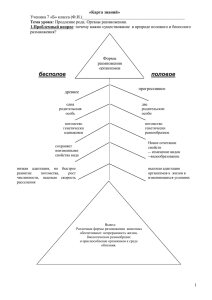
методические рекомендации к эволюционной лаборатории
реклама

МЕТОДИЧЕСКИЕ РЕКОМЕНДАЦИИ К ЭВОЛЮЦИОННОЙ ЛАБОРАТОРИИ Традиционное изучение эволюционной тематики всегда было чисто теоретическим (книжным), потому что природные эволюционные процессы имеют слишком большие масштабы во времени и пространстве для непосредственного восприятия человеческими органами чувств. Теперь компьютерные технологии позволяют моделировать и показывать эти процессы в реальном времени. Эволюционная Лаборатория была задумана как раз для того, чтобы сделать эти явления зримыми и, вдобавок, дать в них вмешиваться. Эволюционная Лаборатория включает два главных инструмента и набор уроков для их внедрения: Анимированный атлас по макроэволюции основан на современных научных сведениях о родственных связях между позвоночными животными. Он представляет собой реальное эволюционное древо, в котором можно наметить любой желаемый маршрут и проиграть его как фильм, показывающий изменения внешности животных на фоне их естественной среды обитания и с бегущей шкалой прошедшего времени. Анимированный атлас позволяет ставить пред учениками новые задачи – прокомментировать и объяснить ту или иную эволюционную цепочку (см. готовые сценарии уроков), найти по словесному описанию соответствующую ветвь на древе, назвать неподписанный узел древа (например, общего предка китов и бегемотов), изобразить промежуточную стадию эволюции, которая на древе не показана и науке не известна), разложить заданный учителем набор картинок из комплекта ЭвоЛаб в правильном эволюционном порядке и т.п. Виртуальный практикум по микроэволюции принадлежит к так называемым моделям «Искусственной жизни» – передовой отрасли применения компьютеров в биологии. В основу модели положена наглядность и быстрота отображения эволюционных процессов. Разветвление видового древа происходит прямо на глазах – как, наверно, его представлял Дарвин. Можно проводить не только лабораторные эксперименты, но и игры – соревнуясь в умении управлять жизненно важными параметрами поселений виртуальных 1 организмов. При всей своей «научности» Виртуальный практикум является настоящей компьютерной игрой. Отсутствие фиксированного набора сценариев и действительно безграничные возможности в выборе игрового поля в сочетании с небольшим набором эффективных регуляторов могут вызвать интерес даже у легкомысленных подростков. Готовые сценарии уроков включают в себя специально подобранные маршруты по Анимированному атласу и лабораторные работы в Виртуальном практикуме, которые можно сразу пустить в ход. В этих сценариях подробно описано, какими способами учитель сможет передать инициативу ученикам, а также организовать работу малыми группами, проектную деятельность и соревнования. В тексте сценариев уроков термины выделены цветом и снабжены всплывающими подсказками из словаря. Полный словарь прилагается. Анимированный атлас и Виртуальный практикум могут использоваться в рамках готовых уроков, независимо от них и порознь друг от друга. Благодаря такой гибкости, инструментарий ЭвоЛаб применим при очной, заочной, дистанционной и самостоятельной формах обучения, а также предоставляет широкие возможности для формирования нового учебного плана, индивидуальных и групповых траекторий обучения, для стимуляции проектной деятельности учащихся, для передачи инициативы от учителя ученикам и для подготовки их в ВУЗы. ФОРМЫ ПОСТРОЕНИЯ ЗАНЯТИЙ на основе ЭвоЛаб могут быть самыми разными (в готовых сценариях уроков применяются все нижеперечисленные варианты): • фронтальные лекции учителя с демонстрацией заготовок (из комплекта или сделанных самим учителем); • семинарские и практические занятия под руководством учителя; • самостоятельная исследовательская и творческая проектная деятельность учащихся в классе и на дому; • соревнования, турниры, олимпиады. 2 РЕКОМЕНДАЦИИ УЧИТЕЛЯМ • Начать с использования готовых сценариев, но потом смело варьировать маршруты в Анимированном атласе и задачи в Виртуальном практикуме в соответствии с профилем и учебной стратегией школы, а также со своими личными предпочтениями и опытом. • Заранее распространять копии ЭвоЛаб среди учеников – чтобы они могли увлечься «эволюционной игрой» и освоить ее принципы еще до официального начала прохождения эволюционной темы. • Пробовать пройти некоторые темы не на уроках, а в виде внеклассных соревнований. • Передавать ученикам инициативу и воспринимать как должное, когда они станут обгонять своего учителя в практическом освоении эволюции. • Если на то имеется техническая возможность – проводить в Виртуальном практикуме многодневные эксперименты, длящиеся миллионы виртуальных лет – как это было в реальной эволюции на Земле. • Сохранять в таких длительных экспериментах промежуточные стадии, чтобы можно было развивать эволюцию в разных направлениях с того или иного места для ответа на вопрос: «А что было бы, если…?» РЕКОМЕНДАЦИИ УЧАЩИМСЯ • Не ограничиваться заданиями учителя. • Не останавливаться на освоении заготовок из комплекта ЭвоЛаб, а скорее переходить к изобретению своих собственных (пускай сперва и наивных) сценариев путешествий в Анимированном атласе и задач в Виртуальном практикуме. • Побольше играть в Виртуальный практикум просто так. Этим путем можно стать настоящим экспертом по микроэволюции. • Почаще загружать в Виртуальный практикум свои собственные картинки и не забывать сохранять выигрышные комбинации биологических параметров для использования в новых поединках. РЕКОМЕНДАЦИИ РОДИТЕЛЯМ • Устраивать домашний показ познавательных «мультфильмов» об эволюции при помощи Анимированного атласа, где зрителям сценарий заранее никогда не известен. • Устраивать в Виртуальном практикуме семейные турниры, в которых свободно могут участвовать даже младшие члены семьи дошкольного возраста, поскольку вся модель изначально ориентирована на интуитивное восприятие наблюдаемых процессов. 3 ПОЛЕЗНЫЕ ССЫЛКИ на русском языке: • Сайт нашей Эволюционной Лаборатории: http://macroevolution.narod.ru/evollab.htm • Печатное учебное пособие в помощь к нашему Анимированному атласу: Черепанов Г.О., Иванов А.О. Палеозоология позвоночных. – М.: Издательский центр «Академия», 2007. – 352 с. • Ссылки на палеонтологические базы данных и программное обеспечение для эволюционных исследований: http://macroevolution.narod.ru/dblist.htm на английском языке: • Сайт с со всеми текстами Дарвина: http://darwin-online.org.uk/ • Сайт о филогенетических связях всех известных организмов: http://www.helsinki.fi/~mhaaramo/ • Палеонтологический сайт с описаниями и изображениями ископаемых останков организмов: http://www.palaeos.com/default.htm • • Сайт с палеобиологической базой данных, по которой проверялось время появления животных, включенных в Анимированный атлас http://paleodb.org/cgibin/bridge.pl?user=Guest&action=beginTaxonInfo Международное общество «Искусственной жизни»: http://www.alife.org/ • Очень простенькая компьютерная игра в эволюцию покровительственной окраски, которая могла бы служить эпиграфом к нашему Виртуальному практикуму: http://www.pbs.org/wgbh/nova/link/evolution.html • Сайт по генетике гаплоидного плесневого гриба Neurospora: http://www.stanford.edu/group/neurospora/home2.html БЛАГОДАРНОСТИ Авторы благодарят за практическую помощь в работе О.А. Белайчука, Е.А. Коблика, Ф.В. Лыкова, В.В. Максимова, П.В. Максимова, А.В. Маркова, И.В. Савельева, И.Д. Селюцкого и С.Э. Сильвестрова. Также авторы благодарят за ценные советы и консультации А.И. Буяновского, А.С. Гамбарян, Н.В. Дмитриеву, Ю.Ф. Ивлева, Д.В. Карелина, В.С. Лебедева, И.В. Муратова, И.М. Никулину, А.К. Панютина, В.С. Фридмана и А.А. Цессарского. 4 ОГЛАВЛЕНИЕ Урок I: «Разнообразие позвоночных животных как продукт эволюции». стр. 6 Урок II: «Жизненный цикл многоклеточных, механизм передачи наследственных признаков и конъюгации хромосом». стр. 11 Урок III: «Цветное зрение человека и обезьян и применение его принципов в практике». стр. 18 Урок IV: «Выживание наиболее приспособленных на примере скрывающей окраски. Роль мутаций в приспособительной эволюции». стр. 23 Урок V: «Эволюционное значение полового размножения». стр. 27 Урок VI: «Половой отбор и эволюционное значение разных способов размножения». стр. 30 Урок VII: «Половой отбор и эволюционное значение разных способов размножения» (продолжение для профильного уровня). стр. 35 Урок VIII: «Принцип дивергенции и экологическое видообразование». стр. 41 Урок IX: «Принцип дивергенции» (продолжение). стр. 48 Урок X: «Симбиоз. Экспериментальная контрольная работа». стр. 55 СЛОВАРЬ ТЕРМИНОВ И НАЗВАНИЙ ЖИВОТНЫХ стр. 60 5 Урок I: «Разнообразие позвоночных животных как продукт эволюции». VII класс – тема «Разнообразие и эволюция живой природы», подтема «Разнообразие позвоночных животных». Содержательная задача – показать, что эволюционное древо росло и ветвилось как настоящее дерево. Методическая задача – научить учеников работать с Анимированным атласом (освоить интерфейс). УЧИТЕЛЬ ОБЪЯСНЯЕТ, КАК РАБОТАТЬ С АНИМИРОВАННЫМ АТЛАСОМ (5-7 мин.) Запустите Анимированный атлас, щелкнув по этому слону. Откроется окно с эволюционным древом позвоночных животных. В нем магистральный путь («ствол») от первых рыб до человека проложен вдоль левого края окна снизу вверх. Для исследования древа пользуйтесь увеличивающей и уменьшающей лупой. Для того, чтобы «оживить» ту или иную ветвь древа, перетащите флажки «Старт» и «Финиш» из Панели инструментов на изображения животных (иконки) на обоих концах интересующей ветви. Как только эти флажки расставлены – маршрут эволюционного путешествия от флажка до флажка подсвечивается. Теперь можно «проиграть маршрут» и увидеть эволюционное превращение одного животного в другое. УЧИТЕЛЬ ПРОВОДИТ СЕРИЮ ДЕМОНСМТРАЦИЙ В АНИМИРОВАННОМ АТЛАСЕ (18-20 мин.) Биология как наука начиналась с изучения человека и потом распространялась все дальше и дальше от него. Эта серия демонстраций построена в аналогичном порядке. Демонстрация № 1: эволюционные корни человека. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «человек разумный», а финиш – на иконку «первые рыбы». Остается проиграть этот маршрут. 6 В последующих демонстрациях старт будет помещаться на показанном в 1-й демонстрации эволюционном стволе – в том месте, где от него отходит очередная «боковая» ветвь. Вначале «старт» будет устанавливаться ближе к вершине (человеку разумному), а в каждой следующей демонстрации – все ниже и ниже по стволу. Демонстрация № 2: эволюция человекообразных обезьян. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «узконосые», а финиш – на боковую иконку «орангутан». Остается проиграть этот маршрут. Объяснение. Предки обезьян с самого начала пошли на решительный отказ от нюха в пользу зрения. Это было связано со сдвигом режима активности на дневное время. Но чтобы уклониться от конкуренции с насекомоядными птицами, обезьянам пришлось приспособиться питаться плодами, даже грубыми и неспелыми, и листьями. Соответственно, они укрупнились в размерах и стали всё больше полагаться при лазании на руки, чем на ноги. Все это происходило поначалу в Африке, а потом многие обезьяны и люди эмигрировали в Евразию. Демонстрация № 3: эволюция лемуров. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «приматы», а финиш – на боковую иконку «лемуры». Остается проиграть этот маршрут. Объяснение. Приматы начали эволюционную карьеру в Северном полушарии, когда там были обширные дождевые леса. Под их пологом они скакали по кустам и ловили ночных насекомых, полагаясь как на нюх, так и на глаза. Затем, при сокращении тропических лесов, полуобезьяны сместились на юг – в Африку, и дальше – на остров Мадагаскар. Тут они достигли особого расцвета в лице разнообразных лемуров, даже дневных – благо настоящие обезьяны сюда не добрались. Демонстрация № 4: эволюция насекомоядных млекопитающих. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «плацентарные», а финиш – на боковую иконку «выхухоль». Остается проиграть этот маршрут. 7 Попросите учеников прокомментировать наблюдаемые превращения. Чем может питаться выхухоль и как она плавает? Демонстрация № 5: эволюция сумчатых млекопитающих. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «млекопитающие», а финиш – на боковую иконку «кенгуру». Остается проиграть этот маршрут. Объяснение. Исходно сумчатые были мелкими и ловкими ночными зверьками – рылись в лесной подстилке и часто лазали по деревьям в поисках насекомых и тому подобной живности. В те времена Антарктида непосредственно соединялась как с Южной Америкой, так и с Австралией, и еще была не на самом Южном Полюсе, так что годилась для жизни. По этому антарктическому «мосту» сумчатые и добрались со своей настоящей родины-Америки – до Австралии. Уже там некоторые сумчатые стали углублять приспособления к жизни на деревьях. Хвост стал хватательным, и пальцы на лапах сгруппировались совершенно особым образом. Дальше некоторые лазающие растительноядные сумчатые оказались способны вернуться на землю и захватить открытые пастбища Австралии. Это кенгуру. С экологической и физиологической точки зрения они подобны жвачным парнокопытным других материков. Демонстрация № 6: эволюция яйцекладущих млекопитающих. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «зверообразные», а финиш – на боковую иконку «яйцекладущие». Остается проиграть этот маршрут. Попросите учеников прокомментировать наблюдаемые превращения. Кто по их мнению тот, кто здесь помечен флажком «старт» – пресмыкающееся или млекопитающее? Демонстрация № 7: эволюция пресмыкающихся и птиц. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «пресмыкающиеся», а финиш – на боковую иконку «птицы». Остается проиграть этот маршрут. 8 Попросите учеников прокомментировать наблюдаемые превращения. Чем птицы похожи на динозавров? Демонстрация № 8: эволюция бесхвостых земноводных. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «мясистолопастные», а финиш – на боковую иконку «бесхвостые». Остается проиграть этот маршрут. Объяснение. Такие хищные, примерно метровые рыбы жили четыреста миллионов лет назад на перегретых застойных мелководьях. Выбираясь из подводных зарослей к поверхности, чтобы наполнить легкие свежим воздухом, они помогали себе мускулистыми парными плавниками и стали на них мало-помалу выползать за добычей на мели. Чтобы продолжить интервенцию берегов этим рыбам пришлось оторвать брюхо от пляжа, подогнув членистые плавники – теперь уже лапы; места сгиба стали локтем и коленом. Для опоры выросли пальцы. Вначале их было штук по восемь на каждой лапе, и «смотрели» они задом наперед, но постепенно обе «недоработки» стали выправляться, а тело – освобождаться от бремени чешуйчатой брони рыб. Наконец, жабры исчезли, а лапы из пассивных костылей сделались правильно согнутым шагающим механизмом о пяти пальцах. Но обмен веществ у земноводных остался почти как у пресноводных рыб – им требуется много воды – даже таким сухопутным земноводным как жабы. А весной есть гораздо более серьезная причина вернуться в воду – нерест. Ведь главная «недоработка» амфибий – что они еще размножаются по рыбьи – икрой! Демонстрация № 9: эволюция костистых рыб. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «костные рыбы», а финиш – на боковую иконку «костистые рыбы». Остается проиграть этот маршрут. Попросите учеников прокомментировать наблюдаемые превращения. Кого на нашей планете больше – рыб, птиц или млекопитающих? Демонстрация № 10: эволюция скатов. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на стволовую иконку «первые рыбы» (на которой 9 был финиш в самой первой демонстрации), а финиш – на боковую иконку «скаты». Остается проиграть этот маршрут. Попросите учеников прокомментировать наблюдаемые превращения. Чем питаются скаты и как они плавают? УЧИТЕЛЬ ПЕРЕДАЕТ ИНИЦИАТИВУ УЧЕНИКАМ (18-20 мин.) Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «первые рыбы», а финиш – на иконку «человек разумный». Учитель ставит перед учениками задачу продемонстрировать другие эволюционные ответвления от помеченного ствола с двумя условиями: 1) Флажок финиша надо переставить на иконку любого современного животного. Чтобы соблюсти это условие, ученику придется вспомнить или угадать, какие животные живут в наши дни, а какие уже вымерли. 2) Флажок старта надо переставить на ту иконку «ствола», которая находится ближе всего к выбранному финишу. Попросите ученика прокомментировать наблюдаемые превращения. В чем их биологический смысл? Знал ли он этих животных раньше? 10 Урок II: «Жизненный цикл многоклеточных, механизм передачи наследственных признаков и конъюгации хромосом». VIII класс – тема «Признаки живых организмов», подтема «Генетика – наука о закономерностях наследственности и изменчивости». Содержательная задача – первый подход к генетике в облегченном варианте. Это особенно важно для тех, кто не пойдет в X-XI классы. Вводятся термины: «ген», «аллель», «хромосома», «оплодотворение», «гаплоидность», «диплоидность», «мейоз» «конъюгация», «рекомбинация». Методическая задача – подготовка к освоению модели Виртуального практикума. ОБЪЯСНЕНИЕ ПРИНЦИПИАЛЬНОЙ СХЕМЫ ЖИЗНЕННОГО ЦИКЛА МНОГОКЛЕТОЧНЫХ ОРГАНИЗМОВ С ПОЛОВЫМ РАЗМНОЖЕНИЕМ (10 мин.) Жизненный цикл многоклеточных растений, грибов и животных состоит из гаплоидной и диплоидной фазы – когда каждый «элементарный наследственный задаток» присутствует, соответственно, в одном единственном или в двух экземплярах – не обязательно идентичных. Можно говорить о двойном и одинарном наборе хромосом или двойном и одинарном наборе аллелей каждого гена. Переход из диплоидной фазы в гаплоидную происходит за счет особого способа деления клеток – мейоза. Переход из гаплоидной фазы в диплоидную происходит путем оплодотворения (половой процесс). У разных организмов многоклеточное существование приходится на разные фазы цикла – иногда на обе, но чаще на какую-либо одну. Генетики изучили оба варианта: 1) Горох и плодовая мушка-дрозофила стали классическими объектами для изучения наследования у тех организмов, которые многоклеточны в диплоидной фазе – как человек. 2) Плесень Neurospora стала классическим объектом для изучения наследования у тех организмов, которые многоклеточны в гаплоидной фазе. У них законы наследственности проявляются проще. Жизненный цикл Neurospora показан на следующем рисунке. 11 ОБЪЯСНЕНИЕ МЕХАНИЗМА РЕКОМБИНАЦИИ (ПЕРЕКОМБИНАЦИИ) НАСЛЕДСТВЕННЫХ ЗАДАТКОВ (15-20 мин.) В качестве примера рассматривается вымышленный организм со следующими свойствами: • Имеются три части тела – красная, зеленая и синяя • Информация о каждой части передается по наследству отдельно, то есть красный, зеленый и синий гены находятся в разных хромосомах. • Разная яркость каждого цвета связана с разными аллелями (вариантами) соответствующего гена. Жизненный цикл такого организма показан на следующем рисунке. Разные аллели условно изображены в виде хромосом разной длины. 12 Данный рисунок позволяет ввести следующие законы генетики: «Закон чистоты гамет» Бэтсона. В диплоидной фазе жизненного цикла два родительских аллеля не смешиваются и не усредняются, а переходят в следующую гаплоидную фазу в исходном чистом виде. Третий закон Менделя. Хромосомы при мейотическом делении распределяются независимо друг от друга, то есть при переходе из диплоидной фазы в гаплоидную аллели тех генов, которые находятся в разных хромосомах, комбинируются в любых сочетаниях. Задача: сколько может быть гаплоидных комбинаций в рассматриваемом случае с тремя разноцветными генами? На следующем рисунке показано, что правильное решение задачи – 8 комбинаций. 13 Общее решение: при переходе из диплоидной фазы в гаплоидную количество возможных гаплоидных вариантов равно 2n, где n – гаплоидное число хромосом. ОБЪЯСНЕНИЕ КОНЪЮГАЦИИ ХРОМОСОМ (15-20 мин.) Перед переходом в гаплоидную фазу «одноименные» хромосомы каким-то образом узнают друг друга и благодаря этому выстраиваются парами – конъюгируют. В нашем вымышленном примере – красная с красной, зеленая с зеленой, синяя с синей (как показано на предшествующем рисунке). Вопрос: для чего этот механизм конъюгации служит? Подсказка: пойдем от противного – представим себе, что, например, аллели синего цвета так сильно различаются между собой, что соответствующие хромосомы не узнают друг друга и не конъюгируют. Что из этого получится? Эту ситуацию легко смоделировать в виде карточной головоломки, показанной на следующем рисунке. 14 Кто вытащит прославленную комбинацию «тройка, семерка, туз» – пират или леди? Ответ – либо оба, либо никто. Размышление над этой головоломкой помогает сообразить, что пират и леди делят «без спору» только сдвоенные карты, а разрозненные карты (тройка и туз синей масти) могут случайно сразу обе достаться какому-то одному игроку, а другому тогда – ни одной. На следующем рисунке ровно то же самое, что было с картами, показано на разноцветных хромосомах. 15 В половине случаев (нижний ряд делящихся клеток) не конъюгировавшая хромосома (здесь синяя) либо вообще не перейдет в гаплоидную фазу, либо перейдет в двух экземплярах вместо одного. И то, и другое является отклонением, которое обычно приводит к патологии (синдром Дауна у человека) или вообще несовместимо с жизнью. Таким образом, конъюгация необходима для правильной комплектации хромосом при переходе в гаплоидную фазу. Задание на дом: для того же примера с тремя цветными хромосомами предлагается обдумать – какова (с учетом третьего закона Менделя) вероятность того, что гаплоидная фаза получит правильный комплект хромосом (по одной каждого цвета), если не произойдет конъюгация двух пар хромосом из трех? А если не конъюгируют все три пары? На эти вопросы легче ответить, разбирая карточные головоломки вроде предложенной выше. Решение: Если не конъюгируют две пары хромосом из трех, то вероятность получения правильного гаплоидного комплекта составляет – 1/4, а если не конъюгируют все три пары – вероятность благополучного исхода 1/8. Иначе говоря, каждая не конъюгировавшая пара хромосом убавляет число жизнеспособных потомков вдвое. Пример нарушения конъюгации – бесплодие мулов. Кто такие мулы? И кто такие ослы и лошади? Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «непарнокопытные», а финиш – на иконку «зебра». Остается проиграть этот маршрут. Объяснение. Все современные представители семейства лошадиных – зебры, ослы и лошади – относятся к одному роду. Мулы и лошаки – это гибриды лошади и осла. Мулы получаются от кобылы с ослом, а лошаки – от жеребца с ослицей (они мельче в соответствие с размером матери). Мулы знамениты трудоспособностью, из-за которой и производились в массовом количестве в эпоху гужевого транспорта. Также знамениты бесплодием – практически полной неспособностью давать яйцеклетки или сперматозоиды. Лошади имеют 32 пары хромосом, а ослы – 31 пару, и их гибриды соответственно получают нечетный комплект из 63 хромосом. Но бесплодие этих гибридов связано с другим. Ведь гибриды домашней лошади и дикой лошади Пржевальского с 33-мя парами хромосом тоже получают нечетный набор – 65, но вполне плодовиты. Суть в том, что хромосомы лошади Пржевальского достаточно похожи на таковые домашней лошади, а ослиные – недостаточно, и не могут 16 попарно конъюгировать и потом аккуратно распределяться при переходе в гаплоидную фазу по сперматозоидам или яйцеклеткам. В результате половые клетки мулов и лошаков получают неправильные комплекты генов, отчего и гибнут – не мулы и лошаки, а их половые клетки. 17 Урок III: «Цветное зрение человека и обезьян и применение его принципов в практике». IX класс – тема «Человек и его здоровье», подтема «Органы чувств, их роль в жизни человека». Содержательные задачи – (1) усвоение иллюзорности нашего цветовосприятия; (2) знакомство с наукой бионикой на примере изобретения цветного экрана. Методическая задача – подготовка к освоению модели Виртуального практикума. ИСТОРИЯ ПРОИСХОЖДЕНИЯ ЦВЕТОВОСПРИЯТИЯ ЧЕЛОВЕКА И ЕГО ПРИНЦИПЫ (20-25 мин.) Демонстрация: прогресс обезьян Старого Света. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «приматы», а финиш – на иконку «орангутан». Остается проиграть этот маршрут. Объяснение. Приматы начали эволюцию в Северном полушарии, когда там были обширные дождевые леса. Там на деревьях по ночам они ловили насекомых, полагаясь уже не только на слух и нюх (как более примитивные млекопитающие), но и на увеличенные, чувствительные к слабому свету глаза. Затем, при сокращении тропических лесов, приматы сместились на юг, где дали две независимые линии обезьян – южноамериканскую и африканскую. И те, и другие отмежевались от полуобезьян в первую очередь благодаря тому, что перенесли режим бодрствования на светлое время суток. Дневной образ жизни предъявляет более высокие требования к зрению, чем к обонянию. Соответственно, нюх у обезьян ослаб, а зрение улучшилось, и особенно радикально – у африканских обезьян, которые приспособились отличать красные цвета от зеленых, к чему обычные млекопитающие не способны. С такими глазами легче находить спелые, уже пожелтевшие, порыжевшие и покрасневшие плоды среди зелени – а именно они стали для обезьян главной едой вместо насекомых. Освоив столь удачный пищевой ресурс, африканские обезьяны во всеоружии вернулись в дождевые леса Евразии. Объяснение цветовосприятия человека и обезьян: «Как они увидели красный цвет?» Видимый свет – это электромагнитное излучение в очень ограниченном диапазоне спектра (с небольшими различиями границ у разных животных). 18 Когда свет прошел через зрачок (отверстие) и хрусталик (линза), он падает на светочувствительное дно глазного яблока – сетчатку. У позвоночных животных в сетчатке бывает два основных типа клеток-приёмников – палочки и колбочки (названия соответствуют форме). Палочки значительно чувствительнее и поэтому лучше подходят для сумеречных животных, чем колбочки. Колбочками можно пользоваться только при хорошем освещении, и поэтому они характерны для дневных животных.. Преимущество колбочек состоит в том, что они бывают разные – для одних более «заметна» одна часть показанной выше радуги, для других – другая. Благодаря этому комплект из колбочек разного сорта может различать не просто яркость света, а разные части радуги и даже смешанные цвета, и тем точнее, чем больше сортов колбочек. Это и есть восприятие цвета. Именно обезьяны и человек являются самыми дневными млекопитающими. Поэтому у них следует ожидать более дифференцированного набора колбочек, чем у остальных. Действительно, было обнаружено, что у обезьян Старого Света к двум старым добавился третий тип колбочек – отсутствующий у подавляющего большинства млекопитающих. О том, как видит это большинство, можно получить представление, посмотрев на правую половину следующего изображения. Левая половина – это то, как видим мы. Впрочем, и среди людей найдутся такие дальтоники (обычно мужского пола), которые не заметят между двумя половинками особой разницы – как не заметили бы ее наши ночные предки, а также зайцы и лисицы. 19 ИСТОРИЯ ПРАКТИЧЕСКОГО ПРИМЕНЕНИЯ ТЕОРИИ ЦВЕТОВОСПРИЯТИЯ ЧЕЛОВЕКА (БИОНИКА) Еще до фактического открытия трех типов колбочек человека была сформулирована так называемая трехкомпонентная теория его цветовосприятия (теория Ломоносова-ЮнгаГельмгольца). В ней за основные цвета были приняты красный, зеленый и синий (RGB), что довольно точно соответствует трем реально существующим типам колбочек, открытым позже. На основе этой теории был изобретен принцип цветного экрана телевизора, а потом и компьютера. Цветной экран является типичным продуктом бионики – прикладной отрасли науки, занимающейся внедрением принципов биологических явлений (в данном случае это трехкомпонентное цветовосприятие человека) в технику. Каждая «ячейка» такого экрана выполнена из трех светящихся элементов – красного, зеленого и синего. Благодаря крошечному размеру их цвета для нашего глаза смешиваются. Каким нам будет представляться смешанный цвет, зависит от интенсивности накала красного, синего и зеленого элементов. В цифровом формате имеется 256 степеней накала: от 0, когда данный элемент полностью потушен, до 255, когда он максимально ярок. Демонстрация этого легко осуществляется путем рассматривания монитора компьютера в лупу. Рекомендуется рассмотреть: белый участок монитора – ярко горят светящиеся элементы всех трех цветов и желтый участок монитора – ярко горят красные и зеленые элементы, а синие потушены Желтый цвет особенно хорошо позволяет уяснить иллюзорность нашего цветовосприятия. Желтый присутствует в радужном спектре, то есть в природе он существует в чистом виде. Но посмотрев на желтый участок монитора в лупу мы убедились, что на цветном экране он не чистый, а смешивается из красного и зеленого. То, что эта смесь подменяет нам чистый спектральный желтый цвет – типичная цветовая иллюзия (обман зрения), базирующаяся на том, что у нас в сетчатке всего три типа колбочек. Если бы их было больше, экран компьютера нас не мог бы так обмануть. Демонстрация цифрового трехкомпонентного программирования цвета средствами Виртуального практикума – куб RGB. Числа, которые кодируют яркость красной, зеленой и синей компонент на мониторе можно отложить по трем координатным осям. Поскольку яркость по каждой координате заключена в диапазоне от 0 до 255 – получится куб. Каждая его ячейка имеет свой цвет. 20 Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. Куб, который появится на экране, можно повертеть при помощи стрелок на клавиатуре или колесика мышки. Отображение произведений живописи в системе координат RGB-куба средствами Виртуального практикума. Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. В одном окне появится картина И.