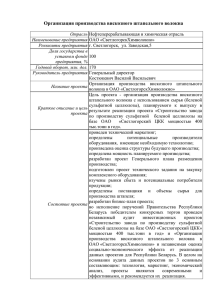
УДК 676.1.054.1 Технологии ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ
реклама

ХИМИЯ РАСТИТЕЛЬНОГО СЫРЬЯ. 2011. №3. С. 183–192. Технологии УДК 676.1.054.1 ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ НА ЭЛЕКТРОПОВЕРХНОСТНЫЕ СВОЙСТВА ЦЕЛЛЮЛОЗНЫХ ГИДРОСУСПЕНЗИЙ © А.С. Смолин1, М. Бисальски2, С. Шабель2, Р.О. Шабиев1 1 Санкт-Петербургский государственный технологический университет растительных полимеров, ул. Ивана Черных, 4, Санкт-Петербург, 198095 (Россия), e-mail: [email protected] 2 Дармштадский технический университет, Дармштадт (ФРГ) В статье приводятся данные влияния времени размола и фракционирования беленой сульфатной лиственной целлюлозы и хлопкового линтера на ζ–потенциал, поверхностный и внутренний заряд. Целлюлозные волокна размалывали в аппарате центробежного действия – мельнице Йокро. Через определенный интервал времени отбирали образец массы и отделяли от него волокнистую мелочь фракционированием. Для каждого образца определяли среднюю длину волокна, относительное водоудержание WRV, общее содержание СООН-групп, ζ–потенциал, поверхностный и внутренний заряд. Оценку поверхностного и внутреннего доступных зарядов проводили методом коллоидного индикаторного обратного титрования растворами катионных полиэлектролитов разной молекулярной массой. ζ–потенциал определяли методом потенциала протекания на установке типа Magendans при высокой общей электропроводности раствора и разной плотности набивки диафрагмы. Содержание СООН-групп устанавливали колориметрическим и алкалиметрическим методами. В результате продолжительного размола обнаружено первоначальное понижение, а затем повышение ζ–потенциала древесной и хлопковой целлюлозы, при разной плотности набивки диафрагмы и электропроводности раствора. Обнаружили зависимость ζ–потенциала от средней длины волокна: чем больше длина фракции, тем больше ζ–потенциал. Поверхностный заряд вырос только на первом этапе размола и затем не изменял своего значения, а внутренний заряд увеличивался постоянно с общим увеличением степени фибрилляции. С размолом снижается содержание СООН-групп длинноволокнистых фракций древесной целлюлозы и незначительно повышается у фракции 200 меш хлопковой. Для объяснения причины наблюдаемых электроповерхностных закономерностей целлюлозных гидросуспензий авторами предложена гипотеза «ориентированной адсорбции сольвента». Выдвинуто предположение, что взаимодействие целлюлозы с катионными полиэлектролитами зависит не столько от наличия СООН-групп, сколько от молекулярной массы полиэлектролитов, а также удельной поверхности и степени фибрилляции самой целлюлозы. Полученные данные можно использовать для оценки действия катионных полиэлектролитов и их адсорбции на этапе подготовке бумажной массы. Ключевые слова: размол, водоудержание, электрокинетический потенциал, двойной электрический слой, карбоксильные группы, коллоидное титрование Выражаем благодарность Министерству образования и науки РФ и немецкой службе академических обменов DAAD за совместную финансовую поддержку. Введение В процессе производства бумаги целлюлозные волокна подвергаются механическому воздействию – размолу. Главная цель размола заключается в улучшении бумагообразующих свойств волокон. При размоле происходит набухание (гидратация), фибрилляция и укорачивание волокон, повышается их гибкость, эти процессы ведут к увеличению удельной поверхности волокна [1]. Набухание и поглощение воды волокнистой структурой связано прежде всего с появлением дополнительных свободных ОН-групп на ее поверхности. Параллельно набуханию происходит образование большого количества волокнистой мелочи, а также имеет место высвобождение и растворение некоторых компонентов клеточной стенки S2 [2]. Хорошо известно, что удельная поверхность мелочи намного превышает удельную поверхность самого волокна, а ее присутствие не только изменяет структурные и механические свойства бумаги, но и отрицательно сказывается на расходе полезных химических вспомогательных веществ. Для отделения непосредственно волокна от мелочи и для разделения массы по длине волокна целлюлозную массу фракционируют. Процесс размола в значи Автор, с которым следует вести переписку. 184 А.С. СМОЛИН, М. БИСАЛЬСКИ, С. ШАБЕЛЬ, Р.