КРЕВЕТКИ-ФУНХАЛИИ (DECAPODA, NATANTIA, PENAEIDAE
реклама

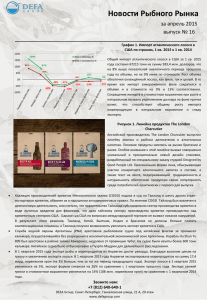
- v. :. ЬЮЛ. МОСК. с-у; .;•••• О-ВА ИСПЫТАТЕЛЕН ПРИРОДЫ. . " ' . A... : . ОТД. ВИОЛ. 1991. Т. 96. ВЫП. 6 У Д К 59 КРЕВЕТКИ-ФУНХАЛИИ (DECAPODA, NATANTIA, PENAEIDAE): СИСТЕМАТИКА, РАСПРОСТРАНЕНИЕ И НЕКОТОРЫЕ ВОПРОСЫ БИОЛОГИИ Р. И. Буруковский, Л. Л. Роменский Креветки-фунхалии (Funchalia s. lato) — м а л е н ь к а я группа пенеидных креветок из пяти видов, широко распространенных в Мировом океане. Информация о них ограничена фаунистическими данными, рассеянными по отдельным работам. Наличие онтогенетической изменчивости таксономических признаков (печеночный шип, редуцирующийся у взрослых особей некоторых видов фунхалий, изменение жаберной формулы) привело к тому, что существуют описания неверно идентифицированных ювенильных особей ( R a m a d a n , 1938; Paulinose, 1974) и д а ж е ошибочно описанных новых видов ( F u j i n o , 1975). Точное определение большинства видов фунхалий пока возможно лишь по особям со сформированными гениталиями. Долгое время оставалось неопределенным само положение Funchalia s. lato в системе пенеидных креветок. Их то с б л и ж а л и с подсемейством Solenocerinae (Bouvier, 1908), то выделяли в отдельную серию F u n c h a l i a e (Balss, 1925), а позднее включили в группу Р е п а е е из подсемейства P e n a e i n a e (Balss, 1925; B u r k e n r o a d , 1936) из-за сходства ж а б е р н ы х формул. В настоящее время фунхалии, подразделенные на два рода ( P e l a g o p e n a e u s Faxon, 1893 и Funchalia Johnson, 1867), остаются в подсемействе P e n a e i n a e (Grippa, 1987). Однако они резко отличаются от всех других пенеидных креветок строением мандибул, переднего отдела ж е л у д к а , яичников и редукцией борозд на карапаксе. На наш взгляд, это дает основание для их выделения в отдельное подсемейство. Отсюда цель нашей работы — уточнение систематического положения группы, создание ее определителя и краткой сводки данных по распространению и некоторым вопросам биологии наиболее массовых видов. МАТЕРИАЛ И МЕТОДИКА М а т е р и а л собирали в 1974—1988 гг. в различных р а й о н а х Атлантического (Северо-Атлантическая конвергенция — Н И С «Профессор Седлецкий», Тропическая Восточная Атлантика — Н И С «Фиолент»; Ю г о - В о с т о ч н а я А т л а н т и к а — рейсы 9 поисковых и исследовательских с у д о в ) , Индийского ( Н И С «Академик К у р ч а т о в » и « П р о ф е с с о р Водяницкий») и Тихого ( С Р Т М 8081) океанов. Количество и места сбора проб приводятся н и ж е при х а р а к т е р и с т и к е отдельных видов. Фунхалий измеряли от основания глазного стебелька до конца тельсона (общ а я длина — О Д ) с точностью до 1 мм. Состояние репродуктивной системы у д в у х наиболее изученных видов оценивали по ш к а л е стадий зрелости, в основу которой п о л о ж е н ы изменения цвета и относительных р а з м е р о в г о н а д ы у с в е ж е п о й м а н н ы х самок и сперматофорных ампул — у самцов. С а м к и. I с т а д и я . Гонада тоненькая, почти п р о з р а ч н а я . Л е ж и т в задней части голоиогруди. П л о х о отличима от других органов. II с т а д и я . Г о н а д а п о л у п р о з р а ч н а я , медово- или я н т а р н о - ж е л т а я , покрытая красными звездочками х р о м а т о ф о р о в — у F. villosa, м у т н о в а т о - б е л а я или с е р а я у F. woodwardi. Л е ж и т в задней части головогруди, о б н и м а я передними р о г а м и ж е л у д о к с боков. 60 III с т а д и я . Гонада с боков п р и к р ы в а е т передними рогами почти весь ж е л у док, средние рога п р и к р ы в а ю т его з а д н ю ю треть. Ц в е т гонады л и л о в а т о - р о з о в ы й у F. villosa, серо-голубой у F. woodwardi. IV с т а д и я . Гонада з а к р ы в а е т почти весь ж е л у д о к с боков и сверху. П е р е д н я я его часть видна. Ц в е т г о н а д ы у F. villosa л и л о в а т о - к о р и ч н е в ы й или коричневый, а у F. woodwardi — синий или сине-фиолетовый. V с т а д и я . П е р е д н и е рога полностью з а к р ы в а ю т ж е л у д о к сверху и с боков, В абдомен з а д н и е рога не з а х о д я т . Ц в е т г о н а д ы у F. villosa — темно-коричневый до вишневого, а у F. woodwardi—черно-фиолетовый. С а м ц ы ( р а з р а б о т а н а на примере /•'. villosa). I стадия. П е т а з м а не с р о с ш а я с я , сперматофорные ампулы в основаниях 5 переопод не различимы. II стадия. П е т а з м а с р о с ш а я с я . А м п у л ы не видны или