.14.6.3393. 212. Fernandes A.P., Holmgren A. Glutaredoxins: glutathione-dependent redox enzymes with functions far beyond a simple thioredoxin backup system // Antioxid. Redox Signal. - 2004. - V. 6. - P. 63–74. 213. Finkel T. Signal transduction by reactive oxygen species // J. Cell. Biol. - 2011. - V. 194. - P. 7–15. 214. Finley J.W. Bioavailability of selenium from foods // Nutr. Rev. - 2006. V. 64. - P. 146-151. 215. Fischer W. A second note on the term “chalcogen” // J. Chem. Educ. - 2001. V. 78. - P. 1333. 216. Fleming A. On the specific antibacterial properties of penicillin and potassium tellurite. Incorporating a method of demonstrating some bacterial antagonisms // J. Pathol. Bacteriol. - 1932. - V. 35. - P. 831–842. 217. Fleury C., Mignotte B., Vayssiere J.L. Mitochondrial reactive oxygen species in cell death signaling // Biochimie. - 2002. - V. 84. - P. 131-141. 218. Flohe L. Glutathione peroxidase brought into focus // Free radicals in biology. 1982. - V. 5. - P.223-254. 219. Flora G., Gupta D., Tiwari A. Toxicity of lead: a review with recent updates // Interdisciplinary Toxicology. - 2012. - V. 5. - P. 47–58. 220. Folz R.J., Crapo J.D. Extracellular superoxide dismutase (SOD3): tissuespecific expression, genomic characterization, and computer-assisted sequence analysis of the human EC SOD gene // Genomics. - 1994. - V. 2. - P. 162–171. 221. Ford C.W., Zurenko G.E., Barbachyn M.R. The discovery of linezolid, the first oxazolidinone antibacterial agent. Curr Drug Targets // Infect. Disord. - 2001. - V. 1. - P. 181–199. 286 222. Fowler B. A. Monitoring of human populations for early markers of cadmium toxicity: a review // Toxicology and Applied Pharmacology. - 2009. - V. 238(3). P. 294–300. 223. Frei B., Stocker R., Ames B.N. Antioxidant defenses and lipid peroxidation in human blood plasma // Proc. Natl. Acad. Sci. USA. — 1988. — Vol.85. — Р. 9748-9752. 224. Fridovich I., Freeman B. Antioxidant defenses in the lung // Annu. Rev. Physiol. - 1986. - V. 48. - P. 693–702. 225. Fu D., Chen J., Zhang Y., Yu Z. Cloning and expression of a heat shock protein (HSP) 90 gene in the haemocytes of Crassostrea hongkongensis under osmotic stress and bacterial challenge // Fish Shellfish Immunol. - 2011. - V. 31. P. 118–125. 226. Fuentes D.E., Fuentes E.L., Castro M.E., Pérez J.M., Araya M.A., Chasteen T.G., Pichuantes S.E., Vásquez C.C. Cysteine metabolism-related genes and bacterial resistance to potassium tellurite // J Bacteriol. - 2007. - V. 189(24). - P. 8953-8960. 227. Fukai T., Ushio-Fukai M. Superoxide dismutases: role in redox signaling, vascular function, and diseases. // Antioxid. Redox Signal. - 2011. - V. 15(6). P. 1583-606. doi: 10.1089/ars.2011.3999. 228. Fullmer C.S. Intestinal calciumand lead absorption: effects of dietary lead and calcium // Environmental Research. - 1991. - V. 54(2). - P. 159–169. 229. Fullmer C.S. Lead-calcium interactions: involvement of 1,25- dihydroxyvitamin D // Environmental Research. - 1997. - V. 72(1). - P. 45–55. 230. Girotti A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems // J. Lipid Res. - 1998. - V. 39. - P. 1529-1542. 231. Gladyshev V.N., Hatfield D.L. Selenocysteine-containing proteins in mammals // J. Biomed. Sci. - 1999. - V. 6. - P. 151-160. 232. Go M.L., Wu X., Liu X.L. Chalcones: an update on cytotoxic and chemoprotective properties // Curr. Med. Chem. - 2005. - V. 12(4). - P. 481-499. 287 233. Gobe G. and Crane D. Mitochondria, reactive oxygen species and cadmiumtoxicity in the kidney // Toxicology Letters. - 2010. - V. 198(1). - P. 49– 55. 234. Goering P. L., Fisher B. R., Noren B. T., Papaconstantinou A., Rojko J. L., Marler R. J. Mercury induces regional and cell-specific stress protein expression in rat kidney // Toxicological Sciences. - 2000. - V. 53(2). - P. 447–457. 235. Gong P., Ogra Y., Koizumi S. Inhibitory Effects of Heavy Metals on Transcription Factor Sp1 // Industrial Health. - 2000. - V. 38. - P. 224–227. 236. Gopalakrishnan M., Sureshkumar P., Thanusu J., Kanagarajan V. Design, synthesis, characterization, antibacterial and antifungal activities of a novel class of 5,7-diaryl-4,4-dimethyl-4,5,6,7-tetrahydropyridino[3,4-d]-1,2,3-selenadiazoles // J. Enzyme Inhib. Med. Chem. - 2008. - V. 23. - N 3. - P. 347-351. 237. Grankvist K., Marklund S.L., Taljedal I.B. CuZn-superoxide dismutase, Mnsuperoxide dismutase, catalase and glutathione peroxidase in pancreatic islets and other tissues in the mouse // Biochem. J. - 1981. - V. 199. - P.393-398. 238. Griendling K.K. NADPH oxidases: new regulators of old functions // Antioxid. Redox Signal. - 2006. - V. 8(9-10). - P. 1443–1445. 239. Griscavage J.M., Wilk S., Ignarro L.J. Inhibitors of the proteasome pathway interfere with induction of nitric oxide synthase in macrophages by blocking activation of transcription factor NF-kappa B // Proc. Natl. Acad. Sci. USA. 1996. - V. 93(8). - P. 3308-3312. 240. Gupta M., Dobashi K., Greene E.L., Orak J.K., Singh I. Studies on hepatic injury and antioxidant enzyme activities in rat subcellular organelles following in vivo ischemia and reperfusion // Mol. Cell Biochem. 1997. V. 176. P. 337-347. 241. Guzik T.J., Chen W., Gongora M.C., Guzik B., Lob H.E., Mangalat D. Calcium-dependent NOX5 nicotinamide adenine dinucleotide phosphate oxidase contributes to vascular oxidative stress in human coronary artery disease // J. Am. Coll. Cardiol. - 2008. - V. 52. - P. 1803–1809. 288 242. Halliwell B. Role of free radicals in the neurodegenerative diseases: therapeutic implications for antioxidant treatment // Drugs Aging. - 2001. - V. 18. P. 685-716. 243. Halliwell B., Gutteridge J.M.C. Free Radicals in Biology and Medicine (3 rd ed.). Oxford University Press, NY, USA, 1999. 244. Halliwell B., Gutteridge J.M.C. Lipid peroxidation, oxygen radicals, cell damage, and antioxidant therapy // Lancet. — 1984. — Р. 1396-1398. 245. Halliwell B., Gutteridge J.M.C. Oxygen free radicals and iron in relation to biology and medicine: some problems and concepts // Arch. Bioch. Bioph. - 1986. - V. 246. - P. 501-514. 246. Hamer D.H. Metallothionein // Ann. Rev. Biochem. - 1986. - V. 55. - P. 913951. 247. Hanschmann E.M., Godoy J.R., Berndt C., Hudemann C., Lillig C.H. Thioredoxins, glutaredoxins, and peroxiredoxins-molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling // Antioxid. Redox Signal. - 2013, doi:10.1089/ars.2012.4599. 248. Hart B.A., Potts R.J., Watkin R.D. Cadmium adaptation in the lung—A double-edged sword? // Toxicology. - 2001. - V. 160. - P. 65–70. 249. Hartl F.-U., Hlodan R., Langer T. Molecular chaperones in protein folding: the art of avoiding sticky situations. // Trends in Bioch.Sci. - 1994. - V. 19. - P. 20 25. 250. Hartwig A. Zinc finger proteins as potential targets for toxic metal ions: Differential effects on structure and function // Antioxid. Redox Signal. - 2001. V. 3. - P. 625–634. 251. Hatfield D.L., Berry M.J., Gladyshev V.N. Selenium: Its Molecular Biology and Role in Human Health, 2nd Ed.; Springer: New York, USA, 2006. 252. He A., Wangi J.A., Guil C., Jiang Y., Sun Y., Chen T. Changes of mitochondrial pathway in hypoxia/reoxygenation induced cardiomyocytes apoptosis // Folia Histochem. Cytobiol. - 2007. - V. 45. - P. 397-400. 289 253. Hell R., Wirtz M. Molecular Biology, Biochemistry and Cellular Physiology of Cysteine Metabolism in Arabidopsis thaliana // American Society of Plant Biologists. The Arabidopsis Book. - 2011. - 19 pages. e0154. doi: 10.1199/tab.0154. 254. Hemaiswarya S., Doble M. Synergistic interaction of phenylpropanoids with antibiotics against bacteria // J. Med. Microbiol. - 2010. - V. 59. - P. 1469–1476. 255. Henderson B.E., Ross R., Bernstein L. Estrogens as a cause of human cancer: the Richard and Hinda Rosenthal foundation award lecture // Cancer Res. - 1988. V. 48. - P. 246–253. 256. Hickman M.J., Petti A.A., Ho-Shing O., Silverman S.J., McIsaac R.S., Lee T.A., Botstein D. Coordinated regulation of sulfur and phospholipid metabolism reflects the importance of methylation in the growth of yeast // Mol. Biol. Cell. 2011. - V. 22. - P. 4192–4204. 