И. Шишкина «Утро в сосновом лесу», сверху которой наложен радужный спектр; Другое окно покажет ту же самую картину и радугу – как они выглядят в кубе RGB (3D график). Можно будет увидеть примерно следующее изображение: Характерные особенности наблюдаемой картины, на которые следует обратить внимание: Радужный спектр изгибается вдоль рёбер RGB-куба (это подтверждение того, что цветной экран устроен так, чтобы обманывать наше зрение). Палитра картины И.И. Шишкина «Утро в сосновом лесу» (и, как правило, других произведений живописи) почти не использует насыщенные цвета, а вытянута от черного угла куба к белому углу вдоль так называемой серой диагонали. Это доказывает, что для нас цвет так и остался менее важным источником информации, чем общий уровень яркости. На этом явлении основано еще одно изобретение бионики, направленное на уменьшение объема цифровых видеозаписей. Один из методов компрессии состоит в том, что информация о 21 яркости записывается в каждом пикселе (элементарной точке) видеокадра, а информация о цвете – через один, что для глаза остается незаметным. 22 Урок IV: «Выживание наиболее приспособленных на примере скрывающей окраски. Роль мутаций в приспособительной эволюции». X-XI классы – тема «Вид», подтема «Элементарные факторы и движущие силы эволюции; их взаимосвязь». ОБЪЯСНЕНИЕ (5-7 мин.) Разбор понятий «скрывающая (покровительственная, маскировочная) окраска», «выживание наиболее приспособленных (естественный отбор)», «мутации (неопределенная изменчивость)» на классическом примере промышленного меланизма у березовой пяденицы в Англии. Эти сумеречные бабочки проводят день, замерев на стволах и ветках берез, где благодаря светлой окраске мало заметны для насекомоядных птиц. В промышленных районах береста покрыта копотью, и там распространились черные (мутантные) особи, как более приспособленные скрываться на черном фоне от глаз птиц. РАБОТА В ВИРТУАЛЬНОМ ПРАКТИКУМЕ (18-20 мин.) Задача: имитация эволюции березовой пяденицы. Условия: фон игрового поля почти черный (как береста в промышленных районах); исходная популяция состоит из почти идеально белых особей (как исходные бабочки). Упрощающее условие: виртуальные организмы, в отличие от березовой пяденицы, не диплоидны, а гаплоидны. Усложняющее условие: у березовой пяденицы степень белизны/черноты кодируется одним геном, а у виртуальных организмов цвет кодируется тремя генами (такой принцип смешения цветов рассматривался на предыдущем уроке применительно к цветовосприятию человека и кодированию цвета на мониторе). Модель естественного отбора, лежащая в основе Виртуального практикума, такова, что поселение виртуальных организмов приобретает в ходе эволюции маскировочную окраску под цвет фоновой картинки (поля). Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. 23 в нем в качестве поля загрузите картинку промышленная_Англия.bmp – тогда в Виртуальном практикуме откроется окно с темно-серым полем; для создания поселения организмов загрузите картинку 100000_белых_пядениц.bmp – тогда в Виртуальном практикуме откроется еще окно с популяцией почти белых организмов; чтобы наблюдать другие аспекты процесса, создайте еще два окна: 1) окно «Адаптация», 2) окно «Гистограмма» – чтобы видеть изменение соотношения в поселении разных вариантов (аллелей), например, красного гена (с тем же успехом можно взять зеленый или синий); в Панели управления присвойте поселению 0.001 % мутаций (это вполне реалистичная величина) – либо вручную, либо загрузив настройки из файла березовая_пяденица.ini; запустите эволюционный процесс и наблюдайте. Пауза автоматически наступит через 100 виртуальных лет. При этом, поскольку стартовое поселение состоит из 100000 организмов (это площадь файла 100000_белых_пядениц.bmp, измеряемая в пикселах), и каждый «год» каждая самка (а их в поселении половина) рождает около 8 потомков, то за назначенные 100 «лет» в эволюционном процессе поучаствует почти 40 миллионов особей (на самом деле несколько меньше благодаря летальным мутациям). Реально перед пользователем предстанут в виде пикселов всего-навсего 10 миллионов особей, потому что те остальные, которые проигрывают борьбу за существование и отсеиваются естественным отбором, на экран не выводятся. Будут наблюдаться следующие явления: в окне, где первоначально было почти белое поселение, будут возникать и распространяться разноцветные и всё более темные (покровительственно окрашенные) мутантные потомки; в окне «Адаптация» эти новые, более адаптированные к среде (полю) организмы будут отображаться все более бледными градациями серого цвета (идеальная приспособленность отображается белым цветом); в окне «Гистограмма» будет наблюдаться типичная последовательность фаз естественного отбора – вначале дестабилизация, затем стабилизация на новом месте. Как и всегда при работе в Виртуальном практикуме, рекомендуется несколько раз повторить описанный выше эксперимент. 24 Для экономии времени можно уже не проходить все описанные выше шаги, а запустить Виртуальный практикум с готовой задачей, щелкнув по этой мушке-дрозофиле. Выводы, к которым приходят учащиеся: Мутации имеют случайный характер (Дарвин называл это неопределенной изменчивостью); это вытекает из наблюдения, что никак нельзя предугадать, в каких точках местообитания поселения они возникнут, и как сильно мутанты будут отличаться от родителей. Мутации в разных генах возникают совершенно независимо друг от друга и редко возникают одновременно; это вытекает из наблюдения, что процесс конкурентного вытеснения светлых особей более темными всегда начинается с появления голубых, розовых и желтых мутантов, а именно эти цвета и соответствуют изменению только какого-то одного из трех генов, кодирующих цвет у виртуальных организмов (красного, зеленого и синего, соответственно). Общее направление эволюции определяется не мутациями, а естественным отбором; это вытекает из того, что через какие бы случайные цветные мутации ни шло вытеснение светлых особей, процесс всегда заканчивается «победой» тёмно-серых. РАБОТА С АНИМИРОВАННЫМ АТЛАСОМ (18-20 мин.) Задача: показать и обсудить реальные примеры эволюции скрывающей окраски у млекопитающих. Пример № 1 – лошадиные. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «непарнокопытные», а финиш – на иконку «зебра». Остается проиграть этот маршрут. Объяснение. Северная Америка пятьдесят пять миллионов лет назад. Это лесной зверек гиракотерий ростом сорок сантиметров. Ест всё – от плодов до мелкой живности. На задних лапах – три копытца, а на передних – четыре, но потом мизинец исчезает, и остается тоже три пальца. В меню начинают преобладать ветки и листья. Спина сохраняет гибкость – признак спринтера. Но вот появляются травянистые прерии. Переход на грубый травяной корм потребовал массивных челюстей, и увеличения роста – до размера пони. Пятнадцать миллионов лет назад боковые пальцы так уменьшились, что перестали опираться на землю. Последующее исчезновение боковых копыт делает ноги еще легче, а спина становится 25 жесткой (уже можно было бы седлать). Это – признаки степных стайеров. Два миллиона лет назад они имели огромный успех. Тогда из Северной Америки табуны однокопытных травоядных двинулись в Южную Америку, Евразию и Африку, заняв все травянистые равнины – прерии, пампасы, степи и саванны. На самом деле зебры, лошади и ослы относятся к одному и тому же роду, то есть имеют более тесное родство, чем, скажем, домовая и полевая мыши. Сейчас время процветания для лошадей закончилось, и следы былого успеха можно лицезреть только в африканских саваннах. Выводы, к которым приходят учащиеся: Скрывающая окраска не обязательно должна быть одноцветной, а может имитировать мелкие детали типичного рисунка окружающей среды. Пятнисто-полосатый рисунок шкуры с «солнечными зайчиками» сменился у предков лошадей на ровную окраску, когда они вышли из лесов на открытые пастбища. Полосатость зебр – тоже особый случай скрывающей окраски: так львам труднее сосредоточить взгляд на какой-то одной зебре в бегущем табуне. Пример № 2 – лисьи. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «псовые», а финиш – на иконку «песец». Остается проиграть этот маршрут. Попросите учеников прокомментировать наблюдаемые превращения. Какие еще млекопитающие белеют на зиму? (заяц-беляк и горностай) 26 Урок V: «Эволюционное значение полового размножения». X-XI классы – тема «Вид», подтема «Элементарные факторы и движущие силы эволюции; их взаимосвязь». ПРИНЦИПИАЛЬНАЯ СХЕМА ЖИЗНЕННОГО ЦИКЛА МНОГОКЛЕТОЧНЫХ ОРГАНИЗМОВ С ПОЛОВЫМ РАЗМНОЖЕНИЕМ; МЕХАНИЗМ РЕКОМБИНАЦИИ ХРОМОСОМ (25 мин.) Закон «чистоты гамет» Бэтсона. Третий закон Менделя о независимом распределении. (повторение урока № II) РАБОТА В ВИРТУАЛЬНОМ ПРАКТИКУМЕ. Задача – доказать, что рекомбинация наследственных задатков, возникающая благодаря половому размножению, ускоряет приспособительную эволюцию. (20 мин.) Организм Виртуального практикума имеют такой же жизненный цикл, как у плесени Neurospora – они проводят жизнь в гаплоидной фазе. У них три хромосомы: в одной локализован ген красной компоненты цвета, в другой – зеленой компоненты, в третьей – синей компоненты. Яркость каждой компоненты зависит от конкретного аллеля соответствующего гена. Условия эксперимента. Чтобы продемонстрировать роль рекомбинации в чистом виде, возникновение мутаций заблокировано (в «Панели управления» процент мутаций установлен на ноль), и все генетическое разнообразие виртуальных организмов остается в рамках разнообразия стартового поселения. Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. Откроются четыре окна: 1) Поле – это картина А.К. Саврасова «Домик в провинции»; она имеет площадь 117200 пикселов – это максимальное число организмов, которые могут на ней жить; 2) Поселение «NE» в реальных цветах – это некая старинная фотография охотников – не цветная и не черно-белая, а то, что называется «сепия»; это и есть стартовое поселение, размер которого 57600 особей (пикселов); 27 3) 3D график этой фотографии в пространстве RGB-куба (с этим типом представления картин учащиеся уже знакомы по уроку № III); 4) Возраст – в нем показан возрастной состав виртуального поселения; возраст каждого пиксела отображается оттенками серого цвета – чем старше особь, тем чернее. Начальное условие задачи – чисто бесполое размножение – клонирование. Запустив эволюционный процесс сосредоточьте внимание на трех окнах: В окне реальных цветов поселения «NE» за счет размножения и естественного отбора покровительственно окрашенных организмов, потомки пикселов старинной фотографии охотников постепенно покроют всю площадь поля и поселение превратится в подобие старинной фотографии «Домика в провинции». В окне «Возраст» будут наблюдаться чёткие белые фронты там, где идет активный естественный отбор, в результате которого старые хозяева территории замещаются молодыми, и мелкая черно-белая мозаика (то есть смесь старых и молодых организмов) в тех местах поля, где ситуация стабилизировалась. За 250 лет ситуация стабилизируется по всему полю и произойдет пауза. В окне «3D график» будет наблюдаться вырождение, потому что естественный отбор уничтожает некоторые клоны, а новых не возникает, ибо мутаций нет. Кода наступит пауза, надо в Панели управления присвоить новое значения в пункте «% клонирования», а именно ноль вместо 100 – чтобы организмы перестали размножаться бесполым путем и начали скрещиваться (в данном примере это будет гермафродитическое размножение). После этого, благодаря скрещиванию организмов, эволюционный процесс примет совершенно другой оборот, причем вначале очень резко. Поэтому перед обычным запуском эволюционного процесса до автоматической паузы рекомендуется несколько раз нажимать на кнопку «запустить на 1 год» – пока процесс не замедлится. Будут наблюдаться следующие явления: В окне «Возраст» вновь появятся белые фронты, показывающие, где естественный отбор активно заменяет старых жильцов молодыми. В окне фенотипа «NE» изображение цвета сепии быстро превратится в не очень точную, но цветную (!) копию «Домика в провинции», то есть окраска виртуальных организмов при половом размножении сделается значительно более покровительственной. 28 В окне «3D график» это появление разнообразия цветов выразится в резком расширении облака точек (вместо наблюдавшегося при бесполом размножении сужения). Главный вывод: поскольку мутации по условию эксперимента не возникают, то появление новых точек на 3D графике вызвано ни чем иным, как рекомбинацией родительских наследственных задатков. 29 Урок VI: «Половой отбор и эволюционное значение разных способов размножения». X-XI классы – тема «Вид», подтема «Элементарные факторы и движущие силы эволюции; их взаимосвязь». Содержательная задача – экспериментальное доказательство влияния способа полового размножения на распространение мутаций и скорость эволюции. ПОСТАНОВКА ПРОБЛЕМЫ С ИСПОЛЬЗОВАНИЕМ АНИМИРОВАННОГО АТЛАСА (20 мин.) Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «баран», а финиш – на иконку «олень» (или наоборот – в данном случае это не принципиально). Остается проиграть этот маршрут. Поскольку оба флажка поставлены на вершины ветвей древа, эволюционное превращение в Проигрывателе вначале идет вспять (к общему предку барана и оленя, иконка которого подписана как «жвачные»), а только потом – от прошлого к настоящему. За изменением направления движения во времени надо следить по ползунку шкалы времени. Объяснение. Вершиной эволюции жвачных парнокопытных стали очень разнообразные антилопы, быки, бараны и им подобные, которых объединяют под названием «полорогие». Кожа на их костяных рогах стала твердым как копыто роговым чехлом. А самые сложные – ветвящиеся – рога приобрели другие жвачные, сохранившие приверженность лесному образу жизни – олени. Кожа с их костяных рогов полностью облезает, но рога так тяжелы, что самки их как правило не носят, а самцы сбрасывают на зиму (и без них тяжко). Ученикам предлагается выдвинуть гипотезы, для чего барану и оленю служат рога. (Для защиты ли от хищников, например волков, или для чего-то еще?) Дополнительные (наводящие) вопросы: У всех ли оленей есть рога? (Обычно их нет у самок). В какое время года олени носят рога? (Во время гона). Выводы: Существуют внешние признаки (такие как огромные, тяжелые рога), появление которых в эволюции нельзя объяснить обычным естественным отбором в направлении прямой 30 приспособленности к внешней среде, и которые даже прямо противоречат этому и вообще затрудняют жизнь. Такие признаки – результат особого отбора, при котором отбирающим агентом является противоположный пол. Дарвин назвал это половым отбором. А внешние различия самцов и самок называются половым диморфизмом. Обсуждение вопроса о том, почему такие половые признаки как рога и яркая окраска чаще наблюдаются у самцов, чем у самок. Учитель подводит учеников к мысли, что обычно половой отбор сильнее сказывается на самцах, потому что они конкурируют между собой за то, кто оплодотворит больше самок, а значит оставит больше потомков, тогда как для самок конкуренция за то, какую из них оплодотворит больше самцов лишена практического смысла, потому что число потомков у самки зависит от числа яйцеклеток, а не от числа брачных партнеров. Самке выгоднее выбрать только одного самца, но самого лучшего с ее точки зрения. Главный вывод: обычно самки выбирают самцов, а вовсе не наоборот. Обсуждение вопроса о том, возможен ли половой отбор у гермафродитов. Рассматривается классический пример гермафродитизма, наблюдающийся у дождевого червя. Половое размножение дождевых червей основано на следующих принципах: Каждая особь является одновременно и самцом, и самкой, но физически не может оплодотворить сама себя – требуется брачный партнер. В отличие от чистого самца, такой гермафродит не может иметь подряд несколько брачных партнеров, потому что уже после первого скрещивания каждый становится матерью. Половые отношения в брачных парах дождевых червей абсолютно равноправны. Вывод: у таких гермафродитов как дождевой червь половой отбор принципиально невозможен – то есть пары сочетаются по случайному принципу. Таким образом половое размножение не обязательно сопряжено с половым отбором. РАБОТА В ВИРТУАЛЬНОМ ПРАКТИКУМЕ. Задача – показать, что при разных типах размножения мутации распространяются с разной скоростью, от которой зависти эволюционный успех. (25 мин.) Вводная информация о том, как устроено половое размножение в модели Виртуального практикума. • Брачные союзы формируются только между ближайшими соседями. • Гермафродитизм смоделирован по образцу дождевых червей. Пары образуются случайным образом. После спаривания каждый гермафродит становится матерью. Он потратил половину сил на исполнение функции отца и поэтому вдвое менее плодовит, чем особь, размножающаяся бесполым путем (без партнера) – у бесполой особи 31 может быть до 8 потомков (см. урок № IV), а при гермафродитизме – до 4 потомков (то есть до 8 на пару). • Раздельнополое размножение смоделировано следующим образом. После скрещивания у пары тоже получается до 8 потомков, но все они рождаются самкой. Зато самец тратит все силы на поиск самок на соседних клеточках поля. Один самец может иметь такой успех у самок, что обретет гарем (до 8 жен), а другой самец может быть, наоборот, отвергнут всеми соседками. Это и есть половой отбор. Самка выбирает одного из ближайших самцов по цвету. Можно задать шесть разных принципов выбора мужа, в том числе – с тенденцией к половому диморфизму и без нее. Условия эксперимента. К чисто черному полю будут адаптироваться, конкурируя за площадь, 4 поселения изначально чисто белых виртуальных организмов (по 40000 особей каждая), имеющие разные способы размножения. Частота мутаций у всех одинаковая – 0.01 % (в 10 раз больше, чем в имитации эволюции березовой пяденицы на уроке № IV). 1) Бесполое поселение будет располагаться под условным именем «тли» в левом нижнем углу поля «SW». 2) Гермафродитическое поселение будет располагаться под условным именем «черви» в левом верхнем углу поля «NW». 3) Раздельнополое поселение, настроенное на половой диморфизм, будет располагаться под условным именем «олени» в правом верхнем углу поля «NE». 4) Раздельнополое поселение, настроенное против полового диморфизма, будет располагаться под условным именем «НЕолени» в правом нижнем углу поля «SE». В этом эксперименте учащиеся впервые познакомятся с окнами «Полового состава». В них виртуальные организмы раскрашены следующими условными цветами: бесполые особи черные, гермафродиты голубые, самцы желтые, самки лиловые. Выполнение эксперимента. Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. В открывшейся заготовке будет 5 окон: 1) окно с черным полем, 2) окно с белыми стартовыми поселениями «тлей» и «оленей», 32 3) соответствующее окно Полового состава «тлей» и «оленей», 4) окно с белыми стартовыми поселениями «червей» и «НЕоленей», 5) соответствующее окно Полового состава «червей» и «НЕоленей». Запустив эволюционный процесс, сосредоточьте внимание вначале на двух окнах, где диагональные пары представлены в реальных цветах. Можно будет видеть, с какой скоростью всё более темные мутации распространяются в разных поселениях: 1) быстрее всего – у «оленей» (правый верхний угол), потому что они настроены на половой диморфизм, а значит каждая самка стремится выбрать самого непохожего на нее саму самца; 2) довольно быстро у гермафродитов «червей», у которых пары образуются случайным образом (левый верхний угол); 3) медленнее всего у лишенных рекомбинации бесполых «тлей» (левый нижний угол) и у раздельнополых «НЕоленей», настроенных против полового диморфизма (правый нижний угол). Следует обратить внимание на то, что у всех трех участников с половым размножением, в отличие от бесполых «тлей», в поселениях возникают мелкие пробелы (беспорядочные точки цвета светло-серой подложки). Это возникает там, где окрестные особи не нашли брачных партнеров и не оставили потомства. У бесполых такой проблемы не возникает, так как они способны размножаться даже в одиночестве. Довольно скоро все поселения настолько хорошо приблизятся к черному цвету, что на глаз их отличия будут неразличимы. Тогда за ходом конкурентной борьбы станет легче следить, глядя на два окна Полового состава. Точное соотношение сил отображается в счетчике очков. Автоматическая пауза произойдет на 300-й «год». К этому времени все важные явления уже произойдут. Обязательно нужно несколько раз повторить описанный выше эксперимент. Сколько раз Вы щелкнете по этой мушке-дрозофиле – столько дублей этого эксперимента и будет запущено. Разумеется – в ущерб скорости работы программы. Поэтому не забывайте выключать ненужные окна программы. 33 Выводы: • Чаще всего побеждают «олени» из правого верхнего угла, потому что при настройке на половой диморфизм мутации распространяются быстрее всего, а значит и приспособительная эволюция имеет более высокий темп. • Но это происходит не всегда – результаты конкурентной борьбы четырех «противников» в разных экспериментах могут существенно отличаться. Это зависит от везения с появлением лучших мутаций, которые появляются случайно (это уже было доказано на уроке № IV). В заключение урока можно предложить ученикам выдвинуть гипотезы о том, почему у «оленей» в конце эксперимента получаются одинаковые черные самцы и самки, несмотря на то, что они настроены на половой диморфизм. Правильное объяснение состоит в том, что естественные отбор в направлении покровительственной окраски здесь преобладает над половым отбором. Таким образом, и в живой природе отсутствие внешних различий между самцами и самками у многих животных вовсе не значит, что у них (в отличие от оленей и баранов) отсутствует половой отбор с тенденцией к диморфизму. 34 Урок VII: «Половой отбор и эволюционное значение разных способов размножения» (продолжение для профильного уровня). X-XI классы – тема «Вид», подтема «Элементарные факторы и движущие силы эволюции; их взаимосвязь». Содержательные задачи – (1) количественно изучить влияние разных способов размножения на скорость эволюции; (2) показать относительность их преимуществ друг перед другом; (3) выяснить смысл чередования полового размножения с бесполым. Методические задачи – (1) овладеть на практике принципами взаимодействия всех пройденных на предыдущих уроках эволюционно значимых параметров; (2) использовать Виртуальный практикум в игровом режиме для повышения мотивации и инициативности учащихся. На данном уроке необходимо выполнить большое количество экспериментов, поэтому для его проведения желательно иметь не менее трёх компьютеров в классе. Турнир можно перенести на внеурочное время. КОЛИЧЕСТВЕННЫЙ ЭКСПЕРИМЕНТАЛЬНЫЙ АНАЛИЗ РАЗНЫХ СПОСОБОВ РАЗМНОЖЕНИЯ (15 мин.) Будут попарно сравниваться в точности те же самые варианты размножения, которые демонстрировались на предыдущем уроке: 1) бесполые «тли», 2) гермафродитические «черви», 3) раздельнополые «олени», настроенные на половой диморфизм, 4) так называемые «НЕолени» – тоже раздельнополые, но настроенные против полового диморфизма. Для сравнения будет использоваться специальный стенд в виде шахматного поля размером 3х10 клеток. Каждая клетка имеет, в свою очередь, размер 50х50 пикселов, то есть на ней может разместиться до 2500 виртуальных организмов. Две белые клетки шахматного поля будут заселены стартовыми поселениями организмов серого цвета, как показано на рисунке: 35 На каждом компьютере учащийся (или группа учащихся) должны провести серию из шести экспериментов и занести их результаты в следующую турнирную таблицу: «тли» «черви» «олени» «НЕолени» «тли» «черви» «олени» «НЕолени» Счет каждого «поединка» измеряется округленным числом шахматных клеток, занятых поселениями «SW» и «NW» к концу эксперимента, который длится 600 лет. Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. В открывшейся заготовке будет представлено пять окон: 1) окно с шахматным полем, 2) окно с серым стартовым поселением «NW» (левый верхний угол), 3) окно с серым стартовым поселением «SW» (левый нижний угол), 4) окно Полового состава поселения «NW», 5) окно Полового состава поселения «SW» (условные цветовые обозначения полов уже знакомы по предыдущему уроку: бесполые особи черные, гермафродиты голубые, самцы желтые, самки лиловые). Теперь требуется задать разные способы размножения в поселениях «SW» и «NW». Для этого надо в Панели управления каждого поселения загрузить один из четырех файлов настроек – тли.ini, или черви.ini, или олени.ini, или НЕолени.ini, но с условием, что одному поселению будут присвоены настройки из одного файла, а другому – из другого. 36 После запуска эволюционного процесса пауза автоматически произойдет через 600 виртуальных лет. Тогда надлежит подсчитать (округленно) число шахматных клеток, занятых тем и другим поселением, и занести этот результат в турнирную таблицу. Чтобы провести следующий эксперимент желательно закрыть предыдущее окно Виртуального практикума и еще раз запустить ту же задачу, щелкнув по этой мушкедрозофиле – но теперь загрузить двум поселениям уже другое сочетание из четырех указанных файлов настроек. В результате наблюдения экспериментов и заполнения турнирных таблиц учащиеся приходят к следующим выводам: • как правило два поселения, распространяясь из своих исходных углов «SW» и «NW», достаточно четко делят между собой черные и белые шахматные клетки, то есть выбирают разные экологические ниши; но иногда какое-то одно из конкурирующих поселений вымирает совсем, отдавая клетки обоих цветов противнику; • результаты одинаковых экспериментов (например, «черви против оленей») могут сильно различаться; это связано не с тем, которому из поселений «SW» и «NW» были назначены те или иные настройки, а с принципиальной случайностью мутационного процесса, то есть с везением; • не существует однозначной линейной иерархии в плане преимущества одного способа размножения над другим – скорее их соотношения напоминают всем известную детскую игру «камень-ножницы-бумага»; • в данном стендовом эксперименте, в отличие от эксперимента, проведенного на предыдущем уроке, чаще побеждают «тли»; таким образом, главный недостаток бесполого размножения – отсутствие рекомбинации – перекрывается следующими двумя преимуществами: 1) бесполый организм может размножаться даже в одиночестве; 2) бесполый организм не расходует сил на выполнение функции самца и потому может дать в каждом помете столько же потомков (в данной модели – до 8), сколько пара организмов при половом размножении – будь они гермафродиты или раздельнополые. Встает вопрос для следующей части урока – в рамках шахматного стенда подобрать такие параметры полового размножения, чтобы устойчиво побеждать «тлей». 37 ПОИСК ОПТИМАЛЬНОГО СПОСОБА РАЗМНОЖЕНИЯ. (10 мин.) Опять запустите шахматный стенд, щелкнув по этой мушке-дрозофиле. Загрузите для одного поселения настройки бесполого размножения из файла тли.ini, а для другого поселения попробуйте подобрать какой-нибудь более выгодный вариант, произвольно присваивая в Панели управления любые параметры в разделе «размножение». При дефиците времени можно просто продемонстрировать учащимся два заготовленных варианта настроек, присвоив их через Панель управления тому поселению, которое противостоит «тлям», и пояснив суть этих настроек: 1) Файл практический_взгляд_самки.ini обеспечивает самкам более практичный выбор мужа, чем у «оленей». Здесь каждая самка ищет среди самцов уже не самого отличающегося по цвету от нее самой, а такого, у которого цвет больше всего похож на цвет той клеточки поля, на которой данная самка проживает. Получается, что она примеривает самцов к своей клеточке на предмет скрывающей окраски, а в результате и ее потомство скорее будет покровительственных цветов. В реальном мире есть много примеров тому, как самки оценивают самцов по их «рабочим качествам». 2) Файл ответ_тлям.ini содержит два других отличия от параметров «оленей», обеспечивающие повышение числа «плодоносящих» особей: (1) доля самцов понижена с обычных 50 % до 40 %; (2) назначено «10 % клонирования» вместо 0 %. Это означает, что каждая особь каждый год, приступая к размножению, с вероятностью 10 % отказывается от скрещивания и размножается сама по себе. В действительности для реальных тлей характерно как раз подобное чередование полового и бесполого размножения, а не «100 % клонирования», как это приписано нашим виртуальным «тлям». Второй вариант поможет учащимся понять, почему в Природе так много живых организмов совмещают половое и бесполое размножение: половое служит для рекомбинации, а бесполое – для плодовитости. Учащиеся должны сами догадаться, что эти два варианта настроек можно совместить – получится еще выгоднее. 38 ТУРНИР МЕЖДУ УЧЕНИКАМИ ПО ЭВОЛЮЦИОННЫМ ШАХМАТАМ. (20 мин.) Запустите попарный_турнир, щелкнув по этой мушке-дрозофиле. Откроются те же знакомые окна с шахматным полем, двумя серыми стартовыми поселениями «SW» и «NW» и окна с их половым составом. Настройки обоих поселений уже хорошо подобраны: 1) У поселения «NW» (в левой верхней шахматной клетке) размножение настроено как в применявшемся в предыдущей части урока файле настроек практический_взгляд_самки.ini, но частота мутаций усилена впятеро – до 0.05 %. 