еле просвечивают сквозь покровы. III стадия. П е т а з м а с р о с ш а я с я . Ампулы в виде розовой вертикальной черточки, просвечивающей сквозь покровы. IV стадия. Ампулы в виде в ы т я н у т ы х к р а с н о в а т о - р о з о в ы х пятен; из половых отверстий могут т о р ч а т ь с п е р м а т о ф о р ы . ТАКСОНОМИЧЕСКАЯ ХАРАКТЕРИСТИКА Подсемейство Funchaliinae subfam. nova Рострум с ж а т с боков, короткий, не достигающий дистальных концов скафоцеритов, вооружен сверху и снизу или только сверху. Тело опушенное или неопушенное. Мандибулы с полностью редуцированным жевательным отростком. Р е ж у щ и й отросток в виде острого шипа, который у большинства видов вытянут в серповидно-изогнутое острие. Мандибулярные пальпы двучленистые. На вторых м а к с и л л а х три зндита. Переоподы укороченные; вытянутые вперед, не заходят за дистальный край карпоцерита. Б а з и с ы и искиумы двух первых переопод вооружены шипами. Н а к а р а п а к с е полностью отсутствуют цервикальная, печеночная и адростральные борозды и соответствующие им кили, но имеются более или менее в ы р а ж е н н ы е разветвляющиеся продольные л а т е р а л ь н ы е кили, закругленные сверху. Они тянутся от печеночной области к а р а п а к са до его заднего края. Плевры 1-го абдоминального сегмента расширены н а з а д и прикрывают переднюю часть плевр 2-го сегмента. На 6-м сегменте абдомена — л а т е р а л ь н ы е кили. Тельсон заканчивается острием, вооружен тремя п а р а м и подвижных субдистальных шипов. Теликум у самок в виде углубления, прикрытого с боков, а у некоторых и сверху более или менее развитыми к л а п а н а м и . П е т а з м а симметричная или асимметричная. Передний отдел ж е л у д к а , в отличие от всех остальных пенеидных креветок, представляет собою тонкостенный мешок, заполненный жидкостью у только что пойманных креветок. З а д н и е рога яичника недоразвиты, не заходят в абдомен, или, если заходят, то лишь в переднюю часть 1-го сегмента. Передние и боковые рога чрезвычайно в а р и а б е л ь ны по строению. Типовой род Funchalia Johnson, 1867. КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ 1. Тело не опушенное. На роструме имеются вентральные зубцы. Антеннальный шип отсутствует. Д и с т а л ь н ы й членик мандибулярных пальп симметричный. Р е ж у щ и й отросток мандибул короткий. Экзоподит уропод с диэрезисом. П е т а з м а симметричная, теликум с большими клапанами, прикрывающими углубление семеприемника сверху. 61 Pelagopenaeus Burkenroad, 1934. Единственный вид P. balboae ( F a x o n , 1893). — Тело опушенное. Рострум вооружен только сверху. Антеннальный шип имеется, дистальный членик мандибулярных пальп асимметричный. Р е ж у щ и й отросток мандибул длинный. Экзоподит уропод без диэрезиса. П е т а з м а асимметричная, боковые к л а п а н ы теликума не прикрывают сверху углубление семеприемника. Funchalia Johnson, 1867. КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА FUNCHALIA 1. У взрослых особей имеется печеночный шип; более 10 ростральных зубцов 2 — У взрослых особей печеночный шип отсутствует; менее 10 ростральных зубцов . 3 2. 6-й сегмент абдомена с коротким килем, расположенным ниже и параллельно длинному среднелатеральному килю; теликум открытый; дистальная часть длинной ветви петазмы не раздвоена, ложковидная. F. woodwardi J o n h s o n , 1867. — 6-й сегмент абдомена лишь с длинным среднелатеральным килем; теликум полуприкрыт боковыми стенками, образующими нечто вроде клапанов; дистальная часть длинной ветви петазмы раздвоена. F. danae B u r k e n r o a d , 1940. 3. П е т а з м а с маленьким треугольным выступом на вентральной поверхности свободной дистальной части длинной ветви петазмы; теликум с маленьким срединным гребнем позади семеприемника. F. villosa (Bouvier, 1905). — П е т а з м а без выступа на вентральной поверхности свободной дистальной части более длинной ветви петазмы; теликум с большим зубообразным выступом позади семеприемника. F. taaningi B u r k e n r o a d , 1940. Характеристика видов Pelagopenaeus balboae (Faxon, 1893) (рис. 1, Л) Атлантический океан: 21.09.82, 23.50, 30°25' с. ш., 28° 18' з. д. (банка Метеор), 1 ?; 7.10.82, 22.30, 6°37 / с. ш., 2 Г 5 3 / з. д. (возвышенность Сьерра-Леоне), 1 ? (лов на поверхности воды накидной сетью); 0°13' ю. ш., ISMS 7 з. д., лов ночью на поверхности: 1 О Д 81 мм, 1 2 О Д 94 мм, со сперматофором; 27.10.81, 5.35—7.40, 7°56' ю. ш., 0°54 / в. д., глубина места 580 м, горизонт