257. Holl V., Coelho D., Silbernagel L., Keyser J.F., Waltzinger C., Dufour P., Bischoff P.L..Prevention of nitrogen mustard-induced apoptosis in normal and transformed lymphocytes by ebselen // Biochem. Pharmacol. - 2000. - V. 60. - N 11. - P. 1565-1577. 258. Hong H., Zeng J.S., Kreulen D.L., Kaufman D.I., Chen A.F. Atorvastatin protects against cerebral infarction via inhibition of NADPH oxidase-derived superoxide in ischemic stroke // Am. J. Physiol. Heart Circ. Physiol. - 2006. - V. 291. - P. 2210–2215. 259. Hu H. A 50-year follow-up of childhood plumbism: hypertension, renal function, and hemoglobin levels among survivors // Тhe American Journal of Diseases of Children. - 1991. - V. 145(6). - P. 681–687. 260. Hudson A.R., Roach S.L., Higuchi R.I. Recent developments in the discovery of selective glucocorticoid receptor modulators (SGRMs) // Curr. Topics Med. Chem. - 2008. – V. 8. - P. 750–765. 261. Ikediobi C.O., Badisa V.L., Ayuk-Takem L.T., Latinwo L.M., West J. Response of antioxidant enzymes and redox metabolites to cadmium-induced 290 oxidative stress in crl-1439 normal rat liver cells // Int. J. Mol. Med. - 2004. - V. 14. - P. 87–92. 262. Imai H., Nakagawa Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells // Free Radic. Biol. Med. - 2003. - V. 34. - P. 145–169. 263. Inglot A.D., Młochowski J., Zielińska-Jenczylik J., Piasecki E., Ledwoń T.K., Kloc K. Seleno-organic compounds as immunostimulants: an approach to the structure-activity relationship // Arch.Immunol.Ther. Exp. (Warsz). - 1996. - V. 44. - N 1. - P. 67-75. 264. Ivanina A.V., Cherkasov A.S., Sokolova I.M. Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin // The Journal of Experimental Biology. 2008. - V. 211. - P. 577-586. 265. Jacob C., Arteel G. E., Kanda T., Engman L., Sies, H. Water-soluble organotellurium compounds: catalytic protection against peroxynitrite and release of zinc from metallothionein // Chem. Res. Toxicol. - 2000. - V. 13. - Р. 3–9. 266. Jadhav S. H., Sarkar S. N., Patil R. D., Tripathi H. C. Effects of subchronic exposure via drinking water to a mixture of eight water-contaminating metals: a biochemical and histopathological study in male rats // Archives of Environmental Contamination and Toxicology. - 2007. - V. 53(4). - P. 667–677. 267. Jamier V., Ba L.A., Jacob C. Selenium- and tellurium-containing multifunctional redox agents as biochemical redox modulators with selective cytotoxicity // Chemistry. - 2010. - V 16. - Р. 10920–10928. 268. Jang J.H., Aruoma O.I., Jen L.S., Chung H.Y., Surh Y.J. Ergothioneine rescues PC12 cells from β-amyloid-induced apoptotic death // Free Radic. Biol. Med. 2004. - V. 36. - P. 288-299. 269. Jeong E. M., Moon C. H., Kim C.S. Cadmium stimulates the expression of ICAM-1 via NF-kappaB activation in cerebrovascular endothelial cells // Biochemical and Biophysical Research Communications. - 2004. - V. 320. - P. 887–892. 291 270. Jezek P., Hlavata L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism // Int. J. Biochem. Cell Biol. - 2005. - V. 37. - P. 24782503. 271. Jiang F., Zhang Y., Dusting G.J. NADPH oxidase-mediated redox signaling: Roles in cellular stress response, stress tolerance, and tissue repair // Pharmacol. Rev. - 2011. - V. 63. - P. 218–242. 272. Jiang Y., Cheng F., Zhou Y., Xia X., Mao W., Shi K., Chen Z., Yu J. Hydrogen peroxide functions as a secondary messenger for brassinosteroids- induced CO2 assimilation and carbohydrate metabolism in Cucumis sativus // Biomed. & Biotechnol. - 2012. - V. 13. - N 10. - P. 811-823. 273. Jin J.Y., JuskoW.J. Pharmacodynamics of Glucose Regulation by Methylprednisolone. II. Normal Rats // Biopharm. Drug Dispos. - 2009. - V. 30(1). - P. 35–48. doi: 10.1002/bdd.642. 274. Johri N., Jacquillet G., Unwin R. Heavy metal poisoning: The effects of cadmium on the kidney // Biometals. - 2010. - V. 23. - P. 783–792. 275. Jones D.P. Radical-free biology of oxidative stress // Am. J. Physiol. Cell Physiol. - 2008. - V. 295. - P.849–868. 276. Jung Y.S., Kim S.J., Kwon do Y., Jun D.S., Kim Y.C. Significance of alterations in the metabolomics of sulfur-containing amino acids during liver regeneration // Biochimie. - 2013. - V. 95(8). - P. 1605-1610. doi: 10.1016/j.biochi.2013.04.015. 277. Kalinina E.V., Novichkova M.D., Scherbak N.P., Solomka V.S., Saprin A.N. GSH-dependent redox regulation and antioxidant enzymes in the formation of resistance to doxorubicin in K562 human erythroleukemia cells // Adv. Exp. Med. Biol. - 2001. - V. 500. - P. 241–244. 278. Kawahara T., Quinn M.T., Lambeth J.D. Molecular evolution of the reactive oxygen-generating NADPH oxidase (Nox/Duox) family of enzymes // BMC Evol. Biol. - 2007. - V. 7. - P. 109. 292 279. Kelly G.E., Nelson C., Waring M.A., Joannou G.E., Reeder A.Y. Metabolites of dietary (Soya) isoflavones in human urine // Clin. Chem. Acta. - 1993. - V. 223. - P. 9–22. 280. Kessi J., Hanselmann K.W. Similarities between the abiotic reduction of selenite with glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrum and Escherichia coli // J. Biol. Chem. - 2004. - V. 279. - P. 50662–50669. 281. Kirkman H.N., Galiano S., Gaetani G.F. The function of catalase-bound NADPH // J. Biol. Chem. - 1987. - V. 262. - P. 660-666. 282. Kirschvink N., Martin N., Fievez L., Smith N., Marlin D., Gustin P. Airway inflammation in cadmium-exposed rats is associated with pulmonary oxidative stress and emphysema // Free Radic. Res. - 2006. - V. 40. - P. 241–250. 283. Kohen R., Nyska A. Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification // Toxicol Pathol. - 2002. - V. 30. - P. 620-650. 284. Kleinschnitz C., Grund H., Wingler K., Armitage M.E., Jones E., Mittal M. Post-Stroke Inhibition of Induced NADPH Oxidase Type 4 Prevents Oxidative Stress and Neurodegeneration // PLoS Biol. - 2010. - V. 8. e1000479.doi:1000410.1001371/journal.pbio.1000479. 285. Klotz L.-O., Kroncke K.-.D, Buchczyk D.P., Sies H. Role of Copper, Zinc, Selenium and Tellurium in the Cellular Defense against Oxidative and Nitrosative Stress // J. Nutr. - 2003. - V. 133. - Р. 1448–1451. 286. Kobayashi Y., Ogra Y., Ishiwata K., Takayama H., Aimi N., Suzuki K.T. Selenosugars are key and urinary metabolites for selenium excretion within the required to low-toxic range // Proc. Natl. Acad. Sci. USA. - 2002. - V. 99. - P. 15932-15936. 287. Koedrith P., Seo Y.R. Advances in Carcinogenic Metal Toxicity and Potential Molecular Markers // Int. J. Mol. Sci. - 2011. - V. 12. - P. 9576-9595; doi:10.3390/ijms12129576. 293 288. Kohanski M., Dwyer D., Hayete B., Lawrence C., Collins J. A common mechanism of cellular death induced by bactericidal antibiotics // Cell. - 2007. V. 130. - P. 797–810. 289. Kohen R., Nyska A. Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification // Toxicol Pathol. - 2002. - V. 30. - P. 620-650. 290. Koppula S., Kumar H., More S.V., Kim B.W., Kim I.S., Choi D.K. Recent advances on the neuroprotective potential of antioxidants in experimental models of Parkinson's disease // Int. J. Mol. Sci. - 2012. - V. 13(8). - P. 10608-10629. doi: 10.3390/ijms130810608. 291. Kopriva S., Mugford S.G., Baraniecka P., Lee B.-R., Matthewman C.A., Koprivova A. Control of sulfur partitioning between primary and secondary metabolism in Arabidopsis // Frontiers in Plant Science. - 2012. - V. 3. - Article 163. 9 pages. doi: 10.3389/fpls.2012.00163. 292. Koziel R., Pircher H., Kratochwil M., Lener B., Hermann M., Dencher N.A., Jansen-Dürr P. Mitochondrial respiratory chain complex I is inactivated by NADPH oxidase Nox4 // Biochem. J. - 2013. - V. 452. - N 2. - P. 231–239. 293. Krause K.H. Tissue distribution and putative physiological function of NOX family NADPH oxidases // Jpn. J. Infect. Dis. - 2004. - V. 57(5). - P. 28–29. 294. Kryukov G.V., Castellano S., Novoselov S.V., Lobanov A.V., Zehtab O., Guigó R., Gladyshev V.N. Characterization of mammalian selenoproteomes // Science. - 2003. - V. 300. - P. 1439–1443. 295. Kumar S., Bjornstedt M., Holmgren A. Selenite is a substrate for calf thymus thioredoxin reductase and thioredoxin and elicits a large non-stoichiometric oxidation of NADPH in the presence of oxygen // Eur. J. Biochem. - 1992. - V. 207. - P. 435–439. 