О. ШАБИЕВ тельной степени зависит от способности волокон набухать. Считается, что чем выше содержание в целлюлозе кристаллической части, тем ниже удельная поверхность всего целлюлозного препарата, и тем ниже его способность к набуханию [3]. Размол влияет и на электроповерхностные свойства целлюлозы. Многие авторы отмечают снижение ζ–потенциала волокнистых гидросуспензий на первом этапе размола [1, 4], замечено и его повышение при продолжительном размоле [5]. Известно, что мелочь и волокно имеют разное значение ζ–потенциала при относительно небольшом различии в их химическом составе, но при значительной разнице в удельной поверхности [2]. Интересным представляется и изучение поверхностного заряда целлюлозы общими аналитическими способами [6, 7]. Титрованием катионными полиэлектролитами разной молекулярной массой можно оценить поверхностный и внутренний доступный заряд целлюлозных волокон [8]. Теория происхождения поверхностного заряда целлюлозы. Можно смело утверждать, что до сих пор не существует точного объяснения природы происхождения отрицательного заряда на поверхности целлюлозных волокон. В классическом случае возникновение двойного электрического слоя происходит вследствие установления адсорбционного равновесия в системе, приводящего к возникновению на поверхности слоя потенциалопределяющих ионов и электростатически связанных с ними противоионов [9]. В целом заряд противоионов уравновешивает заряд диффузионной области. Основные причины возникновения двойного электрического слоя [10]: вследствие преимущественной адсорбции катионов или анионов; как результат диссоциации электролитических активных групп на поверхности; ориентация диполей, входящих в состав дисперсионной среды; дефекты кристаллической решетки [8]. Еще в 30-х гг. прошлого столетия Вильбрандт [9] предположил, что электрохимическое поведение диафрагм целлюлозы и ее производных обусловлено наличием тех или иных химических активных групп на поверхности капилляров. Активными группами для нитроцеллюлозы он считал группы -NO2, а для чистой целлюлозы или ее производных с невысокой степенью замещения – OH-группы. Активность же можно оценивать по величине дипольного момента. Таким образом, он первым выдвинул предположение о причине проявляемых электроповерхностных явлений в гидросуспензиях целлюлозных волокон вследствие ориентации дипольных молекул воды. Тем не менее многие авторитетные ученые, в числе которых Линдстрём [8] и Юрьев [11], полагают, что первопричиной возникновения отрицательного заряда на поверхности целлюлозы в ее гидросуспензиях является ионизация СООН-групп и вообще диссоциация всех электролитических активных групп, как на поверхности, так и внутри волокнистой структуры. Можно отметить, что в литературе приводятся данные [4], что содержание СООН-групп в любых препаратах природной целлюлозы, ничтожно мало для создания столь сильного заряда. Более того, содержание COOH-групп при низких степенях замещения производных целлюлозы снижает по абсолютному значению ζ–потенциал [9, 10]. В результате изучения многочисленных литературных источников по исследованию ζ–потенциала целлюлозы можно считать доказанным роль поляризации поверхности за счет ориентации диполей воды в образовании двойного электрического слоя. Рис. 1. Возможное строение двойного электрического Явление ориентации молекул воды на послоя на волокнах целлюлозы, где iHp – внутренняя верхности целлюлозы можно назвать «ориплоскость Гельмгольдца; iHo – внешняя плоскость ентированной адсорбцией сольвента», авГельмгольдца; δ- –дробный отрицательный заряд δ+ – торское видение строения двойного элекдробный положительный заряд; ↑V – направление трического слоя представлено на рисунке 1. конвективного потока; ↓Е – направление В качестве целых зарядов для возникновеэлектромиграционного потока; ψо – поверхностный ния разницы потенциалов предложено счипотенциал; ψd – потенциал на границе диффузионного и тать гидроксид-ионы (ОН-) и ионы гидроадсорбционного слоя. ксония (Н3О+). ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ … 185 По-видимому, ориентация молекул воды и волокнистая структура влияют и на адсорбцию полиэлектролитов и внутренний заряд целлюлозы. Предположение о наличии потенциала Доннана между общим раствором и волокнистой мембраной вследствие высокого содержания ионногенных групп внутри волокнистой структуры [12] не выдерживает критики, поскольку такие предположения не подтверждаются данными сравнительного химического анализа содержания СООН-групп целлюлозы в гомогенной и гетерогенной среде [13]. Таким образом, можно смело утверждать, что СООН-группы в своем преимущественном большинстве сконцентрированы на поверхности волокна. Экспериментальная часть Для исследования использовали древесные и хлопковые волокна – целлюлозу сульфатную беленую лиственную (далее – ЦСФАЛБ) и хлопковый линтер (далее – линтер). Для проведения испытаний образцы подготавливали, дезинтегрировали и определяли их степень помола в соответствии с требованиями международных и национальных стандартов ISO 5263 и ГОСТ 14363.4. Приготовленные препараты целлюлозы хранились в пластмассовой таре с закрытой крышкой, в специальной холодильной камере при постоянной температуре около 6 оС не более двух недель, после чего они готовились заново. Схема исследования целлюлозы представлена на рисунке 2. Рис. 2. Схема исследования целлюлозы Для размола целлюлозы использовали аппарат центробежного действия – мельницу Йокро, соответствующую стандартам DIN 54360-04 и ГОСТ 14363.4. Процедуру определения водоудерживающей способности при ускорении 3000 g разработали на основе рекомендаций немецкого союза бумажников. Содержание СООН-групп в препаратах неразмолотой целлюлозы определяли двумя методами: колориметрически, при взаимодействии с метиленовым голубым (МГ) индикатором при разной подготовке препарата и алкалиметрическим титрованием избытка щелочи, а препараты размолотой целлюлозы – только колориметрическим способом. Методики определения содержания СООН-групп взяты из литературы [7, 13]. После размола массу фракционировали на классификаторе Бауера-МакНетта (TAPPI T233) в ячейках с разными размерами сит, например 50, 100 или 200 меш, и определяли среднюю длину волокна при помощи анализатора волокна MorFi и микроскопическим анализом при увеличении ×100. Степень фибрилляции осуществляли визуальной оценкой состояния поверхности при увеличении ×200. Электрокинетический потенциал определяли методом потенциала протекания на установке типа Magendans при разной плотности диафрагмы, для исключения влияния поверхностной проводимости измерения проводили при высокой ионной силе раствора 0,01 и 0,02 М. Ионную силу (электропроводность) регулировали добавлением кристаллической соли (NaCl). В данном методе определения ζ–потенциала раствор передвигается под влиянием градиента давления (вакуума) по капиллярам волокнистой диафрагмы, сформированной между электродами измерительной ячейки [10]. На электродах возникает ЭДС, являющаяся разностью потенциалов по обеим сторонам диафрагмы. Вычисление ζ–потенциала производится автоматически согласно формуле Гельгольца – Смолуховски (1) путем измерения общей электропроводности, вакуума и потенциала протекания, остальные значения принимаются исходя из физико-химических показателей воды при 20 °С: ζ = (4πηχ/εP)·E, мВ, (1) А.С. СМОЛИН, М. БИСАЛЬСКИ, С. ШАБЕЛЬ, Р.О. ШАБИЕВ 186 где η – коэффициент вязкости, 0,001Па·с; χ – удельная электропроводимость диафрагмы, мСм/см; ε – диэлектрическая постоянная среды, 80; Р – давление, под которым продавливается жидкость, Па; Е – потенциал протекания, мВ. Поверхностный доступный заряд, наряду с определением содержания СООН-групп, определяли титрованием высокозаряженными катионными полиэлектролитами разной молекулярной массы с ортотолуидиновым голубым (ОТГ) индикатором. Коллоидное титрование, или титрование полиэлектролитами, известно с начала 50-х гг. прошлого столетия, одним из первопроходцев этого метода считается Тераяма [15], и является одним из аналитических способов оценки заряда растворов полиэлектролитов [16]. Для определения поверхностного доступного заряда использовали два высокозаряженных катионных полиэлектролита производства компании Sigma Aldrich – 1,5-диаза-ундекаметилен полиметобромид, торговая марка – полибрен (100 мг/л), и полидиаллилдиметилхлорид аммония, торговая марка – ПДАДМАХ (156 мг/л = 0,001 н). Данные полиэлектролиты имеют узкое молекулярно-массовое распределение. Полибрен используется в медицине для увеличения эффективности заражения клетки ретровирусом и имеет молекулярную массу 8000 г/моль. ПДАДМАХ является стандартным катионным полиэлектролитом, используемым в технологии бумаги для определения катионной потребности на автоматических анализаторах заряда частиц типа SCD (Streaming Current Detector). Проводили эксперимент следующим образом. Отбирали влажный образец (~0,2 г а.