лова 580—540 м, 1 <3 О Д 86 мм. Индийский океан: 24.07.78, 19.20—19.50, 7°07' с. ш„ 89 с 29' в. д., поверхностный лов, 1 о О Д 95 мм; 22.07.78, 7°09 / с. ш., 8 7 ° 1 Г в. д., глубина места 3680 м, трал Айзекса—Кидда, горизонт 80 м, 1 з О Д 102 мм. Тихий океан: Целебесское море у о. Минданао, 2 ?, поверхностный лов ночыо; 27.05.81, 10°23 / ю. ш., 82°24' з. д., над глубиной 4100 м, горизонт 30—6 м, 1 ? (преднерестовая) О Д 116 мм. Г е о г р а ф и ч е с к о е р а с п р о с т р а н е н и е . Атлантический океан: у Бермудских, Антильских и Канарских о-вов, Конго, а т а к ж е 3 0 ° 2 Г ю. ш., 14°02 / з. д.; Индийский океан: Аравийское море, Тихий — Панамский залив и у о. Кокос. Ловится ночью у поверхности над глубинами более 1500 м (Burkenroad, 1936; R a m a d a n , 1938; Crosnier, Fo62 J 3 Рис. 1. Совокупительные органы креветок подсемейства F u n c h a l i i n a e : А — Pelagope пае us balboae (1 — петазма, 2 — т е л и к у м ) ; Б — Funchalia taaningi, В — F. danae, Г — F. villosa, Д — F. woodwardi (1 — п е т а з м а , о б щ и й вид, 2 — д и с т а л ь н а я часть петазмы, 3 — т е л и к у м ) . М а с ш т а б : А — 1 и 2, В — 1 \к 3, Г — 1 и 3, Д — 1, 2 и 3 — 1 мм, прочие — 0,5 мм rest, 1973). Н а ш и материалы в основном подтверждают циркумтропическое распространение этого вида, в Атлантике достигающего субтропиков и на севере, и на юге (30°20 / с. и ю. ш.). Биологическая характеристика. Все исследованные нами особи пойманы только в ночное время на поверхности воды. Самки находились в нагульном состоянии, за исключением наиболее крупной, зрелой. В ж е л у д к а х двух особей найдены крупная полупереварен63 ная нематода, крючья к а л ь м а р а из сем. Enoploteuthidae, остатки рыбы (диаметр хрусталика глаза 0,5 мм) и 4 копеподы длиной 1 мм. Funchalia taaningi Burkenroad, 1940 (рис. 1 , 5 ) , Индийский океан: 23.06.78, 00—01, 0 Г 0 0 ' с. ш., 55°00' в. д., горизонт 80—30 м, трал Айзекса — Кидда, 1 О Д 33 мм. Географическое распространение. Индийский океан: М а д а г а с к а р (22°07 / с. ш., 43°05 / в. д.), несколько севернее экватора ( Г 0 6 ' — Г 0 0 / с. ш., 55—62°05 / в. д.); Тихий океан: в западной части — хребет Кюсю-Палау, в центральной части от 33° 16' с. ш., до 0°14 / ю . ш . и от 99°35' в. д. до 139°39 / з. д.), наиболее многочислен в уловах чуть северней экватора (0°04—0°2Г с. ш., 139°39'—142°28' з. д.) (Burkenroad, 1940; Crosnier, 1985; Baba et al., 1986; W a s m e r , 1989). Вертикальное распределение. Известен в основном из косых ловов незакрывающимися орудиями лова. Так, в Тихом океане из 19 ловов, где был встречен этот вид, 12 выполнены ночью с глубин 300—100 м, известна одна поимка с глубины 85 м. Д н е м обнаружен в уловах с глубин 2600—440 м (в основном 730—440 м) (Wasmer, 1989). По данным Т. Kikuchi, М. Omori (1985), в северо-западной части Тихого океана ночью встречен на глубинах 240—594 м, днем — 483—678 м. Найден в ж е л у д к е Beryx splendens, пойманного на глубине 355 м (Baba et al., 1986). F. taaningi можно считать активным мигрантом, перемещающимся от нижней эпипелагиали ночью до нижней мезопелагиали днем. Funchalia danae Burkenroad, 1940 (рис. 1, В) Атлантический океан: 25.02.76, 10°44 / ю. ш„ 13°05' в. д., 1200— 1160 м, донный трал, 12 (преднерестовая); 3.11.76, 3°28' ю. ш., 5°47 / в. д., из ж е л у д к а Thunnus obesus, 1 с? и 1 ?; 11.06.78, 1 Г 2 8 ' ю. ш., 01°14' в. д., 600—800 м, 2 ? О Д 76 и 64 мм; 11.05.80, 20.40—21.10, 8°00' ю. ш., Г 0 7 / в. д., 490—510 м, 1 О Д 65 мм; 22.01.81, 23.30—24.00, 04°47' с. ш., 9°38' з. д., 798—785 м, 1 экз.; 15.07.87, 05°31' ю. ш„ 0 Г 1 Г в. д., 1 экз.; 27.01.81, 00—00.30, 4°26,8 / с. ш., 9°13,8' з. д., 807—835 м, 2 & 78 и 74 мм. Географическое распространение. В Атлантическом океане известна в треугольнике: Азорские о-ва — о. Св. Елены — Конго (Crosnier, Forest, 1973). Н а ш и материалы собраны в том ж е треугольнике, в основном, над материковым склоном или подводными возвышеннностями в ночное время. Креветки пойманы в косых или горизонтальных ловах незакрывающимися орудиями, но ни разу непосредственно на поверхности. Видимо, среднеглубинный (эпи-мезопелагический) вид, так как кроме Th. obesus им питается т а к ж е Alepisaurus ferox (паши данные), т. е. рыбы, обитающие преимущественно з нижней эпи- и верхней мезопелагиали. Вид известен т а к ж е из Индийского океана: Аравийское море и 16°08 / ю. ш., 97° 14' в. д. (Crosnier, Forest, 1973). Funchalia villosa (Bouvier, 1905) (рис. 