296. La Plante K., Rybak M. Daptomycin - a novel antibiotic against Gram-positive pathogens // Expert Opin Pharmacother. - 2004. - V. 5. - P. 2321–2331. 294 297. Laity J.H., Lee B.M., Wright P.E. Zinc finger proteins: new insights into structural and functional diversity // Curr. Opin. Struct. Biol. - 2001. - V. 11. - P. 39–46. 298. Lapenna D., Degioia S., Mezzetti A., Porreca E., Ciofani G., Marzio L., Capani F., Diilio C., Cuccurullo F. Circadian Variations in Antioxidant Defences and Lipid Peroxidation in the Rat Heart // Free Rad. Res. - 1992. - V. 17. - P. 187194. 299. Larsen P.R., Berry M.J. Nutritional and hormonal regulation of thyroid hormone deiodinases. // Annu. Rev. Nutr. – 1995. – N 15. – P. 323-352. 300. Lecocq F.R., Mebane D., Madison L.L. The acute effect of hydrocortisone on hepatic glucose output and peripheral glucose utilization // J. Clin. Invest. - 1964. V. 43. - P. 237–246. 301. Lee C., Yang W., Parr R.G. Development of the Colle-Salvetti correlation – energy formula into a functional of the electron density // Physical review B. 1988. - V. 37. – N 2. - P. 785-789. 302. Lehuédé J., Fauconneau B., Barrier L., Ourakow M., Piriou A., Vierfond J.M. Synthesis and antioxidant activity of new tetraarylpyrroles // Eur. J. Med. Chem. 1999. - V. 34. - P. 991-996. 303. Levander O., Burk R.F. Selenium // Present knowledge in nutrition / Eds. E.E. Ziegler, L.J. Filer. 7th ed. N.Y.: Acad. Press, 1998. - P. 320-328. 304. Levi S., Santambrogio P., Corsi B., Cozzi A., Arosio P. Evidence that residues exposed on the three-fold channels have active roles in the mechanism of ferritin iron incorporation // Biochem J. - 1996. - V. 317(Pt 2). - P. 467-473. 305. Li W.-H., Shi Y.-C., Tseng I-L., Liao V. H.-C. Protective Efficacy of Selenite against Lead-Induced Neurotoxicity in Caenorhabditis elegans // PLoS ONE. 2013. - V. 8(4). - 9 pages. e62387. doi:10.1371/journal.pone.0062387. 306. Liao C.Y., Zhou Q.F., Fu J.J., Shi J.B., Yuan C.G., Jiang G.B. Interaction of methylmercury and selenium on the bioaccumulation and histopathology in medaka (Oryziaslatipes) // Environ. Toxicol. - 2007. - V. 22. - N 1. - P. 69-77. 295 307. Lillig C.H., Lonn M.E., Enoksson M., Fernandes A.P., Holmgren A. Short interfering RNA-mediated silencing of glutaredoxin 2 increases the sensitivity of HeLa cells toward doxorubicin and phenylarsine oxide // Proc. Natl. Acad. Sci. USA. - 2004. - V. 101. - P. 13227–13232. 308. Lindenblatt N., Schareck W., Belusa L., Nickels R.M., Menger M.D., Vollmar B. Anti-oxidant ebselen delays microvascular thrombus formation in the rat cremaster muscle by inhibiting platelet P-selectin expression // Thromb. Haemost. - 2003. - V. 90. - N 5. - P. 882-892. 309. Lithgow J. K., Hayhurst E. J., Cohen G., Aharonowitz Y., J. Foster S. Role of a cysteine synthase in Staphylococcus aureus // J. Bacteriol. - 2004. - V. 186. P. 1579–1590. 310. Little C., O'Brien P.J. An intracellular GSH-peroxidase interaction with a lipid peroxide substrate // Biochem. Biophys. Res. Commun. - 1968. - V. 31. - P.145150. 311. Liu C.Y., Grant A.L., Kim K.H., Ji S.Q., Hancock D.L., Anderson D.B., Mills S.E. Limitations of ractopamine to affect adipose-tissue metabolism in swine // J. Anim. Sci. - 1994. - V. 72. - P. 62-67. 312. Liu X., Silks L.A., Liu C., Ollivault-Shiflett M., Huang X., Li J., Luo G., Hou Y.M., Liu J., Shen J. Incorporation of tellurocysteine into glutathione transferase generates high glutathione peroxidase efficiency // Angew. Chem. Int. Ed. Engl. 2009. - V. 48. - P. 2020–2023. 313. Lopez E., Arce C., Oset-Gasque M.J., Canadas S., Gonzalez M.P. Cadmium induces reactive oxygen species generation and lipid peroxidation in cortical neurons in culture // Free Radic. Biol. Med. - 2006. - V. 40. - P. 940–951. 314. Lovell M.A., Markesbery W.R. Oxidative damage in mild cognitive impairment and early Alzhelimer's disease // J Neurosci Res. - 2007. - V. 85. P. 3036-3040. 315. Lu T.K., Collins J.J. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy // Proc. Natl. Acad. Sci. USA. - 2009. - V. 106. P. 4629–4634. 296 316. Lubos E., Loscalzo J., Handy D.E. Glutathione Peroxidase-1 in Health and Disease: From Molecular Mechanisms to Therapeutic Opportunities // Antioxidants & redox signaling. - 2011. - V. 15. - Р. 1957-1997. DOI: 10.1089/ars.2010.3586. 317. Lundberg M., Johansson C., Chandra J., Enoksson M., Jacobsson G., Ljung J., Johansson M., Holmgren A. Cloning and expression of a novel human glutaredoxin (Grx2) with mitochondrial and nuclear isoforms // J. Biol. Chem. 2001. - V. 276. - P. 26269–26275. 318. Luparello C., Sirchia R., Longo A. Cadmium as a transcriptional modulator in human cells // Crit. Rev. Toxicol. - 2011. - V. 41. - P. 75–82. 319. Lysell J., Vladic Y.S., Ciarlo N., Holmgren A., Sahlin L. Immunohistochemical determination of thioredoxin and glutaredoxin distribution in the human cervix, and possible relation to cervical ripening // Gynecol. Endocrinol. - 2003. - V. 17. - P. 303–310. 320. Makino J., Kamiya T., Hara H., Adachi T. TPA induces the expression of ECSOD in human monocytic THP-1 cells: involvement of PKC, MEK/ERK and NOX-derived ROS // Free Radic Res. - 2012. - V. 46(5). - P. 637-644. doi: 10.3109/10715762.2012.664841. 321. Mandel J.S., McLaughlin J.K., Schlehofer B., Mellemgaard A., Helmuert U., Linbald R., McCredie M., Adami U. International renal-cell cancer study. IV. Occupation // Int. J. Cancer. - 1995. - V. 61. - P. 601-605. 322. Manikandan R., Beulaja M., Thiagarajan R., Arumugam M. Effect of curcumin on the modulation of alphaA- and alphaB-crystallin and heat shock protein 70 in selenium-induced cataractogenesis in Wistar rat pups // Mol. Vis. - 2011. - V. 17. P. 388–394. 323. Marchetti C. Role of Calcium Channels in Heavy Metal Toxicity // ISRN Toxicology. - V. 2013. Article ID 184360. 9 p. in the http://dx.doi.org/10.1155/2013/184360. 324. Maret W. Cellular zinc and redox states converge metallothionein/thionein pair // J. Nutr. - 2003. - V. 133. - P. 1460S–1462S. 297 325. Maritim A.C., Sanders R.A., Watkins J.B. Diabetes, oxidative stress, and antioxidants: a review // J Biochem Molec Toxicol. - 2003. - V. 17. - Р. 24–38. 326. Marklund S.L. Extracellular superoxide dismutase and other superoxide dismutase isoenzymes in tissues from nine mammalian species // Biochem J. 1984. - V. 222. - P. 649–655. 327. Marklund SL. Extracellular superoxide dismutase in human tissues and human cell lines // J. Clin. Invest. - 1984. - V. 74. - P. 1398–1403. 328. Martin P.M., Horwitz K.B., Rujan D.S., McGuire W.L. Phytoestrogen interaction with estrogen receptors in human breast cancer cells // Endocrinology. 1972. - V. 52. - P. 299–310. 329. Marut W.K., Kavian N., Servettaz A., Nicco C., Ba L.A., Doering M., Chereau C., Jacob C., Weill B., Batteux F. The Organotelluride Catalyst (PHTE)2NQ Prevents HOCl-Induced Systemic Sclerosis in Mouse // Journal of Investigative Dermatology. - 2012. - V. 132. - Р. 1125–1132; doi:10.1038/jid.2011.455. 330. Mastrocola R., Restivo F., Vercellinatto I. Oxidative and nitrosative stress in brain mitochondria of diabetic rats // Journal of Endocrinology. - 2005. - V. 187(1). - P. 37–44. 331. Masumoto H., Kissner R., Koppenol W. H. & Sies H. Kinetic study of the reaction of ebselen with peroxynitrite // FEBS Lett. - 1996. - V. 398. - P. 179–182. 332. Mates J.M., Segura J.A., Alonso F.J., Marquez J. Roles of dioxins and heavy metals in cancer and neurological diseases using ROS-mediated mechanisms // Free Radic. Biol. Med. - 2010. - V. 49. - P. 1328–1341. 333. Mates J.M., Segura J.A., Alonso F.J., Marquez J. Sulphur-containing non enzymatic antioxidants: therapeutic tools against cancer // Front Biosci. (Schol Ed). - 2012. - V. 4. - P. 722-748. 334. Matough F.A., Budin S.B., Hamid Z.A., Alwahaibi N., Mohamed J. The role of oxidative stress and antioxidants in diabetic complications // Sultan Qaboos Univ. Med. J. - 2012. - V. 12(1). - P. 5-18. 298 335. Matovic V., Buha A., Bulat Z., Ðukic- Cosic D. Cadmium toxicity revisited: focus on oxidative stress induction and interactions with zinc and magnesium // Archives of Industrial Hygiene and Toxicology. - 2011. - V. 62. - P. 65–76. 