2) У поселения «SW» (в левой нижней шахматной клетке) размножение настроено как в применявшемся в предыдущей части урока файле настроек ответ_тлям.ini, но частота мутаций усилена втрое – до 0.03 %, а предел долголетия повышен до 25 лет (проверенные естественным отбором «старики» повышают стабильность поселения). Однако, эти настройки есть куда улучшать. За каждое поселение (от каждого угла) может играть один ученик, но рекомендуется играть «команда на команду» – это стимулирует открыто обсуждать и аргументировать дальнейшие действия. Основные действия сторон будут состоять в подборе наилучшего сочетания нескольких параметров в Панели управления своего поселения, а именно: • всех параметров в разделе «размножение» – как в предыдущей части данного урока; • частоты мутаций – как на уроке № IV; • предела долголетия, затронутого на уроке № V; • периода невмешательства. «Период невмешательства» задает, через сколько лет игрок решил вмешаться в свои настройки в следующий раз. Часто вмешиваться выгоднее, чтобы не терять контроль над эволюционным процессом. С другой стороны, за более долгое невмешательство начисляется больше очков при росте поселения, и больше вычитается – при его убыли. Большой период невмешательства – это высокая игровая ставка. То, какую величину игрок назначает периоду невмешательства, отражает его азарт или уверенность в своей способности прогнозировать ситуацию, а то, какой от этого у него реально получается прирост очков (или убыль), помогает корректировать самооценку и исправлять величину периода невмешательства при следующем доступе к параметрам. Таким образом, в турнире надо внимательно следить за Счетчиком лет и Счетчиком очков. 39 Чтобы успеть провести в рамках данного турнира побольше поединков, рекомендуется не дожидаться абсолютной победы (полного вымирания одного поселения), а прекращать поединок по истечение 1000, в крайнем случае 1500 лет от старта, присуждая победу по очкам. Победоносные варианты настроек ученики могут сохранить на будущее в виде ini-файлов. Пока учащиеся соревнуются, можно рассказать об ограниченности модели размножения в Виртуальном практикуме. Объяснение. Помимо того, что вариантов бесполого, гермафродитического и раздельнополого размножения в живой природе – великое множество, есть еще одно крайне важное обстоятельство. Виртуальный практикум позволяет «столкнуть лбами» разные системы размножения (чем в данный момент и заняты учащиеся), но не моделирует такие процессы, как происхождение раздельнополости от гермафродитизма или наоборот. Ведь одно дело – межвидовая борьба раздельнополых организмов и не скрещивающихся с ними гермафродитов (что в данный момент наблюдается на некоторых компьютерах в классе), и совсем другое дело – появление раздельнополых особей в недрах гермафродитной популяции за счет внутривидовой борьбы, которую Дарвин всегда ставил на первое место. Можно вообразить такую природную ситуацию, когда гермафродитный вид после победы над раздельнополым видом сам потом становится раздельнополым. Представьте, что будет, если в популяции дождевых червей заведется червяк, не производящий яйцеклеток: все нормальные черви будут после спаривания честно заниматься деторождением, а этот неотягощенный яйцеклетками «хитрец» поползет оплодотворять следующего соплеменника, потом следующего, и так далее. В результате множественных скрещиваний он передаст свои «аномальные» гены, подавляющие образование яйцеклеток, большему числу потомков, чем ортодоксальные гермафродиты. А когда его сыновья в достаточной степени заполонят популяцию, у оставшихся «честных» гермафродитов автоматически наступит девальвация мужской функции, так как при спаривании с новоявленными самцами сперматозоиды гермафродита уходят впустую, не встретив яйцеклетки у партнера. Следовательно, эффективными станут теперь уже уклонения в сторону «чистой самки», вообще не производящей сперматозоидов – тем более, что такая особа обретает свободу рационально выбирать мужа, ибо стремление спариваться со всеми пропадает вместе со сперматозоидами. 40 Урок VIII: «Принцип дивергенции и экологическое видообразование». X-XI классы – тема «Вид», подтема «Критерии вида (морфологический и биологический). Образование новых видов. Способы видообразования (географическое и экологическое видообразование)». Содержательные задачи – убедиться, что гибридизация не препятствует дивергенции видов; (2) изучить влияние способа размножения на характер процесса видообразования. Для восприятия сложного материала этого занятия ученики должны уже знать и повторить к этому уроку различие морфологического и биологического критериев вида, значение репродуктивной изоляции для видообразования и биологический смысл конъюгации хромосом (урок № II). ОБЪЯСНЕНИЕ ПРИНЦИПА ДИВЕРГЕНЦИИ (5-10 мин.) Дарвин изучал на Галапагосских островах роль географической изоляции для видообразования. Но он также считал, что и без изоляции вид имеет внутреннюю тенденцию к саморазделению. Цитата из Дарвина: «… чем более потомки какого-нибудь вида будут различаться между собою строением, общим складом и привычками, тем легче они будут в состоянии завладеть более многочисленными и более разнообразными местами в экономии природы, а следовательно тем легче будут размножаться.» Последние слова относятся не к скрещиванию между видами, а к росту суммарного поголовья поселения. Суть экологического видообразования сводится к следующему: по мере того, как разные представители первоначально единого вида по-разному приспосабливаются к среде, завладевая разными местами в «экономии природы» (экологическими нишами), они всё больше уклоняются друг от друга (дивергируют), и это расхождение в «строении и привычках» косвенным образом приводит вначале к частичной, а потом и к полной утрате способности разошедшихся представителей давать потомство при скрещивании. Такой процесс видообразования проходит различные «степени бесплодия» (термин Дарвина). Есть примеры видов, которые еще не утратили способность скрещиваться. 41 ПРИМЕР ВОЛКОВ И СОБАК (5-10 мин.) Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «псовые», а финиш – на иконку «серый волк». Остается проиграть этот маршрут. Объяснение. Первые псовые появились около сорока миллионов лет назад в Северном полушарии. Их небольшой размер и повадки охотника-одиночки продержались поныне в эволюционной линии лисиц. И представители линии волков тоже долгое время оставались такими. Но для охоты на добычу превосходящего размера им потребовалось перейти к групповой охоте, да и в собственном размере сильно прибавить. Сейчас серый волк достигает восьмидесяти килограммов, а в прошлом бывали и более крупные виды! Он всегда имел широчайший ареал распространения по всему северному полушарию и не терял тесного контакта с другими представителями рода. Скорее всего домашняя собака произошла именно от серого волка, но так же как с ним, она дает потомство с шакалом и с американским койотом, причем потомство плодовитое (в отличие от мулов – см. урок № II). Таким образом, все эти виды сохраняют генетическую связь. Из-за этого мелкому эфиопскому волку (отдельный вид) даже грозит опасность «раствориться» в местной популяции пастушеских собак. Волки, собаки, шакалы и койоты задержались примерно на той стадии разобщения, которая здесь отмечена красной чертой на знаменитой схеме дивергенции Ф.Г. Добжанского (один из основоположников «биологической концепции вида»). Виртуальные организмы по причине своей примитивной генетической конституции никогда не достигают полной нескрещиваемости, то есть в этом плане не могут зайти намного дальше волков, собак, шакалов и койотов. 42 ВИДООБРАЗОВАНИЕ В ВИРТУАЛЬНОМ ПРАКТИКУМЕ (30 мин.) Вводная информация об этой части модели. Чтобы заработал экологический механизм «экономии природы» в следующих примерах, в отличие от использовавшихся на всех предыдущих уроках, виртуальным организмам присвоена способность питаться. Способ питания виртуальных организмов станет более понятен на следующем уроке, а здесь достаточно знать, что виртуальные организмы питаются цветом клеточек поля, на которых они живут. Таким образом цвет поля будет теперь меняться по мере его выедания. Сам он в свою очередь имеет тенденцию восстановиться до исходного состояния (как трава вновь отрастает на пастбищах рогатого скота). На некоторых местах поля цвет может оказаться не таким, который пригоден в пищу виртуальным организмам, и там они вымирают от голода, как в пустыне. Модель построена таким образом, что биологический критерий вида и морфологический тождественны – плодовитость пары организмов строго зависит от их разницы по цвету. Каждый «год» программа вычисляет видовой состав путем поголовного попарного сравнения организмов по принципу: две особи относятся к одному виду, если они в принципе могли бы дать полное потомство (хотя реально они могут вообще не вступить в контакт – либо потому что живут не рядом, либо потому что оба одного пола или вообще бесполые, либо потому что самка предпочтет другого самца). Плодовито или не плодовито потомство, определяется тем, достаточно ли сходны у сравниваемых особей хромосомы (например с геном красного цвета), смогут ли эти хромосомы «узнать друг друга» и конъюгировать – иначе плодовитость будет меньше нормальной. Порог узнавания/неузнавания хромосом задается в Панели управления регулятором «порог конъюгации». В экспериментах предыдущих уроков разделение поселения на виды не происходило, потому что был назначен максимальный порог конъюгации (255), при котором хромосомы всегда узнавали друг друга и правильно конъюгировали. Преемственность между видами года нынешнего и года предыдущего определяется посредством подсчета – сколько каждому нынешнему виду досталось генов от каждого прошлогоднего вида. Демонстрация видообразования. В данном примере порогу конъюгации присвоено значение 15, а настройки размножения – те же, которые использовались на уроках №№ VI и VII под кодовым названием «олени» (самки настроены на половой диморфизм). 43 Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. В открывшейся заготовке будет 5 окон: 1) поле – уже знакомая картина А.К. Саврасова «Домик в провинции»; 2) стартовое поселение «NE» в правом верхнем углу – опять же уже использовавшаяся на этом поле старинная фотография охотников в тонах сепии (это было на уроке № V, но с другими настройками); 3) окно, представляющее поле в трехмерном RGB-кубе; 4) окно, представляющее поселение «NE» в RGB-кубе; 5) окно происхождения видов в котором вначале имеется только один квадратик, обозначающий стартовое поселение «NE»; в дальнейшем в нем будут отображаться виды численностью не менее 1000 особей. При запуске эволюционного процесса будут наблюдаться следующие явления: Цвет поля будет меняться, потому что виртуальные организмы стали им питаться. Соответствующие изменения происходят и в окне с трехмерным представлением палитры поля – облако точек расползается и образуются новые сгущения. Это своего рода экологические ниши, созданные благодаря пищедобывательной активности самих же организмов. К этому видоизмененному полю организмы теперь и адаптируют свою покровительственную окраску. В основном окне, где организмы представлены в реальном цвете, в некоторых местах обнажается цвет подложки. Это для них пустыня, где не хватает еды. В трехмерном представлении поселения, в разросшемся облаке точек бывшей фотографии охотников появляются характерные щели и проёмы – это границы между видами. Окно происхождения видов начинает бежать как самописец, и в нем на глазах растет и ветвится микроэволюционное древо. Его содержимое будет выглядеть примерно как на следующем рисунке. Обратите внимание на сетчатую структуру связей – как на знаменитой гипотетической схеме Добжанского, показанной выше. 44 Вверху над каждым столбцом кружочков написан номер «года» с момента старта; каждый кружочек – это вид, а цвет его заливки – это усредненный цвет всех особей вида; каждая линия – это связь генетической преемственности; толстые черные линии обозначают преемственность свыше 50 %, а тонкие серые – все более слабые связи; в каждом кружочке имеется 4 числа: 1) верхнее число представляет численность особей в данном виде; 2) правое число – это процент сеголетков среди них; 3) левое число – это точное значение (%) главной связи с предком, то есть толстой черной линии подходящей к кружочку слева; 4) нижнее число – это сколько последних лет подряд главная линия преемственности превышала 90 %. Последние два числа представляют собой показатели обособленности вида – чем она выше, тем числа больше. Когда через 400 лет произойдет автоматическая пауза, следует сохранить результат – его понадобится загрузить на следующем уроке. Как обычно, эти эксперименты надо повторить несколько раз – чтобы отсеять закономерное от случайного. Повторно запускайте ту же самую заготовку, щелкая по этой мушке-дрозофиле. 45 Обсуждение результатов видообразования у «оленей». Настроенность самок на половой диморфизм должна приводить к тому, что они будут при возможности выбирать самцов не из своего, а из других видов. В природе есть тому пример – скрещивание тетеревов и глухарей (разные роды!). Возможно это и связано с их крайне выраженной тенденцией к половому диморфизму (их гибрид называется «межняком»). Вопрос: почему такая тенденция к смешению видов и в Природе, и в Виртуальном практикуме не сводит на нет дивергенцию видов? Ответ: при экологическом видообразовании тоже возникает некоторое пространственное отчуждение видов, хотя и без столь явных внешних барьеров, как при географическом видообразовании. Тому есть три причины: 1) Разные виды занимают разные «места в экономии природы» (экологические ниши), так что самки имеют доступ к чужим самцам только на границах соприкосновения этих мест. 2) Гибридные скрещивания имеют пониженную плодовитость, так что в следующем поколении они все равно сказываются слабее, чем внутривидовые. 3) Они сказываются слабее еще и потому, что гибридный потомок скорее всего получится хуже приспособленным как к экологической нише вида, к которому принадлежала его мать, так и к нише отцовского вида, а это сулит проигрыш в борьбе за существование с «чистокровками». Исследовательская задача. Теперь надо выяснить экспериментально, как влияет тип размножения на характер ветвления древа видов. Для сравнения с уже полученным в предыдущем опыте видообразованием у «оленей» будут использоваться еще 3 варианта настроек размножения, которые на уроках №№ VI и VII получили кодовые названия «НЕолени» (самки настроены против полового диморфизма), «черви» (гермафродиты) и «тли» (бесполые). Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле, а дальше загружайте через Панель управления настройки из файлов виды_у_НЕоленей.ini или виды_у_червей.ini или виды_у_тлей.ini. Для каждого варианта надо повторить опыт несколько раз. 46 Главное, за чем следует наблюдать в окне происхождение видов – это за количество тонких серых линий слабой преемственности, отражающих степень сетчатой гибридизации между видами. Как и следует ожидать, варианты размножения выстроятся по убыванию сетчатости в следующем порядке: 1) «олени» с их тенденцией к гибридизации имеют наиболее густую сеть связей между видами; 2) «черви» со случайным выбором партнера несколько менее перепутаны; 3) «НЕолени», настроенные против диморфизма, а значит и гибридизации, имеют мало серых соединений; 4) «тли», у которых в силу бесполого размножения вообще нет гибридизации, лишены сетчатости – каждый клон это обособленный вид. Однако, у «тлей» все же изредка наблюдаются тонкие серые связи. Вопрос: как это может быть без скрещивания клонов? Ответ: иногда мутации могут привести к конвергентному появлению почти неотличимых клонов, но для этого требуется не только соответствующая мутация, но и сходство условий существования – чтобы естественный отбор такую мутацию поддержал. 