1, Г) Атлантический океан: 31.01.74, 38° 19' с. ш., З Г О К з. д., косой лов с глубины 1200 м, Юс?, 4 5; 2.08.81, 21.00—22.15, 40°33' ю. ш., 10°50' в. д., глубина 5200 м, горизонт лова 0—88 м, 4 экз.; 29.07—25.09.84, ночные ловы 43°58 / —37°53' с. ш., 14—18° з. д., над глубинами 1500— 3000 м, по горизонтам 35—110 м, 278 с? и 2 0 7 ? ; 26.10—05.11.84, дневные и ночные ловы, 36—31° с. ш., 29°—37°02 / з. д., над глубинами 2500—4100 м, косые ловы с 500—800 м, 29 экз. Географическое распространение. Вид известен из Северо-Восточной Атлантики (Burkenroad, 1936), Карибского моря и 64 Мексиканского залива (Grippa, 1987), из района Тристан-да-Кунья (Crosnier, Forest, 1973; Grippa, 1987) и банки Вальдивия (24°09'— 26° 18х ю. т . , 06°25 7 —07° в. д.: Macpherson, 1984), Средиземного моря (Grippa, 1987); Индийского океана (29°52' ю. т . , 34°32' в. д.: Crosnier, 1985; 20°02' ю. т . , 99°41' в. д.: W a s m e r , 1989); Тихого океана юго-западнее о. Лорд-Хау (Dall, 1957; Kensley et al., 1987) и Тасманова моря (Wasmer, 1989). В августе-—сентябре 1984 г. ( Н И С «Профессор Седлецкий») севернее Азорских о-вов F. villosa встречалась к югу от линии 46° с. ш., 29° з. д. — 38° с. т . , 14° з. д. с частотой, близкой к 100%, и была приурочена к зоне субтропической конвергенции. Все другие находки вида в Северной Атлантике т а к ж е тяготеют к этому району. Кроме того, в наших м а т е р и а л а х имеются особи из зоны южно-атлантической субтропической конвергенции (40°33 / ю. ш., 10°50' в. д.). По-видимому, F. villosa •— североцентральный вид в Атлантике и южно-периферический в трех океанах. В е р т и к а л ь н о е р а с п р е д с л е н и е. P. Foxton (1970) установил, что днем вид держится на глубинах 350—700 м, в основном на 450 м, ночыо поднимается в приповерхностный 50-метровый слой, иногда почти до поверхности. По М. Omori (1974), F. villosa — нижнемезопелагический вид. Н а ш и материалы были собраны из 103 ночных ловов с глубин от поверхности до 1200 м. В поверхностных ловах F. villosa не встречена. На глубинах 35—50 м она составляет более 80% уловов и по численности, и по биомассе всех креветок. Д о л я ее несколько падает в диапазоне 50—100 м и очень сильно уменьшается на больших глубинах. Можно полагать,, что поимки в т р а л а х с глубин более 100 м объясняются попаданием вида в трал при выборке. Следовательно, это скорее никто-эпипелагический, но не поднимающийся па поверхность вид. Б иол огичсск а я х а р а к т е р и с т и к а. Размеры креветок 35—73 мм. Самцы (35—70 мм) несколько меньше самок (44—73 мм). Модальные размеры обоих полов совпадают (57 мм). У самок есть второй пик численности: мода 66 мм (рис. 2, Л ) . В исследованный период времени основная часть креветок — впервые созревающие особи; большинство — в нагульном состоянии (II стадия зрелости гонады). Д а ж е самцы (около 3 0 % ) , имеющие сформированные ампулы, не несут сгтерматофоров. Б ы л и встречены лишь две самки с преднерестовыми гонадами. Самки с продвинутыми гонадами ( I I I — I V стадии) составляют второй, меньший пик размерного состава (рис. 2, Г — Е ) . Если допустить, что F. villosa проводит нагульную часть жизни в водах северо-атлантического антициклонического субтропического круговорота, то нерест ее должен происходить поздней осенью в более южных районах: доля самцов и самок с продвинутыми в развитии репродуктивными системами возрастает с северо-запада на юго-восток. Изучение содержимого около 150 ж е л у д к о в (103 из них было с пищей, а 2 2 — п о л н ы х ) , особенно у только что выловленных особей, наводит па мысль, что процесс пищеварения у этого вида своеобразен. У всех креветок ж е л у д о к раздут, напоминая полупрозрачную ягоду. Он заполнен прозрачной, желтоватого цвета жидкостью, но не жиром, как это часто бывает у P a s i p h a e i d a e (Буруковский, 1986). По консистенции она напоминает глицерин или очень жидкое ж е л е . В ней взвешены отдельные косточки рыб, часто размягченные, ж а б р ы и (или) г л а з а эвфаузиид, черные куски, напоминающие по цвету и консистенции ткани миктофид или гоностомовых, щетинки сагитт, крючья эноплотевтидных } Бюллетень биологии, вып. 0 65 Рис. 2. Биологическая характеристика F. villosa: А — суммарный график размерного состава, Б — стадии зрелости гонад у самок, В — стадии зрелости самцов, Г — размерный состав самцов и самок I стадии зрелости, Д — размерный состав самцов II— III и самок II стадии зрелости, Е — размерный