336. Mavelli I., Rigo A., Federico R., Ciriolo M.R., Rotilio G. Superoxide dismutase, glutathione peroxidase and catalase in developing rat brain // Biochem. J. - 1982. - V. 204. - P.535-540. 337. Mbemba F., Houbion A., Raes M., Remacle J. Subcellular localization and modification with ageing of glutathione, glutathione peroxidase and glutathione reductase activities in human fibroblasts // Biochim Biophys Acta. - 1985. - V. 838. - P. 211-220. 338. McCall M.R., Frei B. Can antioxidant vitamins materially reduce oxidative damage in humans? // Free Radic Biol Med. - 1999. - V. 26. - P. 1034-1053. 339. McCord J.M., Fridovich I. Superoxide dismutase: An enzymatic function for erythrocuprein (hemocuprein) // J. Biol. Chem. - 1969. - V. 244. - P. 6049–6055. 340. McGrath N.A., Raines R.T. Chemoselectivity in Chemical Biology: Acyl Transfer Reactions with Sulfur and Selenium // Acc. Chem. Res. - 2011. - V. 44(9). - P. 752–761. doi:10.1021/ar200081s. 341. Meagher E.A., FitzGerald G.A. Indices of lipid peroxidation in vivo: strengths and limitations // Free Radic Biol Med. - 2000. - V. 28. - P. 1745-1750. 342. Mehdi Y., Hornick J.-L., Istasse L., Dufrasne I. Selenium in the Environment, Metabolism and Involvement in Body Functions // Molecules. - 2013. - V. 18. P. 3292-3311. 343. Meinerz D.F., Comparsi B., Allebrandt J., Mariano D.O.C., dos Santos D.B, Zemolin P.A.P., Farina M., Dafre L.A., Rocha J.B.T., Posser T., Franco J.L. Subacute administration of (S)-dimethyl 2-(3-(phenyltellanyl) propanamido) succinate induces toxicity and oxidative stress in mice: unexpected effects of Nacetylcysteine // Springer Plus. - 2013. - N 2. - P. 182-188. 344. Messner B., Bernhard D. Cadmium and cardiovascular diseases: Cell biology, pathophysiology, and epidemiological relevance // Biometals. - 2010. - V. 23. P. 811–822. 299 345. Mills G.C. Hemoglobin catabolism. Glutatione peroxidase is an erythrocyte enzyme which protects hemoglobin from oxidative breakdown // J. Biol. Chem. – 1957. – V. 229. – Р. 189–194. 346. Mills S.E. Biological basis of the ractopamine response // J. Anim. Sci. - 2001. - V. 79. - Suppl. 1. - P. 238. 347. Miranda-Vizuete A., Damdimopoulos A.E., Spyrou G. cDNA cloning, expression and chromosomal localization of the mouse mitochondrial thioredoxin reductase gene(1) // Biochim. Biophys. Acta. - 1999. - V. 1447. - P. 113–118. 348. Mistry H.D., Pipkin F.B., Redman C.W., Poston L. Selenium in reproductive health // Am. J. Obstet. Gynecol. - 2012. - V. 206. - P. 21–30. 349. Młochowski J., Kloc K., Lisiak R., Potaczek P. Developments in the chemistry of selenaheterocyclic compounds of practical importance in synthesis and medicinal biology // Issue in Honor of Prof. Jan Epsztajn. Arkivoc. - 2007. - V. 6. P. 14-46. 350. Mohler M.L., He Y., Wu Z., Hong S.S. Dissociated non-steroidal glucocorticoids: Tuning out untoward effects // Expert Opin. Ther. Patents. - 2008. - V. 17. - P. 37–58. 351. Molina-Navarro M.M., Casas C., Piedrafita L., Belli G., Herrero E. Prokaryotic and eukaryotic monothiol glutaredoxins are able to perform the functions of Grx5 in the biogenesis of Fe/S clusters in yeast mitochondria // FEBS Lett. - 2006. - V. 580. - P. 2273–2280. 352. Molina-Quiroz R.C., Munoz-Villagran C.M., de la Torre E., Tantalean J.C., Vasquez C.C., Perez-Donoso J.M. Enhancing the Antibiotic Antibacterial Effect by Sub Lethal Tellurite Concentrations: Tellurite and Cefotaxime Act Synergistically in Escherichia coli // PLoS ONE. - 2012. – V. 7. - Issue 4. e35452. 353. Monteiro H.P., Winterbourn C.C. Release of iron from ferritin by divicine, isouramil, acid-hydrolyzed vicine, and dialuric acid and initiation of lipid peroxidation // Arch Biochem Biophys. - 1989. - V. 271. - P.536-545. 300 354. Monteiro H.P., Winterbourn C.C. The superoxide-dependent transfer of iron from ferritin to transferrin and lactoferrin // Biochem. J. - 1988. - V. 256. - P. 923928. 355. Moon Y.J., Wang X., Morris M.E. Dietary flavonoids: Effects on xenobiotic and carcinogen metabolism // Toxicol. in Vitro. - 2006. - V. 20. - P. 187-210. 356. Moore M.D., Kaplan S. Identification of intrinsic high-level resistance to rareearth oxides and oxyanions in members of the class Proteobacteria: characterization of tellurite, selenite, and rhodium sesquioxide reduction in Rhodobacter sphaeroides // J. Bacteriol. - 1992. - V. 174. - P. 1505-1514. 357. Muller A., Cadenas E., Graf P., Sies H. A novel biologically active slenoorganic compound: Glutathione peroxidase-like activity in vitro and antioxidant capacity of PZ 51 (Ebselen) // Biochem. Pharmacol. - 1984. - V. 15. - P. 3235– 3239. 358. Murray D.B., Beckmann M., Kitano H. Regulation of yeast oscillatory dynamics // Proc. Natl. Acad. Sci. USA. - 2007. - V. 104. - P. 2241–2246. 359. Nabeebaccus A., Zhang M., Shah A.M. NADPH oxidases and cardiac remodeling // Heart Fail. Rev. - 2011. - V. 16. - P. 5–12. 360. Nair A.R., DeGheselle O., Smeets K., Kerkhove E.V., Cuypers A. CadmiumInduced Pathologies: Where Is the Oxidative Balance Lost (or Not)? // Int. J. Mol. Sci. - 2013. - V. 14. - P. 6116-6143; doi:10.3390/ijms14036116. 361. Narajji C., Karvekar M. and Das K. Biological importance of organoselenium compounds // Indian Journal of Pharmaceutical Sciences. - 2007. - V. 3. - P. 344351. 362. National Toxicology Program, “Health Efects of Low-level Lead Evaluation,” 2012, http://ntp.niehs.nih.gov/?objectid=4F04B8EA-B187-9EF2- 9F9413C68E76458E. 363. Nemmiche S., Chabane-Sari D., Kadri M., Guiraud P. Cadmium chlorideinduced oxidative stress and DNA damage in the human Jurkat T cell line is not linked to intracellular trace elements depletion // Toxicol. In Vitro. - 2011. - V. 25. - P. 191-198. 301 364. Niki E., Yoshida Y., Saito Y., Noguchi N. Lipid peroxidation: Mechanisms, inhibition, and biological effects // Biochem. Biophys. Res. Commun. - 2005. V. 338. - P. 668-676. 365. Nolan C.V., Shaikh Z.A. Lead nephrotoxicity and associated disorders: biochemical mechanisms // Toxicology. - 1992. - V. 73(2). - P. 127–146. 366. Nordberg J., Arner E.S. Reactive oxygen species, antioxidants and the mammalian thioredoxin system // Free Radic. Biol. Med. - 2001. - V. 31. - P. 1287-1312. 367. Nozawa R., Yokota T., Fujimoto T. Susceptibility of methicillin-resistant Staphylococcus aureus to the selenium-containing compound 2-phenyl-1,2benzoisoselenazol-3(2H)-one (PZ51). // Antimicrob. Agents Chemother. - 1989. V. 33. - N 8. - P. 1388-1390. 368. Ogra Y., Anan Y. Selenometabolomics Explored by Speciation // Biol. Pharm. Bull. - 2012. - V. 35(11). - P. 1863–1869. 369. Okado-Matsumoto A., Fridovich I. Subcellular distribution of superoxide dismutases (SOD) in rat liver: Cu,Zn-SOD in mitochondria // J. Biol. Chem. 2001. - V. 276. - P. 38388–38393. 370. Olson J.A. Carotenoids and vitamin A - An overview // Lipid-Soluble Antioxidants: Biochemistry and Clinical Applications. - Basel: Birkhauser Veriag, 1992. - P. 178-192. 371. Ookawara T., Imazeki N., Matsubara O., Kizaki T., Oh-Ishi S., Nakao C., Sato Y., and Ohno H. Tissue distribution of immunoreactive mouse extracellular superoxide dismutase // Am. J. Physiol. - 1998. - V. 275. - P. C840–C847. 372. Otterness D.M., Weinshilboum R.M. Mouse thiopurine methyltransferase pharmacogenetics: monogenic inheritance // J. Pharmacol. Exp. Ther. - 1987. V. 240. - P. 817–824. 373. Ottosson L.-G., Logg K., Ibstedt S., Sunnerhagen P., Kall M., Blomberg A., Warringer J. Sulfate Assimilation Mediates Tellurite Reduction and Toxicity in Saccharomyces cerevisiae // Eukaryotic cell. - 2010. - V. 9(10). - P. 1635–1647. 302 374. Paglia D.E., Valentine W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase // J. Lab. Clin. Med. - 1967. - V. 70. - P. 158-169. 375. Pamenter M.E., Ali S.S., Tang Q., Finley J.C., Gu X.Q., Dugan L.L., Haddad G.G. An in vitro ischemic penumbral mimic perfusate increases NADPH oxidase-mediated superoxide production in cultured hippocampal neurons // Brain Res. - 2012. - V. 1452. - P. 165–172. doi:10.1016/j.brainres.2012.03.004. 