47 Урок IX: «Принцип дивергенции» (продолжение). X-XI классы – тема «Вид», подтема «Критерии вида (морфологический и биологический). Образование новых видов. Способы видообразования (географическое и экологическое видообразование)». Содержательные задачи – (1) экспериментальное доказательство Дарвиновского принципа дивергенции; (2) экспериментальное выяснение приспособительного значения холоднокровности и теплокровности. Методическая задача – освоение возможностей конструирования эволюционного древа виртуальных организмов. ПОСТАНОВКА ЗАДАЧИ (5 мин.) Дарвин объяснял принцип дивергенции так: «Остановимся на примере хищного четвероногого, численность которого достигла, в среднем, того предела, который в состоянии вместить данная страна. Если действию его естественного стремления к размножению будет предоставлен простор, то это размножение может осуществиться на деле (предполагая, что физические условия страны остаются те же), только если его изменившиеся потомки захватят места, занятые другими животными: а это может быть достигнуто или тем, что они приобретут возможность питаться новым родом добычи, живой или мертвой, или тем, что они научатся обитать в новых условиях, лазать на деревья, жить в воде, или тем наконец, что они сделаются менее плотоядными. Чем разнообразнее в своих привычках и строении окажутся эти потомки нашего хищника, тем более будет число мест, которыми они завладеют.» На язык наших виртуальных организмов это переводится следующим образом: «Возьмем пример, когда численность виртуальных организмов достигла, в среднем, того предела, который в состоянии вместить данное поле. Их естественное стремление к размножению может осуществиться на деле (предполагая, что физические условия на поле остаются те же), только если изменившиеся потомки захватят новые клеточки: а это может быть достигнуто тем, что они приобретут возможность питаться цветом поля по новому. Чем разнообразнее в своих пищевых привычках окажутся эти потомки, тем более будет число клеточек на поле, которыми они завладеют.» Это утверждение требуется экспериментально доказать. 48 В качестве примера, «когда численность виртуальных организмов достигла, в среднем, того предела, который в состоянии вместить данное поле» в Виртуальный практикум загружается заготовка, сохраненная на предыдущем уроке № VIII. В окне, представляющем виртуальные организмы в реальном цвете (оно озаглавлено просто «NE») хорошо видно, что некоторые места на поле остаются не заняты – там просвечивает цвет подложки. Там пустыня. Конкретная задача урока состоит в том, чтобы заселить эту пустыню виртуальными организмами, подобрав им подходящие настройки питания. СТРАТЕГИЯ ЗАСЕЛЕНИЯ ГОЛОДНЫХ МЕСТ (10 мин.) На прошлом уроке уже говорилось, что виртуальные организмы питаются цветом клеточек поля, на которых они проживают. Чтобы заселить голодные места, требуется изменить пищевые предпочтения виртуальных организмов, и тогда цвет этих мест поля будет восприниматься уже как достаточное количество пищи. Но такое изменение чревато тем, что пустынями, наоборот, станут ранее заселенные части поля. Поэтому выход один – разным группам виртуальных организмов присвоить разные пищевые предпочтения. Для этого Виртуальный практикум предлагает следующую возможность. Когда Панель управления доступна для редактирования, разрешается выбрать в Панели навигации какойнибудь один вид и возвести его в ранг рода. Тогда он потеряет скрещиваемость со всеми остальными видами, но зато у Вас появится возможность дать новому роду отдельные настройки – и порога конъюгации хромосом, и частоты мутаций, и способа размножения, и долголетия и питания. Пищевая специализация регулируется параметрами условной «пищевой точки». Можно выбрать любой цвет и «приказать», чтобы данный род виртуальных организмов считал его за предел изобилия или, наоборот, за пустыню. Картина радикально поменяется. Второй параметр питания – прожорливость. Он просто задает количество цвета, которое каждый пиксел должен съесть за год, чтобы не умереть с голоду. Прожорливость требует пояснения на реальных биологических примерах. Объяснение. Холоднокровное пресмыкающееся потребляет пищи в 5-8 раз меньше, чем теплокровное млекопитающее того же размера (уместно сравнить варана острова Комодо и южноамериканского ягуара по 100 кг каждый). Из-за низкой прожорливости холоднокровные могут жить там, где теплокровным не хватает еды. Но там, где еды достаточно и теплокровным тоже, борьбу за существование при равной приспособленности выигрывают именно теплокровные, потому что насколько они прожорливее, настолько и 49 активнее. То есть теплокровное существо может быть приспособлено к местным условиям даже несколько хуже, но за счет общей подвижности все равно выиграет у холоднокровного. Этому есть известный пример на Галапагосских островах, где завезенные и одичавшие козы нанесли сильнейший урон местным слоновым черепахам – они быстро сжирают зелень буквально из-под носа у медлительных пресмыкающихся, хотя те в принципе лучше приспособлены к родным условиям. Однако, в Природе на самом деле нет резкой границы между теплокровностью и холоднокровностью. Существует много градаций. Например, уровень обмена веществ утконосов и ленивцев – промежуточный между обменом пресмыкающихся и большинства млекопитающих. Прожорливость сумчатых выше, чем у утконоса, но ниже, чем у плацентарных. А воробьиные имеют еще более высокий уровень обмена веществ, чем остальные птицы. РЕШЕНИЕ ЭКСПЕРИМЕНТАЛЬНОЙ ЗАДАЧИ (30 мин.) Подготовительные операции. Запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле. Загрузите заготовку, сохраненную на предыдущем уроке – откроются все окна, которые там были открыты на момент сохранения. Открыв Панель управления поселением, найдите в Панели навигации многочисленный вид белёсого цвета, ареал которого окружает пустыню над крышей «Домика в провинции». Такой вид обязательно будет присутствовать в заготовке благодаря закономерностям эволюционного процесса, демонстрировавшегося на прошлом уроке. Он будет находиться где-то в начале списка видов и будет выглядеть в окне просмотра примерно вот так: 50 Создайте из этого вида новый род под удобным для себя именем, например, «белые». Чтобы дальше внимательно наблюдать за новым родом, надо открыть для него отдельное окно «Фенотип» – оно будет выглядеть примерно как предшествующий рисунок. Закройте лишние окна, если они были. Оставьте только 4 окна (если все было сделано по инструкции, они уже должны быть на экране): 1) только что созданное окно с новым родом, 2) окно со всем поселением целиком, 3) окно с полем, 4) окно происхождения видов. Теперь надлежит, не запуская эволюционный процесс, просто сохранить достигнутый результат – точно так же, как это делалось на предыдущем уроке. Это требуется для того, чтобы при повторных экспериментах не тратить каждый раз время на описанные выше подготовительные операции, а загружать заготовку уже с созданным родом. После сохранения можно начать эксперименты. 51 Эксперимент № 1. Открыв Панель управления поселением, найдите в Панели навигации вновь созданный род и переставьте ему метку «пищевой точки» с изобилия на пустыню. После присвоения этого изменения пищевые предпочтения рода поменяются на прямо противоположные по отношению к остальным видам поселения, и бывшая пустыня станет для организмов этого рода кормным местом. Запустите и наблюдайте эволюционный процесс: В окне с родом можно будет видеть быстрое заселение пустыни. В окне «Происхождение видов» – в его нижней части – можно будет со временем увидеть, что род, потеряв скрещиваемость с остальными видами поселения, стал уже сам разделяться на новые виды. Они отделены от остальных видов поселения увеличенным проемом, как показано на рисунке. Эксперимент № 2 начинается как 1-й эксперимент; разница в настройках нового рода. Опять запустите Виртуальный практикум, щелкнув по этой мушке-дрозофиле, и загрузите тот же сохраненный результат создания рода – откроются все те же окна, включая окно с новым родом. 52 Открыв Панель управления поселением, найдите в Панели навигации тот же род «белых», но измените ему настройки иначе, чем в 1-м эксперименте. В этот раз надо оставить его «пищевую точку» в статусе изобилия, а вместо этого резко уменьшить прожорливость, например в 10 раз – была равна 90, а надо сделать 9. Таким образом этот род станет по сравнению с остальными организмами холоднокровным и сможет питаться там, где другим пищи не хватает, хотя он и сохранит те же пищевые предпочтения. Запустите и наблюдайте эволюционный процесс. Результат будет в принципе таким же как в 1-м эксперименте – пустыня будет успешно заселена новым родом. Общий вывод. Приведенные в начале урока теоретические рассуждения Дарвина полностью подтвердились в виртуальных экспериментах – страна может прокормить большее число организмов, если они имеют разнообразные потребности и предпочтения. Эксперимент № 3 (в случае нехватки времени задается на дом). Ученикам предлагается самостоятельно испробовать третий способ заселения пустыни тем же самым родом – не меняя ни прожорливости (оставив 90), ни статуса «пищевой точки» (оставив ее в качестве изобилия), а присваивая этой «пищевой точке» новый цвет. Ученик опять запускает Виртуальный практикум и загружает тот же сохраненный результат создания рода. Затем ему надо, открыв Панель управления поселением, найти в Панели навигации тот же род «белых» и присвоить тому новый цвет «пищевой точки». Запустив эволюционный процесс, ученик проверяет, помогло ли роду «белых» заселить пустыню его вмешательство. При неудаче можно повторить эксперимент. Правильным действием будет выставить ползунки так, чтобы подобрать цвет «пищевой точки» как можно ближе к оригиналу картины А.К. Саврасова в том месте, где получилась пустыня, и назначить этот цвет для рода «белых» изобилием. Если учитель сочтет, что данный урок перегружен, можно обойти весь этап подготовительных операций и настроек, запуская прилагающиеся заготовки тех же экспериментов: 53 Загрузите готовый Эксперимент № 1 , щелкнув по этой мушке-дрозофиле. Род уже создан, настроен и открыт в своем окне. Останется только запустить эволюционный процесс и наблюдать. Загрузите готовый Эксперимент № 2 , щелкнув по этой мушке-дрозофиле. Род уже создан, настроен и открыт в своем окне. Останется только запустить эволюционный процесс и наблюдать. 54 Урок X: «Симбиоз. Экспериментальная контрольная работа». X-XI классы – тема «Экосистемы», подтема «Конкуренция, симбиоз». Данное занятие предлагается в качестве итогового по теме «Симбиоз». Предшествующие уроки по этой теме должны были проводиться традиционным способом, а здесь оценивается достигнутое учениками понимание сути явления. Ставится практическая задача – доказать симбиотический характер взаимоотношений между двумя поселениями виртуальных организмов. ОБЪЯСНЕНИЕ ЗАДАЧИ (10 мин.) Итак, далеко зашедший симбиоз (мутуализм) – это когда разные организмы практически не могут жить друг без друга. Один из встречающихся в Природе вариантов такого рода обоюдной зависимости опосредован пищевыми ресурсами. Он наиболее ярко проявляется между копытными африканской саванны. Сейчас это настолько «сыгранная команда», что даже трудно вообразить, что ее «игроки» имеют совершенно разное происхождение. Демонстрация: общий предок белого носорога и жирафа. Запустите Анимированный атлас, щелкнув по этому слону. Старт установлен на иконку «белый носорог», а финиш – на иконку «жираф». Остается проиграть этот маршрут. Попросите учеников прокомментировать общего предка жирафа и носорога. Мог бы он по их мнению найти достойную экологическую нишу среди современных африканских стад? Одни из копытных африканской саванны питаются травой (как белый носорог и зебра), а другие – побегами и листьями кустов и деревьев (как черный носорог и жираф). Когда первые долго пасутся на одном и том же месте, они почти полностью выедают траву и вытаптывают дёрн – тогда на этом месте вырастают кустарники и деревья. Их находят копытные второй категории, и со временем тоже выедают. Тогда здесь вновь вырастает трава, и возвращаются травоядные. И так далее… Такая постоянная циркуляция типов растительности и копытных возможна благодаря двум условиям: 1) прямо противоположные пищевые предпочтения участвующих животных (белому носорогу почти нечего делать в кустарнике, а жирафу – на траве); 55 2) податливость среды обитания (если бы одни не могли уничтожить траву, а другие – кусты, возобновление их излюбленного пищевого ресурса не требовало бы вмешательства другой стороны). В Виртуальном практикуме эти условия легко смоделировать следующим образом: 1) Пускай, например, для организмов одного поселения белый цвет поля будет изобилием пищи, а для организмов другого поселения – пустыней. Соответственно, к черному у них будет обратное отношение. (Подобный вариант противоположных настроек «пищевой точки» уже был освоен на уроке № IX, хотя там она была и не белой). 