состав самцов IV и самок III—V стадии зрелости, 1 — самцы, 2 — самки кальмаров. С к л а д ы в а е т с я впечатление, что это полупереваренные, разж и ж е н н ы е (в состоянии лизиса) ткани жертв. Видимо, д л я фунхалии характерно пищеварение в полости первого отдела ж е л у д к а . Часто содержимое ж е л у д к а представляло собою один или несколько кусков густого ж е л е янтарного цвета. Иногда вместе с этим ж е л е встречались более или менее оформленные части других ж е р т в (рыбы, эвфаузииды). Н а м и исследованы собранные из этих ж е уловов различные кишечнополостные (медузы, сифонофоры), сальпы и пелагические моллюски (Carinaria sp.). Мы полагаем, что ж е л е — это остатки сальп (из-за сходства консистенции и цвета с их внутренними органами) и Carinaria (из-за найденной в ж е л у д к е характерной р а д у л ы ) . Способ питания фунхалии не позволяет оценить р а з м е р ы жертв, но в одном случае были найдены остатки рыбы длиной как минимум 40—50 мм, почти 70% длины тела креветки. Среди эвфаузиид обнаружены остатки хищной Nyctiphanes sp. Ч а щ е всего в ж е л у д к а х попадались эвфаузииды, гетероподы и рыба. Все остальные объекты питания (табл. 1) встречаются значительно реже. По доле в питании абТаблица Состав пищи F. Содержимое желудков Эвфаузииды Carinaria sp. и сальпы Рыба Фораминиферы Копеподы Икра Головоногие Микропланктон Радиолярии Частота встречаемости, % 36,9 32,1 30,1 9,7 6,8 6,8 3,9 2,9 2,9 Значение в пище, % 9,2 59,1 14,1 — — 4,5 — 4,5 — 1 villosa Содержимое желудков Диатомовые Остракоды Мизиды Кишечнополостные Тинтиниды Полихеты Изоподы-бопириды Жидкий жир Всего ж е л у д к о в Частота встречаемости, % 1,9 1,9 1,0 1,0 1,0 1,0 2,9 16,5 103 Значение в пище. % — — — — 4,5 — 4,1 22 солютно преобладают Carinaria и сальпы (разделить их н е в о з м о ж н о ) , составляющие почти 60% среднего объема пищевого комка. Н а втором месте находятся рыбы, на третьем — эвфаузииды. Остальные компоненты пищевого комка п р и н а д л е ж а т , видимо, случайным ж е р т в а м . М о ж н о предполагать, что F. villosa — н а п а д а ю щ и й хищник, специализированный преимущественно на питании мягкотелыми пелагическими животными. С а м а F. villosa служит объектом питания относительно крупным мезопелагическим хищникам. М ы находили ее в желудках Beryx splendens и к а л ь м а р а Ommastrephes bartrami. Funchalia woodwardi J o h n s o n , 1867 (рис. 1, Д) Атлантический океан: 29.07—19.09.84, 21.35—13.50, 43°52'—36°59' с. ш., 26°59 / —14° з. д., над глубинами 1500—3000 м, ловы на горизонтах 40—1050 м, 6 сГ 74—142 мм, 8 S I—III, 70—142 мм; октябрь—декабрь 1975 г., июль и сентябрь 1981 г., октябрь 1984 г., 17—35° ю. ш., над глубинами 400—2500 м, горизонтальные ловы пелагическими и донными т р а л а м и , 4749 экз.; 31.05.69, 08°29" ю. ш., 12°54 / в. д., горизонт 280 м, 1 <?\ 24.10.85, 08° ю. ш„ 00°56' в. д., 1 сГ 92 мм; 1.10.88, 40°28' ю. ш., 53°02 / з. д., косой лов 125—75 м над глубиной более 3000 м, 1 5 74 мм. Тихий океан: 02.01.83, 40°28 / ю. ш„ 163°47 / з. д., 4 сГ, 5 9; ночной лов в поверхностном слое над абиссальными глубинами. 3* 67 Географическое распространение. В Атлантическом океане вид имеет бипериферическое распространение. Северная часть а р е а л а — от Гебридских (Gordon, Ingle, 1956) до Азорских о-вов и М а д е й р ы (Bouvier, 1908) и в западной части Средиземного моря (Relini-Orsi, Costa, 1975; Grippa, 1987). Ю ж н а я часть а р е а л а — с а м а я северная находка — 8° ю. ш. над хребтом Вавилова и в районе ПуэнтН у а р а (Конго), но в основном от Намибии (20° ю. ш.) до южной оконечности Африки, а т а к ж е вокруг южной и юго-восточной частей континента (Kensley, 1977). Н а ш и находки из нотальной зоны Юго-Западной Атлантики и Тихого океана (см. т а к ж е : Буруковский, П о л е ж а е в , 1985) и находки из района о. Лорд-Хау (35 0 36'—35°39' ю. ш., 150°55'—150°56' в. д.: Kensley et al., 1987) з а с т а в л я ю т предполагать, что ю ж н а я часть а р е а л а вида может быть охарактеризована как циркум-южносубтропическо-нот а л ь н а я . Судя по встречаемости в водах Южной Африки (Буруковский, Роменский, 1985) и по биомассе отдельных агрегаций, достигающей почти 1500 т (Miller et al., 1983), это очень массовый вид здесь. Вертикальное распределение. Встречается мористее изобаты 400 м. В ночное время поднимается к поверхности, днем облавливается у дна на глубинах 400—500 м и в толще воды. В верхней части материкового склона на