376. Patching S.G., Gardiner P.H.E. Recent developments in selenium metabolism and chemical speciation: a review // J. Trace Elements Med. Biol. - 1999. - V. 13. P. 193-214. 377. Pendyala S., Gorshkova I.A., Usatyuk P.V., He D., Pennathur A., Lambeth J.D., Thannickal V.J., Natarajan V. Role of Nox4 and Nox2 in hyperoxia-induced reactive oxygen species generation and migration of human lung endothelial cells // Antioxid. Redox Signal. - 2009. - V. 11(4). - P. 747–64. 378. Pendyala S., Natarajan V. Redox regulation of Nox proteins // Respir. Physiol. Neurobiol. - 2010. - V. 174(3). - P. 265–271. doi:10.1016/j.resp.2010.09.016. 379. Peng H., Chen W., Cheng Y., Hakuna L., Strongin R., Wang B. Thiol Reactive Probes and Chemosensors // Sensors. - 2012. - V. 12. - P. 15907-15946; doi:10.3390/s121115907. 380. Perez J., Arenas F., Pradenas G., Sandoval J., Vasquez C.C. Escherichia coli YqhD exhibits aldehyde reductase activity and protects from the harmful effect of lipid peroxidation-derived aldehydes // J. Biol. Chem. - 2008. - V. 283. - P. 7346– 7353. 381. Pérez J.M., Calderón I.L., Arenas F.A., Fuentes D.E., Pradenas G.A., Fuentes E.L., Sandoval J.M., Castro M.E., Elías A.O., Vásquez C.C. Bacterial toxicity of potassium tellurite: unveiling an ancient enigma // PLoS ONE. - 2007. - V. 2(2). – 9 p. e211. doi:10.1371/journal.pone.0000211. 382. Petering D.H., Huang M., Moteki S., Shaw III C.F. Cadmium and lead interactions with transcription factor IIIA from Xenopus laevis: a model for zinc 303 finger protein reactions with toxic metal ions and metallothionein // Mar. Environ. Res. - 2000. - V. 50(1-5). - P. 89–92. 383. Peterson S.A., Ralston N.V., Peck D.V., Van Sickle J., Robertson J.D., Spate V.L., Morris J.S. How might selenium moderate the toxic effects of mercury in stream fish of the western U.S.? // Environ. Sci. Technol. - 2009. - V. 43. - N 10. - P. 3919-25. 384. Petti A.A., Crutchfeld C.A., Rabinowitz J.D., Botstein D. Survival of starving yeast is correlated with oxidative stress response and nonrespiratory mitochondrial function // Proc. Natl. Acad. Sci. USA. - 2011. - V. 108. - P. E1089–E1098. 385. Petti A.A., McIsaaca R.S., Ho-Shinga O., Bussemakerc H.J., Botstein D. Combinatorial control of diverse metabolic and physiological functions by transcriptional regulators of the yeast sulfur assimilation pathway // Molecular Biology of the Cell. - 2012. - V. 23. - P. 3008-3024. 386. Petit-Topin I., Turque N., Fagart J., Fay M., Ulmann A., Gainer E., RafestinOblin M.E. Met909 Plays a Key Role in the Activation of the Progesterone Receptor and Also in the High Potency of 13-Ethyl Progestins // Mol. Pharmacol. 2009. - V. 75. - P. 1317–1324. 387. Pietka-Ottlik M.,Wójtowicz-Młochowska H., Kołodziejczyk K., Piasecki E. New organoselenium compounds active against pathogenic bacteria, fungi and viruses // Chem. Pharm. Bull. (Tokyo). - 2008. - V. 56. - N 10. - P. 1423-1427. 388. Poli G., Leonarduzzi G., Biasi F., Chiarpotto E. Oxidative stress and cell signaling // Curr. Med. Chem. - 2004. - V. 11. - P. 1163–1182. 389. Pounds J. G. Effect of lead intoxication on calcium homeostasis and calciummediated cell function: a review // NeuroToxicology. - 1984. - V. 5(3). - P. 295– 331. 390. Powis G., Montfort W.R. Properties and biological activities of thioredoxins // Annu. Rev. Pharmacol. Toxicol. - 2001. - V. 41. - P. 261–295. 391. Puigserver P., Rhee J., Donovan J., Walkey C.J., Yoon J.C., Oriente F., Kitamura Y., Altomonte J., Dong H., Accili D., Spiegelman B.M. Insulin-regulated 304 hepatic gluconeogenesis through FOXO1-PGC-1alpha interaction // Nature. 2003. - V. 423. - P. 550–555. 392. Puthanveetil P., Wang F., Kewalramani G., Kim M.S., Hosseini-Beheshti E., Ng N., Lau W., Pulinilkunnil T., Allard M., Abrahani A., Rodrigues B. Cardiac glycogen accumulation after dexamethasone is regulated by AMPK // Am. J. Physiol. Heart Circ. Physiol. - 2008. - V. 295. - P. H1753–H1762. doi:10.1152/ajpheart.518.2008. 393. Qi D., Pulinilkunnil T., An D., Ghosh S., Abrahani A., Pospisilik J.A., Brownsey R., Wambolt R., Allard M., Rodrigues B. Single-dose dexamethasone induces whole-body insulin resistance and alters both cardiac fatty acid and carbohydrate metabolism // Diabetes. - 2004. - V. 53. - P. 1790–1797. 394. Rabbani G.H., Saha S.K., Akhtar M., Marni F., Mitra A.K., Ahmed S., Alauddin M., Bhattacharjee M., Sultana S., Chowdhury A.K. Antioxidants in detoxification of arsenic - induced oxidative injury in rabbits // J. Environ. Sci. Health. Part. A Tox. Hazard. Subst. Environ. Eng. - 2003. - V. 38. - N 1. - P. 273287. 395. Radi R., Peluffo G., Alvarez M.N., Naviliat M., Cayota A. Unraveling peroxynitrite formation in biological systems // Free Radic. Biol. Med. - 2001. - V. 30. - P. 463-488. 396. Ralston N.V., Blackwell J.L. 3rd, Raymond L.J. Importance of molar ratios in selenium-dependent protection against methylmercury toxicity // Biol. Trace Elem. Res. - 2007. - V. 119. - N 3. - P. 255-268. 397. Ralston N.V., Ralston C.R., Blackwell J.L. 3rd, Raymond L.J. Dietary and tissue selenium in relation to methylmercury toxicity // Neurotoxicology. - 2008. V. 29. - N 5. - P. 802-811. 398. Ranjard L., Prigent-Combaret C., Nazaret S. and Cournoyer B. Methylation of inorganic and organic selenium by the bacterial thiopurine methyltransferase // J. Bacteriol. - 2002. - V. 184. - P. 3146–3149. 305 399. Ratnam D.V.; Ankola D.D.; Bhardwaj V.; Sahana D.K.; Kumar M.N. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective // J. Control. Release. - 2006. - V. 113. - Р. 189–207. 400. Reddy C.C., Li N.Q., Reddy P.S., Hildenbrandt G.R., Reddy A.P., Scholz R.W., Tu C.-P.D. Selenium-dependent glutathione peroxidase: expression in selenium deficiency // Biological Oxidation systems. - 1990. - V. 1. - P.473-485. 401. Reeves M.A., Hoffmann P.R. The human selenoproteome: recent insights into functions and regulation // Cell. Mol. Life Sci. - 2009. - V. 66. - P. 2457–2478. 402. Reiss U., Gershon D. Comparison of cytoplasmic superoxide dismutase in liver, heart and brain of aging rats and mice // Biochem. Biophys. Res. Commun. 1976. - V. 73. - P.255-262. 403. Reus I.S., Bando I., Andrés D., Cascales M. Relationship between expression of HSP70 and metallothionein and oxidative stress duringmercury chloride induced acute liver injury in rats // Journal of Biochemical and Molecular Toxicology. - 2003. - V. 17(3). - P. 161–168. 404. Reyes J.L., Molina-Jijón E., Rodríguez-Muñoz R., Bautista-García P., DebrayGarcía Y., del Namorado M.C. Tight Junction Proteins and Oxidative Stress in Heavy Metals-Induced Nephrotoxicity // BioMed Research International. – V. 2013. Article ID 730789. 14 p. http://dx.doi.org/10.1155/2013/730789. 405. Rhee S.G. Cell signaling. H2O2, a necessary evil for cell signaling // Science. 2006. - V. 312. - P. 1882–1883. 406. Rhee S.G., Kang S.W., Chang T.-S., Jeong W., Kim K. Peroxiredoxin, a novel family of peroxidases // IUBMB Life. - 2001. - V. 52. - P. 35–41. 407. Rossato J.I., Ketzer L.A., Centurião F.B., Silva S.J., Lüdtke D.S., Zeni G., Braga A.L., Rubin M.A., Rocha J.B. Antioxidant properties of new chalcogenides against lipid peroxidation in rat brain // Neurochem. Res. - 2002. - V. 27. - N. 4. P. 297-303. 408. Rotruck J., Pope A., Ganther H., Swanson A., Hoekstra W. Selenium: biochemical role as a component of glutathione peroxidase // Science. - 1973. V. 179. - P.588-590. 306 409. Rubbo H., Radi R., Anselmi D., Kirk M., Barnes S., Butler J., Eiserich J.P., Freeman B.A. Nitric oxide reaction with lipid peroxyl radicals spares alphatocopherol during lipid peroxidation // J. Biol. Chem. - 2000. - V. 275. - P. 1081210818. 410. Sakamoto M., Murata K., Kakita A., Sasaki M. A review of mercury toxicity with special reference to methylmercury / in Environmental Chemistry and Toxicology of Mercury // G. Liu, Y. Cai, and N. O’Driscoll, Eds. - P. 501–516, John Wiley & Sons, New York, NY, USA, 2011. 411. Sakurai T., Kanayama M., Shibata T., Itoh K. Ebselen, a seleno-organic antioxidant, as an electrophile // Chem. Res. Toxicol. - 2006. - V. 19. - N. 9. P. 1196-204. 