с.в.) фракционированного целлюлозного препарата разной степени фибрилляции и добавляли раствор катионного полиэлектролита. После 0,5 ч отделяли фильтрат (рН=6,5) и аналитически определяли остаточное содержание катионного реагента по формуле (2) титрованием фильтрата раствором полиэтиленсульфоната натрия (PES-Na) до изменения окраски индикатора ОТГ с голубого на розоватофиолетовый цвет. Параллельно проводили титрование холостой пробы. После каждого эксперимента образец целлюлозы фильтровали и определяли точную массу высушиванием в сушильном шкафу при 105 °С. Заряд целлюлозы вычисляли по следующей формуле: С = (Х–Р)·N/m, мкмоль/кг, (2) где Х – расход PES-Na на титрование холостой пробы, мл; Р – расход PES-Na на титрование препаратов целлюлозы, мл; N – концентрация PES-Na, 0,001 н; m – масса целлюлозного препарата, г. Обсуждение результатов Оценка содержания СООН-групп. Результаты всех методов определения содержания СООН-групп показывают снижение их количества с ростом чистоты целлюлозного препарата. Самое низкое значение наблюдается у хлопкового линтера, самое высокое – у механической массы (беленой химикотермомеханической массы БХТММ). Хвойная целлюлоза (сульфатная беленая – ЦСФАХБ) содержит меньше СООН-групп, чем лиственная, вследствие более низкого содержания гемицеллюлоз [1]. Основными источниками СООН-групп в древесной целлюлозе выступают глюкуроновые и галактуроновые кислоты, несравнимо меньшая доля ионногенных кислотных групп приходится на окисленный лигнин и природное содержание самой целлюлозы. Полученные результаты совпадают с литературными данными [2, 8]. Из таблицы 1 следует, что, несмотря на разные цифры содержания СООН-групп, получаемые разными аналитическими методами, результаты определения всех методов демонстрируют одинаковую тенденцию. Следует учитывать, что результаты любого химического анализа содержания СООН-групп целлюлозы в ее гетерогенном состоянии нельзя проверить, так как не существует препарат целлюлозы с точно известным количеством этих групп. Метод определения содержания СООН-групп следует выбирать в зависимости от типа целлюлозных волокон, к примеру, алкалиметрический способ не подходит для анализа полуфабриката высокого выхода (см. табл.1). Таблица. 1. Содержание СООН-групп в разных видах целлюлозных волокон, ммоль/кг Волокнистый полуфабрикат ЦСФАЛБ ЦСФАХБ Линтер БХТММ Алкалиметрическим (гидрокарбонатным) титрованием 29.4 16.9 6.5 38,3 Колориметрическим методом Последовательным разбавлением Центрифугированием 39.7 30.7 19.2 26.6 5.9 4.7 93,3 97,9 ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ … 187 Связь времени размола, степени помола и водоудержания. Приведенные на рисунке 3 результаты показывают, что степень помола целлюлозы постепенно увеличивается со временем размола, причем фракционированная масса набирает степень помола медленнее (фракция 50 меш медленнее фракции 100 меш) по сравнению с общей массой. После набухания и дезинтегрирования лиственной целлюлозы начальная степень помола составила 15 оШР для общей массы (длина волокна l=0,91 мм), 12 оШР для фракции 100 меш (l=0,98 мм) и 11 oШР для фракции 50 меш (l~1,18 мм). Таким образом, степень помола снижается эквивалентно увеличению средней длины волокна фракции. Через 100 мин размола степень помола общей массы достигла 80 oШР, что связано в первую очередь с фибрилляцией основной фракции при одновременном увеличении содержания мелочи и общем снижении средней длины волокна, что видно из данных рисунка 5. Степень помола фракционированной массы, т.е. массы с постоянной средней длиной волокна, выросла незначительно, до 25 oШР для фракции 100 меш (l=0,92 мм) и до 22 oШР для фракции 50 меш (l=1,11 мм) соответственно. Последующий размол только подтверждает продолжение подобной зависимости. Такое действие размола определено исключительно фибрилляцией волокна, поскольку коротковолокнистая фракция и волокнистая мелочь удаляются при фракционировании. Полученные данные подтверждают сильное влияние содержания в массе мелочи на результаты определения степени помола [2] и ошибочность объяснения повышения степени помола только с позиции возрастания фибрилляции. Таким образом, степень помола не является надежным показателем оценки степени фибрилляции и коллоиднохимического состояния массы. Степень помола следует рассматривать только как технологический