2) Цвет поля не должен восстанавливаться сам собой после того, как его съели виртуальные организмы. Для этого в Панели управления параметру «подрост» присвоено нулевое значение. В этой задаче менять его на другое значение запрещено, потому что тогда симбиотические отношения между поселениями распадутся. Любые другие вмешательства разрешаются. Условия задачи. На ровное серое поле в противоположные углы загружены два поселения черных организмов по 10000 особей. Те, которые в левом нижнем углу (поселение «SW»), воспринимают белый цвет как изобилие, а для тех, что в правом верхнем углу (поселение «NE») – это цвет пустыни. Требуется понаблюдать за процессом, а когда наступит автоматическая пауза – активно вмешаться, чтобы каким-то образом доказать, что за время эксперимента эти организмы вступили в симбиотические отношения наподобие копытных из саванны. РЕШЕНИЕ КОНТРОЛЬНЫХ ЗАДАЧ (35 мин.) Задача предлагается в двух вариантах – для общего уровня и для продвинутых учеников. Вариант А (простой). Ученик должен выполнить следующие действия: Запустить Виртуальный практикум, щелкнув по этой мушке-дрозофиле. В открывшейся заготовке будет 4 окна: 1) серое поле; 2) стартовое поселение в левом нижнем углу; 56 3) стартовое поселение в правом верхнем углу; 4) окно полового состава с двумя поселениями сразу; организмы из левого нижнего угла «SW» представлены смесью желтых пикселов (условный цвет самцов) и лиловых (условный цвет самок), а организмы из правого верхнего угла «NE» – голубыми (условный цвет гермафродитов). После запуска эволюционного процесса будут наблюдаться следующие явления: Цвет поля будет податливо меняться: где жили организмы поселения «SW» – в сторону черного (для них это пустыня), а где жили организмы поселения «NE» – в сторону белого (это пустыня для них). Сами организмы не будут менять своего черного цвета, потому что мутаций им в этом варианте не назначено. В окне полового состава будет виден процесс крупномасштабной циркуляции. Именно для этой наглядности одни сделаны раздельнополыми, а другие – гермафродитами. Пока ученики наблюдают этот процесс, учитель может сделать разъяснение по поводу отличия циркуляции африканских копытных и виртуальных организмов. Разница в следующем. Копытные широко перемещаются в течение своей жизни, ища лучшие пастбища, а виртуальные организмы совершенно оседлы, так что у них циркуляция происходит не при помощи ног, а посредством размножения – потомки расселяются вокруг клеточки своей матери (см. урок № IV). По истечение 1000 «лет» процесс автоматически остановится, и ученику надо будет решать – что сделать, чтобы доказать симбиоз поселений. Учитель должен иметь в виду, что в данной задаче по случайному стечению обстоятельств (а эволюции без случайности не бывает) рано или поздно обязательно произойдет вымирание одного поселения, а вслед за ним – и другого. Если это случится в течение установленных 1000 «лет», то ученику придется повторить эксперимент с начала. Но для него это будет не пустой тратой времени, а скорее везением и подсказкой, потому что в этом случае ученик заметит, что за вымиранием одного поселения непосредственно следует вымирание второго, что явно указывает на причинно-следственную связь. Правильное решение задачи состоит в том, чтобы, когда наступит автоматическая пауза, открыть Панель управления и в Панели навигации убить организмы одного поселения (любого), а потом вновь запустить процесс. Вначале будет наблюдаться расцвет оставленного поселения – просто из-за освободившегося места на поле. Однако, очень 57 быстро оставленное в одиночестве поселение тоже вымрет – по причине того, что полностью доест все, что может. Ведь среда совершенно податлива, а никто больше «не вытягивает» ее обратно из состояния, которое для оставленных организмов означает голодную смерть. Общий вывод, который должен в явном виде сформулировать ученик: раз одни без других жить не смогли, значит между ними был настоящий симбиоз. Вариант Б (продвинутый). Он состоит из двух этапов (подзадач). 1-й этап. Имеется два усложняющих условия по сравнению с Вариантом А. Во-первых, тут организмы мутируют, то есть вслед за выеданием цвета фона будет происходить эволюционное приспособительное изменение их собственной окраски. Во-вторых, понижен порог конъюгации (см. урок № VIII), благодаря чему появляющиеся в результате мутаций разноцветные организмы будут образовывать разные виды. Задача запускается щелчком по этой мушке-дрозофиле. Окна появятся другие, чем в Варианте А: • Не будет окна полового состава, так как в данном случае оно не наглядно – в оба угла поля заселены раздельнополые организмы. • Зато для тех и для других представлено окно «Происхождение видов». (Это косвенная подсказка для решения второго этапа задачи). Наблюдаемые после запуска процессы будут существенно многограннее, чем в Варианте А. Самое главное отличие – в том, что кроме крупномасштабных волн циркуляции то там, то здесь будет возникать мелкое чередование организмов из поселений «SW» и «NE» – буквально на уровне отдельных пикселов. На местах, где установилось такое мелкое чередование, цвет поля (то есть состояние среды обитания) будет стабилизироваться. Вообще, это мелкое чередование, являясь результатом приспособительной эволюции цвета организмов, оказывается несравненно устойчивее крупных волн циркуляции, хотя и не столь эффектно. Объяснение этому состоит в том, что при мелком чередовании организмы вступают в более тесный симбиоз на уровне отдельных особей (как у цветкового растения и его насекомого-опылителя), тогда как крупные волны циркуляции отражают симбиоз на уровне «стад» (как у африканских копытных). 58 Как только установится такое тесное взаимодействие, вероятность случайного вымирания поселений сводится к нулю. Дальше этот эксперимент можно продолжать до бесконечности, получая совершенно неповторимые эволюционные процессы. Но для решения задачи достаточно установленного интервала в 1000 «лет». Несмотря на все перечисленные отличия, решение и общий вывод для 1-го этапа Варианта Б – в точности такие же, как для варианта А. 2-й этап. Вновь запускается с самого начала та же эволюционная заготовка. Наблюдая в течение 1000 «лет», что процесс пошел иначе чем при первом запуске, но с теми же закономерностями мелкого чередования, ученик должен поразмышлять над следующим вопросом. Если убить организмы одного симбионта, то как спасти другого при условии, что сдвигать с нуля параметр «подрост» запрещено? Когда через 1000 «лет» наступает автоматическая пауза, ученику предлагается следующее: открыть Панель управления и опять (как в 1-м этапе) целиком убить в Панели навигации поселение одного из симбионтов, но для спасения оставшегося симбионта сразу же каким-то образом изменить настройки в его поселении, чтобы при запуске дальнейшего процесса оно не вымерло от голода. Этот запуск и будет экспериментальной проверкой правильности решения. Правильное решение состоит в том, что надо создать нового симбионта взамен убитого. На практике это осуществляется так: надо выбрать в Панели навигации какой-то вид – лучше для надежности самый многочисленный (первый в списке), сделать его родом (как на уроке № IX) и присвоить ему в Панели управления такую же настройку «изобилие/пустыня», какая была у убитого поселения и, соответственно, противоположную прочим видам оставленного поселения. Жизнь будет спасена. 59 СЛОВАРЬ ТЕРМИНОВ И НАЗВАНИЙ ЖИВОТНЫХ Австралопитек – промежуточный род между человекообразными обезьянами и человеком; вымерший. Адаптация – приспособленность организма к среде обитания, включая другие организмы. Аллель – конкретная вариация гена. Каждый ген может присутствовать у разных организмов в виде множества аллелей. Американские дикобразы – не имеют прямого родства с дикобразами Старого Света; иголки приобрели независимо. Амфибии – латинское название земноводных. Класс позвоночных животных. Ананкус – род мастодонтов, близких к слонам; вымерший. Ареал – географическая территория, где распространен тот или иной вид, род или другая группа организмов. Арсинотерий – рогатый дальний родственник слонов; вымерший. Археоптерикс – древнейший обладатель настоящих птичьих перьев; вымерший. Архозавры – подкласс пресмыкающихся, включающий крокодилов, динозавров и летающих птерозавров. Афротерии – млекопитающих. исконно африканская эволюционная ветвь плацентарных Бабирусса – водолюбивая свинья с островов Юго-восточной Азии. Барапазавр – представитель зауропод – группы крупнейших динозавров. Бегемоты – семейство парнокопытных; родственниками свиней, теперь – китообразных. раньше считались ближайшими Белуха – род зубатых китов, несколько обособленный от настоящих дельфинов. Белый носорог – один из двух родов африканских носорогов; он травоядный и крупнее черного. Бесполое размножение – размножение в чистом виде, когда организм приносит потомство в одиночку, без полового процесса. 60 Бесхвостые – один из трех современных отрядов земноводных. Биологическая концепция вида – виды окончательно разграничены, когда смешанное потомство не получается, нежизнеспособно или неплодовито. Бионика – прикладная отрасль, внедряющая в технику принципы работы живых организмов. Борьба за существование (конкуренция) – следствие неуёмного размножения организмов. Главную роль играет внутривидовая борьба; она острее межвидовой. Бронтотерий – представитель вымершего семейства непарнокопытных; более родственен лошадям, но более похож на носорогов. Вараны – примитивная группа ящериц, дожившая до наших дней. Воробьиные – самая многочисленная и процветающая группа птиц. Выдровая землеройка – не выдра и не землеройка, а представитель исконно африканских насекомоядных зверей. Выдры – полуводные хищники; достигают самых крупных размеров в семействе куньих. Выхухоль русская – полуводный родственник крота, находящийся на грани исчезновения. Гавиал – род, обособившийся от настоящих крокодилов. Гаплоидность – наличие в клетках одинарного набора хромосом и, соответственно, генов. Геликоприон – вымершая хрящевая рыба, известная только по зубной спирали. Ген – элементарный наследственный задаток, представленный отрезком ДНК в хромосоме. Генетика – наука о механизмах наследственности – передачи свойств родителей потомкам. Географическая изоляция – нарушение контактов между поселениями организмов изза внешних преград, например рек. Географическое видообразование – разделение видов из-за появления внешних преград, например горных хребтов. 61 Гепард – самый быстрый бегун; единственная кошка с невтягивающимися когтями; это помогает быстрому разбегу и крутым виражам. Гермафродиты («Гермес плюс Афродита») – организмы, вырабатывающие и сперматозоиды, и яйцеклетки. Обычно сами себя не оплодотворяют. Гибрид (помесь) – потомок родителей, не связанных достаточным родством, например – из разных видов. Гиеновая собака – дикая африканская собака, похожая на пятнистую гиену только окраской. Гиеновые – характерное для Африки самое молодое семейство настоящих хищных млекопитающих. Гиенодон – представитель креодонтов – вымершего отряда хищных зверей. Гипсилофодон – птицетазовый динозавр, стоящий у основания всех динозавров, наделенных клювом. Гиракотерий – один из самых первых представителей отряда непарнокопытных; вымерший. Гистограмма – тип графика, где данные отображаются отдельными столбиками. Глазное яблоко – глаз как таковой, целиком; шаровидное, с прозрачным содержимым. Глухарь – род крупных куриных птиц, характерный для российских лесов. Голый землекоп – представитель африканского семейства грызунов землекопов; живет колониями с царицей и рабочими. Гомфотерии – группа мастодонтов, в которую входят слоны. Горностай – представитель семейства куньих. Грызуны – самый многочисленный отряд плацентарных млекопитающих. Гуаянгозавр – птицетазовый динозавр из Китая с примитивной внешней защитой. Гужевой транспорт передвижения. – различный домашние млекопитающие Дальтоники – люди с недостатками в различении цветов. 62 как средство Даманы – отряд небольших современных млекопитающих, больше всего похожих на предков слонов. Деление клеток – способ размножения клеток; в норме клетка делится за один раз не более чем на две. Дёрн – плотное сплетение корневой системы травянистых растений, препятствующее укоренению семян древесной растительности. Десмостилии – вымерший отряд, который был наиболее родственен с отрядом хоботных. Диапсиды – «настоящие» пресмыкающихся; пресмыкающихся, кроме черепах. включают всех современных Динозавры – два вымерших отряда самых больших наземных животных; относятся к пресмыкающимся. Динотерии – самая обособленная группа в отряде хоботных. Диплоидность – наличие в клетках двойного набора хромосом и, соответственно, генов. Енотовые – характерное для Америки семейство настоящих хищных млекопитающих. Естественный отбор – выживание приспособленных из конкурентов. и более успешное размножение самых Жабры – органы водного дыхания. У рыб жабры внутренние, у личинок земноводных – наружные. Жабы – семейство бесхвостых земноводных. Жвачные – высший подотряд парнопалых. переваривания растительной клетчатки. Имеют сложный желудок для Жизненный цикл – схема развития организма от оплодотворенной яйцеклетки до производства следующего поколения. Зайцеобразные – отряд плацентарных млекопитающих. Зверообразные – отдельная ветвь эволюции пресмыкающихся; включает зверозубых, от которых произошли млекопитающие. Земноводные размножения. – класс наземных четвероногих, 63 сохранивших рыбий способ Зрачок – прозрачное «окошко», пропускающее свет в глаз; снаружи выглядит черным или светящимся. Зубатые киты – подотряд китообразных, питающихся средней и крупной добычей. Игуанообразные – группа ящериц, включающая игуан, агам и хамелеонов. Икра – яйцеклетки первичноводных животных, имеющие водопроницаемую оболочку. Индрикотерий – самый большой сухопутный зверь, относится к носорогам; вымерший. Ихтиозавры – вымерший подкласс чисто морских пресмыкающихся; название переводится как «рыбоящеры». Кайнозой – последние 67 миллионов лет истории Земли. Эта эра связана с расцветом млекопитающих; включает палеогеновый, неогеновый и антропогеновый периоды. Камптозавр – один из двуногих птицетазовых динозавров, имевших плоский клюв. Кариамы – южноамериканские журавлеобразные птицы, убивающие добычу ногами. Киви – маленькая страусообразная птица с Новой Зеландии. Китообразные – отряд морских млекопитающих. Включает самых крупных животных всех времен. Клаудиозавр – вымершее полуводное пресмыкающееся; близких современных родственников не имеет. Клонирование – производство точных копий (клонов) путем бесполого размножения. Койот (луговой волк) – близкий родственник серого волка; живет в Северной Америке, напоминает шакала. Колбочки – светочувствительные клетки глаза, присутствующие в виде нескольких «сортов» для различения цветов. Колючие акулы – группа акул, сохранившая древние шипы на спинных плавниках. Компсогнат – небольшой хищный динозавр, похожий на предка птиц. Конвергенция – приобретение сходств между неродственными организмами при эволюции в сходных условиях. 64 Конкурентное вытеснение – Г.Ф. Гаузе доказал на инфузориях, что виды с одинаковыми потребностями не могут сосуществовать. Конъюгация хромосом – попарное соединение хромосом при мейозе (делении диплоидной клетки на гаплоидные). Копытные – условное название для всех млекопитающих с копытами, независимо от их родства. Копытные в основном растительноядны. Костистые рыбы – высшая группа лучеперых рыб; включает половину современных видов позвоночных. Костные рыбы – одно из двух основных подразделений рыб (второе – хрящевые). Кошачьи – семейство настоящих хищных млекопитающих. Кошкообразные – одна из двух эволюционных ветвей хищных зверей (вторая – собакообразные). Креодонты – отряд хищных млекопитающих, предшествовавший отряду современных хищных и вытесненный им. Критерии вида – морфологический (ученые разделяют виды по строению) и биологический (по пониженной скрещиваемости). Ксенартры – отряд исконных плацентарных Южной Америки с усложненным строением позвоночника. Куньи – семейство настоящих хищных млекопитающих. Куро-гусеобразные – группа отрядов современных птиц. Латимерия – род из отряда актинистий; два вида дожили до нас на противоположных краях Индийского океана. Лемуры – группа мадагаскарских семейств полуобезьян – низшего подразделения отряда приматов. Летальность – смертельность. Летальной может быть мутация, болезнь, яд и т.