412. Salvatore D., Bartha T., Harney J.W., Larsen P.R. Molecular biological and biochemical characterization of the human type 2 selenodeiodinase // Endocrinology. – 1996. – V. 137. - N 8. – Р. 3308-3315. 413. Salvatore D., Low S.C., Berry M., Maia A.L., Harney J.W., Croteau W., St Germain D.L., Larsen P.R. Type 3 Iodothyronine deiodinase: cloning, in vitro expression, and functional analysis of the placental selenoenzyme // J. Clin. Invest. – 1995. – V. 96. N 5. – Р. 2421-2430. 414. Samokyszyn V.M., Thomas C.E., Reif D.W., Saito M., Aust S.D. Release of iron from ferritin and its role in oxygen radical toxicities // Drug Metabol. Rev. 1988. - V. 19. - P.283-303. 415. Sandoval J.M., Arenas F.A., Vasquez C.C. Glucose-6-Phosphate Dehydrogenase Protects Escherichia coli from Tellurite-Mediated Oxidative Stress // PLoS ONE. - 2011. - V. 6(9): e25573. doi:10.1371/journal.pone.0025573. 416. Sartoretto J.L., Kalwa H., Shiroto T., Sartoretto S.M., Pluth M.D., Lippard S.J., Michel T. Role of Ca2+ in the control of H2O2-modulated phosphorylation pathways leading to eNOS activation in cardiac myocytes // PLOS ONE. - 2012. V. 7. - Issue 9. e44627. - 14 p. 307 417. Satarug S. Long-term exposure to cadmium in food and cigarette smoke, liver effects and hepatocellular carcinoma // Curr. Drug Metab. - 2012. - V. 13. - P. 257–271. 418. Satarug S., Garrett S.H., Sens M.A., Sens D.A. Cadmium, environmental exposure, and health outcomes // Environ. Health Perspect. - 2010. - V. 118. P. 182–190. 419. Satarug S., Moore M.R. Adverse health effects of chronic exposure to lowlevel cadmium in foodstuffs and cigarette smoke // Environ. Health Perspect. 2004. - V. 112. - P. 1099–1103. 420. Savolainen H. Superoxide dismutase and glutathione peroxidase activities in rat brain // Res. Commun. Chem. Pathol. Pharm. - 1978. - V. 21. - P.173-176. 421. Schäcke H., Berger M., Hansson T.G., McKerrecher D., Rehwinkel H. Dissociated non-steroidal glucocorticoid receptor modulators: An update on new compounds // Expert. Opin. Ther. Patents. - 2008. - V. 18. - P. 339–352. 422. Schafer F.Q., Buettner G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple // Free Radic. Biol. Med. - 2001. - V. 30. - P. 1191-1212. 423. Scheuhammer A.M., Basu N., Burgess N.M., Elliott J.E., Campbell G.D., Wayland M., Champoux L., Rodrigue J. Relationships among mercury, selenium, and neurochemical parameters in common loons (Gaviaimmer) and bald eagles (Haliaeetusleucocephalus) // Ecotoxicology. - 2008. - V. 17. - N 2. - P. 93-101. 424. Schluter T., Zimmermann U., Protzel C., Miehe B., Klebingat K.J., Rettig R. Intrarenal artery superoxide is mainly NADPH oxidase-derived and modulates endothelium-dependent dilation in elderly patients // Cardiovasc. Res. - 2010. - V. 85. - P. 814–824. 425. Schrauser G.N. The nutritional significance, metabolism and toxicology of selenomethionine // Adv. Food Nutr. Res. - 2003. - V. 47. - P. 73-112. 426. Schwartz G.G., Il’yasova D., Ivanova A. Urinary cadmium, impaired fasting glucose, and diabetes in the NHANES III // Diabetes Care. - 2003. - V. 26. - P. 468–470. 308 427. Schwerdtle T., Ebert F., Thuy C. Genotoxicity of soluble and particulate cadmium compounds: impact on oxidative DNA damage and nucleotide excision repair // Chem. Res. Toxicol. - 2010. - V. 23. - P. 432-442. 428. Sears M.E. Chelation: Harnessing and Enhancing Heavy Metal Detoxification — A Review // Тhe Scientiic World Journal. - V. 2013. Article ID 219840. 13 pages. dx.doi.org/10.1155/2013/219840. 429. Shanker G., Aschner J.L., Syversen T., Aschner M. Free radical formation in cerebral cortical astrocytes in culture induced by methylmercury // Molecular Brain Research. - 2004. - V. 128. - P. 48–57. 430. Shchedrina V.A., Zhang Y., Labunskyy V.M., Hatfield D.L. Structure–function relations, physiological roles, and evolution of mammalian er-resident selenoproteins // Antioxid. Redox Signal. - 2010. - V. 12. - P. 839–849. 431. Shchrauser H.W. Selenium // Elements and their Compounds in the Environment. V. 3. Nonmetals. Wiley-VCH Verlag, 2003. - P. 100-106. 432. Shibata A., Tanabe E., Inoue S., Kitayoshi M., Okimoto S., Hirane M., Araki M., Fukushima N., Tsujiuchi T. Hydrogen peroxide stimulates cell motile activity through LPA receptor-3 in liver epithelial WB-F344 cells // Biochem. Biophys. Res. Commun. - 2013. - V. 433. - N 3. - P. 317-321. 433. Shimada H., Yasutake A., Hirashima T. Strain difference of cadmium accumulation by liver slices of inbred Wistar-Imamichi and Fischer 344 rats // Toxicology In Vitro. - 2008. - V. 22(2). - P. 338–343. 434. Shutt D.A., Cox R.L. Steroid and phytoestrogen binding to sheep uterine receptors in vitro // J. Endocrinol. - 1972. - V. 52. - P. 299–310. 435. Sies H. Oxidative stress: oxidants and antioxidants // Expe. Physiol. - 1997. V. 82. - P. 291-295. 436. Silva J.P., Coutinho O.P. Free radicals in the regulation of damage and cell death – basic mechanisms and prevention // Drug Discoveries & Therapeutics. 2010. - V. 4(3). - Р. 144-167. 437. Sobutskii M.P., Kovanko E.G., Liutinskii S.I., Ivanov S.D. Effect of age and gender on genotoxic and biochemical indexes in animal blood after low doses of 309 radiation-mercury exposures // Advances in Gerontology. - 2007. - V. 20(2). - P. 91–96. 438. Somayajulu M., McCarthy S., Hung M., Sikorska M., Borowy-Borowski H., Pandey S. Role of mitochondria in neuronal cell death induced by oxidative stress; neuroprotection by Coenzyme Q10 // Neurobiol. Dis. - 2005. - V. 18. - P.618-627. 439. Soni D., Gupta P. K., Kumar Y. and Chandrashekhar T G. Antibacterial activity of some unsymmetrical diorganyltellurium(IV) dichlirides // Indian Journal of Biochemistry and Biophyscs. - 2005. - V. 42. - P. 398-400. 440. Speckmann B., Walter P.L., Alili L., Reinehr R., Sies H., Klotz L.O., Steinbrenner H. Selenoprotein P expression is controlled through interaction of the coactivator PGC-1alpha with FoxO1a and hepatocyte nuclear factor 4alpha transcription factors // Hepatology. - 2008. - V. 48. - P. 1998–2006. 441. Spiteller G. Lipid peroxidation in aging and age-dependent diseases // Exp. Gerontol. - 2001. - V. 36. - P. 1425-1457. 442. Stadtman E.R. Role of oxidant species in aging // Curr. Med. Chem. - 2004. V. 11. - P. 1105-1112. 443. Steinbrenner H., Speckmann B., Pinto A., Sies H. High selenium intake and increased diabetes risk: experimental evidence for interplay between selenium and carbohydrate metabolism // J. Clin. Biochem. Nutr. - 2011. - V. 48. - N. 1. - P. 40– 45. 444. Stewart J.J.P. Optimizationofparametersforsemiempiricalmetheds. I. Method // J. Comput. Chem. - 1989. - V. 10. – N 2. - P. 209-220. 445. Stewart J.J.P. Optimization of parameters for semiempirical metheds. II. Applications // J. Comput. Chem. - 1989. - V. 10. – N 2. - P. 221-264. 446. Stinson L.J., Darmon A.J., Dagnino L., D’Souza S.J.A. Delayed apoptosis post-cadmium injury in renal proximal tubule epithelial cells // The American Journal of Nephrology. - 2003. - V. 23(1). - P. 27–37. 447. Stocker R., Frei B. Endogenous antioxidant defences in human blood plasma. In: Sies H. ed. Oxidative stress: oxidants and antioxidants. London: Academic Press. — 1991. — P. 213-243. 310 448. Stralin P., Karlsson K., Johansson B.O., Marklund S.L. The interstitium of the human arterial wall contains very large amounts of extracellular superoxide dismutase // Arterioscler. Thromb. Vasc. Biol. - 1995. - V. 15. - P. 2032–2036. 449. Stults F.H., Forstrom J.W., Chiu D.T.Y., Tappel A.L. Rat liver glutathione peroxidase: purification and study of multiple forms // Arch. Biochem. Biophys. 1977. - V. 183. - P.490-497. 450. Sturtz L.A., Diekert K., Jensen L.T., Lill R., Culotta V.C. A fraction of yeast Cu,Zn-superoxide dismutase and its metallochaperone, CCS, localize to the intermembrane space of mitochondria. A physiological role for SOD1 in guarding against mitochondrial oxidative damage // J. Biol. Chem. - 2001. - V. 276. - P. 38084–38089. 451. Sundbom M., Kaiser C., Björkstrand E., Castro V.M., Larsson C., Selén G., Nyhem C.S., James S.R. Inhibition of 11βHSD1 with the S- phenylethylaminothiazolone BVT116429 increases adiponectin concentrations and improves glucose homeostasis in diabetic KKAy mice // BMC Pharmacology. 