показатель общего состояния бумажной массы. В качестве наиболее доступной оценки удельной поверхности и общего объема пор волокнистого полуфабриката в ЦБП выступает определение относительного водоудержания при центрифугировании (WRV). На рисунке 4 представлена динамика изменения водоудерживающей способности от времени размола для двух типов целлюлозных волокон. Из рисунка 4 следует, что лиственная целлюлоза имеет большую водоудерживающую способность по сравнению с хлопковой. Более высокие значения водоудержания волокна отвечают за более развитую удельную поверхность самой целлюлозы [3]. Как видно из графика, вначале процесса размола превалирует процесс фибрилляции и увеличения водоудерживающей способности, т.е. доступности воде ОН-групп, а затем водоудерживающая способность начинает снижаться, по-видимому, из-за частичного разрушения фибриллярной структуры и макрокапилляров, сильного укорачивания волокна и общей деструкции микрофибрилл. Такое предположение подтверждается данными микроскопических исследований (рис. 6). Физических смысл отсутствия значительного прироста гидрофильности обеих фракций лиственной целлюлозы после 100 мин размола заключается в достижении так называемой точки насыщения (Fiber Saturation Point), при которой степень фибрилляции уравновешивается с потерей прочности волокнистой структуры. Рис. 3. Изменение степени помола ЦСФАБЛ от времени размола в мельнице Йокро: фракционированная (1) и нефракционированная масса (2); фракция 50 меш (▼), фракция 100 меш (▲) Рис. 4. Изменение водоудержания волокнистого полуфабриката от времени размола – линтер (1), ЦСФАЛБ, фракция 100 меш (2а), ЦСФАЛБ, фракция 50 меш (2б) 188 А.С. СМОЛИН, М. БИСАЛЬСКИ, С. ШАБЕЛЬ, Р.О. ШАБИЕВ Оценка средней длины волокна и степени фибрилляции. Как отмечалось ранее, ζ-потенциал мелочи и волокна различается, что, скорее всего, связано с различием их геометрических размеров и удельных поверхностей. Поэтому представляет определенный научный интерес изучить электроповерхностные свойства фракций одинаковой длины, где волокнистая мелочь не будет влиять на плотность диафрагмы и, соответственно, на результаты определения потенциала протекания, а также снижать точность определения поверхностного заряда целлюлозы, повышая общую адсорбцию катионных полиэлектролитов. В качестве объекта исследования была выбрана масса, оставшаяся на сетке с размерами 200 и меньше меш, поскольку общепринято [2], что фракция массы, проходящая через отверстия сита в 200 меш на классификаторе Бауера-МакНетта, составляет волокнистую мелочь, представляющую собой остатки волокон, их фрагменты или просто пучки микрофибрилл. Обычно волокна этой фракции имеют в длину менее 150 мкм с незначительным различием поперечных и продольных размеров, в технологии такую фракцию называют «мельштофом». Приведенные на рисунке 5 данные совпадают со значениями, полученными Кларком [17], и показывают разницу в длине волокна лиственной и хлопковой целлюлозы в зависимости от времени размола и размера сита. За 200 мин размола значительно снижается средняя длина хлопкового линтера от 1,69 до 0,35 мм, по-видимому, за счет рубки еще на первом этапе размола, а затем длина сохраняется на стабильном значении. После размола и фракционирования лиственной целлюлозы средняя длина изменилась незначительно от 0,98 до 0,81 мм, причем длина фракции, не прошедшей через сетку 50 меш, больше, чем через сетку 100 меш, что объРис. 5. Влияние времени размола на длину волокна ясняется различными размерами отверстий сетки 50 а) данные микроскопического анализа; б) данные меш – 0,297 мм против сетки 100 меш – 0,149 мм. анализатора волокна; 1 – линтер; 2 – ЦСФАЛБ Степень фибрилляции оценивали визуально 100 меш; 3 – ЦСФАЛБ 50 меш осмотром поверхностного состояния волокон при увеличении ×200 раз (рис. 6). У линтера перед размолом обнаружили сильное укорачивание и фибрилляцию отдельных волокон с заметным повреждением поверхности. Волокна же лиственной целлюлозы до размола были прозрачными и имели ненарушенную и гладкую поверхность. В результате продолжительного размола в обоих случаях заметно повысилась ширина волокон, появились отдельные волокнистые фрагменты и пучки вырванных микрофибрилл. Одновременно происходило исчезновение контура волокна. По мере продолжения размола многие волокна оказывались все более разорванными, вырванные пучки становились