глубинах 400—500 м встречается с частотой 15,4%, составляя по массе 85% от улова креветок. С глубиной частота встречаемости растет, достигая в уловах с глубин 800—1300 м 62,5%, но биомасса очень м а л а — лишь 0,1% улова (Буруковский, Роменский, 1985). Видимо, активный мигрант, в верхней части материкового склона днем опускающийся на глубины 400—500 м. Биологическая характеристика. Хотя м а т е р и а л ы накапливались в течение многих лет, они, к сожалению, носят разрозненный характер. Их анализ позволяет констатировать следующее. Р а з м е р ы креветок колеблются от 60 до 142 мм, при этом самцы не превышают 130 мм. По м а т е р и а л а м массовых промеров можно судить, что в пелагиали вод Н а м и б и и наиболее мелкие креветки встречаются в ф е в р а л е (мода 70—80 м м ) . В течение года размеры увеличиваются, достигая максимума в ноябре (мода 117 мм, рис. 3, А — 3 ) . В это ж е время над банками Китового хребта обнаружены креветки с модальными р а з м е р а м и 70 мм (рис. 3, И — К ) . Р а з м е р ы первого созревания у самок заметно варьируют: от 64 (район Китового хребта) до 89 мм. Креветки с банок Китового хребта с преднерестовыми гонадами имеют размеры 62—75 мм, а из пелагиали южноафриканских вод — 110—135 мм. Вероятно, нерест приурочен к весенне-летнему периоду, однако неясно, п р и н а д л е ж а т ли креветки с банок Китового хребта и из приконтинентальной пелагиали к одной популяции, будучи разновозрастными особями, или это представители самостоятельных популяций. У 11 самок с гонадами в V стадии зрелости (размер 72—122 мм) подсчитана абсолютная индивидуальная плодовитость: 3000—32000 икринок. Это значительно ниже, чем у других пенеидных креветок сравнимых размеров. У только что пойманной креветки передний отдел ж е л у д к а раздут, как пузырь, и заполнен жидкостью коричневато-янтарного цвета с прож и л к а м и черной слизи, отдельными косточками и чешуйками рыб, а т а к ж е верхними мандибулами (0,5 мм) двух пелагических осьминогов, вероятно, Ocythoe tuberculata (определение М. Липиньского и К. Н. Несиса). П о д а в л я ю щ е е большинство креветок из донных уловов (встречаемость 9 1 % ) имело в ж е л у д к а х обрывки типично донных 68 Рис. 3. Р а з м е р н ы й состав и его д и н а м и к а у F. woodwardi Н а м и б и и (А—3) и Китового хребта (И, К): А — ф е в р а л ь 1984 г. (503 экз.), Б — июнь 1986 г. (359 экз.), В — август 1982 г. (84 экз.), Г —- сентябрь 1981 г. (144 экз.), Д — о к т я б р ь 1984 г. (2459 экз.), Е — состояние г о н а д у с а м о к в о к т я б р е 1984 г. (144 экз.), Ж — н о я б р ь — д е к а б р ь 1975 г. (1090 экз.), 3 — состояние г о н а д у с а м о к в н о я б р е — д е к а б р е 1975 г. (587 экз.), И — ноябрь 1985 г. (110 экз.), К — состояние г о н а д у с а м о к из района Китового хребта (31 экз.) полихет из семейства Terebellidae (определение Н. В. Кучерука) (табл. 2 ) . Они составляли 99,6% объема пищевого комка. Вместе с ними встречались паразитические копеподы Brachiellas, Udonellidae — эктопаразиты полихет (определение А. А. К о в а л е в о й ) . Восстановленные р а з м е р ы полихет достигали 10—60 мм (до 50% длины тела креветки). Это, а т а к ж е наличие песчинок в ж е л у д к а х , говорит о том, что F. woodwardi питалась на дне. В пище креветок из пелагиали по частоте встречаемости преобл а д а л и головоногие моллюски, копеподы, рыба и эвфаузииды. В пищевом комке доминируют рыба ( 3 4 , 4 % ) , головоногие (25,3%) и эвфаузииды ( 2 5 % ) . Следовательно, этот вид действительно мигрант, питающийся как в пелагиали (вплоть до поверхности воды), так и на дне, преодолевая 300—500-метровый слой воды. С а м а F. woodwardi служит объектом питания таких крупных хищников, как к а л ь м а р Todarodes angolensis, рыба Merluccius сарепsis (наши данные) и Thunnus maccoyii ( S h a n n o n , 1986). 69 Таблица Спектр питания F. О б ъ е к т питания и д р у г и е компоненты п и щ е в о г о ком.