2008. - V. 8. - P. 3-12. 452. Sunde R.A. Selenium. In Present Knowledge in Nutrition, 9th ed.; Bowman B.A., Russell R.M., Eds.; ILSI Press Inc: Washington, DC, USA, 2006. - P. 480–497. 453. Sunderman F.W.J.; Barber A.M. Finger-loops, oncogenes, and metals. Claude Passmore Brown Memorial Lecture // Ann. Clin. Lab. Sci. 1988. V. 18. P. 267– 288. 454. Suzuki K.T., Sasakura C., Yoneda S. Binding sites for the (Hg-Se) complex on selenoprotein P // Biochem. Biophys. Acta. - 1998. - V. 1429. - P. 102–112. 455. Szumlanski C., Otterness D., Her C., Lee D., Brandriff B., Kelsell D., Spurr N., Lennard L., Wieben E., Weinshilboum R.M. Thiopurinemethyl transferase pharmacogenetics: human gene cloning and characterization of a common polymorphism // DNA Cell Biol. - 1996. - V. 15. - P. 17–30. 456. Takahashi M. Oxidative stress and redox regulation on in vitro development of mammalian embryos // J. Reprod. Dev. - 2012. - V. 58(1). - P. 1-9. 311 457. Takahashi K., Newburger P.E., Cohen H.J. Glutathione peroxidase protein // J. Clin. Invest. - 1986. - V. 77. - P. 1402-1404. 458. Tamura T., StadtmanT.C. A new selenoprotein from human lung adenocarcinoma cells: purifica-tion, properties, and thioredoxin reductase activity // Proc. Natl. Acad. Sci. USA. - 1996. – V. 93. - N 3. - P.1006-1011. 459. Tappel A.L. Selenium-glutathione peroxidase: properties and synthesis // Curr. Top Cell Regul. - 1984. - V. 24. - P.87-97. 460. Tappel A.L. Selenium-glutathione peroxidase and vitamin E // Am. J. Clin. Nutr. - 1974. - V. 27. - P.960-965. 461. Taverne Y.J.H.J., Bogers A.J.J.C., Duncker D.J. Merkus D. Reactive Oxygen Species and the Cardiovascular System // Oxidative Medicine and Cellular Longevity. V. 2013. Article ID 862423. - 15 p. http://dx.doi.org/10.1155/2013/862423. 462. Taylor E.R., Hurrell F., Shannon R.J., Lin T.K., Hirst J., Murphy M.P. Reversible glutathionylation of complex I increases mitochondrial superoxide formation // J. Biol. Chem. - 2003. - V. 278. - P. 19603–19610. 463. Templeton D.M., Liu Y. Multiple roles of cadmium in cell death and survival // Chem. Biol. Interact. - 2010. - V. 188. - P. 267–275. 464. Thannickal V.J., Fanburg B.L. Reactive oxygen species in cell signaling // Am. J. Physiol. Lung Cell. Mol. Physiol. - 2000. - V. 279. - P. L1005–L1028. 465. Thevenod F. Cadmium and cellular signaling cascades: To be or not to be? // Toxicol. Appl. Pharmacol. - 2009. - V. 238. - P. 221–239. 466. Thijssen S., Cuypers A., Maringwa J., Smeets K., Horemans N., Lambrichts I., Van Kerkhove E. Low cadmium exposure triggers a biphasic oxidative stress response in mice kidneys // Toxicology. - 2007. - V. 236. - P. 29–41. 467. Tomaso G.D., Fedi S., Carnevali M., Manegatti M., Taddei C., and Zannoni D. The membrane-bound respiratory chain of Pseudomonas pseudoalcaligenes KF707 cells grown in the presence or absence of potassium tellurite // Microbiology. 2002. - V. 148. - Р. 1699–1708. 312 468. Trachootham D., Lu W., Ogasawara M.A., Nilsa R.D., Huang P. Redox regulation of cell survival // Antioxid. Redox Signal. - 2008. - V. 10. - P. 1343– 1374. 469. Tremaroli V., Stefano F., Zannoni D. Evidence for a tellurite-dependent generation of reactive oxygen species and absence of a tellurite-mediated adaptive response to oxidative stress of cells of Pseudomonas pseudoalcaligenes KF707. Arch. Microbiol. - 2007. - V. 187(2). - P. 127-135. 470. Tremaroli V., Workentine M.L., Weljie A.M., Vogel H.J., Ceri H., Viti C., Tatti E., Zhang P., Hynes A.P., Turner R.J., Zannoni D. Metabolomic Investigation of the Bacterial Response to a Metal Challenge // Appl. Environ. Microbiol. 2009. - V. 75(3). - P. 719-728. 471. Тrutko S.M., Akimenko V. K., Suzina N.E., Anisimova L.A., Shlyapnikov M.G., Baskunov B.P., Duda V.I., Boronin A.M. Involvement of the respiratory chain of gram-negative bacteria in the reduction of tellurite // Arch. Microbiol. 2000. - V. 173. - P. 178-186. 472. Tu B.P., Kudlicki A., Rowicka M., McKnight S.L. Logic of the yeast metabolic cycle: temporal compartmentalization of cellular processes // Science. 2005. - V. 310. - P. 1152–1158. 473. Turner R.J., Aharonowitz Y., Weiner J., Taylor D.E. Glutathione is a target in tellurite toxicity and is protected by tellurite resistance determinants in Escherichia coli // Can. J. Microbiol. - 2001. - V. 47. - P. 33–40. 474. Turner M.S., Lo R., Giffard P.M. Inhibition of Staphylococcus aureus Growth on Tellurite-Containing Media by Lactobacillus reuteri Is Dependent on CyuC and Thiol Production // Appl. Environ. Microbiol. - 2007. - V. 73(3). - P. 1005-1009. 475. Turner R., Weiner J., Taylor D. Tellurite-mediated thiol oxidation in Escherichia coli // Microbiology. - 1999. - V. 145. - P. 2549–2557. 476. Turrens J.F. Mitochondrial formation of reactive oxygen species // J. Physiol. (Lond.). - 2003. - V. 552. - P. 335-344. 313 477. Tweeddale H., Notley-McRobb L., Ferenci T. Assessing the effect of reactive oxygen species on Escherichia coli using a metabolome approach // Redox Rep. 1999. - V. 4. - P. 237–241. 478. Uemura M., Manabe H., Yoshida N., Fujita N., Ochiai J., Matsumoto N., Takagi T., Naito Y., Yoshikawa T. Alpha-tocopherol prevents apoptosis of vascular endothelial cells via a mechanism exceeding that of mere antioxidation // Eur. J. Pharmacol. - 2002. - V. 456. - P. 29-37. 479. Ufer C., Wang C.C. The roles of glutathione peroxidases during embryo development // Molecular neuroscience review article. - 2011. - V. 4. - P. 1-14. doi: 10.3389/fnmol.2011.00012. 480. Usuki F., Yamashita A., Fujimura M. Post-transcriptional Defects of Antioxidant Selenoenzymes Cause Oxidative Stress under Methylmercury Exposure // The journal of biological chemistry. - 2011. - V. 286(8). - P. 6641– 6649. 481. Valko M., Izakovic M., Mazur M., Rhodes C.J., Telser J. Role of oxygen radicals in DNA damage and cancer incidence // Mol. Cell Biochem. - 2004. V. 266. - P. 37-56. 482. Valko M., Leibfritz D., Moncol J., Cronin M.T., Mazur M., Telser J. Free radicals and antioxidants in normal physiological functions and human disease // Int. J. Biochem. Cell Biol. - 2007. - V. 39. - P.44-84. 483. Valko M., Rhodes C.J., Moncol J., Izakovic M., Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer // Chem. Biol. Interact. 2006. - V. 160. - P. 1-40. 484. Varotto L., Domeneghetti S., Rosani U., Manfrin C., Cajaraville M.P., Raccanelli S., Pallavicini A., Venier P. DNA Damage and Transcriptional Changes in the Gills of Mytilus galloprovincialis Exposed to Nanomolar Doses of Combined Metal Salts (Cd, Cu, Hg) // PLoS ONE. - 2013. - 8(1). e54602. doi:10.1371/journal.pone.0054602. 485. Venkataraman S., Jiang X., Weydert C.J., Zhang Y., Zhang H.J., Goswami P.C., Ritchie J.M., Oberley L.W., Buettner G.R. Manganese superoxide dismutase 314 overexpression inhibits the growth of androgen-independent prostate cancer cells // Oncogene. - 2005. - V. 24. - P. 77–89. 486. Venkataraman S., Wagner B.A., Jiang X., Wang H.P., Schafer F.Q., Ritchie J.M., Burns C.P., Oberley L.W., Buettner G.R. Overexpression of manganese superoxide dismutase promotes the survival of prostate cancer cells exposed to hyperthermia // Free Rad. Res. - 2004. - V. 38. - P. 1119–1132. 487. Verougstraetea V., Lisona D., Hotzb P. Cadmium, lung and prostate cancer: a systematic review of recent epidemiological data // J. Toxicol. Environ. Health. 2003. - V. 6. - P. 227-255. 488. Vila G., Krebs M., Riedl M., Baumgartner-Parzer S.M., Clodi M., Maier C., Pacini G., Luger A. Acute effects of hydrocortisone on the metabolic response to a glucose load: increase in the first-phase insulin secretion // Eur. J. Endocrinol. 2010. - V. 163(2). - P. 225-231. doi: 10.1530/EJE-10-0282. 489. Waalkes M.P., Misra R.R. Cadmium carcinogenicity and genotoxicity / In Toxicology of Metals // Chang L. Ed. CRC Press: Boca Raton, FL, USA, 1996. P. 231-244. 490. Wadaan M.A.M. Effects of mercury exposure on blood chemistry and liver histopathology of male rats // Journal of Pharmacology and Toxicology. - 2009. V. 4(3). - P. 126–131. 491. Wagener F.A.D.T.G., Carels C.E., Lundvig D.M.S. Targeting the Redox Balance in Inflammatory Skin Conditions // Int. J. Mol. Sci. - 2013. - V. 14. - P. 9126-9167. doi:10.3390/ijms14059126. 