все длиннее и выразительнее. В последней точке размола становилось все сложнее и сложнее обнаруживать недеформированные участки поверхности волокна при сохранении средней длины, некоторые волокна претерпели значительное фибриллирование. Из предложенного описания, можно сделать вывод, что волокна лиственной целлюлозы уже после 100 мин размола переходят в высокофибриллированное состояние, а фибрилляция линтера при данном типе воздействия протекает значительно сложнее, нежели его укорачивание. а) б) Рис. 6. Фотографии волокон целлюлозы лиственной (200 мин размола, 100 меш, ×100 раз) (а) и линтера (300 мин размола, 200 меш, ×200) (б) ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ … 189 Влияние времени размола на содержание СООН-групп. Из приведенных на рисунке 7 данных видно, что содержание СООН-групп со временем размола снижается как в лиственной, так и в хвойной фракционированной целлюлозе. Данные изменения содержания СООН-групп в хвойной целлюлозе представлены для сравнения и проверки результатов. Видно, что у хвойной целлюлозы их количество снижается быстрее лиственной. Можно предположить, что это явление вызвано удалением нецеллюлозных компонентов клеточной стенки, которые уходят вместе с мелочью при фракционировании. К примеру, волокнистая мелочь содержит больше гемицеллюлозы и лигнина – основных носителей анионных групп [2]. Волокна хлопковой целлюлозы, наоборот, показывают незначительное повышение содержания СООН-групп со временем размола, что можно объяснить окислением макромолекул кислородом воздуха. Влияние времени размола на доступный поверхностный и внутренний заряд. Как видно из рисунка 8, с размолом целлюлозы увеличивается адсорбция высокозаряженных катионных полиэлектролитов разной молекулярной массы. Адсорбция полибрена, который может проникать в менькие поры, выше адсорбции ПДАДМАХа, который адсорбируется только на поверхности. Причем древесная целлюлоза, имея большую удельную поверхность, поглощает больше полибрена по сравнению с хлопковой, поэтому и заряд у нее выше. То же наблюдается и при исследовании адсорбции полиэлектролита средней молекулярной массы – ПДАДМАХа. Причем до размола адсорбция ПДАДМАХа на хлопковой целлюлозе отсутствует вовсе. Размол повышает адсорбцию ПДАДМАХа, однако только на первом этапе размола, после чего адсорбция обоих типов целлюлозы больше не повышается. По-видимому, из-за большой молекулярной массы ПДАДМАХ не может проникнуть в клеточную стенку, а в отсутствие активных ионногенных центров анионной природы, как в случае с хлопковой целлюлозой, и вовсе не дает возможности задержаться его макромолекулам на поверхности. При размоле, как было показано, общее содержание ионногенных групп не возрастает, разве что в случае хлопковой целлюлозы, зато возрастает степень фибрилляции. Таким образом, адсорбция катионных полиэлектролитов связана только с изменением общей пористости, фибрилляции и общем развитии удельной поверхности, а не только содержанием СООН-групп. Полученные результаты согласуются с литературными данными по колориметрическому определению содержания СООН-групп при размоле волокнистого полуфабриката [14], однако расходятся с данными кондуктометрического титрования [18]. Нгуэн с коллегами [18] считают, что с размолом общий заряд, определенный титрованием полиэлектролитами, не изменяется, а изменяется только поверхностный. Их результат определения общего содержания СООН-групп выглядит несколько завышенным – 134 ммоль/кг, и отличается от поверхностного заряда (49 ммоль/кг) почти в три раза, что не согласуется с данными определения содержания СООН-групп целлюлозы в гомогенной и гетерогенной среде, полученными прямыми химическими методами [13]. Они показывают, что разница между поверхностным и общим содержанием СООН-групп (поверхностный заряд) не превышает 25%, таким образом можно смело утверждать, что большая часть этих групп и вообще всех ионногенных кислотных групп содержится в наружном слое клеточной стенки. Рис. 7. Влияние размола на содержание СООНгрупп фракционированных препаратов целлюлозы (первый столбик ряда ЦСФАЛБ относится к фракции 50 меш, а второй – к фракции 100 меш; первый столбик ряда ЦСФАХБ относится к фракции 30 меш, а второй – к фракции 50 меш; линтер – 100 меш) Рис. 8. Изменение доступного (поверхностного) и внутреннего заряда линтера 200 меш (1) и лиственной целлюлозы 100 меш (2) при размоле в мельнице Йокро при адсорбции катионных полиэлектролитов разной молекулярной массы; (а) C (ПДАДМАХ) 156 мг/л, (б) С (полибрен) 100 мг/л, рН=6,5 А.