ся Встречаемость, % ДКО Полихеты Фораминиферы Рыба Ракообразные: креветки копеподы амфиподы эвфаузниды Головоногие моллюски Щетинкочелюстные Икра Неопределенные остатки Детрит Песок Паразитические черви Сальны Число ж е л у д к о в 91.0 16,2 12,6 6,3 1,8 2,7 0,9 3,6 - - 1,8 0,9 2,7 9,9 5,4 — 111 2 woodwardi пелагиаль 13,0 Доля в о б ъ е м е п и щ е в о г о комкя, % дпо 99,6 пелагиаль 6,0 — 32,6 6,7 42,7 7,5 18,8 42,7 0,4 3,3 1,7 1,2 0,4 — -— — - 34,3 1,0 0,7 25,0 25,0 1,3 0,3 — — — — — — — — 0,4 239 — 40 34 ЗАКЛЮЧЕНИЕ Креветки подсемейства F u n c h a l i i n a e — обитатели пелагиали, освоившие океан от бореальной до нотальной зон. Подсемейство включает обитателя умеренных вод F. woodwardi, субтропическую F. villosa и тяготеющих к тропическим водам, но встречающихся и в субтропиках Pelagopenaeus balboae, F. danae и F. taaningi. Все виды, кроме F. taaningi, встречаются в Атлантическом океане. F. danae известна и из Индийского океана. F. woodwardi и F. villosa имеют в Атлантическом океане антитропические и соответственно циркумнотальный и диркумсубтропический, а P. balboae — циркумтропический ареалы. Пока неясен биотоп фунхалий. Возможно, их отдельные популяции приурочены к конкретным м а к р о м а с ш т а б н ы м круговоротам. Д л я всех видов, кроме F. danae, очевидна связь с поверхностными слоями воды. Сохранивший наибольшее количество анцестральных признаков P. balboae (симметричная петазма, относительно короткий режущий отросток мандибулы, зазубренный нижний край рострума и т. п.), ловится только на поверхности, там ж е д е р ж а т с я ночью F. woodwardi, что неоднократно н а б л ю д а л и с борта судна. F. villosa облавливается в верхнем 50-метровом, a F. taaningi — 100-метровом слоях воды. Д л я большинства видов установлены суточные миграции с опусканием днем до верхней части материкового склона и до мезопелагиали, о чем можно судить как по прямым данным, т а к и по питанию донными объектами — F. woodwardi и находкам в ж е л у д к а х мезопелагических хищников. Об адаптации к жизни в пелагиали говорят и редуцированный рострум, и несколько утоньшенный панцирь, и заметно укороченные переоподы. Но целый ряд признаков, с б л и ж а ю щ и х фунхалии с P e n a e i n a e (сходство ж а б е р н ы х формул и строения петазмы P. balboae с таковой у Penaeus spp.), указывает на то, что эта группа осваивала п е л а г и а л ь 70 независимо от других пенеидных креветок •— S e r g e s t i d a e и Benthesicyminae. Возможно, главной причиной отхода фунхалий от своих донных родственников был переход на питание преимущественно эпипелагическими мягкотелыми животными, что вызвало и уход к поверхности воды, а затем окончательный переход в пелагиаль с такими последствиями, как редукция жевательного отростка мандибул и специфический тип пищеварения. А питание донными полихетами у F. woodwardi (кстати, тоже достаточно мягкотелыми ж и в о т н ы м и ) , видимо, у ж е вторичное явление. Возможно, оно вынужденное и присуще лишь той части вида, что обитает над материковым склоном Южной Африки. Можно полагать, что переход фунхалий в новую экологическую нишу вывел их из-под мощного пресса хищников, из-за чего не произошло взрыва видообразования (как у O p l o p h o r i d a e ) . Результатом этого явилось и уменьшение плодовитости у этой группы креветок по сравнению с P e n a e i n a e , поскольку плодовитость есть мера истребляемости (Шмальгаузен, 1968). СПИСОК ЛИТЕРАТУРЫ Б у р у к о в с к и й Р. Н. О питании креветки Pasiphaea semispinosa/lБиол. науки. 1986. № 7. Б у р у к о в с к и й Р. Н., П о л е ж а е в А. Н. О н а х о д к е в Тихом о к е а н е двух видов креветок, ранее известных из А т л а н т и к и / / Б и о л . моря. 1985. Вып. 2. Б у р у к о в с к и й Р. Н., Р о м е н е к и й Л . Л . Об особенностях р а с п р е д е л е н и я креветок у Атлантического п о б е р е ж ь я Ю ж н о й А ф р и к и / / Б ю л . М О И П . Отд. биол. 1985. Т. 90, вып. 4. Ш м а л ь г а у з е н И. И. Ф а к т о р ы эволюции М., 1968. В a b а К-, H a y аs h i К. I., T o r i y a m а М. D e c a p o d c r u s t a c e a n s f r o m c o n t i n e n t a l shelf a n d slope a r o u n d J a p a n / / J a p . Fish. Res. Cons. Assoc. Tokyo, 1986. B a l s s H. M a c r u r a der Deutschen Tiefsee-Expedition. 2. N a t a n t i a , Teil A . / / E r g e b n . «Valdivia» Exped. 1925. Vol. 20. B o u v i e r E. L. C r u s t a c e s D e c a p o d e s ( P e n e i d e s ) p r o v e n a n t des c a m p a g n e s de r « H i r o n d e l l e » et de la « P r i n c e s s e Alice» ( 1 8 8 6 — 1 9 0 7 ) / / R e s u l t . С о т р . Sci. M o n a c o . 1908. Vol. 33. B u r k e n r o a d M. D. The Aristeinae, S o l e n o c e r i n a e a n d p e l a g i c Pen a e i n a e of the B i n g h a m O c e a n o g r a p h i c C o l l e c t i o n / / B u l l . B i n g h a m o c e a n o g r . Coll. 1936. Vol. 5 (2). C r o s n i e r A. 1985. C r e v e t t e s P e n e i d e s d ' e a u p r o f o n d e recoltee d a n s l'ocean I n d i e n lors des c a m p a g n e s Benthedi, S a f a r i I et II, M D 3 2 / R e u n i o n / / B u l l . M u s . n a t n . Hist. nat. P a r i s , 1985. 