492. Waisberg M., Joseph P., Hale B., Beyersmann D. Molecular and cellular mechanisms of cadmium carcinogenesis // Toxicology. - 2003. - V. 192. - P. 95– 117. 493. Wang L., Wang H., Hu M., Cao J., Chen D., Liu Z. Oxidative stress and apoptotic changes in primary cultures of rat proximal tubular cells exposed to lead // Archives of Toxicology. - 2009. - V. 83(5). - P. 417–427. 315 494. Wang Y., Fang J., Leonard S.S., Rao K.M. Cadmium inhibits the electron transfer chain and induces reactive oxygen species / Free Radic. Biol. Med. - 2004. - V. 36. - P. 1434–1443. 495. Wang Z., Wu Z., Jian J., Lu Y. Cloning and expression of heat shock protein 70 gene in the haemocytes of pearl oyster (Pinctada fucata, Gould 1850) responding to bacterial challenge // Fish Shellfish Immunol. - 2009. - V. 26. - P. 639–645. 496. Watanabe M., Henmi K., Ogawa K., Suzuki T. Cadmium-dependent generation of reactive oxygen species andmitochondrial DNA breaks in photosynthetic and non-photosynthetic strains of Euglena gracilis // Comparative Biochemistry and Physiology C. - 2003. - V. 134(2). - P. 227–234. 497. van de Waterbeemd B, Zomer G, van den IJssel J, van Keulen L, Eppink MH, van der Ley P., van der Pol L.A. Cysteine Depletion Causes Oxidative Stress and Triggers Outer Membrane Vesicle Release by Neisseria meningitidis; Implications for Vaccine Development // PLoS ONE. - 2013. - V. 8(1). 9 p. e54314. doi:10.1371/journal.pone.0054314. 498. Weekley C.M., Aitken J.B., Finney L., Vogt S., Witting P.K., Harris H.H. Selenium Metabolism in Cancer Cells: The Combined Application of XAS and XFM Techniques to the Problem of Selenium Speciation in Biological Systems // Nutrients. - 2013. - V. 5. - P. 1734-1756. doi:10.3390/nu5051734. 499. Weisiger R.A., Fridovich I. Superoxide dismutase. Organelle specificity // J. Biol. Chem. - 1973. - V. 248. - P. 3582–3592. 500. Weitzel F., Ursini F., Wendel A. Phospholipid hydroperoxide glutathione peroxidase in various mouse organs during selenium deficiency and repletion // Biochim. Biophys. Acta. - 1990. - V. 1036. - P. 88–94. 501. Wilcke J.R., Davis L.E. Review of glucocorticoid pharmacology // Vet. Clin. North Am. Small Anim. Pract. - 1982. - V. 12. - P. 3–17. 502. Wilcox D.E., Schenk A.D., Feldman B.M., Xu Y. Oxidation of zinc-binding cysteine residues in transcription factor proteins // Antioxid. Redox Signal. - 2001. - V. 3. - P. 549–564. 316 503. Wilcox J.K., Ash S.L., Catignani G.L. Antioxidants and prevention of chronic disease // Crit. Rev. Food Sci. Nutr. 2004. - V. 44. - P. 275-295. 504. Wingert R.A., Galloway J.L., Barut B., Foott H., Fraenkel P., Axe J.L., Weber G.J., Dooley K., Davidson A.J., Schmid B., Paw B.H., Shaw G.C., Kingsley P., Palis J., Schubert H., Chen O., Kaplan J., Zon L.I. Deficiency of glutaredoxin 5 reveals Fe-S clusters are required for vertebrate haem synthesis // Nature. - 2005. V. 436. - P. 1035–1039. 505. Witkiewicz-Kucharczyk A., Bal W. Damage of zinc fngers in DNA repair proteins, a novel molecular mechanism in carcinogenesis // Toxicol. Lett. - 2005. V. 162. - P. 29-42. 506. Wójtowicz H., Kloc K., Maliszewska I., Młochowski J. Azaanalogues of ebselen as antimicrobial and antiviral agents: synthesis and properties // Farmaco. 2004. - V. 59. - N 11. - P. 863-868. 507. Woolfolk C.A., Whitely H.R. Reduction of inorganic compoundswithmolecular hydrogen byMicrococcus lactilyticus. I. Stoichiometry with compounds of arsenic, selenium, tellurium, transition and other elements // J. Bacteriol. - 1962. - V. 84. - P. 647-658. 508. Yabaluri N., Bashyam M.D. Hormonal regulation of gluconeogenic gene transcription in the liver // J. Biosci. - 2010. - V. 35(3). - P. 473-484. 509. Yamashita M., Yamashita Y., Suzuki T., Kani Y., Mizusawa N., Imamura S., Takemoto K., Hara T., Hossain Md. A., Yabu T., Touhata K. Selenoneine, a Novel Selenium-Containing Compound, Mediates Detoxification Mechanisms against Methylmercury Accumulation and Toxicity in Zebrafish Embryo // Mar. Biotechnol. - 2013. - V. 15. - P. 559–570. 510. Yamashita Y., Amlund H., Suzuki T., Hara T., Hossain A.M., Yabu T., Touhata K., Yamashita M. Selenoneine, total selenium, and total mercury content in the muscle of fishes // Fish Sci. - 2011. - V. 77. - P. 679–686. 511. Yamashita Y., Yabu T., Yamashita M. Discovery of the strong antioxidant selenoneine in tuna and selenium redox metabolism // World J. Biol. Chem. 2010. - V. 1. - P. 144–150. 317 512. Yamashita Y., Yamashita M. Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna // J Biol Chem. - 2010. - V. 285. - P. 18134–18138. 513. Yang L.Y., Wu K.H., Chiu W.T., Wang S.H., Shih C.M. Тhe cadmiuminduced death of mesangial cells results in nephrotoxicity // Autophagy. - 2009. V. 5(4). - P. 571–572. 514. Yeligar S.M., Harris F.L., Hart C. M., Brown L.A.S. Ethanol induces oxidative stress in alveolar macrophages via upregulation of NADPH oxidases // J Immunol. 2012. V. 188(8). P. 3648–3657. doi:10.4049/jimmunol.1101278. 515. Yin Z., Lee E., Ni M., Jiang H., Milatovic D., Rongzhu L., Farina M., Rocha J.B.T., Aschner M. Methylmercury-Induced Alterations in Astrocyte Function are Attenuated by Ebselen // Neurotoxicology. - 2011. - V. 32(3). - P. 291–299. doi:10.1016/j.neuro.2011.01.004. 516. Youn B.W., Fiala E.S., Sohn O.S. Mechanisms of organoselenium compounds in chemoprevention: effects on transcription factor-DNA binding // Nutr. Cancer. 2001. - V. 40. - P. 28–33. 517. Yoon J.C., Puigserver P., Chen G., Donovan J., Wu Z., Rhee J., Adelmant G., Stafford J., Kahn C.R., Granner D.K., Newgard C.B., Spiegelman B.M. Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1 // Nature. 2001. - V. 413. - P. 131–138. 518. Young J. C., Agashe V. R., Siegers K., Hartl F. U. Pathways of chaperonemediated protein folding in the cytosol // Nature Reviews Molecular Cell Biology. - 2004. - V. 5(10). - P. 781–791. 519. Young I.S., McEneny J. Lipoprotein oxidation and atherosclerosis // Biochem. Soc. Trans. - 2001. - V. 29. - P. 358-362. 520. Zannoni D., Borsetti F., Harrison J.J., Turner R.J. The bacterial response to the chalcogen metalloids Se and Te // Adv. Microb. Physiol. 2008. V. 53. P. 1–72. 521. Zelko I.N., Mariani T.J., Folz R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) 318 gene structures, evolution, and expression // Free Radic. Biol. Med. - 2002. - V. 33. - P. 337–349. 522. Zeng H. Selenium as an essential micronutrient: Roles in cell cycle and apoptosis // Molecules. - 2009. - V. 14. - P. 1263–1278. 523. Zeng H., Cao J.J., Combs G.F.J. Selenium in Bone Health: Roles in Antioxidant Protection and Cell Proliferation // Nutrients. - 2013. - V. 5. - P. 97110. doi:10.3390/nu5010097. 524. Zhang Q., Chen L., Guo K., Zheng L., Liu B., Yu W., Guo C., Liu Z., Chen Y., Tang Z. Effects of different selenium levels on gene expression of a subset of selenoproteins and antioxidative capacity in mice // Biol. Trace Elem. Res. - 2013. - V. 154(2). - P. 255-61. doi: 10.1007/s12011-013-9710-z. 525. Zhang L., Maiorino M., Roveri A., Ursini F. Phospholipid hydroperoxide glutathione peroxidase: specific activity in tissues of rats of different age and comparison with other glutathione peroxidases // Biochim. Biophys. Acta. - 1989. V. 1006. - P.140-143. 526. Zhang H.J., Zhao W., Venkataraman S., Robbins M.E., Buettner G.R., Kregel K.C., Oberley L.W. Activation of matrix metalloproteinase-2 by overexpression of manganese superoxide dismutase in human breast cancer MCF-7 cells involves reactive oxygen species // J. Biol. Chem. - 2002. - V. 277(23). - P. 20919–20926. 527. Zhong L., Arner E.S.J., Holmgren A. Structure and mechanism of mammalian thioredoxin reductase: the active site is a redox-active selenolthiol/selenenylsulfide formed from the conserved cysteine-selenocysteine sequence // Proc. Natl. Acad. Sci. USA. - 2000. - V. 97. - P. 5854–5859. 528. Zhou Y.J., Zhang S.P., Liu C.W., Cai Y.Q. The protection of selenium on ROS mediated-apoptosis by mitochondria dysfunction in cadmium-induced LLC-PK(1) cells // Toxicol. In Vitro. - 2009. - V. 23. - P. 288–294.