С. СМОЛИН, М. БИСАЛЬСКИ, С. ШАБЕЛЬ, Р.О. ШАБИЕВ 190 Влияние фракционирования на ζ-потенциал. Несомненно, длина волокна должна влиять на ζ-потенциал, для оценки влияния длины волокна использовали общую массу и фракции 50 и 100 меш лиственной целлюлозы. В таблице 2 приведены результаты определения ζ-потенциала целлюлозных волокон с фракционированием и без него. Приведенные результаты подтверждают, что на значение ζ-потенциала оказывает влияние не только плотность упаковки целлюлозной диафрагмы и электропроводность, но и длина волокна. Чем выше средняя длина волокна, тем выше и ζ-потенциал, причем при одинаковой плотности диафрагмы тенденция сохраняется, но разница заметно снижается. Можно выдвинуть гипотезу, что такая зависимость связана с изменением радиуса капилляров диафрагмы, т.е. длина волокон влияет на форму их упаковывания в волокнистой диафрагме, в то же время не стоит забывать, что волокна 50 меш содержат меньше СООН-групп и являются химически более чистыми. Таблица 2. Влияние длины волокна и плотности диафрагмы на ζ-потенциал – 100 меш Средняя длина волокна, мм 0,91 0,98 50 меш ~1,18 Номер сетки ζ-потенциал, мВ -50,2 -45,4 -52,6 -45,8 -57,4 Электропроводность, мСм/см 1,25 1,25 1,25 1,25 1,25 Плотность диафрагмы, г/см3 <0,2 0,2 <0,2 0,2 <0,2 Влияние времени размола на ζ-потенциал. Оценку влияния времени размола на ζ-потенциал проводили на примере лиственной целлюлозы. После размола целлюлозы определяли ζ–потенциал как общей массы, так и фракционированной массы 100 меш при размоле в мельнице Йокро со сточенной гарнитурой. Фракционировали для отделения длинноволокнистой фракции от мелочи, поскольку они отличаются по свойствам, к примеру, мелочь обладает большей удельной поверхностью, лучше набухает и медленнее обезвоживается. Из-за незначительного различия их поперечных и продольных размеров частички мелочи не обладают волокнистой формой. Хотя стоит отметить их положительное влияние на формование равномерной макроструктуры бумажного полотна. Измерение потенциала протекания проводили как при постоянной плотности диафрагмы, так и при произвольной, заведомо меньшей. Для исключения влияния поверхностной электропроводности использовали массу с высокой общей электропроводимостью. Электропроводимость регулировали добавлением кристаллической соли (NaCl). Данные, представленные на рисунке 9 показывают, что с увеличением времени размола на первом его этапе ζ-потенциал снижается, а затем несколько возрастает по абсолютному значению. Такое поведение можно объяснить следующим образом. При размоле происходит разрыв водородных связей между микрофибриллами целлюлозы (фибрилляция), происходит набухание волокна, увеличивается степень дисперсности гидросуспензии и снижается средняя длина волокна (укорачивание). Расширение гидратационного слоя вызывает перемещение границы скольжения вглубь диффузионного слоя, а такое перемещение снижает ζ-потенциал. Очень интересным является факт увеличеРис. 9. Изменение ζ–потенциала ния ζ-потенциала при длительном времени размофракционированной беленой сульфатной ла. Из-за высокой общей электрической проводилиственной целлюлозы (1, 2, 3) и линтера от мости раствора его очень сложно объяснить только времени размола в мельнице Йокро; 1 – 100 меш, ростом плотности целлюлозной диафрагмы и, соχ=2,2 мСм/см, ρ<0,2 г/см3; 2 – χ=1,25 мСм/см, ответственно, возрастанием вклада поверхностной ρ=0,2 г/см3; 2a – 50 меш, 2b – 100 меш; проводимости в рассчитанное значение электроки3 – χ=1,25 мСм/см, ρ<0,2 г/см3, 3а – 50 меш, 3b – нетического потенциала. Авторы считают, что рас100 меш; 4 – линтер 200 меш, χ=2,2 мСм/см, крытие поверхности увеличивает гидратацию и ρ<0,2 г/см3 приводит к большей ориентации диполей воды на ВЛИЯНИЕ РАЗМОЛА И ФРАКЦИОНИРОВАНИЯ … 191 поверхности целлюлозы и общему росту поверхностного заряда. Таким образом, влияние времени размола (увеличение удельной поверхности) на ζ-потенциал целлюлозы определяется двумя параллельно протекающими и конкурирующими процессами: увеличением гидратации (снижение ζ-потенциала) и дополнительной ориентацией диполей воды (увеличение ζ-потенциала). Плотность набивки только влияет на величину ζ-потенциала, а не на сам характер изменения. Возрастание