4 ser., sect. A. N 4. C r o s n i e r A., F o r e s t J. Les crevettes p r o f o n d e s de 1'Atlantique oriental t r o p i c a l / / F a u n e tropicale. XIX. O R S T O M . P a r i s , 1973. D a 1 1 W. A revision on the A u s t r a l i a n species of P e n a e i n a e ( C r u s t a c e a , D e c a p o d a : Pen a e i d a e ) / / A u s t . J. M a r . F r e s h w . Res. 1957. Vol. 8 ( 2 ) . F o x t o n P. The vertical distrib u t i o n of p e l a g i c d e c a p o d s ( C r u s t a c e a , N a t a n t i a ) collected on the S o n d cruise 1965. II. P e n a e i d e a a n d g e n e r a l d i s c u s s i o n / / J . m a r i n e biol. ass. U. K. 1970. Vol. 50. F u j i n o T. F u n c h a l i a s a g a m i e n s i s sp. nov. f r o m C e n t r a l J a p a n , w i t h discusion of the generic c h a r a c t e r s ( D e c a p o d a , N a t a n t i a , P e n a e i d a e ) / / C r u s t a c e a n a . 1975. Vol. 2 8 ( 2 ) . G o r d o n I., I n g l e R. W. O n a p e l a g i c penaeid p r a w n s , Funchalia woodwardi Johnson, new to the British f a u n a / / J . m a r . biol. ass. U. K. 1956. Vol. 35. G r i p p a G. A revision of gen. Pelagopenaeus B u r k e n r o a d , 1934 a n d Funchalia J o h n s o n , 1867 ( C r u s t a c e a , Decapoda, P e n a e i d e a ) / / I n v . Pesq. 1987. Vol. 51 ( S u p l . 1). K e n s l e y B. The S o u t h African M u s e u m ' s M e i r i n g N a u d e cruises. P a r t 5. C r u s t a c e a , D e c a p o d a , R e p t a n t i a a n d Nat a n t i a / / A n n . of the S o u t h Afric. M u s e u m . 1977. Vol. 7 4 ( 2 ) . K e n s l e y В., Trant e r H. A., G r i f f i n D. J. G. D e e p - w a t e r D e c a p o d C r u s t a c e a f r o m E a s t e r n A u s t r a l i a ( P e n a e i d e a a n d C a r i d e a ) / / R e c o r d s A u s t r . M u s e u m . 1987. Vol. 39. K i k u c h i Т., О т о r i M. Vertical d i s t r i b u t i o n a n d m i g r a t i o n of oceanic s h r i m p s at t w o l o c a t i o n s off the Pacific coast of J a p a n / / D e e p - S e a Res. 1985. Vol. 32. M a c p h e r s o n E. Crustaceos Decapodos del B a n c o Valdivia (Atlantica s u d o r i e n t a l ) / / R e s . Exp. Cient. 1984. Vol. 12. M i l l e r D. G. M., A u g u s t y n L. J., H a m p t o n J. Am u n u s u a l record of the p r a w n Funchalia woodwardi ( C r u s t a c e a : D e c a p o d a ) / / S . Afric. J. m a r . Sci. 1983. Vol. 1. P a ul i n o s e V. T. Decapod C r u s t a c e a f r o m the I n t e r n a t i o n a l I n d i a n O c e a n E x p e d i t i o n : the species of Funchalia (Penaeidae) and their p o s t - l a r v a e / / J . N a t u r . H i s t . 1974. Vol. 8 ( 4 ) . R a m a d a n M. M. C r u s t a c e a : P e n a e i d a e / / J o h n M u r r a y Exped. Sci. Rep. 1938. Vol. 5 ( 3 ) . R e l i n i - O r s i L., C o s t a M. R. S e g n a l a z i o n e di Funchalia woodwardi J o h n s o n in M a r L i g u r e ( C r u s t a c e a D e c a p o d a R e n a e i d a e ) / / B o l l . M u s . e I n s t . biol. Univ. G e n o v a . 1975. Vol. 43. S h a n n o n L. V. The t u n a s of the B e n g u e l a r e g i o n off S o u t h e r n Africa — A s y n t h e s i s / / S C R S (86)31. IKKAT, 1986. W a s m e r R. A. Supple71 i n e n t a r y d e s c r i p t i o n of Funchalia taaningi B u r k e n r o a d , 1940 ( C r u s t a c e a , D e c a p o d a . P e n a e i d a e ) f r o m the C e n t r a l P a c i f i c O c e a n a n d a n e w r e c o r d of F. villosa ( B o u v i e r , 1905) f r o m the E a s t e r n I n d i a n O c e a n / / J . N a t . H i s t . 1989. Vol. 23. АтлантНИРО, Калининград Поступила в редакцию 25.10.89 FUNCHALIA SHRIMPS (DECAPODA, NATANTIA, P E N A E I D A E ) : TAXONOMY, D I S T R I B U T I O N A N D S O M E P R O B L E M S OF BIOLOGY R. N. Burukovsky, L. L. Roinensla/ Summary A c o m p i l a t i o n on the t a x o n o m y , g e o g r a p h i c a l d i s t r i b u t i o n , v e r t i c a l c i b p o s i ' i o n . biology and f e e d i n g in a s m a l l g r o u p of p e l a g i c P e n a e i d a e s h r i m p s , b e i n g v i e w e d a s F i m e b a l i i m i c s u b f a m i l y , is p r e s e n t e d . T h e s e s h r i m p s a r e s h o w n lо l u n c p e c u l i a r nurt u r e of m a n d i b l e s , o v a r y , f r o n t pai'is of the s t o m a c h a n d body c o v e r i n g .