п. Лошадь Пржевальского – единственный сохранившийся в Азии вид диких лошадей в узком смысле слова. Лошак – гибрид от скрещивания жеребца и ослицы; не способен производить ни сперматозоиды, ни яйцеклетки. 65 Лучеперые – одна из двух эволюционных ветвей костных рыб (вторая – мясистолопастные). Макроэволюция – эволюционные процессы, наблюдаемые на уровне родов, семейств и выше. Маммут – род мастодонтов, наименее родственных слонам; подобно мамонту имел шерсть. Мамонт – недавно вымерший шерстистый слон азиатского происхождения. Мангустовые – семейство настоящих хищных млекопитающих. Мандрил – представитель рода павианов. Мара – южноамериканский грызун, внешне похожий на антилопу. Медвежьи – семейство настоящих хищных млекопитающих. Межняк – межродовой гибрид глухарки и тетерева (реже – глухаря и тетерки); практически бесплоден. Мезогиппус – род дальних предков лошади, зебры и осла. Мезозой – предпоследняя эра истории Земли, связанная с расцветом пресмыкающихся; включает триасовый, юрский и меловой периоды, длилась 180 миллионов лет. Мейоз – тип деления клетки, обеспечивающий переход от двойного набора хромосом (диплоидного) к одинарному (гаплоидному). Меланизм – черная или почти черная окраска внешних покровов тела. Мерикгиппус – род близких предков лошади, зебры и осла. Метриоринх – род чисто морских древних крокодилов. Микроэволюция – эволюционные приводящие к их разделению. процессы, происходящие внутри видов и Мозазавр – род гигантских морских ящериц из родственников современных варанов. Мозоленогие – родственники жвачных парнокопытных, но более древние; включают верблюдов и лам. Моноклониус – род однорогих рогатых динозавров. 66 Мул – гибрид от скрещивания осла и кобылы; не способен производить ни сперматозоиды, ни яйцеклетки. Мутант – носитель ярко выраженной мутации. Малозаметные мутации несут в себе практически все организмы. Мутации – случайные изменения в генах при делении клеток. Мутации в половых клетках приводят к отличиям детей от родителей. Мясистолопастные (лопастеперые) – одна из двух эволюционных ветвей костных рыб (вторая – лучеперые). Наземные ленивцы – достигали гигантских размеров и дожили до заселения Америки доисторическими индейцами. Насекомоядные – примитивный отряд плацентарных млекопитающих, из которого недавно исключены африканские представители. Наследование, наследственность – содержащаяся в половых клетках информация, обеспечивающая сходство потомства с родителями. Настоящие тюлени – ластоногие млекопитающие, полностью лишившиеся наружного уха. НЕолени – условное название виртуальных организмов, не делающих разницы между женской и мужской красотой. Неопределенная изменчивость – неожиданные отличия потомства от родителей, которые могут передаться внукам; причина – мутации. Непарнокопытные – отряд плацентарных с тенденцией к развитию основного копыта на одном среднем пальце. Нерест – размножение животных в воде посредством метания икры. Нотозавр – род морских пресмыкающихся, проводивших часть времени на берегу. Обезьяны – наиболее высокоразвитая группа в отряде приматов; отличается дневным образом жизни, включает человека. Обмен веществ – совокупность физико-химических обеспечивающая жизнь; при смерти обмен прекращается. процессов в организме, Оплодотворение – слияние двух половых клеток. Оно противоположно мейозу: из одинарных наборов их хромосом получается двойной. 67 Орангутан – азиатский род человекообразных обезьян. Осетровые – один из древнейших отрядов лучеперых костных рыб; достигают массы в 1000 кг. Офтальмозавр – переводится как «глазастый ящер»; один из самых совершенных морских ихтиозавров. Палеозой – эра в истории Земли, когда появились позвоночные; включает кембрийский, ордовикский, силурийский, девонский, каменноугольный и пермский периоды, длилась около 300 миллионов лет. Палеомастодонт – один из примитивнейших родов отряда хоботных. Палочки – светочувствительные клетки глаза, которых достаточно для черно-белого зрения; работают даже при плохом освещении. Панголины – отряд плацентарных млекопитающих; еще их называют «ящерами» за чешуйчатый покров. Пандерихт – одна из рыб, ближайших к земноводным; почти прямой предок наземных четвероногих. Пантеры – род крупных кошачьих, включающий леопарда, ягуара, льва и тигра. Панцирник – американская панцирная «щука» – пресноводная хищная рыба массой до 80 кг и выше. Паразауролоф – род утконосых птицетазовых динозавров. Парнокопытные – отряд плацентарных млекопитающих, включающий жвачных, свиней, бегемотов; от него произошел отряд китообразных. Парнопалые – группа отрядов плацентарных с тенденцией к развитию основных копыт на двух средних пальцах. Парные плавники рыб – это то, из чего у наземных позвоночных получились ноги, руки, крылья и ласты. Патология – болезненное изменение в строении и функционировании организма. Пекари – небольшие американские родственники настоящих свиней. Первые рыбы – акантодии. Они совмещали признаки хрящевых и костных рыб; характерны шипы на плавниках и между ними. 68 Песец – полярный род лисиц. Пиксел (от английского picture element) – мельчайшая точка на мониторе или на электронной картинке (например, фотографии). Платибелодон – один из самых необычных мастодонтов в отряде хоботных. Плацентарные – высшие млекопитающие; характерно длительное внутриутробное развитие зародыша. Плезиозавры – морские пресмыкающиеся, плававшие без активного участия хвоста. Плесень – так называют различные грибы, напоминающие вату и не имеющие плодовых тел, которые можно набрать в корзинку. Плиозавр – это плезиозавров с увеличенной головой и укороченной шеей. Половой диморфизм – внешне заметное, специально усиленное различие между самцами и самками одного вида. Половой отбор – составляющая естественного отбора, направленная только на самцов или только на самок. Половой процесс – скрещивание двух особей не только для размножения, но и для перетасовки их генов у потомства. Половые клетки (гаметы) – особые клетки для полового процесса; всегда гаплоидны (с одним набором хромосом). Половые признаки – любые признаки, отличающие самцов от самок и самок от самцов. Полорогие – семейство жвачных парнокопытных, включающее быков, баранов и антилоп. Полуобезьяны – низшие представители отряда приматов, включающие лемуров; в основном ночные. Популяция – обособленная часть вида (точный перевод с английского: «население», «поселение»). Пресмыкающиеся – класс или даже приспособившихся размножаться вне воды. несколько классов Приматы – отряд плацентарных млекопитающих, включающий человека. 69 четвероногих, Принцип дивергенции – изобретенный Дарвином механизм разделения видов в результате приспособительной эволюции. Проконсул – вымерший род человекообразных обезьян, близкий к прямым предкам человека. Протозух – один из первых родов древних крокодилов. Протоцератопс – один из первых родов рогатых динозавров – еще небольшой и безрогий. Пситтакозавр – вероятный предок рогатых динозавров, еще ходивший на двух ногах. Псовые – семейство настоящих хищных млекопитающих. Птерозавры – крылатые пресмыкающиеся, жившие одновременно с динозаврами и тоже относящиеся к архозаврам. Птицетазовые – один из двух отрядов динозавров (второй – ящеротазовые); включал только растительноядных ящеров. Пяденицы – семейство бабочек, в основном активных в сумерки и ночью. Раздельнополое размножение – противоположность гермафродитизму: яйцеклетки производят одни особи (самки), а спермии – другие (самцы). Рекомбинация (перекомбинация) – перетасовка родительских хромосом и генов при половом размножении. Репродуктивная изоляция – пониженная возможность скрещиваться; возможны разные причины. Рептилии – латинское название пресмыкающихся. Рогозуб – очень древний австралийский род двоякодышащих рыб, отличающийся мощными парными плавниками. Род – основная систематическая категория, объединяющая ближайшие виды. Обычно роды не могут скрещиваться. Рукокрылые – отряд плацентарных млекопитающих; включает летучих мышей и более крупных крыланов. Саблезубые тигры – древняя эволюционная ветвь кошачьих. Однако, не только кошачьи имели саблевидные верхние клыки. 70 Свинообразные – примитивные парнокопытные. Сеголетки – те, кто родился в этом году. Сепия – цвет коричневатых чернил, вырабатываемых головоногими моллюсками рода Sepia (каракатицы). Сетчатка – слой светочувствительных клеток в глазу. Симбиоз (мутуализм) – взаимовыгодные отношения между организмами разных видов. Такие организмы называют симбионтами. Синдром Дауна – одна из хромосомных патологий, возникающих в половых клетках при мейозе – потомку передается одна лишняя хромосома. Сирены – отряд растительноядных морских млекопитающих, родственных слонам. Скаты – несколько отрядов сплющенных донных акулообразных рыб. Скрывающая (покровительственная, маскировочная) окраска – делает животное незаметным для глаз любимой добычи и/или старых врагов. Скунсы – наиболее обособленная группа куньих. Собакообразные – одна из двух эволюционных ветвей хищных зверей (вторая – кошкообразные). Сперматозоиды (спермии) – половые клетки самца; характеризуются подвижностью при помощи жгутика. Стайер – выносливый бегун, пловец или летун, приспособленный преодолевать длинные дистанции. Стегозавр – род птицетазовых динозавров с характерным гребнем на спине. Стиракозавр – род рогатых динозавров со множеством шипов на голове. Сумчатые – отряд или несколько отрядов млекопитающих; первоначально появились в Америке. Сумчатый лев – крупный древесный представитель сумчатых Австралии с зубами хищника; вымерший. Сцелидозавр – четвероногий растительноядный птицетазовый динозавр с зачатками внешней защиты. 71 Тапир – самый примитивный представитель современных непарнокопытных. Тапиры – отдельное семейство. Тарбозавр – азиатский динозавр, близкий американскому тиранозавру. Это крупнейшие сухопутные хищники всех времен. Теплокровность – постоянно высокий уровень обмена веществ, сглаживающий колебания внешней температуры. Террестризух – род примитивных архозавров, близкий скорее к предкам крокодилов, чем динозавров. Тетерев – род крупных куриных птиц, характерный для российских лугов и полей. Тилакосмилус – крупный саблезубый представитель южноамериканских сумчатых; вымерший. Тли – семейство насекомых, регулярно использующих бесполое размножение: самки дают потомство без помощи самцов. У виртуальных «тлей» самцы также это делают. Тринаксодон – род настоящих зверозубых пресмыкающихся, от которых произошли млекопитающие. Трицератопс – самый известный род рогатых динозавров. Трубкозуб – единственный представитель отдельного отряда исконно африканских плацентарных. Уакари – род широконосых обезьян – исконных обезьян Южной Америки. Узконосые – обезьяны Старого Света. Уранотерии – группа исконно африканских отрядов плацентарных, включая слонов, сирен и даманов. Утконос – род яйцекладущих млекопитающих из Австралии. Ушастые тюлени – ластоногие млекопитающие, сохранившие крошечное наружное ухо. Фенек – маленькая пустынная лисица. Фенотип – «внешний вид», а точнее – все внешние и внутренние признаки организма, развивающиеся на основе унаследованных генов (генотипа). 72 Физиология – наука о жизненных процессах происходящих в организме. Форорак – гигантская нелетающая хищная птица, жившая в Южной Америке в отсутствие крупных хищных млекопитающих. Халикотерии – вымершее семейство из отряда непарнокопытных. Халикотерии имели мощные когти вместо копыт. Холоднокровность – примитивный тип обмена веществ, когда температура тела зависит от температуры среды. Хромосома – находится в клеточном ядре и включает одну молекулу ДНК как носитель наследственной (генетической) информации. Хрусталик – выполняет в глазу функцию фокусирующей линзы; похож на маленькое увеличительное стекло. Хрящевые рыбы – одно из двух основных подразделений рыб (второе – костные). Частота мутаций – мера того, как часто могут возникать мутационные изменения (например – за одно поколение и в одном конкретном гене). Человек разумный – происходит от африканских человекообразных обезьян. Самые древние народы – пигмеи, бушмены и готтентоты. Черепахи – самая обособленная группа современных пресмыкающихся. Шерстокрыл – единственный представитель отдельного отряда плацентарных млекопитающих. Широконосые – обезьяны Нового Света, точнее Южной Америки. Экологическая ниша – роль вида в сообществе окружающих видов. Экологическое видообразование – разделения видов в результате приспособительной эволюции в расходящихся направлениях. Экология – наука о взаимодействиях организмов между собой и с окружающей средой. Эласмозавр – представитель морских пресмыкающихся-плезиозавров с самой длинной шеей. Эналиарктос – древний род ластоногих с североамериканского берега Тихого океана. 73 Эндрюсарх – древний родственник парнокопытных и, вместе с тем, крупнейший сухопутный хищник из всех млекопитающих. Энтелодонты – древние родственники свиней, значительно более крупные. Эузухии – настоящие крокодилы, дожившие до наших дней. Эуплоцефал – род панцирных динозавров. Эустеноптерон – представитель мясистолопастных рыб-рипидистий, от которых в дальнейшем произошли земноводные. Ягуар – южноамериканская пантера. Яйцекладущие (однопроходные) – подкласс Первозвери – исконно австралийская группа примитивнейших млекопитающих. Яйцеклетки (яйца) – половые клетки самки; характеризуются запасом желтка для питания зародыша после оплодотворения. Ящерицы – самая многочисленная группа современных пресмыкающихся. Ящеротазовые – один из двух отрядов динозавров (второй – птицетазовые); включал двуногих хищников и четвероногих растительноядных. RGB (от английского red-green-blue) – система цветопередачи на мониторах в виде красной, зеленой и синей составляющих. 74 ОГЛАВЛЕНИЕ Урок I: «Разнообразие позвоночных животных как продукт эволюции». стр. 6 Урок II: «Жизненный цикл многоклеточных, механизм передачи наследственных признаков и конъюгации хромосом». стр. 11 Урок III: «Цветное зрение человека и обезьян и применение его принципов в практике». стр. 18 Урок IV: «Выживание наиболее приспособленных на примере скрывающей окраски. Роль мутаций в приспособительной эволюции». стр. 23 Урок V: «Эволюционное значение полового размножения». стр. 27 Урок VI: «Половой отбор и эволюционное значение разных способов размножения». стр. 30 Урок VII: «Половой отбор и эволюционное значение разных способов размножения» (продолжение для профильного уровня). стр. 35 Урок VIII: «Принцип дивергенции и экологическое видообразование». стр. 41 Урок IX: «Принцип дивергенции» (продолжение). стр. 48 Урок X: «Симбиоз. Экспериментальная контрольная работа». стр. 55 СЛОВАРЬ ТЕРМИНОВ И НАЗВАНИЙ ЖИВОТНЫХ стр. 60 75