гидратации связано с увеличением доступных для образования водородных связей с молекулами воды ОН-групп, общим увеличением удельной поверхности и снижением содержания СООН-групп. Вкладом поверхностной проводимости можно пренебречь из-за высокого содержания соли и постоянной плотности набивки, хотя поверхностная проводимость должна возрастать со временем размола. Значительное повышение электропроводности раствора (рис. 9, кривые 1, 4) снижает проявление электрокинетических свойств, хотя все равно подтверждает общую зависимость. Результаты влияния времени размола на ζ-потенциал нефракционированной массы были получены ранее [5] и совпадают с приведенными выше. Поскольку характер измерения и даже значения ζ-потенциала в обоих случаях совпадают, то можно сделать вывод, что волокнистая мелочь не оказывает сильного влияния на значения ζ-потенциала. Почти все авторы, которые занимались изучением электрокинетических явлений в целлюлозных гидросуспензиях, отмечали схожую тенденцию снижения ζ-потенциала при размоле, но только на первом его этапе, не проводя таких измерений при продолжительном воздействии размола [1, 13]. Выводы Присутствует тенденция снижения ζ–потенциала со временем размола на первом этапе, и незначительное его повышение при продолжительном воздействии. При этом снижается содержание СООН-групп у фракционированной лиственной целлюлозы и увеличивается у хлопкового линтера. При фракционировании обнаружено, что большей длине волокна соответствует большее значение ζ–потенциала. При размоле также увеличиваются поверхностный и внутренний заряды целлюлозы, определяемые адсорбцией катионных полиэлектролитов разной молекулярной массы, причем внутренний заряд увеличивается непрерывно, а поверхностный – только на первом этапе размола. На основании полученных данных можно заключить, что взаимодействие целлюлозных волокон с полиэлектролитами обусловлено не только действием чисто электростатических сил, но, в том числе, и действием адсорбционных сил и связано с удельной поверхностью целлюлозного препарата. Список литературы 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. Фляте Д.М. Свойства бумаги. Изд. 3-е, исправ. и доп., М., 1986, 680 с. Papermaking science and technology / ed. H. Paulapuro and J. Gullichsen, in 19 v., FAPET/TAPPI, Нelsinki, 1998– 2002. Krässig H.A., Cellulose Structure, Accessibility and Reactivity, Gordon and Breach Science Publishers, South Africa, 1993, 376 p. Аксельрод Г.З., Смолин А.С., Иванов М.А., Фомина М.Л., Ходырева Н.В. О природе электрокинетического потенциала целлюлозы // Cб. трудов ВНИИБа. 1973. Вып. 65. С. 69–77. Смолин А.С., Шабиев Р.О., Яккола П., Исследование дзета-потенциала и катионной потребности волокнистых полуфабрикатов // Химия растительного сырья. 2009. №1. С. 177–184. Scott W.E. Principles of Wet End Chemistry, TAPPI Press, Atlanta, 1996. Оболенская А.В., Ельницкая З.П., Леонович А.А. Лабораторные работы по химии древесины и целлюлозы. М., 1991. 320 с. Lindström T., Eklund D., Paper chemistry, an introduction, Grankulla, Finland, 1991. 306 p. Григоров О.Н. Электрокинетические явления. Л., 1973. 199 с. Никитин Н.И. Химия древесины и целлюлозы. М.; Л., 1962, 708 с. Юрьев В.И. О поверхностном (термодинамическом) потенциале целлюлозных волокон // Межвуз. сб. науч. трудов. Л., 1980. №6. С. 44–46. Hubbe M. Electrokinetic potential of Fibers // Bioresources. 2006. N1. Pp. 116–149. Klemm D., Philipp B., Heinze T. Comprehensive Cellulose Chemistry. Volume 1, Fundamentals and Analytical Methods, 2004, 380 p. Untersuchungen zum chemischen Wasserrückhaltevermögen und zur Trocknungsfähigkeit von Papierstoffen unter besonderer Berücksichtigung der Rolle von chemischen Additiven, Dissertation von K.R. Stumm, TU Darmstadt, 2007. 177 s. Terayama H. Method of Colloid Titration // Journal of Polymer Science. 1951. N8. Pp. 243–253. 192 А.С. СМОЛИН, М. БИСАЛЬСКИ, С. ШАБЕЛЬ, Р.О. ШАБИЕВ 16. Raven A. von, Högerl J. Analytik von Stärke // Wochenblatt für Papierfabrikation. 1991. N4. Pp. 122–127. 17. Кларк Дж. Технология целлюлозы. М., 1983. 456 с. 18. Bhardwaj N.K., Hoang V., Nguyen K.L. Effect of refining on pulp surface charge accessible to polydadmac and FTIR characteristic bands of high yield kraft fibers // Bioresource Technology. 2007. V. 98. Pp. 962–966. Поступило в редакцию 29 июня 2011 г. После переработки 5 августа 2011 г.